
Wheat genetic resources enhancement by the International Maize and Wheat Improvement Center (CIMMYT
46
UNCORRECTED PROOF UNCORRECTED PROOF RESEARCH ARTICLE 1 2 Wheat genetic resources enhancement by the International 3 Maize and Wheat Improvement Center (CIMMYT) 4 Rodomiro Ortiz Hans-Joachim Braun Jose ´ Crossa Jonathan H. Crouch 5 Guy Davenport John Dixon Susanne Dreisigacker Etienne Duveiller 6 Zhonghu He Julio Huerta Arun K. Joshi Masahiro Kishii Petr Kosina 7 Yann Manes Monica Mezzalama Alexei Morgounov Jiro Murakami 8 Julie Nicol Guillermo Ortiz Ferrara J. Iva ´n Ortiz-Monasterio Thomas S. Payne 9 R. Javier Pen ˜a Matthew P. Reynolds Kenneth D. Sayre Ram C. Sharma 10 Ravi P. Singh Jiankang Wang Marilyn Warburton Huixia Wu Masa Iwanaga 11 12 13 Received: 7 April 2008 / Accepted: 25 August 2008 14 Ó Springer Science+Business Media B.V. 2008 15 Abstract The International Maize and Wheat 16 Improvement Center (CIMMYT) acts as a catalyst 17 and leader in a global maize and wheat innovation 18 network that serves the poor in the developing world. 19 Drawing on strong science and effective partnerships, 20 CIMMYT researchers create, share, and use knowl- 21 edge and technology to increase food security, 22 improve the productivity and profitability of farming 23 systems and sustain natural resources. This people- 24 centered mission does not ignore the fact that 25 CIMMYT’s unique niche is as a genetic resources 26 enhancement center for the developing world, as 27 shown by this review article focusing on wheat. 28 CIMMYT’s value proposition resides therefore in its 29 use of crop genetic diversity: conserving it, studying 30 it, adding value to it, and sharing it in enhanced form 31 with clients worldwide. The main undertakings 32 include: long-term safe conservation of world heri- 33 tage of both crop resources for future generations, in 34 line with formal agreements under the 2004 Interna- 35 tional Treaty on Plant Genetic Resources for Food 36 and Agriculture, understanding the rich genetic 37 diversity of two of the most important staples 38 worldwide, exploiting the untapped value of crop 39 genetic resources through discovery of specific, 40 strategically-important traits required for current 41 and future generations of target beneficiaries, and 42 development of strategic germplasm through innova- 43 tive genetic enhancement. Finally, the Center needs 44 to ensure that its main products reach end-users and 45 improve their livelihoods. In this regard, CIMMYT is 46 the main international, public source of wheat seed- 47 embedded technology to reduce vulnerability and 48 alleviate poverty, helping farmers move from subsis- 49 tence to income-generating production systems. 50 Beyond a focus on higher grain yields and value- 51 added germplasm, CIMMYT plays an ‘‘integrative’’ 52 role in crop and natural resource management 53 research, promoting the efficient use of water and 54 other inputs, lower production costs, better manage- 55 ment of biotic stresses, and enhanced system 56 diversity and resilience. A1 R. Ortiz (&) Á H.-J. Braun Á J. Crossa Á J. H. Crouch Á A2 G. Davenport Á J. Dixon Á S. Dreisigacker Á A3 E. Duveiller Á Z. He Á J. Huerta Á M. Kishii Á P. Kosina Á A4 Y. Manes Á M. Mezzalama Á A. Morgounov Á A5 J. Murakami Á J. Nicol Á G. Ortiz Ferrara Á A6 J. I. Ortiz-Monasterio Á T. S. Payne Á R. J. Pen ˜a Á A7 M. P. Reynolds Á K. D. Sayre Á R. C. Sharma Á A8 R. P. Singh Á J. Wang Á M. Warburton Á H. Wu Á A9 M. Iwanaga A10 Centro Internacional de Mejoramiento de Maı ´z y Trigo A11 (CIMMYT), Km. 45 Carretera Me ´xico-Veracruz, A12 Col. El Bata ´n, Texcoco, Edo. de Mexico 56130, Mexico A13 e-mail: [email protected] A14 A. K. Joshi A15 Department of Genetics and Plant Breeding, Institute of A16 Agricultural Sciences, Banaras Hindu University, A17 Varanasi, UP, India 123 Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46 Article No. : 9372 h LE h TYPESET MS Code : GRES951 h CP h DISK 4 4 Genet Resour Crop Evol DOI 10.1007/s10722-008-9372-4 Author Proof
Transcript of Wheat genetic resources enhancement by the International Maize and Wheat Improvement Center (CIMMYT

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
RESEARCH ARTICLE1
2 Wheat genetic resources enhancement by the International
3 Maize and Wheat Improvement Center (CIMMYT)
4 Rodomiro Ortiz Hans-Joachim Braun Jose Crossa Jonathan H. Crouch
5 Guy Davenport John Dixon Susanne Dreisigacker Etienne Duveiller
6 Zhonghu He Julio Huerta Arun K. Joshi Masahiro Kishii Petr Kosina
7 Yann Manes Monica Mezzalama Alexei Morgounov Jiro Murakami
8 Julie Nicol Guillermo Ortiz Ferrara J. Ivan Ortiz-Monasterio Thomas S. Payne
9 R. Javier Pena Matthew P. Reynolds Kenneth D. Sayre Ram C. Sharma
10 Ravi P. Singh Jiankang Wang Marilyn Warburton Huixia Wu Masa Iwanaga111213 Received: 7 April 2008 / Accepted: 25 August 200814 � Springer Science+Business Media B.V. 2008
15 Abstract The International Maize and Wheat
16 Improvement Center (CIMMYT) acts as a catalyst
17 and leader in a global maize and wheat innovation
18 network that serves the poor in the developing world.
19 Drawing on strong science and effective partnerships,
20 CIMMYT researchers create, share, and use knowl-
21 edge and technology to increase food security,
22 improve the productivity and profitability of farming
23 systems and sustain natural resources. This people-
24 centered mission does not ignore the fact that
25 CIMMYT’s unique niche is as a genetic resources
26 enhancement center for the developing world, as
27 shown by this review article focusing on wheat.
28CIMMYT’s value proposition resides therefore in its
29use of crop genetic diversity: conserving it, studying
30it, adding value to it, and sharing it in enhanced form
31with clients worldwide. The main undertakings
32include: long-term safe conservation of world heri-
33tage of both crop resources for future generations, in
34line with formal agreements under the 2004 Interna-
35tional Treaty on Plant Genetic Resources for Food
36and Agriculture, understanding the rich genetic
37diversity of two of the most important staples
38worldwide, exploiting the untapped value of crop
39genetic resources through discovery of specific,
40strategically-important traits required for current
41and future generations of target beneficiaries, and
42development of strategic germplasm through innova-
43tive genetic enhancement. Finally, the Center needs
44to ensure that its main products reach end-users and
45improve their livelihoods. In this regard, CIMMYT is
46the main international, public source of wheat seed-
47embedded technology to reduce vulnerability and
48alleviate poverty, helping farmers move from subsis-
49tence to income-generating production systems.
50Beyond a focus on higher grain yields and value-
51added germplasm, CIMMYT plays an ‘‘integrative’’
52role in crop and natural resource management
53research, promoting the efficient use of water and
54other inputs, lower production costs, better manage-
55ment of biotic stresses, and enhanced system
56diversity and resilience.
A1 R. Ortiz (&) � H.-J. Braun � J. Crossa � J. H. Crouch �
A2 G. Davenport � J. Dixon � S. Dreisigacker �
A3 E. Duveiller � Z. He � J. Huerta � M. Kishii � P. Kosina �
A4 Y. Manes � M. Mezzalama � A. Morgounov �
A5 J. Murakami � J. Nicol � G. Ortiz Ferrara �
A6 J. I. Ortiz-Monasterio � T. S. Payne � R. J. Pena �
A7 M. P. Reynolds � K. D. Sayre � R. C. Sharma �
A8 R. P. Singh � J. Wang � M. Warburton � H. Wu �
A9 M. IwanagaA10 Centro Internacional de Mejoramiento de Maız y TrigoA11 (CIMMYT), Km. 45 Carretera Mexico-Veracruz,A12 Col. El Batan, Texcoco, Edo. de Mexico 56130, MexicoA13 e-mail: [email protected]
A14 A. K. JoshiA15 Department of Genetics and Plant Breeding, Institute ofA16 Agricultural Sciences, Banaras Hindu University,A17 Varanasi, UP, India
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Genet Resour Crop Evol
DOI 10.1007/s10722-008-9372-4
Au
tho
r P
ro
of
UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
57 Keywords Triticum � Biofortification �
58 Climate change � Food safety � Genetic broadening �
59 Modeling � Participatory varietal selection �
60 Plant breeding � Rusts � Scab
Abbreviations
61 AA Association analysis
62 ARI Advanced research institutes
63 BNI Biological nitrification inhibition
64 CAZS-NR Center for Arid Zone Studies-Natural
Resources
65 CGIAR Consultative Group on International
Agricultural Research
66 CIMMYT Centro Internacional de
Mejoramiento de Maız y Trigo
67 CTD Canopy temperature depression
68 CWANA Central and West Asia and Northern
Africa
69 DON Deoxynivalenol
70 FAO Food and Agriculture Organization
of the United Nations
71 FAWWON Facultative and Winter Wheat
Observation Nursery
72 FHB Fusarium head blight
73 FONTAGRO Fondo Regional de Tecnologıa
Agropecuaria
74 GBSSI Granule-bound starch synthase
75 GE Genotype-by-environment
76 ICAR Indian Council of Agricultural
Research
77 ICARDA International Center for Agricultural
Research in the Dry Areas
78 ITPGRFA International Treaty on Plant Genetic
Resources for Food and Agriculture
79 IWIN International Wheat Improvement
Network
80 MEs Mega-environments
81 IWWIP International Winter Wheat
Improvement Program
82 LD Linkage disequilibrium
83 MAS Marker-assisted selection
84 MODPED Modified pedigree/bulk method
85 NARS National Agricultural Research
Systems
86 NIRS Near-infrared spectroscopy
87 NIV Nivalenol
88 OSS Office of Special Studies
89 OSU Oregon State University
90 PCR Polymerase chain reaction
91PVS Participatory variety selection
92QTL Quantitative trait loci
93RWC Rice-Wheat Consortium for the
Indo-Gangetic Plains
94SAGARPA Secretarıa de Agricultura Ganaderıa.
Desarrollo Rural Pesca y
Alimentacion (Mexico)
95SELBLK Selected bulk method
96SHL Seed Health Laboratory
97SMTA Standard Material Transfer
Agreement
98RCT Resource conserving technology
99ZEN Zearalenone
100
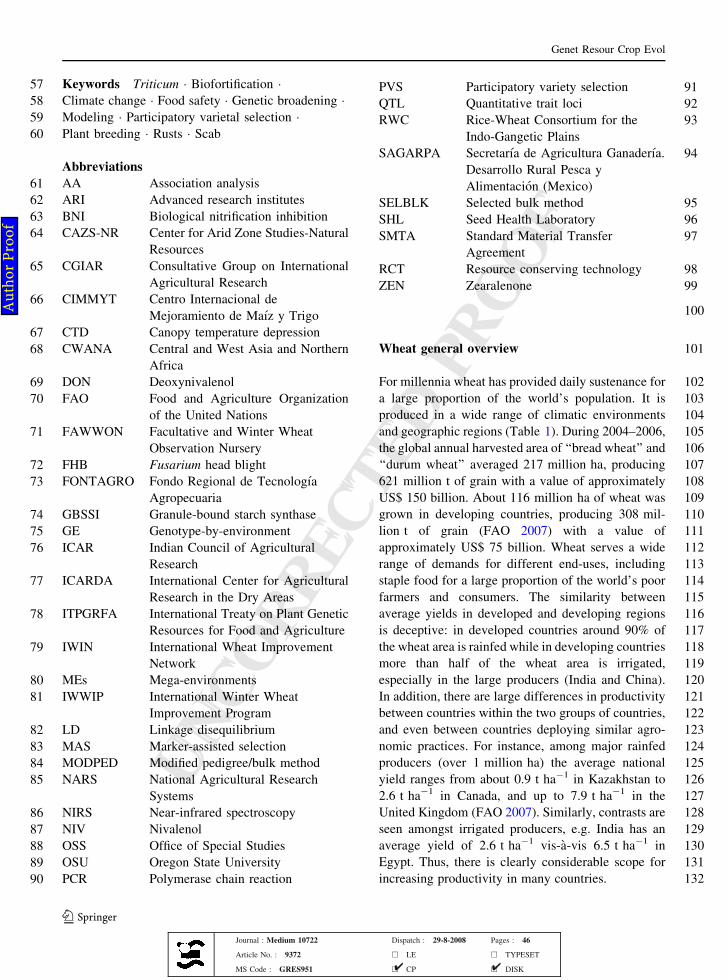
101Wheat general overview
102For millennia wheat has provided daily sustenance for
103a large proportion of the world’s population. It is
104produced in a wide range of climatic environments
105and geographic regions (Table 1). During 2004–2006,
106the global annual harvested area of ‘‘bread wheat’’ and
107‘‘durum wheat’’ averaged 217 million ha, producing
108621 million t of grain with a value of approximately
109US$ 150 billion. About 116 million ha of wheat was
110grown in developing countries, producing 308 mil-
111lion t of grain (FAO 2007) with a value of
112approximately US$ 75 billion. Wheat serves a wide
113range of demands for different end-uses, including
114staple food for a large proportion of the world’s poor
115farmers and consumers. The similarity between
116average yields in developed and developing regions
117is deceptive: in developed countries around 90% of
118the wheat area is rainfed while in developing countries
119more than half of the wheat area is irrigated,
120especially in the large producers (India and China).
121In addition, there are large differences in productivity
122between countries within the two groups of countries,
123and even between countries deploying similar agro-
124nomic practices. For instance, among major rainfed
125producers (over 1 million ha) the average national
126yield ranges from about 0.9 t ha-1 in Kazakhstan to
1272.6 t ha-1 in Canada, and up to 7.9 t ha-1 in the
128United Kingdom (FAO 2007). Similarly, contrasts are
129seen amongst irrigated producers, e.g. India has an
130average yield of 2.6 t ha-1 vis-a-vis 6.5 t ha-1 in
131Egypt. Thus, there is clearly considerable scope for
132increasing productivity in many countries.
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of
UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
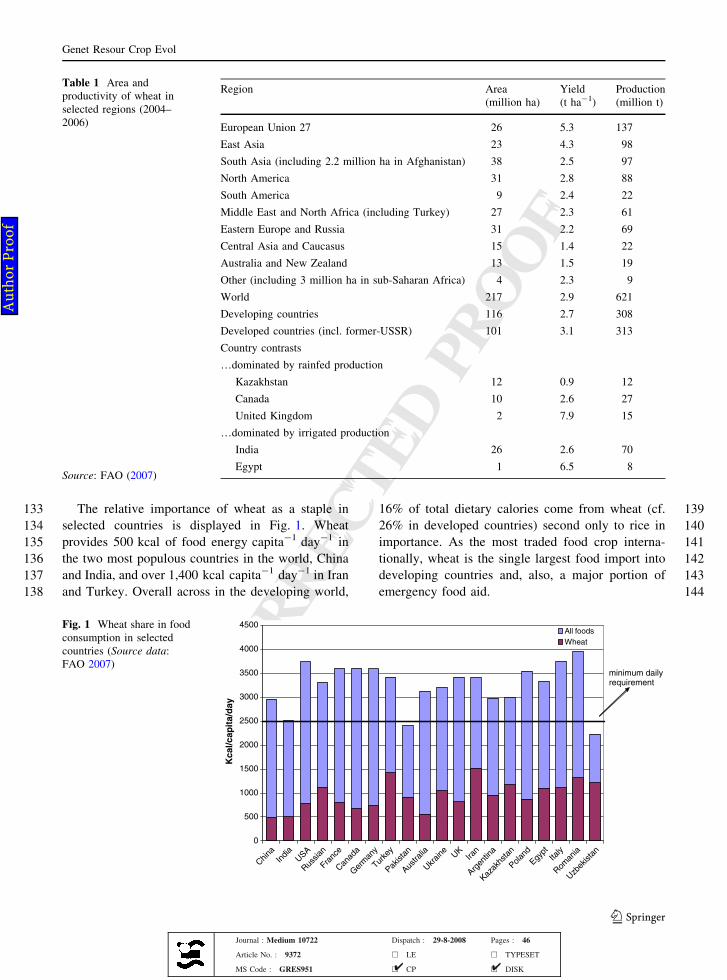
133 The relative importance of wheat as a staple in
134 selected countries is displayed in Fig. 1. Wheat
135 provides 500 kcal of food energy capita-1 day-1 in
136 the two most populous countries in the world, China
137 and India, and over 1,400 kcal capita-1 day-1 in Iran
138 and Turkey. Overall across in the developing world,
13916% of total dietary calories come from wheat (cf.
14026% in developed countries) second only to rice in
141importance. As the most traded food crop interna-
142tionally, wheat is the single largest food import into
143developing countries and, also, a major portion of
144emergency food aid.
Table 1 Area andproductivity of wheat inselected regions (2004–2006)
Source: FAO (2007)
Region Area(million ha)
Yield(t ha-1)
Production(million t)
European Union 27 26 5.3 137
East Asia 23 4.3 98
South Asia (including 2.2 million ha in Afghanistan) 38 2.5 97
North America 31 2.8 88
South America 9 2.4 22
Middle East and North Africa (including Turkey) 27 2.3 61
Eastern Europe and Russia 31 2.2 69
Central Asia and Caucasus 15 1.4 22
Australia and New Zealand 13 1.5 19
Other (including 3 million ha in sub-Saharan Africa) 4 2.3 9
World 217 2.9 621
Developing countries 116 2.7 308
Developed countries (incl. former-USSR) 101 3.1 313
Country contrasts
…dominated by rainfed production
Kazakhstan 12 0.9 12
Canada 10 2.6 27
United Kingdom 2 7.9 15
…dominated by irrigated production
India 26 2.6 70
Egypt 1 6.5 8
0
500
1000
1500
2000
2500
3000
3500
4000
4500
China
India
USA
Rus
sian
Franc
e
Can
ada
Ger
man
y
Turke
y
Pakista
n
Austra
lia
Ukr
aine U
KIra
n
Argen
tina
Kazak
hsta
n
Polan
d
Egypt
Italy
Rom
ania
Uzb
ekista
n
Kcal/cap
ita/d
ay
All foods
Wheat
minimum daily requirement
Fig. 1 Wheat share in foodconsumption in selectedcountries (Source data:FAO 2007)
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
145 Wheat made a significant contribution to the
146 increase in global food production during the past
147 four decades as total production rose steadily through
148 the use of higher-yielding, water and fertilizer-
149 responsive and disease-resistant cultivars supported
150 by strengthened input delivery systems, tailored
151 management practices, and improved marketing
152 (Braun et al. 1998; Dixon et al. 2006) The increased
153 grain production attributable to improved germplasm
154 alone has been valued at up to US$ 6 billion year-1
155 (Lantican et al. 2005). The increased production of
156 wheat (and other staples) led to lower food prices
157 (von Braun 2007), which contributed to a reduction in
158 the proportion of poor in developing countries (Chen
159 and Ravallion 2007). Looking to the future, the
160 global population is projected to steadily increase,
161 albeit at a decreasing rate compared to the past
162 century, to around nine billion in 2050. The food and
163 other needs of the growing population underpin the
164 strong demand for cereals. The demand for wheat,
165 based on production and stock changes, is expected
166 to increase from 621 million t during 2004–2006 to
167 760 million tons in 2020 (Rosegrant et al. 2001), to
168 around 813 million t in 2030, and to more than
169 900 million t in 2050 (FAO 2006, 2007; Rosegrant
170 et al. 2007) implying growth rates of 1.6% for 2005–
171 2020, 1.2% for 2005–2030, and 0.9% for 2005–2050.
172 The International Wheat Improvement Network
173 The history of the International Maize and Wheat
174 Improvement Center (CIMMYT) involvement in
175 wheat improvement begins in the 1940s, more than
176 20 years before it was officially founded as an
177 international organization in 1966 (Ortiz et al.
178 2007b). Its roots reach back to the Office of Special
179 Studies (OSS), a research project sponsored by the
180 Mexican government and the Rockefeller Foundation
181 that was dedicated to improving maize, beans and
182 wheat, and later potatoes. The OSS began as a
183 research and training program focused on Mexico,
184 but soon began collaborating with other countries,
185 especially in South America. The OSS developed the
186 key organizational principles that would eventually
187 become central to the entire network of the Consul-
188 tative Group on International Agricultural Research
189 (CGIAR) centers. The OSS wheat program, led by
190 Nobel Peace Laureate Norman E. Borlaug, did crop
191breeding along with on-farm research and extension
192demonstrations aimed at introducing new technology
193to producers.
194For many decades the global average yield of
195wheat has increased, supported by an effective
196International Wheat Improvement Network (IWIN),
197an alliance of National Agricultural Research Systems
198(NARS), CIMMYT, the International Center for
199Agricultural Research in the Dry Areas (ICARDA)
200and advanced research institutes (ARI). This alliance
201has deployed cutting-edge science alongside practical
202multi-disciplinary applications resulting in the devel-
203opment of germplasm, which has improved food
204security and the livelihoods of farmers in developing
205countries. For example, during the late 1950s and
2061960s, researchers in Mexico, under the leadership of
207Borlaug, developed the improved spring wheat germ-
208plasm, which launched the Green Revolution in India,
209Pakistan, and Turkey (Reynolds and Borlaug 2006).
210Collaboration was broadened during the 1970s to
211include Brazil, China, and other major developing
212country producers, and resulted in wheat cultivars
213with broader disease resistance, better adaptation to
214marginal environments, and tolerance to acid soils.
215During the 1980s, an international collaborative
216partnership between Turkey, CIMMYT, and ICAR-
217DA was established for winter wheat improvement in
218developing countries. The IWIN currently operates
219field evaluation trials in more than 250 locations in
220around 100 countries for testing improved lines of
221wheat in different environments.
222As a publicly-funded international research insti-
223tute, CIMMYT regards its research products as
224international public goods. The main objective for
225regional and global multi-location testing is the
226identification of useful genetic diversity that will lead
227to research products, parental germplasm, or ulti-
228mately cultivars adapted to targeted wheat-production
229environments and systems in the developing world.
230Multi-location testing and data exchange increase the
231selection efficiency of participating wheat breeders.
232Returned data are used to identify parents for
233subsequent crosses and to incorporate new genetic
234variability into advanced lines that are consequently
235able to cope with the dynamics of abiotic and biotic
236stresses affecting wheat farming systems.
237With the growing research capacity of NARS in
238many major wheat-producing countries, the number
239of wheat cultivars released annually by developing
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
240 countries doubled to more than 100 cultivars by the
241 early 1990s (Lantican et al. 2005). The early era of
242 improved cultivars spread rapidly over the high
243 potential production areas in most developing
244 regions. Widespread adoption occurred most rapidly
245 in South Asia, especially in irrigated areas, followed
246 by the rainfed areas of Latin America; adoption has
247 been slower in the Middle East, North Africa, and
248 sub-Saharan Africa because of drier riskier environ-
249 ments and weaker institutions (Evenson and Gollin
250 2003; Lantican et al. 2005). With such widespread
251 adoption accompanied by yield increases, average
252 annual rates of return for investments in wheat
253 research averaged around 50% year-1 (Alston et al.
254 2000). In addition, the urban poor benefited substan-
255 tially as production increases drove down wheat
256 prices.
257 The wheat genetic resources endowment
258 By the 1920s it was acknowledged that wheat
259 cultigens of the genus Triticum L. belonged to three
260 ploidy groups with chromosomes number of
261 2n = 2x = 14 (T. monococcum L.), 28 (T. turgidum
262 L. and T. timopheevii Zhuk.), and 42 (T. aestivum L.
263 em. Thell. and T. zhukovskyi Menabde & Ericz.).
264 However, world wheat production is almost entirely
265 based on two species: T. aestivum—also known as
266 common or bread wheat, which account for about
267 95% of world production, and T. turgidum ssp. durum
268 (Desf.) Husn.—known as macaroni or durum wheat,
269 which accounts for the remaining 5%. The other
270 cultivated species are largely historical relics.
271 Genetic resources in wheat can be categorized into
272 six broad groups (Frankel 1977; FAO 1983), namely
273 modern cultivars in current use, obsolete cultivars—
274 often the elite cultivars of the past and often found in
275 the pedigrees of modern cultivars, landraces, wild
276 relatives of crop species in the Triticeae Dumort. tribe,
277 genetic and cytogenetic stocks, and breeding lines.
278 These genetic resources represent the gene pool
279 potentially available to breeders and other users of
280 collections. This broad pool can be further subdivided
281 into primary, secondary, and tertiary gene pools
282 (Harlan and de Wet 1971). The primary pool consists
283 of the biological species, including cultivated, wild,
284 and weedy forms of the crop, and gene transfer in this
285 group is considered to be easy. The secondary gene
286pool has the coenospecies (or a group of ‘‘allies’’ or
287‘‘relatives’’ to a given taxon) from which gene transfer
288is possible but difficult, while the tertiary gene pool is
289composed of species from which gene transfer is
290possible but only with great difficulty. Clearly, the
291boundaries of these groups are fuzzy and also change
292with changes in technology. Consequently, several
293authors including Smartt (1984) and Konarev et al.
294(1986) have suggested the gene pools concept of
295Harlan and De Wet (1971) be modified to increase the
296number of gene pools from three to four to coincide
297with populations, species, genera, and tribes, respec-
298tively (Merezhko 1998). Unfortunately, even this
299simple concept is difficult to apply in wheat because
300of the lack of an accepted view on the classification of
301wheat species, the genus Triticum, and even the tribe
302Triticeae (von Bothmer et al. 1992; Merezhko 1998).
303The Wheat Genetics Resource Center at Kansas State
304University in the USA provides a comprehensive
305online source of information about wheat taxonomy,
306including a detailed comparison of the most often
307used classifications, as part of the GrainTax project
308(www.k-state.edu/wgrc/). Herein we will follow the
309most recent taxonomic treatment of Triticum and
310Aegilops L. of van Slageren (1994), which updated
311previous research by Hammer (1980) on the taxon-
312omy and nomenclature of the genus Aegilops.
313The cultivated species of Triticum and their
314genomic constitution are given in Table 2. It should
315be noted that there are two valid biological species at
316each ploidy level. The diploid T. monococcum L. has
317both cultivated and wild forms, while T. urartu
318Tumanian only exists in the wild. Both tetraploid
319forms exist in both cultivation and in the wild, while
320both hexaploid species only exist in cultivation.
321The distribution of these species is described by Gill
322and Friebe (2002). Aegilops is the most closely related
323genus to Triticum and has been widely used in
324wheat improvement. All Aegilops are annuals. The
325genus consists of 11 diploid species and 12 poly-
326ploid species, including tetraploids and hexaploids
327(Table 3). Their taxonomy and distribution is
328discussed by van Slageren (1994). Dasypyrum
329[Haynaldia] villosum (L.) Cand. is among the Triti-
330ceae species and is also a genetic resource for wheat
331breeding. It is an annual with a V genome and is easily
332hybridized to durum or bread wheat. Each of the
333chromosomes was added to common wheat by the late
334E. Sears (Global Crop Diversity Trust 2007).
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
335 In addition to Aegilops, a host of more distantly
336 related annual and perennial members of related
337 genera in the Triticeae have potential as sources of
338 germplasm in wheat breeding including cultivated
339 rye and barley and their near relatives, as well as a
340 host of perennial grasses. The bulk of the perennial
341 Triticeae species have been difficult to exploit in
342 wheat improvement primarily because their genomes
343 are non-homologous to those of wheat, and genetic
344 transfers cannot be made by homologous recombi-
345 nation. However, gene transfer is possible via
346 complex cytogenetic protocols. Over the last three
347 decades hybridization per se has become less of a
348 problem in inter-specific hybridization between
349 Triticum species and more distantly related genera,
350 although achieving timely practical outcomes using
351 cytogenetic techniques is difficult in genera other
352 than Secale L. and Thinopyrum (Mujeeb-Kazi and
353 Rajaram 2002). Nevertheless, variation for econom-
354 ically-important traits such as host plant resistance to
355the cereal rusts, salt-tolerance, and resistance to
356barley yellow dwarf virus have been transferred from
357perennial wild species into bread wheat. The disease-
358resistant genes have been used in modern wheat
359cultivars. Mujeeb-Kazi and Hettel (1995) provided a
360comprehensive account of interspecific hybridization
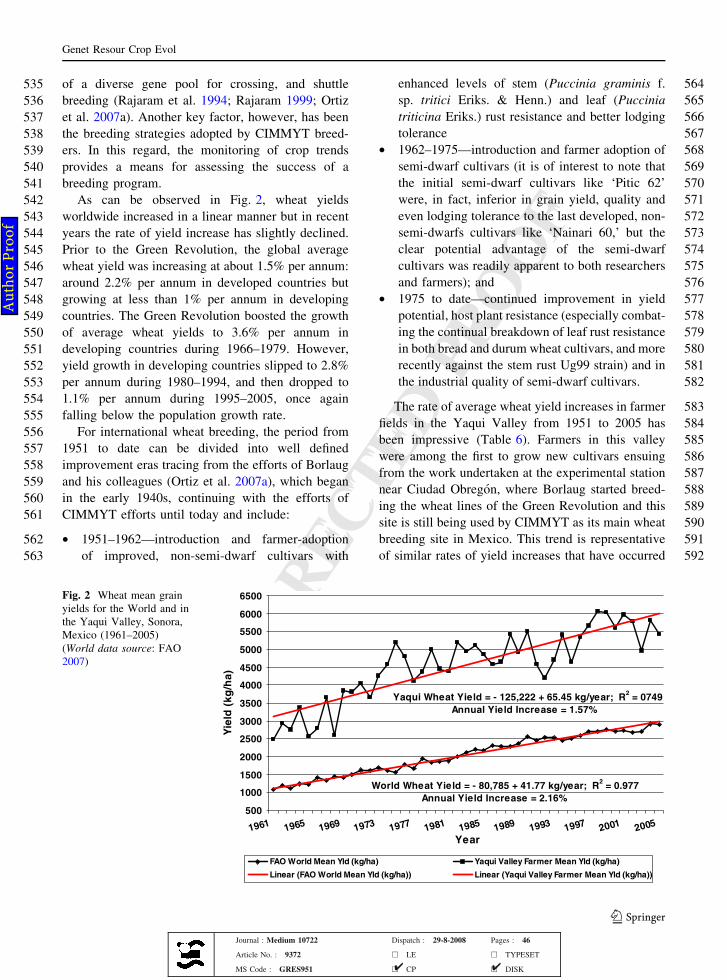
361in the Triticeae. The perennial genera of the tribe
362Triticeae of interest in wheat improvement are given
363in Table 4 along with their genome designations and
364ploidy levels. All the genomes of the perennial
365Triticeae have been combined with the A, B, and D
366genomes of bread wheat (Mujeeb-Kazi 1995).
367Wheat ex situ conservation strategy
368Due to the strategic importance of wheat in food
369security and trade in many countries, and the critical
370importance of breeding in ensuring national industries
371remain competitive, over 80 autonomous germplasm
Table 2 Species of genusTriticum and their genomicconstitution (After Gill andFriebe 2002)
a Related to S-genomespecies, cf. Table 3
Species Genomic constitution
Nuclear Organellar
Triticum aestivum L. ABD B (rel. to S)a
Triticum aestivum ssp. aestivum (common or bread wheat)
Triticum aestivum ssp. compactum (Host) Mackey (club wheat)
Triticum aestivum ssp. macha (Dekapr. & A. M. Menabde) Mackey
Triticum aestivum ssp. Spelta (L.) Thell. (large spelt or dinkel wheat)
Triticum aestivum ssp. sphaerococcum (Percival) Mackey (Indian dwarf wheat)
Triticum turgidum L. AB B (rel. to S)
Triticum turgidum ssp. carthlicum (Nevski) A. Love & D. Love (Persian wheat)
Triticum turgidum ssp. dicoccoides (Korn. ex Asch. et Graebn.) Thell. (wild emmer)
Triticum turgidum ssp. dicoccon (Schrank) Thell. (emmer wheat)
Triticum turgidum ssp. durum (Desf.) Husn. (macaroni or durum wheat)
Triticum turgidum ssp. paleocolchicum (Dekapr. et Menabde) Mac Key ex Hanelt
Triticum turgidum ssp. polonicum (L.) Thell. (Polish wheat)
Triticum turgidum ssp. turanicum (Jakubz.) A. Love & D. Love (Khorassan wheat)
Triticum turgidum ssp. turgidum (pollard wheat)
Triticum zhukovskyi Menabde & Ericz. AtAmG A (rel. to S)
Triticum timopheevii (Zhuk.) Zhuk. AtG G (rel. to S)
Triticum timopheevii ssp. Armeniacum (Jakubz.) Slageren (wild form)
Triticum timopheevii ssp. timopheevii (cultivated form)
Triticum monococcum L. Am Am
Triticum monococcum ssp. aegilopoides (Link) Thell. (wild form)
Triticum monococcum ssp. monococcum (einkorn or small spelt wheat)
Triticum urartu Tumanian ex Gandilyan (wild form) A A
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
372 collections holding an estimated excess of 800,000
373 accessions have been established globally. These
374 collections vary in size and coverage; the largest have
375 over 100,000 accessions and the smallest a few
376 hundred. They also vary greatly in coverage. Most
377 collections evolved from breeders’ working collec-
378 tions and carry predominantly local or regional
379 cultivars—advanced, obsolete or landrace—as well
380 as introduced cultivars of interest to national or
381 regional breeders. There is often substantial duplica-
382 tion within, and certainly between these sorts of
383 collections. Virtually every wheat collection in the
384 world would carry common popular cultivars such as
385 ‘Marquis’ and ‘Bezostaya 1.’ However, there are also
386 numerous small specialist collections of wild wheat
387 relatives and genetic stocks.
388An important issue in developing a global strategy
389for the conservation of wheat genetic resources is
390deciding on the diversity of accessions to be included in
391the strategy (Merezhko 1998). One extreme view
392would be to limit the network to the primary gene
393pool—the cultivated species and the closely-related
394species with which they can be readily hybridized. The
395other extreme is that in themodernworld of transgenics
396all biological species are potential genetic resources for
397wheat breeding and the concepts of primary, second-
398ary, and tertiary gene pools are quaint and outmoded. It
399is suggested here, following Merezhko (1998), that we
400should restrict our focus toTriticum species and related
401genera of the Triticeae. This coverage aligns with the
402intention of the International Treaty on Plant Genetic
403Resources for Food and Agriculture (ITPGRFA).
Table 3 Aegilops species and their genomic constitution (After Gill and Friebe 2002 and modified as per chromosome pairingand DNA analysis following Dvorak 1998)
Species Genomic constitution
Nuclear Organellar
Aegilops bicornis (Forssk.) Jaub. & Spach Sb Sb
Aegilops biuncialis Vis. UM (UMo) U
Aegilops markgrafii (Greuter) K. Hammer (known as Aegilops caudata auct non L.) C C
Aegilops columnaris Zhuk. UM (UXco) U2
Aegilops comosa Sm. in Sibth. & Sm. spp. heldreichii (Holzm. ex Boiss.) Eig M M
Aegilops crassa Boiss. Dc1M
c(D
c1Xc) D
2
var. glumiaristata Eig Dc1Dc2M
c(D
c1Dc2Xc) –
Aegilops cylindrica Host DcCc
D
Aegilops geniculata Roth (syn. Ae. ovata auct. non L.) UM (UMo) M
o
Aegilops juvenalis (Thell.) Eig DMU (DcXcUj) D
2
Aegilops kotschyi Boiss. US (US1) S
v
Aegilops longissima Schweinf. & Muschl. S1 S12
Aegilops mutica Boiss. T T,T2
Aegilops neglecta Req. ex Bertol. (syn. Ae. triaristata) UM (UXn) U
var. recta (Zhuk.) Hammer UMN (UXtN) U
Aegilops peregrina (Hack. in J. Fraser) Maire & Weiller (syn. Ae. variabilis) US (US1) Sv
Aegilops searsii Feldman & Kislev ex Hammer Ss Sv
Aegilops sharonensis Eig Ssh S1
Aegilops speltoides Tausch S S,G,G2
Aegilops tauschii Coss. var. tauschii, var. strangulata (Eig) Tzvel. D D
Aegilops triuncialis L. UCt
U,C2
Aegilops umbellulata Zhuk. U U
Aegilops uniaristata Vis. N N
Aegilops vavilovii (Zhuk.) Chennav. DMS (DcXcSv) D
2
Aegilops ventricosa Tausch DvNv
D
Underlined genomes are modified at the polyploidy level; those in brackets were deduced from DNA analysis
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
Table 4 The nuclear genome of the perennial species of the tribe Triticeae (after Mujeeb-Kazi and Wang 1995)
Species Genome Species Genome
Agropyron cristatum (L.) Gaertn. PP Leymus angustus (Trin.) Pilger NNNNNNXXXXXX
Agropyron cristatum PPPPPP Leymus arenarius (L.) Hochst. NNNNXXXX
Agropyron desertorum (Fisch. ex Link)
Schult.
PPPP Leymus chinensis (Trin.) Tzvelev NNXX
Agropyron fragile (Roth) P. Candargy PP Leymus cinereus (Scribn. & Merr.) A. Love NNXX
Agropyron michnoi Roshev. PPPP Leymus innovatus (Beal) Pilger NNXX
Agropyron mongolicum Keng PP Leymus mollis (Trin.) Pilger NNXX
Australopyrum pectinatum (Labill.) A. Love WW Leymus racemosus (Lam.) Tzvelev NNNNXXXX
Elymus abolinii (Drobow) Tzvelev SSYY Leymus salinus (M.E. Jones) A. Love NNXX
Elymus alatavicus (Drobow) A. Love SSYYPP Leymus triticoides (Buckl.) Pilger NNXX
Elymus arizonicus (Scribn. & J. G. Sm.)
Gould
SSHH Pascopyrum smithii (Rydb.) A. Love SSHHNNXX
Elymus batalinii (Krasn.) A. Love SSYYPP Psathyrostachys alatavicus NN
Elymus canadensis L. SSHH Psathyrostachys fragilis (Boiss.) Nevski NN
Elymus caninus (L.) L. SSHH Psathyrostachys huachanica Keng NN
Elymus ciliaris (Trin.) Tzvelev SSYY Psathyrostachys junceus (Fisch.) Nevski NN
Elymus dahuricus Turcz. ex Griseb. SSHHYY Psathyrostachys kronenburgii (Hackel) Nevski NN
Elymus drobovii Turcz. ex Griseb. SSHHYY Pseudoroegneria deweyii Jensen, Hatch & Wipff SSPP
Elymus gmelinii (Ledeb.) Tzvelev SSYY Pseudoroegneria tauri (Boiss. & Balansa) A. Love SSPP
Elymus grandiglumis (Keng) A. Love SSYYPP Pseudoroegneria libanotica (Hackel) D. R. Dewey SS, SSSS
Elymus kamoji (Ohwi) S. L. Chen SSHHYY Pseudoroegneria spicata (Pursh) A. Love SS, SSSS
Elymus kengii (Tzvelev) A. Love SSYYPP Pseudoroegneria stipifolia (Czern. ex Nevski) A. Love SS, SSSS
Elymus longearistatus (Boiss.) Tzvelev SSYY Pseudoroegneria strigosa (M.Bieb.) A. Love SS, SSSS
Elymus panormitanus (Parl.) Tzvelev SSYY Secale montanum = Secale strictum (Presl) Presl. RR
Elymus parviglume(Keng) A. Love SSYY Thinopyrum bessarabicum (Savul. & Rayss) A. Love JJ
Elymus pendulinus (Nevski) Tzvelev SSYY Thinopyrum caespitosum C. Koch) Barkw. et
D. R. Dewey
EESS
Elymus shandongensis B. Salomon SSYY Thinopyrum curvifolium (Lange) D. R. Dewey JJJJ
Elymus sibiricus L. SSHH Thinopyrum distichum (Thunb.) A. Love JJEE
Elymus strictus (Keng) A. Love SSYY Thinopyrum elongatum (Host) D. R. Dewey EE
Elymus tsukushiensis Honda SSHHYY Thinopyrum intermedium (Host) Barkworth &
D. R. Dewey
JJJJSS, JJEESS,
EEEESS
Elymus ugamicus Drobow SSYY Thinopyrum junceiforme (A. & D. Love) A. Love JJEE
Elymus vaillantianus (Wulfen ex Schreb.) K.
B. Jensen
SSHH Thinopyrum junceum (L.) A. Love p.p. JJJJEE
Elytrigia repens (L.) Desv. ex B. D. Jackson SSSSHH Thinopyrum nodosum [= Lophopyrum nodosum
(Nevski) A. Love]
EESS
Hordeum bogdanii Wilensky HH to
HHHHHH
Thinopyrum ponticum (Podp.) Z.-W. Liu &
R.-C. Wang
JJJJEEEEEE
Hordeum brevisubulatum Link HH to
HHHHHH
Thinopyrum sartorii (Boiss. & Heldr.) A. Love JJEE
Hordeum iranicum (Bothmer) Tzvelev HH to
HHHHHH
Thinopyrum scirpeum (C.Presl) D. R. Dewey EEEE
Hordeum jubatum L. HH to
HHHHHH
Thinopyrum scythicum EESS
Hordeum violaceum Boiss. & Hohen. HH to
HHHHHH
Thinopyrum turcicum JJJJEEEE
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
404 The growing size and sophistication of genetic and
405 molecular stock collections is testimony to their
406 increasing contributions to enable the effective utili-
407 zation of the variation conserved in ‘‘traditional’’
408 germplasm collections. The role of genetic stock
409 collections in the global conservation effort of wheat
410 germplasm should be re-evaluated and these should
411 be given a higher priority in a rationalized system
412 than they had in the past.
413 The Global Wheat, Rye, and Triticale Conservation
414 Strategy (Global Crop Diversity Trust 2007) suggested
415 that the following criteria are essential for an efficient
416 and effective global system for the conservation of
417 wheat genetic resources: globally or regionally-impor-
418 tant, accessible under the internationally agreed terms
419 of access and benefit sharing provided for in the
420 multilateral system as set out in the ITPGRFA,
421 committed to the long-term conservation of the unique
422resources it holds, well-managed and in conformity
423with agreed upon scientific and technical standards of
424management, maintaining effective links to users of
425plant genetic resources, and an indicatedwillingness to
426act in partnership with others to achieve a rational
427system for conserving wheat genetic resources.
428Twenty-three private, national, and global collections
429that fulfilled these criteria where identified as key
430partners for a global wheat conservation network
431(Table 5).
432The proposed wheat conservation strategy focuses
433on the conservation and use of the full spectrum of
434the genetic resources of wheat with the exception of
435the perennial wild relatives. Modern and obsolete
436improved cultivars are generally well-conserved in
437global wheat germplasm collections because many
438such collections either were derived from breeders
439working collections or were primarily established to
Table 5 Collections of a global network of wheat genetic resources
Country Institute No. ofaccessions
Global CIMMYT, El Batan, Mexico 111,681
USA USDA-ARS, National Small Grains Facility, Aberdeen, Idaho 56,218
Russia N.I. Vavilov Research Institute of Plant Industry (VIR), St. Petersburg 39,880
Global ICARDA, Aleppo, Syria 37,830
India National Bureau of Plant Genetic Resources (NBPGR), New Delhi 32,880
Australia Australian Winter Cereals Collection, Tamworth 23,917
France INRA Station d’Amelioration des Plantes, Clermont-Ferrand 15,850
Iran National Genebank of Iran, Genetic Resources Division, Karaj 12,169
Czech Republic Research Institute of Crop Production, Prague 11,018
Ethiopia Plant Genetic Resources Centre, Institute of Biodiversity Conservation and Research, Addis Ababa 10,745
Bulgaria Institute for Plant Genetic Resources ‘‘K. Malkov’’, Sadovo 9,747
Germany Genebank, Institute for Plant Genetics and Crop Plant Research (IPK), Gatersleben 9,633
United Kingdom Department of Applied Genetics, John Innes Centre, Norwich 9,584
Cyprus National Genebank (CYPARI), Agricultural Research Institute, Nicosia 7,696
Japan Genetic Resources Management Section, NIAR (MAFF), Tsukuba 7,148
Switzerland Station Federale de Recherches en Production Vegetale de Changins, Nyon 6,996
Turkey Plant Genetic Resources Department, Aegean Agricultural Research Institute, Izmir 6,381
Netherlands Centre for Genetic Resources, Wageningen 5,529
Canada Plant Gene Resources of Canada, Winnipeg 5,052
USA Wheat Genetics Resource Center, Kansas State University, Manhattan 5,000
Japan Plant Germplasm Institute, Graduate School of Agriculture, Kyoto University 4,378
Spain Centro de Recursos Fitogeneticos, INIA, Madrid 3,183
Sweden Nordic Gene Bank, Alnarp 1,843
Total 23 institutes 434,358
Source: Global Crop Diversity Trust (2007)
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
440 service local or regional breeding programs, and
441 these were the accessions most sought by breeders. In
442 fact, many important cultivars are conserved in the
443 majority of national and international collections. The
444 major focus of a global strategy for this category of
445 genetic resource should be to reduce redundancy in
446 the global set of collections to free up resources for
447 other priorities.
448 Landraces have received priority for collection,
449 conservation, and documentation in recent years,
450 supported by the efforts of FAO, the CGIAR, and
451 others because of the increasing threat to their
452 continued existence by the spread of improved
453 modern cultivars. Nevertheless, such cultivars are
454 poorly represented in world collections compared to
455 modern and obsolete cultivars and should remain
456 a priority for the global strategy, both to ensure the
457 collection of material that is not in collections but
458 that still exists in the field and that the long-term
459 conservation of collected material is in line with
460 agreed upon international standards.
461 The wild relatives of wheat are also generally
462 poorly represented in global wheat germplasm col-
463 lections. There are several reasons for this. First, wild
464 relatives are seldom used in conventional breeding
465 programs as compared to cultivars of the same
466 species and usually require an extensive period of
467 germplasm enhancement. Wild species tend to be
468 collected and used by the small number of specialist
469 institutes concerned with interspecific hybridization.
470 Second, they are more difficult to seed increase and
471 maintain because of their tendency to shatter their
472 seed as compared crop cultivars. For this reason also,
473 the distribution and use of some wild species is
474 limited because of their potential as weeds. Finally,
475 wild species, because of their capacity to self-
476 reproduce in nature, have been seen as under less
477 threat of extinction than the cultivated landraces
478 (Global Crop Diversity Trust 2007).
479 Unfortunately, many populations of the annual
480 wild relatives of wheat, particularly those at the
481 extremes of their distribution that are of special
482 interest for breeding purposes, are under threat
483 because of changing patterns of land use and global
484 warming. At the same time, new technologies have
485 made the use of the annual wild relatives as a
486 germplasm source easier, which has generated an
487 interest and need for representative collections of
488 annual wild relatives to be maintained in accessible
489collections. For these reasons the annual wild rela-
490tives should clearly be afforded a greater priority in
491the global wheat germplasm collections than they
492have had in the past. This is not to suggest that all or
493many collections need to move to collect or conserve
494the wild relatives of wheat, but rather, that those with
495the specialized knowledge and capacity to undertake
496the collection and conservation of this category of
497germplasm should be given priority support.
498As noted above, it can also be argued that defined
499genetic stock collections should receive greater
500priority in a balanced global effort to conserve and
501make available for use the genetic resources of wheat.
502Again, because specialists need to develop and
503reliably maintain genetic stocks as true-to-type
504accessions, it is expected that defined genetic stocks
505will be maintained by specialized institutes. The
506emphasis will be to support those institutes to develop
507a coordinated system that replaces the largely ad hoc
508system that has operated to date for the conservation
509of genetic stocks so that valuable material once
510developed and in the public domain is available on a
511continuing basis for all who need it.
512The perennial wild relatives of wheat were not seen
513as a priority for conservation in the collective global
514wheat germplasm system. Again, there are several
515reasons for this. The first, and perhaps most important,
516is that collections of many of these species are
517maintained in perennial grass collections for use in
518breeding programs as grazing species or for other
519uses. Second, despite the number of perennial wild
520relatives of wheat that exist, their extensive global
521spread, and the extensive research that has taken
522place, the number of examples of commercially-
523successful gene transfer from perennial wild relatives
524to wheat remains modest. Third, the perennial wild
525relatives, like their annual counterparts, require spe-
526cialized seed increase knowledge and facilities, which
527is only likely to be available in specialized collections.
528A brief account of wheat breeding at CIMMYT
529and its impact in grain yield
530The global impact of the wheat breeding program of
531CIMMYT has been significant and well-documented
532(Rajaram 1999; Trethowan et al. 2001). Many factors
533have contributed to CIMMYT’s success, such as
534breeding targeted to mega-environments (MEs), use
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of
UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
535 of a diverse gene pool for crossing, and shuttle
536 breeding (Rajaram et al. 1994; Rajaram 1999; Ortiz
537 et al. 2007a). Another key factor, however, has been
538 the breeding strategies adopted by CIMMYT breed-
539 ers. In this regard, the monitoring of crop trends
540 provides a means for assessing the success of a
541 breeding program.
542 As can be observed in Fig. 2, wheat yields
543 worldwide increased in a linear manner but in recent
544 years the rate of yield increase has slightly declined.
545 Prior to the Green Revolution, the global average
546 wheat yield was increasing at about 1.5% per annum:
547 around 2.2% per annum in developed countries but
548 growing at less than 1% per annum in developing
549 countries. The Green Revolution boosted the growth
550 of average wheat yields to 3.6% per annum in
551 developing countries during 1966–1979. However,
552 yield growth in developing countries slipped to 2.8%
553 per annum during 1980–1994, and then dropped to
554 1.1% per annum during 1995–2005, once again
555 falling below the population growth rate.
556 For international wheat breeding, the period from
557 1951 to date can be divided into well defined
558 improvement eras tracing from the efforts of Borlaug
559 and his colleagues (Ortiz et al. 2007a), which began
560 in the early 1940s, continuing with the efforts of
561 CIMMYT efforts until today and include:
562 • 1951–1962—introduction and farmer-adoption
563 of improved, non-semi-dwarf cultivars with
564enhanced levels of stem (Puccinia graminis f.
565sp. tritici Eriks. & Henn.) and leaf (Puccinia
566triticina Eriks.) rust resistance and better lodging
567tolerance
568• 1962–1975—introduction and farmer adoption of
569semi-dwarf cultivars (it is of interest to note that
570the initial semi-dwarf cultivars like ‘Pitic 62’
571were, in fact, inferior in grain yield, quality and
572even lodging tolerance to the last developed, non-
573semi-dwarfs cultivars like ‘Nainari 60,’ but the
574clear potential advantage of the semi-dwarf
575cultivars was readily apparent to both researchers
576and farmers); and
577• 1975 to date—continued improvement in yield
578potential, host plant resistance (especially combat-
579ing the continual breakdown of leaf rust resistance
580in both bread and durum wheat cultivars, and more
581recently against the stem rust Ug99 strain) and in
582the industrial quality of semi-dwarf cultivars.
583The rate of average wheat yield increases in farmer
584fields in the Yaqui Valley from 1951 to 2005 has
585been impressive (Table 6). Farmers in this valley
586were among the first to grow new cultivars ensuing
587from the work undertaken at the experimental station
588near Ciudad Obregon, where Borlaug started breed-
589ing the wheat lines of the Green Revolution and this
590site is still being used by CIMMYT as its main wheat
591breeding site in Mexico. This trend is representative
592of similar rates of yield increases that have occurred
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
5500
6000
6500
1961 1965 1969 1973 1977 1981 1985 1989 1993 1997 2001 2005
Year
Yie
ld (
kg
/ha
)
FAO World Mean Yld (kg/ha) Yaqui Valley Farmer Mean Yld (kg/ha)
Linear (FAO World Mean Yld (kg/ha)) Linear (Yaqui Valley Farmer Mean Yld (kg/ha))
World Wheat Yield = - 80,785 + 41.77 kg/year; R2 = 0.977
Annual Yield Increase = 2.16%
Yaqui Wheat Yield = - 125,222 + 65.45 kg/year; R2
= 0749
Annual Yield Increase = 1.57%
Fig. 2 Wheat mean grainyields for the World and inthe Yaqui Valley, Sonora,Mexico (1961–2005)(World data source: FAO2007)
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
593 in many other wheat production areas such as India,
594 Pakistan, and China, among many others (especially
595 irrigated production regions), notably since the
596 introduction and adoption of semi-dwarf cultivars
597 with resistance to the various rust diseases (Reynolds
598 and Borlaug 2006). Clearly, the tendency has been
599 towards a reduced rate of yield increases over time.
600 This is a fact of considerable concern and is likely not
601 restricted to the Yaqui Valley as shown by the global
602 trends (Fig. 2). It would appear that factors associated
603 with the declining rate of yield increases in wheat
604 include relatively slow increases in private sector
605 investments during the last decade, and lower appli-
606 cations of production inputs as oil prices drove up the
607 cost of fertilizer and pumping irrigation water while,
608 until very recently, the price of wheat gradually fell.
609 In addition, a lack of attention to crop management
610 and the degradation of resources including soil
611 fertility and quality of water for irrigation, combined
612 with an increasing frequency of droughts.
613 Genetic enhancement of spring bread wheat
614 The spring bread wheat germplasm developed at
615 CIMMYT is targeted for its adaptation to diverse
616 wheat production environments in the developing
617 world. The breeding program is based in Mexico and
618 shuttles germplasm between two contrasting environ-
619 ments: Ciudad Obregon (280 N, 32 m.a.s.l.) in
620 northwestern Mexico and Toluca (180 N, 2,640
621 m.a.s.l.) in the highlands of Central Mexico, thereby
622 achieving two generations a year (Braun et al. 1996).
623 This shuttle breeding exposes wheat materials to
624 diverse photoperiod and temperatures and to a range
625 of important diseases. The lines developed through this
626 process are then tested widely around the world and
627selected materials, based on international perfor-
628mance, are identified for continued crossing. There
629are two major breeding thrusts at present: germplasm
630for irrigated areas, and germplasm for rainfed areas.
631Breeding objectives
632Traits of foremost importance in spring wheat
633improvement include: (1) grain yield potential, stabil-
634ity, and wide adaptation, (2) potential for durable
635resistance to diseases such as stem, leaf, and yellow
636(Puccinia striiformis West.) rusts, Septoria tritici
637blotch, Fusarium head blight or scab, and root rots,
638(3) water-use efficiency and water productivity,
639(4) heat tolerance, and (5) end-use quality character-
640istics. Breeding objectives and schemes are
641continually modified to maintain the efficiency and
642effectiveness of germplasm products. For example, as
643water resources continue to decline, wheat will have to
644be produced with less water. This requires the devel-
645opment of high-yielding cultivars that are also efficient
646in water use for irrigated areas or have improved
647performance under drought for rainfed areas. Expan-
648sion of resource-conserving technologies, for example
649zero-tillage in many countries not only reduces
650production costs but also increases long-term sustain-
651ability. However, it is evident that breeding objectives
652must be modified to develop a different kind of
653germplasm that has better emergence and growth
654characteristics and resistance to those diseases and
655pests that survive on residues (Joshi et al. 2007a).
656Yield potential, yield stability, and wide
657adaptation
658The yield potential of semi-dwarf wheat cultivars,
659irrespective of their origin, has continued to increase at
Table 6 Annual rates of increase in average wheat yield in farmer fields in the Yaqui Valley, Sonora, Mexico (1951–2005)
Periods of cultivar development Yield increaseyear-1 (%)
Yield increaseyear-1 (kg ha-1)
Coefficient of determination(year versus yield)
From the first improved non-semi-dwarfs to date 2.36 81 0.857
Improved non-semi-dwarfs 5.20 110 0.808
First generation semi-dwarf cultivars 3.00 111 0.569
Second generation semi-dwarf cultivars 0.15 9 0.011
Further semi-dwarf cultivar development withmodest farmer adoption
-0.43 -23 0.040
Second generation semi-dwarf cultivars to date 0.80 44 0.337
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
660 the rate of about 1% annually. Yield stability and wide
661 adaptation are important traits that must be present
662 together with yield potential to ensure that a genotype
663 maintains its superiority in diverse environments,
664 management practices, and biotic and abiotic stresses.
665 Breeding for specific adaptations has not been very
666 successful because in most areas, temperatures and
667 rainfall patterns shift annually. The wide adaptation
668 and stable performance of CIMMYT-derived wheat
669 lines and cultivars are largely due to shuttle breeding
670 in Mexico where segregating populations are selected
671 in two contrasting environments under diverse dis-
672 eases and abiotic stresses, followed by international
673 multi-location testing of advanced lines. This
674 approach is capable of identifying the best stable
675 performers in a single year of testing.
676 Various studies have shown that increases in yield
677 potential are mainly associated with increased bio-
678 mass, kernel number, and harvest index (Sayre et al.
679 1997). Yield components, such as grain size and
680 number, or harvest index in more recent germplasm,
681 have made relatively little or no contribution in
682 explaining the increases in yield potential. This
683 would mean that selection for increased yield
684 potential and higher kernel weight can proceed
685 simultaneously. Large kernel size continues to be
686 an important trait in local markets of various
687 developing countries and appears to be associated
688 with better emergence under poor management.
689 Some of the recent wheat germplasm developed at
690 CIMMYT has not only shown increased grain yield
691 potential but also kernel weight as high as 60 mg in
692 northwestern Mexico, compared with about 40 mg
693 for most of the wheat germplasm developed during
694 the 1980s and 1990s.
695 Although the early increases in yield potential of
696 semi-dwarf wheat cultivars came from the incorpo-
697 ration of dwarfing genes, subsequent progress can be
698 attributed to additive genes. It is likely that intense
699 breeding efforts during the last three decades in the
700 post-Green Revolution era had already selected for
701 larger effect additive genes. If that is the case, then
702 further progress is expected from selecting genes that
703 have much smaller effects, thus making it necessary
704 to modify the commonly used traditional breeding
705 schemes. We began utilizing a single-backcross
706 crossing approach that was initially aimed at incor-
707 porating resistance to rust diseases based on multiple
708 additive, minor genes (Singh and Huerta-Espino
7092004). However, it soon became apparent that the
710single-backcross approach also favored selection of
711genotypes with higher yield potential. The reason
712why single backcross shifts the progeny mean toward
713the higher side is that it favors retaining most of the
714desired additive genes from the backcross or recur-
715rent parent, while simultaneously allowing the
716incorporation and selection of additional useful
717small-effect genes from the donor parent.
718A selected bulk-breeding scheme was introduced
719in bread wheat improvement in the mid-1990s.
720According to Singh et al. (1998b) selection schemes
721have little or no effect on the performance of progeny
722lines, the choice of parents determines the progeny
723response. In all segregating generations until F5 or F6,
724one spike from each of the selected plants is
725harvested as bulk and a sample of seed is used in
726growing the next generation. Individual plants or
727spikes are harvested in the F5 or F6 generation. This
728scheme allows retaining a larger sample of selected
729plants and was found to be highly efficient in terms of
730operational costs. Moreover, retaining a large sample
731of plants in segregating populations increases the
732probability of identifying rare segregates that carry
733most desired genes.
734Introgression of new genetic diversity from unre-
735lated wheat germplasm, including inter-specific
736hybridization, can create a new genetic pool and
737bring in large or small-effect genes that may not be
738present in wheat germplasm commonly used in a
739breeding program. Alien translocation T7DS.7DL-
7407Ae#1L from Thinopyrum elongatum (Host) D.R.
741Dewey that carries leaf and stem rust resistance genes
742Lr19 and Sr25, respectively, has been shown to
743increase yield potential ranging from almost non-
744significant levels to over 15%, depending on genetic
745background under irrigated conditions through
746increased biomass production (Singh et al. 1998a)
747associated with increased spike fertility and photo-
748synthetic rate (Reynolds et al. 2001). Thus, its
749widespread incorporation is underway and we expect
750a quantum jump in yield potential in some of the
751resultant lines.
752Breeding to safeguard wheat crops
753from important diseases
754One or more of the three rust diseases of wheat
755continue to pose major breeding challenges worldwide
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
756 due to the pathogens’ ability to evolve continuously, to
757 migrate long distances, and to overcome the deployed
758 race-specific resistance genes. Breeding approaches to
759 control stem rust are described in the below section
760 ‘‘Ug 99 stem rust as a global emerging threat to wheat
761 (food) supply ‘‘ A major genetics and breeding
762 emphasis during the last decades was given to
763 accumulate slow-rusting, minor-resistance genes with
764 additive effects against leaf and yellow-rust patho-
765 gens. High diversity exists in CIMMYT spring wheat
766 lines for such genes and new wheat lines that show
767 negligible disease severity at maturity often carry
768 between four and five slow-rusting resistance genes.
769 Recent studies have found that some of the studied
770 slow-rusting resistance genes have pleiotropic effects
771 on multiple diseases. Most known pleiotropic genes
772 are Lr34/Yr18/Pm38/Bdv1 and Lr46/Yr29/Pm39. Past
773 experience deploying cultivars with slow-rusting
774 resistance has shown that such resistance is durable.
775 CIMMYT initiated breeding for resistance to
776 Septoria tritici blotch, caused by Mycosphaerella
777 graminicola (Fuckel) J. Schrot. (anamorph Septoria
778 tritici)), in semi-dwarf wheat in early 1970 and steady
779 progress has been made since then. Currently, several
780 high-yielding semi-dwarf wheat lines with good
781 resistance are available. Resistance in these wheat
782 genotypes is derived from diverse sources, including
783 re-synthesized wheat lines. Two high rainfall sites
784 (Toluca and Patzcuaro at Michoacan, Mexico), were
785 used for selection. Some re-synthesized wheat lines
786 developed at CIMMYT have shown excellent resis-
787 tance that appears to be leading towards immunity to
788 the disease. These sources offer new genetic diversity
789 of resistance originating from durum wheat or
790 Aegilops tauschii Coss. A high level of resistance
791 from original re-synthesized wheat parents was
792 successfully transferred to derived high-yielding
793 lines.
794 Sources of resistance to scab have been divided
795 into three groups: China and Japan, Argentina and
796 Brazil, and Eastern Europe. More recently, additional
797 sources, including some hexaploid-derived lines from
798 re-synthesized wheat parents have also been identi-
799 fied to carry moderate resistance. Earlier genetic
800 analysis indicated that a few additive genes confer
801 resistance in Chinese and Brazilian wheat lines, and
802 genes present in Chinese sources are different from
803 those in Brazilian sources. Although several genomic
804 regions are now known to contribute quantitative
805resistance (Anderson et al. 2001; Buerstmayr et al.
8062002), a gene from the Chinese cultivar ‘Sumai 3’ in
807the short arm of chromosome 3B has shown the
808largest and most consistent effect in reducing disease
809severity and mycotoxin accumulation (Anderson
810et al. 2001). Further progress in enhancing the level
811of resistance beyond the current level can come from
812a breeding strategy that would favor the accumulation
813of multiple minor genes from various sources into a
814single genotype. CIMMYT is pursuing this strategy at
815present and its outcome will be known in the next
8163–4 years. Some of the recent research advances are
817given below in the section ‘‘Food safety and fighting
818wheat mycotoxins.’’
819Breeding for water-use efficiency and drought
820tolerance
821Wheat is increasingly being grown on marginal lands
822and in farming systems where inputs are limited. In
823most irrigated areas wheat is grown under insufficient
824irrigations. More water-efficient or drought-tolerant
825cultivars can mitigate the effects of changing pro-
826duction environments to some extent. Understanding
827of the genetic basis of drought tolerance is poor.
828Nevertheless, considerable progress has been made in
829yield improvement under drought in recent decades
830using the wheat gene pool and selecting under
831drought stress (Trethowan et al. 2002). The opportu-
832nity exists to improve the tolerance further if new
833genetic variability can be combined with existing
834variability and if the underlying genetic control of
835tolerance can be better understood.
836The re-synthesized wheat lines—developed by
837crossing modern durum wheat with Ae. tauschii, the
838probable donor of the D-genome in hexaploid
839wheat—have introduced new genetic variation into
840the wheat gene pool for many characters. Not
841surprisingly, re-synthesized wheat lines have also
842been a source of variation for drought and heat
843tolerance (Trethowan et al. 2002). Some advanced
844materials derived from re-synthesized wheat lines
845have improved adaptation worldwide, especially in
846drought-stressed environments.
847To improve the breeding efficiency for drought
848tolerance, the CIMMYT strategy is to ensure that
849drought-tolerant germplasm also maintains respon-
850siveness if more moisture becomes available in a
851season. The high yield potential and tolerance to
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
852 drought stress are not mutually exclusive and can be
853 bred simultaneously by selecting segregating popu-
854 lations under favorable environments and drought-
855 stress environments in alternate generations—a
856 practice used at CIMMYT. Moreover, to generate a
857 more precise drought stress at different growth
858 stages, a drip-irrigation system has been installed to
859 irrigate 17 ha of an experimental field near Ciudad
860 Obregon, where rainfall during the crop season in
861 most years is negligible. This system allows applica-
862 tion of exact amounts of water at chosen growth
863 stages to generate different drought scenarios repre-
864 senting different parts of the world.
865 Drought tolerance is a complex trait that is also
866 influenced by root diseases. Healthier roots use the
867 available soil moisture more efficiently. Fortunately,
868 resistance to nematodes and some root pathogens is
869 often simply inherited and molecular markers are
870 available to assist selection. CIMMYT uses these
871 markers to incorporate resistance in drought-tolerant
872 wheat materials.
873 Breeding for heat tolerance
874 The genetic control of heat tolerance, like drought
875 tolerance, is poorly understood. Nevertheless, signif-
876 icant variation for heat tolerance exists in the wheat
877 gene pool (Pfeiffer et al. 2005). In many environ-
878 ments, late planting can expose the crop and breeding
879 nurseries to high temperatures from flowering
880 onwards, giving wheat breeders the opportunity to
881 select lines with high levels of heat tolerance. At
882 CIMMYT, lines are selected during the segregating
883 phase for adaptation to heat by planting late. A
884 gravity table is used to separate bulk populations into
885 those that can maintain seed weight under high
886 temperature; the derived lines are then tested under
887 heat stress in yield trials. Physiological tools, such as
888 the infrared thermometer that measures canopy
889 temperature depression (CTD), are also available to
890 assist the plant breeder in discriminating among
891 progenies (Reynolds et al. 1998). Some details on
892 heat screening are also provided in the below section
893 on ‘‘Climate change adaptation and mitigation.’’
894 Heat avoidance or early maturity is an extremely
895 important trait to circumvent effects of high temper-
896 ature at grain filling. All popular cultivars currently
897 grown in the eastern Gangetic Plains are earlier-
898 maturing than cultivars popular in the northwestern
899Gangetic Plains. A simultaneous improvement of
900heat tolerance and yield potential of earlier-maturing
901germplasm is the best option to increase production
902in heat-stress environments and is being practiced
903(Joshi et al. 2007d).
904Breeding for end-use quality
905Bread wheat is generally milled into flour (both
906refined and whole meal) and made into leavened
907breads, flat breads, biscuits, and noodles. The quality
908of proteins, which have a large effect on end-use
909quality, is controlled by known high and low
910molecular weight glutenins and gliadins. A number
911of rapid, indirect quality tests are available that can
912be applied in the early generations to increase the
913probability of identifying progeny with the desired
914quality profiles. Dough rheological properties can be
915measured in different ways; some methods are time-
916consuming but accurate, e.g., the Alveograph, and
917others are faster, less expensive, but slightly less
918accurate, e.g., the Mixograph. Parents for crossing are
919chosen carefully for quality characteristics. Because
920the primary objective of CIMMYT’s breeding pro-
921gram is to enhance yield, quality tests are done in
922advanced generations or after yield testing. A high
923emphasis is being given to improve the leavened and
924flat bread quality characteristics. About a third of
925improved spring wheat materials developed and
926distributed in recent years have excellent to accept-
927able leavened and flat bread making characteristics.
928The section ‘‘Grain quality for adding value in the
929commodity chain’’ provides further details on this
930research area.
931Improving winter/facultative and high-latitude
932wheat
933In the late 1970s and early 1980s CIMMYT research-
934ers realized that winter/facultative wheat breeding for
935the developing world remained largely un-addressed.
936Small efforts to breed facultative wheat during the
937winter cycle in Toluca were primarily based on
938selection from the germplasm introduced from East-
939ern Europe and the USA. However, the winter in
940Toluca was not cold enough for the development of
941competitive lines. The target region for winter/
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
942 facultative wheat was in the Central and West Asia
943 region covering 15–20 million ha of the crop in
944 Turkey, Iran, Central Asia, and the Caucasus. The
945 early work in Toluca resulted in the identification of
946 good winter parents and some competitive lines that
947 were used mainly for spring 9 winter crosses with
948 Oregon State University. This germplasm was not
949 sufficiently adapted and neither was it on a scale to
950 provide the winter wheat breeding programs in the
951 target region with competitive material. Thus, CI-
952 MMYT established a winter/facultative wheat
953 breeding program outside of Mexico and directly in
954 the region. Turkey was chosen due to its diversity of
955 environments and because it is a major winter wheat
956 producer in the region. The agreement signed in 1981
957 between the Government of Turkey and CIMMYT
958 anticipated the development of new winter/faculta-
959 tive germplasm through a cooperative breeding
960 program.
961 The newly established program operated through
962 several key research institutions in Turkey: the
963 Central Field Crop Research Institute in Ankara, the
964 Anatolian Agric. Research Institute in Eskisehir, and
965 the Bahri Dagdas International Agric. Research
966 Center in Konya—the latter being established spe-
967 cifically to work on winter wheat breeding. The initial
968 breeding efforts were based on screening the large
969 collection of Turkish, East European, and US culti-
970 vars and making crosses. At the same time,
971 Spring 9 Winter Program operated at Oregon State
972 University (OSU) by Prof. W. Kronstad, supplied
973 F3–F4 populations originating from crosses between
974 Mexican spring wheat lines and winter wheat lines.
975 The lines selected from introduced germplasm in
976 Toluca were also sent to Turkey. All the populations
977 and lines from CIMMYT and OSU, germplasm from
978 Eastern Europe and the USA were screened in Turkey
979 and the best ones selected for distribution through
980 winter/facultative international nurseries. The Tur-
981 key-CIMMYT winter wheat program was joined by
982 ICARDA in 1999 to form the International Winter
983 Wheat Improvement Program (IWWIP). Eventually,
984 Toluca-based winter wheat activities were discontin-
985 ued. The winter/facultative germplasm presently
986 distributed from Turkey combines the germplasm
987 developed by IWWIP through its breeding program
988 in Turkey and Syria as well as new cultivars and
989 breeding lines from all cooperators who are willing to
990 share germplasm. The programs in Eastern Europe,
991the USA, and Central Asia routinely submit the
992germplasm that goes through a selection procedure
993and the best material is included for international
994distribution. This is a specific feature of the winter
995wheat program that is highly appreciated by the
996cooperators. Facultative and Winter Wheat Observa-
997tion Nursery (FAWWON) is distributed globally to
998over 100 cooperators in 50 countries. Additionally,
999yield trials are distributed to the target region of
1000Central and West Asia.
1001In retrospective, the establishment of a winter/
1002facultative breeding program in Turkey was a well-
1003justified move resulting in closing the gap in the
1004provision of germplasm for Central and West Asia.
1005The information on the release of cultivars originat-
1006ing from IWWIP has proved its success in developing
1007germplasm which is well-adapted to the region and
1008competitive with local material. By January 2008, 39
1009cultivars originating from IWWIP had been released
1010in 10 countries of Central and West Asia, including
1011Afghanistan, Iran, and Turkey. Because these culti-
1012vars are competitive in grain yield, they possess
1013higher levels of genetic protection against dominating
1014diseases and especially yellow rust. The genetic
1015diversity of these new cultivars is very broad as their
1016pedigree incorporates not only CIMMYT parents but
1017also a wide range of genetically non-related winter
1018wheat lines from Turkey, Iran, Russia, Ukraine,
1019Romania, Bulgaria, Hungary, and the whole diversity
1020of the US winter wheat.
1021The Turkey-based winter wheat program has now
1022been proven successful both in developing new
1023cultivars as well as in serving as a vehicle for
1024international germplasm exchange and will evolve
1025into a modern program able to efficiently address the
1026upcoming challenges. These are resistance to stem
1027rust, specifically against the Ug99 strain, resistance
1028and tolerance to seed and soil-born pathogens, and
1029bread-making quality, which still needs improve-
1030ment. These will be enhanced through wider
1031application of double haploids and marker-aided
1032breeding. The fundamental difference between winter
1033and spring wheat breeding at CIMMYT is one
1034generation of breeding per year versus two. This
1035should be compensated by the wider use of modern
1036breeding tools, e.g. doubled-haploids.
1037CIMMYT breeding efforts for high latitude spring
1038wheat were initiated in 1999 in order to address one
1039specific challenge: resistance to leaf rust. The vast
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1040 steppe region of northern Kazakhstan and western
1041 Siberia grows around 20 million ha of short season
1042 low input wheat in the environment ranging in
1043 precipitation from 250 to 450 mm. The cultivars
1044 currently grown still represent extensive, tall, and
1045 day-length sensitive type. The average yield ranges
1046 from 1.3 to 1.8 t ha-1. However, even in the years
1047 with sufficient precipitation the response in yield is
1048 limited due to widespread occurrence of leaf rust
1049 (Morgounov et al. 2007). Essentially, all the major
1050 cultivars grown in the region are highly susceptible to
1051 the pathogen. The shuttle breeding program between
1052 Kazakhstan/Siberia and Mexico had its major objec-
1053 tive to incorporate leaf rust resistance while
1054 maintaining the broad adaptability and superior
1055 bread-making quality. Most crosses are made in
1056 Mexico between the Kazakh/Siberian material and
1057 the best Mexican lines which are back- or top crossed
1058 by Kazakh/Siberian or Canadian parents. The result-
1059 ing F2 and F3 are screened under leaf rust pressure in
1060 the area with extended artificial light to allow
1061 selection of day-length sensitive genotypes. The
1062 selected F4 and F5 bulks are sent to two key sites in
1063 Kazakhstan (Astana) and Siberia (Omsk) for selec-
1064 tion under local conditions. The best populations are
1065 then distributed to all the breeding programs in the
1066 region through the Kazakhstan-Siberia Network for
1067 Spring Wheat Improvement (KASIB). By 2008
1068 several batches of germplasm had been sent to the
1069 region. It is obvious that the shuttle program managed
1070 to reach its objective and there are presently lines in
1071 the breeding trials that are competitive in yield while
1072 providing good leaf rust resistance.
1073 The future of CIMMYT breeding for high
1074 latitude regions of Kazakhstan and Siberia may lie
1075 in a gradual transfer of its key activities into the
1076 region. Mexico, by its geographic position, is not a
1077 place to breed for long days. Therefore, special
1078 efforts are needed to extend the day length, which is
1079 expensive and limits the scale of the breeding. Once
1080 the short-term priority of developing the leaf rust
1081 resistance is reached—the program may evolve into
1082 longer-term efforts of improving grain quality and
1083 resistance to Septoria and other pathogens, with
1084 most of its activities based in the region and aligned
1085 with one of the breeding programs. However,
1086 CIMMYT will play a clear role from Mexico, e.g.
1087 making crosses and providing valuable parental
1088 material.
1089Broadening the genetic base of,
1090and re-synthesizing wheat with available wild
1091and landrace genetic resources
1092Interspecific hybrization, embryo rescue, plant regen-
1093eration, cytological diagnostic, breeding methods,
1094stress screening, and the assessment of the stability of
1095the advanced derivatives due to homozygosity, are
1096the tools used by CIMMYT to utilize the wealth of
1097the wheat genetic endowment beyond the cultigen
1098pool (Mujeeb-Kazi and Rajaram 2002). ‘‘Capturing’’
1099wild grass diversity requires more time and effort for
1100a sequential production from F1s, amphiploids and
1101addition lines to translocation lines. Only transloca-
1102tion lines are useful for wheat breeding, but
1103intermediate products (amphiploids and addition
1104lines) are useful to evaluate the presence of useful
1105genes or traits. Also, certain amphiploids may be
1106propagated as new man-made crops, e.g. triticale
1107(9Triticosecale Witmm. ex. A. Camus—an amphi-
1108ploid between wheat and rye).
1109The germplasm sources can be classified into three
1110groups (primary, secondary, and tertiary genepools)
1111according to ease of exchanging genetic material with
1112wheat by meiotic recombinations (Jiang et al. 1994).
1113The primary gene pool is the species that have high
1114frequency rates of recombination with wheat, includ-
1115ing local landraces, wide forms of tetra and hexaploid
1116wheat, as well as diploid species of A and D
1117genomes. The secondary gene pool consists of
1118species that have less homology and reduced recom-
1119bination rates such as S (&B) genome species. The
1120tertiary gene pool consists of alien species that have
1121no recombination with wheat chromosomes under
1122normal conditions. They can be useful because they
1123sometimes possess high levels of host plant resistance
1124to biotic stresses, but the most used sources of genetic
1125enhancement for wheat come mainly from the
1126primary gene pool; for example species containing
1127the A and D genomes are sources of alleles that can
1128recombine directly with their respective genome
1129partners in the cultigen pool.
1130Mujeeb-Kazi et al. (1995) gave details of the early
1131years of research at CIMMYT for utilizing wild grass
1132diversity in wheat improvement through interspecific
1133hybridization. CIMMYT has produced F1 hybrids
1134with many genera in Triticeae including Aegilops,
1135Thinopyrum, Secale, Agropyron Gaertn., Elymus L.,
1136Leymus Hochst., Hordeum L. and Psathyrostachys
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of
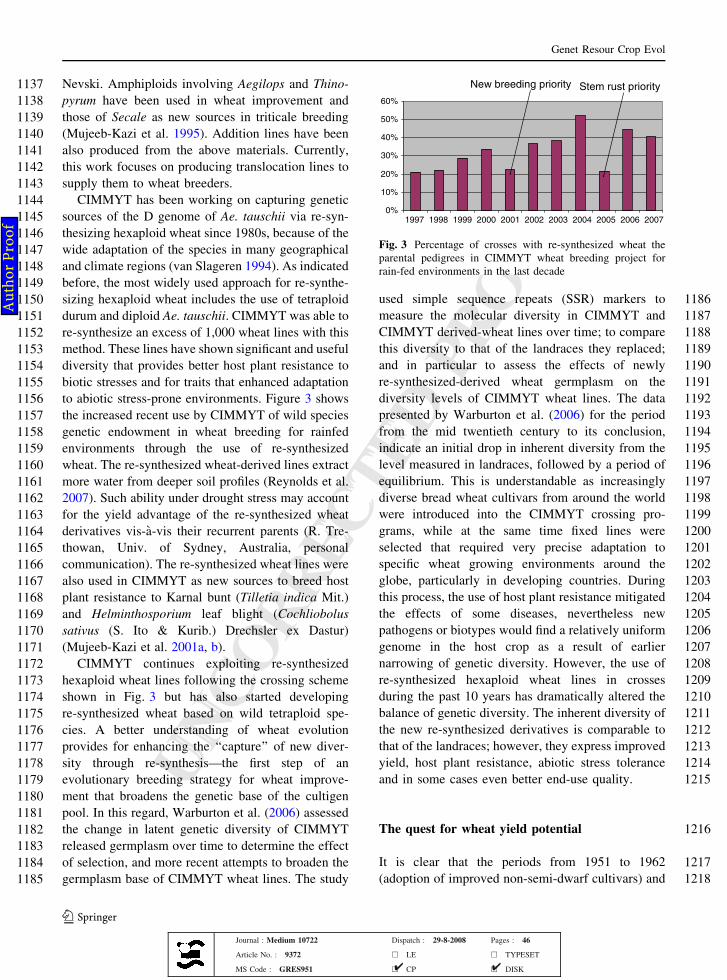
UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1137 Nevski. Amphiploids involving Aegilops and Thino-
1138 pyrum have been used in wheat improvement and
1139 those of Secale as new sources in triticale breeding
1140 (Mujeeb-Kazi et al. 1995). Addition lines have been
1141 also produced from the above materials. Currently,
1142 this work focuses on producing translocation lines to
1143 supply them to wheat breeders.
1144 CIMMYT has been working on capturing genetic
1145 sources of the D genome of Ae. tauschii via re-syn-
1146 thesizing hexaploid wheat since 1980s, because of the
1147 wide adaptation of the species in many geographical
1148 and climate regions (van Slageren 1994). As indicated
1149 before, the most widely used approach for re-synthe-
1150 sizing hexaploid wheat includes the use of tetraploid
1151 durum and diploid Ae. tauschii. CIMMYT was able to
1152 re-synthesize an excess of 1,000 wheat lines with this
1153 method. These lines have shown significant and useful
1154 diversity that provides better host plant resistance to
1155 biotic stresses and for traits that enhanced adaptation
1156 to abiotic stress-prone environments. Figure 3 shows
1157 the increased recent use by CIMMYT of wild species
1158 genetic endowment in wheat breeding for rainfed
1159 environments through the use of re-synthesized
1160 wheat. The re-synthesized wheat-derived lines extract
1161 more water from deeper soil profiles (Reynolds et al.
1162 2007). Such ability under drought stress may account
1163 for the yield advantage of the re-synthesized wheat
1164 derivatives vis-a-vis their recurrent parents (R. Tre-
1165 thowan, Univ. of Sydney, Australia, personal
1166 communication). The re-synthesized wheat lines were
1167 also used in CIMMYT as new sources to breed host
1168 plant resistance to Karnal bunt (Tilletia indica Mit.)
1169 and Helminthosporium leaf blight (Cochliobolus
1170 sativus (S. Ito & Kurib.) Drechsler ex Dastur)
1171 (Mujeeb-Kazi et al. 2001a, b).
1172 CIMMYT continues exploiting re-synthesized
1173 hexaploid wheat lines following the crossing scheme
1174 shown in Fig. 3 but has also started developing
1175 re-synthesized wheat based on wild tetraploid spe-
1176 cies. A better understanding of wheat evolution
1177 provides for enhancing the ‘‘capture’’ of new diver-
1178 sity through re-synthesis—the first step of an
1179 evolutionary breeding strategy for wheat improve-
1180 ment that broadens the genetic base of the cultigen
1181 pool. In this regard, Warburton et al. (2006) assessed
1182 the change in latent genetic diversity of CIMMYT
1183 released germplasm over time to determine the effect
1184 of selection, and more recent attempts to broaden the
1185 germplasm base of CIMMYT wheat lines. The study
1186used simple sequence repeats (SSR) markers to
1187measure the molecular diversity in CIMMYT and
1188CIMMYT derived-wheat lines over time; to compare
1189this diversity to that of the landraces they replaced;
1190and in particular to assess the effects of newly
1191re-synthesized-derived wheat germplasm on the
1192diversity levels of CIMMYT wheat lines. The data
1193presented by Warburton et al. (2006) for the period
1194from the mid twentieth century to its conclusion,
1195indicate an initial drop in inherent diversity from the
1196level measured in landraces, followed by a period of
1197equilibrium. This is understandable as increasingly
1198diverse bread wheat cultivars from around the world
1199were introduced into the CIMMYT crossing pro-
1200grams, while at the same time fixed lines were
1201selected that required very precise adaptation to
1202specific wheat growing environments around the
1203globe, particularly in developing countries. During
1204this process, the use of host plant resistance mitigated
1205the effects of some diseases, nevertheless new
1206pathogens or biotypes would find a relatively uniform
1207genome in the host crop as a result of earlier
1208narrowing of genetic diversity. However, the use of
1209re-synthesized hexaploid wheat lines in crosses
1210during the past 10 years has dramatically altered the
1211balance of genetic diversity. The inherent diversity of
1212the new re-synthesized derivatives is comparable to
1213that of the landraces; however, they express improved
1214yield, host plant resistance, abiotic stress tolerance
1215and in some cases even better end-use quality.
1216The quest for wheat yield potential
1217It is clear that the periods from 1951 to 1962
1218(adoption of improved non-semi-dwarf cultivars) and
0%
10%
20%
30%
40%
50%
60%
1997
New breeding priority Stem rust priority
2007200620052004200320022001200019991998
Fig. 3 Percentage of crosses with re-synthesized wheat theparental pedigrees in CIMMYT wheat breeding project forrain-fed environments in the last decade
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1219 from 1963 to 1975 (the celebrated period of the
1220 introduction and adoption of reliable semi-dwarf
1221 cultivars) were both eras of remarkable increase in
1222 wheat yields in farmer fields (Table 6). Just as the
1223 impact of the Green Revolution can be attributed
1224 mostly to improved partitioning of the products of
1225 photosynthesis to grain yield, progress in yield of
1226 irrigated wheat since the development of semi-dwarf
1227 lines, as pointed out by Sayre et al. (1997), is also
1228 most strongly associated with improved harvest index
1229 (HI). However, from 1976 to 2005 it was noted a
1230 comparably large decrease in the rate of yield
1231 increase in farmer fields to about 1% year-1, a rather
1232 disturbing observation. There are several reasons,
1233 which are related to both breeding as well as crop
1234 management issues. Previous studies in the Yaqui
1235 Valley have concluded that about 50% of farmer
1236 wheat yield increases have been associated with
1237 genetic improvements in yield potential and 50% has
1238 been associated with improved crop-management
1239 practices, e.g. higher fertilizer rates, better irrigation
1240 management, and the use of bed planting systems
1241 (Bell et al. 1995; Sayre et al. 1997).
1242 The rate of increase in bread and durum wheat
1243 genetic yield potential was high for cultivars bred
1244 from the mid-1960s to the mid-1980s but has subse-
1245 quently declined over the past 10 years. However,
1246 under conditions where pests (rusts in particular) are
1247 not controlled, yield potential for both bread and
1248 durum wheat—bred until the 1990s, showed increases
1249 reflecting the investment made in breeding for
1250 potential durable host plant resistance to rusts. Unlike
1251 the Green Revolution cultivars that relied on single
1252 gene rust resistance—which can easily be eroded as
1253 new pathogen races evolve, more modern cultivars
1254 have multi-gene resistance that can be expected to
1255 stand the test of time (Sayre et al. 1998).
1256 Wheat improvement at CIMMYT expanded the
1257 genetic base of modern wheat lines using conven-
1258 tional breeding as well as cytogenetic-led approaches
1259 for introgressing alleles from wild relatives. For
1260 example, the highly successful ‘Veery’ lines bred in
1261 the early 1980s ensued from the cross of a winter
1262 wheat parent containing the 1B/R rye translocation.
1263 Approximately 3,170 different crosses were made
1264 among 51 individual parents originating in 26
1265 countries around the world to develop the Veery
1266 lines, of which 62 of them were grown annually on
1267 about 3 million ha from Chile to China. These widely
1268adapted Veery lines had an outstanding yield
1269potential and other interesting physiological traits,
1270e.g. the cultivar ‘Seri 82’showed superior leaf
1271photosynthetic rate, stomatal conductance, and leaf
1272greenness relative to a set of hallmark varieties
1273developed both before and after its release (Fischer
1274et al. 1998). The success of lines possessing the
1275chromosome substitution T1BL.1RS may also be
1276related to increased tolerance to stresses (Villarreal
1277et al. 1997). Last but not least, the use of
1278winter 9 spring crosses for wheat improvement at
1279CIMMYT, which began in the early 1970s, contrib-
1280uted to increasing the genetic diversity of the spring
1281wheat gene pool.
1282CIMMYT ideotype research using modem semi-
1283dwarf spring wheat cultivars, representing a range of
1284yield potential, supports the idea that genes confer-
1285ring yield, through improved adaptation to the crop
1286environment, are associated with a less competitive
1287phenotype (Reynolds et al. 1994a). One important
1288implication of these results is that improvement in
1289yield potential would appear to be more a function of
1290improved adaptation to canopy microenvironment,
1291rather than macro-environmental factors such as
1292climate. Reynolds et al. (1999) reviewed the physi-
1293ological and genetic changes in irrigated wheat in the
1294post-Green Revolution period. Their assessment
1295shows that morphological traits associated with
1296increased yield potential in CIMMYT-derived wheat
1297cultivars released between 1962 and 1988 include
1298grain number and HI (Reynolds et al. 1999).
1299Although increased HI seems to account for improve-
1300ments in genetic yield potential, even in the aftermath
1301of the Green Revolution, it should be taken into
1302account with caution due to theoretical limit to HI,
1303estimated at 60% (Austin et al. 1980), e.g. if HI could
1304be raised to 60% from its current maximum value
1305(approximately 50%), it implies that grain yields
1306could only be increased by a further 20% using HI as
1307a selection criterion, unless total crop biomass is also
1308raised.
1309As indicated previously, significant increase in
1310yield and biomass has been observed in several
1311backgrounds when alien chromatin associated with
1312Lr19 was introgressed from Th. elongatum (Reynolds
1313et al. 2001). Lr19—transferred to wheat through
1314the 7DL.7Ag translocation—was associated with
1315increases in yield (average 13%), final biomass
1316(10%), and grain number (15%) in all backgrounds
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1317 studied. Lrl9 was associated with an increased
1318 partitioning of biomass to spike growth at anthesis
1319 (13%), a higher grain number per spike, higher
1320 radiation-use efficiency and flag-leaf photosynthetic
1321 rate during grain filling (Reynolds et al. 2005).
1322 The hypothesis that photosynthesis and RUE may
1323 respond directly to a larger number of grains per
1324 spike was tested experimentally by imposing a light
1325 treatment during boot stage. The treatment was
1326 associated with a small increase (5%) in the propor-
1327 tion of biomass invested in spike mass at anthesis,
1328 reflected by on average three extra grains per spike at
1329 maturity. The treatment was associated with 25%
1330 more yield and 22% more biomass than checks, while
1331 carbon assimilation rate measured on flag-leaves
1332 during grain-filling was 10% higher than checks. The
1333 results suggest that RUE can be increased indirectly
1334 by increasing sink strength and a major yield-limiting
1335 factor in modern high-yielding spring wheat is still
1336 the determination of kernel number (Reynolds et al.
1337 2005). This research illustrates the advantage of
1338 introgressing wild relatives’ genes for improving
1339 grain yield potential in wheat.
1340 The power of in silico breeding to define wheat
1341 genetic enhancement approaches
1342 The strategies used by CIMMYT breeders have
1343 evolved. Pedigree selection was used primarily from
1344 1944 until 1985. From 1985 until the second half of
1345 the 1990s the main selection method was a modified
1346 pedigree/bulk method (MODPED) (van Ginkel et al.
1347 2002), which successfully produced many of the
1348 widely adapted wheat cultivars now being grown in
1349 the developing world. This method was replaced in
1350 the late 1990s by the selected bulk method (SEL-
1351 BLK) (van Ginkel et al. 2002) in an attempt to
1352 improve resource-use efficiency. The major differ-
1353 ences between MODPED and SELBLK are outlined
1354 below.
1355 The MODPED method begins with pedigree
1356 selection of individual plants in the F2 generation
1357 followed by three bulk selections from F3 to F5, and
1358 pedigree selection in the F6; hence the name modified
1359 pedigree/bulk. In the SELBLK method, spikes of
1360 selected F2 plants within one cross are harvested in
1361 bulk and threshed together, resulting in one F3 seed
1362 lot per cross. This selected bulk selection is also used
1363from F3 to F5, while pedigree selection is used only in
1364the F6. A major advantage of SELBLK compared
1365with MODPED is that fewer seed lots need to be
1366harvested, threshed, and visually selected for seed
1367appearance. In addition, significant savings in time,
1368labor, and costs associated with nursery preparation,
1369planting, and plot labeling ensue, and potential
1370sources of error are avoided (van Ginkel et al. 2002).
1371Although some small-scale field experiments have
1372been conducted comparing the efficiencies of CI-
1373MMYT wheat breeding methods (Singh et al. 1998b),
1374the efficiency of SELBLK, compared with that of
1375MODPED, remained untested on a larger scale until
1376the use of the breeding simulation tool of QuLine.
1377The breeding and simulation tool of quLine
1378QU-GENE is a simulation platform for quantitative
1379analysis of genetic models, which consists of a two-
1380stage architecture (Podlich and Cooper 1998). The
1381first stage is the engine (referred to as QUGENE), and
1382its role is to define the genotype 9 environment (GE)
1383system (i.e., all the genetic and environmental
1384information of the simulation experiment), and
1385generate the starting population of individuals (base
1386germplasm). The second stage encompasses the
1387application modules, whose role is to investigate,
1388analyze, or manipulate the starting population of
1389individuals within the GE system defined by the
1390engine. The application module will usually represent
1391the operation of a breeding program. A QU-GENE
1392strategic application module, QuLine, has therefore
1393been developed to simulate the breeding procedure
1394for deriving inbred lines. Built on QU-GENE,
1395QuLine (previously called QuCim) is a genetics and
1396breeding simulation tool that can integrate various
1397genes with multiple alleles operating within epistatic
1398networks and differentially interacting with the
1399environment interaction, and predict the outcomes
1400from a specific cross following the application of a
1401real selection scheme (Wang et al. 2003; Wang et al.
14022004). It therefore has the potential to provide a
1403bridge between the vast amount of biological data
1404and breeders’ queries on optimizing selection gain
1405and efficiency. QuLine has been used to compare two
1406selection strategies (Wang et al. 2003), to study the
1407effects on selection of dominance and epistasis
1408(Wang et al. 2004), to predict cross performance
1409using known gene information (Wang et al. 2005),
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1410 and to optimize marker-assisted selection to efficient
1411 pyramid multiple genes (Kuchel et al. 2005; Wang
1412 et al. 2007a, b; Ye et al. 2007).
1413 Comparison of breeding efficiencies for different
1414 selection strategies through simulation
1415 The genetic models developed have accounted for
1416 epistasis, pleiotropy, and genotype-by-environment
1417 interaction. The simulation experiment comprised the
1418 same 1,000 crosses, developed from 200 parents, for
1419 both breeding strategies. A total of 258 advanced
1420 lines remained following 10 generations of selection.
1421 The two strategies were each applied 500 times on 12
1422 GE systems.
1423 Genetic gain in yield from MODPED and SELBLK
1424 The average adjusted gains were 6.7 t ha-1 for no
1425 epistasis, 5.4 t ha-1 for di-genic epistasis, and
1426 5.7 t ha-1 for tri-genic epistasis, which indicates that
1427 epistasis will reduce the adjusted gain. The adjusted
1428 gain associated with the absence of pleiotropy is also
1429 higher than that for the presence of pleiotropy. These
1430 results suggest that the increase in gene number and
1431 the presence of epistasis and pleiotropy make it more
1432 difficult for a breeding strategy to identify the trait
1433 performance level of the best genotype in the defined
1434 GE system. When the experimental factors are
1435 considered individually, the adjusted gain from
1436 SELBLK is always significantly higher than that
1437 from MODPED, except in the absence of pleiotropy,
1438 indicating SELBLK is at least equivalent to or better
1439 than MODPED.
1440 The average adjusted genetic gain on yield is
1441 5.8 t ha-1 for MODPED and 6 t ha-1 for SELBLK, a
1442 difference of 3.3% (Fig. 4a). This difference is not
1443 large and therefore unlikely to be detected using field
1444 experiments (Singh et al. 1998b). However, it can be
1445 detected through simulation, which indicates that the
1446 high level of replication (50 models by 10 runs in this
1447 experiment) possible using simulations can better
1448 account for the stochastic properties of a run of a
1449 breeding strategy and for the sources of experimental
1450 errors. The average adjusted gains for the two yield
1451 gene numbers 20 and 40 are 6.83 and 5.02, respec-
1452 tively, suggesting that genetic gain decreases with
1453 increasing yield gene number.
1454Number of crosses remaining after selection
1455The same 1,000 crosses were made for both breeding
1456strategies and 258 advanced lines were selected after
1457a breeding cycle, regardless of the GE system used.
1458The number of crosses remaining after one breeding
1459cycle is significantly different among models and
1460strategies, but not among runs. The number of crosses
1461remaining from SELBLK is always higher than that
1462from MODPED, which means that delaying pedigree
1463selection favors diversity. On average, 30 more
1464crosses were maintained in SELBLK (Fig. 4b).
1465However, there is a crossover between the two
1466breeding strategies (Fig. 4b). Prior to F5 the number
1467of crosses in MODPED is higher than that in
1468SELBLK. The number of crosses becomes smaller
1469in MODPED after F5 when pedigree selection is
1470applied in F6. Among-family selection from F1 to F51471in SELBLK is equal to among-cross selection, and
1472results in a greater reduction in cross number for
1473SELBLK compared to MODPED in the early gener-
1474ations. In general, only a small proportion of crosses
1475remain at the end of a breeding cycle (11.8% for
1476MODPED and 14.8% for SELBLK); therefore,
1477intense among-cross selection in early generations
1478is unlikely to reduce the genetic gain. On the
1479contrary, breeders will tend to concentrate on fewer
1480but ‘‘higher-probability’’ crosses. That just a few
1481crosses of the many generated remain after the final
1482yield trial stage is common in most breeding
1483programs. Since more crosses remain in SELBLK,
1484the population following selection from SELBLK
1485may have larger genetic diversity than that from
1486MODPED. In this context, SELBLK is also superior
1487to MODPED.
1488Resource allocation
1489Since the number of families and selection methods
1490after F8 are basically the same for both MODPED and
1491SELBLK, only the resources allocated from F1 to F81492are compared. The total number of individual plants
1493from F1 to F8 was calculated to be 5,155,090 for
1494MODPED and 3,358,255 for SELBLK (Fig. 4c).
1495Assuming that planting intensity is similar, SELBLK
1496will use approximately two thirds of the land
1497allocated to MODPED. Furthermore, SELBLK pro-
1498duces a smaller number of families compared to
1499MODPED (Fig. 4d). From F1 to F8, there are 63,188
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of
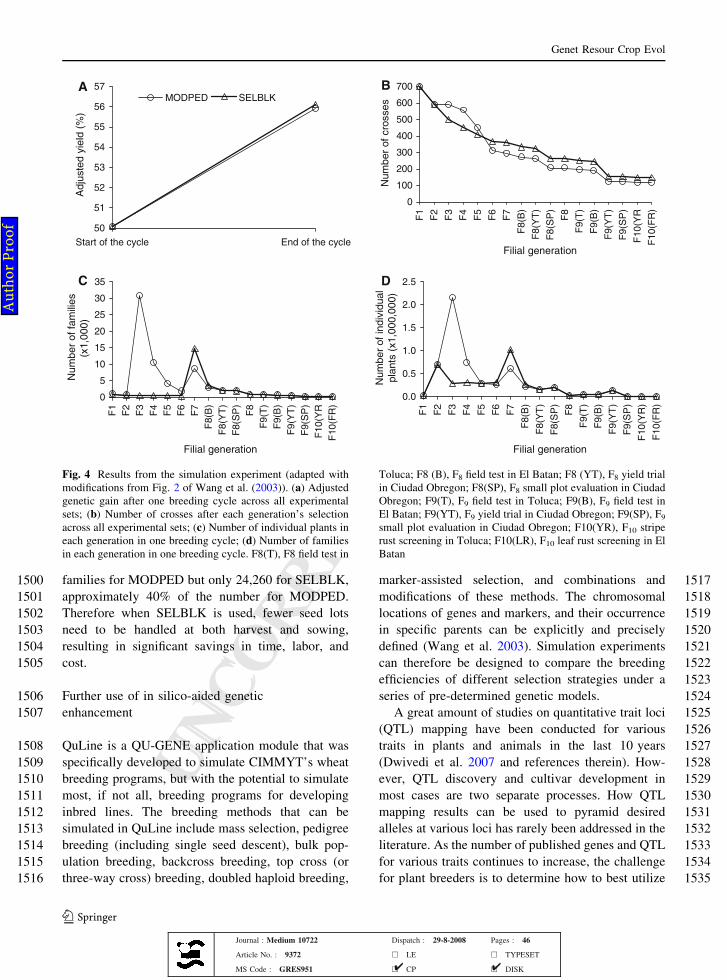
UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1500 families for MODPED but only 24,260 for SELBLK,
1501 approximately 40% of the number for MODPED.
1502 Therefore when SELBLK is used, fewer seed lots
1503 need to be handled at both harvest and sowing,
1504 resulting in significant savings in time, labor, and
1505 cost.
1506 Further use of in silico-aided genetic
1507 enhancement
1508 QuLine is a QU-GENE application module that was
1509 specifically developed to simulate CIMMYT’s wheat
1510 breeding programs, but with the potential to simulate
1511 most, if not all, breeding programs for developing
1512 inbred lines. The breeding methods that can be
1513 simulated in QuLine include mass selection, pedigree
1514 breeding (including single seed descent), bulk pop-
1515 ulation breeding, backcross breeding, top cross (or
1516 three-way cross) breeding, doubled haploid breeding,
1517marker-assisted selection, and combinations and
1518modifications of these methods. The chromosomal
1519locations of genes and markers, and their occurrence
1520in specific parents can be explicitly and precisely
1521defined (Wang et al. 2003). Simulation experiments
1522can therefore be designed to compare the breeding
1523efficiencies of different selection strategies under a
1524series of pre-determined genetic models.
1525A great amount of studies on quantitative trait loci
1526(QTL) mapping have been conducted for various
1527traits in plants and animals in the last 10 years
1528(Dwivedi et al. 2007 and references therein). How-
1529ever, QTL discovery and cultivar development in
1530most cases are two separate processes. How QTL
1531mapping results can be used to pyramid desired
1532alleles at various loci has rarely been addressed in the
1533literature. As the number of published genes and QTL
1534for various traits continues to increase, the challenge
1535for plant breeders is to determine how to best utilize
A
50
51
52
53
54
55
56
57
elcyc eht fo dnEelcyc eht fo tratS
Ad
juste
d y
ield
(%
)
MODPED SELBLKB
0
100
200
300
400
500
600
700
F1
F2
F3
F4
F5
F6
F7
F8(B
)
F8(Y
T)
F8(S
P)
F8
F9(T
)
F9(B
)
F9(Y
T)
F9(S
P)
F10(Y
R
F10(F
R)
Filial generation
Num
ber
of
cro
sses
C
0
5
10
15
20
25
30
35
F1
F2
F3
F4
F5
F6
F7
F8(B
)
F8(Y
T)
F8(S
P)
F8
F9(T
)
F9(B
)
F9(Y
T)
F9(S
P)
F10(Y
R
F10(F
R)
Filial generation
Nu
mb
er
of
fam
ilie
s(x
1,0
00)
D
0.0
0.5
1.0
1.5
2.0
2.5
F1
F2
F3
F4
F5
F6
F7
F8(B
)
F8(Y
T)
F8(S
P)
F8
F9(T
)
F9(B
)
F9(Y
T)
F9(S
P)
F10(Y
R)
F10(F
R)
Filial generation
Num
ber
of
indiv
idual
pla
nts
(x1
,00
0,0
00
)
Fig. 4 Results from the simulation experiment (adapted withmodifications from Fig. 2 of Wang et al. (2003)). (a) Adjustedgenetic gain after one breeding cycle across all experimentalsets; (b) Number of crosses after each generation’s selectionacross all experimental sets; (c) Number of individual plants ineach generation in one breeding cycle; (d) Number of familiesin each generation in one breeding cycle. F8(T), F8 field test in
Toluca; F8 (B), F8 field test in El Batan; F8 (YT), F8 yield trialin Ciudad Obregon; F8(SP), F8 small plot evaluation in CiudadObregon; F9(T), F9 field test in Toluca; F9(B), F9 field test inEl Batan; F9(YT), F9 yield trial in Ciudad Obregon; F9(SP), F9small plot evaluation in Ciudad Obregon; F10(YR), F10 striperust screening in Toluca; F10(LR), F10 leaf rust screening in ElBatan
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1536 this multitude of information in the improvement of
1537 crop performance. QuLine provide an appropriate
1538 tool that can combine different types and levels of
1539 biological data such that the complex and voluminous
1540 data is turned into knowledge that can be applied in
1541 breeding.
1542 Information management and knowledge sharing:
1543 the wheat phenome atlas
1544 A useful new tool for crop genetic improvement is
1545 association analysis (AA), which is the direct selec-
1546 tion of polymorphic markers associated with
1547 phenotypic variation for important traits and identi-
1548 fied by the linkage disequilibrium (LD) between loci
1549 (Thornsberry et al. 2001; Flint-Garcia et al. 2003).
1550 This LD is determined by their physical distance
1551 across chromosomes and has proven to be useful for
1552 dissecting complex traits because it offers a fine scale
1553 mapping due to historical recombination (Lynch and
1554 Walsh 1998). However, covariance between markers
1555 and traits not due to physical distance can arise due to
1556 population structure caused by admixture, mating
1557 system, and genetic drift, or by artificial or natural
1558 selection during evolution, domestication, or plant
1559 improvement. These factors create subpopulation
1560 structure in a linkage disequilibrium study, which
1561 leads to LD between loci that are not physically
1562 linked and cause a high rate of false positives when
1563 relating polymorphic marker to phenotypic trait
1564 variation. However, LD due to physical linkage can
1565 be differentiated from LD due to population structure
1566 using statistical methods such as those suggested by
1567 Pritchard et al. (2000) and Yu et al. (2006). In
1568 addition to controlling population structure, success-
1569 ful application of AA requires comprehensive
1570 phenotypic data, including replicated field trials for
1571 modeling GE interaction. An advantage of AA is that
1572 large amounts of historical phenotypic data can be
1573 used, as demonstrated by Crossa et al. (2007). This
1574 can decrease the need for additional, expensive and
1575 time consuming phenotyping.
1576 CIMMYT, Cornell Univ (Ithaca, NY, USA), and
1577 the Univ. of Queensland (Brisbane, Australia)
1578 recently launched the Wheat Phenome Atlas initiative
1579 which aims for enabling technologies to link geno-
1580 type to phenotype across a wide range of agronomic
1581 traits of high priority to wheat farmers across the
1582world. The Wheat Phenome Atlas will facilitate more
1583rapid development of molecular breeding tools,
1584increased understanding of genotype-by-environment
1585interaction, and will lead to increased precision and
1586scope of targeted breeding impacts. The Phenome
1587Atlas Toolbox will be developed as an open source
1588resource for wheat genetics, breeding, pathology,
1589physiology, and biological research. The initiative is
1590based on a unique database (amongst crop plants)
1591developed by CIMMYT and national partners over
1592the past half century, based on the field evaluation of
1593more than 15,000 elite breeding lines in more than
1594100 locations across the world—at a cost of about
1595US$ 0.5 billion. All seed from these breeding lines
1596has been preserved in the CIMMYT germplasm bank
1597and can now be subjected to whole genome
1598genotyping.
1599The proof-of-concept in this area was recently
1600carried out by researchers from the CIMMYT Crop
1601Research Informatics Laboratory (an initiative of the
1602CIMMYT-IRRI Alliance), the University of Queens-
1603land, and Cornell University with the biotech
1604company Triticarte, who have developed the high
1605throughput genotyping protocol—diversity array
1606technology or DArT for short (Crossa et al. 2007).
1607The Wheat Phenome Atlas initiative is based around
1608the ICIS (International Crop Information System), an
1609open-source informatics platform used by CGIAR,
1610NARS, and private sector germplasm banks,
1611researchers, and breeders across the world. All data
1612and resources developed through this initiative will
1613be available to wheat researchers and breeders across
1614the world through public access databases and
1615portals. It is envisaged that additional international
1616collaborators will join the initiative, including
1617advanced research institutes from the USA, Europe,
1618and Australasia, strong NARS from Latin America
1619and Asia, plus private sector breeding companies
1620from across the world.
1621Establishing the analytical tools to deal with data
1622sets of over 15,000 lines 9 40 years 9 80
1623traits 9 100 locations 9 2,000 DNA data points
1624(approximately 10 billion data points) will involve
1625the development of new and powerful bioinformatics
1626tools and webpage visualization software. The
1627Phenome Atlas Toolbox will facilitate the identifica-
1628tion of gene blocks having beneficial effects on high
1629priority agronomic traits such as rust resistance,
1630drought tolerance, and yield. From these outputs it
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1631 will be possible to develop molecular tools for
1632 rapidly introgressing these elite gene blocks into
1633 new cultivars. The Wheat Phenome Atlas database
1634 will also be used for modeling and simulation studies
1635 to predict cultivar performance in a range of global
1636 environments, including future environments pre-
1637 dicted by climate change models (CIMMYT 2007). It
1638 is envisaged that the Phenome Atlas Toolbox will
1639 develop dynamically with input from wheat research-
1640 ers worldwide. The Phenome Atlas Toolbox will be
1641 generic and applicable to all biological systems,
1642 whether plant or animal.
1643 Safe and legal movement of wheat germplasm
1644 worldwide
1645 CIMMYT has a global mandate for the improvement
1646 of wheat and maize and it has also the responsibility
1647 of conserving the germplasm of these crops.
1648 CIMMYT’s germplasm improvement programs rely
1649 heavily on the free international exchange of maize
1650 and wheat seed. All concerned institutions, cooper-
1651 ators, and regulating authorities must have confidence
1652 in the safety of both imported and exported seed to
1653 facilitate such exchange. CIMMYT is fully commit-
1654 ted to maintaining fundamental health standards in its
1655 worldwide operations. These standards are dictated at
1656 different levels by the International Plant Protection
1657 Convention (FAO 1997 https://www.ippc.int/IPP/En/
1658 default.jsp), by IT-PGRFA (FAO 2002), and by the
1659 CGIAR (1999).
1660 CIMMYTpolicies and practices and its SeedHealth
1661 Laboratory (SHL) control and ensure the implemen-
1662 tation of international and national phytosanitary
1663 regulations for material that is distributed every year.
1664 CIMMYT SHL has operated since 1998 under the
1665 approval of the Mexican Ministry of Agriculture
1666 (SAGARPA) (Norma Oficial Mexicana 036-FITO-
1667 1995), and since April 2007 with the accreditation
1668 under norm ISO/IEC NMX-EC-17025-IMNC-2005
1669 General Requirements for the Competence of Testing
1670 and Calibration Laboratories. These essential legal
1671 recognitions guarantee to the Mexican phytosanitary
1672 authorities that CIMMYT seed exchange activities do
1673 not jeopardize the internal Mexican or international
1674 phytosanitary situation and that the quarantine proce-
1675 dure carried out on seed introductions strictly adheres
1676 to Mexican phytosanitary regulations. Likewise, they
1677reassure our collaborators that CIMMYT procedures
1678are standardized, internationally recognized, and con-
1679trolled constantly through internal and external audit
1680processes.
1681The principal functions of CIMMYT SHL are to:
1682certify the viability and health of maize and small
1683grain cereal seed for international shipments, con-
1684trol the safety of seed arriving to CIMMYT and
1685apply Mexican quarantine regulations, detect the
1686unintentional presence of transgenes on maize
1687introductions from risky countries, maintain the
1688relationship with, and act as spokesman with
1689Mexican phytosanitary authorities, supervise the
1690seed-treating procedures, inspect field multiplication
1691and introduction plots, and ensure that chemical
1692prophylaxes against quarantine disease are applied
1693in the multiplication plots, in storehouses, and in
1694seed preparation areas. The SHL represents the
1695point of entry and exit of all seed shipments to and
1696from CIMMYT headquarters.
1697Seed samples are delivered by programs for
1698analyses or are received from collaborators; upon
1699completion of the testing process the seed is
1700‘‘released’’ and ready to be shipped abroad or planted
1701in CIMMYT experimental stations. According to
1702Mexican phytosanitary regulations, when the inter-
1703ception of a pathogen of quarantine importance
1704occurs both on outgoing or incoming material, the
1705SHL must inform Mexican phytosanitary authorities,
1706who will indicate the measures to follow to prevent
1707the spread of the pathogen.
1708During 2007 the SHL carried out quarantine
1709procedures on 62 seed introductions of small grain
1710cereals (wheat, triticale, and barley) proceeding from
171120 countries and analyzed 822 samples. From this
1712number of samples, which included 41,257 seeds, two
1713interceptions of the quarantined pathogen Tilletia
1714indica were made on two seed introductions that were
1715incinerated.
1716Karnal bunt is considered a disease of moderate
1717economic importance (CABI 2005), for example in
1718Mexico, where Karnal bunt appears regularly, direct
1719losses are not very significant and do not exceed 1%.
1720However, the indirect costs to the Mexican economy
1721are more significant due to quarantine measures,
1722which have to be applied for grain exports (Brennan
1723et al. 1992). The same happens in the USA (Rush
1724et al. 2005). In Europe and Australia the disease is
1725still considered a high risk (Bartlett 2000; Murray and
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1726 Brennan 1998), in spite of the strict environmental
1727 requirement that the pathogen has for developing and
1728 affecting wheat and triticale (Jones 2007). The
1729 disease is given political recognition in many coun-
1730 tries. In terms of its phytosanitary importance, the
1731 pathogen is quarantined in more then 40 countries in
1732 the world (Rush et al. 2005). Hence, CIMMYT is
1733 obligated to devote a considerable amount of
1734 resources in applying a zero-tolerance policy against
1735 this disease. Besides T. indica, particular care is also
1736 taken by CIMMYT to prevent infections by Xantho-
1737 manas translucens (Jones et al.) Vauterin et al. pv.
1738 undulosa, the Barley Stripe Mosaic virus, and the
1739 Wheat Streak Mosaic virus through inspection of the
1740 seed before and after planting.
1741 In the case of outgoing germplasm, 2,468 samples
1742 were tested at the SHL before the shipments were
1743 sent in 2007. The phytosanitary quality of outgoing
1744 seed is particularly high due to the procedure
1745 followed in the multiplication plots that are planted
1746 in an internationally recognized Karnal bunt free area
1747 in northwest Mexico and that are submitted to a
1748 prophylaxis against Karnal bunt and other foliar
1749 diseases, which includes 3–4 fungicide treatments
1750 with propiconazole and tebuconazole during the
1751 cropping cycle and insecticide to protect the crop
1752 from aphid- and mite-borne viruses. Therefore, the
1753 range of pathogens detected on the seed mainly
1754 consists of saprophytes or minor seed-borne patho-
1755 gens that do not affect seed quality, such as fungi
1756belonging to the genus Alternaria, Cephalosporium,
1757Fusarium, Bipolaris, Curvularia, and Cladosporium.
1758Starting 1 January 2007, the CGIAR Centers had
1759to use the Standard Material Transfer Agreement
1760(SMTA) for all shipments, which was adopted in
1761the first session of the Governing Body of the
1762IT-PGFRA. It was agreed upon through the Statement
1763of the CGIAR Centers regarding Implementation of
1764the Agreements between the Centers and the govern-
1765ing body of the International Treaty on Plant Genetic
1766Resources for Food and Agriculture. CIMMYT
1767started using the SMTA as of 14 January 2007, but
1768did not distribute any material prior to that date.
1769During 2007, 565 shipments were sent under the
1770SMTA and three were rejected.
1771Table 7 provides a summary of the number of
1772international nursery seed sets sent during 2007 from
1773CIMMYT headquarters. This table includes bread,
1774durum wheat, triticale, and barley. The seed is
1775distributed mainly to governmental research institu-
1776tions and national programs in Asia and Central and
1777West Asia and Northern Africa (CWANA) through
1778the CIMMYT International Wheat and Improvement
1779Network that every year makes available 14 nurseries
1780adapted to different environments. Table 8 provides a
1781list of nurseries available and whose seeds can be
1782requested by any party that agrees with the SMTA
1783which regulates the use and benefit sharing of the
1784germplasm according to the IT-PGRFA. This activity
1785constitutes the backbone of the breeders’ research
Table 7 Number of international nurseries sets of small grain cereals (wheat, barley and triticale) sent by CIMMYT during 2007to different regions
2007
Region Barley Bread wheat Durum wheat Special nurseries Triticale Total by region
Africa 24 85 10 1 12 132
Asia 47 205 41 16 309
Caribbean 2 2
CWANAa 133 325 123 52 633
Europe 19 110 46 30 205
North America 21 113 27 1 19 181
Oceania 1 2 1 4
South America 30 119 27 3 28 207
Total by crop 277 957 276 5 158 1,673
a CWANA: Central/West Asia and North Africa, which include Algeria, Egypt, Ethiopia, Eritrea, Libya, Mauritania, Morocco,Sudan and Tunisia (North Africa and Nile Valley), Bahrain Iraq, Jordan, Kuwait, Lebanon, Palestine, Qatar, Saudi Arabia, theSultanate of Oman, Syria, Turkey, the United Arab Emirates and Yemen (Middle East), Afghanistan, Armenia, Azerbaijan, Georgia,Iran, Kazakhstan, Kyrgyzstan, Tajikistan, Turkmenistan and Uzbekistan (Central Asia and Caucasus)
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of
UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1786 programs due to the valuable information sent back by
1787 the collaborators that feeds into the breeding program.
1788 Such feedback allows for a constant and prompt
1789 improvement of wheat germplasm according to the
1790 users’ needs and to the biotic and abiotic stresses that
1791 threaten wheat production in developing countries.
1792 CIMMYT also distributes a great number of lines
1793 that are developed under specific projects between
1794 CIMMYT and several collaborators in the public
1795 and private sector. These are called miscellaneous
1796 shipments and as it is shown in Table 9 the greatest
1797 part is sent to Africa, Asia, and North America.
1798 Miscellaneous shipments include also germplasm
1799requested by genebanks to absolve specific scientific
1800needs and also for conservation of duplicates in other
1801genebanks.
1802International CIMMYT wheat nursery systems
1803CIMMYT-improved germplasm is dispatched through
1804nurseries targeted to specific spring and winter type,
1805irrigated, high-rainfall, semi-arid, and heat-tolerant
1806requiring agro-ecological environments to a network of
1807wheat researchers worldwide (http://www.cimmyt.org/
1808english/wps/obtain_seed/frmrequesttrialswheat.htm).
Table 8 Name of thenurseries distributed fromMexico by CIMMYT everyyear to the InternationalWheat ImprovementNetwork
a ICARDA-CIMMYTBarley program, distributedonly until 2007
Name of the nursery Crop
Elite Spring Wheat Yield Trial Spring bread wheat
Semi-Arid Wheat Yield Trial Spring bread wheat
High-Rainfall Wheat Yield Trial Spring bread wheat
International Bread Wheat Screening Nursery Spring bread wheat
Semi-Arid Wheat Screening Nursery Spring bread wheat
High-Rainfall Wheat Screening Nursery Spring bread wheat
High-Latitude Wheat Screening Nursery Spring bread wheat
(Fusarium) Scab Resistance Screening Nursery Spring bread wheat
Stem Rust Resistance Screening Nursery Bread wheat
International Durum Yield Nursery Durum wheat
International Durum Screening Nursery Durum wheat
International Triticale Yield Nursery Triticale
International Triticale Screening Nursery Triticale
International Barley Yield Trial z Barley
International Barley Observation Nurserya Barley
Early Maturity Barley Screening Nurserya Barley
Hull-less Barley Screening Nurserya Barley
Table 9 Number of small grain cereals lines distributed as miscellaneous shipment during 2007 to different regions in the world
Region Bread wheat Durum wheat Triticale Barley Genebank Total by region
Africa 12,138 4,953 430 415 132 18,068
Asia 8,548 503 198 304 717 10,270
Caribbean 6 3 1 10
Europe 672 984 47 21 716 2,440
Middle Easta 1,485 52 27 61 1,238 2,863
North America 1,766 13 12 1,460 17,735 20,986
Oceania 3,267 546 33 1 28 3,875
South America 5,006 6 28 2,679 1,185 8,924
Total by crop 32,888 7,080 775 4,942 21,751 67,436
a Bahrain, Iraq, Jordan, Kuwait, Lebanon, Palestine, Qatar, Saudi Arabia, the Sultanate of Oman, Syria, Turkey, the United ArabEmirates and Yemen
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1809 In exchange, they provide the data from their trials to
1810 CIMMYT for further cataloging and analysis. The use
1811 of the international trial data assists in the improve-
1812 ment of wheat breeding approaches by CIMMYT. For
1813 example, this information has proven useful for
1814 establishing associations among stress environments
1815 and irrigation systems in the screening fields at
1816 CIMMYT’s main wheat breeding sites in Mexico
1817 (Ciudad Obregon, Toluca, and El Batan), and inter-
1818 national test sites for spring bread wheat.
1819 Two basic types of nurseries are distributed, yield
1820 trials containing 30–50 of the best selected advanced
1821 lines and materials nominated by national program
1822 partners; and, screening nurseries containing up to
1823 200 advanced lines selected for specific targeted traits
1824 and elite genetic diversity. Nurseries are distributed
1825 for spring bread wheat, spring durum wheat, spring
1826 and facultative triticale by CIMMYT from Mexico
1827 (Table 8), and winter and facultative bread wheat
1828 from the Turkey/CIMMYT/ICARDA cooperative
1829 program.
1830 Full pedigree and respective selection history are
1831 known for each line and their phenotypic data cover
1832 yield, agronomic, pathological, and quality traits.
1833 Appropriate data analysis for CIMMYT nurseries
1834 tested worldwide or regionally help to understand
1835 genotype-by-environment interactions (GEI), broad
1836 and specific adaptation and their influence on grain
1837 yield and other traits (Ortiz et al. 2007c and references
1838 therein). The recoding of environmental factors, host
1839 plant resistance to biotic stresses and adaptation to
1840 abiotic stresses provides additional means to account
1841 for the GEI. Such information coupled with an
1842 enhanced knowledge regarding the adaptation and
1843 stability of wheat breeding lines allows a better
1844 targeting of bred-germplasm to specific environments,
1845 particularly in stress-prone areas where reducing the
1846 risk of crop failure and better use of inputs are needed
1847 to ensure maximum performance.
1848 Wheat genetic resources enhancement addressing
1849 global challenges
1850 As the world food situation is being transformed by
1851 new driving forces (von Braun 2007), wheat farmers
1852 and researchers are confronted with major challenges
1853 but also emerging opportunities. It may be that the
1854 ‘‘easy gains’’ from wheat research have been
1855exhausted. Clearly, past impacts from wheat research
1856have been greater in high input farming systems,
1857where semi-dwarf cultivars responded well to the
1858increased use of fertilizers and irrigation. Later,
1859spillovers accumulated as improved cultivars spread
1860from irrigated to higher potential rainfed areas, and
1861then progressively into lower potential rainfed areas
1862(Dixon et al. 2006). Looking to the future, will
1863changing consumer preferences and strengthening
1864market value chains create adequate new markets for
1865quality wheat that will justify increased attention to
1866breeding for quality? Will molecular breeding
1867improve the efficiency of field breeding and acceler-
1868ate the release of dramatically more productive lines
1869and cultivars? Does genetically-modified (GM) wheat
1870have significant potential benefits for the industry and
1871consumers? Will the impact of global climate change
1872require major shifts in wheat research and breeding
1873objectives? Are there improved soil and crop man-
1874agement technologies which would enable farmers to
1875obtain the full benefit of new wheat cultivars, while
1876conserving the resource base for future generations of
1877wheat farmers? Are there proven models of integrated
1878‘‘germplasm enhancement—improved crop manage-
1879ment—or more favorable policy environment’’
1880approaches that might be replicated in major wheat-
1881producing areas?
1882Climate change adaptation and mitigation
1883Climate change could strongly affect the wheat crop
1884that accounts for 21% of food and 200 million ha of
1885farmland worldwide (Ortiz et al. 2008b). The Inter-
1886Governmental Panel on Climate Change (2001)
1887already concluded that ‘‘the model of cereal crops
1888indicated that in some temperate areas potential
1889yields increase with small increases in temperatures
1890but decrease with larger temperature changes’’. As a
1891result of possible climate shifts in the Indo-Gangetic
1892Plains—currently part of the favorable, high poten-
1893tial, irrigated, low rainfall mega-environment, which
1894accounts for 15% of global wheat production—as
1895much as 51% of its area might be reclassified as a
1896heat-stressed, irrigated, short-season production
1897mega-environment (Hodson and White 2007). This
1898shift would also represent a significant reduction in
1899wheat yields, unless appropriate cultivars and crop
1900management practices were offered to and adopted
1901by South Asian farmers.
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1902 To adapt and mitigate the climate change effects
1903 on wheat, supplies for the poor, heat-tolerant wheat
1904 germplasm should be bred. Likewise, wild relatives
1905 of wheat as potential sources of genes with inhibitory
1906 effects on soil nitrification must be assessed.
1907 Adaptation to heat and water stresses
1908 Wheat yield in most tropical and subtropical loca-
1909 tions will decrease due to global warming, which may
1910 be further affected by water scarcity or drought (Ortiz
1911 et al. 2007c). Drought and heat are therefore key
1912 factors with high potential impacts on crop yield
1913 (Barnabas et al. 2008) One approach to solve these
1914 heat- and water-related constraints is improving
1915 wheat germplasm with higher tolerance to stresses
1916 associated with these environments. Hence, wheat
1917 breeders should start genetically enhancing the crop
1918 to maintain yield under higher temperatures or water
1919 scarcity using all available means in the tool kit.
1920 Recognizing water productivity and water-use
1921 efficiency as priorities for wheat, CIMMYT research-
1922 ers disaggregated grain yield under water stress into
1923 distinct components to apply these findings to the
1924 genetic enhancement of this crop (Reynolds and
1925 Borlaug 2006). Ongoing research is providing a
1926 better understanding of traits with major effects on
1927 water productivity in dry land wheat areas (Reynolds
1928 et al. 2007). These include root architecture and
1929 physiological traits, resistance to soil-borne pests and
1930 diseases, tolerance to heat and salinity, and zinc-
1931 deficient and boron toxic soils. The combination of
1932 improved germplasm, the Center and partners’
1933 expertise in drought physiology, soil-borne diseases,
1934 agronomy, and the availability of DNA markers for
1935 various traits place CIMMYT in a unique position to
1936 develop water-productive wheat with resistance to the
1937 important stresses for use by partners throughout the
1938 developing world.
1939 Some important attributes for drought-prone envi-
1940 ronments are available in the wild relatives of wheat
1941 (Reynolds et al. 2007). Re-synthesizing hexaploid
1942 wheat with wild ancestors has been used at CIMMYT
1943 for tapping this useful variation and incorporating
1944 such genetic resources into wheat-bred germplasm
1945 (Dreccer et al. 2007). Recently, Ogbonnaya et al.
1946 (2007) found that such lines deriving from re-synthe-
1947 sizing wheat yielded 8–30% higher than the best local
1948 check in multi-site trials across diverse regions of
1949Australia. Their results reinforce previous research
1950conducted at CIMMYT that lines derived from
1951synthetic wheat have the potential to significantly
1952improve grain yield across environments.
1953In addition to the above undertakings, transgenic
1954approaches for incorporating stress-inducible regula-
1955tory genes that encode proteins such as transcription
1956factors (e.g. DREB1A) into the wheat cultigen pool
1957are also being pursued (Hoisington and Ortiz 2008).
1958The DREB1A gene was placed under the control of a
1959stress-inducible promoter rd29A from Arabidopsis
1960rd29A gene and inserted via biolistic approach into
1961bread wheat (Pellegrineschi et al. 2004). Plants
1962expressing the DREB1A gene demonstrated substan-
1963tial resistance to water stress in comparison with
1964checks under experimental greenhouse conditions
1965manifested by a 10-day delay in wilting when water
1966was withheld. Severe symptoms (death of all leaf
1967tissue) were evident in the controls after 15 days
1968without water. The transgenic wheat lines started to
1969show water-stress symptoms only after 15 days. The
1970greenhouse pot experiments based on severe desic-
1971cation stress do not however represent typical field
1972conditions and, therefore, the plants may not be
1973exhibiting a response that would be valuable in
1974farmers’ fields. Hence, CIMMYT researchers shifted
1975their attention to evaluating transgenic wheat lines in
1976contained field trials. Preliminary results showed that
1977the DREB1A gene in wheat significantly lowers
1978canopy temperature compared with the control in
1979these trials mimicking unpredictable mid-season
1980(vegetative phase) drought (Ortiz et al. 2007a). The
1981gene DREB1A driven by abiotic stress inducible
1982promoter rd29A in wheat seemed to delay develop-
1983ment in the transgenic plants and did not result in
1984better grain yields than the control under both
1985irrigated and water-stress conditions. The growth
1986retardation observed on the over-expression of
1987AtDREB1A using 35S CaMV constitutive promoter
1988was overcome using inducible promoter rd29A in
1989transgenic Arabidopsis (Kasuga et al. 1999) and
1990tobacco (Kasuga et al. 2004). However, in some of
1991the transgenic wheat lines with rd29A:DREB1A,
1992growth retardation was observed, which could be
1993due to background expression of this promoter in
1994wheat. We are now pursuing transgenic wheat plants
1995with DREB1A under different inducible promoters,
1996especially promoters with low background, which
1997only switch on when plants are under certain specific
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
1998 stress conditions. For the evaluation of transgenic
1999 wheat plants, pot evaluation is important at early
2000 stage, but it is of great importance to assess
2001 transgenic material in the field as early as possible
2002 to investigate the impact on both plant growth and
2003 grain yield under various appropriate drought-stress
2004 profiles. It is also of interest to investigate at what
2005 stages of growth and in which genetic backgrounds
2006 the transcription factors like DREB1A may have their
2007 most significant effects.
2008 Heat stress already affects wheat plant senescence
2009 and photosynthesis, thereby influencing grain filling.
2010 Wheat cultivars capable of maintaining high 1000-
2011 kernel weight under heat stress appear to possess
2012 higher tolerance to hot environments (Reynolds et al.
2013 1994b). Physiological traits that are associated with
2014 wheat yield in heat-prone environments are canopy
2015 temperature depression, membrane thermo-stability,
2016 leaf chlorophyll content during grain filling, leaf
2017 conductance, and photosynthesis (Reynolds et al.
2018 1998). Amani et al. (1996) used canopy temperature
2019 depression to select for yield under a hot, dry,
2020 irrigated wheat environment in Mexico, whereas
2021 Hede et al. (1999) found that leaf chlorophyll content
2022 was correlated with 1,000-kernel weight while
2023 screening Mexican wheat landraces. Such sources
2024 of alleles coupled with some of the above traits that
2025 defined a new wheat ideotype can provide means for
2026 genetically-enhanced wheat by design in heat-prone
2027 environments.
2028 As indicated above, there are two main wheat
2029 environments in the Indo-Ganges: mega-environment
2030 1 is a favorable, irrigated, low rainfall environment
2031 with high yield potential. In contrast, mega-environ-
2032 ment 5 is a heat-stressed environment (early and late
2033 season heat stress) with available irrigation, but in its
2034 humid and hot areas, the fungi Cochliobolus sativus
2035 cause spot blotch and the Pyrenophora tritici-repentis
2036 (Died.) Drechsler inducing tan spot are pathogens
2037 responsible for leaf blight (Joshi et al. 2007c). These
2038 two major wheat mega-environments in the sub-
2039 continent have been differentiated on the basis of
2040 coolest quarter minimum temperature ranges (3–11�C
2041 for ME-1 and 11–16�C for ME5). In some of the
2042 mega-environment five areas poorer infrastructure,
2043 socio-economic factors, and crop management cou-
2044 pled with the stresses, brought the shortened
2045 vegetative phase and leaf blight ensuing from heat
2046 stress, particularly after flowering stage, which leads
2047to low yield in wheat, the quality of which may be
2048also affected by grain shriveling. Recent data com-
2049piled in the eastern Gangetic plains over 6 years have
2050shown that the higher average temperature observed,
2051especially during the night, was related to higher spot
2052blotch severity and partly explained lower yield
2053performances (Sharma et al. 2007). Disease severity
2054which increases with growth stage depends on crop
2055resilience to heat stress (Duveiller et al. 2005). Thus,
2056improvement of spot blotch resistance in these areas
2057implies a crop physiology adapted to stressed envi-
2058ronments (Sharma et al. 2004). Such observations
2059support, that under global warming, there will be a
2060significant decrease of the most favorable and high
2061yielding mega-environment 1 area due to heat stress,
2062thereby leading to likely yield losses of the wheat
2063grain harvest. Unless appropriate improved germ-
2064plasm, crop husbandry, and resource management are
2065deployed, about 200 million people (based on current
2066population), whose food intake rely on crop harvests
2067in mega-environment 1, will become more vulnerable
2068due to this heat stress affecting the wheat-cropping
2069systems. In this regard, better host plant resistance to
2070leaf blight has been achieved by crossing genetic
2071resistance sources or wild relatives to high-yielding
2072cultivars (Duveiller 2004). Germplasm improvement
2073methods centered on regional partnerships are now
2074more specifically addressing the needs of warmer
2075areas since it is possible to improve host plant
2076resistance of local wheat cultivars based on selective
2077breeding using resistant and agronomically-superior
2078genotypes (Sharma et al. 2004).
2079Mitigation to global warming
2080Nitrous oxide (N2O) is a potent greenhouse gas
2081generated through the use of manure or nitrogen (N)
2082fertilizer and susceptible to de-nitrification, thus often
2083unavailable for crop uptake and utilization. In many
2084intensive wheat-cropping systems common N fertil-
2085izer practices lead to high fluxes of N2O and nitrous
2086oxide (NO). Some plant species show biological
2087nitrification inhibition (BNI) ability that would result
2088in less flux of N2O and NO and retention of N
2089fertilizer for longer time in the soil, which would lead
2090to reducing greenhouse gas. However, wheat, rice,
2091and maize—the most important cereal crops, do not
2092possess BNI (Subbarao et al. 2007b). Leymus
2093racemosus (Lam.) Tzvelev—a wild relative of wheat,
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2094 was found to be a source for inhibiting or reducing
2095 soil nitrification by releasing inhibitory compounds
2096 from roots and suppressing Nitrosomonas bacteria
2097 (Subbarao et al. 2007a). A Leymus chromosome
2098 containing the relevant gene(s) was introduced into a
2099 wheat line that thereafter showed BNI. New research
2100 undertakings are underway to characterize and quan-
2101 tify the biological nitrification inhibition (BNI)
2102 ability from this Leymus-wheat introgression lines
2103 in field trials that may open the path for genetically
2104 enhancing BNI ability in the cultigen pool using this
2105 wild relative as a source for this trait.
2106 Food safety and fighting wheat mycotoxins
2107 Fusarium head blight (FHB) or scab is an important
2108 fungal disease that affects wheat by reducing kernel
2109 weight, yield, and flour-extraction rates in many
2110 important wheat-growing areas in North and South
2111 America, China, and Europe. Several Fusarium
2112 species are associated with scab: Fusarium grami-
2113 nearum Schwabe is the major pathogen worldwide
2114 while F. culmorum (Wm.G. Sm.) Sacc. tends to
2115 predominate in maritime regions (Gilchrist and Dubin
2116 2002). The Fusarium species associated with scab
2117 produce mycotoxins that contaminate the grain. A
2118 number of these compounds have been shown to be
2119 harmful to human and animal health. The mycotoxins
2120 of primary concern with respect to FHB are the
2121 trichothecenes. The most common trichothecene in
2122 grain affected by scab is the mycotoxin known as
2123 deoxynivalenol (DON) produced by F. graminearum
2124 and F. culmorum (Nicholson et al. 2007). A second
2125 trichothecene produced by certain isolates of these
2126 two Fusarium species is nivalenol (NIV). Another
2127 mycotoxin, produced mainly by F. culmorum,
2128 F. graminearum, and F. cerealis (Cooke) Saccardo
2129 is zearalenone (ZEN), a metabolite that is less acutely
2130 toxic and often occurs with trichothecenes (Desjar-
2131 dins 2006; Nicholson et al. 2007). On wheat seedling
2132 tests, the addition of trichothecenes (DON, NIV, and
2133 T-2) to spore suspensions increases lesion size
2134 dramatically, unlike ZEN or mycotoxins produced
2135 by Fusarium species inducing disease on other
2136 cereals. This suggests that trichothecenes play an
2137 important role in lesion development by Fusarium
2138 species associated with FHB (Jiro Murakami,
2139 CIMMYT, unpublished). In recent years, the imple-
2140 mentation of new and more stringent regulations
2141limiting the authorized level of DON in grain and
2142food products has prompted wheat researchers and
2143growers to give more attention to the need to produce
2144wheat with lower amounts of mycotoxins (Ortiz et al.
21452008a).
2146Research toward improved resistance against FHB
2147has been conducted at CIMMYT for more than
214820 years and collaboration with institutes in China,
2149Japan, and Brazil led to the incorporation of superior
2150levels of scab resistance into high-yielding genotypes
2151(Dubin et al. 1996; Ban et al. 2006). However, since
2152the resistance is limited, research is ongoing to
2153expand this resistance base through the identification
2154and validation of QTL associated with field resistance
2155and low levels of DON. In practice, germplasm
2156screening, phenotyping of mapping populations, and
2157detection of novel sources of resistance is conducted
2158under strictly standardized field conditions at El
2159Batan, where CIMMYT is located near Mexico City,
2160under artificial inoculation of F. graminearum iso-
2161lates for which the DON chemotype has been
2162confirmed by the polymerase chain reaction (PCR).
2163At harvest, due to the high cost of mycotoxin
2164detection, only specific research materials and sam-
2165ples of elite wheat germplasm consistently showing a
2166low visual Fusarium head blight index are ground to
2167determine the DON level in the whole grain flour.
2168Analysis for DON content is carried out by means of
2169a commercially available immunoassay test (Rida-
2170screen Fast DON, Biopharm, Germany). CIMMYT is
2171also involved in research aiming at reducing myco-
2172toxin testing costs in particular if a method is used by
2173national research programs. Thus, for selected studies
2174in collaboration with partners in South America,
2175CIMMYT assesses and validates low-cost protocols
2176to determine DON content based on fluorometry
2177(Fuoroquant, Rohmer, Austria). Further applied
2178research includes the validation of quantitative PCR
2179methods aimed at quantifying DON based on the
2180fungal biomass in the grain and the presence of the
2181Tri-5 gene responsible for DON production, analyz-
2182ing the correlation with FHB field symptoms. Other
2183attempts to speed the detection of low DON content
2184in advanced wheat lines included the evaluation of a
2185lateral-flow colloidal gold-based immunoassay for
2186the rapid detection of deoxynivalenol with two
2187indicator ranges (De Saeger et al. 2008).
2188One of the objectives of CIMMYT’s wheat
2189breeding program in coming years is to distribute
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2190 high-yielding wheat germplasm with acceptable end-
2191 use quality and with resistance to FHB which also
2192 harbor a low DON content. Thus, efforts are being
2193 made to add the information on the DON content in
2194 each line when nurseries are distributed (Lewis et al.
2195 2007). Since the main obstacles to provide reliable
2196 quantitative information on resistance to mycotoxins
2197 (i.e., DON) in wheat germplasm are the high field
2198 evaluation and assay costs, understanding the factors
2199 affecting the correlation between phenotypical data
2200 and mycotoxin content is paramount to improve data
2201 accuracy and information reliability.
2202 Grain quality for adding value in the commodity
2203 chain
2204 Globalization, the continuous increase in urbaniza-
2205 tion, and improvements in the income of people in
2206 developing countries have been generating a large
2207 demand for wheat crops possessing specific quality
2208 traits. For example, consumption of western style
2209 food such as pan bread and cookies are increasing
2210 rapidly in large wheat-producing countries such as
2211 China and India, and consumption of flat breads and
2212 flour noodles are becoming popular as convenience
2213 foods in the western hemisphere. In addition, the
2214 demand for high quality traditional products such as
2215 noodles, steamed bread, flat breads, pasta or couscous
2216 among others, is also increasing quickly. Hence,
2217 grain-processing quality and quality uniformity are
2218 becoming more important breeding issues in devel-
2219 oping countries. In wheat breeding, improving wheat
2220 productivity, disease resistance, and tolerance to
2221 biotic stresses and wheat quality attributes need to
2222 be addressed holistically to develop wheat cultivars
2223 satisfying both the producer and the consumer and in
2224 order to maintain competitive and sustainable pro-
2225 duction systems.
2226 End-use quality traits
2227 Flour yellowness is undesirable for producing breads
2228 and most Asian flour noodles (Liu et al. 2003), except
2229 for yellow alkaline flour noodles. In contrast, in
2230 durum wheat semolina, a high concentration of
2231 yellow carotenoids (mainly lutein) is highly desir-
2232 able, for it results in bright, yellow pasta products.
2233 Sprouted grain shows high alpha-amylase activity,
2234 which negatively affects the processing quality of
2235bread-, cake- and cookie-making. Sound wheat may
2236present different levels of poly-phenol oxidase activ-
2237ity, which promote the time-dependent darkening of
2238cooked Asian noodles (Anderson and Morris 2003),
2239and of lipoxygenase, which promotes the oxidation of
2240carotenoids reducing pasta yellowness (Leenhardt
2241et al. 2006).
2242Hard wheat produces flours with much higher levels
2243of starch damage and water absorption as compared to
2244soft wheat. This difference in water-absorption capac-
2245ity makes soft wheat (and not hard wheat) suitable for
2246cookie making and hard wheat (and not soft wheat)
2247suitable for bread making (Souza et al. 2002). Grain
2248hardness is influenced mainly by allelic variations and
2249is closely linked to puroindoline genes. Cane et al.
2250(2004) observed that some allelic variations of Pina
2251show differential effects on milling yield.
2252The main factor determining wheat end-use is the
2253gluten protein. Gluten is composed of monomeric
2254gliadins and polymeric glutenins. Allelic variations
2255controlling gliadin and glutenin composition (mainly
2256at the Glu-1, Glu-3, and Gli-1 loci), are responsible
2257for most variation in dough properties (strength and
2258extensibility), dough-mixing properties, and bread-
2259and Chinese noodle-making quality (Branlard et al.
22602001; He et al. 2005; Liu et al. 2005). In durum
2261wheat, the best pasta-cooking quality is achieved
2262from wheat possessing Glu-B3 LMW-2 type alleles
2263(Pena and Pfeiffer 2005 and references therein).
2264Granule-bound starch synthase (GBSSI) is respon-
2265sible for amylose synthesis in the grain. Grain
2266amylose levels are affected by allelic variations of
2267the Wx gene complex (Geera et al. 2006). Starch
2268pasting viscosity affects the eating quality of wheat
2269flour noodles, particularly Japanese white noodles,
2270which are smooth, soft, and slightly elastic (Crosbie
22711991). Increased starch-swelling power and desirable
2272noodle softness have been associated with the
2273absence of GBSSI controlled by the Wx-B1a gene
2274on chromosome 4A (Ross et al. 1996).
2275Quality testing methodology
2276CIMMYT addresses wheat quality improvement by
2277examining and defining several factors associated
2278with wheat processing and end-use quality, and by
2279applying diverse molecular and conventional analyt-
2280ical and screening tools required to achieve the
2281improvement of relevant grain quality-related traits.
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2282 Ongoing research in China and Mexico are examples
2283 of this approach.
2284 Knowledge about the genetic control and the
2285 influence of allelic variants of main traits (Table 10)
2286 is fundamental for efficient quality improvement.
2287 However, we cannot ignore that some of these quality
2288 traits may be strongly influenced by genotype-by-
2289 environment (heat, drought, high humidity) interac-
2290 tions (Eagles et al. 2002; Spiertz et al. 2006) and
2291 nutrient availability, particularly at the grain-filling
2292 stage (Dupont et al. 2006).
2293 At CIMMYT electrophoresis (SDS-PAGE) is
2294 commonly used to identify allelic variations at Glu-
2295 1, Glu-3, and Gli-1 in parental lines. In addition,
2296 rapid, small-scale tests including marker-assisted
2297 selection (MAS), near-infrared spectroscopy (NIRS),
2298 SDS-sedimentation, and dough rheology, are applied
2299 at different stages of the breeding process. MAS
2300 offers the best option for specific traits controlled by
2301 single genes because it is performed on leaf tissue
2302 before seed-setting. The flour sedimentation test,
2303 NIRS, and the mixograph, on the other hand, allow
2304 rapid estimation of important physical, composi-
2305 tional, and functional grain and flour factors closely
2306 associated with milling and wheat processing quality.
2307 Finally, the use of flour and dough-testing method-
2308 ologies (milling yield, dough viscoelasticity, starch
2309 pasting properties, and actual end-product quality)
2310 allow characterizing more specifically for processing
2311 and end-use quality attributes of advanced lines of
2312 spring and winter/facultative germplasm.
2313 Breeding strategy, quality testing, and screening
2314 In setting breeding priorities and strategies, one must
2315 determine: the cultivar’s intended end-uses and the
2316demands of the targeted market, specific quality traits
2317to breed for, and genotype-by-environment-by-man-
2318agement interactions that may influence the quality of
2319the resulting cultivar.
2320Information on glutenin and gliadin subunit com-
2321position helps in the designing of crosses aimed at
2322achieving allelic combinations known to contribute
2323positively to dough properties required for producing
2324leavened and flat breads, flour noodles, cookies, and
2325pasta. Screening may initiate in F4–F5 applying MAS
2326for desirable quality-related genes or allelic varia-
2327tions. When possible, the use of rapid, high-
2328throughput conventional small-scale tests (e.g. SDS-
2329sedimentation and NIRS) should be used in addition
2330to MAS to screen for complex traits under multigenic
2331control. Because MAS and conventional small-scale
2332quality tests explain end-use quality only partially
2333(Graybosch et al. 1999; Kuchel et al. 2006), it is
2334advisable to apply quality screening based on more
2335specific processing traits (dough viscoelasticity and
2336mixing properties, starch pasting properties, baking
2337performance) and end-product quality attributes in
2338advanced breeding stages; i.e., F6–F9 (Pena et al.
23392002; Souza et al. 2002). Finally, multi-location yield
2340trials exposing advanced elite lines to environmental
2341variation (and farmers’ crop management practices)
2342are necessary to identify the few genotypes combining
2343stable yield and quality attributes across locations.
2344China and CIMMYT have had a very strong and
2345close collaboration regarding germplasm exchange,
2346human resource development, and in the establish-
2347ment of diverse analytical tests efficient to improve
2348bread and noodle quality attributes. ‘Jinan 17’ and
2349‘Jimai 19’ are former leading cultivars in northern
2350China, and ‘Jinmai 20’ and ‘Yumai 34’ are good pan-
2351bread making quality and leading cultivars in the
Table 10 Main wheatquality traits and theirgenetic control
Quality trait Loci Chromosome
Grain hardness (puroindolins) PinA-D1, PinB-D1 5DS
Alpha amylase Amy-1, Amy-2 Groups 6 and 7
Poly phenol oxidase Ppo-A1, Ppo-D1 2AL, 2D
Protein content Pro-1, Pro-2 5D
Glutenins Glu-1, Glu-3 1A, 1B, 1D
Gliadins Gli-1, Gli-2 Groups 1 and 6
Secalins Sec-1 1R
Starch granule-bound synthase Wx-A1, Wx-B1, Wx-D1 7AS, 4AL, 7DS
Yellow Pigment PsyA1, Psy-B1 7A, 7B
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2352 Yellow and Huai Valleys. They derived from
2353 CIMMYT-bred germplasm. Improvement of gluten
2354 quality is the top priority for quality breeding in
2355 China. SDS sedimentation value, mixograph param-
2356 eters, and high- and low- molecular weight glutenin
2357 subunits are primarily used for characterization of
2358 crossing parents and selection in early generations.
2359 Selection of desirable grain hardness Pin-B1 alleles
2360 (Pinb-D1b) is highly desirable. Grain hardness, SDS
2361 sedimentation, peak viscosity, polyphenol oxidase
2362 (PPO) activity and yellow pigment content, can be
2363 used to screen for Chinese white noodle quality in the
2364 early generation of a wheat breeding program;
2365 molecular markers for selection of PPO activity (He
2366 et al. 2007), yellow pigment content (He et al. 2008),
2367 and high starch viscosity (Briney et al. 1998) are also
2368 available for screening. Molecular markers have been
2369 routinely used to characterize crossing parents and for
2370 confirmation of the presence of targeted genes in
2371 breeding programs, both in Mexico and China. Two
2372 multiplex PCR assays targeting improvement of
2373 bread-making quality (including five genes) and of
2374 noodle quality (including three genes) were also
2375 developed to improve the efficiency and reduce the
2376 costs of applying MAS in wheat breeding programs
2377 (Zhang et al. 2008).
2378 Future challenges in quality improvement
2379 The future challenge is to improve dough extensibility
2380 and quality stability. The combination of high yield
2381 potential and good quality and integration of diverse
2382 quality donors from wheat relatives is advisable.
2383 Wheat breeders still face the challenge of unveiling
2384 the complex mechanisms involved in genotype-by-
2385 environment-by-management interactions affected by
2386 heat, drought, erratic climate, nutrient availability
2387 during grain development, among other factors that
2388 significantly influence yield and quality stability
2389 (Dupont et al. 2006; Eagles et al. 2002; Geera et al.
2390 2006; Spiertz et al. 2006). Furthermore, an excess of
2391 three billion people in the world are affected by
2392 micronutrient deficiencies that cause serious health
2393 problems. In addition, obesity and associated illnesses
2394 are becoming serious public health problems in many
2395 parts of world. Breeders need to immediately start
2396 serious undertakings to improve wheat nutritional
2397 value and health-related issues associated with the
2398 consumption of wheat-based foods.
2399Human nutrition/health and wheat biofortification
2400Zinc deficiency is implicated in health problems
2401throughout the world, especially across a wide band
2402of countries in West Asia, North Africa, and the
2403South Asian subcontinent where more than half of
2404inhabitants’ daily calories come from wheat (CI-
2405MMYT 2004). In South and West Asia, millions of
2406heavy wheat consumers are also iron deficient.
2407Women and children are particularly prone to zinc
2408and iron malnutrition.
2409The health of poor people may be enhanced by
2410breeding staple food crops that are rich in micronu-
2411trients, a process referred to as biofortification. In
24122003, the CGIAR launched the Challenge Program
2413HarvestPlus with the aim of breeding and dissemi-
2414nating crops for better nutrition. Within this global
2415alliance undertaking, CIMMYT is developing high-
2416yielding wheat cultivars with grain containing
241730–50% more iron and zinc (Ortiz-Monasterio et al.
24182007). The potential impact is dramatic given that
2419wheat cultivars bred by CIMMYT and its partners
2420cover 80% of the global spring wheat area.
2421The best sources of these micronutrients are wild
2422species that do not cross easily with modern wheats
2423(Cakmak et al. 2000). Researchers have therefore
2424developed a ‘‘bridge’’ line by crossing one such grass
2425(Ae. tauschii) with a high-micronutrient primitive
2426wheat (Triticum dicoccon Schrank). The resulting
2427‘‘bridge’’ lines combine readily with modern wheat
2428cultivars, producing lines whose grain contains more
2429iron and zinc than modern wheat. Partners in India
2430and Pakistan are using this approach to develop high-
2431yielding, disease-resistant, biofortified wheat for
2432South Asia.
2433In Turkey, home to pioneering research on zinc
2434deficiency and wheat, wheat landraces and cultivars
2435that take up and use zinc more efficiently are being
2436combined with wheat cultivars that have resistance to
2437yellow rust and root diseases (CIMMYT 2004).
2438CIMMYT researchers, with partners from labs else-
2439where, are identifying molecular markers for genes
2440that control grain iron and zinc levels, to facilitate
2441their transfer to new cultivars.
2442Recent research undertaken by CIMMYT on iron
2443and zinc losses in milling and cooking, suggests that
2444milling reduces Fe and Zn levels in the flour but
2445hydrate is reduced even more which results in a more
2446favorable Zn/hydrate ratio, which is highly correlated
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2447 with bioavailability (Ortiz-Monasterio unpublished
2448 results). A flour extraction rate of 80% is best to
2449 optimize function of increasing bioavailability versus
2450 decreasing content with decreasing flour extraction.
2451 This and the center’s research on bio-availability will
2452 help determine exactly how much it helps to eat
2453 biofortified daily bread.
2454 Empowering rural people through participatory
2455 wheat improvement and seed systems
2456 Three-quarters of the world’s poorest people live
2457 below US$ 1 a day of which 37% are in South Asia.
2458 In this region, particularly in the eastern Indo-
2459 Gangetic Plains, there is a high level of poverty,
2460 malnutrition, and food insecurity. The number of
2461 poor people is expected to increase still further as
2462 population growth exceeds that of agriculture pro-
2463 ductivity. Wheat, covering an area of some
2464 37 million ha, is one of the economic mainstays in
2465 the region. However, the productivity of wheat
2466 cropping systems lags far behind its potential. Hunger
2467 is inextricably linked to poverty and vulnerability.
2468 The millennium development goal of the elimination
2469 of extreme poverty and hunger can only be met by
2470 increases in agricultural productivity, particularly by
2471 resource-poor farmers, which is fundamental to
2472 growth and poverty reduction in the region.
2473 South Asia produced 101.3 million t of wheat in
2474 2007. India and Pakistan harvested 74.9 and 23.3 mil-
2475 lion t, respectively. In spite of this output, the region
2476 is projected to have a wheat deficit of 21 million t by
2477 2020 (Agcaoili-Sombilla and Rosegrant 1994). If
2478 these projections are correct, Pakistan will have the
2479 highest deficit with 15 million t, followed by Bangla-
2480 desh with 4 million t. India would have to increase its
2481 production from the current 74.9 million t to about 90
2482 (Joshi et al. 2007b) or 109 million t by year 2020
2483 (Nagarajan 2005). To achieve this output, the average
2484 national wheat yield would have to increase from its
2485 current 2.63 t ha-1 to almost 4 t ha-1 in the next
2486 12 years. This is a significant challenge given that
2487 resources for research in these countries are shrinking
2488 and that the present estimated yield gap between
2489 farmers and experimental yields is about 1.8 t ha-1
2490 (Ortiz-Ferrara et al. 2007a).
2491 Surveys conducted in the region suggest that one
2492 of the main causes of low yields is the predominance
2493 of older (at least 1.5 decades) wheat cultivars. These
2494cultivars are genetically inferior to more recently
2495bred-germplasm, and are increasingly susceptible to
2496diseases. Seed replacement is around 10% and there
2497is significant lack of access to seed of appropriate
2498cultivars, especially for those that are adapted to
2499marginal environments. The varietal adoption trends
2500described above are in large part due to the poor
2501extension and weak seed production systems pre-
2502valent in most of these countries. Hence, a system for
2503dissemination of new technologies and for develop-
2504ing community seed industry by local farmers and
2505their groups is important to empower rural people. It
2506is essential to promote new materials and to also
2507make crop production more profitable for farmers.
2508CIMMYT, in partnership with the Center for Arid
2509Zone Studies-Natural Resources (CAZS-NR, Bagor,
2510Wales), has been collaborating with farmers, national
2511programs, and other South Asian partners to promote
2512improved wheat cultivars and new resource-conserv-
2513ing technology (RCT) options in farmers’ fields.
2514Participation fostered among farmers, scientists,
2515extension specialists, non-governmental organiza-
2516tions (NGOs), and the private sector includes
2517participatory variety selection (PVS), and participa-
2518tory evaluation of agronomic practices. Through
2519PVS, several farmer-preferred technologies have
2520been identified including wheat cultivars for adverse
2521conditions in eastern Uttar Pradesh, India (Singh et al.
25222007), and for boron deficiency in some locations of
2523Nepal. There has been considerable improvement in
2524farmers’s access to new cultivars and technologies in
2525the rural areas. Yield increases (15–70%) have been
2526achieved by resource-poor farmers through the
2527adoption of new cultivars and RCT. The farmers
2528have also made substantial cost savings and achieved
2529higher yields through resource-conserving agronomic
2530techniques such as zero- or reduced-tillage. Seed of
2531the new farmer-selected cultivars has been multiplied
2532by groups of collaborating farmers and widely
2533distributed.
2534Following this participatory research, several
2535thousands of farming households have rapidly
2536adopted cultivars and benefited from them. Seed
2537saving by farmers and farmer-to-farmer seed spread
2538of new cultivars has been evident which has brought
2539sustainable impact. Strengthening and capacity-build-
2540ing of local institutions including community-based
2541seed production and distribution has also been taking
2542place. Both men and women farmers have been
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2543 participating in this partnership in various activities
2544 addressing gender perspective and empowerment
2545 issues. They are willing to collaborate with all of
2546 the partners to improve the returns to wheat cropping
2547 systems through participatory approaches.
2548 Partnerships for development, dissemination, and
2549 adoption of improved technologies for improving
2550 wheat productivity
2551 Participatory research and development has been
2552 reported as an efficient approach for up-scaling new
2553 agricultural technologies (Ortiz Ferrara et al. 2001;
2554 Witcombe et al. 2001). It is capable of better
2555 addressing farmers’ problems that very often are
2556 not recognized due to the complexity of farmers’
2557 different situations and the vast diversity in farmers’
2558 fields. Participatory research has been used to com-
2559 plement ongoing research to help farmers by
2560 providing them with a wider choice of options to
2561 evaluate under their own conditions (Witcombe et al.
2562 1996, 2005).
2563 The ‘‘mother-baby’’ trial system, which was suc-
2564 cessfully used for maize in sub-Saharan Africa
2565 (Snapp 1999; de Groote et al. 2002), proved to be
2566 an efficient approach for developing and disseminat-
2567 ing new cultivars and RCT options through close
2568 collaboration with farmers. Under this approach, a
2569 large number of ‘‘mother-baby’’ trials are grown in
2570 different villages to provide better options for farm-
2571 ers. The ‘‘mother’’ trial consists of 12–15 elite wheat
2572 lines or recently released cultivars, which include as
2573 check the most popular cultivars grown by farmers at
2574 each site. A number of ‘‘baby’’ trials, consisting of
2575 just one of the elite cultivars in the ‘‘mother’’ trial
2576 plus a local check, are grown around the ‘‘mother’’
2577 trial in the same year. Also, seed multiplication plots
2578 of all the genotypes in the ‘‘mother’’ and ‘‘baby’’
2579 trials are planted simultaneously to harvest clean
2580 seeds. Bred-lines and cultivars in the ‘‘mother’’ and
2581 ‘‘baby’’ trials are assessed in a collaborative and
2582 consultative mode by farmers. Quantitative feedback
2583 is analyzed and used to ascertain farmers’ prefer-
2584 ences. Clean seed of the farmer-selected cultivars is
2585 distributed to collaborating farmers. In the following
2586 year, farmers’ selected genotypes are tested in large
2587 fields along with the farmers’ local cultivar. Farmers
2588 are trained on seed production to harvest quality seed
2589 from the large plots in the second year. In this way,
2590farmers have clean seed of their preferred cultivars
2591for further planting. Often the participating farmers
2592harvest more seed than they need. The additional seed
2593is sold to other farmers in the community. Thus the
2594new cultivars are spread from farmer to farmer.
2595Experiences in improving food security and income
2596generation
2597In Nepal, a total of 26 wheat cultivars were tested in
259863 villages in seven districts from 2002 to 2005.
2599These cultivars were tested in a wide range of
2600environments using two different sets of PVS
2601‘‘mother’’ trials. As a result, about seven bred-lines
2602or cultivars were identified by participating farmers.
2603Three released cultivars are under extensive seed
2604multiplication by the public seed industry. Farmers in
2605the southern lowlands and in the mid-central hills of
2606the Kathmandu Valley are also involved in commu-
2607nity seed production. These adoption trends have
2608helped farmers in those areas to increase food
2609sufficiency (i.e., households with sufficient food for
2610the year) from 65% in 2002 to 71% in 2005. Also,
2611there were households with food sufficiency for
2612longer period in 2005 compared to 2002. These gains
2613in food sufficiency are primarily attributed to three
2614factors. The farmers reported that of the gains in food
2615sufficiency, 41.3% was due to the cultivar, 44.4%
2616from crop management including fertilizers, and
261714.3% from irrigation. Farmers in Bangladesh have
2618identified at least three new wheat cultivars since
26192005. Farmers who have adopted them have
2620increased their income (Table 11). The yield increase
2621over the widely popular cultivar ‘Kanchan’ was
2622705 kg, which translates into additional income of
2623about US$ 164 ha-1. Varietal diversification and
2624income generation has also increased in eastern India
2625as a result of collaboration with the Indian Council of
2626Agricultural Research (ICAR) and the Rice-Wheat
2627Consortium for the Indo-Gangetic Plains (RWC).
2628Many farmers in Uttar Pradesh are successfully
2629engaged in community seed production. At one of
2630the sites, around one-third of the area came under
2631seed production after 3 years of PVS. In general,
2632farmers at the PVS sites recorded greater profits than
2633at non-PVS sites. Around 83% of the farmers at the
2634PVS sites responded that they were making a profit of
2635at least US$ 67 ha-1. The profit increase was mainly
2636due to increases in productivity, decreases in the cost
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2637 of cultivation, and improved earnings due to quality
2638 seed production of farmer-preferred cultivars. Based
2639 on this new outcome, participatory seed production
2640 was carried out at all the locations where PVS
2641 activities were conducted with the objective of
2642 equipping and training farmers for seed production
2643 of high-yielding cultivars. As a result, they could
2644 multiply seed of their preferred cultivars and not be
2645 dependent on outside sources. This approach was
2646 expected to enhance farmers’ profitability through
2647 lowered seed costs, lower costs of production due to
2648 the adoption of RCT, and surplus grain from higher
2649 production. This concept could be extended to other
2650 crops. The participatory seed production was found
2651 extremely useful for participating farmers, since at
2652 almost all the locations they were able to learn to
2653 produce their own high quality seed. The impact of
2654 participatory seed production is being realized for the
2655 first time on such a wide scale in the area and several
2656 farmers’ societies for seed production have been
2657 established in eastern Uttar Pradesh and the bordering
2658 districts of Bihar. This farmer-participatory approach
2659 has helped resource-poor farmers improve their
2660 livelihoods, while ensuring staple food for the region
2661 through the adoption of new cultivars and other RCT
2662 options.
2663 Ug 99 stem rust as a global emerging threat
2664 to wheat (food) supply
2665 Stem or black rust of wheat is historically known to
2666 cause severe devastation; however, it remained under
2667 control for almost 40 years through the use of host
2668 plant resistance. In 1998, severe stem rust infections
2669 were observed on wheat in Uganda, and a race,
2670 commonly known as Ug99 (TTKSK on North
2671 American nomenclature system) with virulence on
2672resistance gene Sr31, which was initially transferred
2673from rye to wheat, was identified. Race Ug99 was
2674subsequently detected in Kenya and Ethiopia in 2005,
2675in Sudan and Yemen in 2006, and in Iran in 2007. It
2676is predicted that Ug99 will continue to migrate to
2677North Africa, the Middle East, South Asia, and
2678beyond through winds or other means. The most
2679striking feature of Ug99 is that it not only carries
2680virulence to gene Sr31 but also this unique virulence
2681is present together with virulence to most of the genes
2682of wheat origin, and virulence to gene Sr38 intro-
2683duced into wheat from Aegilops ventricosa Tausch.
2684and bred in several European and Australian cultivars
2685and a small portion of new CIMMYT germplasm
2686(Singh et al. 2006). This virulence combination might
2687have accounted for the wide-spread Ug99 suscepti-
2688bility in wheat cultivars worldwide. New variants of
2689this race with virulence to Sr24 and Sr36 were
2690detected in Kenya in 2006 and 2007, respectively. It
2691is anticipated that mutation toward more complex
2692virulence will likely occur as the fungal population
2693size increases and selection pressure is placed on the
2694population by cultivars protected by additional race-
2695specific resistance genes. It is estimated that between
269690 and 95% of the global wheat area is planted with
2697wheat cultivars that are susceptible to Ug99 or its
2698new variants.
2699The Borlaug global rust initiative
2700Reducing the area planted to susceptible cultivars in
2701primary risk areas of East Africa, the Arabian
2702Peninsula, North Africa, the Middle East, and West/
2703South Asia is the best strategy if major losses are to
2704be avoided. The ‘‘Borlaug Global Rust Initiative’’
2705(www.globalrust.org), launched in 2005, is using the
2706following strategies to reduce the possibilities of
Table 11 Income change in villages at Daulatpur, Hatiari and Jogda (Dinajpur, Bangladesh) due to farmer-preferred wheat cultivarsand adoption of recommended practices after participatory varietal selection (2002–2005)
Wheat Cultivar Area(ha)
Yield(kg ha-1)
Yieldincrease(kg ha-1)
Seedsaved(kg ha-1)
Seedsaved in52% area(kg)
Price of Additionalincome
Add prod(91,000 Tk)a
Saved Seed(91,000 Tk)
91,000Tk
Tk/ha
Old popular cultivar ‘Kanchan’ 67.71 2,274
Farmer-preferred cultivars 40.96 2,979 705 80 1,704 361.0 27.26 388.26 9,479
Total 108.67 2,540 266 4,521 361.3 72.34 433.64 3,990
a Tk = Taka local currency, i.e., US$ 1 = Tk 58
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2707 major epidemics: monitoring the spread of race Ug99
2708 beyond eastern Africa for early warning and potential
2709 chemical interventions, screening of released varie-
2710 ties and germplasm for resistance, distributing
2711 sources of resistance worldwide for either direct use
2712 as varieties or for breeding, and breeding to incor-
2713 porate diverse resistance genes and adult plant
2714 resistance into high-yielding adapted cultivars and
2715 new germplasm. An aggressive strategy to identify
2716 and promote high-yielding, resistant cultivars is the
2717 only viable option to reduce potential losses as
2718 resource-poor and commercial farmers in most of
2719 Africa, the Middle East, and Asia cannot afford
2720 chemical control or may not be able to apply chem-
2721 icals in the event of large scale epidemics due to their
2722 unavailability for timely application. Reduction of
2723 susceptible cultivars throughout the primary risk area
2724 should reduce wind dispersal of spores from these
2725 areas to the secondary risk areas.
2726 Strategy to breed race-specific resistance genes
2727 The high frequency of the highly resistant wheat
2728 materials from South America, Australia, the USA,
2729 and CIMMYT was identified through the 2005 and
2730 2006 screenings with Ug99 in Kenya. They possess
2731 Sr24, located on the Th. elongatum translocation on
2732 chromosome 3DL together with the leaf rust resis-
2733 tance gene Lr24. The presence of race TTKST with
2734 Sr24 virulence at low frequencies in Ug99 lineage
2735 during 2006 resulted in a rapid buildup sufficient to
2736 cause an epidemic on the Sr24-carrying cultivar
2737 ‘Kenya Mwamba’ in 2007, which occupied about
2738 30% of the Kenyan wheat area. The situations
2739 described above have once again reminded us of
2740 the consequence of dependence on single race-
2741 specific genes in the control of stem rust in areas
2742 where rust is endemic.
2743 Diverse sources of effective race-specific resis-
2744 tance genes, mostly derived from wheat relatives, are
2745 available for breeding. Genes Sr25 and Sr26, derived
2746 from Th. elongatum, have been previously used
2747 successfully in developing cultivars. Other genes,
2748 practically not used in wheat improvement but that
2749 may have good chances of succeeding are Sr27 of rye
2750 origin, and Sr22 and Sr35 derived from T. monococ-
2751 cum. Although alien resistance genes Sr29, Sr32,
2752 Sr33, Sr37, Sr39, Sr40, and Sr44 have not been used
2753 widely in breeding, preliminary experience has
2754indicated their association with unwanted agronomic
2755traits and therefore sizes of alien chromosome
2756segments must be reduced before they can be used
2757successfully. The undesignated resistance genes
2758SrTmp (wheat origin), SrR and Sr1A.1R (rye origin),
2759and a few other uncharacterized sources originating
2760from re-synthesized hexaploid lines or bread wheat
2761offer further diversity.
2762The fastest way to reduce the susceptibility of
2763important wheat cultivars and the best new germ-
2764plasm is to systematically incorporate diverse sources
2765of resistance through limited or repeated backcross-
2766ing. Because most of these Ug99-effective genes are
2767of alien origin, co-segregating molecular markers for
2768some of them are already available and being used for
2769selection at CIMMYT. To avoid fast breakdown, the
2770best strategy is to use race-specific resistance genes in
2771combinations. Molecular markers provide a powerful
2772tool to identify plants that carry combinations of
2773resistance genes. Markers for other genes need to be
2774developed to facilitate their utilization.
2775One major issue remains in that various currently-
2776effective resistance genes are already present in some
2777advanced spring breeding materials that are being
2778tested in various countries to mitigate the immediate
2779threat from Ug99. Whether of not they should they be
2780deployed until their combinations are developed, is a
2781difficult issue to resolve, and there is a wide range of
2782opinions on the matter. This has provoked the
2783CIMMYT wheat improvement group to focus their
2784breeding efforts towards breeding minor genes based
2785on adult plant resistance, especially for areas consid-
2786ered to be under high risk and where survival of the
2787pathogen for several years is expected due to the
2788presence of susceptible hosts and favorable environ-
2789mental conditions. It is thought that this strategy will
2790allow other areas of the world, especially facultative
2791and winter wheat growing regions to use race-specific
2792resistance genes more successfully in their breeding
2793programs.
2794Identification and breeding for complex adult plant
2795resistance
2796Durable stem rust resistance of some of the older US,
2797Australian, and CIMMYT spring wheat lines and
2798cultivars is believed to be due to the deployment of
2799Sr2 in conjunction with other unknown minor,
2800additive genes that could have originated from
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2801 ‘Thatcher’ and the ‘Thatcher’-derived line ‘Chris’
2802 (Singh et al. 2006). Sr2 can be detected through its
2803 complete linkage with pseudo-black chaff phenotype.
2804 Genotypes with negligible expression of pseudo-
2805 black chaff can be selected in breeding materials. Sr2
2806 does not confer adequate resistance under high
2807 disease pressure when present alone. It was detected
2808 in several highly-resistant old, tall Kenyan cultivars,
2809 including ‘Kenya Plume’ and the CIMMYT-derived
2810 semi-dwarf wheat lines ‘Pavon 76,’ ‘Kritati,’ and
2811 ‘Kingbird.’ ‘Pavon 76’ and ‘Kiritati’ were resistant
2812 since the initiation of rigorous screening in 2005 at
2813 Njoro (Kenya) with maximum disease scores of
2814 20MR-MS. ‘Kingbird,’ a new advanced line, is at
2815 present the best known source of adult plant
2816 resistance in semi-dwarf wheat with maximum score
2817 recorded to be 5 MR-MS during the same period.
2818 Because these wheat lines are susceptible as seedlings
2819 with Ug99, their resistance is speculated to be based
2820 on multiple additive genes where Sr2 is an important
2821 component.
2822 With the exception of Sr2, little is known on the
2823 genes involved in durable adult plant resistance.
2824 However, earlier work done by Knott (1982),
2825 knowledge on durable resistance to leaf and yellow
2826 rusts (Singh et al. 2004), and observations made on
2827 breeding materials and a F6 mapping population
2828 involving ‘Pavon 76,’ indicate that the rate of rust
2829 progress is a function of the cumulative effect of the
2830 number of minor genes present in a genotype and the
2831 individual effects of each gene. Accumulation of four
2832 to five slow-rusting, minor genes is therefore
2833 expected to retard disease progress to rates that result
2834 in negligible disease levels at maturity under high
2835 disease pressure, described as ‘‘near-immunity’’ by
2836 Singh et al. (2000).
2837 Because a large portion of CIMMYT high-yielding
2838 spring wheat germplasm does not carry effective
2839 race-specific stem rust resistance genes to Ug99 and
2840 several lines were identified to carry at least moderate
2841 levels of resistance, we view this as a perfect
2842 opportunity to reconstitute high levels of adult plant
2843 resistance in new wheat materials. Due to the lack of
2844 molecular markers for adult plant resistance genes
2845 and the absence of the Ug99 race in Mexico, a shuttle
2846 breeding scheme between two Mexican sites and
2847 Njoro was initiated in 2006 to transfer or accumulate
2848 high levels of adult-plant resistance identified in
2849 semi-dwarf CIMMYT wheat lines to a range of
2850important wheat germplasm. We expect that the
2851frequency of advanced lines which carry high yield
2852potential and maintain wide adaptation, end-use
2853quality characteristics, and high levels of resistance
2854to all three rusts, will increase over time through the
2855use of the Mexico-Kenya shuttle breeding.
2856Replacing susceptible cultivars in Africa, the Middle
2857East, and Asia
2858Screening in Kenya during 2005, 2006, and 2007 has
2859identified a few resistant released cultivars or
2860advanced breeding materials at various stages of
2861testing in most of the countries that submitted their
2862materials for screening. One strategy is to find ways
2863to ensure that the best, high-yielding resistant mate-
2864rials occupy at least 5% of total wheat area
2865distributed throughout the wheat region and are
2866readily available for use as seed in the case that
2867Ug99 establishment is evident in a particular country.
2868To occupy a large area the resistant cultivars must
2869have superior yields to current popular cultivars. This
2870objective is achievable as most of the current popular
2871cultivars were bred during early- to mid-1990s, and
2872the yield potential of current CIMMYT spring wheat
2873germplasm has progressed significantly since then.
2874Yield performances of 14 new high-yielding, Ug99-
2875resistant wheat lines together with current cultivars
2876were determined during the 2006–2007 crop season
2877at 27 sites in India, Pakistan, Nepal, Afghanistan,
2878Iran, Egypt, Sudan, Syria and Mexico. Results show
2879that at least three lines, possessing race-specific or
2880adult plant resistance, in each country showed yield
2881superiority in the range of 10–15% over the current
2882local cultivars. These superior lines are being tested
2883more extensively during the 2007–2008 season for
2884their performance in the above countries and seed is
2885being multiplied simultaneously. Testing of addi-
2886tional wheat materials, mostly with adult plant
2887resistance, is also underway.
2888Fitting bred-genotypes into sustainable
2889and improved crop management practices
2890Crop system management research adds value to
2891improved germplasm by increasing the degree to
2892which yield potential is achieved—about half of the
2893increase in wheat productivity at the farm level is
2894often attributed to crop system management and more
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
2895 in stressed environments—and there is often synergy
2896 between these effects (the G 9 System interaction).
2897 CIMMYT holds a unique comparative advantage for
2898 such research at the interface between crop improve-
2899 ment and crop system management.
2900 Figure 5 shows the important role of crop man-
2901 agement regarding maximizing yield potential. The
2902 lower line shows the yields of a historical set of
2903 wheat cultivars bred from 1962 to 1988 based on
2904 applying the current crop management practices that
2905 were used at that time by station management for the
2906 fully irrigated yield trials to determine the yield
2907 potential for new CIMMYT materials. The upper line
2908 presents the yields for the same set of lines but
2909 following improved crop management practices that
2910 incorporated the use of deep chiseling to break up
2911 existing soil compaction zones, combined with the
2912 use of the Sesbania spp. summer green manure crop
2913 and chicken manure to try to rapidly enhance the
2914 extremely low level of soil organic matter content.
2915 This trial shows therefore the benefits of deep
2916 chiseling (about 50–70 cm chisel depth, with shanks
2917 50 cm apart) and of using organic matter (green and
2918 chicken manure).
2919 As can be seen in Fig. 5, almost as much progress
2920 in improving wheat yield potential—after 26 years of
2921crop breeding, ensued by improving crop manage-
2922ment as occurred by 26 years of breeding. In
2923addition, two other factors are apparent. Firstly, the
2924linear slopes were different (36 kg ha-1 year-1 for
2925normal trial management versus 52 kg ha-1 year-1
2926for the improved trial management) and the estimated
2927annual rate of yield increase was different (0.52%
2928year-1 for normal trial management versus
29290.68% year-1 for the improved trial management.
2930Secondly, one can observe the contrasting interacting
2931yield performance of the cultivars developed between
29321981 and 1987. For the normal trial management,
2933yields went down for these cultivars whereas for the
2934improved trial management they went up, which
2935could indicate the need for improving phenotyping
2936for yield potential and the appropriate crop manage-
2937ment practices for station field testing.
2938Wheat global partnership network builds national
2939capacity through human resources development
2940More than 40 years history of international wheat
2941nursery trials, underpinning the voluntary collabora-
2942tion of primarily public wheat improvement
2943institutions, has been accompanied in parallel by
2944intensive capacity-building efforts. More than 2000
Comparison of the yield performance of a historical set of cultivars for the
normal CIMMYT, irrigated yield trial management in Obregon in 1988 versus
agronomically improved trial management
5000
5500
6000
6500
7000
7500
8000
8500
9000
1962 1964 1967 1973 1976 1980 1982 1986
Year of Cultivar Release
Yie
ld (
kg
/ha
)
Normal CIMMYT Yield Trial Management in 1988
Agronomically Improved Yield Trial Management
Linear (Agronomically Improved Yield Trial Management)
Linear (Normal CIMMYT Yield Trial Management in 1988)
Fig. 5 Wheat grain yieldusing normal andagronomically improvedyield trial management(Obregon, Mexico, 1987–1988)
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF2945 scientists and research technicians from developing
2946 countries have participated in CIMMYT’s in long
2947 term hands-on courses organized in research stations
2948 in Mexico and a similar number of visiting scientists
2949 have come to CIMMYT to work shoulder to shoulder
2950 with CIMMYT scientists in the research and farmers
2951 fields, laboratories, and in the genebank (Table 12).
2952 Several hundreds of graduate students also had
2953 opportunity to conduct their thesis research with
2954 financial or technical support from CIMMYT. A
2955 number of short term special courses have been
2956 organized annually in collaborating countries partic-
2957 ularly since the 1980s.
2958 Recently however, there has been increasing con-
2959 cern globally about who will prepare the crop breeders
2960 of the twenty first Century as the number of public
2961 breeders, who produce new cultivars, is steadily
2962 declining. This pattern is not limited to the developed
2963 world, but has been reported from many developing
2964 countries as well (Guimaraes et al. 2006a, b, 2007).
2965 The genetic enhancement of wheat is not an
2966 exception, but maybe a more difficult case. Wheat
2967 breeding remains primarily in the domain of the
2968 public sector. Skilled wheat breeders in developing
2969 countries prepared in 1960s and 1970s are now either
2970 retiring or have been promoted to higher administra-
2971 tive positions in their institutions and are not actively
2972 in charge of active breeding programs. They are
2973 being replaced by scientists involved in more basic
2974 genetic studies, with less field experience. This shift
2975 is fueled by the perception that private-sector breed-
2976 ing efforts are adequate to meet cultivar needs. Also
2977 cuts in university resources have led to reduced
2978 support of field programs, and this has pushed the
2979 current public plant breeders to shift their activities
2980 toward fundamental or basic studies that can be
2981 supported by federal grants and the private sector.
2982 The loss of plant breeding programs is of great
2983 concern to both the USA plant breeding industry and
2984 the international community (Hancock 2006).
2985Furthermore, an effective and close partnership
2986with national researchers remains as one of the major
2987challenges to the successful mission of international
2988crop breeding. Their resources have been diminishing
2989in terms of human and infrastructural capital to
2990enable this to occur. However, some national partners
2991have strengthened their research capacity signifi-
2992cantly (e.g. Brazil, China, India, Mexico, South
2993Africa) over the past 40 years and their needs for
2994capacity building are less, and they work more in
2995equal partnership with the international breeding
2996programs. Hence, one of the overriding challenges for
2997many national partners will be to include in their
2998national institutional frameworks, investments in
2999research-for-development to ensure that the level of
3000science and resources are enough to provide basic
3001support and allow key outputs to be achieved. It is
3002clear that without sufficient national capacity the use
3003of international public goods produced in partnership
3004with international breeding programs, such as CI-
3005MMYT, will be restricted, and the mission therefore
3006compromised.
3007In order to develop the strategies to prevent further
3008erosion of institutional plant breeding training capac-
3009ity, several broadly attended meetings have been
3010conducted recently. Based on its broad scale study of
3011national capacities in plant breeding, the United
3012Nations of the Food and Agriculture Organization
3013(FAO) launched the Global Partnership Initiative for
3014Plant Breeding Capacity Building in 2006. In accor-
3015dance with its strategy and in order to address the
3016above described tendency, CIMMYT will continu-
3017ously support capacity-building of wheat scientists in
3018NARS through manifold ways—basic and advanced
3019courses on wheat improvement, visiting scientist
3020stays, and the support of degree thesis research.
3021Outlook: seeding innovations … nourishing hope
3022These are some of the issues that research managers
3023and wheat scientists must now confront in order to
3024select an optimal portfolio of strategic wheat genetic
3025enhancement research for the coming years, which
3026will have an impact on the ground during the coming
3027decades. Until the dramatic expansion of demand for
3028maize biofuels and the weather-induced supply
3029problems in the past few years, the prospects for a
3030reversal of the steady fall of the real prices of cereals
Table 12 Number of benefitting countries, long-term grouptrainees and visiting researchers on wheat improvement atCIMMYT
Region Countryof origin
Grouptrainees
Visitingresearchers
Developing world 85 995 1,391
Developed world 30 39 608
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
3031 including wheat appeared poor. Recent projections
3032 suggest a long-term increase in the real price of wheat
3033 along with other cereals (von Braun 2007). There are
3034 a number of trends and predicted key factors on
3035 which to base decisions: for example, the growing
3036 world population needs more food, more energy, and
3037 more feed grain to supply an ever increasing global
3038 demand for animal products; decreasing water sup-
3039 plies for agriculture and the effects of climate change
3040 are increasing the levels of abiotic stress across major
3041 wheat-producing areas; the application of biotech-
3042 nologies for better use of wheat genetic resources in
3043 the betterment of the crop is likely to offer new
3044 opportunities to increase yield, providing the private
3045 sector is also sufficiently engaged.
3046 As shown by this article, CIMMYT and research
3047 partners worldwide are conserving the wheat genetic
3048 endowment and improving crop yields and stability
3049 across the major cropping systems where wheat
3050 thrives. Likewise, this manuscript illustrates how
3051 such genetic resources are addressing new global
3052 challenges to both wheat production and demand,
3053 especially for the still growing population of the
3054 developing world. Surely, the wealth of genetic
3055 resources available in the wheat gene pools (includ-
3056 ing wild species) will be among the important sources
3057 available to plant breeders in their quest for high and
3058 stable yielding wheat cultivars that meet the end use
3059 quality demands at a time of limited resources and
3060 global warming.
3061 Acknowledgement The authors thank Ms. Allison Gillies3062 (CIMMYT, Mexico) for editing an early version of this3063 manuscript. We acknowledge the kind support of the CGIAR3064 members for wheat improvement at CIMMYT through3065 unrestricted funding and other resources brought by special3066 project grants in recent years from Australia, Belgium, Bill &3067 Melinda Gates Foundation, Canada, China, Denmark,3068 European Commission, FAO, FONTAGRO, Generation3069 Challenge Program, Germany, HarvestPlus, India, Iran, Italy,3070 Japan, Mexico, Norway, Republic of Korea, Rockefeller3071 Foundation, Sasakawa Africa Association, Spain, Sweden,3072 Turkey, United Kingdom, United States of America and the3073 World Bank, as well as the in-kind contributions of national3074 partners elsewhere.
3075 References
3076 Agcaoili-Sombilla MC, Rosegrant MW (1994) World supply3077 and demand projections for cereals, 2020. International3078 Food Policy Research Institute, Washington
3079Amani J, Fischer RA, Reynolds MP (1996) Canopy tempera-3080ture depression association with yield of irrigated spring3081wheat cultivars in a hot climate. J Agron Crop Sci 176:3082119–1293083Anderson JV, Morris CF (2003) Purification and analysis of3084wheat grain polyphenol oxidase (PPO) protein. Cereal3085Chem 80:135–1433086Anderson JA, Stack RW, Liu S, Waldron BL, Fjeld AD, Coyne3087C, Moreno-Sevilla B, Mitchell FJ, Song QJ, Cregan PB,3088Frohberg RC (2001) DNA markers for Fusarium head3089blight resistance QTLs in two wheat populations. Theor3090Appl Genet 102:1164–11683091Austin RB, Bingham J, Blackwell RD, Evans LT, Ford MA,3092Morgan CL, Taylor M (1980) Genetic improvement in3093winter wheat yields since 1900 and associated physio-3094logical changes. J Agric Sci (Cambridge) 94:675–6893095Ban T, Lewis JM, Phipps EE (eds) (2006) The global Fusarium3096initiative for international collaboration. CIMMYT,3097Mexico3098Barnabas B, Jager K, Feher A (2008) The effect of drought and3099heat stress on reproductive process in cereals. Plant Cell3100Environ 31:11–383101Bartlett PW (2000) New pests and diseases in European3102Community plant health legislation. In: The BCPC con-3103ference-pests and diseases, 2000 9B-4, pp 1159–11653104Bell MA, Fischer RA, Byerlee D, Sayre K (1995) Genetic and3105agronomic contributions to yield gains: a case study for3106wheat. Field Crops Res 44:55–653107Branlard G, Dardevet M, Saccomano R, Lagoutte F, Gourdon J3108(2001) Genetic diversity of wheat storage proteins and3109bread wheat quality. In: Bedo Z, Lang L (eds) Wheat in a3110global environment. Kluwer Academic Publishers,3111Dordrecht, pp 157–1693112Brennan JP, Warham EJ, Byerlee D, Hernandez-Estrada J3113(1992) Evaluating the economic impact of quality-reduc-3114ing, seed-borne diseases: lessons from Karnal bunt of3115wheat. Agric Econ 6:345–3523116Briney A, Wilson R, Potter RH, Barclay I, Crosbie G, Appels3117R, Jones MG (1998) A PCR marker for selection of starch3118and potential noodle quality in wheat. Mol Breed 4:427–31194333120Buerstmayr H, Lemmens M, Hartl L, Doldi L, Steiner B, Sti-3121erschneider M, Ruckenbauer P (2002) Molecular mapping3122of QTLs for Fusarium head blight resistance in spring3123wheat. I. Resistance to fungal spread (type II resistance).3124Theor Appl Genet 104:84–913125CABI (2005) Crop protection compendium, global module.3126CAB International, Wallingford3127Cakmak I, Ozkan H, Braun HJ, Welcj RM, Romheld V (2000)3128Zinc and iron concencetrations in seeds of wild, primitive3129and modern wheats. Food Nutr Bull 21:401–4033130Cane K, Spackman M, Eagles HA (2004) Puroindoline genes3131and their effects on grain quality traits in southern Aus-3132tralian wheat cultivars. Aust J Agric Res 55:89–953133CGIAR (1999) CGIAR Center statements on genetic resources,3134intellectual property rights, and biotechnology. May 1999.3135Center Directors and Center of Board Chairs of CGIAR,3136Washington3137Chen S, Ravallion M (2007) Absolute poverty measures for the3138developing world, 1984–2004. Proc Natl Acad Sci (USA)3139104:16757–16762
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
3140 CIMMYT (2004) Adding value for development: CIMMYT3141 annual report 2003–2004. CIMMYT, Mexico3142 CIMMYT (2007) Medium-term plan 2008–2010: translating3143 strategic vision to a vibrant work plan. CIMMYT, Mexico3144 Crosbie GB (1991) The relationship between starch swelling3145 properties, paste viscosity and boiled noodle quality in3146 wheat flours. J Cereal Sci 13:145–1503147 Crossa J, Burgueno J, Dreisigacker S, Vargas M, Herrera S,3148 Lillemo M, Singh RP, Trethowan R, Franco J, Warburton3149 M, Reynolds M, Crouch JH, Ortiz R (2007) Association3150 analysis of historical bread wheat germplasm using3151 additive genetic covariance of relatives and population3152 structure. Genetics 177:1889–19133153 De Groote H, Siambi M, Friesen D, Diallo A (2002) Identi-3154 fying farmers’ preferences for new maize varieties in3155 eastern Africa. In: Bellon MR, Reeves J (eds) Quantitative3156 analysis of data from participatory methods in plant3157 breeding. CIMMYT, Mexico, pp 82–1023158 De Saeger S, Kolosova AY, Sibanda L, Dumoulin F, Lewis J,3159 Duveiller E, Van Peteghem C (2008) Lateral-flow col-3160 loidal gold-based immunoassay for the rapid detection of3161 deoxynivalenol with two indicator ranges. In: Workshop3162 on rapid methods for food and feed safety and quality, 21–3163 23 January 2008, Noordwijkerhout, The Netherlands3164 Desjardins AE (2006) Fusarium mycotoxins: chemistry,3165 genetics and biology. APS Press, St Paul3166 Dixon J, Nally L, Aquino P, Kosina P, La Rovere R, Hellin J3167 (2006) Adoption and economic impact of improved wheat3168 varieties in developing countries. J Agric Sci (Cambridge)3169 144:489–5023170 Dreccer MF, Borgognone MG, Ogbonnaya FC, Trethowan3171 RM, Winter B (2007) CIMMYT-selected derived syn-3172 thetic bread wheats for rainfed environments: yield3173 evaluation in Mexico and Australia. Field Crops Res3174 100:218–2283175 Dubin HJ, Gilchrist L, Reeves J, McNab A (1996) Fusarium3176 head scab: global status and future prospects. CIMMYT,3177 Mexico3178 Dupont FM, Hurkman WJ, Vensel WH, Tanaka C, Kothari3179 KM, Chung OK, Altenbach SB (2006) Protein accumu-3180 lation and composition in wheat grains: effects of mineral3181 nutrients and high temperature. Eur J Agron 25:96–1073182 Duveiller E (2004) Controlling foliar blights of wheat in the3183 rice-wheat systems of Asia. Plant Dis 88:552–5563184 Duveiller E, Kaudel YR, Sharma RC, Shrestha SM (2005)3185 Epidemiology of foliar blights (spot blotch and tan spot)3186 of wheat in the plains bordering the Himalayas. Phyto-3187 pathology 95:248–2563188 Dvorak J (1998) Genomic analysis in the Triticum-Aegilops3189 alliance. In: Slinkard (ed) Proceedings of the 9th inter-3190 national wheat genetics symposium, viol 1. University3191 Extension Press, University of Saskatchewan, SK, pp 8–3192 113193 Dwivedi SL, Crouch JH, Mackill D, Xu Y, Blair MW, Ragot3194 M, Upadhyaya HD, Ortiz R (2007) Molecularization of3195 public sector plant breeding: a synthesis of progress and3196 problems. Adv Agron 95:163–3183197 Eagles HA, Hollamby GJ, Eastwood RF (2002) Genetic and3198 environmental variation for grain quality traits routinely3199 evaluated in southern Australian wheat breeding pro-3200 grams. Aust J Agric Res 53:1047–1057
3201Evenson RE, Gollin D (2003) Assessing the impact of the3202Green Revolution: 1960 to 2000. Science 300:758–7613203FAO (1983) Commission on plant genetic resources. Resolu-3204tion 8/83 of the 22nd Session of the FAO conference.3205Food and Agriculture Organization, Rome3206FAO (2002) The international treaty on plant genetic resources3207for food and agriculture. Food and Agriculture Organi-3208zation, Rome3209FAO (2007) Commission on genetic resources for food and3210agriculture. Food and Agriculture Organization, Rome.3211http://www.fao.org/ag/cgrfa/itpgr.htm (January 2007)3212Fischer RA, Rees D, Sayre KD, Lu Z-M, Condon AG, Larque3213Saavedra A (1998) Wheat yield progress associated with3214higher stomatal conductance and photosynthetic rate, and3215cooler canopies. Crop Sci 38:1467–14753216Flint-Garcia SA, Thornsberry JM, Buckler ES (2003) Structure3217of linkage disequilibrium in plants. Annu Rev Plant Biol321854:357–3743219Frankel OH (1977) Natural variation and its conservation. In:3220Muhammed A, von Botstel RC (eds) Genetic diversity of3221plants. Plenum Press, New York, pp 21–243222Geera BP, Nelson JE, Souza E, Huber KC (2006) Granule3223bound starch synthase I (GBSSI) gene effects related to3224soft wheat flour/starch characteristics and properties.3225Cereal Chem 83:544–5503226Gilchrist L, Dubin HJ (2002) Fusarium head blight. In: Curtis3227BC, Rajaram S, Gomez Macpherson H (eds) Bread wheat:3228improvement and production. Food and Agriculture3229Organization, Rome, Italy. Plant Prod Protect Ser 30:279–3230283, 285–2993231Gill B, Friebe S (2002) Cytogenetics, phylogeny and evolution3232of cultivated wheats. In: Curtis BC, Rajaram S, Gomez3233Macpherson H (eds) Bread wheat improvement and pro-3234duction. Food and Agriculture Organization, Rome. Plant3235Prod Protect Ser 30:71–883236Global Crop Diversity Trust (2007) Global strategy for the3237ex situ conservation with enhanced access to wheat, rye3238and triticale genetic resources. Global Crop Diversity3239Trust, Rome, http://www.croptrust.org/main/strategies.3240php?itemid=373241Graybosch RA, Peterson CJ, Hareland GA, Shelton DR,3242Olewnik MC, He H, Stearns MM (1999) Relationships3243between small-scale wheat quality assays and commercial3244test bakes. Cereal Chem 76:428–4333245Guimaraes EP, Bedoshvili D, Morgounov A, Baboev S, Iska-3246kov A, Muminjanov H, Kueneman E, Paganini M (2006a)3247Plant breeding and related biotechnology competence in3248Central Asia and recommendations to strengthen regional3249capacity. Agromeridian 2(3):137–1433250Guimaraes EP, Kueneman E, Carena MJ (2006b) Assessment3251of national plant breeding and biotechnology capacity in3252Africa and recommendations for future capacity building.3253HortScience 41:50–523254Guimaraes E, Kueneman E, Paganini M (2007) Assessment3255of the national plant breeding and associated biotech-3256nology capacity around the world. Crop Sci 47:S262–3257S2733258Hammer K (1980) Zur Taxonomie und Nomenklatur der3259Gattung Aegilops. Feddes Repert 91:227–2473260Hancock JF (2006) Introduction to the symposium: who will3261train plant breeders? HortScience 41:28–29
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
3262 Harlan JR, De Wet JMJ (1971) Toward a rational classification3263 of cultivated plants. Taxon 20:509–5173264 He ZH, Liu L, Xia XC, Liu JJ, Pena RJ (2005) Composition of3265 HMW and LMW glutenin subunits and their effects on3266 dough properties, pan bread, and noodle quality of Chi-3267 nese bread wheats. Cereal Chem 82:345–3503268 He XY, He ZH, Zhang LP, Sun DJ, Morris CF, Furerst EP, Xia3269 XC (2007) Allelic variation of polyphenol oxidase (PPO)3270 genes located on chromosomes 2A and 2D and develop-3271 ment of functional markers for the PPO genes in common3272 wheat. Theor Appl Genet 115:47–583273 He XY, Zhang YL, He ZH, Wu YP, Xiao YG, Ma CX, Xia XC3274 (2008) Characterization of a phytoene synthase 1 gene3275 (Psy1) located on common wheat chromosome 7A and3276 development of a functional marker. Theor Appl Genet3277 116:213–2213278 Hede A, Skovmand B, Reynolds MP, Crossa J, Vilhelmsen AL,3279 Stølen O (1999) Evaluating genetic diversity for heat3280 tolerance in Mexican wheat landraces. Genet Resour Crop3281 Evol 46:37–453282 Hodson DP, White JW (2007) Use of spatial analyses for3283 global characterization of wheat-based production sys-3284 tems. J Agric Sci (Cambridge) 145:115–1253285 Hoisington D, Ortiz R (2008) Research and field monitoring on3286 transgenic crops by the Centro Internacional de Mej-3287 oramiento de Maız y Trigo (CIMMYT). Euphytica. doi:3288 10.1007/s10681-007-9633-x3289 Inter-Governmental Panel on Climate Change (2001) IPCC3290 Third assessment report—climate change 2001. IPPC,3291 Geneva, http://www.ipcc.ch/3292 Jiang J, Friebe B, Gill BS (1994) Characterization of wheat-3293 alien translocations conferring resistance to diseases and3294 pests: current status. Euphytica 73:199–2123295 Jones DR (2007) Arguments for a low risk of establishment of3296 Karnal bunt disease of wheat in Europe. Eur J Plant Path3297 118:93–1043298 Joshi AK, Chand R, Arun B, Singh RP, Ortiz R (2007a)3299 Breeding crops for reduced-tillage management in the3300 intensive, rice-wheat systems of South Asia. Euphytica3301 153:135–1513302 Joshi AK, Mishra B, Chatrath R, Ortiz-Ferrara G, Singh RP3303 (2007b) Wheat improvement in India: present status,3304 emerging challenges and future prospects. Euphytica3305 157:431–4463306 Joshi AK, Ortiz-Ferrara G, Crossa J, Singh G, Alvarado G,3307 Bhatta MR, Duveiller E, Sharma RC, Pandit DB, Siddique3308 AB, Das SY, Sharma RN, Chand R (2007c) Associations3309 of environments in South Asia based on spot blotch dis-3310 ease of wheat caused by Cochliobolus sativus. Crop Sci3311 47:1071–10813312 Joshi AK, Ortiz-Ferrara G, Crossa J, Singh G, Sharma R, Chand3313 R, Prasad R (2007d) Combining superior agronomic per-3314 formance and terminalheat tolerance with resistance to spot3315 blotch (Bipolaris sorokiniana) in the warm humidGangetic3316 plains of south Asia. Field Crop Res 103:53–613317 Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki Y, Shino-3318 zaki K (1999) Improving plant drought, salt, and freezing3319 tolerance by gene transfer of a single stress-inducible3320 transcription factor. Nat Biotech 17:287–2913321 Kasuga M, Miura S, Shinozaki K, Yamaguchi-Shinozaki Y3322 (2004) A combination of the Arabidopsis DREB1A gene
3323and stress-inducible rd29A promoter improved drought-3324and low-temperature stress tolerance in tobacco by gene3325transfer. Plant Cell Physiol 45:346–3503326Kuchel H, Ye G, Fox R, Jefferies S (2005) Genetic and3327genomic analysis of a targeted marker-assisted wheat3328breeding strategy. Mol Breed 16:67–783329Kuchel H, Langridge P, Mosionek L, Williams K, Jefferies SP3330(2006) The genetic control of milling yield, dough rhe-3331ology and baking quality of wheat. Theor Appl Genet3332112:1487–14953333Knott DR (1982) Multigenic inheritance of stem rust resistance3334in wheat. Crop Sci 22:393–3993335Konarev VG, Gavriljuk IP, Gubareva NK, Peneva TI (1986)3336Proteins as genetic markers in solving the problems of3337applied botany, genetics and plant breeding. Bull Agric3338Sci 12:45–503339Lantican MA, Dubin MJ, Morris ML (2005) Impacts of3340International Wheat Breeding Research in the developing3341world, 1988–2002. CIMMYT, Mexico3342Leenhardt F, Lyan B, Rock E, Boussard A, Potus J, Chanliaud3343E, Ramsey C (2006) Genetic variability of carotenoid3344concentration, lipoxygenase and peroxidase activities3345among cultivated wheat species and bread wheat varieties.3346Eur J Agron 25:170–1763347Lewis J, Ban T, Ward R, Duveiller E (2007) Development of3348CIMMYT’s 11th scab resistance screening nursery. In:3349Proc. 2007 National Fusarium Head Blight Forum, Kansas3350City, Missouri, USA, 2–4 Dec. 2007, p 1923351Liu JJ, He ZH, Zhao ZD, Pena RJ, Rajaram S (2003) Wheat3352quality traits and quality parameters of cooked dry white3353Chinese noodles. Euphytica 131:147–1543354Liu L, He ZH, Yan J, Zhang Y, Pena RJ (2005) Allelic vari-3355ation at the Glu-1 and Glu-3 loci, presence of 1B/1R3356translocation, and their effect on mixgraphic properties in3357Chinese bread wheats. Euphytica 142:197–2043358Lynch M, Walsh B (1998) Genetics and analysis of quantita-3359tive traits. Sinauer Assoc, Sunderland3360Merezhko AF (1998) Impact of plant genetic resources on3361wheat breeding. In: Braun H-J, Altay F, Kronstad WE,3362Beniwal SPS (eds) Wheat: prospects for global3363improvement. Kluwer Academic Publishers, Dordrecht,3364pp 361–3693365Morgounov A, Roseeva L, Koishibayev M (2007) Leaf rust of3366spring wheat in Northern Kazakhstan and Siberia: inci-3367dence, virulence and breeding for resistance. Aust J Agric3368Res 58:847–8533369Mujeeb-Kazi A (1995) 15 Years of progress in wheat wide3370crosses at CIMMYT. In: Mujeeb-Kazi A, Hettel GP (eds)3371Utilizing wild grass biodiversity in wheat improvement:337215 years of wide cross research at CIMMYT. CIMMYT3373research report 2. CIMMYT, Mexico, pp 1–43374Mujeeb-Kazi A, Hettel GP (eds) (1995) Utilizing wild grass3375biodiversity in wheat improvement: 15 years of wide3376cross research at CIMMYT. CIMMYT research report 2.3377CIMMYT, Mexico3378Mujeeb-Kazi A, Rajaram S (2002) Transferring alien genes3379from related species and genera for wheat improvement.3380In: Curtis BC, Rajaram S, Gomez Macpherson H (eds)3381Bread wheat improvement and production. Food and3382Agriculture Organization, Rome, Plant Production and3383Protection Series 30:199–215
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
3384 Mujeeb-Kazi A, Wang RCC (1995) Perennial and annual3385 wheat relatives in the Triticeae. In: Mujeeb-Kazi A, Hettel3386 GP (eds) Utilizing wild grass biodiversity in wheat3387 improvement: 15 years of wide cross research at CI-3388 MMYT. CIMMYT research report 2. CIMMYT, Mexico,3389 pp 5–133390 Mujeeb-Kazi A, Cano S, Rosas V, Cortes A, Delgado R3391 (2001a) Registration of five synthetic hexaploid wheat and3392 seven bread wheat lines resistant to wheat spot blotch.3393 Crop Sci 41:1653–16543394 Mujeeb-Kazi A, Fuentes-Davila G, Villareal RL, Cortes A,3395 Rosas V, Delgado R (2001b) Registration of 10 synthetic3396 hexaploid wheat and six bread wheat germplasms resistant3397 to Karnal bunt. Crop Sci 41:1652–16533398 Murray GM, Brennan JP (1998) The risk to Australia from3399 Tilletia indica, the cause of Karnal bunt of wheat. Aus3400 Plant Path 27:212–2253401 Nagarajan S (2005) Can India produce enough wheat even by3402 2020? Curr Sci 89:1467–14713403 Nicholson P, Gosman N, Draeger R, Thomsett M, Chandler E,3404 Steed A (2007) The Fusarium head blight pathosystem:3405 status and knowledge of its components. In: Buck HT,3406 Nisi JE, Salomon N (eds) Wheat production in stressed3407 environments. Springer, Dordrecht, pp 23–363408 Ogbonnaya FC, Ye G, Trethowan R, Dreccer F, Lush D,3409 Shepperd J, van Ginkel M (2007) Yield of synthetic3410 backcross-derived lines in rainfed environments of Aus-3411 tralia. Euphytica 157:321–3363412 Ortiz R, Iwanaga M, Reynolds MP, Wu H, Crouch JH (2007a)3413 Overview on crop genetic engineering for drought-prone3414 environments. J Semi-Arid Trop Agric Res 4,3415 http://www.icrisat.org/journal/SpecialProject/sp3.pdf3416 Ortiz R, Mowbray D, Dowswell C, Rajaram S (2007b) Norman3417 E. Borlaug: The humanitarian plant scientist who changed3418 the world. Plant Breed Rev 28:1–373419 Ortiz R, Trethowan R, Ortiz Ferrara G, Iwanaga I, Dodds JH,3420 Crouch JH, Crossa J, Braun H-J (2007c) High yield3421 potential, shuttle breeding and new international wheat3422 improvement strategy. Euphytica 157:365–3843423 Ortiz R, Ban T, Bandyopadhyay R, Banziger M, Bergvinson D,3424 Hell K, James J, Jeffers D, Lava Kumar P, Menkir A,3425 Murakami J, Nigam SN, Upadhyaya HD, Waliyar F3426 (2008a) CGIAR research-for-development program on3427 mycotoxins. In: Leslie JF, Bandyopadhyay R, Visconti A3428 (eds) Mycotoxins: detection methods, management, pub-3429 lic health and agricultural trade. CABI Publishing,3430 Wallingford, pp 415–4243431 Ortiz R, Sayre KD, Govaerts B, Gupta R, Subbarao GV, Ban T,3432 Hodson D, Dixon JM, Ortiz-Monasterio JI, Reynolds M3433 (2008b) Climate change: can wheat beat the heat? Agr3434 Ecosyst Environ. doi:10.1016/j.agee.2008.01.0193435 Ortiz Ferrara G, Bhatta MR, Pokharel TP, Mudwari A, Thapa3436 DB, Joshi AK, Chand R, Muhammad D, Duveiller E,3437 Rajaram S (2001) Farmer participatory variety selection3438 in South Asia. In: Research highlights of the CIMMYT3439 wheat program, 1999–2000. CIMMYT, Mexico, pp 33–373440 Ortiz-Ferrara G, Joshi AK, Chand R, Bhatta MR, Mudwari A,3441 Thapa DB, Sufian MA, Saikia TP, Chatrath R, Witcombe3442 JR, Virk DS, Sharma RC (2007) Partnering with farmers3443 to accelerate adoption of new technologies in South Asia3444 to improve wheat productivity. Euphytica 157:399–407
3445Ortiz-Monasterio I, Palacios-Rojas N, Meng E, Pixley K,3446Trethowan R, Pena RJ (2007) Enhancing the mineral and3447vitamin content of wheat and maize through plant3448breeding. J Cereal Sci 46:293–3073449Pena RJ, Pfeiffer WH (2005) Breeding methodologies and3450strategies for durum wheat quality improvement. In: Royo3451C, Nachit MN, Di Fonzo N, Araus JL, Pfeiffer WH, Slafer3452GA (eds) Durum wheat breeding: current approaches and3453future strategies. Food Product Press–The Haworth Press3454Inc., New York, pp 663–7723455Pena RJ, Trethowan RM, Pfeiffer WH, van Ginkel M (2002)3456Quality (end-use) improvement in wheat. Compositional,3457genetic, and environmental factors. In: Basra AS, Randh-3458awa LS (eds) Quality improvement in field crops. Food3459Product Press–The Haworth Press Inc., New York, pp 1–373460Pfeiffer WH, Trethowan RM, van Ginkel M, Ortiz-Monasterio3461I, Rajaram S (2005) Breeding for abiotic stress tolerance3462in wheat. In: Ashraf M, Harris PJC (eds) Abiotic stresses:3463plant resistance through breeding and molecular approa-3464ches. The Haworth Press Inc., New York, pp 401–4893465Podlich DW, Cooper M (1998) QU-GENE: a platform for3466quantitative analysis of genetic models. Bioinformatics346714:632–6533468Pritchard JK, Stephen M, Donnely P (2000) Inference on3469population structure using multilocus genotype data.3470Genetics 155:945–9593471Rajaram S (1999) Historical aspects and future challenges of3472an international wheat program. In: van Ginkel M, McNab3473A, Krupinsky J (eds) Septoria and Stagonospora diseases3474of cereals: a compilation of global research. CIMMYT,3475Mexico, pp 1–173476Rajaram S, van Ginkel M, Fischer RA (1994) CIMMYT’s3477wheat breeding mega-environments (ME). In: Proceed-3478ings of the 8th international wheat genetics symposium.3479China Agricultural Scientech, Beijing, pp 1101–11063480Reynolds MP, Borlaug NE (2006) International collaborative3481wheat improvement: impacts and future prospects. J Agric3482Sci (Cambridge) 144:3–173483Reynolds MP, Acevedo E, Sayre KD, Fischer RA (1994a) Yield3484potential in modem wheat varieties: its association with3485aless competitive ideotype. Field Crops Res 37:149–1603486Reynolds MP, Balota M, Delgado MIB, Amani J, Fischer RA3487(1994b) Physiological and morphological traits associated3488with spring wheat yield under hot, irrigated conditions.3489Aus J Plant Physiol 21:717–7303490Reynolds MP, Singh RP, Ibrahim A, Ageeb OA, Larque-Sa-3491avedra A, Quick JS (1998) Evaluating physiological traits3492to compliment empirical selection for wheat in warm3493environments. Euphytica 100:85–943494Reynolds MP, Sayre KD, Rajaram S (1999) Physiological and3495genetic changes in irrigated wheat in the post Green3496Revolution period and approaches for meeting projected3497global demand. Crop Sci 39:1611–16213498Reynolds MP, Calderini DF, Condon AG, Rajaram S (2001)3499Physiological basis of yield gains in wheat associated with3500the Lr19 translocation from Agropyron elongatum. In:3501Bedo Z, Lang L (eds) Wheat in a global environment.3502Kluwer Academic Publishers, Netherlands, pp 345–3513503Reynolds MP, Pellegrineschi A, Skovmand B (2005) Sink-3504limitation to yield and biomass: a summary of some3505investigations in spring wheat. Ann Appl Biol 146:39–49
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
3506 Reynolds MP, Dreccer F, Trethowan R (2007) Drought adap-3507 tive traits derived from wheat wild relatives and landraces.3508 J Exp Bot 58:177–1863509 Ross AS, Quail KJ, Crosbie GB (1996) An insight into struc-3510 tural features leading to desirable alkaline noodle texture.3511 In: Wrigley CW (ed) Cereals ’96. Royal Australian3512 Chemical Institute, Australia, pp 115–1193513 Rush CM, Stein JM, Bowden RL, Riemenschneider R, Bora-3514 tyniski T, Royer MH (2005) Status of Karnal bunt of3515 wheat in the United States 1996 to 2004. Plant Dis3516 89:212–2233517 Sayre KD, Rajaram S, Fischer RA (1997) Yield potential3518 progress in short bread wheats in northwest Mexico. Crop3519 Sci 37:36–423520 Sayre KD, Singh RP, Huerta-Espino J, Rajaram S (1998)3521 Genetic progress in reducing losses to leaf rust in CI-3522 MMYT-Derived Mexican spring wheat cultivars. Crop3523 Sci 38:654–6593524 Sharma RC, Duveiller E, Gyawali S, Shrestha SM, Chaudhary3525 NK, Bhatta MR (2004) Resistance to Helminthosporium3526 leaf blight and agronomic performance of spring wheat3527 genotypes of diverse origins. Euphytica 139:33–443528 Sharma RC, Duveiller E, Ortiz-Ferrara G (2007) Progress and3529 challenge towards reducing wheat spot blotch threat in the3530 Eastern Gangetic Plains of South Asia: is climate change3531 already taking its toll? Field Crops Res 103:109–1183532 Singh RP, Huerta-Espino J (2004) The use of ‘single-back-3533 cross, selected-bulk’ breeding approach for transferring3534 minor genes based rust resistance into adapted cultivars.3535 In: Black CK, Panozzo JF, Rebetzke GJ (eds) Proceedings3536 of the 54th Australian cereal chemistry conference and3537 11th wheat breeders assembly, Canberra, Australia, 21–243538 September 2004. Publishers Cereal Chemistry Division,3539 Royal Australian Chemical Institute, Victoria, Australia,3540 pp 48–513541 Singh RP, Huerta-Espino J, Rajaram S, Crossa J (1998a) Agro-3542 nomic effects from chromosome translocations 7DL.7Ag3543 and 1BL.1RS in spring wheat. Crop Sci 38:27–333544 Singh RP, Rajaram S, Miranda A, Huerta-Espino J, Autrique E3545 (1998b) Comparison of two crossing and four selection3546 schemes for yield, yield traits, and slow rusting resistance3547 to leaf rust in wheat. Euphytica 100:35–433548 Singh RP, Huerta-Espino J, Rajaram S (2000) Achieving near-3549 immunity to leaf and stripe rusts in wheat by combining3550 slow rusting resistance genes. Acta Phytopathlogica3551 Hungarica 35:133–1393552 Singh RP, William HM, Huerta-Espino J, Rosewarne G (2004)3553 Wheat rust in Asia: meeting the challenges with old and3554 new technologies. In: New directions for a diverse planet:3555 proceedings of the 4th international crop science congress,3556 Brisbane, Australia, 26 September–1 October 2004,3557 http://www.cropscience.org.au./icsc2004/symposia/3/7/3558 141_singhrp.htm (accessed on November 29, 2007)3559 Singh RP, Hodson DP, Jin Y, Huerta-Espino J, Kinyua M,3560 Wanyera R, Njau P, Ward RW (2006) Current status,3561 likely migration and strategies to mitigate the threat to3562 wheat production from race Ug99 (TTKS) of stem rust3563 pathogen. CAB reviews: perspectives in agriculture, vet-3564 erinary science. Nutr Nat Resour 1:543565 Singh RP, Huerta-Espino J, Sharma R, Joshi AK, Trethowan R3566 (2007) High yielding spring wheat germplasm for global
3567irrigated and rainfed production systems. Euphytica 157:3568351–3633569Smartt J (1984) Gene pools in grain legumes. Econ Bot 38:24–3570353571Snapp S (1999) Mother and baby trials: a novel trial design3572being tried out in Malawi. TARGET Jan. 1999. CI-3573MMYT, Harare3574Souza EJ, Graybosch RA, Guttieri MJ (2002) Breeding wheat3575for improved milling and baking quality. In: Basra AS,3576Randhawa LS (eds) Quality improvement in field crops.3577Food Product Press–The Haworth Press Inc., New York,3578pp 39–743579Spiertz JHJ, Hamer RJ, Xu H, Primo-Martin C, Don C, van der3580Putten PEL (2006) Heat stress in wheat (Triticum aes-
3581tivum L.): effects on grain growth and quality traits. Eur J3582Agron 25:89–953583Thornsberry JM, Goodman MM, Doebley J, Kresovich S,3584Nielson D et al (2001) Dwarf8 polymorphisms associate3585with variation in flowering time. Nat Genet 28:286–2893586Trethowan RM, Crossa J, van Ginkel M, Rajaram S (2001)3587Relationships among bread wheat international yield3588testing locations in dry areas. Crop Sci 41:1461–14693589Trethowan RM, Van Ginkel M, Rajaram S (2002) Progress in3590breeding for yield and adaptation in global drought3591affected environments. Crop Sci 42:1441–14463592van Ginkel M, Trethowan R, Ammar K, Wang J, Lillemo M3593(2002) Guide to bread wheat breeding at CIMMYT.3594Wheat special report 5. CIMMYT, Mexico3595van Slageren MW (1994) Wild wheats: a monograph of Ae-3596gilops L. and Amblyopyrum (Jaub. and Spach) Eig3597(Poaceae). Wageningen Agricultural University Agricul-3598tural Papers, Wageningen, pp 331–3333599Villarreal RL, Banuelos O, Mujeeb-Kazi A (1997) Agronomic3600performance of related durum wheat (Triticum turgidum
3601L.) stocks possessing the chromosome substitution3602T1BL.1RS. Crop Sci 37:1735–17403603Von Bothmer R, Seberg O, Jacobsen N (1992) Genetic3604resources in the Triticeae. Hereditas 116:141–1503605Von Braun J (2007) The world food situation: new driving3606forces and required actions. International Food Policy3607Research Institute, Washington3608Wang J, van Ginkel M, Podlich D, Ye G, Trethowan R, Pfeiffer3609W, DeLacy IH, Cooper M, Rajaram S (2003) Comparison3610of two breeding strategies by computer simulation. Crop3611Sci 43:1764–17733612Wang J, van Ginkel M, Trethowan R, Ye G, DeLacy I, Podlich3613D, Cooper M (2004) Simulating the effects of dominance3614and epistasis on selection response in the CIMMYT wheat3615breeding program using QuCim. Crop Sci 44:2006–20183616Wang J, Eagles HA, Trethowan R, van Ginkel M (2005) Using3617computer simulation of the selection process and known3618gene information to assist in parental selection in wheat3619quality breeding. Aust J Agric Res 56:465–4733620Wang J, Wan X, Li H, Pfeiffer W, Crouch J, Wan J (2007a)3621Application of the identified QTL-marker associations in3622rice quality improvement through a design breeding3623approach. Theor Appl Genet 115:87–1003624Wang J, Chapman SC, Bonnett DB, Rebetzke GJ, Crouch J3625(2007b) Application of population genetic theory and3626simulation models to efficiently pyramid multiple genes3627via marker-assisted selection. Crop Sci 47:580–588
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPR
OOF
UNCORRECTEDPR
OOF
3628 Warburton ML, Crossa J, Franco J, Kazi M, Trethowan R,3629 Rajaram S, Pfeiffer W, Zhang P, Dreisigacker S, van3630 Ginkel M (2006) Bringing wild relatives back into the3631 family: recovering genetic diversity of CIMMYT bread3632 wheat germplasm. Euphytica 149:289–3013633 Witcombe JR, Joshi A, Joshi KD, Sthapit BR (1996) Farmer3634 participatory crop improvement. I. Varietal selection and3635 breeding methods and their impact on biodiversity. Exp3636 Agric 32:445–4603637 Witcombe JR, Joshi KD, Rana RB, Virk DS (2001) Increasing3638 genetic diversity by participatory varietal selection in high3639 potential production systems in Nepal and India. Euphy-3640 tica 122:575–5883641 Witcombe JR, Joshi KD, Gyawali S, Musa A, Johansen C, Virk3642 DS, Sthapit BR (2005) Participatory plant breeding is3643 better described as highly client-oriented plant breeding. I.
3644Four indicators of client-orientation in plant breeding. Exp3645Agric 41:1–213646Ye G, Moody D, Emebiri L, van Ginkel M (2007) Designing3647an optimal marker-based pedigree selection strategy for3648parent building in barley in the presence of repulsion3649linkage, using computer simulation. Aust J Agric Res365058:243–2513651Yu J, Pressoir G, Briggs WH, Bi IV, Yamasaki M et al (2006)3652A unified mixed-model method for association mapping3653that accounts for multiple levels for relatedness. Nat3654Genet 38:203–2083655Zhang XK, Liu L, He ZH, Sun DJ, He XY, Xu ZH, Zhang PP,3656Chen F, Xia XC (2008) Development of two multiplex3657PCR assays targeting improvement of bread-making and3658noodle qualities in common wheat. Plant Breed. doi:365910.1111/j.1439-0523.2007.01442
3660
Genet Resour Crop Evol
123
Journal : Medium 10722 Dispatch : 29-8-2008 Pages : 46
Article No. : 9372 h LE h TYPESET
MS Code : GRES951 h CP h DISK4 4
Au
tho
r P
ro
of




















