
Why cells move messages: The biological functions of mRNA localization
7
Seminars in Cell & Developmental Biology 18 (2007) 171–177 Review Why cells move messages: The biological functions of mRNA localization Tung-Gia Du, Maria Schmid, Ralf-Peter Jansen ∗ Department of Chemistry and Biochemistry, Gene Center, Ludwig-Maximilians-University Munich, Feodor-Lynen-Str. 25, D-81377 Munich, Germany Available online 6 February 2007 Abstract RNA localization is a widespread mechanism that allows cells to spatially control protein function by determining their sites of synthesis. In embryos, localized mRNAs are involved in morphogen gradient formation or the asymmetric distribution of cell fate determinants. In somatic cell types, mRNA localization contributes to local assembly of protein complexes or facilitates protein targeting to organelles. Long-distance transport of specific mRNAs in plants allows coordination of developmental processes between different plant organs. In this review, we will discuss the biological significance of different patterns of mRNA localization. © 2007 Elsevier Ltd. All rights reserved. Keywords: Morphogen; Cell fate determinant; -Actin; RNA transport; Asymmetric cell division Contents 1. mRNA localization and the formation of protein gradients ................................................................... 172 2. mRNA localization and asymmetric distribution of cell fate determinants ..................................................... 172 3. mRNA localization and local assembly of protein complexes ................................................................ 173 4. mRNA localization to the vicinity of organelles ............................................................................ 174 5. Long distance transport of mRNAs and small RNAs in plants ................................................................ 175 6. Outlook ................................................................................................................ 175 Acknowledgements ..................................................................................................... 175 References ............................................................................................................. 175 Why do cells target specific mRNAs to particular cytoplas- mic domains? It seems obvious even to the non-specialist that localization of an mRNA should result in local synthesis of the encoded protein at the target site. However, experimental evi- dence for the necessity to target a certain mRNA to a distinct site has been achieved only for a limited number of cases. This is in part due to the use of high-throughput methods to identify localized mRNAs [1,2]. For many localized mRNAs that were identified in high-throughput approaches or by co-purification with known RNA-localizing proteins we still lack functional data, mainly because disturbing their localization did not result ∗ Corresponding author. Tel.: +49 89 2180 76903; fax: +49 89 2180 76948. E-mail address: [email protected] (R.-P. Jansen). in a detectable phenotype (e.g. Ref. [3]). In other cases, localized mRNAs were identified in organisms in which the biological function of RNA localization cannot easily be addressed because they miss the elaborate genetic tools that are at hand in model organisms. Due to these restrictions most – but not all – of our knowledge on the function of mRNA localization stems from analysis of localized mRNAs in mouse, Drosophila or yeast. In this review, we will focus on the functional significance of mRNA localization. For detailed description of the mechanisms of mRNA localization or signals that confer localization we refer to the review articles by Palacios (page 163) and Bullock (page 194) in this issue. Why localize mRNAs? In his first survey of intracellular mRNA localization, Daniel St Johnston has already given several possible answers [4]. Reducing transport costs is one of them. In multiple rounds of translation, one mRNA molecule can give rise 1084-9521/$ – see front matter © 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.semcdb.2007.01.010
-
Upload
uni-tuebingen -
Category
Documents
-
view
5 -
download
0
Transcript of Why cells move messages: The biological functions of mRNA localization

A
etob©
K
C
mledsiliwd
1d
Seminars in Cell & Developmental Biology 18 (2007) 171–177
Review
Why cells move messages: The biologicalfunctions of mRNA localization
Tung-Gia Du, Maria Schmid, Ralf-Peter Jansen ∗Department of Chemistry and Biochemistry, Gene Center, Ludwig-Maximilians-University Munich,
Feodor-Lynen-Str. 25, D-81377 Munich, Germany
Available online 6 February 2007
bstract
RNA localization is a widespread mechanism that allows cells to spatially control protein function by determining their sites of synthesis. Inmbryos, localized mRNAs are involved in morphogen gradient formation or the asymmetric distribution of cell fate determinants. In somatic cellypes, mRNA localization contributes to local assembly of protein complexes or facilitates protein targeting to organelles. Long-distance transportf specific mRNAs in plants allows coordination of developmental processes between different plant organs. In this review, we will discuss theiological significance of different patterns of mRNA localization.
2007 Elsevier Ltd. All rights reserved.
eywords: Morphogen; Cell fate determinant; �-Actin; RNA transport; Asymmetric cell division
ontents
1. mRNA localization and the formation of protein gradients . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1722. mRNA localization and asymmetric distribution of cell fate determinants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1723. mRNA localization and local assembly of protein complexes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1734. mRNA localization to the vicinity of organelles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 174
5. Long distance transport of mRNAs and small RNAs in plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1756. Outlook . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 175. . . .
. . . .
imftoka
mot1
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Why do cells target specific mRNAs to particular cytoplas-ic domains? It seems obvious even to the non-specialist that
ocalization of an mRNA should result in local synthesis of thencoded protein at the target site. However, experimental evi-ence for the necessity to target a certain mRNA to a distinctite has been achieved only for a limited number of cases. Thiss in part due to the use of high-throughput methods to identifyocalized mRNAs [1,2]. For many localized mRNAs that weredentified in high-throughput approaches or by co-purificationith known RNA-localizing proteins we still lack functionalata, mainly because disturbing their localization did not result
∗ Corresponding author. Tel.: +49 89 2180 76903; fax: +49 89 2180 76948.E-mail address: [email protected] (R.-P. Jansen).
mpm
084-9521/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.oi:10.1016/j.semcdb.2007.01.010
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 175
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 175
n a detectable phenotype (e.g. Ref. [3]). In other cases, localizedRNAs were identified in organisms in which the biological
unction of RNA localization cannot easily be addressed becausehey miss the elaborate genetic tools that are at hand in modelrganisms. Due to these restrictions most – but not all – of ournowledge on the function of mRNA localization stems fromnalysis of localized mRNAs in mouse, Drosophila or yeast.
In this review, we will focus on the functional significance ofRNA localization. For detailed description of the mechanisms
f mRNA localization or signals that confer localization we refero the review articles by Palacios (page 163) and Bullock (page94) in this issue.
Why localize mRNAs? In his first survey of intracellularRNA localization, Daniel St Johnston has already given several
ossible answers [4]. Reducing transport costs is one of them. Inultiple rounds of translation, one mRNA molecule can give rise

1 evelo
tsepmrpgcatilmspftetp
1g
Tdapmiietpbtdtoptdrgtbatgi(gsos
co
celfejoo[tf
2c
runremidi
fmcicbttiitciccdacraeiTf
72 T.-G. Du et al. / Seminars in Cell & D
o several protein molecules. Therefore, localization of mRNAhould be more cost-efficient than protein transport. This mightxplain why e.g. specific mRNAs have been detected in cyto-lasmic areas surrounding the nucleus (perinuclear region) oritochondria (perimitochondrial region; see below). Another
eason why mRNA rather than protein is localized is that therotein must not be translated at any other location than the tar-et site. One such example is myelin basic protein (MBP), aomponent of the myelin sheath of oligodendrocytes that wrapsround axons of neurons [5]. As MBP sticks to membranes,ransport of the protein via the secretory pathway would resultn a collapse of the intracellular membrane system. Local trans-ation of MBP from its mRNA that has been targeted to the cell’s
yelinating processes circumvents this problem. Localized ver-us non-localized translation also allows spatial fine-tuning ofrotein synthesis and distribution. This might be especially use-ul if protein synthesis has to respond rapidly to signals, as it ishe case for postsynaptic sites in dendrites of neurons (see Dahmt al. in this issue). Furthermore, mRNA localization can serveo set up protein gradients, or to establish and maintain cellularolarity or cell diversification.
. mRNA localization and the formation of proteinradients
Gradients of morphogens pattern the embryo body plan [6].hey can act as regulators of gene expression that switch onifferent sets of genes dependent on their concentration alongdefined embryonic axis. The first molecularly defined mor-
hogen, Bicoid in Drosophila embryos is encoded by a localizedRNA (for a recent review see Ref. [7]). Bicoid mRNA is local-
zed during oogenesis to the anterior tip of the oocyte wheret is anchored. After fertilization, it is translated and a gradi-nt of Bicoid protein forms with the highest concentration athe anterior. Bicoid can then enter the nuclei in the anteriorart of the embryo and switch on its target genes [8]. Propericoid mRNA targeting is of crucial importance in this sys-em since all mutants with affected bicoid localization showefects in embryonic patterning [7]. While bicoid localiza-ion specifies the anterior end of the embryo, localization ofskar mRNA to the opposite pole initiates formation of polelasm at the posterior of the embryo [9]. The pole plasm con-ains several other localized mRNAs that are essential for laterevelopment of the primordial germ cells [10]. One of the poste-iorly localized mRNAs is nanos mRNA, which upon translationives rise to a second morphogen gradient that defines pos-erior structures of the embryo [11]. However, in contrast toicoid, nanos mRNA is not exclusively localized to the posteriornd gradient formation is mainly achieved by restricting nanosranslation to the posterior end [12]. A third localized mRNA,urken, defines the dorsoventral axis of the embryo (reviewedn Ref. [13]). gurken encodes a transforming growth factor �TGF�) like signalling protein. During late oocyte development
urken is targeted by a two-step mechanism to the anteriodor-al corner of the oocyte adjacent to the cell nucleus. Secretionf the protein made from gurken mRNA at the anteriodor-al corner induces a secondary signal in the adjacent folliclepna
pmental Biology 18 (2007) 171–177
ells that eventually establishes the dorsoventral axis of theocyte.
Localized mRNAs cannot only induce the formation of intra-ellular protein gradients but can also facilitate the formation ofxtracellular signal gradients. In the Drosophila embryo, Wing-ess is a secreted glycoprotein that functions to establish cellates during segmentation of the ventral epidermis [14]. Inter-stingly, in cells that express wingless (wg), the mRNA localizesust below the surface of the apical membrane [15]. Mistargetingf wg mRNA to the basal cell membrane results in an embry-nic phenotype similar to that observed upon loss of wg function15]. This suggests that mRNA localization to the apical side ofhe cell facilitates proper secretion and extracellular gradientormation.
. mRNA localization and asymmetric distribution ofell fate determinants
Asymmetric cell divisions are widespread in the bacte-ial and eukaryotic kingdoms [16–18]. Generally this achievesnequal distribution of different cellular components includingucleic acids and proteins. Numerous mRNAs are asymmet-ically distributed upon cell division, especially during earlymbryogenesis. For many, however, the reason for their asym-etric partitioning is not understood (see, e.g., Ref. [19]). Even
n cases where the encoded protein functions as a cell fateeterminant, the biological significance of the localization ofts mRNA localization is not immediately obvious [20].
The currently best-understood example that provides a directunctional link between mRNA localization and cell fate deter-ination can be found in the budding yeast Saccharomyces
erevisisae (Fig. 1A). Here, the Ash1 protein has a key functionn the regulation of mating-type switching [21]. This occurs afterell division in only one of the two resulting cells and is initiatedy the site-specific restriction endonuclease HO that cleaves inhe actively expressed mating type locus. HO expression is con-rolled temporally and spatially [21]. The protein is made onlyn mother (i.e. the cell that has given rise to the bud) but notn daughter cells (i.e. the mature buds). Ash1p is a transcrip-ional repressor that blocks expression of HO in the daughterell and thus controls spatial distribution of HO protein. ASH1s expressed prior to cell division and accumulates in daughterell nuclei due to the localization of its mRNA to the daughterell tip [21–23]. ASH1 mRNA mis-localization results in equalistribution of the protein in nuclei of mother and daughter cellsnd, consequently, both cells acquire an identical fate [21,22]. Inontrast to ASH1 there are a series of localized mRNAs whoseeason for localization we do not understand. In total, 25 mRNAsre localized to the bud or daughter cell tip by a common machin-ry [3,24]. However, for most of them targeting to the bud tips not necessary to achieve asymmetric protein distribution [3].his indicates that mRNA localization in these cases might either
acilitate protein sorting or functions as backup mechanism.
A similar backup strategy seems to exist in the case ofrospero and inscuteable mRNAs in asymmetrically dividingeuroblasts in Drosophila embryos [20,25]. Neuroblasts usu-lly divide along the apical-basal axis into a ganglion mother
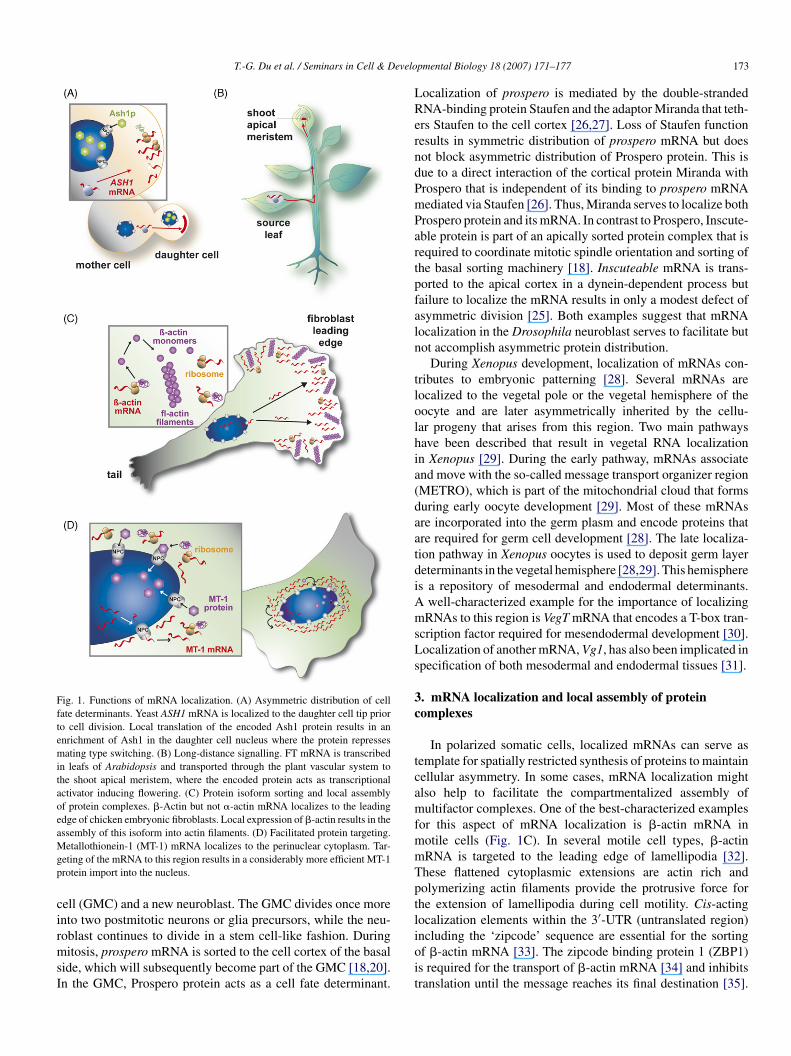
T.-G. Du et al. / Seminars in Cell & Develo
Fig. 1. Functions of mRNA localization. (A) Asymmetric distribution of cellfate determinants. Yeast ASH1 mRNA is localized to the daughter cell tip priorto cell division. Local translation of the encoded Ash1 protein results in anenrichment of Ash1 in the daughter cell nucleus where the protein repressesmating type switching. (B) Long-distance signalling. FT mRNA is transcribedin leafs of Arabidopsis and transported through the plant vascular system tothe shoot apical meristem, where the encoded protein acts as transcriptionalactivator inducing flowering. (C) Protein isoform sorting and local assemblyof protein complexes. �-Actin but not �-actin mRNA localizes to the leadingedge of chicken embryonic fibroblasts. Local expression of �-actin results in theassembly of this isoform into actin filaments. (D) Facilitated protein targeting.Mgp
cirmsI
LRerndPmPartpfaln
tlolhia(daatdiAmsLs
3c
tcamfmmTptl
etallothionein-1 (MT-1) mRNA localizes to the perinuclear cytoplasm. Tar-eting of the mRNA to this region results in a considerably more efficient MT-1rotein import into the nucleus.
ell (GMC) and a new neuroblast. The GMC divides once morento two postmitotic neurons or glia precursors, while the neu-
oblast continues to divide in a stem cell-like fashion. Duringitosis, prospero mRNA is sorted to the cell cortex of the basalide, which will subsequently become part of the GMC [18,20].n the GMC, Prospero protein acts as a cell fate determinant.
ioit
pmental Biology 18 (2007) 171–177 173
ocalization of prospero is mediated by the double-strandedNA-binding protein Staufen and the adaptor Miranda that teth-rs Staufen to the cell cortex [26,27]. Loss of Staufen functionesults in symmetric distribution of prospero mRNA but doesot block asymmetric distribution of Prospero protein. This isue to a direct interaction of the cortical protein Miranda withrospero that is independent of its binding to prospero mRNAediated via Staufen [26]. Thus, Miranda serves to localize bothrospero protein and its mRNA. In contrast to Prospero, Inscute-ble protein is part of an apically sorted protein complex that isequired to coordinate mitotic spindle orientation and sorting ofhe basal sorting machinery [18]. Inscuteable mRNA is trans-orted to the apical cortex in a dynein-dependent process butailure to localize the mRNA results in only a modest defect ofsymmetric division [25]. Both examples suggest that mRNAocalization in the Drosophila neuroblast serves to facilitate butot accomplish asymmetric protein distribution.
During Xenopus development, localization of mRNAs con-ributes to embryonic patterning [28]. Several mRNAs areocalized to the vegetal pole or the vegetal hemisphere of theocyte and are later asymmetrically inherited by the cellu-ar progeny that arises from this region. Two main pathwaysave been described that result in vegetal RNA localizationn Xenopus [29]. During the early pathway, mRNAs associatend move with the so-called message transport organizer regionMETRO), which is part of the mitochondrial cloud that formsuring early oocyte development [29]. Most of these mRNAsre incorporated into the germ plasm and encode proteins thatre required for germ cell development [28]. The late localiza-ion pathway in Xenopus oocytes is used to deposit germ layereterminants in the vegetal hemisphere [28,29]. This hemispheres a repository of mesodermal and endodermal determinants.
well-characterized example for the importance of localizingRNAs to this region is VegT mRNA that encodes a T-box tran-
cription factor required for mesendodermal development [30].ocalization of another mRNA, Vg1, has also been implicated inpecification of both mesodermal and endodermal tissues [31].
. mRNA localization and local assembly of proteinomplexes
In polarized somatic cells, localized mRNAs can serve asemplate for spatially restricted synthesis of proteins to maintainellular asymmetry. In some cases, mRNA localization mightlso help to facilitate the compartmentalized assembly ofultifactor complexes. One of the best-characterized examples
or this aspect of mRNA localization is �-actin mRNA inotile cells (Fig. 1C). In several motile cell types, �-actinRNA is targeted to the leading edge of lamellipodia [32].hese flattened cytoplasmic extensions are actin rich andolymerizing actin filaments provide the protrusive force forhe extension of lamellipodia during cell motility. Cis-actingocalization elements within the 3′-UTR (untranslated region)
ncluding the ‘zipcode’ sequence are essential for the sortingf �-actin mRNA [33]. The zipcode binding protein 1 (ZBP1)s required for the transport of �-actin mRNA [34] and inhibitsranslation until the message reaches its final destination [35].
1 evelo
Ttp
mtcbmndottptmasadtc
tBmcaAlmmabefaeedfd�ar
litwelst[t
phdca
4
owsttp
lm(lcmi1ts3mopMaT1meg
ttstpmtptiamtta
74 T.-G. Du et al. / Seminars in Cell & D
hese results are consistent with a recent publication which forhe first time showed directly that �-actin protein is translatedrimarily at the sites of mRNA localization [36].
The first indications for a physiological function of �-actinRNA targeting came from studies in which motile cells were
reated with antisense oligonucleotides directed against the zip-ode. Treatment resulted not only in loss of mRNA localizationut also in impaired cell morphology and motility [37]. Pointutations in the mRNA zipcode or over-expression of domi-
ant negative ZBP1 fragments had the same effect [32,34]. Whyoes �-actin mRNA delocalization cause loss of cell morphol-gy and motility? A decreased local concentration of �-actin athe leading edge due to mislocalized �-actin mRNA could havewo possible consequences. Reduced efficiency of actin filamentolymerization could slow down the formation of new cell pro-rusions. Alternatively, the rate of actin polymerization itself
ight be unaffected yet the location of actin synthesis may beltered. That indeed the second suggested mechanism is respon-ible for cell polarity and directional motility was suggested byn elegant series of experiments [38]. Loss of cell polarizationue to impaired �-actin mRNA targeting can have other dele-erious effects such as the gain of metastatic potential in tumorells [39–41].
It is evident from the observations discussed above that mis-argeting of �-actin mRNA results in abnormal cell phenotypes.ut this does not directly address the question why �-actinRNA instead of �-actin protein is localized. RNA localization
an create a microenvironment in which the newly synthesizedctin monomers are available at much higher concentration.s �-actin translation occurs in a restricted cell compartment
ike the thin lamellipodium [36], high local levels of �-actinonomers can considerably enhance the dynamics of actin poly-erization [32]. Additionally, mRNA localization can easily
llow for sorting of specific actin isoforms. Since only �-actin,ut not �- or �-actin mRNAs, is targeted to the cell periph-ry in moving cells [33], its local translation might preventormation of unwanted isoform heteromers. Interestingly, somectin-binding proteins that are believed to function at the leadingdge preferentially interact with the �-isoform and its specificnrichment might consequently support these interactions (foretailed discussion see Ref. [32]). Recently, another exampleor protein isoform specific sorting on the level of mRNA wasescribed. Integrin �3 but not other integrin � isoform mRNAs or-subunit mRNAs localize to focal adhesion sites [42]. However,s integrins form obligate �/�-heterodimers, the physiologicalelevance for �3 mRNA localization remains unclear.
�-Actin mRNA is not the only transcript targeted to theeading edge in migrating cells. Several other mRNAs encod-ng motility-related proteins contain the consensus sequence ofhe �-actin zipcode element and are likely to be localized asell [38]. The actin-related protein 2/3 (ARP2/3) complex is
ssential for the nucleation of actin filament assembly and isocated at leading protrusions in crawling cells [43]. A recent
tudy demonstrated that the mRNAs of all seven subunits ofhe ARP2/3 complex are localized to lamellipodia in fibroblasts44]. Importantly, this study provides the first case in which allranscripts encoding a multimeric complex are targeted to its[
ht
pmental Biology 18 (2007) 171–177
lace of function and potential assembly site. The advantage foretero-multimer assembly offered by mRNA localization is evi-ent since high local concentrations of all subunits can facilitateomplex assembly. Such compartmentalized synthesis wouldlso avoid any random and unwanted protein interactions.
. mRNA localization to the vicinity of organelles
mRNA localization can also facilitate protein sorting torganelles. Generally, protein sorting is based on peptide signalsithin the protein to be sorted. However, increasing evidence
uggests that localization of mRNAs encoding proteins destinedo organelles like ER, mitochondria or nuclei supports proteinargeting or allows refined targeting that cannot occur on therotein level (see also Dahm et al., this issue).
Vimentin, �-actin, and �-actin mRNAs display perinuclearocalization [33,45,46]. In addition, transcripts coding for
etallothionein-1 (MT-1), cellular retinoic acid binding protein ICRABPI) and transcription factors c-FOS and c-MYC accumu-ate at the nuclear periphery and associate with the perinuclearytoskeleton [47–50]. The physiological role of perinuclearRNA localization might be to enhance the efficiency of nuclear
mport of proteins encoded by these mRNAs (Fig. 1D). MT-is a predominantly cytosolic protein that is imported into
he nucleus upon G1/S phase transition of the cell cycle. In atudy where cells were transfected with MT-1 mRNA whose′UTR was replaced by a heterologous RNA, the hybrid MT-1essage did no longer take up a perinuclear position. Simultane-
usly, MT-1 protein did not localize to the nucleus during G1/Shase transition [51]. Interestingly, cells with non-localizingT-1 mRNA were more susceptible to stress, DNA damage
nd showed increased apoptosis after exposure to mutagens [52].hese observations indicate that perinuclear localization of MT-mRNA is necessary for the protein to be fully functional, whichight also be true for c-FOS, c-MYC or CRABPI. It is, how-
ver, not yet clear to what extent this functional model can beeneralized.
A number of mRNAs are targeted to the rough ER throughhe nascent chain emanating from a translating ribosome. Onop of this general targeting, certain transcripts are directed topecific subcompartments of the ER. In developing rice seeds,his is the case for two mRNAs encoding different seed storageroteins, prolamines and globulin-like glutelins [53]. ProlamineRNAs localize to the surface of ER containing prolamine pro-
ein bodies, so called PB-ER. In contrast, glutelin RNAs arereferentially targeted to cisternal ER (C-ER). Both storage pro-eins are synthesized on the surface of cortical ER, translocatednto the ER lumen and then further sorted. Targeting of stor-ge protein mRNAs in the rice seeds provides an interestingechanism that facilitates trafficking of their corresponding pro-
eins to distinct endomembrane compartments. Consequently,his allows a spatial separation of protein synthesis to preventnon-productive mixing of the two disparate storage proteins
54–56].During the evolution of the eukaryotic cell, mitochondria
ave lost the majority of their initial genome, which rendershese organelles dependent on the delivery of proteins encoded

evelo
btnpf[TArbmtat
5i
cimtstsaOtplktaltfpssird(tfAimca
otGtt
mLnalRtPtfAtdoosdiasMhsosismml
6
tBgtacfnp
A
F
R
T.-G. Du et al. / Seminars in Cell & D
y nuclear genes. Although there is a large body of evidence thathese proteins are targeted to mitochondria post-translationally,ew data suggest that sorting of a subset of nuclear-encodedroteins to mitochondria involves mRNA localization. In yeast,or instance, the 3′-UTR of ATM1 [57] and ATP2 mRNAs58] direct these transcripts to the vicinity of mitochondria.he latter transcript encodes a subunit of the mitochondrialTP synthase. Interestingly, impaired ATP2 mRNA sorting cor-
elates with a severe respiratory deficiency, indicating a linketween mRNA localization and protein function. However, theechanistic details underlying the transport of mitochondrially-
argeted mRNAs remain elusive. Local enrichment of transcriptst mitochondrial-bound polysomes may ensure an effective co-ranslational import into these organelles [59].
. Long distance transport of mRNAs and small RNAsn plants
Less is known about RNA localization in the plant kingdomompared to animal cells but a role for RNAs as long-distancenformation molecules is emerging. The framework for such a
echanism is provided by the plant vascular system. In con-rast to animals, higher plants have evolved a unique vascularystem that consists of xylem and phloem. Xylem cells allowhe delivery of water and minerals, whereas phloem, which con-ists of living but enucleated sieve elements, transports photossimilates and nutrients to distantly located tissues and organs.ver the last decade, there is a growing body of evidence that
he phloem translocation stream also acts as an informationipeline, enabling coordination of developmental and physio-ogical processes at level of the whole plant. Plant phloem isnown to deliver a wide range of signalling molecules like pro-eins and phytohormones, but strikingly also RNA moleculesre transported. In terms of systemic RNA silencing, it hasong been known that RNA-based silencing signals are able toravel throughout the plant to spread systemically. Small inter-ering RNAs (siRNAs) are likely candidates to exert this form ofost-transcriptional gene silencing [60]. The phloem-dependentystemic transmission and amplification of RNA-based defenceignals may help to alert developing tissues to the presence ofnvasive and pathogenic genetic elements and is therefore ofteneferred to as the plant immune system [61]. In addition, longistance transport was also observed for non-coding microRNAsmiRNAs). In plants, miRNAs have emerged as important post-ranscriptional regulators and many of them have implicatedunctions in plant development or cell differentiation [62,63].ccordingly, the detection of a set of different miRNA species
n phloem sap suggests that in response to environmental cuesiRNAs may serve as bona fide systemic signals to exert non-
ell-autonomous control over diverse processes in plant growthnd development [64].
Support for the concept that also mRNAs are translocatedver long distances was initially provided by the identifica-
ion of a unique set of mRNAs isolated from phloem sap [65].rafting experiments and in situ RT-PCR analysis revealed thathe phloem stream has the capacity to mediate long distanceransport of several messenger RNAs to the apical meristem,
pmental Biology 18 (2007) 171–177 175
oving from the plant rootstock into a grafted scion [65–67].ong distance transport requires that mRNAs can traverse thearrow but dynamic channels of plasmodesmata (PD), whichre pores connecting adjacent cells. The passage via PD high-ights an important prerequisite for cell-to-cell movement ofNA [68]. The up- and unloading of RNAs into and out of
he phloem translocation stream, also appears to occur throughD [67,69]. How long-distance trafficking of mRNA is func-
ionally linked to developmental events was recently shownor the model plant Arabidopsis thaliana [70,71] (Fig. 1). Inrabidopsis day length is a major environmental determinant
hat controls the time of flowering, especially in spring, whenay length and ambient temperature increase. The photoperi-dic perception of day-length in the leaf leads to expressionf the mobile stimulus Flowering locus T (FT). Although tran-cribed in leafs, FT protein functions within the shoot apex, aistantly located meristematic region where floral developments stimulated. Here we can find a situation where expression of
signal and its site of function are spatially and temporarilyeparated. A mobile FT signal could solve the spatial paradox.
ovement of FT mRNA along the phloem was shown usingeat shock promoters to locally induce expression of FT in aingle Arabidopsis leaf [71]. After induction, increased levelsf the transgenic FT transcript appear at the shoot apex. Thisuggests that the pulse of FT expression can move from leaf tots destination where it triggers flowering. Although the compo-ition of the flowering signal is still elusive and specific proteinsediating the transport processes remain to be determined, FTRNA transport nicely demonstrates that mRNAs can serve as
ong-distance developmental signals in the plant kingdom.
. Outlook
mRNA localization serves as a sophisticated cellular toolo optimize protein expression on a post-transcriptional level.esides well-understood functions like generating morphogenradients or sorting determinants, mRNA localization seemso fulfil a variety of additional roles ranging from facilitatedssembly of protein complexes to long-distance signalling andoordination of developmental processes. In addition, newunctions have emerged like modulating synaptic plasticity ineurons or maximizing protein targeting to subcellular com-artments.
cknowledgements
R.P.J. and T.G.D. are supported by grants from the Deutscheorschungsgemeinschaft (DFG JA696/4-4 and SFB646 TPA5).
eferences
[1] Claussen M, Horvay K, Pieler T. Evidence for overlapping, but not identical,
protein machineries operating in vegetal RNA localization along early andlate pathways in Xenopus oocytes. Development 2004;131(17):4263–73.[2] Tomancak P, Beaton A, Weiszmann R, Kwan E, Shu S, Lewis SE, et al.Systematic determination of patterns of gene expression during Drosophilaembryogenesis. Genome Biol 2002;3(12) [RESEARCH0088].

1 evelo
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
76 T.-G. Du et al. / Seminars in Cell & D
[3] Shepard KA, Gerber AP, Jambhekar A, Takizawa PA, Brown PO, HerschlagD, et al. Widespread cytoplasmic mRNA transport in yeast: Identificationof 22 bud-localized transcripts using DNA microarray analysis. Proc NatlAcad Sci USA 2003;100(20):11429–34.
[4] St Johnston D. The intracellular localization of messenger RNAs. Cell1995;81(2):161–70.
[5] Boggs JM. Myelin basic protein: a multifunctional protein. Cell Mol LifeSci 2006;63(17):1945–61.
[6] Wolpert L. Positional information and the spatial pattern of cellular differ-entiation. J Theor Biol 1969;25:1–47.
[7] Ephrussi A, St Johnston D. Seeing is believing: the bicoid morphogengradient matures. Cell 2004;116:143–52.
[8] Driever W, Nusslein-Volhard C. A gradient of bicoid protein in Drosophilaembryos. Cell 1988;54:83–93.
[9] Ephrussi A, Dickinson LK, Lehmann R. oskar organizes the germplasm and directs localization of the posterior determinant nanos. Cell1991;66:37–50.
10] Mahowald AP. Assembly of the Drosophila germ plasm. Int Rev Cytol2001;203:187–213.
11] Wang C, Lehmann R. Nanos is the localized posterior determinant deter-minant in Drosophila. Cell 1991;66:637–47.
12] Crucs S, Chatterjee S, Gavis ER. Overlapping but distinct RNA ele-ments control repression and activation of nanos translation. Mol Cell2000;5:457–67.
13] Nilson LA, Schupbach T. EGF receptor signaling in Drosophila oogenesis.Curr Top Dev Biol 1999;44:203–43.
14] Klingensmith J, Nusse R. Signaling by wingless in Drosophila. Dev Biol1994;166(2):396–414.
15] Simmonds AJ, dosSantos G, Livne-Bar I, Krause HM. Apical local-ization of wingless transcripts is required for wingless signaling. Cell2001;105:197–207.
16] Cowan CR, Hyman AA. Asymmetric cell division in C. elegans: corticalpolarity and spindle positioning. Annu Rev Cell Dev Biol 2004;20:427–53.
17] Piggot PJ, Hilbert DW. Sporulation of Bacillus subtilis. Curr Opin Micro-biol 2004;7(6):579–86.
18] Yu F, Kuo CT, Jan YN. Drosophila neuroblast asymmetric cell divi-sion: recent advances and implications for stem cell biology. Neuron2006;51(1):13–20.
19] Sardet C, Dru P, Prodon F. Maternal determinants and mRNAs in the cortexof ascidian oocytes, zygotes and embryos. Biol Cell 2005;97(1):35–49.
20] Broadus J, Fuerstenberg S, Doe CQ. Staufen-dependent localization ofprospero mRNA contributes to neuroblast daughter cell fate. Nature1998;391:792–5.
21] Cosma MP. Daughter-specific repression of Saccharomyces cerevisiae HO:Ash1 is the commander. EMBO Rep 2004;5(10):953–7.
22] Bobola N, Jansen R-P, Shin TH, Nasmyth K. Asymmetric accumulationof Ash1p in postanaphase nuclei depends on a myosin and restricts yeastmating-type switching to mother cells. Cell 1996;84:699–709.
23] Long RM, Singer RH, Meng X, Gonzalez I, Nasmyth K, Jansen R-P. Matingtype switching in yeast controlled by asymmetric localization of ASH1mRNA. Science 1997;277:383–7.
24] Gonsalvez GB, Urbinati CR, Long RM. RNA localization in yeast: movingtowards a mechanism. Biol Cell 2005;97(1):75–86.
25] Hughes JR, Bullock SL, Ish-Horowicz D. Inscuteable mRNA localizationis dynein-dependent and regulates apicobasal polarity and spindle lengthin Drosophila neuroblasts. Curr Biol 2004;14(21):1950–6.
26] Schuldt AJ, Adams JHJ, Davidson CM, Micklem DR, Haseloff J, St John-ston D, et al. Miranda mediates asymmetric protein and RNA localizationin the developing nervous system. Genes Dev 1998;12:1847–57.
27] Shen CP, Knoblich JA, Chan YM, Jiang MM, Jan LY, Jan YN. Mirandaas a multidomain adapter linking apically localized Inscuteable andbasally localized Staufen and Prospero during asymmetric cell divisionin Drosophila. Genes Dev 1998;12(12):1837–46.
28] King ML, Messitt TJ, Mowry KL. Putting RNAs in the right placeat the right time: RNA localization in the frog oocyte. Biol Cell2005;97(1):19–33.
29] Kloc M, Etkin LD. Two distinct pathways for the localization of RNAs atthe vegetal cortex in Xenopus oocytes. Development 1995;121:287–97.
[
pmental Biology 18 (2007) 171–177
30] Zhang J, Houston DW, King ML, Payne C, Wylie C, Heasman J. The role ofmaternal VegT in establishing the primary germ layers in Xenopus embryos.Cell 1998;94(4):515–24.
31] Melton DA. Translocation of a localized maternal mRNA to the vegetalpole of Xenopus oocytes. Nature 1987;328:80–2.
32] Condeelis J, Singer RH. How and why does beta-actin mRNA target? BiolCell 2005;97(1):97–110.
33] Kislauskis EH, Li Z, Singer RH, Taneja KL. Isoform-specific 3′-untranslated sequences sort alpha-cardiac and beta-cytoplasmic actinmessenger RNAs to different cytoplasmic compartments. J Cell Biol1993;123(1):165–72.
34] Ross AF, Oleynikov Y, Kislauskis EH, Taneja KL, Singer RH. Charac-terization of a beta-actin mRNA zipcode-binding protein. Mol Cell Biol1997;17(4):2158–65.
35] Huttelmaier S, Zenklusen D, Lederer M, Dictenberg J, Lorenz M, MengX, et al. Spatial regulation of beta-actin translation by Src-dependent phos-phorylation of ZBP1. Nature 2005;438(7067):512–5.
36] Rodriguez AJ, Shenoy SM, Singer RH, Condeelis J. Visualization of mRNAtranslation in living cells. J Cell Biol 2006;175(1):67–76.
37] Kislauskis EH, Zhu X, Singer RH. Beta-actin messenger RNA local-ization and protein synthesis augment cell motility. J Cell Biol1997;136(6):1263–70.
38] Shestakova EA, Singer RH, Condeelis J. The physiological significance ofbeta -actin mRNA localization in determining cell polarity and directionalmotility. Proc Natl Acad Sci USA 2001;98(13):7045–50.
39] Condeelis J, Singer RH, Segall JE. The great escape: when cancer cellshijack the genes for chemotaxis and motility. Annu Rev Cell Dev Biol2005;21:695–718.
40] Shestakova EA, Wyckoff J, Jones J, Singer RH, Condeelis J. Correlationof beta-actin messenger RNA localization with metastatic potential in ratadenocarcinoma cell lines. Cancer Res 1999;59(6):1202–5.
41] Wang W, Wyckoff JB, Wang Y, Bottinger EP, Segall JE, Condeelis JS.Gene expression analysis on small numbers of invasive cells collected bychemotaxis from primary mammary tumors of the mouse. BMC Biotechnol2003;3:13.
42] Adereth Y, Dammai V, Kose N, Li R, Hsu T. RNA-dependent inte-grin alpha3 protein localization regulated by the Muscleblind-like proteinMLP1. Nat Cell Biol 2005;7(12):1240–7.
43] Machesky LM, Gould KL. The Arp2/3 complex: a multifunctional actinorganizer. Curr Opin Cell Biol 1999;11(1):117–21.
44] Mingle LA, Okuhama NN, Shi J, Singer RH, Condeelis J, Liu G. Localiza-tion of all seven messenger RNAs for the actin-polymerization nucleatorArp2/3 complex in the protrusions of fibroblasts. J Cell Sci 2005;118(Pt11):2425–33.
45] Hill MA, Gunning P. Beta and gamma actin mRNAs are differentiallylocated within myoblasts. J Cell Biol 1993;122(4):825–32.
46] Lawrence JB, Singer RH. Intracellular localization of messenger RNAs forcytoskeletal proteins. Cell 1986;45(3):407–15.
47] Dalgleish G, Veyrune JL, Blanchard JM, Hesketh J. mRNA localization by a145-nucleotide region of the c-fos 3′-untranslated region links to translationbut not stability. J Biol Chem 2001;276(17):13593–9.
48] Levadoux-Martin M, Li Y, Blackburn A, Chabanon H, Hesketh JE. Perinu-clear localisation of cellular retinoic acid binding protein I mRNA. BiochemBiophys Res Commun 2006;340(1):326–31.
49] Mahon P, Partridge K, Beattie JH, Glover LA, Hesketh JE. The 3′ untrans-lated region plays a role in the targeting of metallothionein-I mRNA tothe perinuclear cytoplasm and cytoskeletal-bound polysomes. BiochimBiophys Acta 1997;1358(2):153–62.
50] Veyrune JL, Campbell GP, Wiseman J, Blanchard JM, Hesketh JE. A local-isation signal in the 3′ untranslated region of c-myc mRNA targets c-mycmRNA and beta-globin reporter sequences to the perinuclear cytoplasmand cytoskeletal-bound polysomes. J Cell Sci 1996;109(Pt 6):1185–94.
51] Levadoux M, Mahon C, Beattie JH, Wallace HM, Hesketh JE. Nuclear
import of metallothionein requires its mRNA to be associated with theperinuclear cytoskeleton. J Biol Chem 1999;274(49):34961–6.52] Levadoux-Martin M, Hesketh JE, Beattie JH, Wallace HM. Influence ofmetallothionein-1 localization on its function. Biochem J 2001;355(Pt2):473–9.

evelo
[
[
[
[
[
[
[
[[
[
[
[
[
[
[
[
[
T.-G. Du et al. / Seminars in Cell & D
53] Choi SB, Wang C, Muench DG, Ozawa K, Franceschi VR, Wu Y, et al.Messenger RNA targeting of rice seed storage proteins to specific ERsubdomains. Nature 2000;407(6805):765–7.
54] Crofts AJ, Washida H, Okita TW, Ogawa M, Kumamaru T, Satoh H. Target-ing of proteins to endoplasmic reticulum-derived compartments in plants.The importance of RNA localization. Plant Physiol 2004;136(3):3414–9.
55] Okita TW, Choi SB. mRNA localization in plants: targeting to the cell’scortical region and beyond. Curr Opin Plant Biol 2002;5(6):553–9.
56] Hara-Nishimura I, Matsushima R, Shimada T, Nishimura M. Diver-sity and formation of endoplasmic reticulum-derived compartments inplants. Are these compartments specific to plant cells? Plant Physiol2004;136(3):3435–9.
57] Corral-Debrinski M, Blugeon C, Jacq C. In yeast, the 3′ untranslatedregion or the presequence of ATM1 is required for the exclusive local-ization of its mRNA to the vicinity of mitochondria. Mol Cell Biol2000;20(21):7881–92.
58] Margeot A, Blugeon C, Sylvestre J, Vialette S, Jacq C, Corral-DebrinskiM. In Saccharomyces cerevisiae ATP2 mRNA sorting to the vicin-ity of mitochondria is essential for respiratory function. EMBO J2002;21(24):6893–904.
59] Margeot A, Garcia M, Wang W, Tetaud E, di Rago JP, Jacq C. Why are
many mRNAs translated to the vicinity of mitochondria: a role in proteincomplex assembly? Gene 2005;354:64–71.60] Baulcombe D. RNA silencing in plants. Nature 2004;431(7006):356–63.61] Lucas WJ, Yoo BC, Kragler F. RNA as a long-distance information macro-
molecule in plants. Nat Rev Mol Cell Biol 2001;2(11):849–57.
[
[
pmental Biology 18 (2007) 171–177 177
62] Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function.Cell 2004;116(2):281–97.
63] Kidner CA, Martienssen RA. The developmental role of microRNA inplants. Curr Opin Plant Biol 2005;8(1):38–44.
64] Yoo BC, Kragler F, Varkonyi-Gasic E, Haywood V, Archer-Evans S, LeeYM, et al. A systemic small RNA signaling system in plants. Plant Cell2004;16(8):1979–2000.
65] Ruiz-Medrano R, Xoconostle-Cazares B, Lucas WJ. Phloem long-distancetransport of CmNACP mRNA: implications for supracellular regulation inplants. Development 1999;126(20):4405–19.
66] Kim M, Canio W, Kessler S, Sinha N. Developmental changes due to long-distance movement of a homeobox fusion transcript in tomato. Science2001;293(5528):287–9.
67] Xoconostle-Cazares B, Xiang Y, Ruiz-Medrano R, Wang HL, Monzer J,Yoo BC, et al. Plant paralog to viral movement protein that potentiatestransport of mRNA into the phloem. Science 1999;283(5398):94–8.
68] Lucas WJ, Lee JY. Plasmodesmata as a supracellular control network inplants. Nat Rev Mol Cell Biol 2004;5(9):712–26.
69] Kim JY, Rim Y, Wang J, Jackson D. A novel cell-to-cell trafficking assayindicates that the KNOX homeodomain is necessary and sufficient forintercellular protein and mRNA trafficking. Genes Dev 2005;19(7):788–93.
70] Blazquez MA. Plant science. The right time and place for making flowers.Science 2005;309(5737):1024–5.
71] Huang T, Bohlenius H, Eriksson S, Parcy F, Nilsson O. The mRNA of theArabidopsis gene FT moves from leaf to shoot apex and induces flowering.Science 2005;309(5741):1694–6.




















