
W5¥3H - OSTI.GOV
169
: h^esw%> W5¥3H MHffiA v 9 —
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of W5¥3H - OSTI.GOV

: h^esw%>
W5¥3H
MHffiA v 9 —

ii t A
A'7/Hs8rj8W)E$iei: & f mmKr n.
W®cV>0*@8rE^roS16*tt. #lVMSH&SAtoft±t*ti££x.Z>m¥mtg0)M&T$>->fc.
t LT4-. m£&Mcowim!fok izvmm. tt^EMtcffdx^to
ZtitzfctX, 4$SI-
CkiiggTit^V'.Sia-tS^fciOSrfc^rJftSreiSfckfcfc, ^<0$S$kL
tWtl^Lftroyx^fSgSL, Wff$-e$>5 -5 . ■tlXtvmmizJ:
9iE^®«tAnrt±B*»D/<7yxci)E#ut%Sh-etL(:J:^r<>itr^ SiiS3E5IL7tti^695eSK S4-ri> i fc C0#»T'?)I. i; ® tT .
IMfflTtt, mcSM®SS@<7)®®l: <fc 9 rssiti«t-j3(t?,jtFlSES%T-'7<7)%gp$j S5IEL, *05f6g»&¥E4^3Mi=tkib, 34f-V^*gf-7i ttS£L*. fi t> iliSixii it •£B9-C'*5®^k%tbti1Z<r>-tfi Xh~>tz.
-k<7)J;t$r«SE«T, i«ll@llg»i • ti«ia«8flEEiSr □ y i ? h !: LX r%#x
4 *#S»)SJEiniS8i<OE^M%j fr'EOAtf £>it. SStBi4¥E4¥l 2^170. Stx*;t^' - • ozt£%-?*:> $toyi? Mtx
f-7®3y-fethfc^-SctSWASr^-f, X^K-. BHtcoSEfciS■fhhnxhh. tcoBE<7)]$Efct4,WZhhiiK A'4*S»k-ecomi$Sffi<7)l/<;UOiE>->j£Ei6«fct5V^Ttt|9iS%-tS<Ofca>§*)L
^T-vf&O. *v>^5ES»$to#5tL5„i«ofgs*t4, ¥E4^® ■ g #-->?& & riss
P$E%J k LTfi-3/iA'4*M<7)K#enii]fcro't"65:EM3ES:ik»,)71t,i7)-ei m#(4^7f K fconT^EffSfficoEEkKtoto^^rt-b'^UXh^^E-^TV^.
s:*s^#><oE%siii<7)*MKi6iitT, 4i^7d-;i^ s#m^m###+k^tu4$^r&&.
ll). 8.-7
StfflSA AUmtSE-fey*- £: ft fi t @ I

B
If g :
x¥<7)~sjm& ...........................................................................sjn i 1
SIS : ^X-fF^flljiKfh
* v dxxf- ht.................................3
s 2 S : ^Xf - K <9 •&)£&#
ga«<9X^£j£ 1 3
Total Synthesis of Nephritogenic Glycopeptide, Nephritogenoside
■ • II 2 2
TOTT-tfC j: & y^Xf • • • S @81. »jkS9! 26
S3S :
Stereoselective Routes toward the Synthesis of Unusual Amino Acids
■ • ■ Xf)8S® 3 5
i&3t^«§«<9£)£k7r4 XXS*/t-XA.ofiJS
• • • «JH X, B# S 4 2
#%#% a yo ax ...................................^fss 51
Chemistry of Lantibiotics .................................................$ 5 9
S4S: Kmmmam
FAB*#»#(9%l#k^Xf-M • $a«-<k1*iiS#f^<9KJ6
• • •BUS*, TBSW 8 5$«ga*<9xxTy ■ .........................##*% 9 o
9 7
S 5 S : ^.Xf F (0
±»K*

6 *: f - h t mm%m
......................................................................_h® » 10 9
A low-cost,high-efficiency solar cell based on dye-sensitized
colloidal Ti02 films • ■ ■ B.0’Regan & M.Gratzel 12 0
Development of Specific Immunoadsorbent Containing Immobilized Synthetic
Peptide of Acetylcholine Receptor for Treatment of Myasthenia Gravis
■ ■ ■ 12 3
im: JHEEE. mm&tnm 121


Lecture^x‘7> KX^dWiEti
Jll
1. liCSblc
(1, 21#E©##R#^>BV(j: r#f{b#J (D%6LTm f#$tlTl')'5o .^B tc, jrf-jrik 1 Tj^eM.tc bti B <t , #
fE£lS:St L C tl££-E L T V < Ci(t $jlg©£-E&1W ^oLT&gsm&Br^cL/:#
7#mKm©^#%*^)6b^Bo igtc#o6 Wk#J (i, 7©E57FiES®t?tc7b-y x/1/-t5<:£&btiT
U66i^=k9o 7CT, C© =k 3^#:#(c^A#B#r
Lv##m^7#m©%6LT r<77KX#J ^B
Etc, *y&%xmi, ^©I6Iiolf{k-i
#LTU6o u^L, 7©M^ti,
<77FX#^C©^>8%X#^bmmLT. ,t%hot Lxmn^tiho zamm
&&&&=?, mmmm
il'ofc, «fc?)I^/$:;£@^©gF^^lT'**K <b
MftZtlSo9 >em%^6<77 Kn:^©i6 A©is l '
Am©2o#^©7; /^m^A^-tftcm^fia^fc^ L, ; /0&5Wa#7 ; y ^Synthon^<-x6 LT£&£EM©#=£C B#<bLT&(), t;LB##%4fI#©-$m6LT &g77bYiBA(c^Bo
<77 %#^tc^m L j; 9 6^(1%£r>tzlZfr'O’Ob'O^ *(DnJM&lZU%%a<DM£fii&Vo Lt^L, Ki^<-xt;tl>iiv^t1iy > 6MX¥<Di£ffi&JWln£Jc(Dtt LTv< c6*<pi^T&b). emm^mm4#©±^©± tc,B0
2. <7°7 KX^O#gfc1) 7 ; yiSil'9 ^ St4 © rSi V synthon £fij B T" S
Bo7 : y E^EtcHti L T tiEtcM^'E2: LTfc' D,
Lvm#g^#c7 ; y#ef ^#t^BC6(Cj: *9, <77H7}7#:(cim^^^m©#A^EIBC6:
7^'T§ Bo o^t9> 7 ; y S££fcB§^JiON-(synthon) tLT. ^/c, itSfX
LT. #A^7 ; y 0&^&<77 K©$B5§;7 b©%$1 b"BtEt ?l t9oo&Bo
ttliiiTdBoA#t©#8(i20#M©7 ^ y#tc =k(9#6%$^iBo
C n^lOHSo/j: v/CYd't j-eio1
&Bo M7; yi^ffli'til^ btc^StttijA^ Bo V^cef^tcmL/:efT^B^<m#$fl
Bo3) <77 K^A^v-p#—#A#^tcJ;^#E$fiB
C6J:0x #itBE%[/AE;l)-h©m-
M#T&Bo$o —~C <£j B C. B t)' b0:!&©flfill;£:-tp-—©
#&(CZ *5BE L?#B^Jtltt»1^ti7c0 WS£E{b
#mtc C O j; 7 Af&f B-#&©& Bjl- htint: <, 7©®^b t>^$nBSiii'x«k
7o7f h V >/?7^ ©SM±fC^*i
^<77K^BEL, 7©####^^^89(c$tmf B^WFodorbtCefcoT^$nTVB1)o $/:. it &?Z#*m'BC6(c^T##T^#%<77 K 0 7^ 75 'J —©BEt^'B^ET’^) B0 7 7 — v©itfx;
10'-'©#^c/c##©<77 K^#BC6A<W#E6
4) <77 KI%©#A\ cbB Vti 2 o©<77 K^©tc B o
<77 K^#E7B7 ; KMAti, ^S •6#%%##-Bo %r,T, <77 K (iBE#©^#^#T&BC(!:^b.##6 LT##fBC6 t^T^Bo
5) ^^fSa^^fiE L-HVo<77 K(t ^7^7]tcj: g E#A##^#-Oo
C<D£otlft&&%mttUf—o©<77 K5>7tc7
<T©##&##&-&B©T(i%<, v<o^\©^7 (c^mrLy:##^mALT^f/=^##^#$-(YBC
<h^#&6^Bo C©«k97C#^(i, AX^vl/#7 7©#m6 l)#AB 4)©T^Bo
6) Bo
- 1 -

cfii$^mm$(DQrmmm%%mtz>±rc\mi6xn.
■So G tl b {iJtzI 15spatial arrange mentornJtl <h
faboT&ao^mT^a^^c^-o/:^ iaef%#=(i/<y^FA5%<Z)^3—c(D^#/j:v-;l/T$,6o coj:9%##
3 6 u 92flTU5o Tirrell(j:C(Z)j:9^#^0m(:#Ll^
L l' a") oCappello
f KM^otl'55)c co«k9Cx Mfi©itoj£ Kic ji-DTmmL j:9 a^ti
3. ^7°5F KOfiJffl^S
c L c «k *9.A<&ao
2 ) ###%##(!: LTO#mLT^ (9.
C a C 6 C j: «9a CL <h ^s'"5JtETab ao t. tzHttSOoS^^spacial a rrangementC X *9 ^ < 0)L#8^##^^#T # j: 9
3) LT<mm
c$i# Lr-^a c i^nrtiT*a0 #mau)
4) LTof#/<yf LT^m^f^
eg#^9C6^pI#T&ao C<Dj:9%mE#/W LTt^jmpj#T&ao
5 ) £#W41 lT(D%m/<KCa##Tj5aC6x
&#c# LT#fl/:m%#^-#@$-t±/= *9 x £.ft<DWtL ^ f a C 6 *<T # a o
teRT#!tt£$&Tt'aa<. unt LrtiMT&aG
6. a c 6 ef^iitfe5Ci^b> i:/d1;/d9f3£<h LTLiifiSSC
^u/:6caT&ao -eom^o/zAc^t LTfiJffl LTV < ±T(DESM?©to35L^ill a Ztfr^Xt'£>6o
4. s<-?3- KX^OEiUM:/<Tf K%#(D##R#6 LT(i
1)2) #^7 ; V3)
4) KoA^cK#s)6)T)8) ^TfFc^a^mm^mm^K#9) Kc=wa^?##m10)
0##^±(f^fiao LLmmL/:i)(DT(i^v AL 4%C0^&mii[LTV<±T^;RT^^I)(D(d:^
*9T£>(K SSA'IX*9SWS£iiao #l\ B*T(i
W6LTW^cLTV< C^LW^T^a.
1) S.P. A. Fodor.J. Leighton Tead,M.C.Pirrung,L. Stryer,A.T.Lu,D.Solas,Science,251, 767 (1991)
2) S.E.Cwirla.E. A. Peters, R.W. Barrett, and W.J. Dower,Proc.Natl.Acad.Sci.,87, 6378 (1990)
3) J.J.Devlin,L.C.Panganiban.P.E.Devlin,Scien- ce,249, 404 (1990)
4) J.K.Scott and G.P.Smith,Science,249, 386 (1990)
5) J.Cappello and J.VV.Crissman,Polym.Prepr.,30 (1), (1990)
6) K.P.McGrath,D.A.Tirrell,Polym.Prepr.,31(1), 190 (1990)
m&sfm, P 1 - 3, J:
D)
- 2 -

% 1 ^ : KJ? SffllSillSft

* V d>\°77 KOE*14<Dm/r£7i'Ji ft t «*-i h »
Quantitative Analysis and Prediction of Hydrophobicity of Oligopeptides
Miki Akamatsu* and Toshio Fujita*
The log P value used for the hydrophobicity parameter, where P is the partition coefficient in a 1-octanol/pH 7.0 buffer system, of a number of di—pentapeptides was analyzed with physicochemical parameters for the side chain substituent of component amino acids. The log P value was shown to be governed not only by the “intrinsic” hydrophobicity but also by the steric effect of side chain substituents as well as intramolecular-type solvation and “polar proximity" effects for polar side chains. The £-turn conformational parameters of compo
nent amino acids devised from the Chou-Fasman propensity index were nicely applied for the analysis of log P value of tetra- and pentapeptides. Using the results of the correlation analyses for log P of oligopeptides, we proposed a new effective hydrophobicity scale. na, for unionizable amino acid side chains. This scale and the hydrophobicity of oligopeptides would be useful not only for the process of peptide synthesis but also for the
quantitative analysis of the structure-activity relationship of bioactive peptides.Key words : Oligopeptides ; Hydrophobicity ; Parti
1. GUAM:
mao## f v >2>, am####*
RGD ^77 K3’& V
m Li'EE, mmogijBWK*7?- ea c&t o, ^-/7 Korn*
it 2
t-r 1 mef-mmmczX Z> -g-E & 77
$-762 «kl:Z i)^77 7--tf^7n7 7--tf06aTk55‘mnmjzfctmm l-c^77
* (7606 stiff
* Faculty of Agriculture. Kyoto University (Kitashira- kawa Oiwake-cho, Sakyo-ku, Kyoto 606, Japan)
coefficient; Peptide synthesis ; Drug design.
mmizxz^&xit, !>'>*')>, j&mxji'Xszv*?9 78§®77o
$ -76 2 h (£,^ N ^5®till,
ilS 20-30 n&fr ^^6^777^77 L,
^77 Foam#### 2 Mb#
A'7?s#jtcr^iiLTv^0 -#c, m76EUtifl*Otfc£-!$££-j&U
<^#077 < >< giWt76 2 a
$>hio mwmiz£76 ^tlteli&xhh 2 17VMM $ ti, Hansch *£8\
- 3 -

( 41 ) 837
Free-Wilson </>y $ tifijfflgtvCS/Jo ^yy mn^&swbs-^-eiz, 2<7)Z 3fc£iMyy7WjftoT«jgig1±ttM#$r£fTV\ y
> L.tz&&, nmt$> z^imm Lfzmm • &&<%#%#####m#L-C#TV^6o flfcLT,'/V >m##MY-5561"», K,,,*fc’^tftbti^»o tkfcZJz&^ztLXfrbftm&xmm'f
^yy^yy K<7)5H-£te*5Ztr
ffiOSM^Bbkttysmy*^±(7)2^;)^, ^yy zc/f(omm&omkts
tffi'&tcr>r%%*&%TL, #my6mmij$r#< C«bli z/j^yy y<dm-&.3&*, ffifcr
< / t migm:&v'T##f & - 2: iz,^>s*?n<n-$:ffim<D:?m*>, ^yy r^ztr^>/<* % t tkfcm t <nfcs.im<vmm c# lt<>T*6o /<yy K^ya#<7)^*1±t:cv'
CKliZay^ <^yy K^2#^±(7)##me# l-c^ih * >^m-cOTEl, L7)'t
* - * <OS^BET% o/jt5^ xhhOo 8&, a*iz^>?/<yy k z T-^jt^fitjESMo^yy ttah l-*9 9 J - Jl'/7k%'C<Dfr8ii&8c P <W& logp y. yotmT < /
MI]iE:S< ^<h£tzn~u)<r>X-, foigco^tcitc^LZzv^Sdo
2. ^vdx-yy k<7)-£/&
^yyKfcztrycmg&zznzsffl&yfflV'TDCC/HOBt&15», N-L Kn*->y?'>W $ K,6) ^ p-r. h □ 7 jl y -;v17,a:<7)f^t4xx-r;vE, iE-g-ge
^(0 * -/ y V > y&C Z 0 £-/& $ tLZ:0 £- j&nmz, ^yy ke<t)J£5^t $ v&bb?ijuik?flt y# <mfkLZ:(7)T, 86A##&4Aa#8= &&#L/Jo fSSMZiltB HPLC H Z. t) *To /Jo
3. log P (7)#%r:&&43<Z[f/xyy K IZOuxTCDSO^iS
ik^#(7) iogp i)-em$K e&sx (7)j^tR14£& * t LT^SI£*i£19,o CCTlog Pn IZ&#% ®<7)#"C&&o
zr=log Px—log P„ (1)
stiz^m. , ^<7)Zo&'#'f&&#c
#60 cozo^#^, m%fk^#(7)logp ®JM<D logP ^ (7)#i%fO^ Lxmfi>£ti2>0fiP *> jta i£ t£ t6? j£ AL-f b <r> x, —A&ny^ifb^o logP(±(^:2)t:Zc xnmmx&z>20)o
log P=log PH+X>X ( 2 )
L^L, X tffr+ftcon+ffixklzmtZS-z.fz t), #91 # e c y#^n iz, 53-bb^ l
xm^hftz>mmt<DfemftizttLxim*5-z.z>o m A(f#-E#^>-E>(7)#^l:(i, cl
logP yItz i) thbomwzZ^m L^ittuz^ ^ ^WD0 7 S >='<>, i^yTy^’^^Afoli^
#<7)#^timmT*6o lot, log p ii, log p„ t mmm x <D“*%:<Dnmfcms z try/r^<7)^^:t<7)fnuz oT(^:3)C7)Z -9 lzmt>2ili>
z. t i:i^a,22)0log P=aB^x + 23b,E, + log P„ ( 3 )
E, IZ, ETrtiLn<7)^myWy6fi^^7U^-^^/< 7^-^t*. v>rn<> h tmmiztZo a, b, tiyyy
-e* & log P, IZ3E%]^ a LT% I) m6fi, 6: W:y#&%(7)^#(7)mm logp lx, gy-^
l:#y<sm^^ftcX^XJfrtbbiiZo (±a (i 1 C#L< ZVXniZLX&frtLfzttmjZfrb, y^fk^#(7)iogp ifettiko
7 5 ymm$m&&<ry nm.t lx, y^s
y <c L*^E%f k^#<7)#aq logp^^gtnztitzM^t, ^yy K#%7)^z6 #^(-o.22r^(7)#emv'/:o ei Km^zm^tz nm^mLtzo ^yy Kti^^< <b <> 2M&±.<nm&& e#c/:&!), pH HZ i) E7)4t(7)^K#%^ikyi,'"o #mAn&v^ ph 5-7 #i5-eti^yy tLT^BBL, logP (i(i(Z-^(7)#y &6(7)"C, *%(7)
pHy#mAc#w7HB^L-cmsEL/zo ttz, mm &<?)'{ *>&j&- j ^m&frbmti/zpH TNZ logP IZ&9&%.12-tfztb, 'i *>&mo.l <Dv >m-r h v>A#maymv\ *9 9X, #im±(:#^$r#/:^v^7 < y&<Dfrfrbffif&2 ti/:v — s<> f^yy K(7)^BB#% P y 25°c UTzP.iJ
— 4 —

838 ^49#^9^ (1991) ( 42 )
5ELt'"o
4. y-, b'J'*?? K<7)EtK14
t-f, l gp*>Gly, Ala, Val, Leu, lie, Phe 20 i@O v
h V ^7+ KCOV'T, #&7 < /T,7C&£ 3 / < 7X, h >1^7+ KlcitL 1, 7^7+H:#L0(7)#e^ Ip«p &#*«&£
#&C&(±-C#&d'c/:o ^7+K^ilVal, ^9
'C^viS'n (Cit^T logP <Dl0/L$ v^d
ri'ofco(7)T*, (5£4)l:^$*i*£a&t:&#fcffiMr*l#5£S:£< ZttfVgf.iz>
log P=1.03 E^—0.78 Ipep+0.52 E'SC(RN)+0.34 [Esc(Rm)+Esc(Rc)]_4.07
n=20 s=0.11 r=0.99 ( 4 )n. s, r i±zti?ixit&®)&, mmmm, mm&x-
hho rn. rc cj£m<d, r«Kt:j3it&**g5<7)7 $ /mgmmemfo e^cr,, i=n,M,c)(i, zti^tuD&mz&it2>T$ymmm<Da-
Table 1 Hydrophobicity scale, steric parameter, and fi- turn potential index of amino acid side chains.
Amino Acid 7Z3 E,,cb logfj+*c
Gly 0.00 0.00 0.31Ala 0.32 -0.20 -0.37Val 1.27 -1.29 —0.46Leu 1.81 -1.44 -0.40lie 1.81 -1.81 -0.57Phe 1.95 -0.90 -0.17Tyr 1.20 -0.90 0.09Trp 1.92 -0.86 0.10Met 0.61 -1.03 -0.85Ser -1.49 -0.48 0.12Thr -1.18 -0.73 -0.09Gin -1.41 -0.82 -0.35Asn -1.95 -0.98 0.33Pro 0.86 - -0.23
a From ref. 12 and 13.b From ref. 12 and 13. The reference point is shifted so that Eic (H)=0.
c Calculated from ref. 28.
?"C, 32:^69< tahn)0 m 11:7< ymmm<vE'scig£*to (5$ 4)
iz&itz Zxm<D&tkl± l t:iili^L<, diui, '%i&<Dnmfc&tZ1z£M(r>mz]:-<D log? iztf-fh$h%ztf,mzit£L\zfrn.ztitzz.t 1^^#%li^7+ K*S£-i5j:[>#a-CHg<7> x-stc^gs-t^o m
NH7, coo-(DCONH ^
^7+ K^<7), *<£ b^SUSV' + ^y-A'^toiS
l±/j'$ < E'sc(RN)a<O^Sr|±, Rm, Rc(DZiUZStgpt, N t
OZ 0±#V'o d illi NH7 <h, COO" CONH £blZ5-z.h, mm<r)$}%:<r>±&2i)m%z>ztZ>o Kt(),Jbtudf-e-?) logP liWfr&OX'&Zo (£4)£(£;3)t
Jtirftuf, (1.03E*-0.78Eb.E's^R,)^^^^^^, ^^-4.07 lid
3 Gly-Gly CO logP ICtiS 1*^0 5/0, BP^Try.Trp,
Met, Ser, Thr 7+ K39i@^AoA,
eff C^^ d 6, V'f iuo@#7 5 / LT t.zuhvxftit, (^4)^1$
t A a'M-PXzt 5)^4# <b tl^,z)olog p=0.96 E^-0.64 Ipep+0.56 E^(RJ
+0.34 Esc(RM)+0.26 Esc(Rc)+0.17Iy +0.35 Iw+0.64I„ + 1.67(Is+It)-3.91
n=59 s=0.14 r=0.98 ( 5 )
ly, Iw, Im. Is, It li, ^7'+ K^Try, Trp, Met, Ser,
Thr &#A-CV'fUff (O&C&C-C 1,2, - <0#^%^Z><DX\ fflil
±t:SlfcS£W1'S7 5 y @E$r#tr^7,+ Kli, &
Asn, Gin
lifg^Ei-#*"C, #&&%!!3E#m= e#6da^T&C:a+li, Asn, Gin £ & ^
Ztltz N-7 + + ;Vv-, h U^r+ K7 5 K<OlogP t’
5) b \Hmm<DT l y MSM
rt iz&^&m<0‘&m%&%££<v!$.iL-tz> ztzmbfr iz ttzn)o tztz L±SSis X Z/^SP<7)S14SliV'rtL 1* 7 $ K&-C&&(0-C. E;^(0#^li Rn, Rm, RcT'lit
- 5 -

( 43 ) 839
Table 2 Regression coefficient of indicator variable terms „
Amino Acid Side Chain RegressionCoeffcient
Corrected1Coefficient nb
Ser -CH2OH 1.67 0.8-1.1 2Thr -CH(CH,)OH 1.67 0.8—1.1 2Met -CH2CH2SCH3 0.64 0.64 3Asn -ch2conh2 1.75c 0.9—1.2c 3Gin -ch2ch2conh2 1.16c 0.3—0.6C 4
Trp "TOH
0.35 0.35 4
Tyr -CH2-^~~^-QH 0.17 0.17 6
“The value is “corrected" by subtracting the intramolecular bridgingsolva- tion factor.
bThe number of bonds separating the polar hetero atom in the polar group from theg-carbon of the peptides.
c The value for N-acety 1-peptide-amides.
Util* aiimt), E2 tCli Asn, Gln##(:oV'TT
W7$ PM&mftJ: Ztit, #(: Ser, Thr, Asn, Gin Offlgg d tUiUSjl OH CONH,
I I I .,*ORHj*---CH*---- Ox Hj:--- CHj----0xH Hj:--- CHj—osf=° ROH Q=Q ~«-o( R0H c=o-—h—o;'_NH — L Jh
I I IFig. I Intramolecular bridging-solvation of the hydroxyl
group of Ser residue and the carboxyl group on the backbone. R is either l-Oct or H.
b*]o Z(D? j T’co^ATKS^-S-li, CO^aifijl^COGln CtiVt-mSSn-CV'S26’,,
log P *&*#*>& d t t±o-7-tf^7x ;-)\'tct,<nmx-&b1xx^b0 ^rti
log P IZ LT 0.6-0.9 tflffibtiZ2,25>ZOJ: 7
izmatZo ztLii&tena&'gm&Tbm l <
ac 0,
Kcoiogp \±,
7- T 5 y
sxizx *)iagztiz>ztrfmb frttc^/Zo
5. rh7- K0EtK1±
m±<7)Z')i:, y-. p roiogpemo # 6 c au
T&'t-r h-7-, KO logP li, V-, hYf- y-f *ftm;tE9K*§&V'o v'-, h V KTiJe io^3>^y-v3>=$r
iE(iiSdL^&tbtiTv^o yrDiiLSiili.
EMtcS^LTSl),mt, 4&B(o?m(DmikmtcofflX'Bf&zti2>7km&'£riz&'3< isVXhho ft*l±-T V?-, ^
kw 4 <Dmmizvr 9 &LtiSii£ffMLT£t), f <o#?m*m#a'%;%d'iog P
IC&CI4LTV'* £#*.£<, * LT#r <0 M L$ £Ei"/*v * — 9 b LT, Chou <h Fasman CD#r
i) m L#afm#m&=' e am f 3 :tc l/jo^h,^tiSitgeno291@(7)y 0igL#ai:ttiart&T 5 smz&vmmfrb, 'E-tL'E-'tuor; y@E
i=i~i+3 <0{£Er ,£: i) jg LSiitg
— 6 —

840 m 49 9 (1991) ( 44 )
Fig. 2 /9-Turn structure (Type I).Reproduced from Dickerson, R.E., Takano, T., Eisenberg, D., Kallai, O.B., Samson, L., Cooper, A. and Margoliash, E.,Biol. Chem, 246, 1511 (1971) by permission of The American Society for Biochemistry & Molecular Biology
ft1£ii&f, m# Lt 4 B&Oiff l) IS L#%mo
0#(f, • f1+1 • f1+, • fl+J)t LT/eS£L/Jo 20Zo^i#%0#0##[E logf,(i = i~i+3)]£z7^*y- ’>3 >
t 26, to<7)^(i(^:5)^:($(3r-g:L, :3>*y-v3> cwa&maa: ea <2 17)?-e £ Zj14,o 22T*fflt^:^7^-^tl±, 4100
^i;7 5 ymsSoi£E2-<ht: f,(h 2 6, log f,+, #E log f,(i=i-i+3)] 1:E 2ttfhfrotzo BP L ^ 4 %&^iff I) e B
fijc-r h t -rtitf, 3#g mmoiff % is<)*§v>2<b7)?b75^Zc29,o 202 tit
■’Pli *), Chou t Fasman OtEtiti L T V > & iff 1) IS UiH ia
4 ii §f t-^T oisE t: s it 6 7 <##%O#%l0g f. £, logf,(i = i-i-F3)tO%KII%im
OWfC&OT, logf[+* tOWHot tAV'2 t
-h'b i>mwztitzo ei i: iogf,+, ome^fo j^±o
ffi'fjf h 7-, log?6 oe#m L <c iffuf<c 6 <cv\ 21, BP> #1:3 #1) IS h',logPt)^# < t£hZ.btfWbfrtt£'ltzU)o
$6l: Pro K£Pro ommi±##a"C Pro l:#i"6 E'sc 6 2
Pro Of&E^^Yf- KO
2 £0/-(^;
6)"'ologP=0.94E^-0.58lpeP+0.55 E^(RJ
+0.30[EE;c(Rm) + Esc(Rc)]+ 0.52 Iturn + 0.76 logf1 + 2 + 0.14 IY+0.38Iw+0.66Im + 1.58(Is + It)—0.81 Ip(N)—0.35IP( # pep)—3.87
n=124 s=0.21 r=0.97 ( 6 )
itum &=m#o&&T h7-, KCjlt LT 1 (Dmzt hWg.’tk,Ip(N), IP(#pep)(i^tL^tLN^C Pro 6#&, Pro 6m^Xhbo 2 ti 6OM^&JlOjtRfcld:5timbi)'X\± &v>0 Pro KO^TK14{:oV'-ai<t 0##
nbixfz&mmemwr, iff 0 ISV'6%.>t*7 r 'J >0 logP 2 6, HMCiSv'fii&WbtlfZo
1^TC^V'Tiffi)iSL^it^S^;U’Cv^^^^75^E mtzfztbizcD(nz:fei4)o^st^ffoZjo i-t-^ ^ y - ?§ ?£ * T- o CD o iM £ i: li @ E t>?^!® £ ii /j O -e, foitt,!) t LTCD#jmi:Z6 h V yjl'tnzc.? y -;i/(TFE)5rffi Wj0 MO^y + KOTFE^K ^^^EEB^OCD StL/^26, El3i:^fzociffi)mL#mL
t MfrtlZ Ile-Pro-Gly-IIe (i, TFE *PX? J 7* ni:&v#me&6^, iffi)lSLi#i££Bf£U:< v> He- Ala- Ala-IIe Itiff l) IS L#^fkO@&^&V'2 t,
t Zct’frt)fr^tzo m4iz&mm%?'r?<nVTonsL^it,—<h -dTvv^v^-^-o CD X^y h ;v$r^fo 2 d LT+ h 7-, '+Kii, ty ^ y-;i/^•e TFE * t < iff I) is Lti
^OZ:Ai:^7K14o*§ <
i^$tiZ:u,o^7*+ KO%*#l:^L, ny^y-va >L
TV'32 t SEO^y-+ K*C(i a-^ V y y X £SJ&"t6 nJilb1475?& 0, Q-^\ V -;UO SJ& L<"f$t%7k1±6:OKimem^6 2t()i *6T-7-C*6")o ^P^O#*i±7)'6, ^7'+K O^m^TOn >*y-'>3 >(:#-f6l#W4#6K 6 jr-ttudf, m7k#o^#](i#mm#o^^6f. ^14^7*+ Kofgii37+.y-vH §#"C*6t#x.6'C*67o
- 7 -

( 45 ) 841
[0] x 10’3
£0] x 10'
Be-Ala-Ala-Ile
240 nm wavelength
240 nm. wavelength
Fig. 3 CD Spectra of Ile-Pro-Gly-Ile and lie-Ala-Ala-He.(------) in TFE, (........) in aqueous 0.1 M sodiumphosphate/phosphoric acid buffer (pH 7)
240 nm wavelength
Fig.4 CD Spectra of peptides, type H /5-turn (------ ),gramicidin S-type /5-turn (.......), and randomcoil (------ ).[Referred from Blout, E.R. et al. ,J. Am. Chem. Soc.. 84. 3193 (1962) and Bush, C.A. et al.. Biochemistry. 17. 4951 (1978)]
e. T5/ ummr/tv/-* % moaa a
/t"7 * — £ £<7Xti:ix
U&iS*C£>£WCt COJ: n K 0^*14£¥iflij1"Z> Z £ l±mJjL-e l ±* V>t .&*>ft* t‘ (> 4V'0 'E-Z-ca^ti, (^: ^
y - % £, (jV)tzSo'v'tgaiL;:H,o
Xa = l“'&%<Onx] + \.3Ll&!b%:(S E^c)]+[#%$(?)
ffa<7)iti3i:gto b ')'<’?* KcDlogP<7)
£ £#S Lfz±X~, ;ra Killco$bM£<D;Bfn £ £ #x. titf <fc v*0 &ES£W
K7 $ K<D logPtfHgJ&SlJri'fc log? zmtz Z t o h7^7f Ki
(ftLlf&t&V'o t K(D log P
*C# tzWia-, XaHill log P t > 7h y - 3
z.(D,tmnfrb'<7'T Kcor^ii^-
?#lf 6 nT#g#7)^i%7 $ yMliM<Di^7K14(:ii^80
±*C. COZi
TTo ^2fi±-CCmm$fi"CV'3#(7)7 < y ME
0^14/N*7 y -* fcwifcffcfcfTofco #(D#7K#/<7 y-^U, <mv^ft6(DKyte-Doolittle COEtKH^^3", ^ Z (7%^?)'# #1: L/j(DWolfenden 6(D#!##!& R 3fC^#RH <D7kffi£%ffi<Dffl<7)fr&j)'bi££>bti-f27l<.m+;f-> '> 7 A'321£ ®Chotia (7)^ ? > / M<0=&7C&&M' b
:t# $ fit ON-T-t^^r z y7 5 K <7) log P 75' b fpiffi $ tl tz Fauchdre-PliSka <7)
- 8 -

842 ^ 49 ## 9 (1991) ( 46 )
Table 3 Hydrophobicity scales of amino acid side chains.
AminoAcid
?VHI(K.D.)d HP(W.)e HS(C.)f K (F.P.)? Ap'CN.T.y w.y
Nb MC=
Gly 0 0 0 0 0 0 0 0Ala 0.19 0.24 2.2 -0.45 0.02 0.31 0.5 0.53Val 0.49 0.82 4.6 -0.40 0.18 1.22 1.5 1.46Leu 0.92 1.28 4.2 -0.11 0.10 1.70 1.8 1.99lie 0.72 1.17 4.9 -0.24 0.22 1.80 - 1.99Phe 1.35 1.57 3.2 -3.15 0.14 1.79 2.5 2.24Tyr 0.78 1.01 -0.9 -8.50 -0.38 0.96 2.3 1,70Trp 1.72 1.93 -0.5 -8.27 -0.12 2.25 3.4 2.31
Met 0.67 0.93 2.3 -3.87 0.05 1.23 1.3 1.08Ser -0.08 0.04 -0.4 -7.45 -0.21 -0.04 -0.3 -0.56
Thr 0.07 0.25 -0.3 -7.27 -0.19 0.26 0.4 -0.26
Gin -0.51 -0.30 -3.1 -11.77 -0.71 -0.22 - -1.09
Asn -0.51 -0.26 -3.1 -12.07 -0.48 -0.60 - -1.05
Pro j j -1.2 - -0.30 0.72 — 1.01
* From ref. 14, calculated from eq. 6. bFor //-terminal residues. c For central and C-terminal residues.d Hydropathy index of Kyte-Doolittle (31). Reference was shfted to Gly.'Hydration potential of VVolfenden et al. (32). f Logarithm of hydrophobic scale of Chothia (33).8 7t value of Fauchere-Pliska (34) from log P values of //-acetyl amino acin amides.h Relative free energy of transfer from either ethanol or dioxane to water (kcal/mole) of Nozaki-Tanford (35).1 f value of Rekker (36).j?ra(location, number of residues) of proline ;ra(N, 2) : 0.35, ,t„(MC, 2) : 1.16,7ra(N, 3) : 0.00,-a(MC, 3) : 0.81, /-« (N, 4) : -0.34, ^.(MC, 4) : 0.46.
HI(KD.) HP (W.) HS (C.)
•• ••0
0
. @
• • • «*
0.4
©
• • * •
«* # • • • •___
___
• •
•• .
0.0 • • *
•• •
•
•
-10« -0.4 •
•
-1 0 1 2-1 Jta(MC)
1 2 -1 Jta(MQ
0 1 2 *a(MC)
X(FJ>.) AH0(N.T.) f(R.)
• ©L *
4' © •
2
©
l 1 • •
•v#e
•
•
2
•V• 0
• • • • • \
0 • •*
-2 ,
-1 0 1 2 -1 ita(MC) ° 1 Xa(MC)2 _1 0 1 Xo(MC) 2
Fig.5 Relationships between na(MC) and other hydrophobicity parameters.
— 9 —

( 47 ) 843
ggO* < Nozaki-Tanford O^rSc351, ©Rekker <7)7 V ^ ^
7 #3 C^fL^CT)#^, itzjeat<7)#^eE)5
% ^©-®<7)#[i^<c I) <D
®~©<7)liLti:£>2
^ Cl^tiio -E-ZT *0 ^#<7)##f(:#6<)/<7 ^ ^
emv'-c, -E-<7)M^:, *zw^?ssm c^(t6 ^sscmcfv^-cmm $ ft zz©~©coiiii, 0 5 7 na t X T
i ;m.mx^x^^^ti)^t>i)^tz0 (D<7)Wolfenden <7)#(i7 < S &mm<7)7km&'k&5-i±^tzifx, ®<7)chotia<Dm±mm<nsLvm%;ttksm£-&
*a li^L^&rt'oZzoZtUt'£<7)J: 7 izmmx^io fi|Hb Wolfenden <7)#(±
<7)7Kt:#f <##LT
Gd'ot#, Z 6%m<kA-ATV'6 Z ^ (:mftft6o Chotia <7)#t, %*
X D litf L57k#m:l^LX(D7km£&i%-5-i£lz Z6##7)^#v^ 3L^^«<7)^75'ib,
Ke-S-1£ ^SfnStStc^-t h 7kmt*m''Z%z>tz£>, *>'*'7nftuizm£>&zix*? -f<££CL£iLb UV'io :<?)ilf(7)f$:##i:L X^h<DX, Kyte-Doolittle <7)E 3zL&5MJg t *S*S
^###%m^zz(j-emft $ ft* 0 %4r <7)#m LX:# lv't 5 j mmmk#/<7 *-*?
xamt. Kmc^if6#$m<7)m7K%^##LX: i> <nxh 9, #(C7kf9;»'tbWm8z''<7)^7> K<7)#m^, '>yy;w<yy K<7)i 9 ^mL%s^my*^yy k<d
6o mt - jgm%7 $ /^Cov'Tli, >#LcM±
m L -cgij # c -e <7)m^i± e 6 o, %###^-0*6^, 7;l/f->#$at:cv'-Cli, >m<7)
v>#im<7)^ t >mt:jt%L-c%#%t:Av' (*14 SK*lii$<7)^)& <L,
ftcc*3o
7.
(^3)<7)mjR^»e^m%i±#(BP)<7)^&#cg#
%A-c, ±m@i±cox;# $<7)mfb^m%<7)#m±<Daf<D^fb<7)^#kL LTE^-tL Hansch EtZ-ftB^ft
*£smiiisi±m<7)^mi, mmsktct-ziZi
*')
KfrWtXfc <rSffl$ftTv>;E>(5t8)8)o
log(BR) = a^+/oa+d‘S+const. ( g )
*, a,Sti^ftW6bK14,a, p, S li^rtl^tl<D%]%:<7)
&5-<Dmm*mt>t&%.x& , m/jcim&c zamig
#^#(:ZcT$^^ft*o Am^#<7)±#$<7)mib$- 6 Bf-C Z ^ Free-Wilson %
£fflvt-c Nisato ib7)fl/->ES#Je##LZz#T, Zas
lavsky btfx.y try t V L^<7)<7), ±g?£i4^yy KO^fiWtl
a###l%#M&V', ccDi&H&li, ^y
<7 y - ^ <7)^#7)%mT* ^ zz c L, & ZLTS^L Zc6H^%:7 ^->a>
l^oZz-ils ^-e**&mftft*. L^L##::"7v^y(i, £t*ti^yy K<o«*t/<7 ^ ^ e#F#f 6 2 t LZzo yc-en>yy-'>3 ytfhi h V
^yy K<7)^i$^(:^y6^m%m#e^*zzo #*t7'*7X-^i:Ltli, HffiiJ^tiZz logP t, &mU' ^y#j$ftzz#Lzm^tztzh, log?aX lffr=f-<o£:m*lz #V'(3 t*mim-f Zti'7 #i$* z> %
^mhiitz9mo logp <%>###tTmmt *w\m<n;< ?*-9t LXWVm^X, #MZj%rtmfrtifzZ £fr b, ^Si]{iy5MogP <Dmzffiz.Z> i> <F>Xhh <ht'
TZtfrXZZo^<yy K<7)n - y 3 XOWfCM LTfi, Si5
2>tm\zx^x, iZzA^## NMR <7)%E(i, ^yy K<7)#%fC(7)3 >y y - '> 3 >##(:y#^%#je^ZzLTv^. ft* It
Bacillus brevis <DM£. t Z>, xZz^yy K"C^) 6 y 7 $ ’> S/ > S(GS)(Val-Orn-Leu-D-Phe-Pro), ^m#:<7)%7k%Lfg#L<7)^#l:^V'T#%e^^-cv^
cs <7)%m@i±(i, mttmt
Val Leu
> Leu ' Val
Orn > D-Phe
D-Phe
Fig.6 Structure of gramicidin S(GS).
-10-

844 #49 ##9-# (1991) ( 48 )
i)*m < MfroTiJ 0, 0 6 <7) J: -9 H o-Phe-Pro <7)Ufr
x-vrtim*z>tti>iz(VT*)j&Lmm). /?-->-Mfit £ t z> z. 1tJ*tS14S££k t o •ctg-e*& t v'frti/cv'&«>„ %4r li Orn £ Lys (:, D-Phe £ $ $ g£ & d-7
logPtamsmt<DR#(:owy^# L7LLC5, n >*;<-'> a£ 925%L&v'&6t logP <h^i(?i^i
LZ:41)o CC1:^L^:Z -) tc,±mmi±^yyel
v^gjE^tOT-if'f L-e^)5 ^ 0
8. &fc>UlL
^yy mmfc&zmfcT i(Drmiz&^XMtifL, 7 X y gESoE Lv^m 7k#/<7 ^ *„ Srittiti LtLo rcO/x-7^ tztifrb^MZtitz^TfT- KcOj^Tk-Eti, ^<yy >
bT, E^it^yy KcoBtWc&ttSn >*,< --> B >60#^, ^ L-c
* - * & <£laota'iw, fr-T->&nzm'''btL&x7iz%z>z.L^mLTV^o
(%3#5^ 13 B#m)
£ m1) J. Hughes, T.W. Smith, H.W. Kosterlitz, L.A.
Fothergill, B.A. Morgan, H.R. Morris, Nature, 258, 577 (1975)
2) A.N. Starratt, B.E. Brown, Life Sci., 17, 1253 (1975)
3) E. Ruoslahti, M.D. Pierschbacher, Science, 238, 491 (1987)
4) K, Nakanishi, R. Matsuno, Eur.J. Biochem., 161, 533 (1986)
5) /Kama, #3ik, 49, 42 (1991)6) D.V. Goeddel, D.G. Kleid, F. Bolovar, H.L.
Heyneker, D.G. Yasura, R. Crea, T. Hi rose, A. Kraszewski, K. Itakura, A.D. Riggs, Proc. Natl. Acad. Sci. USA, 76, 106 (1979)
7) M. SjdstrOm, S. Wold, A. Wieslander, L. Rilfors, EMBOJ., 6, 823 (1987)
8) C. Hansch, T. Fujita, /. Am. Chem. Soc., 86, 1616 (1964)
9) S.H. Free, Jr., J.W. Wilson, J. Med. Chem., 7, 395 (1964)
io) msism, Kv?
/ffoj, it^mK 1986,P.129
id mm- #, 9, 571(1984)12) M. Akamatsu, Y. Yoshida, H. Nakamura, M.
Asao, H. Iwamura, T. Fujita, Quant. Struct. -Act. Relat,. 8, 195 (1989)
13) M. Akamatsu, S. Okutani, K. Nakao, N.J. Hong, T. Fujita, ibid., 9, 189 (1990)
14) % # cf, M. Akamatsu, T. Fujita, J. Pham. Sci., Quantitative Analyses of Hydrophobicity of Di- to Pentapeptides Having Unionizable Side Chains with Substituent and Structural Parameters
15) W. Kdnig, R. Geiger, Chem. Ber. 103, 788 (1970)16) G.W. Anderson, J.E. Zimmerman, F.M. Calla
han, J. Am. Chem. Soc., 86, 1839 (1964)17) M. Bodanszky, V. du Vigneaud, ibid., 81, 5688
(1959)18) J.R. Vaughan, Jr., R.L. Osato, ibid., 74, 676
(1952)19) T. Fujita, J. Iwasa, C. Hansch, ibid., 86, 5175
(1964)20) C. Hansch, A.J. Leo, “Substituent Constants for
Correlation Analysis in Chemistry and Biology”, John Wiley and Sons, Inc., New York, 1979, p.13
21) T. Fujita, Prog. Phys. Org. Chem., 14, 75 (1983)22) C. Takayama, M. Akamatsu, T. Fujita, Quant.
Struct. -Act. Relat., 4, 149 (1985)23) C. Yamagami, N. Takao, T. Fujita, ibid., 9, 313
(1990)24) H. Terada, Chem. Pham. Bull., 23, 1960 (1975)25) D.J. Abraham, A.J. Leo, PROTEINSStructure.
Function, and Genetics, 1987, 13026) G. Fermi, M.F. Perutz, B. Shaanan, R. Fourme.
/. Mol. Biol., 175, 159 (1984)27) C.M. Venkatachalam, Bipolymers, 6, 1425 (1968)28) P.Y. Chou, G.D. Fasman, /. Mol. Biol., 115, 135
(1977)29) ###&. mmW;, #18 0#^##R5/>f
1990, p.31730) K. Nakai, A. Kidera, M. Kanehisa, Protein En
gineering, 2, 93 (1988)31) J. Kyte, R.F. Doolittle, J. Mol. Biol., 157. 105
(1982)32) R.V. Wolfenden, P.M. Cullis, C.C.F. Southgate,
Science, 206, 575 (1979)33) C. Chothia, J. Mol. Biol., 105, 1 (1976)34) J.-L. Fauch6re, V. PliSka, Eur. J. Med. Chem.,
18. 369(1983)35) Y. Nozaki, C. Tanford, /. Biol. Chem., 246, 2211
(1971)36) R.F. Rekker, “The Hydrophobic Fragmental
Constant,” Elsevier Scientific Publishing Company, Amsterdam, 1977
-11-

( 49 ) 845
37) D. Nisato. J, Wagnon. G. Callet, D. Mettefeu, J.-L. Assens. C. Plouzane. B. Tonnerre. V. PliSka. J.-L. Fauchere. J. Med. Chem.. 30. 2287 (1987)
38) B. Yu. Zaslavsky. N.M. Mestechkina, L.M. Miheeva. S.V. Rogozhin. G. Ya. Bakalkin. G.G. Rjazhsky, E.V. Chetverina. A.A. Asmuko. J.D. Bespalova, N. V. Korobov, O.N. Chichenkov,
Biochem. Pharrn., 31, 3757 (1982)39) M. Asao, H. Iwamura, M. Akamatsu, T. Fujita,/.
Med. Chem., 30. 1873 (1987)40) M. Aoyagi, S. Lee, N. Izumiya, /. Mol. Graphics,
5. 35 (1987)41) JVlij 1991
mi. P.i98
-12-

2«: ''WKCD&aas*

^9- v<D#n • ^Stie^isjRiwcfc-t), 7 ~ j& 50 mm
gg©^^? K$, ^OfS^JrOffl^r^fiScrtSJrai^ortfco $6C, JStiJ
#=f* 10 k gjs©ae«fc@^T»a(o<i:c'fe<o*s^$n,
^<7> Yzmmz^m-rzro&mvm&tis&fDiaw
mmvmw, m&goxt: y-7'7-,
TC'So•//////////////////////////////////////////////////////// t////////////////S"ftft"//,////,/S///////////tSSttfS/t/'////////////////////////t///.
tiD&C: Hb#o#2c<o#im
&9cttms:<&mz*iXgtz<> ^7'f Hb#:±,
9,
fb#^:6"oNm#%o'e>t^<0 DNAEF##ci3^om^MO—^
SBoS^ScoH^, SMDNAguioI^II
yf- Ffb#(c#rLv'#§§*r 5 ftgtcjJbtz^xflp
L*Cl»«oiot, KJi, likA,Z<D&£. t$L
®:J:o-CMc,riti'fc»
f- K{b#c^Tc^i'^m,
iEV'SW^itick&ti'&MXb 0,
Ko^f^SIr^-Cto/^
-7‘(t, r <
-^, @sE-en-<7>z.tmmx&o,
ffi3£i>'&tttbiiX£ti0 &#&%%##, 7 ; / ### * I
frtzZi)vm.&.ft^mi%HX%tz0 tiz, a&ffi©HPLC
!t C %»<b "j" 6 FAB (fast atom bom
bardment) ^+k*7!l -"EMi&tfrEft<h*
z <;, @SS"t"n y t- k-c
B61j^oEE^^oFF®^bJ6E> ft o T §-
iz0 K^i'O^o
£ o-Ci'fcliggK.Mi-S^ScS^V'5
$ P>tc-r>f ^ p a v-
.Ho Z 5 i^ft-o fto t5‘ <( L"C, —ES-'' y■?• K-B"
OXZZ.kttZ'itZo
$n.ft*m%-$ziz>tzibo!fcM, f?Mmm<om#T®tz2>
041g^7YKit07tP/, #afb###a<D
tztbo^T/'>"<??- k,
Ky» >f yft<F, ##C##ft-<yf- Kj&'»#fi! ft WffEtzfSl'bizXi'
60 c<7 z $ y^ m
SSrcJgt5 fcift^tz^ tit,
y^ Kfb#©m^-e&S. UT, @ffi£tSo'too4,
Saburo Aimoto, (t 565 ^lEiFrtfcEBtfTlllEHIr 3 - 2) [Institute for Protein Research, Osaka University, Yamadaoka, Suita, Osaka 565, Japan]
Chemical Synthesis of ProteinI K«y word | [$65l<D{b^£-/£] t^fr K-£/£]
-13-

it 215
f- k, S6So<i:^:#^i'^'OV'-c:^l^-?"oo
ESttf'J
1. S^#fiBW2fe(r«fc«Sa^^,51 KO^fig
3§,:&ffi''' Ppix'Ci'cioi t jl56t)r KO
#f£&it, ^^ ^ (Boc) fk7 ? / Eaf/r!
v v 2 @/fg£-c£> o o c *i!Z, El l z 5 CD &E
ggoKJSI, © #=%, © 7 ; y < D
igLOO, KlIlMt‘^7 ; y
itiAU -<7> Ki^#Sc-«:tl'<^£-C'|.oo
c o#fp;z, *'9 y.#A^
I'-cmfTf&aL, tfc^SEoirE^B^rfTES^-C
i'Oo ^E^7E, yfkzKas&awt h 'J7/^-
p y y y#;Z6""C^E^7'f- K^jjjl^&ll L,
^U:^7>K^MU:Ot), ^i"o0
&&*%&*>** b-r<t¥&)&n<D£mi&*#Ji t -t
6^7> nc^i'T—< i
^ft®?ij^o%o^iti§&-e&*§o hplc -coSl
s^sn^y?- ki^k itm •40—50 gii|07 ; y EZ 0 tz2^7’3- Kiwis' I' X
it, 7)>ft 9 oE^t'86-7>-?-£-<7> K&ZfZfo&e
KO^iwii,
<, Boc
#ve -% ^EE*‘^E-C, ^7> K^mS/"5“bO%
D fcti Lie#*7 y <k7KSO Z 5 £ V 7^ V'
9-7 />/h i/ — /v y h ❖ -7 /✓ TP - /u (Fmoc) it 7 i y gj
®IEo&ES^7
< y EoS-n^Eog&fkSo^aiwZ o, Boc &6MI
#C^#-<7'f- KO#gt75»-C15i5C/i:ot tf-1-3^
iiois 70 d' P) 100 ^^@^0 -<y^ KOft#-& E'iwitV't-e^Ti'^ D o v xy^r^9a B5:±'Ci-t, v< K^ifibixAo, 7 ; smfr&r
o -cv' 2mgx$> ^xa>, e#;c#
SEcffli'P>ii*Z 5 ftSI&So ioo£E^JiZ||g|i:::z
^rne<bX 7^V',IllcSrT^il: *( /"£ <5 I-f^c o ~C, Boc JS^ol'IZ Fmoc ^
d»^i-:-zaZMZhftti^fzO, &2i'!t7 ; y EOSXttfE£ kit <9, 7'f- YcDft&Stmi'X £<it
ztk'&m'mf£kit2mm<vx>k'3kLxz>tft>
H2a <<-/*- KiwZo-C!t, -<7-^ K$iO#S^fFO^
±-e, 3:387 ; y SoEEg^.t.&f-l&gE $ Hit <ft9,
g^y'f- K^;± c A < # t a#e 4) & a, m2OE0:t, '<7*V&mffid'!b&0tti'fk&<D%m£
<DT'5t±ttffiM k bn 5 = Tam P>OgB
#9EC Z 0 0 Oii^75»A britti}'*, z ii^-^Xitit
v^o SSOEBIZ, -<7'f- K^mgd'%)g&mLA&6 3„ as&d:# < ft a;-%v\ g#j-<y> Kir
AotfftL, Zn&m&i-oZ >ii^S^iwSS>/-to«>
tzkXiZ, 100 KO-S-Ei-^V'*C, g
6tJ>i-^-<7> Kd;m&fx#0%*'C^LLd'#:&L-C
V'Aii'Z 5ttW,&, M^lfrzgt'OwifDt'^oZ 9
^%m-Cg%#1r%#Z <%Dffi-f w 6iZt^-CSH
"C65.
Boc-NHCHR„CO-OCH2-PAM-®E ®%ESO$5E . I
CF 3COO_ • +NH3CH R * CO-OCH*-P AM-g-51 ©❖fa i
nh2chr„co-och2-pam-we©7 5/Km#* i
Boc-NHCHRn_1CO-NHCHRnCO-OCH2-PAM-^,^©, ©, ®(D < 0 jg L ’
Boc-NHCHRiCO-NHCHR;CO-NHCHRjCO............. NHCHR„-,CO-NHCHR„CO-OCH2-PAM-^S©Sffia^ibOSzOdjL I®B^#OE8 i©«$3 I
NH2CHR1CO-NHCHR2CO-NHCHR3CO-NHCHR4CO--NHCHRn-2CO-NHCHRm-ICO-NHCHR„COOH
0 1. Boc 7 $ /SE^mV'S-tyT K0@t8-£j£08Ei&PAM-8tIg : -QH4CH2CONHCH2C«H4-mm, ®«MOS5J?g : CF,COOH/CH2Cl2, ©❖?□ : ^< 7^0 ^xf WT i 7, @7 i /EO^-S-: Boc-NHCHR2COOH+Ar, Ar,-y v7 ->yV*/UrKv < i K + l-H Ko#^V'/ h U 7 '/-*, ©WSB^^OSt OttiL : M7K7y it&m, ®gROSSIS : 7 2 J B**f, ©fVK = 7*A 7U7h/y7^-, HPLC, 'ff'/^^n7l>/77-<-,
-14-

216
Lfca'»oT:, t-r—&^y* K^^-^v£-e^u,
B##o*-<<,
ffno KicAfoB^y^ k&i^klS&^J: <^tti'< ^yf&ihj-fcJo p-7 h ^7 7 >f -, HP-
LC, -f v3c^^ p ~v h y 7 7 >f —^ISisL'C, ^'OS
SoMEo-<7> K,^,h*iitiW6iic.^ar^i"»o1 70 7b>£>
100^SSSo-<7> K% 1 @^EL, -CJ1-C NMR <D
§znk-fz>zkfrx%z>£?) tmmo&fe g a
c k ti'X^tztz b, 9 gf@0#G^cjSf
<h
teoe^wfH-, (IL-8, 72 ^S)
^vWXb'^'iZtzb 5 5)o
CES5£‘J] Hiv-I 7^7^yiypf7-f (HI-
v-i y p y 7 - o o-g-sM^St'OlgKogsSci'M'ti-? i
SESf&'/fTVSO'/t, Kent 0/;u-7*-e^)5,
$£, Applied Biosystems %t <D 430 A
inx., $ &C#8moB%a&?7 ; / gtogA^f^ESal, Boc 7 z / m
2 iZyji-f- 99 B3E ® 7 i S&£ 9 ?£%> HIV-1 (human
immunodeficiency virus-1) <0 7 7./< 7 ^r vg^y p t 7
--e(Hiv-i ypf 7-4f) ^^L^6)e -<yy F#
1 15Pro-Gin-Ile-Thr-Leu-Trp-GIn-Arg-Rro-Leu-Val-Thr-Ile-Arg-lle-
30Gly-Gly-GIn-Leu-lys-Glu-Ala-Leu-Leu-Asp-Thr-Gly-Ala-Asp-Asp-
45Thr-Val-Leu-Glu-Glu-Met-Asn-leu-Pro-Gly-Lys-Trp-Lys-Pro-Lys-
60Met-Ile-Gly-Gly-lle-GIy-Gly-Phe-lle-Lys-Val-Arg-GIn-Tyr-Asp-
67 75Gin-Ile-Pro-Val-Glu-Ile-Cys-Gly-His-Lys-Ala-Ile-Gly-Thr-Val-
90Lcu-Val-Gly-Pro-Thr-Pro-Val-Asn-IIc-IIc-Gly-Arg-Asn-Lcu-Lcu-
95 99Thr-GIn-IIc-Gly-Cys-Thr-Leu-Asn-Phe
0 2. yU7-l (HIV-l) £077/-?7^y®yDr7-y<7)7 $ / m&n
mas #e g %
B-e^TLfcOt,, ^S^y^- Kmi
^*7 -y-fk^KS-e^SL,
Sephadex G-50 HPLC "C^L, $?>}-
HIV-1 ypf7--t’^f|c); kiZ&
$jLtz0 ^«yp / 5 i
025^6tl^0t5>,*f l%? -e& a Hiv-i
y p 7 7 - -fcf^ z 5 , ^®rnM^Mi±t'K'ti'tzb 1to®X:Z>bt)''ik$S}b\z,tZ, L 4'g'J&ilS
e>»#e>.tLfcS6S!t, 55aso gag lUjE&oEM-^fL
y p T 7 - 11,, ttz&!&mnz%^tznm^b, cogami, gt07;y^a:fe^
1 Z>*^/y-r^->^m’<zftfc-?b&&-C^7'y Klg^SJE
i"o C £75‘i£jE>£'<h frofco Nutt #E7-^:t#
57)o
Kent e>ii, e 6i- glSO^SH'afWSi-i"57^-
67 95 {50*77 7^ L-a-7 ; / gSE(Aba)
^S^LfcBS#ar-^E Lfcs)0 ^
mi^rn-to 0. C O [Aba 67,
95] HIV-l y p ?7 —eo 99 $ZLfz0 $ P> ir. d" V b fcf ^ — (Ac-Thr-Ile-Nle-F [CH2-
NH]-Nle-Gln-Arg*7 ? Y') k, £ <DESftBS£OE
SEt-* ar V ;510>o ffio-f yt
Vol. 37 No. 3 (1992)
mU [Aba67,95]-HIV-l yo77—
EMBSX
-tyf F
(^mol)i£S&a)-aiaeie mem
1 phe-mm 300 - = 100% -
3 ^yf F-etsi 211 - 70% -
4 154 51% = 100%5 Sephadex G-50 0.146 #& HPLC 15 0.29 5% 10%6 1.007 -f O
7^*77>f-5*» l.OQc) 1.8% 3. 5%
a ) = : Abz-Thr-Ile-Nle-Phe(NO);-Gln-Arg • 7; K (Abz ; 2-7 Z / ^'s 7 -f 7U)
SifilCE : pH 6. 5, 0. 1 M MES, 1 M NaCi0b) ^ 50 mg0c ) f-7^—/«-& : ^ 5/f^o
-15-

(Ac-Ser-Leu-Asn-Phe-#- [CH(OH)-CH2N]-Pro-Ile- Val-OMe, Val-Ser-Gln-Asn-Leu-F[CH(OH)-CH2]-
Val-Ile-Val) > <D ###^0 MW'1’™Nagai j^MgLfc^y*- K© j5 *-yfcg|Lfc- - -yhx»^4o HIV-1 y p y y --e oSfS#irE L14\
gf§i*fc£S:^/£$-£a&S!'t(> £ » o * p*r h if =7-7 4-±tr^-e^^V'il^E%75s comi#"eiELV'#a
^J£t5'*C'£-f i-£tS£L*I 0, --CMf£5 l-^o^Dtoc HIV-1 7ppt7-
•fc'OJ: $ i- 2 fi&2fSEi~ amaHi-M '-at, 2## p>t-S5S*»»^|6-c*ofcfi6 5,
66ES© HIV-1 7 = 77'-f^fgi;
(i, Kent i> 2 Z> Z kt£fi>b,Z.CDX o fafj§S$±©ftj.5tk ' L fa © *at fa /) > 6 5 /5>0
2. @S£r^u/:^7,f rtyy> h^muas
aK©@*§£E-ai, 30ESS
mo^y'rrx'&h'.z, -be, mm. e#©^fii©@>&»&*>. ta^ESEfai^g© getj^y^- Ktfto:^*‘,
vt-e:twriE-e^,oe <> L@tgE-c-BELfa so ESS
go^yf- K«t/^ •/ h^SHISifcot), t < mef a c 6 ^-e t *lm:\ so-ioo ESSEo-<y y K-e&iiti*. fa B' c a>jgi'setj^/V'-BEt- ?o!t-fy^a, #%, sfcfe-e&e^a^yy K-ty
/ y h l DA^fto^yy K^mm-e&a i ? i 9 lio^yf K-t
y/ po-be^
^#E>faa-e&6^. c©i $ic,
l, $^-ky/ y hBE©^fL$4r##L,
Ei-S^ £ fa aM£-C#-fc y / y h^fEB-e-tir-CSd r^toc^>t^n(i ^^famam©ib^BE
£>faa!t-f-e£a0C © 1 5 fa-BEE^Ei" ^ < , Kaiser oy/u-y
(t±4r y AV' 6 Giralt ©y/U-yttFmoc -SiCX^^rC tbLlc), Li © yx, - y (*;£$$* /u.-K
yy/us^r^m-ji-iE&ik-tal 17), mamolb^B-Eic^^l-a'ao Ld'L,
E^B h #8351 L-ci • a BEQftssSriS it-? a a{@tSl$!] Bacillus stcar other mo philas © DNA |£
-Bmam chbS) ©-bei9)
E^att, K-ty > y h
Kjfay ? / S©^m^##cSfax.a^& ^ gg # l,BPTI < -BE-C^ a C BB;^Lfa19)0 Lri'LE
y^Eoig^BtjfstBfbBfTfa 5 c k25‘"C^fa^*ofafa£>, SSI© * y/vS^ro
72}c^SL*C^='< MMfi'bofao -r©faa6c,
fai-t, 2-0©gJIj@^£ofa0 floras*!-*, y:^ yx^E© &Senfa&JB© -< y y K-e y >= v
^m-c^mi-afzoc, **#EBB afa'd^ttY^fa^fac k-Cabo M2©&g MA:t, y ^y<7^y^y/>^ ; ySS^t^< <>o -t y > y h *at, ^^CO^TKi^oi^i'^ES *r & -o c act a fa *, ^yf * o. mmohplc -eo-t=y/ y iomw>>mmt^bzk^^&
kX'&bo
—^, Li a^y^-^^y'^y-fcy/t y hE-S-S (i,
* yu.-K-yjiEtR^jS&<b> v' 5 ^icjaV'-c, ES8 ftLfrft’C&ofcT&s SI*trtSL-CV'fco y-zh*/u,-H^
•y^^o^-7r-^^:t, y ; / Sc^#S^#ATaK
6, y ; @Mb$TVC^^-ra75»®g^c>tjgc-r. co^%##oy; /miC(t, ^ - /US i: C $ /£ V '^tK y h 9
3 yit*iA-e|a-> h 7
^rx.^ < ftofc, cnttgjic^fLy
X'$>Z>tz2>, HPLC %<b'O0B=&#T
-emmi-ac6^-e#-t<^^-cL$-?/c. tfc, -n ■7 = -/uibe^i-cva^^^^rBi^oWESSic^-ta
Ec#< T^cyuao e aic^^y i / go &ES<-, y 1 7 3 - <b 512, / f- /> X /L-.-h p /'-— h 41 y ^7 /L'.-H — $
1 151Met-Asn-Lys-Thr-Glu-Leu-Ile-Asn-Ala-Va!-Ala-Glu-Thr-Ser-GIy-
16 30
Leu-Ser-Lys-Lys-Asp-Ala-Thr-Lys-Ala-Val-Asp-Ala-Val-Phe-Asp-
31 I 45Ser-Ile-Thr-Glu-Ala-Leu-Arg-Lys-Gly-Asp-Lys-Val-Gln-Leu-1le-
4G 601Gly-Rhe-Gly-Asn-Phe-Glu-Val-Arg-Glu-Arg-Ala-Ala-Arg-Lys-Gly-
61 75Arg-Asn*Pro-Cln*Thr-Gly*Glu-Glu-Met*Glu-11e-Pro-AIa-Ser-Lys-
76 90Val-Pro-Ala-Pfie*Lys-Pro-Gly-Lys-Ala-Lcu-Lys-Asp-Ala-Val-Lys
El 3. Bacillus stearothermophilus (D DNA-B$an (hbs) <D71 / min
I Enu. -try/ y hHSB-efi/j:-5tztiLmZfr'to
217
—16 —

218 a H m m. mm Vol. 37 No. 3 (1992)
Sit. 0.1m NaOH
$jic>z5s
7- K O ux i~ X Lz>'T2~f,
^^"C'^Oo t-i^Trfi/^Sr-Zr'y
^Sitfb^Sjj- 4T^-e£ 0,
itfi: $ nrx 9 2nzK#^ £ -H,7- f
-trC-C,
^ ^.t: ^
|g£fc£ J: 5, Li p>^ 7 7 7*7
arSXL,
^|gt©:iiLfc, 4U SB
'<-7j7~ F* *S~7 /U 4r /U 7 zh
^XT/u^r Boc 7Wl?:|i'
■OC07Z —x77;)l|g7 7 y-C
<fS^b$TLTLit, Li P>
O^gcDjttfrafBL,
^oggA^f^-c^m-e# % it-f
-e&6.
LIT, K 5-7
7 7- 7 7 7 — x 7 /> O ##& <5:,
•Ol'-C, 90 i y E t D
^3 dna m-a-mam chbS) o
L-CxMa,
HBs <D 7 i / gtiE^Jzf El 3 i-
4 - methy I benzhyd ry I am i ne #g# (NHa-^fii)1 Boc-Gly-SCHaCHaCOOH + DCC
Boc-Gly-SCH2CH2C0NH-&fliABI 430A^7*TK£j£$£Z77AV7 F 9%7 ver. 1.40 NMP/HOBt f-Boc
Boc-Asp(OcHex)-Lys(ClZ)-VaI -Gin-Leu-11e-GIy-Phe-GIy-Asn-Phe-GIu(0BzI)- Val-Arg-GIu(0Bzl)-Arg(Tos)-Ala-Ala-Arg(Tos)-Lys(ClZ)-Gly-SCH2CH2C0NH-8Jii
4 55U V7;u^-DMz^ikyTb> l iox4-NT^^/tx>; y/igftyfi/>4 /Noc-ONp/DMF
z"Noc-Asp(OcHex)-Lys(CIZ)-VaI-GIn-Leu-1le-Gly-Phe-Gly-Asn-Phe-Glu(OBzl)- Va I-Arg-G I u(0Bzl) -Arg(Tos)-A I a-AI a-Arg(Tos)-Lys(CIZ)-G I y-SCH2CH2CONH-£fls
4 HF^lgz’Noc-Asp-Lys-Val -Gin-Leu-1Ie-GIy-Phe-GIy-Asn-Phe-GIu-VaI-Arg-GIu-Arg- A la-Ala-Arg-Lys-G I y-SCH2CH2CONH2
4 HPLCCcke#^4 Boc-ONSuZ F V x7;L7 3. >
z‘Noc-Asp-Lys(Boc)-Val -Gin-Leu-1 le-Gly-Phe-Gly-Asn-Phe-Glu-Val-Arg-Glu- Arg-Ala-Ala-Arg-Lys(Boc)-Gly-SCH2CH2CONH2
z'Noc-[Lys(Boc)41,59]-HBs(40-60)-SCH2CHaC0NH2 (3)m 4. F-77^x7/uo-g-f5*i&
DCC : X, X'-v -y y □ ^ 7 v A ytvU n-Ny'f $ K, cHex : y y a ^ 7 y A, C1Z : 2-y n □ vN y -y A/tj- *> ^7 A— ;L, Bzl : ^tyyyv, Tos : F yA, /Noc
: < V — n 7 — yU 7 7 ’> 7 /l— /l, ONp : / "y — F a 7 * —/L0
Boc-Met-Asn-Lys(Boc)-Thr-Glu-Leu-l le-Asn-Ala-Val-Ala-Glu-Thr-Ser-Gly- SCH2CH2C0NH2
Boc-[Lys(Boc)3]-HBs(l-15)-SCH2CH2C0NH2 (1)
z‘Noc-Leu-Ser-Lys(Boc)-Lys(Boc)-Asp-Ala-Thr-Lys(Boc)-Ala-Val-Asp-Ala-Val- Phe-Asp-Ser-Ile-Thr-Glu-Ala-Leu-Arg-Lys(Boc)-Gly-SCH2CH2CONH2
/Noc-[Lys(Boc)18,19,23,38] -HBs(16-39)-SCH2CH2 C0NH2 (2)
jjrf0 %aiz£}®y-t i itisPK'tzzb, Z v y yZ/‘7 /utHt- y^vsix
Sr^SJL, K-t:
y'y y F <h Lfz0
/Noc-Asp-Lys(Boc)-Val-Gln-Leu-lle-Gly-Phe-Gly-Asn-Phe-Glu-Val-Arg-Glu- Arg-Ala-Ala-Arg-Lys(Boc)-Gly-SCH2CH2CONH2
z.Noc-[Lys(Boc)41,59]-HBs(40-60)-SCH2CH2C0NH2 (3)
Arg-Asn-Pro-GIn-Thr-Gly-Glu-Glu-Met-Glu-1 le-Pro-Ala-Ser-Lys(Boc)-Val-Pro- Ala-Phe-Lys(Boc)-Pro-Gly-Lys(Boc)-Ala-Leu-Lys(Boc)-Asp-Ala-Val-Lys(Boc)
[Lys(Boc)75,80,83,86,90]-HBs(61-90) (4)
0 5. -ty'y y F'3f$#&^x’7 Kf-±-XT/>
o-S-EB-^EI 4 ii/Tto ^y>
F"#0#^;t 7 7* 7 -f K • z ^ * y X 7 X x' til <0
430A§)^^mV', Boc 7;y«l'o DCC+
HOBt -cfe-CfTfcofc, 7 Kf
±^x7^^ 10-15%
KO£•/£«£ OETEvmK -<77
S C* Noc) ifTT^Xf
hplc ct vmmi-zztX'X'Z
tz-3 HPLC ##0 7 ; yli: Boc-ONSu
-C Boc
ofc, ts:&, i Noc Sit, HF ^
itz,
F'7tt — xt/v:t, y v — = 7-At=Fy^ .̂7* — /v
m#&T, uss-csas-t
fr>^.ri/Z7 7 KU'!b4 El 6 ;x/7x7ET,
—17 —

it 219
(1) (2) (3) (4)(Boc) 1 (Boc) 4 (Boc) 2 (Boc) 5
Boc ll .</ -Til SR /Noc ■■■■"•■<39] SR iNoc -»~60l SR H |6l'^;y;;^90| OH
HE
AgNCh-fHONp AgNOj-rHONp
/Noc EE
AgNCh-rHONp
TFA, HPLCHI OH (5)
TFA.
I Boc-ONSu J Zn/AcOHHPLC
iNoc |16'/-; '< . .,'/ lo]OH (6)Boc-ONSuZn/AcOH
TFA, HPLCHI OH (7)
m 6. HBs (1-90)
KLGco-C-tfV y -<7>
7-^3 ti, b P7 = y-/^4T, i"
VO'<7 - h p y ^K0t£{£-*7vw*, -<7> K
4 6 me L, & E HBs (40-90) , KJfri
hplc -cm# tt, c 6
EZhtZo £.f&%d.kO Boc B9-7
#Z" HPLC 50% <0[sl4%$-C&of:,
Boc g^SA LfcOt), * Noc 2£*$i&/gfcgtt'&Si
L, F2 LE'B-S-tir, b"6&*#%0 commoitz$:i$ti 60% -e&ofc, e6i'-*<y
f- K 1 45% -C^y^- K 7 [HBs (1-
90)] t^X'^fZo -<y*K 7 <D fcfcfg&Z b
V 7/Vzh pLtzi><D<D HPLC oSiti^y-
7 &*gO HPLC, -f 7T y£&7 t=" b y
7 7 -f -<7> b'4S^FL L-C, 10%
mo JfcSF-C HBs (1-90, 40 mg) 6
Gtfti&(iT i / g£##r, %% HPLC, 'f ± y%£|7 p T
b /77 / -, 1H-NMR x^^tm-o^m^L
tlol' 7b‘ Boc-[Lys (Boc) 3, IS, 19,23,38]-
HBs (l-39)-SCH2CH2CONH2 (39 ES) 6 [Lys(Boc)
41,59,75,80,83,86,90]-HBs (61-90) (51 S^) O ^
7> K6?
%&, tmoUl' HBs
(1-90) 6(1Scfc^E1*S2:*C
100 mES^Olgd^irELl ) 6-to6,
-b y / y b gl
HBs (1-90)
(^)m 7. HBs (1-90) <D HPLC
il 7 A : Cosmosil 5 C18, 4. 6 mm X250 mm, iSttifS : T -t b - b ') A, / 0.1% b ') 7 ;u * 3 SESStKzSIS, Sig : 40’Co
S^ibO^EoBfifrU, COL 5 5 icil,
—51D 0^7> K£j£©(lgfc&.&^6i"S*'e&Oo &
^-<yf- b'&mBf A#%Pn@oo6o;i,
<rop7. b-c*%. m$Ti%-<7> v^mx-tzms
(l4)t,c/LOC 6, Ifog
6, 2,000 Rtf' p, 177F311
ffiiO &t5 L-C^’ < So ttz, l^C
20 v* 100
-18-

CO
g
I-CO
£
28
n
oooooooooo oooooooooo oooooooooo oooooooooo oooooooooo oooooooooo • ©<>♦□■<]*«>► oooooooooo oooooooooo oooooooooo
CJ X) 0-0 <U M-H box
8Io§
8
oooo 88
o oo oo oooooo ooooooo oooooooo ooooooo oooooo ooooo oo oo 000
AI
t\
20
8o
-I
0 OO 0o 0 0 o o0oooooooo# o oooooooooooooeo o oooooo0000000*00 o ooooooo000000*000 o oooooooo00000*0000 o ooooooo0000*00000 o oooooo000*000000 o ooooo00*0000000 o O 0 o o0*00000000 o o o oo#000000000 A o o o1 ~o ort x> u -o <u u-. box) *h in tfs
V\2y <
I 4 y Ik m y Xiv Q y y 6tV It =x m N JS y HE % •4
-y4nL<_
7cdVtvu
<X
Ny
V4?m
H KekQ >0
8E
U-I4<y
4 YQ8
%4a
4;
li
AJ5%
b
4t
•wIty
£ <X y 11 St 0 % m «IN >0 y •m prV US lO 5 4a A v if >0 V G
A)y V H u u. J, Q .¥ •R y % if Q AK> tK ■Vf •si AN It Mr Rti BE *< <: u 4 -sH •TO V y !i Kvc3 "A AH •vt M J A$N w iO >J
4a K 0 v4n to \If V AH Q M H g j, CiU- g K NaBig AO r•I'X y Q *J 4a #g iQ e m 4
*14lit •R >k 0 Q 4v g 4 >v o if y if K %*4. UK w _) ira AO y 10 f: % y s~s if 0
E y if V l £ e E % Au R V A E xz 4<It. & y V HE 4 Tx E •JJ y ■X Q n •y m
s X xz if if•VH 7 u E w >J •R y Q yy A •H '( -R # Ira V JJ iW 4a tf\ VJ *9 In 4 -j AJYjgrt 1 V 4a y N h 4 E ->p s ‘•R w y •RV < ■H u y if # 0 y ti # AJ •E % 4a m
oT 4 to m g <3 u. m >J E y Q if Qoo . <Q II 4a 8 N g J ik K> R A u. &
if AN w m "f a % 3 if > o 0: /-X A y eJ U- Q if O s k w iB Y 4<? if h w *AJ« y Q
\k HE g <X E -4 if if N e y U- & Q X
u% y N y y 1*3 1 Q 2K 0 L> AQ Al tv 3
S’ o Q 10 AJ y if >J 8S *£N if y Ilk % „ HE®i iO o y M AI y ‘x „ Q -N H u Q y 104x< •R \ •E ih K> A M -W E O ne r\VU if
■R M 4a & y y y $ W E Q xz m u y1 ) o if y u _j X n b 1 DM •V y CO CO '•R «
•ii 'ju eK if iO AJ Cx •It !i 0 y E -VJ e•Cl 4a AJ M S AJ y #J u. •ii y AH he sNK '-R AJ "IN it >J n J, y 6 AJ 11 y rO e
Se y X J «fx E •*-t y tK 4R dll y HI y #y 0 Q O' 'f if ‘if X Q Cx IR if wm HE O 4a -VJ y 0 m aQ 4a y 0 m-
4a *0 lis co 4H y 'if s y 52 tO 4a4a i Q AJ •R y iO Q « y 52 4a s E tK» y y j?
52 aQ z- m Rl d? M 4a y f # Q ■K »j •CN HE y K xz y if iD ■E ns E %E „ •*-< yQ Q It ASU y U. •<IU Wl S9 % i Q m lO M? 0(«E 4< -VJ u y aQ Q 4I,D| U- y >J J, o HIm y 4a k s Q y if <cx Rl "-R s y Wi A;< AJ y „ •Fn
an # X « vO ? m 0 y w 6 Q y m iO-vj < J if fiS y % y g f4 U- Y K? w AH AO YfO •R V *rx AK H y m y iO % 8K < m
§ y 5 J) 'll AJ J »v AH *rx u k(N y 89 m y 4a 0 E A % i
AJ V? 5N 4a J, •*-t J!p< H Y s y it? ‘•R 1 y
y y "f K 0 iK 4a 4% y -J 1R >v y yB #4 o >j y y Y Q Q * Q it m ><? . j Yg if lO AO K a\o „ #R li < rO rt if 4a j m y 4 e iO■N \u y if X «rx £ K A y y % AO•>/ ik y Q y y 4a A 1 K 4 >0 o 4a 4a if AJh y y H? if B it m H tv B K 4a XlU y u if a #3, 5y Y AO y y ii 4a Q y AJ tx •' Q li m y *AJ Cl,
*tiU- if o tm
V Q •R •R „ #9 •R # 1 Q m 318 Q y ik g CQ oE E ej H y m y B •A CO % g 4a E Q y 4 i)
if 89 -,P AJ e y 8k Q B iO M g *x i> 44 Y tO >Vm y HI e HI
•lr* 4% « 318 g y N n # e m • j tx
B9 44 Ex Tx Q 4a -iAu # *x '•If :r u K E X O 4 K r< J8k m xz iO ’5 A if $ Q 4a -v! h E y -rP In y Q H
B ,<Q u -R -R
'•it A U- y 0 N AJ Q if y 4a ¥: 33 m fiif B
m “H y •R AJ 4x -j K> li y m 4X '< # # ex Q if5^ '•R •*-< a y tM y •R 1K Q u Q if A Q y. AK W Se
-VI y Y HE £ ti y «rx *x y 3I8 O m y „ A m o i|y l|N 4a f HI y if 9% $ Q y »r AH # m iO ik y i IJxiO ‘•R y iO % •V: Q 318 it N •—t m u. AJ y H AH 3* hAiU HI •4x y y Q AJ y •Vf # y Q cO A ik u. m •Ify A if H -,P y K y u. •R 0 1 y ik m "A: o 8
HE UN u *tn if n 4a B .< \k J3 DC; tv Y y y CQ CQ
if y 13 •it an if Q Q <4: if y if \ Y i) li
y # s y Q y U- g y 4% Y ii y 0 Q Q xv n 4 AJB 1 u m >j *AJ *x y Ax u y <rx H y HI Q aQ
to

221
-v -4 {7 t= =2 v fcf — & *C redW $ fl, 96 ^ 7 h
6 Ert-g-J&'C #- 6 £rfclz£ zl"C i'%. 20 ^So&SL^SiE^r K 96 SEar 1
SW@m-cmB±^5,0 -C7> L#*b,~
*5Lvizkkst 10 *oKKf-e^U-C/w
F#0##mfF^TLf2
ot), W5I<h, K
'&%:<D£&Wj&&mz.it'<,
zi^vk i/io #mc#[#-T(f 6c 6
&t> 9 C
&Jgs-* 0, 10 K EEo/MSdK^E^ralo
5 2}-^L,
Lri>L,6T51-it,
5 ftSsSESuri'1 ttzt il-UMXh 5 o -t: y / v hirS-a
5 fc, BSSarfO^L/zS^EE^y^-
PS&oarS^5:&frt jitt*/c P)7^i'-e^,6 5»
S^7> KcoHM^^v-cst, fH-, ^(D-<yy K^r,
t'e cl 5 tcfco~Ci~tz0 i5v'4?
5(E. ,|''^Bf^tl7>f 7‘7 >J - i t"CfiJ^L, 9f % %ii*bS £ <k^Bjlg ftzh-ZtbbDo -r
z><k
c$fi, -rofijffi^'^^b-t^it',
LxmmzKtz^&n-c<mmKm£*3\
fc§gf3jg^,tH'in&.ZzkA'-smktzZX'&ZS,,
^fPtcaiu:!), #-6mznvB^o
m&no&m^f&w-tztzbb\^&w£?-& t\ x owmi-n
bftZX 5 K:%a-0&6 5,
4#6
ciat:, 4b#e^mam^^og^m^c%.om
6H14i§^oSJfcti *£nom3r£J 9giK££>
£ ^
1) Chun, R., Glabe, C. G., Fan, H. : J. Virol.,
64, 3074-3077 (1990)2) W u, C.-R., XV ade, J. D., Tregear, G. W. :
Int. J. Peptide Protein Res., 31, 47-57 (1583)3) Khomura, M., Nio, N., Ariyoshi, Y. : Agric.
Biol. Chem., 54, 1521-1530 (1990)4) Tam, J. P., Heath, W. F., Merrified, R. B. :
J. Am. Chem. Soc., 105, 6442-6455 (1983)5) Clark-Lewis, I., Moser, B., Walz, A., Bag-
giolini, M., Scott, G. J., Aebersold, R. : Biochemistry, 30, 3128-3135 (1991)
6) Schneider, J., Kent, S. B. H. : Cell, 54, 363- 368 (1988)
7) Nutt, R. F., Brady, S. F., Darke, P. L., Cic- carone, T. M., Colton, C. D., Nutt, E. M., Rodkey, J. A., Bennett, C. D., Waxman, L.H. , Sigal, I. S., Anderson, P. S., Veber, D. F.: Proc. Natl. Acad. Sci. USA, 85, 7129- 7133 (1988)
8) Wlodawer, A., Miller, M., Jaskolski, M., Sathyanarayana, B. K., Baldwin, E., Weber,I. T., Selk, L. M., Clawson, L., Schneider, J., Kent, S. B. H.: Science, 245, 616-621 (1989)
9) Kent, S. B. H., Alowood, D., Andrews, J.L., Bergman, D., Brinkworth, R., Jones, A.: in Peptides 1990 (eds. Giralt, E., Andreu, D.),pp. 172-173 (A* 7. 9--- b y v' a V'^C^f), ESCOMScience Publishers, Leiden (1991)
10) Miller, M., Schneider, J., Sathyanarayana, B.K. , Toth, M. V., Marshall, G. R., Clawson,L. , Selk, L., Kent, S. B. H.: Science, 246, 1149-1152 (1989)
11) Swain, A. L., Miller, M. M., Green, J., Rich,D. H., Schneider, J., Kent, S. B. H., Wlodawer, A. : Proc. Natl. Acad. Sci. USA, 87, 8805-8809 (1990)
12) Jaskolski, M., Tomasseli, A. G., Sawyer, T. K., Staples, D. G., Henrikson, R. L., Schneider, J., Kent, S. B. H., Wlodawer, A.: Biochemistry, 30, 1600-1609 (1991)
13) Nagai, U., Sato, K.: Tetrahedron Lett., 26, 647-650 (1985)
14) Baka, M., Alawood, P. F., Kent, S. B. H.: z« Peptides: Proceedings of Twelfth American Peptide Symposium (ed. Smith, J. A.),
15) Kaiser, E. T., Mihara, H., Laforet, G. A., Kelly, J. W., Walters, L., Findeis, M. A., Sasaki, T. : Science, 243, 187-192 (1989)
16) Grandas, A., Albericio, F., Josa, J., Giralt,E. , Pedroso, E., Sabatier, J. M., Van Riet- schoten, J. : Tetrahedron, 45, 4637-4648 (1989)
17) Yamashiro, D., Li, C. H.: Int. J. Peptide Protein Res., 31, 322-334 (1988)
18) Hojo, H., Yoshimura, S., Maegawa, C., Ai- moto, S.: in Peptide Chemistry 1990 (ed. Simonishi, Y.), pp. 23-28, Protein Research Foundation, Osaka (1991)
19) Aimoto, S., Mizoguchi, N., Hojo, H., Yoshi-
-20-

999 S6S as & s Vo 1. 37 No. 3 (1992)
mura, S.: Bull. Chem. Soc. Jpn., 62, 524-531 (1989;
20) Fodor, S. Read, J. L-, Pirrung, M. C., Stryer, L., Lu, A. T., Solas, D.: Science, 251, 767-773 (1991)
21) Houghten, R. A.: Proc. Natl. Acad. Sci. USA, 82, 5131-5135 (1985)
22) Ruggeri, Z. M., Houghten, R. A., Russell, S. R., Zimmerman, T. S.: Proc. Natl. Acad. Sci. USA, S3, 5708-5712 (1986)
23) Gey sen, H. M., Melon, R. H., Barteling, S. J.: Proc. Natl. Acad. Sci. USA, SI, 3998- 4002 (1984)
24) Geysen, H. M., Barteling, S. J., Melon, R. H.: Proc. Natl. Acad. Sci. USA, S2, 178- 182 (1985)
25) Papsidero, L. D., Shue, M., Ruscetti, F. W.: J. Virol., 63, 267-272 (1989)
26) Schnorrenberg, G., Gerhardt, H.: Tetrahedron, 45, 7759-7764 (1989)
-21-

Tetrahedron Letters, VoL 33, No. 3, pp. 363-366,1992 Printed in Great Britain
0040-4039/92 S3.00 + .00 Pcrgamon Press pic
TOTAL SYNTHESIS OF NEPHRJTOGENIC GLYCOPEPTIDE, NEPHRITOGENOSIDE
Tadashi Teshima, Kiichiro Nakajima, Minor! Takahashi, and Tetsuo Shiba*
Protein Research Foundation, 4-1-2 Ina, Minoh, Osaka 562, Japan
Key Words: nephritogenoside; glycopeptide; allyloxycarbonyl group; palladium complex; nephritogenic activity
Abstract: Nephritogenoside (1), proposed as a nephritogenic triglycosylhenicosapeptide with asparagine N-a-
glycoside linkage, was first synthesized by the coupling of triglycosyldipeptide active ester with the nonadecapeptide
(3-21) using allyloxycarbonyl group as the final protecting group. A {3-anomeric glycopeptide was also prepared.
Nephritogenoside was isolated as an active principle of nephritogenicity from the basement membrane
of rats by S. Shibata in 19811X The structure was determined by S. Shibata and his collaborators in 1988 to be
a glycopeptide in which the trisaccharide composed of three glucose moieties is linked to the peptide of 21
amino acids via N-glycoside bond on the asparagine residue.2) Although syntheses of the trisaccharide part connected with an amino acid or a short peptide have been reported,3) a total synthesis of whole structure of
nephritogenoside has not yet been accomplished. In order to confirm the proposed structure and to elucidate
the biological activity in molecular level, we performed the first total synthesis of nephritogenoside.
10 15 20lle-Ala-Gly-Glu-Asp-Gly-Pro-Thr-Gly-Pro-Ser-Gly-lle-Val-Gly-GIn-OH
Structure of Nephritogenoside (1)
In our synthetic strategy, allyloxycarbonyl (Aloe) group4) which is removable by palladium complex
under neutral conditions was chosen as the final protecting group of amino and hydroxyl groups, taking
account of general instability of glycopeptide under acidic and basic conditions. No existence of basic amino
acid residues in the peptide chain makes possible the free peptide to be coupled with glycosyl moiety. The
-22-

364
peptide used in this study was synthesized by ABI 430A peptide synthesizer. Boc group was used as protection for a-amino group, and cyclohexyl ester for co-carboxyl group of Asp and Glu. The final deprotec
tion and cleavage from the resin were carried out by the treatment with HF - p-cresol (8:2). After purification
by HPLC, the peptide obtained was directly used for the coupling with the glycosyl moiety.
Heptaacetylisomaltosyl fluoride (2) was prepared from isomaltose by peracetylation followed by treatment with 60% HF in pyridine, and then coupled with 2,3,4-triacety Iglucopyranosy 1-a-azide (3)^b) to give
trisaccharide azide 4a as shown in Fig. 1. First, a formation of N-glycoside linkage of this trisaccharide with
asparagine moiety was attempted. Thus the compound 4a was reduced by catalytic hydrogenation using Pd- C, and then coupled with Aloc-Asp(OH)-(yBu5) to give an anomeric mixture of N-glycosides 5a and 5(3.
Under reduction conditions, an anomerization at C-l carbon occurred to give a mixture of diastereomers which
were separated and used for the following synthetic steps respectively.Replacement of acetyl group in compound 5a or 5(3 with Aloe group gave 6a or 6(3, which was
converted to N-hydroxysuccinimide (OSu) ester 7a or 7(3 by TFA treatment followed by active esterification
as shown in Fig. 2. However, the active ester 7a or 7(3 did not react with the eicosapeptide (2-21) at all but
cyclized itself to give the succinimide derivative.In order to avoid the succinimide formation of the active ester 7a or 7(3 during the coupling reaction,
we next tried a coupling reaction of glycosyldipeptide active ester with free nonadecapeptide (3-21) as shown in
1) Hj/Pd-C
2) Aloc-Asp(OH)-OtBu HOBt,WSCD*HCI
-O
OAc
,NHCO\h2
IAloc-HNCHCOOf Bu
Aloc-HNCHCOO/Bu (5tt) 13.3% (5(3) 68.3%
Fig.l
-23-

365
AlocO
OAlocAlocO
1) EfeNMeOH-H,0
OAloc1) TFA
OAloc 2) HOSu.NHCO WSCO-HC1
2) Aloc-CI DMAP a: 33.9% p: 23.9%
AlocO OAloc
OAloc | Aloc-HNCHCOOf Bu
Peptide(2-21)
r—oNHCO
No Coupling Product1 ch2 ------OAloc |
Aloc-HNC HCOOSu
(7a,7P)
NH-AlocOAloc
Fig.2
Fig. 3. The glycosyl dipeptide can be prepared from glycosylasparagine and the protected proline residue by another coupling method rather than the active ester method. Thus, the compound 5a was treated with TFA to
remove r-butyl ester and then coupled with H-Pro-OrBu by l-(3-dimethylaminopropyl)-3-ethylcarbodiimide
hydrochloride (WSCD-HC1) and 1-hydroxybenzotriazole (HOBt) method. Acetyl group of triglycosyldi- peptide (8a) thus obtained was replaced with Aloe group to give 9a. r-Butyl ester group in 9a was cleaved
-O
5a,5p1) TFA
2) H-Pro-OfBu HOBtWSCD-HCI OAc
Aloc-HNC HCO-Pro-O t Bu(8a, 8P)
1) TFA
2) HOSu, WSCD-HCI
NHCOI
ch2
a: 97.1% p: 97.4%
(9a, 9(3) OAlocAloc-HNCHCO-Pro-OfBu
1) Et3N MeOH-H20
----------------- »2) Aloc-CI
DMAP
a: 42.0% p: 40.9%
•O
-NHCOI
ch2OAloc |Aloc-HNCHCO-Pro-OSu
(10a,10p)
1) Peptide (3-21)-------------------------- Nephritogenoside and its p-Anomer2) Pd(PPh3)4. PPha a: 33.7%
dimedone p:23.5%
Fig. 3
-24-

366
with TFA and converted to the active ester (10a). Finally, 10a was reacted with the nonadecapeptide (3-21)
to give us a desirable protected nephritogenoside without cyclizadon at asparagine residue as in compound 7.
After deprotection of all Aloe groups by palladium complex, the crude product was purified by HPLC to give free nephritogenoside.6)
Moreover, the (3-anomer of nephritogenoside7) was also synthesized from 5(3 by the same manner as
that of nephritogenoside. Biological tests of nephritogenoside and its (3-anomer are now being undertaken.
In this study, we established the general strategy for the synthesis of glycopeptide with N-glycosyl
bond of asparagine which is linked to a relatively long peptide chain: (i) The glycosyl moiety is first coupled
with asparagine residue, (ii) The glycosylasparagine in a protected form is coupled with amino acid or peptide9), (iii) The glycosylpeptide thus obtained is coupled with the longer peptide by the active ester method,
(iv) The Aloe group is used as the final protecting group.
REFERENCES AND NOTES
1) S. Shibata and K. Miura, J. Biochem., 1987, 89, 1737-1749.2) a) S. Shibata, T. Takeda, and Y. Natori, J. Biol. Chem., 1988, 263, 12483-12485. b) T. Takeda, M.
Sawaki, Y. Ogihara, and S. Shibata, Chem. Pharm. Bull., 1989, 37, 54-56.3) a) T. Ogawa, S. Nakabayashi, and S. Shibata, Carbohydr. Res., 1980, 86, C7-C10. b) T. Takeda, Y.
Sugiura, C. Hamada, R. Fujii, K. Suzuki, Y. Ogihara, and S. Shibata, Chem. Pharm. Bull., 1981, 29, 3196-3201. c) T. Takeda, A. Utsuno, N. Okamoto, Y. Ogihara, and S. Shibata, Carbohydr. Res., 1990, 207, 71-79. d) A. J. Ratcliffe, P. Konradsson, and B. Fraser-Reid, J. Am. Chem. Soc., 1990, 112, 5665-5667.
4) a) H. Kunz,'Angew. Chem. Int. Ed. Engl., 1987, 26, 294-308. b) Y. Hayakawa, H. Kato, M. Uchiyama, H. Kajino, and R. Noyori, J. Org. Chem., 1986, 51, 2400-2402.
5) Aloc-Asp(OH)-OrBu was prepared from H-Asp(OBzl)-OH through three steps. [1) Aloc-Cl, NaHC03, 2) (CH3)2C=CH2, H2S04, 3) NaOH, total yield: 75.4%]
6) Amino acid analysis**3), Asp(2): 2.00, Thr(l): 0.92, Ser(l): 0.91, Glu(2): 1.96, Pro(3): 2.90, Gly(6):5.91, Ala(l): 0.96, Val(l): 0.58, Ile(2): 1.56, Leu(l): 1.00, Phe(l): 0.98, NH3(2): 2.35; PD-MS^b),M+H : 2471.3, M+Na : 2493.7; iH-NMR&O, 0.78-0.95 (24H), 1.05-1.23 (3H), 1.19 (3H, d, 6.3 Hz), 1.37 (3H, d, 7.3 Hz), 1.35-1.60 (5H), 1.70-2.48 (24H), 2.75-4.55 (59H), 4.92 (1H, d, 3.7 Hz), 5.60 (1H, d, 5.4 Hz), 7.20-7.39 (5H); HPLC&U: 15.7 min.
7) Amino acid analysis83), Asp(2): 2.00, Thr(l): 0.95, Ser(l): 0.97, Glu(2): 2.04, Pro(3): 3.03, Gly(6):5.87, Ala(l): 1.04, Val(l): 0.57, Ile(2): 1.50, Leu(l): 1.01, Phe(l): 1.00, NH3(2): 2.67; PD-MS8W,M+H : 2471.6, M+Na : 2493.9; iH-NMR&O, 0.78-0.95 (24H), 1.05-1.20 (3H), 1.19 (3H, d, 6.4 Hz), 1.33 (3H, d, 7.1 Hz), 1.35-1.60 (5H), 1.65-2.50 (24H), 2.75-4.55 (59H), 4.89 (1H, d, 3.9 Hz), 4.98 (1H, d, 9.0 Hz), 7.20-7.39 (5H); HPLC8d): 15.7 min.
8) a) Hydrolysis conditions: 6M HC1,110°C, 22 h. Under this condition, only about 60 % of Ile-Val in the peptide sequence was hydrolyzed. Theoretical values of amino acid ratios were shown in parentheses, b) Plasma desorption mass spectrometry. The calculated molecular weight, M+H: 2470.5, M+Na: 2492.5. c) 8(ppm)(270 MHz *H-NMR in D20). The peak of HDO was used as the reference (5=4.70 ppm), d) Retention time of HPLC (Cosmosil 5Cig, 10 x 250 mm, CH3CN-H20 containing 0.1% TFA, gradient 10 - 60 % CH3CN (25 min), flow rate: 3 ml/min, detection: 210 nm).
9) The C-terminal amino acid of the glycosylpeptide is recommended to be either Gly or Pro to avoid the racemization in further fragment condensation.
(Received in Japan 17 October 1991)
-25-

430
*4¥£XM 63(10), 430—437 (1989)
# m*, s ^5**
TU5o CCD#A, y bX&6ttb\C,*wra, mmm©^^y7 ncwu-c, >e©-;eM>H:B:tiv££Eir6
ammmy°fT--u, -e©mrn#e%3a /06
u/:o c©Kfsi±, 2 mm 1 ^E#©KfGtf3C6^T&, $/:, ccDBI&te. MS W'S-teS/vXKjjVg£$TM^-Sti:k
o--jj(D?m£K)bmiicftirzmmjm:-'fti£-x$'^t^r)<$®iZtt^r\,'tzo
1 12 C&ic
yofr-t'ii, xck, 7 ncfpmur, ^t"7>6 O c tl t X 1C, ■ 7" n 7- r —Hr' #&& 5 ^ ©.Wtfffll'bnt^;©(i !5<hA£fr* *<Dl})\\&m\\ UcbOX&Zo tztx.lt, (W, SM]©fi^
7 T ~b'©?W-1>, 7 -X© ®[ & i£&iiiti& i/ y- y3) ^Mfei/y - y*\ f-vutfijc
f y/<^ 3J£E$i)|$/jlh (DtzidCD chillproofing enzyme 6 U©^vq y©#m»\ < left###
UT©yofT-4:©#m^
u, <e©a!Rf5-e*5^siS^jffl-rn{i, r^i
A^^X7 K&&9&MT6 LT,
7 K©^EA<"maw, c(Dm\w, &&<Dftffx (i. m^©&,4, %##%, m*ui<Diji&xte. 6bmm^yf vn&mtzicGWi 3-%t\ct£z±m&ztiz>o
7°DX7—\£fr, &ZftM(D&ftTXffi&5Lm*fto ■- (h (2 IV < Xn 6 til b ^1T & *5.» 1901 ^1% Sawya low7’ fi,'' y '< V rei(nmit'&\c-f a 7 7 --t/£ jjlI;l.TiNli tX'kif
Ictifcob, t°-? x7^ y (huf-tdftioiyl^?-©? y/<"7^3 C U/Co -Er L"C, Bergmann
bit, yiCct^t, N- 'O1'/4 yb-7" y -y^i^T■=■ y K8) *5, $/:, 4-7 h y -f'y yfciot, N-^y
7* 4 /1/-L-7 p y -y;i/T- V K9) c
-/y 7-7-f y/J-:EK^;/j^7"7 KfrEixlC -Cf65C6^JiEWcoi:©m, X4XhyXyy/fy 11' *-©@#7 ; / 5 Arg63 t l- y 'y y, h61'lt l- h y 7” h 7 r v£©3W\ 7“7 7f4 ylxj-K
L/cfjo^O*. 7 yy 4 7 y -y y v,jP t hM'f y -y
^ y y^^©^ss&^7°7 k©^ei3_15) tetficb.c©mAK^WJMTr&5C60, 7°nr7 —\£
u j; a -(x 7 K^EA<-&«h g $ ^15
&i\Ol-oX%fZoMitsa4i7,DT7-^fi,
il7#Df7-fef{Cj;5y'^X7 K©#^K^(C#f5
2 7*0x7-if©ti07X^##m6,^-B-fPffl
#%E© ^ 7° 7 le ft? ^ 7s D r 7 - -te' © #,$U (C^r<k7ic, 6mAK1&<k u 7 E#
ll,-C0-NH-Rz + Hz0 =z=R,-COOH + H2N-R2 ( | )
z(Dm7kfrBixfctm&Kfcfrb&tz¥ffife$t.$:, 7nt\* Ahyd cb^cond <h ^3 ^ <h , 5C 2 •p •S o
- 11 [R,-C00H][R2-NH,1y Aeon,, [R,-C0-NH-R2] [Hz0] u;
t Condensation Reaction of Dipeptides by Proteases
* Tetsuo MURO ±WtfttLJLmVK)T\ &VHt¥3HWfteL St® (536 7E1 - 6-50)Yoshio Tominaca f-1 .h $8fi JSii?
$W-tXM Vol.63
— 26

431
[R.-CO-NH-R*] [R,-COOH3[R2-NH2][H20]
[R.-COOH] [R2-NH2][H20] • Khyci
Ri-COOH t R2-NH2/>>6lh
3SLT, j: D T Ri-CO-NH-R: eWUX
'&Z>tzl6\at, cftb<Dg£’E<DS$£3E*bzttblt,
2.1
;d* 4 y /7 yt/ d"' -=• ;u -L-T vi/4- —vu -L-a 4 i/ jut i K©
Oj&n#, d66u(i, +^& b y 7°-y y-^-y--^ y y y;cz3N-^< y yl/d" 4 y y: — vtz-L-7 i-^T 7 — yu-L-n>r yvu-/y y yo^jgJcBSicfctvr,
L/c (^ Do
1
# ;u d< 4- yb1c^> (M)
T ; y&%# (M)
(#)
Z-Arg Leu-NHg b y yy y=) yx°yx°yf yb>
0.05 0.05 0 00.05 0.2 5 80.05 0.5 11 160.05 1.0 40 490.05 2.0 49 51
Z-Phe Leu-Gly -f-t yyyc) ^ 4= b y7°y yd)
0.05 0.1 14 —0.05 1.0 64 —0.1 0.1 — 140.1 0.2 — 180.1 0.4 — 270.1 1.2 — 52
a) [E>0.1mM, M^-?H >l»ISffiiR (pH6.5), 37°C.208.? rase
b) CED=10aiM, KS?@«iK (pH5.5), 0.05M KCN, ImM EDTA. 37°C, 20l$|S]JglS
c) [E3=10jt/M. h V a-?L/4 >86@«?K (pH7.0). 37°C,2on5rase
d) CE] = 0.2mM. h V CpH7.0). 37°C.20D5iaiSlS
wmf§#0WOK j; 5ck l'AmtC"Ol/^T(^, 5o /c<k7.(dL Laskowski,Jr. 4-* b y Ay yic«fcs N-^yyvi/^ + yt> yud< —yU-L- b y 7° h 7 T —A/-? 'J y^T S K ©
m 2 y f;u*;uAT ; K©»1#)
y > f yt/d;yt/AT ; K
($^)
-&I&# (^)
b v 7°y y=) /N°VX°zf yb)
0 ii 1610 17 2620 34 3730 39 5250 46 7660 20 6
* yu,-K 4. •;/)&©• (0.05M Z-Atr)T < (0.5M LCU-NH2)a) [E3=0.1mM. b V >KSffi«c (pH6.5).
37°C. 20B$iaiSlCb) [E3=10^M. fttWraffiiR (pI-IS.5). 0.05 M KCN.
ImM EDTA. 37°C. 20n?|^Sl5
7%m! (85 0 1,4-7'f yyM--yi/®mjjoT(iizabr^mtu<0, £ c
6^L/:o $/:, bVAyy&5^(i/<^4 y\c£Z> N-^yyjujrJr'yftju#—ju-l-ta'# =-jU-\-u4 i/J^Ti K CD^KlLlC&iSft’WW^to®
mm6 L. 50# y / -7- /M-. yu a r • K
(Dtr-tt-e. zommucttM l-c 4 ~ 5 ir^c^^w^-LTuo ca 2)0
octit, titfM,
2-2 tb7'1M
< ff 7 fz a6 ic (tg#S<DjliR t> m#/Cy bT&O, 6#
7>4 FOW, -eK A&^^#^4 X. 6
£ t bft5o
Schechter <h Berger20*21} (ft yaB%^Te:7‘
-r&^M y£Djb07K^BfFffl^wtifi‘5K&fz o, mm
b6(,'7#!E±&#L&tsal/co to#,7'nfT-trOjjOA^fPffi^b, ^-4 ©S SP ft JC
^•y5l/7>4 b2)###%6/MC3tl/c"''^o iztTLit, ** b y 7ey ycD-y-7>4 b s% », ^ /
W L, b 'J 7°y y0t^t4 bSj «, #&#% ^ %
(C#«h^7t /:, ^7f'J’>y BPN’, x 7 x —fef, ^7ey y/iiZ
(i, {pfs. < tb, ^7>4 b Si^S4 cDdfe^kO^ti-:/)^ WZ&'O, ctucftfcirZ'*??- KSjtiESchch^lcjfjoA
Vol.63
— 27 —

432
C ft bCDlJUTkfrMfcfcr-^7^4 h 0## (5, *(DZt&f&Kfc\tbMmirZ>zt&-?:£6Zt&
N—<77J\s3r 4- 7# jutf—jl/ -L-7 i-^77-^ -L-O'f 7vl/
7 i KO^-J&tCftt/'T, a- +"& b V 7°7 7(5, N-^‘ 7 V5 vl/;t 4- -7* vl/# — VW L -7 JC. — VL/T 7 — y^kti VI/.-K4-
7vi/^6UT*@^K^fT7^, %7'f V77 BPN' (5, -eft#T&%v\, tfz, •>', b -;, fhv^fK
T, m&$.fc'Z(Dti vurf-:+ 7vi/^0##0 ^ ^ 6 £, <*-4^ h V7°77 T(5^yf- KgmS©WR(5E #> bfttel Vv*. 7 7'f IJ ->7 BPN’ JP^7P-7 7|Cft(/'>-m, e o (cm^ <3 ft a, o iio^KiU^f5^Jvjc^'FjO££ 17-Sco $/:, «-4-c- b V 7° -7 7C0-S-EMIG(C3 -tf-7> /f b S,' CD^ft171
WK^mg)irft^6j:<-%LTftU, It^mi! 7k#T ( / m^##=T ; / (#
3)0 C(D£?\C, ZBrnofS^tiACbtf5-y-7>>f b0##& ft£ 5; < —mf
3 Ij <S a-4- -t b V 7° -7 70-tf-7'lM b Si' 0#%"'
(0.1M)T ; 7J&4>
(0.1M) (^)
Z-Phe Gly-NH2 <10Z-Phe Ala-NH2 12Z Phe Pro-NH2 0Z-Phe Val-NH2 70Z-Phe Leu-NHg 71Z-Phe Ile-NH2 75
[E3=0.2mM, h V *—v u«f >B6S®$( (pH7.0). 37"C. 20S5U3StS
C05C"5(C, SISoiS&Mlc&tt 5-tf-7>4 b0#
-e0^RfS(c^:#ttiZWbfr x&za Ltztt^x, 0 1 «k3 (c, 7^yf b'
(X—Y) &Z>, ZOX.mfoX&Zf- Vy^XT- K (X -Y-X-Y)< &1&ir2>K(5, @m0^7'tb4
b s2<h Si, Si'6 s2 o-eftfft, 6&vx5W
(COOH)X — Y
Protease
0 1 /nr7-k'OgWi^^7,f F (X—Y) 0&&S5;
Vol.63
-%&, x-y (cjm Lxmmvmwj&tti-zc t ti&mT636#^.^ftao C0j:9^#%[:0-;yf b'0A-gc tcmufc-v-y-'M b^<rr-y-%fif^^^ftft(5.'p (C a 0T t #4^0 £ l £ 5 o
3 7‘nf7-t'MI©^i
o f T--W:®R^6L/T, flli'iLl., /v% v-
4 (TA)£;vy7*4:'f7^^
%T6UTmV:f 7,<?###% (PA)(TA/PA) &7$<.l&tz0 C0l±(5, 7 7'*?%f
1 -flU4£>/c <9©^7°f- b'^B%#0#&^i£%& & 0-c, C0li:0A fr-^Mfiimi U
"C, Streptomyces cellulosae Jk\\\lZrtM\'$(£>,AL9\iX o V u -rT--tf(5, {ti>O^M<S®©^ilt^ LT, : "/ ? *-b' 4 7 < , 7^0, "X"kZ 7 7 / ' 7-------7" 7 7
t07)#% $ ft, vf'lMiCffl V VcSMB®0 7°nf 7 —t: £
f- K^-EtCEl/'C, *S©M2T 2> 7s D ^ 7 —b' 0#2ii %A#0^^0Mf4#J: <0 <dtW&t'iiizJx.\c%^Bf^'^mmnc^-mm\czxm^ u, cft^7pof 7—fe' SCC tfo%Ltz0
c 20 -
^ 16 -
^ 12 -
6 24MCWfm (h)
0 2»rDf7-(r'0Ml
# : Streptomyces cellulosae ©££/•??"73 7o r r — -t' (6G£!ti6r!»&). O : «-#* Ml
A : t:' 7 y 7 — -tf {Bacillus subtilis var. Biotecus). ▲ : 7of-f A {Streptomyces No. 1033)
-28

433
4 h'cj^ftST'pt-7—a see <d
•fu 7-7—2 see ©-?7°f- K£j£fCfcttS&l£i|#fS.
~f* a-7 b
fy7<-lc£K>frVrLtzo 4 iCTjkt^om#, (%
; y m JfWlC l~2S1(D-y t: K V yR#%CDx^ -y h
at,7sf- KT&6 Phe-Val, Val-Phe. &j:[f Leu-Met VZ
*S?:^©fPiTJ{C<LoTR«(i SL< #f&bizo Phe-Val *>b£j&L/:f±##icouT, r ^ y EIEE, N^ffir^yiic
«r;yio»i', *CD Sit (Phe-Val) 2 T & D , ttz, Val-Phe, fc«fctf Leu-Met^ bCD^#(i, (Val-Phe)2, (Vr.l-Phe)3 <h (Leu-Met) 2, (Leu-Met) 3 7r&Z> C £. frtdfr
o/;0 —-Jj, SizKter ; y ( y mtt^UK, /c6x.li, Leu-Gly^Sgicm W:#A
Kli, (Leu-Gly): (Plgi) t (Leu-Gly)3(PLG2) && lt(Dm&, ±f5©£7«^ft®ld:£< M&b fZo
#C(C, Plgi t Plg2 J<ot^
T,. flbCDiBgiCD/afr —if CD-ttl <h b /c flf &
E5 tc^-to c®ET, PA id^yiM #-tf4 y^-^fc mwcii$(Df (Plgi+plg2)
E 5 &#&mcD7°of T--t:(C=ka Plgi <h Plg2 <D-fW&
7° n -f T - -t: (PA)(U/mZ)
PLGl + PLG2b)
(/iM/min)
PLG1 + PLG2
PA
7°of7 —if SCC (5. cellulosae) 25 250 10
a-4^r h V 7°'7y 1,000 10 1.0X10"2
yN°y%M y 1,000 4.2 4.2X10-'
f-—t u y y 1,000 13 1.3X10"2
7° d -tf yf A (iStreptomyces No.1033) 1,000 49 4.9X10"2
If 7° 7 —if (S. subtilis var. Biotecus) 5,000 0 0
% 7'f- V y y bpn* 10,000 0 0
7 id V 7° o T T ■—tf (5. griseus) 10,000 0 0
7,af7—If A (A. oryzae) 10,000 0 0
[Ml:7°off —tf (5. griseus) 10,000 0 0
a) i n-b'-t >*i£gtc/?H7i£ £©*b) 0.2M Leu-Gly #© Plgi t Plg2 t©SfiKU!)a&©:to£5vf .
4 2.5% -7^7°^ K(C^y,7enTT--tf
sec <Dfffl
ff-<7'? K
Leu-Gly 2 -Leu -Ala 1 -Leu-Ser 2 -Leu-Val 1 -Leu-Met 1 +Gly-Leu 2 -Gly-Ile 1 -Gly-Phe 2 -Ala-Leu 1 -Phe-Val 1 +Val-Phe 1 +
a) DThynilcai't, Ki:*OMat-^5 7 S y ffijkW© - V t F V >PStt©*to,±Et^®$ir.
b) SIC 1 rf|Hl&iz:*5 it 2 SISK©33 •) ©Wto
Id:, ^^KPCDTn^^n (Plgi + Pi.r,2)/
PA CDMK^I^T-^y <h, -f ot-T-2 SCC
tefi&TFb;! vi/7 yj-tf ^ 1 #{&&/: D CDe 5 C f:o
5 yo^T-iz scc
MrmiA<>mm'r'm\i.<Dis'<-7-y- &,%huc;
tCch-oT^r ©—6Cd: b /: o
co«k9^w 6 aamost&am ic&^s^y
f#ill Vol.63
— 29 —

434
b ©#&£Bj!
5.1 Leu-GIy
Leu-Gly(C*#m^fpm $-£6£ (Leu-Gly)2 (Plgi) t (Leu-GIy)3 (PlG2> 17e£.J&'f t, Leu-GIy
frb Leu6 Gly
TftZo c n b ©WRBOjSS^zB!i^1"£ <h,
(iA07K^#KfLZ 0 & £fricsg-pfricitfr-ft>ij^tzo %ic, cnb©mmis:ic*i-r£p$mM
©^*^,#L/:o ^C©^, PLGI 6 PLG2 0/km/J i/ctiu £ ti&ttmoffi&Kfc&focD
^m&t^kTLht, 0 3 OZ-pIC rc (1 Leu-GIy©# miCfcftLrisio -7 7'* 4 K#OAm^U/:o
0.2 0.4 0.6
0.2 0.4 0.6[Leu-GIy] (M)
121 3 (Leu-GIy);, (Leu-GIy),, Leu & <LCf Gly © $LJ&V)Mfeltto&\Zt Leu-GIy ©#B©S9,i
/fa(tf:«i/fb)=0.65M. 7fi,(*k Id:/f,) =0.016 M, #310= y=2.4XlO*aM-s-1 5£(4)Kt£-3T$iO
A : (Leu-GIy); ©£mz)ii®, • : (Leu-GIy),©^®®®:. O : (Leu-GIy); £ (Leu-GIy), t ©£z3t3J38g©fn (vc).□ : Leu Sf:li Gly ©£M$®, [E]=1.4xl0"5 M
Leu—Gly Leu — Gly
Protease SCC
El 4 7°OTT--<f SCC ©r£t£M (D-7>'f b) (C&M-& Leu-GIy ©£r&$&i£
Vol.63
%(C, C©vci Leu-GIy CD #[£<!: £
\Chlz K), ^©tW^L^to 5 AT 2 ^-T-© Leu-GIy
( A) d#5B©@#am(C# A L, #A#T&£ Leu-GIy ©rSft (Plgi) £6^7 tf vi/ (g) 4)
AE
CCT, Ei A(i, Leu-GIy^43, Ka It, Leu-Gly(A) ©#^©^%%{t© «b7 ^
bS; iS, $/:,
maic xu (a, A7)#^©^7'^-f bs.'6 s,'
©% Vc (i.
btlo o
vc = K[A]V{( K, + [A])( Ab+ [A])J (4)
cc-c, yo, *[el icm'i-#-6o ([E].
El 3 © Vc IcmtZ&iWJ)', c© 5EU IC&XlttZ /AS-fMF.I*£© 1C, ^©—O©%GlC^lj-C^-
7u£o© [A]>Xb, £ /c Id: [A]>Ka 0 v:o7f;]t
[A]>Kb©6&,
[A]/ VC = ( /<a + [ A] ) / V (5)
[A]>i^a©<Lt,
[A]/ Vc = (/Ci, + [A])/y (6)
t, ifi *-') 7.—y yf- yEfC/Xb), [A]/vc
chca]i©no(cmmwwfT^a» lwi#EOT, El 5 a ICTfvir£ 7 (CEtz>SroS!^([ A]>0.13 M)7? [A]/vc~[A]7°o -7 ##©3cA<L0^a(t/GMb)=0.65M<h^$n/Co
© [AK/Ca, £/cte [A]«b«Diz:-7E©S!E
[A]<Ka©6#,
[A]2/ vc = (Aa/y)-(Ab + [A]) (7)
tfcte, [A]<Kb©6 &,
[A]2/ t/c = (tfb/KMtf.+ [A]) (8)
©-rnt[a]2/vc 6 [A]<h©^icmmR#^El 5b IC^fJy)
1C, i£t'&MMX$([A] <0.13M)"C, [A]Vvc-[A]
30-

^5<
[ A ] (M)
5 b(a)6m'jmi&mmit&vz [Aivrc-uiyo .7 Kb)[E]=1.4xlO'5M
WTMfL/:
7°d .y hzctifiTp.£ft, ##© % A «k 0
Kb (t/cMa)=0.016M <hfAlS$tl/CoEl 3 (cXsi VC, 'JL-ISi'C/j; Ltc. vc ojjj — ~/ii, 5£4 (C
Ka(t/c(iKb) = 0.65M, Kb(£ft(iKa) = 0.016
M, V=2.4 x 10”6 M-s"1 £ © 7 ±;E©{5£© b t 'C'&
Gft/Kii'i&JIJl'-CMiW^PlUHullilST^ «9 . llll^/^-Jl-'/iVlc,-k < —% LTI'O C ch o,L/c-^t Gftfto U^L, Leu-Gly
©MSracliltli, Ka (£/cte Kb)=0.65M 6Kb (^^liKa)-0.016M
t>, Wfit UT Leu-Gly ^©5^0, <_©20©M-
m >iz % © m ^Ka 6K,,©^5k)(cm-^6©X)\ k v > /;> xtui, /K@m©mm4:'C^^ $ t? ^ s ^#j©^7>4
b a & ©7?* a y> Mc-r a ci ^©0 t-c-c, a/csbicJaiT©^
£fl o/Co
100 200 300
o.oi r
100 200 300
[Leu-Hllg] ( mM )
El 6 Leu-Gly-Leu-NH2, Leu Gly ©^felKLeu-NHz ©&&©^@
# : Leu-Gly-Leu-NHz 0!il!S.ViWSL (vc),O : Leu 3f:li Gly <D%.&$)&!$. (,vH).CE]=2.1 x 10'5M, [Leu-Gly]=6.25mM
5.2 Leu-Gly £ Leu-NH2 <b©iSo-^IC
0.13M Leu-Gly t. 0.2M Leu-NHz
#"C&a Leu-Gly-Leu-NHz (Plgln) ^r6k ti£ft a6, Leu-Gly LT Leu
£ Gly a Jjli/K^MKit c AWilcmn-Tan cn^^miL-c. 5.1 -c ft g ft ft:.')U) Leu-Gly ©MSKiftt (Ka £ Ki.) ©
mm ^sa#i©tb7'-v-i b icgwTa^e
©to;sa: ic a ± ; i -t wvt ft © « © ^ i: o
tr, {£©K (6.25 mM) ©
Leu-Gly (Kb!T(C&©"C, Plgln ©-frKt/J
&££ (vc) {C&(3:ft Leu-NHz © #ljl ©!£ El 6 ICtf^ <£-?!<:, 50 mM £1
rommmmv, vc £Leu-NH2 cD-mst<tommi, ;ii x 1J x —/ y T y ?? J©iXiTtlilhS^^ Lft^, 100 mM
a h©,V:iril^mM'c(l^,iti:il',lf©^%^^©GnZ:.. - #, -fc$m<D Leu-NHz m:. vc IC%(Z
^Leu-Gly ©2^©^e^<ft£G6, ft£x_(i, 50mM Leu -NHz lr,T: K'C(i, 25 mMU bWMGlUre Vc £ Leu-Gly ©^Kl!<h©[IlKcLb^llW^©
(El 7)u
50 100 150
0.01 _
50 100 150[Leu-Gly] ( mM )
El 7 Leu-Gly-Leu-NH2, Leu &<£££ Gly (D{\:J)})i %}5&;M (vc 6 vH) Leu-Gly ©
[I.eu-NHzl : 25mM (O), 50mM (#). 100mM (A).200mM (□), 300mM (A)
CE3=2.1 x 10*5M
J£C'<2, Vc 6 Leu-NH2 Leu-Gly ©#@]
"Clb 7*if'f b SziS.IC Leu-Gly^, ib7'ib -< b Si’ \C Leu-NHz^, LTiFi^r'S, a Leu-Gly-Leu-NHz ^gic-ra t © 7 a^ra©i$ss:<h LTit 9 ^^naQ
f4'T-i -TS Vol.63
— 31 —

436
V c = V [A] + [A]) (lu + [B])} (9)
CCX, Kit K2(i, Leu-Gly 6 Leu-NH2 #
±iE©ft^$a£t-otztts(Dim’̂ nr*5o[A]6[B]{i, ^n-c'n, Leu-Gly 6 Lcu-NHZ ©JR
y(±,C©^a(C&T(i$
a fca6 {c^©nocD^{t^^T^x. 5 o
® [A]#—&©&£, 3C9 ti.
l/yc={(/C, + CA])/(K[A])}-(i +A2/[B]) (10)
titi*') x-y %f D. i/vc t Vc B ] 6 ©^icmmm# & a, »
X,6 ^6©m
© [B]#-dZT, UpfcCA]<2C, mw>i-e(5, a 9 (i,
yc = (F[B]/{Kl(K2 + [B])}) -[A] (11)
tte*), ccogj&it, vc tiAitortucm&Bsmmttx^Zo tfc, Kt tvtvmmw
teWf iz -Xs 0 X, ©©tiifL 50mM PJLK© Leu -NH2#%TT l/«rc—1/[ B ] 7° □ 7 b frWJfo ttZV, m «l©£&«fc*5 K2=0.008M S$#©3ciU(jai2
(K, +[A])/V [A] = 6.70 x 106M"' -min (12)
c!CT'[A]=6.25X10-3M "££>£„ — ©©#A, uc
£[A]6©RmcJt^§8»^&&ftrt: (07)o t/c, S
HBW^BSb btlteKB] =50mM ©6&, ##©#&
j:»9s:i3*^t»nfco
V[B]/(A,(A:2 + [B])}=2.08x 10"5 min"1 (13)
Ltz&^X, a 1263: 13J; 0 Leu-GlyA\6 LT 0.6M 6U9#^#^fiy=oC© Kx (Dfitit,
5. 1 XyS^tzm&Kfc^lOnbtltz 2~D<Dfm%& K& £tz\t Kh ©fi|(0.65M6 0.016M) ©9 %><D 0.65M(C
Protease SCC
Leu —GlyLeu — Gly
K, = 0.65 M Kb = 0.016M
El 8 7°of 7-4f SCC ©S*fcfe®{4 (^7*tM b) (CMT5 Leu-Gly© ^£t£a 6 ^©W-gBE^C
Vol.63
SWii!rC£oo ^©/IW^cLD.El 8 ^©ffittfm©t7't-( h S26 S,tC Leu-Gly
A'il(=A']) (i 0.65M, tv'+H bS,’6 SV(C Leu-Gly 0.016M X J5 o C 6 /jiHJJ G /j' 6 o tz o
cn b©Srt: £ D,© 7 7'4J- -i b S|’6 S2\ (±, V 7" b S26 Si (Ctiz<T Leu-Gly 6 61' C 6 frfrfr0, COJCtfr. (Leu-Gly)2^ (Leu-Gly)3
c: a c 6 ©^%ma-c^ a 6#/i
iotltZo
6 is tit) ic& ;miG&amir 6 y ° T 7
$6©/:o
6 6 %o„ m%> ^7 7*Df7—tf©m#t. %tiz>&mmbtz-mmn®
Hitnte 6: (cm a a $ns0
(t:a^l'4 JJ2GI l-SPIl)
X M1 ) K.Hayashi, M.Terada, K.Mogi, Agric. Biol.
Ckem., 34, 627 (1970)2) H.Sekine.. ibid., 40, 703 (1976)3 ) B.Foltmnnn, “Methods in Fnzymology,” Vol. 19,
p. 421 (1970, Academic Press, New York)
4 ) K.Arima, S.Iwasaki, G.Tamura, Agric. Biol.Chem., 31, 540 (1967)
5 ) L.Wallerstein, U.S. Patents 995, 824 (1911)
6) xruiE, vof7-b'" , p.205(1983, '->-£,41®* V9 -)
7 ) W.W. Sawyalow, Pflujers Arch., 85, 171 (1901)8 ) M.Bergmann, H.Fraenkel-Conrat, J. Biol. Chem.,
119, 707 (1937)9) M.Bergmann, J.S. Fruton, ibid., 124, 321 (1938)
10) R.W. Sealock, M.Laskowski Jr., Biochemistry, 8, 3703 (1969)
11) M. Fujimnki, S. Aral, M.Yamashita, Protein, Nucleic Acid and Enzyme, 20, 927 (1975)
12) M.Yamashita, S. Arai, S.Tanimoto, M. Fujimaki, Biochim. Biophys. Acta, 358, 105 (1974)
13) Y. Isowa, M.Ohmori, M.Sato, K.Mori. Bull. Chem. Soc. Jpn., 50, 2766 (1977)
14) K.Morihara, T.Oka, H.Tsuzuki, Nature, 280, 412
-32-

437
(1979)15) K.Morihara, T.Oka, H.Tsuzuki, Y.Tochino, T.
Kanaya, Biochem. Biophys. Res. Commun., 92,
396 (1980)16) H.Tsuzuki, T.Oka, K.Morihara, J. Biochem., 88,
669 (1980)17) T.Oka, K.Morihara, ibid.., 84, 1277(1978)
18) T.Oka, K.Morihara, ibid., 88, 807(1980)
19) G.A. Homandberg, J.A. Mattis, M.Laskowski Jr.,
Biochemistry, 17, 5220 (1978)20) I. Schechter, A.Berger, Biochem. Biophys. Res.
Commun., 27, 157 (1967)
21) I. Schechter, A. Berger, ibid., 32, 898 (1968)
22) J.S. Fruton, Adv. Enzymol., 53, 239 (1982)
23) K.Morihara, ibid., 41, 175 (1974)
24) K.Morihara, Seikagaku, 46, 949 (1974)25) K.Morihara, T.Oka, J. Biochem., 89, 385 (1981)26) K. Morihara, T.Oka, FEBS Lett., 33, 54 (1973)27) K.Morihara, T.Oka, Arch. Biochem. Biophys., 178,
188 (1977)28) T.Muro; Y.Tominaga, S.Okada, Agric. Biol.
Chem., 48, 1223 (1984)29) -T.Muro, Y.Tominaga, S.Okada, ibid., 48, 1231
(1984)30) T.Muro, Y.Tominaga, S.Okada, J. Biochem., 99,
1625 (1986)31) T.Muro, Y.Tominaga, S.Okada, Agric. Biol.
Chem., 51, 2657 (1987)
-33

mas:

360 Reprinted from Accounts of Chemical Research, 1992, 25.Copyright © 1992 by the American Chemical Society and reprinted by permission of the copyright owner.
Stereoselective Routes toward the Synthesis of Unusual AminoAcids
Yasufumi Ohfune
Suntory Institute for Bioorganic Research, Shimamoto-cho, Osaka 618, Japan
Received. January 22, 1992
More than 700 amino acids that are the so-called unusual, unnatural, or nonproteinous amino acids have been found in nature in the free zwitterionic form or as constituents of peptides. These amino acids have attracted much attention from scientists due to their important biological activities as antibiotics, metal chelators, neurotoxins, enzyme inhibitors, etc.1 Because in many cases only minute quantities have been isolated and because their structures are unique, they are interesting synthetic targets.2-4
A feature common to many of these unusual amino acids is the 1,2- or 1,3-amino hydroxyl system. In particular, such amino acids are often found as constituents of peptides. Our research group has been involved in the development of new methods for the elaboration of such 1,2- and 1,3-amino hydroxyl systems. Our chief focus has been on the total synthesis of the peptide antibiotics echinocandins (l)5,6 and ga- lantin I (2: the structure has been revised to 5 as a result of our synthesis),7-11 which have a variety of new amino acids. Echinocandins, isolated from Aspergillus ruglosus or nidulans, exhibit potent antifungal and anti-yeast activities.5 The structures of la—c are composed of a highly hydrophilic cyclic peptide and a hydrophobic linoleyl moiety. Galantin I is a metabolite of Bacillus pulvifaciens with antibacterial activity.7 Galantin I has two new amino acids, named galanti- namic acid (Glm, 3a assigned as its primary structure) and galantinic acid (Gla, 4b: isolated as an anhydro form 4a) (Figure l).11
In recent years, great progress in the asymmetric synthesis of both the 1,2- and 1,3-amino hydroxyl systems has been reported. The available methods for the synthesis of the 1,2-amino hydroxyl system are the following: (i) nucleophilic opening of an epoxide with amines, isonitriles, and azides; (ii) reduction of amino ketones or hydroxyl imines; (iii) nucleophilic addition of organometallic reagents to amino ketones; and (iv) coupling of an achiral or a chiral glycine equivalent with aldehydes.3,12 However, several basic problems regarding stereocontrol, racemization, protecting groups, etc. still remain.
Our approach to the 1,2- and 1,3-amino hydroxyl systems is the diastereoselective introduction of a hydroxyl group into the allyl or homoallyl amines 6-11, which are readily available from commercial a-amino acids. These unsaturated amines can be viewed as
Yasufumi Ohfune was bom in Asahikawa, Japan, in 1948. He received his B.Sc. Degree from Hokkaido University in 1971 and his Dr.Sc. degree from the same university in 1976, where he worked with Professor Takeshi Matsumoto. After a postdoctoral position with Professor Paul A. Grieco at the University of Pittsburgh, he was appointed to the position of Researcher at the Suntory Institute for Bioorganic Research in 1979 and is currently Chief Researcher. His research interests include the development of new synthetic methods for bioactive amino acids and peptides, biomime tic reactions. and studies on excitatory amino add receptors in mammalian CNS.
Scheme IEpoxidation of 2-Amino-3-butenol DerivativesNHR MCPBAAXjQ? NHR
OH
6a R-Boc 12a (syn) 12b (anti)6b 2R epemef R - Boc: synZanti - 400
H - Ts: syrvanti -1.6/1 R - COCFj: $yn/arei - 2/1
NHSoc6dR-COCFj
NHBocH-O
NHBocTBSO'
TBSO'
CO/I
useful synthetic building blocks because of the presence of chirality and appropriate functionalities accessible to a variety of chemical transformations. Using the above synthons, their diastereoselective conversions to syn and anti 1,2- and 1,3-amino hydroxyl systems (methods a-f) were examined as summarized in Figure 2. The total synthesis of lc and the right-half equiv-
(1) (a) Amino acids. Peptides and Proteins; The Chemical Society; Cambridge, 1968-1991; Vols. 1—22. (b) Hunt, S. Chemistry and Biochemistry of the Amino Acids; Barrett, G. C-, Ed.; Chapman and Hall; London, 1985; p 55. (c) Wagner, I.; Musso, H. Angew. Chem., Int. Ed. Engl. 1983, 22, 816.
(2) Coppola, G. M.; Shuster, H. F. Asymmetric Synthesis, Construction of Chiral Molecules Using Amino Acids; John Wiley & Sons: New York, 1987; pp 127, 204, and 267.
(3) Williams, R. M. Synthesis of Optically Active a-Amino Acids; Pergamon Press: London, 1989; pp 1, 167, 186, 208, and 304.
(4) a-Amino Acid Synthesis; Tetrahedron Symposium-in-Print; O’Donnell, M. J., Ed. Tetrahedron 1988, 44, 5253.
(5) (a) Benz, F.; Knusel, F.; Nuesch, J.; Treichler, H.; Voser, W.; Ny- feler, R.; Keller-Schierlein, W. Helv. Chim. Acta 1974, 57, 2459. (b) Keller-Juslen, C.; Kuhn, M.; Loosli, H.-R.; Petcher, T. J.; Weber, H. P.; von Wartburg, A. Tetrahedron Lett. 1976, 4147.
(6) (a) Kurokawa, N.; Ohfune, Y. J. Am. Chem. Soc. 1986,108,6041. (b) Kurokawa, N.; Ohfune, Y. J. Am. Chem. Soc. 1986, 108, 6043. (b) Evans, D. A.; Weber, A. E. J. Am. Chem. Soc. 1987, 109, 7151.
(7) Shoji, J.; Sakazaki, R.; Wakisaka, Y.; Koizumi, K.; Mayama, M.; Matsuura, S. J. Antibiot. 1975, 28, 122.
(8) (a) Ando, T.; Terashima, Sn; Kawata, M.; Teshima, T.; Wakamiya, T.; Shiba, T. Peptide Chemistry 1980; Okawa, K., Ed.; Protein Research Foundation: Osaka, Japan, 1981; p 113. (b) Wakamiya, T.; Ando, T.; Teshima, T.; Shiba, T. Bull. Chem. Soc. Jpn. 1984,57,142. (c) Wakamiya, T.; Terashima, S.; Kawata, M.; Teshima, T4 Shiba, T. Bull. Chem. Soc. Jpn. 1988, 61, 1422.
(9) Ohfune, Y.; Kurokawa, N. Tetrahedron Lett. 1984, 25, 1587.(10) Hori, K.; Ohfune, Y. J. Org. Chem. 1988, 53, 3886.(11) (a) Sakai, N.; Ohfune, Y. Tetrahedron Lett. 1990, 31, 3183. (b)
Sakai, N.; Ohfune, Y. Tetrahedron Lett. 1991, 32, 4151. (c) Sakai, N.; Ohfune, Y. J. Am. Chem. Soc. 1992, 114, 998.
(12) Zurczak, J.; Golebiowski, A. Chem. Rev. 1989, 89, 149. Other references are cited therein.
0001-4842/92/0125-0360S03.00/0 © 1992 American Chemical Society
-35-

Stereoselective Synthesis of Unusual Amino Acids Acc. Chem. Res., Vol. 25, No. 8, 1992 361
Ecfwocandins (1)
(R,-R,.O0 "YY« '(R.. OUR,-M) W \o
stn<tjf* o* gaiantn I:2*{n-3). 2£>(n-4)
Oh OH O CH, M o ICM*L
galantmamc aod. Gim (3a)
Revised srvcure of gXansn I5e(n-3).5b(n-4)
XXUrU.
paiamwwc aod: Gu (**)
acsotute svuaure of Gim (3b)revised srucaxe of Gla (4b) Soe(43): fevsed secuence
Figure 1.
5/nZAea/# eZ Ij-omlno frydrojyi Bymtmm.
R . OtpCHjRn. R* • O of OCOPK Boe - CO-hBv. TBS - SA4^-i-Bu
Syr*th0*i9 of 7,3-emfno hydroxyl •yxtom.
NHC0,TBSNH, OH
(1L (SI Kcidxcn with MCP8A: (c) S*2 tyw cydc catfiamti tocneon: (dl (•) Sew typ* eye he araerm* kxm*on: (!) Mduomeam
Figure 2.
alent of la,b and 5 has been accomplished on the basis of these methods.
Epoxidation of 2-Amino-3-butenol DerivativesBoth enantiomers of N-(ter£-butoxycarbonyl)-2-
amino-3-butenol (6), a masked form of chiral serine, have been synthesized from L- or D-methionine.13 14 15 * Compound 6a, upon treatment with 3-chloroperoxy- benzoic acid (MCPBA), underwent stereoselective epoxidation to give syn-epoxide 12a (syn/anti = 40/1). The reaction likely proceeds through an internal chelation of MCPBA with the primary hydroxyl group, since epoxidation after protection of the hydroxyl group of 6a with the tert-butyldimethylsilyl (TBS) group required a prolonged reaction time (~1 week) and resulted in reduced syn selectivity (syn/anti = 3/1).14,15 The bulky t ert-butoxy car bony 1 (Boc) group may hinder the undesired chelation with the amino group (A).
The syn-epoxide 12a proved to be useful as a precursor for the synthesis of various /3-hydroxy a-amino acids. For example, the reaction of the acetate 13 with the appropriate cuprate resulted in regioselective nucleophilic opening of the epoxide to give syn-1,2-amino alcohols 14 and 16, respectively, which were converted to /3-hydroxyhomotyrosine 156* and anhydro Gla 4a,9,16
(13) Ohfune, Y.; Kurokawa, N. Tetrahedron Lett. 1984, 25, 1071.(14) (a) Rao, A. S.; Paknikar, S. K.; Kirtane, J. G. Tetrahedron 1983,
39, 2333. (b) Narnia, A. S. Tetrahedron Lett. 1981, 22, 2017.(15) (a) Umbreit, M. A.; Sharpless, K. B. J. Am. Chem. Soc. 1977,99,
5526. (b) Sharpless, K. B.; Verhoeven, T. R. Aldrichimica Acta 1979,12,63.
Scheme IIEpoxidation of Hydroxymethyl Allylamines
PhCH7OCH2/Xv_r/\C18 (syn/anti - 1/25)
R'^NBoc
7b R, -H. R, - TBS 20 R, - R2 - acetomde
^—NBoc
22
R,"NBoc
V19 Rn - H, R2 - TBS (syn/anti - 40/1) 21 R, - R2 - acetonida
NBoc
U\A^S/C o
23 (syn/anti - 1/1)
MCPBA
(C) R, - H. Rj - TBS orMCPBA
which are constituents of 1 and 2, respectively (SchemeI).
Epoxidation of Hydroxymethyl (Z)-AllylaminesEpoxidation of the allyl alcohol 17, which has a
methyl group at C4, has been shown to give anti-epoxide 18 with high stereoselectivity (syn/anti = 1/25).17 The mechanism involves less hindered side attack (re face on C3) of an internal chelate complex of MCPBA on the C-C double bond (Scheme II, B). Contrary to this, epoxidation of the hydroxymethyl (Z)-aUylamine 7b yielded syn-epoxide 19a (syn/anti = 40/1), stereo- selectively.10,18 This example indicated that MCPBA attacked from the more hindered si face on C3. The epoxidation of 20, in spite of the lack of an amide hydrogen, was also syn selective to afford syn-epoxide 21 exclusively. The protection of the hydroxyl group of 20 with the TBS group resulted in a decrease in both yield (>20%, 3 days) and product ratio (~3/l). Therefore, the high syn selectivity in the epoxidation of hydroxymethyl (Z)-allylamine was attributed to the fact that the epoxidation proceeded through a chelation complex C. The (E)-allyl alcohol 22 provided a 1/1 mixture of syn- and anti-epoxides 23.llc Thus, epoxidation of hydroxymethyl (Z)-allylamine proved to be a potential method for the preparation of syn-1,2- amino alcohols.
Sn2 Type Cyclic Carbamate Formation from tert -B utyldimethy lsily 1 Carbamate
iV-tert-Butoxycarbonyl (Boc) and iV-benzyloxy- carbonyl (Z) groups are the most common amino protecting groups used for the synthesis of amino acids, amino sugars, and peptides.19 These groups can be transformed into the N-(tert-butyldimethylsilyl)oxy-
(16) (a) Golebiowski, A.; Kozak, J.; Juiczak, J. J. Org. Chem. 1991,56, 7344. (b) Ikota. N. Heterocycles 1991, 32, 521. (c) Takahata, H.; Banba, Y.; Tajima, M.; Momose, T. J. Org. Chem. 1991, 56, 240. (d) Kano, S.; Yokomatsu, T.; Shibuya, S. Heterocycles 1990, 31,13. (e) Golebiowski, A.; Kozak, J.; Jurczak, J. Tetrahedron Lett. 1989, 30, 7103.
(17) Nagaoka, N.; Kishi, Y. Tetrahedron 1981, 37, 3873.(18) Kogen, H.; Nishi, T. J. Chem. Soc., Chem. Commun. 1987, 311.(19) (a) Bodanszky, M.; Bodanszky, A. The Practice of Peptide Syn
thesis; Springer-Verlag: Berlin, 1984; pp 7 and 151. (b) Greene, Y. W. Protective Group in Organic Synthesis; Wiley: New York, 1982; pp 232 and 239.
-36-

362 Acc. Chem. Res., Vol. 25, No. 8, 1992 Ohfune
RNHCOj-t-Su (Boc)
RNHCOzCHzPh (Z)
RNHCOjSid-BuJMe,
Scheme IIITBDMSOTV2.6-lulidlne
------------ RNHCOySid-BuJMej
TBSH/PeiOAc), (equivalent to RNHCOj )
H,O.Of MeOHRNHj
RNHCOzR-FTX(R-. ikyt, benzyl: X - Cl. Br)
PhCHjC
Scheme IVScn- Cyclic Carbamate Formation
oNHCOjTBS
PhCHjO ^ Pt>CHjO
»X-Cl: |$yrvmti. 3,1. 76%)'AgF/P0(U) (lyrvim, .8/1. 81%J*
27»(syn)
O
776 (anti)
33 X - OBz: nBu«NF/Pd(0) (lyrvant. - 9/1, 78%)'
(1) TBSOTt/2.6-lut«5meMBoc-L-Val-OMe (24) ------------------------------------ N-Z-L-Val-OMe (2S. 85%)
(2) n-Bu.NF/PnCHjS/
27b (anti) t O
26b (anti) OM$
28c 3fl ap«mar of 28a
carbonyl group20 by treatment of the N-Boc derivative with tert-butyldimethylsilyl trifluoromethanesulfonate (TBSOTf) in the presence of 2,6-lutidine and the N-Z compound with tert-butyldimethylsilane (TBSH) in the presence of a catalytic amount of Pd(OAc)2, respectively.21 The silyl carbamate is stable under ambient conditions and can be activated by fluoride ion to undergo various electrophilic substitution reactions. In the presence of an alkyl halide, the silyl carbamate yielded the corresponding alkyl carbamate. For example, the N-Boc amino ester 24 was converted, efficiently, to the corresponding N-Z amino ester 25 via the silyl carbamate. Thus, the silyl carbamate prepared from an N-Boc or N-Z compound is a useful intermediate which can be converted to a variety of urethane-type compounds.21
Furthermore, the silyl carbamate can be trapped by an internal electrophile with complete inversion of configuration. Treatment of the syn-mesylate 26a with n-Bu4NF gave the cyclic carbamate 27b having an anti-amino hydroxyl system. The anti-mesylate 26b gave the syn-carbamte 27a. This method proves to be useful when the Mitsunobu reaction is not effective.22 23 For example, under Mitsunobu conditions the conversion of the syn-amino alcohol 28a to the corresponding anti isomer 28c was not effective due to the bulky nature of the neighboring iV-Boc group. However, the silyl carbamate method was successfully applied to this conversion to give the desired anti cyclic carbamate 29 (Scheme III).10,23
ScN' Cyclic Carbamate Formation via Silyl Carbamate
Because of its high reactivity, a silyl carbamate can be viewed as an N-carboxylate ion equivalent. Thus, intramolecular trapping of this reactive species in an
manner24 provides a stereoselective method for the
(20) Breederveld, H. Reel. Trau. Chim. Pays-Bas 1962, 81, 276.(21) (a) Sakaitani, M.; Ohfune, Y. Tetrahedron Lett. 1985, 26, 5543.
(b) Sakai tani, M.; Hori, K.; Ohfune, Y. Tetrahedron Lett. 1988,29, 2983.(c) Sakaitani, M.; Ohfune, Y. J. Org. Chem. 1990, 55, 870.
(22) Mitsunobu, 0. Synthesis 1981, 1.(23) (a) Sakaitani, M.; Ohfune, Y. J. Am. Chem. Soc. 1990,112. 1150.
(b) Sakaitani, M.; Ohfune, Y. Tetrahedron Lett. 1987, 28, 3987.(24) Stork, G.; Schoofs, A. R. J. Am. Chem. Soc. 1979, 101, 5081.
I
OHstatin# (32)
*2 #Qu<v of A;FOi3CN*2 aquiv AgF. 0.1 aquw of atytoal»dium(H) Chbnd# 0.3 #otw PfyP/CH^CN c1.2 #Qixv of n-Bu«NF. 0.1 eqmv of (Ph)P)«Pd(0)/THF
synthesis of the 1,2- and 1,3-amino hydroxyl systems as a cyclic carbamate. AgF was chosen in order to activate both the silyloxycarbonyl and the allyl chloride groups. Compound 9 with AgF underwent cyclic carbamate formation in an ScN/ manner to give a mixture of syn-27a and anti-2Tb (AgF, syn/anti = 3/1). The use of AgF in the presence of a Pd(II) catalyst (0.1 equiv of allylpalladium(II) chloride dimer and 0.3 equiv of triphenylphosphine) was superior to AgF in view of its syn selectivity (syn/anti = 8/1). As an additional example, treatment of the allyl chloride 30 with AgF or AgF/Pd(II) gave cyclic carbamate 31 (AgF, syn/anti = 5/1; AgF/Pd(II), syn/anti = 15/1). The carbamate 31 was readily converted into statine (32), a constituent of pepstatine, which is a well-known potent renin inhibitor.23,25
Allyl esters are more attractive than allylic clorides as internal electrophiles in view of their ease of preparation and stability.26 Also, the use of stoichiometric amounts of AgF can be avoided. Thus, treatment of the benzoate (Bz) 33 with a catalytic amount of tetra- kis(triphenylphosphine)palladium(0) and 1 equiv of n-Bu4NF gave syn-27a as the major product (syn/anti = 9/1) (Scheme IV).
The syn selectivity using AgF can be understood from the examination of the hypothetical cyclic intermediates D-l and D-2. Due to the presence of severe allylic strain2' in the transition-state conformer D-2, the reaction proceeds via the thermodynamically more favored transition-state D-l to give syn-27a. Using AgF/Pd(II) or n-Bu4NF/Pd(0), the major isomer 27a could be derived via (T-allyl)palladium complex F-l from the thermodynamically more favored conformer E-l, which has less allylic strain than E-2 (rate-determining step) (Scheme V).28 It is noted that the proline derivative 34 gives a mixture of cyclic carbamates 35a,b [4,5-trans (syn)/4,5-cis (anti) = 3/2] in good yield. The decreased syn selectivity can be attributed to the reduced steric bulkiness of the conformationally constrained CH2 group in a five-membered ring (G-l and G-2) when compared to the freely rotating CH2R of 33
(25) (a) Umezawa, H.; Aoyagi, T.; Morishima, H.; Matsuzaki, M.; Hamada, H.; Takeuchi, T. J. Antibiot. 1970, 23, 2569. (b) Rich, D. H. Proteinase Inhibitors; Barrett, A. J.; Salvensen, G., Eds.; Elsevier. New York, 1986; p 179. (c) Omura, S.; Inamura. N.; Kawakita, K.; Mori, Y.; Yamazaki, Y.; Masuma, R.; Takahashi, Y.; Tanaka, H.; Huang, L.-Y.; Woodruff, H. B. J. Antibiot. 1986, 39, 1079.
(26) (a) Trost, B. M. Angew. Chem., Int. Ed. Engl. 1989,28,1173. (b) Trost, B. M.; Verhoeven, T. R. Comprehensive Organometallic Chemistry; Wilkinson, G., Sir, Stone, F. G. A., Abel, E. W., Eds.; Pergamon Press: Oxford, 1982; p 799.
(27) (a) Johnson, F. Chem. Reu. 1968, 68, 375. (b) Hoffmann, R. W. Chem. Rev. 1989, 89, 1841.
(28) Anh, N. T.; Eisenstein, 0. Nouv. J. Chim. 1977, 1, 61.
-37-

Stereoselective Synthesis of Unusual Amino Acids Acc. Chem. Res., Vol. 25, No. 8, 1992 363
Scheme VProposed Mechanism of ScN- Cyclic Carbamate Formation
(0-1)
H Cl(D-2)
27* (syn)
R - CHjOCHjPh
rale of syrvantiAgF: 27a/27b . 371AgFZPd(ll). X - O: 27»/27b - 8/1nBu«NF/Pd(0). X - OBz: 27a/27b - 9/1
27b (ami)
AgF/Pd(ll) of n8u4NF/Pd(0)
X-ClofOBz
35a 4.5-trans (syn) 35b 4.5-hs (ami)
G-1 G-2
reduced alfyic strain compared to E-1
(the structure is shown in Scheme IV).29The use of secondary allylic esters is of interest since
there are four possible products considering both the ring stereochemistry and double-bond geometry. From the (S/S) isomer 36, the 4,5-trans (E) product (syn) 37a was produced with high stereoselectivty (37a/37b = 20/1), while (SJl)-38 gave the 4,5-cis (E) adduct (anti) 39b as the major product (39b/39b = 7/1). In each case, the other stereoisomers were not detected. Formation of the syn adduct 37a from the (S,S) isomer 36 and primarily the anti adduct 39b from the (SJl) isomer 38 suggests that these transformations proceed via double inversion (net retention): (1) the leaving group and silyloxycarbonyl group are placed on the same face in the ground-state conformers (36a and 36b, 38a and 38b) (the major or exclusive isomer is produced from the conformers, 36a and 38b, having the least A1-3 strain); (2) palladium(0) attacks from the back side of the leaving group; and (3) attack of the N-carboxylate ion species on the (x-allyl)palladium complex gives the cyclic carbamates 37a and 39b, respectively. These transformations provide stereoselective access to the syn- or anti-1,2-amino hydroxyl systems (Scheme VI).29
The present method was also employed for the synthesis of a 1,3-amino hydroxyl system (method e, Figure 2). The chloromethyl homoallylamine 10 produced a mixture of six-membered cyclic carbamates 40a,b. The syn/anti ratio was ~ 1/1. However, greater syn selectivity was observed when the reaction was applied to the peptide system (vide infra) (Scheme VII).23
Scheme VICyclic Carbamate Formation from Secondary Allyl Esters
TBSO—'
37a/37b » 20/1
37*: 4,5-trans (syn)-E 37b: 4.5-cis (anli)-Z
TBSO—'
39a: 4,5-trans (syn)-Z 39b: 4.5-cis (anti)-E
Scheme VIISix-Membered-Ring Cyclic Carbamate Formation
NMCOjTBS A,
40a (syn) R.CO/4, AgF (syrVanti . 3/2. 63%)
AgFZPd(ll) (syn/anti. 1/1.72%)
O
40b (anti)
Syntheses of Glm (3b) and Gla (4b)Before this work, only the primary structure of Glm
(3a) was reported.8 The structure determination of 3a required the synthesis of eight diastereomers from L- or D-lysine. The methods elaborated in our laboratory (Figure 2a-d) were used for the synthesis of the key intermediates 43 and 44. The hydroxymethyl (Z)-al- lylamine 41 on epoxidation (method b, Figure 2) gave the syn-epoxide 42, which upon subsequent reduction with LiAlH4 afforded, regioselectively, the desired syn amino diol 43. Upon S^2 type cyclic carbamate formation using method c (Figure 2), the mesylate 28b provided the anti cyclic carbamate 44, exclusively.
Each isomer was then converted to the corresponding Z and E unsaturated esters 45a-d using a Wittig or Horner-Emmons type olefination.30 Upon osmium tetraoxide oxidation of 45a-d, each gave a 1/1 mixture of diols 46a-h, a total of eight diastereomers. Thus, eight diastereomers with unambiguous stereochemistry were prepared. Spectroscopic comparisons of the triacetates 47a-h prepared from 46 with the natural product 3b indicated that (2S,3E,5E,6S)-47 derived from 45d was identical with the natural triacetate in all respects except the sign of its optical rotation. Thus, the absolute structure of Glm was confirmed to be (2f?,3S,5S,6f?)-3b. Finally, the synthesis of the natural form 3b was accomplished in a straightforward manner starting from D-lysine (43 — 44 —- 45d —*• 3b) (Scheme VIII).10
(29) Spears, G. W.; Nakanishi, K.; Ohfune, Y. Tetrahedron Lett. 1990,31, 5339. (30) Maryanoff, B. E.; Reitz, A. B. Chem. Rev. 1989, 89, 863.
-38-

364 Acc. Chem. Res., Vol. 25, No. 8, 1992 Ohfune
Scheme VIIISynthetic Structure Determination of Glm (3b)
Scheme XHalolactonization of 2-Substituted 4-Pentenoic Acids
L-Lysine . ZHN(CH,)«x/=> ZHNlCt^L
«7e-h R - Ac
Scheme IXSynthesis of Revised Structure of Gla (4b)
(IM-BuOOWTttonB
(1) Tfa>vdmso(2)P)TBSO
revised structure ol Gla (4b)
Sie R . Me Of Pn SlbR-OHorOCHjPh 11a R - NHBoe 116R-NHZ 11C R - NWTs Hd R - NPnt
52 R - Me or Pn. X - I. ca/eans - 3/253 R - OH orOCHjPn. X - Br. os/tfans - 2/154 R . NHBoe. X . Br. ds/Vans - an (82%)55 R - NHZ. X- Br. ds/trans - 8/1 (80%)56 R - NHTs. X - Br. co/vans - 8.8/1 (100%)57 R . NPrn. X - Br. dsdans - 6n (81%)
Scheme XIR
~° (H-1)
(1) TFA(2) Ba(OH),
'NHBoe
(-)-Bulgecinine (61)
° 59a R - CHjOH59b R - CHjOTBS
60 R - CH,OH
HO,C
62aR-CH7OH 62b R . CHtOTBS
63a R . CH^O
AgOTl/THF/ ' 2.6-luidine
63b R . CH^C (Z isomer)
59a (78%. ds/trans - 6.3/1) 59b (88%. ds/trans -17/1)
O -*0"*
BU)
(92% from 63a) (73% from 63b)
oer64 d&ftrans - 6fi
Synthesis of the reported structure of galantin I (2) and comparison with the natural product showed that the two components were not identical. The structure of galantin I was then revised to 5 with the change of the structure of Gla to 4b from 4a.lu,c
The revised structure of Gla (4b) and its C3 epimer was synthesized, stereoselectively, starting from epoxide 21. The epoxide was converted to the unsaturated 5- lactone 48, which upon epoxidation followed by reductive cleavage of the resulting epoxide31 32 33 regio- and stereoselectively gave the alcohol 49. The desired 3S isomer 50 was obtained from 49 by an oxidation/reduction sequence.3233 Removal of the protecting groups with trifluoroacetic acid (TFA) afforded Gla (4b) (Scheme IX).
Electrophilic Lactonization of 2-Amino-4-pentenoic Acid Derivatives
Halolactonization of 3-substituted 4-pentenoic acid derivatives has been shown to be an efficient entry for the stereoselective construction of either 3,4-syn or -anti stereochemistry.34 35 36 However, 2-substituted 4-pentenoic acid derivative 51a produced a mixture of 2,4-disub- stituted 7-butyrolactones 52 with poor stereoselectivity (cis/trans = ~3/2).33,36 On the other hand, Witkop
(31) Miyashita, M.; Suzuki, T.; Yoshikoshi, A. Tetrahedron Lett. 1987, 28, 4293.
(32) (a) Mancuso, A. J.; Swem, D. Synthesis 1981,165. (b) Smith, A. B., Ill; Levenberg, P. A. Synthesis 1981, 567.
(33) Hausler, J. Liebigs. Ann. Chem. 1983, 982.(34) (a) Dowle, M. D.; Davis, D. I. Chem. Soc. Reu. 1979, 171. (b)
Bartlett, P. A.; Richardson, D. R; Myerson, J. Tetrahedron 1984, 40, 2317. (c) Chamberlin, A. R; Dezube, M.; Dussault, P.; McMills, M. C. J. Am. Chem. Soc. 1983, 105, 5819.
(35) Bartlett, P. A.; Myerson, J. J. Am. Chem. Soc. 1978, 100, 3950.(36) Ohfune, Y.; Kurokawa, N. Unpublished results.
et al. reported that 2-amino derivative lib gives cis-y- butyrolactone 55 as the major isomer (cis/trans = 4.6/1).37 Since this method appeared to be potentially useful for the synthesis of the anti-1,3-amino hydroxyl systems (method f, Figure 2), we reexamined the bro- molactonization of 2-amino-4-pentenoic acids with IV-bromosuccinimide (NBS) in order to optimize the reaction conditions with respect to improving both the yield and stereoselectivity. The use of anhydrous tet- rahydrofuran (THF) as the solvent afforded satisfactory results (cis/trans = >8/1, >80%). The product ratio was independent of the nature of the N protecting group (Scheme X).6*’38,39
It has been proposed that the reaction proceeds through an initial bromination of the amino group and a subsequent internal bromonium ion transfer to the C-C double bond (Scheme XI, H-1 via path A) to give cis-7-butyrolactone as the major product.37 However, despite the fact that the N-phthaloyl group of lid cannot be brominated, the cis selectivity remains unchanged (cis/trans = 6/1). This fact led to an alternative mechanism (path B) in which the C2 amino group would stabilize, stereoelectronically, the putative halonium species of the cis transition state (H-2).
In order to determine the favored pathway, the halolactonization of 5-substituted (Z)-allylglycine 58 was examined: the reaction of 58 would produce the 7-bu- tyrolactone (5JR)-59a via H-1 (path A) or (5S)-60 via H-2 (path B). Determination of the stereochemistry of the resulting lactone should give an answer.
(37) Izumiya, N.; Witkop, B. J. Am. Chem. Soc. 1963, 85, 1835.(38) Ohfune, Y.; Kurokawa, N. J. Synth. Org. Chem., Jpn. 1986, 44,
647.(39) Ohfune, Y.; Hori, K.; Sakaitani, M. Tetrahedron Lett. 1986, 27,
6079.
-39-

Stereoselective Synthesis of Unusual Amino Acids Acc. Chem. Res., Vol. 25, No. 8, 1992 365
Treatment of the (Z)-aUyl alcohol 58 with NBS in THF gave a mixture of cis 2,4-disubstituted 7-butyro- lactone (either 59a or 60) and its trans isomer (cis/trans = 8.8/1, 95%). The structure of the major isomer was determined by its conversion to the amino acid bulge- cinine (61), a constituent amino acid of the glycopeptide bulgecine.39-41 Thus, the structure of the lactone obtained from 58 was shown to be 60. It was concluded that the bromolactonization of 2-amino-4-pentenoic acid derivatives proceeded via H-2 (path B). This mechanism was further supported by the halolactonization of 62a having a 5(E)-substituent. This yielded cis-7- lactone 59a as the major product (cis/trans = 6.3/1), which indicated that the reaction proceeded through intermediate I. Protected alcohol 62b showed greater cis selectivity to afford 59b (cis/trans = 17/1).39
In addition to a bromonium cation, an allyl cation could also be stabilized by the C2 amino group. Treatment of either the (E)- or (Z)-allyl chlorides 63a,b with silver trifluoromethanesulfonate (AgOTf) gave the same cls-4-vinyl 7-butyrolactone 64 as the major product (cis/trans = 6/1). The mechanism of this transformation appears to involve intermediate J.39 Thus, halolactonization of 2-amino-4-pentenoic acid derivatives proved to be useful for the synthesis of the chiral lactones which are equivalent to the anti-1,3- amino hydroxyl systems.
hH? %-nh ) NHCOC„H„
H^-= _
HN Me —
o^S,-5W^..=h
ngrg hafl thpepde of echtixcand^i B and C
Scheme XIIcydic cart&mate
N.NHCOrl-Bu
-- ■ :> hn m«HN Me
otbs^l^'OTBS
65taaonizabon
66» R, - H. R, . TBS66b R, - Na. R, - TBS66c R, -Na. R, - H
Stereoselective Conversion of a Simple Tripeptide to the Echinocandin Right Half Equivalent
Diastereoselective synthesis of the constituent amino acids of echinocandins (1) followed by the coupling of these constituents has led to the successful total synthesis of echinocandin D (lc).6 41 In conjunction with this study, we also examined the synthesis of the right half equivalent (tripeptide 65) from a simplified tripeptide 66. Our idea was to examine the applicability of methods e and f (Figure 2) in the peptide system. Moreover, it was expected that this novel strategy might provide information concerning chemo- and stereoselectivity induced by the peptide functionality and/or peptide conformation.42 The key transformation was a stereoselective introduction of the requisite 7-hydroxyl group into both the N- and the C-termini of allylglycyl moieties.
Halolactonization of 66 was highly dependent on the reactivity of the COOH vs CONH group. Using the free carboxyl compound 66a, peptide bond cleavage occurred at the N-terminal of threonine to give cis-7- butyrolactone 54 and H-threonyl-N-Boc-allylglycine- OH. On the other hand, the sodium salt 66b, which is more nucleophilic than the free carboxyl, provided the desired cis-7-butyrolactone 67a, exclusively." The cis selectivity (>99/1) was much greater than that of N- Boc-allylglycine 11 (cis/trans = 8/1). On the other hand, unprotected 66c showed a decrease in cis selectivity (cis/trans = 4/1). In order to examine the stereochemical outcome of this transformation, we exam
(40) Imada, A.; Kin taka, K.; Nakao, M.; Shinagawa, S. J. Antibiot. 1982, 35, 1400.
(41) (a) Wakamiya, T.; Yamanoi, K.; Nishikawa. M.; Shiba, T. Tetrahedron Lett. 1985, 26, 4759. (b) Bashyal, B. P.; Cho, H.-F.; Fleet, G. W. J. Tetrahedron Lett. 1986, 27, 3205.
(42) (a) Izumiya, N.; Francis, J. E.; Robertson, A. V.; Witkop, B. J.Am. Chem. Soc. 1962, 84, 1702. (b) Wilchek, M.; Patchornik, A. J. Am.Chem. Soc. 1962, 84, 6079.
ined halolactonization of the structurally rigid analogue 68 in which the N,0-acetonide constrained the structure to be U-shaped (7-turn conformation).43 The reaction provided cis-lactone 69 as the major product (cis/trans. = 30/1). These results suggested that the bulky silyloxy group of 66b constrained the conformation to be U- shaped (Scheme XII, K), where the bromonium cation was stabilized by a cooperative stereoelectronic effect of the neighboring amide and carbonyl groups. Unprotected 66c might have a linear conformation which results in a decrease of cis selectivity. The desired 67a was then converted to the protected tripeptide 70.
Although the silyl carbamate method e (Figure 2) was shown to give poor 1,3-syn selectivity (10, syn/anti = —1/1, Scheme VII), this method (e) was applied to introduce a hydroxyl group into the allyl chloride 71 prepared from 70. Surprisingly, successive treatment of 71 with TBSOTf and AgF gave the desired syn cyclic carbamate 65 as the major product (cis/trans = 12/1). The AgF/Pd(II) system was not effective in this case. Both participation from the proximal functional groups and steric effects derived from the peptide conformation at the reaction site may have contributed to the high stereoselectivity observed. Thus, the conversion of a simple tripeptide 66 into the right half equivalent of echinocandins 65 was accomplished.44
Conclusions
The basic stereochemical consequences of hydroxy- lation of allyl- or homoallylamines which lead to the syn and/or anti 1,2- and 1,3-amino hydroxyl systems have been described. The synthetic methods developed in our group have also been used for the structure determination and synthesis of several unusual amino acids. In spite of these advances, more efficient methods are
(43) Chon, P. Y.; Fasman, G. D. J. Mol. Biol. 1977, 115, 135.(44) Sakaitani, M.; Ohfune, Y. Tetrahedron Lett. 1989, 30, 2251.
— 40 —

366
still required for the synthesis of such amino acids due to their important biological activities and medicinal interests.45 Furthermore, it is suggested that a hydroxyl group placed on the backbone of an amino acid is an active site when bound to a receptor protein or a biomembrane. When such amino acids are incorporated into peptides, the hydroxyl group plays an essential role in constraining the peptide structure into a specific conformation through intramolecular or external hydrogen bonding.la,b,2s Recent interest in the family of
(45) (a) Shimamoto, K.; Ohfune, Y. Tetrahedron Lett. 1988, 29, 5177.(b) Sakaitani, M.; Hon, K; Ohfune, Y. Tetrahedron Lett. 1988,29, 2983.(c) Spears, G. W.; Nakanishi, K.; Ohfune, Y. Synlett 1991, 91.
unusual amino acids is focused not only on chemistry but also on the impacts on interdisciplinary scientific fields. These amino acids are expected to function as useful probes to investigate molecular mechanisms of a variety of biological functions.1,46
It is a pleasure to acknowledge the contribution of my colleagues: their names are recorded in the references. I am grateful to Professor Koji Nakanishi for his continuous encouragement. The financial support of a grant-in-aid from the Ministry of Education, Sciences, and Culture, Japan, is appreciated.
(46) (a) Axelrod, J. Science 1971, 173, 598. (b) Williams, D. H. Acc. Chem. Res. 1984,17, 364. (c) Shimamoto, K.; Ishida, M.; Shinozaki, H.; Ohfune, Y. J. Org. Chem. 1991, 56, 4167.
-41

182
7 T ><r ^ X?
mn\ ie\ B3# m”
1. It 10 &6 10
S. 4#^i LT4m, 6 + 7 & 6.
L /:a^(±m^T/Z (,'#
-e 9 T-wnidf§i&©£# A l m'cttz Vltmfr'Z&Z. '&'iTg=$itMt>K>(D&Z*7 t A#:Hb##lcffaic^LT(a, {tft&fe&W-X
rn^iwe, 4-acc it m)>*av *(MNi)CD<^.»iil—CD^—S^SliSirab^. *#T
{iiWftt^MNi (Dmu^m^com^totuctom^ oJIEit /<£ -7 izJt&r&WMK co 7 7 4 y*r it>^*
LTCD&HJIC-ot^T&^S. && MNi ©M mmm<Dwm it o i t&&<Dtim*&m ^ntz^l\
2. frLl'MNi ; TA-NaBr-MRNi-U
2.1 m¥£<Dft&MNi \t-v^^tm.m.mt*mtt±fcw\<D 2 0®
1963 ££ ItS £ Itti/te^6^j 11*^j&jg-e& s. c
tmmt&, $.mt¥, tom&itssc6 d> %a#d' ^#6 ^fi/t$#
^^gmit, fmm%cD&&ic&tiT£tz. &&fm
* Tadashi Kikukawa (#)± — K@f ^StTSEStIS-
S^ff±MIS^> CS»^E) 19761976^9^ xn y-y y^®^, 1979^ ; ->/f y^5.
i985^m^#m#i#±. csm) wmr# fk#. c@% n cs^5t) 66o/Ediem@s^0j 2-6-1 (gj^)
** Akira Tai ^SS5:ig^XS^S^SB^S. m¥i$
±C*»^g] l956^*|g*^St^2£3l-
(#P9] W&£&tk¥. Bm*X Cjg&ft) 678-12&mm%mSt±2PBT&£lil£ (»«F5fe)
TKSik^B, #5#- NaBr
^E^-MLSRNi (TA-NaBr-MRNi-U)
St? It/X o tz. b, MNi Itm(@5#: TA)
Ni, m#im@ AihtiX^tz. TA-NaBr-MRNi-U (2#KSiJ6Iig^E SiJ0«<fct)^SWltSn^< , gfSi8ii»b^
6 ® Ti±^ v # 9 6 a it §a%
Ztltzk>(DX&Ss\ TA-NaBr-MRNi-U
&mttLx&*mg&m?Lx\,'z>yt&fls&z-1 ko
+ 1. 3- ^-yuo^cS^Mlt^:ftfi«jlc^iJffl$tlSt?KU^X^tz.
TA-NaBr-MRNi-U WK$kft, BJL&ftomfrK<lcfrlSftSfeg-e, l^TltxG
^ssis^T-esss,< 6C6 ^£1 6AC6.
2.2 TA-NaBr-MRNi-U 0»2.2.1 S#&4!3:S 7 T' — Ni (RNi - U)
y*-&&am7 7 4 Ni/Al =48/52)(40g)^NaOH*%%(NaOH 90g^i%/f ^y *400 ml Iti8#)ltjg%%0;ia. &&(D?&1jn%;T'&, %
B*±T 1 #%##, ^%RNi m^E5T@SL±S %^M#ltZ0^<. ^>*( 100 ml) 66^itHA 7 7 x 3 ( iooo mi) k& l . m&fDmw&m&tit
48 kHz) ^T3^iE#&K^ff^.
ftmm^j%/f^y*(ioomi)-c2a%^f5. w±oE
^M@#fT^M(t 2 t 6lt Zip RNi-U^ms.
2.2.2 wmoissy(a,R)-@5@K^#^3/:(i(S,f)-@:50 (20
g)6NaBr(200g), WH d" y *(2000 ml )ltiW, & tllC 1 N NaOH^Qx.T pH 3.2 icm^HtS. tOig^f
#8ocitw<##itm'a.
42-

Vol. 34 No. 3 1992 7 zd y^r $ yjvtzx^coftjffi 183
2.2.3 RNi-UOSB£#1CH|®J LfcRNi-U!CSn&LfcBtfr?£®¥S (I000
mi) &tinz-i%m7kmcpt' so ^ ni tm#um&ic^&, mzimzisM^-yTK(lOOmi)T|Hj—mfEZftj. ^bft/cSiigTA-NaBr-MRNi-U
fi, &&4 y tK( 100 ml) 7f‘ 1 0, /f/-vi/(100ml) 3@, 7Kmibicm^6^m(THF$/c(±yob'^y#y
f-JU, 100 ml )T 2 0^7#%, ^(100 ml)*T^#f
5.liUL©^fPT' 40 g (Da&frbft.1 16.8 g OTA-NaBr-
MRNi-Uri^bilS. T&^txDTA-NaBr-MRNi-U
3. TA-NaBr-MRNi-U(cj:51,3-v> h>
(Dimmit- •>'*—;u
3.1l. 3 -it^Y y E 1C 6 ©T
*#fka#ic@!ga-#A-#-a c uv\ hitt±fvi/7-t h y(Dm7C(Co^T##:^llC^5. (R, R)~ TA-NaBr-MRNi-UOTHF###(8A#16.8g, THF
ioomi)^i/^-- y-yicAn,(loog)*mK tKSEiookg/cm3, ioox:-eit^Tic WS, ItSfiOTkS^®JRznzt o. msi&T'&im&p&i. mm&%£-tzztT~(R,R)-2.4- ^yf y^^-^CPD)
fl6.
7Ks<b(i-mTSff u, ib
^OTA-NaBr-MRNiTli^-ei^^TSL,U<£ 1' C <ho Tcri*. TA-NaBr-MRNi-U
* ft 6 6#ic, &p £?£&£< &££>#<# 9, #lc 2. 6 - vVfvi/- 3.5-
Table1 icmmmt&mvKfco&gkzttit tr^-r.
3.2 1,3- i/Tt—Ji'mvmmLTmeso- -;u£/j>g
#tr. LT2f&mo y d V7*D t7u
x-xAdc&jg, - 10CT»^%S^56%##K=%#%(#, a-RD ^#^6 ^ocit*
&Z&CS, 5)-TA-NaBr-MRNi-U fcTkStffcbites, s)-PD&'&btiZ>.
TA-NaBr-MRNi-U0_h<k me so M^WSOiSTl?,&<0±L/: (Table 1).
4. mtl,3->y-^jfflbfc7 70y a
^#89%#^ 1.3- ^EUMNi am#
fto ft4). tpT:-b\ #A^m^74zyvi/7-k h y^b^i^na PDti, c,^
ltj£ < wa^icfijffl ztizx^ic /X o ft. tj3T? t>S kj£i§® icJgK L ft#3%(±^!C^$
tia^M ^ v ^ a 7 4z f y 7 %x i'*^\lffljQfcfcT?Z<D&Mt¥lt&irZVmtm3Z fesn-cv a6).
Table 1 Comparison of (/?, /^)-TA-NaBr-MRNi and (/?, R)-TA-NaBr-MRNi-U in the enantio- differentiating hydrogenation of 1,3-diketones.
OO H, OH OR/^^R
n
H2 OIi OH
5*)(meso)
OH OH+ r *
(R*. /?*)V
R-^^R step-1
I
step-2 R(.Rm
Reaction Product E.e of Isolated yieldconditions^ ratio6) y<=) of optical!
Substrate pure r/d)R Catalyst 77 C t/h i n in n $ * Configuration
Me TA-NaBr-MRNi 100 24 o CO O O -O i
O ;
90 21 (2R. 4R)TA-NaBr-MRNi-U 100 4 0: 7: 7:86 91 60 (2R, 4R)
(Me)jCH TA-NaBr- MRNi 100 192 1:17:16:66 85 32 (35,55)110 48 c)
TA-NaBr-MRNi-U 100 60 0: 6:22:72 90 59 (35, 5.S)110 12
00cnoCD 89 62 (35.55)
») Reaction conditions: substrate (100 g); tetrahydrofuran THF (100 cm3); catalyst (15.9 g); initial H2 pressure 100 kPa. b) Determined by means of GC. c) Determined by polarimetry, [ct] d20—54.7‘(CIO, EtOH) for (2R, 4R)-pcntanediol and [ct] D20—64.5‘(C1.0, MeOH) for (35 55)-2,6-dimethyIpentane-3,5-diol after separation of each component by MPLC. d) Two succesive recrystallizations of the reaction products from diisopropyl ether gave optically pure IV. e) Dehydration of II took place under these conditions.
43

184 mi ie, h# m
pd
cttxic,-/S&vvnfr <b <D*&fficDm£&# SlcSSTS, ZCt ie*#
J&**S(53;(1)). Fig. 1 ICg-B^c
cn^omi^nm
T^S^O—&\t LTxtd^ K dvi/^e y rn^cDlulK^fc Inhoffen-
Lythgoe diol (1) <$:5Sl
(2)IC^f.PD - 7-te?
KSiJ^D^Lx (i±
KI) $fre^aosm-m).
PD jo=kCX^O%m^ir&5 2. 6- i/ / •fyu- 3, 5 - ^7°7 y
-yv(DMHD) -;t/
t LTT'4' < StfcottSTgHic ^ALT^iE^^Txxy^-KSU
OH H "
x0H “CH3 Ph
'CgHn
xo:s0H
CHS CsHn
OH #H 7)
'CbHusXc
OHN.cX
CgHi?
h<^oh‘Me
Fig. 1 Diastereo differentiating nucleophilic addition to PD-acetal.
Jk +Ri J?2 OH OH
JUL
'SiEts
-0 0 -
Ri Rz
ho^Xr,
Nu'-Rg
-^YYi
»<t Nuy.
"A._ riA
1) PCC2) Base
y
2) TiCl4
1
pjc 3) AcgO
1
Nu Rz
H2/Lindlar
OAcH'^H
OAc
1) HCHO/BFs-EtaO
2) H2/PtOg u\v v u3) aq KOH/MeOH QH
H .H y^oH
Inhoffen - Lyt hgoediol (1)
Cl)
(2)">
Ph
Me Li (3)">
(4)")
(5) 16)
— 44 —

Vol. 34 No. 3 1992 TO—7 r >f y<r ituux^cD^m 185
94n-CnHga * 7-bit*
Br2AlH .9-° —V'
' c„h2, c„h23
iH2S
0<5C„h2S
(6) 17)
xr/-
b°LiNEt2
0 0 ---------- - O O
OH
(7) 18)
O OH CHsI/ZnEt2K-'-rY"*
O OH
'CH2)n or 4HgOAc
"VV'0 OH
Cl
. 1) LiAlH,
R=Me(PD) or t-Pr(DMHD)
1) Na/MeOH
2) Hg(OAc)2
CN 2) BuONO/H +
VyHgOAc
1) Na/MeOH
2) £7
(8) 19)
(9) 20)
(10) 21)
(+) -T7 'J /7/-A, (2)
WJ&tt:*. DMHD •>* ay‘a f MbT-tite# 100# CD d.e
(8)).y ^ o7°o tv^x-f-yutiig
V h y#MK#
m ft ftmTsmts® * £LTmR-e£ z(^(9)).
ccDfcfezmm lt (+) - t 7V * / -M2) ic^t*(3:(10)). .
1C C, 1,3-/ y<r ; ^;i/%
TA-NaBr-MRNi-U(CZ6 1. 3 - is*-^cDUffl^ESc
<Dt&t>tl&.
5. TA-NaBr-MRNi-Ufc <£ 5 3 -7"VUO*S<b <h S3- b K n 4-
'>g£€>'£/&
5.1 3- ^-4y^x^yuo>kS^bS*6^)fc(i 1. 3- y<r h y CD7kS4b<hIn] L:T-&5
Table 2 IC^-§| 3 - t + V®x7
TVUCDTkSib^miCo^TTA-NaBr-MRNi - U t IB
^OTA-NaBr-MRNiObb^^^f. TA-NaBr- MRNi-UTO-vi/Ty -? $ /clteioS^ & rTtglc&ojt.
Jcl-F*t -fe ^KDTKS'fboMWJ^rlEY. ftS
— 45 —

186 mu ie, a# m
Table 2 Comparison of (R, /?)-TA-NaBr-MRNi and (R, .R) -T A-NaBr-MRNi-U in the enantio-differentiating hydrogenation of methyl 3-oxoalkanoates.
0 0 OH 0R '/^v^SOMe H2 OMe
E. e. ofs / ca) Reaction1^ product*^
Substrate Catalyst (g) (g) time/ h (*) ConfigurationMe TA-NaBr-MRNi 100/16 5 80 (a)
TA-NaBr-MRNi-U 100/16 2.5 86 (/?)15000/160 8 81 (^)
n~ C7H18 TA-NaBr-MRNi 10/1.6 144 83d) (/?)TA-NaBr-MRNi-U 10/1.6 36 89 (/?)
n-CeH,e TA-NaBr-MRNi 10/1.6 192 86d) (a)TA-NaBr-MRNi-U 10/1.6 48 91 (R)
n~C]] Hj3 TA-NaBr-MRNi 10/1.6 192 86 d) (/?)TA-NaBr-MRNi-U 10/1.6 48 94 (/?)
8) S: substrate, C: catalyst. b) Reaction conditions: THF as solvent, initial pressure of H2 100 kPa: temperature 100’C. c) Determined by polarimetry [a] D20—22.4 (neat) for (^?)-Methyl 3-hydroxy- butanoate. d) Determined by NMR spectroscopy with a chiral shift reagent.
a 50 / *- h 7 Is-71C(R, R)~TA-NaBr-MRNi-U
C160 g) , THF( 10 /) , T-t hm#/ ^;i/(15kg)^m
m□*., Fgs£LTfrt,mnr,
100 kg/cm2, lOOCTTkSft^n^. ^ 8
©7Ks©Mt;mna. ij, fern*1 K □ 4 y y 7 y#7
^/i/(81—83% e. e) <&f#a.5.2 3-bTA-NaB r- MRNi- U 1C J: a TkSfbTl# £ n a 3-
t + 81 -95 %
6 % a.b«L/:7 ; ya"\
ttmmt LT3 -t Kn + y7'7 y^©^ico^r
&-<a. 7kSfbT-t#bn^ 81%e.e O 3 - b Ku4^y7'
7 y#/ •?• vi/(2.5 kg) £-7 7 7 — (5 /) L, S
fwfioNaOH'SEtR(i /) yfbf
6. a Na i££E@L, &£PTStM©zgii@eT
6. ^fn^lCTHF/AcCNCl/l, 35 /) CDM&%) £frl*.NaCl £tfrfcB, ZPgUf 5. P%lcy-<yyvi/T ;
y (4.i7kg) &w.,
^t-'Scb 97.5 % e. e ©3-b Ko + y^y»BBa
Ef-a. C©#^#mAcCNT##^f 6 ftt£j*£t£a (3- b K o * y 7f y@g*^©[glR# 75
%). t y^{itfM©NaOH7K^<h^SLiSM-f ar; y^@JK, g&tcr-b h y£fln*.a<h 100%
e.e © 3 - b K o 4- y y 4 y h V 9
na. y h u 9CT#%#©0$ %
yyi/ic^#z<mmT^a.
b Kd + ->^S©7; yj^tiHETEfO
6. 7 r4 >y s t ur©3- b k a ^
3- b Fo4#M(6"- m LT#&u, cn
6%©3!z:iS^b(i, 4- o -y 7 *a^li7T7x yyKSiJ^lD©7F^Fz@<h LT7 7 d yy ; y yt/7^6%H*4£ LT##^fb^#re&a.
6.1 ^»®3-b Ka + '«£&£>a 3- b Ko 4 y#^g^r^n^^#©C'< —
S^Fig.2 Chb©rt^^*'>y(3)., /<7F+y y (4), v b- K A (5) tii-TlCMRNiLT
b Kn + yi^b^^nn>a. 3-b
Kni©I^tl(8) Ve## y 7 x y -y 7 ^^4 i l ra g $ nr ^ a88).
6.2 ^^^-#-^7 0 y y b LT©>3- b KP*•>»«
3-b k d 4^ y E ic {i y 7 - a tKES <h<tkicts/uy + yjum, cn5rg&7^yy# ©wies^wl, cn^^imf a c 6-c&@©%3%
a. Fig. 3 & J;t>'Fig. 4 IC(iftifti3- b Fo + yyy y#%cx-e©4to - 7'©
*yu4'4-ywg^-fij^ L /cA-6%fb#©M
7'V -b^- b* V 7 y©M$K#T6 a 7 7 h y(9) , £l£ ##3 y h yy-yu(io), 7'7^a?4 yy-Ai(ii),
— 46 —

Vol. 34 No. 3 1992 7 r d v'r $ 187
OHOMyrmicacin (3)2S)
OAcO© ©
00 OCHjCHgN (CHj)s • Cl
Pahutoxin (4) 245
0 -(0-CH-CH2-C£(0-CH-CH2-C]H
Empossfilin (7) z7) Biopolymer (8) 28)
Fig. 2 3-Hydroxyalkanoic acid moiety in natural products.
Fig. 3 Various optically active natural products derived from 3-hydroxybutanoic acid.
-47

188 mu iE, ffl# m f& m
OH Ochs(ch2)„/V^qh
Fig. 4 Natural products derived from 3-hydroxyalkanoic acids.
I47iOty®773i'l'7'(KJ(12)I |>-71/(13), t-fe ^ -yu(i4) tr<h'(t(50 - 3-1 k»
*y7> y^£, — #(/?)- f-yS2(15),
x#-^ y ycoffl&ftttez 4-r-fe $A(16)W, *fcO?) /mic^ h y (it) &mztiz.
3- b + D^y(18), (19), /<? X V T7L/f K-A(20) , 6tSt£^7*
^ F###(21)W^^$aTU6.3 - t F a + >> i x x 7 D ^>'
ft7A/*vi/<E©RET rf- 7A/*A/{|:£tt3. 3- b K
d^yy7 y^xx-ryi/£cz- y fyy- - jb*J:?>'x 'Jxo-2-yf-7i/-3- t K a 4-
y7> yi^iC7 7/'^'f ®i7x#,(55(11)),ftT(^£43). $/: 3- b KD + yixxr^i/^i L
fc&ETvi/*aSeebach 60#^^ T-E5:$nT^^44).
6.3 T5§Figi IT0 3- b h'P4r*>®! ^#mic%#7Z3- b Ko^y 7'7 y#&c/c#^m
70 LTf# bft£ 1. 3 - 7'7 y y * - Mi 4 STxjW^ c2
^4% 1. 3 - yy* — vi/ <k |o|#7-fe 7 — vi/^F^z^<h LTffll/'
b#,6. ftmM^Tlc^f(^:(12), (13)). ia±7^#rS%3- b
^Ticj5:sB@icfijm^n, LT^y#mHcD^WJffllcSoTuVc. L^Lco^Slii)EF0
^bW## ItetftittK b /a V '#, y -
OH CHjIA/CO2BU --------- -
Base
OHA/CO:Bu
OHA/CO%Bu
OH ORVH + xUL
O OH. X
%^\o
BrZnCHzCOzEtXX^'CH2OH + n - C7H15CHOOH TiCh
CM
CH3 CHs OH
CHS CH3
RNuo^H
— /k/C02H
x;; -
R-C7H15
22a)
22b)
C7H15
OHxk_%COsEt
(11)^
(12)4^
(13)4"
— 48 —

Vol. 34 No. 3 1992 ? 7 4 v<r S 189
yu*> 1^ ©x >y- Vut V r (£SB-ra V \ 3- t KLT#(cmm^#fF^'#6 ux
L'MRNi (cZ6TKS-ft(Ccka' 1Cmnistiz.
7. *; *? y ic
TA-NaBr-MRNi-U©^^T:Ca 1, 3- ^Z-
^E, 3- t Kd.+.i/#E©AJ5%#g#K:#o fz. cft
©SI£V't>£t\ $S b (i C © Wx - - XicjS*., ZD$iic 5«k 9 7*0rt£ii&T^£.mmc## < mm
l t-t.
X E
1) b# m, mm, 29,953 (1991)2) A. Tai, T. Harada, “Tailored Metalcatalysts”,
ed. Y. _ Iwasawa, Reidel Publishing Company (1986), p. 265
3) A. Tai, T. Kikukawa, T. Sugimura, Y. Inoue, T. Osawa, S. Fujii, Chem. Commun., 795 (1991)
4) K. Ito, H. Harada, A. Tai, Bull. Chem. Soc. Jpn., 53, 3367 (1980)
5) A. Alexakis, P. Mangeney, . Tetrahedron: Asymmetry, 1, 477 (1990)
6) P. A. Bartlett, W. S." Johnson, J. D. Elliott, J. Am. Chem. Soc., 105, 2088 (1983)
7) W. S. Johnson, R. Elliott, J. D. Elliott, ibid., 105, 2904 (1983)
8) J. D. Elliott, V. M. F. choi, W. S. Johnson, J. Org. Chem., 48, 2295 (1983)
9) A. Mori, K. Ishihara, H. Yamamoto, Tetrahedron Lett., 27, 987 (1986)
10) A. Mori, J. Fujihara, K. Maruoka, H. Yamamoto, ibid., 24, 3367 (1983)
11) S. D. Lindell, J. D. Elliott, W. S. Johnson, ibid., 25, 3947 (1984)
12) A. Ghriki, A. Alexakis, J. F. Normart, ibid., 25, 3038 (1984)
13) W. S. Johnson, J. D. Elliott, G. Hanson, J. Am. Chem. Soc., 106, 1138 (1984)
14) T. Hosokawa, T. Yagi, Y. Ataka, S. Mura- hashi, Bull. Chem. Soc. Jpn., 61, 3380 (1988)
15) K. Funakoshi, N. Togo, K. Sakai, Tetrahedron Lett., 30, 117 (1989)
16) M. Kaino, K. Ishihara, H. Yamamoto, Bull. Chem. Soc. Jpn., 62, 3736 (1989)
17) A. Mori, H. Yamamoto, J. Org. Chem., 50, 5444 (1985)
18) M. Yoshikawa, T. Sugimura, A. Tai, Chem.
Lett., 1003 (1990)19) T. Sugimura, M. Yoshikawa, T. Futagawa, A.
Tai, Tetrahedron, 46, 5955 (1990)20) T. Sugimura, T. Futagawa, I. Ryu, A. Tai,
Chem. Commun. {in press)21) T. Sugimura, T. Futagawa, A. Tai, Chem.
Lett., 2291, 2295 (1990)22) a) T. Kikukawa, Y. Iizuka, T. Sugimura, T.
Harada, A. Tai, Chem. Lett., 1267 (1987); b) N. Nakahata, M. Imaida, H. Ozaki, T. Harada, A. Tai, Bull. Chem. Soc. Jpn., 55, 2186 (1982)
23) H. Schildknecht, K. Koob, Angew. Chem. Int. Ed., 10, 124 (1971)
24) M. Yoshikawa, T. Sugimura, A. Tai, Agric.■ Biol. Chem., 53, 37 (1989)
25) s mx, 46,.501 (1988)..
26) D. G. Bishop, et ah, J. Lipid. Res., 4, 81 (1963)
27) W. Herz, R. P. Sharma, J. Org. Chem., 41,: ^ .1015 (1976)
28) IE, B3# % it*£tTLM, 44, 1740 (1991)
■±imr£, 38, 141 (1990)29) A. I. Meyers, R. A. Amos, J. Am. Chem.
Soc., 102, 870 (1980)30) H. Tsutsui, O. Mitsunobu, Tetrahedron Lett.,
25, 2159 (1984)31) W. Seidel, D. Seebach, Tetrahedron Lett., 23,
159 (1982)32) T. Kitahara, K. Koseki, K. Mori, Agric. Boil.
Chem., 47, 389 (1983)33) K. Mori, Tetrahedron, 37, 1341 (1981)34) K. Mori, K. Tanida, Tetrahedron, 37, 3221
(1981)35) P. J. Maurer, M. J. Miller, J. Am. Chem. Soc.,
105, 240 (1983)36) 61-1879137) A. Griesbeck, D. Seebach, Helv. Chim. Acta,
70, 1320 (1987)38) G. Frater, Helv. Chim. Acta, 62, 2829 (1979)39) T. Kikukawa, A. Tai, Chem. Lett., 1935
(1984)40) M. Kiso, Y. Ogawa, S. Tanaka, H. Ishida, A.
Hasegawa, J. Carbohydr. Chem., 5, 621 (1986)
41) B. Lammek, W. Neugebaner, D. Perkowska, G. Kupryszewski, Poly. J. Chem., 52, 756 (1978)
42) A. Tai, N. Morimoto, M. Yoshikawa, T. Sugimura, T. Kikukawa, Agric. Biol. Chem., 54, 1753 (1990)
43) EB4J B#r, Nippon Nogeikagakukaishi, 64, 1741 (1990)
44) D. Seebach, S. Roggo, J. Zimmermann,
-49-

190 mu ie, bb# m d* m
“Stereochemistry of Organic Transformations”, eds. W. Bartmann, B. Sharpless, VCH, Weinheim (1987), p. 55
45) D. Seebach, R. Imwinkelried, G. Stucky,
Helv. Chim. Acta, 70, 448 (1987)46) T. Basile, E. Tagliovini, C. Trowbini, A
Umani-Ronchi, J. Chem. Soc., Chem. Com- mun., 596 (1989)
Asymmetrically Modified Heterogeneous Catalyst; Its Application for the Synthesis of Optically Active Fine Chemicals
Tadashi KIKUKAWA* and Akira TAI***NARD Institute, LTD,
2-6-1, Nishinagasucho, Amagasaki, Hyogo 660 Japan **Faculty of Science, Himeji Institute of Technology,
1479-1 , Kanaji, Kamigori, Hyogo 678-12, Japan
Tartaric acid-NaBr modified Raney nickel catalyst(MNi) is an unique heterogeneous catalyst for the enantio-differentiating hydrogenation of prochiral ketones. This catalyst system has recently been significantly improved by the use of ultrasound irradiated Raney nickel catalyst. In the first part of this report, preparation procedures of the new MNi and methods to obtaine optically pure 1, 3-diols of C^ symmetry and 3- hydroxyalkanoic acids by the use of MNi were mentioned and in the last part, utilizations of optically active 1,3-diols and 3- hydroxyalanoic acids for the synthesis of fine chemicals were reviewed.
(© 1992 Catalysis Society of Japan)
-50-

SFilE
1 . (i C »icS«£gftoo«-^54i «
%@£ LT S 7c. -g-Mitf ‘J '*■'71- K £ f flCDffllZIZ-o^X&li £ i> £ , f^'6% 2o#*mt-c&o *. -s-s;^ >/ -<7"^- K l:ov>t^U -y y x ic ftS? tij (Kfltffitjia'fiiiSS ti. 4 vy\*y RtfO^c r^ £ L xf v -f/'f x<9#itf l*. & fcS^ratsttStiico^T/y->--yyx^'-S-SKctl. ^jSa«OSWIc£$^$|lJ6$7i: L?t. £ ca%)' 1 9 8 Of ftz)' 4iSG?a@A&*ag£tj^>/<ysi^#g®i:^sl. M^wg&^ycitca^t* S * J: 7 tc» -?/i. f £LA,lcc^TXm$sA*#. n m r ^cOfiSiiJSt/rf 5#5K$iigcoJff yfz)'»$n5 i 7 fc£ * £ . 6 (if XT )Vt LXcDrf- 'I ^TT Kfi-ec9&g!j£3*i. c5 &
73*^ J: 7 ic#-?*. CiO|«|6i(iSSTt ^is^uTvS =t 7 T\ ffffgf iiifiiifi £A£' SS-Xib^M-xcO—
£££>i'cy)»f- #lcTX V A©fi‘Sft^#$4J'C,'ic LT'±ft^«0«*SJt*£ ITLTtf5JSS5^S4MSi'ffi^ic*-3-CS*. ty y
o-f LT^S! L. $■attflsTt £*j>'4«W-rft -5. @%*icf *f67kf-#£ u 7 a> «> JSifcc iu
v'd Tt-S-tit;S"Si{k-B-®g$-4'fk¥toS!fc;lc#i(i $ -if Jfi'T # & # 4ti(i¥^{b¥«0#%^&{b#&m#£ LT#4&Af£ax-6T-&£,7. jo b ^> ^Hb^KfoUfS&.^
7 i<D:4'!i £ A£‘T* 9. i 7
tc(±T#rv>*v>. i*£®c £x)i4»$^yf.giSt%icMLT$$r-*6S6r-^*5. li)*l. st <isffT5£. $ &%-&j'JSi; £2>'*jJ--pT £ *.
frtibixttl O^mmA'b. Afc'lclifftliV'Ti /# (#AM‘TS y$r) S-S-JSL.
4fl4 &/-/£-<y-^- Kf f V ''VT'-f- KO-S-titS-Tf-yT v> 6. zcD^titz J: 9 . #% lc£tii£fE6&$$tf iM7’f K(c#Af 6 C £ yr # . «tc .Jfcx 4 ^f-f cO »*J £M#y-5«Sg£»A-C&b Z £ sJ'#j>>7> *. *7!fC<DW55££;i7*9Ui. #»TSy®
&mv^6$(oa#$$ lts< mmixa b?. *«icv'd ^sscose^i-esrassatb^mt^ix^tz. ft$. xic^-<^8£titcM%$n, #a«;t 5 y
coWtSHb-S-^SgitAn?, ii -o £ <j fngttcOS VST* 4 £fx6ii(c^
-3 A. t%bb. l-®{fc¥<7)8ifigtt. £tttt£4ffc¥<9«8fc£ £*fcS5x.fc*/rLw/N-'f *• y Sett’S:*, £ 1-ft (£ Tables 2> £cT)ISi$£7#o J: 7 ic K
-3 7tsRT-£> 5.
;®yVJ!y7ATU:. (1) 55SK#A£ST 5 y & A ftf-g-iSf V ^AA HO*
-1-
— 51 —

(2) (g? yy^SO-S-fiS) . je
tty (3) #XS>T5 /&£$iJg£1"5*/ ? D-^-;vEffff5i!<7)%#lc-ov'ri$-x-S.
2. x v^vi- K<7)gijE»simii$fML7i%fesas($<7)fies
-S-i&tf '>'<71- b'(t<kcD <t 7 ^#%£ <j -?TV'S. ( 1 ) JKii Wtc liffS«7 S /®y-yvxSftod!'J^7'f Ki'MtUii. (2) jSig&S V'iiEff't'far -^') v 9 X% £cDMM®'&£'&t) 7 j; £. (3) Sir»±*S5i|tt$ 5
it. (4) 3 L-XT V -7 ?0A^L C £. C<7)
#%£%y L. *v-^yy l£g$ftctL£xifF-r-sr.ilse^ij$-£3 C <!: ZitSULT £ ft. 0 1 ic*-fa^ £«*!(: /&£•£-<£U -fflA, i^8«7S y Set £&b-y-T* V^y-f- K£f#*t--5. C tiz t 0 . 3ttb^
i"<n'X-ffl®x-z zytmtx'i ^7i-t-£-&-«% u A.
Nonnaturat Amino Acids Carrying Aromatic Groups: HjNCHCOOH
R
o iojnnr Co
0 1 . £-)£? s ;&
2-1. mi'ISZ9t|LA:^*#M^£#-7 4; V^yf K - - ®jgx*;irdf-eiift:- -
77 y^. e u — >v^ * ^ tW±T v h >; ;ir*d9 J: 7 W^£i#-7->-->-vx45 V
^77 M'-S-fiS-Ctx^ii-cfcTcWtaiilSe^lLTi^ff^^feSW-g-ffi LT. «su^:. r> t v;ir$£t#
o-> — 7 'sx V ^77 K (I)i±. !§ffi4IT-0 2<X)«t7CTVh V V ^7-f ISdO-^V 7 2xiCf&oTa|iJWi;ffi5i|LTVAi»y. Rlfett (CD) x--<? b ##z)' beRb it'ic $ jl7t.
- 2“
-52-

NH-CH-CO-hr--K-NH-CH-CO
poly(Lys(Z)2 -ant Ala 1
(I)
@2.
V^7'f A).
xfc&o oti^: t&ftVZetifco
2 - 2. ^ >/x h o-A^ r#Bir
c? 'b ------
Tt: or - -x V v ^ X^ V ^<7"f K (H) flit. C DX"<^ h y ^ / 3 7#rtf((D%£3k, El 3 CO «£ 1 ^$ tilt.
-3-
-53-

m = 1
m = 2
0 3 . i>y 7-1V7-') y 8 - b-1/- i <J otf V -<7'f K
*i8iS$P £/<A'X*(!8!N LtzikconfrbP ■ Ste./z. D tP k&5.5AMtlX^Z%>& (m=0, 2) . 1 0 7 COX-f-X-tb 0 ,9.4Aiyy,-§- (m= 1 ) 1 0scOX-?'-X'a5Z Z k&ty b i'tz %-3 tz. i fz m = 1 J: 0 ra = 2^,-it ')^7i- pcofivXZKn.T&WmKZffitctiS'b.Ltzi.co.TtiZ < . Sfifl £iS l ft <J OT& i> Z k&VRb*'lztt -D it.
■ j: o at < aa wt-. o t p
<0^S®e$$-8ALft (III. 04) „ 7i-)I . t7f^. t'7i-)K 7it'/X'J1). t;4LSliS»*(±/>UT-(±*a^*# < * i, x t *<
ftti'-o tz.
(Ill)
04. i/y v / 8 - f-yf ;i-8- f p —;i^ 8 confess $$
-4-
— 54 —

m±<o£ 7 iznmvgiz kcsal. -en^s-
—&<o&t&E&nz&&-i-z*&x-zz. v "c/^-
jp-SIt. *4 ia(± Static J: oT«L*Sf«SJL«StoSK£ # ^ Ett l i§-fi£f £ z id'^agf &&.
%&g£$:A&A. tct&x-ztia.KMlc-3%^6MM%)'^TnKK» 7 (05).
05. ^0
J: 0 DNAc7)#$<yfi@l:S±7- I-' y«M®S©'>iK 3 r yJ»A L. J t rnai;#x87S / JC ilci-ot. i y&£&Xt Z C j:x)'-p§ i;ii'l 9 8 9 ^StorAcoDFS&Stc J: -ot^gc *1*. f dj@a$:0 6 (ZTfi-f.
*-(>) (i#±3 K'/UAGSI5i81"d t RNA (t7Vy t-tRNA) !c . #A*STsySeSottrc-yyyi/^A K &»*Wi=iH•^n^ste'r^^ y/<y ??•&•$*
lc*Af 6 A(=$ 3. ZCO^atit *'8:.ft)£4'lc£> 9 . H)K«S7S /SiSfATS 5SPSlc(±Sl-ri,'»VA #lc. #5e»TS /8ftt t RN AtiS?iifcttMt-'J t: V - A tc $ I) *). 75 yBwaSS'ba'-f- zx-ft bti-C'.'Z'S!&&&$>%.-ecr-TS / ScO^fSJS i: ylTa^«#»7S y»&@#L Ak-^-D -7-f yyrtn y-^-s-SL. -etp v *y -
a-xO*S-& (y y/tyK^-S-fiE^BS) SPH/c. f (p^3*0 7 *a$&(#/:. ;ti
d'» WS^^fSiMaTiSftfiSC J: o T It & XX S £^Tngt±;4'& £ C
-5-
— 55 —

Amber Codon
Plasmid
In Vitro Protein Biosynthesis
suppressor tRNA-aa*
A
Alloprotein Containing aa*
T4 RNA ligase
suppressor tRNA(-CA)A
Transcription
p(dC)pA-aa*zk
deprotection
coupling of aa*-Boc
DNA Synthesis Synthesis of
protected
p(dC)pA-3'0H
Synthesis and Optical
Resolution of Non-
Natural Amino Acid, aa*
^6.
-6-

hich3)2
0 7 . yV7tD7C j: 6 f
(of# (BSetii t Sti^-cors y swv ,1-: v-Aicitss met )
3. 5 S&itilgti-Z*/ ? n-y/L-Kf*:
#xsts it. tftbztgffit z*s ?v-i-ji'}ji#cDfflZziijf-otui. Kin i 4 tc 6^*6 nr v>5»'^-ck@ \<di7 ySi'SA?fti i: <£ 9 Kjgtix)1® < SStiST-**. V'dv^»#X%?T
S y K6#Mtoicl§S61-Z,ElSii.r-y <, S66Ett(ofiil$$® Lr. c 6z)'r-# 6. «
x(£. T'/-<y-fc'y$£iSo7 5 y ®£-irtf^7f- KSStiSi itt / ?D-t/«^ ffMlft. c <o6tff(i. h 7 yxTV-^yyySSIgffil.. 2a-S-;S&K = 1 07(07-y"-
-Vg-S-tifr. i'XttttilS ,h A-iTSS-S-LSv. EjSS^EfSo^ic J:5*s-g- • *?@t£osf@^!r-^ z z <ti'7K$n* (08) . umtA^Ksoa. a-^co^sss
(t)9l*t»crv^(9T. tn$-±$«i#f 6c tr% tx75 yttttiyi'^o*
M»-f i> ify IS ft * t a x. X V> S .
-7-
— 57 —

4.
K87St't. ? >;<? SlcftEsn^ S*ttiS^S-34*iS^"T-. $ b£lifi<kmmm'&£&-?■£{$?<Oi,<Di: <Dte£i.£ftZ> zt&VZ 5. AIM^V/'7R (m.?y;<7 (Ft) SSMIfSffiTS/ *&#ALTfF6C g (S»t®l) $ 2 OUSSOTi ASA v> 7 4%%60#^Sx.rfFl) ttit"c AA'Fr-*2. (0 9) .$t£«S§1-£ *£ ( A:A. i8^ v.^? 5!tffl??<0 ^ -7 h t Lt 4).•? A ^ “Tflgtt<7)ffiv^<7)T-S>2.
-8-
— 58 —

Chemistry of Lantibiotics
19
Tetsuo ShibaPeptide Institute, Protein Research Foundation,4-1-2 Ina, Minoh, Osaka 562, Japan
SUMMARY: Lantibiotics are biologically active lanthionine peptidesisolated as products of microorganisms. The first lantibiotic, nisin, was found in 1928 and is now in practical use as a food preservative. Another type of lantibiotic was recognized in ancovenin, which is an angiotensin I converting enzyme inhibitor. We elucidated the chemical structures of nisin and ancovenin by chemical analyses. The structure of lanthiopeptin with an antiviral lantibiotic was also determined in our laboratory. It was confirmed that lanthiopeptin is the same compound as cinnamycin and Ro 09-0198 whose structures had not been established at that time. A classification of all lantibiotics in view of those structures is presented.
We achieved the first total chemical synthesis of nisin of 34 amino acids involving five lanthionine or methyllanthionine loops and three dehydroamino acids. In this study, new methodologies for the preparation of lanthionine peptide and dehydroamino acid peptide are presented.
Related to the biosynthesis of lantibiotics, a novel preparative method of lanthionine peptide from synthetic precursor by enzyme for coupling of thiol group with double bond is introduced.
1. INTRODUCTIONNature sometimes shows rather peculiar behavior in peptide chemistry.
A disulfide linkage in cystine residue is an obligatory clamp to fix a peptide chain to the definite conformation of proteins in plants and animals. In some microorganisms, a monosulfide linkage is utilized plausibly for the same purpose. A thioether amino acid corresponding to cystine is called lanthionine, which was originally found in wool.1) On the other hand
-59

430 T. Shiba
lanthionine peptides have been found as microbial products possessing various biological activities. Recently, it has been proposed that a group of biologically active lanthionine peptides should be called lantibiotics.
An antibiotic-like activity of nisin was first reported in 1928 slightly earlier than the discovery of penicillin,2) but it was not isolated until1947.3) Since then, more than ten different kinds of lantibiotics have been
found, all from microorganisms. Because of the multiple biological activity and unique structure of lantibiotics, much attention has been paid from many points of view, particularly concerning biosynthesis, chemical structure, chemical synthesis, active mode, fermentative production and even practical application.
During the past ten years, we performed structural studies on nisin, ancovenin and lanthiopeptin as well as the synthetic study of nisin at Osaka University and the Protein Research Foundation. The results are summarized and presented herein.
2. STRUCTURE OF NISIN, ANCOVENIN, AND LANTHIOPEPTINThe establishment of the molecular structure of the lantibiotic is
necessary for the further development of biology and chemistry of this unique bioactive peptide. It must be based on a precise analysis of the components including stereochemistry and an exact determination of the amino acid sequence involving the lanthionine ring moiety.
The structural study of nisin was initiated by Gross and Morell in1971.4) They proposed the whole structure of nisin (Fig. 1) which is composed of 34 amino acids including five thioether linkages of meso- lanthionine and f/zreo-p-methyllanthionine as well as two residues of
dehydroalanine and one residue of dehydrobutyrine. Similar structural features were soon recognized in another lantibiotic, subtilin (Fig. 2), although this has 32 composite amino acids.5)
A B C D E
I teJfHisYVajYD h aflys) ohProjfGlv [AsnjrMet
Fig. 1 Structure of nisin.
-60-

Chemistry of Lantibiotics 431
A B C D E
Fig. 2 Structure of subtilin.
Before starting our synthetic study of nisin, we were forced to reinvestigate the structure of this lantibiotic, because details of itsstructure determination had never been published. We purifiedcommercially available nisin6) by preparative HPLC method (Nucleosil 7Cig,
35 % acetonitrile-0.3 % HCOONH4 (pH 4.5)) to obtain the completely pure sample. The amino acid analysis confirmed the proposed amino acid composition of nisin. FAB-MS gave a molecular ion peak of 3352.7 (M+H+) corresponding to the proposed structure.
1) OH2) H+
+Ph-NH-C = NH
Sx CH-RmC
11O
CH3 RnNH3 + 0=C-C0-NH-CH-C0----
Fig. 3 Edman degradation of dehydroalanine peptide.
-61-

432 T. Shiba
For sequential analysis of nisin including dehydro amino acid, it must be kept in mind that, as shown in Fig. 3, the Edman degradation does not go over the dehydro amino acid residue by loosing an amino terminal.
Furthermore, when phenyl isothiocyanate reacts with the counterpart of lanthionine residue, the thiazolinone ring after cleavage of the peptide bond, links to another part of the lanthionine moiety by thioether linkage, and will be liberated after Edman degradation proceeds to this residue, as shown in Fig. 4.
Ri R,Ph-N=C=S + H2N-CH-COj-NH-CH-CO.........-jNH-CH-COtNH-CH-CO.........
CH-R V______
Ph-NH-C = NH I IS CH-CH-R x/ k____C
iiO
CH-R'J
1) OH2) H+
h2n-ch-co-
(R, R'=H or CH3)
R„
—-NH-CH-CO-NH-CH-CO.........ICH-R'
_______ J
S=C—NH I I
Ph-N CH-CH-R w k____C
11o
HN—C=S I I
R'-CH-CH N-Ph ___V x /
9o
(not detected)
R„+ H2N-CH-CO-------
uNext jCydeJ
Fig. 4 Edman degradation of lanthionine peptide.
In fact, we first reduced the intact molecule in the presence of palladium black under 8-9 atm of hydrogen at 55 °C to obtain the reduced nisin in which the double bonds in Dha (dehydroalanine) and Dhb (dehydrobutyrine) were saturated, and meso-Lan (me50-lanthionine) was converted to two Ala (alanine) residues by desulfurization, and MeLan (p- methyllanthionine) to Ala and Abu (a-aminobutyric acid) residues. The
-62-

reduced nisin was subjected to Edman degradation, which proceeded as far as 15 steps, as shown in Fig. 5.
Chemistry of Lantibiotics 433
Ile-Abu-Ala-Ile-Ala-Leu-Ala-Abu-Pro-Gly-Ala-Lys-Abu-Gly-Ala- - -
Fig. 5 Edman degradation of reduced nisin.
D E ABC
25 min
Cosmosil 5C18 (4 x 125 mm)
Fig. 6 Elution pattern of BrCN degradation products of nisin on HPLC.
We then tried a fragmentation of the original molecule of nisin by means of cyanogen bromide degradation to obtain mainly two fragments cleaved at methionine residues which were separated as shown in Fig. 6.
63-

EntryPeptide Modification
1/2g 1/2g 1/2g 1/2g 1/2g 1/2g
I I I 5 10 I I 15 IH-Ala-Val-GIn-Ala-Ala-Dha-Phe-Gly-Pro-Leu-Abu-Trp-Ser-Ala-Asp-Gly-Asn-Abu-Lys-OH
Ancovenin Intact
1 Reduction of Dha
9 Addition of HSCH2COOCH32 to Dha
o Proline Specific Endopeptidase d Digestion
4 Reduction and Desulfurization
5^ Ala
5 X 10
/ Mem \ \%=Cys )
10
5 Ala Cha 10 15
Fig. 7 Sequence analysis of intact and modified ancovenin.
434

Chemistry of Lantibiotics 435
One corresponds to the A-B-C ring part where ring C is opened at the methionine position, and the other to the D-E cumulative ring part. These two fragments were analyzed sequentially after reduction as mentioned before. The results of the sequence analysis were quite satisfactory being coincident to the proposed sequence. An assignment of thioether linkages at lanthionine and methyllanthionine was carried out according to the same principle mentioned later in the structure determination of ancovenin. Consequently, a correctness of the structure of nisin proposed by E. Gross was assured by our cautious reinvestigation of the structural analysis.7)
Parallel to the structural study of nisin, we attempted a determination of the structure of a novel lantibiotic, ancovenin. Ancovenin was isolated from a culture broth of Streptomyces sp No. A647P-2 as an inhibitor of angiotensin I converting enzyme at Fujirebio Inc., Japan in 1983.8) IC50
value of the inhibition activity for a rat lung angiotensin I converting enzyme is 8.7 x 10-7 M, which is between those of the natural potentiator C
and the synthetic antihypertensive agent, captopril, in strength.Although this lantibiotic exhibits quite different biological activity from
that of nisin, ancovenin also contains Lan, Melan and Dha in the molecule, which is composed of 19 amino acid residues. The sequential analysis of ancovenin was carried out according to the same principle as that for nisin.9) The results are summarized in Fig. 7.
The Edman degradation to an intact molecule of ancovenin gave only phenylthiohydantoin derivatives of the 2nd and 3rd amino acids and stopped there (Fig. 7). Therefore we reduced ancovenin under 1 kg/cm2
hydrogen atmosphere in the presence of Pd at room temperature, thenrepeated the Edman degradation (Entry 1). In this experiment, Ala appeared at the 6th position indicating Dha in the original form, but the reaction stopped at the 7th residue. Next we treated ancovenin withmethyl thioacetate which was added to the double bond of Dha. The Edman reaction of this product went as far as the 10th position (Entry 2) showing a Pro residue at the 9th residue. Taking advantage of this information, we digested the original molecule with proline-specific endopeptidase to secure a new amino terminus for further sequencing(Entry 3). The Edman degradation now gave us whole amino acid residues as far as the C-terminus. In order to make sure of the sequential assignment, we finally attempted a drastic reduction under 10 kg/cm2
hydrogen atmosphere in the presence of Pd at 50 °C to saturate all double bonds and remove sulfur atoms from thioether linkages. The result is shown in Entry 4.
— 65 —

436 T. Shiba
1/23 1/25 1/25 1/23 1/23 1/23
H-Ala-Val-GIn-Ala-Ala-Dha-Phe-Gly-Pro-Leu-Abu-Trp-Ser-Ala-Asp-Gly-Asn-Abu-Lys-OH
Entry 1 |.....................................Dns derivative B
Entry 2 —------ ------ — j .......... Dns derivative A
Entry 3 —------ ------ ------- j --- Dns derivative B
: Edman degradation | : Dansylation
Fig. 8 Determination of position of meso-lanthionine.
The determination of the positions of thioether linkages for Lan and MeLan moieties was carried out as follows (Fig. 8). First, the molecule of ancovenin was dansyl(Dns)ated followed by acid hydrolysis to give Dns- thre6>-p-MeLan (Dns derivative B in Fig. 8) indicating that the counterpart
of 1st Ala residue should be Abu (Entry 1). Second, after three steps of Edman degradation, the same procedures as above were applied to give Dns-meso-lanthionine (Dns derivative A) showing that the counterpart of the 4th Ala residue should be Ala (Entry 2). Finally, after four steps of Edman degradation followed by dansylation and then hydrolysis gave Dns- threo-fi-MeLan again (Entry 3). It was thus concluded that the 14th Ala
residue is clearly linked to the 4th Ala residue.
Ancovenin
Deamination and cleavage of prolyl peptide bond
1) NaN02 2) proline specific endopeptidase digestion
Edman degradation ( ——)
1/23 /i/ag 1/23 ^ 1/23
H-Hpr-Val-GIn-Ala-Ala-Dha-Phe-Gly-Pro- Leu-Abu-Trp-Ser-Ala-Asp-Gly-Asn-Abu-Lys-OH\ 4 ...............s-....................-*T~ 14
........................... s.................................... *Hpr: 2-hydroxypropionic acid
fhreop-methyllanthionlne disappeared
4H-A|a-Val-Gln-Ala-Aja-Dha-Phe-Gly-Pro-Leu-Ayu-Trp-Ser-Ala-Asp-Gly-Asn-A^u-Lys-OH
14
Fig. 9 Assignment of sulfide bridge belonging to fhreo-p-methyllanthionine.
— 66 —

Chemistry of Lantibiotics 437
Assignment of the remaining two thioether linkages of MeLan was rather tricky. First the original molecule was treated with NaNOz to change the amino terminus to hydroxyl group. We then applied proline endo- peptidase digestion followed by Edman degradation (Fig. 9).
Acid hydrolysis of the product after the second step of the Edman degradation gave no more (3-MeLan. This means that Ala5 and Abu18 are
not linked to each other, otherwise both residues would give intact (3- MeLan after these treatments. The conclusion is that Ala1 must be linked to Abu18 and Ala5 to Abu11, as shown in Fig. 9.9)
1
HO-fLysAlalAla
SerTTrp
Fig. 10 Structure of ancovenin.
Fig. 11 Conformation of ancovenin.
-67-

438 T. Shiba
Steric configurations of all component amino acids were determined gas-chromatographically 10) on chiral capillary column1O for TFA-amino
acid isopropyl esters obtained from acid hydrolyzate of either ancovenin or desulfurized ancovenin.9) The results showed that all amino acids except
Lan and MeLan are of L-forms, one Ala residue of Lan and two Abu residues are of D-forms. Therefore, we deduced that ancovenin has one
Smeso-Lan and two MeLan of L-Ala D-Abu form. The whole structure of ancovenin was thus determined as shown in Fig. 10.9)
This structure of ancovenin is quite unique and peculiar having triply cumulative sulfide rings. Conformation analysis of this lantibiotic was performed by NMR measurements including COSY and NOESY in H%0 and D 2O. For an elucidation of the tertiary structure, distance geometry calculations were used. One of the conformations of the lowest value of target function is shown in Fig. 11.12)
The structure-activity relationship of this particular compound is worthy of attention. We attempted specific cleavages of rings B and C by 2-(2-nitrophenylsulfenyl)-3-methyl-3-bromoindole and proline-specific endopeptidase respectively (Fig. 12). Surprisingly both products lacking either ring B or C maintained significant inhibitory activity comparable to that of the original ancovenin. Based to this result, only ring A seems to be responsible for the exhibition of enzyme inhibition.
IC50 = 6.8 x 10*6 M
*Oia: oxyindolylalanine
Fig. 12 Ring opening of ancovenin.
-68-

Chemistry of Lantibiotics 439
Fig. 13 Structure of lanthiopeptin.
The third lantibiotic we studied was an antiviral compound, lanthiopeptin, found at Bristol-Myers Research Institute, Japan.13) Lanthiopeptin was isolated from Streptoverticillium No. L337-2 and shown to be effective against Herpes simplex virus I and some other bacteria. An amino acid analysis showed again the presence of meso-Lan and threo-$- MeLan. Besides these, erythro-p-hydroxyaspartic acid (Asp(OH)) and
lysinoalanine were found as unique amino acid components. FAB-MS and Edman degradation were carried out in a manner similar to that for nisin or ancovenin. The structure of lanthiopeptin deduced from the structural analysis was shown to be very similar to that of ancovenin (Fig. 13). The only significant differences are in a replacement of Asp with Asp(OH) and the existence of a lysinoalanine residue which corresponds to the positions of Dha6 and Lys19 in ancovenin.14)
H-Ala-Arg-GIn-Ala-Ala-Phe-Phe-Gly-Pro-Ala-Abu-Phe-Val-Ala-Asp-Gly-Asn-Abu-Lys-OH
Fig. 14 Structure originally proposed for Ro 09-0198.
-69-

440 T. Shiba
Meanwhile, the structure of an immunopotentiating lantibiotic termed Ro 09-0198 was proposed by Kessler et al,15) as shown in Fig. 14. As seen in this structure, this lantibiotic is quite similar to that of lanthiopeptin except for the position of a counterpart of Ala residue of lysinoalanine. We compared samples of lanthiopeptin and Ro 09-0198 directly by NMR and HPLC and found that both are exactly the same compound.I4) At the same time Kessler et al. revised the structure of Ro 09-0198 to the correct one, making it identical to that of lanthiopeptin. 16)
Furthermore, cinnamycin, another antibiotic isolated by Dworidi et al. in 1954 but of unknown structure,17) was found to be the same compound as
lanthiopeptin and Ro 09-0198. In addition it was soon realized that duramycin18) has a very similar amino acid composition to lanthiopeptin.
The only difference was found at position 2. In duramycin, Arg2 of lanthiopeptin is replaced by Lys2. Therefore, a similar amino acid sequence was again suggested.14) Leucopeptin was also found to be identical to duramycin.19)
In summary, the structures of the lantibiotics so far found may be classified into four groups with respect to structural features,20) as shown in Table 1.
Table 1 Classification of lantibiotics.
Number of Amino Acids Number of Thioether Rings
Nisin Tvpe
Nisin 34 5Subtilin 32 5
Ancovenin Tvpe
Ancovenin 19 3Cinnamycin ZLanthiopeptM \Ro 09-0198 )
19 3
Duramycin(Leucopeptin)
Epidermin Tvpe20)
19 3
Epidermin 22 3Gallidermin 22 3
Others201Pep 5 34 3Mersacidine 20 4Actagardine 19 4
-70

Chemistry of Lantibiotics 441
3. SYNTHESIS OF NISINFor the synthetic study of lantibiotics, synthetic methods for the
preparation of lanthionine loop and introduction of dehydroamino acid in the peptide chain must be exploited and established. As for the synthetic route of lanthionine peptide, there are three possible ways, as shown in Fig. 15:
(a) NHCHCO..... OH HI
ch2----------s—
NHCHCO-I
—CHR
(b) ..... NHCHCO........................... NHCCOI II
CH2—SH CHR
--NHCHCO-- -—NHCHCO.....|ch2------- —s—
|-------- CHR
,
-S
(c) ..... NHCHCO........... ............ NHCHCO
CH2—SH HS—CHR
Oxidation NHCHCO.......................NHCHCOI ICH2------S-------- S--------CHR
Fig. 15 Synthetic routes for lanthionine peptide.
Route (a) seems to be the most accessible way to build the lanthionine peptide, and this route was actually employed by Photaki et al.21) However, this route may encounter severe difficulty in the selection and control of protection of four functional groups in lanthionine residue.
Route (b) is interesting from the viewpoint of biosynthesis of lantibiotics.22) A crucial point in this route is the inevitable disadvantage
that the addition of a thiol group to dehydroamino acid may not give a desirable enantiomer as a single product but a mixture of diastereomers. In contrast, route (c) seems to be very promising, since the cyclic disulfide peptide is readily available according to the usual peptide synthesis using the protected cysteine residues. For desulfurization from cystine peptide by a particular reagent, P(NEt2)3 may be employed.23*24)
-71-

442 T. Shiba
NH2l *CH2I £
—NHCHCO—
A2pr
/\ HCHO
NaBH3CN
McI/KHC03
NMe2 I *CH2I *
—NHCHCO —
\(_NMe3
Mel(leq) /
KHCO3
CH2— NH-C^CO-
H
CH2II
—NHCCO-
Dha
Fig. 16 Synthesis of dehydroalanine peptide through Hofmann degradation,
A-S-f---------0--------- \
H-lle-Dhb-o-Ala-lle-Dha-Leu-Ala
(1 -7)
Segment I
! B Ct------------s--------- ,
iD-Abu-Pro-Gly-Ala-Lys o-Abu-Gly-Ala-Leu-Met-Gly-Ala-Asn-Met
1 (8 - 12) (13 - 21)
• Segment II Segment III
D
Lys-o-Abu-Ala-o-Abu-Ala-His-Ala
(22 - ^8)~
Segment IV
Ser-lle-His-Val-Dha-Lys-OH
(29 - 34) Segment V
Fig. 17 Five segments for the total synthesis of nisin.
Much caution is required for introducing a dehydroamino acid whose double bond may cause degradation or polymerization of the peptide. We exploited a new synthetic method for dehydrtialanine peptide from a,p-
diaminopropionic acid (A2pr) peptide by application of Hofmann degradation as shown in Fig. 16.25,26) Dehydrobutyrine (Dhb) residue was successfully prepared by dehydration of threonine residue in peptide with carbodiimide-CuC1.27)
-72-

Chemistry of Lantibiotics 443
D-Cys lie
Boc-
Boc
Boc-A
Boc
Boc-
LTrt HCI
ONSu H-
Trt/
ra
Boc-
A2pr Leu Cys
Boc
Boc- 6
HCI ONSu H-
Boc"~GNSu H
Boc-
c HCI ONSu H-
Z
L
L
L
L
V
kl
Acm -^•OMe
Acm ■^-OMe
HC1/THF Acm ^OMe
Acm ^OMe
HC1/THF Acm^OMe
HC1/THF
12/MeOH
A-A
P(Et2N)3
Acm ^-OMe
Acm OMe
Acm ^-OMe
a-OMe
■OMe
Fig. 18 Synthesis of the lanthionine intermediate of ring A in segment I.
For the total synthesis of nisin, we adopted a synthetic strategy in which five segments containing A, B, C, D-E rings and C-terminal chain as shown in Fig 17 are successively coupled. According to this strategy, we can check the identity of synthetic coupled segments I-II and IV-V with authentic samples of nisin (1-12) and nisin (22-34), respectively, which are obtained as degradative products from the whole nisin molecule either by enzymatic or BrCN degradation.
-73-

444 T. Shiba
Boc-D-A(a-lle-A2pr(Z)-Leu-Ala-OMe 1) Pd/HCQ2NH4
2) HCHO/NaBH3CN90% (2 steps)
Boc-D-Aia-lle-A2pr(Me2)-Leu-Ala-OMeMMIMHtMMHtHIHI
Mel/KHCCh tDMF (+MeOH)
91%
r _S----------^ 1)TFA f----------S-----------\Boc-o-Ala-lle-Dha-Leu-Ala-OMe -- TT„1/w ^ HCtH-D-Ala-lle-Dha-Leu-Ala-OMe
...... 2)HCl/MeOH
Z-lle-Thr-OH
EDC/HOBt DMF-DMSO (2:1)
83% (3 steps)
rZ-lle-Thr-o-Ala-lle-Dha-Leu-Ala-OMe
EDCHCl/CuCl
DMF-CH2C12 (1:1)
74%
f S \ 1m NaOH/DioxaneZ-lle-Dhb-o-Ala-lle-Dha-Leu-Ala-OMe -------------------------- •
........ 50%
------------------------- ------------------------ <1 f-------s--------v
Z-lle-Dhb-o-Ala-lle-Dha-Leu-Ala-OH
Fig. 19 Synthesis of segment I containing ring A.
Synthesis of the intermediate to segment I was carried out as shown in Fig. 1825) by elongation of the peptide chain followed by desulfurization of cystine derivative to lanthionine derivative. For removal of the NP-Z group
of A2pr residue, catalytic transfer hydrogenation was applied to give a free (3-amino group, which was then permethylated using DMF as solvent in the final step of the methylation with CH3I (Fig. 19). p-Elimination of the
quaternary ammonium intermediate in Hofmann degradation was accelerated by the addition of methanol to the reaction mixture to afford Dha derivative. After removal of the Boc group, the ring A part was coupled with Z-Ile-Thr-OH. Dehydration at Thr residue was carried out by 1 -ethyl-3-(3-dimethylaminopropyl)carbodiimide (EDC)*HCl/CuCl and finally the segment I of TV-terminal part was obtained after saponification.
-74-

Chemistry of Lantibiotics 445
Synthesis of segment II containing ring B was performed as shown in Fig. 20.28) For this synthesis, r/zreo-3-methylcysteine is required as the
starting amino acid. This thioamino acid was prepared from D-Thr through aziridine ring intermediate stereospecifically, as shown in Fig. 21,29) The original stereoconfigurations in D-Thr were maintained in the final product of 3-methylcysteine since double Sn2 reactions occurred both at ring closure and opening steps.
threo3-Me-D-Cys
Boc-
Boc-
\Jn^-OSu H-
/Tn 87%
Boc-
Pro Gly
-OH Boc-
-OH Boc-
Cys
TrtBqc—/- PCC/HOSu
zTrt
•OSu HCIH-
■OSu HCIH-
82%
Acm*—OBz\
Acm*-—OBz\
HCl/AcOH Acm Z-OBzl
Boc-o-Abu
Boc-
-s------------- s-
-------------s------------P(NEt2)3
Acm 77 %
OBzl
90%
a-OBzl
41%
•OBzl
1) HF-Anisole (9:1)y 74%
B
T roc-o-Abu-Pro-Gly-Ala-OH + HCIH-Lys(Z)-OBir
EDC/HOBt in DMF 90%
Troc-D-Abu-Pro-Gly-Ala-Lys(Z)-OBuf
l)Zn/AcOH2)HCiyTHF
86%
8f------S------ \ 12HCI-H-D-Abu-Pro-Gly-Ala-Lys(Z)-OBir
Fig. 20 Synthesis of segment II containing ring B.
75

446 T. Shiba
Desulfurization reaction proceeded satisfactorily to give methyl- lanthionine derivative (Fig. 20). In this desulfurization, two plausible routes as shown in Fig. 22 are considered. However, actually, the reaction went through via route (a) to lead to the desired threo form, perhaps due to an electron donating effect of the methyl group; otherwise an inversion may occur via route (b) to give the wrong configuration. Saponification of the methyl ester at C-terminus resulted in an undesired decomposition. Therefore, we chose benzyl ester for C-terminal blocking in this case. After the simultaneous removal of Boc and Bzl groups, the amino terminus was reprotected with Troc group. Finally, the ring B part with Lys moiety gave us the segment II derivative, as shown in Fig. 20.
H
/1) CH3NH2/CH3OH
2) I2
Trt = (C6H5)3C , Ms = CH3SO2, Z = C6H5CH2OCO
Fig. 21 Synthesis of //zreo-3-methylcysteine from D-threonine.
— 76 —

447
(2S.3S)
(a•S
lch2
aXHjC-C-H
—NHCHCO- -NHCHCO-—(ft)
threo -3-Methyl-D-cystelne(b)
P(NEy,
P(NEt2)3
H-iC—C—H
■NHCHCO NHCHCO
—NHCHCO.........NHCHCO-(2S.3S) (R)
fhred-Methyllanthionine
--NHCHCO........NHCHCO —-(2S,3fl) (R)
e/ylh/t>-Methyllanthionine
Fig. 22 Plausible desulfurization pathways in ring B formation.
C
Boc-d-
1) HF-Anisol (9:1)2) TrocOSu 80%
rTroc-o-Abu-Gly-Ala-Leu-Met-Gly-Ala-OH
Nps-Asn(Mbh)-Met-OBuf
HC1 in THF 96%
HCIH-Asn(Mbh)-Met-OBuf
EDC/HOBt in DMF 90%
z----------------S---------------- \Troc-D-Abu-Gly-Ala-Leu-Met-Gly-Ala-Asn(Mbh)-Met-OBu‘
1) Zn/AcOH; 2) HC1 58%
13/--------------- S---------------- \ 21HCIH-D-Abu-Gly-Ala-Leu-Met-Gly-Ala-Asn(Mbh)-Met-OBu t
Fig. 23 Synthesis of segment III containing ring C.
-77-

448
threo threo3-Me-D-Cys Ala 3-Me-o-Cys Cys His Cys
I2/CH2Cl2-CF3CH2OH=10:1
Boc-AbU
OH Hl)BDP
IIJTsCI
90% MeOHBoc-AbU
P(NEt2)3 in Benzene
Boc-D-Abu-Ala-D-Abu-Ala-His(Tos)-Ala-OMe
1) TFA; 2) HCl/MeOH
78%
quant
91 %
quant
70%
78%
Boc-Lys(Z)-OHr---------o \
HCIH-D-Abu-Ala-D-Abu-Ala-His(Tos)-Ala-OMe v-------- s--------- /
EDC/HOBt in DMF 69% (3 steps)
Boc-Lys(Z)-o-At^u-Ala-o-Abu-Ala-His-Ala-OMe
NH2NH2H20 89%
Boc-Lys(Z)-D-Abu-Ala-D-Abu-Ala-His-A)a-NHNH2 22 V------ s------- J 28
Fig. 24 Synthesis of segment IV containing ring D-E.
-78-

Chemistry of Lantibiotics 449
In a similar way, segment III involving ring C was synthesized as shown in Fig. 23.3°) An acid amide group of Asn residue was protected by the 4,4'-dimethoxybenzhydryl (Mbh) group. Removal of the Troc group gave the desired segment III.
A cumulative D-E ring moiety was prepared by a simultaneous one- step desulfurization reaction from the corresponding biscystine peptide as shown in Fig. 24.31) After coupling with Lys moiety, the segment IV
derivative was obtained, which was converted to a hydrazide for coupling with segment V by azide method.
29Ser lie
Boc-
Boc
Boc
Boc-
Boc-
^BZ'k—OH H-
Bzl
kiBzl
His Val Dha
Boc-
Boc
•OH H-
Boc-
Boc-
Boc-
TosOH H-
/Jos
Jos
Jos
34Lys
ZlJos
Jos
HOBt (6eq)
/Troc josOH- Boc-A2pr-OH H-^OBzl
Boc-
-OH H-
DCC/HOBt
/
i) Zn/AcOH
Dha
i) DCC/HOBt
i) DCC/HOBt
i) DCC/HOBt
Troc
Troc
Troc
ii) Mel
ii) TosCl
ii) TosCl
ii) TosCl
^-OBzl
Z 75% ^OBz
Z 73% OBzl
Z 82% l^-OBzl
^OBzl
Z 72% ^OBzl
^OBzl
Z 80% k—OBzl
OBzl
z 62% k—OBzl
Z 81% OBzl
Fig. 25 Synthesis of segment V of C-terminus.
-79-

450 T. Shiba
Segment V, corresponding to the C-terminal linear hexapeptide was synthesized as shown in Fig. 25.32) Dha residue in this part was also
prepared from A%pr residue by Hofmann degradation.Couplings of the five segments I to V prepared above were carried out
as shown in Fig. 26.32) The synthetic intermediate obtained by the
coupling of segment I and II was confirmed by comparison with benzyloxycarbonyl derivative of nisin (1-12) which was derived from natural nisin by trypsin digestion. The coupling product of segment IV and V was also identified with the authentic sample of nisin (22-34), the BrCN degradation product from the original nisin, after deprotection. The final coupling of the synthons I-II-III and IV-V followed by deprotection with HF in the presence of anisole afforded the crude product which was purified by HPLC. The synthetic pure product thus obtained was completely identical to natural nisin in all respects including 1H-NMR, FAB- MS, HPLC, and antibacterial activity.
1 (--------°-------- siZ-lle-Dhb-D-Ala-lle-Dha-Leu-Ala-OH
Segment I
8/ ° \ 12 , HCIH-D-Abu-Pro-Gly-Aia-Lys(Z)-OBir
Segment II
l)EDC/HOBtinDMF
2)TFA55% l)EDC/HOBt
in DMF
2)TFA40%
13^---------------o--------------- \ 21 ,HCIH-D-Abu-Gly-Ala-Leu-Met-Gly-Ala-Asn(Mbh)-Met-OBur —
Segment III
22 f------ S------- \ 28 Isoamyl nitriteBoc-l_ys(Z)-D-Abu-Ala-D-Abu-Ala-His-Ala-NHNH2 ----------
Segment IV
29 34Boc-Ser(Bzl)-lle-His-Val-Dha-Lys(Z)-OBzl
HC1/THFin DMF
TFA
1) TEA2) Boc20
Segment V
l)EDC/HOBtinDMF
2)HF-Anisole(9:1)
10%
Nisin
3) TFA4) HC1
25% (4 steps)
Fig. 26 Couplings of segments I to V for the total synthesis of nisin.

Chemistry of Lantibiotics 451
Thus we achieved the first total synthesis of one lantibiotic, nisin, sixty years after its discovery. This synthetic work also provides a novel synthetic method for the preparation of dehydroalanine peptide as well as lanthionine peptide.
In a recent study, we investigated the structure-activity relationship of nisin employing NMR conformational analysis.33) Besides several synthetic
segments for total synthesis of nisin, we prepared more than ten kinds of fragments or analogs for this purpose. Biological tests for those compounds suggested that the minimum structure required for the exhibition of antibacterial activity is the N-terminal part (1-19). Furthermore, Dhb2 and
Dha9 cannot be replaced with the corresponding saturated amino acids
without loss of the activity.Although we succeeded in the total synthesis of the typical lantibiotic
nisin, tremendous efforts were needed particularly for the preparation of the sterically pure methyl cysteine, desulfurization step, and introduction of the double bond in amino acid residue. Such disadvantages strongly inhibit the further development of the chemistry and biology of lantibiotics by limiting the supply of necessary samples.
N
N
N
Fig. 27 Biosynthesis of lantibioitics.

452 T. Shiba
On the other hand, a biosynthesis of the lantibiotics has been studied in relation to that of other peptide antibiotics. However, it was earlier suggested that the lantibiotic is not formed by a multiple enzyme system like usual peptide antibiotics, but synthesized on a ribosome from a precursor peptide as in protein biosynthesis.34*35) Lanthionine or methyl-
lanthionine part is formed between cysteine and serine or threonine residues on propeptide, which is connected to a leader peptide at the N- terminal side. After a thioether ring formation in the propeptide between cysteine and serine or threonine, and also dehydration at serine and threonine residues, a process to remove the leader peptide will occur to complete the biosynthesis of the lantibiotics.20)
We are now attempting to utilize this concept of biosynthetic process as a tool for the preparation of the desired peptide. The idea is to achieve an enzymatic synthesis of lanthionine peptides from artificial precursor peptides possessing cysteine, serine and threonine at appropriate positions but without the leader peptide part. Such precursor peptides can be easily prepared using techniques for conventional peptide synthesis.
In fact, we investigated an enzymatic formation of lanthionine or methyllanthionine from synthetic peptide substrates involving cysteine and serine or threonine using cell-free extracts of 35 different kinds of microorganisms. As a result, some species of Streptomyces and Str eptov erticillium certainly produced 3-methyllanthionine in the hydrolyzate of a product from the substrate corresponding to the nisin precursor. We are now developing this strategy to establish a novel and general preparative method for the lanthionine peptide in collaboration with Prof. H. Yamada, Faculty of Agriculture, Kyoto University.
AcknowledgmentsThe author expresses sincere gratitude for the invaluable contributions
of excellent collaborators to this study. They are Dr. T. Wakamiya, Dr. K. Fukase, Dr. T. Teshima, Dr. K. Inami, and many students at Osaka
University.
References1. N. J. Horn, D. B. Jones, and S. J. Ringel, J. Biol. Chem., 138, 141 (1941).2. L. A. Rogers and E. O. Whittier, J. Bad., 16, 211, 321 (1928).3. A. T. R. Mattik, and A. Hirsh, Lancet, ii, 5 (1947).4. E. Gross, and J. L. Morell, J. Am. Chem. Soc., 93, 4634 (1971).
— 82 —

5. E. Gross, H. H. Kiltz, and E. Nebelin, Hoppe-Seyler's Z. Physiol. Chem., 354, 810 (1973).
6. The sample of natural nisin was produced in Aplin & Barrett Ltd., Beaminster, Dorset, U. K. and supplied by the laboratory of the late Dr. Gross.
7. K. Fukase, Doctor's Thesis, Faculty of Science, Osaka University, 1987.8. Y. Kido, T. Hamakado, T. Yoshida, M. Anno, Y. Motoki, T. Wakamiya, and
T. Shiba, J. Antibiot., 36, 1295 (1983).9. T. Wakamiya, Y. Ueki, T. Shiba, Y. Kido, and Y. Motoki, Bull. Chem. Soc.
Jpn., 63, 1032 (1990).10. E. Bayer, E. Gil-Av, W. A. Konig, S. Nakaparksoin, J. Oro, and W. Parr, J.
Am. Chem. Soc., 92, 1738 (1970).11. T. Saeed, P. Sandra, and M. Verzele, J. Chromatogr., 186, 611 (1979).12. M. Nishikawa, T. Teshima, T. Wakamiya, T. Shiba, Y. Kobayashi,
T. Okubo, Y. Kyogoku, and Y. Kido, Peptide Chemistry 1987, pp 71-74.13. N. Naruse, O. Tenmyo, K. Tomita, M. Konishi, T. Miyake, and
H. Kawaguchi, J. Antibiot., 42, 837 (1989).14. T. Wakamiya, K. Fukase, N. Naruse, M. Konishi, and T. Shiba,
Tetrahedron Lett., 29, 4771 (1988).15. H. Kessler, S. Steuernagel, D. Gillessen, and T. Kamiyama, Helv. Chim.
Ac(a,70, 726 (1987).16. H. Kessler, S. Steuernagel, M. Will, G. Jung, R. Kellner, D. Gillessen, and
T. Kamiyama, Helv. Chim. Acta, 71, 1924 (1988).17. W. Dworidi, O. L. Shotwell, R. G. Benedict, T. G. Pridham, and
L.A. Lindenfelser, Antibiot. Chemother., 4, 1135 (1954).18. O. L. Shotwell, F. H. Stodola, W. R. Michael, L. A. Lindenfelser, R. G.
Dworschak, and T. G. Pridham, J. Am. Chem. Soc., 80, 3912 (1958).19. S. Kondo, M. Sezaki, M. Shimura, K. Sato, and T. Hara, J. Antibiot., 17,
262, (1964).20. G. Jung, Angew. Chem., 103, 1067 (1991); Angew. Chem. Int. Ed. Engl.,
30, 1051 (1991).21. I. Photaki, S. Caranikas, I. Samonilidis, and L. Zervas, J. Chem. Soc.
Perkin 1, 1980, 1965 .22. L. Ingram, Biochem. Biophys. Acta, 224, 263 (1970).23. D. N. Harpp and J. G. Gleason, J. Org. Chem., 35, 3259 (1970).24. D. N. Harpp and J. G. Gleason, J. Am. Chem. Soc., 93, 2437 (1971).25. T. Wakamiya, K. Shimbo, A. Sano, K. Fukase, and T. Shiba, Bull. Chem.
Soc. 56, 2044 (1983).26. S. Nomoto, A. Sano, and T. Shiba, Tetrahedron Lett., 1979, 521.27. M. J. Miller, J. Org. Chem., 45, 3131 (1980).
Chemistry of Lantibiotics 453
-83-

454 T. Shiba
28. K. Fukase, T. Wakamiya, and T. Shiba, Bull. Chem. Soc. Jpn., 59, 2505 (1986).
29. T. Wakamiya, K. Shimbo, T. Shiba, and K. Nakajima, Bull. Chem. Soc. Jpn.,55, 3878 (1982).
30. K. Fukase, M. Kitazawa, T. Wakamiya, and T. Shiba, Bull. Chem. Soc.63, 1838 (1990).
31. K. Fukase, Y. Oda, A. Kubo, T. Wakamiya, and T. Shiba, Bull. Chem. Soc.63, 1758 (1990).
32. K. Fukase, M. Kitazawa, A. Sano, K. Shimbo, H. Fujita, S. Horimoto, T. Wakamiya, and T. Shiba, Tetrahedron Lett..,29, 795 (1988).
33. D. E. Palmer, D. F. Mierke, C. Pattaroni, M. Goodman, T. Wakamiya, K. Fukase, M. Kitazawa, H. Fujita, and T. Shiba, Biopolymers, 28, 397 (1989).
34. A. Hurst, Adv. Appl. Microbiol., 27, 85 (1981).35. C. Nishio, S. Komura, and K. Kurahashi, Biochem. Biophys. Res. Commun.,
16, 751 (1983).
-84-

Sg48

FAB4*va0^ ft 1®———
FAB K • E6M
e* • S6@f • Tffi Sot
1 . (1UAC
11 Z 0, SfLvW t >
^t: ^ o T # "C ^ 6. ^(7)ST,
fb^55h#f!t Fast Atom Bombardment (FAB) %###&')
i), ^tT.b<Dffimm$tizij:<mi'htLr\,'Z> ^'tix-hh. mz, -m&n, ^zh- F<D->m&mvt(ZtiV^-CIi, DNA^SSB^J^ibSS^ tL/jS6H<7)
t ^ /se8efij<7)B992)-3),5e4)-5), «%%##, sasmiRBtm
<DffiW6)-7) izmti)Xhh. itz, SST-ti,
(^>TAHfi^tt) em
V'Z:®%f514,fk^§: (collisionally activated dissocia
tion (CAD)) / linked-scan ££tC «£ &z<7’-f* K<7)7 5 V
(Df& (± 2 ^ <c 6 t LT&B ^S^TV^. CCTIi, FAB
cv'T^mf 6 ^
2 . FAB MIM
m 1 fi FAB 4 * >M£gMLfz~mz$inmfrtit
-eofMli, 4 *y®, Mi%, m#,
4 c(7)gB^6a:o, m^n^nMmma ±cSB#tnaas,
6. (#&B^^fL-cw&
SfCS^flrlt (JMS-HX110) Xlt, Eit¥S72cm,
fiDilEElOkV -Cyjil^flliM.OOOT*^^. ) FAB 4
f y^-C, 6keV (ih'-etDilLT-j^
( + t/y#)
Application of fast atom bombardment mass spectrometry to analysis of protein primary structures.
Toshifumi TAKAO, Yasutsugu SHIMONISHI Institute for Protein Research. Osaka University
01 fab nMfrmt<D&*&m.
txvtv y#<7)7 h v ?tizgm<vmmzj:io]E'{* y (NH+) &V'L#d':f y (MB") t
l vg/M\ <vmm-cm%i±<D mmizm^L, 1 ^zm-yx\±, mo-lOOpmol (DWfrZ-Z'^ti-Z.
tfz, ##%&##-?#.#%% (tfV^uyXVn-A^)
•loE, oi^
o. 8M5e^lU±, k: - * <0 s/N Jt,SENSES i)\ nw2000&TX~&io Jl-E-i 5-10mu
CAD/linked-scan &C <t 4 * y?j?.SCT*< ty^7t-^7%yy xgf&co n u y 3 >
-b^rtT'^V 2 <b 1:ioT-wu frttzwm.i%(n&2 (e) «bm#
<7)msss (B) (Mtir-TElz&hZfrbWi^Zmi ~t h ifllj5e f£ (B/E linked-scan &) l~ i *), #5e<?)SC
Kt5^^ELZ:7 7X7 y h ■{6. cfoiiijS'eii, -tetc,
#>< (DK# (lOOpmol-) L, iSiJ^oT
m&, ^^Z-E"2500u
6.
3. E6H, ^*7> K<7)-*imSpyf
sen,2 iT,
ZW:,^ * ;v ,-h'4^ y ^ y-f- *f~ -tf (CPase) cz^mfL^
ft'ifziivii-sL^zfc txmm l, nwftmzxt)^ tub<7)^Sx^<^ h;u^$ij^i-^>.
(40kDa~)
85-

(42) # # # Vol.32 No. 1 (1992)
(esi
iBnil^r-?-K3i£Hi| |^t+k|
(FABKl-StfrCAD/linked- iceni£))
T1 h/V- 1
ONAttZKiT"'-,7J( nytfju-
—v< >-/-<■/T|
( FABWagJtftW)/"'rTTtii'fi'""..
/ toting{?VC x-f-HX')
i|7C*«gl
Ik • gQSto-ixnufi
iZ:, Kt-&2:KTV'6C t &&&C)•C, o'? ^‘77^ -#HZ 19
K9 v\
7<
dnai&mm&burn$utz7< ;mz^J % ii t iz, o > \£ j. — ? — u Z 0 j$l$,, [uj/E L, 7^ 7.^-7 < k&£-£
Wo IK fd f( co <7)11111^1? I i, vx/1/7 < Kft'i^fi*#LZ:^7'f- KMteOEHvxh^ K&f^
LT&CZ,? >/>'»
UEi]2;IV^,2<h/M0, 0>-f-\H'y7)U7 1 b'&£r<7)iZmi)ikfctZ>ZttfX'£2>. ifz, # ifiT'li, 7.U7 hvXT’l'-'ft'sIt (ESI) H5b> WJ) HZ 0, ititf-f-SIK6HotH-fi£uWMM-
£ <)uThE-C%I9, fUfZ 0#^^%
>(<: iH’f 6 2 <h 7)?-C£ 6. —~Jj, ip.—K, &6 v>
li, CAD/linked-scan
?J;-t: J;l77/^H'Z ii'i^EiWf 6 2 t *C7
ttz, h ;i/ <7) j$#r H & v > T-ci^^^FAB
f(S5>rrnzofomimafuat6. mmh/:tt!:7;7^M, &6V'W, 7;
viKE^jz of^^^fL-cw^^emL^Ksail, c(D#e#c7:3
>t*a-y-f:J: 0 nTH^x^Mf^JOaiL,
DNA^m@6^em^3c <bnzo-cf C07 $ j ma.
UZXlZM;-f62<h fcftl
$ ti6 ma^<7)-^#mn
fab^M^rrnz
n^mzt£ht^k.bnh. (asn,
W%H Z o T^JMT/M-: $ ixtz E M>y-7j.o>-aA(D h V 7v >mik#(7) FAB HV5)
frTFLZ:. E#l£ iitz v ;u£ ^ > t? .x - ? -v'-c, cDNAtii^fld^j^ibS^snrv^ k: h-r>
y-7i07-aAW7; / ^Sd^jH##f 6 2 <kH
z0, -xffi&tDmfcZftvfz.5. CAD/linked-scan ;i(C J; KC7^7g|
6E*|J>*5£CAD/linked-scan £fc*C (i, FAB 'f OftClil't
500 1000 1500 2000 2500 3000 3500
03 Z;IHHI'Cj?g£fe.£;fiZ: t h 4 > if - y x n y-ah <nh V Z-> >mib^<o fab KSU^? h ;u5).
— 86 —

fab k
Kr LTAaEf ^ 77yyyh7fy
62^:^T#6. %c"C, |H]Lm^:'e%Kf^(fX:7
>Mt>(±, -ted, WZfi&#mc Z 0EI4 (:
7^-rio n^^$ti-cv^^10), mmzii&y? y
y y h7 tycoSE, £J$7 7 >sii^7,7 KcotiH CZoT^^ 0(O#7)%6.
El 5 tCftL^sE ^D7'J 7 L Sj|<7) Achromobacter 7°0 T 7--tf#<l:#^6#^ LXz^T'T K<9 CAD/linked - scan 7^7 h 71/LtL. ±(C, y, b v V -X(D7yyyyMfymWSK
^uc, Esj^nzzvyFWNS (Thr-Ser-Thr-Ser-), &CF,
(-Thr-Ser-Pro-Xle-Val-Lys) frb<DT l J Z> z ttfX' £, i>t<D'<'f1- Y<DT 5
y 6*65?!] $r Thr-Ser-Thr-Ser-Pro-Xle-Val-Lys <h $: ^L^lU. (Xle Id Leu t tz\± lie -TtVb 2o(7)7 5 y^MnSMCS^-czcox^y
*J yj Zj X; y? Z; X, y, z,
aJ i>, c, a} b3 c2 a, W c.
0 4 CAD/linked-scan &(C.tittS^77 Y<F>17 7*
UEyiJ'CSt'V^. ; V
7yyyyh7^y^zo%mt:%#f6B
%-c,Lid; Uli'fflv'ibtLTv^. #$:%I:#btizr-
-MMfrmtTj: o# a tvt-) A-e#<g-£>.
e. maLTdj I),
#(:, MS6H^(7)Sll<7)W2)-l3) ^SE7-a^x.$6W^MjS mx± tzmmymtmm&vMm5' c fab
At:mv^KTv7)L Rdjv'-ai,
k#%i±mc (16: AfammmTmmzigibfL.
3i e SteT hZ.t frX # 6 * ■XfetbX ;fiTMT&£v'
x.6. L^L, K<0?WMrC(i. /J#AmTl#7
bmmmfnii^mth -t nw«^ -1 t^wa
3tifB5-e<7)zfliJ^^"TiigC^O, ^#%e^tf7< y%. *&V4i, ^7*
ti£ i> 11£g#K%T6tcSM£f#
62L^f-CS6OTimSMS/c^J: 0??£,CffA
Z>. LIT, t'/MGTP^!56H (hyyxra- y y) z^yjL--/ h ictiit>5>^BS^<7))WrrM) coi'T$-<£.
(43)
Thr-Ser-Thr-Ser-Pro-Xle-Val-Lys (204 —211)
200 400 600 800 m/z0 5 7 O 7 V y L iA<D Achromobacter 70 t 7 —tf
iH(L$t)'J|tE L/J^77 F (204-211) (7) CAD/ linked-scan ^ ;i/"\
-87-

1 ̂u-c
V'^ma^T, e(39k), /?(35k), y(8k) 030(7)"thT'^L— v hri'bte'DX^'i). a V-3. — v h (±, jfc
/? y4fyjL-v h^T-CGTP^^mc^
/?, y 4fy:L^ -y Mcoo-Cfi,
mmcnoLT^^Qq^A^v^, y-v-yjL
-•y hdtirSHM • TFrSItM («tri-7 KcW
6GTP^^#ei%mt6^1±) <7)2#(7)^f-7!,%
#5*14S!, ^F?5i4S! / -tfT'jL - v h -cii, ‘frtibnS. aureus V 8 ^O f 7 - HPLC
(44) £ #
PG-C3;%/<7?-K7 7/;<>!' (Leu- Lys-Gly-Gly-Cys) IZ&75<M.\,'t: $ tl, y-t%%- v
h(7)C^%7 < y Cys-70^<cA^T)'(7)^B
2<7)C*%^y^K
# (m / z : 681.438) T'7- K Srtil/Si't" -5) 5 £§5£c0
7<y%z5^:t#:$fL6# (476.242) Z 1)205.196#^
< rnwzK Cisiizs
tpTiz 1/ x T □ - iW'jE'fri&MUXfeftXft birh
# # Vol.32 No. 1 (1992)
681.438
Leu- Lys- Gly-Gly-Cys
205.196476.242
681.438
Sesquiterpene(Farnesyl)
1 10 20 Pro-Val-Ile-Asn-1le-Glu-Asp-Leu-Thr-Glu-Lys-Asp-Lys-Leu-Lys-Met-Glu-Val-Asp-Gln-
30 I------- 1 40Leu-Lys-Lys-Glu-Val-Thr-Leu-Glu-Arg-Met-Leu-Val-Ser-Lys-Cys-Cys-Glu-Glu-Phe-Arg-
50 60Asp-Tyr-Val-Glu-Glu-Arg-Ser-Gly-Glu-Asp-Pro-Leu-Val-Lys-Gly-Ile-Pro-Glu-Asp-Lys-
Asn-Pro-Phe-Lys-Glu-Leu-Lys-Gly-Gly-Cys
0 6 7 ->8!IKGTP$a6-3ieff ( h 7 2Xf *-'>>)r + ya-7 hcOC*^^7-/- K (66-70) O FAB
h^14’, %(7. y-try's*--v ho
-88-

FAB 5%### (ogm ^ -/f- K - m a R (45)
fctZZtizJ: V) z.7. £t?*JlZ
mw,(:^A6<7)T, 2t, 10;
^ K3 2 ^ ^
b, K, sagtF
4-%, $ 6
I:, (fmol) (7)ma^(7)7
±CZ3^m^^(S(D-@(7)%m75##$ft6. ^
2 LTff 9 ^
X.h^-7'^- Y(D7 5 smm&7Z, ESI Hfi^-rrltt:-eur, ^i»o7i>
^77^-tllSl/: FAB *6V4i ESI WS^tfrltmij^coamkcz6^#i#i:ij(7)@m,
M'T, Cti6<7)rrLv^S^rr»f<7)fUffl i>tyz.X < i)<Dt&£>tlZ>. ,
x m
1 ) Barber. M„ Bordoli, R. S„ Sedgwick. R. D., Tyler. A.N. (1981) J. Chem. Soc. Chem. Commun, 325-327.
2 ) Takao. T.. Hitouji, T.. Shimonishi, Y.. Tanabe, T„ In-ouye, S.. Inouye, M. (1984) J. Biol. Chem. 259, 6105 -
6109.3 ) Gibson, B. W„ Biemann. K. (1984) Proc. Natl. Acad.
Set. U. S. A. 81. 1956-1960.4 ) Fukuhara, K„ Tsuji. T.. Toi, K„ Takao. T„ Shimonishi.
Y. (1985) J. Biol. Chem. 260. 10487-10494.5 ) Takao, T„ Kobayashi. M„ Nishimura. 0.. Shimonishi.
Y. (1987) J. Biol. Chem. 262. 3541-3547.6 ) Carr. S. A., Biemann. K. (1984) “Methods in Enzymolo-
gy", ed. by Wold. F„ Moldave, K„ Academic Press, London 106. 29-58.
7 ) Takao. T„ Yoshino, K.. Suzuki, N.. Shimonishi, Y. (1990)Biomed. Environ. Mass Speclrom. 19. 705-712.
8 ) Biemann. K., Martin, S. A. (1987) Mass Spectrom. Rev.6. 1-76.
9 ) Fenn, J. B., Mann. M„ Meng. C. K„ Wong. S. F., White-house. C. M. Science 246, 64-71.
10) Roepstorff, P„ Fohlman. J. (1984) Biotned. Mass Spectrom., 11. 601.
11) Takao. T.. Hori. H., Okamoto. K„ Harada, A.. Kamachi, M., Shimonishi, Y. (1991) Rapid Commun. Mass Speclrom. 5.312-315.
12) Carr, S. A.. Roberts. G. D. (1986) Anal. Biochem. 157,396-406.
13) Nishimura. IL, Kawabata, S., Kisiel, W„ Ease. S., Ike- naka, T., Takao, T.. Shimonishi, Y., Iwanaga. S. (1989) J. Biol. Chem. 264, 20320-20325.
14) Fukada, Y., Takao, T„ Ohguro, H„ Yoshizawa, T„ Aki- no. T., Shimonishi. Y. (1990) Nature 346. 658-660.
15) Fukada. Y„ Ohguro, H.. Saito, T., Yoshizawa. T„ Aki- no. T. (1989) J. Biol. Chem. 264. 5937-5943.
-89-

-Pa 'f * > «fc 5 SSffc t f %##-
*e*^8¥® ft m *& je
A molten globule state has been assumed to be a major intermediate of protein folding. The conformational stability of the acidic molten globule state of several proteins and the corresponding state of a synthetic model polypeptide is determined by the balance of the electrostatic repulsive forces, which favor the extended unfolded state, and the opposing forces mainly hydrophobic interactions, which favor the compact molten globule state.
molten globule state/protein folding/acid denaturation/cytochrome c/amphiphilic polypeptide
ti$t%x.btiX£fz.vs&mx, ta'ccm
VfWtfr b X: )U 7- > • V o fcf j. — )U (molten globule)vim t wifti"'" m 6 nvmxkfc&vi&ffiz&zm t
^(7) (±1983%, f- h ? n A c-f h ^ a A c (/) pH 2
A3/f
317- h ? a A c t 6.
EB(±fM%#j&<7)^f(7)3 >/<? h
t'jL-;u^Ǥtu^A/in.
(lit LX. XJUf> • ?'□
L/c^tiSiS t v* o tz 4 ? - >'Xhh i.*7? br^rz • zkm
biZX l)g£lC1975%3U 07?F711/7; >'Cli43i4pH r(7)^iim(:Z 6^11(7)^
^i±pH • 7 a bf jl -
Molten Globule State of Globular Proteins
Yuji GOTODepartment of Biology. Faculty of Science. Osaka University.
btlt>. (CD) Xh-y7,h7D-ifc'trffli/'fa 7 ? h
o, sje
ii&ZtZTFLfz. i£lzX)Uf-> • ?u\zJ.-)Uffi
&&S£KE>t;t V od'<75gS»-CilSffl!|$ti,Ztifz.
%&X)Ut> • ?*o tf JL-;i/C7)£^i±%3fc0£&ft
*0'L'(:9f%iiS»b ibnr S tztf, &<tiz> tZ<7)mmi>&&X£>Z>Z ttftoii'vXbtz. ttzX)V7-y . ya tf * - ;u<73#K fc L-CtMttfc&gg
-eLT, f-h?oA c (DSHt^ramitld:, ^JnzXr^T's ■ 'fu\£x.-7i't^nmmm^v.tz\z<>frfrhb-r, tsm&BWt4 7.: ^6 ;u-r> • rnbTjL-;KC(i^itL*
LtJ‘L. ##(±%^%(:itf
$ 2 $'Z&@%7):*6&•?, f- h ? n Ac 7Ux > • ?*ntf i-; u<o*E>#{;A4i:#x.'CV'&.7; K^n h ><7)3i “-&7C H-NMR £E
txjux> • if-^mi2f7)fZtLi T'C 0 7? h T7U7'$ >5), 7^ 5
*yn ¥>*', f- h ? o l*c' X-&L&2titz. Ztib <V9Z&n<7)*JUT'S • ya tf JL-7i/*i&(:
< t itz^m
-90-

(123)hf.x-;u:tfyg
*> izwm-r 6 a rm&v-'kh-nmr f-
h ? a a c\ V '/f--a9) & <k%'
< ffMSfL* 2
a^$fLTvi6. f- h ^ n A c
a K# £ i o X V .
lx, t?-%.%&<»&C # T^CE^il^-C # tV'-)7l/-
a 7 - ? ^et5 titv>ht)?4>, -e;i/f > • ?*
o h: LT#%t6C <k (iCOtf ;i/t-&tZ.
3c ^<cctzzvmiya
<7)^ti fS^$6H<7)^14EtE>ld:^ <B t C&ftt'C # 6 EE>T-& 0,
t> • yui'J.-)i,ffi&imt}%:Mlzii, £*)&%%
9tlz$&tLtz£ i IZ-‘%HZ*)\st's • ^n tf jL-;u«ji
Privalov • yo b'jL-^ti, Et£#cSk v> b -?? n ^^a-e<7) 5?o&#i&##mmoczcTAi;a ^
• jZnbf
frvm&m&nvztm&V't * ym.mzm- lx±§ < -t;u-f> • ^o
h'jl-
2. ^xccfc5g?^t£ sa
v >£5^8i-s6£S'e&s/?7^? T - -tf L-c%t # <IttZ:12’. B. cereus <Dp7? 9-7-H
It pH 2, #/f f ^^COAf^Trii? >/A3-f /P
#(:%Cl±L-CV'6df, ^ (0.5M KCl) t€^f>-/ok:'a-«»ti. -f- h ^ o a c -C-KEC#2f R LW&X$>6.^i:7*‘U/of011,m, ^ >^-7jlo>
Wavelength ( nm I
0 1 B&oZ:z8$<7)£®E41'C-<7)7v • -f- h ^ □ Ac(7)SgmSBCD*^? h ;u. 2013. &ti li^&OzSS (iaf£limM)£,ivt. £&<i*m£$6S(pH7.5).
/151 * ^-c t -b fi6
c t tffr, $ tt,- ft**) -&&zmmx$> hzt #t> d'ot. ttz, a 7? bT^ZTl XnmS.^ fl'T > • ^o ss#
&i:®mtz>t m#E ^ l /j m it fgf k * *-t z t i>m%ztifzv.
Ztib<D&&m&%\ifemz£^Xi>m&$tit$:& z-fzttfhfr^t™. mk\£, &mmx>izmx&%.
izm&2-£tz?- b ? V j+ C IZ2 biziz&irlmkz, t,
m&migmv^ji'f-y • ^n ^jl-$
ns (Eii). z(Dt$x>mm$zitit£.iz£z>i><Dt £ < tttrv^Zj. pK, (i 3 Z I)*§v>(ot*, (pH2) (cm^^x.PHeTIfT t #^(7)^(1^^ % & V\ cordis
js,
-r> • k' a. - ;ufSit<7)$^k7i?B^ *>IZ£^?
£SSC<k£vWL/_\
^OAc (:)tf &##(?)JS,(02) sj:cf/\A(7)%%%
^Efkz *)jt^LZ:T @E,
;Ut> • 7aeJ.-j\,m&7!>''MJ&$iifz. LfrL, ffimn®i:mz-tx)ilz&mbm'( *jlgcz oT±& < #C:oZ:. t'k<nWBE*C o Zz.
ferricyanide3" >ferrocyanide2'5" >sulfate1'5* >
ccbcoo* >scn‘ >cio4‘ >r > NO3" >cf3coo* >Br‘ >C1' ( 1 )
zzxm&^itpMxm *>x> -Mnn^t7 i/T yf&J * > ttict&'f * ><nmi*W£X(Dm
-91

024) ± # # m Vol.31 No.4 (1991)
HC10TCAHTFAH
NaCIO
.TFANa
TCANa
1000
0 2 222nm t (A) tS (B) HI • 7 h ? Die otiitlrf£. 20r. 18mM £-77*7 h D ? ^C UK^*V'{±a«rflnx./j.
ti CD ^7^; t/ok:
fen
a*6w(i^$rAnf 6 af f#mn#f
6<bv'<c^(0%W'^r^T*6"'.
im&asa-nj:&&*%. b)£il%i (Dcbyc-Htickcl OiSEM) <D 2 O 7> nTfigtl
rthh. 2 hlZc) J 7kco^jZin>tf
6%-^emi:-email 6et <,
til k> lL"C v' Z>.
etVbH^6 3 oco^mco^ -tb, b) (7)%bmii±n
a) tc)
c) (0%#(7) -f 7 > 1C I 6 j§ V' 11 Hofmeister #^ij t LX X
(£n £) titr v > 6.SO.*2" >phosphate2'>F" >CI">Br" > 1 > ClOi'>SCN' (2)
fkf6. -^ja)m?Utm(:.&6C<k7!r'C&6. L"C H V^7;i/7 ; ><o
Sillltt (electro-selectivity) ll^KOMR^ij (-% 9 .
-92-

E6
'MLL:l (lid#)+ ‘(OI-S'CHd) mW&v'M
&C0 J7D4U • ^ -titi UZIZIW M + ^rJ. Z m
o^itirvzm -^n5 y □ 4H +o?mm
iSW-O-^SiBNZ
SMG9M9S ‘3-ntSii^5vnz>i
f!%$ N ' Q /$ n ^ y a <= 4 4r
. z.4
JISZU -9^ 9 Of $^Ba/ • <)3IJ.C. / > z ';ri|^aai
<Z^Xc'Z:&^'t< £>'CfiC(<24-mt-)Vrlr?&'%:% '0^9^BM-G)?-T.3
a/ •
O vin4H ^qHr-£4Z 'E
•^zn^ ?
r 9 °- g >93
a a/; • <^4rgr•9^a^9 3M-3-j? G ^ :?-£#*G>< * ymx\ ?lf
- <^ira- '%<%*
(SET)
a.<: • <^'ifi.'/iiilWrH n0.^:5if»#3i$
ssn ‘Ctffxi4^<fi?)#&Q><*ym?&z$gm
'WZ a n
3,mmnwmzc'ctyziv*
G)9"if-3-?9Y^^3@:i^9^?f:ir^ '9^0.1#
( p ) on — x n
•9?^Ic-T:i^<4>^1^94-?(rSr?^ (ON) E#
'V-TJ n.4 • <^-/if9r '? ^%^O^M&Z94-?
^9?^ (n) emom9m%m
•9 fail:) i TG>¥m&B15#iG>M9SB3E
^9T:K^mr^ ■9vUi^?:9'U^
BG>#3#E# Q T:1?T 9^.^^5%^49#my<
-t>^G)*^^MB3IG)MgS?)?ra?r "9^%
O'Ojfamr) ((E) teW) %%#9_^^:)§m%^ (i)
9'A
ZL & 3 Ti & G> fiiW Jd)si3uij0H flZLtlort^ -£ >G?B
- ' > %,? i)< -K: ^< 4: >#( E ) _J = .003€HD< JO
<^8<%N<_ I <.NOS< JOO<^OS<_z^OS
BOftof — ^ Pi njf • <^./if3rG)g8@:#&

(126) A % # m Vol.31 No. 4 (1991)
Ztz.
pH 7 XX> CD 7^? h ;M±S#n.
7-tf ffe^. &
% * * >5£jS'C<0itS*'t-CD %/<? h ;Hi7-fef-
a^'J -/
m 4 A Ii222nm
LT-/n v h Lfzi>vx-bh.20t *C*li£j 7 ffiCDT-t-T-JlsItlZ >• 7'D
kr J. - $ tL/j.HI 4 B ItjE^gmXmtXJU-r > • ?*o
ttmv-ytmtiRjzLxnnLtz. *>u-r> • yo
(AG) T%
£. 7-k-f AGx&iztitz. &&<Dm£frbffii&-mM*)<ntfrn& &K%7jli, IOC (:& wCli460cal/mol. 50t T* li
-10 -
Number of Acetylated c-Ami no Groups
J2 o -
Number of Acetylated c-Amino Groups
04 f- h 9 n A C OT-b^MtCi £222nm <Of»FW.$ iL (A) £ *;vf- > • ro^i-;u«i2Jf?f£OdlfH
(D). 18mM HCI (pill.8)im#lilO (Z). 20 (O). 30 (O). 40 ([]). 50t (^).
280cal/mol X2b S 2 b. bfrb ta o fz.
4. c & # a w^
g6J?<05L&fiiili$ t
(^tepH, gE#^JO#;#^ET) i:&v>
-C£ftSii<OE1±t: 5 xm-f 2>±$%mb £>'£ 6 2t <)75$tvrv^161. 2(0J: -9
*7&‘«bi>&^x&z>j)\ %mn&x>n WZ'D^'XliZfBfiX£>Z>. T-t^Juit-f-hfn&cV)
5-k.h. ^h?DAc<0PH7 +
8-C^S. pH 2 h ? □ A c <7)^^(i24t* * & <D XM W 8 CO ft? ti li 1610 7 -fc f-;i/ fk $ ti tz ft ?-l:#^f 6. EJ4 B i 'ombfrbZ 1 C167-fe-7;u itr Y^uuc itJ±ffl&jizx)i'-r> • yo kfi -MS
pH 2 tpH 7 £&-</.:£ £, pH 7 C
v\ h ? da c
5. Lys^Leu^b^^"Ex;u^';^7*^K(Dmi x>izj:zm'&&&7)
zX)Vt's • V n t: jl- ;HS it HTH-
(Acetamidomethyl-Cys) - (Lys-Lys-Leu-Leu)s- Lys- Lys~ Asn-Pro-Gly- (Leu-Leu-Lys-Lys^-Tyr-NHa O
6E?iJ tii'DXrii'** V ^y-f- K(0^j&$rK*X:. 2(0 ^h" V ^ 7*^" K li Lys-Lys-Leu-Leu 7)^ 0 j§ LSS^'Jt)' b%*) 41 'L' I- P 9~'sL#V^ Asn-Pro-Gly X> BZPj&S'tf. ty^rxum5 iz7jxtJ:i^2
Anion
0 5 ^f)b+;,J^7'f KdPM f > C (Al'f L /_f «'. v n6 yz^miZiEf?. □ < ■> > ^ xx.
-94-

6$^fk t ^rvrnm- (127)
v ?
Z&tfctZZt imftLtz.^ KdlOmM h V Xt&mSflrB (pH7.5)
c#-ei±7 > /An /( ;i/T*6 C <b^CD h
bbfr^tz. frZlzMi<Vtz,
ce#ut.
f#ii$rf£;W1± pH EE>
%-Dtz. mj * ><D^$j)1&l±M$L<&Mi&’g.X-'&b1xtz )\m (1) tJK-SL*. 2<7)^(i, -£t;v*’V^7‘f- K7)?4,'t± pH fffMti o V -y ^ X##(lg
;i/T> •ii-C£»), Z ^ T^fb$ fiTw6 2 &
-;^-t> K(D#%(±%*1±7
^xlFmcm^Tv^C <bl±%6^-C*6. LZzt)^
Tgfi^c7)^;u-r > • /a t: jL-/l/#ac6%7k## ICfEffi7)^4L T v > &aj 1^147)^5v\ $6So^;vf > • /o l:
') y -f 7 9 U >X;U7 * >g£7)r*££-f h Z t, ifzb ') 7* b 7 r {> 2 £ $:
h. ^)Uf-> • /n
2<hliSS&t$E-C&^.
&fc>U (:
- /nfcrJU->M?|j££ «fc
t;i/-f> • /ntfA-
f ;i/-r> • 7‘ofi-^E AE
^-^TliCrL'
frt%z.btiZ>. Lfr L, />%#] & (i• /o i:
t>75'oTk'£V'^fi7^V\ f ^oAc
K^CjtmSZ
)FMf6m8%emv'T,
m'<xftinmfz%mffltf&btiz><7)x-{t%'s'frt%ATV'3.
::mt/:^-gji±* v 7 f ;i/-7±#a. l. Fink mtn)~li), #*E
bis±,7) t <D%HWfftv&z z fti-m use l it.
£ 6$
1 ) Ogushi. M, Wada. A. (1983) FEBS Letters 164. 21-24.2 ) Ptitsyn, 0. B. (1987) /. Prot. Chem. 6.273-293.3 ) Kuwajima. K. (1989) Proteins 6.87-103.4 ) Kim, P. S.. Baldwin, R. L. (1990) Ann. Rev. Biochem.
59. 631-660.5 ) Baum. J., Dobson. C. M„ Evans. P. A_ Hanley. C. (1989)
Biochemistry 28. 7 -13.6 ) Hughson. F. M, Wright, P. E.. Baldwin, R. L. (1990) Science
249. 1544-1548.7 ) Jeng, M. -F.. Englander, S. W„ Elove. G. A.. Wand. A. J..
Roder. H. (1990) Biochemistry 29. 10433-10437.8 ) Roder. H.. Elove. G. A.. Englander. S. W. (1988) Nature
335. 700-704.9 ) Miranker. A.. Radford, S'. E.. Karplus. M.. Dobson, C.
M. (1991) Nature 349. 633-636.10) Griko. Y. VM Privalov. P. L_ Venyaminov, S. Y, Kutyshenko.
V. P. (1988) J. Mol. Biol. 202. 127-138.11) Privalov. P. L, Tiktopulo. E. I.. Venyaminov. S. Y.. Griko.
Y. V.. Makhatadze. G. I.. Khechinashvili, N. N. (1989) J. Mol. Biol. 205. 737-750.
12) Goto. Y.. Fink. A. L. (1989) Biochemistry 28. 945-952.13) Goto. Y.. Calciano. L. J.. Fink. A. L. (1990) Proc. Natl.
Acad. Sci. U. S. A. 87. 573-577.14) Goto, Y.. Takahashi. N.. Fink. A. L. (1990) Biochemis
try 29. 3480-3488.15) Arakawa, T, Hsu. Y. -R.. Yphantis. D. A.(1987)Biochemistry
26. 5428-5432.16) Kawata, Y„ Goto. Y_ Hamaguchi. K_ Hay ash i. F., Kobayashi.
Y.. Kyogoku. Y. (1988) Biochemistry 27. 348-350.17) Goto. Y„ Aimoto. S. (1991) J. Mol. Biol. 218. 387-396.
-95-

5S:

(m 3 kx^w5S6)
F h -E-CDMiaM#±# ##@P#sfk## _h!f&£
1. tiXtotc
^7- KA^X E®^4Sffiitffl4'A'T*> ffi^tc^X-dSfeilSttttrti.ST-fe*
SATfc'DTiA if-6ibA'i:i'Atfe%if5SrojitLfc^»(cM-rs. -£©gliti\ ti.ltttffi
C@#Ri#=f;axa) •I®i9 4’41StttiI(:f^T, Sx AS©#®*!
#Jf>SLgEStttc-e^.l$k'S<M3U4>i,'k5g-xe>ATi.'fc3 ktcS-5.
tttiift., &^©A^%-fl:#©#%©m#ck 4,» tiltroE^SiSHiKSSAEIfc^
v, m#a%t%k©3± • m'i*•?■ ti6$zus<ay!©tt^SL®
A 6Sf U Ir'ESSS-S^MSs AooS5 ©T-, 3 3 TH-e ©#%& X'5C#mX
5^i:(ct5.
2. %^#Yb#%#&©#&
X<%6ATt'6j:3C, H'©SX#tti9F35®W©-o
t&cfcA^ ^©WSS®gT-ffl^^f-Kffl#ffiS*©i(i^ttti«©®l;9EtiAg&»Si&#
xfc„ 3©#%A'6, (i##) f A6
©&aaaA#mc^#gj%Tu6. jm®r# • #g# • mm# • m## • %
S#&' i: tc Affl S AT v' S AX CA6©m%C*T&AafimW:?%T&6. -fxKX>®e
»i:©-g;©#g#A%A#JM%©A©^-^v h t LtftltJlfc. itiu;ti, TA6©w%&mcT*e• *v3&#Amm?sjmz-sti'*.?
A KAS#A^###C*X^>#&##F: LTfc'tttAti, fft&yvftk l/tSIJJiSit
tc&ofc.
XfflW5EiS@T-ti3 Si^t)KfflWJSffljiisi:fet'*oT, {&£t»A*4gX-S^affittt)
K6S5fcto©jgilS^4#)t^$&ro#S5 ATSfcz ^ ^SATtt&'^&'V'.
3. m#k%&#©MmA*t=AWT6m#&%V#<mm@©T##T&6. Sit-*4*.5t'ttf4i«
A & 5 ® @ © T SEA' AX 4'BT«fb#&m#:%»7ti'0-^C j: V
3A6©##CM#X^&a%##KASn< cA'AiSAT W6. 3 A'Dffl&'A'tc&'tti’A
K#{bo'#l4i'^'& AT A U \ B'Jx.tSFerrichrome-’f' A Jf ASt© <fc 3 & s iderophoretl###:
•r'0>&M'i A>ffl®ai:lf4'SII4SCTt'5.1
-97-

chemical determinant t L Ti£# < & o -?:tl^ 6D#IE(c &^^ #mm t m# ^ ^6 ^#' 6,
S©T% ^-fphytotoxinCEE^ciOUTj&^S A A -£tl £ (D o (Dhz-o
V'T$E l£t"5o
host-selective
-4 5A -* 3E# —■► $£Ai
i
Fig.i ^
3 — 1. fittefSij@illffl?B3£#IH4(Host-specificity of plant pathogenic fungi)3>1. to®i: !RilC®4® <9^tc 5 „
B»J) $;<3t+F-f: -f 4-
7 : ISSlfS
2. 1: < C#i#to®T'W:#M#C#LTl%—to®Tt##i± (SSttsusceptible, S)®#1: jSSttt (resistant, R) -5 „
fi*J) th :>■$$# : -+ttE (S) , e+BE (R)V l/=l|S*i?*#i : f>K - f'J i>7% (S) , C$ (R) x.yiXV Victoria blight# : -? h U 7 (P c 2 , S),
#S ( P c 3 8 , R)3 • ssttts#:
4. eistts#: efflffiMttxe^s^mfimtuewms=»HST, ■y-^b-yfl—^KSiSStSBcD^ih7>?SdEMSSCS§tS*A':fea cOT'ti* V'*'?
2
-98-

3 — 2 . %### (Phytotoxin)
M) >, 7 V 7#, Marasmin, Tentoxin, Marformin, Cyl 2
Host-specific toxin(Host-selective toxin)
2.
3.
5. HSffl) AM-toxin1, 2) : U 7 ternaria mail)
AK-toxin1, 2) : 7 t'MM.JHiAlternaria kikuchiana)HMT-toxinu : h^-^nniyzr^ imtl^KHelminthosporium maydis race T) HC-toxinn : h 77:0 3 v Zil£1,'£$itlfff(Helminthosporium carbonum)
4. 6phytotoxin (^F#|li$LTd, Vt/% - n^f K - 7^377X7 - $0# (X^7U 7)
t&ftW (7 hf - 4V 3) T(Csst"-5phytotoxinT1 Xx7 K^
4 — 1 . CoronatineA## : Pseudomonas syringae pv. coronafaciens
vibotoxine<5 coronafacic acid, coronamic acidXif Vitti:& 0
coronatine
3
99-

4—2. Rhizobitoxine: Rhizobium japonicum (X4 X(DW&W)
: irreversible inhibitor of ethylene biosynthesis
: 7^ JMCDX-J £ tc&i
V\i K L =
H NH
H NH
rhizobitoxine
4 — 3. TabtoxintkWtW : Pseudomonas syringae pv. tabaci
^ — LactamSieHN
*OH
CH.
CH.
RICHOH
h2n-ch-co-nh-ch-co2h
R=CH3 tabtoxin
R=H (2-serine)tabtoxin
4 — 4. PhaseolotoxinAz## : Pseudomonas syringe pv. phaseolicola
: irreversible inhibitor of carbamoyl transferase
7ho
: phosphosulfamyl tripeptide
?N H-P(0H)-0-S 02 NH2
(CH,)]
H2N-CH-CO-a la -h.arg
Phaseolotoxin
4
-100-

5. 5phytotoxinA) #######5—1. Fusaric acidk Picolic acid A## : Fusarium oxysporum f. sp. lycopersici
Pyricularia oryzae
ho2c
R
R = (CH2)3 ch3
R = (CH2)2 CH = CH2
R = (CH2)2 CH(OH)CH3
R = H
C02H
5 — 2. Marasmin: Fusarium oxysporum f. sp. lycopersici
A###:
NH —- CH2----CH — - R
ch2—-CH NH
co2h C02H r2
R, R2C02H ch2conh2
co2h CH2CH(NH2)C02H
co2h ch2co2h
co2h H
H CH(C02H)CH2C02H
C02H CH(C02H)CH2C02H
5 — 3. Tentoxin 411 : Alternaria tenuis
: chloroplast(D%f
Me N ---- CH C0I L
HN ---- CH
Lco
CO---- C H 2 ----- NH CO
H
N Me
Ph
5
-101-

5 — 4. Malformin: Aspergillus niger
(malformation)
I 0 0 L D L |
NH — CH — CO — NH — CH — CO — NH — CH — CO — NH — CH — CO — NH — CH CO
C H-> — S — 5 — CH- CH/ \
CH, CH,
CH,
CH/ X
CH, CH,
C ,HC
5-4. Cyl-2: Cylindrocladium scoparium
yy * ^ — i> 3 y tetentoxinC8n{£l UTt'S. HC-toxinCD^l$:Mck 0
b)
5 5 . AM toxin1, 2) Biological activity of AM-t ox ins and synthesized analogs
%/f~(CH2)rr_C—H (D
\
^ H
ch3 ■/ch3
XCH-C-
IH(4) X
X (3) H—C—-R ' % /
X(2)
X ,0
X Y z n R Toxicity(ug/ml)
1 AM-toxin I 0 =CH2 OCH 3 ch3 0.002-0.0002
2 AM-toxin II 0 =CH2 H 3 ch3- 0.02
3 AM-toxin III 0 =CH2 OH 3 ch3 . 0.02-0.002
4 (L-Ala2)AM-toxin I0 -ch3,--h och3 3 C„3 >100
5 (D-Ala2)AM-toxin I
(L-Tyr-Me\) AM-toxin I
(L-Tyr- Me]L-Ala2)AM-toxin I
(L-Tyr-Me]o-Ala2)AM-toxin I
(L-Amb^)AM-toxin I
(L-Amh^)AM-toxin I
0 -H,--CH3 och3 3 ch3 5-10
6
7
8
0
0
0
=ch2
-CH^ H
-H,—CH_
,och3
• 0CH3
0CH3
1
1
1
CH3
CH3
ch3
10-20
>100
>100
9 0 <H2 0CH3 2 CH3 0.01
10 0 =CH2 0CH3 4 ch3 ,1.0
11
12
(l-VaV)AM-toxin I (L-Tyr-Me]L-Val^)AM-toxi n I'
NH
NH
<H2
=ch2
och3
och3
3
1
ch3
ch3
0.1
0.1
13 (L-Abu )AM-toxin I, *3
0 -CH2 och3 3 CH3CH2 0.1-0.01
14 (L-Leu )AM-toxin I 0 =ch2 0CH3 3 ch2ch2ch(ch3)2 0.01
15 (L-Phe )AM-toxin I 0 : =ch2 0CH3 3 CHgPh 0.001
6
-102-

43= B : V y^M&MMffi(AIternaria mali)
4sstt: m®m (K+omm)*«
5 — 6 . M-toxin1, 214Se® : i- is(A1 ternaria kikuchiana)
4SSE: StttiStoTSC, M««8 (K'fflttiS) ©%#AK-toxin I i: II Alternaria kikuchiana(AIternaria alter-nata
Japanese pear pathotype)^4#4(c jt o X (la) t (2 a) fflJdCMAUfc11. ■?-©«> 4#6(=J: V,T'Si-f ?3’S SDR®© A. fragariae(A. alternata Strawberry pat ho- type) /r h= AF- toxin I , II it HIyAz U >brown spotifilE© A. citri (A. alternata Tangerin pathotype)A'e,ACT-toxin I b k D b ;Wgi£ tt, -itl <b ( 2 a) (2b) (2 c) t (3 a) (3 b) m «k o ttfc2'31 „ SI 1 tibfe XdfiffliHifi
X ti 6 ©S4#Sto#Stt*SX*^S5»cZ)MEEti5Ad-' 5 =b ©©
V'Tttt) (8A( 9S)-8-hydroxy-9,10-epoxy-9-methyl-decatrienoic acid 8 fii©7Ka-y 5 VEifc-M'tiia-k h'n*S/i£Sfi»&T-x;t =f Mt£
&#Lxt'&. -£©fc®, ttxv'3*!4-’\ 5C jt6 ©#4##%#m©&K#m©^fBKX,&toiri$SF%6BI?fiL4-51, aTfcifi^SJt t) itStttft
AK-toxin t-tttSh Ux>*;b^>^©^f5JSE«tt [£,Z,£] (C@$S tlT »\ AF- toxinXHmM#i@^#4 6 [E,E,Z] , [i,Z,£] , [£,£,£] © 3 #© jltt&rikiaie S *1
Stt©%$kS$ TKSXtCig-g-Ufcy 5 Kn*©###©*mc(J:44 V©g4i:##@#m©6A, z©gm©^4##
Sto4a«St$©%$tcM4-UTt'-51:#xe.tL5„3ffli)4'l,Sii'8, ;b it t);h-t2AK-toxin II (lb) U _hlf T> ^®(t¥6)7
* 1 (c)n44%#%©mm46
1) t kd+ b ux> • vx> • h udt
itB^LTt'5.2) *S©A;i/xK*i>;b$6xxy;HbLXt.gi4{cH:E4ti4tt„ x©x it
^5"5T|gtiE65HSUTV'd„3) gtt©%31tctt, k h'n^i>x,K*J/A;i/tJ<>®m8,9fit©l6>tfSaS 0?,S) ti-K-itT-
7
—103 —

$ •£> „4) y$ v^gES-ffl2’ffi<y#6*fS5Eti^ST-tt»v'0
5)
o
COOH
(la) AK-toxin I : R = CH3(lb) AK-toxin II : R = H
R'0
COOH
(2a) AF-toxinl :R = C0CH(0H)C(CH3)20H(2b) AF-toxin II : R = H(2c) AF-toxin III : R = COCH(OH)CH(CH3)2
R OH 0
COOH
(3a) ACT-toxin lb : R = OH (3b) ACT-toxin lib : R = H
Ei 3 . n h ^ ?i jb iK tj it t zF n — V <nmi#,#) (^^)
8
-104

1.
R 2 R i Configurationof C i and C 2
Toxicityto Plant
Ac-L-Phe -CH=CHC00Me= R, S +Ac-D-Phe -CH=CHC00Me' R, S +Ac-L-Phe -CH=CHC00Me' R, R —Ac-D-Phe -CH=CHC00Me' R, R —Ac-L-Phe -(CH=CH)2C00Me’ R, S +Ac-D-Phe -(CH=CH)2C00Me’ R, S +Ac-L-Phe -(CH=CH)2C00Me* R, R —Ac-D-Phe -(CH=CH)2C00Me‘ R, R —Ac-L-Phe -(CH=CH)3C00Me“ R, S + +Ac-D-Phe -(CH=CH)3C00Me- R, S + +Ac-L-Phe -(CH=CH)3C00Me' R, R — 'Ac-D-Phe -(CH=CH)3C00Me* R, R —Ac-L-Phe -(CH=CH)3C00Me‘ " R, S + +Ac-D-Phe -(CH=CH)3C00Me'* R, S + -PAc-L-Phe -(CH=CH)3C00Me‘‘ R, R —Ac-D-Phe -(CH = CH)3C00He' ‘ R, R —
( ~P ) til 1 0 - 3 m o 1, (—P ) til 1 0 ~4mo1necrosis^f^^ti- 3 0
( * ) tiitrans.trans-transfc'cfct/trans-trans-trans&o
(1 1) ti: trans-c i s-t rans& 0
9
-105

5 — 7. HC-toxinQcMMh 0 ^Enn is ££&$itlM(Helminthosporium carbonum)
H v
CH,H .0
N"A-c'
/ H/ \
: H H C-\ /
N : H A,
/zC—C
/°\
HC-toxin
revised by
Walton ./ of. (l982) Gross •> o/.^1982)
Pope 11 <?/. ^19 8 3 j Kawal •/ o/.^l 9 83^
Rich ./ ol. (1983)
VTv\
CH,—C-H\ v
0 | HCH,
H — C — (CH,)j-C — CH
AHC - toxin
proposed by ./ ol. (19 8 2)
\ ' U /\H — C-(CHjJj— C — CH — CHj
A
OCH,
Cyl-2
Hlrota *1 ol. ^19 7 3 j
V /
of"-c'H \
N'
CH,— C • H H C—(CH,) —C-CH
\
,c~9
CH,Chlamydocln
Closse •' oi.
Y ° ,»\ , \ li / \H —C —(CH, J, - C — CH — CH,
Cyl-1
Takayama «r ol. ^1984 j
CH,
kv’I'C/
CH CH,- C-H
/ \CH,
C//
CH,
\ ° 0 \ , x " /
H — C — (CH,),- C — CH-
AVH WF 3161
Umeha hare
(1083)
OCH,
1 0
—106 —
CH,
Llesch
\-CH,
(1974)
CH,
« I ol.

5 — 8. Victor in (HV-toxin): x y^Victoria blight^S(Helminthosporium victoriae)
:glyoxylic acidnP##*U ^V'>12) ==>04#®Victorin C-receptor CD-^.]H13)
125I-Victorin CCDbinding protein(lOOKDa) £: R, Sv dz -y ^ ^ ^ nT
(?)
HO-C-OHCOOH
NH Me'
CHCI
1 Victorin C ECjq = 37 nM
6 . (D*ffe
6 — 1 . bennzoxazinone#^£phytoalexin
avenalunim I (benzoxazinone)
dianthalexin(benzoxazinone)X; — ^ —3 > (i"T
1 1
107-

6-2.
HERBICIDES containinmg C-P bond
O
HO-P-CH2-NH-CH2-COOH
OHglyphosate synthesis
inhibitor of 5-enolpyruvylsikimate(EPSP) synthase0
ho-p-ch2-ch2-ch-cor1 z z I
OH NH2
bialaphos analogs natural products
inhibitor of glutamine synthase(GS)ch3 ch3
R= NH-CH-CO-NH-CH-COOH bialaphos Streptomyces hygroscopics
R = OH L-isomer : phosphinothricin (active phytometabolite)DL-form : glufosinate (active phytometabolite)
H3C, .CH3 CH
ch3 ch2
R = NH-CH-CO-NH-CH-COOH phosalacine Kitasatosporia phosalacinea
OCoHcO-P-O+NH,' synthesis
CONH2
fosamine
O OHHoC-P-CH-COOH synthesis
J 1ch3
inhibitor of keto-acid reductoisomerase
7. tfTTf
#### (phytotoxin) & d Z.TM&'n'&CD
1) fb#, 39, 161-167(1984)2) 1 9 8 6^8^#, 72-83^-^.3) (lb#-^#fil 1 4) 13-22(1988).4) R, P, Scheffer and R. S. Livingston, Science, 223, 17-21(1984).5) R.D.Durbin ed., Toxin in Plant Disease, Academic Press(1981)
1 2
-108

me*:

D992.12.3. mm/cyf
_tum a#
1. it DA 1C
0%*u:agf ^t#ma#a?KaaTsih'-css. i,m:A 6®&#)%mg#mf i±&#
H*tc#«t>&$ttfc» >/■?»*, a*, ^jiaie^E^y^AtcE^js^fcs
V, ^#1C$&*j»U<, 100%tcEv')6eg#ia*$:mU, L*'fe
$$T-65rt;i6'jiD&hTV'5. Ch.g,l±A%®*aE*#f »±T@A
v® P - n Jg£ 4 mat i ■> U 3 y *» «»li 10-18% ®%*&##*!c k if to T fc'U, 3X ImcttSiKItK’]®?:/.) left*)■$.AS«%«KiMffl3 ft* ic ItSo "O'
*v\ EoT, x*;i/^-StSi:i6«ltai«iaiHKeEVO'5W4-, ^^68%6:<
x;i/t t a * M&* * cc k14 *?«. A'o«gfc»« k *
o TV' 5.
)t6j$*Tlt9no7.ut/Zf'jtS$#lc8%S$£8:E&®fcLTV'S. *«T
li9nn7^;v®#&ttSkt*Sgtc&BU-7c>, >ti£Ji!WRT»b:h,TV'a)fc*®®
®fe e.t Lfcigx, c*vtTlc&* €,*vfcy6-&fi8Si* tfcfcAX©««g*mtcc5
V'T, **g*mg*m®S>)'68EBiU 3 e>tc4fcffl®»&iB'<*.
2. icfe'It5 it
2.1. 7>tA63R
» n D 7 W /!/ a (Chi a) tt@ 1 ffl#ifi&% *>, 7t 7 7-k DT#
6U 662 nra ICjRitUiA&feo. UA'Ufctf 6, 7>yA Chi a lt»>X9®k
#£L, 0 2 ICjRI-J: e, lC»#©6^#&#l&DT#tf »tA#iR*lt®%®&/
7 HU, A^KNtlKA^oTt y#<©*&%%f &Ck A^T3&". tfc,
-&*®»%#A&&A'%10 nm T-Di/7 hUTfcy, MU*-'*? MU k -f )fc IV* 7 HI/ k 6'd t < %% y, ForsterS®x*;i/¥—#S6A'5Sll*j; <@Z3 t7lC<Ro
-109-

-on-
¥-0 B iMD
3 OSHQ :?94»»3i¥ '30^ng#3iT## 20“S %gi*a-3»%###0?
• 5 I M3 |Of3##OSMO • 5 TO ? (DVAd) c*i*c* ‘33 <C>/^0##9'V##4M 6>/Y:0
* a. 1*9«<?#!»<•£**/? •* 113 9^*f#%n ?#^#0 e 113 c.%5¥$W3igt# "™ S89 '3* (1) S09'(*m*04M 6>/¥<A6## Z(OSHd-® 113) #e#c»9¥
“u 689 stf * ii3 a.*Sf®9-l? (oshq) }i/s-*sMr*<ir■**/;-¥ % A/A OS ‘¥1 ,i1*c*1*c* Z(0'H-« 113) 8 1134^31
(8) S013^^1® OOAd C.%?¥S»»31 ®u OOi ¥1<«/C—/|O?0C|Z»B)|
9S*0»mi$v4%»0>$:iq)Y)'X^ *> A™i*¥l3Bf»9«0*
?ft3J*W0*in»4f? nv/3H/oil /1*¥4'S >,S <*a:'Y5¥ia.#»9^0l.m »S6K ‘*¥?*»3ijfH'i$ (38) ‘-3'*3fS9^ st^JtA -a ¥t<s9*E-"s*A5{«F
"9if $#B^?7 93a.^##g*K0'l-4YfH <( n A :1 $ $ 1* ? 311«( 9 Ba 311 T 9 ® 31 #i(( * ? ? 1* ? g30 Tf 3Tja ¥ 3 Cl ^ a ■=■ ¥■
31 *(f #*¥©-* >4 (38) /If>/:nn<?4fi4<?>.<'<!'*3ia- "4¥##3i (i)
t-B3i*3g05t* *4*31 (B) tBa.g%ii#9#il@0*4x< fin m^3iiuim 'M2S0iHF#95WW^39fit 3U<*9 Bit3##*##¥*=0*3. ##3
'YS^tHFigW 'T%W3i#*E#N!Y?q)##0#ff WSfS^'l? stPUlA 8 #«^¥9H!$l„9"l»l"BBtB0*S( • (9 3i<*mW
«) 94m93i%i¥-^#m03jn*»3i^#^#^f3##gjrm94#M# P-c.$<i •94^9?ftTfar-i* "17@!> ¥$%%####¥ "19#^ -
B mo f Y?1*¥IT? OOAd #¥3 089d ¥131####
/\i y can <C0'*51S z z
'9<11C.%23 3##/W4&0*n 8#3132%:*94* n d /?*ffi¥<?y/3in ^#M0a<?$■/< *v<Mr yc n n <f
4:4<^93$#3iv-/:a n ^93J.##%#0#WW^¥9# ‘9t6¥t,ti*<* i*c* -^1*$%<*?::c3iM#¥4<z ,ftJfa&«>fi(iKi®9'¥WM
<?^X0|D82g93lc.¥4¥3l (1) 8M¥MH <CxY¥/403: '9fit%$)^K%<?y< 6/ e no C35/1M <?5vif)!h®0 («) eBiA-?)##? * 113 ^n»S!3KUA0 (X a/a 9) #^31##>p0 (VS8) < $ .Cdf ZOT1*¥I ceW^C*
m«na^(f > ¥3¥¥t3-'it33ii<#0/i/yc n n a
44<Z '4)g3lZ0%W-04H tf^mqi»0^/-caa/J4i<Z0###S! •9’iia^55i3lC.¥?'39 0W3V)'*iySXOO131$1¥l-*'ir*T¥=Vniili® ‘(<33-

(Chi a• 2H20)n , ### Chi a
mVtz. CA&toESlc4k1ti*h&iS<ktfE# Ifc KjfcSEE
4k k
3.
3.1 7 □ a 7 y ;i/ CSS3tE?6]
n-SiNI&k UT*0 6ft4 ZnO SffitC'O'tf *-V X t- UTt#6*lfc
Chi a Sfl$&ttabT7yT7-7nn7^;t/&«LfcSS)te?a»' Trlbutsch k
Calvin9’ 1:j=?T$a#g^t (0 6) . Miyasaka 6 181 ti Chi a LB
tt&ani n-SnOa 12% T^ESgS:toifflUfe.
3.2 i/nn7^b [*££■»• y K-f y ■fDgEflS]
Tang k Albrecht11 ’ l*jfc«ES±lcESi!6TSgS ftfc (Chi a1 2HaO)n »®
®JhKttSM8iro/.hSfc A1 S: XSSS ic * ^ T«J8 Ufc-fryK'fy *13-lz;i/ Ag/
Chl/Al SrflMKbfc. Dodelet6,8lli Ag/Chl/Al "b7l/®7 a n 7-f 71/7 it--A0je
AE 77-0 Chi 8 (Chi a - 2KaO)n 0*^iev$
*)tb*)ti;3,i±-S'*ri<vt'-;i/7;i/3-;i/7d’;i/A,k 7-feh>g®»'6>*7xh
UTtlfc Chi a »Hk®a*S-y-> K-f yf-tlb Au/PVA/Chl a/Au &(#RL, 8
a^fflkJgeEA&aaibfc. chi aZPVA#0l:/\f (
07)
3.3 c-igjgE***^]
mmTm^f/A**mA^Ea#0#kLT#%T*4:k&mA60T,a
l&E«k;ES#<bS*-eW6K;16as#t,4d(Ufcfn-;i/, rKU-N-x^;vkTn-;i/,
jKU (N-k'-;t/*;i/z<V-;t/) , dfUf* 7%y, >tfv O-x-Tyt/f-*7iy) »
if®a«®±c*SE*Si? a-jf5>B^7jcy»77->»kT06#&aa L, se.tc ai &Kaa»vTsarsa^EaasiF' auzis*bv6szai &#«Lfc,4>. C#ve.®a^liv'-flT.t)*ST-C Au EffiKE®EE&RliDUfctil^a*
fokfc4B*fttii$:jftLfc. *fcz;ti6cye&aif1-4k, *®m#*AlKNa» <#tc Au E&fftfE, Al E«0A'Ak 4)16iBEE8: £ b fc. L6'U»»6>, E SEttE^a* tc l*E£ k |iWc$#E«*7 - * y Tifc 4 CIO. - -T * y # K -/t y hk LTBtV&SftTfc'tJ, k l: «k ? T6$& 4 w* AlkSJiSf 4r ktJ:oTfiEa®«(fc^ Al Effi®«A$:5lser L,EL<mTg-K-Cn4k#A.6ft,618’. 4kV, K-.rty hOEttE^b^WKK -yicjt^Tae.vfckk4, -3 vraK-^uferifu o-xfvt/f x-7%y) (
-in-

PMeT) fcV'T 1" CtvT V 4 k k A'*> A'o fc.
081C PMeTtoRF-y (K-7) (R
B) &mn6^0#?Au/PMeT/RB/Al ®Ba@88)tiS*E 860 mV, ft
570 nA (*SI*Ii=1.5 mV cm"2) , 7/71/77 9* - 0. 14, 0.0048
XfejRkbT, 5-X^7l/-10-/<>i7;i/-5, 10-i7fc K □ 7 x7-j7>»aitt: 1 k 5 &
9) . kfte>®*El±«MTT'**fcS*ftftfli«:;itL (010) , #BtJt r. r. k L
T-t-A-fiv 1,200 at ±4 V ££tf 12,000 at ±2 V k H 7*t*jt*8E#6SEk b
ktfl?#, PMeT k®$£WB-e&fcfc)fc«g#Htt&5tLfc,T’. MElK^eeE
650 mV, @#*#zft 190 nA OftSMElr=0. 17 mV cm'2) , 7 -f 71/7 7 7 4> - 0. 25,
***#&* 0.018 x fc#jtfc. ^fefflv'fcfREttSflBllSjeeeE 560 mV,
56 nA OfegftKlt=0. 17 mV cm 2) , 7 .<71/77 9#- 0.23,
%$ o.ooi x -efty, *****E-kurtt i®#?i:)tx-c*6A\ #$«SEk . 011 (a,b) ®77i/a >7./<57 h7V®Jti<6A'6,
_L®gEtt PMeTZ(l) 5.®*ETl±,
PMeT/(§) IFB®l$A>i: (5)/Al JWfcWSk bTtt< fcft*£-4'#jfttf;$:.§.jtfck
#*.€>** (012) . k®Cktt^^fia%®e^(RlMA>#@lC<i:^rv'5kkA'
*E ® j£8ft toih K ## U T v' * k k & 5t«1" 5.
4. fcfcVlc
S: ^ r7V k f 5 « «$#3ilE® l± * 72-e to tl ic o V'te « A> U T fe 5
A', <|jR*S<%#At0#S#t.5B^@Tafe5. 6A;bA&3:##?®@2,66
#g&«mi//<7i/?®%9;#m&#gu, -tht#
S6Kttl$:ie3|K<t^r««StS*fc
ltS«tt»^ai«tcKiRi • BS-r 5 k v> d jsrnttrem®, & $» * 7 k lt v> 5.
LB®Si$KA*.5$tis^-T®iB5>E#Slk«SSffiA®EB • BI$i£®B8%A'S*ft*. $ * OTi#6ft *&*m&&?® #8k• /U
=f#EA®j&mtMRA'tT&. *n*>Al±DlK-yS*vA:««tt{bS-&g$tcW#
60t*S3*UMrlMlUfcS#r.»#|iBA^il6*«*K*®#i?65kk6JlfllL fcA<, k®#E&*^^^® 100 x R#$®t®K
f a&AKK, %#®#0&#RbT»A'»Wft«»6»t\ kfte>®«f&tt&$
toKBUi'^ra«#®M«$fkt.A:)t.®T6a.
-112-

1) C. S. French et al. , Plant Physiol. 49, 421 (1972).
2) K. Uehara et al. , Chem. Lett. 1M, 1643; Photochem.Photobiol. £8, 725
(1988).
3) K. Uehara et al. , Chem. Lett. 1991. 909.
4) J. Deisenhofer et al. , Nature 318, 618 (1985).
5) Y. Yasuda et al. , Thin Solid Films 210/211, 733 (1992).
6) L. L. Shipman et al. , Proc. Natl. Acad. Sci. U. S. A. 73, 1791 (1976).
7) K. Uehara et al. , Chem. Lett. 1982. 1445; Photochem. Photobiol. 53, 371
(1991).
8) K. Uehara et al. , Chem. Lett. 1985. 897.
9) H. Toribuch 1 and M. Calvin, Photochem. Photobiol. U, 95 (1971).
10) T. Miyasaka et al. , J. Am. Chem. Soc. 100, 6657 (1978).
11) C. W. Tang and A. C. Albrecht, J. Chem. Phys. 62, 2139 (1975).
12) J. P. Dodelet et al. , Photochem. Photobiol, 29, 1135 (1979).
13) K. Uehara et al., Chem. Lett. 1984. 1499.
14) K. Uehara et al., Chem. Express 2, 129 (1987); 1987, 2236;
J. Polym. Scl. Part C Poylm. Lett. 26, 95 (1988).
15) K. Uehara et al., Denki Kagaku 57, 1121 (1989); 47, 909
(1990).
16) K. Uehara et al., Proceeding of 2nd Internatinal Simposlum on
Chemistry of Functional Dyes (1992.8. Kobe) in press.
17) K. Uehara et al., Thin Solid Film 215, 123 (1992).
-113

COOR COOR
R =0 ' 0
CH3H ch3h
(Phytyl)
01 9 0 0 7 4 ©(b##iS
A: Chl a (r3 = ch3) ; Cbl fe (Rs = CHO)
B : A 9 T II is 9 O O 7 4 b (BC);<9rV*7x*74?-y (BP) It Mg 2@©H-eB#3
Wavelength,nm
677,2 8.8Width
669.4 9.8
683.6 9.9649.2 11.7
702.5640.9 12.6
Error X8.I9
—,—1------1—|—1--------1 j—* J f ------ —1—160 155 150 145 140
Wavenumber, cm'1 XIO"3
02 Scenedesmus <h 0 □ P ~f 7 X b fr 6 *§ 6 *1 tz 7 > T 'f 0 a D 7 A &&ft & £ 0
h;i/'5
-114-
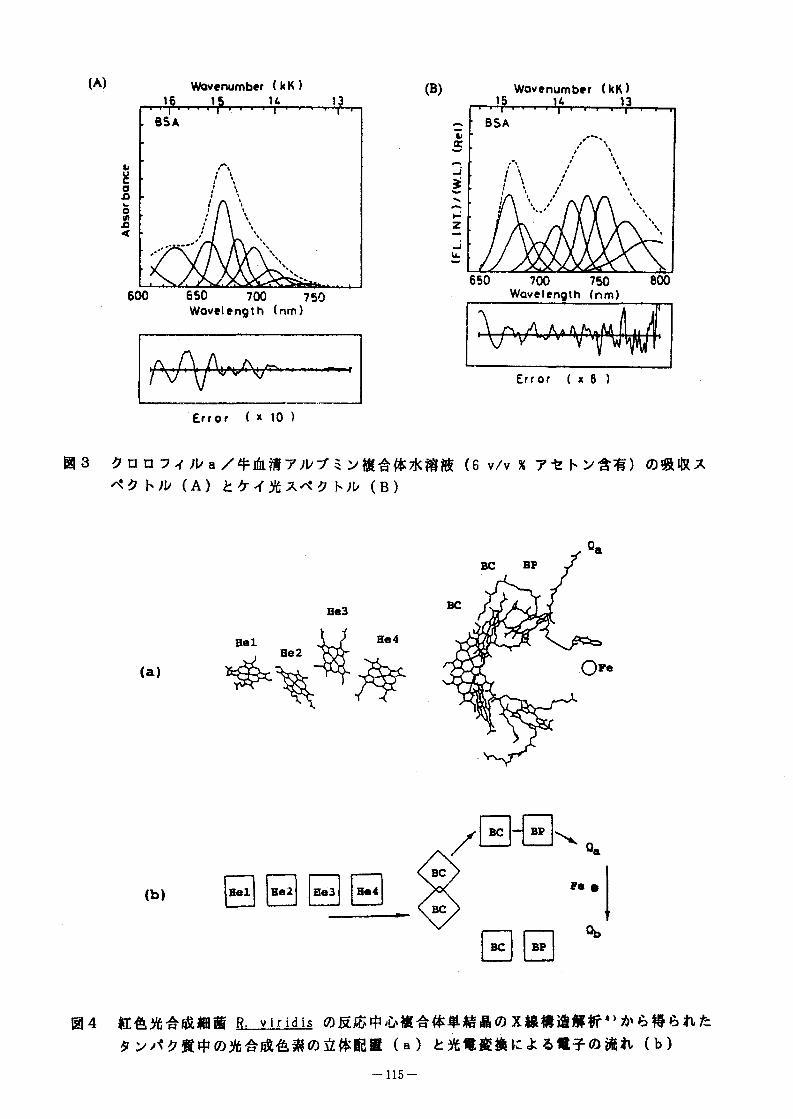
(A) Wavenumber (kK)
. BSA
650 700Wavelength (nm)
( x 10 )Error
(B) Wavenumber (kK)
700 750Wavelength (nm)
Error ( x 6 )
03 9 a n 7 -HI/ a /4=A#7;i/7'S >*K^**MK (6 v/v X 7-fe h©»(SIX *0 HI/ (A) kWjfcX/ti/ HI/ (B)
14 R. v i r id is JIO XttWiflWff *'*> 6 ft fc9 >/VK*mjt^A&S©S»e* (a) t##e#K,t&*?(&*ft (b)
—115 —

I
0 5 Rfatyfoi) n n 7 -f(a) P700 Katz6" ®t f 71/ (R-=H)(b) P680 6 A-683 ffl*r7l/7> (* = Mg)
A E
VBChi
SEMICONDUCTOR ELECTROLYTE
06 ma?oo
Ch!: if a a yjju&¥, vb: ##?#, cb: E81F, Ep: 7x7i/$1//<;!/, Red: t//<7P, Ox: BUbAi®*?
-116-

Transparent Electrode(CELEC®)
Chi a
Glass Plate
Chi a
@7 Chi a/pvA t^t A (Sl%)
*Et4N*
undoping--------►<-------doping
xEt4NCl04
08 PMeT 0l*fbWRK-y (K-f)
09 i/'n \i ay (1-5) PMeT/a-g) -te;u®W8
-117-

INI8<a
i0.2 0.4
0.6 0.8' Bias voltage on Ati / V
@10 Au/PMeT/ (1)/A1 ±)]/ (A) £ Au/PMeT/ (5)/Al ± )V (B) CD- SEtlfemi: BIST, «fS: ^MMT
♦ ••# e
Wavelength / nm Wavelength / nm
@11 Au/PMeT/(1)/A1 (a) k Au/PMeT/(5)/A 1 (b) b/1/PMeT #*&: (l) k (5) ®«iRx^» K/v
O: A1 e: Au fflfr 6 jfcSM
—118 —

(a)
A1
------- ---------- _ ^
(b)\
\
Au PMeT 5
H 12 Au/PMoT/ (1)/A1 (a) t Au/PMeT/(5)/Al (b) 0) x* ;i/-f V 7 A (iSES^)
—119 —

LETTERS TO NATURE
10“7 S m2 J"' for H2P and ZnP respectively at room temperature. The presence of the central zinc atom therefore seems to increase the conductive properties of the solid slightly.
In Fig. 2, (Acr/D)0 is plotted as a function of temperature. For H2P, a single, sharp decrease occurs at the transition from the solid to the isotropic liquid. No conductivity could be detected in the isotropic phase. For ZnP, the conductivity decreases abruptly, but does not disappear completely, at the solid to liquid-crystal transition temperature. The conductivity in the mesophase is a factor of ~3 less than in the solid with (Acr/D)n = 0.5x 10-7 S m2 J-1. This increases slightly with further increase in temperature in the mesophase and then suddenly drops to zero at the clearing point of ZnP.
We conclude that the long-range columnar order of porphyrin moieties in the solid and liquid-crystalline phases is essential for the conductive properties observed. In the isotropic liquid phase, any molecular organization that might occur must be of such a limited temporal or geometrical extent that it cannot support charge conduction. The results provide definitive evidence, therefore, for the general supposition that electrons and/or holes can move rapidly along the axis of column-stacked Tr-systems5,6. The decrease in conductivity on going from the crystalline solid to the mesophase is attributed to a lower charge mobility. This is probably due to an increase in positional disorder of the porphyrin moieties7 on melting of the hydrocarbon mantle.
We have found a similar decrease in the radiation-induced conductivity at the solid to mesophase transition of octa-alkoxy- substituled phthalocyanincs. An isotropic liquid phase of these compounds is not, however, attainable within the temperature range of our equipment. They could not therefore be used to carry out the conclusive experiment of completely melting the material to sec if long-range order was essential for conduction. The experiments on octa-n-alkoxy substituted phthalocyanincs have, however, shown that the lifetime of the conductivity transient increases exponentially with the length of the alkyl tails*. This is taken to be strong evidence that the mobile charge carriers responsible are restricted to rapid diffusional motion along the phthalocyanine axis of a stack, with eventual charge recombination requiring stack-to-stack electron tunnelling
. through the hydrocarbon mantle.The absolute value of (Acr/D)0 is related to the mobilities,
ft, of the charge carrcrs, [^ = W-) + M+)), and the average energy required to produce one charge-carrier pair, Ep (in cV), by3-< . ..
(A cr/D)0>=X/i/£p (1)
For organic compounds, the total initial yield of electron-hole pairs corresponds to ja value of Ep of ~25 eV for high-energy radiation9"11. Only a fraction of the initial electrons and holes will escape rapid (subnanosecond) geminate recombination* 9 and diffuse to separate columns, thus being observed in the present experiments. Using a value of Ep = 25 cV together with the experimentally determined value of (Acr/ D)a will therefore yield a lower limit to the charge-carrier mobilities. For the solid phase of ZnP at room temperature, assuming el-ectrons and holes to have almost equal mobilities'2, this gives jjl>
2.6 x 10-A m2 V-1 s-' and for the mesophase, it gives fi> 0.6 x 10~6 m2 V-' s-1.
This order of magnitude of the mobility indicates small polaron motion'3 corresponding to the hopping of a more or less localized charge between neighbouring sites with an average jump time between sites of r-r The mobility measured in the present, randomly orientated columnar materials is related to7) by
r} = edj/6k0T^ ' (2)
where d-t is the distance moved along the columnar axis per jump (4.9 A for the present systems). The minimum values of the carrier mobilities given above correspond therefore toNATURE • VOL 353 - 24 OCTOBER 1991
maximum jump times of 0.6 and 2 ps for solid ZnP at room temperature and the mesophase at 100 °C, respectively.
Our results confirm that the transport of charge can occur very rapidly within an organized self-assembly of porphyrin moieties even in a relatively flexible, liquid crystalline phase. Such peripherally hydrocarbon substituted compounds may therefore be considered to provide model systems which crudely mimic the channelling of energy and charge found in the highly varied antenna chlorophyll arrays of pholosynthctio membranes. • ' ■ . ‘ • □
Received 9 July, occcpicd 13 September 1991.
1. Gregs. 0. A. Fox. M. A. & Bard. A. J. J Am. chcm. See. Ill, 3024-3029 (1989).2. Weber. P. Coition. D. & Skoulios. A. lie CrysL 9. 369-382 11991). ' ''3. War man. J. M. & do Haas. M. P. in Pulse Radiolysis. Ctv 6. (ed. Tabala. YJ 101-133 (CRC, Does
Raton. 1991).4. War man. J. M. & do Haas. M. P. Radial Ptrys. Chem 34, 581-586 (1989V5. Elcy.O. D. Motet Crysl lie Crysl 171, 1-21 (1989). .6. Simon. J. & Andrt. J.J. Molecular Scrruconckx:Iors (Springer. Berlin. 198517. Pietro. W. J. Marks. T. 1 & Rainer. M. A. i Am. chem. Soc 107, 5387-5391 (1985).8. War man. J. M. Oc Haas.M. P-van Ocr Pol. 1 F.irDtcnltvW. Chem. Ptrys led 164, 581-586 (1989).9. War man J. M. In The Study of Fast Processes and Transient Species by Electron Poise Rndiolysis
(eds Baxcndale. J. K & Bust FJ 433-533 (Rcidcl. Dordrecht. 19821.10. Schmidl W. F. & Alien. A. 0.2 Pnys. Chem 72. 3730-3736 (1968). " ' ■■' ''11. Shmsaka. K. «. Freeman. 6. R Can. X chem 52. 3495-3506 (1974). . ; ' .12. Cos. C. A. & Knight P. C. 1 Phys. C 7. 146-156 (1974).13. Moll N. F. Conduction in Non-Crystalline Materials Ch.6 (Clarendon. Oxford. 1987).
ACKNOWLEDGEMENTS. We thank M_ Norlhotl and S. J. Picked (AKZ0 Research laboratories. Arnhem) ' lor X-ray Attraction analysis ol the ZnP compound and lor helpful discussions. The synthetic work was spodsored by ihe Texas Advanced Technology Program. •
A low-cost, high-efficiency solar cell based on dye-sensitized colloidal TiOz filmsBrian O’Regan* & Michael Gratzelt
Institute of Physical Chemistry. Swiss Federal Institute of Technology, CH-1015 Lausanne, Switzerland ■
The large-scale use of photovoltaic devices for electricity generation is prohibitively expensive" at present: generation from existing commercial devices costs about ten times more than conventional methods'. Here we describe a photovoltaic cell, created from low- to medium-purity materials through low-cost processes, which exhibits a commercially realistic energy-conversion efficiency. The device is based on a 10-p.m-thick, optically transparent film of titanium dioxide particles a few nanometres in size, coated with a monolayer of a charge-transfer dye to sensitize the film for light harvesting. Because of the high surface area of the semiconductor film and the ideal spectral characteristics of the dye, the device harvests a high proportion of the incident solar energy flux (46%) and shows exceptionally high efficiencies for (he conversion of incident photons to electrical current (more than 80%). The overall ligh(-to-clcctric energy conversion yield is 7.1-7.9% in simulated solar light and 12% in diffuse daylight. The large current densities (greater than 12 mA cm"2) and exceptional stability (sustaining at least five million turnovers without decomposition), as well as the low cost, make practical applications feasible.
Solar energy conversion by photoelectrochcmical cells has been intensively investigated2*". Dye-sensitized cells differ from the conventional semiconductor devices in that they separate the function of light absorption from charge carrier transport. In the case of n-type materials, such as Ti02, current is generated when a photon absorbed by a dye molecule gives rise to electron injection into the conduction band of the semiconductor, Fig! 1. To complete the circuit, the dye must be regenerated by
* Present address: Department ol Chemistry. University ol Washington. Seattle. Washington 98195.' USA. .t To whom correspondence shotid be addressed.
737
-120-

LETTERS TO NATURE
electron transfer from a redox species in solution which is then reduced at the counter electrode. The monochromatic current yield
T,(A) = LHE(A) x </>injX 7)c (1)
where LHE (light harvesting efficiency) is the fraction of the incident photons that arc absorbed by the dye, is the quantum yield for charge injection and 77, is the efficiency of collecting the injected charge at the back contact, expresses the ratio of measured electric current to the incident photon flux at a given wavelength. The photovoltage A V in Fig. 1, generated by the cell, corresponds to the difference between the Fermi level in the semiconductor under illumination and the Nernst potential of the redox couple in the electrolyte.
Although attempts to use dye-sensitized photoelectrochcmical cells in energy conversion have been made before, the efficiency of such devices has been extremely low and practical applications have seemed remote. One problem is that of poor light harvesting. On a smooth surface, a monomolccular layer of sensitizer absorbs less than 1 % of incident monochromatic light. Attempts to harvest more light by using multilayers of dye have in general been unsuccessful. The remaining option is to increase the roughness of the semiconductor surface so that a larger number of dye molecules can be adsorbed directly to the surface and can simultaneously be in direct contact with the redox electrolyte. Matsumura el al,12 and Alonso el al.xi have used sintered ZnO electrodes to increase the efficiency of sensitization by rose bcngal and related dyes. Willig, Parkinson and colleagues14 have reported high quantum yields for the dye sensitization of SnS2. But the conversion yields from solar light to electricity remained well below 1% for these systems. In addition, the instability of the dyes employed presented a severe practical drawback. By using semiconductor films consisting of nanometer-sized Ti02 particles, together with newly developed charge-transfer dyes, we have improved the efficiency and stability of the solar cell.
High-surfacc-arca Ti02 films were deposited on a conducting glass sheet from colloidal solutions. A transmission electron micrograph of the colloid is shown in Fig. 2. Electronic contact between the particles was produced by brief sintering at 450 °C. The size of the particles and pores making up the film is controlled by the size of the particles in the colloidal solution. The internal surface area of the film is determined by the size of the particles and the thickness of the film. These parameters were optimized to obtain efficient light harvesting while maintaining a pore size large enough to allow the redox electrolyte to diffuse
semiconductor dye electrolyte conducting glasscounterelectrode
FIG. 1 Schematic representation ot the principle ot the dye-sensitized photovoltaic cell to indicate the electron energy level in the different phases. The cell voltage observed under illumination corresponds to the difference. AK between the ouasi-Fermi level of Ti02 under illumination and the electrochemical potential of the electrolyte. The latter is equal to the Nernst potential of the redox couple (R/FT) used to mediate charge transfer between the electrodes. S, sensitizer; S*. electronically excited sensitizer;S+. oxidized sensitizer.
738
easily. Films of 10 pan thickness consisting of particles with an average size of 15 nm gave linear photocurrent response up to full sunlight and were used throughout. A cubic close packing of 15-nm-sizcd spheres to a 10-p.m-lhick layer is expected to produce a 2,000-fold increase in surface area.
Absorption spectra obtained for such nanostructurcd Ti02 films arc shown in Fig. T films tin*■trunspnrent.jajid colourless displaying the fundamental absorption edge of anatase (band -gap 3.2 eV) in the ultraviolet region. Deposition of a monolayer of the trimcric ruthenium complex15-1'’, RuL2(p.- (CN)Ru(CN)L2)2, 1, where Lis 2,2'bipyridine-4,4'-dicarboxylic acid and L' is 2,2'-bipyridine, results in deep brownish-red coloration jafjhc film. The absorption onsctls shTTted'fb~750TmT, tRe ffghtTiarvesting efficiency reading almost 100% in the whole visible region below 550 nm.Integration 01 the spectral overlap ^ctween-a snlur^emission of AM 1.5 and this absorption band shows that 46% of the incident solar energy flux is harvested by the dye coated film (AM = 1/sin a where a is the angle of incidence of the solar rays at the Earth’s surface).
The optical density of the film at 478 nm corrected for the absorption by the conducting glass support was 2.45. Dividing by the extinction coefficient"' of 1 (e47* = 1.88 x 107 cm2 mol-1) yields the dye surface concentration, F = 1.3 x 10-7 mol cm-2. As each dye molecule occupies an area16 of 1 nm2, the inner
■surface of the film is 780 cm2 for each 1 cm2 of geometric surface. Thus, the roughness factor is 780, which is smaller than the predicted value ofJLQOO.. The difference is attributed to necking between Ti02 particles. In addition, the large size of 1 prevents its access to very small pores, reducing the apparent surface area.
The photocurrent action spectrum obtained with the dye- coated Ti02 film is also shown in Fig. 3. It closely matches the absorption spectrum, indicating that the current is due to electron injection from 1 into the conduction band of TiOz. The photocurrcnt yield measured at 520 nm was found to depend
FIG. 2 Transmission electron micrograph of Titi2 particles used in thin film production. The scale bar represents 10 nm. Particles were prepared by hydrolysis of titanium tetraisopropoxide18-19 followed by autoclaving for 12 h at 200 °C. To form films the sol was concentrated by evaporation of water in vacuum at 25 *C until a viscous liquid was obtained. Carbowax M-20,000 (40% by weight of Ti02) was added and the viscous dispersion (Ti02 content 20% by weight) was spread on the conducting glass support (Asahi glass, fluorine-doped Sn02 overlayer, transmission 85% in the visible, sheet resistance 80/square) to give a membrane of 10 p.m thickness. This was heated under air for 30 min at 450 °C. High-resolution scanning electron microscopy revealed Ti02 films to be composed of a three-dimensional network of interconnected nanoscale particles19. Transmission electron micrographs of Ti02 particles were taken before heat treatment; the annealing of 450 °C did not induce significant changes in particle size. Before dye coating, a few monolayers of Ti02 were electrodeposited onto the colloidal Ti02 film from a Ti(lll) solution. Detailed description of this procedure will be published elsewhere (L. Haven. B.O'R., A. Kay and M.G.. manuscript in preparation).
NATURE • VOL 353 • 24 OCTOBER 1991
-121-

LETTERS TO NATURE
on the counter ion of the iodide/triiodide redox electrolyte, increasing from 68% for.tctrapropvlammonium to 84% for Li*. After correction for the light absorption by the conducting glass.the yields are 80% and 97%, respectively. This shows that thenanostructured TiO-> films used in conjunction with suitable charge transfer dyes can achieve quantitative conversion ofvisible light photons into electric current"
Figure 4 shows the current-voltage characteristics obtained • with the thin layer cell under illumination by simulated AM 1.5 solar light. The conversion efficiencies for oni tenth and full sunlight are 7.9% and 7.12%, and the fill factors (maximum output power of cell+-[short circuit current x open circuit voltage]) are 0.76 and 0.685, respectively. Similar yields were obtained under direct sunlight (measurements performed in June early in the afternoon on the roof of the institute). Under diffuse daylight the efficiency increased to 12%, indicating that under such conditions the cell performance is better than that of conventional silicon devices. This is because the spectral distribution of diffuse daylight overlaps more favourably with the absorption spectrum of the dye-coated Ti02 film than direct sunlight. The.fill factor of the cell remains above 0.7 even at very low light intensity (<5 W m~2). Conventional photovoltaic cells have a much smaller fill factor (<0.5) under these conditions. This indicates that loss mechanisms such as recombination, normally encountered in semiconductor photoconversion, have been minimized. This result might appear surprising in view of the disordered structure of our film giving rise to defects. But the role of the semiconductor in a dye-sensitized device is merely to conduct the injected majority charge carriers. There arc no minority carriers involved in the photoconversion process. Therefore, surface and bulk recombination losses due to lattice defects, encountered in conventional photovoltaic cells, are not observed in such a device.
The long-term stability of cell performance was tested by illuminating the thin TiQ, film loaded with 1 with visible (A > 400 nm) light foi 2 months] The change in the photocurrcnt was less than 10% over this period, during which a charge of 62,000 C cm-2 was passed through the device, corresponding to a turnover numbcr_of 5 x IQ6 for the sensitizer. This implies that if any dye degradation had occurred its quantum yield (<£,„.) is less than 2 x 10~*. As d>dcc = A'dce/2k, the rate constant, /cdcc s~‘, for excited-state decomposition (due to processes such as ligand loss) must be at least 10”* times smaller than Zt, the sum of rate constants for all channels of dye deactivation. Because charge injection is the predominant channel, this sum is practi-
• Cell potential (V)
FIG. 4 Photocurrent-voltage characteristics of a cell, based on a colloidal Ti02 film sensitized by 1; the film, supported on a conducting glass sheet, was used in a sandwich-type configuration. The size of the dye coated Ti02 photoanode was 0.5 cm2. The counterelectrode, consisting of conducting glass coated with a few monolayers of platinum, was placed directly on top of the working electrode. A thin layer of redox electrolyte is attracted into the intra-electrode space through capillary forces. The cell was exposed to simulated sunlight with AM1.5 spectral distribution, a, light intensity 83 W m-2, electrolyte: 0.5 M tetrapropylammonium iodide +0.04 M Iodine in a mixture of ethylene carbonate (80% by volume) with acetonitrile. Fill factor was 0.76: surface area 0.5 cm2 (before multiplication by roughness factor). Conversion efficiency was 7.9%. b, light intensity 750 W m“3. electrolyte: 0.5 M tetrapropylammonium Iodide. 0.02 M HI xO.04 M l2 in the same solvent. Fill factor was 0.684: conversion was 7.12%.
ui o.4o
Wavelength (nm)
FIG. 3 Absorption and photocurrent action spectra of Ti02 films supported on conducting glass. A. absorption efficiency of the bare Ti02 film corrected for conducting glass background: B, absorption efficiency of the same film coated with a monolayer of 1; full circles, monochromatic current yield at short circuit as a function of excitation wavelength. Yield is corrected for 15% loss of incident photons through light absorption and scattering by the conducting glass support.
NATURE • VOL 353 • 24 OCTOBER 1991
cally equal to the rale constant for charge injection, which exceeds 1012 s-1 in the case of 1. Therefore, the upper limit for A',lcc is 2 x 104 s-1, which agrees with the known photophysics of this class ol transition metal complexes17. The very fast electron injection observed with dyes such as 1, combined with high chemical stability, renders these compounds very attractive for practical development. □
Received 19 July: accepted 20 August 1991.
1. Bucher. K. & Frlcke. 1 fhys. Zeit 21. 237-244 (1980).2. Honda. K. & Fujishlma. A. Native 238, 37-39 (1972)..3 Tufts. 0. J. et at. Nature 326. 681-683 (1987).4. Gerischcr. K Elcctrochim Acta 35,1677 (1990).5. Uchl. S. Modes. G.. Tennc. R. A Manasscn. J. Nature 326. 863-804 (191)7).6. Heller. A. Ace chew. ties. 14. 154-162 (1981).7. Nozk. A. J. Phil. Trans, ft Soe Lent A295, 453-470 (19801.8. Trtoulsch. K & OcnocL 1 C J eiectroanaL Chem 81. 97 (1977).9. Wrighton. M. S. Ace 'chem. Res. 12. 303-310 (1979).
10. Bard. A. J. Science 207. 139 (1980).11. Memmlng. R. Phil. Tech. Rpv. 38.160 (1979).12. Matsumura. M_ Nomura. Y. A Tsubomura. H. Butt, chcni Soe Japan 50. 2533 (1977).13. Alonso. N- Oetey. V. M_ Char tier. P. A Em. V. Rev. Ptiys. Appi 16, 5 (1981).14. Willig. F. Ekhbcrgcr. R_ Sundaresan.N. S. A Parkinson. 0. A. 1 Am chem Soe 112. 2702-2707
(1990).15. Amodclll. R_ Argazzl. R_ OlgnozzL C. A. A Scandola. F. i Am chem Soe 112. 7099-7103 (1990).16. Nazceruddin. M. K. Uska. P. Moser. J. Vlachopoulos. N. A GrSlzcl. M. Hetv. clvm Acta 73,
1788-1803 (1990).17. Axis. A. Oatrani, V. Oariglclti. F_ Campagna. S. Oclzcr. 0. Coord. Chem Rev. 84. 85 (1900).
739
-122-

Proceeding of 4th International Congress of the World Apheresis Association, 1992. 6. 3-5, Sapporo, in press
Nakaji S, et al.
DEVELOPMENT OF SPECIFIC IMMUNOADSORBENT CONTAINING IMMOBILIZED SYNTHETIC PEPTIDE OF ACETYLCHOLINE RECEPTOR FOR
TREATMENT OF MYASTHENIA GRAVIS
Shuhei Nakaji, Kiichiro Oka, Masao Tanihara,Koichi.Takakura, Masaharu Takamori*Kuraray Co., Ltd., Kurashiki, Japan
Kanazawa University School of Medicine, Kanazawa, Japan
KEY WORDS: Myasthenia gravis, Immunoadsorption, Acetylcholine receptor, Synthetic peptide, Blocking antibody,
IntroductionMyasthenia gravis (MG) is an autoimmune disorder in
which neuromuscular transmission is impaired by antibodies against the nicotinic acetylcholine receptor (AChR) in skeletal muscle. In order to remove anti-AChR antibodies, plasma exchange and double filtration plasmapheresis have currently been used. However, these methods have the disadvantages that loss of useful plasma components is inevitable, and supply of the replacement fluid such as fresh frozen plasma or albumin preparations is required. Therefore, recently the clinical significance of the immunoadsorption has increasingly been recognized.
Anti-AChR antibodies are classified into two subclasses; binding antibody and blocking antibody. These antibodies cause the acceleration of AChR degradation, complement- mediated lysis of post-synaptic membrane and blockade of ACh-binding with AChR. The blocking antibody is known to prevent the ACh-binding with AChR, thereby inducing MG.Takamori et al.1>,2> reported that the (X183-200 segment of the Torpedo californica AChR is the ACh-binding site recognized by a blocking antibody.
Based on their study, we have developed the new immunoadsorbent column for MG treatment (Medisorba MG) by using the synthetic peptide (Torpedo CX183-200) as an affinity ligand to remove specifically the blocking antibodies.
Materials and MethodsAdsorbent : The synthetic peptide Torpedo californica a
183-200 (Fig. 1) synthesized by a solid-phase procedure was used as a ligand. The peptide was covalently immobilized on porous cellulose beads (dia. 250pm). The ' amount of the immobilized peptide was 35mg/50ml beads.
-123-

Nakaji S, et al.
In vitro study : (Blocking antibody) The patient plasma 200pl was treated with 50mg of peptide-bound adsorbent for 3hr at 37 * C. A glycine-bound adsorbent was used as a control. The blocking antibody titer was measured by a- bungarotoxin binding inhibition assay. (Plasma protein) 5ml of healthy human plasma was treated with 250mg of the pep tide-bound adsorbent for 3hr at 37 * C, and then the plasma protein levels were measured.
Immunoadsorbent device : Immunoadsorbent Medisorba MG consists of a small column, packed with 50ml of peptide-bound adsorbent and sterilized by autoclaving. The safety of the device has been confirmed by various toxicity tests including acute and subacute toxicity, cytotoxicity, mutagenicity and immunogenicity tests.
Clinical evaluation : 77 treatments of plasmaimmunoadsorption were performed for 19 patients with MG. The immunoadsorption treatment was carried out 3 times/week.31
183H-Lys-Lys-Gly-Trp-Lys-His-Trp-Val-Tyr-Tyr-Thr-Cys-Cys
200-Pro-Asp-Thr-Pro-Tyr-Leu-Asp-Lys-Lys-Gly-OH
Figure 1 Amino acid sequence of the synthetic peptide corresponding to Torpedo at 83-200
Results and DiscussionIn Fig. 2 and Table 1 are shown the results of in vitro
study on adsorption of blocking antibody and on adsorption of plasma proteins, respectively. The clinical results of changes in blocking antibody obtained with the 36 immunoadsorption treatments are represented in Fig. 3. From both in vitro study and clinical evaluation, it was demonstrated that the peptide-bound adsorbent removed specifically blocking antibody without significantly reducing plasma proteins such as albumin and IgG. The removal rates obtained with clinical studies are as follows: blocking antibody 40.2%, binding antibody 12.4%, total proteins 5.7%, albumin 2.4%, IgG 10.2%. Fig. 4 shows the changes in blocking antibody (anti-peptide antibody) and IgG level over the period of 35 days. Blocking antibody titer was reduced remarkably after the treatments contrary to the slight change in IgG level, and remained at low level for the observation period. It is suggested that this immunoadsorption treatment provides promise to the long-term effect without occurrence of rebound. Clinical treatments improved effectively the myasthenic state. The improvement was found in 78% of the cases, and no adverse effects were observed in any case of treatments.
-124-

Nakaji S, et al.
75
&B 50
E
.g 25
10
0
g Control
Adsorbent
12 3 4Patient serum
Cut off value
Figure 2 Changes in blocking antibody titer by adsorption treatment(in vitro) Adsorbent : synthetic peptide Torpedo a 183-200-bound to
porous cellulose beadsControl : glycine bound to porous cellulose beads
Table 1 Effect of adsorption treatment on plasma protein levels (in vitro)
Adsorbent Control
Alb (g/dl) 3.8 3.7IgG (mg/dl) 1060 1120IgA (mg/dl) 207 196IgM (mg/dl) 113 105C3 (mg/dl) 60 60C4 (mg/dl) 19 22TP (g/dl) 5.6 5.6A/G 2.08 2.01
Adsorbent : Synthetic peptide Torpedo a183-200 bound to porous cellulose beads
Control : Without adsorbent
ConclusionsWe have developed the new immunoadsorbent containing
immobilized synthetic peptide of AChR (Torpedo (X183-200)
for MG treatment. Medisorba MG showed the specific removal of blocking antibody, one of the antibodies involved in the pathogenesis of MG, without reducing significantly plasma protein levels. The clinical usefulness of the immunoadsorbent has been demonstrated for the MG treatment.
References1. Takamori M, Okumura S, Nagata M, et al.. Myasthenogenic
significance of synthetic a-subunit peptide 183-200 of Torpedo californica and human acetylcholine receptor. J
-125-
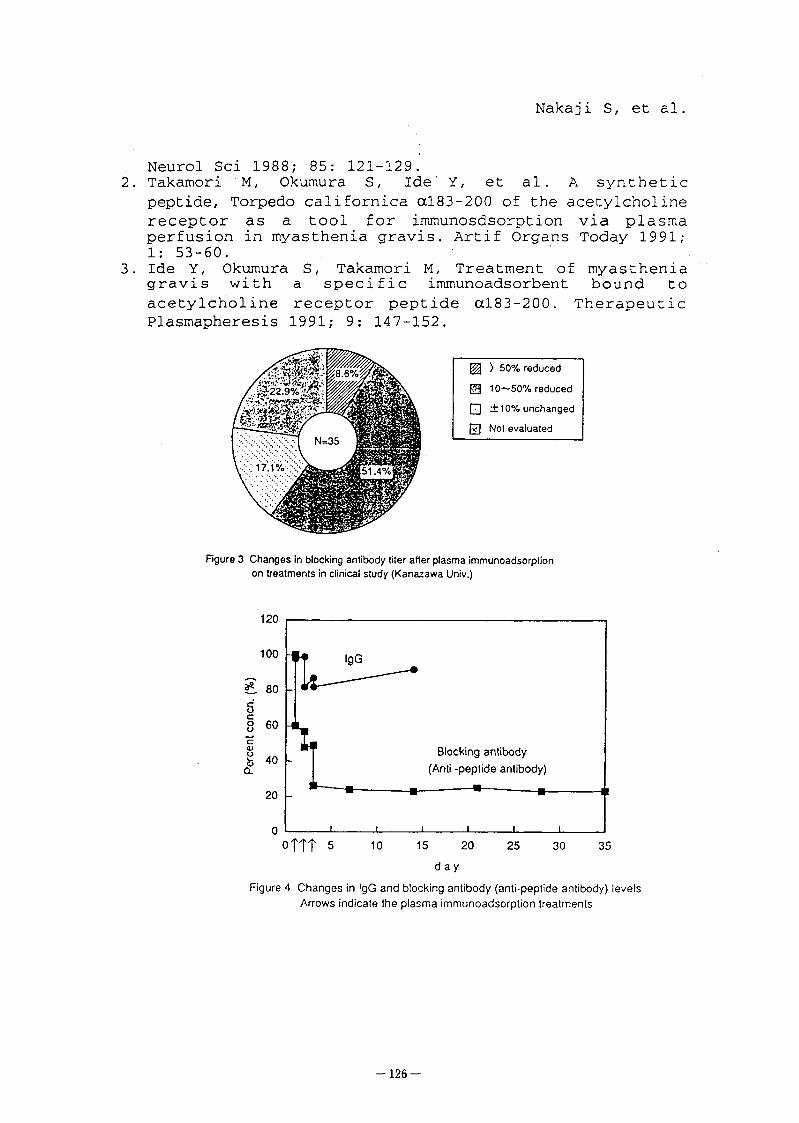
Nakaji S, et al.
Neurol Sci 1988; 85: 121-129.2. Takamori M, Okumura S, Ide' Y, et al . A synthetic
peptide, Torpedo californica CX183-200 of the acetylcholine receptor as a tool for immunosdsorption via plasma perfusion in myasthenia gravis. Artif Organs Today 1991; 1: 53-60.
3. Ide Y, Okumura S, Takamori M, Treatment of myasthenia gravis with a specific immunoadsorbent bound toacetylcholine receptor peptide al83-200. Therapeutic Plasmapheresis 1991; 9: 147-152.
Figure 3 Changes in blocking antibody titer after plasma immunoadsorption on treatments in clinical study (Kanazawa Univ.)
Blocking antibody (Anti -peptide antibody)
Oftt 5 10 15 20 25 30 35
day
Figure 4 Changes in IgG and blocking antibody (anti-peptide antibody) levels Arrows indicate the plasma immunoadsorption treatments
-126-

ZiM^zr? KCDIWmm

-IZl
'aw
5zw^m§sz5a3s*tBes?}$$ •??awt)?oiv<i>m?i!it«i(<" >£?!aa#i+±3® 'c ^ °wn9vtav9w%w2sw-a-a
MPE;Hii3iw§SSto±gs?K— ' § an ill-tl#;iB2 S n •t/v8M«aBIIsl° ^ <~ u xn
Me$(tiD 2 nan Jewish assess'isamvys;!. mm ¥.av • #gg 'nm#•■n0:iisi«'?v: 9%a##u^#*n:) rsifii
<>,q^av-yjflwsgsrvqiij^ziy^Y '¥? •■miiMWEi
?g?o?t?^-4^'i^^r]TicisS/5ao e °¥n :)#92*=1jLn#/Sao ET1H14- '>yu%n G
mic&m® 'z
° ■? V <? ? # n W ^ ? 7 -*r 3 »* W<? <? M D
jliW.^/vlY ' G EfaiS:53IMMB^fl„?7^W.4 Yavwirtfl- °¥n:)?a9A tghYavs^wnmanngm im '^9%?##^m#w)V7ann$M5«Y
_an* °9%a# 'YS»v^/v«SISi® '3>i#®WBes»3#l$© ' PS W <? ¥av^¥B28M W
%%(D '.K?W "?V9?#^Y-axawjgifiW^o^ninW:mfilW,4 YYVS3I1:
r?on?! ■ t
e BSiiif
-4=^^BS^fri3l¥(jf)
2^ 3# g# GO w

3 . K
$a»«l£T 5 JW.fr YXhtfr,VI ^) e
3 - 1 . Ky)K?iJ*OT(8f)$6*E9i%B^(cES5itrv^ . $a®£1rtr£^r^-KtOK^JA^TRSrm
cs-r, rsysim oo-isos-f-yfc lt£*' <3 *>(:** Ltns<oWJ.3 OMT(± 1 3 7 9 8T$-o%.
3-2. J: 6^-m
3 0rsyi?JJlTO5?M^rf-Kiil 3 7 98$, 9 ,
S6«(i»to$6, SirSBk LT*@§iiSi3^. ##AtM=f &<%u#Lv^ ±«s
ttrs-s-r$voiisg*s<)5r*&r$hfr, two sari,. ;;tiiiif«c, sou r c e T$&A#8(:j;Ck cut.
<( li_k@A'4,^. ( kingdom) , M ( phylum) , 8 (class) , @ (order) , f4
(family) . # (genus) . 8 (species) COjfr$$.& (taxon) C<£ 9 £ ft. T V i ,
fi)fiT'f±S<7)-hf3®,'S:k L"t_hfr ( super kingdom) SriUt, ;ft i < 3 OiC^itT V> h ,C cft^SSCfic-0 TL/C . %a#*r Hco sourceCO8 (species) (O'S&^.'Z (CSx'4". ^7f K0 source k LT^SS£ft.TW6A#lf)8<ftmT$ 9 . ^7"f ^7*f-K0f5£»M*k
ixco$iV>imz-o\cx(owmm%t>tiz.acfc^rf-FS:±ft8C:J: -?-C#3I-f t#^M(=WR-taxont'»mf<@T
mm±&4lil/<A'##&6taxon&#mL%;5#EM-C$&. (lt)gaH9f%?5:Eii><3. K/K9g$:& 2 0%/;>-?' ( rt 3 oiispecies-ecO
teo) tcjj-ltrSaLTviSco-e, f«7;k-ey/5'SfflLt. 7 tLT«S'J
Ztit:. $a»$r*txr^-KeiS:&tX3 0T5 y®mT<0^.7‘4- H(7)E^S3lcS-t„
d«r-yt±, A%#^m%M#ktr-oTV\&t%8vomm^M^gr^LTVvak#z.
TSV^fc/,5. h^%Akf6#%##A*y)#l*<Em*K:$v\. ,iy)f*k
human> E. coI i x S. cerevisiae Mammal iav Prokaryotax Fungi C-OS&^tC
Efc*$jirv^5„ 3 0T5 yBEiyTyxr^KCov^Tti. ff?U«*{EmWlz^^coit K(ft8^kRET$5*% 7okx.
-128-

3 - 3 . H iO U X h
ae^o&B® ^ k j: o t r s y sots y$JiiT«o^^^7f H<0 l)X h £^4 KLfc. CX) V X H4#a(0±##%HK4 kA?#^Lt. C
iL^O^r^b'OTS /ISK?IJ. ZStK. keyTOrd^fcOtlTii^JfcKasfl-LfcXn -y t‘-
T1 X?lzimztix^$. S4toEcOlt^liXES^^XiT'. fA
V'„
4 . afSTW^KEWtctoTffi^W^ft^^tV^V^T^-K
8 ii 5 xt*^ K T-. ga®M7 5/lW«t>fci9>I.Sg7;y
@£irtf, rn&ELt j:
■rtvsxxrf-Hco^E#-^. ««r fflaicro-t-E.v^< zfrcoamzmeizv;-?'3 - 3^i; [§]«(;. satixn V f-Td xy (cEliSATV'5,.
5 . joi^DiC
Eigtt^rf- wtmffigcofctbcomgwm i spst lx, mam 3 o T s x #mT(OA%
KXJ^EESSrffvx P'Jrco&mi LTfcO-SiaS-f^fiSL*.
LTK^^SUc^ 0 . #@k LT4 k 4
^otLioft#, ±8ti, ElgK|iJS9f%7)-toC:5:jitfSV'X'Z)5.
-129

mi.
Km- 3 0 13 7 9
3 0-40 5 6 04 0-50 5 0 75 0-60 8 1 16 0-70 8 5 47 0-80 8 2 68 0-90 8 6 390-100 10 5 9
100-150 6 0 5 3150-200 3 7 6 7200-250 3 3 10250-300 2 8 9 8300-350 2 6 7 7350-400 2 4 6 2400-450 19 5 2450-500 19 5 1500-550 14 5 9550-600 9 4 1600-650 7 3 2650-700 5 6 3700-750 4 9 2750-800 3 8 1800-850 3 0 1850-900 3 2 9900-950 2 3 2950-1000 2 17
1000-1500 9 4 51500-2000 2 5 62000-2500 1 4 52500-3000 4 63 0 0 0 - 7 3
A #1 3 9 0 4 1
130-

2. ^7°-f* K <7) source
Super Kingdom (3) ± FFKingdom (6) n
Phylum (55) P!Virus 7 / : 451 NematodaProkaryota IM Nematomorpha
Prokaryomycota m##E^ AcanthocephalaArchebacteria "SlfflS 29 KinorhychaMycoplasma 7-f37" 7X' "7 7 MolluscaBacteria SfflE 242 SipunculoideaMyxobacteria %%#E 3 PriapuloideaSpirocheta Xt* D'W 2 AnnelidaActinomycota SxSS E 43 Onychophora
Prokaryophyta LinguatulidaCyanophyta 19 TardigradaGlaucophyta 0 Arthropoda
Eukaryota CrustaceaFungi E^ Arachnoidea
Myxomycota ^BE 2 InsectaAcrasinomycota rnrn^m 2 EctoproctaLabyrintulomycota 7t' yyf%7 0 PhoronideaOomycota mm 0 BrachiopodaHyphochtridiomycota ftiT-' 7*' tit' 0 Chaetognathachytridiomycota 7-+' tit' 0 EchinodermataZygomycota 7 HemichordataAscomycota 63 PogonophoraBasidiomycota E 12 Prochordata
Lichenes VertebrataLichenobionta 0 Agnatha
Plantae ChondrichthyeRhodphyta 3 OsteichthyesCryptphyta 7V7" 0 AmphibiaDinphyta 1 ReptiliaChromophyta 3 AvesVaucheriophyta +' -7V7#% 0 MammaliaEugrenophyta 2Chrolophyta 170
AnimaliaProtozoa 42Porifera 0Cnidaria 13Ctenophora 0Plathelminthes 5Nemertinea mB#% 1Endoproca 0Gastrotricha 0Rotifera ^B#% 0
mBrnm :4
:0WjfyjMfjfy) :0%
:32MnMb&J : 4
: 0saBmm :io
:0
:0
:05P :Ho
:0#&#% :0
:0 :0 :15
###% : 0 :0 :2:594
Ran
e#m
K ^OsourceCO^^^OM (species) <7)lfc$:7jvf~0
-131-

*3.
KE$t 3 ors smut
o^rf- Kj&^7 'i )VX Virus 2 9 4 0 3 4
Prokaryota 3 7 0 8 9 4Eukaryota
#-# Fungi 10 6 5 3 3#3^# Lichenes 0###- Plantae 10 8 9 3 8##-# Animal ia
MA## Protozoa 2 6 0####) Nematoda 64 8
#(#;#% Mollusca 1 0 4 6 9Arthropoda 7 84 1 8 0
#### Vertebrata
F3 ditt Agnatha 2 1 4Chondrichthyes 7 0 3 5Osteichythyes 24 3 8 9Amphibia 2 8 8 10 9Kept ilia 2 4 6 5 6
S3S Aves 5 7 3 4 8Mammalia 7 8 7 4 5 1 1
2 3 0 7 1t h 3 3 3 3E . coli 13 9 1S . cerevisiae 4 6 0
3(taxon) .
*1 i kii-^u&n.
-132-

$kA. SE^F^IE^J£ J: o T T 5 /ME?'] O&tf)3 0 7$/il^iT^^rf K (7n7e-fa; SEQ30NS)
#::1 LIST OF PEPTIDES WITH LESS THAN 30 AMINO ACIDS.#::2 F0RMAT=LIT.N0.:TAXON TAG:PEPTIDE765024A:Agnatha:fibrinopeptide A1205260A:Agnatha:luliberin1414306A:Agriatha:somatostatin 14 variant1406259A:Agnatha:vasotocin1516329A:Amphibia:alpha melanotropin710343A:Amphibia:alytesin0904250A:Amphibia:angiotensin I0510184A:Amphibia:angiotensin,crinia1718263A:Amphibia:antimicrobial peptide PGQ1413199A:Amphibia:atrial natriuretic factor 241414243A:Amphibia:atrial natriuretic-like peptide1509286A:Amphibia:basic tryptophyllin1608110A:Amphibia:bioact hydrophobic peptide A11608110B:Amphibia:bioact hydrophobic peptide B9710343B:Amphibia:bombesin720133A:Amphibia:bombesin730327A:Amphibia:bombesin1610281A:Amphibia:bombesin related peptide PG-L1714191A:Amphibia:bombesin-like neuropeptide700810A:Amphibia:bombinin1103176A:Amphibia:bradykinin650766A:Amphibia:bradykinin690622A:Amphibia:bradykinin701177A:Amphibia:bradykinin1103176B:Amphibia:bradykinin 5-9680356A:Amphibia:bradykinin,1-Val,6-Thr0510365A:Amphibia:bradykinin,3-Hyp0504194A:Amphibia:bradykinin,6-Thr1103176C:Amphibia:bradykinin,des-9-Arg730327B:Amphibia:bradykininyl-Gly-Lys-Phe-His1703232A:Amphibia:C type natriuretic peptide680326A:Amphibia:caerulein0310191A:Amphibia:caerulein,2-Asn,5-Leu1508203A:Amphibia:deltorphin1510394A:Amphibia:deltorphin I1510394B:Amphibia:deltorphin II1515244A:Amphibia:dermenkephalin0704273A:Amphibia:dermorphin1810298A:Amphibia:dermorphin1810298B:Amphibia:dermorphin0704274A:Amphibia:dermorphin,6-Hyp0311228A:Amphibia:granuliberin R0812283A:Amphibia:histone H31511130A:Amphibia:hydrin 11511130B:Amphibia:hydrin 21606259A:Amphibia:hydrin 20801196A:Amphibia:hylambatin0712211A:Amphibia:hypothalamic peptide0308208A:Amphibia:kassinin0801196B:Amphibia:kassinin,2-Glu 5-Pro730327C:Amphibia:kinin 0680668A:Amphibia:kinin peptide III1104233A:Amphibia:litorin751393A:Amphibia:litorin0605192A:Amphibia:litorin,2-Glu(OEt)0311238A:Amphibia:litorin,2-Glu(OMe)640051A:Amphibia:mesotocin 1602196A:Amphibia:neuromedin U 1811353A:Amphibia: neurotensin 610522A:Amphibia:oxytocin 610522C:Amphibia:oxytocin 690622B:Amphibia:phyllocaerulein 660391A:Amphibia:phyllokinin 0910155A:Amphibia:phyllolitorin
133-

701175A:Amphibia:phyllomedusin 640455A:Amphibia:physalaemin 0605192B:Amphibia:physalaemin like peptide 1803428A:Amphibiarranakinin 680668B:Amphibiazranakinin N 0504194B:Amphibia:ranakinin R 1711287A:Amphibia:ranatachykinin A 1711287B:Amphibia:ranatachykinin B 1711287C:Amphibia:ranatachykinin C 1711287D.-Amphibia:ranatachykinin D 701177B:Amphibia:ranatensin 0504195A:Amphibia:ranatensin R 1112220A:Amphibia:somatostatin1610281B:Amphibia:tachykinin related peptide PG-KI 1610281C:Amphibia:tachykinin related peptide PG-KII 1610281D:Amphibia:tachykinin related peptide PG-KIII 1610281E:Amphibia:tachykinin related peptide PG-SPI 1610281F:Amphibia:tachykinin related peptide PG-SPII 742295A:Amphibia:thyroliberin 690954A:Amphibia:toxin peptide 4a alpha 690954B:Amphibia:toxin peptide 4a beta1204212A:Amphibia 1208379L:Amphibia 1208379A:Amphibia 1208379B:Amphibia 1208379D:Amphibia 1208379C:Amphibia 1208379E:Amphibia 1208379G:Amphibia 1208379H:Amphibia 1208379F:Amphibia 1208379J:Amphibia 1208379K:Amphibia 1005217A:Amphibia 1104246A:Amphibia 1104246B:Amphibia 1104246C:Amphibia 1005217B:Amphibia 1104246D:Amphibia
tryptophyllin 13 tryptophyllin 13 tryptophyllin 4 tryptophyllin 4 tryptophyllin 4 tryptophyllin 4 tryptophyllin 4 tryptophyllin 5 tryptophyllin 5 tryptophyllin 5 B tryptophyllin 5 P tryptophyllin 7 tryptophyllin TPH4 tryptophyllin TPH4 tryptophyllin TPH4 tryptophyllin TPH4 tryptophyllin TPH5 tryptophyllin TPH5 tryptophyllin TPH7
A1A2MlM3
Q
1104246E:Amphibia:751003A:Amphibia:uperolein1803252A:Amphibia:vasotocin600015A:Amphibia:vasotocin610522E:Amphibia:vasotocin610522F:Amphibia:vasotocin640487A:Amphibia:vasotocin671037B:Amphibia:vasotocin1803252B:Amphibia:vasotocin-related peptide731315A:Amphibia:xenopsin1302299A:AnimaliaO:antho RFamide1612407A:AnimaliaO:antho RNamide1805361A:AnimaliaO:antho-RIamide I1414251A:AnimaliaO:antho-RWamide0910246A:AnimaliaO:casomorphin beta1604340A:AnimaliaO:cionin1210338A:AnimaliaO:egg assocd peptide A1210338B:AnimaliaO:egg assocd peptide B0712245A:AnimaliaO:morphogenetic peptide1412363A:AnimaliaO:mosact1404258A:AnimaliaO:neuropeptide1817216A:AnimaliaO:neuropeptide1817216B:AnimaliaO:neuropeptide1717144A:AnimaliaO:neuropeptide Antho-KAamide1302184A:AnimaliaO:neuropeptide,pennatulid1001132A:AnimaliaO:peptide Al,sperm activating1507171A:AnimaliaO:Pol-RFamide
-134-

1808186A:Animalia0 1101334A:AnimaliaO 1711480A:Animalia0 1711480B:AnimaliaO 0804270A:AnimaliaO 1708180A:AnimaliaO 1708180B:AnimaliaO 1708180C:AnimaliaO 1708180D:AnimaliaO 1708180E:AnimaliaO 1708180F:AnimaliaO 1708180G:AnimaliaO 1708180H:AnimaliaO 1708180J:AnimaliaO 1708180K:AnimaliaO 1708180L:AnimaliaO 1713215A:AnimaliaO 1713215K:AnimaliaO 1713215B:AnimaliaO 1713215C:AnimaliaO 1713215D:AnimaliaO 1713215E:AnimaliaO 1713215F:AnimaliaO 1713215G:AnimaliaO 1713215H:AnimaliaO 1713215J:AnimaliaO 0705150B:AnimaliaO 0705150A:AnimaliaO 1502213A:AnimaliaO 1502214A:AnimaliaO 1508189A:AnimaliaO 1508189B:AnimaliaO 1616199B:AnimaliaO 1616199T:AnimaliaO 1616199H:AnimaliaO 1616199G:AnimaliaO 1616199P:AnimaliaO 1616199V:AnimaliaO 1616199R:AnimaliaO 1616199F:AnimaliaO 1616199L:AnimaliaO 1616199M:AnimaliaO 1616199A:AnimaliaO 1616199C:AnimaliaO 1616199Q:AnimaliaO 1616199U:AnimaliaO 1616199W:AnimaliaO 1616199E:AnimaliaO 1616199D:AnimaliaO 1616199N:AnimaliaO 1616199K:AnimaliaO 1616199J:AnimaliaO 1802280A:AnimaliaO 0402232A:AnimaliaO
Pol-RFamide resactSALMFamide 1 SALMFamide 2 speractsperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide III sperm activating peptide TG-1 sperm activating peptide TG-10 sperm activating peptide TG-2 sperm activating peptide TG-3 sperm activating peptide TG-4 sperm activating peptide TG-5 sperm activating peptide TG-6 sperm activating peptide TG-7 sperm activating peptide TG-8 sperm activating peptide TG-9 sperm-activating 9-peptide sperm-activating peptide sperm-activating peptide sperm-activating peptide sperm-activating peptide b sperm-activating peptide d sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I sperm-activating peptide I, sperm-activating peptide I, sperm-activating peptide SV-1
(2,5-Thr,3-Leu,4-Pro,7,10-Ser) (2-Phe(Br))(2-Phe(Br),3-Asn)(2-Tyr,3,5-Ser,8-Ala,10-Asp) (2-Tyr,3-Asn,5-Gly,9-Ile,10-Asp) (2-Tyr,3-Asn,7,10-Asp,8-Arg,9-Ile (3,5-Gly)(3,5-Ser)(3-Ala,5-Gly)(3-Asn)(3-Asn,4-Ile,5-Gly)(3-Asn,5-Ser)(3-Glu,4-Met,5-Gly,7-Thr)(3-Gly,5-Ser,9-Ile)(3-Ser,5-Gly)(5-Gly)(5-Thr)Des-6,7-Gly (3-Ala,5-Asp)Des-6,7-Gly (3-Ala,5-Asp,10-Asn)
toxin III1201191A:Arthropoda:adipokinetic hormone 1204166A:Arthropoda:adipokinetic hormone 1515421A:Arthropoda:adipokinetic hormone 1613147A:Arthropoda:adipokinetic hormone 1613152A:Arthropoda:adipokinetic hormone 1708296A:Arthropoda:adipokinetic hormone 1810246A:Arthropoda:adipokinetic hormone 1813270A:Arthropoda:adipokinetic hormone 763125A:Arthropoda:adipokinetic hormone 1109158A:Arthropoda:adipokinetic hormone II 1109158B:Arthropoda:adipokinetic hormone II 1203209A:Arthropoda:adipokinetic hormone II
-135-

1203209B:Arthropoda:adipokinetic hormone II 1708195A:Arthropoda:adipokinetic hormone III 1402182A:Arthropoda:adipokinetic peptide AKH-G 1413346A:Arthropoda:AKH family neuropeptide 1413258A:Arthropoda:AKH like brain factor BI 1515171A:Arthropoda:allatostatin 1718377A:Arthropoda:allatostatin 1511156A:Arthropoda:allatostatin 1 1511156B:Arthropoda:allatostatin 2 1511156C.-Arthropoda:allatostatin 3 1511156D:Arthropoda:allatostatin 4 1707324A:Arthropoda:allatostatin ASB2 1505417A:Arthropoda:allatotropin 670258A:Arthropoda:apamine 1511072A:Arthropoda:apidaecin lb1010431A:Arthropoda:Balbiani ring 2 gene beta repeat C1010431E:Arthropoda:Balbiani ring 2 gene beta repeat SR1103246A:Arthropoda:bombolitin I1103246B:Arthropoda:bombolitin II1103246C:Arthropoda:bombolitin III1103246D:Arthropoda:bombolitin IV1103246E:Arthropoda:bombolitin V1412231A:Arthropoda:bombyxin II A1412231B:Arthropoda:bombyxin II B1503112A: Arthropoda .'bombyxin IV A1503112B:Arthropoda:bombyxin IV B1309327A:Arthropoda:bradykinin,6-Thr754921A:Arthropoda:bradykinin,Ala-Arg1309327B:Arthropoda:bradykinyl-Lys-Ala,6-Thr1807377A:Arthropoda:calliFMRFamide 11807377K:Arthropoda:calliFMRFamide 101807377L:Arthropoda:calliFMRFamide 111807377M:Arthropoda:calliFMRFamide 121807377N:Arthropoda:calliFMRFamide 131807377B:Arthropoda:calliFMRFamide 21807377C:Arthropoda:calliFMRFamide 31807377D:Arthropoda:calliFMRFamide 41807377E:Arthropoda:calliFMRFamide 51807377F:Arthropoda:calliFMRFamide 61807377G:Arthropoda:calliFMRFamide 71807377H:Arthropoda:calliFMRFamide 81807377J:Arthropoda:calliFMRFamide 91807377P:Arthropoda:calliMIRFamide1302325A:Arthropoda:cardioactive peptide CCAP1601326C:Arthropoda:chemotactic peptide721419A:Arthropoda:color change hormone1511082A:Arthropoda:corazonin1805363A:Arthropoda:corazonin,7-His1009229A:Arthropoda:crabrolin1401236C:Arthropoda:Cs gene1811325A:Arthropoda:diapuase hormone1803186A:Arthropoda:diuretic peptide Mas-DP21508172A:Arthropoda:FMRFamide like neuropeptide1413303A:Arthropoda:FMRFamide like peptide 31413303B:Arthropoda:FMRFamide like peptide 41616410A:Arthropoda:FMRFamide-related peptide1805364A:Arthropoda:FMRFamide-related peptide1402211A:Arthropoda:ganglion peptide1802220A:Arthropoda:gastrin/cholecystokinin-like peptide 1709276A:Arthropoda:growth blocking peptide 0712190A:Arthropoda:histamine releasing peptide 0712190B:Arthropoda:histamine releasing peptide 1304184A:Arthropoda:hypertrehalosemic factor II 1211206A:Arthropoda:hypertrehalosemic hormone 1607342A:Arthropoda:hypertrehalosemic neuropeptide 1612381A:Arthropoda:hypertrehalosemic neuropeptide
—136 —

1613146A:Arthropoda:hypertrehalosemic neuropeptide1810301A:Arthropoda:hypertrehalosemic peptide1810301B:Arthropoda:hypertrehalosemic peptide1610160C:Arthropoda:hypertrehalosemic peptide Bld-HrTH1610160D:Arthropoda:hypertrehalosemic peptide Cam-HrTH-II1610160A:Arthropoda:hypertrehalosemic peptide Pea-CAH-I1610160B:Arthropoda:hypertrehalosemic peptide Pea-CAH-II1515421B:Arthropoda:hypotrehalosemic hormone681037A:Arthropoda:kinin1301220A:Arthropoda:leucokinin I1301220B:Arthropoda:leucokinin II1402209A:Arthropoda:leucokinin V1402209B:Arthropoda:leucokinin VI1402210A:Arthropoda:leucokinin VII1402210B:Arthropoda:leucokinin VIII1210374A:Arthropoda:leucosulfakinin1211204A:Arthropoda:leucosulfakinin II1304197A:Arthropoda:leukopyrokinin762531A:Arthropoda:light adapting hormone1510214A:Arthropoda:locusta peptide A1510214B:Arthropoda:locusta peptide B1804432A:Arthropoda:locustakinin1801363A:Arthropoda:locustamyoinhibiting peptide1612380A:Arthropoda:locustamyotropin1713291A:Arthropoda:locustamyotropin II1706255A:Arthropoda:locustapyrokinin1605188A:Arthropoda:locustatachykinin I1605188B:Arthropoda:locustatachykinin II1704380A:Arthropoda:locustatachykinin III1704380B:Arthropoda:locustatachykinin IV1709386A:Arthropoda:Lom-AG-myotropin1208383A:Arthropoda:mast cell degranulating peptide1312417A:Arthropoda:mast cell degranulating peptide690923A:Arthropoda:mast cell degranulating peptide0510099A:Arthropoda:mastoparan0607189A:Arthropoda:mastoparan1601326B:Arthropoda:mastoparan1708155A:Arthropoda:mastoparan1009229B:Arthropoda:mastoparan C0710225A:Arthropoda:mas toparan M0510182A:Arthropoda:mastoparan X670043A:Arthropoda:melittin730710A:Arthropoda:melittin751015A:Arthropoda:melittin0405213A:Arthropoda:melittin F720299A:Arthropoda:melittin II1709388A:Arthropoda:myotropin1607220A:Arthropoda:neuropeptide Aeal1607220B:Arthropoda:neuropeptide Aeall1414190A:Arthropoda:neuropeptide hez-HrTH1606261A:Arthropoda:neuropeptide led CCI1606261B:Arthropoda:neuropeptide led CCII1711289A:Arthropoda:neuropeptide mem-CC1810245A:Arthropoda:neuropeptide mem-CC1011173A:Arthropoda:neuropeptide MI1011173B:Arthropoda:neuropeptide Mil1301199A:Arthropoda:neuropeptide,hypertrehalosaeraic1603305C:Arthropoda:omega agatoxin IIA1107279A:Arthropoda:ORF 1 C term1103308B:Arthropoda:ORF HB2750746A:Arthropoda:paragonial peptide PS1713343A:Arthropoda:paralytic peptide I1713343B:Arthropoda:paralytic peptide I1713343C:Arthropoda:paralytic peptide I1713343D:Arthropoda:paralytic peptide II1713343F:Arthropoda:paralytic peptide II
-137-

1713343G:Arthropoda:paralytic peptide II 1713343E:Arthropoda:paralytic peptide III 0401151A:Arthropoda:peptide Cp.silk fibroin 1501209A:Arthropoda:peptide HR1 1501209B:Arthropoda:peptide HR2 1010330A:Arthropoda:periplanetin CC1 1010330B:Arthropoda:periplanetin CC2 1601433A:Arthropoda:perisulfakinin 1804200A:Arthropoda:pheromonotropic neuropeptide 1304257A:Arthropoda:pigment dispersing factor 1109281A:Arthropoda:pigment dispersing hormone 1211218A:Arthropoda:pigment dispersing hormone 754921B:Arthropoda:polisteskinin R 1601383A:Arthropoda:polyphemusin I 1601383B:Arthropoda:polyphemusin II 1610280A:Arthropoda:proctolin 754183A:Arthropoda:proctolin1812292A:Arthropoda:red pigment-concentrating hormone 1413346B:Arthropoda:RPCH family neuropeptide 0405213B:Arthropoda:secapin 1002241A:Arthropoda:secapin .1501299A:Arthropoda:tachyplesin1601383C:Arthropoda:tachyplesin II1615222A:Arthropoda:tachyplesin III0605179A:Arthropoda:tertiapin0902216A:Arthropoda:toxic 8-peptide1005287B:Arthropoda:toxin Bot V N term1601342B:Arthropoda:toxin like peptide II1104330A:Arthropoda:tropomyosin C term1314162A:Arthropoda:vasopressin like diuretic hormone764671A:Arthropoda:vespakinin M771747A:Arthropoda:vespakinin X760232A:Arthropoda:vespulakinin760232B:Arthropoda:vespulakinin1601326A:Arthropoda:waspkinin1612286A:Aves:angiotensin I730699A:Aves:angiotensin I0410203A:Aves:antigen,major histocompatibility 1413198A:Aves:atrial natriuretic factor 0910250A:Aves:brain peptide 1203416A:Aves:bursin1704143A:Aves:C type natriuretic peptide 1003829A:Aves:c-myc gene0504213A:Aves:collagen alphal(I) N term,pro 1007220A:Aves:collagen alphal(II) CB14,pro 0407229A:Aves:fibrinopeptide A 1110284A:Aves:fibrinopeptide A 1110284B:Aves:fibrinopeptide B 1715216A:Aves:galanin0701228A:Aves:gastric proventricular peptide1403328A:Aves:glucagon1714255A:Aves:glucagon721287A:Aves:glucagon721913A:Aves:glucagon753726A:Aves:glucagon1604250B:Aves:histone HI1506448B:Aves:histone H2AF assocd ORF730530A:Aves:insulin C peptide0810231A:Aves:luliberin0906295A:Aves:luliberin1007246A:Aves:luliberin II0904299A:Aves:luliberin,8-Gln701182A:Aves:mesotocin1403440B:Aves:neurokinin A1718372A:Ayes:neuromedin U0608263A:Aves:neurotensin
138-

1310315A:Aves:neurotensin0903220A:Aves:neurotensin related 6-peptide1314219A:Aves:neurotensin related peptide1313235A:Aves:ornitho kinin1313235C:Avesrornitho kininogen600454A:Aves:oxytocin640486A:Aves:oxytocin750800A:Aves rprotamine0311203A:Aves:pyruvate carboxylase biotin peptide0701221A:Aves:secretin0507258A:Aves:somatostatin1403440A:Aves:substance P,3-Arg753696A:Aves:vasoactive intestinal peptide600456A:Aves:vasopressin,Arg600456B:Aves:vasotocin701182B:Aves rvasotocin1412334A:Aves:xenopsin related peptide720476A:Chondrichthyes:aspartocin720633A:Chondrichthyes:aspartocin741686B:Chondrichthyes:corticotropin like interm lobe peptide1206345A:Chondrichthyes:endorphin beta1401182A:Chondrichthyes:gastrin releasing peptide1112169A:Chondrichthyes:glucagon1305260A:Chondrichthyes:glucagon1516142B:Chondrichthyes:glucagon650080A:Chondrichthyes:glumitocin650768A:Chondrichthyes:glumitocin671036A:Chondrichthyes:glumitocin1301248A:Chondrichthyes:insulin A1412225A:Chondrichthyes:insulin A1211262B:Chondrichthyes:insulin C peptide1709219A:Chondrichthyes:luliberin1815266A:Chondrichthyes:luliberin1817182A:Chondrichthyes:luliberin I700801A:Chondrichthyes:melanotropin alpha741687A: Chondrichthyes .-melanotropin alpha741687B:Chondrichthyes:melanotropin beta741687C:Chondrichthyes:melanotropin beta690930A:Chondrichthyes:oxytocin650096A:Chondrichthyes:oxytocin,3-Ser 8-Ile1301239A:Chondrichthyes:relaxin A1301239B:Chondrichthyes:relaxin B1206256A:Chondrichthyes:scyliorhinin I1206256B:Chondrichthyes:scyliorhinin II1412223A:Chondrichthyes:scyliorhinin II 3-181112170A:Chondrichthyes:somatostatin1703277A:Chondrichthyes:somatostatin 14,5-Ser1806387A:Chondrichthyes:urotensin II720476B:Chondrichthyes:valitocin720633B:Chondrichthyes:valitocin1311236A:Chondrichthyes:vasoactive intestinal peptide610242B:Chondrichthyes:vasotocin764683A:Fungi:alpha factor1612305A:Fungi:arg-2 assocd ORF1104318B:Fungi:ATPase 9,F01309198B:Fungi:carbamoylphosphate synthetase ORF1407245B:Fungi:gene nucl ORF 11407245C:Fungi:gene nucl ORF 21407245D:Fungi:gene nucl ORF 3290004A:Fungi:glutathione721924A:Fungi:inhibitor,galactose oxidase0401178A:Fungi:mating factor1302211A:Fungi-.mating factor a21302211B:Fungi:mating factor a31208294A:Fungi:mating factor alpha762484A:Fungi:mating hormone alphalB
-139-

1206200A:Fungi:mating pheromone alpha ski1206200B:Fungi:mating pheromone alpha sk31112155A:Fungirmetallothionein,Cu1201202A:Fungi:metaliothionein,Cu1408283B:Fungi:0RF 11103229A:Fungi:ori region ORF 31109166B:Fungi:oxidase I,cytochrome c1109166C:Fungi:oxidase III,cytochrome c1003184A:Fungi:pheromone alpha sk2,mating0812287F:Fungi:protein URFd1109166D:Fungi:protein varl0504158A:Fungi:rhodotorucine A0502206A:Fungi:sillucin0707174A:Fungi:substance alphaK771273A:Fungi:substance IA,alpha762484B:Fungi:substance IB,alpha0601162A:Fungi:tremerogen A100705139A:Fungi:tremerogen al31005183A:Fungi:tremerogen A9291I1817362A:IS0TYPE=A:Mammalia:relaxin1805261A:IS0TYPE=II:Arthropoda:myotropin1008163A:Mammalia:acrosin light chain1104244A:Mammalia:adrenal medullary peptide precursor0911276A:Mammalia:adrenorphin0410472A:Mammalia:albumin prepiece1710319A:Mammalia:albuminamide1608236A:Mammalia:alpha neo-endorphin732148A:Mammalia:ameletin1102244A:Mammalia:amidorphin C term0804298A:Mammalia:amylase alpha570121A:Mammalia:angiotensin1210206A:Mammalia:angiotensin converting enzyme inhibitor560088A:Mammalia:angiotensin I560148A:Mammalia:angiotensin I670366A:Mammalia:angiotensin I720249A:Mammalia:angiotensin I560149A:Mammalia:angiotensin II0411243A:Mammalia:anorexigenic peptide0803236A:Mammalia:antiarrhythmic peptide0901253A:Mammalia:antigen,spermatozoal0604199A:Mammalia:antigonadotropic 3-peptide1805147A:Mammalia:antioncogene protein pt270601225A:Mammalia:ATPase accelerating peptide1001137A:Mammalia:atria peptide1004222A:Mammalia:atrial natriuretic factor1010154C:Mammalia:atrial natriuretic factor III1010154D:Mammalia:atrial natriuretic factor IV1002183A:Mammalia:atrial natriuretic peptide alpha1007202A:Mammalia:atrial natriuretic/vasoactive peptide1001244A:Mammalia:atriopeptin I1005179A:Mammalia:atriopeptin I1005179B:Mammalia:atriopeptin I,des-1,2-Ser1005179C:Mammalia:atriopeptin I,des-1-Ser1005179D:Mammalia:atriopeptin I,des-21-Ser1001244B:Mammalia:atriopeptin II1005179E:Mammalia:atriopeptin II1005179F:Mammalia:atriopeptin III1409308A:Mammalia:bactenecin1701465A:Mammalia:beta prepro-tachykinin 117-126 0906234A:Mammalia:bombesin like peptide 0906234B:Mammalia:bombesin like peptide 0906234C:Mammalia:bombesin like peptide 1011314A:Mammalia:bombesin like peptide 600196A:Mammalia:bradykinin 620203A:Mammalia:bradykinin 1207241A:Mammalia:bradykinin,3-Ala Lys
140 —

1407164A:Mammalia:bradykinin,3-Hyp1414313A:Mammalia:bradykinin,3-Hyp650157A:Mammalia:bradykinin,Met-Lys1402188A:Mammalia:bradykinin,N-Lys 3-Hyp1404380A:Mammalia:brain natriuretic peptide1609081A:Mammalia:C type natriuretic peptide0311203B:Mammalia:carboxylase biotin peptide,pyruvate1101255A:Mammalia:casein 6-peptide,immunostimulating0511270A:Mammaliacasomorphin beta1815292A:Mammalia:cationic Cys-rich peptide1815292C:Mammalia:cationic Cys-rich peptide1406215A:Mammalia:CCK-PZ 81012338A:Mammalia:cerebellin753732A:Mammalia:chemotactic factor753732B:Mammalia:chemotactic factor1011260A:Mammalia:cholecystokinin1103329A:Mammalia:cholecystokinin 221108290A:Mammalia:cholecystokinin 220604474A:Mammalia:cholecystokinin 41304169A:Mammalia:cholecystokinin 50409339A:Mammalia:cholecystokinin 81108290B:Mammalia:cholecystokinin 81408277A:Mammalia:cholecystokinin 81812203A:Mammalia:cholinergic neurostimulating peptide1005178A:Mammalia:chromaffin granule peptide1005178B:Mammalia:chromaffin granule peptide1005178C:Mammalia:chromaffin granule peptide precursor1809219A:Mammalia:chromogranin A-derived peptide WE141104274A:Mammalia:collagen alphal III signal peptide1105272A:Mammalia:collagen II,pro N term1413205A:Mammalia complement C3f754907A:Mammalia contraceptive peptide1718167C:Mammaliacorticostatic peptide 30611192B:Mammalia:corticotropin like interm lobe peptide740695A:Mammalia:corticotropin like interm lobe peptide0707220A:Mammalia corticotropin releasing 7-peptide0406187A:Mammalia:CRF active 14-peptide0310165A:Mammalia:decarboxylase fragment,dopa1112214A:Mammalia:defensin HNP11112214B:Mammalia:defensin HNP21112214C:Mammalia:defensin HNP31603276B:Mammalia:defensin NP30411144A:Mammalia:delicious taste peptide0412267A:Mammalia:delta sleep inducing peptide1211230A:Mammalia:DFT stimulating peptide0712247A:Mammalia:dynorphin0601259A:Mammalia:dynorphin 130811243A:Mammalia:dynorphin 240610285A:Mammalia:dynorphin related 8-peptide750521A:Mammalia:encephalitogenic peptide M761141A:Mammalia:endorphin alpha0705147A:Mammalia:endorphin alpha,neo0705146A:Mammalia:endorphin beta,neo764744A:Mammalia:endorphin gamma1412332A:Mammalia:endothelin0702167A:Mammalia:enkephalin BAM22P1011313A:Mammalia:enkephalin derived peptide,prepro1002185A:Mammalia:enkephalin derived peptide,pro753272A:Mammalia:enkephalin,Leu761375A:Mammalia:enkephalin,Leu762848A:Mammalia:enkephalin,Leu753272B:Mammalia:enkephalin,Met761375B:Mammalia:enkephalin,Met762848B:Mammalia:enkephalin,Met0708293A:Mammalia:enkephalin,N-Ac Leu0610284A:Mammalia:enkephalinyl-Arg,Leu
141-

0601256A:Mammalia:enkephalinyl-Arg,Met 0601260A:Mammalia:enkephalinyl-Arg-Phe,Met 1109269A:Mammalia:epidermal growth factor receptor 1005225A:Mammalia:ferritin H0504173A:Mammalia:fibrinogen alpha N term fragment1006279A:Mammalia:fibrinogen alpha signal1006279B:Mammalia:fibrinogen beta signal1006279C:Mammalia:fibrinogen gamma signal0912289A:Mammalia:fibrinogen N term Bern II1001180A:Mammalia:fibrinopeptide A1001180B:Mammalia:fibrinopeptide A1001180C:Mammalia:fibrinopeptide A1105261A:Mammalia:fibrinopeptide A650770A:Mammalia:fibrinopeptide A650770B:Mammalia:fibrinopeptide A650770C:Mammalia:fibrinopeptide A650770D:Mammalia:fibrinopeptide A650771A:Mammalia:fibrinopeptide A650771B-.Mammalia:fibrinopeptide A650771C:Mammalia:fibrinopeptide A650771D:Mammalia:fibrinopeptide A650771E:Mammalia:fibrinopeptide A650771F:Mammalia:fibrinopeptide A650771G:Mammalia:fibrinopeptide A650771H:Mammalia:fibrinopeptide A650771J:Mammalia:fibrinopeptide A650771K:Mammalia:fibrinopeptide A650771L:Mammalia:fibrinopeptide A650771M:Mammalia:fibrinopeptide A650771N:Mammalia:fibrinopeptide A650771P:Mammalia:fibrinopeptide A650771Q:Mammalia:fibrinopeptide A650771R:Mammalia:fibrinopeptide A650771T:Mammalia:fibrinopeptide A660906A:Mammalia:fibrinopeptide A660906B:Mammalia:fibrinopeptide A660906C:Mammalia:fibrinopeptide A670960A:Mammalia:fibrinopeptide A670960B:Mammalia:fibrinopeptide A670960C:Mammalia:fibrinopeptide A670960D:Mammalia:fibrinopeptide A670960E:Mammalia:fibrinopeptide A671044B:Mammalia:fibrinopeptide A671044C:Mammalia:fibrinopeptide A690946A:Mammalia:fibrinopeptide A701211A:Mammalia:fibrinopeptide A701218A:Mammalia:fibrinopeptide A721948A:Mammalia:fibrinopeptide A721948C:Mammalia:fibrinopeptide A732189A:Mammalia:fibrinopeptide A732189B:Mammalia:fibrinopeptide A754929A:Mammalia:fibrinopeptide A1004217A:Mammalia:fibrinopeptide A Louisville1001180D:Mammalia:fibrinopeptide B1001180E:Mammalia:fibrinopeptide B1105261B:Mammalia:fibrinopeptide B1105261C:Mammalia:fibrinopeptide B630449A:Mammalia:fibrinopeptide B650770E:Mammalia:fibrinopeptide B650770F:Mammalia:fibrinopeptide B650770G:Mammalia:fibrinopeptide B650771AA:Mammalia:fibrinopeptide B650771AB:Mammalia:fibrinopeptide B650771AC:Mammalia:fibrinopeptide B650771AD:Mammalia:fibrinopeptide B650771AE:Mammalia:fibrinopeptide B
-142-

650771AF:Mammalia:fibrinopeptide B 650771U:Mammalia:fibrinopeptide B 650771V:Mammalia:fibrinopeptide B 650771W:Mammalia:fibrinopeptide B 650771X:Mammalia:fibrinopeptide B 650771Y:Mammalia:fibrinopeptide B 660906D:Mammalia:fibrinopeptide B 660906E:Mammalia:fibrinopeptide B 670960F:Mammalia:fibrinopeptide B 670960G:Mammalia:fibrinopeptide B 670960H:Mammalia:fibrinopeptide B 670960J:Mammalia:fibrinopeptide B 670960K:Mammalia:fibrinopeptide B 671044E:Mammalia:fibrinopeptide B 671044F:Mammalia:fibrinopeptide B 690946B:Mammalia:fibrinopeptide B 690946C:Mammalia:fibrinopeptide B 701211B:Mammalia:fibrinopeptide B 701218B:Mammalia:fibrinopeptide B 721948D:Mammalia:fibrinopeptide B 721948F:Mammalia:fibrinopeptide B 721948G:Mammalia:fibrinopeptide B 721948H:Mammalia:fibrinopeptide B 732189C:Mammalia:fibrinopeptide B0807231A:Mammalia:fibronectin Cys containing region1112273A:Mammalia:FMRFamide like 18-peptide1112273B:Mammalia:FMRFamide like 8-peptide1001157A:Mammalia:galanin1712179A:Mammalia:galanin1718373A:Mammalia:galanin1802436A:Mammalia:galanin1815257A:Mammalia:galanin701192A:Mammalia:gastric peptide1413347A:Mammalia:gastrin640126A:Mammalia:gastrin660134A:Mammalia:gastrin680320A:Mammalia:gastrin680320B:Mammalia:gastrin690052A:Mammalia:gastrin690262A:Mammalia:gastrin0802193A:Mammalia:gastrin little0503136A:Mammalia:gastrin mini1009184A: Mammalia .-gastrin N term,pro0510270A:Mammalia:gastrin releasing peptide1202335A:Mammalia:gastrin,littie1405288A:Mammalia:gelsolin1003207A:Mammalia:globin epsilonlll1003207B:Mammalia:globin epsilonlV1110283A:Mammalia:glucagon1207205A:Mammalia:glucagon1606260B:Mammalia:glucagon1616407A:Mammalia:glucagon570104A:Mammalia:glucagon710325A:Mammalia:glucagon711649A:Mammalia:glucagon720483A:Mammalia:glucagon720967A:Mammalia:glucagon1512256A:Mammalia:glucagon like peptide 10806160A:Mammalia:glycentin related peptide1304348A:Mammalia:gonadotropin releasing peptide730306A:Mammalia:growth modulating 3-peptide1805415A:Mammalia:growth/mitosis inhibitory peptide1805411A:Mammalia:guanylin0711246A:Mammalia:head' activator peptide
1408207C:Mammalia:histatin 51010306A:Mammalia:Ig C deltal fragment
—143 —

1101448A:Mammalia:Ig D/JH1101448B:Mammalia:Ig D/JH0407221A:Mammalia:Ig L-V extra peptide0407221B:Mammalia:Ig lambda C extra peptide770551A:Mammalia:Ig lambdal N Term extra1011444C:Mammalia:Ig VH E4.151011444D:Mammalia:Ig VH E4psi1807240A:Mammalia:indolicidin0412251A:Mammalia:inhibitor PI,alphal proteinase0806130A:Mammalia:inhibitor,angiotensin converting enzyme1312309A:Mammalia:insulin1312309B:Mammalia:insulin560164B:Mammalia:insulin1506296A:Mammalia:insulin A1506296B:Mammalia:insulin B0510475A:Mammalia:insulin B,24/25-Leu680495A:Mammalia:insulin C peptide711102A:Mammalia:insulin C peptide720389A:Mammalia:insulin C peptide720389C:Mammalia:insulin C peptide740063A:Mammalia:insulin C peptide1008138A:Mammalia:insulin releasing peptide1008138B:Mammalia:insulin releasing peptide610207A:Mammalia:kallidin II1301340A:Mammalia:kinase,B creatine1103228A:Mammalia:kinase,phosphoglycerate1107242A:Mammalia:kinase.phosphoglycerate1206205A:Mammalia:kinetensin0905156A:Mammalia:kinin T0505388A:Mammalia:kyotorphin0811241A:Mammalia:kyotorphin,neo0905239A:Mammalia:kyotorphin.neo710434A:Mammalia:luliberin720377A:Mammalia:luliberin1714374A:Mammalia:luliberin,9-Hyp0806461A:Mammalia:lymphocyte stimulating peptide1510156B:Mammalia:M assocd ORE1609099A:Mammalia:macrophage cheraotactic factor1516181A:Mammalia:melanin concentrating hormone710420A:Mammalia:melanostatin I711502A:Mammalia:melanostatin I720300A:Mammalia:melanostatin II0609249A:Mammalia:melanotropin 9-18,9-Tyr570116A:Mammalia:melanotropin alpha590325A:Mammal!a:melanotropin alpha600123A:Mammalia:melanotropin alpha610131A:Mammalia:melanotropin alpha630479A:Mammalia:melanotropin alpha560090A:Mammalia:melanotropin beta570098A:Mammalia:melanotropin beta590120A:Mammalia:melanotropin beta610131B:Mammalia:melanotropin beta610263A:Mammalia:melanotropin beta630479B:Mammalia:melanotropin beta0703205A: Mammalia-.melanotropin beta, des-1-Asp0810171A:Mammalia:melanotropin delta0807304A:Mammalia:melanotropin gammal0706201A:Mammalia:melanotropin gamma30807304B:Mammalia:melanotropin gamma30707206A:Mammalia:melanotropin like peptide,gammal0506561A: Mammalia .-melanotropin potentiating factor0702463A:Mammalia:melanotropin,des-Ac0512240A:Mammalia:melanotropin,N,0-(Ac)20909223A:Mammalia:mesotocin1504308A:Mammalia:Met enkephalin like peptide 0512482A: Mammalia-.Met enkephalin releasing peptide
-144

0912288A:Mammalia:metorphamide 0904334A:Mammalia:motilin 1105286A:Mammalia:motilin 730210A:Mammalia:motilin 740052A:Mammalia:motilin1702149A:Mammalia:myelin peptide amide pMPA141411251A:Mammalia:nephritogenoside0908308A: Mammalia-.neurokinin alpha0908308B:Mammalia:neurokinin beta0908190A:Mammalia:neuromedin B1003162A:Mammalia:neuromedin C1011992A:Mammalia:neuromedin L1008139A:Mammalia:neuromedin N1411284A:Mammalia:neuromedin U1414195A:Mammalia:neuromedin U1612383A:Mammalia:neuromedin U1709387A:Mammalia:neuromedin U 251709387B:Mammalia:neuromedin U 81109148A:Mammalia:neuromedin U251711482A:Mammalia:neuromedin U251406325A:Mammalia:neuropeptide gamma1413348A:Mammalia:neuropeptide K 1-241612231A:Mammalia:neuropeptide Y 12-361208283A:Mammalia:neurotenisin,7-Ser0605222A:Mammalia:neurotensin1716411A:Mammalia:neurotensin750525A:Mammalia:neurotensin1306331A:Mammalia:neurotensin related peptide1809273A:Mammalia:neutral thyroliberin-like peptide1809273B:Mammalia:neutral thyroliberin-like peptide1811319A:Mammalia:neutrophil granule peptide HP10811159A:Mammalia:opioid 4-peptide0704330A:Mammalia:opioid peptide,adrenal0706309A:Mammalia:opioid peptide,adrenal1512206A:Mammalia:0RF 1520009A:Mammalia:oxytocin530009A:Mammalia:oxytocin580093A:Mammalia:oxytocin580181A:Mammalia:oxytocin590227A:Mammalia:oxytocin600454B:Mammalia:oxytocin640049A:Mammalia:oxytocin690929A:Mammalia:oxytocin730938A:Mammalia:oxytocin1004251A:Mammalia:pancreatic 20-peptide1301201A:Mammalia:pancreatic 20-peptide1003237A:Mammalia:pancreatic 20-peptide C term0801264A:Mammalia:pancreatic islet peptide720699A:Mammalia:pancreatic polypeptide0411198A:Mammalia:pepsinogen I/II major glycopeptide1003823A:Mammalia:peptide 3b,Pro rich0405230A:Mammalia:peptide A12d,glyco0405230B:Mammalia:peptide A17c,glyco0405230C:Mammalia:peptide B12,glyco0610176A:Mammalia:peptide BAM12P1707342B:Mammalia:peptide His-Ile1109251A:Mammalia:peptide 0A24b0712238A:Mammalia:peptide PHI1010243A:Mammalia:peptide PHI1011215A:Mammalia:peptide PHI1610159A:Mammalia:peptide PHI 1-271109144A:Mammalia:peptide SCP31109144B:Mammalia:peptide SCP41109144C:Mammalia:peptide SCP51109144D:Mammalia:peptide SCP60405258A:Mammalia:peptide,carbohydrate carrying
—145 —

0405258B:Mammalia:peptide,carbohydrate carrying711641A:Mammalia:peptide,conditioned avoidance response711078A:Mammalia:peptide,glyco763904A:Mammalia:peptide,inhibitory1310217A:Mammalia:peptide,mitosis inhibiting0712205A:Mammalia:peptide,salivary low MW0712205B:Mammalia:peptide,salivary low MW0909223B:Mammalia:phenypressin1208305A:Mammalia:physalaemin like peptide1312257A:Mammalia:physalaemin like peptide0509287A:Mammalia:placental lactogen precursor1617235A:Mammalia:pneumadin720638A:Mammaliaposterior pituitary peptide1603334A:Mammalia:prepro-tachykinin C flanking peptide1603334B:Mammalia:prepro-tachykinin C flanking peptide0906230A:Mammalia:preprocalcitonin derived peptide1706329A:Mammalia:pro-bursin 14-peptide1412264A:Mammalia:pro-enkephalin A 209-2390501205A:Mammalia:prolactin fragment,pre1503284A:Mammalia proliferation inhibiting 4-peptide0905217A:Mammalia:protein B,alpha2HS glyco711659A:Mammalia:protein,glyco0409335A:Mammalia protein,myeloma,IF3 H chain fragment1508220A:Mammalia:Pyr-Glu-Pro-NH20401165A:Mammalia:redox active peptide1503186A:Mammalia:relaxin A1010328B:Mammalia:renin precursor N terra0705052A:Mammalia:rigin0811244A:Mammalia:rimorphin0811266A:Mammalia:rimorphin1708211A:Mammalia:salivary His rich peptide721151A:Mammalia:scotophobin0706234A:Mammalia:secretin1111240A:Mammalia:secretin1311331A:Mammalia:secretin1508173A:Mammalia:secretin1608239A:Mammalia:secretin1707342C:Mammalia:secretin700579A:Mammalia:secretin1009234A:Mammalia:secretin,pro1310205A:Mammalia:seminal peptide 201509222A:Mammalia:seminal plasma peptide,major1509222B:Mammalia:seminal plasma peptide.minor710685B:Mammalia:somatoliberin1007198A:Mammalia:somatostatin730308A:Mammalia:somatostatin760556A:Mammalia:somatostatin0706466A:Mammalia:somatostatin 140701278A:Mammalia:somatostatin 250701278B:Mammalia:somatostatin 281101392A:Mammalia:somatostatin 280602201A:Mammalia:somatostatin,N term extended710685A:Mammalia:somatotropin releasing 10-peptide0703632A:Mammalia:spinal cord 3-peptide0804312A:Maramalia:spinal cord 3-peptide1305396A:Mammalia:steroidogenesis activator protein1516350A:Mammalia:substance P710757A:Mammalia:substance P730714A:Mammalia:substance P1102255B:Mammalia:superoxide dismutase N term,Cu/Zn 0612159A:Mammalia:synaptosomal 3-peptide 0612159B:Mammalia:synaptosomal 4-peptide 1109246C:Mammalia:T cell receptor alpha TA20 1106231B:Mammalia:T cell receptor beta2 1107197B:Mammalia:T cell receptor D/Jbeta 1107197C:Mammalia:? cell receptor D/Jbeta
-146

1209256R:Mammalia:T cell receptor J alpha 1209256T:Mammalia:T cell receptor J alpha 1209256V:Mammalia:T cell receptor J alpha 1209256W:Mammalia:T cell receptor J alpha 0401182A:Mammalia:thymic factor 1408159A:Mammalia:thymic humoral factor gamma2 1714203A:Mammalia:thymocyte growth peptide 0503215A:Mammalia:thymosin alphal 770552A:Mammalia:thymosin alphal 690932A:Mammalia:thyroliberin 700037A:Mammalia:thyroliberin 0608648A:Mammalia:toxin,uremic1002226A:Mammalia:transferase Munich,HG phosphoribosyl 0311213A:Mammalia:trypsinogen 2nd activation peptide 0507223B:Mammalia:trypsinogen N term fragment,cationic 732142A:Mammalia:tuftsin1814294A:Mammalia:tumor invasion inhibiting factor 20409242A:Mammalia:uremic 3-peptide0509213A:Mammalia:uremic 5-peptide0410178A:Mammalia:uremic 7-peptide0901251A:Mammalia:uremic peptide0810210A:Mammalia:urokinase A11111231A: Mammalia-.valosin0603334A:Mammalia:vasoactive 11-peptide0603334B:Mammalia:vasoactive 5-peptide0603334C:Mammalia:vasoactive 5-peptide0601216A:Mammalia:vasoactive intestinal peptide1007235A:Mammalia:vasoactive intestinal peptide1012267A:Mammalia:vasoactive intestinal peptide1106144A:Mammalia:vasoactive intestinal peptide1109226A:Mammalia:vasoactive intestinal peptide1301342A:Mammalia:vasoactive intestinal peptide1301342B:Mammalia:vasoactive intestinal peptide1616408A:Mammalia:vasoactive intestinal peptide1707342A:Mammalia:vasoactive intestinal peptide1809387A:Mammalia:vasoactive intestinal peptide720473A:Mammalia:vasoactive intestinal peptide0909628A:Mammalia:vasopressin metabolite0611243A:Mammalia:vasopressin,2-Phe530215A:Mammalia:vasopressin,Arg580093B:Mammalia:vasopressin,Arg580181B:Mammalia:vasopressin,Arg590227B:Mammalia:vasopressin,Arg600454C:Mammalia:vasopressin,Arg600454D:Mammalia:vasopressin,Arg610025A:Mammalia:vasopressin,Arg640049B:Mammalia:vasopressin,Arg681070A:Mammalia:vasopressin,Arg690929B:Mammalia:vasopressin,Arg730938B:Mammalia:vasopressin,Arg0909223C:Mammalia:vasopressin, Lys520042A:Mammalia:vasopressin, Lys530011A .'Mammalia: vasopressin, Lys610024A:Mammalia:vasopressin,Lys681070B:Mammalia:vasopressin, Lys690929C:Mammalia:vasopressin, Lys0603289A:Mammalia:vasopressin,N-Ala-Gly Arg0603289B: Mammalia-.vasopress in, N-Val-Asp Arg701174A:Mammalia:vasotocin1806498A:Mammalia:VIP1616409A:Mammalia:xenopsin precursor1614424A:Mammal!a:xenopsin-related peptide1817263N:Mammalia:Zn finger protein ZNF601508175A:Mollusca:achatin I1712115A:Mollusca:achatin I0910276A.-Mollusca: alpha bag cell peptide
-147

1412184A:Mollusca:alpha conotoxin SI 1718180A:Mollusca:alpha conotoxin SIA 1605128A:Mollusca:APGWamide 1711282A:Mollusca:APGffamide 1411339A:Mollusca:buccalin 1203283B:Mollusca:califin S 1607138A:Mollusca:cardio-excitatory peptide 0901287A:Mollusca:cardioactive peptide 0309321A:Mollusca:cardioexcitatory peptide 1312259A:Mollusca:catch relaxing peptide 1806388A:Mollusca:cephalotocin 1607264A:Mollusca:conantokin T 1314217A:Mollusca:conopressin 1314217B:Mollusca:conopressin 0706261A:Mollusca:conotoxin GI 0706261B:Mollusca:conotoxin GIA 0706261C:Mollusca:conotoxin GII 1108256A:Mollusca:conotoxin GIIIA 1108256B:Mollusca:conotoxin GIIIB 1108256C:Mollusca:conotoxin GIIIC 1011179A:Mollusca:conotoxin GVIA omega 0812169A:Mollusca:conotoxin MI • 1501345A:Mollusca:egg laying hormone precursor 1207245A:Mollusca:egg laying peptide A-AP 630171A:Mollusca:eledoisin1003242A-.Mollusca 1003242B:Mollusca 1006817A:Mollusca 0811378A:Mollusca 1201209A:Mollusca 1309313A:Mollusca 1309313B:Mollusca 1714188A:Mollusca 1504408A:Mollusca 1310373A:Mollusca 1718371A:Mollusca 1407114A:Mollusca 1810269A:Mollusca 1810269B:Mollusca 1810269C-.Mollusca 1810269D:Mollusca 1810269E:Mollusca 1815162A:Mollusca 1815162K:Mollusca 1815162B:Mollusca 1815162C:Mollusca 1815162D:Mollusca 1815162E:Mollusca 1815162F:Mollusca 1815162G:Mollusca 1815162H:Mollusca 1815162J:Mollusca 1506519B:Mollusca 1506427A:Mollusca 1602255B .'Mollusca 1203403A:Mollusca 1816140A:Mollusca 1101325A:Mollusca 1304344A:Mollusca 0906220A:Mollusca 0906220B:Mollusca 1803237B:Mollusca 1803237C:Mollusca 1803237A:Mollusca
1610245B:Nematoda 1813281A:Nematoda
enkephalin,Leu enkephalin,Met enkephalinyl-Arg-Phe,Met FMRFamide like peptide FMRFamide like peptide FMRFamide like peptide 2 FMRFamide like peptide 3 fulicinmolluscivorous conus toxinmyomodulinmyomodulin Bmytilus inhibitory peptidemytilus inhibitory peptidemytilus inhibitory peptidemytilus inhibitory peptidemytilus inhibitory peptidemytilus inhibitory peptideMytilus inhibitory peptide A1Mytilus inhibitory peptide A10Mytilus inhibitory peptide A2Mytilus inhibitory peptide A3Mytilus inhibitory peptide A4Mytilus inhibitory peptide A5Mytilus inhibitory peptide A6Mytilus inhibitory peptide A7Mytilus inhibitory peptide A8Mytilus inhibitory peptide A9neuropeptide RISbetapedal peptidepeptide Ipeptide II-peptide PYFsleeper peptidesmall cardioactive peptide Atoxin I.geographutoxin Il.geographutoxin TxIAtoxin TxIBtoxin TxIIAceh-12 geneFMRFamide
—148 —

1611406A:Nematoda:neuropeptide AF1 1813281B:Nematoda:SADPNFLRFamide 1109265A:Nematoda:vitellogenin 1 1109265B:Nematoda:vitellogenin 2 1109265C:Nematoda:vitellogenin 4 1109265D:Nematoda:vitellogenin 6 0403176A:Osteichthyes:angiotensin I1510162A:Osteichthyes:angiotensin-converting enzyme inhibitor 1805355A:Osteichthyes:angiotensin-converting enzyme inhibitor B-l 1805355B:Osteichthyes:angiotensin-converting enzyme inhibitor B-2 1805355C:Osteichthyes:angiotensin-converting enzyme inhibitor B-3 1717175A:Osteichthyes:ANP1515184A:Osteichthyes:atrial natriuretic factor 1613139A:Osteichthyes:BNP like natriuretic peptide 1709312A:Osteichthyes:carassin 721464A:Osteichthyes:clupeine Y20803150A:Osteichthyes:corticotropin like interm lobe peptide0508149A:Osteichthyes:endorphin0604188A:Osteichthyes:endorphin II0708424A:Osteichthyes:glucagon1105264A:Osteichthyes:glucagon1205373A:Osteichthyes:glucagon1307182B:Osteichthyes:glucagon1716265A:Osteichthyes:glucagon1709218B:Osteichthyes:glucagon-like peptide1206278A:Osteichthyes:glycoprotein 7,antifreeze1206278B:Osteichthyes:glycoprotein 7,antifreeze1206278C:Osteichthyes:glycoprotein 7R,antifreeze1206278D:Osteichthyes:glycoprotein 8R,antifreeze690924A:Osteichthyes:insulin1412252A:0steichthyes:insulin A1412252B:Osteichthyes:insulin B1603264A:Osteichthyes:insulin C peptide1718169A:Osteichthyes:intestinal peptide EIPP620022A:Osteichthyes:isotocin650767A:Osteichthyes:isotocin681068A:Osteichthyes:isotocin690928A:Osteichthyes:isotocin701183A:Osteichthyes:isotocin765600A:Osteichthyes.'learning induced brain peptide 765600B:0steichthyes:learning induced brain peptide 0905260A:Osteichthyes:luliberin0910249A:Osteichthyes:melanin concentrating hormone0612177A:Osteichthyes:melanotropin0511280A:Osteichthyes:melanotropin alpha0511280B:Osteichthyes:melanotropin beta0608228A:Osteichthyes:melanotropin II alpha671037A:Osteichthyes:mesotocin701188A:Osteichthyes:mesotocin1814320C:Osteichthyes:neurokinin A1403319A:Osteichthyes:oxyntomodulin610242A:Osteichthyes:oxytocin1208325A:Osteichthyes:peptide,pro-aPY1412186A:Osteichthyes:polysialo glycoprotein repeat seq1412186B:0steichthyes:polysialo glycoprotein repeat seq1313241A:Osteichthyes:pro-GIF derived peptide SI1313241C:Osteichthyes:pro-GIF derived peptide S21209204A:Osteichthyes:protamine1209204B:Osteichthyes:protamine0601202A:Osteichthyes:somatostatin1012240A:Osteichthyes:somatostatin 221210254A:Osteichthyes:somatostatin 251101339A:Osteichthyes:somatostatin 281305261A:Osteichthyes:somatostatin 281010245A:Osteichthyes:somatostatin 28 II0605221A:Osteichthyes:somatostatin I
-149

1103330A:Osteichthyes:somatostatin precursor1414253A:0steichthyes:somatostatin/glucagon related peptide1414253B:0steichthyes:somatostatin/glucagon related peptide0503186A:Osteichthyes:stellin A764668A:0steichthyes:sturine B1814320A:Osteichthyes:substance P1814320B:Osteichthyes:substance P753677A:Osteichthyes:thyroliberin0902237A:Osteichthyes:urotensin I1701464B:0steichthyes rurotensin I N-terminal flanking peptide0610250A:Osteichthyes rurotensin II1817244A:0steichthyesrurotensin II1007166A:Osteichthyes:urotensin Ilalpha1007166B:Osteichthyes rurotensin Ilbeta1007166C:Osteichthyes rurotensin Ilgamma0909285A:Osteichthyesrurotensin UIIA0909285B:0steichthyesrurotensin UIIB610017ArOsteichthyesrvasotocin610023Ar0steichthyes:vasotocin610203ArOsteichthyes rvasotocin610242Cr0steichthyesrvasotocin650767B:Osteichthyesrvasotocin681068B:0steichthyesrvasotocin690928B:Osteichthyesrvasotocin701183B:Osteichthyesrvasotocin1603248ArOsteichthyesrVIP related peptide1601305ArPlantaerACE inhibitory peptide FLP 11601305B:PlantaerACE inhibitory peptide FLP 21601305C:PlantaerACE inhibitory peptide FLP 31508168A:Plantaeragglutinin S1810244A:Plantae:antimicrobial peptide 11810244B:Plantae:antimicrobial peptide 21709506A:Plantae:cycloleonurinin671044A:Plantae:fibrinopeptide A671044D:Plantaerfibrinopeptide B1108239E:Plantae:gliadin gamma1103178B:Plantae:inhibitor CPGTI-I,trypsin1103178C:Plantae:inhibitor CPTI-II,trypsin1103178F:Plantae:inhibitor CSTI-IV,trypsin0708183A:Plantae:inhibitor III,trypsin0903201A:Plantae:inhibitor LTD I,trypsin iso0903201B:Plantae:inhibitor LTD III,trypsin iso750673ArPlantaermelanotropin beta1603356CN:PlantaerORF 231603356CD:Plantae:ORF 281603356N:Plantae 1506382A:Plantae 1405344C:Plantae 1010307E:Plantae 1211248A:Plantae 1503172C:Plantae 1503172A:Plantae 1503172B:Plantae 1715381A:Plantae 1314174A:Plantae 1314174B:Plantae 1702179A:Plantae 1801181A r Plantae 1801181B:Plantae 1817218A:Plantae 1817218B:Plantae 1817218C:Plantae 1111187A:Plantae 1111187CrPlantae
ORF 29protease inhibitor ribosomal protein L2 ribosomal protein S19'S rich 2S protein S Ser protease inhibitor MCEI-I Ser protease inhibitor MCTI-I Ser protease inhibitor MCTI-II systemintrypsin inhibitor trypsin inhibitor trypsin inhibitor trypsin inhibitor 1 trypsin inhibitor 2 trypsin inhibitor I trypsin inhibitor II trypsin inhibitor IIIURF 601
: URF 6031414347B:Prokaryota:aceB gene assocd ORF 1109139B:Prokaryota:amylase alpha
-150

1708222A:Prokaryota:anantin/des-Phe analog 1616302A:Prokaryota:appA assocd ORF X 1512237B:Prokaryota:bfr assocd ORF 24’1702271B:Prokaryota:cinnamycin1312339D:Prokaryota:colicin E9 lysis gene1004221A:Prokaryota:dehydrogenase fragment K,glucose1616249A:Prokaryota:delta-like toxin1501381C:Prokaryota:dicB operon ORF1702271A:Prokaryota:duramycin1702270A:Prokaryota:duramycin B1702270B:Prokaryota:duramycin C1101217A:Prokaryota:enterotoxin,heat stable1111219A:Prokaryota:enterotoxin,heat stable1201224A:Prokaryota:enterotoxin,heat stable1314236C:Prokaryota:F sex factor traD1208313B:Prokaryota:flavoprotein1414238A:Prokaryota:gallidermin1310302B:Prokaryota:gene cotB1308167H:Prokaryota:gene F1313282B:Prokaryota:gene glgA1111234B:Prokaryota:gene glnL1309264A:Prokaryota:gene nodD1309261B:Prokaryota:gene orf191108230B:Prokaryota:gene tmpB1302279A:Prokaryota:gene trpE1307194A:Prokaryota:gene trpL1109252A:Prokaryota:germ specificity regulating peptide1504256J:Prokaryota:H translocating ATPase beta1506318A:Prokaryota:halo toxin1813209A:Prokaryota:heat-stable enterotoxin0608214A:Prokaryota:hemolysin delta1005204A:Prokaryota:hemolysin delta1812252A:Prokaryota:lad gene1302189B:Prokaryota:leader peptide ermG1302189C:Prokaryota:leader peptide ermG1810255A:Prokaryota:marinestat in Cl1810255B:Prokaryota:marinostatin C21805264A:Prokaryota:marinostatin D1109142A:Prokaryota:microcin 71105311B:Prokaryota:ORF1413245C:Prokaryota:ORF 21502355C:Prokaryota:ORF 21803344B:Prokaryota:ORF 2 in noncoding region of sra gene 1209230C:Prokaryota:ORF 31803344C:Prokaryota:ORF 3 in noncoding region of sra gene1209230D:Prokaryota:ORF 41807289B:Prokaryota:ORF 7K1305262A:Prokaryota:ORF E1010255A:Prokaryota:ORF in ori1012216A:Prokaryota:ORF N term,toluate operon1615170D:Prokaryota:pdhA assocd ORF1202257G:Prokaryota:peptide 11202257H:Prokaryota:peptide 21101257A:Prokaryota:pheromone cADI.bacterial sex1502267B:Prokaryota:phoP gene0504198A:Prokaryota:phosphorylase fragment,maltodextrin1010260B:Prokaryota:phycobiliprotein N term1109172A:Prokaryota:phycocyanin beta1402269F:Prokaryota:plasmid virD1504241B:Prokaryota:PQQ gene assocd ORF1810428B:Prokaryota:pqqA gene1002248A:Prokaryota:protein araE,regulatory0812230A:Prokaryota:protein fragment,replication initiation1002280B:Prokaryota:protein H,reaction center1002280C:Prokaryota:protein L,reaction center1002280D:Prokaryota:protein M,reaction center
-151-

1501278D:Prokaryota 1501278E:Prokaryota 1807309C:Prokaryota 1807238A:Prokaryota 1403333B:Prokaryota 1712183A:Prokaryota 0907240E:Prokaryota 1516355H:Prokaryota 1501251B:Prokaryota 1612377A:Prokaryota 1210251A:Prokaryota 1413289A:Prokaryota 1708286A:Prokaryota 1708286B:Prokaryota 1616276A:Prokaryota 1313358C:Prokaryota 1203213B:Prokaryota 1012306A:Prokaryota 0709232A:Prokaryota 0902149A:Prokaryota 0902181A:Prokaryota 0906203A:Prokaryota 1812252B:Prokaryota 1704313A:Prokaryota 1513305A:Prokaryota
pSPA12 gene pSPA13 gene pufK genepyrrolo-quinoline-quinone synthesis gene repAX genereplication locus ORE 30ribosoraal protein S17ribosomal protein S8rpoB gene assocd ORErRNA methylase assocd OREsex pheromone cAM373sex pheromone cCFlOsex pheromone inhibitor iADlsex pheromone inhibitor iADl downstream OREshl leader peptidestreptomycin syn gene aphDsynthase,enolpyruvylshikimate phosphatesynthetase N term,Gintoxin,heat stable enterotoxin,heat stable enterotoxin,heat stable enterotoxin,heat stable enterotraE upstream ORE TTS1trp operon leader peptidetrpEG attenuator peptidetrpGDC operon leader peptide1607231A:Prokaryota
1805362A:Reptilia:alpha MSH 0402224A:Reptilia:angiotensin 0402224B:Reptilia:angiotensin 0402224C:Reptilia:angiotensin710784A:Reptilia:angiotensin converting enz inh V2 710784B:Reptilia:angiotensin converting enz inh V6I 710784C:Reptilia:angiotensin converting enz inh V6II 710784D:Reptilia:angiotensin converting enz inh V7 710784E:Reptilia:angiotensin converting enz inh V8 710784F:Reptilia:angiotensin converting enz inh V9 1002212A:Reptilia:angiotensin I0903145A:Reptilia:bradykinin potentiating peptide1005285A:Reptilia:bradykinin potentiating peptide1208380A:Reptilia:bradykinin potentiating peptide690149A:Reptilia:bradykinin potentiating peptide A700017A:Reptilia:bradykinin potentiating peptide B710067A:Reptilia:bradykinin potentiating peptide C690149B:Reptilia:bradykinin potentiating peptide D700471A:Reptilia:bradykinin potentiating peptide E1617159A:Reptilia:bradykinin potentiating peptide S3-.11617159D:Reptilia:bradykinin potentiating peptide S4-.l-.2-1617159B:Reptilia:bradykinin potentiating peptide S4-,3,l-1617159C:Reptilia:bradykinin potentiating peptide S4-.3.2-1617159H:Reptilia:bradykinin potentiating peptide S5-.1-1617159G:Reptilia:bradykinin potentiating peptide S5-.2-1617159F:Reptilia:bradykinin potentiating peptide S5-.3-1617159E:Reptilia:bradykinin potentiating peptide S5-.4-700202A:Reptilia:bradykinin potentiating peptide V3A1607186A:Reptilia:bradykinin,6-Thr701176A:Reptilia:bradykinin,6-Thr721948B:Reptilia:fibrinopeptide A721948E:Reptilia:fibrinopeptide B1612382B:Reptilia:glucagon1709419A:Reptilia:lethal toxin I1711481A:Reptilia:luliberin I1711481B:Reptilia:luliberin II690927A:Reptilia:mesotocin721912A:Reptilia:mesotocin610522B:Reptilia:oxytocin660031A:Reptilia:peptide A,venom
152-

660031B:Reptilia:peptide B,venom 1208381A:Reptilia:peptide POL236 1410356A:Reptilia:sarafotoxin S6al 1410356B:Reptilia:sarafotoxin S6b 1410356C:Reptilia:sarafotoxin S6c 1515248A:Reptilia:sarafotoxin SRTXd 610522D:Reptilia:vasotocin 680069A:Reptilia:vasotocin 69092TB:Reptilia:vasotocin 721912B:Reptilia:vasotocin1813189A:SUBUNIT=A chain:ISOTYPE=II:Mollusca:insulin-related peptide 1803251A:SUBUNIT=A chain:Reptilia:insulin1810291A:SUBUNIT=A:IS0TYPE=III:Mollusca:insulin-related peptide1715201A:SUBUNIT=A:Mammalia:relaxin1712127A:SUBUNIT=A:0steichthyes:insulin1812293A:SUBUNIT=A:Osteichthyes:insulin1803251B:SUBUNIT=B chain:Reptilia:insulin1715201B:SUBUNIT=B:Mammalia:relaxin1812293B:SUBUNIT=B:Osteichthyes:insulin1715168M:SUBUNIT=ganuna:IS0TYPE=1:Reptilia:crotoxin A11715168N:SUBUNIT=gamma:IS0TYPE=2:Reptilia:crotoxin A11715168P:SUBUNIT=gamma:IS0TYPE=3:Reptilia:crotoxin A11715168Q:SUBUNIT=gamma:IS0TYPE=4:Reptilia:crotoxin A11716290A:SUBUNIT=heavy chain:Mammalia:Ig J1 segment1716290B:SUBUNIT=heavy chain:Mammalia:Ig J2 segment1716290C:SUBUNIT=heavy chain:Mammalia:Ig J3 segment1716290D:SUBUNIT=heavy chain:Mammalia:Ig J4 segment1303320A:Virus 1615267B:Virus 1304195A:Virus 1414220D:Virus 1304292E:Virus 1604374K:Virus 1813170G:Virus 1206374B:Virus 1806326E:Virus 1712189A:Virus 0403209A:Virus 1210381A:Virus 1412351B:Virus 1408236A:Virus 1710262B:Virus 1709308B:Virus 1301338X:Virus 1112178M:Virus 1209333D:Virus 1309330A:Virus 1715306B:Virus 0908286L:Virus 1202223A:Virus 1512364C:Virus
art gene exon 1capsid proteinearly leader proteinG proteingene 1.5gene 5.1gene Jgene Lgene VIIgenome linked protein VPginternal peptide VIIOREOREORE 1ORE 26ORE 30ORE 58.3ORE 7ORE IIB satellite RNA5 ORE satellite RNA ORE,5'-terminal protein 1.5 protein Cprotein gp51
0902231AQ:Virus:protein orf28 0908283B:Virus:protein VPg 1105312L:Virus:protein VPg 1011278A:Virus:protein VPx N term 1009209A:Virus:RNA 5 1402304A:Virus:RNA polymerase 1403393C:Virus:stp gene ORE B 1505419B:Virus:tat gene 1412350B:Virus:terminal protein 1504354C:Virus:upstream ORE
* Animal ia O <59fa"f"o
*707 t‘—t ^ x^(;fi:7;i77<7 MKtz
-153-

^^r-f- F (7O7t-fa^STR30N)
1 9 > ? !R S E 7 5 / E T t£ V T $ / E £ is /l /d>" 7* tK 93.2 strn2 (DNA (c 3 - ^ntl^il^7'f K CD V x h )
3 mm)
5 n=«i£ tilgiffifc jl S&g#* S x D=D-T ; 7 E££t/6 LIST OF PEPTIDES CONTAINING NON PROTEIN AMINO ACIDS7 FORMAT=LITERATURE NO:NAME:COMMENT(structure,component,propery)
0608217 :Aad-Ala-Val Aad-Ser-Val Aad-Ser-IsodehydroVal:1305532 :ACE Inhibitor K13:Cyclo(Tyr-Isodityrosine) Deriv 1305533 :ACE Inhibitor WF10129:MeCH(OH)(CH2)2C0CH2CH(COOH)-Ala-Tyr 1306580n:Actinoidin:Glycopeptide Antibiotic 0512246n:Adenochrome:Fe Binding Peptide 0602198n:Amaninamide:1412598n:Amphibine:Cyclopeptide Alkaloid1103492D:Ancovenin:AVQaAXFGPLXWSADGNXK a=D-Ala1610381 :Ancovenin:Lanthionine,Abu,DehydroAla1815450 :Anthelmintic PF1022A:Cyclo(Lac-MeLeu-3-Ph Lac-MeLeu)21819804n:Antibiotic A10255B/G/J:Thiopeptide1508428n:Antibiotic A42867:Vancomycin Family1612612n:Antibiotic A54145:Lipopeptide1409561n:Antibiotic A83586C:Depsipeptide1011404 :Antibiotic BMY28160:Cyclodepsi-10-Peptide Dbu(Ac),Val,Phe,Leu,Ser0907210 :Antibiotic Bu2470:Dbu-8-Peptide1612607D:Antibiotic FR112123:Lys(Gly-Glu)-D-Ala1714591n:Antibiotic GE2270A:Thiazolyl Peptide0906226P:Antibiotic K582:Arg-Arg(0H)-0rn-Thr-0rn-Lys-Tyr1717651 :Antibiotic MM55266/M55268:Glyco-7-Peptide0701171n:Antibiotic Nol907:0701170n:Antibiotic P168:1008327 :Antibiotic P168:9-Peptide 6-0H-4-Me-8-0xo Aminodecanoate,3-OH Ile.Aib 1816436 :Antibiotic P168:Aib-9-Peptide MePro,Hyl,Aib,Leu,beta Ala.AHMOD 1403592 :Antibiotic Sch37137:Ala-Gin(X) X=Epoxy Dicarboxylic Acid Monoamide 0907171 :Antitumor Cyclo-6-Peptide:1506745P:Aphostatin:Ile-Ile-Ser(Phospho)-Gln-Glu 1211448 :Argiopine:C6H3((OH)2)CH2CO-Arg-Polyamine-Arg 1313442 :Argiotoxin:Arg-Polyamine-Asn 1803716 :Aureobasidin:Cyclodepsi-9-Peptide 1803717 :Aureobasidin:Cyclodepsi-9-Peptide1718512 :Aureobasidin:MeVal,Phe,MePhe,Pro,alle,MeVal,Leu beta-OH MeVal 1307361D:Avellanin:Cyclo(D-MePhe-D-Ala-Ile-o-Abe-Pro)1803768P:Axinastatin:Cyclo(Asn-Pro-Phe-Val2-Pro-Val)1202461D:Azinothricin:3-0H Leu,D-Thr,D-MeAla,HO-Ser(Me) D/L-Piperazic Acid1406382 :Azotobactin:D/L-Ser,3-0H D-threo-Asp,D,G,Hse,D-Citrulline,D-Orn(Ac,OH)1305415 :B1625 FA2betal:Thr-Val-Sar-MeVal Deriv1201380D:Bacillomycin:beta AA,D-Tyr,D/L-Asn,Gin,Pro,Thr0709212D:Bacillomycin:beta AA,Asp,D-Tyr,D-Asn,Pro,Glu D-Ser,Thr0509590D:Beauverilide:Cyclo(Hmd-Val-Phe-D-alle)1212537n:beta Lactam Potentiator:0 Sulfated Glycopeptide0808325P:beta MAPI:CO(Phe}-Arg-Val-Phe-H1903606 :Bistratamide:Cyclo(Ala-Tzl-Val-Tzl-Val-Ozl)1903606 :Bistratamide:Cyclo(Ozl-Val-Ozl-Val-Tzl-Val-mOzn)1410632n:Blastolysin:1503540 .-Blue Green Algae Toxin I/II:Cyclo(Ala-Leu-MAsp-Arg-Adda-Glu-Mdha) 1815454n:Bottromycin:0707285 :Brevigellin:Cyclodepsi-5-Peptide 1818672 :BZR Cotoxin:Depsi-9-Peptide 1705341P:Cadystin:(Glu(Cys))n-Gly n=2/3 1010441P:Cadystin:Glu(Cys-Glu(Cys-Glu(Cys-Gly)))1807606D:Calophycin:Cyclo(AUNQGRJPXV) U=D-Asp,J=MeAsn X=Aminopalmitate Deriv0907212P:cAMP Phosphodiesterase Inhibitor:Acyl-Glu-Leu2-Val-Asp-Leu2 Lactone1310405P:Caricastatin:Ac-Leu-DL-Arg-H1803667P:Cd Binding Peptide:((Glu(Cys))4-Gly0612201 :Celenamide:4-Peptide1110365 :Cephabacin:Ala-Ser-0rn-Val-0rn-Val1008320 :Cephalosporin:4-Peptide0808184 :Cirratiomycin:7-Peptide
#5 .
-154-

1610297 :Citropeptin:Cyclo-Depsi-6-Peptide Antibiotic 1603866P:Cleromyrine:Cyclo(Ala-Gly-Pro-Ile-Val-Phe)1711651n:Conagenin:1006359 :Cortinarin:Bicyclodepsi-10-Peptide 1715556 :Cortinarin:Cyclic 10-Peptide1403721D:Cyanogenosin:MeDehydroAla,D-Ala,Arg,D-Asp(3-Me),Arg beta-AA,D-IsoGlu 1202488 :Cyanoginosin:Cyclo-7-Peptide1011415D:Cyanoginosin:D-Ala,Leu,beta-Me D-Asp,Ala,Adda,D-Glu N-Me DehydroAla 1312673 :Cyanoviridin:Cyclo(Ala-Arg-Masp-Arg-Adda-Glu-Mdha)1805429P:Cyclo(APFWGGP):1805429P:Cyclo(EAEWGEVP):1305652P:Cyclo(Hyp-Leu):1505628P:Cyclo(Hyp-Phe):1305652P:Cyclo(Hyp-Pro):1805429P:Cyclo(LIATGTF):1805429P:Cyclo(LLPYGSP):1503521P:Cyclo(PNSXNYNQ):0706215P:Cyclo(Thr-Val2-Leu) N-Acyl Deriv:1306545P:Cycloaspeptide:Cyclo(Ala-Phe-Leu-Tyr-X)0809267D:Cyclosporin:Cyclo-10-Peptide MeVal,MeLeu,D-Ala 1212534D:Cytostatic Peptide WF3161:Cyclo(Leu-Pip-Aoe-D-Phe)1516458D:D-Asp(Gly):1502533 zDecadienoic Acid Deriv:Cyclic 7-Peptide Toxin 1616628 :Depsidomycin:Cyclodepsi-7-Peptide1409638 :Depsipeptide FR900359:Ala,MeAla,Leu(beta-OH),Thr(Me),MeDehydroAla1809743D:Desferri Ferribactin Pyoverdin:D-Ala,Dbu,Glu,Lys,Orn,D-Ser,Thr,D-Tyr0710282 :Destruxin:Depsi-6-peptide,MeAla,MeVal,Pipecolic Acid0710238 :Detoxin Complex Congener:lie,Val,Phe,Detoxinine1005339P:Diprotin:Ile-Pro-Ile,Val-Pro-Leu1212595 :Discarin:3-0H Leu,MeIle,Trp,p-0H Styrylaraine1104476D:Discodermin:MeGln,t-Leu,D-Trp,D-Cys(OSH),D-Leu1012457D:Discodermin:Thr,Asn,Sar D-Ala,D-Pro,D-Trp,D-Cys,D-Leu D/L-tLeu,MeGln 1811428 :Discokiolide:Phe,Pro,Asn,MeIle,Asp,Discokiic Acid 1407508 :Dityromycin:MeVal,Pro,Val,Gly(Ph),Me2Thr,DehydroAA,Epoxide Deriv 1212543D:Dolastatin:Cyclo(Pro-D-Leu-Val-D-Thz-Thz)1314416 :Dolastatin:Cyclo(Val-Pro-Leu-(gln)Thz-(gly)Thz)1512544n:Dolastatin:Depsipeptide1602473 :Dolastatin:Gly2,Ala2,MeLeu,MeTyr(Me),MeVal,Aminopentanoate1604691 :Dolastatin:Me2Val-Val-MeVal-Pro2-Hiv-Dpy0803206n:Dolastatin:Thiazole Containing Cyclopeptide0507218 :Echinocandine:Cyclo-6-Peptide1111420 :Edeine:Gly(Ph)-beta-Ala(OH)-Dbu-Dha-Gly-Spe0806152n:Efrapeptin:1811540 :Efrapeptin:15-Peptide1706616 :Efrapeptin:Pip,Aib,beta-Ala,Iva,Ala,Leu,Gly1010384D:Empedopeptin:Depsi-9-Peptide D/L-Pro,D-Ser,Arg,3-Hyp,D/L-beta-OH Asp1805543D:Endothelin Antagonist:Cyclo(D-AA-Leu-D-Trp-D-Glu-Ala)1804602D:Endothelin Binding Inhibitor:Cyclo(D-Glu-Ala-D-Val-Leu-D-Trp)1817583n:Enniatin:Cyclodepsipeptide1804729 :Enopeptin:Cyclodepsi-5-Peptide0901283 :Ergobutyrine:Aib,Pro,Leu1901570 :Eurystatin:Leu,Orn,3-Amino-2-Oxobutyric Acid 1806831 :Fellutaraide:3-OH Decanoyl-Asn-Gln-Leu-H 1501727P:Fenestin:Cyclo(Pro2-Leu-Ile),Cyclo(Pro-Val-Pro-Leu-Ile) 0608265P:Feruloyl-Gly-Phe:1112362D:Fluoroactinomycin:(MeVal-Sar-X-D-Val-Thr)2 Deriv1301475 :Folyl Poly-Glu:Abe-(Glu)n n=2-61201394 :Foroxymithine:For-0rn(For,OH)-Ser-Cyclo-2-Peptide0906301 :Fosfazinomycin:Val-Arg-NHNMePO(OH)CH(OH)COOMe1412598n:Frangufoline:Cyclopeptide Alkaloid1808459n:Galacardin:Glycopeptide Antibiotic1212564 :gamma-Glu Peptide:Glu(Cys(Me)-beta-Ala),Glu(Orn(Ac))1606469n:Geodiamolide:Cytotoxic Cyclodepsipeptide 0502218P:Globomycin:Cyclo(alle-Ser-aThr-Gly-X-MeLeu)0712212 :Globomycin:Cyclodepsi-6-Peptide 1505533P:Glu(Asp):
—155 —

1211549 :Glu(D-Hph):Hph=3-Ph-3-Aminopropanoic Acid 1413528D:Glycopeptidolipid Antigen:X-D-Phe-aThr(X)-D-Ala-Ala-X 1801429D:Glycopeptidolipi :D-Phe-D-aThr-D-Ala-Alaninol 1007292n:Gramicidin S2/S3:1505427P:GSH Analog:Glu(Ser-Gly)1603557 :Halocyamine:His-Phe((OH)2)-Gly-Tryptamine Deriv1707360 :Harmaomycin:aThr,lie,3-Me Phe,4-Propenyl Pro 3-Nitrocyclopropyl Ala 0804347 :HC Toxin:Cyclo-4-Peptide 9,10-Epoxy-8-0xo Aminodecanoate Ala2,Pro 0901322 :HC Toxin:Cyclo-4-Peptide Ala,Pro,Aminoepoxyoxodecanoic Acid 1709567n:Helvecardin:Glycopeptide Antibiotic 1515700 :Hepatotoxin:Arg-isoAsp-Arg-Adda-isoGlu-mdhAla-Ala 1703716D:Hepatotoxin:Cyclic 7-Peptide D-Ala,Leu,D-MeAsp,D-Glu,Arg 1112392D:Herbicolin:X-Thr-D-aThr-D-Leu-Gly-D-Gln-Gly-Y-Arg 1210453P:Homophytochelatin:Glu(Cys)n-beta Ala n=2-71605356D:Hormaomycin:D-aThr Pro(Propenyl) threo Phe(Me) Nitro Cyclopentyl Ala 1810793D:Hormothamnin:D-Phe,D-Leu,lie,D-alle,Leu,Gly,D-Baoa Hse,Dhha,Hyp,Hse 1104477 :Host Specific Plant Toxin:Cyclo(Pro-Ala-Gly-Epd)0908203 :Host Specific Toxin:Cyclo(Pro-Ala2-0ed)1508276n:HV Toxin M:Cyclodepsipeptide 1009308 :Hypelcin:Aib-19-Peptide 1516538n:Hypeptin:Peptide Antibiotic 0802177 :Imacidin:10-Peptide Lactone1612548 rlmmunoreactive Lipopeptide WS1279:XNSGGS X=Acyl-Cys(Glyceryl)0811220D:Immunoreactive Peptide FK156:D-Lac-Ala-D-Glu(Dpm-Gly)0909205D:Isariin:Cyclo(X-Gly-Val-D-Leu-Ala-AA)0711220D:Isariin:Cyclodepsi-6-Peptide1305530 :Iturin:Asp3,Glu,Pro,Ser,Tyr Heterogeneous beta-AA 0808326 :Iturin:beta-AA,Asn3,Tyr,Gin,Ser 1006405D:Iturin:beta-AA,Pro,Ser,D-Asp,Asp,Glu,D-Tyr 1412491P:Iturin:SNPQNYNX1614572D:Janthinocin:lie,Thr,Ser,Phe,D-Ser,D-Orn beta-OH Trp,DehydroAbu, Hyl 1207499 :Jasplakinolide:Cyclodepsipeptide Ala.MeTrp(Br).beta Tyr 1409560 :Katanosin:Cyclic 11-Depsipeptide Asp,Leu,Phe,beta-OH Deriv 1801564 :Keramamide:Cyclo(Phe-MeCht-Leu2-Lys-C0-Phe) MeCht=MeTrp(Cl,OH)1801515 :Keramamide:Pro,Orn,lie,Abu,Nva,Trp(2-Br,5-0H),2-0H-3-Me Pentanoate 1819805 :Keramamide:Thiazole,DehydroTrp,IsoSer lie,Ala,Dpr,Aminohexanoate 1716594 :Konbamide:Cyclo((CO-Leu)-Lys-Ala-Leu-MeLeu-BhTrp)1708462 :L156602:3-OH Leu,D/L-Peperazic Acid,HO-Ala Tetrahydropyranylpropionate1509414P:Lanthibiotic:TAGPAIRASVKQCQKTLKATRLFTVSCKGKNGCK1414692 :Lantiopeptin:aRQaaXFGPFUFVadGNUX U=Abu d=3-0H Asp1109340 :Lavendomycin:6-Peptide erythro-Dbu.Me Arg,DehydroAbu,Pipecolic Acid1818556 :Laxaphycin:Ade,Aoc,Dhb,Hse,Hyp Leu(OH),Asn(beta-OH),MeIle1815462 :Leualacin:Leu,MePhe,beta Ala,R/S-Leucic Acid1302409 :Leucinostatin:9-Peptide Antibiotic1001277 :Leucinostatin:FA-Pro(Me)-X-HyLeu-Aib-Leu2-Aib2-Y1306583 :Leucinostatin:Pro(Me)-X-Hyl-Aib-Leu2-Aib2-beta-Ala-R0802173 :Leucinostatin:Pro(Me)-X-HyLeu-Aib-Leu2-Aib2-beta-Ala1308304D:Lipopeptide Antibiotic:R-WNDTGOrnD(A)DG(S)MeGluKyn ()=D-AA1505632 :Lipopeptide:3,4-(OH)2 HomoTyr,3-0H Gin,Hyp(Me),Hyp Thr,4,5-(OH)2 Orn0612271P:Lipopeptin:Cyclodepsi-8-Peptide Thr,Asp,Ser,MeAsn,MePhe,HyGln,Glu1611703 :Lissoclinamide:Cyclo-7-Peptide1404691D:Lophyrotomin:Bz-(A)(F)VI(D)D(E)Q ()=D-AA0909245D:Lophyrotoxin:Bz-Ala-Phe-Val-Ile-Asp-Asp-Glu-Gln D-AA at 1,2,5,7 1505693 :Lysobactin:3-0H Phe,3-0H Leu,3-0H Asn 1414667P:Maculosin:Cyclo(Pro-Tyr)1413479 :Maduraferrin:beta-Ala,Gly,Ser,MeOrn(OH)1003303 :Majusculamide:Cyclodepsi-9-Peptide 1818673 :Malaysiatin:Cyclo-7-Peptide 1803819n:Malformin:0606153 :Malonichrome:Cyclo-6-Peptide Ala,Gly,0rn(0H)1706617 :Melanostatin:MePhe-NHCH(C00H)CH2-His1815457n:Mersacidin:Lantibiotic1307321 :Methanofuran b:(gamma-Glu)4 Moiety1713741 :Microcystin XY:beta-MeAsp,Asp,Glu,Ala,Leu,Arg1903686n:Microcystin:1903687n:Microcystin:
-156

1903684D:Microcystin:7-Dha,3-D-Asp,7-Dha,3-D-Asp,7-Ser Deriv 1605607n:Microcystin:Cyclic Peptide Hepatotoxin 1903685 :Microcystin:Cyclo-7-Peptide1818675D:Microcystin:D-Ala,D-3-Me Asp,Adda,D-Glu N-Me DehydroAla1807605D:Microcystin:D-Ala,Leu,D-MeAsp,Arg,D-Glu1801655 :Microcystin:Toxic Cyclo-7-Peptide1207498 :Moroidin:Tricyclic 8-Peptide1810794 rMotuporin:Cyclo-5-Peptide1304500n:Mulundocandin:Lipopeptide Antibiotic0909236D:Mycoplanecin:Cyclodepsi-10-Peptide Me-D-Leu MeThr 4-Me/Et Pro 1803821P:Nazumamide:Bz((OH)2)-Arg-Pro-Ile-Abu1003314P:Neopeptin:Cyclodepsi-8-Peptide MeAsn,beta-MeGlu,beta-OH Gln.MePhe1203424P:Neopeptin:Ser,Asp,MeAsp,MePhe beta-Me Glu,beta-OH Glu1003248n:Neoviridogrisein Cls:Cyclodepsipeptide1405484 :Nephilatoxin:Lys,Asp,Orn,Putreamine,Cadavarine1504432 :Nephilatoxin:X-Orn-Asn-Cad-(Ptn)n-Y1306582 :Nitropeptin:Leu-Glu(beta-N02)1101617 :Nummularine:14/13 Merabered Cyclopeptide 1606455 :Nummularine:Cyclo-13-Peptide Alkaloid l307439n:Nummularine:Cyclopeptide 1402611n:Nummularine:Cyclopeptide 1412598n:Nummularine:Cyclopeptide1801514 :Orbiculamide:Pro,Ala,Abu,Orn,Trp(2-Br,5-OH),Theonalanine,Theoleucine1409563 :P Containging Antibiotic:Gly-Leu-Aminosiopropenyl Phosphonate1105414n:Paracelsin:Aib-Peptide1309443n:Parvodicin:Glycopeptide Antibiotic0806221 :Patellamide:lie,Val,Ser,Thr,Thiazole0912210n:Peptide Alkaloi :1506823P:Peptide Lactone:RCH(OH)CH2C0-Glu-Leu2-Val-Asp-Leu2-Leu2 0911233n:Peptide Monobactam SQ28332:1307406 :Peptidoglycan:GlcNAc-MurNAc-Ala-Glu-Dpm-Ala0912239 :Phomopsin:Cyclo-6-Peptide 3-OH lie,Dehydrolle,DehydroPro1302323P:Phytochelatin:(Glu(Cys))3-Gly1111493P:Phytochelatin:(Glu(Cys))n-Gly n=3-71004352D:Phytotoxic Cy:Cyclo(D-Tyr(Me)-Ile-Pip-Aoe)1203426 :Phytotoxic Cy:Epoxide AA 8-0xo-9,10-Epoxy Arainodecanoate 1813497 :Phytotoxin:Cyclodepsi-10-Peptide1311674 :Platelet Activating Factor Inhibitor:Cyclo(Met-Trp) Analog 1207405D:Plipastatin:E(0)Y(t)E(A)PQ(Y)I ()=D-AA t=aThr 0=0rn
1815456P:Plusbacin:aThr-Ala-Hyp-Arg-X-Ser-Hyp-X X=3-0H Asp 1311706P:Poly(Glu(Cys))-Gly:1809662 :Polydiscamide:A-Phe(Br)-P-tLeu-Ile(beta-Me)-WR-Cys(03H)-T-MeGln-VPN 1718514 :Poststatin:(Val)2-NHCH(Et)COCO-D-Leu-Val 1605493 :Probestin:2-0H-4-Ph-3-Aminobutanoyl-Leu-Pro2 1211482P:Propioxatin:Acyl-Pro-Val0503225P:Protease Inhibitor alpha-MAPI:CO(Phe)-Arg-Val-Phenylalaninal 1614500D:Pseudobactin 589A:Asp,Lys,3-OH D-Asp,D-Ser,Thr D-Ala,D-Glu,Ser,Orn(OH) 1717844D:Pseudobactin:D-Orn(OH),Ala,D-aThr,beta-OH Asp,Lya 1211430D:Pseudobactin:X-Ser-Ala-Gly-Ser-Ala-Y-aThr-Z 1819869 :Puwainaphycin:Cyclo-10-Peptide 1810795 :Pyoverdin:13 Membered Cyclic Peptide 1507670D:Pyoverdin:D-Ser-Arg-D-Ser-0rn(X)-Lys-0rn(X)-Thr2 0912294D:Pyoverdin:D-Ser-Arg-D-Ser-X-Thr2-Lys-X X=0rn(OH)1704446D:Pyoverdin:UKSUTUXTUY U=D-Ser1704630 :Pyoverdin:X-Ala-Lys-Gly2-H0Asp-Y-Ser-Ala-H00rn1505634 :Quinoraycin Like Antibiotic:3-OH Quinaldic Acid N-Me Me-Cyclopropane AA 0704279n:Quinoxaline Cyclodepsipeptide:1812415 :RA XI/XII/XIII/XIV:Antitumor Bicyclic 6-Peptide 1211454 :RA-VI/III/II/I:Cyclo-6-Peptide 1804515 :RA1-III/VI:Bicyclic 6-Peptide1505636 :Ramoplanin:3-0H Asp,D,aThr3,G,A,L,F Gly(Ph(Cl,OH),(Gly(Ph(OH)))5 0rn2 1801513 :Ramoplanose:A,D,OH Asp,G,L,Orn2,F,N,Gly(Ph(Cl,OH)),Gly(Ph(OH) )4,aThr 1410560 :Rhizocticin:AA-Arg-APPA APPA=Araino Phosphono Pentenoic Acid 0903222P:Rhizonin:Cyclo(aIle-Val2-X-Leu-MeAla-X)0503208n:Ristocetin:Glycopeptide 0810168n:Ristomycin:Glycopeptide
-157-

1808460?:Ro091679:Fumaryl-Arg-Leu-Arg0510199?:Roseotoxin:Cyclo(Hpt-Pro(Me)-Ile-MeVal-beta-Ala-MeAla1005336 rRoseotoxln:Cyclodepsi-6-Peptide 3-Me Pro,lie,MeVal,beta Ala.MeAla1816438 :Rotihibin:6-Peptide1812408 :Rotihibin:Dbu,Ser,Cit,beta-0H Asn,Asparaginol,aThr1501689 :Rugosanine:For-MeAla,Val,Hyp,Leu as Component1603680 :Sandramycin:Cyclodepsi-10-Peptide1713718n:Sanjoinenine:Cyclopeptide1101618 :Sativanine:13 Membered Cyclopeptide1207453 :Sativanine:13 Membered Cyclopeptide1207478 :Sativanine:13 Membered Cyclopeptide1207479n:Sativanine:N-For Cyclopeptide Alkaloid1809578DrSerrawettin:Cyclo(Hdc-D-Leu-Ser-Thr-D-Phe-Ile)0701240n:Siomycin:0702208n:Siomycin:1309378 :Spider Toxin NSTX:Acyl-Asn-Cadaverine(Arg-Putreanyl)1305203 :Staccopin:Val4-Phe-H,Val4-Tyr-H1011438 :Stenothricin:D-Cys,D-aThr,D/L-Ser,Dpr,Val,Lys,Sar D-DehydroAbu 1405656 :Sulfomycin:Oxazole AA.Thiazole AA,DehydroAla,Thr 1802558n:Surfactin:1106354n:Suzukacillin:Aib-Peptide1607769 :Syringomycin:DehydroThr,Thr(Cl),Asp(Hy)1515556 :Syringomycin:Ser2-Dab2-Arg-Phe-Dhb-Thr(Cl)-X1717623 :Syringopeptin:Dhb,Pro,Val,Ala,aThr,Ser,Dbu,Tyr Dhb=DehydroAbu1810723 :Syringostatin:Cyclo-9-Depsipeptide1705343 :Syringotoxin:Cyclodepsi-9-Peptide Phytotoxin1818627 :Tawicyclamide:Cyclo-8-Peptide Thiazoline x31513586n:Teicoplanin Related Substance:Glycopeptide Antibiotic1302408n:Teicoplanin:1506742n:Teicoplanin:Glycopeptide1701485 :Tentoxin:Cyclo-4-Peptide N-Me DehydroPhe,Gly,MeAla,Leu 1210387 :Tentoxin:Cyclo-4-Peptide0901321 :Tentoxine:Cyclo-4-Peptide N-Me DehydroPhe,Gly,MeAla,Leu1512600 :Thaxtomin:Cyclo(MeTrp-MePhe) Deriv1507564 :Theonellamide:Bicyclic 11-Peptide1210553D:Theonellamine:Me0Ac-Cyclo-13-Peptide D/L-MeAA1307362D:Theonellapeptolide:13-Peptide Lactone D-AA MeAA beta-Ala alle1711811D:Theonellapeptolide:D-MeLeu,beta Ala,D-Leu,Melle D-alle,MeVal,D-Mealle1713573D:Theonellapeptolide:D-MeLeu,D-Leu,beta Ala,D-alle Melle,MeAla,D-Mealle1208453D:Theonellapeptolide:Melle,MeAla,MeVal,D-MeLeu D-Leu,D-alle,D-Mealle0702208n:Thiostrepton:0907225D:Toxin BE4:Cyclo(D-Ala-Leu-X-Ala-betaAA-D-iso-Glu)1711499 :Trialaphos:MP02HCH2CH2CH(NH2)C0-Ala3 0708199 :Trichopolyn:RC0-Pro-X-Ala-Aib2-AA-Ala-Aib2-Tcd1304492 :Trichorzianine:Ac-XA2XYQX3SLXPVXIQ2-Pheol X=Aib,Y=Ive,Pheol=Pheol1611798 :Trichorzianine:Ac-XAAXUQX3SLXPVXIQQ-Pheol X=Aib U=Iva1409564 :Trichosporin:Aib-20-Peptide1310496P:Trypsin Inhibitor:For-His-Val1505423P:Tyrostatin:Isovaleryl-Tyr-Leu-Tyr-H1611628P:UDP-Disaccharide 5-Peptide:Glu-Ala-Lys(gamma Glu)-Ala1404628 :Vancomycin:Me2-Ala-Tyr(3-C1)-Gln-Tyr4 Analog1608276n:Variapeptin:Cyclo Depsipeptide1612608 :Variapeptin:Depsi-6-Peptide1301460 :Verlamelin:Cyclo-Depsi-7-Peptide1301443 :Victorin:Cyclo-5-Peptide0812205 :Virginiamycin:6/7-Peptide Lactone0812205 :Viridogrisein:6/7-Peptide Lactone1816622n:Westiellamide:Cyclic Oxazoline Peptide1812598 :Xanthostatin:Cyclodepsi-6-Peptide
—158 —

f ^ 5 f 3 ^
mm - ma&A ^ -T550 ;APArfi@|X W^SJ 1-8-4
T E L 06(443)5321
FAX 06(443)5319
mmmm




















