
VOLUME IV - UEPA
561
1 ANAIS TRABALHOS COMPLETOS – 2019 VOLUME IV ISSN: 2316-7637
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of VOLUME IV - UEPA

1
ANAIS TRABALHOS COMPLETOS – 2019
VOLUME IV
ISSN: 2316-7637

UNIVERSIDADE DO ESTADO DO PARÁ
RUBENS CARDOSO DA SILVA
Reitor da Universidade do Estado do Pará
CLAY ANDERSON NUNES CHAGAS
Vice-Reitor da Universidade do Estado do Pará
RENATO DA COSTA TEIXEIRA
Pró-Reitora de Pesquisa e Pós-Graduação
CARLOS JOSÉ CAPELA BISPO
Pró-Reitor de Gestão e Planejamento
ALBA LÚCIA RIBEIRO RAITHY PEREIRA
Pró-Reitora de Extensão
ANA DA CONCEIÇÃO OLIVEIRA
Pró-Reitora de Graduação
ELIANE DE CASTRO COUTINHO
Diretora do Centro de Ciências Naturais e Tecnologia
ALTEM NASCIMENTO PONTES
Coordenador do Programa de Pós-Graduação em Ciências Ambientais

REALIZAÇÃO
Programa de Pós-Graduação em Ciências Ambientais
Centro de Ciências Naturais e Tecnologia
Universidade do Estado do Pará
COORDENAÇÃO DO SIMPÓSIO
Prof. Dr. Altem Nascimento Pontes

COMISSÃO ORGANIZADORA DOCENTE
Prof. Dr. Altem Nascimento Pontes (Coordenador) – UEPA
Profa. Dra. Ana Lúcia Nunes Gutjahr (Vice – Coordenadora) – UEPA
Profa. Dra. Jéssica Herzog Viana – UEPA
COMISSÃO ORGANIZADORA DISCENTE
Alcione Pinheiro de Oliveira
Ana Beatriz Neves da Silva
Ana Flávia Santos de Brito
Danielle Nazaré Salgado Mamede Pantoja
Emile Lebrego Cardoso
Enilde Santos de Aguiar
Gysele Maria Morais Costa
Isabela Rodrigues Santos
Jaqueline Fontel de Queiroz
Lucila Pereira Da Silva
Manuella Almeida Raiol da Silva
Marcelo Coelho Simões
Marcos Felipe Bentes Cansanção Pereira
Marcus Vitor Almeida Campos
Marinele Maria Saraiva Rodrigues
Mayara Gomes da Silva
Mônica Moraes Ribeiro
Ronaldo Darlan Gaspar Aquino
Thamires Beatriz dos Santos Caitano
Wilson Figueiredo de Lima
Yuri Freitas da Silva

COMISSÃO CIENTÍFICA DOCENTE
Prof. Dr. Alfredo Kingo Oyama Homma – UEPA
Prof. Dr. Alisson Rangel Albuquerque – UEPA
Prof. Dr. Altem Nascimento Pontes – UEPA
Profa. Dra. Ana Cláudia Tavares Martins – UEPA
Prof. Me. Antônio Bisconsin Junior – IFRO
Prof. Dr. Argemiro Midones Bastos – IFAP
Prof. Dr. Carlos Elias de Souza Braga – UEPA
Profa. Dra. Carmelita de Fátima Amaral Ribeiro – UEPA
Profa. Ma. Claudeth de Souza Pinto – SEDUC-PA
Profa. Dra. Cláudia Viana Urbinati – UEPA
Prof. Dr. Edmir dos Santos Jesus – UEPA
Profa. Dra. Fernanda da Silva Mendes – UEPA
Profa. Dra. Flávia Cristina Araújo Lucas – UEPA
Profa. Dra. Hebe Morganne Campos Ribeiro – UEPA
Prof. Dr. Hélio Raymundo Ferreira Filho – UEPA
Prof. Dr. Jefferson Dos Santos Marcondes – IFPA
Profa. Dra. Jéssica Herzog Viana – UEPA
Prof. Dr. José Augusto Carvalho de Araújo – UEPA
Prof. Dr. José Moacir Ferreira Ribeiro – UEPA
Profa. Dra. Leila de Fátima Oliveira de Jesus Robert – UEPA
Profa. Dra. Luiza Helena da Silva Martins – UFRA
Prof. Dr. Madson Alan Rocha de Sousa – UEPA
Prof. Dr. Marcelo José Raiol Souza – UEPA
Prof. Dr. Marcos Adami – INPE
Prof. Dr. Marcos Augusto Eger da Cunha – UEPA
Profa. Dra. Norma Ely Beltrão – UEPA

Prof. Dr. Pablo Luis Baia Figueiredo – UEPA
Prof. Dr. Rafael de Paiva Salomão – MPEG
Prof. Dr. Raimundo Sérgio de Farias Junior – UEPA
Prof. Me. Renan Coelho de Vasconcellos – IFPA
Profa. Dra. Veracilda Ribeiro Alves – IEC
Prof. Dr. Werner Damião Morhy Terrazas – UEPA
COMISSÃO CIENTÍFICA DISCENTE
Alcione Pinheiro de Oliveira
Emile Lebrego Cardoso
Evelyn Lebrego Cardoso
Enilde Santos de Aguiar
Fernanda Neves Ferreira
Gilson Celso Albuquerque Chagas Junior
Gysele Maria Morais Costa
Isabela Rodrigues Santos
Jaíra Thayse Souza Batista
Jaqueline Fontel de Queiroz
Jefferson Inayan de Oliveira Souto
Lucila Pereira Da Silva
Manuella Almeida Raiol da Silva
Marcelo Coelho Simões
Marcos Felipe Bentes Cansanção Pereira
Marcus Vitor Almeida Campos
Marinele Maria Saraiva Rodrigues
Mayara Gomes da Silva
Mônica Moraes Ribeiro
Paulo Weslem Portal Gomes
Thamires Beatriz dos Santos Caitano
Wilson Figueiredo de Lima
Yuri Freitas da Silva

EDITORAÇÃO
Prof. Dr. Altem Nascimento Pontes
Emile Lebrego Cardoso
Enilde Santos de Aguiar
Isabela Rodrigues Santos
Jaqueline Fontel de Queiroz
Lucila Pereira Da Silva
Manuella Almeida Raiol da Silva
Marcus Vitor Almeida Campos
Marinele Maria Saraiva Rodrigues
Thamires Beatriz dos Santos Caitano
Wilson Figueiredo de Lima
Yuri Freitas da Silva

APRESENTAÇÃO
No período de 11 a 13 de dezembro de 2019, o Programa de Pós-Graduação em
Ciências Ambientais – em nível de mestrado acadêmico – da Universidade do Estado do
Pará (UEPA) promoveu o VIII Simpósio de Estudos e Pesquisas em Ciências Ambientais
na Amazônia, no Auditório do Centro de Ciências Naturais e Tecnologia da UEPA. Nesta
oitava edição, o tema abordado foi: Consciência Ambiental: Desafios Na Relação Ser
Humano-Natureza.
Em 2019, o Simpósio recebeu 676 inscrições. Os inscritos submeterem 203
resumos e 281 trabalhos completos. Após o processo de avaliação, realizado pela
Comissão Científica do Simpósio, foram aceitos 178 resumos e 219 trabalhos completos.
Todos os trabalhos aceitos foram disponibilizados nos Anais do evento e estarão à
disposição dos simposistas no site do Simpósio.
Externo meus agradecimentos pela participação de todos no VIII Simpósio de
Estudos e Pesquisas em Ciências Ambientais na Amazônia, e expresso aqui meu convite
para reencontrá-los na nona edição do evento.
Prof. Dr. Altem Nascimento Pontes Professor e Pesquisador da Universidade do Estado do Pará

Belém (PA), 11 a 13 de dezembro de 2019
SUMÁRIO
ANÁLISE DO MERCADO VAREJISTA DE ORQUÍDEAS NO MUNICÍPIO DE BELÉM (PA) .............13
ACIDEZ ATIVA E POTENCIAL DE UM LATOSSOLO AMARELO SOB INTEGRAÇÃO LAVOURA-
PECUÁRIA NO MUNICÍPIO DE PARAGOMINAS-PA......................................................................23
ANÁLISE DO POTENCIAL PRODUTIVO DO FRUTO DE CUPUAÇU DOS PRODUTORES ASSOCIADOS
A COOPERATIVA AGRÍCOLA MISTA DE TOMÉ-AÇU (CAMTA), PARÁ ...........................................31
AVALIAÇÃO DE CARACTERÍSTICAS SOCIOECONÔMICAS DE COMERCIANTES E DE
ABASTECIMENTO DE HORTALIÇAS EM FEIRAS LIVRES DE BELÉM – PA ......................................42
AVALIAÇÃO FENOLÓGICA DA ESPÉCIE Cissus sicyoides L. ASSOCIADA COM A PRECIPITAÇÃO
PLUVIOMÉTRICA ........................................................................................................................48
CARACTERÍSTICAS SOCIOECONÔMICAS E DE CONSUMO DOS FREQUENTADORES DE FEIRAS
LIVRES DO MUNICÍPIO DE MARABÁ-PA .....................................................................................53
EFEITO DA INOCULAÇÃO DE BACTÉRIAS PGPR EM ARROZ (Oryza sativa L.) ..............................59
EFEITO DO PASTEJO NAS PROPRIEDADES QUÍMICAS DO SOLO NO PROJETO DE
ASSENTAMENTO SÃO FRANCISCO, ELDORADO DOS CARAJÁS, PA .............................................71
O EFEITO DA FREQUÊNCIA REGA COM BIOFERTILIZANTES EM PARÂMETROS BIOMÉTRICOS,
TROCAS GASOSAS E FLUORESCÊNCIA DA CLOROFILA A EM MUDAS DE AÇAIZEIRO ..................79
PLANEJAMENTO SUSTENTÁVEL DO USO GLIFOSATO NA AGRICULTURA ...................................89
PRODUÇÃO HIDROPÔNICA DE ALFACE INTEGRADA À PRODUÇÃO DE PESCADO COMO FORMA
DE TRATAMENTO DO EFLUENTE ..............................................................................................101
QUALIDADE FITOSSANITÁRIA E GERMINATIVA DE SEMENTES DE MILHO SUBMETIDAS A
TERMOTERAPIA ........................................................................................................................109
SELEÇÃO DE ISOLADOS DE Trichoderma sp. PROMOTORES DE CRESCIMENTO DE MUDAS DE
COQUEIRO ................................................................................................................................117
A DENSIDADE DA MADEIRA É RELACIONADA AO TEOR DE EXTRATIVOS? ESTUDO COM QUATRO
ESPÉCIES AMAZÔNICAS ............................................................................................................127
ANÁLISE COMPARATIVA DE FOCOS DE QUEIMADAS ENTRE MUNICÍPIOS DO ESTADO DO PARÁ
.................................................................................................................................................136
AVALIAÇÃO DAS ESTRUTURAS DIAMÉTRICAS EM FLORESTAS EXPLORADAS SEM AUTORIZAÇÃO,
SUBMETIDAS AO LICENCIAMENTO AMBIENTAL PARA PRODUÇÃO MADEIREIRA NO ESTADO DO
PARÁ ........................................................................................................................................146
AVALIAÇÃO DE TEOR DE UMIDADE EM CASTANHA-DE-SAPUCAIA ..........................................156
CARACTERIZAÇÃO DE PROPRIEDADES FÍSICAS E ANATÔMICAS DA MADEIRA DE Qualea albiflora
Warm, MANDIOQUEIRA PARA CONSTRUÇÃO CIVIL .................................................................165
CONJUNTURA DO MERCADO DA FIBRA DE BURITI (Mauritia flexuosa L.) NO ESTADO DO PARÁ
ENTRE OS ANOS DE 1997 A 2017 .............................................................................................172

Belém (PA), 11 a 13 de dezembro de 2019
DINÂMICA DO TRABALHO NA ATIVIDADE EXTRATIVISTA DA Carapa guianensis Aubl.
(ANDIROBA) EM PROJETOS DE ASSENTAMENTOS AGROEXTRATIVISTAS (PAEX) DAS VÁRZEAS
DO MUNICÍPIO DE IGARAPÉ-MIRI, PARÁ ..................................................................................182
DESCRIÇÃO DOS FATORES ORGANIZACIONAIS E AMBIENTAIS DE UMA EMPRESA MADEIREIRA:
ESTUDO DE CASO NO DISTRITO DE ICOARACI, BELÉM/PA .......................................................191
EQUAÇÕES DE CRESCIMENTO EM ALTURA DOMINANTE PARA UM PLANTIO CLONAL DE
Tectona grandis Linn F. ............................................................................................................199
IDENTIFICAÇÃO BOTÂNICA EM PLANOS DE MANEJO FLORESTAL SUSTENTÁVEL: O CASO DO IPÊ
.................................................................................................................................................209
PRODUÇÃO DE BIOMASSA FRESCA ACIMA DO SOLO EM UMA ÁREA DE PMFS LICENCIADO NO
MUNICÍPIO DE SANTARÉM - PA ...............................................................................................219
SERAPILHEIRA COMO UM INDICADOR PARA AVALIAÇÃO DE ECOSSISTEMAS EM RESTAURAÇÃO
APÓS MINERAÇÃO DE BAUXITA NA AMAZÔNIA ORIENTAL .....................................................228
VARIAÇÃO RADIAL DO ASPECTO ANATÔMICO DA FIBRA DA MADEIRA DE CALYCOPHYLLUM
SPRUCEANUM BENTH K. SCHUM .............................................................................................239
A IMPORTÂNCIA AMBIENTAL DO SISTEMA AGROFLORESTAL DE TOMÉ-AÇÚ – SAFTA,
LOCALIZADO NO NORDESTE DO ESTADO DO PARÁ .................................................................251
ANÁLISE COMPARATIVA DO USO DO PROTEINADO E A INCLUSÃO PARCIAL DA TORTA DE
DENDÊ (ELAEIS GUINEENSIS) EM DIETAS PARA RUMINANTES DA RAÇA NELORE: CRESCIMENTO
E OBTENÇÃO DE GANHO DE PESO ...........................................................................................262
AVISTAMENTO DA AVIFAUNA NO MUNICÍPIO DE JURUTI, BAIXO AMAZONAS, ESTADO DO
PARÁ, BRASIL ............................................................................................................................273
CARACTERÍSTICAS FÍSICO-QUÍMICAS E BIOMASSA DO MICROFITOPLÂNCTON EM DOIS
MUNICÍPIOS DO NORDESTE PARAENSE ...................................................................................282
COMPOSIÇÃO DE ROTIFERA NO PARQUE ESTADUAL DO UTINGA (BELÉM, PARÁ, BRASIL)......293
DINÂMICA DA COMUNIDADE ZOOPLANCTÔNICA E SUA RELAÇÃO COM A QUALIDADE DA ÁGUA
NO PORTO DE VILA DO CONDE (PARÁ, BRASIL) .......................................................................303
DISTRIBUIÇÃO ESPACIAL DA REGENERAÇÃO NATURAL EM RELAÇÃO À ÁRVORE MATRIZ DE
Euxylophora paraensis Huber EM FRAGMENTO FLORESTAL NO MUNICÍPIO DE PARAGOMINAS-
PARÁ ........................................................................................................................................314
EFEITOS DA ADIÇÃO DE NUTRIENTES SOBRE O FITOPLÂNCTON DO RESERVATÓRIO DE
ABASTECIMENTO BOLONHA (BELÉM-PARÁ) ............................................................................325
ANÁLISE DA EFICIÊNCIA DA COLETA DE MATERIAL BOTÂNICO PARA A MELHORA DA
IDENTIFICAÇÃO CIENTÍFICA DAS ESPÉCIES ARBÓREAS NA FLONA DO TAPAJÓS ......................335
LEVANTAMENTO ETNOBOTÂNICO DE PLANTAS MEDICINAIS UTILIZADAS NA PASTORAL DA
SAÚDE NO MUNICÍPIO DE MOJU-PA ........................................................................................342
LEVANTAMENTO E IDENTIFICAÇÃO DE ESPÉCIES COMESTÍVEIS E ORNAMENTAIS COM
POTENCIAL TÓXICO NO MUNICÍPIO DE SALVATERRA-PARÁ ....................................................352

Belém (PA), 11 a 13 de dezembro de 2019
PERFIL QUÍMICO E ATIVIDADE ANTIOXIDANTE DA PLANTA VERÔNICA (Dalbergia monetaria L.)
COLETADA NO ARQUIPÉLAGO DO MARAJÓ - PA ......................................................................363
PROBLEMÁTICA NA ASSOCIAÇÃO DE NOMES VERNACULARES E CIENTÍFICOS: O CASO DO CAJU
.................................................................................................................................................372
MAPEAMENTO E LEVANTAMENTO DAS COLEÇÕES DE ESPÉCIES ORNAMENTAIS DO HERBÁRIO
FELISBERTO CAMARGO REGISTRADAS PELO BRAHMS .............................................................379
COMPORTAMENTO ALIMENTAR DE ESTRELAS-DO-MAR EM AMBIENTE RECIFAL CONTROLADO
.................................................................................................................................................389
LEVANTAMENTO BIBLIOGRÁFICO SOBRE A MARRECA-CABOCLA Dendrocygna autumnalis
(ANSERIFORMES: ANATIDAE): TEMAS ESTUDADOS E HISTÓRICO DOS TRABALHOS ................399
PRIMEIRO REGISTRO DO MOLUSCO Polymesoda caroliniana (BOSC, 1801) (BIVALVIA:
CYRENIDAE) NO LITORAL BRASILEIRO ......................................................................................410
CARACTERIZAÇÃO MORFOLÓGICA DAS CEPAS DE Microcystis LBARP-1 e Microcystis LBARP-2
ISOLADAS DO RIO PARÁ (BARCARENA, PARÁ). .........................................................................419
CIANOBACTÉRIAS COMO INDICADORAS DA QUALIDADE DAS ÁGUAS USADAS PARA O
ABASTECIMENTO DE COMUNIDADES TRADICIONAIS NA RESERVA BIOLÓGICA RIO TROMBETAS
(PARÁ) ......................................................................................................................................428
COMPOSIÇÃO FLORÍSTICA E FITOSSOCIOLOGIA ESPACIAL DE COMUNIDADES VEGETAIS EM UM
AMBIENTE RIPÁRIO ANTROPIZADO NO NORDESTE DO ESTADO DO PARÁ, BRASIL .................440
DESEMPENHO DE MACRÓFITAS AQUÁTICAS SOBRE INTERFERÊNCIA DE ESPÉCIES NATIVAS E
INVASORAS...............................................................................................................................451
EFEITOS DO MANEJO DE IMPACTO REDUZIDO SOBRE A ESTRUTURA HORIZONTAL E
DIVERSIDADE FLORÍSTICA DA ÁREA DE CONCESSÃO FLORESTAL MAMURU-ARAPIUNS, OESTE
PARAENSE ................................................................................................................................460
FENODINÂMICA DA POPULAÇÃO DE Euxylophora paraensis Huber EM FRAGMENTO FLORESTAL
NO MUNICÍPIO DE PARAGOMINAS-PARÁ ................................................................................469
FITOSSOCIOLOGIA E CARACTERIZAÇÃO DE ESPÉCIES EM ÁREA DE VEGETAÇÃO SECUNDÁRIA NO
ECOSSISTEMA DE CAPOEIRA DO MUNICÍPIO DE BRAGANÇA (PA) ...........................................480
IMPACTOS NA BIODIVERSIDADE COM OS FOCOS DE QUEIMADAS NA AMAZÔNIA PARAENSE490
ÍNDICES DE EMERGÊNCIA DE PLÂNTULAS DE ACMELLA OLERACEA (L.) R.K. JANSEN E ACMELLA
CILIATA (KUNTH) CASS .............................................................................................................502
REGISTROS REPRODUTIVOS DA MARRECA, DENDROCYGNA AUTUMNALIS, NO ENTORNO DA
RESEX MARINHA DE TRACUATEUA-PA .....................................................................................511
INFLUÊNCIA CLIMÁTICA NA FLUTUAÇÃO SAZONAL DE INSETOS CERATOPOGONÍDEOS
CAPTURADOS NA ÁREA DE PROTEÇÃO AMBIENTAL ILHA DO COMBU, BELÉM, PARÁ .............521
DEPOSIÇÃO DE SERAPILHEIRA E NUTRIENTES EM ÁREA DE RESTAURAÇÃO APÓS MINERAÇÃO
DE BAUXITA NA AMAZÔNIA ORIENTAL ....................................................................................533
PROMOÇÃO DE CRESCIMENTO DE MUDAS DE JAMBÚ INOCULADAS COM TECNOLOGIA
MICROBIANA ............................................................................................................................541

Belém (PA), 11 a 13 de dezembro de 2019
USO DE MICRORGANISMOS PROMOTORES DE CRESCIMENTO ASSOCIADOS À ADUBAÇÂO
MINERAL EM UROCHLOA BRIZANTHA ......................................................................................550

13
Belém (PA), 11 a 13 de dezembro de 2019
ANÁLISE DO MERCADO VAREJISTA DE ORQUÍDEAS NO MUNICÍPIO DE
BELÉM (PA)
¹Maria Vitória Queiroz Almeida1; Ana Caroline Palheta De Farias²; Tainara Miranda Da
Silva³; Eliude Navegantes Do Carmo4; Fabrício Khoury Rebello5.
1 Graduanda em engenharia florestal/UFRA. [email protected]
²Graduanda em agronomia/UFRA. [email protected]
³Engenheira Agrônoma/UFRA. [email protected]
4Engenheira Agrônoma/UFRA. [email protected] 5Doutor em ciências agrárias/UFRA. [email protected]
RESUMO
O Brasil é reconhecido mundialmente pela riqueza de sua biodiversidade, em que se
destacam as plantas ornamentais e as flores, como orquídeas e as bromeliáceas, sendo a
floricultura considerada uma atividade econômica de grande relevância no agronegócio
nacional e internacional. Dentro dessa produção, destacam-se as orquídeas, pois são
muito requeridas no mercado de plantas ornamentais. Com isso, o presente estudo teve
como objetivo analisar o mercado varejista de orquídeas no município de Belém (PA)
com o intuito de analisar sua importância no mercado das plantas ornamentais. Para
coletar os dados foram aplicados questionários online e presenciais, em 16
estabelecimentos e divididos em três partes: caracterização do estabelecimento comercial,
identificar os aspectos do mercado varejista e identificar as espécies comercializadas.
Utilizou-se a estatística descritiva para avaliar os dados obtidos. A orquídea Phalaenopsis
envasada(?) está presente em 81,25% dos estabelecimentos e é, também, a mais
comercializada. Dos entrevistados que comercializam orquídeas, 46,15% consideram que
a venda aumentou nos últimos três anos. A pesquisa revela que existem muitos pontos
fortes e potencialidades para expansão do mercado de orquídeas no município de Belém,
porém há grandes entraves a serem superados para que este mercado venha a se
desenvolver de forma significativa na economia paraense.
Palavras-chave: Comércio de orquídeas. Pará. Plantas Ornamentais.
Área de Interesse do Simpósio: Agronomia

14
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Atualmente, a floricultura é considerada uma atividade econômica de grande
relevância no agronegócio nacional e internacional, devido, principalmente, à criação de
um elevado número de empregos diretos e indiretos e, também, pelo valor de sua
produção e comercialização (CANÇADO JÚNIOR et al., 2005).
O agronegócio de flores e plantas ornamentais vem se expandindo no Brasil e com
isso ganhado notoriedade na agricultura nacional. Isso pode ser constatado a partir dos
dados do Instituto Brasileiro de Floricultura (IBRAFLOR) o qual mostra que o comércio
de flores obteve um faturamento de R$ 5,7 bilhões, em 2014, e de R$ 6 bilhões, em 2015,
sendo que a expectativa para o ano de 2016 que era o crescimento de 8% para o ano todo,
já havia sido atingida no primeiro semestre (ALENCAR, 2016).
A família Orchidaceae é uma das maiores famílias botânicas do mundo, com
aproximadamente 20.000 espécies, das quais 2.462 ocorrem no Brasil; no Pará registram-
se 421 espécies (AFONSO; KOCH; COSTA, 2016). As orquídeas estão entre as dez
plantas ornamentais mais comercializadas no mercado internacional e são consideradas
as flores de negociação do século XXI (PAKYONG, 2013). Segundo Suzuki (2014), o
comércio exterior das orquídeas aumenta a cada ano desde 2004, quando o mesmo saltou
de US$ 376 mil para mais de US$ 10,8 milhões, em 2013.
Segundo Suzuki (2014) o mercado interno de orquídeas gera grande circulação
monetária, entretanto, não há informações disponíveis sobre a movimentação financeira
decorrente dessa cadeia produtiva. O mesmo autor afirma que em discussão com
colecionadores e membros de associações orquidófilas, foi possível determinar que a
comercialização de orquídeas, realizada somente nas exposições no Brasil, movimenta
anualmente mais de R$ 10 milhões.
O presente estudo teve como objetivo analisar o mercado varejista de orquídeas
no município de Belém (PA).
2. MATERIAL E MÉTODOS
O estudo foi realizado no município de Belém, capital do estado do Pará. Segundo
dados do Instituto Brasileiro de Geografia e Estatística (IBGE, 2016), Belém destaca-se
por possuir o maior centro populacional do estado do Pará, estimado em 1.446.042

15
Belém (PA), 11 a 13 de dezembro de 2019
habitantes, o que mostra a sua representatividade para a avaliação da situação do mercado
varejista de flores e plantas ornamentais na economia paraense.
O trabalho se enquadra na pesquisa de natureza aplicada, pois objetiva gerar
conhecimentos de finalidade prática, ou seja, dirigidos à solução de problemas específicos
(GIL, 2008; SILVA, 2005). No caso, o motivo inicial de sua elaboração foi o
questionamento sobre o panorama atual do mercado de flores e plantas ornamentais no
município de Belém, mais especificamente em relação às espécies da família
Orchidaceae. O procedimento técnico utilizado foi o levantamento de campo, já que
envolveu, para sua produção, a interrogação direta das pessoas cujo comportamento se
deseja ter conhecimento. Procede-se à solicitação de informações a um grupo
significativo de pessoas acerca do problema estudado para em seguida, mediante análise
quantitativa, obter as conclusões correspondentes dos dados coletados (GIL, 2008;
SILVA, 2005).
O estudo englobou as floriculturas situadas no município de Belém, não incluindo
os autosserviços (como supermercado) ou outra forma de comercialização. Para obter os
resultados, realizou-se uma busca online, em sites de divulgação, com vista a ampliar o
rol de empresas a serem pesquisadas. Na sequência, efetuou-se o levantamento de campo,
a partir da aplicação de questionários e entrevistas junto às empresas em atividades do
ramo e com pessoas chaves.
Para coletar os dados foram aplicados questionários no período de 17 a 24 de
fevereiro de 2017, em um total de 16 estabelecimentos procurando identificar os aspectos
do mercado varejista e as espécies comercializadas. Desta forma, utilizaram-se das
técnicas de analise tabular e de estatística descritiva para interpretação dos dados e
resultados, respectivamente. A tabulação dos dados foi realizada utilizando software
Microsoft Excel 2013.
As informações obtidas com os atores-chave e perguntas de opinião do
questionário foram utilizadas para gerar a estrutura da Matriz Swot (strengths,
weaknesses, opportunities e threats), que em português é conhecida como Matriz Fofa
(Forças, Oportunidades, Fraquezas e Ameaças).

16
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
Dos 13 estabelecimentos que comercializam alguma espécie de orquídea, 53,84%
comercializam duas ou mais espécies, sendo que os demais 46,15% comercializam
unicamente Phalaenopsis. Este é o gênero mais comercializado no mundo e um dos mais
conhecidos da família (PUTINATTI, 2015). Dentre os 81,25% dos estabelecimentos que
comercializam alguma espécie de orquídeas, todos ofertam a espécie denominada
phalaenopsis (Phalaenopsis sp.); 53,84% comercializam denphal (Dendobrium sp.);
46,15% dos estabelecimentos comercializam as espécies conhecidas como pingo-de-ouro
ou chuva-de-ouro (Oncidium sp.); 23,07% comercializam a arundina (Arundina
bambusifolia Lindl.); e apenas 7,79% das empresas comercializam espécies como
orquídea chocolate ou Cattleya, conforme destacado na Figura 1.
Figura 1. Principais espécies de orquídeas comercializadas no município de Belém, 2017.
Fonte: Autores, 2017.
Da amostra estudada, 40,00% comercializavam hastes florais de orquídeas. Dentre
as espécies citadas com finalidade de haste destacam-se a phalaenopsis (Phalaenopsis
sp.) e denphal (Dendobrium sp.), sendo a última de significativa importância para a
comunidade Barreirão, no município de Castanhal, localizado na Região Metropolitana
de Belém (RMB), conforme destacado pela pesquisa de Arévalo, Ponte e Silva (2016).
O preço de compra para x vasos variou de R$11,58 (Arundina) a R$42,11
(Phalaenopsis), média de R$23,67; já para a venda foi R$21,66 e R$82,08, média de
R$43,75, respectivamente para as mesmas espécies. Percentualmente a margem de

17
Belém (PA), 11 a 13 de dezembro de 2019
comercialização de varejo variou de 37,50% (orquídea chocolate) a 48,69%
(Phalaenopsis), média de 45,54% (Tabela 1).
Tabela 1. Preços médios de compra e venda de orquídeas nos 13 estabelecimentos pesquisados.
Espécies Compra
(R$) Venda
(R$) Margem de comercialização do varejo (%)
Arundina 11,58 21,66 46,53
Cattleya 18 35 48,57
Chuva-de-ouro 20,83 39,58 47,37
Denphal 24,5 44,2 44,57 Orquídea
chocolate 25 40 37,5
Phalaenopsis 42,11 82,08 48,69
Fonte: Autores, 2017.
Quanto aos principais fornecedores de orquídeas, dos 13 estabelecimentos que a
comercializam, 61,53% citaram que obtém este produto de apenas um fornecedor,
23,07% citaram dois fornecedores e apenas 7,69% citou três ou mais fornecedores. Foram
identificados 11 fornecedores diferentes, sendo que destes, 54,54% são oriundos de São
Paulo, 36,36% são da própria região do estado do Pará, e apenas 9,09% obtém produtos
do Ceará.
Foi observado, em todos os estabelecimentos, a inexistência de funcionários com
conhecimento técnico específico, 61,5% das empresas afirmaram que a maioria dos
clientes realiza a compra de forma planejada para alguma ocasião especial, por exemplo:
aniversários, casamentos, círio, natal e dia dos namorados; em 92,3% dos
estabelecimentos não é informado nenhum site ou plataforma digital, a receita
proveniente das vendas de orquídeas. Quanto as perdas na comercialização a média,
considerando todos os estabelecimentos, foi de 8,66% do volume (vasos) adquirido.
3.1 PERSPECTIVA DO MERCADO DE ORQUÍDEA NO ESTADO DO PARÁ
Com o objetivo de analisar a situação do mercado de orquídeas no estado, foi
desenvolvida a Matriz Swot, a qual é apresentada na Tabela 2.

18
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 2. Matriz Swot do mercado de orquídeas no estado do Pará, 2017.
Pontos Fortes (Forças) Pontos Fracos (Fraquezas)
AMBIENTE
INTERNO 1. Proporciona embelezamento
de ambientes externos e internos;
2. São associadas à beleza
exótica e sofisticação;
3. Espécies com ciclo de vida
perene.
1. É considerada uma planta de difícil
cultivo;
2. Dados pouco precisos sobre as
preferências dos consumidores;
3. É uma planta com preço
relativamente elevado;
4. Poucas informações sobre cultivo
doméstico na região Norte. AMBIENTE
EXTERNO 1. Crescimento no mercado
paisagístico;
2. Incentivo a pesquisa sobre
espécies nativas;
3. Variedades comerciais
adaptadas ao clima regional.
1. Crise econômica nacional;
2. Baixa renda per capita da
população paraense;
3. Baixa cultura de consumo de flores
e plantas ornamentais na região;
4. Baixo número de centros locais de
referência para divulgação de
informações.
Oportunidades Ameaças
Fonte: Autores, 2017.
Uma das principais características das orquídeas é o embelezamento que
proporcionam a ambientes interiores e exteriores, por isso, este fator torna-se um dos
pontos fortes desse mercado, pois, a família Orchidaceae possui uma gama de exemplares
que podem ser utilizadas para a decoração. Segundo Rocha et al. (2012) boa parte das
pessoas adquirem esta espécie pela beleza e por paixão, porém muitas vezes a falta do
conhecimento adequado de manejo e exigências agronômicas, ocasiona, a perda do
exemplar, e isso pode vir a causar frustração. Esse aspecto abre uma oportunidade
interessante para a oferta de treinamento específicos nessa área, para colecionadores de
orquídeas, ou de se anexar um folheto para aqueles que possuem poucos espécimes e tem
pouco conhecimento do manejo adequado para a boa sobrevivência da planta que
adquiriu.
Uma questão importante para o desenvolvimento desse mercado é o conhecimento
sobre o público alvo o produto, no caso as orquídeas, notavelmente são poucos os dados
confiáveis na nossa região principalmente, sobre produção e comercialização, como tipos
de substratos, defensivos adubação o que acarreta dificuldades para fortalecimento ou
inserção de novos agentes nessa atividade, pois, para se ter êxito, é necessário conhecer
de forma precisa a gama de clientes para quem será ofertada a produção.

19
Belém (PA), 11 a 13 de dezembro de 2019
Outro ponto considerado como fraqueza é o preço de comercialização
relativamente elevado das orquídeas com relação a outras plantas, como kalanchoe, por
exemplo, que chegam a custar de R$ 3,00 a 4,00. É necessário a agregar valor ao produto,
para que esta espécie possa elevar seu nível de comercialização ou até se aumentar a
produção, para que haja a diminuição dos preços, tornando-se mais acessível para outra
parcela da pulação.
Segundo Mattiuz, Rodrigues e Mattiuz (2006) muitas espécies ocorrem em
florestas úmidas de regiões tropicais e subtropicais, e devido às condições edafoclimáticas
presentes no estado do Pará, este poderia ser considerado um local com boa aptidão para
o desenvolvimento do cultivo de orquidáceas. É notável a necessidade de mais estudos
das espécies da família endêmicas da Amazônia, sendo que estes beneficiam a
preservação, já que amplia o entendimento sobre a flora local, assim como apresentam ao
mercado uma gama nova de espécies e possibilidades de desenvolvimento inclusive de
híbridos.
Com relação às oportunidades que são indicadas para esse mercado, tem-se o
potencial para crescimento do mercado paisagístico, pois, segundo Mello e Pantoja
(2014) devido ao grande número de gêneros, as orquídeas possibilitam uma diversidade
de opções no paisagismo urbano e de interiores. Dessa forma, a família Orchidaceae
também pode contribuir de forma significativa para estes segmentos, pois, observa-se que
existem várias espécies com potencial para serem utilizadas.
Ainda que a Phalaenopsis seja o gênero de grande comercialização, é notável que
a mudança de temperatura até a chegada em Belém afete a sua durabilidade, seja por ter
sido cultivada em uma região diferente ou no transporte. O transporte se dá
principalmente por via aérea, em bandejas (quando o transporte é de mudas) e embaladas
em caixas ou (em exemplares adultos) o espécime sem substrato. É interessante divulgar
e incentivar o consumo sobre diferentes espécies, principalmente as regionais, a fim de
reduzir o preço e disponibilizar produtos diferenciados, com qualidade e durabilidade.
Quanto às ameaças, o fato das flores e plantas ornamentais não se enquadrarem
em produtos de primeira necessidade, pode ser um limitante de alta relevância na
expansão desse mercado como um todo, sendo ainda mais representativo para o mercado
de orquídea, pois algumas espécies apresentam custo elevado para o consumidor final,
levando-se em consideração que o preço é um fator de importância para o mesmo.

20
Belém (PA), 11 a 13 de dezembro de 2019
Ainda quanto às ameaças, tem-se um fator bastante expressivo, mas que não se
restringe unicamente ao comércio de orquídeas, que é a baixa cultura do consumo de
flores e plantas ornamentais na região, ainda que não se possa afirmar isso a partir de
dados de mercado. A colocação de Reis (2011) ao citar a ausência do comportamento
semelhante a cultura europeia de se levar flores quando se vai jantar na casa de alguém,
deixa claro a importância do fator cultural para elevação do consumo e cultivo.
Ainda existe dificuldades quanto ao manejo e cultivo no ambiente doméstico,
apesar de haver bastante informações na internet, porém poucas no estado do Pará, sendo
necessário, portanto, que a oferta de cursos, palestras, feiras e eventos pelas
universidades, orquidários públicos e associações afins para ampliar o público com
afinidade por orquídeas, inclusive com ações para que os indivíduos possam conhecer
mais sobre essas espécies.
4. CONCLUSÃO
A espécie Phalaenopsis é a mais comercializada.
O comércio de orquídeas é fraco, em relação ao restante do país, principalmente
pela baixa cultura do consumo de flores e plantas ornamentais na região.
É importante o incentivo o comércio de orquídeas regionais e disponibilizar
informação de manejo.

21
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
AFONSO, E. A. L.; KOCH, A. K.; COSTA, J. M. Flora preliminar de Orchidaceae no
município de Abaetetuba, Pará, Brasil. Biota Amazônia, v. 6, n. 1, p. 107-118, 2016.
ALENCAR, B.; GALERA, V. Mercado de flores atinge faturamento esperado para
este ano, 2016. Disponível em: <http://revistagloborural.globo.com/Noticias/Agricultura
/noticia /2016/06/mercado-de-flores-atinge-expectativa-de-faturamento-para-o-
ano.html>. Acesso em: 15 nov. 2016.
ARÉVALO, M. R.; PONTE, M. X.; SILVA, D. M. A floricultura tropical paraense:
fatores de inovação na produção e comercialização da orquídea de corte. Revista
Interdisciplinar Científica Aplicada, Blumenau, v.10, n.3, p.01-21, 2016.
CANÇADO J.; LOPES F.; PAIVA, B. M.; ESTANISLAU, M. L. L. Perspectivas para
exportação de flores e plantas ornamentais. Informe Agropecuário, v. 26, n. 227, p. 96-
102, 2005.
GIL, A. C. Métodos e técnicas de pesquisa social. 6ª ed. - São Paulo: Atlas, 2008.
INSTITUTO BRASILEIRO DE FLORICULTURA. O ibraflor. Disponível em:
<http://www.ibraflor.com/oibraflor.php>. Acesso em: 5 mar. 2017.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA, Renda domiciliar per
capita 2016, 2017. Disponível em: <ftp://ftp.ibge.gov.br/Trabalho_e_
Rendimento/Pesquisa_Nacional_por_Amostra_de_Domicilios_continua/Renda_domicil
iar_per_capita/Renda_domiciliar_per_capita_2016.pdf>. Acesso em: 18 mar. 2017.
MATTIUZ, C. F. M.; RODRGUES, T. J. D.; MATTIUS, B. Aspectos fisiológicos de
orquídeas cortadas. Revista Brasileira de Horticultura Ornamental. v. 12, n. 1, p. 21
- 30, 2006.
PAKYONG, S. Abstracts cum Souvenir of National Dialogue on Orchid
Conservation & Sustainable Development for Community Livelihood, March 8-9,
2013.
PUTINATTI, R. gêneros: Phalaenopsis. In. Guia como cultivar orquídeas para
iniciantes, São Paulo, v.1, p. 75, 2015.
REIS, J. N. P. Cultivo de orquídeas: uma opção à agricultura familiar? In: ENCONTRO
NACIONAL DA ECONOMIA ECOLÓGICA ,9, Brasília - DF, 4-8 out, 2011.
ROCHA, L. L.; BARROS, I. S; TELES, H. F.; AGUIAR, L. C. O.; ARRAUJO, C. L. P.;
ADORNO, B. V.; JORGE, L. B.; NUNES, M.; GANGA R. M. D. Capacitação em
cultivo de orquídeas e bromélias. UNIVERSIDADE FEDERAL DE GOIAS - UFG,
2012.
RODRIGUES, V. T. Orchidaceae Juss. Aspectos morfológicos e taxonômicos. Instituto
de Botânica de São Paulo, 19 p., 2011.

22
Belém (PA), 11 a 13 de dezembro de 2019
STANCATO, G. C.; BERMELMANS, P. F.; VEGRO, C. L. R. Produção de mudas de
orquídeas a partir de sementes in vitro e sua viabilidade econômica: estudo de caso.
Revista Brasileira de Horticultura Ornamental, Campinas, v. 7, n. 1, p. 25- 33, 2001.
SUZUKI, R. M. Breve análise sobre o comércio exterior de orquídeas no Brasil.
REUNIÃO ANUAL DO INSTITUTO DE BOTÂNICA, 21. Instituto de botânica, São
Paulo, 2014.

23
Belém (PA), 11 a 13 de dezembro de 2019
ACIDEZ ATIVA E POTENCIAL DE UM LATOSSOLO AMARELO SOB
INTEGRAÇÃO LAVOURA-PECUÁRIA NO MUNICÍPIO DE PARAGOMINAS-
PA
Jamil Chaar El-Husny1; Letícia Cunha da Hungria2; Claudio José Reis de Carvalho3;
Cleo Marcelo de Araújo Souza4; Eduardo Jorge Maklouf Carvalho5
1 Doutor em Ciências Agrárias. Embrapa Amazônia Oriental. [email protected]
2 Doutoranda em Agronomia. Universidade Federal Rural da Amazônia.
3 PhD em Ecofisiologia Vegetal. Embrapa Amazônia Oriental.
4 Mestre em Agronomia. Embrapa Amazônia Oriental. [email protected]
5 Doutor em Solos e Nutrição de Plantas. Embrapa Amazônia Oriental.
RESUMO
As alterações ou desequilíbrios provocados pelos impactos de diferentes formas de uso
da terra e sistemas de produção à qualidade do solo demandam ações e o uso de
indicadores que permitam identificar a dimensão das modificações causadas por estes
fatores. Embora os indicadores isoladamente nos estudos sobre qualidade do solo não
sejam completamente elucidativos diante da complexidade do sistema edáfico, a acidez
ativa (pH) e potencial (H+Al) costumam ser amplamente estudadas em função de suas
influências nas propriedades químicas do solo. Neste sentido, o objetivo do estudo foi
avaliar as condições de acidez ativa e potencial de um Latossolo Amarelo sob integração
Lavoura-Pecuária em duas épocas de coleta de solo: abril (período chuvoso) e novembro
(final do período seco) no município de Paragominas-PA. Para isso, foram amostrados
solos nas profundidades 0-5, 5-10, 10-20 e 20-30 cm. O delineamento experimental
utilizado foi inteiramente casualizado, em fatorial 4x2, com quatro repetições. O pH em
água das amostras de solo foi obtido pelo método potenciométrico, enquanto a acidez
potencial (H+Al) foi determinada com extrator acetato de cálcio 0,5 M. Nas áreas sob
pastagem foram registrados pH’s menos ácidos quando comparados a solos sob mata e
plantio de milho, que não diferiram entre si. No período chuvoso (4,11 cmolc dm-3), a
acidez potencial do solo foi superior à encontrada no final do período seco (3,64 cmolc
dm-3). Quando comparados os sistemas, em solos sob plantio de milho foi observada maior
acidez potencial em todas as camadas estudadas, exceto na profundidade 20-30 cm, na
qual não houve diferença entre os sistemas. As pastagens pareceram ter capacidade
natural em elevar o valor do pH (acidez ativa) dos solos.
Palavras-chave: ILP. Panicum maximun. pH.
Área de Interesse do Simpósio: Agronomia

24
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O uso dos solos pela incorporação aos processos produtivos na agricultura,
pecuária ou silvicultura, naturalmente interfere no equilíbrio e na dinâmica de qualquer
ecossistema (GODOI, 2001). As alterações ou desequilíbrios provocados pelos impactos
de diferentes formas de uso da terra e sistemas de produção à qualidade do solo
demandam ações e o uso de indicadores que permitam identificar a dimensão das
modificações causadas pelo uso da terra e sistemas de produção adotados. Dessa maneira,
a avaliação da qualidade do solo como medida componente da sustentabilidade das
atividades agropecuárias é um indicador fundamental, devendo ser necessariamente
considerada.
Embora índices e características considerados como indicadores nos estudos sobre
qualidade do solo não sejam completamente elucidativos diante da complexidade do
sistema edáfico (MOREIRA & SIQUEIRA, 2006), a acidez ativa (pH) do solo costuma
ser amplamente utilizada como indicadora, em função da sua influência sobre a
produtividade dos cultivos e utilização para recomendação de calagem (CAIRES, 2002).
Em adição, a acidez potencial (H+Al) é comumente incluída em análises de
fertilidade, em função de sua influência nas propriedades químicas do solo, com destaque
para a capacidade de troca catiônica (CTC) (ESCOSTEGUY e BISSANI, 1999; GAMA;
PROCHNOW; GAMA, 2002).
Neste contexto, objetivou-se avaliar as condições de acidez ativa e potencial de
um Latossolo Amarelo sob integração Lavoura-Pecuária em duas épocas de coleta de
solo: período chuvoso e final do período seco, no município de Paragominas-PA
2. METODOLOGIA
O estudo foi realizado nas fazendas Nova Paz e Vitória, localizadas no município
de Paragominas (02º 59’ 45’’ S e 47º 21’ 10’’ O), sudeste do estado do Pará. A fazenda
Vitória adota sistemas de integração lavoura-pecuária para renovação de pastagens,
enquanto na fazenda Nova Paz a atividade é a pecuária de leite em pastejo rotacionado.
A região tem um clima tropical chuvoso, com expressivo período de estiagem (Aw,
conforme classificação de Köppen) e início da estação chuvosa em dezembro,
prolongando-se até maio, com menor disponibilidade hídrica entre junho e novembro. A

25
Belém (PA), 11 a 13 de dezembro de 2019
precipitação média anual (média de 30 anos) é de 1.800 mm e a umidade relativa do ar é
bastante elevada com média anual em torno de 82 % (RODRIGUES et al., 2003).
Para o estudo foram considerados e avaliados quatro sistemas de uso do solo,
sendo três na Fazenda Vitória e um na Fazenda Nova Paz referente ao sistema com
braquiária (Tabela 1).
Tabela 1. Descrição das áreas de estudo, Paragominas, PA.
Área Histórico
Mata
(Reflorestamento
natural)
Área de reflorestamento natural com um tempo, aproximadamente, de
30 anos. O uso desta área teve como objetivo servir de referência na
comparação de alterações possivelmente identificadas nos demais
sistemas avaliados.
Plantio de Milho
Cultivo de milho híbrido Pionner 30F80 em sistema convencional
associado ao plantio de braquiária (Brachiaria ruziziensis).
Pastagem de
Mombaça
Pastagem cultivada com mombaça (Panicum maximum) formada a
partir de uma sucessão de plantios de arroz e milho.
Pastagem de
braquiária
Pastagem de Braquiária (Brachiaria brizantha cv. marandu) com
tempo aproximado de dez anos, com antecedente de derruba e queima
da floresta, sendo utilizada com pecuária leiteira.
Fonte: Os Autores (2019).
Foram coletadas amostras nas profundidades 0-5, 5-10, 10-20 e 20-30 cm, em dois
períodos: 1 - Período chuvoso (abril) e 2 - Final do período seco (novembro). Os solos
em todos os sistemas de uso avaliados foram classificados como Latossolo Amarelo
Distrófico coeso, textura muito argilosa, A moderado, com relevo plano a suavemente
ondulado.
O delineamento experimental foi inteiramente casualizado com quatro repetições,
em esquema fatorial 4x2, sendo quatro os níveis do fator sistemas de uso do solo e dois
os níveis do fator época de coleta de solo. Em cada área de tratamento foram estabelecidas
quatro parcelas, distribuídas aleatoriamente, de 6 m x 12 m (72 m2), as quais
corresponderam às quatro repetições. De cada parcela foram retiradas cinco amostras
simples, a cada profundidade, que formaram uma amostra composta por parcela.
O pH em água foi obtido pelo método potenciométrico, enquanto a acidez
potencial (H+Al) foi determinada com extrator acetato de cálcio 0,5 M, conforme
metodologia recomendada (EMBRAPA, 1997).

26
Belém (PA), 11 a 13 de dezembro de 2019
Em relação ao tratamento dos dados, foram realizadas análises de variância (teste
F) para determinar o efeito dos diferentes sistemas de uso do solo, profundidade e época
de amostragem sobre a acidez ativa e potencial do solo. As comparações entre médias
foram realizadas pelo teste de Scott Knott a 5% de probabilidade com o auxílio do
programa SISVAR versão 5.6. (FERREIRA, 2011).
3. RESULTADOS E DISCUSSÃO
Em relação ao pH, somente houve influência do sistema de uso do solo (p<0,05).
Na tabela 2, são encontrados os valores médios obtidos para a variável de pH em água
sob diferentes sistemas da integração Lavoura-Pecuária estudada.
Tabela 2 – Valores médios de pH em H2O, sob diferentes sistemas de uso do solo. Médias seguidas
pela mesma letra não diferem estatisticamente entre si ao nível de 5% do teste de Scott Knott.
Sistema pH
Mata 5,11 b
Plantio de Milho 5,04 b
Pastagem de mombaça 5,50 a
Pastagem de braquiária 5,61 a
Fonte: Os autores (2019).
Nas áreas sob pastagem foram registrados pH’s menos ácidos quando comparados
a solos sob mata e plantio de milho, que não diferiram entre si. Em um estudo sobre
propriedades químicas de um Latossolo Amarelo cultivado com pastagens de Brachiaria
brizantha e Panicum maximum, durante dois anos, os autores observaram um acréscimo
no pH no segundo ano. Este resultado foi atribuído ao aumento da CTC como reflexo do
aumento de cátions decorrente da mineralização da matéria orgânica, esclarecendo que,
como consequência da elevação da CTC e dos cátions trocáveis, verificou-se aumento da
saturação de bases, causando a elevação do pH e consequente diminuição no alumínio
trocável (PEREIRA; VELOSO; GAMA, 2000).
Em adição, Lopes (1998) e Lopes et al. (2015) esclarecem que solos sob mata ou
floresta tendem a ser mais ácidos, ainda que solos cultivados podem se tornar mais ácidos
na medida em que as colheitas removem as bases.
De fato, Gama-Rodrigues et al. (2008) informaram que a introdução de
leguminosas florestais, por exemplo, promovem decréscimo no pH. Santos et al. (2010),

27
Belém (PA), 11 a 13 de dezembro de 2019
comparando mata com pastagem, observaram resultados semelhantes, com pH um pouco
mais elevado na pastagem.
Em relação à acidez potencial, houve efeito do sistema de uso do solo e da
profundidade isoladamente, bem como da interação destes fatores. Além disso, a época
de amostragem do solo provocou alterações na acidez potencial do solo. Na Figura 1 são
apresentados os resultados referentes à acidez potencial em função da sazonalidade (a) e
da interação sistema x profundidade (b).
Figura 1. Acidez potencial (H+Al) em um Latossolo Amarelo em duas épocas de coleta (a) e
(b) Médias seguidas pela mesma letra minúscula, que comparam entre os sistemas na mesma
profundidade, não diferem entre si e médias seguidas pela mesma letra maiúscula, que
comparam as profundidades dentro do mesmo sistema, não diferem ao nível de 5% do teste de
Scott Knott.
Fonte: Os autores (2019).

28
Belém (PA), 11 a 13 de dezembro de 2019
No período chuvoso (4,11 cmolc dm-3), a acidez potencial do solo foi superior à
encontrada no final do período seco (3,64 cmolc dm-3), independentemente do uso solo ou
camada avaliada.
Entre os sistemas, de forma geral, o cultivo de milho contribuiu para a maior
acidez potencial do solo, seguido de solos sob mombaça e mata, que não diferiram entre
si e apresentaram valores superiores em relação ao solo sob braquiária. Entre as
profundidades, independentemente do sistema ou época de coleta, a acidez potencial foi
da seguinte ordem 0-5cm>5-10cm>10-20cm=20-30cm.
Quando avaliado o comportamento da acidez potencial sob os diferentes sistemas
nas camadas, observa-se que para a maioria das profundidades estudadas o cultivo de
milho conduziu aos maiores valores, exceto na última camada onde não se registraram
alterações em função dos sistemas.
A acidez potencial do solo representa os íons H+ e Al3+ existentes nos colóides do
solo e possui relação com o conteúdo de matéria orgânica (MO), considerando que na
quantidade de cargas negativas nos solos dependentes de pH existe grande contribuição
da MO, principalmente em solos tropicais (CARDOSO, 2011). No presente estudo, a
maior acidez potencial em solos sob cultivo de milho pode estar relacionado não só ao
conteúdo como o material de origem da MO. Segundo Camargos (2005) a maior parte do
hidrogênio da acidez potencial provém das cargas negativas dependentes do pH.
4. CONCLUSÕES
1 – As pastagens pareceram ter capacidade natural em elevar o valor do pH (acidez ativa)
dos solos;
2 - Verificou-se o efeito da sazonalidade para a acidez potencial do solo, sendo essa
mudança destacada no período chuvoso em relação ao final do período seco nas condições
climáticas da região de Paragominas;
3 – O plantio de milho contribuiu para a obtenção de maior acidez potencial do solo.

29
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
CAIRES, E. F. et al. Correção da acidez do solo, crescimento radicular e nutrição do
milho de acordo com a calagem na superfície em sistema plantio direto. Revista
Brasileira de Ciência do Solo, Viçosa, v. 26, n. 4, p.1011-1022, dez. 2002.
CAMARGOS, S. L. Conceitos sobre fertilidade e produtividade. Cuiabá: Universidade
Federal do Mato Grosso; Faculdade de Agronomia e Veterinária; Departamento de Solos
e Engenharia Rural, 2005. 41p.
CARDOSO, E. L. Qualidade química e física do solo sob vegetação arbórea nativa e
pastagens no Pantanal Sul-Mato-Grossense. Revista Brasileira de Ciência do Solo, v.
35, n. 2, p.613-622, 2011.
EMBRAPA. Empresa Brasileira de Pesquisa Agropecuária. Centro Nacional de Pesquisa
de Solos. Manual de métodos de análise de solo. 2. ed. Ver. Rio de Janeiro: Embrapa
Solos, 2011. 230 p.
ESCOSTEGUY, P. A. V.; BISSANI, C. A. Estimativa de H + AL pelo pH SMP em solos
do Rio Grande do Sul e de Santa Catarina. Revista Brasileira de Ciência do Solo, v.23,
n.1, 1999.
FERREIRA, D. F. SISVAR: a computer statistical analysis system. Ciência e
Agrotecnologia, v. 35, n. 6, p. 1039- 1042, 2011.
GAMA, M. A. P.; PROCHNOW, L. I.; GAMA, J. R. N. F. Estimativa da acidez potencial
pelo método SMP em solos ocorrentes no nordeste Paraense. Revista Brasileira de
Ciência do Solo, v. 26, n. 4, p.1093-1097, dez. 2002.
GAMA-RODRIGUES, E.F.; GAMA-RODRIGUES, A.C.; PAULINO, G.M. &
FRANCO, A.A. Atributos químicos e microbianos de solos sob diferentes coberturas
vegetais no norte do Estado do Rio de Janeiro. Revista Brasileira de Ciência do Solo,
32:1521-1530, 2008.
GODOI, L. C. L. Propriedades microbiológicas de solos de áreas degradadas e
recuperadas na região de cerrados goianos. 2001. 87f. Dissertação (Mestrado em
Agronomia). Universidade Federal de Goiás, Goiânia. 2001.
LOPES, A. S. Manual internacional de fertilidade do solo. 2. ed. Piracicaba:
POTAFOS, 1998. 177p.
LOPES, E. L. N., FERNANDES, A. R., TEIXEIRA, R. A., SOUZA, E. S., RUIVO, M.
L. P. Soil attributes under different crop management systems in a Amazon Oxisols.
Bragantia, 35, 1127-1139. 2015.
MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e bioquímica do solo. 2. ed.
Lavras: UFLA, 2006. 729 p.

30
Belém (PA), 11 a 13 de dezembro de 2019
PEREIRA, W. L. M.; VELOSO, C. A. C.; GAMA, J. R. N. F. Propriedades químicas de
um Latossolo Amarelo cultivado com pastagens na Amazônia Oriental. Scientia
Agricola, Piracicaba, v.57, n.3, p.531-537, 2000.
RODRIGUES, T. E., SILVA, R. C., SILVA, J. M. L., OLIVEIRA, R. C., Jr., GAMA, J.
R. N. F., & VALENTE, M. A. (2003). Caracterização e classificação dos solos do
município de Paragominas, estado do Pará. Belém: Embrapa Amazônia Oriental. 51
p.
SANTOS, J.T.; ANDRADE, A.P.; SILVA, I.F.; SILVA, D.S.; SANTOS, E.M. & SILVA,
A.P.G. Atributos físicos e químicos do solo de áreas sob pastejo na microrregião do Brejo
Paraibano. Ciência Rural, 40:2486-2492, 2010.

31
Belém (PA), 11 a 13 de dezembro de 2019
ANÁLISE DO POTENCIAL PRODUTIVO DO FRUTO DE CUPUAÇU DOS
PRODUTORES ASSOCIADOS A COOPERATIVA AGRÍCOLA MISTA DE
TOMÉ-AÇU (CAMTA), PARÁ
Bruna Stefanny das Neves de Sousa1; Alfredo Kingo Oyama Homma2
1Mestrado em Ciências Ambientais. Universidade do Estado do Pará.
E-mail: [email protected] 2Doutorado em Economia Rural. Embrapa Amazônia Oriental.
E-mail: [email protected]
RESUMO
O cupuaçuzeiro é uma espécie tropical nativa da região amazônica, com carência de
informações sobre produção disponíveis na literatura. Eeste estudo efetuou análise de
relatórios da produção do cupuaçu em Sistemas Agroflorestais (SAFs), obtidos por
produtores brasileiros e nipo-brasileiros associados a Cooperativa Agrícola Mista de
Tomé-Açu (CAMTA) no ano de 2016, além da visita em campo referente as entrevistas
realizadas junto aos produtores. Foi utilizada a estatística descritiva, com intuito de
verificar a partir da estratificação da produção e número de pés, de dois tipos de cupuaçu beneficiados pela cooperativa (tipo B e em fruta). A pesquisa também fez uso do software
Microsoft Excel 2013 para a tabulação dos dados, verificação de médias e elaboração de
gráficos. Verificou que o cupuaçu em fruta obteve maior produção entre os produtores
(67,78%), assim como, a quantidade de número de pés plantados (77%) em comparação
ao cupuaçu tipo B. Em relação a produção arrecadada por número de produtores no ano
de 2016, a mesma se concentra em produções menores de 30.000 kg, correspondente a
um total de 65%. Em relação a produção por número de pés de cupuazureiros plantados,
os melhores indices produtivos foram adquiridos em áreas no estrato de 3.000 a 6.000 pés
plantados. Conclui-se, que a produção de cupuaçu vem ainda sendo bastante utilizada em
Sistemas Agroflorestais na região de Tomé-Açu, no entanto, segundo os produtores
entrevistados, há uma grande necessidade de pesquisas para o aprimoramento da
produção.
Palavras-chave: Produtores Nipo-brasileiros. Sistema agroflorestal. Produção agrícola.
Área de Interesse do Simpósio: Agronomia.

32
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A produção de cupuaçuzeiro (Theobroma grandiflorum (Willdenow ex Sprengel)
Schumann) na região de Tome-Açu, tornou-se um importante componente em Sistemas
Agroflorestais (SAF’s), devido ser bem adaptado ao sistema, além de proporcionar
benefício econômico na produção (VENTURIERI, 2011). De acordo, com Martins et al
(2013), o cupuaçuzeiro é muito produzido por pequenos produtores, em consórcio, devido
ser bem adaptado ao sombreamento. A transição do cupuaçuzeiro extrativista para o
domesticado, também possibilitou o aumento na produção, permitindo desta forma,
atender a demanda nacional e internacional, que nos últimos anos teve aumento
progressivo (GONDIM et al, 2001).
O consumo de cupuaçu exibe boa aceitação por parte da população regional,
principalmente pela fabricação de bombons, doces, sorvetes, picolés, cerveja, cosméticos
etc, sendo este, muito restrito para Estados fora da Amazônia e no exterior. Segundo
Lopes (2015), o cupuaçu é um fruto que proporciona além da polpa, importante matéria-
prima para indústria alimentícia, a semente possui destino para indústrias de cosmética,
o fruto ao contrário da polpa de açaí, teve sucesso restrito no mercado nacional e
internacional (SOCHA & PINHEIRO, 2016).
O acompanhamento da produção de cupuaçu no cenário econômico no Brasil, é
um tanto dificultoso, em virtude, ao acesso das informações disponíveis ao público.
Conforme, Almudi & Pinheiro (2015) o Instituto Brasileiro de Geografia Estatística
(IBGE), não permite o acesso aos dados de produção do cupuaçu, entretanto, pode ser
acompanhada pelo Grupo de Coordenação de Estatísticas Agropecuárias do Amazonas
(GCEA/PA), que revela a partir de seus dados, que o cupuaçu apresenta grande potencial
para a agricultura (HOMMA, 1996).
A maior produção de cupuaçu no Brasil se concentra na região Norte, onde o
Estado do Pará ocupa o primeiro lugar como maior produtor e comercializador. Contudo,
ressalta-se a ocorrência de perdas dos frutos para comercialização por diversos fatores,
principalmente pelo mau manejo das práticas agrícolas, influenciando de forma negativa
na obtenção de frutos salubres (RAMOS, 2015).
Na região Nordeste do Estado do Pará a CAMTA exerce importante papel para o
crescimento de pequenos produtores rurais, proporcionando melhores resultados na

33
Belém (PA), 11 a 13 de dezembro de 2019
produção e na economia, fundamentado na união e tomadas de decisões entre os mesmos.
A comercialização de polpas na CAMTA ainda é o principal produto comercializado, no
entanto, vem se aperfeiçoando a fabricação de derivados como sorvetes e geleias, como
também em estudos para o beneficiamento da semente, para fabricação de chocolate,
devido apresentar bom mercado de exportação (SOCHA & PINHEIRO, 2016).
Neste sentido, dada a importância ao estudo da produção de cupuaçu, o objetivo
do trabalho visa identificar por meio da análise de produção de cupuaçu em Sistemas
Agroflorestais (SAF’s), obtido pelo produtores nipo-brasileiros vinculados a CAMTA,
no ano de 2016, e por meio de entrevistas junto aos produtores brasileiros e nipo-
brasileiros, realizadas no ano de 2017, a verificação de mudanças nos sistemas de
produção e comercialização.
2. MATERIAL E MÉTODOS
Área de estudo
O estudo foi realizado no município de Tomé-Açu, região Nordeste do Estado do
Pará (Figura 1), de acordo com Valente et al. (2014) a região está localizada entre as
coordenadas 01° 24’ 46.14” e 01° 28’ 4.11” de latitude Sul, e 48° 20’ 04.60” e 48° 20’
31.84” de longitude Oeste de Greenwich. A população em 2010 compreende a 56.518
habitantes, com estimativa para 2017 de 61.709 habitantes (IBGE, 2016).
Figura 1- Localização da área de estudo.
Fonte: Sousa et al. (2017).

34
Belém (PA), 11 a 13 de dezembro de 2019
As características morfoclimáticas da região indicam relevo com cobertura vegetal
de Floresta Ombrófila Densa, clima quente e ajustando-se ao tipo climático Ami, da
classificação de Köppen, caracterizado como chuvoso, porém com pequena estação seca.
Os tipos de solos concentram-se em grande parte, em Latossolo Amarelo e Argilossolo
Amarelo (VALENTE et al, 2014).
Tipo de pesquisa
O presente estudo trata-se de uma pesquisa de abordagem quali-quantitativa.
Conforme Pereira (1997) os dados qualitativos e quantitativos apresentam diferenças entre
si, no entanto, estão diretamente correlacionados, pois dados qualitativos possuem
vantagens sobre os dados quantitativos, por possuir referências menos restritivas e maior
oportunidade de manifestação da subjetividade do binômio pesquisador/pesquisado e se
soma à pesquisa quantitativa na tentativa de explicar mais adequadamente a realidade. O
procedimento técnico utilizado neste estudo para a coleta de dados qualitativos, foram
feitos por meio de levantamento de dados em campo, no período de 18 a 20 de setembro
de 2017, em cinco propriedades de produtores vinculados à cooperativa CAMTA, onde
foram realizadas entrevistas, com perguntas abertas, a respeito das experiências obtidas
na produção de cupuaçu em SAF’s.
Utilizou-se também, o método da estatística descritiva, para análise de dados
quantitativos, com intuito de verificar a partir da estratificação da produção e número de
pés, de dois tipos de cupuaçu beneficiados pela cooperativa (tipo B e em fruta),
provenientes de relatórios da CAMTA, adquirido por 90 produtores nipo-brasileiros, no
ano de 2016. A descrição de informações oriundos de dados obtidos com a pesquisa, é o
objetivo da estatística descritiva, sendo o termo estatística a designação dos próprios dados
derivados (SPIEGEL, 1985).
Análise de dados
Foi utilizado o software Microsoft Excel 2013, para tabulação de dados,
determinação de médias, elaboração de gráficos e tabelas.
3. RESULTADOS E DISCUSSÃO
Atráves da pesquisa em campo, pode-se verificar por meio de entrevistas
realizadas junto aos produtores agrícolas de Tomé-Açu, que o modelo de cultivo do

35
Belém (PA), 11 a 13 de dezembro de 2019
cupuaçuzeiro em consócio com outras culturas realizados pelos emigrantes japoneses,
serviu como modelo adotado por muitos produtores na região. A imigração japonesa
para região Nordeste do Estado do Pará, foi resultado da busca pelo desenvolvimento
econômico da região, servindo como mão-de-obra para desenvolvimento na Amazônia
(SILVA & TAFNER JUNIOR., 2010).
Em se tratando da produção do cupuaçu, são diversas as combinações e
espaçamentos com outras culturas agrícolas e também florestais (Tabela 1). De acordo
com os entrevistados, o plantio da cultivar BRS Carimbó e mudas de pé-franco são mais
comuns cultivadas entre os agricultores na região. A cultivar BRS Carimbó tem como
diferencial, características geneticamente melhoradas, possibilitando ótima capacidade
de produção de frutos e boa resistência à vassoura-de-bruxa (Moniliophtera perniciosa),
assim como, apresentar técnicas semelhantes ao cultivo do cupuaçuzeiro do tipo pé-
franco, como preparo de mudas, instalação e condução de pomares (LIMA, 2012).
Tabela 1- Combinações com cupuaçuzeiros em Sistemas Agroflorestais (SAF’s),
presente na propriedade de um dos produtores entrevistados.
Espaçamento Combinação de cupuaçuzeiros em SAF’s
(7m x 5m) Cupuaçuzeiro + coqueiro
(7m x 5m) Cupuaçuzeiro + castanheira + mogno + mangustanzeiro
(7m x 5m) Cupuaçuzeiro + freijoeiro + samaumeira + açaizeiro
(7m x 5m) Cupuaçuzeiro + pimenteira + maracujazeiro + bananeira+
cacaueiro + paricazeiro + mogno
(7m x 5m) Cupuaçuzeiro + castanheira + mogno + cacaueiro
(7m x 5m) Cupuaçuzeiro + pimenteira + piquiazeiro + pupunheira
(7m x 5m) Cupuaçuzeiro + açaizeiro + castanheira + bacabeira
(7m x 5m) Cupuaçuzeiro + cacaueiro + paricazeiro
(7m x 5m) Cupuaçuzeiro + paricazeiro + piquiazeiro + uxizeiro +
copaibeira + andirobeira + castanheira
(7m x 5m) Cupuaçuzeiro + pimenteira + cacaueiro + pupunheira +
castanheira+ piquiazeiro
(7m x 5m) Cupuaçuzeiro + cacaueiro + mogno africano + paricazeiro +
andirobeira
(6m x 5m) Cupuaçuzeiro + açaizeiro
Fonte: Sousa et al. (2019).

36
Belém (PA), 11 a 13 de dezembro de 2019
Um fator importante que tem possibilitado melhorias no cultivo e também de
retorno econômico aos produtores, consiste no acesso de informações oferecidas pelas
empresas de assistência técnica como a Emater-Pará, Embrapa Amazônia Oriental,
Chamada pública e INCUBITEC (Incubadora Tecnológica de Desenvolvimento e
Inovação de Cooperativas e Empreendimentos Solidários). Essas empresas tem o
propósito de ensinar aos agricultores como desenvolverem sistemas agrícolas de forma
ambiental e sustentável (CASTRO, 2015).
Os dados quantitativos adquiridos dos relatórios da CAMTA mostraram que a
produção do cupuaçu para fins de comercialização da polpa, classificam dois tipos de
cupuaçu, o em fruta e o tipo B, isso ocorre por meio do beneficiamento dos frutos, sendo
feito a classificação do mesmo, de acordo com as qualidades e propriedades que
apresentam (tamanho, sabor, consistência, acidez, etc.). A comercialização da polpa de
cupuaçu é motivada pela viabilidade do mercado, tanto a nível nacional e internacional
(GODIM et al., 2001). Esses fatores estão também relacionados a substituição do cultivo
extrativista para a forma domesticada devido aos bons resultados dos sistemas
consorciados e de forma racional (HOMMA, 1996).
Na Tabela 2, pode-se observar que os dados obtidos referente ao número de
produtores que produzem o cupuaçu em fruta (67,78%), se destaca em relação ao
cupuaçu tipo B (2,22%), sendo que 30% dos produtores não cultivam nenhum dos dois
tipos de cupuaçu analisados neste estudo. O cupuaçu tipo B pode ser considerado como
uma novidade na produção, ainda é produzido em poucas quantidades como parte
integrante em sistemas agroflorestais. A preferêcia pela produção do cupuaçu em fruta,
pode ser justificado por facilitar a comercialização sem necessidade de maiores
cuidados. O melhoramento genético possibilita a obtenção de um genótipo com bons
padrões de produção, com excelente qualidade de polpa para agroindústria, e diferentes
formas de amadurecimento (precoce, mediano e tardio), visando aumentar o período de
colheita e oferta de frutos no mercado (SOUZA, 1966).
37
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 2- Número produtores agrícolas de Tomé-Açu, em relação a obtenção dos tipos de cupuaçu
beneficiados pela cooperativa (CAMTA), no ano de 2016.
Tipos de cupuaçu Nº de produtores Proporção (%)
Cupuaçu em fruta 61 67,78
Cupuaçu tipo B 2 2,22
Não plantam 27 30
Total 90 100,00
Fonte: Sousa et al. (2019).
Com base na produção arrecadada por número de pés plantados pelos produtores,
contata-se que o tipo de cupuaçu em fruta apresentou uma considerada produção de 77%
em comparação a 0,02% do cupuaçu tipo B (Tabela 3). As mesmas observações relatadas
na tabela anterior, refletem na obtenção dos dados da produção. A aquisição de materiais
produtivos, tende ser uma variável importante, pois permite conhecer a produção total
de polpa por árvore, obtida por meio do número de frutos produzidos (GONDIM, 2001).
Tabela 3- Dados médios de produção obtidos pelos produtores agrícolas de Tomé-Açu, no ano de
2016.
Tipos de cupuaçu Produção (kg) Porcentagem (%)
Cupuaçu em fruta 744,26 99,98
Cupuaçu tipo B 149,00 0,02
Total 744,41 100,00
Fonte: Sousa et al. (2019).
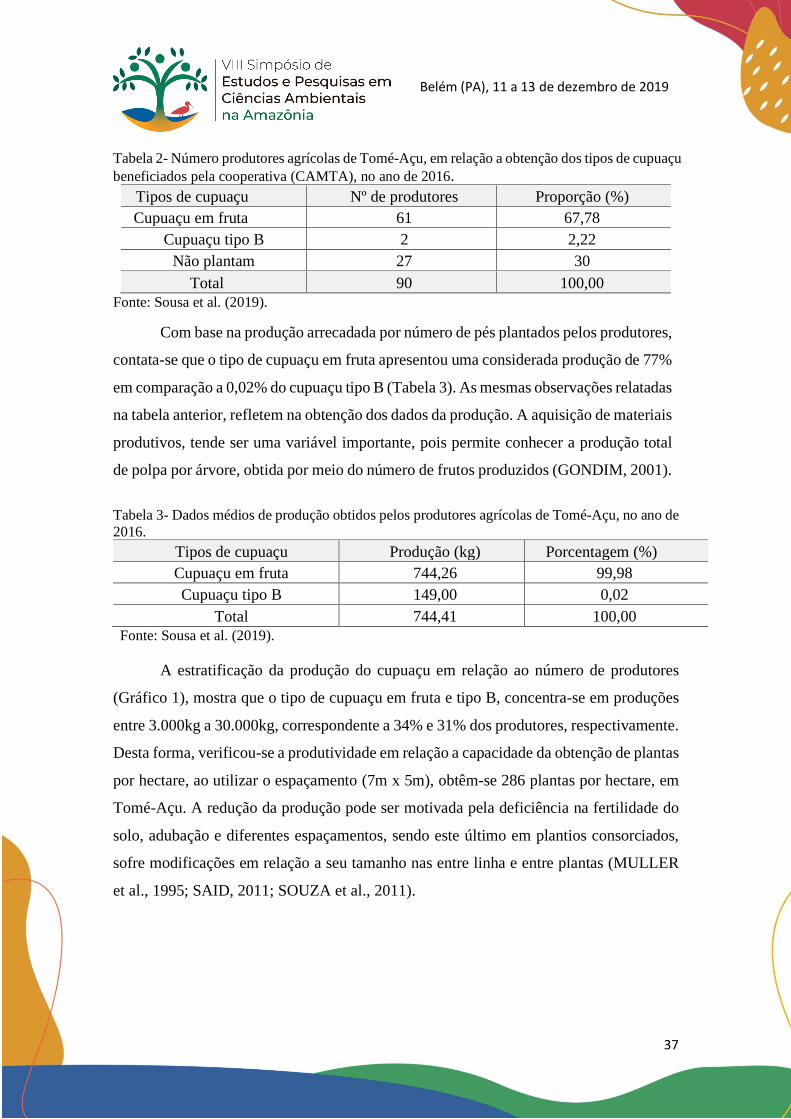
A estratificação da produção do cupuaçu em relação ao número de produtores
(Gráfico 1), mostra que o tipo de cupuaçu em fruta e tipo B, concentra-se em produções
entre 3.000kg a 30.000kg, correspondente a 34% e 31% dos produtores, respectivamente.
Desta forma, verificou-se a produtividade em relação a capacidade da obtenção de plantas
por hectare, ao utilizar o espaçamento (7m x 5m), obtêm-se 286 plantas por hectare, em
Tomé-Açu. A redução da produção pode ser motivada pela deficiência na fertilidade do
solo, adubação e diferentes espaçamentos, sendo este último em plantios consorciados,
sofre modificações em relação a seu tamanho nas entre linha e entre plantas (MULLER
et al., 1995; SAID, 2011; SOUZA et al., 2011).
38
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2 - Estratificação da produção de cupuaçu (kg), obtido pelos produtores agrícolas
vinculados à CAMTA, no ano de 2016.
Fonte: Sousa et al. (2019).
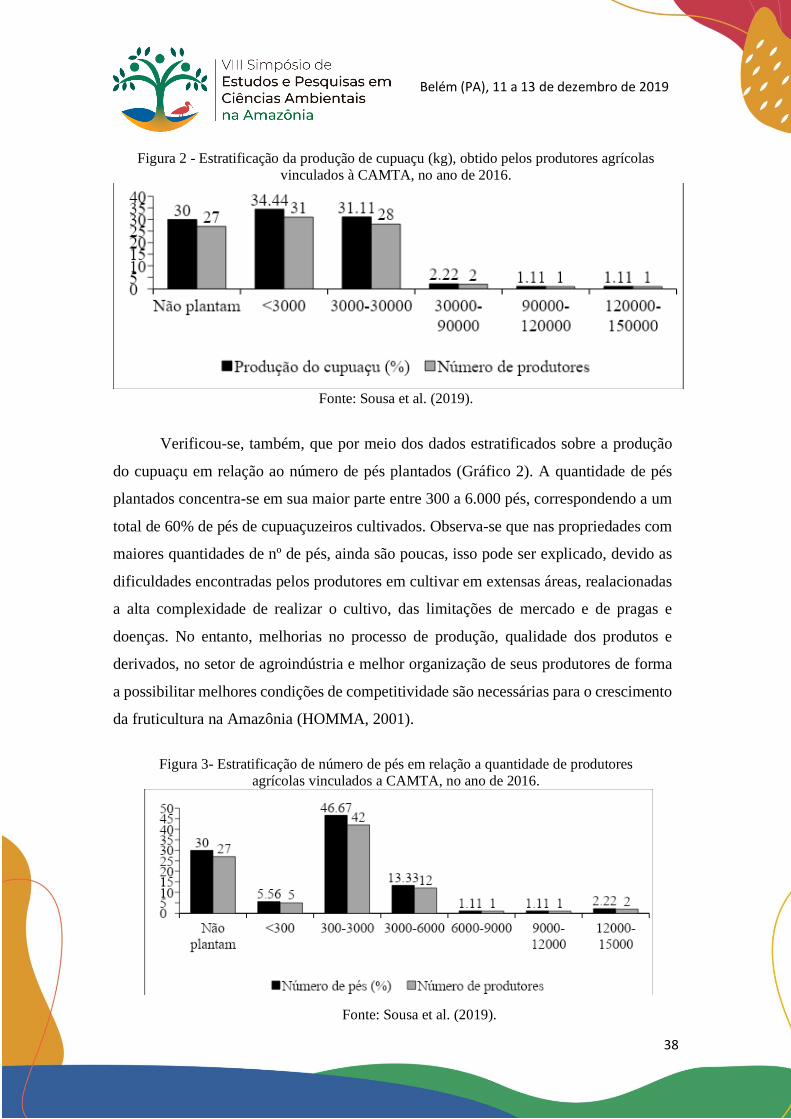
Verificou-se, também, que por meio dos dados estratificados sobre a produção
do cupuaçu em relação ao número de pés plantados (Gráfico 2). A quantidade de pés
plantados concentra-se em sua maior parte entre 300 a 6.000 pés, correspondendo a um
total de 60% de pés de cupuaçuzeiros cultivados. Observa-se que nas propriedades com
maiores quantidades de nº de pés, ainda são poucas, isso pode ser explicado, devido as
dificuldades encontradas pelos produtores em cultivar em extensas áreas, realacionadas
a alta complexidade de realizar o cultivo, das limitações de mercado e de pragas e
doenças. No entanto, melhorias no processo de produção, qualidade dos produtos e
derivados, no setor de agroindústria e melhor organização de seus produtores de forma
a possibilitar melhores condições de competitividade são necessárias para o crescimento
da fruticultura na Amazônia (HOMMA, 2001).
Figura 3- Estratificação de número de pés em relação a quantidade de produtores
agrícolas vinculados a CAMTA, no ano de 2016.
Fonte: Sousa et al. (2019).

39
Belém (PA), 11 a 13 de dezembro de 2019
4. CONCLUSÃO
O início dos primeiros plantios de cupuaçuzeiros ocorreu no municipio de Tomé-
Açu antes restrita a reduzidas árvores em quintais e da produção extrativa na região de
Marabá. O cupuazuzeiro adaptou muito bem em SAFs, porém restrições de mercado e
ataque de pragas e doenças tem limitado a sua expansão. Há necessidade do
desenvolvimento de novas tecnologias visando o aproveitamento da polpa para novos
usos, melhoramento genético e das práticas agrícolas.

40
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALMUDI, T; PINHEIRO, J. O. C. Dados estatísticos da Produção Agropecuária e
Florestal do Estado do Amazonas: Ano 2013. Embrapa, Brasília - DF, 2015. 105p.
CASTRO, N. C. Desafios da agricultura familiar: o caso da assistência técnica e
extensão rural. Boletim regional, urbano e ambiental, v.12, p. 49-59, 2015.
CREMONESE, C.; SCHALLENBERGER, E. Cooperativismo e agricultura familiar na
formação do espaço agrícola do Oeste do Paraná. Tempo da Ciência, v. 12, n. 23, p.
49-63, 2005.
GONDIM, T. M. S.; THOMAZINI, M. J.; CAVALCANTE, M. J.; SOUZA, J. M. L.
Aspectos da produção de cupuaçu- (Embrapa Acre. Documentos; 67), 2001. 46p.
HOMMA, A. K. O. Cupuaçu: potencialidades e mercado, algumas especulações. In:
WORKSHOP SOBRE AS CULTURAS DE CUPUAÇU E PUPUNHA NA
AMAZÔNIA, 1996, Manaus. Anais... Manaus: Embrapa- CPAA. 1996. p. 85-95.
HOMMA, A. K. O. O desenvolvimento da agroindústria no estado do Pará. Saber:
Ciências exatas e tecnologia, Belém, v. 3, p. 49-73, 2001.
Instituto Brasileiro de Geografia e Estatística – 2016. Tomé-Açu. Disponível em:
http://cod.ibge.gov.br/2VREN. Acesso em: 31 de jun. 2019.
LIMA, A. L. Produtores terão nova cultivar de cupuaçuzeiro, a BRS Carimbó. Embrapa,
2012. Disponível em: https://www.embrapa.br/busca-de-noticias/-
/noticia/1463133/produtores-terao-nova-cultivar-de-cupuacuzeiro-a-brs-carimbo.
Acesso em: 9 de set. 2019.
MARTIM, S. R.; CARDOSO NETO, J.; OLIVEIRA, I. M. A. Características físico-
químicas e atividade da peroxidase e polifenoloxidase em genótipos de cupuaçu
(Theobroma grandiflorum Willd ex-Spreng Schum) submetidos ao congelamento.
Ciências Agrárias, v. 34, n. 5, p. 2265-2276, 2013.
MARTINS, V. B. Perfil sensorial de suco tropical de cupuaçu (Theobroma
grandiflorum Schum) com valor calórico reduzido. 2008. Tese (Doutorado em
Alimentos e Nutrição) – Faculdade de Engenharia de Alimentos, Campinas, 2008.
MINATEL, J. F.; BONGANHA, C. A. Agronegócios: A importância do cooperativismo
e da agricultura familiar. Empreendedorismo, Gestão e Negócios, v. 4, n. 4, p. 247- 259,
2015.
MÜLLER, C. H.; FIGUEIRÊDO, F. J. C.; NASCIMENTO, W. M. O. do; GALVÃO, E.
U. P.; STEIN, R. L. B.; SILVA, A. de B.; RODRIGUES, J. E. L. F.; CARVALHO, J. E.
U. de; NUNES, A. M. L.; NAZARÉ, R. F. R. de; BARBOSA, W. C. A cultura do
cupuaçu. Brasília, DF: Embrapa-SPI; Belém: Embrapa-CPATU. 1995. 61p.

41
Belém (PA), 11 a 13 de dezembro de 2019
PEREIRA, I. B. Metodologia Qualitativa nas Pesquisas em Saúde Coletiva:
Considerações teóricas e instrumentais. Faculdade de Saúde Pública, Universidade de
São Paulo, São Paulo. 1997. 21p .
RAMOS, S. N. M. Influência dos microrganismos envolvidos no processo de
fermentação de cupuaçu (Theobroma grandiflorum Schum) na formação do sabor.
Tese (Doutorado em Tecnologia de Alimentos) – Universidade Estadual de Campinas,
Campinas, SP, 2015.
SAGRI. Secretaria de Estado de Agricultura. A fruticultura no Estado do Pará. Pará, 2012.
Disponível em:
http://sedap.pa.gov.br/files/pdfs/SEB_Cartilha_Frutal_18x21cm_OUT11_FINAL.pdf.>.
Acesso em: 09 set. 2019.
SAID, M. M. Aspectos culturais e potenciais de uso do cupuaçu no estado do
Amazonas. Dissertação (Mestrado em Ciências do ambiente e Sustentabilidade na
Amazônia) –PPG/CASA. Ufam-Manaus, 2011.
SILVA, F. C.; TAFNER JÚNIOR, A. W. Cooperativismo e Desenvolvimento Regional
na Amazônia: a contribuição da Cooperativa Agrícola Mista de Tomé-Açu para o
desenvolvimento do Nordeste paraense. Dissertação (Mestrado em Planejamento do
Desenvolvimento) - Universidade Federal do Pará, PA, 2010.
SOCHA; L. B; PINHEIRO, R.B.M. Cupuaçu: A fruta globalizada. Revista Gestão &
Sustentabilidade Ambiental, v. 4, n. 2, p. 554-567, 2016.
SPIEGEL, M. Estatística. 2ª ed. São Paulo: Mc Graw-Hill do Brasil, 1985.
SOUZA, A. G. C, SOUZA. M. G; PAMPLONA. A. M. S. R. Boas práticas na colheita
e pôs colheita do cupuaçu – (Embrapa Amazônia Oriental. Circular técnica), 2011. 8p.
VALENTE, M. A.; WATRIN, O. S.; CASTRO, A. R.C. Mapeamento Detalhado dos
Solos da Fazenda Experimental da Embrapa Amazônia Oriental em Tomé-Açu, PA.
(Embrapa/Documentos; 405), 2014. 33p.
VENTURIERI, G. A. Flowering levels, harvest season and yields of cupuassu
(Theobroma grandiflorum). Acta Amazonica, v. 41, p. 143-152, 2017.

42
Belém (PA), 11 a 13 de dezembro de 2019
AVALIAÇÃO DE CARACTERÍSTICAS SOCIOECONÔMICAS DE
COMERCIANTES E DE ABASTECIMENTO DE HORTALIÇAS EM FEIRAS
LIVRES DE BELÉM – PA
Josiene Amanda dos Santos Viana1; Marcus Vinicius Santiago de Oliveira Silva2; André
Gustavo Campinas Pereira3; Érica Coutinho David4; Letícia Cunha da Hungria5.
1 Graduanda em Agronomia. Universidade Federal Rural da Amazônia.
[email protected]. 2Graduando em Agronomia. Universidade Federal Rural da Amazônia.
[email protected]. 3 Especialista em Geoprocessamento Aplicado à Agroecologia e ao Uso dos Recursos
Naturais. Universidade Federal do Pará. [email protected]. 4 Graduanda em Agronomia. Universidade Federal Rural da Amazônia.
[email protected]. 5 Doutoranda em Agronomia. Universidade Federal Rural da Amazônia.
RESUMO
As hortaliças comercializadas nas feiras livres apresentam uma grande atividade
econômica e mais antiga da história, possuindo alta importância no agronegócio. O
seguinte estudo objetivou realizar um levantamento e avaliar características que compõe
o perfil socioeconômico de comerciantes de hortaliças e a situação de abastecimento em
feiras livres na cidade de Belém-PA. A pesquisa ocorreu no mês de junho de 2018 em
três feiras livres localizadas na região, onde foram aplicados 53 questionários que
contemplavam aspectos sociais abordando gênero, faixa etária e nível de escolaridade.
No seguinte estudo foi possível observar que há a predominância masculina (66%) em
relação a feminina (34%) à frente dos empreendimentos relativos à venda de hortaliças.
Com relação a faixa etária, há distribuição uniforme a partir das idades a 30 anos (92,5%),
com variações menores de 10% entre as faixas. Dentre estas faixas, estão entre 50 a 59
anos (28,3%) e uma pequena participação dos jovens entre 18 a 29 anos (7,5%). Foi
possível concluir que, a CEASA-PA exerce importante papel no fornecimento dos
produtos olerícolas comercializados nas feiras livres de Belém-PA, sendo o principal
fornecedor na região. Também é possível entender que a atividade como comerciante de
hortaliças está ameaçada pela avançada faixa etária dos feirantes em exercício e o
reduzido quadro de novas pessoas no ramo.
Palavras-chave: Socioeconomia dos comerciantes. Feiras livres. Belém.
Área de Interesse do Simpósio: Agronomia.

43
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
As feiras livres se conformam como uma das atividades econômicas mais antigas
da história e fizeram parte tanto da emergência quanto do desdobramento do modelo
capitalista (BERNARDINO, 2013). Apesar da crescente participação dos supermercados
na comercialização de produtos olerícolas, as feiras livres ainda detêm grande
importância neste segmento do agronegócio. SILVEIRA et al. (2017) afirmam que
oferecer produtos de qualidade por preços mais atrativos são objetivos das feiras livres.
Um dos fatores que contribui fortemente para a grande busca na aquisição de
produtos em feiras livres é a relação mais direta entre o produtor e o consumidor,
possibilitando a identificação mais rápida e íntima das necessidades do consumidor, que
influenciam nos aspectos da produção e comercialização (MACHADO & SILVA, 2005).
Afinal, a satisfação do consumidor é um dos fatores que garantem a continuidade do
negócio e para que isso ocorra é necessário haver adequação dos negócios conforme a
demanda. Além disso, a feira tem como característica a venda de diversos produtos, além
disso, a limpeza, as mercadorias bem exibidas e com os preços mais baixos que os
supermercados despertam cada vez mais os consumidores a frequentá-la (SILVEIRA et
al., 2017).
A comercialização de hortaliças, por exemplo, é efetivada não só pelo ponto de
vista nutricional e farmacêutico, mas também econômico, atuando como fator de
movimentação financeira em localidades onde estão sendo comercializadas. No entanto,
a falta de informação ou planejamento na venda pode influenciar o consumo de hortaliças
em feiras. Segundo o Centro de Produções Técnicas - CPT (2019), a queda no consumo
hortaliças pode estar relacionado a diversos fatores como a perecibilidade, a pouca
disponibilidade de pontos de venda com produtos de boa qualidade, receio da origem do
produto e consumo de alimentos contaminados por defensivos agrícolas e o conhecimento
incipiente sobre a dimensão das hortaliças na alimentação.
Neste contexto, dados sobre o perfil dos comerciantes junto à origem e frequência
de abastecimento de hortaliças em feiras podem proporcionar informações valiosas no
melhor planejamento da comercialização, uma vez que os feirantes são agentes
fundamentais na intermediação de mercadoria entre produtor-consumidor.

44
Belém (PA), 11 a 13 de dezembro de 2019
O estudo objetivou realizar um levantamento e avaliar características que compõe
o perfil socioeconômico de comerciantes de hortaliças e a situação de abastecimento em
feiras livres na cidade de Belém-PA.
2. METODOLOGIA
A pesquisa foi realizada durante o mês de junho de 2018 em três feiras livres
localizadas em Belém, nordeste do Estado do Pará, sendo eleitas para o estudo a feira do
ver-o-peso, feira da 25 e feira da pedreira. Para isso, utilizou-se metodologia de visitação
“in loco” para a aplicação de questionários semiestruturados (survey) e entrevistas aos
comerciantes de hortaliças.
Um total de 53 questionários foi aplicado e contemplavam aspectos sociais como
gênero, faixa etária e nível de escolaridade, bem como aspectos de abastecimento de
hortaliças nos estabelecimentos. Para a análise dos dados utilizou-se estatística descritiva
para obtenção de médias e frequências relativas.
3. RESULTADOS E DISCUSSÃO
Nesta pesquisa, registrou-se a predominância da participação masculina (66%) em
relação à proporção feminina (34%) à frente dos empreendimentos relativos à venda de
hortaliças (Figura 1). Em estudo semelhante BERNARDINO (2013) observou que a
maioria dos feirantes deste setor era do sexo masculino (64,2%). Isto pode estar
associado, principalmente, à traços culturais e tradicionais, trazendo geralmente o homem
para atividades de comercialização.
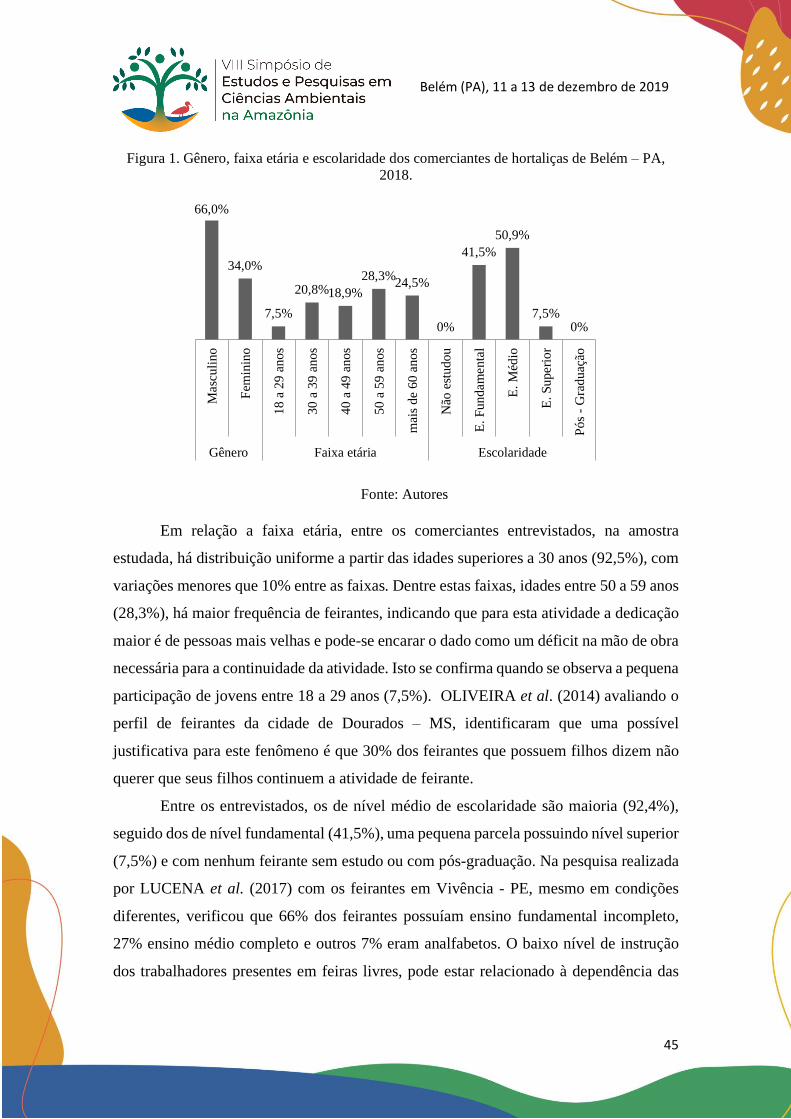
45
Belém (PA), 11 a 13 de dezembro de 2019
Figura 1. Gênero, faixa etária e escolaridade dos comerciantes de hortaliças de Belém – PA,
2018.
Fonte: Autores
Em relação a faixa etária, entre os comerciantes entrevistados, na amostra
estudada, há distribuição uniforme a partir das idades superiores a 30 anos (92,5%), com
variações menores que 10% entre as faixas. Dentre estas faixas, idades entre 50 a 59 anos
(28,3%), há maior frequência de feirantes, indicando que para esta atividade a dedicação
maior é de pessoas mais velhas e pode-se encarar o dado como um déficit na mão de obra
necessária para a continuidade da atividade. Isto se confirma quando se observa a pequena
participação de jovens entre 18 a 29 anos (7,5%). OLIVEIRA et al. (2014) avaliando o
perfil de feirantes da cidade de Dourados – MS, identificaram que uma possível
justificativa para este fenômeno é que 30% dos feirantes que possuem filhos dizem não
querer que seus filhos continuem a atividade de feirante.
Entre os entrevistados, os de nível médio de escolaridade são maioria (92,4%),
seguido dos de nível fundamental (41,5%), uma pequena parcela possuindo nível superior
(7,5%) e com nenhum feirante sem estudo ou com pós-graduação. Na pesquisa realizada
por LUCENA et al. (2017) com os feirantes em Vivência - PE, mesmo em condições
diferentes, verificou que 66% dos feirantes possuíam ensino fundamental incompleto,
27% ensino médio completo e outros 7% eram analfabetos. O baixo nível de instrução
dos trabalhadores presentes em feiras livres, pode estar relacionado à dependência das
66,0%
34,0%
7,5%
20,8%18,9%
28,3%24,5%
0%
41,5%
50,9%
7,5%0%
Masc
uli
no
Fem
inin
o
18 a
29 a
nos
30 a
39 a
nos
40 a
49 a
nos
50 a
59 a
nos
mai
s de
60 a
nos
Não
est
udou
E. F
undam
enta
l
E. M
édio
E. S
uperi
or
Pós
- G
raduaç
ão
Gênero Faixa etária Escolaridade

46
Belém (PA), 11 a 13 de dezembro de 2019
pessoas na busca pelo sustento familiar, muitas vezes atrelado à comercialização, que
acaba limitando seus estudos.
A maior parte do abastecimento dos feirantes (49,1%) advém da CEASA – PA,
juntamente a produtores locais da região metropolitana de Belém, assim como ocorre nas
observações de LUCENA et al. (2018), quando realizam o diagnóstico socioeconômico
dos feirantes de Vicência – PE.
Em adição, a maioria dos estabelecimentos (41,5%) são abastecidos diariamente,
seguidos dos que recebem os produtos três vezes na semana (24,5%). Tal peculiaridade é
facilmente explicada pela alta perecibilidade das hortaliças em geral, fato este que
imprime a essas mercadorias um tempo de prateleira estritamente curto.
Figura 2. Origem e frequência do abastecimento de hortaliças em feiras livres de Belém-PA,
2018.
Fonte: Autores
4. CONCLUSÃO
De tal forma conclui-se que, a CEASA-PA exerce importante papel no
fornecimento dos produtos olerícolas comercializados nas feiras livres de Belém/PA,
sendo majoritariamente o principal fornecedor na região.
Também é possível entender que a atividade como comerciante de hortaliças está
ameaçada pela avançada faixa etária dos feirantes em exercício e o reduzido quadro de
novas pessoas no ramo.
0%
41,5%49,1%
9,4% 9,4%
41,5%
7,5%17,0%
24,5%
Pro
duto
res
Loca
is
CE
AS
A-P
A
CE
AS
A e
Pro
d.
Loca
is Outr
os
Irre
gula
Dia
riam
ente
Um
a vez
por
sem
ana
Duas
vez
es p
or
sem
ana
Trê
s veze
s por
sem
ana
Origem do abastecimento Frequência de abastecimento

47
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BERNARDINO, V. M. P. Perfil dos feirantes e aspectos geohistóricos de
comercialização da feira livre de Leiria – Portugal. In: SIMPÓSIO DE ESTUDOS
URBANOS: A DINÂMICA DAS CIDADES E A PRODUÇÃO DO ESPAÇO, 2, 2013,
Campo Mourão. Anais...Campo Mourão: UEPR, 2013.
SILVEIRA, V. C. da; OLIVEIRA, E. S. de; SILVEIRA, N. F.; MARIANI, M. A. P.
Avaliação da importância das feiras livres e a forma de comercialização adotada pelos
feirantes na cidade de Nova Andradina – MS. In: ENCONTRO INTERNACIONAL DE
GESTÃO, DESENVOLVIMENTO E INOVAÇÃO, 1, 2017, Naviraí – MS.
Anais...Naviraí: UFMS, 2017.
CENTRO DE PRODUÇÕES TÉCNICAS: POR QUE CONSUMIR HORTALIÇAS?
Porque é 10 para a saúde!. Disponível em: < https://www.cpt.com.br/cursos-horticultura-
agricultura/artigos/por-que-consumir-hortalicas-porque-e-10-para-a-saude>. Acesso em:
01 de fevereiro de 2019.
CANELLA, D. S.; LOUZADA, M. L. da C.; CLARO, R. M.; COSTA, J. C.; BANDONI,
D. H.; LEVY, R. B.; MARTINS, A. P. B. Consumo de hortaliças e sua relação com os
alimentos ultraprocessados no Brasil. Revista de Saúde Pública, Rio de Janeiro, v. 52,
p. 50, 3 maio de 2018.
OLIVEIRA, K. V. de; SANJINEZ-ARGANDOÑA, E. J.; CHUBA, C. A. M. Avaliação
do perfil dos feirantes da cidade de Dourados – MS na comercialização de produtos
alimentícios. In: ENCONTRO DE ENSINO, PESQUISA E EXTENSÃO
UNIVERSITÁRIA, 5/8, 2014, Dourados. Anais...Dourados: Universidade Federal da
Grande Dourados, 2014.
LUCENA, T. C.; COSTA, C. M dos S.; PERES, G. M. da S.; SANTOS, S. C. R. dos;
SILVA, W. C. da. Diagnóstico socioeconômico dos feirantes de Vicência - PE. In:
Congresso Internacional das Ciências Agrárias, 3, 2018, João Pessoa – PB. Anais... João
Pessoa: COINTER, 2018.
MACHADO MD; SILVA AL. 2005. Canais de distribuição para produtos da agricultura
familiar. In: SOUZA FILHO HM; BATALHA MO. (org). Gestão integrada da
agricultura familiar. São Carlos: Edufscar. p.95-129.
SILVEIRA, V. C. da; OLIVEIRA, E. S. de; SILVEIRA, N. F.; MARIANI, M. A. P.
Avaliação da importância das feiras livres e a forma de comercialização adotada pelos
feirantes na cidade de Nova Andradina – MS. In: I Encontro Internacional de Gestão,
Desenvolvimento e Inovação. I, 2017, Naviraí-MS.

48
Belém (PA), 11 a 13 de dezembro de 2019
AVALIAÇÃO FENOLÓGICA DA ESPÉCIE Cissus sicyoides L. ASSOCIADA
COM A PRECIPITAÇÃO PLUVIOMÉTRICA
Ruanny Karen Vidal Pantoja Portal Moreira1; Osmar Alves Lameira2; Meiciane Ferreira
Campelo3; Allan Christiam Santos Ramires4; Nilza Araújo Pacheco5
1Discente, Curso de Doutorado do Programa de Pós-Graduação em Biodiversidade e
Biotecnologia da Rede Bionorte, Universidade Federal do Pará, UFPA,
[email protected] 2 Pesquisador da Embrapa Amazônia Oriental, [email protected]
3 Discente, Curso de Doutorado do Programa de Pós-Graduação em Biodiversidade e
Biotecnologia da Rede Bionorte, Universidade Federal do Pará,
UFPA,[email protected]
4 Discente de Agronomia, Universidade Federal Rural da Amazônia,UFRA,
5 Pesquisadora da Embrapa Amazônia Oriental, [email protected]
RESUMO
A espécie Cissus sicyoides L., conhecida popularmente como cipó-pucá, insulina
vegetal sendo muita utilizada na medicina tradicional. O objetivo do trabalho foi de
associar os eventos de floração e frutificação da espécie Cissus sicyoides L. com a
precipitação pluviométrica, tendo como finalidade identificar a época favorável para a
coleta de sementes e utilização de folhas da espécie para uso medicinal. Foram avaliados
dez indivíduos diariamente no período de janeiro de 2015a dezembro de 2018 e registrou-
se a quantidade de dias de floração e frutificação. Ocorreram floração e frutificação em
quase todos os meses do ano. A maior média de floração e frutificação foi registrada no
mês de março, respectivamente, 10 e 11 dias. Os resultados demonstraram que o elemento
meteorológico influenciou o número de dias de ocorrência das fenofases e a coleta de
folhas para uso medicinal deve ser realizada, preferencialmente no mês de março e nos
dias em que ocorrem as fenofases.
Palavras-chave: Cipó-pucá. Fenofases. Elemento meteorológico.
Área de Interesse do Simpósio: Agronomia

49
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Uma das práticas mais antigas utilizadas pela humanidade é a fitoterapia, assim,
as plantas medicinais possuem um elo cultural muito forte com as distintas regiões
brasileiras no efeito e utilização de resultados para o alívio e até a cura dos sintomas,
sendo elas possuidoras de conhecimentos milenares que são repassados de gerações a
gerações (COSTA; MARINHO, 2015).
Dentre as diversas plantas encontradas na medicina terapêutica, encontra-se o
Cissus sicyoides L. pertencente à família Vitaceae, é conhecida popularmente como
insulina vegetal, cipó-pucá e anil trepador. Esta espécie é usada popularmente no
tratamento de diabetes (SANTOS et al., 2008), controle de estados epilépticos (Abreu et
al., 2003), hipotensor e no tratamento de doenças do coração (ALEXANDRE,
SILVEIRA, 2007).
É relevante ressaltar que quando a espécie é utilizada com fins terapêuticos, a
época de colheita inadequada pode acarretar em um material com redução do valor
terapêutico e/ou predominância de princípios tóxicos (CARVALHO, 2012). Desta forma,
estudos sobre fenologia abordam os diferentes eventos biológicos, dentre estes eventos
estão o florescimento e a frutificação (NEVES et al., 2010). E para o desenvolvimento da
planta Bergamaschi (2007) afirma que dentre os principais elementos do clima que
influenciam no desenvolvimento e crescimento das plantas, e consequentemente na sua
atividade fenológica, estão à duração de horas brilho solar do dia, a temperatura do ar e o
regime pluviométrico. Conforme Segantini et al. (2010) cada espécie apresenta
comportamento diferente em função das variáveis do ambiente, sendo necessários estudos
direcionados para cada espécie e em cada local de cultivo.
Com base nisto, este trabalho teve como objetivo associar os eventos de floração
e frutificação da espécie Cissus sicyoides L. com a precipitação pluviométrica, tendo
como finalidade identificar a época favorável para a coleta de sementes e utilização de
folhas da espécie para uso medicinal.

50
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
A pesquisa foi realizada no horto de plantas medicinais da Embrapa Amazônia
Oriental, situada no município de Belém-PA, localizado a 01º 28’ de latitude de sul e 48o
27’ de longitude oeste de Greenwich, com temperaturas médias, máximas e mínimas de
26.4 ºC, 31.8 e ºC, 22.9 ºC, respectivamente, umidade relativa de 84 % e precipitação
pluviométrica de 3.001 mm (BASTOS et al, 2002).
Para as observações das fenofases, floração e frutificação foram monitoradas dez
indivíduos adultos de Cissus sicyoides L. (Figura 1). As avaliações foram realizadas
diariamente pela manhã durante o período de janeiro de 2015 a dezembro de 2018, todos
os dados foram tabulados e processados em planilhas do sistema Microsoft Excel® (2010),
gerando um gráfico para a análise de possível influência do elemento climático sobre as
fenofases em estudo. Para os eventos fenológicos considerou-se número de dias por mês
de ocorrência das fenofases e para a variável precipitação pluviométrica foi utilizado à
média mensal acumulada. Os valores da precipitação pluviométrica foram provenientes
da Estação Climatológica da Embrapa Amazônia Oriental, instalada em Belém-PA.
Figura 1: Espécie cipó-pucá (Cissus sicyoides L.)
Fonte: Portal Moreira et al., (2019)

51
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
De acordo com a figura 2, para o evento fenológico, floração, não houve registro
nos meses de maio a setembro, e a maior média de dias de ocorrência foi no mês de março,
com valor de 10 dias, consequentemente no mês que ocorreu o pico de intensidade da
precipitação pluviométrica (481,00 mm). Para a frutificação, esta não ocorreu nos meses
de junho, setembro e outubro, e como a floração também o maior registro da fenofase foi
observado no mês de maior intensidade chuvoso com 11 dias de registro desta fenofase,
induzindo que esta espécie tem seu ápice fenológico no período intenso do inverno
amazônico. Lameira & Pinto (2008) relatam que o estádio de desenvolvimento da planta
também é muito importante para que se determine o ponto de colheita, e que a coleta de
material vegetal para fins fitoterápicos não deve coincidir com ocorrência dos eventos
fenológicos, pois os locais destes eventos concentram maiores quantidades de princípios
ativos.
Figura 2: Média dos números de dias de fenofases da espécie Cipó-pucá relacionado com a
Precipitação pluviométrica.
Fonte: Portal Moreira et al., (2019).
4. CONCLUSÃO
A precipitação pluviométrica influencia as fenofases da espécie Cissus sicyoides
L., pois, os maiores registros de ocorrência das fenofases coincidem com o maior pico de
regime pluviométrico. Com isso, é recomendado realizar a coleta de material vegetal para
propagação por sementes da espécie podem ser realizada durante a ocorrência das
fenofases e para seu uso medicinal as folhas somente podem ser coletadas nos períodos
de menor registro das fenofases.

52
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ABREU, I.N.D. Propagação In Vivo e In Vitro de Cissus sicyoides, uma planta
medicinal. Acta Amazônica, v.33, n.1, p. 1-7, Novembro de 2003.
ALEXANDRE, F. S. O.; SILVEIRA, E. R. Análise Fitoquímica de Plantas do Ceará:
potencial farmacológico de Cissus verticillata e composição volatil de Myrcia sp.2007.
159 f. Dissertação (Mestrado em Química Orgânica) - Centro de Ciências, Universidade
Federal do Ceará, Fortaleza, 2007.
BASTOS, T. X., PACHECO, N. A., NECHET, D., & Sá, T. D. A. Aspectos climáticos
de Belém nos últimos cem anos. Embrapa Amazônia Oriental-Documentos
(INFOTECA-E). 2002.
BERGAMASCHI, H. O clima como fator determinante da fenologia das plantas. In:
REGO, G.M.; NEGRELLE, R. B.; MORELLATO, L.P.C. (Orgs.). Fenologia como
ferramenta para conservação e manejo de recursos vegetais. Curitiba: EMBRAPA,
p. 291-310. 2007.
CARVALHO, F. R. de. A ecologia no cultivo de plantas medicinais. Revista
Agrogeoambiental, v. 4, n. 1, p. 85- 90. 2012.
COSTA, J.C.; MARINHO, M.G.V. Etnobotânica de plantas medicinais em duas
comunidades do município de Picuí, Paraíba, Brasil. Revista Brasileira de plantas
medicinais, Botucatu, v. 18, n. 1, p. 125-134, Março de 2016.
LAMEIRA, O. A.; PINTO, J. E. B. P. Plantas medicinais: do cultivo, manipulação e
uso à recomendação popular. Belém, PA: Embrapa Amazônia Oriental, p. 264, 2008.
MICROSOFT EXCEL versão 2010. Disponível em: <
http://pt.wikipedia.org/wiki/Microsoft_excel>
NEVES, E. L. das; FUNCH, L. S.; VIANA, B. F. Comportamento fenológico de três
espécies de Jatropha (Euphorbiaceae) da Caatinga, semi-árido do Brasil. Revista
Brasileira Botânica, v. 33, n. 1, p. 155-166, 2010.
SANTOS, H.B. et al. Avaliação do efeito hipoglicemiante de Cissus sicyoides em estudos
clínicos fase II. Revista Brasileira Farmacognosia. João Pessoa, v.18, n.1, p.70-76,
2008.
SEGANTINI, D, M., TORRES, L. ; BOLIANI ,A.C.; LEONEL,S. Fenologia da figueira
da índia em Selviria-MG.Revista brasileira de fruticultura, Jabuticabal – SP, v.32, n.2
, p. 630-636, 2010. Disponível em:.DOI: 10.1590/S0100-29452010005000049.

53
Belém (PA), 11 a 13 de dezembro de 2019
CARACTERÍSTICAS SOCIOECONÔMICAS E DE CONSUMO DOS
FREQUENTADORES DE FEIRAS LIVRES DO MUNICÍPIO DE MARABÁ-PA
André Gustavo Campinas Pereira1; Ana Caroline Palheta de Farias2; Laiane Pinto da
Silva3; Raimara Reis do Rosário4; Danielle do Socorro Nunes Campinas5.
1 Especialista em Geoprocessamento Aplicado à Agroecologia e ao Uso do Recursos
Naturais. Universidade Federal do Pará. [email protected]. 2 Graduanda em Agronomia. Universidade Federal Rural da Amazônia.
[email protected]. 3 Pós-graduanda em Geoprocessamento Aplicado à Agroecologia e ao Uso do Recursos
Naturais. Universidade Federal do Pará. [email protected]. 4 Graduanda em Agronomia. Universidade Federal Rural da Amazônia.
[email protected]. 5 Mestre em Agronomia. Universidade Federal Rural da Amazônia.
RESUMO
Neste estudo, objetivou-se compreender qual é a realidade do município de Marabá,
perante o comércio de hortaliças. Essas informações podem ser usadas como
direcionamentos na tomada de decisão de produtores no momento de escolher quais
culturas serão cultivadas para atender a demanda por produtos hortícolas do município
em questão. Para isso, utilizou-se um questionário semiestruturado, que teve como
objetivo identificar fatores sociais como gênero, faixa etária, nível de escolaridade e renda
mensal, bem como perguntas relacionadas ao hábito de consumo quando considerado os
tipos de hortaliças que costumam ser compradas. A amostragem utilizada foi do tipo
probabilística aleatória simples, sendo a população amostral composta por indivíduos
maiores que 18 anos, a que se teve acesso em pesquisa de campo. A delimitação da
amostra conferiu uma margem de erro de 10% e 90% de nível de confiança. Com base
nos resultados obtidos, foi observada a predominância de mulheres (52,7%), com mais de
sessenta anos (28,2%), possuindo nível médio de instrução (33,7%). Para a variável
correspondente a renda mensal, não houve diferença entre os entrevistados que recebem
até dois salários mínimos e entre três e cinco salários mínimos, totalizando 67,2% dos
entrevistados. Quando considerada a demanda de consumos, foi observada a
predominância pelo consumo do coentro (98,2%), cebolinha (95,5%), couve (95,2%) e
pimentinha verde (81,8%). A compreensão das características de consumo relacionada a
um centro consumidor pode proporcionar redução dos riscos referentes à comercialização
de produtos.
Palavras-chave: Perfil socioeconômico. Consumo de hortaliças. Estudo de mercado.
Área de Interesse do Simpósio: Agronomia.

54
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O mercado brasileiro de hortaliças apresenta uma grande diversificação de
produtos e contribui para a geração de emprego e renda de atividades ligadas direta e
indiretamente a essa cadeia produtiva, colaborando com a segurança alimentar da
população por oferecer alimentos saudáveis e seguros (JUBELINI, 2018).
As pessoas cada vez mais estão buscando uma alimentação saudável e
diversificada, elevando a procura por hortaliças (SILVEIRA et al., 2017). Estas são
encontradas em vários seguimentos de mercado, como sacolões, supermercados e
também nas feiras livres, que por sua vez, representa um dos métodos mais antigos de
comercialização de produtos agrícolas e tem por intuito o oferecimento de mercadorias
de boa qualidade e com preços mais baixos do que o comumente aplicado em
supermercados (SILVEIRA et al., 2017).
Quando avaliada a tendência de crescimento do setor de produção e
comercialização de hortaliças, observa-se que as diversificações de produtos aliados a
preços acessíveis tornam o investimento neste tipo de atividade, significativamente
atraente (SILVA; SANTOS; FERREIRA, 2017). Entretanto, devem-se considerar os
hábitos de alimentação da população de uma determinada região, pois características
como a quantidade consumida, tipo de produto, frequência de consumo e outros aspectos,
podem variar entre diferentes regiões (OLIVEIRA et al., 2018).
A compreensão acerca dos fatores inerentes as características do perfil e demanda
do público consumidor de produtos hortifrutigranjeiros, nas feiras livres, mostra-se como
importante ferramenta necessária para a construção do plano de negócios de produtores,
que obtém um melhor atendimento das demandas do mercado (ALVES; SOUZA;
PONTES, 2014; BRANDÃO et al, 2015).
As feiras livres são um dos principais meios de comercialização e distribuição de
hortaliças no município de Marabá. Este município está entre um dos cinco municípios
paraenses que mais contribuem com o PIB (Produto Interno Bruto) estadual, pertencendo
à região sudeste do estado do Pará, com uma população de aproximadamente 275.086 mil
habitantes (SOUSA et al., 2017). O objetivo deste trabalho foi caracterizar os aspectos
socioeconômicos dos consumidores de hortaliças do município de Marabá – PA.

55
Belém (PA), 11 a 13 de dezembro de 2019
2. METODOLOGIA
O estudo foi realizado na cidade de Marabá, sudeste do Estado do Pará, durante o
mês de outubro de 2018 em duas principais feiras de comercialização a nível local: Feira
da 28 e Feira da Laranjeira. O levantamento de informações para a pesquisa incluiu
visitação “in loco” e aplicação de formulários semi-estruturados (survey) aos
consumidores de hortaliças.
A amostragem utilizada foi do tipo probabilística aleatória simples, cuja amostra
era formada por indivíduos, maiores que 18 anos, a que se teve acesso em pesquisa de
campo. A delimitação da amostra foi realizada conforme metodologia descrita por
Barbeta (2008) e foi considerada a margem de erro de 10% e 90% de nível de confiança.
Foram aplicados 110 formulários que contemplavam questionamentos sociais
como gênero, faixa etária, nível de escolaridade e renda mensal, bem como perguntas
relacionadas ao hábito de consumo quando considerado os tipos de hortaliças que
costumam ser compradas.
Para a análise dos dados quantitativos utilizou-se estatística descritiva para
obtenção de médias e frequências relativas. O grau de importância dos itens citados em
cada questão foi definido considerando-se a classe modal, ou seja, a que mais vezes foi
citada no item, obtendo-se então a frequência relativa percentual.
3. RESULTADOS E DISCUSSÃO
Na figura 1 são apresentadas as informações referentes ao levantamento
socioeconômico dos consumidores de hortaliças que frequentam as feiras livres de
Marabá. Observou-se que as mulheres (52,7%) representaram a maioria do público
entrevistado. Resultados similares foram observados por Hungria et al. (2018), que
identificaram a predominância de mulheres (62,5%) em feiras livres de um distrito de
Belém do Pará.
Quando considerada a distribuição dos entrevistados por faixa etária, a maior
representatividade ocorreu para pessoas com mais de sessenta anos de idade (28,2%),
seguidas pelo público com idade entre dezoito e vinte e nove anos (23,6%). Esses dados
contrastam com as informações obtidas por Hungria et al. (2018), que identificaram para

56
Belém (PA), 11 a 13 de dezembro de 2019
estas mesmas faixas etárias, a presença de 31,7% e 6,7% dos entrevistados,
respectivamente.
Para a variável, nível de escolaridade, o destaque foi dado para pessoas (33,7%)
com nível médio de ensino. O mesmo comportamento das informações foi identificado
por Hungria et al. (2018), que relataram a presença da maioria do público entrevistado
com nível médio de formação (59,6%).
Com relação à distribuição de renda, observou-se que 67,2% dos entrevistados,
apresentam renda que varia entre dois a cinco salários mínimos. Esses resultados foram
superiores ao observado por Hungria et al. (2018), que identificaram a presença de 49,1%
da população estudada, recebendo apenas um salário mínimo.
Figura 1. Caracterização socioeconômica dos consumidores de hortaliças de Marabá – PA.
Fonte: Os autores.
Dentre os principais produtos consumidos em Marabá, podemos citar: coentro
(98,2%), cebolinha (95,5%), couve (95,2%) e pimentinha verde (81,8%) (Figura 2). Os
resultados encontrados neste estudo foram superiores quando comparados aos resultados
obtidos por Hungria et al. (2018), que identificaram a seguinte preferência de consumo:
coentro (79,8%), cebolinha (71,2%), couve (76,9%) e pimentinha verde (67,3%).
47,3%52,7%
23,6%
14,5%
5,5%2,2%
28,2%
13,6%
23,6%
33,7%
23,6%
5,5%0%
28,3%33,6%33,6%
4,5%
Masc
uli
no
Fem
inin
o
18 a
29 a
nos
30 a
39 a
nos
40 a
49 a
nos
50 a
59 a
nos
mai
s de
60 a
nos
Não
est
udou
Ensi
no F
undam
enta
l
Ensi
no M
édio
Ensi
no S
uper
ior
Pós-
Gra
duaç
ão
sem
ren
da
até
1 s
alár
io m
ínim
o
até
2 s
alár
ios
mín
imos
de
3 a
5 s
alár
ios
mín
imos
acim
a de
5 s
alár
ios
mín
imos
Gênero Faixa etária Nível de escolaridade Renda mensal

57
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2. Consumo de hortaliças.
Fonte: Os autores.
4. CONCLUSÃO
As informações aqui fornecidas tornam-se importantes para a melhor
entendimento do mercado hortícola nesse município, para que se possa subsidiar a
elaboração de planos de negócios que apresentem redução de riscos à comercialização de
produtos.

58
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
JUBELINI, G. Produção de frutas e hortaliças no Brasil destaca-se pela geração de
empregos. Revista Cultivar. Disponível em
<https://www.grupocultivar.com.br/noticias/producao-de-frutas-e-hortalicas-no-brasil-
destaca-se-pela-geracao-de-empregos>. Acesso em 20/10/2019.
OLIVEIRA, E. G. C.; PEREIRA, A. G. C.; CUNHA, T. S. N.; BRAGA, G. F.;
HUNGRIA, L. C.; CAMPINAS, D. S. N. Prospecção socioeconômica de
comercializadores de jambu em feiras livres de Belém. In: Jornada de Iniciação Científica
do Programa de Educação Tutorial do Pará, 14, 2018, Belém. Anais... Belém, UFRA,
2018.
SILVA, J. S. S.; SANTOS, M. A. S.; FERREIRA, C. S. S.; COSTA, J. F.; SOUZA, V.
C. Comportamento de preços de hortaliças folhosas na região metropolitana de Belém,
Estado do Pará. Enciclopédia Biosfera, Centro Científico Conhecer, Goiânia, v. 14 n. 26,
p. 2017.
SILVEIRA, V. C.; OLIVEIRA, E. S.; MARIANI, M. A.; SILVEIRA, N. F. Avaliação da
importância das feiras livres e a forma de comercialização adotada pelas feirantes na
cidade de Nova Andradina, MS.
ALVES, R. J. M.; SOUZA, L. P. F.; PONTES, A. N. Perfil socioeconômico e satisfação
dos consumidores de produtos hortifrutícolas do complexo do Ver-O-Peso, Pará.
Enciclopédia biosfera.v.10, n.19, 2014.
BRANDÃO, A. A.; COSTA, C. A.; GALIZONI, F. M.; CAVALCANTE, T. F. M.;
NEVES, A. C. Perfil socioeconômico dos consumidores de hortaliças em feiras livres na
microrregião de Januária. Horticultura Brasileira. v.33, n.1, 2015.
SOUSA, A. M. S.; OLIVEIRA, D. N.; ZEN, E. Z.; FERREIRA, M. L. O.; PEREIRA, R.
M. Economia regional: uma análise dos setores que compõe a economia da microrregião
de Marabá-PA. IN: Simpósio de Engenharia de Produção de Sergipe. 9. Sergipe. Anais...
2017. P. 568-581.
BARBETA, P. A. Estatística Aplicada às Ciências Sociais. 6. ed Florianópolis: Editora
da UFSC, 2008. 340p.
HUNGRIA, L. C.; PEREIRA, A. G. C.; VIANA, J. A. S.; DAVID, E. C.; SILVA, M. V.
S. O.; GUSMÃO, S. A. L. Aspectos socioeconômicos e de compra dos consumidores de
hortaliças em feiras livres da Ilha de Mosqueiro – PA. In: Encontro Amazônico de
Agrárias, 10, 2018, Belém. Anais... Belém, 2018.

59
Belém (PA), 11 a 13 de dezembro de 2019
EFEITO DA INOCULAÇÃO DE BACTÉRIAS PGPR EM ARROZ (Oryza sativa
L.)
Carmona, Marcio Augusto Costa Junior.1; Moraes, Alessandra Jackeline Guedes De2;
Machado, Ricardo Christin Lobato.3; Santos, Gleiciane Rodrigues Dos4; Silva, Gisele
Barata Da5
1 Graduando do curso de Agronomia. Universidade Federal Rural da Amazônia. E-mail:
[email protected] 2 Doutora na área de Fitopatologia. Universidade Federal Rural da Amazônia. E-mail:
[email protected] 3 Mestrando em Agronomia. Universidade Federal Rural da Amazônia. E-mail:
[email protected] 4 Doutorando em Agronomia. Universidade Federal Rural da Amazônia. E-mail:
[email protected] 5 Doutora na área de Fitopatologia. Universidade Federal Rural da Amazônia. E-mail:
RESUMO
O arroz é um dos cereais mais consumidos no mundo. Devido à sua grande importância
econômica, social e a intensa busca no campo pela sustentabilidade, objetivou-se estudar
a inoculação de isolados de bactérias em arroz com premissa na avaliação do seu
desenvolvimento e acúmulo de biomassa. O trabalho foi conduzido no Laboratório de
Proteção de Plantas, localizado na UFRA. As sementes de arroz, da cultivar BRS
Primavera, foram adquiridas na EMBRAPA Amazônia Oriental. Os isolados de bactérias
utilizados no ensaio de promoção de crescimento foram obtidos de rizosfera de açaizeiro,
os quais foram multiplicados em placa de Petri contendo meio de cultura 523 Kado e
Heskett por 48h. Após o crescimento das colônias, foi realizado a suspensão com água
destilada para cada isolado. O ensaio em DIC foi realizado com 10 tratamentos e 4
repetições. Inicialmente, as sementes foram semeadas em copos descartáveis de 200mL
e, após o período de 7 dias, contado a partir da semeadura, fez-se a aplicação da suspensão
bacteriana com 10mL por repetição. O presente trabalhou avaliou comprimento da parte
aérea (CPA), comprimento da raiz (CR), massa de matéria seca da parte aérea (MSPA) e
massa de matéria seca da raiz (MSR). Para CPA e MSPA, observou-se que os isolados
T02, T03, T04, T06 e T09 apresentaram os melhores resultados. Para CR, os isolados T02
e T06 apresentaram os melhores índices em relação aos demais tratamentos. Para MSR,
o isolado 06 apresentou os melhores resultados. Concluiu-se que os isolados T02, T03,
T04, T06 e T09 apresentaram resultados satisfatórios para os parâmetros avaliados neste
estudo (CPA, CR, MSPA e MSR). Os isolados T05, T07, T08 e T10 não diferiram
estatisticamente do tratamento controle.
Palavras-chave: Arroz Desenvolvimento. Bactérias Biomassa. Desenvolvimento
Sustentabilidade.
Área de interesse do Simpósio: Agronomia.

60
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O arroz é uma planta monocotiledônea, da ordem Poales, família Poaceae, do
gênero Oryza, possuindo em torno de 20 espécies, sendo a Oryza sativa L. a mais
cultivada. Este cereal possui grande valor nutricional, sendo altamente energético,
dispondo ainda de sais minerais e vitaminas do complexo B, além de suas características
possibilitarem uma diversidade de formas de preparo, como pratos salgados ou doces,
também podendo ser associado a outros alimentos, de forma a enriquecer ainda mais a
qualidade nutricional (CONAB, 2015), p.18).
O cultivo do arroz ocorre em dois tipos detanto em ecossistema, o de várzeas sob
inundação, como também e o de terras altas, comumente conhecido como arroz sequeiro,
estando entre um dos cereais mais consumidos no mundo, ocupando uma área total de
159 milhões de hectares, e tendo produção em cerca de 740 milhões de toneladas
(FAOSTAT, 2017), p. 118). O Brasil é o nono maior produtor mundial de arroz, tendo
todos os estados como fontes de cultivo. Na safra de 2018 a setembro de 2019, esta cultura
teve uma ocupação de área total em aproximadamente 1,7 milhões de hectares, tendo uma
produção na faixa de 10 milhões de toneladas (CONAB, 2019), p.16; FAOSTAT, 2017,
p. 118).
O estudo a respeito das interações ocorrentes entre plantas e microrganismos vem
se intensificando, com premissa em entender os vários fatores envolvidos na seleção de
estirpes de bactérias eficientes na promoção de crescimento. No arroz, a inoculação de
bioformulados à base de bactérias, seja na semente ou planta, vem mostrando eficácia na
promoção do crescimento, como o aumento do comprimento da raiz e da parte aérea e a
melhoria na biomassa, o que evidencia o aumento da absorção de nutrientes pela cultura
(FERREIRA et al., 2014;, p. 16; SHARMA, 2014), p. 519).
Em outras culturas, como o feijão-caupi (Vigna unguiculata L.) e lentilha (Lens
culinaris Medik) também se obtiveram melhorias no comprimento da raiz e da parte
aérea, além do aumento na absorção de nutrientes (MINAXI et al., 2012;, p. 130; ZAFAR
et al., 2012), p. 854). Estudos afirmam que as bactérias promotoras de crescimento de
plantas fazem parte da população residente das plantas epifíticas ou endofiticas e não são
fitopatogênicas (MARIANO et al., 2004), p.90).
Devido à importância econômica e social do arroz, dentro do contexto da
necessidade de maior produtividade por área desta cultura, com enfoque na

61
Belém (PA), 11 a 13 de dezembro de 2019
sustentabilidade agrícola, torna-se relevante a avaliação do efeito de bactérias promotores
de crescimento vegetal (BPCV) inoculadas a sementes de arroz, em sua fase inicial de
desenvolvimento.
2. METODOLOGIA
O trabalho foi desenvolvido no Laboratório de Proteção de Plantas (LPP) do
Instituto de Ciências Agrárias (ICA), localizado na Universidade Federal Rural da
Amazônia (UFRA). As sementes de arroz (Oryza sativa L.), da cultivar BRS Primavera
foram adquiridas na EMBRAPA Amazônia Oriental, no município de Belém, Pará. Os
isolados utilizados no ensaio de promoção de crescimento foram obtidos de rizosfera de
açaizeiro (Euterpe oleracea) de um plantio comercial localizado no município de Tomé
Açu, Pará. Após a obtenção, os isolados foram purificados e adicionados à coleção de
microrganismos do LPP.
Os isolados foram multiplicados em placa de Petri contendo meio de cultura 523
Kado e Heskett (1970) por 48 h, em temperatura ambiente. Após o crescimento das
colônias foi realizado a suspensão com água destilada para cada isolado, ajustado a uma
concentração de 1,0 X 108 UFC.mL-1, utilizando-se espectrofotômetro ajustado a 550 mm
de absorbância (A550 = 0,1).
As sementes foram semeadas em copos descartáveis de 200 mL contendo terra
preta peneirada. Aos 7 dias, contados a partir da data de semeadura, fez-se a aplicação
dos isolados, via rega, em uma quantidade de 10 mL por repetição. O tratamento controle
recebeu apenas água destilada.
O presente trabalho aAvaliou-se o comprimento da parte aérea (CPA) e o
comprimento da raiz (CR), com o auxílio de uma régua graduada, a massa de matéria
seca da parte aérea (MSPA) e a massa de matéria seca da raiz (MSR), feitas com o auxílio
de estufa de ar forçado a 50ºC por 96h, posteriormente, pesadas com o auxílio de balança
semi-analítica.
O experimento foi realizado em delineamento inteiramente casualizado com dez
tratamentos, sendo nove isolados de rizobactérias (T02; T03; T04; T05; T06; T07; T08;
T09; T10) e o controle (sem rhizobacteria) e quatro repetições. Os dados de comprimento
da parte aérea (CPA), comprimento da raiz (CR), massa seca da parte aérea (MSPA) e
massa seca da raiz (MSR) foram submetidos à análise de agrupamento usando os valores

62
Belém (PA), 11 a 13 de dezembro de 2019
médios de cada indivíduo e o grau de similaridade foi obtido pela distância Euclidiana
padrão. Os grupos foram submetidas à análise de variância (ANOVA) e as médias foram
comparadas pelo teste Student-Newman-Keuls (SNK), P < 0,05, usando o software R
versão 3.1.
3. RESULTADOS E DISCUSSÃO
O efeito da inoculação de bactérias promotoras de crescimento vegetal, na cultura
do arroz, sobre os dados de comprimento da parte aérea (CPA), comprimento da raiz
(CR), massa seca da parte aérea (MSPA) e massa seca da raiz (MSR) foram relevantes ao
nível de 5% de significância. Os tratamentos foram divididos em quatro grupos, pelo
dendograma de similaridade, de acordo com a associação das médias de todos os dados,
separados desde os melhores tratamentos, como os isolados T02 e T06, até os tratamentos
semelhantes ao controle (identificado por T01), sendo os isolados T10, T08, T05, T07
(Figura 1).
Figura 1 – Dendograma de similaridade construído com base nas características fisiológicas das
estirpes de BPCP obtidos de rizosfera de açaizeiro (Euterpe oleracea).
Para o comprimento da parte aérea (CPA), observou-se que isolados T03, T04 e
T09 (grupo 3) apresentaram os maiores índices de parte aérea em relação aos demais
tratamentos (Figura 2). Resultados semelhantes foram encontrados em arroz vermelho
(Oryza Sativa L.), genótipo 405 Embrapa Meio Norte, quando inoculados à
Gluconacetobacter diazotrophicus, as quais apresentaram resultados satisfatórios para o
comprimento da parte aérea em comparação à testemunha (SILVA FILHO, 2016), p. 40).

63
Belém (PA), 11 a 13 de dezembro de 2019
Silva Filho (2016), p. 42), reportou em seu trabalho que bactérias promotoras de
crescimento induzem a transformação do nitrogênio em amônia solúvel em água, que é
utilizado diretamente pela planta, quando ocorre o processo de fixação biológica do
nitrogênio.
Os isolados dos grupos 3 (T03, T04 e T09) apresentaram maior crescimento da
parte aérea em comparação ao tratamento controle. Entretanto, os isolados T05, T07, T08
e T10 não diferiram estatisticamente do tratamento controle. Supõe-se que, devido à
característica de elevada produção de auxina por algumas bactérias, possa ter inibido o
crescimento da parte aérea das plantas respectivas aos tratamentos do grupo 4.
Estudos realizados em arroz irrigado (Oryza sativa L.), e Lactuca sativa afirmam
que a inoculação de rizobactérias de efeito inibidor possuem capacidade de produção, em
excesso, de auxinas, como o ácido indolacético (AIA), as quais provocam uma redução
no crescimento da parte aérea e/ou da raiz; (SARWAR & KREMER, 1995, p. 265;
BARANZANI & FRIEDMAN, 1999, p. 2402).
Figura 2 – Comprimento da parte aérea (CPA) de arroz submetidas a diferentes aplicações de
bactérias rizosféricas. Médias seguidas pela mesma letra não diferem estatisticamente entre si
pelo teste SNK, (p < 0,05). (CPA – CV% = 6,48))
Para o comprimento da raiz (CR), os isolados T02 e T06 não diferiram
estatisticamente, apresentando os melhores resultados para este parâmetro (Figura 3).
Supõe-se que o sinergismo presente entre os mecanismos de ação das bactérias
promotores de crescimento, como solubilização de fosfatos inorgânicos e incrementos na
atividade do óxido nítrico, resultam em maior desenvolvimento radicular (GUIMARÃES,
2018), p. 196).
Trabalhos realizados em cultivares de arroz (Oryza Sativa L.), mostraram que os
tratamentos à base de inoculação de estirpes de bactérias apresentaram valores de

64
Belém (PA), 11 a 13 de dezembro de 2019
comprimento radicular superiores aos observados no tratamento controle. Resultados
semelhantes também foram encontrados na cana-de-açúcar (Saccharum officinarum L.),
em que o comprimento radicular nos tratamentos inoculados foi maior do que nos
tratamentos não inoculados (SOUSA et al., 2019;, p. 143; FERREIRA, 2014), p. 658).
Os isolados T03, T04 e T09 não diferiram estatisticamente entre si. O isolados
T05, T07, T08 e T10 não diferiram estatisticamente do tratamento controle, apresentando
os resultados menos satisfatórios em relação aos demais grupos de isolados (Figura 3).
Figura 3 – Comprimento da raiz (CR) de arroz submetidas a diferentes aplicações de bactérias
rizosféricas. Médias seguidas pela mesma letra não diferem estatisticamente entre si pelo teste
SNK, (p < 0,05). (CR – CV% = 9,56)
Para a massa de matéria seca da parte aérea (MSPA), os isolados T02, T06, T03,
T04 e T09 apresentaram resultados satisfatórios em relação aos demais tratamentos
(Figura 4). Trabalhos realizados em cana-de-açúcar (Saccharum officinarum) mostraram
que a inoculação de rizobactérias resultaram em maior massa de matéria seca das raízes
e da parte aérea. Em arroz vermelho (Oryza sativa L.), resultados semelhantes foram
encontrados para a massa seca da parte aérea, quando inoculadas a isolados de bactérias
promotoras de crescimento. (GIRIO et al., 2015;, p. 39; DUTRA et al., 2014), p. 96)
Estudos realizados em sementes de soja (Glycine max L.) levaram à suposição de
que o ganho de biomassa pode estar associado à capacidade dos isolados em produzir
ácido indolacético (AIA) AIA e solubilizar fosfato (ASSUMPÇÃO et al., 2009), p. 507).
O isolados T05, T07, T08 e T10 não diferiram estatisticamente do tratamento
controle (figura 4). Resultados semelhantes foram encontrados em meloeiro (Cucumis
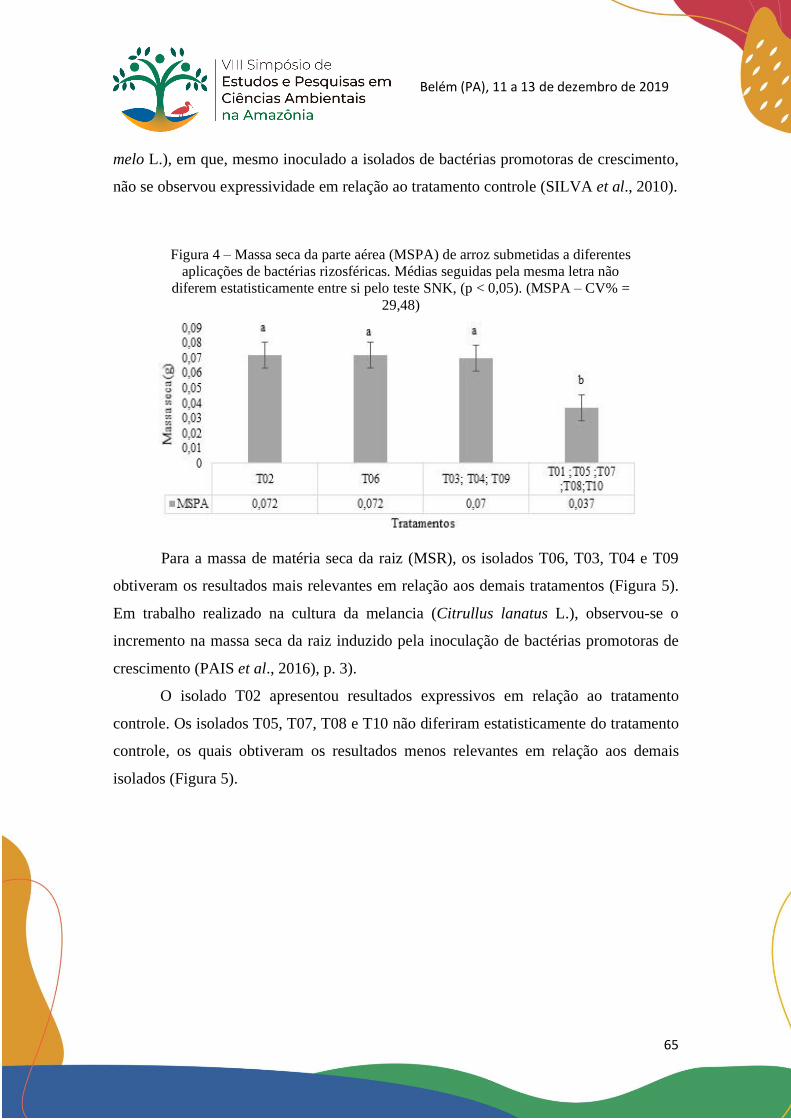
65
Belém (PA), 11 a 13 de dezembro de 2019
melo L.), em que, mesmo inoculado a isolados de bactérias promotoras de crescimento,
não se observou expressividade em relação ao tratamento controle (SILVA et al., 2010).
Figura 4 – Massa seca da parte aérea (MSPA) de arroz submetidas a diferentes
aplicações de bactérias rizosféricas. Médias seguidas pela mesma letra não
diferem estatisticamente entre si pelo teste SNK, (p < 0,05). (MSPA – CV% =
29,48)
Para a massa de matéria seca da raiz (MSR), os isolados T06, T03, T04 e T09
obtiveram os resultados mais relevantes em relação aos demais tratamentos (Figura 5).
Em trabalho realizado na cultura da melancia (Citrullus lanatus L.), observou-se o
incremento na massa seca da raiz induzido pela inoculação de bactérias promotoras de
crescimento (PAIS et al., 2016), p. 3).
O isolado T02 apresentou resultados expressivos em relação ao tratamento
controle. Os isolados T05, T07, T08 e T10 não diferiram estatisticamente do tratamento
controle, os quais obtiveram os resultados menos relevantes em relação aos demais
isolados (Figura 5).

66
Belém (PA), 11 a 13 de dezembro de 2019
Figura 5 – Massa seca da raiz (MSR) de arroz submetidas a diferentes aplicações de bactérias
rizosféricas. Médias seguidas pela mesma letra não diferem estatisticamente entre si pelo teste
SNK, (p < 0,05). (MSPA – CV% = 29,83)
O efeito da inoculação de bactérias promotoras de crescimento vegetal, na cultura
do arroz, sobre os dados de comprimento da parte aérea (CPA), comprimento da raiz
(CR), massa seca da parte aérea (MSPA) e massa seca da raiz (MSR) foram relevantes ao
nível de 5% de significância. Os tratamentos foram divididos em quatro grupos, pelo
dendograma de similaridade, de acordo com a associação das médias de todos os dados,
separados desde os melhores tratamentos, como os isolados T02 e T06, até os tratamentos
semelhantes ao controle (identificado por T01), sendo os isolados T10, T08, T05, T07
(figura Figura 01).
Para o comprimento da parte aérea (CPA), observou-se que isolados T03, T04 e
T09 (grupo 3) apresentaram os maiores índices de parte aérea em relação aos demais
tratamentos (Figura 2). Resultados semelhantes foram encontrados em arroz vermelho
(Oryza Sativa L.), genótipo 405 Embrapa Meio Norte, quando inoculados à
Gluconacetobacter diazotrophicus, as quais apresentaram resultados satisfatórios para o
comprimento da parte aérea em comparação à testemunha (SILVA FILHO, 2016, p. 40).
Silva Filho (2016, p. 42), reportou em seu trabalho que bactérias promotoras de
crescimento induzem a transformação do nitrogênio em amônia solúvel em água, que é
utilizado diretamente pela planta, quando ocorre o processo de fixação biológica do
nitrogênio.
Os isolados dos grupos 3 (T03, T04 e T09) apresentaram maior crescimento da parte aérea
em comparação ao tratamento controle. Entretanto, os isolados T05, T07, T08 e T10 não
diferiram estatisticamente do tratamento controle. Supõe-se que, devido à característica

67
Belém (PA), 11 a 13 de dezembro de 2019
de elevada produção de auxina por algumas bactérias, possa ter inibido o crescimento da
parte aérea das plantas respectivas aos tratamentos do grupo 4.
Estudos realizados em arroz irrigado (Oryza sativa L.), e Lactuca sativa afirmam
que a inoculação de rizobactérias de efeito inibidor possuem capacidade de produção, em
excesso, de auxinas, como o ácido indolacético (AIA), as quais provocam uma redução
no crescimento da parte aérea e/ou da raiz; (SARWAR & KREMER, 1995, p. 265;
BARANZANI & FRIEDMAN, 1999, p. 2402).
Para o comprimento da raiz (CR), os isolados T02 e T06 não diferiram
estatisticamente, apresentando os melhores resultados para este parâmetro (figura Figura
3). Supõe-se que o sinergismo presente entre os mecanismos de ação das bactérias
promotores de crescimento, como solubilização de fosfatos inorgânicos e incrementos na
atividade do óxido nítrico, resultam em maior desenvolvimento radicular (GUIMARÃES,
2018, p. 196).
Trabalhos realizados em cultivares de arroz (Oryza Sativa L.), mostraram que os
tratamentos à base de inoculação de estirpes de bactérias apresentaram valores de
comprimento radicular superiores aos observados no tratamento controle. Resultados
semelhantes também foram encontrados na cana-de-açúcar (Saccharum officinarum), em
que o comprimento radicular nos tratamentos inoculados foi maior do que nos tratamentos
não inoculados (SOUSA et al., 2019, p. 143; FERREIRA, 2014, p. 658).
Os isolados T03, T04 e T09 não diferiram estatisticamente entre si (figura Figura
3). O isolados T05, T07, T08 e T10 não diferiram estatisticamente do tratamento controle,
apresentando os resultados menos satisfatórios em relação aos demais grupos de isolados
(Figura 3).
Para a massa de matéria seca da parte aérea (MSPA), os isolados T02, T06, T03,
T04 e T09 apresentaram resultados satisfatórios em relação aos demais tratamentos
(figura Figura 4). Trabalhos realizados em cana-de-açúcar (Saccharum officinarum)
mostraram que a inoculação de rizobactérias resultaram em maior massa de matéria seca
das raízes e da parte aérea. Em arroz vermelho (Oryza sativa L.), resultados semelhantes
foram encontrados para a massa seca da parte aérea, quando inoculadas a isolados de
bactérias promotoras de crescimento. (GIRIO, et al., 2015, p. 39; DUTRA et al., 2014, p.
96)

68
Belém (PA), 11 a 13 de dezembro de 2019
Estudos realizados em sementes de soja (Glycine max) levaram à suposição de que o
ganho de biomassa pode estar associado à capacidade dos isolados em produzir AIA e
solubilizar fosfato (ASSUMPÇÃO et al., 2009, p. 507).
O isolados T05, T07, T08 e T10 não diferiram estatisticamente do tratamento
controle (figura 4). Resultados semelhantes foram encontrados em meloeiro (Cucumis
melo), em que, mesmo inoculado a isolados de bactérias promotoras de crescimento, não
se observou expressividade em relação ao tratamento controle (SILVA et al., 2010, p.
1121).
Para a massa de matéria seca da raiz (MSR), os isolados T06, T03, T04 e T09
obtiveram os resultados mais relevantes em relação aos demais tratamentos (figura Figura
5). Em trabalho realizado na cultura da melancia (Citrullus lanatus), observou-se o
incremento na massa seca da raiz induzido pela inoculação de bactérias promotoras de
crescimento (PAIS et al., 2016, p. 3).
O isolado T02 apresentou resultados expressivos em relação ao tratamento
controle (figura Figura 5). Os isolados T05, T07, T08 e T10 não diferiram
estatisticamente do tratamento controle, os quais obtiveram os resultados menos
relevantes em relação aos demais isolados (figura Figura 5).
4. CONCLUSÃOÕES
Os isolados T03, T04 e T09 apresentaram os melhores resultados para o
comprimento da parte aérea. Para o comprimento da raiz, os isolados T02 e T06
apresentaram os melhores índices em relação aos demais tratamentos. Os isolados T02,
T03, T04, T06 e T09 não mostraram diferenças significativas entre si, apresentando os
melhores índices para a massa de matéria seca da parte aérea. Para a massa de matéria
seca da raiz, o isolado T06 apresentou os melhores resultados. O grupo quatro, composto
pelos isolados T05, T07, T08, T10 e o tratamento controle apresentaram os resultados
menos satisfatórios em relação aos demais tratamentos.

69
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ASSUMPÇÃO L. C.; LACAVA P. T.; DIAS A. C. F.; AZEVEDO J. L.; MENTEN J. O.
M. Diversidade e potencial biotecnológico da comunidade bacteriana endofítica de
sementes de soja. Pesquisa Agropecuária Brasileira, v. 44, n. 5, p. 503 – 510, maio,
2009.
BARANZANI, O.; FRIEDMAN, J. Is IAA the major root growth factor secreted from
plant-growth-mediating bacteria. Journal of Chemical Ecology, v. 25, n. 10, p. 2397 –
2406, 1999.
COMPANHIA NACIONAL DE ABASTECIMENTO – CONAB. Acompanhamento da
Safra Brasileira de Grãos. Setembro de 2019.
COMPANHIA NACIONAL DE ABASTECIMENTO – CONAB – A cultura do arroz.
2015.
DUTRA, L. M. F.; BARBOSA, F. M.; MELO, A. S.; FERNANDES, P. D.; VIDAL, M.
S.; BALDANI, J. I.; MENESES, C. H. S. G. Inoculação de Gluconacetobacter
diazotrophicus e seu efeito no desenvolvimento de plantas de arroz vermelho. Journal of
Biology & Pharmacy and Agricultural Management, v. 10, n. 4, 2014.
FERREIRA, E. P. B.; KNUPP, A. M.; MARTIN-DIDONET, C. C. G. Crescimento de
cultivares de arroz (Oryza sativa L.) influenciado pela inoculação com bactérias
promotoras de crescimento de plantas. Bioscience Journal, Uberlândia, v. 30, n. 3, p.
655 – 665, maio/junho, 2014.
FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS -
FAOSTAT. Production of cereals and share in world. 2017.
GIRIO, L. A. S.; DIAS, F. L. F.; REIS, V. M.; URQUIAGA, S.; SCHULT, N.;
BOLONHEZI, D.; MUTTON, M. A. Bactérias promotoras de crescimento e adubação
nitrogenada no crescimento inicial de cana-de-açúcar proveniente de mudas pré-brotadas.
Pesquisa agropecuária brasileira. v. 50, n. 1, p. 33 – 43, 2015.
GUIMARÃES, F. V.; BATTISTUS, A. G.; SOUZA, A. K. P.; BULEGON, L. G.;
OFFEMANN, L. C.; ANAGAKI, A. M. Bactérias Promotoras de Crescimento Vegetal:
da FBN à regulação hormonal, possibilitando novas aplicações. CIÊNCIAS
AGRÁRIAS: ética do cuidado, legislação e tecnologia na agropecuária. Cap. 11, 192
– 196, 2018.
MARIANO, R. L. R; SILVEIRA, E. B.; ASSIS, S. M. P.; GOMES, A. M. A.;
NASCIMENTO, A. R. P.; DONATO, V. M. T. S. Importância de bactérias promotoras
de crescimento e de biocontrole de doenças de plantas para uma agricultura sustentável.
Anais da Academia Pernambucana de Ciência Agronômica, Recife, v. 1, p.89 – 111,
2004.

70
Belém (PA), 11 a 13 de dezembro de 2019
MINAXI, NAIN, L.; YADAV, R.C.; SAXENA, J. Characterization of multifaceted
Bacillus sp. RM-2 for its use as plant growth promoting bioinoculant for crops grown in
semi arid deserts. Applied Soil Ecology, 59, 124 – 135, 2012.
PAIS, A. K. L.; SILVA, J. R.; ALENCAR, F. C.; PEIXOTO, A. R.; SOUZA J. C.; PAZ,
C. D. Seleção de rizobactérias como promotoras de crescimento em melancia. Scientia
Plena, v. 12, 2016.
SARWAR, M.; KREMER, R. Enhanced suppression of plant growth through production
of L-tryptophan-derived compounds by deleterious rhizobacteria. Plant and Soil, v. 172,
n. 2, p. 261 – 269, 1995.
SHARMA, A.; SHANKHDHAR, D. Growth promotion of the rice genotypes by pgprs
isolated from rice rhizosphere. Journal of Soil Science and Plant Nutrition, v. 14, n. 2,
505 – 517, 2014.
SILVA J. R.; KARASAWA M.; CARVALHO L. N. J.; PAZ C. D.; OLIVEIRA E. A. S.
Utilização de bactérias na promoção de crescimento de meloeiro através da bacterização
de sementes e substrato. Horticultura Brasileira, v. 28, p. 1119 – 1122, 2010
SOUZA, I. M.; NASCENTE, A. S.; FILIPPI, M. C. C. Bactérias promotoras do
crescimento radicular em plântulas de dois cultivares de arroz irrigado por inundação.
Colloquium Agrariae, v. 15, n.2, p. 140 – 145, março-abril 2019
ZAFAR, M., ABBASI, M.K., KHAN, M.A., KHALIQ, A., SULTAN, T., ASLAM, M.
Effect of plant growthpromoting rhizobacteria on growth, nodulation an nutrient
accumulation of lentil under controlled conditions. Pedosphere, 2012 22, 848 – 859.

71
Belém (PA), 11 a 13 de dezembro de 2019
EFEITO DO PASTEJO NAS PROPRIEDADES QUÍMICAS DO SOLO NO
PROJETO DE ASSENTAMENTO SÃO FRANCISCO, ELDORADO DOS
CARAJÁS, PA
Kelvis Nunes da Silva1; Eduarda Costa Ferreira2; Vanessa Silva Oliveira2; Nathalia
Ingrid Gomes Machado2; Gleidson Marques Pereira3.
1 Graduando em Engenharia Ambiental. Universidade do Estado do Pará.
[email protected] 2 Graduandos em Engenharia Ambiental. Universidade do Estado do Pará.
3Mestre em Agronomia – Solos e Nutrição de Plantas. Universidade do Estado do Pará;
RESUMO
O uso incorreto dos solos altera diretamente suas características físicas, químicas e
biológicas, resultado do desmatamento, queimadas, expansão da pecuária, compactação
e erosão dos solos. O presente estudo tem como objetivo avaliar as alterações nas
propriedades químicas do solo sob pastagem (Brachiaria brizantha) em distintas
condições de manejo. O estudo foi realizado no lote 102 do Projeto de Assentamento São
Francisco, no município de Eldorado dos Carajás (PA). O solo local foi classificado de
acordo com a Embrapa (2007) como Argissolo Vermelho-Amarelo Distrófico com
textura média/argilosa. Para a coleta das amostras de solo, utilizou-se o inteiramente
casualizado, com total de 12 amostras compostas e dividas entre áreas de pastagens de 1
ano, 2 anos e 30 anos sem tratamento. O resultado mostrou que o teor de matéria orgânica
do solo (MOS) bem como o (N) no pasto de 30 anos sem tratamento, obteve média bem
superior em relação aos demais tratamentos (1 ano e 2 anos), enquanto que os
macronutrientes (P) e (K) apresentaram coeficientes mais baixos em relação aos demais
atributos. Os diferentes manejos nos sistemas pastoris alteraram as propriedades químicas
do solo quando comparados. Os sistemas com pastagens que sofreram reformas em
atividade mantiveram seus atributos químicos para os elementos N, P, K e teores de MO
próximos nas médias observados e sem diferenças estatísticas significativas. As
concentrações dos atributos químicos colaboraram para a avaliação da qualidade do solo,
subsidiando a comprovação de manejos de solo mais sustentáveis.
Palavras-chave: Degradação. Pastagem. Nutrientes do solo.
Área de Interesse do Simpósio: Agronomia

72
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Entre as ações antrópicas negativas sobre o meio ambiente, a degradação do solo
constitui uma das mais preocupantes, visto que tem afetado, diretamente, a vida do
homem. A principal causa da degradação resulta do mau uso do solo, tendo como
consequência a redução da matéria orgânica e, por conseguinte, alterações nas
características físicas, químicas e biológicas do solo (JAKELAITIS et al. 2008).
A degradação do solo decorre do seu uso de maneira incorreta, resultado do
desmatamento, queimada, negligência no manejo da pastagem, compactação e erosão
(CUNHA et al., 2008 apud NEVES NETO et al., 2013). A expansão da pecuária é a
principal causa do acelerado processo de desmatamento na região Amazônica
(MATTEDI, 2007 apud SCHLICKMANN; SCHAUMAN, 2007).
Segundo Rivero et al. (2009), baseado nos estudos de Pikkety et al. (2003) e
Rodrigues (2004), no histórico recente da ocupação da Amazônia, a atividade pecuária
tem se expandido continuamente, o que torna a criação de gado a atividade econômica de
maior impacto em toda região.
Desse modo, as práticas da atividade pecuária são consideradas importantes para
a perda dos nutrientes no solo e matéria orgânica. Estudos relacionados às alterações no
solo podem fornecer subsídios para reduzir os impactos causados pela pecuária na Região
Amazônica. Objetivou-se, neste trabalho, avaliar as alterações nas propriedades químicas
do solo sob pastagem (Brachiaria brizantha) em distintas condições de manejo no
Assentamento São Francisco, no município de Eldorado dos Carajás (PA).
2. MATERIAL E MÉTODOS
O estudo foi realizado no Lote 102 do Projeto de Assentamento São Francisco, no
Município de Eldorado dos Carajás (PA), localizado à latitude 05º55’21” (S), longitude
49º11’52” (W) (Figura 01) e altitude média local de 140 m. De acordo com a classificação
de Koppen, a região apresenta clima do tipo quente e úmido com temperatura média anual
de 26,5ºC, e a precipitação média anual de aproximadamente 1.757 mm. O solo do local
foi classificado como Argissolo Vermelho-Amarelo Distrófico com textura argilosa, de
acordo com a metodologia da Embrapa (2007).

73
Belém (PA), 11 a 13 de dezembro de 2019
Figura 01 - Localização da área escolhida para a realização das coletas de amostras de solo.
Fonte: Autores, 2019
O Projeto de Assentamento São Francisco tem como aptidão agrícola a banana
(Musa sapiento) e o cupuaçu (Theobroma grandiflorum) como culturas perenes de
principal importância respectivamente no PA. Porém, a pecuária também é um sistema
agrícola de muita importância no local abrangendo a criação em pastejo de animais de
grande e médio porte com solo caracterizado com relevo ondulado e solo bom e
moderadamente drenado e textura argilosa, admitindo-se também a presença e cascalho
pedregosidade ao longo do perfil (SILVA; SILVA; SAMPAIO, 2002).
Para a coleta das amostras de solo, o delineamento utilizou-se o inteiramente
casualizado, com quatro parcelas (repetições) em cada sistema de manejo com 30 anos
sem reforma, 1 ano sem reforma e 2 anos sem reforma respectivamente, onde os mesmos
foram classificados pelo período em que apresentam ausência de adubação e de
mecanização. Dentro de cada área de sistema de uso da terra realizou-se amostragem, por
meio da coleta com auxílio do trado, coletou-se uma amostra na profundidade 0-5 cm.
Coletou-se 20 sub-amostras aleatórias dentro de cada área de tratamento, perfazendo uma
amostra composta, isto é, uma amostra composta por parcela, esta última escolheu-se
através de sorteio (casualização). Ao serem coletadas, as amostras foram imediatamente

74
Belém (PA), 11 a 13 de dezembro de 2019
acondicionadas em sacos plásticos e identificadas. No total coletaram-se 12 amostras, as
quais submeteram-se às análises de parâmetros químicos do solo no Laboratório de Solos
da Embrapa Amazônia Oriental.
Nas amostras coletadas determinou-se o teor de matéria orgânica (MO),
determinado pelo método de Walkey & Black, além dos componentes totais Nitrogênio
(N), Fósforo (P) e Potássio (K) conforme metodologia empregada pela EMBRAPA
(1997). Os dados foram submetidos à análise de variância, e, quando significativos,
analisados pelo teste de Tukey (p <0,05) utilizando o programa SISVAR 5.6.86
(FERREIRA, 2014). Além disso, realizou-se análise de correlação de Pearson em função
do tempo de reforma de pasto e os atributos químicos do solo adotado no trabalho.
3. RESULTADOS E DISCUSSÃO
Em relação ao teor médio de matéria orgânica, as análises demonstraram (Tabela
01) que o pasto mais antigo (30 anos sem reforma) obteve média de teor de matéria
orgânica bem superior (57,035), em relação aos demais, que obtiveram valores bem
próximos (42,420 e 40,69, respectivamente: pasto de 1 ano sem reforma e 2 anos sem
reforma) e pelo teste de Tukey o pasto de 30 anos diferem entre os outros tratamentos.
Isso concorda com os estudos realizados também na Região Amazônica com a mesma
classificação de solo por Muller et al. (2001), que não observou diferenças importantes
de teor de matéria orgânica, na camada superficial do solo, entre as pastagens com tempos
próximos de reforma.
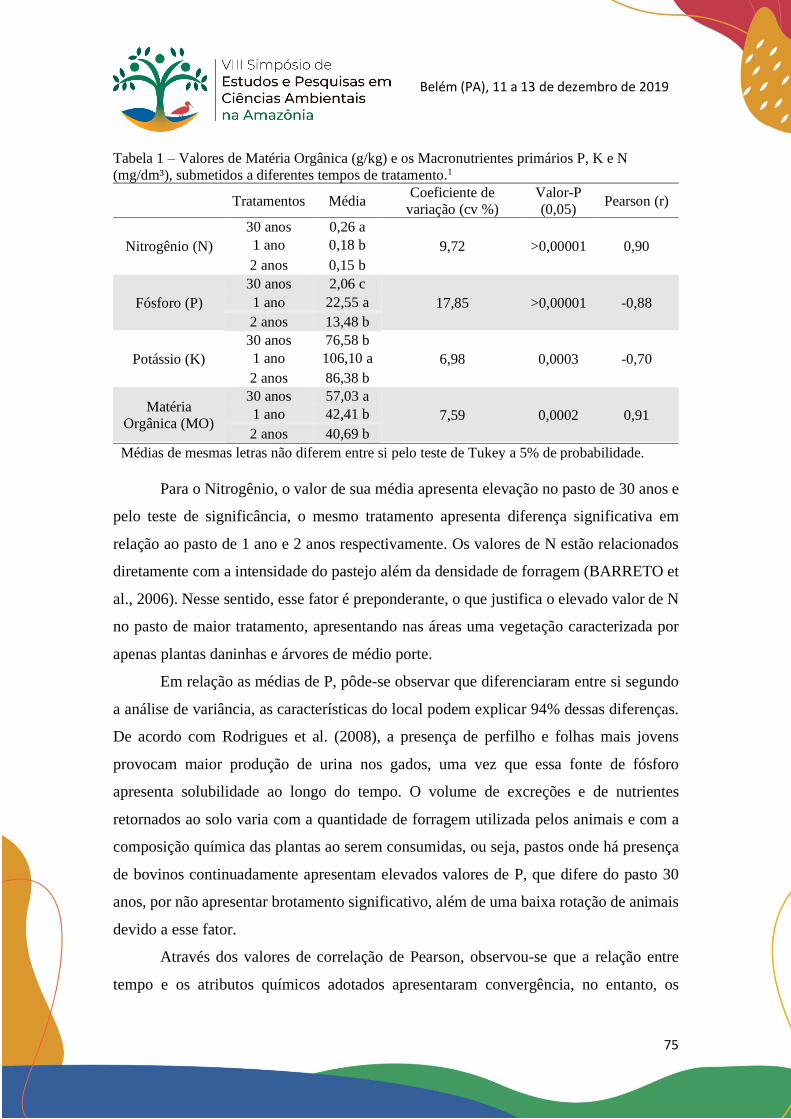
75
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1 – Valores de Matéria Orgânica (g/kg) e os Macronutrientes primários P, K e N
(mg/dm³), submetidos a diferentes tempos de tratamento.1
Tratamentos Média Coeficiente de
variação (cv %)
Valor-P
(0,05) Pearson (r)
Nitrogênio (N)
30 anos 0,26 a
9,72 >0,00001 0,90 1 ano 0,18 b
2 anos 0,15 b
Fósforo (P)
30 anos 2,06 c
17,85 >0,00001 -0,88 1 ano 22,55 a
2 anos 13,48 b
Potássio (K)
30 anos 76,58 b
6,98 0,0003 -0,70 1 ano 106,10 a
2 anos 86,38 b
Matéria
Orgânica (MO)
30 anos 57,03 a
7,59 0,0002 0,91 1 ano 42,41 b
2 anos 40,69 b
Médias de mesmas letras não diferem entre si pelo teste de Tukey a 5% de probabilidade.
Para o Nitrogênio, o valor de sua média apresenta elevação no pasto de 30 anos e
pelo teste de significância, o mesmo tratamento apresenta diferença significativa em
relação ao pasto de 1 ano e 2 anos respectivamente. Os valores de N estão relacionados
diretamente com a intensidade do pastejo além da densidade de forragem (BARRETO et
al., 2006). Nesse sentido, esse fator é preponderante, o que justifica o elevado valor de N
no pasto de maior tratamento, apresentando nas áreas uma vegetação caracterizada por
apenas plantas daninhas e árvores de médio porte.
Em relação as médias de P, pôde-se observar que diferenciaram entre si segundo
a análise de variância, as características do local podem explicar 94% dessas diferenças.
De acordo com Rodrigues et al. (2008), a presença de perfilho e folhas mais jovens
provocam maior produção de urina nos gados, uma vez que essa fonte de fósforo
apresenta solubilidade ao longo do tempo. O volume de excreções e de nutrientes
retornados ao solo varia com a quantidade de forragem utilizada pelos animais e com a
composição química das plantas ao serem consumidas, ou seja, pastos onde há presença
de bovinos continuadamente apresentam elevados valores de P, que difere do pasto 30
anos, por não apresentar brotamento significativo, além de uma baixa rotação de animais
devido a esse fator.
Através dos valores de correlação de Pearson, observou-se que a relação entre
tempo e os atributos químicos adotados apresentaram convergência, no entanto, os

76
Belém (PA), 11 a 13 de dezembro de 2019
macronutrientes P e K apresentam coeficientes mais baixos em comparação com os
demais atributos.
À medida que o pasto não apresenta uma manutenção, os teores citados tendem a
diminuir, isso ocorre devido à alta procura dos animais para com a vegetação que é fonte
de consumo desses nutrientes, diminuindo o estoque dos mesmos no solo.
Além disso, são atributos solúveis e consequentemente arrastados e consumidos
por espécies consideradas invasoras no pasto, ocasionando na diminuição desses
nutrientes e consequentemente a eficiência do pasto.
A concentração de K diverge dos demais nutrientes, pois apresentou significância
com a maior média (106,10) no talhão de 1 ano, e diferindo das demais áreas pelo teste
de Tukey. O K+ possui facilidade em ser lixiviado segundo os autores Lima; Souza; Silva
(2010) que em seu estudo de variabilidade espacial de atributos químicos do solo,
observou que o potássio se encontra em áreas mais baixas, isso é justificado devido ao
arraste superficial pela água da chuva, dos locais altos para locais baixos, sendo
caracterizado como ambiente de deposição.
4. CONCLUSÃO
Os diferentes manejos nos sistemas pastoris alteraram as propriedades químicas
do solo quando comparados. Os sistemas com pastagens que sofreram reformas em
atividade mantiveram seus atributos químicos para os elementos N, P, K e teores de MO
próximos nas médias observados e sem diferenças estatísticas significativas. As
concentrações dos atributos químicos colaboraram para a avaliação da qualidade do solo,
subsidiando a comprovação de manejos de solo mais sustentáveis.

77
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BARRETO et al. Características químicas e físicas de um solo sob floresta, sistema
agroflorestal e pastagem no sul da Bahia. Revista Caatinga. v.19, n.4, p.415-425.
Mossoró: outubro/dezembro, 2006.
FERREIRA, D. F. Sisvar: a Guide for its Bootstrap procedures in multiple comparisons.
Ciências e agrotecnologia. [online], vol.38, n.2, p. 109-112, 2014. Disponível em:
<ISSN 1413-7054. http://dx.doi.org/10.1590/S141370542014000200001>.
JAKELAITIS et al. Qualidade da camada superficial do solo sob mata, pastagens e áreas
cultivadas. Revista Pesquisa Agropecuária Tropical. V.38, n.2, p.118-127. Goiânia:
junho, 2008.
LIMA, J.S.S., SOUZA, G.S., SILVA, S.A. Amostragem e variabilidade espacial de
atributos químicos do solo em área de vegetação natural em regeneração. Revista
Árvore. v.34, n.1, p.127-136. Viçosa: fevereiro/outubro, 2010.
MULLER et al. Degradação de pastagens na Região Amazônica: propriedades físicas do
solo e crescimento de raízes. Pesquisa Agropecuária Brasileira. v.36, n.11, p.1409-
1418. Brasília: 2001.
NEVES NETO et al. Análise espacial de atributos do solo e cobertura vegetal em
diferentes condições de pastagem. Revista Brasileira de Engenharia Agrícola e
Ambiental. v.17, n.9, p.995-1004, Campina Grande: 2013.
RIVERO et al. Pecuária e desmatamento: uma análise das principais causas diretas do
desmatamento na Amazônia. Revista Nova Economia. v.19, p. 41-66, Belo Horizonte:
janeiro/abril, 2009.
RODRIGUES et al. Concentrações e quantidades de macronutrientes na excreção de
animais em pastagem de capim-mombaça fertilizada com fontes de fósforo. Revista
Brasileira de Zootecnia. v.37, n.6, p.990-997. Viçosa: 2008.
ROCHA JUNIOR, P.R.; SILVA, V.M.; GUIMARÃES, G.P. Degradação de pastagens
brasileiras e práticas de recuperação. Enciclopédia Biosfera, Centro Científico
Conhecer. v.9, n.17, p.952. Goiânia: 2013.
SCHLICKMANN, H.; SCHAUMAN, S.A. Pecuária, desmatamento e desastres
ambientais na Amazônia. Revista Ciências do Ambiente OnLine. v.3, n.2, 2007.
Disponível em: <http://www-di.inf.puc-rio.br/~endler/links/opinioes/RCA-OL-2017-
113.pdf>. Acesso em: 14, abr., 2018.
SILVA, B.; Nelson R. da; SILVA, L. G. T.; SAMPAIO, S. M. N. Caracterização
Biofísica e Aptidão Agrícola das Terras em Áreas de Projeto de Assentamento
Oficial: PA São Francisco. Belém: Documentos (EMBRAPA Amazônia Oriental),
2002. 38 p.

78
Belém (PA), 11 a 13 de dezembro de 2019
SOUZA et al. Variabilidade espacial de atributos químicos em um argissolo sob
pastagem. Acta Scientiarum. Agronomy. v.30, n.4, p.589-596. Maringá: 2008.

79
Belém (PA), 11 a 13 de dezembro de 2019
O EFEITO DA FREQUÊNCIA REGA COM BIOFERTILIZANTES EM
PARÂMETROS BIOMÉTRICOS, TROCAS GASOSAS E FLUORESCÊNCIA
DA CLOROFILA A EM MUDAS DE AÇAIZEIRO
Gleiciane Rodrigues dos Santos1; Rafael Rodrigo2; Gledson Luiz Salgado de Castro 3;
Alice Cavalcante 4; Gisele Barata da Silva5
1 Doutoranda em Agronomia. Universidade Federal Rural da Amazônia.
[email protected] 2Mestre em Agronomia. Universidade Federal Rural da Amazônia.
[email protected] 3Doutor em Agronomia. Universidade Federal Rural da Amazônia.
[email protected] 4Graduanda em Agronomia. Universidade Federal Rural da Amazônia.
[email protected] 5 Drª em Fitopatologia e professora. Universidade Federal Rural da Amazônia.
RESUMO
O crescimento lento e o menor vigor das mudas de açaizeiro limitam a expansão dos
plantios comerciais. O objetivo do estudo foi avaliar o efeito da frequência rega com
biofertilizantes na biometria, trocas gasosas e fluorescência da clorofila a em mudas de
açaizeiro. Os tratamentos consistiram das inoculações individuais de cinco isolados de
rizobactérias, Mix de Trichoderma e um controle (inoculados com água) nas raízes das
mudas de açaizeiro. Os dados de biometria foram submetidos a análise de agrupamento, e as médias dos grupos foram comparadas pelo teste SNK (P<0,05). Três grupos foram
formados, sendo o grupo 1 composto pelo controle, o grupo 2 composto pelas
Rizobactérias (R-38, R-46, R-55, R-61 e R-92) e o grupo 3 composto pelo Mix de
Trichoderma (T06, T09, T12 e T52). Os isolados dos grupos 2 e 3 promoveram aumento
no crescimento e melhoria nos parâmetros de trocas gasosas e fluorescência da clorofila
a em comparação ao grupo 1. Os resultados evidenciam que a segunda rega é a mais
eficiente para acelerar o crescimento e aumentar a eficiência fotossintética em mudas de
açaizeiro.
Palavras-chave: Euterpe oleracea. Microrganismos. Bioestimulantes
Área de Interesse do Simpósio: Agronomia

80
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O açaizeiro é uma palmeira nativa da Amazônia e de grande interesse econômico
no Brasil e no mundo. O Brasil é o maior produtor mundial do fruto de açaí, sendo o
estado do Pará o maior produtor nacional (IBGE, 2013). A maior demanda nacional e
internacional pela fruta na última década estimulou a mudança do sistema de produção
extrativista para plantios comerciais em grandes áreas (OLIVEIRA; NETO, 2004).
Entretanto, o crescimento inicial e o menor vigor limitam a produção de mudas nos
viveiros (COSTA; OLIVEIRA; OHAZE, 2004).
Na literatura atual, existem diversos estudos que reportam as possíveis influências
dos microrganismos promovendo o crescimento de mudas de palmeiras, como palma de
óleo (OM et al. 2009) e coco (GEORGE et al. 2013). Os biofertilizantes estimulam o
crescimento das plantas por diversos mecanismos, que incluem a melhoria da
transformação e aquisição de nitrogênio, auxilia na mineralização de fósforo, estimula a
produção de fitohormônios (KUREPIN et al., 2015; RASHID et al., 2016; VIMAL et al.,
2016; BELL et al., 2015; CHAUDHARY; SINDHU 2016).
O uso da tecnologia microbiana em mudas de açaizeiro pode acelerar o
crescimento e diminuir o tempo para obtenção das mudas em viveiros, contribuindo para
aumentar o vigor e diminuir a taxa de mortalidade das plantas no campo. O objetivo do
estudo foi avaliar à influência da frequência rega com os biofertilizantes na biometria,
trocas gasosas e fluorescência da clorofila a em mudas de açaizeiro.
2. MATERIAL E MÉTODOS
2.1 PREPARO DE MUDAS
Sementes de açaizeiro (cultivar BRS-Pará) foram semeadas em bandejas de
plástico contendo 2,5 L de substrato composto de fibra de coco triturada (Golden mix).
Aos 32 dias após a germinação, as plântulas que apresentaram duas folhas expandidas e
altura próxima de 13 cm foram transplantadas para sacos de plástico (15 x 25 cm,
comprimento x altura) contendo substrato composto de 60% de Latossolo e 40% de cama
de aviário curtida.

81
Belém (PA), 11 a 13 de dezembro de 2019
2.2 Isolados de Rizobactérias e Trichoderma asperrellum
Os isolados de Rizobactérias (R 38, R 46, R 55, R61 e R 92) e T. Mix (T06, T09,
T12 e T52) utilizados no ensaio são provenientes da coleção de microrganismos do
Laboratório de Proteção de Plantas (LPP) da Universidade Federal Rural da Amazônia-
UFRA, campus Belém.
2.3 Inoculação das rizobactérias e Trichoderma asperrellum
As rizobactérias (R-38, R-46, R-55, R-61 e R-92) foram cultivadas em meio sólido
523 (Kado; Heskett 1970) durante 48 h, a 28 ºC. As suspensões bacterianas foram
preparadas com água destilada e esterilizada, e a concentração foi ajustada em
espectrofotômetro para A540 = 0,5 (108 UFC). Os isolados de Trichoderma asperrellum
(T06, T09, T12 e T52) foram primeiramente cultivados em meio sólido BDA (batata,
dextrose e ágar) durante 96 h, a 28ºC. A partir dessas placas, foram preparadas suspensões
de conídios, ajustadas em câmara de Neubauer para 1 x 108 conídios mL-1. As plântulas
foram imersas em 500 mL da suspensão da PGPR e Mix de Trichoderma durante 30 min.
As plântulas do controle foram imersas em água destilada e esterilizada. Em seguida, foi
realizada uma segunda rega com 15 dias e uma terceira com 30 dias com 50 mL/plântula
da suspenção da PGPR e Mix de Trichoderma.
2.4 Avaliação do crescimento e biomassa
A promoção do crescimento foi avaliada aos cinco meses após a germinação das
sementes. A altura das plantas e o diâmetro do coleto foi mensurado com paquímetro
digital (precisão de 0,02 mm). O índice de robustez foi calculado pela razão entre a altura
da planta e diâmetro do coleto. O número de folhas foi avaliado através da contagem
direta das folhas.
2.5 Trocas gasosas e fluorescência da clorofila a
As trocas gasosas foram medidas aos três meses após a inoculação da rizobactérias
e do fungo Trichoderma. A assimilação líquida de CO2 (A), condutância estomática ao
vapor de água (gs), concentração intercelular de CO2 (Ci) e taxa de transpiração (E) foram
medidos com o LI-6400XT, LI-COR, Lincoln, NE. A fluorescência da clorofila a foi
determinada simultaneamente com as trocas gasosas utilizando-se uma câmara de

82
Belém (PA), 11 a 13 de dezembro de 2019
fluorescência IG 6400-40; LI-COR Inc. Foram estimados a Fluorescência inicial (Fo),
Fluorescência máxima (Fm), Eficiência fotoquímica efetiva do PSII (Fv’/Fm’), Taxa de
transferência de elétrons (ETR), Coeficientes de dissipação fotoquímica (qP) e
Coeficientes de extinção não fotoquímica(NPQ) de acordo com MAXWELL;
JOHNSON, 2000.
2.6 Análise estatística
O experimento foi realizado em delineamento inteiramente casualizado com sete
tratamentos cinco isolados de rizobactérias, Mix de Trichoderma e o controle (sem
rizobactéria ou Trichoderma) e quatro repetições. Os dados de crescimento foram
submetidos à análise de agrupamento usando os valores médios de cada indivíduo e o
grau de similaridade foi obtido pela distância Euclidiana padrão. Os grupos foram
submetidas à análise de variância (ANOVA) e as médias foram comparadas pelo teste
Student-Newman-Keuls (SNK), P < 0,05, usando o software R versão 3.1.
3. RESULTADOS E DISCUSSÃO
Todos os isolados testados na frequência de rega promoveram o crescimento em
mudas de açaizeiro (Figura 1). A análise de agrupamento revelou a formação de três
grupos na primeira rega, onde o tratamento controle composto pelo grupo 1 foi inferior
para todas as variáveis de promoção de crescimento em relação aos grupos 2 e 3,
composto pelos isolados R-38; R-46; R-55; R-61; R-92 e o T MIX, respectivamente. A
segunda e terceira rega também revelaram a formação de três grupos, onde o tratamento
controle foi inferior para todas as variáveis de promoção de crescimento (Figura 1).
Dentre as regas utilizadas com os biopromotores de crescimento, houve destaque para
segunda rega para todas as variáveis biométrica em mudas de açaizeiro.

83
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1: Avaliação biométrica em mudas de açaizeiro (5 meses), inoculadas com
biopromotores em relação ao controle.
dAvaliações biométricas aRegas bGrupos cIsolados ALT
(cm)
NF DC
(mm)
IR
1X
1 Controle 40,12 c 3,25 b 8,07 b 4,97 b
2 R 46; R 55; R 61; R 92;T. Mix 49,55 b 4,20 a 9,15 ab 5,45 ab
3 R 38 57,25 a 3,75 ab 9,78 a 5,89 a
CV % 6,39 10, 5 8,96 7,46
2X
1 Controle 44,00 b 4,25 a 8,97 b 5,00 b
2 R 38; R 46; R 55; R 61; R 92. 54,94 b 4,31 a 10,65 a 5,19 ab
3 T. Mix 65,25 a 4,5 a 11,19 a 5,83 a
CV % 9,76 8,96 10,18 9,54
3X
1 Controle 44,00 b 4,25 a 9,39 b 4,68 ab
2 R 38; R 46; R 55; R 61; T.MIX. 53,35 a 4,40 a 10,80 ab 5,16 a
3 R 92 42,15 b 4,25 a 11,39 a 4,35 b
CV % 13,70 12,60 12,77 11,24 aFrequência de regas com suspensão dos biopromotores; bgrupos de similaridade de acordo com
a matriz Euclidiana; cIsolados de rizobactérias (R38, R46, R 55, R 61 e R 92) e T. Mix (4 isolados
de Trichoderma asperrellum) e dAvaliações biométricas: Altura em cm (ALT), Número de folhas
(NF), Diâmetro do coleto em mm, (DC) e Índice de robustez (IR).*Médias com letras iguais não
diferem significativamente de acordo com o teste Student-Newman-Keuls (P<0,05).
Na segunda rega todas as variáveis de crescimento apresentaram aumento
significativo, o aumento variou entre 23 e 47% para a altura de planta, em 19 e 32% para
o diâmetro do coleto, o número de folha em 38% e, em 3 e 17% para o índice de robustez
para os isolados dos grupos 2 e 3, respectivamente em comparação com o grupo 1.
O crescimento da parte aérea, assim como do número de folhas observados no
grupo 2 e 3 (Figura 1) pode ser atribuído a capacidade das rizobactérias e do fungo
Trichoderma de induzir a síntese de giberelinas e citocininas que regulam a expansão
foliar e síntese de clorofilas (DODD et al. 2010; KANG et al. 2014). ). Nascente et al,
(2016) reportaram que a aplicação de microrganismos como Serratia sp. (BRM32114),
Bacillus sp. (BRM32110 and BRM32109) e o Mix de Trichoderma asperellum
promoveram um aumento na taxa fotossintética e na biomasssa seca da folha em planta
de arroz.

84
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 2. Avaliação de trocas gasosas em mudas de açaizeiro tratadas com duas regas como
os biofertilizantes.
aGrupos bIsolados
cAvalições de Trocas gasosas
A Ci gs E
1 Controle 11,67 b 357,14 a 0,0563 b 1,3512 b
2 R-38; R-46; R-55;
R-61; R-92 14,17 a 353,62 a 0.0756 ab 1,8483 ab
3 T. MIX 15,61 a 360,05 a 0,0849 a 2,1107 a
CV (%)
12,37 1,41 22,63 20,98
Grupos de similaridade de acordo com a matriz Euclidiana; bIsolados de rizobactérias (R 38,
R 46, R 55, R 61 e R 92) e o Mix de Trichoderma asperrellum (T06, T09, T12 e T52) e cAvaliações de Trocas gasosa: (A) Taxa de assimilação líquida de CO2, (Ci) Concentração
intercelular de CO2, (gs) Condutância estomática ao vapor de água e (E) Transpiração.*Médias
com letras iguais não diferem significativamente de acordo com o teste de Student-Newman-
Keuls (P<0,05).
Os resultados da segunda rega com os biofertilizantes aumentou o desempenho
fotossintético através dos parâmetros de trocas gasosas e fluorescência da clorofila a. Os
aumentos variaram entre 21 e 34% para A, em 34 e 52% para gs, em 19% e 32% para Ci
e, em 36 e 56% para E nos grupos 2 e 3, respectivamente (Tabela 2). Dentre os
biofertilizantes utilizados na avaliação de trocas gasosas o Mix de Trichoderma,
destacou-se com aumento na taxa de assimilação líquida de carbono (A), condutância
estomática ao vapor de água (gs), carbono intercelular (Ci) e taxa de transpiração (E).
A variável de concentração intercelular de CO2 (Ci), não apresentou diferença
estatística em comparação ao grupo 1, corroborando, assim, com os resultados relatados
por Nascente et al (2016). Entretanto, os parâmetros de A e E nos grupos 2 e 3 foram
maiores em relação ao grupo 1, provavelmente essas variáveis foram influenciadas pelo
maior nível de abertura estomática (gs) em comparação ao grupo 1. Zhang et al (2008),
relataram que algumas rizobactérias têm a capacidade de modular as trocas gasosas e o
metabolismo ABA, provavelmente o aumento da densidade de estômatos nessas plantas
inoculadas com grupos 2 e 3 induziu o aumento na taxa de assimilação líquida de carbono
(A) em relação a plantas inoculadas do grupo 1 (Figura 2).
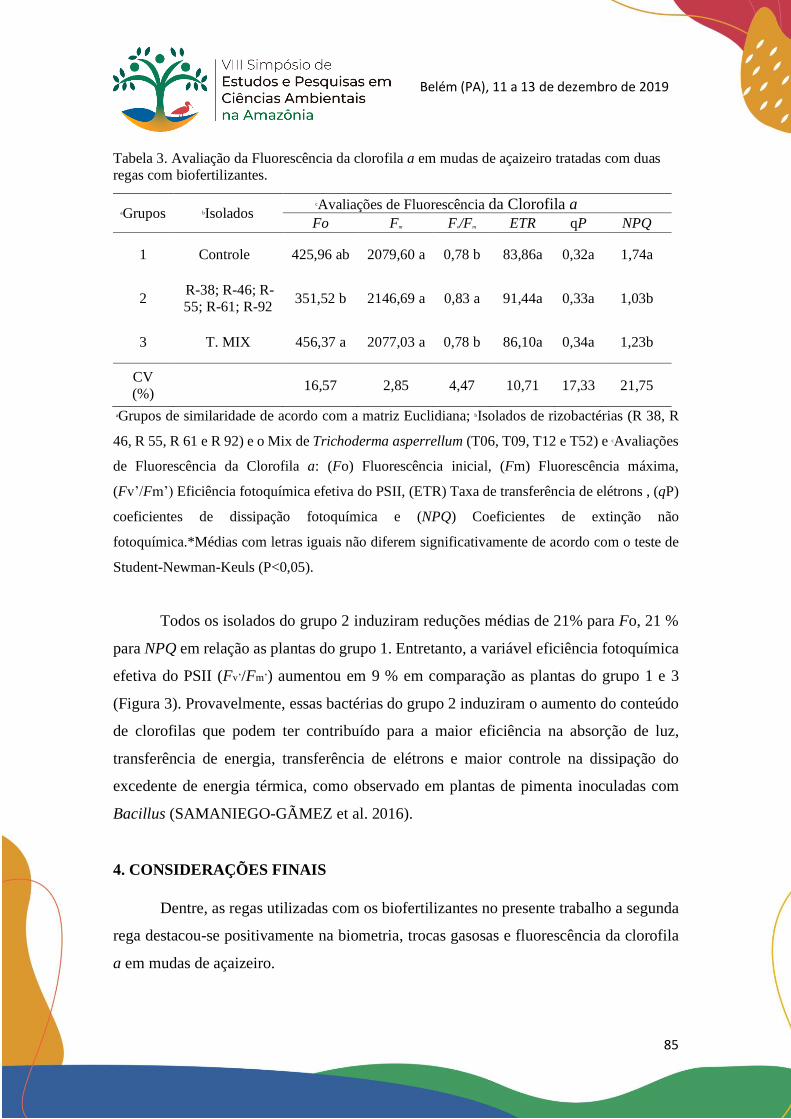
85
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 3. Avaliação da Fluorescência da clorofila a em mudas de açaizeiro tratadas com duas
regas com biofertilizantes.
aGrupos bIsolados cAvaliações de Fluorescência da Clorofila a
Fo Fm Fv/Fm ETR qP NPQ
1 Controle 425,96 ab 2079,60 a 0,78 b 83,86a 0,32a 1,74a
2 R-38; R-46; R-
55; R-61; R-92 351,52 b 2146,69 a 0,83 a 91,44a 0,33a 1,03b
3 T. MIX 456,37 a 2077,03 a 0,78 b 86,10a 0,34a 1,23b
CV
(%) 16,57 2,85 4,47 10,71 17,33 21,75
aGrupos de similaridade de acordo com a matriz Euclidiana; bIsolados de rizobactérias (R 38, R
46, R 55, R 61 e R 92) e o Mix de Trichoderma asperrellum (T06, T09, T12 e T52) e cAvaliações
de Fluorescência da Clorofila a: (Fo) Fluorescência inicial, (Fm) Fluorescência máxima,
(Fv’/Fm’) Eficiência fotoquímica efetiva do PSII, (ETR) Taxa de transferência de elétrons , (qP)
coeficientes de dissipação fotoquímica e (NPQ) Coeficientes de extinção não
fotoquímica.*Médias com letras iguais não diferem significativamente de acordo com o teste de
Student-Newman-Keuls (P<0,05).
Todos os isolados do grupo 2 induziram reduções médias de 21% para Fo, 21 %
para NPQ em relação as plantas do grupo 1. Entretanto, a variável eficiência fotoquímica
efetiva do PSII (Fv’/Fm’) aumentou em 9 % em comparação as plantas do grupo 1 e 3
(Figura 3). Provavelmente, essas bactérias do grupo 2 induziram o aumento do conteúdo
de clorofilas que podem ter contribuído para a maior eficiência na absorção de luz,
transferência de energia, transferência de elétrons e maior controle na dissipação do
excedente de energia térmica, como observado em plantas de pimenta inoculadas com
Bacillus (SAMANIEGO-GÃMEZ et al. 2016).
4. CONSIDERAÇÕES FINAIS
Dentre, as regas utilizadas com os biofertilizantes no presente trabalho a segunda
rega destacou-se positivamente na biometria, trocas gasosas e fluorescência da clorofila
a em mudas de açaizeiro.

86
Belém (PA), 11 a 13 de dezembro de 2019
O biofertilizante a base de Trichoderma asperrellum acelerou o crescimento de
em mudas de açaizeiro e trocas gasosas.
A inoculação do Mix de Trichoderma asperrellum pode contribuir para o manejo
sustentável da produção de mudas de açaizeiro em viveiros.

87
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BELL C W, ASAO S, CALDERON F, WOLK B, WALLENSTEIN M D. Plant nitrogen
uptake drives rhizosphere bacterial community assembly during plant growth. Soil Biol
Biochem. 85: 170–182, 2015.
CHAUDHARY, S. R.; SINDHU, S. S. Growth stimulation of clusterbean (Cyamopsis
tetragonoloba) by coinoculation with rhizosphere bacteria and Rhizobium. Legume
Research - An International Journal, v. 39, n. OF, p. 1003–1012, 2016.
COSTA, M. R.; OLIVEIRA, M. D. S. P. DE; OHAZE, M. M. M. Divergência Genética
No Açaizeiro Com Base Em Marcadores Rapd. REv. Ciênc. Agrár., v. 41, p. 89–95,
2004.
DODD IC, ZINOVKINA NY, SAFRONOVA VI, BELIMOV AA. Rhizobacterial
mediation of plant hormone status. Ann Appl Biol 157:361–379.
doi:10.1111/j.17447348.2010.00439, 2010.
GEORGE P, GUPTA A, GOPAL M, et al. Multifarious beneficial traits and plant growth
promoting potential of Serratia marcescens KiSII and Enterobacter sp. RNF 267 isolated
from the rhizosphere of coconut palms (Cocos nucifera L.). World J Microbiol
Biotechnol 29:109–117. doi: 10.1007/s11274-012-1163-6, 2013.
IBGE. 2013. Produção da Extração Vegetal e da Silvicultura. Disponível em:
<http://www.ibge.gov.br/> Acesso em: 10 de outubro de 2019.
KADO CI, HESKETT MG. Selective Media for Isolation of Agrobacterium,
Corynebacterium, Erwinia, Pseudomonas, and Xanthomonas. Phytopathology 60:969.
doi: 10.1094/Phyto-60-969, 1970.
KANG SM, KHAN AL, YOU YH, et al. Gibberellin production by newly isolated strain
Leifsonia soli SE134 and Its potential to promote plant growth. J Microbiol Biotechnol
24:106–112. doi: 10.4014/jmb.1304.04015, 2014.
KUREPIN L V, PARK M J, LAZAROVITS G, BERNARDS M A. Burkholderia
phytofirmans-induced shoot and root growth promotion is associated with endogenous
changes in plant growth hormone levels. Plant Growth Regul. v 75: 199–207, 2015.
MAXWELL K, JOHNSON GN. Chlorophyll fluorescence--a practical guide. J Exp Bot.
51:659–668. doi: 10.1093/jexbot/51.345.659, 2000.
NASCENTE AS, DE FILIPPI MCC, LANNA AC, et al. Biomass, gas exchange, and
nutrient contents in upland rice plants affected by application forms of microorganism
growth promoters. Environ Sci Pollut Res 24:2956–2965. doi: 10.1007/s11356-016-
8013-2, 2016.
OLIVEIRA, M. D. S. P.; NETO, J. T. D. F. Cultivar BRS-Pará: Açaizeiro para Produção
de Frutos em Terra Firme. Embrapa, Comunicado Técnico, v. 114, n. 1, p. 1–3, 2004.

88
Belém (PA), 11 a 13 de dezembro de 2019
OM AC, GHAZALI AHA, KENG CL, ISHAK Z. Microbial inoculation improves growth
of oil palm plants (Elaeis guineensis Jacq.). Trop Life Sci Res 20:71–77, 2009.
RASHID M A, MUJAWAR L H, SHAHZAD T, ALMEELBI T, ISMAIL I M I, OVES
M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation
in degraded soils. Microbiol Res. 183: 26–41, 2016.
SAMANIEGO-GÁMEZ BY, GARRUÑA R, TUN-SUÁREZ JM, et al. Bacillus spp.
inoculation improves photosystem II efficiency and enhances photosynthesis in pepper
plants. Chil J Agric Res 76:409–416. doi: 10.4067/S0718-58392016000400003, 2016.
VIMAL S R, SINGH J S, ARORA N K, SINGH D P. PGPR—an effective bioagent in
stress agriculture management. In Sarma B K, Jain A (eds.) Microbial Empowerment in
Agriculture: A Key to Sustainability and Crop Productivity. Biotech Books, New Delhi.
pp. 81–108, 2016.
ZHANG K, LIU Z, SHAN X, et al. Physiological properties and chlorophyll biosynthesis
in a Pak-choi (Brassica rapa L. ssp. chinensis) yellow leaf mutant, pylm. Acta Physiol
Plant 39:22. doi: 10.1007/s11738-016-2321-5, 2017.

89
Belém (PA), 11 a 13 de dezembro de 2019
PLANEJAMENTO SUSTENTÁVEL DO USO GLIFOSATO NA
AGRICULTURA
Ruricksson Progênio da Conceição1; Mayke Feitosa Progênio2; Ynglety Cascaes
Pereira3; Yash Brendo Pereira Coelho Guimarães4; Felipe Angolini Karpinski5
1 Engenheiro Agrônomo graduado pela Universidade Federal Rural da Amazônia
(UFRA).
[email protected] 2 Doutorando em Engenharia civil na Universidade Federal do Pará (GAES/GEEPAM).
[email protected] 3 Mestranda na Universidade Federal Rural da Amazônia (PPGEA).
[email protected] 4Mestrando na Universidade Federal Rural da Amazônia (PPGEA).
[email protected] 5 Engenheiro Agrônomo graduado no Centro Universitário Luterano de Santarém.
RESUMO
O herbicida glifosato é amplamente utilizado na agricultura, e tem se tornado uma
preocupação ambiental devida seus efeitos deletérios aos microorganismo do solo e da
água, sendo um potencial causador de desequilíbrio ambiental. Desta forma é importante
a discussão sobre o tema, novas pesquisas e tecnologias a respeito do mesmo, assim como
propor métodos de manejo a fim de reduzir o uso exagerado deste químico. O objetivo
deste estudo foi discorrer, sobre a utilização do glifosato na agricultura, seu
comportamento no solo e possíveis alternativas de manejo para redução de seu uso na
produção agrícola. O presente trabalho utiliza-se de levantamento bibliográfico como
ferramenta metodológica, de produções cientificas relacionadas ao uso do glifosato, seus
efeitos no ambiente, formas de degradação, e técnicas de manejo estudados no intuito de
reduzir sua utilização. As conclusões obtidas foram que apesar de algumas técnicas de
remediação do glifosato no ambiente terem se mostrado promissoras, como a utilização
de biorreatores com bactérias de biofilme e adsorção em carvão ativado, as mesmas são
métodos dispendiosos, e que ainda necessitam de mais pesquisas. Assim reduzir a
utilização do glifosato na agricultura se mostra mais eficiente e possível de ser feito
através de manejo adequado da produção, utilizando da técnica do plantio direto, assim
como o planejamento adequado da safra.
Palavras-chave: Ambiente. Herbicidas. Manejo.
Área de Interesse do Simpósio: Agronomia

90
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O uso de herbicida na agricultura seja para controle de plantas daninhas, ou como
dissecante, é uma técnica de uso generalizado e frequente. Verificando-se, que em
determinadas situações de uso contínuo, há existência de gramíneas resistentes aos
herbicidas, como a Digitaria Insularis também conhecida como capim-amargoso, que
possui resistência ao herbicida glifosato (Ferreira, 2019).
O princípio ativo do herbicida não atinge diretamente o ser humano, mas seu efeito
de contaminação do solo é preocupante, uma vez que, o solo é um sistema vivo e
heterogêneo, muito sensível a mudanças químicas e físicas que podem interferir na sua
microbiota (MADALÃO et al, 2016). Sendo a degradação microbiana o principal
processo que influência na dinâmica do herbicida no solo, o que inclui sua persistência
no mesmo e susceptibilidade de lixiviação, é imperativo a manutenção desta biota
(TEJADA, 2009).
A classificação para os agroquímicos (herbicidas) varia de acordo com seu grupo
químico, formulação, seletividade, índices toxicológicos e mecanismo de ação. Esses
mecanismos podem ser sistêmicos ou por contato, no primeiro a ação ocorre através do
sistema xilema e floema da planta, sendo transportado para os pontos de crescimento,
inibindo a síntese de aminoácidos (PRUDENCI; JÚNIOR, 2019), já no segundo, o
químico causa a destruição dos tecidos atingidos, sendo o principal foco o tecido foliar,
responsável pelas trocas gasosas e pela fotossíntese (ZUCONELLI, 2018).
Neste estado da arte, estão os herbicidas organofosforados que vem sendo
amplamente utilizados nas lavouras. Estes por sua vez tornaram-se uma questão de
preocupação ambiental, devido os efeitos nocivos desses produtos químicos aos
processos biológicos do solo e da água (SVIRIDOV et al, 2015; FRANCESCHI et al,
2017; FARIA, 2018;). Os organofosforados são compostos orgânicos formados de
ligações fosforo-carbono, sendo base da maioria dos herbicidas utilizados na agricultura,
sendo os mais conhecidos o 2,4D e o glifosato (BARBOSA et al,2018). Desta forma, é
muito importante que pesquisas sejam feitas dentro deste tema, buscando novas soluções
e tecnologias a respeito da degradação do glifosato. O objetivo deste trabalho foi
discorrer, sobre a utilização do glifosato na agricultura, sua dinâmica no solo, e possíveis
alternativas de manejo para a redução do uso do mesmo dentro do ambiente de produção.

91
Belém (PA), 11 a 13 de dezembro de 2019
2. METODOLOGIA
O presente artigo trata-se de uma revisão bibliográfica, e para a seleção foram
utilizadas as bases de dados Scielo, Elsevier e Google acadêmico, com publicações do
período de 2015-2019, sendo o tema central: o uso intensivo do herbicida glifosato na
agricultura. Foram utilizados os descritores controlados: dinâmica do glifosato no solo,
biodegradação de herbicidas, processos de adsorção de glifosato, impactos ambientais por
herbicidas, plantio direto, modelagem estocástica de previsão. Sendo realizado o
diagnóstico de estudos já realizados, que descrevem aplicações e resultados obtidos a
respeito do tema.
3. RESULTADOS E DISCUSSÃO
3.1. UTILIZAÇÃO DO GLIFOSATO NA AGRICULTURA BRASILEIRA
O Brasil nos últimos anos tem se tornado um dos maiores produtores e
exportadores de soja, com produção de 123,664 milhões de toneladas, segundo maior
produtor do mundo atrás dos norte-americanos (CONAB, 2018). Toda essa escala de
produção é consequência de maiores investimentos em tecnologia, melhor uso da terra e
também da expansão das fronteiras agrícolas, principalmente para a região norte
(EMBRAPA, 2013).
O sucesso de uma boa produção está intimamente ligado aos tratos culturais
realizados, seja contra pragas ou doenças, assim como plantas oportunistas no caso das
plantas daninhas (MAGALHÃES et al, 2001). As plantas daninhas ou espontâneas, são
espécies vegetais que afloram dentro de uma produção causando prejuízo econômico ou
material dentro da mesma (LORENZI, 1984; MELLONI et al, 2016). Assim, a
metodologia aplicada no manejo dessas invasoras, vai depender do tipo e escala de
produção, assim como o seu tipo. Dessa forma, é comum em áreas muito extensas o uso
mais de métodos químicos do que físicos, devido a facilidade de aplicação por
pulverização, e também a rapidez de ação do químico, os herbicidas organofosforados
podem ser de contato ou sistêmicos, sendo os mais populares o glifosato (N-
fosfonometilglicina) e o 2,4-D (Ácido 2,4-diclorofenoxiacético) (LACERDA et al, 2001;
MACIEL et al, 2005; TOLEDO et al, 2012).

92
Belém (PA), 11 a 13 de dezembro de 2019
Nas principais culturas econômicas, principalmente a soja, o glifosato além de ser
usado como químico de controle de daninhas, também é utilizado como dissecantes pós
emergentes, para evitar atraso de colheita (MARCANDALI et al, 2011). De acordo com
a Associação Brasileira de Produtores de Soja (APROSOJA) a média de aplicação por
hectare de glifosato varia de 2 a 4 litros, considerando a área plantada de soja no Brasil
de 5,89 milhões de hectares prevista para safra de 2018/2019, serão 29,45 milhões de
litros destinados a produção de soja neste ciclo (APROSOJA, 2018). A utilização
exagerada desses químicos além de acarretar em danos ambientais tanto ao solo como a
corpos hídricos, os mesmos podem deixar resíduos na semente prejudicando na sua
qualidade, e deterioramento de seu tegumento, além de ser uma possível fonte de
contaminação (GOUVÊA et al, 2015).
3.2. ADSORÇÃO E BIODEGRADAÇÃO DA MOLECULA DE GLIFOSATO
A grande expansão da agricultura, trouxe consigo maior demanda na utilização de
agentes de controle químico, apenas neste ano, o ministério da agricultura, liberou 262
novos registros de agrotóxicos (MAPA, 2019). Além do fato de algumas moléculas já
apresentarem estudos no exterior sobres seus efeitos maléficos ao meio ambiente, a
flexibilização na utilização desses químicos pode acarretar na perca de eficácia do mesmo
sobre o agente causador (PAPA et al, 2014).
Devido aos efeitos nocivos dos agrotóxicos ao meio ambiente, estudos são
realizados sobre a degradação do mesmo, sua dinâmica no solo, na água e também na
atmosfera (RIBEIRO, 2010). O estudo realizado por Gorza (2012), avaliou a remoção de
glifosato, e metabólitos, em uma planta piloto, associada a pré-oxidação com dióxido de
cloro e adsorção com carvão ativado granular (CAG), onde na concentração de 0,620
mg.L-1 de glifosato ouve a eficiência de remoção de 85% do residual do herbicida
mencionado.
A eficiência de alguns métodos podem ser questionados quanto a remoção do
glifosato, como no trabalho de Lopes (2017), que utilizou de biorreatores de membrana
para remoção de glifosato e 2-4D, onde inicialmente ouve a retenção/adsorção dos
herbicidas no lodo ativado, porém, ao longo do tempo houve a eliminação gradativa do
químico para a solução, sendo assim pouco eficiente neste processo. Entretanto, o método
com biorreatores se mostrou eficaz quando utilizado em conjunto com carvão ativado

93
Belém (PA), 11 a 13 de dezembro de 2019
granular, onde na concentração de 10 mg.L-1 de glifosato ouve a taxa de remoção de 90%.
Vargas et.al (2019), em seu trabalho com a utilização de bactérias do gênero limnoperma
na degradação do glifosato em água, verificou a eficiência da degradação do herbicida
através da bactéria, onde na concentração de 6 mg.L-1 do herbicida ouve a remoção de
grande parte do mesmo assim como sua dissipação no meio aquoso, demonstrando a
eficácia dessas bactérias em degradar o glifosato, porém essa dissipação de nutrientes
decorrente da degradação, pode acelerar o processo de eutrofização dos recursos hídricos.
O glifosato no ambiente quando não absorvido por plantas, tende a forma
complexos com íons de Ca2+ e Mg2+, sofrendo sorção com sedimentos ou partículas
suspensas na água ou no solo, ou sendo biodegradado por microorganismos (SOUZA et
al, 2017). A biodegradação por microorganismos podem ocorrer de maneira direta ou
indireta, envolvendo a oxidação dos compostos intermediários, sendo o catabolismo e o
metabolismo microbiano os principais meios de mineralização (ZOBIOLE et al, 2010).
A rota de degradação do glifosato mais comum envolve a conversão do mesmo em ácido
aminometilfosfônico (AMPA), como metabólito principal através da ação de enzimas
oxirredutases e transaminases, que modificam as ligações entre carbono e fósforo
(TEJADA, 2009).
De acordo com Andrighetti et.al (2014), em seus estudos sobre biodegradação de
glifosato pela respiração da microbiota do solo, mostraram que a capacidade dos
microorganismos edáficos em degradar o mesmo, estavam diretamente relacionados com
a não presença deste a priori no solo e com o intervalo de tempo entre as aplicações. De
acordo com Tang et al. (2019), o glifosato quando utilizado por longos períodos sobre o
mesmo solo, acarreta efeitos adversos a multiplicação dos microorganismos do solo,
diminuindo sua eficácia na degradação da molécula. Desta forma um bom manejo de
aplicação, junto a trocas consecutivas de moléculas químicas, auxilia na redução destes
efeitos e possíveis danos ambientais (SÁ; ROCHA, 2018).
3.3. MÉTODOS E AÇÕES POSSIVEIS PARA REDUÇÃO DO USO DO GLIFOSATO
NA AGRICULTURA
Atualmente muito se é discutido na literatura sobre o uso exacerbado dos
agrotóxicos, seja por seus impactos ao meio ambiente, como também a saúde humana
(ALMEIDA et al, 2017; OLIVEIRA et al, 2018). Entretanto a sua utilização não deveria

94
Belém (PA), 11 a 13 de dezembro de 2019
ser discriminada, esses químicos exercem um papel fundamental dentro do processo de
produção, ao contrário de ser visto como o principal ferramenta de controle, o mesmo
deveria ser usado como mais um, de um conjunto de métodos profiláticos de controle
(SOUZA et al, 2016).
Em pequenas propriedades o método físico pode ser mais aplicado como capinas,
coroamentos, uso de cobertura morta, assim como a queima controlada, são alternativas
eficazes no controle de plantas daninhas (ANJOS et al, 2016). A utilização de cobertura
morta, tem grande eficiência no controle de daninhas, como realizado no trabalho de
Tonin et.al (2017), onde o mesmo utilizou de maravalha e acícula de pinus como
cobertura, obtendo grande eficiência na diminuição da taxa de proliferação dessas
espontâneas, equiparado ao mulching de polietileno.
Se tratando de áreas mais extensas, como na cultura de soja e milho, também pode
ser utilizado o uso de cobertura morta, como no caso do sistema de plantio direto, onde a
cobertura serve de barreira para a passagem da luz, impedindo a germinação do banco de
sementes do solo (MARCHESAN et al, 2016). Este método também pode ser aplicado
em diversas outras culturas, como o feijão e a cana-de-açúcar, os benefícios vão além,
como proteção do solo a erosão e manutenção da umidade (ORDOÑEZ-GÓMEZ;
REÁTEGUI-DÍAZ; VILLANUEVA-TIBURCIO, 2018).
O glifosato também é utilizado como dissecante na cultura da soja, o que está
relacionado diretamente ao tempo que esta cultura passa no campo (SILVA; FENILLI
JUNIOR; LORENZETTI, 2016). Para que a soja seja colhida no tempo certo, são
necessários períodos consecutivos de pouca umidade, ou seja, poucas lâminas de
precipitação pluvial (PINTO et al, 2017). Uma forma de contornar esta situação, se
encontra dentro do planejamento agrícola, onde o produtor, com dados climáticos em
mãos, poderia fazer seu plano de safra diante as adversidades do clima. O método de
estimativa de precipitação diária, vem sendo estudado como uma ferramenta a auxiliar o
desenvolvimento agrícola de uma região, onde através de uma séries de modelos de
probabilidades é possível estimar a precipitação tanto no espaço como no tempo, desta
forma sendo possível plantar e colher no período certo ( CALGARO et al, 2009; BAÚ;
AZEVEDO; BRESOLIN, 2013; TEIXEIRA-GRANDA et al, 2017).

95
Belém (PA), 11 a 13 de dezembro de 2019
De acordo com Progênio (2019), a modelagem estocástica de precipitação
pluviométrica, é uma ferramenta eficaz na definição da probabilidade de ocorrência de
chuvas diárias, em seu trabalho na Região Hidrográfica Tocantins-Araguaia (RHTA),
conseguiu reproduzir as quantidades de dias secos e chuvosos de forma satisfatória
através da cadeia de markov (CM) de 1ª ordem. Assim, em posse das informações obtidas
por estes modelos hidrológicos, o calendário agrícola pode ser otimizado, ou seja, o
processo de colheita será realizado no período de maior probabilidade de ocorrência dos
dias secos. Portanto, a utilização de dissecantes químicos neste caso não serão mais
necessários.
4. CONSIDERAÇÕES FINAIS
A utilização do glifosato na agricultura é uma prática rotineira para a maioria dos
produtores, sejam estes grandes ou pequenos, mudar a percepção destes é importante
atualmente para o desenvolvimento sustentável. O estudo de novas técnicas para lidar
com a contaminação por herbicidas tanto de solos como de recursos hídricos se fazem
necessários, dentre estas a adsorção do glifosato em biorreatores de membrana com
bactérias de biofilme assim como em carvão ativado se mostrou promissores. Todavia
são métodos dispendiosos e que ainda carecem de pesquisa. A microbiota do solo se
mostrou importante no processo de biodegradação deste herbicida, revelando a
necessidade de rotatividade de produtos químicos assim como a sua alternância e
intervalos de uso prolongados para a manutenção da mesma. A necessidade de utilização
do glifosato pode ser reduzida através de métodos como o plantio direto, sendo o mesmo
eficaz no controle de plantas daninhas, além de outras melhorias que o mesmo agrega ao
solo. O plano de safra, é a melhor alternativa para redução do uso do glifosato como
dissecante, aliado a modelagem estocástica de precipitação, pode se torna a forma mais
lucrativa para o agricultor, reduzindo o uso de químicos na sua produção. Porém, são
necessários mais estudos voltados diretamente para as principais culturas comerciais,
principalmente na região Amazônica.

96
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALMEIDA, Mirella Dias et al. A flexibilização da legislação brasileira de agrotóxicos e
os riscos à saúde humana: análise do Projeto de Lei nº 3.200/2015. Cadernos de Saúde
Pública,[s.l.],v.33,n.7,p.16,2017.FapUNIFESP(SciELO)..Disponível..em:..<http://dx.do
i.org/10.1590/0102311x00181016>. Acesso em: 23 set. 2019.
Associação Brasileira de Produtores de Soja (APROSOJA). Glifosato entre vilão e
salvador da safra. Disponível em:
<https://gauchazh.clicrbs.com.br/economia/campoelavoura/noticia/2018/09/glifosatoent
revilaoesalvadordasafracjls4q3jz01ib01pxwcgjbuyj.html> Acesso em 12 de novembro de
2019.
BARBOZA, Henriqueta Talita G. et al. Organophosphorus Compounds and their Role in
Agriculture. Revista Virtual de Química, [s.l.], v. 10, n. 1, p.172-193, 2018. Sociedade
Brasileira.de.Quimica.(SBQ).Disponivél.em:.<http://dx.doi.org/10.21577/19846835.201
80015>. Acesso em: 26 de setembro de 2019
BAÚ, Antonio L.; AZEVEDO, Carlos A. V. de; BRESOLIN, Adriano de A..Modelagem
da precipitação pluvial diária intra-anual da Bacia Hidrográfica Paraná III associada aos
eventos ENOS. Revista Brasileira de Engenharia Agrícola e Ambiental, Campina
Grande, v. 17, n.8, p.883.891, 31 maio 2013 Mensal. Disponível em:
<https://www.researchgate.net/profile/Adriano_Bresolin/publication/260771848_Model
ing_of_daily_intraannual_precipitation_of_the_Parana_III_Basin_associated_with_eve
nts_ENSO/links/54088a3a0cf2718acd381149.pdf>. Acesso em: 23 set. 2019.
BRANDELERO, W; BARBACOVI, A; ROSBACH, M. G. VIEBRANTZ, C; GIRARDI,
L.B; MAYER, A.R; CASASSOLA, A. Vigor e viabilidade de sementes de soja em
resposta a umidade durante o processo de armazenagem. Brazilian Journal Of
Development. Curitiba, v. 5, n.1 p.342-350, 10 dez 2018. Mensal Disponível em:
<http://brjd.com.br/index.php/BRJD/article/view/924/801>. Acesso em: 21 set. 2019.
BÜLOW,,R.L.;CRUZ_SILVA,C.T.A...DESSECANTES.APLICADOS.NA.PRÉ.COLH
EITA.NA.QUALIDADE.FISIOLÓGICA.DE.SEMENTES.DE.SOJA.
.Journal.Of.Agronomic.Sciences, Umuarama, v. 1, n. 1, p.67-75, 05 jul. 2012. Mensal.
Disponível em: <http://www.dca.uem.br/V1N1/07-Bullow.pdf>. Acesso em: 21 set.
2019.
CALGARO, Marcelo et al. VARIAÇÃO ESPAÇO-TEMPORAL DOS PARÂMETROS
PARA A MODELAGEM ESTOCÁSTICA DA PRECIPITAÇÃO PLUVIAL DIÁRIA
NO RIO GRANDE DO SUL. Revista Engenharia Agrícola, Jaboticabal, v. 29, n. 2,
p.196-206, 14 maio 2009 Mensal. Disponível em:
<https://www.alice.cnptia.embrapa.br/bitstream/doc/660938/1/Calgaro.pdf>. Acesso
em: 23 set. 2019.
CONAB, COMPANHIA NACIONAL DE ABASTECIMENTO, safra brasileira de
grãos. Boletim de safra de grãos ano de 2018. Disponível em:
<https://www.conab.gov.br/info-agro/safras/graos> Acesso em 18 de setembro de 2019.
ANJOS, Jamille Santana et al. A experiência empírica de agricultores em áreas de várzea
e de terra firme no Município de São Domingos do Capim, Pará. Cadernos de

97
Belém (PA), 11 a 13 de dezembro de 2019
Agroecologia, [S.l.], v. 10, n. 3, may 2016. ISSN 2236-7934. Disponível em:
<http://revistas.aba-agroecologia.org.br/index.php/cad/article/view/17551>. Acesso em:
25 setembro. 2019.
EMBRAPA SOJA. Tecnologias de produção de soja - Região Central do Brasil, 2012 e
2013. Londrina: EMBRAPA Soja, 261p. 2011. (EMBRAPA Soja, Sistema de Produção,
n.15).
FARIA, M. A. EFEITO DA VINHAÇA E HERBICIDA CONTENDO
TEBUTIUROMSOBRE BACTÉRIAS EM SOLO TOXICIDADE EM ALFACE.
2018.70f.Dissertação(Mestrado)..Curso.de.Ciencias.Biologicas,.Departamento.de.Bioqu
ímica.e.Microbiologia,Universidade.Estadual.Paulista.“jÚlio.de.Mesquita.Filho”,Rio.Cl
aro,2018..Disponível.em:.<https://repositorio.unesp.br/bitstream/handle/11449/155928/f
aria_ma_me_rcla.pdf?sequence=5&isAllowed=y>. Acesso em: 25 set. 2019.
FERREIRA, Silvio Douglas. Resistencia ao glifosato em biótipos de Digitaria Insularis e
nível de dano econômico em soja e milho. 2019. F. Tese (Doutorado em Agronomia) –
Universidade Estadual do Oeste do Paraná, Marechal Cândido Rondon, 2019. Disponível
em: <http://tede.unioeste.br/handle/tede/4466> Acesso em 25 de setembro de 2019
FRANCESCHI, Mauricio et al. Comportamento do 2,4-D + picloram em Latossolo
Vermelho Amarelo. Revista Brasileira de Herbicidas, [s.l.], v. 16, n. 3, p.238-245, 10
set. 2017. Revista Brasileira de Herbicidas. Disponivel em:
http://dx.doi.org/10.7824/rbh.v16i3.524. Acesso dia 26 de setembro de 2019
GORZA, N.L. Remoção de agrotóxicos em uma instalação piloto de tratamento de
águas de abastecimento do tipo convencional, associado a pré-oxidação e adsorção
em carvão ativado granular. 2012. 134f. Dissertação (Mestrado em Engenharia
ambiental) -Universidade Federal do Espírito Santo, Vitória, 2012.
GOUVÊA, A. V. et al. Avaliação da contaminação de amostras de soja com resíduos de
agrotóxicos pelo método QuEChERS acetato com análise por meio de CLAE-EM/EM.
Rev. Inst. Adolfo Lutz, São Paulo, v. 74, n. 3, p. 225-238, 2015. Disponível em:<
https://www.arca.fiocruz.br/handle/icict/14306> Acesso em 21 de setembro de 2019.
LACERDA, A.L.S.; LAZARINI, E.; SA, M.E.; WALTER FILHO, V.V. Aplicação de
dessecantes na cultura da soja: antecipação da colheita e produção de sementes. Planta
Daninha, Viçosa, v.19, n. 3, p.381-390, 2001.
LORENZI, H. Manual de identificação e controle de plantas daninhas: plantio direto e
convencional. Nova Odessa, 1984. 220 p
MACIEL, C. D. G.; FERREIRA, M. A. M.; POLETINE, J. P.; MONDINI, M. L. Uso de
adjuvantes na dessecação da cultura da soja: qualidade fisiológica de sementes. Revista
Científica Eletrônica de Agronomia, Garça, v.5, n.7, 2005.
MADALÃO, João Carlos et al. O herbicida sulfentrazone interfere na biomassa
microbiana e na atividade da microbiota do solo. Revista de Ciências Agrarias -
Amazon Journal Of Agricultural And Environmental Sciences, [s.l.], v. 59, n. 1, p.54-
59, 2016. Editora Cubo Multimidia. Disponivel em: http://dx.doi.org/10.4322/rca.2016.
Acesso em 25 de set de 2019

98
Belém (PA), 11 a 13 de dezembro de 2019
MAGALHÃES, P.C.; SILVA, J.B.; DURÃES, F.O.M.; KARAM, D; RIBEIRO, L.S.
Efeito de doses reduzidas de glyphosate e paraquat simulando deriva na cultura do sorgo.
Planta Daninha, Viçosa, v.19, n.2, p.255-262, 2001.
MARCHESAN, E.d. et al. Integration mulches with atrazine for weed management in
corn. Revista Brasileira de Ciências Agrárias - Brazilian Journal Of Agricultural
Sciences, [s.l.], v. 11, n. 1, p.1-7, 31 mar. 2016. Revista Brasileira de Ciencias Agrarias.
http://dx.doi.org/10.5039/agraria.v11i1a5353.
MELLONI, R; SILVE, E.M; ALVARENGA, M.I.N; MELLONI, E.G; ALCÂNTARA,
E.N. Impacto do controle de plantas espontâneas sobre propágulos de FMAs e
micorrização de cafeeiro. Coffe Science, Lavras, v. 12, n. 207, p.207-215, 05
set..2016..Bimestral.Disponível..em:..<http://www.sbicafe.ufv.br/handle/123456789/86
44>. Acesso em: 21 set. 2019.
OLIVEIRA, Luã Kramer de et al. Processo sócio-sanitário-ambiental da poluição por
agrotóxicos na bacia dos rios Juruena, Tapajós e Amazonas em Mato Grosso, Brasil.
Saúde e Sociedade, [s.l.], v. 27, n. 2, p.573-587, jun. 2018. FapUNIFESP (SciELO).
http://dx.doi.org/10.1590/s0104-12902018170904.
ORDOÑEZ-GÓMEZ, E.s.; REÁTEGUI-DÍAZ, D.; VILLANUEVA-TIBURCIO, J.e..
Total polyphenols and antioxidant capacity of peel and leaves in twelve citrus. Scientia
Agropecuaria, [s.l.], v. 9, n. 1, p.123-131, 28 mar. 2018. Scientia Agropecuaria.
http://dx.doi.org/10.17268/sci.agropecu.2018.01.13.
PAPA, Geraldo et al. Manejo de resistência de pragas a agrotóxicos. In: Embrapa Uva
e Vinho-Artigo em anais de congresso (ALICE). In: WORKSHOP RELAÇÃO
PRODUTIVA ENTRE AGRICULTURA E APICULTURA, 1., 2014, Belo Horizonte.
Anais. Belo Horizonte: SBDA, 2014., 2014.
PINTO, Rosecler Silva et al. Qualidade de grãos de soja em diferentes épocas de colheita.
Sinop Pesquisas Agrárias e Ambientais, Mato Grosso, v. 5, n. 7, p.463-470, 4
nov.2017.Mensal..Disponível..em:.<https://www.alice.cnptia.embrapa.br/alice/bitstream
/doc/1087506/1/2017cpamtsilviabotelhoqualidadegraossojadiferentesepocacolheita.pdf>
..Acesso em: 23 set. 2019.
PROGÊNIO, Mayke Feitosa. Modelagem estocástica de função cumulativa de
probabilidades de precipitação diária na região hidrográfica tocantins-araguaia (RHTA).
2019. 119 f. Dissertação (Mestrado) - Universidade Federal do Pará, Instituto de
Tecnologia, Belém, 2019. Programa de Pós-Graduação em Engenharia Civil. Disponível
em: <http://repositorio.ufpa.br/jspui/handle/2011/11148>. Acesso em 25 de novembro de
2019.
PRUDENCI, Marcelo Falaci; ANDREANI JÚNIOR, Roberto. AÇÃO DE
HERBICIDAS AUXÍNICOS NO CONTROLE DE GUANXUMA (Sida glaziovii) EM
PASTAGEM DE Urochloa decumbens. Nucleus, [s.l.], v. 16, n. 1, p.97-104, 30 abr.
2019. Fundacao Educational de Ituverava. Disponivel em:
http://dx.doi.org/10.3738/1982.2278.2950. Acesso dia 26 de novembro de 2019
SÁ, Klênia Rodrigues Pacheco; ROCHA, Hugo Leonardo da. ANÁLISE DE
CONTROLE QUÍMICO E BIOLÓGICO EM NEMATOIDES NA SOJA. 2018. 29

99
Belém (PA), 11 a 13 de dezembro de 2019
f. Dissertação (Mestrado) - Curso de Agronomia, Centro Universitário de Anápolis,
Anápolis, 2018. Disponível em: <http://repositorio.aee.edu.br/jspui/handle/aee/1006>.
Acesso em: 22 set. 2019.
SILVA, I. F.; FENILLI JUNIOR, A; LORENZETTI, E. Efeito de dessecantes na
produtividade e qualidade fisiológica de sementes de soja. Revista Cultivando O Saber,
Toledo, v. 9, n. 3, p.309-317, 4 set. 2016. Mensal. Disponível em:
<https://www.researchgate.net/profile/Itamar_Silva2/publication/313741415_Efeito_de
_dessecantes_na_produtividade_e_qualidade_fisiologica_de_sementes_de_soja/links/58
a4595caca27206d975b2d6/Efeito-de-dessecantes-na-produtividade-e-qualidade-
fisiologica-de-sementes-de-soja.pdf>. Acesso em: 24 set. 2019.
SOUSA, José Antônio de et al. PERCEPÇÃO DOS PRODUTORES RURAIS QUANTO
AO USO DE AGROTÓXICOS. Revista Brasileira de Agricultura Irrigada, [s.l.], v.
10, n.5, p.976-989, 31 out 2016. INOVAGRI.
http://dx.doi.org/10.7127/rbai.v10n500484.
SOUZA, Marco Aurélio Pessoa de et al. Remoção de agrotóxicos por escoamento
superficial – Princípios e práticas. Caderno de Ciências Agrárias, Planaltinha, v. 9, n.
3, p.119-125, 28 set 2017 Mensal. Disponível em:
<https://www.alice.cnptia.embrapa.br/bitstream/doc/1087792/1/HeleniceRemocaodeagr
otoxicosporescoamentosuperficial.pdf>. Acesso em: 22 set. 2019.
SVIRIDOV, A. V. et al. Microbial degradation of glyphosate herbicides (Review).
Applied Biochemistry And Microbiology, [s.l.], v. 51, n. 2, p.188-195, mar. 2015.
Pleiades Publishing Ltd. Disponivel em :http://dx.doi.org/10.1134/s0003683815020209.
Acesso em 26 de set de 2019.
TANG, Fiona H.m. et al. Microcosm experiments and kinetic modeling of glyphosate
biodegradation in soils and sediments. Science Of The Total Environment, [s.l.], v. 658,
p.105-115, mar. 2019. Elsevier BV. http://dx.doi.org/10.1016/j.scitotenv.2018.12.179.
TEIXEIRA-GANDRA, Claudia Fernanda Almeida et al. Cadeia de Markov no
preenchimento de falhas de dados diários de precipitação no Rio Grande do Sul.
Magistra, Cruz das Almas, v. 28, n. 2, p.157-167, 06 mar. 2017. Mensal. Disponível em:
<https://magistraonline.ufrb.edu.br/index.php/magistra/article/view/301/243>. Acesso
em: 23 set. 2019.
TEJADA, M. Evolution of soil biological properties after addition of glyphosate,
diflufenican and glyphosate+diflufenican herbicides. Chemosphere, 76:365-373, 2009.
TOLEDO, M.Z.; CAVARIANI, C.; FRANCA-NETO, J.B. Qualidade fisiológica de
sementes de soja colhidas em duas épocas após dessecação com glyphosate. Revista
Brasileira de Sementes, Londrina, v.34, n.1, p.134-142, 2012
VARGAS, Rp Flórez et al. Bacterial composition of the biofilm on valves of Limnoperna
fortunei and its role in glyphosate degradation in water. Aquatic Microbial Ecology,
[s.l.], v. 83, n. 1, p.83-94, 6 jun. 2019. Inter-Research Science Center.
http://dx.doi.org/10.3354/ame01907

100
Belém (PA), 11 a 13 de dezembro de 2019
ZOBIOLE, L.H.S.; KREMER, R.J.; OLIVEIRA, R.S. & CONSTANTIN, J. Glyphosate
affects microorganisms in rhizospheres of glyphosate-resistant soybeans. J. Appl.
Microbiol., 110:118-127, 2010.

101
Belém (PA), 11 a 13 de dezembro de 2019
PRODUÇÃO HIDROPÔNICA DE ALFACE INTEGRADA À PRODUÇÃO DE
PESCADO COMO FORMA DE TRATAMENTO DO EFLUENTE
Janete da Silva Souto1; Haroldo. Ferreira de Araújo2; Gessica de Lima Mota3.
1 Aluna do Curso Técnico em Agropecuária, Instituto Federal do Pará
(IFPA);[email protected] 2 Prof. Dr. Instituto Federal do Pará (IFPA), [email protected]
3 Técnico (o) em Agropecuária, Instituto Federal do Pará (IFPA).
RESUMO
Produzir com sustentabilidade é uma das principais prioridades globais da agricultura, e
encontrar métodos sustentáveis para aumentar a produção de alimentos tornou-se o foco
principal de muitas pesquisas agrícolas. Assim, objetivou-se com esta pesquisa avaliar a
viabilidade da produção de alface integrada à produção de pescado, utilizando seu
efluente sem tratamento prévio como solução nutritiva com duas densidades de peixes,
comparando sua eficiência produtiva com o sistema convencional. A pesquisa foi
desenvolvida no Campus experimental do IFPA/Campus Breves região do Marajó.
Utilizou-se bancadas de produção hidropônica alternativa (PVC 50 mm) alocadas em uma
casa de vegetação. Estas bancadas foram abastecidas com água de efluente da
piscicultura, advinda de dois sistemas intensivos de criação de peixes, a exceção do
tratamento testemunha que foi utilizada solução nutritiva convencional. Os sistemas de
produção verso criação funcionaram em circuito fechado, sendo a entrada e a saída de
água, controlada em função da programação de um timer analógico utilizado para o
controle da frequência de irrigação das bancadas hidropônicas. A análise dos dados de produção, mostraram uma média (g/planta) abaixo do padrão comercial, obtendo plantas
estioladas e com baixa qualidade comercial. Além disso, observou-se que a fonte hídrica
utilizada (água coletada do telhado do Instituto) interferiu na qualidade produtiva, visto
que esta não foi tratada previamente antes do uso, bem como, o excesso de excretas dos
peixes, principalmente no tanque com maior densidade de peixes. Assim, diante dos
resultados, concluiu-se que é possível produzir alface integrada à produção de pescado,
utilizando o efluente sem tratamento prévio como solução nutritiva, seja com uma média
de 4 e 6 peixes de 120 g/100 l de água, sendo equiparados a utilização de solução
convencional, desde que sanado os seus inconiventes.
Palavras-chave: Aquoponia. Produção alternativa. Efluente da piscicultura.
Área de Interesse do Simpósio: Agronomia

102
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Produzir com sustentabilidade é uma das principais prioridades globais da
agricultura, e encontrar métodos sustentáveis para aumentar a produção de alimentos
tornou-se o foco principal de muitas pesquisas agrícolas. Contudo, para Petrea et al.
(2016), a maximização do lucro continua sendo o objetivo principal de toda atividade
econômica, vislumbrando assim, a integração de sistemas de recirculação da aquicultura
com diferentes técnicas de aquaponia uma resposta aos dois requisitos mencionados
acima. Vale salientar, que piscicultura intensiva em cativeiro com fornecimento de
alimento a base de proteína (rações) acarreta na geração de efluentes contendo
concentrações elevadas de matéria orgânica e formas reduzidas de nitrogênio,
principalmente, nitrogênio amoniacal e nitrito que podem trazer sérios problemas
ambientais aos mananciais se não forem tratados previamente. Estas formas reduzidas de
nitrogênio são oriundas dos alimentos e excreta dos peixes depositadas na água, deixando
a mesma rica em nutrientes que podem ser utilizados para produção de culturas em
sistemas hidropônicos. Assim, esses sistemas podem ser utilizados como forma de
tratamento da água para posterior reutilização no sistema criação de peixe em circuito
fechado ou aberto (CORTEZ et al., 2009). Para isso, lança-se mão do sistema de
aquaponia que é a combinação das palavras “aquicultura” e “hidroponia” que é uma
técnica milenar de cultivo de vegetais sem o uso de solo, porém com o auxílio da criação
de peixes para suprir a demanda nutricional da cultura. Portanto, de forma simplificada
pode-se dizer que os peixes e minerais presentes na água auxiliam a nutrição das plantas,
as quais mantém a água em boas condições para os peixes (EQUILIBRIUM, 2016).
Apesar da importância que esse sistema pode representar, a literatura ainda é
limitada a cerca de informações concretas sobre a viabilidade produtiva do sistema, razão
pela qual, certamente não é difundido o sistema para tratamento do efluente e produção
agrícola em escala comercial (LOVE et al., 2015). Nessa linha de pesquisa, a EMBRAPA
meio norte (2013) vem pesquisando e lançou o “sisteminha” que consiste em um tanque
de piscicultura, construído artesanalmente, galinheiro, minhocário, hidroponia, abrigo
para compostagem, além de uma horta periférica, funcionando em sistema integrado. Este
sistema engloba uma unidade de produção familiar completa, diferenciando-se desta
pesquisa pelo enfoque dado nesta, que foi o tratamento da água de efluente através da

103
Belém (PA), 11 a 13 de dezembro de 2019
produção hidropônica para posterior reutilização. Sendo assim, observando a inclinação
natural da região do Marajó pela piscicultura intensiva em tanques, haja visto a
disponibilidade água, e consciente dos problemas causados pela liberação do efluente a
natureza, além da falta de produção agrícola na região, pesquisar alternativas que possam
contribuir para suprir essas necessidades deve ser prioridades. Dessa forma, objetivou-se
com a pesquisa avaliar a viabilidade da produção de alface integrada à produção de
pescado, utilizando seu efluente sem tratamento prévio como solução nutritiva com duas
densidades de peixes, comparando sua eficiência produtiva com o sistema convencional.
2. MATERIAL E MÉTODOS
2.1. CARACTERIZAÇÃO LOCAL
A pesquisa foi desenvolvida no Campus experimental do IFPA/Campus Breves
região do Marajó. O tipo climático da região de acordo com a classificação de Köppen, é
Af, pertence ao grupo de clima úmido, com precipitação média anual de 2356 mm bem
distribuídos ao longo do ano, e temperatura média anual de 26,8 °C.
2.2. CARACTERIZAÇÃO EXPERIMENTAL
As bancadas de produção hidropônica foram alocadas em uma casa de vegetação
instalada na orientação Norte-Sul, com área total de 300 m2 (15 x 20 m) e 2,50 m de pé-
direito, construída em madeira serrada. A cobertura em duas águas (tipo londrina com
lanternim de 0,50 m) foi coberta com plástico de polietileno de baixa densidade (PEBD)
de 120 micras e laterais fechadas com tela antiafídeos (Baby Citrus) malha de 50 mesh.
Estas bancadas foram abastecidas com água de efluente da piscicultura, advinda de dois
sistemas intensivos de criação de peixes, a exceção do tratamento testemunha que foi
utilizada solução nutritiva convencional. Utilizou-se como reservatórios de solução
nutritiva e tanques de criação de peixes, três caixas d’água de 100 l, sendo uma com
solução nutritiva previamente preparada, a segunda somente com 4 peixes e a terceira
com 6 peixes, sendo cada peixe contido nessas caixas com aproximadamente 120 g.
Os sistemas de produção verso criação funcionaram em circuito fechado, sendo a
entrada (água limpa) e a saída (efluente) de água, controlada em função da programação
de um timer analógico utilizado para o controle da frequência de irrigação das bancadas
hidropônicas. Assim, a mesma quantidade de água que saía para as bancadas de produção

104
Belém (PA), 11 a 13 de dezembro de 2019
de alface era reposta automaticamente nos tanques de criação de peixes, depois de
passado pelo sistema radicular das plantas.
A programação do timer durante o período diurno (06:00 as 19:00 h) seguiu o
padrão on/off (liga 15 minutos e passa 15 minutos desligada) de acionamento de três
bombas centrífugas de 3/4 cv, uma para cada forma de produção de peixes, além da
bancada convencional. Durante o período noturno (19:01 as 05:59 h), as bombas foram
acionadas somente nos horários de 10, 01 e 03 horas seguindo a mesma programação
on/off.
Os peixes foram alimentados com ração específica (26% de proteína) três vezes
ao dia, com a quantidade fornecida conforme seu crescimento, sendo esta feita
manualmente.
Foi utilizado três bancadas hidropônicas reduzidas construídas com tubos de PVC
de 50 mm, com capacidade para 35 mudas cada, (orifícios de 0,05 m de diâmetro e
espaçamento de 0,20 x 0,20 m), sendo mantida uma vazão total de solução de 7,5
l/min/bancada, distribuído 1,5 l/min a cada 0,20 m ou calha. Cada bancada foi instalada
com uma inclinação de 7% para facilitar o escoamento da solução aplicada.
Utilizou-se mudas da variedade de alface Mimosa SF31 (produção o ano todo),
previamente plantadas em viveiro em substrato oriundo da mistura de duas partes de terra
e 1 parte de esterco bovino curtido, sendo as mesmas transplantadas no dia (27/11/2018)
quando apresentavam três folhas verdadeiras, aproximadamente 25 dias depois da
semeadura. Antes do transplantio as plantas foram avaliadas e transplantas nas bancadas,
sendo reavaliadas a cada 5 dias até o ponto de colheita. As variáveis avaliadas foram: (1)
altura (cm); (2) número de folhas por planta; (3) comprimento e largura da maior folha
(cm) (comprimento da folha do início do pecíolo até o final do limbo foliar). No ponto de
colheita avaliou-se: (4) massa de matéria fresca da parte aérea das plantas (g planta-1); (5)
produtividade (g m-2) (massa de matéria fresca multiplicada pela quantidade de plantas
por metro quadrado).
2.3. TRATAMENTOS EXPERIMENTAIS
Foram avaliados os tratamentos A1 – água do tanque com 4 peixes; A2 – água do
tanque com 6 peixes e A3 – água com solução nutritiva convencional, fazendo uma média
de 4 repetições, sendo cada repetição a média da análise de quatro plantas. Estes
tratamentos foram agrupados e analisados estatisticamente pelo delineamento

105
Belém (PA), 11 a 13 de dezembro de 2019
Inteiramente Casualisados (DIC), com comparação das médias pelo teste de Tukey. No
tratamento testemunha a adubação seguiu a recomendação de Trani et al. (2011) para
produção de alface em solo, mantendo a condutividade elétrica (EC) da solução nutritiva
nos primeiros 10 dias a 0,7 dS m-1 e posteriormente a 1,3 dS m-1 até o final do ciclo.
3. RESULTADOS E DISCUSSÃO
As análises de variância dos dados mostraram que não houve diferença estatística
entre os tratamentos utilizados, contudo, foi realizado o teste de comparação das médias
apresentados na Tabela 01.
Em termos produtivos, o sistema com a utilização de quatro peixes apresentou
produtividade superior aos demais tratamentos em cultivo de 22 dias (0, 938 kg m-2),
porém abaixo do verificado por PINHO et al. (2017), que obtiveram valores de
produtividade média de 1,20 kg m-2 para a mesma variedade em sistema aquoponico e 21
dias de cultivo. Porém próximo ao valor observado por LENZ et al. (2017), que obtiveram
valor de 1,04 kg m-2 em cultivo de 28 dias.
Em termos de padrão comercial, a média verificada ficou muito abaixa dos valores
praticados no mercado, que são de 100 a 300 g/planta (SANTOS et al., 2011). Isso pode
ter ocorrido devido as características climáticas da região que, naturalmente são de alta
temperatura e umidade relativa do ar. Os feitos destas variáveis contribuíram diretamente
para o resultado final da produção, obtendo plantas estioladas e com baixa qualidade
comercial. Além dessas variáveis, a fonte hídrica utilizada (água coletada do telhado do
Instituto) também interferiu, visto que esta não foi tratada previamente antes do uso. Esta
interferência diz respeito a proliferação de microrganismos (algas fotossintéticas - lodo)
que acabam por dificultar a absorção de nutrientes pelas raízes das plantas.
Outro problema verificado foi o excesso de excretas dos peixes, principalmente
no tanque com maior densidade, havendo a necessidade de retirada destas a cada 5 dias.
Mesmo retirando parte destas excretas, parte ficavam acumuladas nas raízes das plantas,
ocasionado a morte das mesmas. Assim, recomenda-se o uso de câmaras de filtragens
antes da injeção da solução do efluente nas raízes das plantas para amenizar esse
problema.

106
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 01 - Médias dos dados das análises das plantas de alface em função do seu
desenvolvimento e tratamentos, Breves - PA.
Tratamentos DAP C.
Folha
(cm)
D.
Folha
(cm)
L.
Folha
(cm)
N.
Folhas
D.
Colo
(mm)
Produção
(g/planta) Produtividade
(g m2)
Convencional
5 6,66 a 4,33 a 1,50 a 6,25 a
10 8,70 a 6,18 a
2,083
a 7,69 a
16 11,31
a 8,045 a 5,409
a 6,72 a
22 10,95
a 9,416 a 2,750
a 8,50 a
29 16,41
a 12,500 a 4,416
a 9,91 a 6,30 a 37,17 a 929,17 a
4 Peixes
5 7,82 a 4,607 a
1,714
a 6,28 a
10 9,32 a 6,700 a
2,100
a 7,46 a
16 11,5 a 7,633 a
4,633
a 8,00 a
22 13,63
a 9,433 a 3,533
a 8,86 a
29 16,40
a 13,733 a 4,733
a 10,53 a 7,57 a 37,53 a 938,33 a
6 Peixes
5 7,83 a 4,666 a
1,733
a 6,00 a
10 10,34
a 7,066 a 2,166
a 7,86 a
16 11,40
a 8,033 a 3,900
a 7,93 a
22 14,13
a 10,800 a 3,100
a 9,53 a
29 15,43
a 12,666 a 4,500
a 10,33 a 7,31 a 33,47 a 836,67 a
DAP: Dias Após o Transplantio; C: comprimento; D: diâmetro; L: largura; N: número; médias
seguidas da mesma letra e mesma coluna e DAP não se diferem estatisticamente pelo teste de
Tukey (p<0,05).
Fazendo uma comparação em termos práticos desse sistema ao “Sisteminha” da
EMBRAPA que integra vários sistemas produtivos, esta pesquisa demonstrou viabilidade
técnica por tratar a água utilizada da piscicultura, que na região do Marajó já é uma
atividade culturalmente praticada. Além disso, foi possível inserir a produção vegetal a
atividade pesqueira regional para viabilizar a produção de folhosas.
4. CONCLUSÃO

107
Belém (PA), 11 a 13 de dezembro de 2019
Diante dos resultados, concluiu-se que é possível produzir alface integrada à
produção de pescado, utilizando o efluente sem tratamento prévio como solução nutritiva,
seja com uma média de 4 e 6 peixes de 120 g/100 l de água, sendo equiparados a utilização
de solução convencional, desde que sanado os seus inconiventes. O sistema pesquisado
apresentou viabilidade técnica em termos práticos, sendo possível a expansão das
atividades produtivas integrada (Peixes e folhosas).

108
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
CORTEZ, G. E. P., ARAÚJO, JAIRO A. C.; BELLINGIERI, P. A.; DALRI, A. B.
Qualidade química da água residual da criação de peixes para cultivo de alface em
hidroponia. Revista Brasileira de Engenharia Agrícola e Ambiental v.13, n.4, p.494–
498, 2009.
EMBRAPA - EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA (MEIO
NORTE - 2013). Disponível em <https://www.embrapa.br/busca-de-publicacoes/-
/publicacao/958146/sistema-integrado-alternativo-para-producao-de-alimentos-
agricultura-familiar>. Acessado em 20 de novembro de 2019.
EQUILIBRIUM – SISTEMA DE AQUOPONIA - Aquaponia, o que é (2016).
Disponível em <http://equilibrium.org.br/ees-aquaponia/sobre/>. Acessado em 22 de
fevereiro de 2018. LENZ, G. L, DURIGON, E. G.; LAPA, K. G.; EMERENCIANO M. G. C. Produção de alface
(Lactuca sativa) em efluentes de um cultivo de tilápias mantidas em sistema BFT em baixa
salinidade. Bol Inst Pesca, n,43., p-614–630. 2017.
LOVE, D. C.; FRY, J. P.; LI, X.; HILL, E. S.; GENELLO, L.; SEMMENS, K.;
THOMPSON, R. E. Commercial aquaponics production and pro fi tability: Findings from
na international survey. Aquaculture, n.435. p.67–74. 2015.
SANTOS, D.; MENDONÇA, R. M. N.; SILVA, S. M.; ESPÍNOLA, J. E. F.; SOUZA,
A. P. Produção comercial de cultivares de alface em Bananeiras. Horticultura.
Brasileira. v.29 n.4, p.609-612, 2011.
PETREA, S. M.; COADÃ, M. T.; CRISTEA, V.; DEDIU, L.; CRISTEA, D.; et al. A
Comparative Cost – Effectiveness Analysis in Different Tested Aquaponic Systems.
Agriculture and Agricultural Science Procedia, n.10, p.555 – 565, 2016.
PINHO, S.M.; MOLINARI, D.; MELLO, G. L.; FITZSIMMONS, K. M.;
EMERENCIANO, M. G. C. Effluent from a biofloc technology (BFT) tilapia culture on
the aquaponics production of different lettuce varieties. Ecological Engineering, n.103.
p-146-153, 2017.
TRANI, P. E.; TIVELLI, S. W.; CARRIJO, O. A. Fertirrigação em Hortaliças, série
tecnologia apta. Boletim Técnico IAC, Campinas, n. 196, 2011.

109
Belém (PA), 11 a 13 de dezembro de 2019
QUALIDADE FITOSSANITÁRIA E GERMINATIVA DE SEMENTES DE
MILHO SUBMETIDAS A TERMOTERAPIA
Amada de Nazaré Rodrigues Pereira 1; Sayure Mariana Raad2; Erick dos Santos Ribeiro
3; Rayanne de Jesus Castro4; Adélia Benedita Côelho dos Santos5.
1 Engenheira Agrônoma. Universidade Federal Rural da Amazônia (UFRA). E-mail:
2 Discente do curso de agronomia da Universidade Federal Rural da Amazônia(UFRA).
E- mail: [email protected]
3Discente do curso de agronomia da Universidade Federal Rural da Amazônia(UFRA).
E-mail: [email protected]
4Discente do curso de agronomia da Universidade Federal Rural da Amazônia(UFRA).
E-mail: [email protected].
5Professora de Fitopatologia da Universidade Federal Rural da Amazônia, Ms. Em
Agronomia, Universidade Federal do Ceará. E-mail: [email protected].
RESUMO
O Brasil encontra-se como o 3º maior produtor de milho no mundo e 2º maior exportador,
sendo, um dos principais produtores mundiais de proteína animal. No entanto, para isso,
se faz necessário ter semente de boa qualidade fisiológica e sanitária para um adequado
estabelecimento das plantas no campo. Com isso, o presente trabalho teve por objetivo
avaliar a eficiência da termoterapia no controle de patógenos de sementes de milho e seu
poder germinativo. O presente trabalho foi desenvolvido no laboratório de Fitopatologia
da Universidade Federal Rural da Amazônia, Campus Belém, PA. Para isso, o
delineamento experimental utilizado foi o inteiramente casualizado (DIC), com 6
tratamentos e cinco repetições, cada repetição foi composta por um lote de 20 sementes.
Os tratamentos térmicos empregados foram por imersão em água aquecida nas seguintes
temperaturas: 40oC, 50oC, 60oC, 70oC e 80oC, sendo denominados, T2, T3, T4, T5, T6 e
T7, respectivamente. Após as sementes serem submetidas aos tratamentos, analisou-se a
sanidade para a determinação da eficiência dos tratamentos na redução fúngica das
sementes e germinação. A termoterapia mostrou-se eficiente em relação a testemunha em
todos os tratamentos na redução fúngica das sementes, dando destaque para os
tratamentos T5 e T6. Na germinação, esse método obteve resultados positivos nos
tratamentos T2 e T3, obtendo 60% e 98% de germinação em relação a testemunha. Com
isso, a termoterapia demostrou ser um método alternativo positivo na redução do
potencial fúngico das sementes de milho e no aumento do poder germinativo em
determinadas temperaturas.
Palavras-chave: Tratamento de sementes. Método alternativo. Germinação.
Área de Interesse do Simpósio: Agronomia

110
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A cultura do milho (Zea mays L.) pertence à família Poaceae, sendo uma das
principais espécies cultivadas no mundo. Apresenta elevada demanda agrícola, tendo seu
uso destinado tanto para consumo na bovinocultura, suinocultura e piscicultura, quanto
para na alimentação humana. A utilização de sementes certificadas está interligada aos
acréscimos tecnológicos em lavouras, nas mais diferentes culturas, essas por sua vez
desenvolvidas e produzidas conforme os exigentes padrões do Sistema Brasileiro de
Sementes e Mudas (MIYAMOTO, 2013).
No decorrer das últimas décadas, o milho alcançou o patamar de maior cultura
agrícola do mundo, sendo a única a ter ultrapassado a marca de 1 bilhão de toneladas,
deixando para trás antigos concorrentes, como o arroz e o trigo. Concomitantemente à
sua importância em termos de produção, a cultura ainda se notabiliza pelos diversos usos.
Estimativas apontam para mais de 3.500 aplicações deste cereal. Além da relevância no
aspecto de segurança alimentar, na alimentação humana e, principalmente, animal, é
possível produzir com o milho uma infinidade de produtos, tais como combustíveis,
bebidas, polímeros, etc. (MIRANDA, 2018).
Da safra de 2000/01 para a de 2017/18, a produção mundial de milho passou de
591 milhões de toneladas para 1,076 bilhão de toneladas (representando um aumento de
82%), por causa principalmente do uso como ração animal para a produção de frangos e
suínos. A produção é relativamente concentrada em poucos países, com destaque para os
Estados Unidos, com 34,5% (371 milhões de toneladas) da produção mundial, seguidos
da China, com 24,5% (263 milhões de toneladas) (CONTINI et al., 2019). No entanto, a
associação de fungos fitopatogênicos a sementes de milho é responsável, muitas vezes,
pela redução da qualidade fisiológica das sementes e baixa produtividade. Para isso, a
eliminação ou redução do inóculo infectivo de fungos em sementes, tem sido
eficientemente alcançada por tratamentos químicos, biológicos e físicos. Dentre os
métodos físicos, a eficácia da termoterapia, que consiste na exposição das sementes à
ação do calor em combinação com o tempo de tratamento, tem sido demonstrada em
vários estudos (Bergman, 1994; Trigo et al., 1998; Machado, 2000).

111
Belém (PA), 11 a 13 de dezembro de 2019
Desta forma neste trabalho, objetivou-se avaliar o tratamento de sementes de
milho por termoterapia em diferentes temperaturas, avaliando-se a sanidade das sementes,
bem como o efeito destes na germinação.
1.1 SANIDADE E GERMINAÇÃO DE SEMENTES
Há um grande número de fatores que afetam a qualidade das sementes. Dentre
eles podemos citar os fatores sanitários, que se caracterizam pelo efeito deletério
provocado pela ocorrência de microrganismos e insetos associados às sementes, desde o
campo de produção até o armazenamento (LUCCA FILHO, 2003). Esses fatores são mais
intensificados em virtude da semente ser um veículo para distribuição e disseminação de
patógenos, os quais podem, às vezes, causar surtos de doenças nas plantas, pois pequenas
quantidades de inóculo na semente podem ter uma grande significância epidemiológica
(PESKE; BARROS, 2003)
A germinação de sementes ocorre sob limites relativamente amplos de
temperatura, sendo que os extremos dependem principalmente da espécie e suas
características genéticas (MATHEUS E LOPES, 2009). Cada espécie apresenta
temperaturas cardeais para a germinação de sementes, ou seja, a temperatura máxima,
mínima e ótima para que o processo aconteça (MARCOS FILHO, 2005). Para sementes
de milho, as temperaturas cardeais são: 9 °C (mínima), 32-35 °C (ótima) e 44 °C
(máxima) (BORBA et al., 1995).
1.1.1 Termoterapia
Zambolim (2004) relata que a termoterapia é uma das técnicas mais citadas e
descritas na literatura para erradicação de fitobactérias localizadas interna ou
externamente nas sementes. O sucesso do tratamento térmico depende do tipo de calor,
seco ou úmido, da temperatura empregada, do período de exposição e da uniformidade
da aplicação do calor (MENDES et al., 2001), devendo ser cuidadosamente ajustados à
espécie, cultivar e lote. Contudo, seja qual for a modalidade empregada, a termoterapia
requer o uso de equipamentos com controle preciso de temperatura, apresentando,
consequentemente, inconvenientes quando aplicada a grandes quantidades de sementes
(ESTEFANI; MIRANDA FILHO; UESUGI, 2007).

112
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
Os ensaios foram conduzidos nos Laboratórios de Fitopatologia da Universidade
Federal Rural da Amazônia (UFRA), no período de 2018. Para o ensaio foram utilizadas
sementes de milho (Zea mays L.), com viabilidade determinada por meio de testes de
germinação, onde se obteve 98% de germinação, teste esse realizado segundo a
metodologia de Brasil (1992).
Para a condução do experimento foi utilizado o delineamento inteiramente
casualizado (DIC), com 6 tratamentos e cinco repetições, cada repetição foi composta por
um lote de 20 sementes. Os tratamentos térmicos empregados foram por imersão em água
aquecida nas seguintes temperaturas: 40oC, 50oC, 60oC, 70oC e 80oC, sendo
denominados, T2, T3, T4, T5, T6 e T7, respectivamente. O tratamento testemunha, que
corresponde ao tratamento T1, não foi submetido a termoterapia. As sementes foram
acondicionadas em sacos de filó, os quais foram dispostos aleatoriamente em banho-
maria com água aquecida nas temperaturas correspondentes aos seus tratamentos em um
tempo de 5 minutos.
Após os tratamentos térmicos, as sementes foram postas para secar sobre papel
toalha a temperatura ambiente, e distribuídas sobre uma folha de papel germitest em caixa
tipo gerbox (11cm x 11cm x 4cm). Após a distribuição das sementes no gerbox,
umedeceu-se o papel filtro com um volume de água equivalente a 2,5 vezes a massa do
papel seco. Para o teste de germinação, as contagens foram realizadas aos quatro e sete
dias após a semeadura; as avaliações foram realizadas de acordo com os critérios
estabelecidos pelas Regras para Análise de Sementes (Brasil,1992). Já para o teste de
sanidade das sementes submetidas aos tratamentos, as avaliações dos fungos associados
às sementes foram realizadas após oito dias de incubação, examinando-se,
individualmente, as sementes ao microscópio estereoscópico. Em alguns casos, a
identificação foi confirmada pela visualização das estruturas morfológicas dos fungos ao
microscópio óptico. Os dados coletados foram submetidos à análise de variância, e as
médias comparadas pelo teste de Tukey, a 5% de probabilidade, utilizando o programa
estatístico SISVAR.

113
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
Os resultados referentes à ocorrência (porcentagem de sementes com fungos) de
sementes de milho submetidas à termoterapia por diferentes temperaturas são
apresentados na figura 1.
Figura 1 –Incidência de fungos associados a sementes de milho sob diferentes temperaturas.
Fonte: Autores (2019).
Na avaliação de sanidade das sementes de milho foram encontrados os seguintes
fungos: Aspergillus flavus, Penicillium sp e Fusarium moniliforme. Para Aspergillus
flavus foram observados efeitos significativos (P≤0,05) em relação as diferentes
temperaturas do tratamento térmico. Em avaliação, é possível observar que com o
aumento das temperaturas, houve diminuição na incidência do fungo, principalmente no
tratamento T6(80oC). Já para o fungo Penicillium sp, o tratamento térmico T3(40oC) e
T5(70oC) mostrou ser mais eficiente no tratamento deste fungo. Avaliando a incidência
de Fusarium moniliforme, as sementes submetidas aos tratamentos, foi possível constatar
a redução da ocorrência do fungo no tratamento térmico T5(70oC) e a sua eliminação nas
temperaturas de tratamento de T4(60oC) e T6(80oC). Mendes et al. (2001) observaram
que sementes de alfafa submetidas a tratamento térmico seco de 60°C e 90°C não
apresentaram desenvolvimento de Fusarium oxysporum.

114
Belém (PA), 11 a 13 de dezembro de 2019
Em trabalhos encontrados na literatura, com outras culturas, o uso da termoterapia
foi eficaz no controle de alguns patógenos, não evidenciando implicações negativas
(ERDEY et al., 1997). No entanto, alguns fatores podem interferir nos resultados da
termoterapia, como o tipo e procedência das sementes, ou seja, efeitos variáveis do
tratamento térmico podem ser observados em relação a diferentes cultivares de uma
mesma espécie ou diferentes lotes de um mesmo cultivar (GROOT et al., 2006).
Na Tabela 1 verificam-se os resultados de percentagem de germinação de
sementes de milho, submetidas à transformação arco seno raiz quadrada de x/100.
Conforme Tabela 1, verificaram-se diferenças significativas entre os tratamentos.
Observou-se que os tratamentos T2(40oC) e T3(50oC) diferiram significativamente dos
demais tratamentos, apresentando percentagem de germinação iguais a 60% e 98% em
relação a testemunha, respectivamente. Outro ponto relevante a se observar é que a partir
do tratamento T4 houve diminuição na porcentagem de germinação da semente estudada.
No tratamento de sementes de milho com água quente, Coutinho et al. (2007) também
relataram a redução significativa do potencial fisiológico de sementes tratadas a 60 °C.
Tabela 1 – Percentagem de germinação (%) de sementes de milho submetidas a diferentes
temperaturas.
Tratamentos T1 T2 T3 T4 T5 T6 % germinação 52 a 60 ab 98 b 32 bc 30 bc 0 c
Fonte: Autores (2019).
4. CONCLUSÃO
A termoterapia mostrou-se eficiente no controle dos fungos das sementes milho,
porém existiu diferença na ação dos tratamentos térmicos sobre os diferentes fungos que
ocorreram. Todos os tratamentos térmicos reduziram os fungos Aspergillus flavus,
Penicillium sp e Fusarium moniliforme. Os tratamentos T4 (60oC) e T6(80o) erradicaram
o fungo Fusarium moniliforme.
Os resultados do presente trabalho também apresentaram boas perspectivas de
utilização dos tratamentos térmico na germinação de sementes de milho, com resultados
positivos nos tratamentos T2(40oC) e T3(50oC), dando destaque a este último que teve o
maior poder germinativo.

115
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BERGMAN, S. Hot treatment of seed-borne fungi on cereals. Seed Pathology and
Microbiology 5:20-21. 1994.
BRASIL. Ministério da Agricultura e Reforma Agrária. Secretaria Nacional de Defesa
Agropecuária. Departamento Nacional de Defesa Vegetal. (1992). Regras para análise
de sementes. Brasília, 1992. 365p.
BORBA, C.S.; ANDRADE, R.V.; AZEVEDO, J.T.; ANDREOLI, C.; PURCINO,
A.A.C. Germinação de sementes de diversos genótipos de milho tropical (Zea mays L.)
em diferentes temperaturas. Revista Brasileira de Sementes, v.17, n.2, p.141-144, 1995.
CONTINI, E.; MOTA, M. M.; MARRA, R.;BORGHI, E.; MIRANDA, R, A.; SILVA,
A. F.; SILVA, D. D.; MACHADO, J. R. A; COTA, L. V.; COSTA, R. V.; MENDES, S.
M. Milho: Caracterização e desafios tecnológicos. Embrapa, 2019.
COUTINHO, W. M., SILVA-MANN, R.; VIEIRA, M. G. G. C., MACHADO, C. F.;
MACHADO, J. C. Qualidade sanitária e fisiológica de sementes de milho submetidas à
termoterapia e condicionamento fisiológico. Fitopatologia Brasileira, Brasília, v. 32, n.
6, p. 458-464, 2007.
ESTEFANI, R.C.C.; MIRANDA FILHO, R.J; UESUGI, C.H. Tratamento térmico e
químico de sementes de feijoeiro: eficiência na erradicação de Curtobacterium
flaccumfaciens pv flaccumfaciens e efeitos na qualidade fisiológica das sementes.
Fitopatologia Brasileira, v.32, n.5, p.434-438, 2007.
ERDEY, D. P.; MYCOCK, D. J.; BERJAK, P. The elimination of Fusarium moniliforme
(Sheldon) infection in maize caryopses by hot water treatment. Seed Science and
Technology, v. 25, n. 3, p. 485-501, 1997.
GROOT, S.P.C.; BIRNBAUM, Y.; ROP, N.; JALINK, H.; FORSBERG, G.;
KROMPHARDT, C.; WERNER, S.; KOCH, E. Effect of seed maturity on sensitivity of
seeds towards physical sanitation treatments. Seed Science and Technology, v.34, p.403-
413, 2006.
LUCCA FILHO, O. A. Patologia de Sementes In.: PESKE, S. T.; ROSENTHAL, M. D.;
ROTA, G. R. M. Sementes : Fundamentos Científicos e Tecnológicos, 1. ed., 2003.
MACHADO, J.C. Tratamento de sementes no controle de doenças. Lavras MG.
Editora UFLA. 2000.
MARCOS FILHO, J. Fisiologia de sementes de plantas cultivadas. Piracicaba:
FEALQ, 1. ed. 2005. 495p.
MATHEUS, M.T.; LOPES, J.C. Temperaturas cardinais para a germinação de sementes
de Erythrina variegata L.. Revista Brasileira de Sementes, v.31, n.3, p. 115-122, 2009.
MENDES, M. S. S.; LIMA, P. M. M. P.; FONSECA, J. N. L.; SANTOS, M. de F.
Erradicação de Fusarium oxysorum em sementes de alfafa utilizando termo e
quimioterapia. Fitopatologia Brasileira, Brasília, v. 30, n. 2, p. 148-152, jun. 2001.

116
Belém (PA), 11 a 13 de dezembro de 2019
MIRANDA, R. A. de. Uma história de sucesso da civilização. A Granja, v. 74, n. 829,
p. 24-27, jan. 2018.
MIYAMOTO, Y. Semente, a mãe da agricultura. Brasília: Anuário Abrasem 2013,
Brasília p. 62-63. 2013.
PESKE, S. T.; BARROS, A. C. S. A. Produção de sementes. In: PESKE, S. T.;
ROSENTHAL, M. D.; ROTA, G. R. M. Sementes: Fundamentos Científicos e
Tecnológicos, 1. ed., 2003.
TRIGO, M.F.O., PIEROBOM, C.R., NEDEL, J.L. & TRIGO, L.F.N. Tratamento térmico
em sementes de cenoura. Pesquisa Agropecuária Brasileira 33:357-361. 1998.
ZAMBOLIM, L. Importância do tratamento de sementes no manejo integrado de
doenças. In: SIMPÓSIO BRASILEIRO DE PATOLOGIA DE SEMENTES, 8, 2004,
João Pessoa, PB. Palestras, João Pessoa: Tropical Hotel Tambaú, p.94-94, 2004.

117
Belém (PA), 11 a 13 de dezembro de 2019
SELEÇÃO DE ISOLADOS DE Trichoderma sp. PROMOTORES DE
CRESCIMENTO DE MUDAS DE COQUEIRO
Aline Figueiredo Cardoso1; Waldiney Xavier Ferreira2; Jennyfer Silva Campos3; Paulo
Manoel Pontes Lins4; Gisele Barata da Silva5
1Doutoranda. Universidade Federal Rural da Amazônia. [email protected]
2Engº Agrônomo. Universidade Federal Rural da Amazônia. [email protected] 3Graduanda do curso de Agronomia. Universidade Federal Rural da Amazônia.
[email protected] 4Doutor. Superintendente Agrícola-fazenda reunidas Sococo. [email protected]>
5Doutora. Professora da Universidade Federal Rural da Amazônia.
4.
RESUMO
O coqueiro (Cocos nucifera L.) é uma palmeira de grande importância no Brasil e no
mundo, principalmente pela diversidade de produtos e geração de emprego ao longo da
cadeia produtiva. Um dos principais problemas encontrados no desenvolvimento inicial
de mudas de coqueiro está na baixa taxa de geminação e obtenção de plantas com
deformação. Assim, o efeito que fungos do gênero Trichoderma podem exercer sobre o
desenvolvimento vegetal, tem papel importante no incremento das características de
crescimento, e pode tornar-se uma estratégia para a promoção de crescimento. O objetivo
do estudo foi selecionar isolados de Trichoderma que promovam o crescimento de mudas
de coqueiro combinado a redução de adubação com NPK. O experimento foi realizado
na fazenda comercial Reunidas Sococo, localizada no município de Santa Isabel-PA, com
três tratamentos: controle (T1), Mix de Trichoderma asperellum (T.06, T.09, T.12 e T.52)
(T2) e Mix de isolados de Trichoderma sp. (T09, T17 e T19) (T3), com sete repetições
cada. As avaliações realizadas foram: altura, diâmetro do coleto, massa fresca e seca de
raiz e parte aérea e teor de água. As mudas de coco tiveram incremento positivo nos dois
tratamentos com Trichoderma, destacando o tratamnto Trichoderma (T09, T17 e T19)
com incrementos de 25% em altura, 17% no diâmetro do coleto, 23% em massa seca da
parte aérea, 18% em massa seca da raiz e 13% em teor de água. O uso de Trichoderma
em mudas de coqueiro em fibra de coco promovem o crescimento das mudas.
Palavras-chave: Crescimento. Sustentabilidade. Cocus nucifera L.
Área de Interesse do Simpósio: Agronomia

118
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O coqueiro é uma planta perene originada no sudeste asiático (PERSLEY, 1992),
da espécie Cocos nucifera L., única do gênero Cocos (TEULAT et al., 2000). A produção
mundial desta cultura é de mais de 60 milhões de toneladas de frutos com uma área
plantada de mais de 110 milhões de hectares. O Brasil está entre os maiores produtores
mundiais, assumindo o quinto lugar em termo de produção, destacando-se o estado do
Pará como o terceiro maior produtor do país (IBGE/LSPA, 2017). Na busca por técnicas
mais eficientes na propagação de plantas, os substratos assumem cada vez maior
importância na área de fruticultura, principalmente como meio de crescimento e suporte
do sistema radicular das plantas e mudas em recipientes (BOSA et al., 2003; KLEIN et
al., 2000; KÄMPF, 2000; CARVALHO e NAKAGAWA, 2012). As características mais
desejadas para mudas frutíferas são homogeneidade, vigor, sanidade e baixa mortalidade
durante o estabelecimento (GALVÃO et al., 2007; NOMURA et al., 2008). A fibra de
coco possui grande durabilidade sem alteração de suas características físicas, baixo custo,
e retenção de água (BARRETO et al., 2011; COSTA et al., 2007).
O uso de fungos do gênero Trichoderma tem sido amplamente estudado pela
capacidade de promover o crescimento de plantas a partir da modulação de hormônios de
crescimento, que por sua vez induzem crescimento de parte aérea e radicular, favorecendo
maior área de contato, maior absorção de água e nutrientes, e consequentemente maior
acúmulo de biomassa (FILHO et al., 2008; SILVA et al., 2011; GAVA e MENEZES,
2012). Possuem também a capacidade em auxiliar no melhor aproveitamento de
nutrientes que podem não estar disponível para absorção pela planta, tais como a
solubilização fosfatos, mineralização da matéria organica e produção de siderofóros
(JALAL et al., 1987; ALTOMARE et al., 1999).
O objetivo do estudo foi selecionar isolados de Trichoderma que promovam o
crescimento de mudas de coqueiro e otimizem a fertilização química.
2. MATERIAL E MÉTODOS
O experimento foi realizado em viveiro comercial de produção de mudas de coco
no município de Santa Isabel-PA.

119
Belém (PA), 11 a 13 de dezembro de 2019
Delineamento experimental
O experimento foi inteiramente casualizado com três tratamentos: Controle (T1),
Mix de T. asperellum + 33% da adubação (T2), e Trichoderma sp.+ 33% da adubação
(T3), com sete repetições.
Obtenção de isolados
Os isolados fazem parte da coleção de microrganismo do Laboratório de proteção
de Plantas-LPP da Universidade Federal Rural da Amazônia-UFRA.
Preparo de mudas de coqueiro
As sementes de coco foram depositadas em germinadouros contendo substrato de fibra
de coco, a germinação ocorreu no período de 90 dias, logo após, foram transferidas para
sacos de polietileno de dimensões 40 x 40 cm, contendo substrato fibra de coco.
Preparo da suspensão
A suspensão de conídios foi preparada com adição de 10mL de água estéril em
placa contendo colônia dos isolado de fungo Trichoderma, todo conteúdo sendo
adicionado em tubo de vidro e homogeneizado com auxílio do vortex, seguido de
diluição, sendo retirado 1mL da suspensão original e colocadas em 9mL de água estéril
até a concentração 10-8 (108 UFC.mL- 1), segundo França et al., (2014).
Preparo das sementes: microbiolização
As sementes foram submersas na suspensão de conídios, para ajuste na
concentração de 10-8 ufc foi utilizada câmara de neubauer, levadas à mesa agitadora a 114
rpm e temperatura de 27°C por 24horas.
Avalições
Foram realizadas avaliações biométricas das mudas, como a altura e diâmetro do
coleto, utilizando um paquímetro, determinação da massa seca (parte aérea e raiz), através
do processo de secagem das amostras em estufa a 65°, seguida da pesagem do material,
e, por fim, a quantificação do conteúdo de água através da diferença entre o peso fresco
e o peso seco das amostras.
Análise estatística
Os dados foram submetidos a análise de variância e comparadas pelo teste de SNK
a 5% de probabilidade (P<0,05) utilizando o software Sisvar versão 5.6.

120
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
O substrato fibra de coco contribuiu para a adaptação dos isolados de Trichoderma
neste estudo, provavelmente em função de suas características físicas e químicas. O que
resultou em mudas com maior desenvolvimento radicular, possibilitando maior absorção
de água e nutrientes pelas mudas de coqueiro (ARAGÃO, 2002; Hernández et al., 2005).
As plantas inoculadas com Mix de Trichoderma de rizosfera de coqueiro
apresentaram incremento em todas as variáveis biométricas. Os incrementos foram em
25% em altura, 17% em diâmetro do coleto, 23% em massa seca da parte aérea e em 18%
em massa seca radicular (Figura 1). Estes resultados estão associados a capacidade do
fungo de colonizar a epiderme das raízes das plantas e alterar a arquitetura do sistema
radicular, aumentando principalmente o número de pêlos radiculares (GONZALES et al.,
2018), possibilitando aumento na absorção de nutrientes, além do aumento no teor de
água como ocorreu neste estudo. O crescimento radicular está relacionado ao maior
estimulo com uso de Trichoderma na síntese de hormônios vegetais, dentre eles o AIA
(Ácido-indol-acético) e da enzima ACC diaminase que diminui a biossíntese de etileno
que são responsáveis pelo alongamento e diferenciação radicular (OLIVEIRA et al.,
2012).
Figura 1. Variáveis biométricas de mudas de coqueiro em fibras de coco. Altura da muda (A),
Diâmetro do coleto (B), Massa seca parte aérea (C), Massa seca da raiz (D), Teor de Água (E).
Controle (T1), T. asperellum + 33% da adubação (T2), e Trichoderma sp.+ 33% da adubação
(T3). * letras iguais não diferem entre si a 5% de probabilidade pelo teste SNK.

121
Belém (PA), 11 a 13 de dezembro de 2019
(C) (D)
O uso de Trichoderma asperellum em planta de Pinus taeda em tubetes, em casa
de vegetação, promoveu o aumento no diâmetro das plantas (PEREIRA., 2017), podendo
ser resultado do aumento nas concentrações de giberelina, hormônio responsável pelo
crescimento de caule e lançamento de folhas novas, o que pode ter ocorrido nas mudas
de coqueiro inoculadas com Trichoderma sp. e com 33% da adubação (CASSÁN et al.,
2003; LUCON, 2009; TAIZ e ZEIGER, 2009), além de que este tratamento demostrou
eficiência no uso eficiente dos nutrientes favorecendo então o maior acúmulo de
biomassa. Segundo Prates et al., (2007), Altomare et al., (1999) e Brotman et al., (2010),
alguns isolados de Trichoderma, principalmente associado com substrato, tem capacidade
mineralizar fósforo (P), solubilizar e, em alguns casos, quelar compostos do substrato ao
qual estão inclusos.
Neste estudo, o substrato fibra de coco possui 92% de matéria orgânica, e a
combinação do Mix de Trichoderma pode ter acelerado o processo de mineralização da
MO (SUGITO et al., 1995). A utilização de T. Harzianum acelerou a degradação de
materiais difíceis como lignina e celulose, resultando em melhora do crescimento e o
rendimento de plantas (AFFANDI, 2001) (Figura 2).

122
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2. Mudas de coqueiro. Controle (A e D), muda com 33% da fertilização química padrão
e inoculada com Trichoderma sp. (B e E), muda com 33% da fertilização química padrão e
inoculada com Trichoderma asperellum (C e F).
Fonte: CARDOSO, 2018

123
Belém (PA), 11 a 13 de dezembro de 2019
4. CONCLUSÃO
O uso de Trichoderma sp. como bioestimulantes em mudas de coqueiro
proporciona maiores ganhos nos parâmetros de crescimento, e pode ser estudado como
estratégia eficiente para redução na adubação e possível redução de custos e impactos
ambientais.
5. AGRADECIMENTOS
Agradecemos a Universidade Federal Rural da Amazônia (UFRA) e o Laboratório de
Proteção de Plantas (LPP) pela estrutura fornecida, e pela parceria e ajuda financeira da
FAPESPA e da empresa Sococo S/A.

124
Belém (PA), 11 a 13 de dezembro de 2019
6. REFERÊNCIAS
AFFANDI, Moch. Diversitas dan visualisasi karakter jamur yang berasosiasi dengan
proses degradasi serasah di lingkungan mangrove. Jurnal Penelitian Medika Ekstra. Vol.
2 No. 1:
39-52. 2001.
ALTOMARE, C., NORVELL, W. A., AND HARMAN, G. E. Solubilization of
phosphates and micronutrients by the plant-growth-promoting and biocontrol
fungus Trichoderma harzianum Rifai. Appl. Environ. Microbiol. 65, 2926–2933. Baker,
R., 1999.
ARAGÃO, W. M. Coco: Pós-Colheita. Série Frutas do Brasil. Brasília: Embrapa, 2002.
BARRETO, C. V. G.; TESTEZLAF, R.; SALVA DOR, C. A. Ascensão capilar de água
em substratos de coco e de pinus. Bragantia, Campinas, v. 71, n. 3, p.385-393, 2011.
BOSA, N.; CALVETE, E.O.; KLEIN, V.A.; SUZIN, M. Crescimento de mudas de
gipsofila em diferentes substratos. Horticultura Brasileira. v. 21, n. 3, p. 514-519.
Brasília. jul-set, 2003.
BROTMAN, R. M., Klebanoff, M. A., Nansel, T. R., Yu, K. F., Andrews, W. W., Zhang,
J., & Schwebke, J. R. Bacterial vaginosis assessed by gram stain and diminished
colonization resistance to incident gonococcal, chlamydial, and trichomonal genital
infection. Journal of Infectious Diseases, 202(12), 1907-1915. 2010.
CARVALHO, N. M.; NAKAGAWA, J. Sementes: ciência, tecnologia e produção.
FUNEP: Jaboticabal, 5.ed., p. 590, 2012.
CASSÁN F, D.; PICCOLI, P.; BOTTINI, R Promoción del crecimiento vegetal por
Azospirillum sp. através de la producción de giberelinas. Um modelo alternativo para
incrementar la producción agrícola. In: Albanesi A, Kunst C, Anriquez A, Luna S,
Ledesma R (eds) Microbiología Agrícola. Un aporte de la investigación en Argentina para
la sociedad. Universidad Nacional de Santiago del Estero, Santiago, pp 1–16. 2003.
COSTA, C. A.; RAMOS, S. J.; SAMPAIO, R. A.; GUILHERME, D. O.; FERNANDES,
L. A. Fibra de coco e resíduo de algodão para substrato de mudas de tomateiro.
Horticultura Brasileira, Brasília, v.25,p. 387-391, 2007.
FILHO, M. R. C.; MELLO, S. C. M. de; SANTOS, R. P. dos; MENÊZES, J. E. Avaliação
de isolados de Trichoderma na promoção de crescimento, produção de ácido
indolacético in vitro e colonização endofítica de mudas de eucalipto. Boletim de
pesquisa e desenvolvimento 226. Brasília, DF: Embrapa Recursos Genéticos e
Biotecnologia, 2008.
FRANÇA, Suenny Kelly Santos et al. Biocontrol of sheath blight by Trichoderma
asperellum in tropical lowland rice. Agronomy for sustainable development, v. 35, n. 1,
p. 317-324, 2015.

125
Belém (PA), 11 a 13 de dezembro de 2019
GALVÃO, R. de O.; ARAÚJO NETO, S. E. de; SANTO S, F. C. B. dos; SILVA, S. S.
da. Desempenho de mudas de mamoeiro cv. Sunrise solo sob diferentes substratos
orgânicos. Revista Caatinga, Mossoró, v.20, n.3, p.144-151, 2007.
GAVA, C. A. T.; MENEZES, M. E. L. Eficiência de isolados de Trichoderma spp. no
controle de patógenos de solo em meloeiro amarelo. Revista Ciência Agronômica, v.
43, n. 4, p. 633-640, out-dez, 2012.
GONZÁLEZ-PÉREZ, E.; ORTEGA-AMARO, M. A.; SALAZAR-BADILLO, F. B.,
BAUTISTA; E., DOUTERLUNGNE, D.; JIMÉNEZ-BREMONT, J. F. The
Arabidopsis-Trichoderma interaction reveals that the fungal growth medium is an
important factor in plant growth induction. Scientific reports, v. 8, n. 1, p. 16427,
2018.
HERNÁNDEZ, A., L.; GASCÓ, A. M.; GASCÓ, J. M.; GUERRERO, F. Reuse of waste
materials as growing media for ornamental plants. Bioresource Technology, v. 96, n.
1, p. 125-131, 2005.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA: Levantamento
Sistemático de Produção Agropecuária. Disponível em:
http://www.agricultura.mg.gov.br/images/Arq_Relatorios/Agricultura/2017/Fev/perfil_c
oc o_fev_2017.pdf. Acesso em: 20 maio 2019.
JALAL, M. A. F., LOVE, S. K; VANDER-HELM, D. Siderophore mediated iron III
uptake in Gliocladium virens (Trichoderma virens). 2. Role of ferric mono- and
dihydroxamates as iron transport agent. Journal of Inorganic Biochemistry, 29: 259-
267, 1987.
KÄMPF, A.N. Seleção de materiais para uso como substrato. In: Substrato para
plantas: A base da produção vegetal em recipientes. Porto Alegre: Genises, 2000a.
p.139-145.
KLEIN, V. A.; SIOTA, T. A.; ANESI, A. L.; BARBOZA, R. Propriedades físico-
hídricas de substratos hortícolas comerciais. Revista Brasileira de Agrociência,
Pelotas, v. 6, n. 3, p. 218- 221. set-dez, 2000.
LUCON, C.M.M. Promoção de crescimento de plantas com o uso de Trichoderma
spp. 2009. Artigo em Hypertexto. Disponível em:
http://www.infobibos.com/Artigos/2009_1/trichoderma/index.htm. Acesso em: 6 jan.
2019.
NOMURA, E. S.; LIMA, J. D.; GARCIA, V.A.; RODRIGUES, D. S. Crescimento de
mudas micropropagadas da bananeira cv. Nanicão em diferentes substratos e fontes
de fertilizante. Acta Scientiarum. Agronomy , Maringá, v. 30, n. 3, p.359-363, 2008.
OLIVEIRA, A.G; JUNIOR, A.F.C; SANTOS, G.R; MILLER, L.O; CHAGAS, L.F.B.
Potencial de solubilização de fosfato e produção de AIA por Trichoderma spp. Rev.
Verde. 2012; 7:149-155.
PEREIRA, F. B. Fungos promotores de crescimento e produção de mudas de Pinus
taeda L. 47f. 2017. Dissertação (Mestrado em Ciências Florestais), Universidade
Estadual do Centro Oeste, Irati, 2017.

126
Belém (PA), 11 a 13 de dezembro de 2019
PERSLEY, G.J. Replanting the Tree of Life: Toward an Internation al Agenda for
Coconut Palm Research. CAB/ACIAR, Wallingford, 156p, 1992.
PRATES, H.S.; JUNIOR, J. L.; ROSSI, M. L. Composição mineral de mudas cítricas
com aplicação de Trichoderma spp. Informações Agronômicas. São Paulo, 2007.
SILVA,V. N; GUZZO, S. D; LUCON, C. M. M; HARAKAVA, R. Promoção de
crescimento e indução de resistência à antracnose por Trichoderma spp. em
pepineiro. Pesquisa Brasilleira Agropecuária, vol.46, n.12, p.1609-1618, 2011.
SUGITO, M. Akusento onsei jiten (Accent pronunciation dictionary). Tokyo: Maruzen,
1995. CD ROM
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. Porto Alegre: Artmed, 4 ed, 2009.
TEULAT, B.; ALDAM, C.; TREHIN, R.; LEBRUN, P.; BARKER, J.H.A.; ARNOLD,
G.M.; KARP, A.; BAUDOUIN, L.; ROGNON, F. Analysis of genetic diversity in
coconut (Cocos nucifera L.) populations from across the geographic range using
sequence-tagged microsatellites (SSRs) And RFLPs. Theoretical Applied Genetics
100: 764-771, 2000.

127
Belém (PA), 11 a 13 de dezembro de 2019
A DENSIDADE DA MADEIRA É RELACIONADA AO TEOR DE
EXTRATIVOS? ESTUDO COM QUATRO ESPÉCIES AMAZÔNICAS
Marília Castro Brasil Duarte¹, Ana Kelly de Carvalho², Igor do Vale Gonçalves³,
Manoela Rodrigues do Nascimento4, Lina Bufalino5
1Mestranda em Ciências Florestais. Universidade Federal Rural da Amazônia. e-mail:
[email protected] 2 Mestranda em Ciências Florestais. Universidade Federal Rural da Amazônia. e-mail:
³Doutor em Ciências Florestais. Universidade Federal Rural da Amazônia. e-mail:
[email protected] 4Academica de Engenharia Florestal. Universidade Federal Rural da Amazônia. e-mail:
[email protected] 5Professora Adjunta I. Universidade Federal Rural da Amazônia. e-mail:
RESUMO
Os extrativos, considerados constituintes secundários da madeira, são compostos
químicos que não fazem parte da estrutura da parede celular. São solúveis em água ou em
solventes orgânicos neutros. Além disso, a quantidade e qualidade de alguns desses
componentes influencia a resistência ao ataque de fungos e insetos, a coloração, o odor,
a permeabilidade, as características físicas e a dureza da madeira. Neste sentido, o
objetivo do trabalho foi avaliar se a densidade da madeira está relacionada aos teores de
extrativos encontrados em quatro espécies florestais arbóreas amazônicas:
Pseudopiptadenia suaveolens, Euxylophora paraenses, Aegiphila integrifolia e
Jacaranda copaia. Foi determinada a densidade básica e os teores de extrativos das
espécies em água quente e água fria. As espécies apresentaram diferenças significativas
entre si em densidade básica e teores de extrativos solúveis em água, contudo a influência
do teor de extrativos na densidade básica das quatro espécies deste estudo não foi
detectada. Entretanto, P. suaveolens apresentou maior densidade e maior teor de
extrativos, tanto em água fria, como em água quente.
Palavras-chave: Coloração da madeira. Extrativos água quente. Extrativos água fria.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal

128
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Os extrativos, classificados como componentes químicos secundários da madeira,
não fazem parte da estrutura da parede celular. São solúveis em água e/ou solventes
orgânicos neutros e estão presentes em maior quantidade na casca. Apresentam baixa
massa molecular e ocorrem em pequenas proporções em relação aos componentes
primários. Englobam óleos essenciais, resinas, taninos, graxas e pigmentos (MORAES et
al., 2005). Além disso, a presença de alguns desses componentes influencia a resistência
ao ataque de fungos e insetos, a coloração, o odor, a permeabilidade e a dureza da madeira
(PETTERSON, 1984).
Na madeira, os extrativos se encontram acumulados nas células de parênquima,
nos espaços entre as células, nos canais secretores e em concentrações mínimas na lamela
média (DIETRICH; WEGENER, 2003). Portanto, os extrativos podem ter um efeito
profundo nas propriedades da madeira influenciando as características físicas e químicas
que as diferentes espécies apresentam (SILVA, 2010).
A densidade da madeira, é uma propriedade a qual pode sofrer forte influência da
quantidade de extrativos (GARCIA; MARINONIO, 2016). Estudos com Tectona grandis
L.f. mostraram diferença significativa na densidade em amostras com e sem extrativos
(OLIVEIRA, 2011). Neste contexto, este trabalho objetivou avaliar se a densidade da
madeira é relacionada aos teores de extrativos encontrados em quatro espécies florestais
amazônicas.
2. MATERIAL E MÉTODOS
2.1 SELEÇÃO DO MATERIAL
A madeira utilizada no estudo é proveniente da supressão vegetal realizada em área
de mineração no município de Paragominas, nordeste do Pará com coordenadas
geográficas 3°15'38"S e 47°43’28”O. A coleta do material foi realizada em 2018, portanto
as toras permaneceram armazenadas durante dois anos em pátios de estocagem a céu
aberto.
Para este estudo, foram selecionadas quatro espécies arbóreas: Pseudopiptadenia
suaveolens (Miq.) J.WGRIMES (timborana), Euxylophora paraenses Huber (pau-
amarelo), Aegiphila integrifolia (Jacq.) Moldenke (tamanqueira) e Jacaranda copaia

129
Belém (PA), 11 a 13 de dezembro de 2019
(Aubl.) D.Don (parapará). As espécies foram escolhidas pelas diferenças em suas
densidades observadas na literatura. J. copaia é considerada de baixa densidade, E.
paraenses e A. integrifolia possuem densidade média, e P. suaveolens tem alta densidade
(IBAMA, 1997; IPT, 1983; OLIVEIRA, 2014). A identificação das espécies foi realizada
pela xiloteca da Embrapa Amazônia Oriental, conforme NIDX 04/2019.
2.3 DETERMINAÇÃO DA DENSIDADE BÁSICA
Os discos de madeira foram retirados da extremidade de cada tora, excluindo-se
os primeiros dez centímetros de espessura, de cada disco foram retiradas cinco amostras
em formato prismático do cerne que foram imersas em água destilada até atingir o ponto
de saturação necessário à medição. Em seguida, cada uma das peças foi mergulhada em
outro recipiente com água para determinação do volume saturado em balança digital
(precisão de 0,01 g) - Princípio de Arquimedes, conforme demonstrado por Trugilho et
al., 1990. A massa seca foi determinada em estufa a 105ºC, após estabilização constatada
em três pesagens consecutivas, realizadas a cada 24 horas de acordo com as diretrizes
especificadas pela norma NBR 11941 (ABNT, 2003a).
2.4 DETERMINAÇÃO DO TEOR DE EXTRATIVOS
Para a determinação do teor de extrativos, foi utilizado o cerne de três discos da
madeira de três diferentes indivíduos. Foram retiradas maravalhas que posteriormente
foram trituradas em moinho analítico, com a serragem obtida da trituração, foi realizada
uma classificação do material retido entre peneiras de 40 mesh e 60 mesh.
O teor de extrativos solúveis em água quente e água fria foram determinados de
acordo com a norma da Associação Brasileira de Normas Técnicas (ABNT NBR 14577,
2003b), com a extração em água fria realizada por 48 horas sem a agitação. As análises
foram realizadas no Laboratório de Tecnologia em Produtos Florestais (LTPF) na
Universidade Federal Rural da Amazônia (UFRA), campus Belém.
2.5 ANÁLISES ESTATÍSTICAS
A diferença da densidade básica e teores de extrativos entre as espécies foi
determinada por análise de variância (ANOVA), seguido por teste post hoc de Tukey.
Para verificar se as diferenças em densidade básica entre as espécies estão relacionadas
aos teores de extrativos, foi realizado duas análises de covariância (ANCOVA), uma para

130
Belém (PA), 11 a 13 de dezembro de 2019
cada método de determinação (água fria e água quente). Por fim, um modelo linear
múltiplo estabeleceu a relação entre a densidade básica da madeira e os teores de
extrativos obtidos nos diferentes métodos de determinação.
3. RESULTADOS E DISCUSSÃO
Houve diferença significativa em densidade básica entre as espécies (ANOVA;
Tabela 1). As espécies P. suaveolens e E. paraenses apresentaram densidade básica
significativamente maior que J. copaia e A. integrifolia; contudo, P. suaveolens e E.
paraenses apresentaram densidade básica similar entre si, assim como J. copaia e A.
integrifolia (Figura 1).
Tabela1.Análise de variância (ANOVA) para determinar as diferenças em densidade básica, teor
de extrativos solúveis em água fria e água quente entre as espécies; e análise de covariância
(ANCOVA) para determinar se a diferença em densidade básica das espécies sofre influência dos
teores de extrativos solúveis em água fria (1) e água quente (2). Análises de
Variância GL SQ MQ F p Análises de
Covariância GL SQ MQ F p ANOVA
Densidade
ANCOVA 1
Espécies 3 0.46 0.15 30.07 0.001 Extrativo em
Água Fria 1 0.17 0.17 54.45 0.01
Resíduos 8 0.04 0.01 Espécies 3 0.30 0.10 32.20 0.01
Água Fria:
Espécies 3 0.01 0.00 1.10 0.445 ANOVA Extrativos em água fria
Resíduo 4 0.01 0.00
Espécies 3 25.64 8.55 44.68 0.001
Resíduos 8 1.53 0.19 ANCOVA 2
Extrativo em
Água Quente 1 0.10 0.10 14.98 0.020
ANOVA Extrativos em água quente Espécies 3 0.36 0.12 17.41 0.01
Espécies 3 23.40 7.80 249.81 0.001 Água Quente:
Espécies 3 0.01 0.00 0.42 0.749
Resíduos 8 0.25 0.03 Resíduo 4 0.03 0.01
Fonte: Autores (2019)
As espécies também mostraram diferenças significativas em teor de extrativos
(ANOVA; Tabela 1). Os teores de extrativos solúveis em água fria foi significativamente
maior na P. suaveolens que nas demais espécies. J. copaia, E. paraenses e A. integrifolia
apresentaram médias similares entre si (Figura 2A). Para os extrativos solúveis em água

131
Belém (PA), 11 a 13 de dezembro de 2019
quente, todas as espécies apresentaram diferença significativa entre si. P. suaveolens,
assim como no teor em água fria, apresentou a maior média, seguida pelas espécies A.
integrifolia, J. copaia e E. paraenses (Figura 2B).
Figura 1. Densidade básica de quatro espécies florestais amazônicas. Cores
diferentes indicam diferenças estatísticas.
Fonte: Autores (2019).
P. suaveolens, espécie com maior densidade, apresentou maior teor de extrativos,
tanto em água fria, como em água quente. O maior teor de extrativos nesta espécie pode
ser percebido pela coloração da madeira, bem mais escura em relação às demais, em tom
castanho-avermelhado (IPT, 1983); pois em madeiras tropicais a cor é relacionada com a
quantidade de extrativos localizados no lúmen e na parede celular do cerne (FPL, 2010).
E. paraenses e J. copaia, que apresentaram baixa concentração de extrativos
solúveis em água, possuem madeira de coloração mais clara. A. integrifólia possui
coloração intermediária, em tom creme-escuro. Esta espécie apresentou baixo teor de
extrativos em água fria, mas teve a segunda maior porcentagem de teor de extrativo em
água quente. A diferença na resposta das espécies entre os dois métodos é compreensível,
visto que em água fria é extraído substâncias como gomas, taninos, açúcares e corantes,
enquanto na água quente, além das substâncias anteriores, os amidos também são
extraídos (WASTOWSKI, 2018).

132
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2. Porcentagem de extrativos solúveis em água fria (A) e
água quente (B) presentes na madeira de quatro espécies florestais
amazônicas. Cores diferentes indicam diferença estatística.
Fonte: Autores (2019).
A densidade básica foi relacionada aos teores de extrativos (em água fria e água
quente) e às diferentes espécies amazônicas estudadas. Contudo não houve influência da
interação entre teor de extrativos e as espécies (ANCOVA 1 e 2; Tabela 1). Este resultado
indica que a variação da densidade básica entre as espécies não é causada pela diferença
nos teores de extrativos. A densidade básica dessas espécies parece sofrer maior
influência de outras características da madeira, tais como espessura de parede celular das

133
Belém (PA), 11 a 13 de dezembro de 2019
fibras ou mudança na proporção das fibras em relação à proporção dos vasos
(OLIVEIRA; SILVA, 2003 & PANSHIN; DE ZEEUW, 1980).
Apesar dos teores de extrativos não terem influenciado na diferença em densidade
básica entre as espécies, os valores densidade mostraram uma alta correlação com os
teores de extrativos solúveis tanto em água fria, como em água quente, incluindo a
interação entre eles (R² ajustado = 0,81; p = 0,001), o que gerou o seguinte modelo de
regressão linear:
Db = 8,81002 – 1,44868x1 – 1,2089x2 + 0.20186x1x2
Onde, Db = densidade básica; x1 = teor de extrativo solúvel em água fria; x2 =
teor de extrativo solúvel em água quente.
4. CONCLUSÕES
A densidade básica não foi influenciada pelo teor de extrativos das quatro espécies
estudadas, porém foi possível estimar a densidade básica pelo teor dos extrativos em água
quente e água fria a partir de um modelo linear múltiplo.
P. suaveolens apresentou alta densidade básica e maior quantidade de teores de
extrativos em água quente e água fria, o que pode indicar que espécies com madeiras mais
escuras possuem maiores teores de extrativos.

134
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ABNT - ASSOCIAÇÃO BRASILEIRA DE NORMAS TÉCNICAS. NBR 11941:
Madeira - determinação da densidade básica. Rio de Janeiro: ABNT, 2003a.
ABNT - ASSOCIAÇÃO BRASILEIRA DE NORMAS TÉCNICAS. NBR 14577: pasta
celulósica e madeira – determinação do material solúvel em água. Rio de Janeiro: ABNT,
2003b.
DIETRICH, F.; WEGENER, G. Wood: chemistry, ultrastructure and reactions. Munich:
VerlagKessel, 2003, 387 p.
FOREST PRODUCTS LABORATORY – FPL. Wood handbook: wood as an
engineering material. Madison: U.S. Department of Agriculture, Forest Service, Forest
Products Laboratory; 2010. 508 p.
GARCIA, RA, MARINONIO, GB. Variação da cor da madeira de teca em função da
densidade e do teor de extrativos. Floresta e Ambiente, v. 23, n. 1, p. 124-134, 2016.
GOMIDE, J. L. et al. Análise de critérios de qualidade da madeira de eucalipto para
produção de celulose kraft. Revista Árvore, V. 34, N. 2, P. 339-344, 2010.
IBAMA. Laboratorio de Produtos Florestais. Páginas: 152p., 1997.
IPT. Madeiras da Reserva Florestal de Curuá-Una, estado do Pará: caracterização
anatômica, propriedades gerais e aplicações. São Paulo: IPT, 1981. 117p.
MORAIS, S. A. L.; NASCIMENTO, E. A.; MELO, D. C. Análise da madeira de Pinus
oocarpa Parte 1 – Estudo dos constituintes macromoleculares e extrativos voláteis.
Revista Árvore, v. 29, n. 3, p. 461-470, 2005.
OLIVEIRA B. R. U. Dendrocronologia e análise da variação radial da densidade do lenho
de árvores de Tectonagrandis L. f., do município de Cáceres, MT [dissertação].
Seropédica: Universidade Federal Rural do Rio de Janeiro; 2011.
OLIVEIRA, G. M. V. Densidade da madeira em Minas Gerais: amostragem,
espacialização e relação com variáveis ambientais [tese]. Lavras: UFLA, 2014.
OLIVEIRA, JTS, SILVA, JC. Variação radial da retratibilidade e densidade básica da
madeira de EucalyptussalignaSm. 2003.
OLIVEIRA, J. T. S., SOUZA, L. C.; DELLA LÚCIA, R. M.; JÚNIOR, W. P. S.
Influência dos extrativos na resistência ao apodrecimento de seis espécies de madeira.
RevistaÁrvore, v. 29, n. 5, p. 819-826, 2005.
PETTERSON, R. C. The chemical composition of wood. In: ROWELL, R. (ed). The
chemistry of solid wood. Washington, American ChemicalSociety, 1984.
SILVA, MECM, Apontamentos de Tecnologia dos Produtos Florestais - Composição
Química da Madeira. UTAD, Vila Real, 2010.

135
Belém (PA), 11 a 13 de dezembro de 2019
TRUGILHO, P.F.; SILVA, D.A.; FRAZÃO, F.J.L.; MATOS, J.L.M. Comparação de
métodos de determinação de densidade básica em madeira. Acta Amazonica,1990, 20
(único): 307-319.
VALE, A. T.; DIAS, I. S.; SANTANA, M. A. E. Relações entre propriedades químicas,
físicas e energéticas da madeira de cinco espécies de cerrado. Ciência Florestal, v. 20,
n. 1, p. 137-145, 2010.
WASTOWSKI, A. D. Química da madeira. Rio de Janeiro: Interciência, 2018.

136
Belém (PA), 11 a 13 de dezembro de 2019
ANÁLISE COMPARATIVA DE FOCOS DE QUEIMADAS ENTRE
MUNICÍPIOS DO ESTADO DO PARÁ
Gabriela da Silva Azevedo1; Sofia Rocha Nascimento Louchard2; Patrícia Mie Suzuki3;
Beatriz Cordeiro Costa4; Breno Pinto Rayol5
1Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected] 2Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
³Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected] 4Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected] 5Engenheiro Florestal e Doutor em Biodiversidade e Biotecnologia. Universidade
Federal Rural da Amazônia. [email protected]
RESUMO
Na região Amazônica, a conversão da cobertura vegetal nativa em diferentes usos do solo
tem alterado o clima local, gerando focos de incêndios. O estudo objetivou realizar uma
análise comparativa quantitativa de focos de queimadas entre os municípios do estado do
Pará no ano de 2018. Os dados foram levantados a partir da plataforma do Instituto
Nacional de Pesquisas Espaciais (INPE) - Programa Queimadas com apoio do Ministério
do Meio Ambiente (MMA) por meio do Banco de Dados de Queimadas “Base 2”, no
período de um ano (01 de janeiro de 2018 a 31 de dezembro de 2018). A maior
concentração de focos de incêndios ocorreu em Altamira, Portel, Pacajá, Novo Progresso
e São Félix do Xingu, uma vez que nestes municípios as atividades agropecuárias,
minerais ou madeireiras são predominantes. Ao contrário do que demonstra os municípios
de Colares, Salinópolis, Benevides, Ananindeua e Marituba, devido suas características
naturais e a predominância de atividades terciárias. Dessa forma, as atividades
econômicas podem influenciar na maior ou menor incidência de focos de incêndios.
Palavras-chave: Amazônia. Desmatamento. Incêndios Florestais.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal

137
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O Brasil com suas extensas áreas florestais e agrícolas apresenta um alto grau de
periculosidade, um baixo nível de informação e comprometimento com a manutenção das
florestas. Na região Amazônica, a conversão da cobertura vegetal nativa em diferentes
usos do solo tem alterado o clima local, por meio da diminuição da taxa de
evapotranspiração (SPRACKLEN et al., 2012; ANDERSON et al., 2017).
A precipitação, a umidade relativa do ar e a temperatura são variáveis
microclimáticas que vem sofrendo alterações extremas. Dessa forma, em períodos de
estiagem, com temperaturas elevadas e umidade baixa, a incidência de focos de calor
tende a aumentar, possibilitando a propagação de incêndios florestais, o que exige atenção
redobrada da comunidade local e das autoridades (ANDERSON et al., 2015; LOPES et
al., 2017).
Historicamente, o processo de ocupação e produção agrícola na Amazônia tem o
fogo como um aliado no preparo da área, tanto para diminuir resíduos quanto para
disponibilizar nutrientes ao solo (MIRANDA, 2002; ANDERSON et al., 2017). Sem as
devidas precauções e controle, o fogo pode avançar para a floresta, transformando-se em
um incêndio florestal, sendo, por isso importante verificar os locais com maiores
quantidades de focos de queima de vegetação para que sejam identificadas as causas e
tomadas as devidas ações (LOPES et al., 2017).
Além disso, as intensas ações antrópicas também podem ser percebidas nas
regiões do “arco do desmatamento”, também conhecido como “arco do fogo”. Nestas
áreas, o avanço das atividades agropecuárias em direção à floresta registra índices
elevados de desmatamento e consequentemente maiores incêndios nas vegetações
nativas. Com isso, o presente estudo objetivou realizar uma análise comparativa
quantitativa de focos de queimadas entre os municípios do estado do Pará no ano de 2018.
2. METODOLOGIA
Foi realizado um levantamento de dados quantitativos de focos de queima de
vegetação por município do estado do Pará que foram computados e disponibilizados pela
plataforma do Instituto Nacional de Pesquisas Espaciais (INPE) - Programa Queimadas
com apoio do Ministério do Meio Ambiente (MMA) através do Banco de Dados de

138
Belém (PA), 11 a 13 de dezembro de 2019
Queimadas “Base 2”. Foram utilizados os dados referentes ao período de um ano (01 de
janeiro de 2018 a 31 de dezembro de 2018). Os dados de foco queima de vegetação do
INPE são obtidos através de imagens de satélite, os quais são: NPP-375, GOES-16,
AQUA_M-T, NOAA-19, METOP-B, TERRA_M-M, NOAA-18, TERRA_M-T,
AQUA_M-M, NOAA-19D, NOAA-15, NOAA-18D, MSG-03, GOES-13 e NOAA-15D.
A análise comparativa foi feita entre os cinco municípios que se destacaram
apresentando maiores índices de focos de queima e os cinco que apresentaram menores
quantidades no estado do Pará, em 2018.
3. RESULTADOS E DISCUSSÃO
Altamira foi o município com maior número de focos de queima de vegetação
(23790 focos), correspondendo a 8,08% do total que existe no estado (Figura 1).
Figura 1 – Os cincos municípios com maiores índices de focos de queima de
vegetação no estado do Pará no período de 2018.
Fonte: Autores, 2019.
No município de Altamira destaca-se a construção exacerbada de estradas e
ramais de forma insustentável durante a década de 60 - com o Plano de Integração
Nacional, o que vem a gerar o padrão de desmatamento conhecido como “espinha de
peixe”, principalmente após a execução do projeto da rodovia Transamazônica
(BECKER, 1998; PINTO et al., 2017). Além disso, essa região apresenta uma boa
expressão no polo madeireiro, tendo seu PIB com participação significativa do setor
agropecuário (STEINBERGER, 2002). Com isso, a grande influência antrópica, assim
como a dimensão do município, favorece a ocorrência de inúmeros focos de queima.

139
Belém (PA), 11 a 13 de dezembro de 2019
Portel é o segundo município com maior número de focos, sendo eles 22783, ou
ainda, 7,74% do total do estado. Isto pode ser devido ao baixo índice de Bem Estar
Humano (BEHum) - na escala do Barômetro da Sustentabilidade (BS) - que assola a
mesorregião do Marajó, sendo Portel, um dos municípios com pior BEHum, conforme a
pesquisa de Quintela, Toledo e Vieira (2018), ao analisar aspectos socioeconômicos e
ambientais. Isso pode se refletir no modo como a população desta ilha lida com o manejo
da terra e o uso do solo, afinal, é mais provável que práticas insustentáveis do fogo sejam
realizadas nessa área, podendo levar ao maior número de focos de incêndio.
O município de Pacajá encontra-se como o terceiro alto índice de focos de
incêndio (20778 focos), equivalentes a 7,06% do total. Serviços, agropecuária e indústria
são os principais setores da economia local, contribuindo, respectivamente, com 53%,
35,1% e 10,9% do Produto Interno Bruto do município no ano de 2010 (IDESP, 2013).
O município apresentou, nos últimos 10 anos, expansão de áreas de pastagem, atividades
agrícolas e madeireiras, contribuindo para o aumento da densidade demográfica
(SANTO-DE-OLIVEIRA; ROCHA, 2011). Tais fatores podem estar colaborando para o
elevado número de incêndios na região, visto que, os maiores números de atividades
antrópicas relacionadas ao desfloramento proporcionam altos índices de queima vegetal.
Novo Progresso se apresentou como o município de quarto maior índice de focos
de incêndio em 2018, com o total de 15060, o que corresponde a 5,12% do total de focos
no estado. Isso ocorre por causa da mudança no uso da terra, em 1970, por meio das
políticas públicas nacionais, quando a rodovia que une Santarém e Cuiabá foi construída
(SCHMITT et al., 2011). Com a estrada, a atividade pecuária se expandiu, assim como
as atividades madeireiras e as que envolvem minérios naturais, sendo as duas últimas as
principais causadoras do elevado número de focos (LOPES et al., 2017). Além disso, em
Novo Progresso, fatores como o desmatamento, estiagem e fatores meteorológicos
também foram observados, uma vez que em períodos mais secos e com menor cobertura
vegetal, o número de focos de queima aumenta.
São Félix do Xingu foi classificado como quinto município com maior número de
focos de incêndio, correspondendo este número a 14439 focos. Isto pode ser explicado
pelo fato de que o município em questão apresenta o maior rebanho bovino do Pará e um
dos três maiores do Brasil. Sendo assim, também é o município que apresenta altas taxas
de desmatamento, apontando a atividade pecuária como uma das responsáveis por este

140
Belém (PA), 11 a 13 de dezembro de 2019
fato. A retirada da cobertura vegetal em conjunto com as próprias características
climáticas da região - temperaturas típicas de latitudes próximas ao Equador - e com solo
apropriado para abertura de pastos levam a maiores focos de incêndio, logo, maiores
possibilidades de queimadas (SILVA et al., 2015).
Percebe-se que esses cinco municípios têm em comum a ocupação desenfreada
das terras, a expansão da agropecuária e o uso do fogo no preparo da terra, com ênfase
nos municípios de Altamira, Pacajá, São Félix do Xingu e Novo Progresso, por estarem
localizados na Mesorregião do Sudoeste Paraense, onde está concentrado o “arco do
desmatamento”. A localização desses municípios no estado do Pará pode ser observada
na Figura 2. Além disso, no segundo semestre do ano levantado, foi registrado o evento
“El Niño”, fenômeno climático o qual reduz a precipitação, aumentando
significativamente a ocorrência de incêndios florestais na região amazônica (ALENCAR,
2004).
Figura 2 – Localização no estado do Pará dos cincos municípios com maiores índices de focos
de queima de vegetação no período de 2018.
Fonte: Autores, 2019.
Ao contrário do que demonstra os cinco municípios com maiores índices de focos
de queima, Colares, Salinópolis, Benevides, Ananindeua e Marituba apresentaram as
menores quantidades de focos, sendo elas, respectivamente, 26, 22, 20, 4 e 1 foco (Figura
3).

141
Belém (PA), 11 a 13 de dezembro de 2019
Figura 3 – Os cincos municípios com menores índices de focos de queima de vegetação no
estado do Pará no período de 2018.
Fonte: Autores, 2019.
O município de Colares está localizado às margens da baía do Marajó e da baía
do Sol, uma área bastante úmida - em média 82% - o que dificulta a ação do fogo.
Ademais, por ser uma ilha, a ocupação antrópica desenfreada, a abertura de estradas e a
mudança do uso do solo são mais difíceis de ocorrer, levando a um menor índice de focos
de incêndio (SILVA et al., 1999).
Salinópolis, também por se tratar de um ambiente costeiro, como Colares,
apresenta a umidade um pouco mais elevada, o que também dificulta a ocorrência de
incêndios, além de ser uma região mais voltada ao turismo do que às atividades
agropecuárias, minerais ou madeireiras. Além disso, na região de Salinas encontra-se
muitos manguezais, os quais apresentam produção de biomassa intensa (LIMA;
GALVANI, 2013; LIMA; TOGNELLA, 2012), logo, grande produção de aerossóis, além
de vapor d’água e sal marinho provenientes também do oceano. Essas partículas
absorvem e espalham grande parte da radiação solar incidente, atenuando os efeitos da
radiação (FERNANDES et al., 2018). Isso pode ser um fato que explica o menor índice
de focos de incêndio na região.
Quanto à Benevides, Ananindeua e Marituba, as quantidades de focos de calor são
bem menores pelo fato de estarem localizados na região metropolitana de Belém, ou seja,
são mais urbanizados e quase não apresentam grandes áreas de floresta. Além disso, por
estarem localizados em área urbana, são exercidas em grande maioria as atividades
terciárias (comércio e serviços) e poucas atividades agrárias. É possível observar a
localização dos cinco municípios com menores focos de queima na Figura 4.

142
Belém (PA), 11 a 13 de dezembro de 2019
Nesse contexto, é possível perceber que as atividades econômicas - serviço,
turismo e comércio - nos cinco municípios com menores quantidades de foco de queima,
diferem das atividades nos cinco que apresentam maiores focos, o que pode explicar, em
parte, o motivo da concentração de focos de incêndios em Altamira, Portel, Pacajá, Novo
Progresso e São Félix do Xingu, uma vez que nestes municípios as atividades
agropecuárias, minerais ou madeireiras.
Figura 4 – Localização no estado do Pará dos cincos municípios com menores índices de focos
de queima de vegetação no período de 2018.
Fonte: Autores, 2019.

143
Belém (PA), 11 a 13 de dezembro de 2019
4. CONSIDERAÇÕES FINAIS
A mudança do uso da terra, abertura de estradas, criação bovina e as mudanças
climáticas das regiões apresentadas, contribuem para maiores quantidades de focos de
calor, logo risco de incêndio.
As atividades econômicas são fatores que influenciam na maior ou menor
incidência de focos de queima. Sendo os municípios com predominância de atividades do
setor agropecuário os cinco com as maiores quantidades de focos de queima. Por outro
lado, as regiões com atividades terciárias (comércio e serviços) demonstraram reduzidos
valores de focos de queima no ano de 2018.
Além disso, o emprego de ferramentas alternativas que as comunidades possam
utilizar em substituição ao uso do fogo, o patrulhamento, a utilização de imagens de
satélites, assim como a própria prevenção da propagação das chamas por meio de
brigadistas treinados podem ser instrumentos que diminuam o índice de focos de calor e
incêndios florestais.

144
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALENCAR, Ane et al. Desmatamento na Amazônia: indo além da" emergência
crônica". Belém/PA: Ipam, 2004.
ANDERSON, L. O.; YAMAMOTO, M.; CUNNINGHAM, C.; FONSECA, M. G.;
FERNANDES, L. K.; PIMENTEL, A.; BROWN, F.; JUNIOR, C. H. L. S.; LOPES, E.
S. S.; MOREIRA, D. S.; SALAZAR, N.; ANDERE, L.; ROSAN, T. M.; REIS, V.;
ARAGÃO, L. E. O. C. Utilização de dados orbitais de focos de calor para caracterização
de riscos de incêndios florestais e priorização de áreas para a tomada de decisão. Revista
Brasileira de Cartografia, v. 69, n. 1, p. 163–177, 2017.
ANDERSON, LIANA O.; ARAGÃO, L.E.O.C.; GLOOR, M.; ARAI, E.; ADAMI, M.;
SAATCHI, S.; MALHI, Y.; SHIMABUKURO, Y.; BARLOW, J.; BERENGUER, E.;
DUARTE, V. Disentangling the contribution of multiple land covers to fi re-mediated
carbon emission in Amazonia during the 2010 drought. Global Biogeochemical Cycles,
v. 28, p.1739–1753, 2015. doi:10.1002/2014GB005008.
BECKER, B. K. Amazônia. São Paulo: Editora Ática, 1998. p. 112
FERNANDES, D.A.J.; VITORINO, M.I.; SOUZA, P.J.O.P.; JARDIM, M.A.G. Efeito da
radiação solar sobre a regeneração natural de manguezal em Cuiarana, Salinópolis, Pará.
RBCIAMB, n.49, p. 108-122, 2018.
IDESP. Síntese econômica, social e ambiental do município de Pacajá. Belém - PA,
2013. 50p.
LIMA, N. G. B.; GALVANI, E. Mangrove Microclimate: A case Study from
Southeastern Brazil. Earth Interactions, v. 17, p. 1-16, 2013.
https://doi.org/10.1175/2012EI000464.1
LIMA, T. M. J.; TOGNELLA, M. M. P. Estrutura e Função dos Manguezais: Revisão
Conceitual. Enciclopédia Biosfera, Goiânia, v. 8, n. 15, p. 1801-1827, 2012.
LOPES, A. C. L.; FREITAS, A. V. M.; COSTA, C. D. O. da; BELTRÃO, N. E. S.;
TAVARES, P. A. Análise da distribuição de focos de calor no município de Novo
Progresso, Pará,” Revista Verde de Agroecologia e Desenvolvimento Sustentável, v.
12, n. 2, pp. 298– 303, 2017.
MIRANDA J. B. Aplicações Do Monitoramento Por Satélite No Controle De
Queimadas. In: IV Encontro Nacional sobre Educação Ambiental na Agricultura, 2002,
Campinas-SP. Anais... Campinas–SP: EMBRAPA/CNPM, 2002. p.26.
PINTO, J. F. S. K. C.; SETZER, A.; MORELLI, F.; GOMES, A. R.; ADAMI, M.;
VENTURIERI, A.; GUIMARÃES, T. de F. P. L. Dinâmica do uso e cobertura do solo
em áreas queimadas de municípios na Amazônia brasileira. Anais do XVIII Simpósio
Brasileiro de Sensoriamento Remoto - SBSR, 2017.

145
Belém (PA), 11 a 13 de dezembro de 2019
QUINTELA, P. D. A.; TOLEDO, P. M. de; VIEIRA, I. C. G. Desenvolvimento
sustentável do Marajó, Pará: uma visão a partir do Barômetro da Sustentabilidade. Novos
Cadernos NAEA, v. 21, n. 1, p. 199-213, 2018.
SANTO DE OLIVEIRA, E. E.; ROCHA, C. G. S. Percepções da Problemática
Ambiental pelos Agricultores Familiares do Sudoeste Paraense, Município de
Pacajá. Sustentabilidade em Debate, v. 2, n. 2, p. 135-148, 2011.
SCHMITT, J.; COSTA, D. P.; Novo Progresso (PA) – Acompanhamento E Análise De
Indicadores Socioambientais Utilizando Geotecnologias. In: Simpósio Brasileiro de
Sensoriamento Remoto (SBSR), 2011, Curitiba. Anais... Curitiba-PR: INPE, 2011.
p.2870.
SILVA, J. M. L. da; GAMA, J. R. N. F.; VALENTE, M. A.; REGO, R. S.; RODRIGUES,
T. E.; SANTOS, P. L. dos; CARDOSO JÚNIOR, E. Q.; SILVA, P. R. O. Avaliação da
aptidão agrícola das terras do município de Colares – Estado do Pará. Doc. 21.
Belém: Embrapa Amazônia Oriental, 1999. p. 26.
SILVA, A. H. L. da; PENA, H. W. A.; SOUZA, I. C. M. de. Análise multivariada e a
identificação dos fatores causadores do desflorestamento no Xingu no estado do Pará,
Amazônia oriental. Revista Delos, v. 8, n. 23, p. 1-14, 2015.
SPRACKLEN, D. V.; ARNOLD, S. R.; TAYLOR, C. M. Observations of increased
tropical rainfall proceeded by air passage over forests. Nature, v. 489, p. 282-285, 2012.
STEINBERGER, M. Trabalho apresentado no XIII Encontro da Associação Brasileira de
Estudos Populacionais, realizado em Ouro Preto, Minas Gerais, Brasil de 4 a 8 de
novembro de 2002.

146
Belém (PA), 11 a 13 de dezembro de 2019
AVALIAÇÃO DAS ESTRUTURAS DIAMÉTRICAS EM FLORESTAS
EXPLORADAS SEM AUTORIZAÇÃO, SUBMETIDAS AO LICENCIAMENTO
AMBIENTAL PARA PRODUÇÃO MADEIREIRA NO ESTADO DO PARÁ
Rafael Lima Araújo Ferreira1; Rivaldo Costa Cardoso Junior 2; Wheriton Fernando
Moreira da Silva 3; Jéssica Cristina Oliveira Maciel 4; Karina Sampaio Martins de
Sousa5
1 Especialista em Educação Ambiental. Secretaria de Meio Ambiente e Sustentabilidade
do Pará. [email protected].
2 Mestre em Botânica Tropical. Secretaria de Meio Ambiente e Sustentabilidade do
Pará. [email protected].
3 Mestre em Ciências de Florestas Tropicais. Secretaria de Meio Ambiente e
Sustentabilidade do Pará. [email protected].
4 Especialista em Georreferenciamento, Geoprocessamento e Sensoriamento Remoto.
Secretaria de Meio Ambiente e Sustentabilidade do Pará. [email protected].
5 Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
RESUMO
A Secretaria de Meio Ambiente e Sustentabilidade do Estado do Pará normatizou o
licenciamento de planos de manejo florestais sustentáveis no Estado em “áreas exploradas
no passado sem autorização”, ou seja, áreas que foram submetidas à exploração empírica
e irregular, sem a conivência do proprietário e que ainda tenham potencial madeireiro,
conforme o período de ocorrência do fato e o NDFI, assim disposto na IN Nº 01/2014 e
IN Nº 05/2015. Este trabalho apresenta a comparação das estruturas florestais de quatro
processos de manejo florestal protocolados na SEMAS, utilizando a distribuição por
classes diamétricas por densidade e área basal, onde duas áreas são em concessão florestal
e as outras duas em áreas que tiveram alguma degradação florestal, que caracterize como
“áreas exploradas no passado sem autorização”. Observou-se que as normas e a
metodologia de análise para estas “áreas exploradas sem autorização” não conseguem
determinar o efeito na estrutura florestal. Desta forma, a partir do resultado da distribuição
da densidade e da área basal nas classes diamétricas, observou-se que a estrutura florestal
pode ser comprometida, com empreendimentos que apresentam áreas com algum tipo de
intervenção que resulte na remoção de indivíduos arbóreos. Desta forma, a SEMAS deve
normatizar os procedimentos de apresentação de projeto dos empreendimentos com áreas
exploradas no passado e de suas análises por parte do corpo técnico da secretaria,
considerando a manutenção da estrutura florestal e seu melhor balanceamento entre as
classes diamétricas.
Palavras-chave: Manejo. Legislação . Florestal.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal

147
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O Governo do Estado do Pará, por meio da Secretaria de Meio Ambiente e
Sustentabilidade do Estado do Pará (SEMAS-PA) no uso de suas atribuições, elaborou
instruções normatizadoras visando os planos de manejo florestais sustentáveis no Estado.
As mais recentes, neste sentido, foram a Instrução Normativa Nº 01 de 2014 e a Instrução
Normativa Nº 05 de 2015. A Instrução Normativa Nº 01 de 2014 estabelece a
obrigatoriedade da Autorização Prévia à Análise Técnica de Plano de Manejo Florestal
Sustentável (APAT), como requisito prévio à prática do manejo florestal sustentável; já
a Instrução Normativa Nº 05 de 2015 trata dos procedimentos técnicos para elaboração,
apresentação, execução e avaliação técnica de Plano de Manejo Florestal Sustentável
(PMFS). (PARÁ, 2014a; PARÁ, 2015)
Algo a se destacar nas referidas Normativas é a possibilidade de
Licenciamento/Autorização de “áreas exploradas no passado sem autorização”. Esta
caracterização de “áreas exploradas no passado sem autorização” refere-se a áreas que
foram submetidas à exploração empírica e irregular, sem a conivência do proprietário e
que ainda contenha “capacidade produtiva”, conforme o período de ocorrência do fato e
o Índice Normalizado de Diferença de Fração (NDFI), uma metodologia criada pelo
Instituto do Homem e Meio Ambiente da Amazônia (IMAZOM) para verificar os níveis
de degradação florestal, regulamentada pela Portaria Nº 63/2014 da SEMAS, para a
utilização da Diretoria de Geotecnologias da SEMAS (DIGEO), baseando-se na análise
temporal de imagens e calculando o nível das ações exploratórias. Considerando todas
estas normatizações e demais ferramentas norteadoras das análises da SEMAS, pouco se
observa de apresentação de estudos de distribuição diamétrica, conforme prevê o artigo
22 da Instrução Normativa Nº 05 de 2015, estes estudos são importantes para o
conhecimentos da estrutura das florestas a serem manejadas.
A distribuição diamétrica, também é denominada de estrutura diamétrica ou
distribuição dos diâmetros, conceitua-se da seguinte forma “[...] é a distribuição do
número de árvores, por hectare e por classe de Diâmetro à altura do peito, por espécie ou
por grupo de espécie“. (SOUZA; SOARES, 2013)
A análise da estrutura diamétrica da florestal é imprescindível para uso dos seus
recursos de forma sustentável, possibilitando o melhor planejamento para o manejo, ela
é uma das análises mais empregadas para retratar o comportamento estrutural de um

148
Belém (PA), 11 a 13 de dezembro de 2019
maciço florestal. Desta Forma, entendendo o comportamento da floresta frente aos
distúrbios naturais e antropogênicos, será possível lançar de medidas mitigadoras
(SANTOS et al., 2016). Ou seja, para a conservação dos serviços ecossistêmicos florestas
é importante antes de qualquer plano de ação, conhecer a composição e estrutura dos
remanescentes, o que é de suma relevância para que se façam intervenções e elaboração
de planos de manejo sustentável (COSTA JUNIOR, et al., 2008).
Este trabalho originou-se da vontade dos autores, que são servidores¹ da SEMAS,
de contribuir e externar suas observações, e assim, com a pretensão de apresentar as
análises de quatro POAs e comparar a estrutura florestal destes empreendimentos, sendo
duas áreas de concessões florestais do Estado do Pará, e as outras duas áreas que tiveram
alguma degradação florestal, que caracterize como “áreas exploradas no passado sem
autorização”, para verificar que as normas e a metodologia de análise para estas “áreas
exploradas sem autorização” não conseguem determinar o efeito na estrutura florestal.
2. METODOLOGIA
Foram selecionadas Autorização de Exploração de Produtos Florestais (AUTEF),
bem como processos com informações de Inventário Florestal a 100% (IF100%), Laudos
Técnicos de Geotecnologia e todas as demais informações do empreendimento e das
análises dos setores da SEMAS. Foram selecionados dois processos em áreas de
concessão florestal, nos Municípios de Monte Alegre e Santarém/Aveiro, que serão
denominados de propriedades D e C, e outros dois processos em áreas particulares e com
caracterização de “áreas exploradas sem autorização no passado”, nos Municípios de
Ulianópolis e Dom Eliseu e que serão denominados de propriedades B e A, conforme a
Figura 01, onde é possível observar as suas localizações no Estado do Pará.

149
Belém (PA), 11 a 13 de dezembro de 2019
Figura 1 – Localização aproximada das áreas dos Processos selecionados no Estado do Pará.
Fonte:Rafael Ferreira, 2019.
Por meio de planilha eletrônica, analisou-se os Inventários Florestais dos quatro
processos, realizando a classificação diamétrica, utilizando como intervalo de classes 10
centímetros de diâmetro, agrupando num total 13 Classes, onde todos os indivíduos a
partir 50 cm, que segundo Pará (2015) é o Diâmetro Mínimo de Corte, à 180 cm de
Diâmetro à Altura do Peito (DAP), com Qualidade de Fuste (QF) de 1 à 3, e com qualquer
destinação (Corte, Remanescente, Porta sementes, Protegidas, Etc.) foram alocados.
Calculou-se os valores de área de secção transversal de cada árvore (g), ainda no IF, onde
o somatório foi agrupado nas classes diamétricas as quais os indivíduos pertenciam.
Para reduzir o efeito da diferença do tamanho das áreas, foram calculadas as
Densidades e as Áreas Basais por classes diamétricas, com as informações dos Laudos
Técnicos emitidos pela DIGEO, especialmente as Áreas de Efetivo Manejo Florestal
(AEMF), que são as áreas indicadas para a realização do Manejo Florestal, descontadas
as áreas como Área de Preservação Permanente (APP), Infraestruturas, Áreas cipoálicas
e outras.
3. RESULTADOS E DISCUSSÃO
O resultado da primeira análise dos inventários, como demonstra a Tabela 1,
apresenta as diferentes características, como o número de árvores inventariadas e as áreas
dos empreendimentos (AEMF). Algumas diferenças são bem marcantes, como exemplo

150
Belém (PA), 11 a 13 de dezembro de 2019
a AEMF do empreendimento “C”, cuja a área é quase 2,5 vezes maior que a somando das
áreas dos outros 3 empreendimentos.
Verificando as médias dos cálculos da Densidade (Nº/ha) e da Área Basal (m²/ha),
observou-se que os empreendimentos A e C apresentaram valores próximos para a
Densidade, quanto B e D apresentaram os extremos, para mais e para menos
respectivamente. Quanto ao resultado da Área Basal, os empreendimentos B e C
apresentaram valores extremamente próximos, assim como A e D, o que demonstra que
visualmente não haveria diferença entre “áreas exploradas no passado sem autorização”
e áreas sem exploração.
Tabela 1 – Resumo das informações dos IF’s e os valores médios de Densidade(N/ha), Secção
transversal (m²) e Área Basal (m²/ha), onde A e B são áreas exploradas e C e D não foram
exploradas.
EMPREENDIMENTOS N AEMF (ha) D(N/ha) g(m²) AB(m²/ha)
A 4182 402,3772 10,3932 1410,6558 3,5058
B 7233 504,4504 14,3384 2454,1710 4,8650
C 47253 3944,7966 11,9786 19434,3393 4,9266
D 5522 686,1503 8,0478 2299,8787 3,3519 Fonte: Ferreira et al. (2019).
Mas ao aplicarmos o teste estatístico de correlação linear nas classes diamétricas,
observou-se uma grande correlação entre os empreendimentos C e D que são as áreas
florestais sem exploração, assim como os empreendimentos A e B, que são as áreas
exploradas, conforme mostra a Tabela 2, produzida com o auxilio do programa
BioEstat5.3 (Ayres et al. 2007).
Tabela 2 – Resultado da análise de Correlação para os valores de Área Basal (m²/ha) por classes
diamétricas. EMPREENDIM. A e B A e C A e D B e C B e D C e D n (pares) = 13 13 13 13 13 13 r (Pearson) = 0.9317 0.8919 0.8765 0.8086 0.8375 0.9809 IC 95% = 0.78 a 0.98 0.67 a 0.97 0.63 a 0.96 0.46 a 0.94 0.53 a 0.95 0.94 a 0.99 IC 99% = 0.69 a 0.99 0.55 a 0.98 0.50 a 0.97 0.30 a 0.96 0.38 a 0.97 0.91 a 1.00 R2 = 0.868 0.7954 0.7683 0.6538 0.7014 0.9622 t = 8.5066 6.5402 6.0391 4.5579 5.0832 16.7417 GL = 11 11 11 11 11 11 (p) = < 0.0001 < 0.0001 < 0.0001 0.0008 0.0004 < 0.0001 Poder 0.05 = 0.9999 0.998 0.9961 0.9717 0.9857 1 Poder 0.01 = 0.9985 0.9861 0.9759 0.8897 0.9344 1
Fonte: Ferreira et al. (2019).

151
Belém (PA), 11 a 13 de dezembro de 2019
Ao analisarmos a Área Basal (m²/ha) por classes diamétricas na Figura 2,
observaremos o reflexo nas classes iniciais (até a 3ª) da exploração não autorizada (A e
B), causadora da grande representatividade dos indivíduos de menor diâmetro na
ocupação destas áreas, devido aos indivíduos ingressantes nestas classes inferiores, que
tiveram a oportunidades, em contra partida, a pouca representatividade nas classes
maiores (a partir da 6ª), provavelmente as classes afetadas com a retirada dos indivíduos
arbóreos.
Figura 2 – Área Basal distribuídas em classes diamétricas de quatro empreendimentos, onde A e
B são áreas exploradas e C e D não foram exploradas.
Fonte: Ferreira et al. (2019).
Analisando a Densidade (Nº/ha) por classes diamétricas, verificou-se na Figura 3
que as “áreas exploradas no passado sem autorização” (A e B) tem maior densidade nas
menores classes diamétricas e que a partir da 3ª classe diamétrica observa-se uma grande
redução. Esta Figura nos permite supor que não existe um balanceamento entre as classes
diamétricas e um processo de restabelecimento da comunidade arbórea, assim como um
baixo quantitativo nas classes acima de 80 cm, que poderia indicar que alguns indivíduos
que deveriam estar nestas classes tenham sido explorados quando ainda estavam nas
classes inferiores, e desta forma, houve o comprometimento nas classes maiores.

152
Belém (PA), 11 a 13 de dezembro de 2019
Figura 3 – Densidades distribuídas em classes diamétricas de quatro empreendimentos, onde A
e B são áreas exploradas e C e D não foram exploradas.
Fonte: Ferreira et al. (2019).
A estrutura da floresta do empreendimento A parece ser a mais comprometida,
pois apresenta pouca representatividade nas classes diamétricas maiores que 100 cm. Esta
informação pode ser correlacionada com o Laudo Técnico Geotecnológico da SEMAS,
que apresenta a UPA deste empreendimento com mais de 70% da sua área com algum
tipo de intervenção no passado. Enquanto que a floresta do empreendimento B apresenta
a sua UPA com cerca de 30% da área do empreendimento com algum tipo de intervenção,
segundo o Laudo deste empreendimento.
Para Oliveira et al. (2013), em seus estudos comparativos entre áreas degradadas
e áreas conservadas, também observou o efeito na estrutura florestal nas áreas degradadas,
que apresentaram uma distribuição descontínua dos diâmetros ao longo das classes,
resultando num desbalanceamento diamétrico dos indivíduos em determinadas classes.
Gonçalves & Santos (2008) apresentaram em seus estudos que houve a manutenção da
estrutura diamétrica, por conta da baixa intensidade de exploração em área corretamente
manejada, assim como Reis et al. (2014), que também observou o aumento na densidade
após colheita, mostrando que ocorre um rápido recrutamento em todas as classes
diamétricas em áreas bem manejadas, mas que se o estoque não for observado, pode

153
Belém (PA), 11 a 13 de dezembro de 2019
comprometer o recrutamento, o que pode prejudicar a recomposição de classes maiores.
Carvalho & Nascimento (2009) observaram uma grande diferença nas maiores classes
diamétricas, relacionando-as ao ”corte seletivo pretérito e à mortalidade proporcionada
logo após a fragmentação” da floresta estudada, e consequente abertura do dossel, que
proporcionou o crescimento de juvenis, alterando a estrutura da floresta.
4. CONSIDERAÇÕES FINAIS
O Licenciamento em “áreas exploradas no passado sem autorização” para a
atividade de manejo florestal mostra-se não atento a estrutura florestal, pois as normas
atuais focam apenas na produtividade, o período de ocorrência das explorações sem
autorização e na ausência de indivíduos nas classes inferiores ao diâmetro de corte.
O presente trabalho demonstra que outros parâmetros também devem fazer parte
de futuras normas, assim como as análises técnicas da SEMAS, que deverão atentar para
a manutenção de comunidades florestais, justamente num momento de recuperação de
suas estruturas, como no caso das “áreas exploradas no passado sem autorização”.

154
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
AYRES, M., AYRES JÚNIOR, M., AYRES, D.L. & SANTOS, A.A. 2007. BioEstat 5.3:
Aplicações estatísticas nas áreas das ciências biomédicas. Ong. Mamiraua. Belém, PA.
CARVALHO, F. A.; NASCIMENTO, M. T. Estrutura diamétrica da comunidade e das
principais populações arbóreas de um remanescente de Floresta Atlântica Submontana
(Silva Jardim-RJ, Brasil). Revista Árvore, vol. 33, núm. 2, abril, 2009, pp. 327-337.
Universidade Federal de Viçosa, Viçosa, Brasil.
COSTA JUNIOR, R. F.; FERREIRA, R. L. C.; RODAL, M. J. N.; FELICIANO, A. L. P.
F; MARANGON, L. C.; SILVA, W. C. Estrutura fitossociológica do componente arbóreo
de um fragmento de floresta ombrófila densa na mata sul de Pernambuco, nordeste do
Brasil. Ciência Florestal, Santa Maria, vol. 18, núm. 2, p. 173-183, abr.-jun., 2008
GONÇALVES, F. G.; SANTOS, J. S. Composição florística e estrutura de uma unidade
de manejo florestal sustentável na Floresta Nacional do Tapajós, Pará. Acta Amazonica.
vol. 38(2) 2008: 229 - 244.
OLIVEIRA, D. G.; FERREIRA, R. A.; MELLO, A. A.; ALMEIDA, E. S. Estrutura
diamétrica da vegetação arbustivo-arbórea no entorno de nascentes da bacia hidrográfica
do rio Piauitinga, Salgado – SE, Brasil. Revista Biotemas, 26 (2), junho de 2013.
PARÁ. Instrução Normativa Nº 001/2014, de 14 de janeiro de 2014a. Estabelecer a
obrigatoriedade da Autorização Prévia à Análise Técnica de Plano de Manejo Florestal
Sustentável – APAT, como requisito prévio à prática do manejo florestal sustentável de
uso múltiplo, Secretaria de Meio Ambiente e Sustentabilidade do Pará (SEMAS), Belém,
PA, Nov 2019. Disponível em: <https://www.semas.pa.gov.br/2014/01/14/in-0012014-
de-10-de-janeiro-de-2014-publicada-no-doepa-no32563-de-14012014-caderno-5-
paginas-6-7-8/>
PARÁ. Portaria Nº 063/2014-Gab/SEMAS de 15 de janeiro de 2014b. Instituir a
metodologia de análise de geoprocessamento, no âmbito desta Secretaria de Estado de
Meio Ambiente do Pará – SEMA/PA, para áreas exploradas no passado sem autorização
do órgão ambiental competente, que forem objeto de pedido de Autorização Prévia à
Análise Técnica de Plano de Manejo Florestal Sustentável – APAT , com a finalidade de
avaliar o nível de degradação florestal e definir as áreas com cobertura florestal passíveis
de exploração com aplicação de boas práticas de manejo, Secretaria de Meio Ambiente e
Sustentabilidade do Pará(SEMAS), Belém, PA, Nov 2019. Disponível em:
<https://www.semas.pa.gov.br/2014/01/14/in-0012014-de-10-de-janeiro-de-2014-
publicada-no-doepa-no32563-de-14012014-caderno-5-paginas-6-7-8/>
PARÁ. Instrução Normativa Nº 005/2015, de 10 de setembro de 2015. Procedimentos
técnicos para elaboração, apresentação, execução e avaliação técnica de Plano de Manejo
Florestal Sustentável – PMFS nas florestas primárias, exploradas ou não, e suas formas
de sucessão, no Estado do Pará, Secretaria de Meio Ambiente e Sustentabilidade do
Pará(SEMAS), Belém, PA, Nov 2019. Disponível em:
<https://www.semas.pa.gov.br/2015/09/11/in-05-de-10092015-publicada-no-doe-
32969-de-11092015-paginas-de-37-57/>

155
Belém (PA), 11 a 13 de dezembro de 2019
REIS, L. P.; RUSCHEL, A. R.; SILVA, J. N. M.; REIS, P. C. M.;CARVALHO, J. O. P.;
SOARES, M. H. M. Dinâmica da distribuição diamétrica de algumas espécies de
Sapotaceae após exploração florestal na Amazônia Oriental. Rev. Cienc. Agrar., v. 57,
n. 3, p. 234-243, jul./set. 2014.
SANTOS, R. O.; ABREU, J. C.; LIMA, R. B.; APARÍCIO, P. S.; SOTTA, E. D.;LIMA,
R. C. Distribuição diamétrica de uma comunidade arbórea na Floresta Estadual do
Amapá, Brasil. Biota Amazônia . Macapá, v. 6, n. 2, p. 24-31, 2016
SOUZA, A. L.; SOAREAS, C. P. B. Florestas Nativas: estrutura, dinâmica e manejo.
Viçosa, MG: Ed. UFV, 2013.

156
Belém (PA), 11 a 13 de dezembro de 2019
AVALIAÇÃO DE TEOR DE UMIDADE EM CASTANHA-DE-SAPUCAIA
Soany Elen Palheta da Conceição1; Elizane Alves Arraes Araújo2; Noemi Vianna
Martins Leão³; Elizabeth Santos Cordeiro Shmiszu4; Vanilza Marques Miranda5
1Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 2Discente do Mestrado em Ciências Florestais. Universidade Federal Rural da
Amazônia. [email protected].
³Doutora em Ciências Florestais. Universidade Federal Rural da Amazônia.
[email protected] 4Mestre em Ciências Florestais. Universidade Federal Rural da Amazônia.
[email protected] 5 Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
RESUMO
A castanha de sapucaia (Lecythis pisonis Cambess) é uma castanha nativa do Brasil,
pertencente à família Lecythidacea. A espécie pode substituir, em igualdade de condições,
as nozes, amêndoas ou castanhas comuns (Bertholletia excelsa e Anarcadium occidentale
L.). Para alcançar sementes de alta qualidade, é fundamental considerar os muitos
aspectos. Dentre esses aspectos, o grau de umidade é considerado primordial. Diante do
exposto, o presente trabalho teve por objetivo determinar o teor de umidade de sementes
de sapucaia, em quatro tratamentos distintos. O estudo foi realizado no Laboratório de
Sementes Florestais (LASF) da EMBRAPA Amazônia Oriental, com frutos de duas
matrizes de Lecythis pisonis Cambess, coletadas em Julho de 2015. Os frutos foram
colocados por 10 dias, em lona, para a secagem sob condições ambientais de Belém. A
determinação do grau de umidade das sementes realizou-se com o lote das duas matrizes,
15 dias após a colheita, por meio da verificação do peso úmido e do peso seco, obtidos
após secagem em estufa a ±105°C, durante 24 horas. No tratamento das sementes inteiras,
na repetição 4 foi verificado o maior teor de umidade (58,06%) e o menor valor
encontrado foi na repetição 3 da semente macerada (28,41%). A maior média do teor de
umidade encontrada foi no tratamento inteiras (44,11%), seguida da macerada (37,57%),
corte horizontal (35,97%) e corte vertical (35,94%). Esse trabalho permitiu avaliar
quantitativamente o teor de umidade presente nas sementes recém colhidas da Lecythis
pisonis Cambess, bem como ressalta que a metodologia de avaliação de teor de umidade
do lote de sementes não difere significativamente o processo.
Palavras-chave: Sapucaia. Teor de umidade. Sementes.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal.

157
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A castanha de sapucaia (Lecythis pisonis Cambess) é uma castanha nativa do
Brasil, pertencente à família Lecythidacea, originária de regiões de floresta pluvial
atlântica. A árvore de sapucaia é de grande porte, podendo atingir de 20 a 40 metros de
alturaa (SOUZA et al., 2014). A espécie pode substituir, em igualdade de condições, as
nozes, amêndoas ou castanhas comuns (Bertholletia excelsa e Anarcadium occidentale
L.), atualmente os maiores consumidores de castanha de sapucaia em florestas são os
animais, principalmente morcegos e macacos, destacando sua importância para o
equilíbrio da fauna (LORENZI, 1995).
A floração da L. pisonis ocorre nos meses de setembro a outubro, as flores são de
cor roxa e branca (SOUZA et al., 2014). A frutificação acontece de junho a setembro,
possuem frutos de casca muito dura e uma tampa que se solta quando maduros, liberando
as sementes que têm coloração marrom claro, sendo comestíveis e muito saborosas e com
propriedades medicinais, os frutos por sua vez pode ter uso como recipientes
(OLIVEIRA, 2002). Possui frutos lenhosos resistentes e pesados, com produção média
de 20 castanhas por fruto, além de apresentar aproximadamente 180 sementes por
quilograma (OLIVEIRA, 2002). É uma castanha aromática e oleaginosa que pode ser
consumida em natura ou assada (CARVALHO; NAKAGAWA, 2012).
A “sapucaia” é importante no tratamento do prurido (SILVA et al., 2011), possui
ação eficiente no trato do bacilo da hanseníase. As cascas arbóreas são adstringentes, e
ajudam no tratamento de diarreias quando submetidas à infusão, as folhas por sua vez,
são usadas como Toni cardíacas e diuréticas (OLIVEIRA, 2002), outro uso é no banho,
combatendo irritações cutâneas (FRANCO; BARROS, 2006).
Sementes, nozes e castanhas comestíveis de plantas têm recebido atenção especial
de pesquisadores uma vez que são fontes naturais de vitaminas, minerais, proteínas e
ácidos graxos essenciais, podendo assim contribuir para a alimentação humana
(CARVALHO, 2012).
Para alcançar sementes de alta qualidade, é fundamental considerar os muitos
aspectos no decorrer dos processos que essas sementes são sujeitadas, principalmente
antes de aplicá-las nos plantios. Dentre esses aspectos, o grau de umidade é considerado
primordial, visto que o entendimento desse atributo possibilita a utilização de métodos

158
Belém (PA), 11 a 13 de dezembro de 2019
mais pertinentes no momento da colheita, secagem, beneficiamento e armazenamento,
garantindo a proteção da qualidade física, fisiológica e sanitária (NERY et al., 2004).
Do momento da colheita da semente até a sua utilização em plantios, é importantes
que se determine regularmente o grau de umidade, justamente para possibilitar o
reconhecimento de problemas que decorram, durante os diversos estágios de
processamento, ajudando em questões de armazenamento e viabilidade, permitindo assim
o emprego das devidas ações para resolvê-los. (MARCOS FILHO; CICERO; SILVA,
1987; NERY; CARVALHO; OLIVEIRA, 2004).
A diminuição do poder germinativo durante o armazenamento é motivado
principalmente pelo elevado grau de umidade das sementes. Nessa circunstância, a taxa
respiratória aumenta juntamente com a atividade de microrganismos e com graus de
umidade acima de 20% é possível o aquecimento da massa de sementes a uma
temperatura mortal. (DESAI et al., 1997; GARCIA et. al., 2008).
Em testes oficiais de qualidade de lotes de sementes, é imprescindível que o teor
de água de sementes seja determinado, visto que interfere exatamente nos muitos fatores
da qualidade fisiológica da semente. Além disso, afeta sua maturação, longevidade e
armazenamento, em diferentes pré-tratamentos indispensáveis em testes de germinação,
bem como no período ideal de colheita, peso das sementes e a predisposição a danos por
calor, fumigação, pragas e injúrias mecânicas. (GRABE, 1989).
Conforme Nascimento e Moraes (2011), teores de umidade superiores a 25%
durante o armazenamento, abriria espaço para o aumento de fungos, acentuando assim
putrefação de sementes por conta da consumação de reservas do embrião e da liberação
de toxinas, impossibilitando o estoque por períodos extensos.
Segundo Tresena et al. (2010), para preservar a conservação das sementes e
postergar sua deterioração é preciso que sua taxa de água seja reduzida, visto que em
conformidade com Barbedo e Bilia (1998) e Popiningis (1997), uma taxa de água grande
propicia o aumento da temperatura da massa de sementes, resultante do crescimento de
sua ação biológica, sendo capaz de ocasionar alterações degenerativas e estímulo dos
micro-organismos.
Diante do exposto, o presente trabalho teve por objetivo determinar o teor de
umidade de sementes de sapucaia, em quatro tratamentos distintos. Com isso, temos a
seguinte questão: Os teores de umidade das sementes de L. pisonis alteram em diferentes

159
Belém (PA), 11 a 13 de dezembro de 2019
tratamentos? A hipótese é de que se há tratamentos distintos, há diferença da taxa de
umidade entre eles.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
O estudo foi realizado no Laboratório de Sementes Florestais (LASF) da
EMBRAPA Amazônia Oriental. Foram selecionadas sementes provenientes de 87 frutos
de duas matrizes de L. pisonis, coletadas em Julho de 2015 e procedentes da Fazenda São
Marcos no município de Mãe do Rio, no estado do Pará. A coleta dos frutos foi feita
diretamente do chão e a distância entre as matrizes eram maiores que 30m. Os frutos
chegaram ao LASF no dia seguinte à coleta, e foram colocados por 10 dias, em lona, para
a secagem sob condições ambientais de Belém (umidade relativa do ar, em média 84% e
temperatura média de 27°C).
2.2 EXPERIMENTO
A determinação do grau de umidade das sementes foi realizada com o lote das
duas matrizes, 15 dias após a colheita, por meio da verificação do peso úmido e do peso
seco, obtidos após secagem em estufa a ±105°C, durante 24 horas. Para a avaliação, foram
dispostas duas sementes por cápsula de alumínio (massa de aproximadamente 2,5g), com
quatro repetições para cada tratamento, totalizando quatro distintos: sementes inteiras,
com corte vertical ao meio, com corte horizontal ao meio e sementes maceradas. Por fim,
para aferir o grau de umidade adotou-se a equação descrita pela Regra de Análise de
Sementes (BRASIL, 2009): % de Umidade = 100* (peso úmido - peso seco) /peso úmido.
2.3 ANÁLISE DOS DADOS
A avaliação de porcentagem de umidade foi através de ANOVA, através da
variável umidade entre os tipos de corte e as amostras foram compostas por duas
sementes, sendo 4 o total de repetições por tratamento. Para testar a hipótese, foi utilizado
o auxílio do software SISVAR 5.6, considerado 5 % de probabilidade e quando
necessário, realizou-se o teste de Tukey, ao nível de 5 % (p<0,05) para a análise de
diferenças estatísticas. O teste aplicado para a normalidade foi SHAPIRO-WILK

160
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
Verificou-se no teste, ao nível de 5 % (p<0,05) que as variações foram
consideradas pequenas nas repetições entre os tratamentos, porém, no tratamento das
sementes inteiras, na repetição 4 foi verificado o maior teor de umidade (58,06%) e o
menor valor encontrado foi na repetição 3 da semente macerada (28,41%), conforme o
Quadro 1.
Tabela 1 – Teores de umidade (%) de sementes de sapucaia em quatro tratamentos distintos.
Tratamento Repetição 1 Repetição 2 Repetição 3 Repetição 4
Inteiras 43,09 35,01 40,30 58,06
Corte Vertical 46,25 31,27 33,39 32,87
Corte Horizontal 30,22 37,02 38,81 37,81
Macerada 38,35 41,81 28,41 41,72
Fonte: Autores (2019).
No que diz respeito à tecnologia de sementes, é essencial que a taxa de umidade
seja estabelecida, preferencialmente nas várias fases no processo de produção, como no
armazenamento, isto porque a taxa de umidade interfere nas características físicas e
bioquímicas de sementes (CARVALHO, 2005).
Carvalho et al. (2006) verificaram que o grau de umidade nas sementes de L.
pisonis recém-beneficiadas no estado de Minas Gerais foi alto, por volta de 21%.
Conforme Oliveira et al. (2006) a temperatura e a umidade relativa do ar no local de
estudo exercem grande influência na umidade da semente, pois as sementes são
higroscópicas, ou seja, elas possuem o poder de absorver, ceder ou reter água do
ambiente. Isso explica os elevados teores de umidade encontrados nesse trabalho, devido
à região de estudo apresentar alta umidade relativa do ar.
A maior média do teor de umidade encontrada foi no tratamento inteiras com
((44,11%)), seguida da macerada (37,57%), corte horizontal (35,97%) e corte vertical
(35,94%) (Figura 1).
Carvalho e Nakagawa (2000) afirmam que a qualidade de sementes pode ser
afetada pelo desequilíbrio do teor de água, o que dificulta o manejo e a eficiência do

161
Belém (PA), 11 a 13 de dezembro de 2019
beneficiamento, além de ser um fator limitante no armazenamento. (CARVALHO;
NAKAGAWA, 2000).
Figura 1 – Gráfico de médias do teor de umidade de sementes de sapucaia.
Fonte: Autores (2019).
As médias passaram pela Anova e apresentaram um CV de 18,45% e os
tratamentos não apresentaram diferenças significativas quando submetidos ao teste
p<0,05. Ou seja, as diferentes metodologias apresentaram comportamentos semelhantes
no processo de secagem ao qual foram submetidas, afirmando assim, que a hipótese foi
rejeitada, conforme o teste empregado.
Campos e Tillmann (1996) e Ramos et al. (2000) viram que conforme o método
empregado para estabelecimento da umidade, pode ocorrer uma alta ou baixa alteração
dos resultados.
Tabela 2 – Tabela de análise de variância para os dados de teor de umidade da Castanha. FV GL SQ QM FC
Tratamento 3 181,208919 60,4020973 1,203
Erro 12 602,331375 50,194281
CV(%)= 18,45
Média Geral: 38,3993750 Nº de observações: 16
GL- Grau de Liberdade; SQ- Soma dos quadrados; QM-
Quadrado das médias; FC- fator calculado; CV – Coeficiente de
variação.
Fonte: Autores (2019).

162
Belém (PA), 11 a 13 de dezembro de 2019
4. CONCLUSÃO
Este trabalho permitiu avaliar quantitativamente o teor de umidade presente nas
sementes recém-colhidas da Lecythis pisonis Cambess. A média de teor de umidade no
tratamento inteiras, macerada, corte horizontal e corte vertical foram: 44,11%; 37,57%;
35,97% e 35,94% respectivamente. O estudo trouxe a importância de se determinar o teor
de umidade nas sementes, justamente para assegurar a qualidade delas nas variadas etapas
de beneficiamento.
O presente trabalho ressaltou que a metodologia de avaliação de teor de umidade
do lote não diferiu significativamente o processo, uma vez que as médias apresentaram
valores aproximados.

163
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BARBEDO, C. J.; BILIA, D. A. C. Evolution of research on recalcitrant seeds. Scientia
Agrícola, Piracicaba, v. 55, p. 45- 48, 1998. Número especial.
BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de
sementes. Brasília: Mapa/ACS, 2009. 399p.
CAMPOS, V. C.; TILLMANN, M. A. A. Comparação entre os métodos oficiais de estufa
para determinação do grau de umidade de sementes. Revista Brasileira de Sementes,
Brasília, v. 18, n. 1, p.134-137, 1996.
CARVALHO, L.R.; SILVA, E.A.A.; DAVIDE, A.C. Classificação de sementes florestais
quanto ao comportamento no armazenamento. Revista Brasileira de Sementes, v.28,
n.2, p.15-25, 2006.
CARVALHO, N. M.; NAKAGAWA, J. Sementes: ciência, tecnologia e produção.
Jaboticabal: FUNEP, 588 p. 2000.
CARVALHO, N.M.A secagem de sementes. Funep, São Paulo, 184 p. 2005.
CARVALHO, N.M. e NAKAGAWA, J. Sementes – ciência, tecnologia e produção.
FUNEP, Jaboticabal, 5.ª ed, 590 p. 2012.
DESAI, B.B; P.M. KOTECHA; D.K. SALUNKHE. Seeds Handbook, Biology,
Production, Processing and Storage. Marcel Dekker Inc., USA. 1997.
FRANCO, E. A. P.; BARROS, R. F. M. Uso e Diversidade de plantas Medicinais no
Quilombo Olho D'água dos Pires, Esperantina, Piauí. Revista Brasileira de Medicina,
n.8, p.78-88, 2006.
GARCIA, D. E. et al. Nutritional characterization of six fodder species foliage with
emphasis in their polyphenolic profiles. Revista Científica, FCV-LUZ, v. 18, n.2, p. 188-
196, 2008.
GRABE, D. F. Measurement of seed moisture. In:STANWOOD, P. C. and McDONALD,
M. B (Eds).Seed Moisture. Madison: The Crop Science Society of America, p.69-92,
1989.
LORENZI, H.. Arvores Brasileiras: Manual De Identificação E Cultivo De Plantas
Arbóreas Nativas Do Brasil. Plantarum, Nova Odessa, SP ,v.1, n.1, 1995.
MARCOS FILHO, J.; CICERO, S. M.; SILVA, W. R.Avaliação da qualidade das
sementes. Piracicaba: FEALQ, 1987. 230 p.

164
Belém (PA), 11 a 13 de dezembro de 2019
NASCIMENTO, W.M.O; MORAES, M.H.D. Fungos associados a sementes de açaí:
efeito da temperatura e do teor de água das sementes durante o armazenamento. Revista
Brasileira de Sementes, vol. 33, n. 3, p. 415-425, 2011.
NERY, M. C.; CARVALHO, M. L. M.; OLIVEIRA, L. M. Determinação do grau de
umidade de sementes de ipê-docerrado Tabebuia ochracea ((Cham.) Standl.) pelos
métodos de estufa e forno de microondas. Ciência e Agrotecnologia, Lavras, v. 28, p.
1299-1305, 2004.
OLIVEIRA, F.L. Ficha da Biodiversidade. Bio trilhas. Disponível em:
<http://www.toligado.futuro.usp.br/envio/exibir.php?atividade=bio_trilhas&cód=502.
Acesso em: 27 out. 2018.
OLIVEIRA, J. A.; GUIMARÃES, R. M.; ROSA, S. D. V. F. Processamento de sementes
pós-colheita. Informe Agropecuário. Belo Horizonte, v.27, n.232, p.52-58, 2006.
POPININGIS, F. Fisiologia de sementes. Brasília: AGIPLAN, 1997. 315 p.
RAMOS, F. N.; SOUZA, A.F.; LOUREIRO, M.B.; CRUZ, A.P.M.; ANDRADE, A.C.S.
Comparação entre métodos de secagem na determinação do grau de umidade em
sementes de Parkia multijuga Benth. (Leguminosa Mimonoideae). Revista Árvore,
Viçosa, v. 24, n. 2, p. 175-179, 2000.
SILVA, L.L., GOMES, B.S., SOUSA-NETO, B.P., OLIVEIRA, J.P.C., FERREIRA,
E.L.F., CHAVES, M.H., OLIVEIRA, F.A. Effects of Lecythis pisonis Camb.
(Lecythidaceae) in a mouse model of pruritus. Journal of Ethnopharmacology. V. 139,
p 90–97, 2012.
SOUZA, A. S.; MARGALHO, L.; PRANCE, G. T.; GURGEL, E. S. C.; GOMES, J. I.;
CARVALHO, L. T.; SILVA, R. C. V. M. Conhecendo Espécies de Plantas da
Amazônia: Sapucaia (Lecythis pisonis Cambess. – Lecythidaceae). Comunicado
Técnico 250 Embrapa. Belém-PA, 2014.
TRESENA, N. L.; CAVALCANTI MATA, M.E.R.M; DUARTE, M.E.M.; MORAES,
A.M. Determinação do teor de água limite para crioconservação das sementes de ipê
amarelo (Tabebuia chrysotrica (Mart. Ex. DC.) Standl.). Revista Cerne, Lavras, v.16,
n.2, p.171-175, abr./jun, 2010.

165
Belém (PA), 11 a 13 de dezembro de 2019
CARACTERIZAÇÃO DE PROPRIEDADES FÍSICAS E ANATÔMICAS DA
MADEIRA DE Qualea albiflora Warm, MANDIOQUEIRA PARA
CONSTRUÇÃO CIVIL
Cinthia Manuella Pantoja Pereira; Alice Ferreira Cardoso2; Ewerton Alves Nazareno 3;
Aliça Dayane Araújo Abreu4; Marcela Gomes da Silva 5
1 Graduanda do curso de Engenharia Florestal. Universidade Federal Rural da
Amazônia, campus Belém-PA, E-mail: [email protected]. 2 Graduanda do curso de Engenharia Florestal. Universidade Federal Rural da
Amazônia, campus Belém-PA. E-mail: [email protected];
³ Graduando do curso de Engenharia Florestal. Universidade Federal Rural da
Amazônia, campus Belém-PA. E-mail: [email protected]. 4 Graduando do curso de Engenharia Florestal. Universidade Federal Rural da
Amazônia, campus Belém-PA. E-mail: [email protected]. 5 Professora Doutora em Tecnologia da madeira, pertencente ao Instituto de Ciências
Agrárias da Universidade Federal Rural da Amazônia, campus Belém-PA, .E-mail:
RESUMO
A Amazônia possui uma diversidade madeireira relevante no cenário mundial e o Brasil
possui um grande potencial em ser um produtor de móveis de competitividade global,
porém sua posição de 31°lugar no ranking dos exportadores de móveis é desconfortável.
Dentre as espécies madeireiras da flora amazônica, destaca-se a espécie, Qualea albiflora
Warm popularmente conhecida como Mandioqueira e que surge como forte potencial
para exportação e construção civil. Diante do exposto, o objetivo do trabalho é determinar
as características físicas, no que diz respeito à densidade; e anatômicas, referente à
macroscopia da madeira da espécie em questão, visando fornecer subsídios para fortalecer
sua aptidão tecnológica. Foi coletado pranchão da espécie e foram confeccionados corpos
de prova a partir desse pranchão. Foram feitas amostras de dimensões de 2x3x5 cm
medidas estas que foram tomadas com base na NBR 7190 que é referência para esse tipo
de procedimento. As amostras foram submetidas à saturação e posteriormente levadas a
estufa a 103ºC até saturar sua massa. Após a saturação, foram feitas as análises da espécie
em estudo, primeiramente, para as análises anatômicas macroscópicas e posteriormente
feitas as avaliações da densidade básica e da retratibilidade do indivíduo através da
relação entre massa seca pelo volume saturado. Após os resultados do estudo, constatou-
se que a densidade básica para Qualea albiflora Warm obteve um valor médio de 0,78
g/cm³ valor este obtido devido às variações nas propriedades físicas e macroscópicas
dessas espécies.
Palavras-chave: Densidade básica. Propriedades físicas. Macroscopia.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal

166
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A Amazônia possui inestimáveis riquezas de essências florestais como: látex,
óleos, frutos e castanhas, além de outros produtos de origem da madeira. Desta forma o
Brasil possui um grande potencial em ser um produtor de móveis de competitividade
mundial, porém essa abundância florestal não é diretamente proporcional a sua posição
de 31°lugar no ranking dos exportadores de móveis (SOARES; CARVALHO, 2013).
Qualea albiflora Warm, surge com forte potencial para a exportação.
Vulgarmente conhecida como Mandioqueira na região amazônica, é da família
Vochysiaceae, e tem origem no norte do Brasil, com altura comercial de 12,9m
(LUCHTEMBERG, 2013). Segundo CASTRO (2014) a Qualea albiflora Warm é
utilizada na construção civil como ripas, forros, guarnições, cordões, rodapés, móveis
standard e partes internas de móveis, compensados, embalagens e paletes.
A diversidade de nomes vulgares utilizados no mercado dificulta a
comercialização, resultando em prejuízos tanto para revendedores quanto para
compradores (REIS et al., 2014). Desta forma, ressalta-se a importância de identificação
anatômica da madeira com base em análises macroscópicas na qual é possível visualizar
o parênquima axial, vasos, fibras, raios, entre outras características, as quais são de grande
utilidade ao setor madeireiro para garantir ao consumidor um certificado autêntico das
espécies desejadas (REIS et al., 2014). Tão importante quanto à descrição anatômica
macroscópica, outro índice de qualidade da madeira utilizados na indústria florestal é a
densidade básica, por ser uma das características da madeira que melhor expressam sua
qualidade no beneficiamento industrial.
Tendo em vista toda está problemática, a mandioqueira (Qualea albiflora Warm)
foi escolhida para ser estudada dentro das várias espécies florestais comercias que vem
ganhando grande demanda de mercado na região amazônica por apresentar grande
resistência física as intempéries temporais e apresentar preço altamente competitivo no
mercado moveleiro na construção civil (Garcia, 2013).
Diante do exposto, objetivou-se descrever as características físicas, no tocante a
densidade básica; e anatômicas do lenho, referente à macroscopia da madeira da espécie
Qualea albiflora Warm, visando fornecer subsídios para fortalecer sua aptidão
tecnológica para construção civil.

167
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
Esse estudo foi realizado em 2017, a madeira de Qualea albiflora Warm foi
coletada em produtora de madeira serrada para construção civil no município de Belém,
proveniente de plano de manejo, situada na divisa dos estados do Pará e Amapá, através
do projeto FLONA Jari. O indivíduo abatido foi desdobrado em pranchões centrais, nas
dimensões de 1,40 m de comprimento por 0,2 m de espessura visando facilitar o
transporte até o Laboratório de Tecnologia e Produtos Florestais (LTPF) localizado na
Universidade Federal Rural da Amazônia campus Belém-PA.
Para a determinação da estrutura anatômica, foram confeccionados corpos de
prova com tamanho de 2cm x 3cm x 5cm em 10 repetições. Para a melhor visualização,
foi utilizada uma lupa conta fios de 10x de aumento.
Além da análise macroscópica, foi determinada a densidade básica.
Primeiramente, as amostras passaram pelo processo de saturação, após esse
procedimento, determinou-se o volume saturado utilizando-se como base o princípio de
Arquimedes no qual o volume do corpo de prova era determinado pelo deslocamento do
volume de água.
Após a determinação do volume as amostras foram colocadas em estufa à 103º ±
2 para a secagem. De posse da massa seca e do volume saturado foi possível determinar
a densidade básica da espécie em estudo seguindo as normais da ABNT NBR 7190 que
é btida pela seguinte fórmula:
Db=MsVs
Onde, Db= densidade básica (g/cm³); Ms= massa de amostra absolutamente seca, Vs=
volume saturado em (cm³).
A análise dos dados estatísticos foi feita com a utilização do software Excel a
partir dos cálculos da média, pontos de máximo e mínimo, desvio padrão, erro padrão e
coeficiente de variância a fim de facilitar a interpretação desses dados.
3. RESULTADOS E DISCUSSÃO
3.1 ANÁLISE ANATÔMICA MACROSCÓPICA
No que diz respeito à descrição anatômica macroscópica de Qualea albiflora
Warm verificou-se a ocorrência do parênquima axial visível a olho nu, do tipo aliforme e
losangular e também linear; poros pouco visíveis a olho nu, com solitários predominantes,

168
Belém (PA), 11 a 13 de dezembro de 2019
mas com ocorrência de múltiplos de 2 a 4, em disposição diagonal e com presença de
depósito branco. Os raios transversais são bem visíveis a olho nu e não estratificados
(Figura 1). As camadas de crescimento são pouco distintas, e a madeira foi classificada
como medianamente pesada e de textura média, além de cerne de cor bege claro, brilho
moderado, cheiro e gosto imperceptíveis.
Figura 1. Macroscopia da madeira de Qualea albiflora Warm em faces
Transversal (A), Longitudinal Tangencial (B) e Longitudinal Radial (C).
Fonte: Autores (2017).
Resultados semelhantes foram encontrados por Reis et al. (2014) com a madeira
do mesmo gênero, apresentando camada de crescimento indistintas, poros visíveis a olho
nu, difusos, com arranjo tendendo a diagonal, apresentando contorno angular;
parênquima axial visível a olho nu, paratraqueal aliforme de extensão linear; raios em
observação macroscópica foram considerados contrastados em seção radial, com largura
variando entre 1 a 3 células.
De acordo com a Organização Internacional das Madeiras Tropicais (2017) com
base no livro Catálogo das Madeiras da Amazônia (1968), destacou para a espécie de
mesmo gênero, Qualea sp, o parênquima relativamente abundante, visível a olho nu,
aliforme, chegando a formar arranjos oblíquos ligando alguns poros. Poros visíveis a olho
nu, solitários, agrupados em 2-3 poros, vazios, embora alguns sejam obstruídos por tílos.
Linhas vasculares facilmente visíveis. Raios no topo são finos e numerosos, porém
visíveis sob lente; madeira considerada pesada. ZENID (2013) descreveu Qualea
albiflora Warm como uma madeira com cerne na cor bege-claro levemente rosado e de
brilho moderado; cheiro e gosto imperceptíveis, densidade média, moderadamente dura
ao corte, textura média.

169
Belém (PA), 11 a 13 de dezembro de 2019
3.2 AVALIAÇÃO DA DENSIDADE BÁSICA
A espécie Qualea albiflora Warm apresentou média de densidade básica igual a
0,78 g/cm³ (Tabela 1). No estudo feito por Garcia (2013), que visava à obtenção do
rendimento volumétrico de Qualea albiflora Warm em serrarias, a espécie mostrou
densidade básica média de 0,68 g/cm³ a qual foi classificada como madeira
moderadamente dura ao corte. Entretanto, o Catálogo de Madeiras Brasileiras para
construção civil do Instituto de Pesquisa Tecnológicas de São Paulo que padroniza a
densidade para as espécies nacionais, definiu como moderadamente dura as madeiras com
densidade a partir de 0,70 g/cm³ ou 700 kg/m³.
Tabela 1. Análise estatística descritiva e da densidade básica
da Qualea albiflora Warm. Espécie Estatística descritiva DB
Qualea albiflora Warm
Mínimo 0,76
Máximo 0,79
Média Aritmética 0,78
Desvio Padrão 0,01
Erro Padrão 0,002
Coeficiente de Variância 0,01
Fonte: Autores (2017).
A Qualea albiflora Warm obteve densidade básica de 683 kg/m³. Segundo o
Catálogo de Madeiras Brasileiras para Construção Civil do Instituto de Pesquisa
Tecnológica - IPT de São Paulo, esta espécie é considerada de densidade moderadamente
média a dura, com densidade aparente a 15% de 820 kg/m³. Os valores encontrados no
presente estudo estão abaixo aos descritos por esse catálogo, reforçando a aptidão da
madeira para a construção civil, devido à diferença das condições de umidade relativa e
ao ponto de saturação das fibras PSF, que no caso do estudo foi a abaixo deste ponto.
Segundo Rocha et al (2003), a densidade básica da madeira varia em função da
taxa de crescimento, local, espaçamento, idade, procedência, espécie, e varia ainda na
mesma espécie e dentro da mesma árvore. A autora afirma também que a variação
individual evidencia o potencial de seleção das espécies. Outras características
influenciam nas diferenças encontradas na densidade básica da madeira em espécies
diferentes, haja vista que as características da madeira são o resultado da combinação

170
Belém (PA), 11 a 13 de dezembro de 2019
entre fatores genéticos e ambientais (Rocha et al, 2003). O tamanho e espessura de seus
elementos celulares na madeira também influenciam na densidade básica, especialmente
em se tratando de fibras e microfibrilas que são o resultado da rica composição química
intracelular.
.
4. CONCLUSÃO
Analisando os resultados da densidade básica e o comportamento dimensional da
espécie em estudo, pôde-se observar que a espécie Qualea albiflora Warm, conhecida na
região amazônica como mandioqueira, apresentou densidade 0,78 g/cm³, devido a fatores
genéticos, ambientais e anatômicos. A espécie foi classificada como moderadamente dura
para a construção civil e sendo apta para construções temporárias, partes internas de
construção civil tais como molduras, forros, rodapés, lambris e similares.

171
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
CASTRO, A. P. et al. Características das Madeiras para Construção Civil. Instituto
de Pesquisas Tecnológicas (IPT), São Paulo, 2013.
CATÁLOGO DE MADEIRAS BRASILEIRAS PARA A CONSTRUÇÃO CIVIL -
Coordenação Augusto Rabelo Nahuz - SÃO PAULO- Instituto de Pesquisas
Tecnológicas do Estado de São Paulo - ITP, 2013.
ITTO - International Tropical Timber Organization. Espécies menos usadas. 2017.
Disponível em: < http://www.tropicaltimber.info/pt-br/specie/mandioqueira-qualea-
paraensis/#lower-content> Acesso em: 7 set. 2017.
LOUREIRO, A. A; Silva, M. F. Catálogo das Madeiras da Amazônia, Ed. SUDAM,
1968. V. 2
LUCHTEMBERG, P.H.Q. Resistências natural de dez espécies de madeiras Amazônicas
submetidas ao ataque de fungos apodrecedores em ensaio de laboratório, Trabalho de
Conclusão de Curso apresentado ao Departamento de Engenharia Florestal da
Universidade de Brasília como parte das exigências para obtenção do título de Engenheiro
Florestal. 2013.
MAURI, Renata et al. Wood density of clones of Eucalyptus urophylla x Eucalyptus
grandis in different conditions of growth. Floresta, v. 45, n. 1, p. 193-202, 2014.
REIS, A. R. S. et al. Caracterização anatômica da madeira de mandioqueira (Qualea
Aubl.) comercializada no mercado madeireiro do estado do Pará. Enciclopédia
Biosfera, Centro Científico Conhecer - Goiânia, 2014. V.10, n.19; p. 448
ROCHA, Maria das Graças de B. et al. Variação da densidade básica e correlações entre
caracteres progênies jovens de Eucalytus grandis Hill ex- Maiden em duas etapas de
crescimento. Revista árvore. Vol 7. Sociedade de Investigações Florestais nº2, visoça
MG, 2003.
SOARES, M. H. M.; CARVALHO, J. O. P. D. Importância ecológica das espécies
Bagassa guianensis (tatajuba), Copaifera multijuga (copaíba), Dipteryx odorata
(cumaru). Hymenaea courbaril (jatobé), e Tabebuia serra tifolia (ipê-amarelo) em
Floresta Natural na Amazônia Brasileira. Embrapa. Pará, 2013. p. 1-3.

172
Belém (PA), 11 a 13 de dezembro de 2019
CONJUNTURA DO MERCADO DA FIBRA DE BURITI (Mauritia flexuosa L.)
NO ESTADO DO PARÁ ENTRE OS ANOS DE 1997 A 2017
Adson Eluan da Silva1; Francisco Valente Braga2; Patrícia Mie Suzuki3; Welton dos
Santos Barros4; Fabrício Khoury Rebello5
1 Graduando em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 2 Graduando em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 3 Graduando em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 4 Graduando em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 5 Professor orientador. Universidade Federal Rural da Amazônia.
RESUMO
A biodiversidade amazônica vem diminuindo devido o avanço das fronteiras agrícolas,
fato que leva à exaustão dos seus recursos. Com isso, a fibra do buriti, como produto
florestal não madeireiro (PFNM), apresenta-se como uma alternativa sustentável e
economicamente viável para os pequenos produtores. O objetivo do trabalho foi analisar
os efeitos da cadeia produtiva da fibra do buriti no mercado (preço e produção) durante
os anos de 1997 a 2017 no estado do Pará. Os dados coletados para o desenvolvimento
deste trabalho foram retirados do Sistema IBGE de Recuperação Automática (SIDRA). Foi realizada a análise da taxa média geométrica de crescimento anual em percentagem
(TCG%). Para corrigir efeito da inflação do decorrer dos anos, foi realizado a correção
inflacionária por meio do Índice Geral de Preços Disponibilidade Interna (IGP-DI) da
Fundação Getúlio Vargas (FGV). No estado do Pará, os municípios com maior produção
são: Igarapé-Miri, Abaetetuba, Cametá e Barcarena. O preço da fibra cresceu 397,06%
entre os períodos de 1997 a 2017, no entanto, a produção, no mesmo período, reduziu
cerca de 18,67%. Dentre os principais atores da cadeia produtiva da fibra de buriti,
destacam-se os extrativistas e extrativistas-artesãos, atravessadores-proprietários de
barcos, artesãos - de brinquedos e de paneiros, comerciantes e, por fim, os consumidores
regionais, nacionais e internacionais. O apoio e incentivo do governo com implantação
de políticas públicas devem ser desenvolvidas, para que haja a continuidade da produção
por meio de manejos sustentáveis das palmeiras e uma ampliação do mercado para os
produtos derivados da fibra de buriti. Visto que é uma cadeia produtiva que sustenta várias
famílias e apresenta uma grande importância na tradição e na cultura das comunidades
paraenses.
Palavras-chave: Extrativismo. Produto Florestal não Madeireiro. Cadeia Produtiva.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal

173
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Com o decorrer dos anos, o avanço das fronteiras agrícolas põe em risco cada vez
mais os ecossistemas florestais, a biodiversidade e a conservação dos mesmos. Neste
contexto, as populações que vivem tradicionalmente desses recursos tem um papel
fundamental na manutenção da floresta, pois muitos apresentam rendas originárias do
extrativismo de produtos florestais não madeireiros - PFNM (FIEDLER et al., 2008).
Partindo-se da premissa do desenvolvimento sustentável, por meio da exploração
dos PFNM, encaixa-se nesse contexto a utilização da Mauritia flexuosa L., palmeira
nativa de áreas alagadas, conhecida popularmente como, buriti, miriti, carandá-guaçu. O
buriti tem uma ampla distribuição geográfica, sendo encontrado no Cerrado, Amazônia e
Caatinga (FLORA DO BRASIL, 2019). As pequenas comunidades localizadas nessas
regiões usufruem, tradicionalmente, dos diversos produtos originários da palmeira nativa,
apresentando elevada importância social, cultural e econômica.
O buritizeiro pode chegar até 35 m de altura e produz entre 400 a 500 frutos por
cacho, contudo apenas a palmeira fêmea é capaz de produzir frutos, necessitando da
polinização da palmeira macho (SOUZA; VIANA, 2018). Do buritizeiro quase tudo é
aproveitado pelas comunidades locais, desde os frutos para o consumo até a
comercialização de sucos e polpas, contribuindo para a segurança alimentar da família,
servindo as folhas para extração das fibras para produção de artesanatos e brinquedos e o
caule para fazer vinhos (ALMEIDA; SILVA, 1994).
A fibra do buriti é um dos principais produtos da palmeira, sendo o estado do Pará
responsável pela maior produção do Brasil (IBGE, 2017). A matéria-prima da fibra é o
pecíolo, onde a parte externa do pecíolo é utilizada para confeccionar paneiros e a parte
interna (medula) apresenta uma textura esponjosa, utilizada na fabricação de brinquedos.
Da folha mais jovem, vulgarmente conhecida como “linho” ou “seda” do buriti, são
confeccionados bolsas, cordas, toalhas, chapéus, entre outros (SARAIVA, 2009;
CATTANI; BARUQUE- RAMOS, 2014).
O artesanato é uma forma de manifestação cultural e obtenção de renda das
pequenas comunidades. Vale ressaltar que nos últimos anos os artefatos de miriti,
sobretudo os brinquedos, passaram a ser conhecidos e comercializados em outros países
do mundo, como Japão, Estados Unidos, França, Alemanha e Portugal, por meio de feiras

174
Belém (PA), 11 a 13 de dezembro de 2019
públicas e exposições culturais, estimulando a exportação desses produtos (MORAES,
2013).
A fibra do buriti tem uma importância social e cultural relevante para os pequenos
produtores e para o estado do Pará, porém poucas são as informações a respeito da cadeia
produtiva desse produto, prejudicando a criação de políticas públicas para dinamizar a
comercialização dessa fibra. Com isso, este trabalho teve como objetivo de analisar os
efeitos da cadeia produtiva da fibra do buriti no mercado (preço e produção) durante os
anos de 1997 a 2017, no estado do Pará.
2. MATERIAL E MÉTODOS
Os dados utilizados neste trabalho foram coletados do Sistema IBGE de
Recuperação Automática (SIDRA). Os dados são referentes ao período de 1997 a 2017.
As variáveis estudadas foram: quantidade da produção anual de fibra de buriti no estado
do Pará em toneladas e o preço médio do buriti em R$/toneladas. O preço foi obtido por
meio da razão entre o valor monetário da produção e pela quantidade produzida, conforme
a equação 1.
𝑃 = 𝑉/𝑄 Eq. 1
Em que:
P = preço em R$/tonelada;
V = valor anual da produção (R$);
Q = quantidade anual produzida (toneladas).
A análise da taxa média geométrica de crescimento anual em percentagem
(TGC%) foi realizada ao nível de 5% de significância, por meio de regressão entre as
variáveis ano, quantidade produzida em toneladas, valor bruto da produção e valor real.
Seguido da aplicação da equação 2 no coeficiente b da variável independente.
𝑇𝐺𝐶 (%) = (𝐴𝑛𝑡𝑙𝑜𝑔 𝑏 − 1) 𝑥 100 Eq. 2

175
Belém (PA), 11 a 13 de dezembro de 2019
Para corrigir o efeito da inflação, foi realizado a correção inflacionária por meio
do Índice Geral de Preços Disponibilidade Interna (IGP-DI) da Fundação Getúlio Vargas
(FGV). O software utilizado para a análise dos dados foi o Microsoft Office Excel.
3. RESULTADOS E DISCUSSÃO
No estado do Pará, segundo IBGE (2017), os maiores produtores da fibra do buriti
foram os municípios de: Igarapé-Miri, com 89,5% da produção em relação ao Pará e
46,84% em relação ao Brasil; Abaetetuba e Cametá, ambos com 3,9% em relação ao Pará
e 2,04% em relação ao Brasil; Barcarena com 1,2% em relação ao Pará e 0,61% em
relação ao Brasil; correspondendo os 4 municípios a 98,5% da produção estadual e
51,53% da produção brasileira (Tabela 1).
Tabela 1 - Produção da fibra de buriti por município do estado do Pará em 2017.
Local Produção (toneladas) Percentual (Brasil) Porcentagem (Pará)
Brasil 491 100% -
Pará 257 52,34% 100%
Igarapé-Miri 230 46,84% 89,49%
Abaetetuba 10 2,04% 3,89%
Cametá 10 2,04% 3,89%
Barcarena 3 0,61% 1,17%
Oeiras do Pará 2 0,41% 0,78%
Tracuateua 2 0,41% 0,78%
Fonte: SIDRA (2019).
No município de Igarapé-Miri, além da elevada produção da fibra do buriti,
também é responsável pela maior produção de açaí no estado, tendo em vista que
apresenta grandes áreas com açaizais nativos nas ilhas. Dessa maneira, os buritizais
correm o risco de substituição por plantio de açaizais, os quais podem gerar uma maior
renda aos produtores locais, tendo em vista que apresentam uma elevada demanda, tanto
nacional quanto internacional (ARAÚJO; NAVEGANTES-ALVES, 2015). Além disso,
a substituição dos buritizais por açaizais também se dá devido aos financiamentos
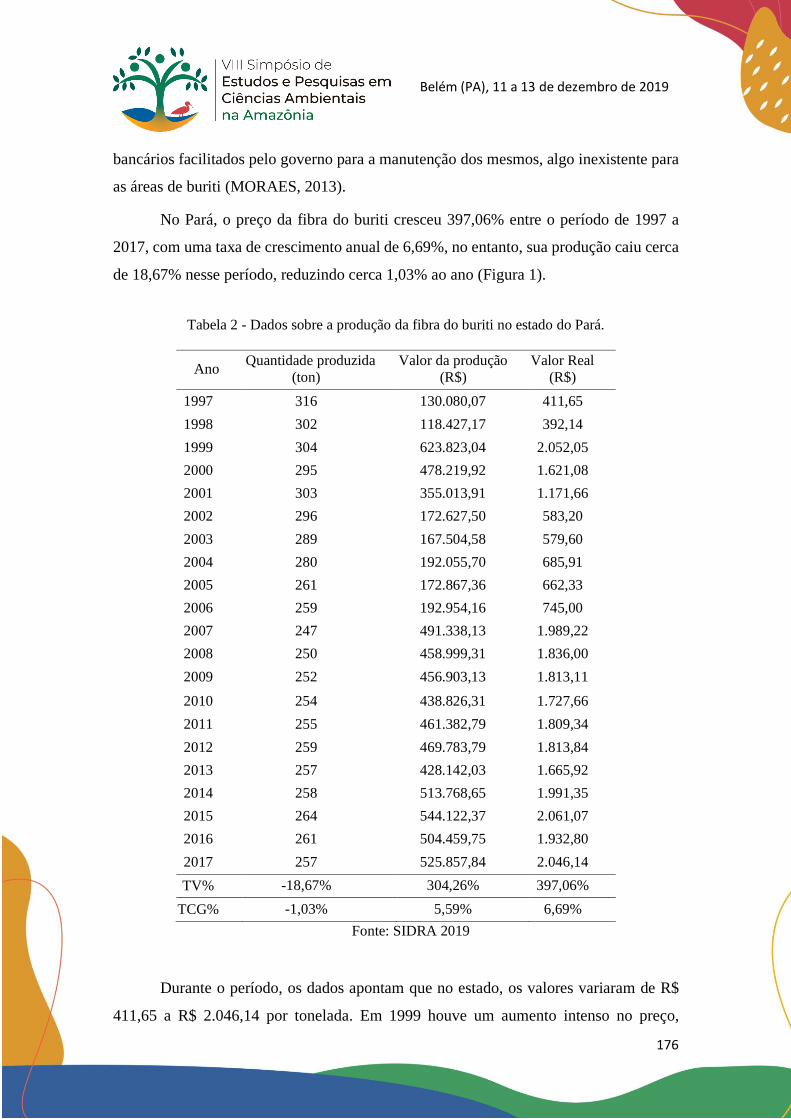
176
Belém (PA), 11 a 13 de dezembro de 2019
bancários facilitados pelo governo para a manutenção dos mesmos, algo inexistente para
as áreas de buriti (MORAES, 2013).
No Pará, o preço da fibra do buriti cresceu 397,06% entre o período de 1997 a
2017, com uma taxa de crescimento anual de 6,69%, no entanto, sua produção caiu cerca
de 18,67% nesse período, reduzindo cerca 1,03% ao ano (Figura 1).
Tabela 2 - Dados sobre a produção da fibra do buriti no estado do Pará.
Ano Quantidade produzida
(ton) Valor da produção
(R$) Valor Real
(R$)
1997 316 130.080,07 411,65
1998 302 118.427,17 392,14
1999 304 623.823,04 2.052,05
2000 295 478.219,92 1.621,08
2001 303 355.013,91 1.171,66
2002 296 172.627,50 583,20
2003 289 167.504,58 579,60
2004 280 192.055,70 685,91
2005 261 172.867,36 662,33
2006 259 192.954,16 745,00
2007 247 491.338,13 1.989,22
2008 250 458.999,31 1.836,00
2009 252 456.903,13 1.813,11
2010 254 438.826,31 1.727,66
2011 255 461.382,79 1.809,34
2012 259 469.783,79 1.813,84
2013 257 428.142,03 1.665,92
2014 258 513.768,65 1.991,35
2015 264 544.122,37 2.061,07
2016 261 504.459,75 1.932,80
2017 257 525.857,84 2.046,14
TV% -18,67% 304,26% 397,06%
TCG% -1,03% 5,59% 6,69%
Fonte: SIDRA 2019
Durante o período, os dados apontam que no estado, os valores variaram de R$
411,65 a R$ 2.046,14 por tonelada. Em 1999 houve um aumento intenso no preço,

177
Belém (PA), 11 a 13 de dezembro de 2019
seguido de uma sequência de quedas nos anos 2000 e 2001, e em 2007, novamente uma
acentuada elevação no preço da tonelada, sofrendo pouca variação desde então (Figura
1).
Figura 1 - Variação do preço da tonelada da fibra do buriti entre os anos de 1997 a 2017.
Fonte: SIDRA 2019
Diversos fatores podem ter influenciado no aumento repentino do preço da fibra
do buriti na década de 90. Hammet (1999) afirma que um dos fatores que pode ter
alavancado os PFNMs, tenha sido a Conferência das Nações Unidas sobre o Meio
Ambiente e Desenvolvimento, ocorrida em 1992 por meio da Agenda 21, onde
identificava os PFNMs como produtos da sustentabilidade, necessitando futuramente de
medidas apropriadas para melhor aproveitamento desses produtos.
Outro fator que também pode ter influenciado o aumento repentino do valor da
fibra do buriti, pode ter sido a “efêmera” crise monetária brasileira em 1999, na qual a
taxa de câmbio efetiva do real atingiu cerca de 36,6% (MURTA et al., 2003).
Em relação ao ano de 2007, um dos motivos que pode ter influenciado no aumento
contínuo do preço da fibra do buriti seria o decreto n° 6.040, instituindo a Política
Nacional de Desenvolvimento Sustentável dos Povos e Comunidades Tradicionais, que

178
Belém (PA), 11 a 13 de dezembro de 2019
reconhecia povos e comunidades como usuários dos recursos naturais para subsistência e
desenvolvimento econômico, assegurando que os recursos naturais para reprodução
cultural, física e econômica (SARAIVA, 2009). Além disso, a constância do preço
elevado da fibra do buriti entre o período de 2007 a 2017, pode estar relacionado com a
implantação dos produtos extrativistas na Política Nacional de Garantia de Preços
Mínimos (PGPM), que garantia aos produtores extrativistas manter a oferta, mesmo com
oscilações constantes no mercado (SARAIVA, 2009).
A queda na produção da fibra do buriti, ocorrida ao longo dos anos, pode estar
relacionado à sua cadeia produtiva. De acordo com Cattani (2016) devido o buriti ser uma
espécie de crescimento lento, a intensa atividade extrativista não respeita o tempo de
recuperação, que varia de 6 a 8 meses, tendo como resultado a morte inevitável dessa
palmeira. Por isso, Souza e Perpétuo (2016) sugerem que, para assegurar a sobrevivência,
a extração deve acontecer obedecendo a periodicidade para não danificar ou até mesmo
matar a palmeira.
A prática do extrativismo para obtenção da matéria prima pode influenciar
diretamente na sua oferta. Devido ao esgotamento dos buritizais nas localidades próximas
às comunidades, ocasionada pela falta de manejo adequado, tem como resultado a morte
dos indivíduos, fazendo com que seja necessário a extração em áreas mais distantes,
encarecendo e desestimulando a produção da fibra (FERNANDES-PINTO, 2006;
ALMEIDA et al., 2009), confirmando a teoria de Homma (2014), sobre as limitações do
extrativismo para atendendimento das demandas crescentes de mercado.
4. CONCLUSÃO
Com o aumento da dificuldade e distância para se chegar até as palmeiras de buriti,
para a extração das folhas, tem-se o crescente risco de substituição das áreas de buritizais
nativas por plantios de açaizais, tendo em vista que essas que proporcionam uma maior
renda e segurança alimentar para a população local. Estes fatores comprometem a
produção da fibra do buriti no estado do Pará, tendendo a redução considerada da
produção. O apoio e incentivo do governo com implantação de políticas públicas devem
ser desenvolvidos, para que haja a continuidade da produção por meio de manejos
sustentáveis das palmeiras e ampliação do mercado para os produtos derivados da fibra

179
Belém (PA), 11 a 13 de dezembro de 2019
de buriti, visto que é uma cadeia produtiva que sustenta várias famílias e apresenta uma
grande importância na tradição e na cultura das comunidades paraenses.

180
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALMEIDA, A. N.; BITTENCOURT, A. M.; SANTOS, J. A.; LOYOLA, C. E.;
SANTOS, V. S. Evolução da produção e preço dos principais produtos florestais não
madeireiros extrativos do Brasil. Revista Cerne, Lavras, v. 15, n. 3, 2009.
ALMEIDA, S. P; SILVA, J. A. Pequi e buriti: importância alimentar para a população do
Cerrado. Planaltina: EMBRAPA-CPAC, 1994.
ARAÚJO, C. T. D de; NAVEGANTES-ALVES, L. F de. Do extrativismo ao cultivo
intensivo do açaizeiro (Euterpe oleracea Mart.) no estuário amazônico: sistemas de
manejo e suas implicações sobre a diversidade de espécies arbóreas. Revista Brasileira
de Agroecologia, nº 10, pg. 12-23, 2015.
CATTANI, I. M. Fibra de Buriti (Mauritia flexuosa Mart.): Registro em comunidade
local (Barreirinhas - MA), caracterização fisico-quimica e estudo com impregnação
de resinas. 213f. Dissertação (Mestrado) Curso de pós-graduação em Têxtil e moda,
universidade de São Paulo, São Paulo - SP, 2016.
CATTANI, I. M.; BARUQUE-RAMOS, J. Fibra de buriti (Mauritia flexuosa Mart.) e
aplicações em produtos têxteis. Escola de Artes, Ciências e Humanidades –
Universidade de São Paulo – EACH-USP - São Paulo – Brasil – 03828-000, 2014.
FERNANDES-PINTO, É.; SARAIVA, N. A. Fauna silvestre associada a buritizais –
etnoecologia e conservação em uma região do semi-árido Maranhense. In: VII Congresso
Internacional sobre Manejo de Fauna Silvestre na Amazônia e América Latina, 2006,
Ilhéus. Resumo...Ilhéus: 2006.
FIEDLER, N. C.; SOARES, T. S.; da SILVA, G. F. Produtos Florestais Não Madeireiros:
Importância e Manejo Sustentável da Floresta. Revista Ciências Exatas e Naturais,
Vol.10 nº 2, 2008.
Flora do Brasil 2020 em construção. Jardim Botânico do Rio de Janeiro. Disponível
em: http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB15723. Acesso em: 31 Jan.
2019
HAMMET, T. Special forest products: identifying opportunities for sustainable
forestbased development (part 1). Virginia Forest Landowner Update, v. 13, n. 1, 1999.
HOMMA, A. K. O. Extrativismo vegetal na Amazônia: história, ecologia, economia e
domesticação, Brasília, DF: Embrapa, 2014.
MORAES, Y. S. Brinquedo de miriti e o desenvolvimento local no município de
Abaetetuba. 2013. 120f. 2017. Tese de Doutorado. Dissertação (Mestrado), Núcleo do
Meio Ambiente, Universidade Federal do Pará, Belém, 2013.
MURTA, L. R.; BRASIL, G. H.; SAMOHYL, R. W. Crise monetária brasileira de 1999:
uma análise econométrica da desvalorização do real em 1999. XXXV SBPO, 2003, Natal.

181
Belém (PA), 11 a 13 de dezembro de 2019
SARAIVA, N. A. Manejo Sustentável e Potencial Econômico da Extração do Buriti
nos Lençóis Maranhenses. 2009. 129 f. Dissertação (Mestrado) - Universidade de
Brasília, Centro de Desenvolvimento Sustentável.
SARAIVA, N. A. Manejo Sustentável e Potencial Econômico da Extração do Buriti
nos Lençóis Maranhenses, Brasil. 2009. Dissertação de Mestrado. Centro de
Desenvolvimento Sustentável. Universidade de Brasília, Brasília, DF.
SOUZA, N. F. S.; VIANA, D. S. F. Aspectos ecológicos e potencial econômico do buriti
(Mauritia flexuosa). Revista Agrarian Academy, Centro Científico Conhecer - Goiânia,
v.5, n.9, p. 535, 2018.

182
Belém (PA), 11 a 13 de dezembro de 2019
DINÂMICA DO TRABALHO NA ATIVIDADE EXTRATIVISTA DA Carapa
guianensis Aubl. (ANDIROBA) EM PROJETOS DE ASSENTAMENTOS
AGROEXTRATIVISTAS (PAEX) DAS VÁRZEAS DO MUNICÍPIO DE
IGARAPÉ-MIRI, PARÁ
Aline Dias Brito1; Carlos Anderson Sousa de Carvalho2; Tayse Fernanda do Amaral
Silva3 Louise Ferreira Rosal4
Roberta de Fatima Rodrigues Coelho5
1Engenheira Agrô[email protected] 2 Engenheiro Agrônomo. [email protected] 3Engenheira Agrônoma. [email protected]
4Engenheira Agrônoma. IFPA. [email protected] 5Engenheira Florestal. IFPA. [email protected]
RESUMO
Este estudo tem como objetivo analisar a dinâmica do trabalho na atividade extrativista
da Carapa guianensis Aubl. (andiroba) em Projetos de Assentamentos Agroextrativistas
(PAEX) das várzeas do Município de Igarapé-Miri, PA. A coleta de dados aconteceu nos
PAEX das ilhas Mamangal, Mutirão, Buçu e Jarimbu, através de questionário usando a
metodologia snowball ou “Bola de Neve”, que permitiu alcançar uma base de dados de
14 extrativistas. Sobre o trabalho com a andiroba, verificou- se 4 (quatro) são coletores
de sementes, 4 (quatro) são coletores de sementes e extratores de óleo e 6 (seis) são
extratores de óleo, sendo a maioria dos extrativistas constituída por mulheres
correspondendo a 11 dos entrevistados e apenas 3 (três) homens. A principal organização
social de trabalho para comercialização de sementes é Cooperativa Agrícola dos
Empreendedores Populares de Igarapé-Miri (CAEPIM), sendo o óleo comercializado de
forma individual. A atividade de coleta de sementes para venda e para a extração do óleo
é uma alternativa viável de renda, mais, faz-se necessário ações para potencializar o
comercio de óleo para além das várzeas.
PALAVRAS-CHAVE: Produtos não madeireiros; Organização social; Extrativismo.
Área de Interesse do Simpósio: Unidades de conservação

183
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A Carapa guianensis Aubl. (andiroba) possui grande valor econômico atribuído
a madeira e o óleo extraídos das sementes. O óleo de andiroba é um produto florestal não
madeireiro (PFNM) bastante utilizado pela população amazônica, sendo um dos
principais óleos considerados tradicionais na região norte do país para fins medicinais e
cosmecêutica, tornando-o um óleo de alto interesse econômico (LOURENÇO et al.,
2017).
Nas áreas de várzeas do município de Igarapé-Miri, Região do baixo Tocantins,
apresentam grande potencial para a produção florestal durante a entressafra do açaí,
principal cultura de consumo e renda dos extrativistas. Nesse contexto, a andiroba possui
grande potencial para o manejo e uso sustentável, através da atividade de coleta de
sementes para a extração de óleo, podendo promover o desenvolvimento local e renda aos
extrativistas. Além disso, a atividade de coleta de sementes e extração de óleo, promove
a conservação da biodiversidade, comparado a extração de madeira e outros usos da terra,
os quais implicam em conversão da floresta.
Existem poucas informações sobre as formas de trabalhos com a andiroba nas
várzeas de Igarapé-Miri, e por essa razão, tornam-se necessários estudos visando
potencializar o reconhecimento, a disseminação e reprodução de dados consistentes sobre
andiroba. Diante disso, este trabalho tem como objetivo analisar a dinâmica do trabalho
na atividade extrativista da Carapa guianensis Aubl. (andiroba) em Projetos de
Assentamentos Agroextrativistas (PAEX) das várzeas do Município de Igarapé-Miri, PA.
2. MATERIAL E MÉTODOS
A pesquisa ocorreu nos projetos de assentamentos agroextrativistas (PAEX) das
ilhas Mamangal, Mutirão, Buçu e Jarimbu (Figura 01), localizada no município de
Igarapé-Miri, localizado na região do Baixo Tocantins, mesorregião do nordeste
paraense, a 78 Km da capital do estado do Pará.

184
Belém (PA), 11 a 13 de dezembro de 2019
Figura 1 - Mapa de localização do PAEX no município de Igarapé-Miri, PA.
Fonte: elaborada pelo autor, 2018
Os PAEX são destinados à exploração de área dotadas de riquezas extrativas,
através de atividades economicamente viáveis, socialmente justas e ecologicamente
sustentáveis partindo da lógica do uso coletivo da terra (SANTOS et al., 2017). Para
Silva et al. (2017), a regularização por meio do uso coletivo da terra garante a proteção
dos direitos territoriais das famílias agroextrativista, surgindo novas oportunidade de
produção agrícola, manutenção da diversidade e dignidade as populações tradicionais.
A pesquisa foi de cunho qualitativo com auxílio de questionários, os entrevistados
foram selecionados por amostragem não probabilística, através da metodologia snowball
ou “Bola de Neve” apresentada por Vinuto (2014), em que os participantes iniciais de um
estudo, indicam novos participantes ao pesquisador, que por sua vez indicam novos
participantes e assim sucessivamente até que seja alcançado o objetivo proposto. Esta
metodologia permitiu alcançar, alcançar 14 extrativistas que exercem a atividade com a
andiroba.
As entrevistas foram gravadas e para o consentimento na participação da pesquisa,
foram disponibilizados aos respondentes o Termo de Consentimento Livre e
Esclarecimento (TCLE) de acordo com as diretrizes estabelecidas pela resolução nº
466/12 (BRASIL, 2012).
A pesquisa foi cadastrada na plataforma Sistema de Autorização e Informação em
Biodiversidade (SISBio) regida pela instrução Normativa nº 03 de 01 de setembro de

185
Belém (PA), 11 a 13 de dezembro de 2019
2014. Os dados obtidos nos questionários foram tabulados em planilha no programa Excel
2010, e analise de estatística descritiva.
3. RESULTADOS E DISCUSSÃO
Verificou- se que dos 14 entrevistados, 4 (quatro) são coletores de sementes,
fazem apenas a venda de sementes de andiroba como uma oportunidade de renda, uma
vez que, não possuem conhecimento e a prática de extração de óleo; 4 (quatro) são
coletores de sementes e extratores de óleo, coletam as sementes tanto para a venda, quanto
para extração do óleo e o óleo extraído é destinado para consumo e venda, desta maneira,
visam o aumento da renda familiar praticando as duas atividades, e 6 (seis) são extratores
de óleo, coletam as sementes apenas para a extração do óleo que são destinados a venda
e ao uso pessoal.
O núcleo familiar dos entrevistados, possui em média 6 (seis) pessoas por unidade
de produção familiar (UPF). A dinâmica do trabalho nessas ilhas é desenvolvida a partir
da divisão social de trabalho de homens e mulheres (Gráfico 1 e 2). A maioria dos
extrativistas que trabalham com andiroba são mulheres correspondendo a 11 dos
entrevistados e apenas 3 (três) são homens.
Os homens são os principais responsáveis pelo extrativismo da Euterpe oleraceae
Mart. (açaí), desenvolvendo paralelamente atividades secundárias como produção de
hortaliças, pesca e participam da coleta de andiroba. As mulheres desempenham uma
força de trabalho diversificada dentro do núcleo familiar extrativista, conciliando o
trabalho doméstico a outras atividades dentro e fora das UPF, sendo as principais
responsáveis pela coleta de sementes e extração de óleo da andiroba.
O trabalho realizado pelas mulheres é em função do conhecimento tradicional
que elas adquiriram através da oralidade com seus familiares, se destacando neste cenário
como um elemento essencial para preservação das andirobeiras nos sistemas de produção
familiar (SPF).

186
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2- Principais atividades desenvolvidas pelos homens na UFP do PAEX no Municipio
de Igarapé-Miri.
Fonte: autores.
Figura 3- Principais atividades desenvolvidas pelas mulheres na UFP do PAEX no Municipio
de Igarapé-Miri.
Fonte: autores.
Estudos realizados por Mota et al. (2014) entre as catadoras de mangaba no estado
de Sergipe e Soares et al. (2017) em Salvaterra no Marajó no estado do Pará, destacam
que são as mulheres as principais responsáveis pelo extrativismo, e através da organização
de trabalho, controlam os recursos da comercialização das sementes utilizados no sustento
da família, denotando o exercício de autonomia sobre essas atividades. Santos (2013)
destaca que a prática de trabalho associado a andiroba realizado pelas mulheres, ocorre
em função dos conhecimentos acerca dessa espécie, sendo também as principais
responsáveis pela transmissão de conhecimentos a geração futura. Nesse sentido, as
mulheres contribuem para a gestão dos recursos do ecossistema local, através do manejo
desta espécie dentro das unidades produtivas que é feito exclusivamente pelos homens,
seja marido ou filhos.
A organização social de trabalho nas várzeas de Igarapé-Miri é bem diversificada.
A maioria dos extrativistas estão articulados através de cooperativas e associações como

187
Belém (PA), 11 a 13 de dezembro de 2019
estratégia para fortalecer o trabalho coletivo e, assim, viabilizar a comercialização de
produtos. Dentre as organizações sociais, a Cooperativa Agrícola dos Empreendedores
Populares de Igarapé-Miri (CAEPIM) realiza a comercialização das sementes de andiroba
para uma empresa multinacional. Essa cooperativa tem um papel importante para o
fortalecimento da cadeia de produtos da sociobiodiversidade da várzea mirense
contribuindo para economia tradicional local.
Outras organizações importantes destacadas pelos extrativistas foram a
Associação dos Minis e Pequenos Produtores Rurais de Igarapé-Miri - Mutirão, que
segundo Reis (2008) é uma entidade originariamente agrícola, constituída por
trabalhadores rurais, com objetivos e finalidades de buscar melhores condições de vida
da população e das comunidades, a partir do usufruto de direitos políticos, institucionais
e civis; a colônia dos pescadores denominada pelos ribeirinhos CACIPESCA, um projeto
voltado para o fortalecimento dos acordos de pesca como iniciativas de gestão
comunitária de recursos aquáticos e o Sindicato dos Trabalhadores e trabalhadoras Rurais
(STTR) de Igarapé-Miri.
Apesar de várias organizações sociais, nenhuma faz a facilitação do comercio do
óleo, dessa forma cada extrator vende sua produção de forma individual. Uma parte do
óleo extraído é destinado para uso familiar e em alguns casos é destinado a doação a
parentes e amigos, e a outra parte é vendida.
O litro do óleo de andiroba é vendido entre R$ 10,00 a R$ 40,00 diretamente ao
consumidor, que na maioria são os próprios moradores da região, venda para atravessador
e olarias. Dessa maneira, pode-se perceber que a produção de óleo de andiroba nas
várzeas tem contribuído significativamente para geração de renda e conservação
ambiental.
4. CONCLUSÃO
A atividade de coleta de sementes para venda e para a extração do óleo é uma
alternativa viável de renda dos ribeirinhos nas várzeas do município de Igarapé-Miri no
período que o açaí está na entre safra. Essa atividade é desenvolvida principalmente pelas
mulheres que neste cenário fazem presente como força de trabalho, sedo elemento
fundamental para a conservação dessa espécie na várzea miriense.

188
Belém (PA), 11 a 13 de dezembro de 2019
Apesar de várias organizações sociais, apenas uma facilita a comercialização de
sementes e o óleo passa a ser comercializado de forma individual, sendo necessário ações
que partam dos próprios extrativistas para potencializar o comercio de óleo para além das
várzeas e fortalecer a produção, já que a demanda por esse produto é grande.

189
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BRASIL. Resolução n. 466, de 12 de dezembro de 2012. Aprova diretrizes e normas
regulamentadoras de pesquisas envolvendo seres humanos. Saudelegis: Diário Oficial
da União, Brasília, v.24, 15p, 2012.
GUIMARÃES, S. T. Mulheres e florestas: um estudo sobre comunidades tradicionais no
PE da Serra do Mar, Núcleo Santa Virginia (PESM-NSV), estado de São Paulo, Brasil/
Women and Forests: a study of traditional communities (...). Caderno de Geografia, v.
24, n. 42, p. 264-286, 2014.
LOURENÇO, J. N. DE P. et al. Produção, biometria de frutos e sementes e extração
do óleo de andiroba (Carapa guianensis Aublet.) sob manejo comunitário em
Parintins, AM. EMBRAPA, Amazônia Oriental, 2017. Disponível em:
<https://www.infoteca.cnptia.embrapa.br/infoteca/bitstream/doc/1100708/1/Doc138.pdf
>. Acesso em: 27 abr. 2019.
MENEZES, A. J. E. A. O histórico do sistema extrativo e a extração de óleo de andiroba
cultivado no município de Tomé-Açu, estado do Pará. 2005. Disponível em:
https://www.alice.cnptia.embrapa.br/alice/bitstream/doc/576188/1/142.pdf. Acesso em:
15 nov. 2018.
MOTA, D. M. D.; SCHMITZ, H., SILVA JÚNIOR, J. F. D.; RODRIGUES, R. F. D. A.
O trabalho familiar extrativista sob a influência de políticas públicas. Revista de
Economia e Sociologia Rural, v 52, 189-204. 2014.
REIS, A. A. Estratégias de desenvolvimento local sustentável da pequena produção
familiar na várzea do município de Igarapé-Miri (PA). 2008. 128 p. Dissertação
(Mestrado em Desenvolvimento sustentável) - Universidade Federal do Pará., Belém,
2008.
SANTOS, M. Conhecimento Ecológico Local sobre as Andirobeiras e a Extração
Artesanal do Óleo de Andiroba em Uma Área de Proteção Ambiental, Floresta de
Várzea Periurbana. Macapá. 2013. 110p. Dissertação (Biodiversidade Tropical) -
Universidade Federal do Amapá.
SANTOS, R. O.; LIMA, R. C.; CARMO LIMA, R.; BORGES LIMA, R., APARICIO,
P. S., ABREU, J. C. Florística e estrutura de uma comunidade arbórea na floresta
estadual do Amapá, Amazônia Oriental, Brasil. Nativa, v.5, n. 05, p.529-539, 2017.
SILVA, V. D. A. D., CARDOSO, M. C., BENTES, A. J. M., & VIEIRA, T. A.
Assentamento agroextrativista e a dinâmica de desenvolvimento na Amazônia: um
estudo de caso da comunidade São Braz no PAE eixo forte em Santarém/pa. 2017.
SOARES, I. D. S.; OTOBO, A. O.; RIBEIRO, C. F. A.; ESPIRITO SANTO, K. R.A.
Caracterização socioeconômica e de aspectos produtivos do extrativismo de sementes
oleaginosas amazônicas na mesorregião do Marajó-Município de Salvaterra, Pará, Brasil.
Revista Observatorio de la Economía Latinoamericana, v2, p. 1-23, 2017.

190
Belém (PA), 11 a 13 de dezembro de 2019
VINUTO, J. A amostragem em bola de neve na pesquisa qualitativa: um debate em
aberto. Temáticas, v.22, n. 44, p. 203-220, 2014.

191
Belém (PA), 11 a 13 de dezembro de 2019
DESCRIÇÃO DOS FATORES ORGANIZACIONAIS E AMBIENTAIS DE UMA
EMPRESA MADEIREIRA: ESTUDO DE CASO NO DISTRITO DE ICOARACI,
BELÉM/PA
Rodrigo Corrêa de Castro1; Caroline das Graças dos Santos Ribeiro2; Lucas Silva
Pereira3
1Especialista em Perícia Ambiental. Faculdade Estácio de Belém. E-mail:
2Especialista em Educação Ambiental. Faculdade Estácio de Belém. E-mail:
3Graduando em Engenharia Sanitária e Ambiental. Universidade Federal do Pará. E-
mail: [email protected]
RESUMO
A pesquisa buscou mapear o processo organizacional e ambiental das atividades
exploratórias de uma empresa madeireira localizada no Distrito de Icoaraci, Belém/PA.
As empresas madeireiras visam se adequar às normas organizacionais e ambientais frente
à exploração excessiva e indiscriminada dos recursos florestais. O progresso do
desenvolvimento sustentável está associado a crescente e contínua sensibilização da
sociedade. Logo, necessita-se aprender e elaborar boas estratégias organizacionais,
econômicas e ambientais com assessoria de técnicas e ferramentas, as quais alinhem o
desempenho da empresa às demandas sociais como o planejamento, o controle das
unidades de manejo florestal e a qualificação dos seus colaboradores. Logo, é importante
criar e manter um cronograma de treinamentos focados na segurança e qualificação
técnica dos profissionais. Assim, o objetivo do estudo é identificar o processo produtivo;
quantificar e classificar as ferramentas e técnicas utilizadas; e verificar a contribuição da
empresa quanto ao desenvolvimento sustentável no setor madeireiro. A pesquisa
apresenta metodologia quantitativa e método exploratório. As informações pertinentes à
execução da pesquisa foram obtidas in loco. E as visitas técnicas na empresa ocorreram
entre os meses de agosto e setembro de 2018. Então, delimitou-se a área de estudo para
mapear a empresa de forma mais eficiente. A empresa demonstrou preocupação em
planejar suas atividades exploratórias, pois além de cumprir todas as normas estabelecidas
na Legislação Ambiental, apresentou um Plano de Manejo Florestal Sustentável próprio
de suas áreas de exploração, cuja aprovação ocorreu por intermédio da Secretaria de Meio
Ambiente e Sustentabilidade (SEMAS). A indústria madeireira é comercializada
conforme os parâmetros e regras exigidos pela SEMAS. Identificou-se que a empresa
possui fundamentos e práticas de desenvolvimento sustentável em suas atividades
exploratórias como fator de minimização aos impactos no segmento madeireiro. Ela
inclui em sua estrutura organizacional atividades de planejamento, responsabilidades
práticas e procedimentos que visam manter a Política Ambiental compreensível.
Palavras-chave: Atividades Exploratórias. Recursos Florestais. Desenvolvimento
Sustentável.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal.

192
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Para Woitchunas e Silva (2008), a Revolução Industrial é o marco histórico da
substituição da força de trabalho humano pelo emprego das máquinas. Isso gera grande
transformação na organização e divisão do trabalho, visto que é necessário simplificar e
padronizar as atividades produtivas para que se adequem aos processos organizacionais
e gestacionais de tarefas das grandes empresas que surgiram no século XVIII.
De acordo com a assertiva de Brum (1996), a diversidade dos recursos naturais
propiciou ao Brasil estabelecer uma relação predatória entre o homem e a floresta, o que
comprometeu a resiliência do meio ambiente. À vista disso, a utilização indiscriminada
da madeira é efetivada e identificada desde a descoberta da América por Cristóvão
Colombo.
Dean (1994) diz que a divisão de terras por sesmarias possibilitou o processo de
devastação da mata nativa, cuja intensificação abriu espaço para a exploração de outros
recursos naturais e, assim, gerou um ciclo de economia. A devastação das florestas
primárias ocorria sem dificuldade, pois os subprodutos eram doados ou trocados, a fim
de estabelecer uma relação de comércio simplificada.
Segundo Dean (1996), apesar do histórico de excessiva exploração indiscriminada
das empresas madeireiras na Amazônia Legal, busca-se adequar os seus padrões de
produção às normas e leis ambientais através dos Sistemas de Plano de Manejo Florestal
e Licitação de Florestas Públicas. Essas ações requerem um processo organizacional
planejado, desde suas atividades pré-exploratórias até suas atividades pós-exploratórias.
Consoante com Machado (2002), a colheita florestal é definida como um conjunto
de operações efetuadas no maciço florestal, o qual visa preparar, extrair e transportar a
madeira através do uso de técnicas e padrões estabelecidos com a finalidade de
transformá-la em produto manufaturado. Essa etapa é a mais importante do ponto de vista
técnico-econômico, pois ela é composta por uma série de ações que geram o produto final.
Dessa forma, o manejo florestal sustentável é aplicado como um processo de
gerenciamento e planejamento para propor as melhores intervenções socioeconômicas e
ambientais a médio e longo prazo.

193
Belém (PA), 11 a 13 de dezembro de 2019
O objetivo do estudo é identificar o processo produtivo; quantificar e classificar
as ferramentas e técnicas utilizadas; e verificar a contribuição da empresa quanto ao
desenvolvimento sustentável no setor madeireiro no Distrito de Icoaraci, Belém/PA.
2. METODOLOGIA
Em relação à natureza da pesquisa, Cervo e Bervian (2004) dizem que ela é
quantitativa, pois o investigador é movido pela vontade de contribuir para fins práticos,
buscando investigar e solucionar problemas concretos. E o tipo de pesquisa é classificado
como exploratório, haja vista que se utiliza o estudo de caso como método de
investigação.
As informações necessárias à execução da pesquisa foram obtidas in loco, por
meio de análise de dados oriundos de registros históricos da Indústria Madeireira em
questão, que foram produzidos pelo proprietário responsável e os demais colaboradores,
os quais contêm livros de registro em seu domínio.
As visitas técnicas na empresa ocorreram entre os meses de agosto e setembro de
2018. Elas foram acompanhadas pelos responsáveis que exercem atividades exploratórias
de floresta manejada e, além disso, contêm uma ampla visão sobre os processos
organizacionais que envolvem o manejo florestal.
De tal maneira, delimitou-se a área de estudo para mapear a empresa de forma
mais eficiente. Logo, evidenciou-se o sistema de exploração florestal; o sistema pré
exploratório; a construção de infraestrutura de colheita, corte e derrubada direcionada; o
arraste de tora; a operação de pátio, logística e transporte; a organização, comando,
coordenação e controle.
3. RESULTADOS E DISCUSSÃO
A empresa demonstrou preocupação em planejar suas atividades exploratórias,
pois além de cumprir todas as normas estabelecidas na Legislação Ambiental, apresentou
um Plano de Manejo Florestal Sustentável próprio de suas áreas de exploração, cuja
aprovação ocorreu por intermédio da Secretaria de Meio Ambiente e Sustentabilidade
(SEMAS).

194
Belém (PA), 11 a 13 de dezembro de 2019
O sistema praticado pela empresa envolve práticas relacionadas aos processos de
colheita de madeira e a regeneração natural para garantir as produções futuras. A
exploração florestal da empresa tem como prática: o planejamento da safra e uso de mapas
logísticos para auxiliar na derrubada das árvores e no arraste das toras, assim como a
orientação necessária para realizar as técnicas operacionais.
A seguir, listam-se algumas das atividades que compõe o sistema de exploração
da empresa, as quais são de fundamental importância para compreender o estudo de caso:
• Inventário pré-exploratório
Ele consiste na demarcação das unidades de trabalho e identificação, classificação,
medição e localização de todas as árvores de espécies comerciais e potencias acima de 40
cm de diâmetro a altura do peito (DAP). Nesta etapa são coletadas informações da
topografia do terreno e condições adversas da floresta como: áreas com muitos cipós e
clareiras. Todas as informações são tratadas e servirão para a realização de todo o
planejamento da colheita.
• Construção da infraestrutura de colheita
Toda infraestrutura é construída respeitando os limites ambientais e o prévio
planejamento realizado com base no mapeamento resultante do inventário pré-
exploratório, o qual evita o cruzamento de áreas de preservação permanente e árvores
protegidas.
• Corte e derrubada direcionada
O operador de motosserra e seu auxiliar, de posse dos mapas, encontram as
árvores indicadas para a derrubada. Nesse momento, o operador e seu auxiliar iniciam os
procedimentos de verificação de possibilidade de derrubada da árvore, seguindo os
seguintes procedimentos:
• Fazer o teste de oco;
• Verificar a direção de queda natural da árvore;
• Definir a direção de queda desejável (derruba direcionada), baseado em critérios
de direcionamento de queda;
• Abrir os caminhos de fuga;
• Executar a derrubada direcionada da árvore.
São derrubadas as árvores destinadas à produção madeireira, cujos atributos sejam
coincidentes com os critérios de seleção de corte. No momento em que uma destas árvores

195
Belém (PA), 11 a 13 de dezembro de 2019
não for cortada por motivos identificados no momento da derruba, ocorre a substituição
por outra árvore assinalada, sendo obrigatoriamente da mesma espécie.
• Arraste de toras
É realizado com tratores skidder e, eventualmente, por tratores de esteira adaptada
para o arraste, cujos planejamentos seguem as trilhas sinalizadas com fitas coloridas para
diferenciar as trilhas principais das trilhas secundárias. Ressalta-se que cada trator deve
trafegar em uma trilha distinta para evitar o seu alargamento.
• Operação de pátio
Consiste no processo de traçado, identificação e romaneio de toras. Neste
momento, inicia-se o controle por meio de volume real, por isso é muito importante que
seja realizada corretamente para evitar prejuízos de ordem econômica e ambiental. Os
erros nesta etapa podem ocasionar mau aproveitamento de toras.
• Logística e transporte
O transporte é realizado através de carretas que movimentam a matéria-prima
entre as Unidades de Produção Anual (UPA) e os portos. Ela fica estocada até nova
movimentação que ocorre por via fluvial, com origem na balsa até o local de
beneficiamento da indústria.
Nas movimentações entre a floresta e os portos; e entre os portos e destino final,
o transporte somente ocorre após a emissão do romaneio de toras, Guia Florestal (GF) e
Nota Fiscal (NF). No baldeio a movimentação ocorre sem a necessidade de
documentação, visto que é realizado dentro da UPA.
• Organizar
Para a empresa, organizar é o processo de identificar, dividir e alocar o trabalho.
Nesse sentido, ela identifica, divide e grupa o trabalho a ser realizado; define
responsabilidades e autoridades, assim como estabelece as relações entre os grupos, de
modo a possibilitar que as pessoas trabalhem de maneira eficaz para atingirem os
objetivos.
• Comandar
A empresa entende que comandar significa interpretar os planos para as pessoas e
dar as instruções e orientações sobre como executá-los, assim como garantir o alcance
dos objetivos. A empresa acredita que é necessário definir os objetivos; traçar as
estratégias para alcançá-los; estabelecer o planejamento e estruturar a organização. As

196
Belém (PA), 11 a 13 de dezembro de 2019
atividades e os processos de direção são os meios adequados para o sistema funcionar de
maneira eficaz.
• Coordenar
A empresa entende que coordenar é ligar, unir, harmonizar todos os atos e esforços
para que se chegue aos objetivos traçados. Por isso ela coordena suas ações através de
planejamentos de todas as suas etapas no processo de exploração da madeira.
• Controlar
Na empresa são controlados os documentos de registros de treinamentos, os
relatórios produzidos etc. As etapas da colheita serão controladas através de registros
manuais, os quais serão transcritos ao sistema de planilhas eletrônicas para apuração dos
dados e emissão de relatórios gerenciais e de produção. Eles serão divulgados à diretoria,
sob responsabilidade do Engenheiro Florestal da empresa. Não obstante, menciona-se que
todos os arquivos, físicos ou eletrônicos, serão preservados por um período mínimo de
três anos.
4. CONSIDERAÇÕES FINAIS
A indústria madeireira é comercializada conforme os parâmetros e regras exigidos
pela SEMAS. Identificou-se que a empresa possui fundamentos e práticas de
desenvolvimento sustentável em suas atividades exploratórias como fator de minimização
aos impactos no segmento madeireiro. Ela inclui em sua estrutura organizacional
atividades de planejamento, responsabilidades práticas e procedimentos que visam
manter a Política Ambiental compreensível.
De acordo com as funções organizacionais descritas, constata-se que a empresa
tem como funções administrativas: prever, organiza, comandar, coordenar e controlar
suas atividades de exploração etc. No entanto, mesmo com a inserção de fundamentos e
práticas organizacionais e ambientais adequados, é preciso que a empresa tenha mais
conhecimento sobre os impactos positivos e negativos sobre a atividade de mercado que
ela está inserida.
Portanto, faz-se necessário contratar profissionais capacitados na área ambiental
para promover o gerenciamento necessário à evolução da empresa, assim como contratar
profissionais de marketing para que a empresa ganhe destaque no cenário econômico e,

197
Belém (PA), 11 a 13 de dezembro de 2019
assim, invista em políticas empresariais e socioambientais consoantes com a realidade
local.

198
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BRUM, A. J. O desenvolvimento econômico brasileiro. 16. ed. Petrópolis: Vozes, 1996.
CERVO, A. L.; BERVIAN, P. A. Metodologia científica. 5.ed. São Paulo: Prentice Hall,
2004
DEAN, W. A ferro e fogo: A história da devastação da mata atlântica. São Paulo:
Companhia das Letras, 1997.
MACHADO, C. C. Colheita Florestal. Ed. Viçosa, MG: UFV, 2002. 468 p.
RONDOBEL MADEIREIRA. Resumo Público do Plano de Manejo Florestal
Sustentado: Praticando Manejo Florestal para Manter a Floresta Viva. Belém: Marina
Silva, 2012. 19 p.
RONDOBEL MADEIREIRA. Sistema de Cadeia de Custódia Florestal: Procedimento
Operacional. Belém: João Paulo Lima, 2009.
WOITCHUNAS, F. L.; SILVA, M. Fundamentos da gestão organizacional. Ed. Unijuí,
2008. 128 p.

199
Belém (PA), 11 a 13 de dezembro de 2019
EQUAÇÕES DE CRESCIMENTO EM ALTURA DOMINANTE PARA UM
PLANTIO CLONAL DE Tectona grandis Linn F.
Welton dos Santos Barros1, Mario Lima dos Santos2; Beatriz Cordeiro Costa3; Dione
Dambrós Raddatz4; Walmer Bruno Rocha Martins5
1 Graduando em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 2 Doutorando em Ciências Florestais. Universidade de Brasília.
[email protected]. 3 Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 4 Doutorando em Ciências Florestais. Universidade de Brasília. Universidade Federal
Rural da Amazônia.
[email protected]. 5 Professor orientador. Universidade Federal Rural da Amazônia.
RESUMO
Este trabalho teve por objetivo ajustar e selecionar o modelo que melhor expressar a
tendência de crescimento de um plantio clonal de Tectona grandis Linn F. (teca), no
município de Capitão Poço, PA. O estudo foi realizado em um plantio pertencente à
empresa Tietê Agrícola LTDA, localizada no município de Capitão do Poço. Foram
utilizados dados coletados nos anos de 2011, 2012, 2013 e 2014, sempre nos meses de
julho, sendo mensuradas 1125 árvores anualmente. As variáveis dendrométricas
coletadas foram: altura total (Ht) e circunferência a altura do peito (CAP) que
posteriormente foi transformado em diâmetro a altura do peito (DAP) e idade. Foram
ajustados modelos de crescimento em altura dominante, sendo os indivíduos dominantes
selecionados pelo conceito de Asmamm. A seleção do melhor modelo ajustado foi
determinada pelo coeficiente de determinação ajustado (R²aj.%), erro padrão residual da
estimativa em percentagem (Syx%), erro padrão residual recalculado (Syxr%),
diagnóstico da distribuição dos resíduos em percentagem e do desvio médio percentual
(DMP%). Todos os modelos ajustados apresentaram F significativo, a 95% de
probabilidade (p < 0,05), evidenciando a existência de regressão, ou seja, as variáveis
respostas podem ser estimadas através da(s) variável(is) independente(s) dos diferentes
modelos ajustados. Na análise dos seis modelos que envolveram as variáveis altura
dominante e idade, é possível utilizar todos os seis modelos ajustados; porém, o modelo
4, representado pela equação Id²/Hdom= -0,0909 + 0,1997 * Id + 0,03531 Id² foi o modelo
mais recomendado, sendo o mais prático para a estimativa da altura dominante em função
da idade dos indivíduos.
Palavras-chave: plantio. teca. reflorestamento.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal

200
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A espécie Tectona grandis Linn F. vulgarmente conhecida como teca, pertencente
à família Lamiaceae, tem se destacado mundialmente no setor florestal. É uma espécie de
elevado valor comercial, bastante utilizada na construção de móveis finos e na construção
naval, devido sua madeira de alta qualidade, podendo ser comercializada desde os
primeiros desbastes, afirma Pelissari et al. (2014). É uma espécie natural do sul e sudeste
do continente asiático, onde possui uma boa adaptação no Brasil, na qual encontrou
condições consideradas ideais para o seu desenvolvimento (OLIVEIRA, 2003;
BEHLING, 2009).
Segundo o Relatório da Indústria Brasileira de Árvores, “IBÁ” (2015), a espécie
T. grandis apresentou um crescimento em área plantada de 22.059 hectares durante os
anos de 2010 a 2014, demostrando a crescente expansão e importância do cultivo de tal
espécie nacionalmente, onde atualmente possui 87.499 hectares plantados. Possivelmente
o valor real de áreas plantadas seja superior pelo fato dos relatórios serem realizado com
dados apresentados por empresas vinculadas ao IBÁ, onde uma parcela não é
contabilizada.
A Associação Brasileira de Produtores de Florestas Plantadas “ABRAF” (2013),
atesta que Brasil cultiva teca em escala comercial nos estados do Mato Grosso,
Amazonas, Acre, Roraima e Pará, sendo o estado da região centro-oeste o maior produtor
do país desta espécie.
Para Tonini et al. (2009), apesar de existirem diversos estudos sobre a espécie no
mundo, estes ainda são escassos no Brasil. As práticas de manejo adotadas em plantios
de teca no Brasil, geralmente são baseadas em conhecimentos insuficientes e inadequados
(PELISSARI et al., 2014). Por este fato, devido à crescente implantação de povoamentos
com teca, em especial na Região Norte, torna-se imprescindível o conhecimento do
crescimento e produção desta espécie em cada sítio florestal.
Sendo assim, a predição do crescimento de povoamentos é importantíssima para
o gerenciamento florestal, sendo necessárias para compreender os processos de
crescimento da floresta, uma vez que é aconselhável que manejo seja determinado em sua
maioria pela prognose de produções futuras a partir de informações correntes,
favorecendo-se assim, o planejamento e rentabilidade do empreendimento. Os modelos
de regressão devidamente ajustados estimam o crescimento a partir de atributos em nível

201
Belém (PA), 11 a 13 de dezembro de 2019
de povoamento, como idade, área basal e índice de local, de acordo com Campos & Leite,
(2006), o que justifica tal estudo.
Dentre as variáveis dendrométricas, a altura dominante geralmente independe do
espaçamento adotado no plantio e é menos afetada pela manipulação da densidade, a não
ser em casos extremos. Os indivíduos com altura dominante possuem maior probabilidade
de permanecerem durante toda a rotação da cultura, possibilitando-se assim, análises
continuas dos mesmos. Por outro lado, as variáveis diâmetro, área basal e volume são
bastante influenciadas por fatores externos, onde esta última, é a mais utilizada em se
tratando de predição de crescimento e produção (SCOLFORO, 1994).
Além do fato da altura dominante sofrer menos influência dos tratamentos
silviculturais, esta medida de crescimento, demostra uma satisfatória correlação com os
fatores ambientais inter-relacionados, assim com a produção total em volume, além de
ser considerado o índice de sítio de um local. Em comparação com o volume, a altura
dominante é mais facilmente obtida, além de proporcionar maior praticidade na aplicação
(MENEGOL, 1991).
Este trabalho teve por objetivo ajustar e selecionar o modelo que melhor expressar
a tendência de crescimento de um plantio T. grandis, localizado no município de Capitão
Poço, PA.
2. MATERIAL E MÉTODOS
O presente estudo será realizado na Fazenda São Luiz, pertencente à empresa
Tietê Agrícola LTDA, localizada no município de Capitão do Poço, na margem esquerda
da rodovia BR – 010, no sentido Belém – Paragominas, pertencendo a mesorregião do
nordeste paraense e microrregião do Guamá.
2.1 ÁREA DE ESTUDO
O plantio clonal foi realizado no ano de 2010, onde possui uma área de
aproximadamente 54 hectares, com espaçamento de 4 m x 4 m entre plantas, perfazendo
625 indivíduos/ha. Para monitoramento do plantio foram implantadas 5 parcelas
permanentes quadradas com área de 3600 m2 cada, com processo de amostragem aleatório
simples.
De acordo com a classificação climática de Köppen, a região da bacia do Guajará
está enquadrada no clima tipo Am. Este tipo define um clima quente e úmido clima

202
Belém (PA), 11 a 13 de dezembro de 2019
tropical chuvoso com curta estação seca (BRASIL, 2009). O subclima seco associa-se ao
regime anual das precipitações e corresponde ao inverno seco, com precipitação média
do mês mais seco inferior a 60 mm.
As áreas que compõem a vegetação da Fazenda São Luiz pertencem à região da
floresta densa, sub-região dos altos platôs do Pará Maranhão e estão constituídas de
floresta densa dos terraços e secundária latifoliada. Na região Amazônica a denominação
Floresta Densa pode ser considerada sinônimo de floresta ombrófila tropical (BRASIL,
1973).
Na região existe uma predominância de Latossolos Amarelos e Plintossolos
(BRASIL, 1973, VIEIRA, 1988). Na área da fazenda, existem duas classes predominante
de solos, classificado conforme o Sistema Brasileiro de Classificação de Solos, conforme
Santos et al. (2013), como Latossolo Amarelo Distrófico Petroplíntico (LAd), Latossolo
Amarelo Distrófico Típico (lad) e Plintossolo Pétrico Concrecionário (FFc).
O relevo plano a suave ondulado, sendo que poucas áreas podem ser classificadas
como de relevo ondulado, ocupadas apenas com pastagem, onde as declividades são
identificadas como suave, e suave ondulado chegando a próxima de 5% (FOLONI et al.,
2010).
2.2 COLETA DE DADOS
Foram utilizados os dados coletados nos anos de 2011, 2012, 2013 e 2014, sempre
nos meses de julho, sendo mensuradas 1125 árvores anualmente. As variáveis coletadas
foram: Altura Total (Ht), Circunferência a Altura do Peito (CAP) que posteriormente foi
transformado em Diâmetro a Altura do Peito (DAP) para identificar os indivíduos
dominantes e idade (Id).
Para cada ano foi calculada a Altura dominante (Hd) por parcela através da média
das alturas dos 36 indivíduos mais grossos presentes em 3600 m², metodologia esta
adotada seguindo o conceito de Assman (MACHADO & FIGUEIREDO FILHO, 2014).
Os instrumentos necessários na coleta de dados são: Clinômetro Haglof para a
Altura Total (Ht) e dominante (Hd), fita métrica para o CAP, trena de 30 m, GPS
(Garmim), ficha de campo, lápis e borracha.
ANÁLISE DE DADOS

203
Belém (PA), 11 a 13 de dezembro de 2019
Foi realizada uma análise quantitativa dos dados coletados em campo. Serão
ajustados modelo de crescimento envolvendo dados de Hd em função da Idade (Id).
Utilizando-se o software Microsoft Excel 2013, serão realizados os ajustes dos modelos
de crescimento, geração da tabela resumo com as medidas de precisão, geração de
gráficos da distribuição dos resíduos e estimar os valores de altura dominante (Hd) para
os anos posteriores do plantio.
2.3 MODELOS MATEMÁTICOS DE REGRESSÃO
Os modelos de crescimento selecionados para ajuste são compostos pela Altura
dominante (Hd) como sendo a variável dependente ou de interesse, e a Idade (Id) com
suas transformações matemáticas, como variáveis independentes. Foram ajustados e
avaliados 1 modelo linear e 5 modelos linearizáveis de crescimento, como mostrados na
Tabela abaixo.
Tabela 1 - Modelos de crescimento selecionados para ajuste. Onde: Hd= Altura
Dominante; βi= Parâmetros da regressão; Id= Idade; εi= erro.
Nº MODELOS
1 Hd= β0 + β1 Id + β2 Id² + β3 Id³ + εi
2 Hd = β0 Id β1 + εi
3 Hd = β0 e β1 Id + εi
4 Hd = Id ²/(β0 + β1 Id + β2 Id2 ) + εi
5 Hd = e (β0 + β1 1/Id) + εi
6 Hd = β0 e (β1 Ln Id + β2 (Ln Id)²) + εi
Fonte: Os autores, 2019.
2.3.1 Critérios de seleção dos modelos ajustados
Após os ajustes dos modelos de regressão pelo método dos mínimos quadrados,
foi gerada a análise de variância (ANOVA) obtendo-se o valor do Teste F a 95% de
probabilidade, com o intuito de verificar a existência de regressão entre a variável
resposta e a(s) variável(eis) independente(s) de cada modelo. A seleção do melhor modelo
foi feita através da análise dos seguintes critérios estatísticos: o maior coeficiente de
determinação ajustado (R² aj.%); o menor erro padrão residual da estimativa em
percentagem (Syx %) e recalculado (Syxr %); e diagnóstico da distribuição dos resíduos
através da análise gráfica e Desvio Médio Percentual (DMP%).

204
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
Na tabela 2 são apresentados os seis modelos ajustados aos dados do plantio clonal
com as suas respectivas medidas de precisão.
Tabela 2 - Modelos ajustados aos dados do plantio clonal de T. grandis.
Nº MODELOS R²aj % Syx% Syxr% DMP%
1 Hd= b0 + b1 Id + b2 Id² + b3 Id³ 92,93 6,14 - -0,36
2 Ln Hd= Ln b0 + b1 Ln Id 93,09 - 5,99 -0,19
3 Ln Hd= Ln b0 + b1 Id 90,31 - 7,02 -0,26
4 Id²/Hd= b0 + b1 Id + b2 Id² 99,18 - 6,35 -0,34
5 Ln Hd= b0 + b1 1/Id 89,15 - 8,02 -0,29
6 Ln Hd= Ln b0 + b1 Ln Id + b2 (Ln Id)² 92,74 - 6,12 -0,18
Fonte: Os autores, 2019.
Todos os modelos ajustados apresentaram F significativo, a 95% de probabilidade
(p < 0,05), evidenciando a existência de regressão, ou seja, as variáveis respostas podem
ser estimadas através da(s) variável(is) independente(s) dos diferentes modelos ajustados.
As análises estatísticas dos modelos foram as seguintes:
• Coeficiente de determinação (R²aj.) dos seis modelos matemáticos, variou de
89,15% à 99,18%;
• Erro padrão da estimativa (Syx) variando de 5,99 % à 8,02 %;
• Desvio médio percentual (DMP) variando de -0,36 à -0,19%;
• As distribuições gráficas dos resíduos mostraram-se não tendenciosas para os seis
modelos ajustados.
Em relação aos coeficientes de determinação, destacaram-se os modelos 2 e 4 (,.
O melhor resultado mostra que 99,17% de todas as variações que ocorrem na variável
dependente são de responsabilidade deste modelo.
Quanto ao erro padrão da estimativa em percentagem (Syx%) e o recalculado
(Syxr%), percebe-se que todos os modelos apresentaram valores inferiores a 10%, sendo
o modelo 2 com melhor resultado de Syxr . Portanto, verificou-se que os valores de altura
dominante estimados através do modelo 2 oscilam, em média, na faixa de 5,99% em

205
Belém (PA), 11 a 13 de dezembro de 2019
relação aos valores reais de altura dominante, denotando que o plantio clonal de T.
grandis aos quatro anos de idade, está se desenvolvendo dentro de um limite adequado
de homogeneidade. Sendo assim, as diferenças na estimativa da variável altura total
tendem a serem menores.
Para AHRENS (1982), a análise visual dos gráficos da distribuição dos resíduos é
de natureza subjetiva, embora em algumas verificações, seja considerável para uma
adequada avaliação. Por este motivo, além da análise visual dos gráficos analisou-se
também o Desvio Médio Percentual (DMP%) para verificar a existência de
tendenciosidade nas estimativas das alturas dominantes das árvores do plantio clonal de
teca.
Analisando-se numericamente, é possível afirmar que o modelo 6 foi o que
apresentou melhor distribuição residual, havendo uma subestimativa, sendo que em
média, 0,18% das alturas dominantes são estimadas abaixo dos valores reais.
Na comparação e escolha do(s) melhore(s) modelo(s) para estimativa da(s)
altura(s) dominante(s) das árvores do presente estudo, verificou-se que através das
medidas de precisão (R²aj., Sxy%, DMP% e Distribuição dos Resíduos) todas as equações
analisadas apresentaram bom ajustamento aos dados do plantio clonal de Tectona grandis
Linn F., sendo que, através da hierarquização seria indicado o modelo 2, seguido pelos
modelos 6, 4 e 1, respectivamente.
Em relação ao erro padrão da estimativa em percentagem os quatro modelos
apresentam valores em torno de 6%; em relação ao desvio médio percentual as diferenças
são pequenas entre os valores, sendo todos menores do que 0,36%, em relação a análise
gráfica da distribuição dos resíduos. Verificou-se a não tendenciosidade para os modelos
ajustados; o coeficiente de determinação para o modelo 4 foi bastante superior aos
demais; e em relação a facilidade de aplicação pelo usuário final, os modelos que não
envolvem transformações logarítmicas são mais simples para a sua utilização.
Portanto, apesar do resultado indicado pela hierarquização, recomenda-se uso do
modelo 4 para estimativa da variável altura dominante em função da idade do plantio
clonal de T. grandis aos 4 anos de idade

206
Belém (PA), 11 a 13 de dezembro de 2019
4. CONCLUSÃO
Na análise dos seis modelos que envolveram as variáveis altura dominante e idade,
é possível utilizar todos os seis modelos ajustados. Porém o modelo 4, representado pela
equação Id²/Hd= -0,0909 + 0,1997 * Id + 0,03531 Id² é o modelo mais recomendado por
ser também o mais prático para a estimativa da altura dominante em função da idade dos
indivíduos do povoamento de T. grandis localizado no município de Capitão Poço, PA.

207
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ABRAF – Associação Brasileira de Produtores de Florestas Plantadas. Anuário
Estatístico da ABRAF 2013, ano base 2012. Brasília: ABRAF, 2013. 55 p.
AHRES, S. Importância da distribuição de resíduos de regressão na seleção de
equações de volume. Documentos URPFC Nº 10. Contribuição da URPFC ao 4º
Congresso Florestal Brasileiro. Curitiba-PR: Embrapa, julho,1982. p. 7-19.
BEHLING, M. Nutrição, partição de biomassa e crescimento de povoamentos de teca
em Tangará da Serra-MT. 2009. 176 f. Tese (Doutorado em Solos e Nutrição de
Plantas) – Universidade Federal de Viçosa, Viçosa – MG.
BRASIL. Ministério de Minas e Energia. Departamento de Produção Mineral. Projeto
RADAM. Folha SA. 23 São Luiz e parte da folha SA. 24 Fortaleza; geologia;
geomorfologia, solos, vegetação e uso potencial da terra. Rio de Janeiro, 1973.
BRASIL. Sistema de Proteção da Amazônia. SIPAM. Centro Regional de Belém. CR-
BE. Zoneamento Climático. Relatório Preliminar. Zoneamento Ecológico Econômico
do Pará. Rio de Janeiro, 2009.
FOLONI, José Marcos; HACK, Edson; TZIBOY, Edgar. Levantamento da capacidade
de uso do solo das Fazendas São Luiz e Veneza para fins silviculturais da Teca
(Tectona Grandis L.f.). Solo Engenharia Agroflorestal. Cuiabá, MT, 2010.
MACHADO, S. A.; FIGUEIREDO FILHO, A. Dendrometria. Guarapuava,
UNICENTRO, 2. ed., p. 316, 2014.
MENEGOL, O. Índice de sítio e relação entre altura dominante e teores nutricionais
das acículas em povoamentos de Pinus elliottii var. elliottii no segundo planalto
paranaense. 74 f. Dissertação (Mestrado em Ciências Florestais) - Setor de Ciências
Agrárias, Universidade Federal do Paraná, Curitiba, 1991.
MONTEIRO, L. A. da S. Equações de volume geral e para dez espécies dominantes
em uma floresta primária no sudeste do Pará. 2009. 65 f. Dissertação (Mestrado em
Ciências Florestais) – Universidade Federal Rural da Amazônia, Belém, PA, 2009.
OLIVEIRA, J. R. V. de. Sistema para cálculo de balanço nutricional e recomendação
de calagem e adubação de povoamentos de teca – Nutriteca. 2003. 89 f. Dissertação
(Mestrado em Solos e Nutrição de Plantas) – Universidade Federal de Viçosa, Viçosa –
MG.
PELISSARI, A. LI.; GUIMARÃES, P. P.; BEHLING, A.; EBLING, Â. A. Cultivo da
teca: Características da espécie para implantação e condução de povoamentos florestais.
Agrarian Academy, Centro Científico Conhecer – Goiânia, v.1, n.01; p.128, 2014.
RODOVANSKI, E. Modelos estatísticos para previsão de crescimento de plantações
florestais. Curitiba, 2003. Dissertação (Mestrado), Universidade Federal do Paraná.
SANTOS, H. G.; JACOMINE, P. K. T.; ANJOS, L. H. C.; OLIVEIRA, V. A.;
LUMBRERAS, J. F.; COELHO, M. R.; ALMEIDA, J. A.; CUNHA, T. J. F.; OLIVEIRA,
J. B. Sistema Brasileiro de Classificação de Solos. Embrapa, 353 p., 2013.

208
Belém (PA), 11 a 13 de dezembro de 2019
SCOLFORO, José Roberto. Modelos para expressar o crescimento e a produção
florestal: Parte 1. Lavras, ESAL/FAEPE, 1994.
SILVA, E. N. da. Estimação de modelos volumétricos para espécies florestais de uso
comercial em Paragominas-Pa. 2011. 66 f. Dissertação (Mestrado em Ciências
Florestais) - Universidade Federal Rural da Amazônia, Belém, 2011.
THOMAS, C.; ANDRADE. C. M.; SCHNEIDER. P. R.; FINGER. C. A. G. Comparação
de equações volumétricas ajustadas com os dados de cubagem e analise de tronco.
Revista Ciência Florestal, Santa Maria, v. 16, n. 3, p. 319-327, 2006.
TONINI, H.; COSTA, M. C. G. C.; SCHWENGBER, L. A. M. Crescimento da teca
(Tectona grandis) em reflorestamento na Amazônia Setentrional. Pesquisa Florestal
Brasileira, n. 59, p. 05–14, 2009.
VIEIRA, L. S. Manual de Ciência do solo com ênfase aos solos tropicais. São Paulo:
Ed. Agronômica Ceres, 1988. 464p.

209
Belém (PA), 11 a 13 de dezembro de 2019
IDENTIFICAÇÃO BOTÂNICA EM PLANOS DE MANEJO FLORESTAL
SUSTENTÁVEL: O CASO DO IPÊ
Rivaldo Costa Cardoso Junior1; Wheriton Fernando Moreira da Silva2; Rafael Lima
Araújo Ferreira³; Karina Sampaio Martins de Souza4; Jessica Cristina Oliveira Maciel5
1M.Sc. Botânica Tropical. Secretaria de Estado de Meio Ambiente e Sustentabilidade -
SEMAS. [email protected] 2M.Sc. Em Ciências de Florestas Tropicais. Secretaria de Estado de Meio Ambiente e
Sustentabilidade – SEMAS. [email protected]
³Especialista em Educação Ambiental. Secretaria de Estado de Meio Ambiente e
Sustentabilidade - SEMAS. [email protected] 4Graduada em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected] 5Especialista em Georreferenciamento, Geoprocessamento e Sensoriamento Remoto.
Secretaria de Estado de Meio Ambiente e Sustentabilidade – SEMAS.
RESUMO
Tabebuia é muito conhecido pelo setor florestal, pois apresentava todas as espécies
comumente identificadas como ipês. Recentemente, através de estudos filogenéticos,
dividiram Tabebuia em três gêneros: Tabebuia, Handroanthus e Roseodendron. Com a
confirmação, os principais ipês que ocorrem na Amazônia brasileira passaram a serem
identificados com o táxon Handroanthus. As Autorizações de Exploração Florestal –
AUTEF, as quais são emitidas pela Secretaria de Estado de Meio Ambiente e
Sustentabilidade – SEMAS, na maioria das vezes apresenta apenas uma espécie de “ipê”
para a exploração madeireira. Sem ferramentas adequadas e técnicas especializadas, a
SEMAS fica incapacitada de monitorar e fiscalizar a ocorrência e identificação correta
dessas espécies. Nesse sentido, este trabalho objetivou realizar revisão das AUTEF
emitidas no período de 2017 a 2019 e verificar a identificação de ipês, com base no banco
de dados do Sistema de Monitoramento e Licenciamento Ambiental – SIMLAM, além de
fazer levantamento do gênero para a região do oeste do Pará, em principal nos municípios
de Santarém, Óbidos, Juruti, Alenquer, Prainha, Mojuí dos Campos, Monte Alegre e
Oriximiná. Além disso, verificar o nome científico mais usado nas autorizações. Esta
pesquisa foi realizada no escritório da SEMAS.Quantificou-se 53 AUTEF e dentre essas,
51 autorizações foram apresentadas com uma única espécie para os ipês nas áreas dos
Planos de Manejo Florestal Sustentável – PMFS. O nome científico mais utilizado foi
Tabebuia serratifolia. O município com maior número de AUTEF emitidas ede
indivíduos de ipês identificadosfoi Juruti, com 86% apresentando uma única espécie.
Portanto, métodos empíricos na identificação de inventários florestais podem ocasionar
agrupamentos de espécies em PMFS. Isso pode acarretar perdas irreparáveis na
manutenção das espécies e com isso, medidas mais cautelares devem ser tomadas pela
SEMAS para tentar diminuir a perda de diversidade em floresta manejada.
Palavras-chave: Identificação. Manejo. Ocorrência.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal

210
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O gênero Tabebuia, pertencente à Família Bignoniaceae, já sofreu inúmeras
alterações taxonômicas ao longo do tempo. Tabebuia ficou muito conhecido pelo setor
florestal, pois apresentava, originalmente, todas as espécies comumente identificadas
como ipês incluídas no gênero (GROSE & OLMSTEAD, 2007). Entretanto, Mattos
(1970) dividiu as espécies de “ipês” em dois grupos, Tabebuia e Handroanthus, este
criado, especialmente para os ipês brasileiros.
Por sua vez, Gentry (1972, 1992), após forte crítica a Mattos, reposicionou todas
as espécies de “ipês” para Tabebuia. Todavia, recentemente, Grose &Olmstead (2007)
com base em estudos filogenéticos, dividiram Tabebuia em três gêneros: Tabebuia,
Handroanthus e Roseodendron. Isso confirmou em definitivo a segregação proposta por
Mattos (1970) e a existência de linhagens diferentes dentro do grupo (SANTOS, 2017).
Com a confirmação em definitivo, os principais ipês que ocorrem na Amazônia
brasileira, que são também consideradas uma das espécies madeireiras mais
comercializadas no Brasil (MARTINS-DA-SILVA et al, 2014, “SERVIÇO
FLORESTAL BRASILEIRO, 2019”), passaram a serem identificadas corretamente com
táxon a nível genérico, Handroanthus.
Ocorre que as Autorizações de Exploração Florestal – AUTEF, as quais são
emitidas pela Secretaria de Estado de Meio Ambiente e Sustentabilidade – SEMAS –
órgão gestor do Estado do Pará, na maioria das vezes apresenta apenas uma espécie de
“ipê” para a exploração madeireira, com o nome Tabebuia serratifolia(Vahl) G.
Nicholson. Considerando a legislação vigente (RESOLUÇÃO DO CONAMA nº
406/2009; IN/SEMAS nº 05/2015 e IN/MMA nº 05/2006), toda a informação prestada ao
órgão gestor ambiental, neste caso a SEMAS, é de inteira responsabilidade do interessado
pelo empreendimento florestal.
Dessa forma, sem ferramentas adequadas, técnicas especializadas de identificação
botânica e quadro funcional suficiente, a SEMAS fica incapacitada de monitorar e
fiscalizar a ocorrência e identificação correta da espécie “ipê” nos Planos de Manejos
Florestais Sustentáveis – PMFS, que são protocolados no órgão.
Sem o monitoramento e adequação dos PMFS, a identificação dos ipês fica
comprometida, pois, conjectura-se, que nos PMFS há o agrupamento de espécies. Isso
ocorre quando a identificação é feita através de conhecimento empírico, que

211
Belém (PA), 11 a 13 de dezembro de 2019
consequentemente, a identificação passa a ser a nível genérico, ou seja, onde ocorre duas
ou mais espécies do mesmo gênero, é, então, agrupado como uma única espécie.
De acordo com Procópio & Secco (2008), o conhecimento empírico de nativos,
conhecidos como “mateiros”, é um método usual de inventários florestais, nos quais esses
profissionais adotam nomes vernaculares ou populares para determinação das espécies.
Isso pode omitir a verdadeira ocorrência de diferentes espécies de “ipês” na região, o que
afeta a diversidade local dessas espécies.
Diante o exposto, o objetivo deste trabalho é realizar uma revisão das AUTEF e
verificar a identificação das espécies “ipês”, com base no banco de dados do Sistema de
Monitoramento e Licenciamento Ambiental – SIMLAM – da SEMAS. Procedendo-se
com as seguintes etapas: realização de um levantamento do gênero para a região do oeste
do Estado do Pará; verificação do nome científico utilizado nas autorizações; elaboração
de uma lista das espécies de “ipês” autorizadas e verificação de possível agrupamento de
espécies de ocorrência naquela região.
2. MATERIAL E MÉTODOS
Esta pesquisa foi realizada no escritório da SEMAS, localizado na Travessa
LomasValentinas, 2717, Belém, Pará, no qual foi realizado levantamento das AUTEF no
período de 2017 a junho de 2019, no acervo do banco de dados do sistema SIMLAM,
para o oeste do Pará, em principal nos municípios de Santarém, Óbidos, Juruti, Alenquer,
Prainha, Mojuí dos Campos, Monte Alegre e Oriximiná.
O banco de dados do SIMLAM encontra-se com o seu acervo informatizado,
baseado no catálogo de árvores do Brasil – Laboratório de produtos florestais – LPF. O
banco de dados do SIMLAM é adicionado informação de acordo com a necessidade,
principalmente quando do surgimento de novas espécies ou atualizações, tal como a dos
ipês.
As informações foram conferidas em arquivo PDF, as quais foram baixadas do
sistema, consultando, diretamente as AUTEF e procedendo-se as anotações do número
da autorização, nome científico do “ipê” empregado no título emitido, número de
indivíduos autorizados e respectivos municípios de ocorrência. A seguir, essas
informações foram importadas para uma planilha do excel.

212
Belém (PA), 11 a 13 de dezembro de 2019
Posteriormente a filtragem das espécies foi criada uma lista dos municípios
paraenses de ocorrência da espécie “ipê”.
A identificação das espécies do referido gênero foi baseado na classificação
taxonômica do sistema de APG IV (2016).
3. RESULTADOS E DISCUSSÃO
Foram quantificadas 53 Autorizações para Exploração Florestal – AUTEF,
distribuídas nos municípios selecionados no período de 2017 a junho de 2019. Em 51
autorizações foram apresentadas apenas uma única espécie para representar os ipês nas
áreas dos Planos de Manejo Florestal Sustentável – PMFS.
No geral, as autorizações emitidas no período representaram aproximadamente
96% dos ipês identificados como uma única espécie. Apenas duas AUTEF apresentaram
identificação de mais de uma espécie do gênero. Somando-se, nesse caso, 55
identificações.
Considerando todas as autorizações, o nome científico mais utilizado foi Tabebuia
serratifolia(Vahl) Nichols., seguidos por Handroanthus serratifolius (Vahl) S.O. Grose,
Handroanthus impetiginosus(Mart. ex DC.) Mattos e Tabebuia impetiginosa(Mart. ex
DC.) Atandl (Figura 1).
Figura 1 – Relação dos nomes científicos utilizados na identificação botânica nas Autorizações
para Exploração Florestal – AUTEF, e a quantidade de vezes que foram adotadas no período de
2017 a junho de 2019, Pará, Brasil.
Fonte: autores.

213
Belém (PA), 11 a 13 de dezembro de 2019
É válido lembrar que T. serratifoliae T. impetiginosasão sinonímias de H.
serratifolius e H. impetiginosus, respectivamente, e que aquelas não são mais usuais, de
acordo com a nova classificação taxonômica em vigor (APG IV, 2016). Isto é, T.
serratifoliae H. Serratifolius se tratam da mesma espécie, assim como T. impetiginosae
H. impetiginosus. De acordo com esses resultados, constatou-se que apenas duas espécies
têm ocorrência na região dos municípios selecionados.
Dos municípios selecionados, os que apresentaram maior número de AUTEF
emitidas foi Juruti, Santarém e Prainha. Em relação ao município de Juruti, esse
apresentou 86% das autorizações com identificação de uma única espécie. Na Tabela 1,
é possível observar o número de AUTEF por município e seus respectivos usos dos nomes
científicos para os ipês.
Tabela 1 – Quantificação do número de AUTEF emitidas pela SEMAS, por município e seus
respectivos usos dos nomes científicos para os ipês. Espécies
Municípios H.
impetiginosus H.
serratifolius T.
impetiginosa T.
serratifolia Total
geral
Alenquer 1 1
Almeirim 1 1
Juruti 1 2 14 17
Mojuí dos
Campos 1 2 3
Óbidos 1 2 4 7
Oriximiná 1 1
Prainha 4 5 9
Santarém 2 4 10 16
Total geral 3 12 2 38 55
Fonte: autoes.
Com relação ao número de indivíduos de ipês identificados, os quais foram
apresentados nas autorizações, o município com maior destaque foi Juruti. Na figura 2,
tem-se a relação dos municípios e seus respectivos números de indivíduos por espécie de
ipê.

214
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2 – Relação do número de indivíduos de ipês e suas respectivas identificações
botânicas, por municípios, de acordo com a análise das AUTEF emitidas pela SEMAS, Pará,
Brasil.
Fonte: autores.
Com relação à identificação botânica dos ipês no que tange a ocorrência da espécie
para o Estado do Pará, em principal, os municípios selecionados, a verificação de apenas
duas espécies, era o resultado esperado, pois a SEMAS não dispõe de profissional
habilitado e nem ferramentas adequadas para monitorar as informações referentes à
identificação correta das espécies a serem manejadas.
Segundo o sítio reflora.jbrj.gov.br (2019), são ocorrentes no Estado do Pará,
considerando o gênero Handroanthus, as seguintes espécies de “ipês”, H. barbatus (E.
Mey) Mattos, H. capitatus (Bureau e K. Schum.) Mattos, H. impetiginosus (Mart. ex DC.)
Mattos, H. incanus (A.H. Gentry) S. Grose e H. serratifolius (Vahl.) S. Grose. Em relação
ao sítio splink.org.br (2019), além dos citados acima, apresentou a espécie H. ochraceus
(Cham.) Mattos como também de ocorrência no Estado.
É importante informar que o sítio splink.org.br disponibiliza dados de herbários
do mundo inteiro, com isso foi possível filtrar as espécies do gênero Handroanthus para
o Estado do Pará e assim classificá-los de acordo com a ocorrência nos municípios
selecionados neste estudo. Dessa forma, verificaram-se as seguintes espécies de “ipês”
ocorrentes e seus respectivos municípios, tais quais: H. barbatus, H. ochraceus, H.
impetiginosus e H. serratifolius, de ocorrência no município de Santarém; H. ochraceus
(Monte Alegre); H. serratifolius (Oriximiná); e H. capitatusem Almeirim.
Dados como os apresentados pelos sítios reflora e splink revelam a ocorrência de
várias espécies para o gênero Handroanthus na região. Nesse sentido, pode-se levantar a

215
Belém (PA), 11 a 13 de dezembro de 2019
hipótese de possíveis agrupamentos de espécies de “ipês” em PMFS. Isto é, a forte
possibilidade de existir indivíduos de outras espécies de “ipês”, os quais ficam omissos
devido ao método de identificação utilizado.
Procópio & Secco (2008), constataram agrupamentos de espécies para indivíduos
de “tauarí” em duas florestas exploradas nos municípios de Paragominas e Belterra, em
2003. Em Belterra, de 112 indivíduos de “tauarí” identificados como Couratari
guianensis, pela empresa que manejou a área, apenas um era C. guianensis, sendo 107 C.
stellatae quatro C. oblongifolia; Enquanto que em Paragominas apresentou 33 indivíduos
identificados como C. guianensis, entretanto, no inventário taxonômico realizado pelos
pesquisadores encontraram o agrupamento de duas espécies: C. guianensise C.
oblongifolia.
Estudos como o de Procópio & Secco podem corroborar com a idéia de que,
provavelmente, possa existir a ocorrência de mais de uma espécie de “ipês” nos PMFS
protocolizados na SEMAS. Dessa maneira, pode-se inferir novamente que aqueles
resultados podem refletir num possível agrupamento de espécies de “ipês” nos PMFS
liberados pelo órgão gestor.
De acordo com Martins-da-Silva et al (2003), a obtenção da nomenclatura
científica requer metodologia específica, a fim de minimizar erros. A identificação
botânica correta é um dos pré-requisitos para que se tenha êxito na implantação de um
plano de manejo florestal, sendo fundamental que se conheçam as espécies utilizadas, a
fim de se planejar o seu uso, de forma a garantir a continuidade das espécies.
Para ter entendimento da conseqüência da identificação incorreta nos inventários
florestais, de acordo com a legislação, é assegurado a manutenção do número de
indivíduos arbóreos selecionados para colheita florestal (corte), chamado de índice
manutenção, que é de 10% e/ou 3 indivíduos a cada 100 ha de área licenciada para
exploração, por espécie.
Considerando um Manejo Florestal ideal com duas espécies que pertencem ao
mesmo gênero, A e B. A primeira possui dez indivíduos, a B três. Aplicando apenas os
10% do índice de manutenção, tem-se a seguinte relação: espécie A = 9 indivíduos para
corte e 1 para remanescente; espécie B = 0 para corte, pois não existe 0,3 indivíduos e 3
remanescentes. Isso significa que no final será explorado apenas 9 indivíduos da espécie
A. Mantendo em pé na floresta, como manutenção, quatro árvores das espécies A e B.

216
Belém (PA), 11 a 13 de dezembro de 2019
No caso do agrupamento das espécies, onde A+B = 13 indivíduos, aplicando-se
apenas os 10%, ter-se-ia 11 indivíduos para corte e 2 remanescentes. Issoprovavelmente
poderá refletir diretamente na estrutura da floresta, podendo ter um impacto local na
dispersão de sementes das espécies que não tiveram indivíduos remanescentes.
No caso da hipótese em questão, a figura 2, apresenta um número gigantesco de
indivíduos autorizados para uma única espécie no período abordado, podendo
possivelmente, ter comprometido a floresta quanto à perpetuação da espécie e
consequentemente na continuidade dos aspectos ecológicos que determinadas espécies
podem oferecer para se ter equilíbrio ambiental nos ecossistemas florestais.
4. CONSIDERAÇÕES FINAIS
O uso de métodos empíricos na identificação de inventários florestais pode
ocasionar agrupamentos de espécies em PMFS. O agrupamento pode acarretar perdas
irreparáveis quanto a manutenção das espécies nas florestas.
Medidas mais cautelares para tentar monitorar as florestas manejadas devem ser
realizadas pela SEMAS, apesar das dificuldades do órgão. Isso é importante para tentar
diminuir a perda de diversidade em decorrência do agrupamento, não só das espécies de
“ipês”, mas de outras espécies potencialmente comerciais de ocorrência no Estado do
Pará.

217
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BRASIL, Resolução do CONAMA nº 406, de 02 de fevereiro de 2009. Estabelece
parâmetros técnicos a serem adotadas na elaboração, apresentação, avaliação técnica e
execução de Plano de Manejo Florestal Sustentável - PMFS com fins madeireiros, para
florestas nativas e suas formas de sucessão no bioma Amazônia. Diário Oficial da União,
Brasília, n. 26, fev. 2009.
BRASIL, Instrução Normativa nº 05, de 11 de dezembro de 2006. Dispõe sobre
procedimentos técnicos para elaboração, apresentação, execução e avaliação técnica de
Planos de Manejo Florestal Sustentável - PMFS nas florestas primitivas e suas formas de
sucessão na Amazônia Legal, e dá outras providências. Diário Oficial da União, Brasília,
n. 238, dez. 2006.
BYNG, J.W. et al. An update of the Angiosperm Phylogeny Group classification for the
orders and families of flowering plants: APG IV. Botanical Journal of the Linnean
Society, n. 181, p. 1-20, 2016.
GENTRY, A.H. Handroanthus (Bignoniaceae): a critique. Taxon, n. 21, p. 113-114,
1972.
GENTRY, A.H. Bignoniaceae. Part II (Tribe Tecome). Flora Neotropica, n. 25, p. 1-
370, 1992.
GROSE, S.O.; OLMSTEAD, R.G. Taxonomic revision in the polyphyletic genus
Tabebuias.l. (Bignoniaceae). SystematicBotany, v. 32, n. 3, p. 660-670, 2007.
MARTINS-DA-SILVA, R.C.V.; HOPKINS, M.G.; THOMPSON, I.S. Identificação
Botânica na Amazônia: Situação Atual e Perspectivas. Embrapa Amazônia Oriental,
Belém, Documento 168, jun. 2003.
MARTINS-DA-SILVA, R.C.V.; SILVA, A.S.L.; FERNANDES, M.M.; MARGALHO,
L.F. Noções morfológicas e taxonômicas para a identificação botânica. Embrapa,
Brasília. 2014.
MATTOS, J.R. Handroanthus, um novo gênero para os “ipês” do Brasil. Loefgrenia, n.
50, p. 1-4, 1970.
PARÁ, Instrução Normativa nº 05, de 10 de setembro de 2015. Dispõe sobre
procedimentos técnicos para elaboração, apresentação, execução e avaliação técnica de
Planos de Manejo Florestal Sustentável - PMFS nas florestas nativas exploradas ou não
e suas formas de sucessão no Estado do Pará, e dá outras providências. Diário Oficial do
Estado, Belém, n. 32969, p. 37-57, set. 2015.
PROCÓPIO, L.C.; SECCO, R. de S. A importância da identificação botânica nos
inventários florestais: o exemplo do “tauari” (Couratari spp. e Cariniana spp. –
Lecythidaceae) em duas áreas manejadas no Estado do Pará. Acta Amazonica, v. 38, n.
1, p. 31-44, 2008.

218
Belém (PA), 11 a 13 de dezembro de 2019
REFLORA – Plantas do Brasil: Resgate Histórico e Herbário Virtual para o
Conhecimento e Conservação da Flora Brasileira. Disponível em:
<HTTP://reflora.jbrj.gov.br/reflora/listaBrasil/>acesso em: 20 de set. 2019.
SANTOS, S.R. DOS, A atual classificação do antigo gênero Tabebuia (Bignoniaceae),
sob o ponto de vista da anatomia da madeira. Balduinia, n. 58, p. 10-21, julho. 2017.
SERVIÇO FLORESTAL BRASILEIRO. Espécies Florestais – Sistema Nacional de
Informações Florestais. Disponível em: <HTTP://www.florestal.gov.br/snif/recursos-
florestais/especies-florestais> acesso em: 20 set. 2019.
SPECIES LINK. Disponível em: <HTTP://splink.org.br> acesso em: 24 de set. 2019.

219
Belém (PA), 11 a 13 de dezembro de 2019
PRODUÇÃO DE BIOMASSA FRESCA ACIMA DO SOLO EM UMA ÁREA DE
PMFS LICENCIADO NO MUNICÍPIO DE SANTARÉM - PA
Wheriton Fernando Moreira da Silva¹; Rafael Lima Araújo Ferreira²; Rivaldo Costa
Cardoso Junior3; Karina Sampaio Martins de Sousa4; Jéssica Cristina Oliveira Maciel5
1Mestre em Ciências de Florestas Tropicais. Secretaria de Meio Ambiente e
Sustentabilidade do Pará. [email protected]
2 Especialista em Educação Ambiental. Secretaria de Meio Ambiente e Sustentabilidade
do Pará. [email protected]
3 Mestre em Botânica Tropical. Secretaria de Meio Ambiente e Sustentabilidade do
Pará. [email protected]
4 Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
5 Engenheira Florestal. Secretaria de Meio Ambiente e Sustentabilidade do Pará.
RESUMO
O setor florestal é um dos que mais gera renda no estado do Pará, principalmente pelo
grande potêncial produtivo da floresta Amazônica. Com isto, o manejo florestal é a
ferramenta mais viável para a mitigação dos processos degradativos da exploração
madeireira. O produto madeira é inerente ao que chamamos de biomassa, este por
conseguinte apresentando uma relação direta em sua composição com o elemento
carbono, este sendo liberado após a derrubada de árvores. Tais medidas de quantificação
do Carbono são importantes na discussão das mudanças climáticas, e saber o quantitativo
da biomassa é a base de todo estudo relacionado. Neste enfoque o trabalhou objetivou
quantificar a biomassa fresca com o uso de uma equação global para uma área de manejo
florestal sustentável no oeste do estado do Pará. Usou-se as informações do inventário
florestal 100%, de um empreendimento localizado em Santarém, que passou pelo
processo de licenciamento florestal sendo deferido pelo orgão responsável. Por fim,
detectou-se uma alto número de espécies na área, sendo a base da identificação o apelo
comércial destas, a floresta apresentou características estruturais de uma floresta em
equlíbrio populacional e com sua distribuição normal. Por se tratar de uma floresta densa,
os valores de biomassa fresca foram considerados elevados e dentro do esperado para o
garnde porte das árvores, dando destaque para a biomassa das árvores exploradas, que foi
aproximadamente 30 % do valor da comunidade árborea inventariáda. O trabalho
quantificou a biomassa fresca de forma satisfatória para a propriedade, e dentro dos erros
amostrais existentes, dessa forma contribuindo para o aprofundamento de estudos
envolvendo modelos alométricos globais e a quantificação de biomassa fresca a cima do
solo.
Palavras-chave: FlorestasTropicais. Manejo Florestal Sustentável. Inventário Florestal
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal

220
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Uma das maiores reservas de biodiversidade do mundo, a maior floresta tropical
continua e maior fonte de recursos naturais do mundo, a Amazônia é o centro das
atenções globais no que se refere ao seu uso, proteção e papel no ciclo biogeoquímico
mundial.
O uso do solo Amazônico, assim como os demais usos da floresta vem sendo alvo
de discussões em diversos setores da sociedade, principalmente no que se refere a sua
degradação ambiental. Fearnside (2013) enfatiza que a forma tradicional de
implementação de pequenas atividades agrícolas com queimadas, o alto apelo ao
empreendimento pecuarísta e a exploração madeireira de forma não sustentável são as
principais formas de alteração do cenário natural e de grande potencial degradatório, e
muito comuns na região.
As alterações ocorrentes na florestas agem diretamente no balanço de biomassa
presente, sendo a biomassa (massa biológica) conceituada como o conjunto da matéria
orgânica animal ou vegetal disposta em um ambiente natural. Essa massa apresenta uma
relação direta com a quantidade de carbono (C) presente na constituição da matéria, e
com essa relação (biomassa - carbono), quantidade de biomassa e variáveis como taxa de
desmatamento, é possível estimar o percentual/quantidade de C emitido para a atmosfera
em formato de CO2 (HIGUCHI e CARVALHO JR, 1995; SILVEIRA, 2008).
Pela elevada quantidade de biomassa presente em sua formação, a Amazônia é
considerada o maior sumidouro de carbono do mundo, o mau uso de sua cobertura vegetal
pode afetar diretamente nas emissões de CO2 locais, regionais e global e assim interferindo
nos processos climáticos existentes. Essa perspectiva coloca a floresta tropical
Amazônica no centro das atenções globais no que se refere as mudanças climáticas
globais e uso sustentável de recursos naturais.
Visando o uso sustentável, o manejo florestal surge como uma ferramenta para a
obtenção de produtos madeireiros e não madeireiros de forma organizada, produtiva e de
baixo impacto para o ambiente natural em que é aplicado. Além deste, o manejo inseri
um uso legal e jurídico da propriedade, passando por diversas etapas de licenciamento de
órgãos competentes e responsáveis.

221
Belém (PA), 11 a 13 de dezembro de 2019
Com as premissas descritas, este trabalho objetivou a quantificação estimada da
biomassa fresca acima do solo, de um projeto de manejo florestal sustentável licenciado
no Oeste do Estado do Pará, além de sua relação com o carbono presente na área.
2. MATERIAL E MÉTODOS
Com base na lei nº 12.527/2011 que regulamenta o direito constitucional às
informações públicas, criando mecanismos para que pessoas físicas ou jurídicas possam
receber informações dos órgãos e entidades publicas, sem que haja um motivo
institucional ou oficial, este trabalho usou como banco de dados, informações de um
inventário florestal 100% (IF 100%) protocolado na Secretaria de Meio Ambiente e
Sustentabilidade do Pará (SEMAS), com o objetivo da exploração florestal em regime de
manejo florestal sustentável. Assim, por se tratar de um PMFS empresarial, as
informações referente a empresa serão mantidas em sigilo, por não serem parte
fundamenta do trabalho. Ressalto que estas informações foram solicitadas em nome de
pessoas física, pelo documento nº 2019/40165 protocolado na SEMAS - PA.
2.1. ÁREA DE ESTUDO
Este empreendimento está localizado no município de Santarém, no Oeste do
estado do Pará, Santarém é destaque por ser o principal polo econômico, social e humano
da região, principalmente por ser a 3ºcidade mais populosa do estado e estar
geograficamente inserida as margens do Rio Tapajós e às proximidades do Rio
Amazonas.
O imóvel rural possui uma área total aproximada de 2.348,84 ha, o manejo
florestal foi implementado em toda a propriedade, assim a área de manejo florestal (AMF)
possui o mesmo valor da área total. Históricamente, essa propriedade foi fracionada em
2 UPA’s (unidade de produção anual), sendo a UPA 1 licenciada e explorada no ano de
2018 e a UPA 2 para o ano de 2019. A UPA 1 apresentava uma área de 849,0614 ha, e a
UPA 2 de 680,3172 ha porém para a exploração florestal atual a UPA 2 apresenta a área
de efetivo manejo com 590,7886 ha. No processo de licenciamento, a área de efetivo
manejo é oriunda dos dedutivos referentes a massas d’águas, APP’s, construções e etc.,
sendo incluso apenas a área totalmente coberta por floresta tropical. Por fim, a UPA 2 foi

222
Belém (PA), 11 a 13 de dezembro de 2019
fragmentada em 7 UT’s (unidades de trabalho), a menor escala no planejamento em
planos de manejo. Para fins de estudo, esse trabalho usou apenas os dados referentes a
UPA 2.
2.2.COLETA DE DADOS
Os dados foram obtidos a partir do processo protocolado na SEMAS, para
realização de manejo florestal sustentável empresarial, sendo seu inventário florestal
100% totalmente de responsabilidade do detentor do projeto. Essas informações são parte
inerente do Sistema Integrado de Monitoramento e Licenciamento Ambiental
(SIMLAM), base de informações da secretária. De tal forma, os dados foram obtidos por
equipes de campo profissionais do detentor. Algumas informações obtida: identificação
botânica, valor de CAP/DAP, qualidade do fuste, altura comercial, localização
geográficas e etc.
2.3. ANÁLISE DE DADOS
Como dado primário, usou-se o DAP (diamêtro a altura do solo - 1,30 m) haja
visto que está variável é a principal fonte de inferência em estudos florestais, por ser de
fácil obtenção, arguição e correlação com a estrutura florestal e volumétrica. O inventário
contia árvores com DAP minimo de 30 cm e máximo de 280 cm, estando classificadas
em “Explorar” e “Remanescente”, conforme sua destinação final.
O produto por se tratar de estimativas e baseado em inferências, os dados passaram
por análises de normalidade com o teste paramétrico de Kolmogorov-Smirnov a nível de
significância em 95% (p = 0,05), haja vista a necessidade do uso da estatística descritiva
e paramétrica.
Para a obtenção dos valores de biomassa fresca acima do solo, usou-se a equação
desenvolvida e ajustada por LUCAS et al. (2014) para florestas tropicais úmidas, modelos
gerais desenvolvidos com uma ampla base de dados são alternativas para as estimativas
de biomassa fresca de forma global, desde que as características básicas e os parâmetros
dondrométricos do ambiente sejam observados (CHAVE et al, 2014; LIMA, 2015) vista
que o O modelo alométrico usado está descritos a seguir:

223
Belém (PA), 11 a 13 de dezembro de 2019
BFAS = 0,509 x DAP2 x h - Floresta Tropical úmida (FTU)
Onde:
DAP: Diâmetro a altura do peito em centímetros (1,30m em relação ao solo)
h: Altura total ou comercial em metros
BFAS: Biomassa fresca a cima do solo
3. RESULTADOS E DISCUSSÃO
Com base no censo florestal desenvolvido na área, foram obtidos os dados de
21.282 árvores, estas distribuidas em 126 espécies, com amplitude de DAP variando entre
31 e 281 cm minímo e máximo respectivamente. Para o uso estátístico e de estimativas,
a melhor forma encontrada foi a estratificação em UT’s, já presente no planejamento de
exploração da área de estudo.
Por se tratar de uma área de manejo florestal, os princípios da sustentabilidade
devem estar implícitos em todas as etapas da colheita, principalmente no planejamento.
De tal modo, a distribuição diamétrica é uma das ferramentas para avaliar a estrutura
populacional além do estoque remanescente para um novo ciclo de corte (35 anos). Para
este trabalho, a floresta demonstrou características de uma floresta tropical inequiánea, e
com estoque presente para uma nova exploração (figura 1).
Figura 1- Distribuição diamétrica da comunidade arbórea inventariada com ênfase na destinação
de uso no projeto de PMFS.
Fonte: SILVA et al., 2019

224
Belém (PA), 11 a 13 de dezembro de 2019
Observando a distribuição por classes de tamanho, constatou-se o padrão de uma
exponêncial negativa, o denominado “J-Invertido”. Esse detalhe é observado com maior
ênfase para o grupo das “árvores remanescentes”, assegurando a base do estoque para
uma futura intervensão. Essa informação indica que o projeto tem base inicial para ser
licenciado como um PMFS.
Verificando os dados obtidos no IF 100%, e aplicando à equação alométrica para
FTU, testou-se a normalidade a partir do teste de Kolmogorov-Smirnov, constatando que
os dados relativos a BFAS apresentam distribuição normal (p=0,00000078), sendo
possível o uso de inferências paramétricas.Vale ressaltar que para a obtenção dos valores
de BFAS nesse trabalho foram usados apenas o matéria comercial de cada árvore (fuste).
Para a Comunidade florestal inventariada da área de estudo, foi encontrado um
valor total de biomassa de 69.044,9778 Toneladas de material fresco, sendo seu
representativo em 116,8691 TON. ha-1 (BFAS), esse valor referência a projeção do
material madeira sem que haja a retirada de água, extratos e exçudados. Como valor
médio para cada árvore em: BFAS = 3,2444 TON.
Abordando as destinações de uso, tivemos valores significativos de BFAS
corroborando com a distribuição populacional, e sendo superior no grupo das
remanescentes em comparação com as exploradas, respectivamente 85,3 e 31,5TON. ha-
1, tais valores são alusivos a toda a comunidade inventariada, fazendo a comparação com
o mesmo número de classes de tamanho, o valor das remanescentes cai para 70,7234
TON. ha-1. Esse resultado expressa a base da produção de biomassa da floresta, para que
haja a recuperação das características produtivas e de acumulação de material vegetal, em
consideração ao estado inicial da floresta no período pré-exploratório, Essa é uma das
características à serem observada para inferências num “status de floresta recuperada”
após um ciclo efetivo de exploração florestal.
Ao estimar a massa de BFAS em função das UT’s encontramos os valores médios
para as exploradas de 4,5094 ± 1,80 TON. ha-1 e para as remanescentes de 12,1861 ± 5,13
TON. ha-1,dentre as UT’s constatou-se pouca variação dos valores de BFAS (tabela 1),
haja vista que há uma boa distribuição de árvores de grande porte dentro da AMF.

225
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1 - Tabela com os valores de BFAS em função da Unidade de trabalho em PMFS
UT BFAS Exp. (TON . ha-1) BFAS Rem. (TON . ha-1)
1 4,6097 15,0400
2 6,4241 20,2112
3 6,3086 11,9703
4 6,4981 16,6139
5 2,9677 9,4799
6 1,9106 3,8889
7 2,8473 8,0988
Total geral 31,5660 85,3032
Média 4,5094 12,1862
Desvio Padrão 1,9477 5,5539
Int. de confiança (95%) 1,8013 5,1365
Fonte: SILVA et al. (2019)
Com os dados de inventário florestal do projeto licenciado, confirmou-se o que já
é descrito por diversos autores e de diversas áreas, de que florestas tropicais são
reconhecidas por apresentarem um elevado valor de biomassa estocada. Na exploração
florestal parte dessa biomassa é transformada, deixando uma lacuna na floresta, no
entanto, essa variação deverá ser preenchida através dos processos de regeneração natural
e crescimento da floresta, buscando a estabilidade do ambiente. Quanto menor for o
impacto provocado, mais rápido a floresta tenderá a recuperar suas características naturais
de acumulo de BFAS, sendo otimizado pelo processo inicial de crescimento após a
intervenção.
Nesse patamar as florestas tropicais e principalmente a Amazônia, são
classificados como os principais sumidouros de carbono, além de possuir a maior
quantidade deste elemento estocado. Para as discussões referentes ao clima, o bom uso
dos recursos florestais são vitais para o planejamento de novas medidas de controle e
otimização dos processos produtivos, nisso o manejo florestal deve ser aprimorado para
minimizar emissões, já que o produto madeira, acaba por liberar carbono gradativamente
para a natureza. Quanto maior a área explorada, maior será a perda de biomassa e por
conseguinte o nível de carbono lançado futuramente na atmosfera.

226
Belém (PA), 11 a 13 de dezembro de 2019
4. CONCLUSÃO
O trabalho mostrou um elevado estoque de biomassa florestal a cima do solo, na
área licenciada para o manejo florestal. Por se tratar de um PMFS, o inventário da
comunidade arbórea se assemelhou ao padrão de uma floresta tropical com a distribuição
diamétrica em um “J-invertido”, e quando observou-se a distribuissão de biomassa por
UT’s não se verificou uma discrepância elevada entre elas, hava vista que as populações
da área são próximas em estrutura e características dendrométricas. Para fins de
discussões globais, as estimativas de biomassas são importantes para a quantificação do
carbono e criação de estratégias para minimizar o impacto na atmosfera, assim como
medidas de contenção para diversos problemas ambientais.

227
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
CHAVE, J.; RÉJOU-MÉCHAIN, M.; BÚRQUEZ, A.; CHIDUMAYO, E.; COLGAN,
M. S.; DELITTI, W. B. C.; DUQUE, A.; EID, T.; FEARNSIDE, P. M.; GOODMAN, R.
C.; HENRY, M.; MARTI´NEZ-YRÍZAR, A.; MUGASHA, W. A.; MULLER-
LANDAU, H. C.; MENCUCCINI, M.; NELSON, B. W.; GOMANDA, A. N.;
NOGUEIRA, E. M.; ORTIZ-MALAVASSI, E.; PÉLISSIER, R.; PLOTON, P.; RYAN,
C. M.; SALDARRIAGA, J. G.; VIEILLEDENT, G. Improved allometric models to
estimate the aboveground biomass of tropical trees. Global Change Biology v.20, p.
3177–3190, 2014.
FEARNSIDE, P. M.; BARBOSA, R. I.; PEREIRA, V. B. Emissões de gases do efeito
estufa por desmatamento e incêndios florestais em Roraima: fontes e sumidouros.
Agroambiente On-line, [S.I.], v. 7, n. 1, p. 95-111. 2013.
HIGUCHI, N.; CARVALHO JÚNIOR, J. A. Fitomassa e conteúdo de carbono de
espécies arbóreas da Amazônia. Em: Emissão x Seqüestro de CO2. Uma Nova
Oportunidade de Negócios para o Brasil. Manaus, p. 125-153. 1995.
LUCAS, C. M.; SHONGART, J.; SHEIKH, P.; WITTMANN, F.; PIEDADE, M. T. F.;
McGRATH, D. G. Effects of land-use and hydroperiod on aboveground biomassand
productivity of secondary Amazonian floodplain forests. Forest Ecology and
Management 319, p 116–127. 2014.
LIMA, R. C. Equações para estimativas de biomassa de uma floresta tropical úmida
do Amapá. 2015. Dissertação (mestrado em Ciências Florestais) Universidade Federal
Rural de Pernanbuco - UFRPE. Recife. 2015.
SILVEIRA, P; KOEHLER, H. S.; SANQUETA, C. R.; ARCE, J. E. O estado da arte na
estimativa de biomassa e carbono em formações florestais. FLORESTA, Curitiba, PR,
v. 38, n. 1, p. 186-206. 2008.

228
Belém (PA), 11 a 13 de dezembro de 2019
SERAPILHEIRA COMO UM INDICADOR PARA AVALIAÇÃO DE
ECOSSISTEMAS EM RESTAURAÇÃO APÓS MINERAÇÃO DE BAUXITA NA
AMAZÔNIA ORIENTAL
Luana Rodrigues Vieira; Walmer Bruno Rocha Martins2; Gracialda Costa Ferreira3; Victor
Moreira Barbosa4 Francisco de Assis Oliveira5
1 Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected] 2 Mestre em Ciências Florestais. Universidade Federal Rural da Amazônia.
[email protected] 3 Dra. Profa. da Universidade Federal Rural da Amazônia. [email protected]
4 Biólogo, Analista Ambiental da Mineração Paragominas SA. [email protected] 5 Prof. da Universidade Federal Rural da Amazônia. [email protected]
Resumo
O objetivo deste trabalho foi avaliar o estoque de serapilheira e micronutrientes em
ecossistemas anteriormente degradados pela mineração e atualmente em processo de
restauração florestal com diferentes idades e compará-los com uma floresta nativa,
utilizada como ecossistema de referência. Para isso, foi realizada coleta de serapilheira
em dois períodos, chuvoso e menos chuvoso, utilizando-se um gabarito de 0,5 m x 0,5 m
(0,25 m²) e 0,10 m de altura. Foi feita uma amostragem aleatória contendo 6 unidades de
amostras em cada parcela, totalizando 72 no ecossistema com 8 anos de restauração e 48
amostras nos demais ecossistemas. Após as coletas, a serapilheira foi triada em duas
frações, lenhosa e não lenhosa, as quais foram secas em estufa a 70 °C durante 72 horas. Depois, o material foi pesado em balança analítica de precisão de 0,01 g e a porção não
lenhosa foi triturado em um moinho de lâminas do tipo Willey com peneira de 0,85 mm
de abertura para análise química. Os resultados mostraram que é abrangente a presença
de espécies pioneiras, característica de florestas em estágio de regeneração. Os teores de
micronutrientes contidos na serapilheira apresentaram diferenças nítidas para os
elementos Fe e Cu. Além disso, pode-se concluir que, os valores de massa seca ainda se
encontram inferiores aos da floresta natural, mas os valores de Zn e Mn são semelhantes
aos encontrados na floresta natural e, portanto, esses já demostram restaurados. Diferente
disso, o Fe e Cu da serapilheira advinda dos ecossistemas em restauração precisam de
mais tempo para que possam se equiparar a ecossistema de referência.
Palavras-chave: Degradação ambiental. Restauração florestal. Estoque de
micronutrientes.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal.

229
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A mineração é uma atividade que contribui significativamente para a economia
brasileira, gerando, aproximadamente 2,2 milhões de empregos diretos (LIMA; NEVES,
2014). É uma atividade que vem se fortalecendo na Amazônia brasileira, e dentre os
vários produtos explorados, um dos principais é a bauxita, principalmente para obtenção
de alumínio. Neste contexto, o estado do Pará se destaca por ser o principal produtor de
bauxita no país, abrangendo cerca de 90% da produção nacional, o que gerou 33,61
milhões de toneladas no ano de 2015 (ABAL, 2017). Entretanto, mesmo com todo esse
potencial, os impactos ambientais gerados são muitos e quase que inevitáveis, dentre
alguns deles estão a supressão vegetal, revolvimento das camadas superficiais e
subsuperficiais do solo, além do afugentamento da fauna silvestres (LIMA et al., 2016)
Após o processo da atividade mineradora, é fundamental que haja a restauração
do ambiente degradado, especialmente aqueles que se encontram em ecossistemas de
florestas naturais. Para que isso ocorra, faz-se necessário utilizar de métodos eficazes. O
método mais utilizado e mais eficiente é o plantio de mudas arbóreas nativas. Entretanto,
realizar somente o plantio não assegura êxito na recomposição florestal, é necessário que,
atrelado a ele exista um sistema de monitoramento para acompanhar o progresso desse
ecossistema.
Um dos indicadores, que podem ser utilizados para monitorar o processo de
restauração florestal é a serapilheira, formada pela deposição de folhas, galhos, material
reprodutivo e carcaças de animais (RODRIGUES; MARTINS; LEITE, 2010). O aumento
da deposição de serapilheira e dos nutrientes contribui de forma positiva para acelerar o
processo de desenvolvimento do ecossistema (MACHADO et al., 2015).
A serapilheira contribui para a formação e manutenção da fertilidade dos solos
através do processo de decomposição, especialmente nas florestas amazônicas, que são
consideradas pouco férteis naturalmente (QUESADA et al., 2011). Outrossim, a camada
de serapilheira em ecossistemas em início de restauração florestal impede ou minimiza
os processos erosivos (VILLA et al., 2016), que são bastante comuns em áreas mineradas,
principalmente na região amazônica, onde o período chuvoso é intenso.
A serapilheira é, essencialmente, importante por exercer na superfície do solo, a
função de um sistema de entrada e saída, ou seja, sustentando entradas via vegetação e,
por sua vez, decompondo-se e fornecendo ao solo e as raízes nutrientes e matéria

230
Belém (PA), 11 a 13 de dezembro de 2019
orgânica, tornando-se vital na restauração da fertilidade do solo em áreas que se
encontram no início do processo de sucessão ecológica (MACHADO et al., 2019).
A serapilheira é uma grande aliada e uma importante via de transferência de
carbono orgânico, de macronutrientes e micronutrientes (Fe, Zn, Mn e Cu), ajudando no
processo de nutrição das plantas (CALDEIRA et al., 2017). Os micronutrientes têm um
importante papel nessa nutrição, diferindo os teores e absorção de acordo com a
necessidade das plantas e a mobilidade de cada nutriente, apresentando-se de forma
diferente em cada região da planta.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
A pesquisa foi desenvolvida na empresa Mineração Paragominas S.A. (MPSA),
do grupo Hydro, localizada no nordeste do estado do Pará, no Platô Miltônia 3 (3°15'38"S
e 47°43’28” O) a altitude de 150 m, a 70 km da sede municipal de Paragominas.
O clima da região é do tipo “Aw”, segundo a classificação de Köppen-Geiger,
caracterizado como quente e úmido com estações de chuva e de seca bem definidos. A
temperatura média é de 26,3 °C com índice pluviométrico anual de aproximadamente
1.800 mm, sendo que, o período mais chuvoso vai de janeiro a maio, com umidade
relativa do ar em torno de 81% (ALVARES et al., 2013).
A empresa Mineração Paragominas S.A., após a exploração de bauxita do subsolo,
realizou a reconformação do terreno e o espalhamento do topsoil (≅ 20 primeiros
centímetros do solo superficial retirado logo após a supressão de uma floresta natural).
Após este procedimento, foi realizado a restauração florestal com a utilização do método
de plantio de mudas de espécies arbóreas florestais nativas.
2.2 ECOSSISTEMAS DE ESTUDO
Foram selecionadas quatro áreas para avaliação dos indicadores de restauração.
Destes, três estão em processo de restauração e um quarto foi classificado como floresta
ombrófila densa, escolhido como ecossistema de referência para comparação dos
resultados.

231
Belém (PA), 11 a 13 de dezembro de 2019
Nas áreas em restauração foi utilizado o método de plantio de mudas de espécies
florestais arbóreas nativas, plantadas no espaçamento de 3 m x 3 m, implantadas nos anos
de 2009, 2010, 2011, onde foram plantadas inicialmente 104, 89 e 109 espécies
respectivamente. As características das três áreas estão descritas abaixo, na Tabela 1.
Tabela 1- Características específicas dos três ambientes em processo de restauração florestal e a
floresta em áreas de mineração de bauxita, Paragominas, Pará, Brasil.
Áreas
Ano
de
Plantio
Restauração
(Anos)
Área
Hectare Parcela
Subsolagem e
Adubação
pré-plantio
Adubação de plantio
pós-plantio
1 2009 9 71,44 8
Fosfato
natural:
33% P2O5
total, 10%
solúvel ácido
cítrico
200 g.cova-1 de NPK
06-30-06 (0,5% B,
0,5% Cu, 0,5% Zn) e
2,5 kg.cova-1 de adubo
orgânico de cobertura
(calcário dolomítico:
CaO-28%, MgO-12%)
2 2010 8
61,69 8 ----------
Adubação de cobertura
– calcário dolomítico:
CaO-28%, MgO-12%
3 2011 7
123,23 12 ----------
Em covas - 200 g.cova-
1 de NPK 06-30-06
(0,5% B, 0,5% Cu,
0,5% Zn) e 2,5 kg.cova-
1 de adubo orgânico
Fonte: Autores, 2019.
2.3 MÉTODOS
a. Estoque de serapilheira e nutrientes
A coleta de serapilheira estocada no solo foi realizada de maneira aleatória, com
auxílio de um gabarito de 0,5 m x 0,5 m (0,25 m²) e 0,10 m de altura. Para a amostragem,
foram coletadas no interior de cada parcela, 6 amostras, totalizando 72 no ecossistema
com 8 anos de restauração e 48 amostras nos demais ecossistemas. As coletas foram

232
Belém (PA), 11 a 13 de dezembro de 2019
realizadas em dois períodos, março e novembro, ambos em 2018, sendo considerados os
meses mais e menos chuvosos respectivamente.
Em seguida, após a coleta, a serapilheira foi triada em duas frações, lenhosa
(ramos, sementes e material enrijecido) e não lenhoso (folhas e material não enrijecido)
e seca em estufa a 70 °C por 72 horas. Posteriormente o material foi pesado em balança
analítica de precisão de 0,01 g e a porção não lenhosa foi triturado em um moinho de
lâminas do tipo Willey com peneira de 0,85 mm de abertura para análise química de Sódio
(Na), Ferro (Fe), Manganês (Mn), Zinco (Zn) e Cobre (Cu) no Laboratório da Embrapa
Amazônia Oriental - PA, seguindo o método descrito por Malavolta et al. (1997).
2.4 ANÁLISE DE DADOS
Os valores do estoque de serapilheira e nutrientes foi submetido ao teste de
normalidade de Shapiro Wilk e atendidos a esses pressupostos foi realizado a Análise de
Variância para comparação entre os quatro ecossistemas e o teste “t” para comparação do
mesmo ecossistema em períodos diferentes, mais chuvoso e menos chuvoso. As análises
estatísticas foram realizadas no programa estatístico R versão 3.4.3 (R Core Team
Development, 2017).
3. RESULTADOS E DISCUSSÃO
Em relação as espécies predominantes dos ecossistemas em restauração: no
ecossistema com 7 anos de restauração destacaram-se, Croton matourensis Aubl.,
Cecropia sciadophylla Mart., Vismia guianensis (Aubl.) Choisy, já no ecossistema com
8 anos de restauração as espécies que mais se destacaram foram, Croton matourensis
Aubl., Vismia guianensis (Aubl.) Choisy, Guatteria punctata (Aubl.) R.A. Howard, no
ecossistema com 9 anos de restauração, Vismia guianensis (Aubl.) Choisy, Croton
matourensis Aubl., Talisia microphylla Uittien foram as espécies que mais se destacaram,
e por fim, no ecossistema de floresta natural, as espécies foram, Rinorea flavescens
(Aubl.) Kuntz, Inga alba (Sw) Willd., Lecythis idatimon Aubl.
De maneira geral, C. matourensis e V. guianensis foram as espécies que
apresentaram maior representatividade, sendo elas oriundas exclusivamente da
regeneração natural, e estando presente em todos os ecossistemas, exceto a floresta

233
Belém (PA), 11 a 13 de dezembro de 2019
natural, por serem pioneiras que se estabelecem em ambientes perturbados e com ciclo
de vida curto, assistindo na ocupação rápida e, por conseguinte na ocupação do solo
(PARROTTA; KNOWLES, 2001).
3.1 ESTOQUE DE SERAPILHEIRA E NUTRIENTES
O estoque de serapilheira foi maior no ecossistema de floresta natural, nos dois
períodos de coleta, embora, no mês de março, considerado o mais chuvoso, não diferiu
estatisticamente dos ecossistemas com 9 e 8 anos de restauração, diferindo apenas do
ecossistema com 7 anos de restauração (Fig. 1). Por outro lado, no mês de novembro o
estoque de serapilheira no ecossistema de floresta natural, foi maior do que os
ecossistemas com 9 e 7 anos de restauração, não diferindo do ecossistema com 8 anos de
restauração, essa não diferença provavelmente deva ter ocorrido em função da elevada
variabilidade dos valores do ecossistema em restauração (Fig. 1).
Já entre os dois meses para cada ecossistema, o ecossistema com 9 anos de
restauração, foi o único que diferiu entre os meses de março e novembro, sendo que
apresentou maior valor em março, isso pode ter se dado devido a diferença de precipitação
pluviométrica, já que em março há uma intensidade de chuva maior que em novembro.
Figura 1- Estoque de serapilheira no mês de março e novembro de 2018 em ecossistemas com
diferentes idades de restauração florestal após mineração de bauxita e uma floresta de referência,
Paragominas – PA, Brasil. Médias seguidas de seus respectivos erros padrões com a mesma letra
minúscula entre os ecossistemas no mesmo mês não diferem estatisticamente pelo teste de Tukey
(p<0,05) e letras maiúsculas iguais entre os dois meses para cada ecossistema não diferem
estatisticamente pelo teste de t (p<0,05).
Fonte: Autores, 2019.

234
Belém (PA), 11 a 13 de dezembro de 2019
A serapilheira é a principal fonte natural de nutrientes, onde, após o processo de
decomposição, estes retornam para o solo e deste para as plantas, fazendo a ciclagem de
nutrientes. A concentração desses elementos é diretamente afetada pelas condições
climáticas, sobretudo a precipitação pluviométrica (MARTINS et al., 2018) e também
pela matéria orgânica do solo, o que pôde ser observado no presente estudo,
principalmente para o Fe, Mn, Zn e Cu, como mostra a (Fig. 2).
Figura 2. Concentração de micronutrientes na serapilheira estocada no solo em ecossistemas em
processo de restauração florestal após mineração de bauxita com 9, 8 e 7 anos de idade em uma
floresta testemunha. Médias seguidas de seus respectivos erros padrões com a mesma letra
minúscula entre os ecossistemas no mesmo mês não diferem estatisticamente pelo teste de Tukey
(p<0,05) e letras maiúsculas iguais entre os dois meses para cada ecossistema não diferem
estatisticamente pelo teste de t (p<0,05). ns = diferença não significativo.
Fonte: Autores, 2019.

235
Belém (PA), 11 a 13 de dezembro de 2019
A ordem decrescente de concentração dos micronutrientes observada no gráfico
acima foi a seguinte: Zn>Cu>Mn>Fe, que foi diferente da ordem encontrada em outros
artigos como (GODINHO et al., 2014). Essa diferença pode ter se dado por vários
motivos, como o estágio sucessional das diferentes florestas e as espécies nelas
encontradas, como também diferença do material coletado, já que muitos fatores
influenciam na concentração e qualidade do mesmo, como fatores ambientais, clima,
temperatura, umidade e qualidade do material de origem, entre outros (GODINHO et al.,
2014).
Na Fig 2 A, o Fe não apresentou diferença estatística entre os ecossistemas de 7 e
8 anos de restauração no período mais chuvoso, sendo que o ecossistema com 9 anos de
restauração apresentou maior teor de Fe, e este diferiu de todos os ecossistemas, inclusive
a floresta, já no período menos chuvoso a floresta e os ecossistemas com 7 e 8 anos de
restauração foram iguais, diferindo apenas do ecossistema com 9 anos de restauração, que
por sua vez apresentou maior teor de ferro. Entretanto esse valor foi maior no período
mais chuvoso, ou seja, março.
Como mostra a Fig. 2 B, o ecossistema com 7 anos de restauração apresentou
maior de Mn, entretanto não apresentou diferença estatística entre os ecossistemas e a
floresta, no período mais chuvoso. Já no período mais seco, o ecossistema com 7 anos de
restauração foi igual aos demais e o ecossistema com 8 anos de restauração apresentou
maior valor de Mn, entretanto se comparado entre os períodos, todos os ecossistemas
foram iguais, não havendo diferença significativa.
O teor de Zn foi igual em todos ecossistemas, em ambos os períodos, porém se
comparado entre os períodos, o ecossistema com 8 anos de restauração diferiu entre os
dois, sendo que apresentou maior teor de Zn no período mais seco (Fig. 2 C).
Já para Cu, o teor não apresentou diferença estatística entre os ecossistemas com
7, 8 e 9 anos de restauração, no período mais chuvoso, diferindo apenas da floresta, já no
período mais seco o ecossistema com 8 anos de restauração foi igual a todos os outros
ecossistemas, se comparado entre os meses, o ecossistema com 8 anos de restauração foi
o único que diferiu, com a floresta apresentando maior valor de Cu em ambos os períodos,
sendo maior no período menos chuvoso. (Fig. 4 D).

236
Belém (PA), 11 a 13 de dezembro de 2019
4. CONCLUSÃO
Após a análise das amostras, conclui-se que a serapilheira e os micronutrientes
são considerados indicadores eficientes de restauração florestal de áreas degradadas,
principalmente nos ecossistemas mais antigos, sendo ela responsável por vários serviços
na floresta, como ciclagem de nutrientes, cobertura do solo, controle da temperatura e
manutenção da microfauna contribuindo de forma significativa para manutenção do sítio.

237
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ABAL. 2017. Associação Brasileira de Alumínio. Bauxita no Brasil: mineração
responsável e competitividade. São Paulo, ABAL. 66p. Disponível em:
<http://abal.org.br/publicacao/bauxita-no-brasil-mineracao-responsavel-e-
competitividade/> Acesso em 20 de outubro de 2019.
ALVARES, C. A. et al. Köppen's climate classification map for Brazil. Meteorologische
Zeitschrift 22, 711-728, 2013.
CALDEIRA, M. V. W. et al. Quantificação De Serapilheira E De Nutrientes – Floresta
Ombrófila Mista Montana – Paraná. Revista Acadêmica: Ciência Animal, v. 5, n. 2, p.
101, 2017.
GODINHO, T. DE O. et al. Quantificação de biomassa e nutrientes na serapilheira
acumulada em trecho de Floresta Estacional Semidecidual Submontana, ES. Cerne, v.
20, n. 1, p. 11–20, 2014.
LIMA, A. T. et al. The legacy of surface mining: Remediation, restoration, reclamation
and rehabilitation. Environmental Science and Policy, v. 66, p. 227–233, 2016.
LIMA, T. M.; NEVES, C. A. R. Departamento Nacional de Produção Mineral. Sumário
Mineral. Brasília: DNPM, 2014. 152 p.
MACHADO, D. L. et al. Ciclagem de nutrientes em diferentes estádios sucessionais da
Mata Atlântica na bacia do rio Paraíba do Sul, RJ. Bioscience Journal, v. 31, n. 4, p.
1222–1237, 2015.
MACHADO, D. L. et al. Organic matter and soil fertility in different successional stages
of seasonal semidecidual forest. Revista Caatinga, v. 32, n. 1, p. 179–188, 2019.
MALAVOLTA, E.; VITTI, G. C.; OLIVEIRA, S. . Avaliação do Estado Nutricional
das Plantas. 2. ed. Piracicaba: Associação Brasileira para Pesquisa de Potassa e do
Fosfato, 1997.
MARTINS, W. B. R. et al. Deposição de serapilheira e nutrientes em áreas de mineração
submetidas a métodos de restauração florestal em paragominas, Pará. Floresta, v. 48, p.
67–76, 2018.
PARROTTA, J. A.; KNOWLES, O. H. Restoring tropical forests on lands mined for
bauxite: Examples from the Brazilian Amazon. Ecological Engineering, v. 17, n. 2–3, p.
219–239, 2001.
QUESADA, C. A. et al. Soils of Amazonia with particular reference to the RAINFOR
sites. Biogeosciences, v. 8, n. 6, p. 1415–1440, 2011.
R Development Core Team. R: A Language and Environment for Statistical Computing;
R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online:
http://www.R-project.org (acesso em 21 de novembro de 2019).
RODRIGUES, B. D.; MARTINS, S. V.; LEITE, H. G. Avaliação do potencial da
transposição da serapilheira e do banco de sementes do solo para restauração florestal em
áreas degradadas. Revista Arvore, v. 34, n. 1, p. 65–73, 2010.

238
Belém (PA), 11 a 13 de dezembro de 2019
VILLA, E. B. et al. Aporte de serapilheira e nutrientes em área de restauração florestal
com diferentes espaçamentos de plantio. Floresta e Ambiente, v. 23, n. 1, p. 90–99, 2016.

239
Belém (PA), 11 a 13 de dezembro de 2019
VARIAÇÃO RADIAL DO ASPECTO ANATÔMICO DA FIBRA DA MADEIRA
DE CALYCOPHYLLUM SPRUCEANUM BENTH K. SCHUM
João Vitor Farias Paiva Cohen1; Rafaelle de Nazare Queiroz Nascimento2; Marcelino
Carneiro Guedes3; Fernanda Ilkiu Borges de Souza4; Madson Alan Rocha de Sousa5
1Acadêmico de Engenharia Florestal. Universidade do Estado do Pará.
2Acadêmico de Engenharia Florestal. Universidade do Estado do Pará.
3PhD em Recursos Florestais. Embrapa Amapá. [email protected]
4Dra. em Ciências Biológicas. Embrapa Amazônia Oriental. [email protected]
5Mestre em Biodiversidade Tropical. Universidade do Estado do Pará.
RESUMO
O presente estudo teve como objetivo caracterizar o elemento de fibra do lenho de
Calycophyllum spruceanum visando encontrar indicativos de marcadores de transição da
madeira juvenil para adulta no sentido medula-câmbio. Esse tipo de estudo é importante,
pois em virtude de o xilema produzido nos primeiros anos da árvore até certa idade
cambial receber a denominação de lenho juvenil e apresenta propriedades anatômicas,
físicas e mecânicas pouco apropriadas para uso no âmbito industrial quando considerado
o uso da espécie com a finalidade de madeira serrada. Amostra de lenho foi retirada a
altura do DAP, com a demarcação e retirada de corpos de prova de 1 cm² de seção
transversal no sentido medula-câmbio. Assim, para cada posição foi realizado o processo
de maceração do lenho, com 30 medições por posição para as variáveis comprimento de
fibra e espessura da parede da fibra. Desse modo, os valores médios encontrados para
comprimento de fibra foi de 1.703,42 μm e 6,93 μm para a espessura média da parede das
fibras. Por conseguinte, conclui – se que a madeira de Calycophyllum spruceanum com
12 anos de idade e 9 cm de raio não apresentou padrão de estabilização para esas
variáveis, indicando a ausência de lenho adulto.
Palavras-chave: Lenho juvenil. FibraPau mulato. Várzea.
Área de Interesse do Simpósio: Recursos Florestais e Engenharia Florestal.

240
Belém (PA), 11 a 13 de dezembro de 2019
1.INTRODUÇÃO
Os estudos de anatomia da madeira tem de forma contínua contribuído para o
entendimento das amplas relações que essa matéria prima possui com a taxonomia
(MARCELO-PEÑA et al, 2019; AYALA-USMA et al, 2019), ecologia (APARECIDO
et al, 2019), ecofisiologia (CAMP et al, 2018) e indústria (SONG et al, 2018).
A madeira é um conjunto de tecidos especializados e formados por diferentes tipos
de células xilemáticas produzidas pelo câmbio vascular, com características especificas
que formam um sistema complexo, inteligente e eficiente (EVERT, 2013). Apresentam
funções vitais, como por exemplo, condução, armazenamento de substâncias e
sustentação da árvore. Dessa forma, para entender o comportamento da madeira, é
necessário o conhecimento da estrutura e organização dos elementos anatômicos que
compõem o lenho, pois possuem influência em suas propriedades físicas, químicas,
mecânicas e anatômicas (BURGER; RICHTER, 1991; OLIVEIRA et al., 1982). Para
Pimenta (2010) os critérios de qualidade de uma madeira são feitos pelas suas
características anatômicas e composições químicas.
De maneira geral, há uma dificuldade de caracterização da madeira quanto o seu
lenho juvenil e adulto, pois, segundo Gonçalves (2007), esse material é originário de um
sistema biológico complexo e suas propriedades físico-mecânicas variam entre espécies,
entre árvores de uma mesma espécie e nas diferentes partes de uma mesma árvore, como
no sentido medula-casca ou mesmo nos anéis de crescimento. A madeira juvenil se
caracteriza por apresentar menor massa específica, maior ângulo das microfibrilas na
camada S2 da parede celular, fibras mais curtas, menor contração transversal e maior
longitudinal, maior volume de lenho de reação, menor proporção de lenho tardio, paredes
celulares mais finas e menor resistência mecânica em relação à madeira adulta, a qual
possui propriedades mais preferíveis em função da qualidade (ZOBEL, 1984; SENFT;
BENDTESEN; GALLIGAN, 1985; LARSON et al., 2001; NIGOSKI, 2005;
LEONELLO; PALMA; BALLARIN, 2008).
Entende-se que o comprimento das fibras são os parâmetros mais significativos
na diferenciação entre madeira juvenil e madeira adulta, tendo o lenho juvenil o
comprimento de fibra inferior ao do lenho adulto. Além disso, constata-se que com o
aumento da idade a espessura da parede do lenho juvenil é menor que a do lenho adulto
(VIDAURRE, 2011). Segundo Mady (2007), o estudo da anatomia do lenho tem grande

241
Belém (PA), 11 a 13 de dezembro de 2019
importância nas regiões tropicais, pois, se ainda existe um elevado número de espécies
que tem suas características pouco conhecidas, caracterizar essas madeiras é essencial
para indicar seu uso mais adequado.
Estudos que avaliem a variação radial são importantes também e se destacam por
poder indicar e relacionar como as caracteristicas anaômicas do lenho estão relacionadas
com fatores genéticoas, com o clima, solo e outros fatores ambientais (LONGUI et al.,
2017).
Dessa forma, decidiu-se estudar a madeira de Calycophyllum spruceanum
(Benth.) Hook. f. ex K. Schum (pau mulato) pela importância econômica de sua
exploração florestal e uso entre as famílias ribeirinhas no estuário do rio amazonas.
O pau mulato é uma espécie florestal que pertence à família Rubiaceae. A árvore
atinge 20-35 m de altura e 0,7-1,8 m de diâmetro à altura do peito. O tronco retilíneo é
ramificado somente no ápice. A casca fina, quando nova é de coloração esverdeada,
tornando-se pardacenta até castanho-escura. O fruto é uma cápsula elipsoidal deiscente
de 10 mm de comprimento, com lenticelas, pelos esparsos e duas valvas. As sementes
numerosas possuem em torno de 4 mm de comprimento e 0,8 mm de largura e apresentam
coloração parda escura; são aladas em ambas as extremidades (ALMEIDA, 2003).
No âmbito comercial de madeira oriundas de várzea, no estuário amazônico,
Calycophyllum spruceanum conquista o seu espaço na economia, com representação de
20% desse montante. Isso significa que por ano a comercialização da sua madeira gera
R$ 1.140.857 nas microsserrarias (pertencentes às famílias extrativista) e R$ 3.249.672
nas estâncias que vendem madeira serrada nas cidades de Macapá e Santana.
(CASTILHO, 2013). Além do aspecto econômico o pau mulato encontra-se entre as
espécies com importância social, pois atende demanda de muitas microsserrarias
familiares existentes na várzea do estuário amazônico, sendo o destino da madeira
explorada voltado, sobretudo ao atendimento do consumo local e regional (OLIVEIRA
JR et al., 2005).
Assim, este estudo teve como objetivo caracterizar o elemento de fibra do lenho
de Calycophyllum spruceanum visando encontrar indicativos de marcadores de transição
da madeira juvenil para adulta no sentido medula-câmbio.

242
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
As árvores foram coletadas em floresta de várzea do estuário amazônico, no
município de Mazagão, estado do Amapá (00° 14’ 35’’ S e 51° 22’ 59’’ O), em um
afluente do rio Amazonas denominado de igarapé Bispo, área situada em um campo
experimental da Embrapa Amapá. O clima da região é do tipo Ami, equatorial
superúmido, conforme a classificação de Köppen. A temperatura média mínima é de 23°C
e a média máxima de 38°C, com umidade relativa média de 80%. A precipitação
pluviométrica anual oscila em torno de 2.500 mm (GAZEL FILHO et al., 2008). O solo
é classificado como Gleissolo Háplico (SANTOS & TARDIN, 2003). A vegetação
predominante da região é a Floresta Ombrófila Densa Aluvial, também conhecida por
floresta de várzea (IBGE, 2012).
Para a retirada das amostras foram abatidas cinco árvores de pau-mulato aos 12
anos de idade, das quais de cada uma, a 1,30 m de altura do solo, foram retirados discos
de 5 cm de espessura para confecção dos corpos de prova. Esse material foi preparado na
Xiloteca da Embrapa Amazônia Oriental, com a demarcação e retirada de corpos-de-
prova de 1 cm² de seção transversal no sentido medula-câmbio. Para fins de apresentação
dos resultados parciais da pesquisa, selecionou-se uma árvore, que totalizou nove
posições no sentido radial, com cada posição distante 1cm entre si (posição 1 mais
próxima à medula, posição 9 mais próxima ao câmbio). Foram medidos as dimensões de
comprimento de fibra (CF) e espessura da parede da fibra (Epf), com 30 repetições para
cada posição.
Para tais análises microscópicas foram preparadas lâminas com material
macerado. O material macerado foi obtido a partir da inclusão de fragmentos
longitudinais de madeira, oriundos dos corpos de prova, em solução de acético e peróxido
de hidrogênio na proporção 1:1, levados a estufa a 60⁰C por 24 h. Após esse período, o
material já dissociado foi lavado em água destilada e corado com safranina hidro-etílica
(1%) para melhor observação dos elementos celulares (vasos e fibras) em lâminas
temporárias., As mensurações para o comprimento de fibra e espessura da parede foram
realizadas mensurados de acordo com orientação da International Association of Wood
Anatomists – IAWA Committee (1989).

243
Belém (PA), 11 a 13 de dezembro de 2019
Análise de variância foi testada para verificar diferenças significativas entre as
posições, com posterior teste de Tukey para identificar quais são as diferenças, ambas a
0,05% de significância.
3. RESULTADOS E DISCUSSÃO
A estatística descritiva para os valores de comprimento da fibra e espessura da
parede encontra-se na tabela 1. Os valores encontrados para essas variáveis são
condizentes com resultados apresentados por Baldin e Marchiori (2015), que descreveram
a espécie para definição de sua melhor posição taxonômica na família Rubiaceae, com
fibra de comprimento médio de 1441 µm ± 366, e espessura da parede média de 7 µm ±
2. Com relação ao aspecto da espessura da parede o pau mulato apresentou valor próximo
a Aspidosperma polyneuron com 7,36 µm e maior espessura da parede que outras
espécies, como a Pterodon pubescens com 5,79 µm, a Cedrela fissilis com 3,87 µm,
Myroxylon balsamum com 5,91 µm e a Anadenanthera macrocarpa com 5,49 µm
(Benites, 2015).
Tabela 1 – Parâmetros anatômicos usados na determinação da madeira juvenil e madeira adulta Estruturas da fibra Média Mínimo Máximo Desvio padrão Coeficiente de
Variação (%)
Comprimento (µm) 1.703,42 1.000 2.500,00 214,80 10,30
Espessura da parede (µm) 6,93 5 12,5 1,28 17,36
Fonte: Autor
O comprimento de fibra apresentou muita variação, e segundo a classificação
sugerida pelo IAWA, elas podem ser definidas como longas, com os valores > 1600. A
espessura da parede da fibra de acordo com a classificação da IAWA apresenta dimensões
de espessura fina para grossa, lume da fibra menor que três vezes a espessura da parede.
A análise de variância para os valores de comprimento da fibra e espessura da
parede encontra-se na tabela 2 e 3, respectivamente. Existiu diferenças significativas para
ambas características em função da posição radial no fuste.
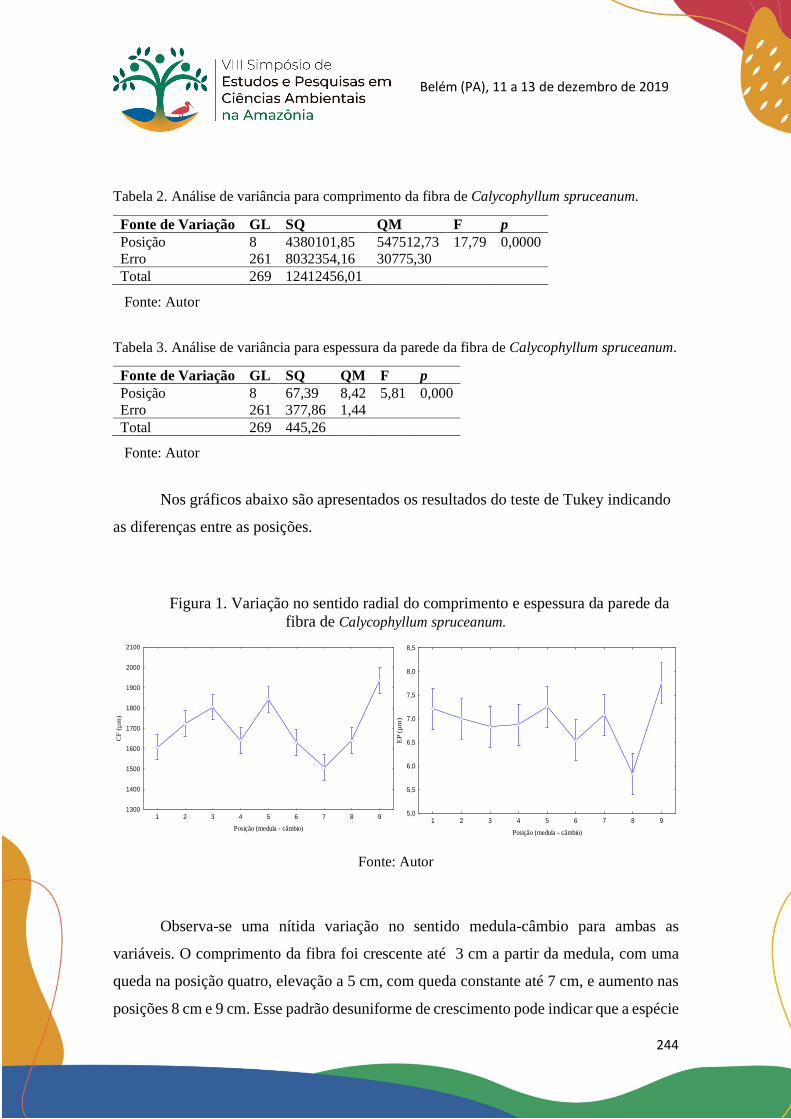
244
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 2. Análise de variância para comprimento da fibra de Calycophyllum spruceanum.
Fonte de Variação GL SQ QM F p Posição 8 4380101,85 547512,73 17,79 0,0000 Erro 261 8032354,16 30775,30
Total 269 12412456,01
Fonte: Autor
Tabela 3. Análise de variância para espessura da parede da fibra de Calycophyllum spruceanum.
Fonte de Variação GL SQ QM F p Posição 8 67,39 8,42 5,81 0,000 Erro 261 377,86 1,44
Total 269 445,26
Fonte: Autor
Nos gráficos abaixo são apresentados os resultados do teste de Tukey indicando
as diferenças entre as posições.
Figura 1. Variação no sentido radial do comprimento e espessura da parede da
fibra de Calycophyllum spruceanum.
Fonte: Autor
Observa-se uma nítida variação no sentido medula-câmbio para ambas as
variáveis. O comprimento da fibra foi crescente até 3 cm a partir da medula, com uma
queda na posição quatro, elevação a 5 cm, com queda constante até 7 cm, e aumento nas
posições 8 cm e 9 cm. Esse padrão desuniforme de crescimento pode indicar que a espécie
Posição; LS Means
Wilks lambda=,41702, F(24, 751,78)=11,025, p=0,0000
Effective hypothesis decomposition
Vertical bars denote 0,95 confidence intervals
1 2 3 4 5 6 7 8 9
Posição (medula - câmbio)
1300
1400
1500
1600
1700
1800
1900
2000
2100
CF
(µ
m)
Posição; LS Means
Wilks lambda=,41702, F(24, 751,78)=11,025, p=0,0000
Effective hypothesis decomposition
Vertical bars denote 0,95 confidence intervals
1 2 3 4 5 6 7 8 9
Posição (medula - câmbio)
5,0
5,5
6,0
6,5
7,0
7,5
8,0
8,5
EP
(µ
m)

245
Belém (PA), 11 a 13 de dezembro de 2019
ainda não tenha formado madeira adulta, pois não houve estabilização para o
comprimento da fibra. Fato sustentado pela afirmação de Palermo et al (2013), que
destacam o comprimento da fibra como principal indicador para o ponto de transição de
madeira juvenil para adulta. Esses resultados são semelhantes ao encontrados por
Barreiros (2016).
De forma similar, a espessura da parede da fibra também não apresentou padrão
de variação. Embora, tal qual para o comprimento, o maior valor tenha sido encontrado
na região próxima ao câmbio (7,75 µm), não houve estabilização da medida indicando a
presença de madeira adulta. Vidaurre (2011) aborda que as propriedades da madeira não
são uniformes no sentido medula-casca, e que nos primeiros anéis de crescimento a
madeira apresenta menor comprimento de espessura da parede. Em anéis sucessivos,
partindo do centro da árvore, verifica-se o aumento das características anatômicas, como
comprimento, diâmetro do lúmen e espessura da parede da fibra, porque a espessura da
parede celular do lenho juvenil é menor do que a do lenho adulto e isso é observado com
o aumento da idade. No entanto, tais características não se apresentaram de forma
ordenada para o lenho de pau mulato, fato que pode indicar que aos 12 anos de idade a
espécie ainda não apresenta madeira adulta, todavia, essa consideração deve ser
ponderada pelo fato de os resultados parciais apresentados serem de uma única árvore.
Longui et al. (2017) estudando a influência de fatores ambientais e genéticos na
variação radial de características da madeira de Myracrodruon urundeuva Allemão
(Anarcadiaceae) encontraram um padrão de variação crescente no sentido medula-câmbio
apenas para o comprimento de fibra, com a espessura da parede não apresentando
regularidade; os autores concluem que o comprimento de fibra é muito mais associado a
fatores genéticos do que ambientais, pois analisando o lenho da espécie oriunda de
sementes de duas procedências diferentes, o comprimento de fibra apresentou o mesmo
padrão na região de instalação do experimento.
4. CONCLUSÃO

246
Belém (PA), 11 a 13 de dezembro de 2019
A árvore Calycophyllum spruceanum analisada apresenta variação radial na
dimensão do comprimento e espessura da parede de fibra do lenho aos 12 anos de idade
e 9 cm de raio. A madeira foi caracterizada como juvenil, não havendo um ponto de
estabilização para estimar a transição entre a madeira juvenil e adulta. Novas ferramentas
estatísticas serão realizadas com o aumento da amostragem de árvores pars estabelece de
forma definitiva a presença ou não de madeira adulta.

247
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALMEIDA, M. C. Aspectos Ecofisiológicos da Germinação de Sementes de Mulateiro
(Calycophyllum spruceanum Benth.) – Rubiaceae. 2003. 116 p. Tese (Doutorado em Ciências
Biológicas). Universidade Estadual Paulista “Júlio de Mesquita Filho”, Rio Claro, 2003.
APARECIDO, L.M.T. et al. Relevance of wood anatomy and size of Amazonian trees in the
determination and allometry of sapwood area. Acta Amazônica, v. 49, 2019.
AYALA-USMA, D.A.; LOZANO-GUTIÉRREZ, R.E.; ARANGO, C.G. Wood anatomy of two
species of the genus Chrysochlamys (Clusiaceae: Clusioideae: Clusieae) from the northern Andes
of Colombia. Heliyon, v. 5, 2019.
BALDIN, T.; MARCHIORI, J.N.C. Anatomia da madeira de Calycophyllum spruceanum
(BENTH.) K. SCHUM. e Calycophyllum spruceanum f. brasiliensis K. SCHUM. Balduinia, n.
50, p. 16-23, 30-XI, 2015. BALDIN, T.alita. Anatomia do lenho do gênero calycophyllum a.Dc. (rubiaceae). 2015. 136
f. Dissertação (Mestrado em Engenharia Florestal) - Universidade Federal De Santa Maria, Santa
Maria, 2015.
BARREIROS, RicardoR.. Estudo quantitativo da variação estrutural morfológica na madeira de
calophyllum brasiliense. Cerne, Lavras, vol. 22, núm. 1, 2016, pp. 77-83, 2016. Universidade
Federal de Lavras, Lavras, Brasil.
BENITES, P.atrícia. Caracterização anatômica das fibras de oito espécies florestais do Cerrado
de Mato Grosso do Sul para a produção de papel. Ciência da Madeira, Pelotas, v. 6, n. 2, 2015.
BURGER, L. M.; RICHTER, H. G. Anatomia da madeira. São Paulo: Nobel, 1991. 154 p.
CAMP, J.V. et al. Cambial pinning relates wood anatomy to ecophysiology in the African tropical
tree Maesopsis eminii. Tree Physiology, v. 38, 2017.
CASTILHO, Núbia TathianeFurtado. Manejo da regeneração natural e produção de madeira de
pau mulato em floresta de várzea do estuário amazônico. Dissertação Programa de Pós-
Graduação em Biodiversidade Tropical. AmapáUniversidade Federal do Amapá, 2013. 96p.
EVERT, RAY FRANKLIN. Anatomia das plantas de Esau: meristemas, células e tecidos do
corpo da planta: sua estrutura, função e desenvolvimento [livro eletrônico] / Ray F.Evert ;
Ccoord.enação e tradução de Carmen Regina Marcati. – São Paulo: Bluecher, 2013. 728 p.;
GAZEL FILHO, A. B. et al. ; YARED, J. A. G.; MOURÃO JR., M.; SILVA, M. F.; CARIM, M.
J. V.; JARDIM, M. A. G.; MEDEIROS, T. D. S. Composição florística e estrutura de floresta de
várzea no município de Mazagão, estado do Amapá, Brasil. Scientia Forestalis, Piracicaba, v.
36, n. 79, p. 191-201, 2008.
GONÇALVES, M. P. M. et al. Variação radial da densidade básica e comprimento das fibras da
madeira de Tectona grandis L. Floresta e Ambiente, Rio de Janeiro, v. 14, n. 1, p. 70-75, 2007.
IBGE - INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Manual técnico da
vegetação brasileira. 2.ed. Rio de Janeiro, p. 271, 2012.
LARSON P. R. et al. Formation and properties of juvenile wood in southern pines. Madison:
Forest service, 2001. 46 p.
LEONELLO, E. C.; PALMA, H. A. L.; BALLARIN, A. W. Delimitação da madeira juvenil e
adulta de Eucalyptusgrandis. Revista Forestal Venezolana, Merida, v. 52, n. 1, p. 93-98, 2008.

248
Belém (PA), 11 a 13 de dezembro de 2019
LONGUI, E.L. et al. Genetic Versus Environmental Influence on Radial Variation in
Myracrodruon urundeuva Wood. Floresta e Ambiente: 24, 2017.
MADY, F.T.M. Técnicas para Microscopia da Madeira. Manaus: Editora da Universidade
Federal do Amazonas (EDUA); 2007. 80 p.
MARCELO-PEÑA, J.L.; JR SANTINI, L.; TOMAZELLO FILHO, M. Wood anatomy and
growth rate of seasonally dry tropical forest trees in the Marañón River Valley, northern Peru.
Dendrochronologia, v. 55, 135-145, 2019.
NIGOSKI, S. Espectroscopia no infravermelho no estudo de características da madeira e
papel de Pinus taeda L. 2005. 160 f. Tese (Doutorado em Ciências Florestais) - Universidade
Federal do Paraná, Curitiba, 2005.
OLIVEIRA JR., de P. H. B.; MACEDO, D. S.; NOGUEIRA, E. L. S.; GUEDELHA, C. Relatório
Complementar: Caracterização da produção, comercialização e o potencial de certificação
de madeira nas florestas de várzea. Manaus, 2005.51p.
OLIVEIRA, J. B.; VIVACQUA FILHO, A.; GOMES, P. A. Produção de carvão vegetal: aspectos
técnicos. In: PENEDO, W.R. (Org.). Produção e utilização de carvão vegetal. Belo Horizonte:
CETEC, 1982. p. 60-73. (Série de Publicações Técnicas, 8).
PALERMO, G. P. de M. et al; LATORRACA, J. V. de F.; SEVERO, E. T. D.; NASCIMENTO,
A. M. do; REZENDE, M. A. Delimitação entre os lenhos juvenil e adulto de Pinus elliottii
Engelm. Revista Árvore, Viçosa, v. 37, n. 1, p. 191-200, 2013.
PIMENTA, A. S.; BARCELLOS, D. C.; OLIVEIRA E. Curso de carbonização. Viçosa:
Universidade Federal de Viçosa, Laboratório de Painéis e Energia da Madeira. 2010. 94p.
SANTOS, V. F.; TARDIN, A. T. Projeto zoneamento ecológico-econômico do setor costeiro
estuarino do Estado do Amapá: diagnóstico socioambiental, relatório técnico de solos. Macapá:
IEPA, p. 22, 2003.
SONG, J. et al. Processing bulk natural wood into a high-performance structural material. Nature,
Letter, v. 554, 2018.
TREVISAN, Rômul. Dimensões das fibras e sua relação com a idade de transição entre lenho
juvenil e adulto de eucalyptus grandis w. Hill ex maiden. Ciência Florestal, Santa Maria, v. 27,
n. 4, p. 1385-1393, out.-dez., 2017.
VIDAURRE, G.; LOMBARDI, L. R.; OLIVEIRA, J. T. S.; ARANTES, M. D. C. Lenho juvenil
e adulto e as propriedades da madeira. Floresta e Ambiente, v. 18, n. 4, 2011.
ALMEIDA, M. C. Aspectos Ecofisiológicos da Germinação de Sementes de Mulateiro
(Calycophyllum spruceanum Benth.) – Rubiaceae. 2003. 116 p. Tese (Doutorado em Ciências
Biológicas). Universidade Estadual Paulista “Júlio de Mesquita Filho”, Rio Claro, 2003.
BALDIN, T.; MARCHIORI, J.N.C. Anatomia da madeira de Calycophyllum spruceanum
(BENTH.) K. SCHUM. e Calycophyllum spruceanum f. brasiliensis K. SCHUM. Balduinia, n.
50, p. 16-23, 30-XI, 2015.
BALDIN, T.alita. Anatomia do lenho do gênero calycophyllum a.Dc. (rubiaceae). 2015. 136
f. Dissertação (Mestrado em Engenharia Florestal) - Universidade Federal De Santa Maria, Santa
Maria, 2015.

249
Belém (PA), 11 a 13 de dezembro de 2019
BARREIROS, RicardoR.. Estudo quantitativo da variação estrutural morfológica na madeira de
calophyllum brasiliense. Cerne, Lavras, vol. 22, núm. 1, 2016, pp. 77-83, 2016. Universidade
Federal de Lavras, Lavras, Brasil.
BENITES, Patrícia. Caracterização anatômica das fibras de oito espécies florestais do Cerrado de
Mato Grosso do Sul para a produção de papel. Ciência da Madeira, Pelotas, v. 6, n. 2, 2015.
BURGER, L. M.; RICHTER, H. G. Anatomia da madeira. São Paulo: Nobel, 1991. 154 p.
CASTILHO, Núbia TathianeFurtado. Manejo da regeneração natural e produção de madeira de
pau mulato em floresta de várzea do estuário amazônico. Dissertação Pós-Graduação. Amapá,
2013. 96p.
EVERT, RAY FRANKLIN. Anatomia das plantas de Esau: meristemas, células e tecidos do
corpo da planta: sua estrutura, função e desenvolvimento [livro eletrônico] / Ray F.Evert ;
coordenação e tradução de Carmen Regina Marcati. – São Paulo: Bluecher, 2013. 728 p.; PDF
GAZEL FILHO, A. B.; YARED, J. A. G.; MOURÃO JR., M.; SILVA, M. F.; CARIM, M. J. V.;
JARDIM, M. A. G.; MEDEIROS, T. D. S. Composição florística e estrutura de floresta de várzea
no município de Mazagão, estado do Amapá, Brasil. Scientia Forestalis, Piracicaba, v. 36, n. 79,
p. 191-201, 2008.
GONÇALVES, M. P. M. et al. Variação radial da densidade básica e comprimento das fibras da
madeira de Tectonagrandis L. Floresta e Ambiente, Rio de Janeiro, v. 14, n. 1, p. 70-75, 2007.
IBGE - INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Manual técnico da
vegetação brasileira. 2.ed. Rio de Janeiro, p. 271, 2012.
LARSON P. R. et al. Formation and properties of juvenile wood in southern pines. Madison:
Forest service, 2001. 46 p.
LEONELLO, E. C.; PALMA, H. A. L.; BALLARIN, A. W. Delimitação da madeira juvenil e
adulta de Eucalyptusgrandis. Revista Forestal Venezolana, Merida, v. 52, n. 1, p. 93-98, 2008.
Mady FTM. Técnicas para Microscopia da Madeira. Manaus: Editora da Universidade Federal
do Amazonas (EDUA); 2007. 80 p.
NIGOSKI, S. Espectroscopia no infravermelho no estudo de características da madeira e
papel de Pinus taeda L. 2005. 160 f. Tese (Doutorado em Ciências Florestais) - Universidade
Federal do Paraná, Curitiba, 2005.
OLIVEIRA JR., de P. H. B.; MACEDO, D. S.; NOGUEIRA, E. L. S.; GUEDELHA, C. Relatório
Complementar: Caracterização da produção, comercialização e o potencial de certificação
de madeira nas florestas de várzea. Manaus, 2005.51p.
OLIVEIRA, J. B.; VIVACQUA FILHO, A.; GOMES, P. A. Produção de carvão vegetal: aspectos
técnicos. In: PENEDO, W.R. (Org.). Produção e utilização de carvão vegetal. Belo Horizonte:
CETEC, 1982. p. 60-73. (Série de Publicações Técnicas, 8).
PALERMO, G. P. de M.; LATORRACA, J. V. de F.; SEVERO, E. T. D.; NASCIMENTO, A.
M. do; REZENDE, M. A. Delimitação entre os lenhos juvenil e adulto de Pinus elliottii Engelm.
Revista Árvore, Viçosa, v. 37, n. 1, p. 191-200, 2013.
PIMENTA, A. S.; BARCELLOS, D. C.; OLIVEIRA E. Curso de carbonização. Viçosa:
Universidade Federal de Viçosa, Laboratório de Painéis e Energia da Madeira. 2010. 94p.

250
Belém (PA), 11 a 13 de dezembro de 2019
SANTOS, V. F.; TARDIN, A. T. Projeto zoneamento ecológico-econômico do setor costeiro
estuarino do Estado do Amapá: diagnóstico socioambiental, relatório técnico de solos. Macapá:
IEPA, p. 22, 2003.
SENFT, J. F.; BENDTESEN, B. A.; GALLIGAN, W. L. Weakwood. Journa lof Forestry,
Bethesda, n. 83, p. 476-485, 1985.
TREVISAN, Rômul. Dimensões das fibras e sua relação com a idade de transição entre lenho
juvenil e adulto de eucalyptus grandis w. Hill ex maiden. Ciência Florestal, Santa Maria, v. 27,
n. 4, p. 1385-1393, out.-dez., 2017.
VIDAURRE, Graziela. Lenho Juvenil e Adulto e as Propriedades da Madeira. Floresta e
Ambiente. Espírito Santo, 2011. Disponível em:
file:///C:/Users/Sansung/Downloads/Vidaurre.et.al.2011.Lenho%20juvenil%20e%20adulto%20
e%20as%20propriedades%20da%20madeira.pdf. Acesso em: 19/10/2019.
ZOBEL B. J. The changing quality of the world woodsupply. Wood Science and Technology,
New York, v. 18, n. 1, p. 1-17, 1984.

251
Belém (PA), 11 a 13 de dezembro de 2019
A IMPORTÂNCIA AMBIENTAL DO SISTEMA AGROFLORESTAL DE
TOMÉ-AÇÚ – SAFTA, LOCALIZADO NO NORDESTE DO ESTADO DO
PARÁ
Joice Machado Martins1; Carla Lyzandra Linhares2; Adler Henrique Rodrigues Alves³;
Paulo Rick Soares Rodrigues⁴
1 Graduada em Engenharia Ambiental e Energias Renováveis. Universidade Federal
Rural da Amazônia. [email protected]
2 Graduada em Engenharia Ambiental e Energias Renováveis. Universidade Federal
Rural da Amazônia. [email protected]
³ Mestrando no programa de pós-graduação em aquicultura e recursos aquáticos
tropicais. Universidade Federal Rural da Amazônia. [email protected]
⁴ Graduando em Engenharia Ambiental e Energias Renováveis. Universidade Federal
Rural da Amazônia. [email protected]
RESUMO
Visando a redução da agricultura convencional que faz uso de práticas predatórias é
interessante pensar em formas de plantio mais eficazes que levem em consideração os
conceitos sustentáveis de uso do solo e dos recursos naturais. Sendo assim, o objetivo
deste estudo é destacar a importância ambiental dos sistemas agroflorestais e incentivar
o seu emprego em qualquer bioma visto as suas inúmeras vantagens. Para isso, utilizou-
se como exemplo prático o sistema agroflorestal de Tomé-Açú – SAFTA – e buscou-se
verificar como o SAFTA e a cooperativa mista de Tomé-Açú – CAMTA – afetam
positivamente o cenário ambiental e econômico da região. A metodologia utilizada foi a pesquisa de campo e bibliográfica. Na ocasião, duas fazendas foram visitadas com o
intuito de analisar de perto os SAF’s e coletar informações de agricultores. Além disso,
visitou-se também a cooperativa mista de Tomé-Açú – CAMTA com a finalidade de
conhecer a sua organização e seus processos de produção. Diante ao exposto, pôde-se
constatar que os SAF’s trazem inúmeros benefícios para a localidade onde é implantado,
no âmbito ambiental ele proporciona grande biodiversidade e maior produção de matéria
orgânica, promovendo a manutenção do bem-estar dos ecossistemas, enquanto que no
âmbito social ele permite e união dos cooperados e a integração de mulheres no processo
de cultivo, já no contexto econômico, por meio da CAMTA, é proporcionado um
aumento da rentabilidade dos agricultores, melhorando a qualidade de vida e a estrutura
do município.
Palavras-chave: sistemas agroflorestais. CAMTA. Modos de produção sustentáveis.
Área de Interesse do Simpósio: sistemas agroflorestais

252
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Nas últimas décadas notou-se diretamente as consequências ambientais negativas
que o modo de produção predatório derivado da agricultura convencional pode ocasionar
no bioma Amazônia. A floresta vem perdendo extensão e biodiversidade devido a retirada
de madeira legal e ilegal, queimadas em grande escala que desestabilizam a fauna e a
flora, além disso, tem-se a contaminação do meio aquático e do solo por produtos
agrícolas ou de mineração. Somado a estes fatores vê-se ainda a retirada de grandes
extensões de cobertura vegetal cujo terreno residual é direcionado para a criação de gado
e plantação de monoculturas.
Neste cenário, segundo o boletim de desmatamento da Amazônia de 2019,
elaborado pelo sistema de alerta de desmatamento (SAD) do Imazon, que em seu
calendário atual considerou o período de agosto de 2018 a julho de 2019, destacou que o
desmatamento na Amazônia legal foi de 5.054 Km², correspondendo então, a um aumento
de 15% em relação ao calendário anterior (IMAZON, 2019). Visando a redução do
desmatamento e da degradação da floresta amazônica é interessante pensar em formas de
plantio mais eficazes que leve em consideração a manutenção da qualidade de seus
recursos naturais. Atualmente vê-se muitos incentivos à agricultores que empregam
técnicas verdes em seus modos de cultivo, além de obterem um bom reconhecimento
perante seus consumidores, acrescentando assim, um diferencial aos seus produtos.
Levando isso em consideração, o emprego de sistemas agroflorestais – SAF’s – é
muito viável e interessante tanto para quem produz quanto para o meio ambiente. Este
sistema de plantio consiste em atividades extrativistas sustentáveis associadas a
reconstituição de regiões degradadas. Apresenta como principal objetivo o
aperfeiçoamento do uso da terra, incentivando a produção florestal juntamente com a
produção agrícola, evidenciando a importância dos sistemas agroflorestais com os
processos dinâmicos dos ecossistemas naturais uma vez que o intuito nos SAF’s é a
formação de vários andares de vegetação, imitando uma floresta natural, tendo as árvores
e arbustos como os elementos estruturais básicos e principais para a estabilidade do
sistema, respeitando o tempo natural de produção das culturas (KONAGANO et al.,
2016).
Sendo assim, o objetivo deste estudo é destacar a importância ambiental dos
sistemas agroflorestais para a manutenção da qualidade do bioma Amazônia e incentivar

253
Belém (PA), 11 a 13 de dezembro de 2019
o seu emprego em qualquer bioma do país visto as suas inúmeras vantagens. Para isso,
utilizou-se como exemplo prático o sistema agroflorestal de Tomé-Açú – SAFTA – e
buscou-se verificar como o SAFTA e a cooperativa mista de Tomé-Açú – CAMTA –
afetam positivamente o cenário ambiental e econômico desta região.
2. MATERIAL E MÉTODOS
Com o intuito de observar os SAF’s de perto, acompanhar as metodologias dos
agricultores, ouvir seus relatos, identificar quais as principais culturas da região e realizar
registros fotográficos, optou-se por realizar visitas de campo, caracterizando-se então,
como pesquisa exploratória. Na oportunidade, duas fazendas foram visitadas, a primeira,
é propriedade do senhor Mitinori Konagano e a segunda, do senhor Kozaburo Mineshita,
os dois são contribuintes da Cooperativa Mista de Tomé-Açú (CAMTA), que também foi
ponto de estudo. Dessa forma, pôde-se observar como a CAMTA é organizada e
estruturada. Por fim, fez-se uso também de referencial teórico de fontes confiáveis.
2.1 ÁREA DE ESTUDO
A área objeto deste estudo situa-se na região nordeste do estado do Pará, o
município denomina-se Tomé-Açu, sua extensão territorial é de 5.179 Km2, comporta
cerca de 61.095 habitantes. Possui um clima tropical chuvoso com estação seca bem
definida, além disso, apresenta precipitação média anual de 2.500 mm, com distribuição
mensal irregular, tendo de novembro a junho uma maior intensidade de chuvas. Sua
temperatura média anual é de 26ºC e umidade relativa do ar de 85%. Os solos são
representados principalmente por latossolo amarelo distrófico, gley pouco úmido
eutrófico e distrófico e aluvial eutrófico e distrófico, em associação. A vegetação original,
atualmente alterada, correspondendo a floresta ombrófila densa (DE BARROS et al.,
2009; GOVERNO DO ESTADO DO PARÁ, 2011; EMBRAPA, 2001)

254
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
Este tópico será destinado à exposição dos resultados obtidos através das
conversas com os produtores rurais das duas fazendas mencionais e com os técnicos
presente nas visitas, bem como, das observações feitas acerca dos SAF’s.
2.1 SISTEMAS AGROFLORESTAIS DE TOMÉ-AÇÚ – SAFTA
Os SAF’s correspondem a um conjunto de sistemas e tecnologias de uso da terra
onde o cultivo final pode derivar de cultivos anteriores, de forma sequencial ou rotativa,
podendo ser culturas anuais, semi-perenes ou perenes, também podem ser derivadas de
pastagens, reflorestamento, dentre outros (DE BARROS et al., 2009). Dubois (2009)
divide os SAF’s em três tipos, levando em consideração os aspectos estruturais e
funcionais, por primeiro, apresenta-se os sistemas agrossilviculturais, no qual ocorre a
combinação de cultivos agrícolas anuais com árvores, por conseguinte tem-se os sistemas
agrossilvipastoris que além das combinações já citadas, abrange a criação de animais, por
fim, tem-se os sistemas silvipastoris, aqui, associa-se árvores à criação e animais.
Os SAF’s geralmente apresentam diferentes composições de espécies, com
variadas formas e tamanho, tais aspectos vão alternando de acordo com as necessidades
e objetivos de cada agricultor. Dessa forma, torna-se um modelo base de produção
diversificada cuja biodiversidade é seu principal ponto positivo (KONAGANO et al.,
2016).
No Sistema Agroflorestal de Tomé – Açú – SAFTA, o cacau e o cupuaçu são as
duas culturas perenes predominantes devido sua forte participação na rentabilidade final
dos agricultores. Além disso, observou-se plantações de pimenta-do-reino em muitas
propriedades, isso ocorre devido a tradição e também por trazer retorno financeiro em
curto período. Segundo Konagano et al (2016) os SAF’s de Tomé-Açú são formados
sobretudo pelo cultivo de açaí, cupuaçu, cacau e pimenta-do-reino, que podem estar
dispostas juntas e/ou com espécies frutíferas ou florestais.
2.2 FAZENDA KONAGANO
A fazenda Konagano é propriedade do Sr. Mitinori Konagano, possui uma
extensão territorial de 850 hectares, sendo 230 ha de áreas de SAF’s produtivos, divididos
entre culturas de cacau e açaí (160 ha), como mostra a Figura 1, cupuaçu (55 ha) e

255
Belém (PA), 11 a 13 de dezembro de 2019
pimenta-do-reino (15 ha), além da criação de aves, peixes e suínos. O plantio de cacau
em consórcio com açaí (Euterpe Oleracea) ocupa a mesma área e ocorre em períodos
intercalados do ano, sendo o cacau de maio a outubro e o açaí de outubro a maio. O cacau
(Theobroma cacao L.) é a principal cultura explorada nos SAF’s do município, estando
pressente em 81% das propriedades, devido a sua alta rentabilidade no sistema
(KONAGANO et al., 2016).
Figura 1 – SAF’s de cacau e açaí
Fonte: autores (2019)
Na Fazenda Konagano, a pimenta-do-reino ocupa a menor área do SAF, 15ha,
tendo sua safra nos períodos de julho a dezembro, na fazenda usa-se o espaçamento
2mx2mx5m no cultivo dessa cultura. Para exemplificar a rentabilidade da pimenta-do-
reino, o Sr. Mitinori esmiuçou como ocorre o processo dinâmico dela: o preço do Quilo
atualmente custa em torno de R$ 7,00, ou seja, com a venda de uma tonelada, o lucro
bruto é de R$ 70 mil reais. Com esse valor, o agricultor consegue comprar 40 toneladas
de adubo químico para abastecer 50 ha de cacau, no qual irão gerar um faturamento bruto
de R$ 525.000 ao agricultor. Logo, para os agricultores de Tomé-açu, a pimenta do reino
é vista como uma poupança ou um “coringa”, pois mesmo com as baixas de preço sempre
haverá mercado e seus lucros são convertidos em investimentos em outras culturas. Além
dos cultivos já citadas, a fazenda Konagano produz outras culturas anuais em menor
escala como: melânica, abóbora, milho e macaxeira, sendo um complemento a renda.
Na fazenda existem dois tanques de criação de peixes. O proprietário comentou
sobre a intenção de criar um sistema de irrigação que aproveitasse a água dos tanques
para as plantações, diminuindo então, a utilização de água limpa. Quanto a produção de
suínos, o Sr. Mitinori criou um sistema nos chiqueiros, no qual as casas de porcos são

256
Belém (PA), 11 a 13 de dezembro de 2019
construídas sobre solos compostos de 80% de serragem, 10% de terra da mata e 10% de
carvão e sal mineral. Essa mistura foi desenvolvida especialmente pelo proprietário para
que, após certo período de uso os dejetos dos animais sejam misturados ao composto do
solo, servindo como adubo natural para as culturas da área. Além disso, a fazenda tem a
produção de seu próprio fertilizante: o biofertilizante, que é gerado através da mistura de
folhas de bananeira, esterco animal, “caldo” fermentativo do cacau e outros componentes,
essa mistura passa por vários dias de fermentação até poder ser utilizado nos plantios
(Figura 2).
Figura 2 (a e b) – Produção de biofertilizante
A B
Fonte: autores (2019)
O Sr. Mitinori busca maximizar o uso dos serviços ecossistêmicos a seu favor,
com técnicas de agricultura alternativa que não utilizam agrotóxicos, herbicidas ou
fertilizantes inorgânicos, para assim, garantir o melhor aproveitamento de sua
propriedade.
2.3 FAZENDA MINESHITA
A segunda fazenda visitada é propriedade do Sr. Kozaburo Mineshita, abrange
uma extensão territorial de 15 ha divididos entre as culturas de cupuaçu, açaí, pupunha e
cacau. Segundo o proprietário, a fazenda apresenta uma produção anual de 10 toneladas
de cacau, 2 toneladas de pimenta-do-reino, 20 toneladas de açaí e 20 toneladas de
pupunha.

257
Belém (PA), 11 a 13 de dezembro de 2019
Em decorrência das dimensões da fazenda, o Sr. Mineshita aproveita ao máximo
sua área através de um espaçamento adequado, intercalando as culturas de modo que não
haja competição entre as espécies.
Além das culturas já citadas, foi observado a presença de limoeiros, cupuaçuzeiros
e árvores de mangostão. As árvores de grande porte têm como principal objetivo o
sombreamento para a cultura de cacau. O Sr. Mineshita explicou que até a formação atual
de seu SAF, houve testes com várias espécies arbóreas, inclusive mogno africano, porém
por ser uma espécie muito competitiva acabou por prejudicar as outras culturas.
Atualmente a fazenda é administrada pelo filho do Sr. Mineshita que está trabalhando em
novas combinações de culturas
Uma técnica de manejo utilizada na fazenda é a poda e posterior incorporação dos
resíduos no solo. Muito utilizada por agricultores, a poda é uma técnica de manejo que
possibilita a entrada de luz no sistema, assim como proporciona matéria orgânica e
nutrientes. Logo, as culturas contribuem para a proteção do solo na estação seca e suas
podas após a colheita fornecem nutrientes para o solo através da decomposição e
participação na ciclagem de nutrientes (EMBRAPA, 2001).
2.4 COOPERATIVA MISTA DE TOMÉ-AÇÚ – CAMTA
As cooperativas agroindustriais, segundo a organização das cooperativas
brasileiras: são aquelas nas quais os produtores rurais se associam para a execução das
etapas que envolvem a agricultura ou a pecuária, tais como a compra de sementes, o
beneficiamento, o armazenamento da produção e o escoamento da safra. Essas tarefas
podem ser executadas pela cooperativa, resultando em maior probabilidade de sucesso na
comercialização do produto (SANTOS, 2011). Nesse sentido, tem-se a cooperativa Mista
de Tomé-açu que está localizada na Rodovia PA 140, nº 13083, Tomé-Açu – Pará. Possui
489 cooperados, segundo o cooperado José Rômulo. Atualmente a cooperativa comporta
mais de 200 cooperados e mais de 2.000 pequenos produtores rurais, além de gerar mais
de 10.000 empregos, diretos e indiretos.
A produção de pimenta-do-reino no município, sendo de responsabilidade da
CAMTA, perfaz 600 toneladas anuais. A estrutura do processamento da pimenta-do-reino
ocorre em quatro estágios: Primeiramente, a pimenta chega seca e faz-se a sua limpeza,
posteriormente ela é peneirada (Figura 3a) para a retirada de talos e impurezas. Em

258
Belém (PA), 11 a 13 de dezembro de 2019
seguida, ocorre a separação dos grãos, para então, partir para o processo de ventilação
(Figura 3b) e posteriormente para o ensacamento. E por fim, o último estágio refere-se ao
depósito de estocagem (Figura 3 c) no qual a vida útil do insumo é de 2 anos.
Figura 3- Processamento. (a) peneiragem; (b) ventilação; (c) ensacamento
A B C Fonte: autores (2019)
Quanto ao cacau, a cooperativa possui uma produção de 800 toneladas e 70%
dessa produção é exportada para o Japão. Segundo o cooperado Rômulo, o cacau gira em
torno da bolsa a uns 7/8 reais, porém poderá variar dependendo do produtor, vale ressaltar
que o seu custo de produção está em torno de 6 reais.
A CAMTA produz e fornece as mudas para os produtores juntamente com
assistência técnica, realizando o acompanhamento no plantio, colheita e processamento,
mantendo boa qualidade no produto. No que tangue a produção total da cooperativa,
quase 90% é exportado para os Estados Unidos, Canadá, Alemanha, Japão e Argentina.
Outros produtos da cooperativa como açaí, abacaxi, cacau, cupuaçu, mandioca e dendê
também possui um bom mercado consumidor local e para a exportação.
Após ser explanado todo processo de produção da pimenta, foi abordado a
produção de óleo vegetais, no qual o principal comprador é a natura, com uma produção
60 toneladas. A extração de óleo ocorre principalmente no cupuaçu, maracujá e andiroba,
o preço do óleo varia de R$ 20,00 a R$ 30,00 e possui uma produção diária de 500 kg de
óleo.
A CAMTA trabalha com 14 tipos de polpa em 3 tipos de qualidade: popular,
médio e especial. Entre os 14 tipos de poupa, existe 4 linhas especiais: açaí, acerola,
abacaxi e cupuaçu. O processo de produção de poupa de fruta começa com a chegada da
fruta in natura. Em seguida é pesada e alocada em câmeras frigorificas, no qual são
acumuladas por uma semana ou até obter-se um grande volume considerável para seu

259
Belém (PA), 11 a 13 de dezembro de 2019
processamento. Posteriormente são encaminhados para tubos de congelamento para ao
final ocorrer o empacotamento. A agroindústria possui dois tubos de congelamento (25
toneladas) e três câmeras frigorificas. São processadas 50 toneladas de polpa diariamente,
de acordo com a demanda e safra do produto. Em relação ao mercado da polpa, a maior
demanda é externa, de 20% a 30% são exportados para o Japão.
A agroindústria de poupa tem como resíduos gerados os efluentes provenientes da
lavagem das frutas, bem como, dos locais de processamento. Foi informado que a
empresa regularmente faz análises dos seus efluentes gerados. Por se tratar de matéria
orgânica, despeja-se diretamente nos rios próximos, apenas com o tratamento preliminar.
Entretanto, as partes das frutas que não são processadas são dispostas a céu aberto em um
terreno da empresa. O intuito da empresa é a compostagem com esses resíduos, que é um
processo biológico aeróbio de tratamento e estabilização de resíduos orgânicos para a
produção do composto, nome dado ao fertilizante orgânico assim produzido. Durante a
compostagem, a matéria orgânica é decomposta principalmente através da ação de
microrganismos e enzimas, resultando na fragmentação gradual e oxidação dos detritos
(BUDZIAK et al., 2004). Entretanto a forma como essa compostagem está sendo
realizada não é ambientalmente adequada, visto o efluente gerado, além de gases. Vale
ressaltar que a agroindústria e o ponto de disposição de resíduos, localizam-se em locais
afastado da população residente. Dessa forma, teoricamente não os atingindo diretamente,
mas indiretamente pela degradação do bem comum a todos.
Diante ao exposto, ficou evidente os benefícios ambientais que os SAF’s agregam
às propriedades e ao município de Tomé-Açú, pois permite a estabilidade da qualidade
dos ecossistemas uma vez que não faz uso de herbicidas, comportam grande
biodiversidade, pois tenta imitar uma floresta natural, possibilita a grande geração de
matéria orgânica, permitindo maior umidade e nutrientes no solo, as árvores de maior
porte podem auxiliar na mitigação das mudanças climáticas por meio do sequestro do
carbono, melhora ainda, a qualidade da água e diminui as chances de erosão e
assoreamento uma vez que o solo fica protegido.
A CAMTA por meio da assistencia que provém aos seus cooperados, permite que
esses indivíduos aprendam novas técnicas de cultivo, além disso, incentiva seus estudos
sobre agricultura no Brasil e no Japão, possibilita a integração da mulher no campo,
realizando a colheita ou ajudando-os em processos burocráticos, além disso, aproxima os

260
Belém (PA), 11 a 13 de dezembro de 2019
cooperados, acarretando a união e o sucesso da instituição. Já no âmbito econômico o
sucesso da cooperativa é visível no estado do Pará, estando presente nas maiores redes de
supermercados do estado e de outros países. A CAMTA possibilita maior geração de
renda, melhorando a qualidade de vida de seus cooperados e a estrutura do município.
4. CONSIDERAÇÕES FINAIS
Com base nas informações dispostas pôde-se constatar que o SAF’s é um sistema
que agrega grande importancia social, ambiental e econômica. Os produtores rurais
podem agir em unidade ou formar cooperativas, como a CAMTA, possibilitando uma
maior captação de recursos econômicos, além disso, no que tange ao âmbito ambiental,
as técnicas utilizadas levam em consideração a sustentabilidade do planeta e o respeito a
capacidade de uso do solo, também, prezam pela adubação verde, protegendo o solo
contra eventuais contaminações por produtos químicos inorgânicos, outro fator
importante é que possibilita elevada biodiversidade, podendo alcançar características
naturais da área. Já no contexto social, o SAF agrega uma maior participação da mulher
no processo de produção rural uma vez que elas podem trabalhar na colheita ou
formulando metas para alcançar maiores mercados, permitindo assim, uma participação
direta e essencial do gênero feminino. Este tipo de sistema traz grande visibilidade para
as localidades que os escolhem e os produtos gerados tem grande aceitação perante seus
consumidores, isto ocorre, porque como já mencionado, provoca impactos ambientais
relativamente pequenos se considerados aos meios de produção convencionais e
atualmente, a população mundial está mais interessadas em questões ambientais e em
produtos cujo seu processo leve em consideração o bem estar dos recursos naturais,
Por fim, ressalta-se que atualmente é de extrema relevância estudos direcionados
ao uso racional dos recursos naturais, já que a manutenção da qualidade do meio ambiente
é essencial para a atual e futuras gerações. Por isso, outros acadêmicos podem dar
continuidade a este trabalhando, mostrando organizações de SAF’s em diferentes biomas,
abordando diferentes técnicas de sucessão ecológica, mostrando outras espécies que
podem ser utilizadas para sombreamento, ou ainda, sobre como preparar o solo para um
futuro SAF’s.

261
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BUDZIAK, Cristiane R. et al. Transformações químicas da matéria orgânica durante a
compostagem de resíduos na indústria madeireira. Embrapa Florestas-Artigo em periódico
indexado (ALICE), 2004.
DE BARROS, Andréa Vieira Lourenço et al. Evolução e percepção dos sistemas agroflorestais
desenvolvidos pelos agricultores nipo-brasileiros do município de Tomé-Açu, estado do
Pará. Embrapa Amazônia Oriental-Artigo em periódico indexado (ALICE), 2009.
DUBOIS, J. C. L. Sistemas agroflorestais na Amazônia: avaliação dos principais avanços e
dificuldades em uma trajetória de duas décadas. Alternativa agroflorestal na Amazônia em
transformação. Brasília: EMBRAPA, p. 171-217, 2009.
EMBRAPA. Caracterização e classificação dos solos do município de Tomé-Açú, PA. ISSN
1517-2201. Belém, PA. 2001.
GOVERNO DO ESTADO DO PARÁ. Estatística municipal: Tomé-Açu. 2011.
IMAZON. Boletim do desmatamento da Amazônia legal – SAD. Imazon.org.br. 2019. Acesso:
15 de outubro de 2019.
KONAGANO et al. Sistema Agroflorestal de Tomé-Açu, Pará – SAFTA: Resumo de
Palestra. X Congresso Brasileiro de Sistemas Agroflorestais (XCBSAF) Cuiabá, outubro de
2016.
SANTOS, Ercílio. Criação de valor econômico em cooperativas agroindustriais. Organizações
Rurais & Agroindustriais, v. 4, n. 1, 2011.

262
Belém (PA), 11 a 13 de dezembro de 2019
ANÁLISE COMPARATIVA DO USO DO PROTEINADO E A INCLUSÃO
PARCIAL DA TORTA DE DENDÊ (ELAEIS GUINEENSIS) EM DIETAS PARA
RUMINANTES DA RAÇA NELORE: CRESCIMENTO E OBTENÇÃO DE
GANHO DE PESO
Halison Felipe Pimenta Almeida1; Adriano Silva de Souza2; Ana Carolina Assmar de
Lima Rabelo3; Hellem Cristina Teixeira Rodrigues4; Carla Lyzandra Linhares5
1 Engenheiro Ambiental. Faculdade Estácio de Belém. [email protected]
2 Engenheiro Ambiental. Faculdade Estácio de Belém. [email protected]
3 Doutoranda em Desenvolvimento Sustentável dó Trópico Úmido (NAEA/UFPA).
Faculdade Estácio de Belém. [email protected]
4 Discente da Especialização em Geotecnologias Aplicadas ao Planejamento Ambiental
e Desenvolvimento Territorial Rural. [email protected]
5 Engenharia Ambiental e Energias Renováveis. Universidade Federal Rural da
Amazônia - UFRA. [email protected]
RESUMO
A pecuária bovina é de grande importância na economia brasileira, considerando que o
Brasil possui um dos maiores rebanhos bovino do mundo, sendo o maior exportador e
segundo maior produtor, entretanto, uma das principais deficiências encontrada na
exploração da pecuária, referente à produção animal, são decorrentes do manejo
alimentar, sanitário e reprodutivo inadequado. Por tanto se torna de fundamental
importância a implementação de inovações tecnológicas e alternativas nutricionais que
possam potencializar o crescimento e obtenção de ganho de peso de maneira eficiente e
saudável, aumentando a produtividade animal. O presente trabalho foi desenvolvido a fim
de realizar uma análise comparativa do uso do Proteinado e a inclusão parcial da Torta de
Dendê em dietas para ruminantes da raça nelore. Foram utilizadas 10 novilhas da raça
nelore com idade entre 12 a 24 meses em um delineamento experimental, no período de
227 dias, com divisão de 2 grupos para a nutrição dos animais utilizando Proteinado e
capim (Brachiaria), Torta de Dendê, Proteinado e Capim (Brachiaria), onde o 1º grupo
utilizou (400g/dia) para suplementação de Proteinado, 2º grupo (300g/dia) Torta de
Dendê e Proteinado, todos os subprodutos aplicados nos grupos foram complementados
com 200 g/dia de sal mineral e capim (Brachiaria). Aplicação dos elementos Proteinado,
Torta de Dendê e sal mineral influenciou positivamente na engorda dos bovinos da
espécie nelore, sendo que as proporções foram consideradas satisfatórias em termo de
economia, quando associadas à utilização da Torta de Dendê.
Palavras-chave: Dietas. Produção Animal. Subprodutos
Área de Interesse do Simpósio: Zootecnia

263
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A pecuária bovina é de grande importância na economia brasileira, visto que o
Brasil possui o segundo maior rebanho bovino do mundo, segundo o Instituto Brasileiro
de Geografia e Estatística (IBGE, 2017).
As principais deficiências encontradas no rebanho paraense são decorrentes de
erros no manejo alimentar, sanitário e reprodutivo adotado nos criatórios e são
intensificados ainda mais pela falta de políticas públicas para os produtores, dificultando
o desenvolvimento da pecuária na Amazônia e no restante do país (AZEVÊDO et al.
2010; MINERVINO et al. 2008;).
Neste cenário, considerando que os gastos com a alimentação de ruminantes em
regimes de confinamento e semi-confinamento, seja um fator limitante para o
desenvolvimento, o aproveitamento de subprodutos de agroindústrias, pode contribuir
para a redução dos gastos com a alimentação em regiões produtoras como o Estado do
Pará (LOUSADA JUNIOR et al. 2005).
Nesse pressuposto, após uma análise realizada por Martha Jr. et al. (2012)
constatou se que esse aumento na produção de bovinos de corte da-se pela eficiência no
aumento de produtividade através de melhorias nos índices zootécnicos. A melhoria
nesses indicies podem ser alcançadas com práticas de manejo, melhorias na nutrição e
genética dos animais.
De acordo com WanZarari e Alimon (2004), com a intensificação da
dendeicultura no Estado do Pará, o aproveitamento de subprodutos como a torta de dendê
surge como alternativa econômica na produção animal, pois pode melhorar o custo-
benefício da formulação de dietas para bovinos, promovendo ainda a conservação
ambiental, ao minimizar os efeitos negativos gerados pelo acúmulo desse resíduo no
ambiente.
Dentre os subprodutos aproveitados na alimentação animal, a torta de dendê
apresenta grande potencial levando em consideração não só a sua rica composição em
nutrientes e teores de proteína e fibras que podem suprir parte dos nutrientes na
alimentação animal, como também a sua disponibilidade durante o ano e o seu baixo
custo, principalmente em regiões de alta produção como as regiões Norte e Nordeste do
Brasil, (COSTA et al. 2006; VIEIRA et al. 2011).

264
Belém (PA), 11 a 13 de dezembro de 2019
Dito isso, este trabalho se propôs a realizar uma analise comparativa do uso do
Proteinado e a inclusão parcial da Torta de Dendê em dietas para ruminantes da raça
nelore, considerando a grande problemática ambiental acerca da adequada destinação dos
subprodutos da enorme produção de óleo, observando o crescimento e engorda dos
animais, com ênfase para a digestibilidade aparente, consumo voluntário e desempenho
de ruminantes criados sob condições tropicais típicas do Nordeste Paraense e
posteriormente verificar a viabilidade econômica sobre o melhor custo-benefício para a
alimentação dos ruminantes entre a utilização da ração de proteinado tradicional e a
proposta de inclusão parcial da torta de dendê.
2. MATERIAL E MÉTODOS
Este trabalho foi realizado na Chacára Colinas, propriedade privada localizada na
PA 150, KM 110, no município de Moju – PA, com as coordenadas geográficas:
“2º40’11.22” W de latitude e “49º5’15.36” S de longitude (Figura 1).
Figura 1 – Mapa de Localização do Município de Moju -PA.
Fonte: Autores, (2019).

265
Belém (PA), 11 a 13 de dezembro de 2019
2.1 TIPO E MÉTODO DE PESQUISA
O trabalho baseou-se em uma profunda pesquisa bibliográfica com o intuito de
obter conhecimentos relativos ao assunto proposto. Também utilizou se de pesquisa de
campo, permitindo extrair dados e informações diretamente da realidade do objeto de
estudo. Para a complementação da pesquisa requereu-se ainda de exame descritivos,
qualitativos e quantitativos. O método de pesquisa utilizado foi exploratório, com intuito
de obter maior proximidade com o universo do objeto de estudo, favorecendo o acesso a
informações que permite orientar a formulação das hipóteses da pesquisa.
2.2 PROCEDIMENTOS ADOTADOS
Na fase experimental da pesquisa, foram utilizados 10 animais da raça Nelore. Os
animais foram divididos em dois grupos de diferentes níveis de peso, com a mesma faixa
etária (12 a 24 meses), cada um com uma combinação de alimentação diferente, sendo:
Grupo 1 alimentado por Proteinado e capim; e Grupo 2 alimentado por Torta de Dendê,
Proteinado e capim. Todos os animais foram identificados com brincos numerados
seqüencialmente de 01 a 10 (Tabela 1) e submetidos à pesagem consecutivamente. Ao
total, cada animal passou por duas pesagens, uma no início (06/10/2018) do experimento
e outra ao final (12/05/2019), após sete meses. A pesagem visa verificar os parâmetros de
ganho de peso para cada grupo.
Tabela 1 – Apresentação dos grupos. Grupo Nº identificação Quantidade
G1- Proteinado 01 – 05 5
G2- Proteinado e Torta de Dendê 06 – 10 5
Fonte: Autores, (2019).
No 1º grupo de ruminantes, com brincos de identificação de 01 a 05 com peso
inicial de 1300 kg e uma média por animais de 260 kg, a alimentação foi conduzida
através de proteinado (sulfato de amônio, flor de enxofre, uréia, sal mineral) e capim,
utilizando uma quantidade de 400grama/dia.
No 2º grupo, com brincos de identificação de 06 a 10 com o peso inicial de 1250
kg e uma média por animal de 250 kg, os animais foram alimentados com torta de dendê,
proteinado e capim, com dosagem de 300 kg/dia.

266
Belém (PA), 11 a 13 de dezembro de 2019
Durante todo o tempo do experimento, cada grupo de ruminantes com cinco
animais esteve separado por piquetes com dimensões de 200x200m (Figura 2).
Importante ressaltar que, para não haver divergência entre os dados, os dois grupos
analisados utilizaram a mesma pastagem do tipo Brachiaria na sua alimentação e ambos
tiveram a sua alimentação complementada com 200 g/dia de sal mineral.
As formas de alimentação praticadas nos grupos 1º (Proteinado) e 2º (Proteinado
e Torta de dendê) são utilizadas com maior freqüência pelos pecuaristas na região norte,
por conta da rentabilidade e qualidade proporcionada por esses subprodutos, por ser uma
região tropical esses tipos de alimentação se adaptou de forma excepcional.
Figura 2 – Mapa de caracterização da Área de Estudo.
Fonte: Autores, (2019).

267
Belém (PA), 11 a 13 de dezembro de 2019
A seguir, apresentam-se comentários gerais dos dois tipos de alimentação
praticados em cada grupo:
GRUPO 1 - Alimentação através de Proteinado: A mistura múltipla conhecida como
proteinado, é a alternativa de suplementação que costuma ter a melhor relação custo-
benefício e adquirido em lojas agropecuárias da região. Em pastagens com boa
disponibilidade forrageira e lotação de 1 UA/ha, possibilita ganhos de peso em torno de
200 a 400 g/cabeça/dia. Exemplos de formulações de sal mineral com uréia, tendo como
base o sal mineral tradicional Ingrediente Formulação 1 Formulação 2 Sulfato de Amônio
(%) 3 - Flor de Enxofre (%) - 1 Ureia (%) 30 30 Sal Mineral (%) 67 69 Total 100 100.
GRUPO 2 - Alimentação através da intercalação de Pastagem, Proteinado e
Torta de Dendê: a pastagem composta por capim é volumosa devido ser natural e de
fácil manejo, é considerada uma fonte econômica de nutrientes para alimentação destes
animais, porém com algumas limitações quanto ao clima, relevo, qualidade do solo e
acesso, além de requerer cuidados ao selecionar as forrageiras a serem cultivadas, pois
precisam ser analisados todos os fatores anteriormente citados, de acordo com as
informações da Secretaria Municipal de Mojú.
3. RESULTADOS E DISCUSSÃO
Na Tabela 2, realizou se a comparação dos principais parâmetros sobre
crescimento e engorda bovina. Portanto foi executada a junção do peso inicial e final para
obter um amplo conhecimento de cada grupo, onde é possível observar o comportamento
e o desempenho de cada grupo após a aplicação dos subprodutos (Proteinado; Proteinado
e Torta de Dendê), evidenciando resultados sobre adequação dos animais.
Os resultados alcançados (Tabela 2) apontam que o peso médio inicial de cada
bovino era 260 e 250kgs nos grupos 1 e 2, respectivamente. Ao final do experimento, o
peso médio por bovino foi 286 e 300kgs nos grupos 1 e 2, respectivamente.

268
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 2 – Comparação dos Grupos.
Grupo Nº
identificação Dosagem
(kg/bovino.
Dia)
Média inicial
(kg/bovino) Média final
(kg/bovino) Peso total
do grupo
inicial (kg)
Peso total
do grupo
final (kg)
G1-
Proteinado 51 – 56 400 260 286 1300 1430
G2-
Proteinado e
Torta de Dendê
61 – 65 300 250 300 1250 1500
Fonte: Autores, (2019).
Figura 3 – Comparação do peso médio (Inicial/Final) de cada bovino.
Fonte: Autores, (2019).

269
Belém (PA), 11 a 13 de dezembro de 2019
Figura 4 – Comparação do peso total (Inicial/Final) dos grupos.
Fonte: Autores, (2019).
De acordo com as Figuras 03 e 04, é notável que ambos os grupos apresentaram
ótima adequação aos subprodutos aplicados, no entanto, o último grupo (Grupo 2)
expressou uma adequação maior, potencializando o crescimento e ganho de peso dos
bovinos de maneira eficiente, saudável e sustentável. A análise econômica das opções
avaliadas apontou que a torta de dendê é mais viável economicamente quando comparada
ao proteinado, pois expressa um valor unitário em torno 40 reais para 60 Kg, enquanto, o
proteinado custa 85 reais para 30 Kg (Tabela 3).
Tabela 3 - Analise Econômica
Ração Saco (kg) Valor unitário (R$)
Proteinado 30 85
Torta de dendê 60 40
Sal mineral 25 25 Fonte: Autores, (2010).
A Tabela 4 apresenta o ganho de peso diário e mensal dos bovinos. O grupo 1
sujeito a aplicação de proteinado e sal mineral apresentou um rendimento de ganho de
peso diário de 0.73 Kg/dia e um ganho de peso mensal de 21.9 Kg/mês. Como o estudo

270
Belém (PA), 11 a 13 de dezembro de 2019
foi realizado em um período de constantes chuvas, os animais não apresentaram uma boa
disgestibilidade em relação ao subproduto (Proteinado), devido o mesmo ficar umedecido
no cocho.
O Grupo 2, sujeito a aplicação dos subprodutos, Proteinado e Torta de Dendê,
expressou um rendimento surpreendente, evidenciando um desempenho excepcional,
com ganho de peso diário de 1, 10 kg/dia por bovino. Como foi realizado em uma área
de estudo de regime a pasto, notou se que o grupo 2 aferiu um rendimento aproximado
ao regime de confinamento, onde o mesmo ganha em média 1, 50kg/dia, devido os
ruminantes se alimentarem sobre condições adequada para ganho de peso.
Tabela 4 – Ganho de Peso.
Grupo Nº
Identificação Ganho de peso
(kg/dia) Ganho de peso
(kg/mês)
G1- Proteinado 51 – 55 0, 73 21, 9
G2- Proteinado e Torta de
Dendê 61 – 65 1, 10 33
Fonte: Autores, (2019).
De acordo com uma breve comparação entre os grupos 1 e 2 (Tabela 4), é notório
que o grupo 2 expôs um desempenho maior. O mesmo apresentou um rendimento no
ganho de peso diário de 1, 10 Kg/dia e um ganho de peso mensal de 33 Kg/mensal,
evidenciando que o período de chuvas constantes não influenciou na adequação dos
animais a inclusão parcial da torta de dendê, a inclusão do mesmo favoreceu no
comportamento dos animais, deixando eles mais dóceis, por conta do agradável cheiro da
torta, ostentando uma boa digestibilidade, demonstrando que esse subproduto pode ser
aplicado pelos pecuaristas sem nenhum receio.
4. CONCLUSÃO
A inclusão da torta de dendê incrementada para nutrição bovina comparada com
o proteinado (Fonte energética convencional) expressou resultados bastante significativos

271
Belém (PA), 11 a 13 de dezembro de 2019
e promissores em relação ao custo com a alimentação e ganho de peso dos ruminantes.
Uma vez que a inclusão da torta de dendê mostrou ser viável economicamente e pode ser
mais uma alternativa presente para pequenos e grandes produtores rurais. Com isso
realizando a destinação adequada dos resíduos gerados pelas grandes indústrias extratoras
de óleo vegetal, promovendo uma iniciativa de desenvolvimento sustentável, atendendo
aspectos em termos econômicos, social e ambiental.

272
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
AZEVÊDO, D. M. M. R. et al. A ovinocultura no mundo e no Brasil: uma realidade,
2010. Disponível em. Acesso em: 15/03/2019
COSTA, D. A. Avaliação nutricional da torta de dendê para suplementação alimentar de
ruminantes na Amazônia Oriental. 2006. 60 f. Dissertação (Mestrado em Ciência Animal)
- Centro de Ciências Agrárias, UFPA. Universidade Federal do Pará, Belém, 2006.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA (IBGE) – Censo
Agro2017.Disponívelem:https://censos.ibge.gov.br/agro/2017/templates/censo_agro/res
ultadosagro/agricultura.ht ml?localidade=15&tema=76288. Acesso em 16/03/2019
LOUSADA JUNIOR, J.E.; MIRANDA NEIVA, J.N.; RODRIGUEZ, N.M. et al.
Consumo e digestibilidade de subprodutos do processamento de frutas em ovinos. Revista
Brasileira de Zootecnia, v.34, n.2, p.659-669, 2005. Acesso em 16/03/2019
MARTHA Jr, G. B.; ALVES, E.; CONTINI, E. Land-saving approaches and beef
production growth in Brazil. Agricultural Systems, v.110, p. 173-177, 2012.
VIEIRA, C. A.; USO DA TORTA DE DENDÊ NA ALIMENTAÇÃO DE
RUMINANTES, 2011. Disponível em: . Acesso em: 16/03/2019
WAN ZAHARI, M.; ALIMON, A. R. Use of palm kernel cake and oil palm by-products
in compound fedd. Palm Oil developments, V.8, n.40, P.5-9, 2004. Acesso em:
10/09/2018.

273
Belém (PA), 11 a 13 de dezembro de 2019
AVISTAMENTO DA AVIFAUNA NO MUNICÍPIO DE JURUTI, BAIXO
AMAZONAS, ESTADO DO PARÁ, BRASIL
Neuder Wesley França da Silva
Mestre em Saúde e Produção Animal na Amazônia. Secretaria de Estado de Saúde
Pública. E-mail: [email protected]
RESUMO
O município de Juruti, por localizar-se na mesorregião do baixo Amazonas, estado do
Pará, possui atributos ambientais favoráveis para conter inúmeras espécies de aves.
Detentor de grande área de várzea, além da área de terra firme, a região alberga variada
concentração de aves nativas e visitantes. Desta feita, objetivou-se identificar as
principais espécies da avifauna através do registro visual, o qual corresponde em método
distinto e complementar de censo por observação direta em estudos ornitológicos. Assim
sendo foi realizado estudo durante 10 dias no período de estiagem em 2013 e 10 dias no
período de maior pluviosidade em 2014, abrangendo área de terra firme e de várzea na
região, por meio de registro visual (a olho nu) e fotográfico em máquina digital. Desta
forma, obteve-se registro de 51 espécies de aves de 29 famílias, com representantes de
aves aquáticas, comumente presentes em leitos de rios e várzea da região e que atuam
como indicadoras da qualidade da água, bem como a presença de aves residentes e
visitante sazonal como Pandion haliaetus, que indicam potencial e qualidade da avifauna
do município de Juruti. Destaca-se ainda a presença comum de aves de hábitos florestais
em áreas de ocupação antrópicas e realizando forrageamento próximos de animais de
produção como bovinos e suínos.
Palavras-chave: Avifauna. Espécies. Baixo Amazonas.
Área de interesse do Simpósio: Biodiversidade.

274
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O Brasil apresenta uma das mais ricas avifaunas do mundo, com estimativas de
1.731 espécies, aproximadamente 10% ameaçadas de extinção, sendo a Amazônia com
maior número de espécies (MARINI; GARCIA, 2005; SICK, 1993). A identificação de
espécies contribui para o entendimento do potencial ecológico e qualidade do
ecossistema, sendo o registro visual um método distinto e complementar de censo por
observação direta e convencional em estudos ornitológicos de reconhecimento qualitativo
(RODRIGUES et al., 2005; STRAUBE; URBEN-FILHO, 2005). Neste sentido foi
realizado registro visual para identificar a diversidade da avifauna no município de Juruti,
Estado do Pará, Brasil.
2. MATERIAL E MÉTODOS
O estudo foi desenvolvido no município de Juruti, mesorregião do Baixo
Amazonas, Estado do Pará, durante 10 dias do mês de novembro de 2013,
correspondendo ao período de estiagem e 10 dias do mês de março de 2014, referente à
período maior pluviosidade na região, embora a pluviosidade seja irregular durante o ano
no município (PARÁ, 2016).
Os registros abrangeram área de terra firme e áreas de várzeas que apresentam
variações significativas de inundações, bem como determinadas áreas de ocupações
antrópicas. Para tanto, utilizou-se metodologia tipo visual (a olho nu), com registro
fotográfico em máquina digital Nikon P510 de 16 megapixels de resolução e zoom óptico
de 42X.
Registraram-se apenas indivíduos e ninhos acessíveis visualmente, sem qualquer
aproximação física que produzisse perturbação animal, bem como facilitasse a captura de
imagens sem desfoco fotográfico para auxiliar na identificação de espécies. Os métodos
de deslocamentos foram através de caminhadas e por meio de lanchas (áreas ribeirinhas)
utilizadas no período diurno.
A identificação das espécies foi realizada por intermédio de referências básicas
sobre aves encontradas por SICK (1997); GWYNNE et al. (2010) e BINI (2009). Para a
sistematização de nomenclatura das espécies, bem como status de ocorrência, seguiu-se
a lista das aves do Brasil, organizada pelo Comitê Brasileiro de Registros Ornitológicos
de 2015 (REVISTA BRASILEIRA DE ORNITOLOGIA, 2015).

275
Belém (PA), 11 a 13 de dezembro de 2019
Embora qualquer espécie avistada tenham importância do ponto de vista
ecológico, destacou-se nesse estudo a descrição de algumas espécies de importância no
aspecto de geodistribuição.
3. RESULTADOS E DISCUSSÃO
Foram registradas 51 espécies de aves, as quais estão distribuídas em 29 famílias,
sendo que 8 famílias (27,58%) representam aves aquáticas (Tabela 1). Apenas um
indivíduo foi classificado pelo gênero Sprophika, bem como nenhuma espécie registrada
no estudo consta na lista Estadual da Fauna Brasileira Ameaçada de Extinção (COEMA,
2007).
Tabela 1: Lista de espécies de aves registradas visualmente no município de Juruti, Pará.
Família, espécie, nomes populares e status (R= evidências de reprodução no país
disponíveis; VS = Visitante sazonal oriundo do hemisfério norte). Família Espécie Nome popular Status
Accipitridae
Buteo nitidus gavião-pedrês R
Busarellus nigricollis gavião-belo R
Heterospizias meridionalis gavião-caboclo R
Rupornis magnirostris gavião-carijó R
Alcedinidae
Chloroceryle amazona martim-pescador-verde R Megaceryle torquata martim-pescador-grande R
Chloroceryle americana martim-pescador-pequeno R
Aramidae Aramus guarauna carão R
Ardeidae
Ardea alba garça-branca-grande R
Pilherodius pileatus garça-real R
Egretta thula garça-branca-pequena R
Ardea cocoi garça-moura R
Bubulcus ibis garça-vaqueira R
Tigrisoma lineatum socó-boi R
Bucconidae Monasa nigrifrons chora-chuva-preto R
Nonnula rubecula macuru R
Cathartidae Coragyps atratus urubu-da-cabeça-preta R
Charadriidae Vanellus chilensis quero-quero R
Ciconiidae Mycteria americana cabeça-seca R Columbidae Columbina passerina rolinha-cinzenta R Cteridae Gymnomystax mexicanus iratauá-grande R Dendrocolaptidae Nasica longirostris arapaçu-de-bico-comprido R
(continua)

276
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 2: Lista de espécies de aves registradas visualmente no município de Juruti, Pará.
Família, espécie, nomes populares e status (R: evidências de reprodução no país
disponíveis; VS: Visitante sazonal oriundo do hemisfério norte). Família Espécie Nome popular Status
Falconidae Caracara plancus caracará R
Milvago chimachima carrapateiro R
Furnariidae Furnarius minor joãozinho R Galbulidae Galbula galbula ariramba-de-cauda-verde R Hirundinidae Tachycineta albiventer andorinha-do-rio R Jacanidae Jacana jacana jaçanã R Pandionidae Pandion haliaetus águia-pescadora VS Phalacrocoracidae Phalacrocorax brasilianus Biguá
Picidae
Dryocopus lineatus pica-pau-de-banda-branca R Colaptes punctigula pica-pau-de-peito-pontilhado R
Campephilus melanoleucos pica-pau-de-topete-vermelho R
Psittacidae Forpus passerinus tuim-santo R Psittacara leucophthalmus periquitão R
Ramphastidae Pteroglossus aracari araçari-de-bico-branco R Rynchopidae Rynchops niger talha-mar R Sternidae Phaetusa simplex trinta-réis-grande R
Thraupidae
Tangara episcopus sanhaço-da-amazônia R
Sporophila lineola bigodinho R
Sicalis columbiana canário-do-amazonas R
Paroaria gularis cardeal-da-amazônia R
Tangara mexicana saíra-de-bando R
Sporophila sp. - R
Threskiornithidae Theristicus caudatus curicaca R Tityridae Tityra semifasciata anambé-branco-de-máscara-negra R Trochilidae Amazilia fimbriata beija-flor-da-garganta-verde R Troglodytidae Troglodytes musculus corruíra R
Tyrannidae
Pitangus sulphuratus bem-te-vi R
Tyrannus savana tesourinha R
Suiriri suiriri suiriri-cinzento R
Fonte: Revista Brasileira de Ornitologia, 2015
Dos avistamentos, observou-se Pandion haliaetus em março de 2013, no ponto
mais elevado de uma torre de telefonia localizada na região urbana de Juruti. Pandion
haliaetus (Figura 1, A) é visitante sazonal oriundo do hemisfério norte que realiza seu
movimento migratório para “invernar” no Brasil (REVISTA BRASILEIRA DE
ORNITOLOGIA, 2015). As demais espécies avistadas são consideradas residentes no
país, como as presente na Figura 1 (B), (C), (D) e (E).

277
Belém (PA), 11 a 13 de dezembro de 2019
Figura 1- Espécies de aves registradas em Juruti-Pará. A: Pandion haliaetus; B: Nasica
longirostris; C: Dryocopus lineatus; D: Tityra semifasciata; E: Phalacrocorax
brasilianus
Fonte: autoria própria, 2013, 2014
Conforme SICK (1993) noventa e dois por cento das aves brasileiras são espécies
residentes, sendo apenas 8% espécies migrantes, sendo que o maior número de espécies
de aves residentes (1.300) e a maior taxa de endemismo (20%) ocorrem na Amazônia
(MITTERMEIER et al., 2003).
Em relação a ave migratória, comumente avistou-se Tyrannus savana, encontrada desde
Uruguai, Argentina, Brasil, Bolívia, até Venezuela e Colômbia, no extremo norte da
América do Sul (ZIMMER, 1937; PINTO, 1944), tendo sua reprodução durante a
primavera e verão no centro e sul de sua área de distribuição geográfica, migrando para
o norte da América do Sul, onde encontra sua área de invernada (ZIMER, 1937;
NEGRET; NEGRET, 1981; SICK, 1983, 1997; CHESSER, 1997). Outra ave aquática
migratória avistada foi Rynchops niger.
Em área ribeirinha identificou-se uma colônia de Bubulcus ibis com poucos
indivíduos Phalacrocorax brasilianus (Figura 2), e B. ibis associado a forrageamento de

278
Belém (PA), 11 a 13 de dezembro de 2019
gado. Heterospizias meridionalis foi registrado em momento de acasalamento em
vegetação de margem várzea (Figura 2).
Figura 2- A: Colônia de Bubulcus ibis com presença de Phalacrocorax brasilianus; B:
Heterospizias meridionalis em acasalamento, área de várzea, Juruti-Pará.
Fonte: autoria própria, 2013, 2014
Furnarius minor foi avistado em nidificação, e um filho de Coragyps atratus em
oco de árvore na ausência de seus pais. Diversos Sicalis columbiana foram avistados em
ambientes abertos associados à várzea local e comumente próximos de habitações
antrópicas.
Registraram-se vários grupos de Forpus passerinus, usualmente presente em área
de várzea aberta e próximo de habitações antrópicas e Nasica longirostris que possui
hábito florestal, porém foi encontrado em comunidade antrópica em área de várzea, sendo
que o avistamento de Mycteria americana se justifica pela sua preferência por terrenos
alagados com gramíneas, conforme descrito por MORRISON et al (2008).
4. CONCLUSÃO
A avifauna observada no município de Juruti é ampla e diversificada. A presença
de visitante sazonal indica o potencial e qualidade da avifauna local, havendo a
necessidade de estudo de monitoramento para avaliar o padrão de extinção e colonização
de aves residentes no município, principalmente de aves aquáticas que são
significativamente representantes dos leitos de rios e várzea da região, bem como
indicadoras da qualidade da água, possivelmente pela oferta de pesca para as aves.
Ademais evidenciou-se a aproximação de aves de hábito florestal em áreas de ocupação

279
Belém (PA), 11 a 13 de dezembro de 2019
antrópica, além de espécies forrageando próximos de animais de produção,
principalmente com bovinos e suínos.

280
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BINI, E. Aves do Brasil: Guia Prático. Lages: Homem-pássaro publicações, 233p, 2009.
CHESSER, R. Terry. Patterns of Seasonal and Geographical Distribution of Austral
Migrant Flycatchers (Tyrannidae) in Bolivia. Ornithological Monographs, [s.l.], n. 48,
p.171-204, jan. 1997. University of California Press. http://dx.doi.org/10.2307/40157534.
Disponível em: <https://www.jstor.org/stable/40157534?seq=1>. Acesso em: 06 out.
2019.
GWYNNER, John A. et al. Aves do Brasil: Pantanal & Cerrado. São Paulo: Horizonte,
2010. 322 p.
MARINI, M.A.; GARCIA, F.I. Conservação de aves no Brasil. Megadiversidade, v.1,
n.1, p. 95-102, 2005.
MITTERMEIER, R. A. et al. Wilderness and biodiversity conservation. 2003. Disponível
em: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC193557/pdf/10010309.pdf. Acesso
em: 01 out. 2018.
MORRISON, R. I. G., et al. Aves migratórias no Pantanal: distribuição de aves limícolas
neárticas e outras espécies aquáticas no Pantanal. Brasília: WWF Brasil, 2008.
NEGRET, A. J.; NEGRET, R. As aves migratórias do Distrito Federal. Instituto
Brasileiro de Desenvolvimento Florestal, Brasília, 1981.
PARÁ. Fundação Amazônia de Amparo A Estudos e Pesquisas. Diretoria de Estatística
e de Tecnologia e Gestão da Informação. Estatísticas Municipais Paraenses: Juruti.
Belém: [s.n.], 2016. 59 p. Disponível em:
http://www.fapespa.pa.gov.br/upload/Arquivo/anexo /1265.pdf?id=1536470490. Acesso
em: 01 out. 2018.
PINTO, O. M. Catálago das aves do Brasil e lista dos exemplares existentes na coleção
do Departamento de Zoologia - 2ª parte. Publicação do Departamento de Zoologia, São
Paulo, 1944.
RESOLUÇÃO COEMA, nº 54 de 24 de outubro de 2007. Homologa a lista de espécies
da flora e da fauna ameaçadas no Estado do Pará. Disponível em:
https://www.semas.pa.gov.br/ 2007/10/24/ 10059/. Acesso em: Acesso em: 01 out. 2019.
REVISTA BRASILEIRA DE ORNITOLOGIA. Pará: [s.n.], v. 23, 2015. Disponível em:
www.revbrasilornitol.com.br › BJO › issue › download › pdf_888. Acesso em: 06 out.
2019.
RODRIGUES, Marcos et al. Aves do Parque Nacional da Serra do Cipó: o Vale do Rio
Cipó, Minas Gerais, Brasil. Revista Brasileira de Zoologia, [s.l.], v. 22, n. 2, p.326-338,
jun. 2005. FapUNIFESP (SciELO). http://dx.doi.org/10.1590/s0101-
81752005000200005.

281
Belém (PA), 11 a 13 de dezembro de 2019
SICK, H. Birds in Brazil: A natural history. Princeton University Press. Princeton, EUA,
1993.
SICK, H. Ornitologia brasileira. Nova Fronteira, Rio de Janeiro, 1997.
STRAUBE, F. C.; URBEN-FILHO, A. Avifauna da Reserva Natural Salto Morato
(Guaraqueçaba, Paraná). Atualidades Ornitológicas, 124: 12-32, 2005.
ZIMMER, J. T. Stuties of peruvian birds: Note on the genera muscivora, Tyrannus,
empidonomus, and sirustes, with furth notes on kipolegus American Museum Novitates,
New York City, p.1-28, 1937.

282
Belém (PA), 11 a 13 de dezembro de 2019
CARACTERÍSTICAS FÍSICO-QUÍMICAS E BIOMASSA DO
MICROFITOPLÂNCTON EM DOIS MUNICÍPIOS DO NORDESTE
PARAENSE
Ariel Romeiro Pereira¹; Rosildo Santos Paiva².
1Graduanda em Licenciatura em Ciências Biológicas. Universidade Federal do Pará.
2Professor Doutor. Universidade Federal do Pará. [email protected]
RESUMO
Estuários são caracterizados como zonas de transição localizadas entre o ecossistema
limnético e marinho, onde persistem condições de instabilidade, com períodos de maior
influência terrestre e outros de maior influência oceânica. São ambientes altamente
produtivos pela disponibilidade de sais nutrientes e produtores primários, sendo em sua
grande maioria a comunidade fitoplanctônica. O estudo visa determinar a variação físico-
química da água e biomassa das comunidades fitoplanctônicas. Foram obtidos em cinco
pontos distintos no estuário rio Pacamorema (Curuçá – PA) e sete pontos no rio
Mocajubaju (São João da Ponta – PA), entre setembro e outubro de 2018, no período
menos chuvoso das regiões. A biomassa foi calculada a partir da determinação da
densidade fitoplanctônica (org/L) segundo o método de Utermohl. Foram medidos in situ,
com auxílio de uma sonda HANNA HI9828, a temperatura, salinidade, condutividade
elétrica, pH, sólidos totais dissolvido e oxigênio dissolvido em porcentagem (%) e PPM.
O parâmetro com maior diferença entre os dois municípios foi o de Oxigênio Dissolvido
em PPM, onde em Curuçá a máxima e mínima foram de 10,43 PPM e 9,67 PPM
respectivamente, e em São João da Ponta a máxima foi de 0,85 PPM e a mínima 0,21
PPM, ambas tendo sua mínima no ponto inicial. Nos outros parâmetros não houveram
diferenças significativas, a máxima de Salinidade em nos dois locais foram de 14,704 PSI
em Curuçá e 14,662 PSI em São João da ponta. As médias de temperatura foram de 29ºC,
as máximas de pH variaram entre 7,91 em Curuçá e 7,28 em São João da Ponta. O
parâmetro de Sólidos Totais Dissolvidos teve máxima de 509 nos dois rios de estudo e a
Condutividade Elétrica variou pouco entre os locais, sendo máxima de 1019 e mínima de
1008 no Rio Pacamorema e máxima de 1018 e mínima de 1015 em São João da Ponta. A
densidade do fitoplâncton teve máxima de 39.704 org/L e mínima de 28.489 org/L em
Curuçá e máxima de 56.148 org/L e mínima de 49.555 org/L em São João da Ponta.
Apesar do parâmetro de Oxigênio Dissolvido em São João da Ponta ter sido menor que o
de Curuçá, observamos uma maior densidade do microfitoplâncton nesse local, podendo
estar relacionados com a disponibilidade de oxigênio na água. Usando testes estatíscos
fFoi observado diferenças significativas entre as áreas nos parâmetros de Densidade do
Fitoplâncton e Oxigênio Dissolvido, (p<0,05).
Palavras-chave: Estuário. Fitoplâncton. Produtividade.
Área de Interesse do Simpósio: Biodiversidade

283
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Os estuários são caracterizados como zonas de transição localizadas entre o
ecossistema limnético e marinho, onde persistem condições de instabilidade, havendo
ocorrência de constantes alterações determinadas pelos parâmetros físicos e químicos,
com períodos de maior influência terrestre e outros de maior influência oceânica
(Eskinazi-Leça et al, 2004). O conceito de estuário vem de Pritchard (1967) que define
“um corpo de água costeiro, semifechado que apresenta conexão livre com o mar aberto,
e no interior do qual a água do mar é mensuravelmente diluída pela água doce, derivada
da drenagem terrestre”.
São consideradas áreas altamente produtivas, tanto em função da grande
disponibilidade de sais nutrientes como em função da presença de vários produtores
primários, destacando-se dentre estes a comunidade fitoplactônica. Este ambiente pode
contribuir tanto para a conservação da biota aquática como para a subsistência de seres
humanos que têm no estuário e no manguezal a sua sobrevivência (Cardoso, 2009).
O fitoplâncton pode ser definido como um aglomerado de organismos
fotossintetizantes em que a capacidade de locomoção na coluna d’água é praticamente
inexistente ou quando esta ocorre, é de modo deficiente.Esses organismos são
encontrados em todos os lagos, rios de baixa correnteza, estuários e oceanos. Os
organismos que formam o fitoplâncton são provavelmente os organismos
fotossintetizantes mais comuns no mundo (Horne, 1994).
A comunidade fitoplactônica é amplamente utilizada em estudos de
biomonitoramento de corpos hídricos em função da sua capacidade ecológica em
responder rapidamente a qualquer estímulo externo, como por exemplo, aos processos de
poluição e contaminação do meio aquático (Carvalho, 2003). Estes organismos são muito
utilizados como bioindicadores da qualidade ambiental (Aquino et al., 2011; Lopes, 2007;
Pereira, 2007). A distribuição dos organismos fitoplanctônicos nos estuários é
influenciada por fatores bióticos (herbivoria) a abióticos tais como luz, temperatura,
salinidade, PH e disponibilidade de nutrientes que interferem no seu crescimento e
fisiologia (MELO-MAGALHÃES 2004).

284
Belém (PA), 11 a 13 de dezembro de 2019
As alterações na composição das comunidades fitoplanctônicas, assim como na
densidade ou volume dos organismos, são bons indícios de alterações no ambiente, posto
que a distribuição das espécies no ambiente reflete aspectos da variação da qualidade
ambiental. Por isso, os estudos de parâmetros bióticos avaliam mudanças ambientais ao
longo do tempo, ao contrário das medidas físico-químicas, que somente refletem o
momento da coleta (Salmaso et al., 2012; Carraro, 2009; Margalef, 1983). Com isso, a
determinação do comportamento e constituição do fitoplâncton é um método eficaz para
se avaliar a conservação do ambiente, pela resposta dos organismos às mudanças físico-
químicas da água (Round,1973).
O objetivo geral do presente estudo apresenta a variação da densidade visa
determinar a composição da microficoflora planctônica (org/l) e das características físico-
químicas da água dos estuários dos rios Pacamorema (Curuçá) e Mocajuba (São João da
Ponta) - Pará.E os objetivos específicos são determinar a variação espacial dos parâmetros
físico-químicos e a variação espacial da biomassa fitoplanctônica (org/l) das águas dos
estuários dos rios Pacamorema (Curuçá) e Mocajuba (São João da Ponta) – Pará.
2. METODOLOGIA
Os estuários objetos de estudo estão localizados em dois municípios: São João da
Ponta - tem uma área de 195,90 km², sendo que destes, 57,50 km² são florestas. A sede
do município localiza-se entre os paralelos 00º50‟59” Lat. S / 47º55‟12” Long. W.
Localiza-se na mesorregião do Nordeste Paraense e microrregião do Salgado, está a 132
km de Belém. Como unidade de conservação o município possui a Reserva Extrativista
Marinha de São João da Ponta. O clima no município é equatorial amazônico, com
temperaturas elevadas, média anual de 26º C, com chuvas intensas de janeiro a junho. Na
região foram utilizados cinco pontos de coletas (QUADRO 1) ao longo do rio Mocajuba,
sendo suas localidades:

285
Belém (PA), 11 a 13 de dezembro de 2019
Quadro 1 – Localização dos Pontos de Coleta do Rio Mocajuba, São João da Ponta.
Fonte: Rosildo Santos Paiva, Ariel Romeiro Pereira.
Curuçá - tem uma área de 672,70 km², sendo que destes 200,70 km² são florestas.
A sede do município localiza-se entre os paralelos 00º43‟48” Lat. S / 47º51‟06” Long.
W. Localiza-se na mesorregião do Nordeste Paraense e microrregião do Salgado, está a
132 km de Belém. Possui a unidades de conservação Reserva Extrativista Marinha Mãe
Grande de Curuçá. O município possui clima equatorial amazônico, com temperatura
média de 27º C, baixa amplitude térmica e precipitações pluviométricas abundante, sendo
os primeiros seis meses do ano os mais chuvosos. Na região foram utilizados cinco pontos
de coleta (QUADRO 2) ao longo do rio Pacamorema, sendo suas localidades:
Quadro 2 – Localização dos pontos de coleta do Rio Pacamorema, em Curuçá
Fonte: Rosildo Santos Paiva, Ariel Romeiro Pereira.
Os parâmetros físico-químicos da água: temperatura, potencial hidrogeniônico,
sólidos totais dissolvidos, condutividade elétrica, saturação de OD (%), Oxigênio
Dissolvido (ppm) e salinidade foram medidos in situ com o auxílio de uma sonda
multiparamétrica HANNA HI 9828®.
Para à adeterminação da densidade, foram coletadas amostras da subsuperfície da
água, em seguida, acondicionadas em frascos de polietileno com capacidade de 250ml e
preservadas com formol neutro a uma concentração final a 4%. A análise quantitativa foi
realizada segundo o método de sedimentação de Uthermöhl (1958). As amostras foram

286
Belém (PA), 11 a 13 de dezembro de 2019
sedimentadas por um período de 24 horas e analisadas em microscópio invertido e os
resultados expressos em número de organismos por litro. Na análise dos dados, para
buscar diferenças significativas, foi aplicado um teste estatístico T. Os organismos
unicelulares, filamentos, tricomas, colônias e cenóbios foram considerados como um
único indivíduo.
3. RESULTADOS E DISCUSSÃO
O estudo foi feito com um momento de coleta em cada um dos dois municípios,
ambas durante o período menos chuvoso da região, as coletas feitas ocorreram durante o
segundo semestre de 2018. Os dois municípios em estudo são Curuçá e São João da Ponta.
Os resultados serão apresentados na forma de gráficos nas figuras 1 e 2.
A maior diferença entre os dois municípios foi no parâmetro de Oxigênio
Dissolvido em PPM, enquanto em Curuçá a máxima foi de 10,43 PPM, a mínima 9,67
PPM e o desvio padrão 1,2, em São João da Ponta a máxima foi 0,85, a mínima 0,21, com
um desvio padrão de 0,22. A Salinidade não sofreu variação uma grande muito
significante entre os dois locais, a máxima no rio Pacamorema e no rio Mocajuba
atingiram 14,704 PSI e 14,662 respectivamente, enquanto as mínimas variaram entre
14,681 em Curuçá e 14,646 em São João da Ponta, em ambas o desvio padrão foi igual a
0 (zero). A Condutividade Elétrica variou pouco entre os dois estuários, em Curuçá a
máxima foi de 1019 e a mínima 1008 e em São João da Ponta foi de 1018 e 1015, no
entanto, o desvio padrão obteve diferença significativa, sendo 3,54 no estuário do Rio
Pacamorema e 1,02 no estuário do rio Mocajuba, mostrando que em São João da Ponta
houve uma maior homogeneidade dos dados. O parâmetro de Sólidos Totais Dissolvidos
teve máxima de 509 nos dois rios e mínima de 504 no rio Pacamorema, com um desvio
padrão de 1,62, e 507 no rio Mocajuba, com um desvio padrão de 0,75. O pH sofreu
variação de 7,91 a 7,79 em Curuçá e 7,28 e 7,15 em São João da Ponta, ambas com desvio
padrão igual a 0 (zero). Enquanto a densidade fitoplanctônica em Curuçá teve máxima de
39.704 org/L, mínima de 28.489 org/L e desvio padrão de 3.956, e em São João da Ponta
teve máxima de 56.148 org/L, mínima de 49.555 org/L e desvio padrão de 2.284.
Em seu estudo, na foz do rio Guamá, em Belém – PAd, a Rocha Neto et. al. (2016)
constataramou que, apesar de seus dados não obterem uma variabilidade

287
Belém (PA), 11 a 13 de dezembro de 2019
variaçãosignificativa, a maioria dos parâmetros apresentaram maiores valores durante o
período de menor pluviosidade e, o seu estudo, realizado na foz do rio Guamá, em Belém
– PA, constataram salinidade menor que um (1,0) durante todo o período de estudo, com
valores mais elevados no mês de setembro, no período de menor regime de chuvas. No
presente estudoNeste estudo, realizado nos estuários do rio Mocajuba (São João da Ponta)
e rio Pacamorema (Curuçá) a salinidade se manteve entre 14 PSI, podendo também ter
sido influenciada pelo baixo regime de chuvas. Estudos realizados por Pinheiro (1987) e
Paiva (2006) constataram que, no estuário da região amazônica, as altas descargas fluviais
obstruem de maneira significativa a entrada de águas salobras vindas do oceano. Portanto,
além da localidade dos dois estuários do presente estudo, o baixo índice de pluviosidade
da região no período de coleta influenciou na alta salinidade de ambos.
A maior diferença entre os dois estuários se deu no parâmetro de Oxigênio
Dissolvido (PPM). Segundo o estudo que Macêdo & Costa (1978) desenvolveram, onde
criaram um sistema de classificação parar qualificar a qualidade hídrica segundo as taxas
de oxigênio, determinando as categorias: zona supersaturada (> 100%); zona saturada (75
a 100%); zona de baixa-saturação (50 a 75%); zona semipoluída (25 a 50%) e zona
poluída (< 25%); as águas do estuário do rio Mocajuba se qualificam como poluídas,
sendo a máxima de 0,85 PPM (11,3%) e as águas do estuário do rio Pacamorema como
supersaturada, sendo a máxima 10,43 PPM (139,4%).
No entanto, no estuário do rio Mocajuba, as taxas de densidade do
microfitoplâncton foram maiores do que no estuário do rio Pacamorema. Estudar a
biomassa primária de uma região é relevante, pois, a partir da sua mensuração, é possível
determinar a quantidade de matéria orgânica disponível em um determinado local e traçar
um perfil sobre o caráter ecológico da região (PASSAVANTE & FEITOSA, 1989).Em
seu estudo, Paiva (2006) atribuiu grande importância à influência direta ou indireta da
precipitação pluviométrica sobre a biomassa fitoplanctônica.Realizado na baía do
Guajará e foz do rio Guamá, os valores de organismos por litro tiveram máxima de
4.790.000 org/L, mostrando a influência fluvial que é um estímulo para o crescimento de
algas. A salinidade no rio Guamá é consideravelmente menor que nos rios Mocajuba e
Pacamorema, podendo assim ter sido grande determinante na menor quantidade de
organismos por litro nesses dois estuários.

288
Belém (PA), 11 a 13 de dezembro de 2019
Além da densidade, dois parâmetros que foram maiores no estuário do rio
Mocajuba foram os de condutividade elétrica e sólidos totais dissolvidos, o que pode ter
influenciado na maior quantidade de microalgas no ambiente.
FIGURA 1- A – Oxigênio Dissolvido (PPM); B – Salinidade (PSI); C – Temperatura
°C; D – pH; E – Sólidos Totais Dissolvidos (PPM); F – Condutividade Elétrica.
Fonte: Ariel Romeiro Pereira.
7.029.7 10.01 10.43 9.67
0.21 0.31 0.56 0.42 0.85
0
5
10
15
1 2 3 4 5
Pontos de Coleta
A
Curuçá S. J. da Ponta
14,682 14,688 14,69514,704
14,681
14,646 14,646 14,65114,662 14,661
14,60014,62014,64014,66014,68014,70014,720
1 2 3 4 5
Pontos de Coleta
B
Curuçá S. J. da Ponta
29.32
29.8 29.68 29.69
30.06
29.38 29.38 29.44 29.48 29.57
28.5
29
29.5
30
30.5
1 2 3 4 5
Pontos de Coleta
C
Curuçá S. J. da Ponta
7.88 7.79 7.797.91 7.86
7.23 7.28 7.22 7.15 7.19
6.5
7
7.5
8
1 2 3 4 5
Pontos de Coleta
D
Curuçá S. J. da Ponta
509
506507 507
504
509 509508 508
507
500
502
504
506
508
510
1 2 3 4 5
Pontos de Coleta
E
Curuçá S. J. da Ponta
1019
10121013 1014
1008
1017
10181016 1016 1015
1000
1005
1010
1015
1020
1 2 3 4 5
Pontos de Coleta
F
Curuçá S. J. da Ponta

289
Belém (PA), 11 a 13 de dezembro de 2019
No que se refere a densidade fitoplanctônica, no estuário do rio Mocajuba, as taxas
de densidade foram maiores do que no estuário do rio Pacamorema. Em seu estudo,
realizado na baía do Guajará e foz do rio Guamá, Paiva (2006) atribuiu grande
importância à influência direta ou indireta da precipitação pluviométrica sobre a biomassa
fitoplanctônica, cujos valores de organismos por litro apresentarammáxima de 4.790.000
org/L, mostrando a influência fluvial que é um estímulo para o florescimento das
microalgas. A salinidade no rio Guamá é consideravelmente menor que nos rios
Mocajuba e Pacamorema, podendo assim ter sido grande determinante na menor
quantidade de organismos por litro nesses dois estuários.
FIGURA 2 – Variação fitoplanctônica dos dois rios.
Fonte: Ariel Romeiro Pereira.
Além da densidade do microfitoplâncton, dois parâmetros que foram mais
elevados no estuário do rio Mocajuba, a condutividade elétrica e sólidos totais
dissolvidos, o que pode ter influenciado na maior biomassa de microalgas no ambiente.
4. CONSIDERAÇÕES FINAIS
Apesar da pouca diferença com relação aos parâmetros físico-químicos, a
diferença entre a densidade do microfitoplâncton foi consideravelmente alta, podendo ter

290
Belém (PA), 11 a 13 de dezembro de 2019
sofrido grande influência da diferença de Oxigênio Dissolvido na água, que foi a maior
entre os dois pontos, sendo tão significativa quanto a variação do fitoplâncton.
Os parâmetros de São João da Ponta foram os que mais tiveram homogeneidade
nos dados, tendo os menores valores de desvio padrão, o que pode ter influenciado na
produtividade primária do local, pois é onde houve uma menor variação entre os pontos
coletadospontos coletados.
O T mostrou diferenças significativas entre as áreas (p<0,05)Ao fazer um teste
estatístico (teste T) com os parâmetros de maior variação, que foramnaDdensidade do
fitoplâncton (org/L)e Oxigênio Dissolvido, foram observadas diferenças significativas
entre as áreas (p<0,05).

291
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
AQUINO, E. P.; Oliveira, E. C. C.; FERNANDES, U. L.; LACERDA, S. R.
Fitoplâncton de uma lagoa de estabilização no nordeste do Brasil. Brazilian Journal
of Aquatic Science and Technology, 15, n.1, p. 71-77, 2011.
CARDOSO, F. F. Dinâmica da comunidade microfitoplanctônica Relacionada com
os parâmetros físico químicos do estuário guajara-mirim(Vigia-Pa). Dissertação,
Instituto de Ciências Biológicas, Universidade Federal do Pará,Belém, 2009. 10-13 p.
CARRARO, F. G. P. Estrutura do fitoplâncton e sua utilização como indicador de
condições ecológicas no reservatório de Pedra, Bahia. Dissertação de Mestrado,
Universidade Federal Rural de Pernambuco, Departamento de Pesca, Recife, PE,
2009.59p.
CARVALHO, M. C. Comunidade fitoplanctônica como instrumento de
biomonitoramento de reservatórios no Estado de São Paulo. Tese (Doutorado)
Faculdade de Saúde Pública. Departamento de Saúde Ambiental. Universidade de São
Paulo, São Paulo, 2003. 83p.
DA ROCHA NETO, Orlando Dias; DA SILVA, Bruno Melo; PAIVA, Rosildo Santos.
VARIAÇÃO DOS PARÂMETROS FÍSICO-QUÍMICOS, COMPOSIÇÃO E
BIOMASSA FITOPLANCTÔNICA EM UMA ESTAÇÃO FIXA NA FOZ DO RIO
GUAMÁ, BELÉM, PARÁ-BRASIL. Boletim Técnico Científico do Cepnor/Tropical
Journal of Fisheries and Aquatic Sciences, v. 16, n. 1, p. 19-28, 2017.
ESKINAZI-LEÇA, E.; KOENING, M. L.; SILVA-CUNHA, M. G. G. Estrutura e
dinâmica da comunidade fitoplanctônica.2004. 353-373p.
HORNE, A. J. & GOLDMAN, C. R. Limnology, 2nd Ed. Library of Congress Catalogin-
in-Publication Data.1994. 576p.
LOPES, A. G. D. Estudo da comunidade fitoplanctônica como bioindicador de
poluição em três reservatórios em série do Parque Estadual das Fontes do Ipiranga
(PEFI), São Paulo, SP. Dissertação (Mestrado) - Faculdade de Saúde Pública.
Departamento de Saúde Ambiental Universidade de São Paulo, São Paulo, 2007. 116 p.
MACEDO, SJ de; COSTA, K. M. P. Estudo ecológico da região de Itamaracá
Pernambuco-Brasil, condições hidrológicas do estuário do rio Botafogo. Ciência e
Cultura, v. 30, n. 7, p. 346-368, 1978.
MARGALEF, R. Limnologia. Ediciones Omega, Barcelona, 1983. 1010p.
ROUND, F.E. Biologia das algas. 2 ed. Rio de Janeiro, Guanabara dois. 1973. 212-228p.
MELO-MAGALHAES, MARINHO, E.; MARIA, L. K.; SANT’ANNA, C. L.
Fitoplancton e variáveis ambientais nos canais do sistema estuarino lagunar
Mundali/Manguaba, Alagoas, Brasil.Hoenea, 2004.

292
Belém (PA), 11 a 13 de dezembro de 2019
PAIVA, Rosildo Santos et al. Considerações ecológicas sobre o fitoplâncton da baía do
Guajará e foz do rio Guamá, Pará, Brasil. Boletim do Museu Paraense Emílio Goeldi
Ciências Naturais, v. 1, n. 2, p. 133-146, 2006.
PASSAVANTE, J. Z de O. & FEITOSA, F. do N. Hidrologia e plâncton da plataforma
continental de Pernambuco 2. Biomassa primária do fitoplâncton. Anais do Encontro
Brasileiro de Gerenciamento Costeiro 3. Fortaleza: 1989.
PEREIRA, A. C. A. Bioindicadores fitoplanctônicos da qualidade da água no estuário
do rio Catu, Aquiraz, Ceará.Dissertação (Mestrado). Universidade Federal do Ceará,
Fortaleza, 2007. 77p.
PRITCHARD, D. W. What is an estuary: physical viewpoint. American Association
for the Advancement of Science, 1967.
SALMASO, N.; NASELLI-FLORES, L.; PADISÁK, J. Impairing the largest and most
productive forest on our planet, how do human activities impact
phytoplankton?.Hydrobiologia, 698: 375-384. 2012.

293
Belém (PA), 11 a 13 de dezembro de 2019
COMPOSIÇÃO DE ROTIFERA NO PARQUE ESTADUAL DO UTINGA
(BELÉM, PARÁ, BRASIL)
Raissa de Cassia Serrão de Souza1 , Marcos Adriano Pereira Paiva2, Lisbethe Melo
Sckyr Ahndrew3, Samara Cristina Campelo Pinheiro 4.
1Graduando em Licenciatura em Ciências Biológicas - Universidade Federal do Pará –
[email protected] 2Graduando em Licenciatura em Ciências Biológicas - Universidade Federal do Pará -
[email protected] 3Mestrado em Ecologia Aquática e Pesca – Instituto Evandro Chagas -
[email protected] 4Doutorado em Saúde Pública (Orientador)– Instituto Evandro Chagas -
RESUMO
O Parque Estadual do Utinga (PEUt) abastece com água potável a cidade de Belém e 70%
da Região Metropolitana e vem sofrendo alguns processos de degradação ambiental
crescente, devido às ações antrópicas e as retiradas da vegetação aquática. Diante disso,
este trabalho teve como objetivo caracterizar a população de Rotifera, considerados
excelentes bioindicadores dos processos de alterações ambientais, durante os processos
de retirada das macrófitas. A amostragem foi realizada em setembro/2017, janeiro/2018
e maio/2018, nos períodos antes, durante e após a retirada da vegetação, respectivamente.
As amostras foram coletadas com auxílio de um balde graduado (10L) em rede plâncton
(45µm), fixadas em formol 4%. Análises qualitativas e quantitativas foram realizadas
através de sub-amostras (1mL). Foram calculadas a densidade total e frequência de
ocorrência, as quais foram correlacionadas com as variáveis ambientais. Foram
registradas 26 espécies e nove famílias de Rotifera. Os valores de densidade variaram de
3.750 org/m³, durante (maio/18), a 102.000 org/m³ (setembro/17) antes da retirada da
vegetação. Entre as espécies muito frequentes (100% FR) antes da retirada das macrófitas,
destacam-se: Brachionus mirus, Brachionus zahniseri, Keratella americana, Keratella
cochlearis, Keratella lenzi, Filinia longiseta e Filinia terminalis. Durante o processo e
após, Brachionus caudatus, Brachionus dolabratus e Brachionus mirus, foram as
espécies que mais se destacaram. Entre as variáveis ambientais, o pH (Anova, F= 17,79
e p= 0,0) e Alcalinidade (Anova, F=22,05 e p= 0,0 ) apresentaram variações significativas
entre os períodos analisados. Não houve correlação entre as densidades de Rotífera e
variáveis densidade de Rotifera durante os processos ambientais. De acordo com
resultados obtidos, foi possível observar que os processos de retirada das macrófitas
aquáticas podem alterar as variáveis ambientais e consequentemente a densidade da
população de Rotifera. Desse modo, faz-se necessário biomonitoramento desses
organismos, com ênfase nas espécies biondicadoras das condições tróficas do Parque
Estadual do Utinga (PEUt).
Palavras-chave: Bioindicadores; Macrófitas; Modificações Tróficas.
Área de Interesse do Simpósio: Biodiversidade

294
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O Parque Estadual do Utinga (PEUt) é um complexo de conservação que possui
grande diversidade de espécies de fauna e flora, o PEUt é um reservatório de decantação
e sistema hídrico que abastece 70% da Região Metropolitana de Belém (PARÁ, 2013).
Com o avanço da urbanização, o complexo tornou-se vulnerável a processos de alterações
ambientais, em virtude do lançamento de rejeitos domésticos e industriais.
A área do Parque tem sofrido com a degradação ambiental em razão das obras de
ampliação da Avenida João Paulo II e a eminente pressão urbana. Com o crescimento das
comunidades habitacionais nas proximidades, o parque padece com a ação antropológica,
visto que, a maioria destas moradias não possui um sistema de saneamento básico
adequado para rejeitar os resíduos produzidos (PARÁ, 2013).
Entre os componentes da comunidade zooplanctônica, os rotíferos são
microrganismos muito importantes nas teias alimentares de ambientes aquáticos, se
alimentam de algas, protozoários, bactérias e detritos, servindo como presas para
predadores invertebrados, principalmente peixes. Participam da reciclagem de nutrientes
e serem úteis como indicadores de qualidade ambiental (SANTOS ET AL, 2013).
Diante disso, é necessário o monitoramento desses efeitos externos no ambiente,
utilizando abordagens ecológicas com foco nas comunidades biológicas para analisar as
modificações ambientais provenientes destes efeitos. Por isto, este estudo optou por
observar as comunidades zooplanctônicas, visto que, esses organismos mostram-se
excelentes bioindicadores devido à alta sensibilidade a mudanças ambientais, podendo
alterar tanto em sua densidade quanto composição (COELHO-BOTELHO, 2002).
Desse modo, este trabalho teve como objetivo caracterizar a comunidade de
Rotifera durante os processos de retirada das macrófitas aquáticas do Parque Ambiental
do Utinga (PEUt), sistema de abastecimento público da região metropolitana de Belém
(Pará-Brasil).
2. MATERIAL E MÉTODOS
2.1. AMOSTRAGEM
As amostragens foram realizadas em diferentes períodos: antes (Setembro/2017),
durante (Janeiro/2018) e depois (Maio/2018) dos processos de retirada das macrófitas

295
Belém (PA), 11 a 13 de dezembro de 2019
aquáticas. Em três pontos: PT 01 (S 01°25’15.3” W 048°26’02.2”); PT 02 (S 01°25’31.3”
W 048°26’00.9”); PT 03 (S 01°24’59.7” W 048°25’47.2”), distribuídos ao longo do
Parque Estadual do Utinga (PEUT) (Figura 01).
Figura 01. Mapa de localização da área de estudo com as estações de coleta no Parque Estadual
do Utinga (Pará – Brasil).
Fonte: SOUSA, 2019
Amostras para o estudo qualitativo e quantitativo dos rotíferos foram realizadas
com auxílio de um balde graduado (volume 10L) em uma rede de plâncton (45 µm de
abertura de malha), próximas às vegetações de macrófitas aquáticas (principalmente do
gênero Eichhornia). Todo o material foi fixado em solução de formol a 4% e
acondicionado em frascos plásticos de polipropileno de 250 mL.
Os parâmetros temperatura (°C), potencial hidrogeniônico (pH), condutividade
elétrica (CE), sólidos totais dissolvidos (STD), salinidade (SA) e oxigênio dissolvido
(OD) foram realizados in situ, utilizando um medidor multiparamétrico portátil (HI9828
- HANNA®).
2.3. ANÁLISE LABORATORIAL
No Laboratório de Biologia Ambiental (LBA/SAMAM/IEC), para as análises
qualitativas foram utilizadas sub amostras em placas de Petri, e para as análises
quantitativas foram utilizadas 7 mL da amostra total de cada estação, corados com rosa
de bengala 1%, em lâminas de Sedgewick Rafter (1 mL), em seguida, observadas em

296
Belém (PA), 11 a 13 de dezembro de 2019
microscópio invertido (Axiovert 40C - Carl Zeiss). A determinação sistemática das
espécies foi baseada nas bibliografias especializadas.
As variáveis: turbidez (TRB), sólidos totais em suspensão (STS) e demanda
química de oxigênio (DQO) foram determinadas por Espectrofotometria de UV-VIS.
Para a determinação da demanda bioquímica de oxigênio (DBO) se utilizou técnica de
incubação por cinco dias (APHA, 2005). As análises dos íons N-nitrito (N-NO2-), N-
nitrato (N-NO3-), nitrogênio amoniacal (N-NH4) e fósforo total (Ptotal) foram realizados
por Cromatografia de Íons (ICS DUAL 2000-DIONEX).
2.4. TRATAMENTO DOS DADOS
A frequência de ocorrência (F) das espécies foi determinada pela seguinte
equação: F= (PA/P)*100. Onde: F= frequência da espécie A nas amostras; PA= número
de amostras onde a espécie A ocorreu e P= número total de amostras analisadas. Seguindo
a seguinte classificação: muito frequentes (>70%), frequentes (≤70% e >40%), pouco
frequente (≤ 40% e >10%) e esporádico (≤10%). A abundância relativa (AR) foi calculada
a partir da relação entre o número total de espécie, pelo número total de espécies na
amostra.
As análises de variância (ANOVA) e os testes de correlação, coeficiente de
correlação de Spearman R, foram realizados para comparar a densidade de Rotifera e
variáveis abióticas (transparência, temperatura, pH, OD, TDS, Salinidade, Alcalinidade,
Turbidez, STS, DQO e nutrientes), utilizando-se o programa STATISTICA 6.0 (2001).
Os cálculos das concentrações de clorofila-a foram realizados segundo Strickland &
Parsons (1972).
3. RESULTADO E DISCUSSÃO
3.1. VARIÁVEIS AMBIENTAIS
Durante o período do estudo, as concentrações de clorofila-a variaram de 4,59
(PT03, Setembro/2017) a 24,01 (PT02, Maio/2018), a transparência da água variou de 42
cm (PT02, Maio/2018) a 120 cm (PT03, Janeiro/2018). A temperatura da água teve uma
variação de 28,79º (PT02, Maio/2018) a 30,4º (PT02, Setembro/2017). O pH apresentou
valores mínimo e máximo no PT03, em períodos sazonais diferentes, com 5,06 em

297
Belém (PA), 11 a 13 de dezembro de 2019
setembro/17 e 6,5 em janeiro/18. A concentração de oxigênio dissolvido (OD) variou
entre 2,4 mg/L (PT03, Setembro/17) a 7,76 mg/L (PT01, Maio/18). Os valores de TDS
apresentaram uma variação de 37mg/L (PT02, Maio/18) a 64mg/L (PT02, Setembro/17).
A salinidade apresentou valor mínimo (0,03%) em janeiro e maio de 2018 e
máximo (0,06%) no período de setembro/2017. A alcalinidade variou de 15 ppm (PT01,
Janeiro/18) a 60 ppm (PT01 de Maio/18). A turbidez da água apresentou variações de 5
NTU (PT03, Maio/18) a 25 NTU (PT02, Janeiro/2018). O STS variou entre 5,59 mg/L
(PT03, Maio/18) e 20 mg/L (PT02, Setembro/17). A DQO variou de 0,6 mg.L-1 ( PT02,
setembro/17) a 66 mg.L-1 (PT02, maio/18) (Tabela 01).
As concentrações de Fosfato variaram de 0,76 (PT03, Maio/18) a 0,81 (PT03,
Janeiro/18). O Nitrito variou de 0,0072 (PT03) a 0,015 (PT02), ambos no período de
janeiro/18. As concentrações de sulfato variaram de 0,87 (PT03, maio/18) a 3,9 (PT03,
janeiro/18).
Entre as variáveis ambientais, o pH (Anova, F= 17,79 e p= 0,0) e Alcalinidade
(Anova, F=22,05 e p= 0,0) apresentaram variações significativas entre os períodos
analisados. Em média, a alcalinidade foi maior no período depois (50±10) da retirada. O
pH foi maior nos períodos durante (6,23±0,25) e depois (6,23±0,25) das retiradas de
macrófitas, caracterizando ligeiramente a água como ácida. Isso pode está relacionado
com o revolvimento da água proveniente dos processos de retiradas das macrófitas no
lago do PEut, contribuindo para o possível aumento da concentração de nutrientes na
água.
Tabela 01. Variáveis hidrológicas registradas no reservatório do Parque do Utinga, de
setembro/2017 a maio/2018.
Setembro/2017 Janeiro/2018 Maio/2018
Antes Durante Depois
PT01 PT02 PT03 PT01 PT02 PT03 PT01 PT02 PT03
Clorofila-a 8,5 10,5 4,6 7,2 13,1 3,3 11,8 24,0 4,7 Transparência 50,0 44,0 90,0 110,0 60,0 120,0 55,0 42,0 70,0 Temperatura 30,1 30,4 30,4 28,8 28,8 28,9 29,1 28,8 29,1 pH 5,4 5,1 5,1 6,0 6,2 6,5 6,0 6,5 6,2 OD 3,8 5,0 2,4 4,0 3,8 5,0 7,8 5,2 3,1 TDS 60,0 64,0 63,0 39,0 39,0 44,0 40,0 37,0 41,0 Salinidade 0,1 0,1 0,1 0,0 0,0 0,0 0,0 0,0 0,0 Alcalinidade 30,0 25,0 25,0 15,0 19,0 19,0 60,0 40,0 50,0 Turbidez 11,0 22,0 7,0 14,0 25,0 22,0 7,0 9,0 5,0

298
Belém (PA), 11 a 13 de dezembro de 2019
STS 8,0 20,0 5,6 7,0 13,0 9,0 6,0 8,0 5,6 DQO 1,6 0,6 1,6 2,0 1,6 1,6 64,0 66,0 64,0 Fosfato 0,3 0,0 0,4 0,7 0,7 0,8 0,1 0,8 0,8 Nitrito 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 Sulfato 3,0 3,0 0,2 3,5 3,2 3,9 0,9 1,3 0,9
Fonte: Laboratório de Toxicologia, SAMAM, Instituto Evandro Chagas (2018).
Segundo Cavalcante (2010), um dos fatores biológicos que pode afetar o pH e a
alcalinidade da água é a fotossíntese. Com a remoção de CO2 da água durante a
fotossíntese, há elevação do pH da água pelo consequente consumo de íons H+, uma vez
que foi observado um aumento da concentração de clorofila-a após a retirada das
macrófitas.
3.2. COMPOSIÇÃO TAXONÔMICA
O Filo Rotifera, durante o período estudado, foi composto por 26 espécies,
pertencem a nove famílias, sendo Brachionidae a mais representativa, destacando-se
ainda Trochosphaeridae (4 espécies) e Trichocercidae (5 espécies) (Tabela 2).
3.3. DENSIDADE TOTAL
A densidade total de Rotifera variou entre os pontos amostrados e períodos
estudados, com valores mínimos (3.750 org/m³) registrados maio/18 (PT 02), depois da
retirada das macrófitas, e máximo (102.000 org/m³), PT 01, antes da retirada
(setembro/17). No presente trabalho, foi observada, que em setembro/2017, uma elevada
densidade de Rotiferas, no entanto diminuiu consideravelmente após o início do processo
de retirada da vegetação. Este fato pode estar relacionado à influência das macrófitas
sobre a zooplanctônica, e sobre os impactos nos fatores ambientais. Além disso, as
macrófitas oferecem um refúgio para o zooplâncton contra peixes planctívoros, bem
como contra a predação por invertebrados.
Trabalhos como o de Margalef (1983), ressaltaram que o número de espécies de
rotíferos tende a aumentar em ambientes menos mineralizados e eutróficos; porém,
diminui quando o grau de eutrofização aumenta. Alguns autores enfatizam que a
predominância dos rotíferos não está relacionada somente ao estado trófico dos corpos
hídricos, mas coincidente a outros fatores, como origem do biossistema e interações
biológicas.

299
Belém (PA), 11 a 13 de dezembro de 2019
3.6. FREQUÊNCIA DE OCORRÊNCIA
As espécies classificadas como muito frequentes (>70%), durante todo o período
do estudo, foram: Brachionus mirus, Filinia longiseta e Filinia terminalis, os quais
ocorreram em 100% das amostras.
No período antes da retirada das macrófitas (setembro/2017), as espécies
classificados como muito frequentes (>70%) foram: Brachionus mirus, Brachionus
zahniseri, Keratella americana, Keratella cochlearis, Keratella lenzi, Filinia longiseta e
Filinia terminalis.
No mês de janeiro/2018 (durante a retirada das macrófitas) foram observadas
como muito frequentes: Brachionus caudatus, Brachionus dolabratus, Brachionus mirus,
Brachionus zahniseri, Keratella americana, Keratella cochlearis, Filinia longiseta e
Filinia terminalis. Já no mês de maio/2018 (após a retirada das macrófitas) organismos
que permaneceram como muito frequentes: Brachionus caudatus, Brachionus mirus,
Filinia longiseta e Filinia terminalis. (Tabela 2).
Assim como em outros reservatórios, os rotíferos mostraram-se frequentes e
abundantes, (SENDACZ ET AL., 1985; ALMEIDA ET AL., 2006; MEIRINHO &
POMPÊO, 2015), pois são indivíduos oportunistas capazes de se adaptar às diferentes
condições ambientais assim como desequilibrar a estrutura da comunidade
zooplanctônica devido à competição por recursos alimentares. (ALLAN, 1976;
LOUREIRO ET AL., 2011).
Tabela 02. Composição Taxonômica e Frequência de Ocorrência do Filo Rotifera, registrados
durante o período do estudo, do Parque Estadual do Utinga (Belém, Pará)
TÁXON
Setembro/17 Janeiro/18 Maio/18 FR%
Antes Durante Depois
PT
01 PT
02 PT
03 PT
01 PT
02 PT
03 PT
01 PT
02 PT
03
FILO ROTIFERA
Classe Eurotatória Ordem Ploima Família Asplanchnidae Asplanchna sieboldi Leydig,
1854 PF 11%
Família Brachionidae

300
Belém (PA), 11 a 13 de dezembro de 2019
Brachionus caudatus Barrois
And Daday, 1894 FR FR MF MF MF MF MF MF 89%
Brachionus dolabratus
Harring, 1914 FR FR MF MF MF 56%
Brachionus falcatus
Zacharias 1898 PF PF FR FR 44%
Brachionus mirus Daday,
1905 MF MF MF MF MF MF MF MF MF 100%
Brachionus patulus Müller,
1786 FR FR 22%
Brachionus patulus var
macracanthus Daday, 1905 PF 11%
Brachionus zahniseri
Ahlstrom, 1934 MF MF MF FR FR FR FR 78%
Brachionus sp. PF PF 22%
Keratella americana Carlin,
1943 MF MF MF MF MF MF FR FR 89%
Keratella cochlearis Gosse,
1851 MF MF MF MF MF MF FR FR 56%
Keratella lenzi Hauer, 1953 MF MF MF PF FR FR 44%
Família Lecanidae Lecane curvicornis Murray,
1913 PF FR FR 11%
Lecane proiecta Hauer, 1956 FR PF 78%
Família Lepadellidae Lepadella sp. FR 11%
Família Synchaetidae Polyarthra remata Skorikov,
1896 FR FR FR FR FR 56%
Família Trichoceridae Trichocerca sp. PF PF 22%
Trichocerca cylindrica
Imhof, 1891 ES ES ES
Ordem Flosculariaceae Família Trochosphaeridae Filinia longiseta Ehrenberg,
1834 MF MF MF MF MF MF MF MF MF 100%
Filinia opoliensis Zacharias,
1898 FR FR FR
Filinia terminalis Plate, 1886 MF MF MF MF MF MF MF MF MF 100% Filinia sp PF PF 22%
Família: HEXARTHRIDAE Hexarthra fennica Levander,
1892 FR FR 22%
Hexarthra intermedia
Wiszniewski, 1929 FR FR 22%
Hexarthra polyodonta Hauer,
1957 PF 11%
Fonte: Laboratório de Biologia Ambiental, SAMAM, 2018.

301
Belém (PA), 11 a 13 de dezembro de 2019
3.5. ABUNDÂNCIA RELATIVA
Entre as famílias de Rotifera registrada, Brachionidae foi mais abundante, em
todos os períodos analisados, com valores mais elevados no PT 01 em janeiro/18 (30,8%),
durante a retirada das macrófitas, e no PT 01 (26,9%) e PT 02 (26,9%) do mês de
setembro/17, antes da retirada (Figura 02).
Figura 02. Abundância relativa das famílias de Rotifera, registradas durante o período do
estudo, no Parque Estadual do Utinga (Pará – Brasil)
Fonte: Laboratório de Biologia Ambiental, SAMAM, Instituto Evandro Chagas, 2018.
3.6.CORRELAÇÃO DE SPEARMAN-R.
Não houve correlação entre as densidades de Rotifera e variáveis abióticas
4.CONCLUSÃO
No presente estudo, levando em consideração que as modificações antrópicas
realizadas no Parque do Utinga (PEut) podem alterar os padrões da composição e
estrutura de Rotifera, apesar de não se ter obtido correlação entre as variáveis abióticas e
densidade, foi possível observar o aumento das concentrações de pH e alcalinidade, e a
diminuição da densidade de Rotifera durante os processos de retiradas das macrófitas
aquáticas no reservatório do PEut. Desse modo, faz-se necessário o monitoramento
continuo desses organismos, com ênfase nas espécies biondicadoras das condições
tróficas e consequentemente na qualidade da água superficial do Parque Ambiental do
Utinga (PEUt).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
PT 01 PT 02 PT 03 PT 01 PT 02 PT 03 PT 01 PT 02 PT 03
Antes Durante Depois
Setembro/2017 Janeiro/2018 mai/18
Asplanchnidae Brachionidae Lecanidae
Lepadellidae Synchaetidae Trichoceridae
Hexarthridae Trichotriidae Trochosphaeridae

302
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALLAN, J. D. Life history patterns in zooplankton. American Naturalist. v. 110, p. 165-
176. 1976.
ALMEIDA, V. L. S. et al. Rotífera das zonas limnéticas e litorânea do reservatório
de Tapacurá, Pernambuco, Brasil.Iheringia, Série Zoologia, v.96, n.4, p.445-451,
2006.
ANDREOLI, C. V.; CARNEIRO, C. (2005). Gestão integrada de mananciais de
abastecimento eutrofizados.Curitiba: Sanepar/ FINEP.
PARÁ. Revisão do Plano de Manejo do Parque Estadual do Utinga. Secretaria de
Estado de Meio Ambiente. Belém: SEMA; Belém: IMAZON, 2013. 376 p.
CAVALCANTE, D. H.; SÁ, M. V. C. Efeito Da Fotossíntese Na Alcalinidade Da Água
De Cultivo da Tilápia do Nilo. Revista Ciencia Agronomica. v. 41, n. 1, p. 67-72. 2010.
COELHO-BOTELHO, M. J. Influência da transposição das águas do reservatório
Billings para o reservatório Guarapiranga (São Paulo) na comunidade zooplanctônica. I.
Período chuvoso (1997 a 2001). In: Resumos do XXIV Congresso Brasileiro de
Zoologia, Itajaí-SC, 17 a 22 de fevereiro de 2002.
LOUREIRO, B. R.; COSTA, S. M.; MACEDO, C. F.; HUSZAR, L. M.; BRANCO, C.
W. C. Comunidades zooplanctônicas em sistemas de criação de peixes. Boletim Instituto
de Pesca de São Paulo, v. 37, n. 1, p. 47-60. 2011.
MARGALEF, R. Limnologia.Barcelona: Omega. 1983
MEIRINHO, P. A.; POMPÊO, M. 2015. Histórico de estudos sobre a comunidade
zooplanctônica do reservatório Rio Grande ao longo do tempo e sua
heterogeneidade espacial. In: POMPÊO, M.; MOSCHINI-CARLOS, V.; NISHIMURA,
P.; CARDOSO-SILVA, S. & LOPEZ-DOVAL, J. C. (Eds.) Ecologia de reservatórios e
interfaces. São Paulo, Instituto de Biociências da Universidade de São Paulo. 460p.
SANTOS, T. M.; MELO JÚNIOR, H. N. Comportamento limnológico da coluna de água
da piscicultura em tanque rede no açude rosário Lavras da Mangabeira–CE. In: XIII
Reunião Científica do Instituto de Pesca. São Paulo. 2013. Acesso:
http://www.pesca.sp.gov.br/11recip2013/resumos/11a_ReCIP_R54_173-175.pdf
SENDACZ, S. et al. Zooplankton Biomass Of Reservoirs In Different Trophic Conditions
In The State Of São Paulo, Brazil. Brazilian Journal of Biology, v. 66, p. 337-350, 2006.

303
Belém (PA), 11 a 13 de dezembro de 2019
DINÂMICA DA COMUNIDADE ZOOPLANCTÔNICA E SUA RELAÇÃO COM
A QUALIDADE DA ÁGUA NO PORTO DE VILA DO CONDE (PARÁ,
BRASIL)
Marcos Adriano Pereira Paiva¹; Raissa De Cássia Serrão De Souza²; Lisbethe Melo
Sckyr Ahndrew3; Samara Cristina Campelo Pinheiro4
1Graduando em Licenciatura Plena em Ciências Biológicas (ICB). Instituto Evandro
Chagas (IEC) [email protected] 2Graduando em Licenciatura Plena em Ciências Biológicas (ICB). Instituto Evandro
Chagas (IEC) [email protected] 3Mestre Ecologia Aquática e Pesca.Instituto Evandro Chagas (IEC)
[email protected] 4Doutora em Saúde Coletiva (Orientadora). Instituto Evandro Chagas (IEC)
RESUMO
A comunidade zooplanctônica, base da cadeia alimentar aquática, é considerada um
importante elo na transferência energética entre produtores primários e os demais níveis
tróficos. Além disso, algumas espécies zooplanctônicas são sensíveis ou tolerantes aos
processos de alterações ambientais. Levando em consideração a potencialidade desses
organismos como bioindicadores, este estudo tem como objetivo caracterizar a
comunidade zooplanctônica e relacioná-los com as variáveis ambientais no Porto de Vila
do Conde, no município de Barcarena (Pará). A amostragem foi realizada em dois pontos
de coleta distribuídos ao longo do porto, durante os meses de dezembro/2017, março,
junho e setembro/2018. Foram realizados arrastos horizontais na sub-superfície da água,
com o auxilio de rede de plâncton (45 µm). Análises qualitativas e quantitativas foram
realizadas através de sub-amostras (1mL), em placas de Sedgwick rafter, com o auxílio
de microscópio invertido e identificadas em literatura especializada. Foram calculadas
densidade total e frequência de ocorrência, as quais foram correlacionadas com as
variáveis ambientais. A comunidade zooplanctônica foi representada por 7 filos, sendo
Rotifera o mais representativo. A densidade total dos organismos variou de 309 org./m³
(março/2018) a 8105 org./m³ dezembro/2017), nas marés enchente e vazante,
respectivamente. Os grupos zooplanctônicos que mais se destacaram, em frequência de
ocorrência, foram: Rotifera (48%), seguido pelos Cladocera (28%) e Copepoda (24%). A
estrutura da comunidade zooplanctônica esteve correlacionada com a sazonalidade, onde
foi observado elevada densidade de organismos durante o período mais chuvoso,
principalmente pelas fases iniciais de naúplio e copepodito. No entanto, o padrão de
variação desses organismos pode estar relacionado às condições ambientais da região,
historicamente impactado pelo lançamento de efluentes e rejeitos industriais. Desse
modo, faz-se necessário biomonitoramento da comunidade zooplônctonica, com ênfase
nas espécies biondicadoras das condições tróficas das regiões portuárias amazônicas.
Palavras-chave: Zooplâncton, Bioindicadores, Biologia ambiental.
Área de Interesse do Simpósio: Biodiversiade

304
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O porto de Vila do Conde, localizado na cidade de Barcarena (Pará), agrega um
importante pólo industrial e portuário da região Paraense, o qual recebe um elevado fluxo
de navios que transportam tanto matéria prima (bauxita, alumina), como cargas
beneficiadas (lingotes de alumínio) (PIRATOBA, 2017).
Devido às intensas atividades portuária e industrial esta região passa por constantes
processos de alterações ambientais, como consequência da introdução de compostos
gerados pelas atividades desenvolvidas na região e desde modo gerando impactos ao
ecossistema aquático (PINHEIRO et al., 2019).
Com a intensificação da atividade na zona portuária e seu funcionamento tornam
crescente a preocupação ecológica com o ambiente aquático, uma vez que a descarga de
substancias também aumenta, as quais podem afetar de maneira negativa o ambiente. Os
subprodutos produzidos no beneficiamento da bauxita e caulim e efluentes domésticos
podem afetar a qualidade do ambiente para os organismos aquáticos, ou mesmo a saúde
humana, por meio do consumo de águas contaminadas (AZEVEDO; CHASIN, 2003).
O estudo do zooplâncton, base da cadeia alimentar aquática, pode auxiliar no
monitoramento dos efeitos nocivos de despejos domésticos e industriais, pois esses
organismos possuem grande sensibilidade ambiental e respondem a diversos tipos de
impactos, tanto pela alteração na sua quantidade como na composição e diversidade da
comunidade (COELHO-BOTELHO, 2002). Considerados excelentes biondicadores das
condições tróficas do ambiente, fornecendo dados relevantes sobre processos que
interagem no meio, sendo influenciados pelas condições abióticas e bióticas
(NEUMANN-LEITÃO et al., 1991).
O zooplâncton é formado por organismos de diversos táxons, dentre estes, o filo
Rotifera e os microcrustáceos Cladocera e Copepoda são os mais representativos,
constituindo a maior parte dos invertebrados planctônicos (CARDOSO et al., 2008). Este
trabalho teve como objetivo analisar as possíveis correlações entre a comunidade
zooplanctônica (Rotifera, Copepoda e Cladocera) e as características tróficas do estuário
do rio Pará e avaliar o emprego desta comunidade como bioindicadora da qualidade da
água região Portuária Paraense.

305
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
O porto de Vila do Conde está localizado no município de Barcarena - PA, na
margem direita do rio Pará, a aproximadamente 3 km de distância de Vila do Conde. O
estuário do rio Pará sofre grande influência de águas fluviais em ambas as margens, com
salinidade reduzida, mas a influência de maré é registrada até vários quilômetros adentro
do continente, caracterizando esse estuário como uma região de transição fluvio-marinha
sob o impacto de marés semidiurnas (GREGÓRIO; MENDES, 2008). Pelo porto transita
diversas cargas de bauxita, alumina, lingotes de alumínio, cargas gerais, soda caustica e
óleo combustível, recebendo navios graneleiros, navios com contêineres e navios de
granéis líquidos (CDP, 2010).
De acordo com Ramos (2004), a área de estudo está inserida em uma microrregião
com características bem peculiares em relação ao clima amazônico. Devido a sua posição
geográfica próxima ao Oceano Atlântico, sua baixa altitude e sua vegetação densa, lhe
conferem características distintas daquelas comuns à região amazônica. Caracterizado
pelas amplitudes térmicas menores que 200 C, porém nas horas de maior insolação
(durante o meio dia) podem-se registrar temperaturas acima de 30o C. Quanto às condições
atmosféricas, podem-se destacar meses muito secos, além de não existir uma distribuição
uniforme das chuvas durante o ano, ocorrendo uma sensível diminuição na precipitação
pluviométrica a partir do mês de junho atingindo índices mínimos por volta do mês de
outubro a novembro.
2.2. AMOSTRAGEM
As coletas foram realizadas nos meses de dezembro/2017, março/2018, junho/2018
e setembro/2018, durante os períodos de maré enchente e vazante. As amostragens foram
realizadas em dois pontos (PT 02 e PT 06), distribuídos ao longo do porto de Vila do
Conde (Figura 1), onde foram efetuadas mediações in situ dos fatores físico-químicos da
água (Temperatura (T°), Potencial Hidrogeniônico (pH), Condutividade Elétrica (CE),
Salinidade, Oxigênio Dissolvido (OD), Demanda Bioquímica de Oxigênio (DBO)) e
parâmetros biológicos (zooplâncton).
Os organismos zooplanctônicos foram obtidos através de arrastos horizontais na
sub-superfície da coluna d’água, utilizando-se redes de plâncton (45 µm). O material

306
Belém (PA), 11 a 13 de dezembro de 2019
coletado foi retirado do copo coletor e colocado em frascos de polipropileno de 250 ml,
devidamente etiquetados com informação do ponto de amostragem, contendo formalina
4% neutralizada com tetraborato de sódio.
Figura 1. Mapa da área de estudo com a delimitação com os pontos de amostragem na
área de abrangência do Porto de Vila do Conde (Pará-Brasil).
Fonte: Trindade, 2013
2.3. ANÁLISE LABORATORIAL
No Laboratório de Biologia Ambiental (LBA/SAMAM/IEC) foi realizada a
quantificação dos organismos zooplanctônicos após a homogeneização e sub-amostragem
de cada amostra. As amostras foram analisadas quantitativamente em câmaras de
Sedgewick Rafter de 1mL, onde foram realizadas três sub-amostragens (1 mL cada) e
observadas em um invertoscópio, com câmera fotográfica digital acoplada (contagem e
registro fotográfico). Para as amostras qualitativas, foram preparadas três sub-
amostragem (2 mL cada), em placas de petri analisadas em microscópio invertido. A
determinação sistemática das espécies foi baseada em literatura especializada.
As análises das variáveis ambientais (Temperatura (T), Potencial Hidrogeniônico
(pH), Condutividade Elétrica (CE), Salinidade, Oxigênio Dissolvido (OD), Demanda
Bioquímica de Oxigênio (DBO) foram realizadas no Laboratório de Toxicologia
Ambiental (LT / SAMAM/ IEC).

307
Belém (PA), 11 a 13 de dezembro de 2019
2.4. TRATAMENTO DOS DADOS
A densidade foi determinada pela equação: N= (Vt*x) /Vc. Onde: N= densidade
total (org./m³); Vt= volume total de diluição; Vc= volume da sub-amostra; x= número de
organismos de cada táxon, na subamostra. A abundância relativa (AR) foi calculada a
partir da relação entre o número total de espécie, pelo número total de espécies na
amostra. A frequência de ocorrência (F) das espécies é determinada pela seguinte
equação: F= (PA/P) *100. Onde: F= frequência da espécie A nas amostras; PA= número
de amostras onde a espécie A ocorreu e P= número total de amostras analisadas, e
posteriormente as espécies são classificadas de acordo com o resultado do cálculo da (F):
esporádicos (FR ≤ 10%), pouco frequente (10% < FR ≥ 30%), frequentes (30% < FR ≥
70%) e muito frequente (FR > 70%).
As análises estatísticas foram realizadas com o auxílio do programa Statistica 7.0
(StatSoft, 2004). A normalidade e homocedasticidade das variâncias foram testadas, e
quando necessário os dados foram transformados (log x+1 ou Raiz Quadrada) para se
obter distribuições próximas da normalidade. Para valores fora da normalidade foi
utilizado o teste de Kruskal- Wallis. A correlação entre físico-químicos e os valores de
densidade foram testados por correlação de Spearman.
3. RESULTADOS E DISCUSSÃO
3.1. VARIÁVEIS AMBIENTAIS
Foi possível observar, entre os períodos sazonais analisados, que a precipitação
pluviométrica foi um fator determinante nas variações de alguns parâmetros ambientais.
O pH apresentou maior valor (10,87) no mês de junho/2018 (maré enchente, PT 06) e
menor (7,4) no mês de dezembro/2017 (maré enchente, PT 02). Carvalho et al. (2000)
mencionam que com o aumento das chuvas, o pH tende a subir e aproximar-se da
neutralidade, pois ocorre maior diluição dos compostos dissolvidos e escoamento mais
rápido. Isso é causado pelo aumento no volume de água que faz com que a acidez da água
diminua.
A temperatura não apresentou elevada variação durante o período do estudo, com
valores mínimo (28,92 ºC), durante o mês de março/2018 (maré enchente, PT 02) e
máximo (29,97 ºC) em setembro/2018, (maré enchente, PT 02) (Tabela 1). Apesar de

308
Belém (PA), 11 a 13 de dezembro de 2019
ser um fator importante na distribuição das espécies, as variações de temperatura em
zonas tropicais são mínimas, desse modo não é considerada como um fator limitante ao
crescimento e distribuição dos organismos (SANTOS, 2008).
A condutividade elétrica (CE) apresentou valor máximo (104 μS/cm) durante mês
de junho/2018 (maré vazante, PT 02) e valor mínimo (66 μS/cm) registrado no mês de
dezembro/2017 (maré enchente, PT 06). Os valores elevados de CE podem estar
relacionados à composição iônica dos corpos d'água, e influenciados pelos lançamentos
de efluentes e rejeitos industriais (MEDEIROS, 2018) e pelo volume de chuvas na região
de estudo.
Tabela 1: Variáveis Ambientais registrados durante o período do estudo, pontos e
período de marés, enchente (EN) e vazante (VZ), no Porto de Vila do Conde (Pará-
Brasil).
Variáveis
Ambientai
s
DEZEMBRO/2017 MARÇO/2018 JUNHO/2018 SETEMBRO/2018 PT 02 PT 06 PT 02 PT 06 PT 02 PT 06 PT 02 PT 06
EN VZ EN VZ EM VZ EN VZ EN VZ EN VZ EN VZ EN VZ
pH 7,42 7,63 7,76 7,7 8,55 7,95 8,99 8,6 9,94 9,58 10,8
7 9,29 9,31 9,47 9,83 10
Temp. 29 29,2
8 29,0
2 29,
3 28,9
2 29,0
7 29,0
3 29,1
1 29,4
4 29,8
1 29,3
6 29,7
6 29,9
7 29,7
1 29,7
8 29,6
6 CE 92 74 66 70 80 77 74 73 97 104 84 88 103 116 89 100 OD 7,47 7,7 7,5 7,7
2 7,47 7,39 7,49 7,34 7,23 7,26 7,13 7,23 7,41 7,3 7,19 7,28
DBO <L
Q 3 1,1 2 1,9 <LQ 1,3 0,9 <LQ <LQ <LQ <LQ <LQ <LQ <LQ <LQ
DQO 4 36 12 25 12 18 16 25 66 46 64 33 3,6 <LQ <LQ 3,5 Salin. 0,03 0,03 0,03 0,0
3 0,03 0,03 0,03 0,03 0,04 0,04 0,03 0,04 0,04 0,05 0,04 0,04
Fonte: Paiva et. al. (2019)
3.2. COMUNIDADE ZOOPLANCTÔNICA
A comunidade zooplanctônica, durante o período do estudo, foi representada por 7
filos, 18 famílias, 19 gêneros e 57 espécies, sendo o filo Rotifera o mais representativo
em número de espécies registradas e identificadas, seguido por Cladocera e Copepoda.
A densidade total dos organismos variou de 309 org./m³ (PT02, março/2018) a
8105 org./m³ (PT06, dezembro/2017), nas marés enchente e vazante respectivamente. Foi
observado diferença significativa das densidades entre os meses de março/2018 e
dezembro/2017 (ANOVA, F=5,44; p= 0,01) (Figura 2).
A densidade de Copepoda apresentou seus maiores valores no mês de
dezembro/2017 (maré vazante, PT06) e menores no mês de março/2018 (maré vazante,
PT 06). Foi obervado diferença significativa entre a densidade e os meses amostrados

309
Belém (PA), 11 a 13 de dezembro de 2019
(ANOVA, F=5,17; p= 0,01), onde o mês de março/2018 foi diferente dos meses de
dezembro/2017 e junho/2018. As elevadas densidades de Copepoda foram atribuídas a
uma maior contribuição das formas iniciais de naúplios e copepoditos, os quais podem
estar relacionados a uma estratégia adaptativa em condições adversas da região de estudo,
uma vez que a produção de muitos descendentes garante que uma pequena parte deles
alcançam a idade adulta (PINHEIRO et al., 2019).
A densidade de Cladocera apresentou elevados valores no mês de dezembro/2017
(maré vazante, PT 02) e menor no mês de junho/2018 (maré vazante, PT 02). Foi
observada variação significativa entre a densidade de Cladocera e os meses amostrados
(Anova, F= 7,51; p=0,004) e sazonalidade (F=8,7; p= 0,01). A densidade dos Rotifera
apresentou valores máximos no mês de dezembro/2017 (maré vazante, PT02) e mínimo
em junho/2018 (maré vazante, PT02). Entre estas densidades foi observada diferença
significativa entre os meses amostrados (Anova, F=14,18; p= 0,0030) e sazonalidade
(Anova, F=6,84; p=0,02).
De forma geral, foi observado que as maiores densidade zooplanctônica foram
registradas no mês de dezembro/2017, considerado como período chuvoso, onde há uma
maior descarga de águas provenientes dos afluentes da Baía do Marajó, bem como o
aumento do volume de água, precipitação pluviométrica, o qual aumenta o contato do rio
com as margens, além dos processos de lixiviação, o que pode acarretar o aumento das
concentrações de nutrientes, aumentando a oferta de alimento, além de criar maior
número de habitats (CAMPOS et al., 1996).

310
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2. Densidade total dos organismos zooplanctônicos registrados durante o período
do estudo no Porto de Vila do Conde (Pará-Brasil).
Fonte: Paiva et. al. (2019).
3.2.2. Frequência de Ocorrência (FR)
Os grupos zooplanctônicos que mais se destacaram foram: Rotifera (48%), seguido
pelos Cladocera (28%), Copepoda (24%) (Figura 3).
Os organismos classificados com muito frequente (≥ 70%), em ambos os períodos
sazonais, foram: copepodito de Calanoida (100%), Copepodito de Cycloploida (100%),
naúplio (100%), Diaphanosoma birgei (100%), Moina minuta (100%), Filinia terminalis
(100%), Keratella americana (100%), Keratella cochelaris (100%), Brachionus mirus
(81%) e Filinia opoliensis (75%). Estas espécies são comumente registradas para a região
portuárias paraenses, destacando os gêneros Keratella, Filinia e Brachionus, os quais são
característicos de ambientes em processos de alterações ambientais (COSTA et al., 2016).
Figura 3. Frequência de Ocorrência da comunidade zooplanctônica registrada durante o
período do estudo no Porto de Vila do Conde (Pará-Brasil).
Fonte: Paiva et. al. (2019)

311
Belém (PA), 11 a 13 de dezembro de 2019
3.2.3. Correlações
Em relação à correlação de Spearman, não foi observado correlações significativas
entre as densidades totais de organismos com os fatores físico-químicos da água. No
entanto, a densidade de Cladocera apresentou correlação negativa com o pH (R= - 0,64),
T° (R= -0,50) e CE (R= -0,52), demonstrando que esse grupo é altamente sensível a
mudanças ambientais, uma vez que os valores de condutividade indicam a interferência
de atividades antrópicas como citado por Medeiros et al (2018). E correlação positiva
com o OD (R= 0,7). Para a densidade de Rotifera foi registrado uma correlação negativa
com pH (R= -61) e positiva para OD (R= 0,65). Para os demais fatores não foi registrado
correlações significativas.
4. CONSIDERAÇÕES FINAIS
A estrutura da comunidade zooplanctônica, durante o período do estudo, esteve
correlacionada principalmente com a sazonalidade, onde foi observado elevada densidade
de organismos durante o período mais chuvoso, principalmente pelas fases iniciais de
naúplio e copepodito. No entanto, o padrão de variação dos organismos pode também
estar relacionado às condições ambientais da região, com constantes pressões antrópicas.
Vale ressaltar que o Porto de Vila do conde está situado numa zona de intensa atividade
portuária e industrial, historicamente impactado pelo lançamento de efluentes e rejeitos.
Desse modo, faz-se necessário o biomonitoramento da comunidade zooplônctonica, com
ênfase nas espécies indicadoras das condições tróficas e consequentemente na qualidade
da água superficial das regiões portuárias amazônicas.

312
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
AZEVEDO, F. A.; CHASIN, A. A. M. Metais: gerenciamento da toxicidade. São
Paulo: Atheneu, 2003.
CAMPOS, J. R. C., LANSAC- TÔHA, F. A., NUNES, M. A., GARCIA A. P. P.;
PRADO, F. R. Composição da comunidade zooplanctônica de três lagoas da ilha de
Porto Rico na planície de inundação do alto rio Paraná. Acta Limnologica
Brasiliensia, v. 8, p. 183-194, 1996.
CARDOSO, L. S.; RAMOS, I. D, MELLO, H. O. O. Composição, densidade e
abundância das populações de Cladocera, Copepoda e Rotifera de áreas de proteção
permanente do Rio Uberlandinha. Em extensão v. 7, p. 95-107. 2008.
CARVALHO, A. R.; SCHLITTLER, F. H. M.; TORNISIELO, V. L. Relações da
atividade agropecuária com parâmetros físicos químicos da água. Química Nova,
cap. 23, v.5, p. 618-622. 2000.
CDP - COMPANHIA DOCAS DO PARÁ. Atualização do plano de desenvolvimento
e zoneamento do porto de Vila do Conde, situado no Município de Barcarena,
Belém/Pará. Belém: CDP, 2010. Disponível em:
http://www2.cdp.com.br/arquivo/PDZ/pdz_pvc.pdf. Acesso em: 01 fev. de 2019.
COELHO-BOTELHO, M. J. Influência da transposição das águas do reservatório
Billings para o reservatório Guarapiranga (São Paulo) na comunidade
zooplanctônica. I. Período chuvoso (1997 a 2001). Em: Resumos do XXIV Congresso
Brasileiro de Zoologia, Itajaí-SC, 17 a 22 de fevereiro de 2002.
COSTA, B. N. S.; PINHEIRO, S. C. C.; LIMA, M. O.; AMADO, L. L.
Microzooplankton as an indicator of environmental quality at an industrial complex
in the Brazilian Amazon. Ecological Indicators, v. 66, p. 220–229, 2016.
GREGÓRIO, A. M.; MENDES, A. Characterization of sedimentary deposits at the
confluence of two tributaries of the Pará River estuary (Guajará Bay, Amazon).
Continental Shelf Research, v. 29, p. 609-618, 2008.
MEDEIROS, W. M. V., SILVA, C. E., LINS, R. P. M. Avaliação sazonal e espacial
da qualidade das águas superficiais da bacia hidrográfica do rio Longá, Piauí,
Brasil Revista Ambiente & Água [online]. v.13, n.2, 2018.
NEUMANN-LEITÃO, S., MATSUMURA-TUNDISI, T. & CALIJURI, M. C.
Distribuição e aspectos ecológicos do zooplâncton da Represa do Ribeirão do Lobo
(Broa) São Paulo. In: ENCONTRO BRASILEIRO DE PLANCTON, 4., RECIFE,.
Anais. Recife, UFPE, p. 393-413, 1991.
PINHEIRO, S.; LIMA, M.; CARNEIRO, B.; TAVARES, V. C.; CÂMARA, V. Effects
of a shipwreck on the zooplankton community in a port region of the Amazon.
Environmental Science and Polluition Research, v. 26, p. 5738-5750, 2019.

313
Belém (PA), 11 a 13 de dezembro de 2019
PIRATOBA, A. R. A.; RIBEIRO, H. M. C.; MORALES, G. P.; GONÇALVES. W. G.
Caracterização de parâmetros de qualidade da água na área portuária de
Barcarena, PA. Brasil Ambiente & Água - An Interdisciplinary Journal of Applied
Science, vol. 12, p. 436-456, 2017.
RAMOS, J. F. F. Poluição e contaminação da orla de Belém-PA. Em : A questão da
água na grande Belém. Casa de Estudos Germânicos (Fundação Heinrich Boll), p. 121-
148. 2004.
SANTOS, T. G. Zooplâncton como indicador da qualidade ambiental dos estuários
dos rio Carrapicho e Botafogo, Itamaracá – PE. Dissertação (Mestrado em
Oceanografia) – Universidade Federal de Pernambuco, Recife, 2008.

314
Belém (PA), 11 a 13 de dezembro de 2019
DISTRIBUIÇÃO ESPACIAL DA REGENERAÇÃO NATURAL EM RELAÇÃO
À ÁRVORE MATRIZ DE Euxylophora paraensis Huber EM FRAGMENTO
FLORESTAL NO MUNICÍPIO DE PARAGOMINAS-PARÁ
Ariel Barroso Monteiro¹; Joyce Ananda Paixão Duarte²; Gracialda Costa Ferreira³;
Jakeline Prata de Assis Pires 4
1 Graduanda em Engenharia Florestal, Universidade Federal Rural da Amazônia,
2 Doutoranda, Programa de Pós Graduação em Ciências Florestais, Universidade
Federal Rural da Amazônia. [email protected]
3 Professora do Instituto de Ciências Agrárias; Universidade Federal Rural da
Amazônia, [email protected]
4 Professora Assistente I, Departamento de Biologia, Pontifícia Universidade católica do
Rio de Janeiro, [email protected]
RESUMO
A fragmentação florestal é uma ameaça presente em todos os ecossistemas florestais do
planeta. Dentre as consequências ocasionadas pela fragmentação, destaca-se a diminuição
da riqueza de espécies e a densidade destas, podendo levar muitas espécies a extinção
local. Euxylophora paraenses Huber, conhecida como pau-amarelo, pertence a família
Rutaceae, com ocorrência restrita no norte da Amazônia e ameaçada de extinção como
“criticamente em perigo”. O estudo objetivou avaliar a dispersão e o alcance da
regeneração natural de Euxylophora paraensis Hub. em relação árvore matriz no
fragmento florestal localizado no município de Paragominas-Pará. O trabalho contou com a avaliação de 10 matrizes da espécie, verificando a organização do recrutamento de
mudas e arvoretas em parcelas com raio de 20 metros. Dessa forma, observou-se que há
uma tendência relação inversamente proporcional entre o tamanho da copa e o número de
indivíduos recrutados próximo à copa que está relacionada à disponibilidade de luz, à
sobrevivência e competição intraespecífica e à uma dispersão secundária das sementes,
após a barocoria.
Palavras-chave: Copa de árvores. Espécie em extinção. Fragmentação.
Área de Interesse do Simpósio: Ecologia e Biodiversidade

315
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A fragmentação florestal é uma ameaça presente em todos os ecossistemas
florestais do planeta. Este evento é ocasionado principalmente pelo aumento contínuo da
população e pressões econômicas geradas no decorrer da história, ou seja, a expansão das
atividades socioeconômicas tem sido a principal responsável pela modificação (PIRES et
al., 2006).
Dentre as consequências ocasionadas pela fragmentação, destaca-se a diminuição
da riqueza de espécies e a densidade destas. Mesmo que uma espécie rara esteja presente
num dado fragmento quando este é isolado, sua população local será tão pequena que terá
poucas chances de sobrevivência em longo prazo, o que pode levar ao declínio
populacional, redução da biodiversidade e espécies à extinção local (LAURENCE &
VASCONCELOS, 2009).
Euxylophora paraensis Huber pertence a família Rutaceae, é monoespecífico do
gênero com ocorrência restrita no norte da Amazônia (PIRANI, 2016; CNCFLORA,
2012). É uma espécie arbórea de grande porte que alcança o dossel da floresta, variando
entre 40 e 50 metros de altura (MARGALHO et al., 2014). A espécie produz madeira
pesada e de boa qualidade (NEGRÃO et al., 2013) o que ocasionou uma intensa
exploração no passado, tornando-a potencialmente suscetível ao impacto e levando a
redução populacional (MARTINI et al., 1998). Devido a intensa exploração, associada
com a distribuição restrita, a baixa densidade populacional e o intenso desmatamento,
esta entrou na lista de espécies ameaçadas de extinção classificadas como criticamente
em perigo” (IBAMA, 2014).
Desta forma, tem-se buscado estudar a ecologia desta espécie de forma a subsidiar
informações que promovam o manejo adequado desta espécie em fragmentos florestais,
como a identificação da distribuição espacial da regeneração natural em relação à árvore-
matriz. Conhecer a dispersão da regeneração natural em fragmentos florestais tem grande
relevância, pois estes são os novos indivíduos que irão substituir a vegetação antiga,
promovendo o rejuvenescimento e a perpetuação da floresta (RODRIGUES &
NASCIMENTO, 2006).

316
Belém (PA), 11 a 13 de dezembro de 2019
Desta forma, o presente estudo buscou avaliar a dispersão e o alcance da
regeneração natural de Euxylophora paraensis Huber em relação árvore matriz no
fragmento florestal localizado no município de Paragominas-Pará.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
O estudo foi conduzido em um fragmento florestal, localizado em uma área com
atividade minerária, denominada Platô Miltônia III, pertencente à empresa Mineração
Paragominas S.A. que, atualmente, está sob a administração do grupo Hydro. A área está
localizada no município de Paragominas na mesorregião do Sudeste Paraense, nas
coordenadas 3°15'50"S e 47°44'19"W (Figura 2). A mineração de bauxita em
Paragominas fica a 64 km da área urbana e 350 km da capital Belém.
Figura 1: Mapa de localização da área de estudo e das matrizes de Euxylophora
paraensis Huber no fragmento florestal no município de Paragominas - Pará.
Fonte: Joyce Duarte, 2019
A região apresenta clima equatorial do tipo quente e úmido, temperatura média
anual de 26,3ºC, umidade relativa do ar média de 81% (BASTOS et al. 2006). A
pluviosidade média anual é de 1.800 milímetros, com o período mais chuvoso de
dezembro a maio, e o mais seco de junho a novembro, evidenciando o período de estiagem

317
Belém (PA), 11 a 13 de dezembro de 2019
(BASTOS et al., 2006). O relevo é plano a suavemente ondulado, com altitude média de
200 metros em relação ao nível do mar. O solo de Paragominas é do tipo latossolo amarelo
distrófico, com alto grau de intemperismo, profundos, ácidos e ricos em alumínio
(RODRIGUES et al. 2003).
2.2 COLETA DE DADOS
Foram selecionadas dez matrizes com indivíduos regenerantes próximo à árvore-
matriz. A metodologia empregada para análise da regeneração natural foi uma adaptação
utilizada por Cetnarski-Filho e Nogueira (2004), utilizando uma parcela circular, em cada
matriz, com 20 metros de raio tendo como centro da parcela o fuste da árvore. Foi
amostrada uma área total de 1,26 hectares.
A regeneração foi identificada, marcada e contabilizada em cada matriz. Para avaliar
o efeito da distância da regeneração natural em relação à matriz, foi obtida a localização
de cada indivíduo pelas coordenadas cartesianas as quais foram, posteriormente,
transformadas em coordenadas geográficas.
Cada árvore-matriz teve sua área de projeção da copa mensurada com uma trena a
partir do centro da parcela, a cada 45º em relação a matriz, totalizando oito medições para
cada árvore. A partir da obtenção desses dados foi possível plotar e calcular a área da
copa.
A localização da regeneração natural e a obtenção das distâncias em relação à árvore
matriz à matriz, bem como o cálculo da área da projeção da copa foram obtidas com o
auxílio do programa ArcGis 10.1.
2.3 ANÁLISE DE DADOS
Foi calculado o grau de correlação de Pearson entre o número de indivíduos em
relação a área de copa e o distanciamento da árvore matriz. Para isto foram utilizadas as
variáveis: número de indivíduos, distância da regeneração natural e área de copa.

318
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
Nas dez parcelas medidas foram registrados 594 indivíduos regenerantes vivos
encontrados no monitoramento. O maior número de indivíduos foi encontrado na matriz
15 e a menor abundância foi encontrada na matriz 72 (Tabela 1).
Tabela 1: Densidade, área de copa, distância mínima e média da regeneração natural de
cada árvore-matriz de Euxylophora paraensis Huber do fragmento no município de
Paragominas - Pará.
Matriz N°
Indivíduos
Área de Copa
(m²)
Distância mínima
(m)
Distância média
(m)
M1 53 229,9 4,43 13,4
M2 46 197,9 8,47 14,6
M3 129 73,3 1,33 11,9
M4 118 38,2 8,45 13,7
M5 54 71,4 4,32 12
M6 59 79,7 5,75 15,7
M7 27 92,1 7,22 14,7
M8 5 509,4 9,52 12,9
M9 38 90,8 7,48 16,5
M10 65 43,6 3,32 9,9
Média 59,4 161,9 5,77 13,2
Fonte: Joyce Duarte, 2019
O número de plantas localizadas ao redor da matriz em relação a área de copa
apresentou correlação negativa e inversamente proporcional (r = -0,607; p < 0,05). Em
matrizes com maior número de regenerantes, a copa é reduzida em área. A matriz com
maior área de copa tinha apenas cinco indivíduos da regeneração natural (Figura 2). O
ambiente situado sob as copas de árvores é influenciado pelo tamanho de área e
arquitetura da copa que estas apresentam (MELO et al., 2015), interferindo na entrada de
luz no sub-bosque ou sobre o solo que, consequentemente, afetam a temperatura
superficial do solo, a germinação e o estabelecimento de espécies (HUBELL, 1999;
GUARIGUATA & OSTERTAG, 2001; AMIR, 2012). Por criarem sob si distintas
condições abióticas e bióticas, limitariam em maior ou menor grau o recrutamento, a
abundância, a sobrevivência e a distribuição espacial das espécies dispersas sob suas
copas (GANDOLFI, 2003; MELO ET AL., 2015).

319
Belém (PA), 11 a 13 de dezembro de 2019
A distância mínima entre a árvore e a regeneração natural foi de 1,33 metros,
sendo este um indivíduo com altura menor que 1,50 metros e localizado sob a copa da
árvore matriz (Tabela 1; Figura 2). Apenas 5,63% das mudas estavam sob a copa das
matrizes e o restante situa-se fora da projeção da copa. A distância média, considerando
as dez árvores matrizes foi de 13,5 metros. Esse resultado pode ser explicado pela alta
taxa de predação das sementes próximo à planta-mãe, pelo sombreamento sob a copa,
dormência de sementes e o tipo de dispersão da espécie (CORRÊA & MOURA, 2011).
De acordo com a teoria de Escape postulada por Janzen-Connell (JANZEN, 1970;
CONNELL, 1971) afirmam que através da dispersão, sementes e plântulas podem escapar
da alta mortalidade, causada por patógenos, predadores de sementes ou herbívoros, que
ocorre debaixo de plantas-mães. Desta forma, indivíduos distantes da planta-mãe tem
mais chances de sobrevivência no ambiente. As sementes dispersas muito além das copas
parentais podem representar indivíduos de maior qualidade de dispersão, pois estes têm
maior probabilidade de sobrevivência até a idade adulta devido escaparem da mortalidade
dependente da distância (CRAMER et al., 2007).

320
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2: Projeção de copa e localização da regeneração natural e projeção das árvores matriz de
Euxylophora paraensis Huber do fragmento no município de Paragominas - Pará.
A
D
B
F E
Matriz 1 Matriz 3
Matriz 2
Matriz 4
Matriz 7
Matriz 8
Matriz 5
C

321
Belém (PA), 11 a 13 de dezembro de 2019
Fonte: Joyce Duarte, 2019
Apesar da síndrome de dispersão primária de E. paraensis ser barocórica, em que
as sementes são dispersas pela força da gravidade tendem a cair próximo à planta mãe
(BRANDÃO, et al., 2018), algumas espécies podem apresentar dispersão secundária por
animais que podem promover um distanciamento das sementes da árvore-mãe
(ZIMMERMAN et al., 2015). Apesar deste estudo não ter avaliado os tipos de dispersores, foi
observado durante o monitoramento, a presença de araras e macacos no período de
frutificação de E. paraensis. Silva (2015) registrou cavidades nos troncos de E. paraensis
na região do mosaico Carajás-Pará sendo utilizadas como ninho para a reprodução de
Anodorhynchus hyacinthinus - Arara azul, sendo esta uma espécie que pode contribui de alguma
forma na dispersão das sementes de E. paraensis sendo necessários estudos que comprovem esse
evento.
H G
I J Matriz 6
Matriz 10
Matriz 9
Matriz 5

322
Belém (PA), 11 a 13 de dezembro de 2019
Além disso, em estudo fitoquímico, Arruda & Silva et al. (2012), detectaram
presença de cumarinas na espécie, um metabólito que segundo Souza-filho & Alves
(2000) tem sido apontado como um metabólito com potencial alelopático o que pode ser
um fator que pode interferir no recrutamento da regeneração próximo a planata-mãe. A
alelopatia, segundo Ferreira & Áquila (2000), é a capacidade de atrair ou repelir outros
organismos mediante a liberação de compostos químicos no ambiente.
4. CONCLUSÃO
A regeneração natural de Euxylophora paraensis Huber tem maior número de
indivíduos distante e fora da projeção da copa da árvore matriz. Foi identificado que a
copa influencia na distribuição da regeneração natural, onde quanto menor a copa, maior
o número de indivíduos e isso se deve, provavelmente, ao sombreamento que esta
promove no sub-bosque, interferindo no recrutamento de novos indivíduos.

323
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ARRUDA, A. C.; DA CRUZ SILVA, S. Fitoquímica e quimiossistemática de
Euxylophora paraensis (Rutaceae). Quim. Nova, v. 35, n. 11, p. 2119-2124, 2012.
AMIR, A. A. Canopy gaps and the natural regeneration of Matang mangroves. Forest
Ecology and Management, v. 269, p. 60-67, 2012.
BRANDÃO, A. D. D. S. Análise espacial de euxylophora paraensis Huber na
Amazônia Oriental. 2018. Tese de Doutorado. UFRA.
CONNELL J. H. On the role of natural enemies in preventing competitive exclusion in
some marine animals and in rain forest trees. In: Boer PJ, Gradwell GR, editores.
Dynamics of populations. Wageningen: Center for Agricultural Publishing and
Documentation; 1971.
CNCFlora. Euxylophora paraensis in Lista Vermelha da flora brasileira versão 2012.2
Centro Nacional de Conservação da Flora. Disponível em
<http://cncflora.jbrj.gov.br/portal/pt-br/profile/Euxylophora paraensis>. Acesso em 22
outubro 2019.
DE MELO, A. C. G.; DARONCO, C.; RÉ, D. S.; DURIGAN, G. Atributos de espécies
arbóreas e a facilitação da regeneração natural em plantio heterogêneo de mata ciliar.
Scientia Forestalis/Forest Sciences, p. 333-344, 2015.
FERREIRA, A. G.; AQUILA, M. E. A. Alelopatia: uma área emergente da ecofisiologia.
Revista Brasileira de Fisiologia Vegetal, v. 12, n. 1, p. 175-204, 2000.
GANDOLFI, S. Regimes de luz em florestas estacionais semideciduais e suas possíveis
conseqüências. Ecossistemas brasileiros: manejo e conservação (VC Sales, org.).
Expressão Gráfica e Editora, Fortaleza, p. 305-311, 2003.
GUARIGUATA, M. R.; OSTERTAG, R. Neotropical secondary forest succession:
changes in structural and functional characteristics. Forest ecology and management, v.
148, n. 1-3, p. 185-206, 2001.
IBAMA. (2019) INSTITUTO BRASILEIRO DO MEIO AMBIENTE E DOS
RECURSOS NATURAIS RENOVÁVEIS. Lista de Espécies Ameaçadas de Extinção.
<https://www.ibama.gov.br/flora-e-madeira/especies-ameacadas-de-extincao/especies-
da-flora-ameacadas-de-extincao. Acessado em: outubro de 2019>. Acessado em: 21 Out
2019.
JANZEN, D. H. Herbivores and the number of tree species in tropical forests. American
Naturalist 1970; 104(940): 501-528. http://dx.doi.org/10.1086/282687.
LAURANCE, W. F.; VASCONCELOS, H. L. Conseqüências ecológicas da
fragmentação florestal na Amazônia. Oecologia brasiliensis, 2009.
MARGALHO, L. F., GURGEL, E. S. C., GOMES, J. I., GROPPO, M., MARTINS-DA-
SILVA, R. C. V., DE CARVALHO, L. T., DE SOUZA, A. S. Conhecendo espécies de
plantas da Amazônia: pau-amarelo (Euxylophora paraensis Huber-Rutaceae). Embrapa
Amazônia Oriental-Comunicado Técnico (INFOTECA-E), 2014.

324
Belém (PA), 11 a 13 de dezembro de 2019
MARRA, D. M. Distância da planta-mãe e estabelecimento de plântulas de Scleronema
micranthum (MALVACEAE) em uma floresta de terra firme na Amazônia Central. EFA-
Ecologia da Floresta Amazônica, 2008.
MARTINI, A.; DE ARAÚJO ROSA, N.; UHL, C.. Espécies de árvores potencialmente
ameaçadas pela atividade madeireira na Amazônia. IMAZON, Instituto do Homem e
Meio Ambiente da Amazônia, 1998.
NEGRÃO, E. N., DOS SANTOS, T. F., PANTOJA, M. V., & MAUÉS, M. M.
Características sobre a biologia da polinização e dispersão de sementes de espécies
prioritárias para o manejo florestal no estado do Pará. In: Embrapa Amazônia Oriental-
Resumo em anais de congresso (ALICE). In: CONGRESSO NACIONAL DE
BOTÂNICA, 64.; ENCONTRO REGIONAL DE BOTÂNICOS MG, BA E ES, 23.,
2013, Belo Horizonte. Anais... Belo Horizonte: SBB, 2013.,
PIRANI, J.R.; GROPPO, M.; DIAS, P. Rutaceae in Flora do Brasil 2020 em
construção. Jardim Botânico do Rio de Janeiro. Disponível em:
<http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB620>. Acesso em: 23 Out. 2019.
PIRES, A. S.; FERNANDEZ, F. A.; BARROS, C. S. Vivendo em um mundo em pedaços:
efeitos da fragmentação florestal sobre comunidades e populações animais. Biologia da
conservação: essências, p. 231-260, 2006.
RODRIGUES, P. J. F. P.; NASCIMENTO, M. T. Fragmentação florestal: breves
considerações teóricas sobre efeitos de borda. Rodriguésia, p. 63-74, 2006.
RIBBENS, E.; SILANDER JR, J. A.; PACALA, S. W. Seedling recruitment in forests:
calibrating models to predict patterns of tree seedling dispersion. Ecology, v. 75, n. 6, p.
1794-1806, 1994.
SILVA, G. F. Aspectos da biologia reprodutiva da arara-azul Anodorhynchus
hyacinthinus (Latham, 1790) no mosaico Carajás-PA. 2015.
SOUZA FILHO, A. P. S.; ALVES, S. de M. Potencial alelopático de plantas de acapu
(Vouacapoua americana): efeitos sobre plantas daninhas de pastagens. Embrapa
Amazônia Oriental-Artigo em periódico indexado (ALICE), 2000.

325
Belém (PA), 11 a 13 de dezembro de 2019
EFEITOS DA ADIÇÃO DE NUTRIENTES SOBRE O FITOPLÂNCTON DO
RESERVATÓRIO DE ABASTECIMENTO BOLONHA (BELÉM-PARÁ)
Samara Letícia dos Santos Pinto1; Paola Vitória Brito Pires2; Bruno Santana Carneiro3;
Eliane Brabo de Sousa 4
1 Graduação em Biologia. Universidade Federal do Pará. [email protected];
2 Especialista em Biologia Ambiental. Universidade Rural da
[email protected]; 3 Doutorado em Saúde Coletiva. Instituto Evandro Chagas. [email protected];
4 Doutorado em Saúde Coletiva. Instituto Evandro Chagas. [email protected]
Orientadora
RESUMO
A dinâmica do fitoplâncton em ambientes artificiais controlados com diferentes
quantidades de nutrientes pode sugerir um possível cenário de Bloom algal em
reservatórios. Para verificar os efeitos da adição de nutrientes sobre o desenvolvimento
do fitoplâncton na ausência de macrófitas aquáticas foram feitos experimentos em
ambientes simulados (microcosmo). A coleta de água foi realizada no reservatório
Bolonha (Belém, Pará) no mês de novembro/2018, referente ao período seco. As águas
foram distribuídas em quatro experimentos com tréplicas, cada, os quais se diferenciaram
pelas concentrações na proporção nitrogênio e fósforo: experimento controle;
experimento E1 (nitrogênio > fósforo), experimento E2 (nitrogênio = fósforo) e
experimento E3 (nitrogênio < fósforo). Os experimentos foram acondicionados em
câmara de crescimento de microalgas sob contínua aeração, temperatura 28 ± 2 °C,
luminosidade de 100 µmol.fótons.m2s-1 e fotoperíodo de 12 horas por 40 dias. A cada 5
dias foram retirados 250 mL de amostras para análise físico-química, concentrações de
nutrientes e fitoplâncton. Foram identificadas 42 espécies com dominância das classes
Euglenophyceae, Cyanophyceae, Bacillariophyceae e Chlamydophyceae. A baixa
relação nitrogênio e fósforo não promoveu a floração de cianobactérias, mas de
Chlamydomonas sp., alga verde que cresce em condições de elevada quantidade de
nutrientes, transparência e águas paradas. Sugere-se que em ambientes naturais a espécie
Chlamydomonas sp. tem seu crescimento limitado pelas macrófitas aquáticas nos
aspectos da competição por nutrientes e pelo sombreamento das águas, diminuindo a
entrada de luz e reduzindo a fotossíntese deste organismo. Sugere-se que o manejo de
macrófitas deve ser associado ao prévio conhecimento da comunidade fitoplanctônica
com e sem a presença destes vegetais.
Palavras-chave: Reservatório. Abastecimento. Microcosmos.
Área de Interesse do Simpósio: Ecologia e Biodiversidade

326
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Os principais mananciais de abastecimento da Região Metropolitana de Belém
são os lagos Bolonha e Água Preta, lagos artificiais que captam água do Rio Guamá,
corpo hídrico de grande porte que margeia parte da cidade de Belém.
Os lagos apresentam vestígios de eutrofização artificial representados,
principalmente, pela presença de macrófitas aquáticas na superfície dos lagos. Estes
vegetais são utilizadas como indicadores da qualidade da água (KLUMPP et al., 2002),
devido seu potencial em absorver nutrientes e até bactérias fecais (MEES et al., 2009),
logo sua intensa proliferação indica que o ambiente se encontra eutrofizado, sendo os
nutrientes, principalmente nitrogênio e fósforo, oriundos do esgoto doméstico, visto que
a cidade possui um baixo índice de saneamento básico.
Também como consequência da eutrofização está o crescimento das
cianobactérias que em altas densidades liberam, para o ambiente aquático, substâncias
tóxicas ao homem pondo em risco a saúde da população consumidora. Embora o lago
Bolonha apresente-se eutrofizado não se identificou o crescimento de cianobactérias. Isto
se deve possivelmente as macrófitas que competem com o fitoplâncton (incluindo
cianobactérias) pelos mesmos recursos. Logo, a remoção das macrófitas pode propiciar
as florações de cianobactérias.
A dinâmica destes organismos em diferentes condições de nutrientes pode sugerir
um possível cenário de Bloom algal para o lago Bolonha e direcionar as estratégias de
monitoramento e gerenciamento deste reservatório. Portanto, este trabalho tem como
objetivo verificar os efeitos da adição de nutrientes (nitrogênio e fósforo) sobre o
desenvolvimento do fitoplâncton, em ambientes simulados (microcosmo) sem a presença
das macrófitas.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO E ESTRATÉGIA AMOSTRAL
O lago Bolonha apresenta uma forma alongada, com 2,10 x 106 m3 de água
acumulada, 512.540 m2 de lâmina d’água, profundidade máxima em torno de 7,64 m e
uma área total de 1.790.000 m2 (COSANPA, 2004).

327
Belém (PA), 11 a 13 de dezembro de 2019
Neste lago foram coletados 25 a 30 litros de água no ponto de captação de água
bruta para a Estação de Tratamento de água- ETA São Brás (Figura 1) no mês de
novembro de 2018, referente ao período seco, em garrafões de polipropileno de 5 L os
quais foram resfriados até a chegada do laboratório para diminuir o metabolismo dos
organismos.
Figura 1. Reservatório Bolonha (Belém, Pará) mostrando o ponto de coleta na ETA
São Braz, imagem mostra a foto do ponto de coleta e nota-se a proliferação de
gramíneas e macrófitas sugerindo a eutrofização e assoreamento do reservatório.
Fonte: Sousa et al. (2017)

328
Belém (PA), 11 a 13 de dezembro de 2019
2.2 ENSAIOS EM MICROCOSMOS
Foram montados quatro experimentos com tréplicas, cada, os quais se
diferenciaram pela concentração na proporção nitrogênio e fósforo, totalizando 12
microcosmos:
Controle- experimento controle: 6 L de amostra mantidos em água do ambiente
natural, divididos em três erlenmeyers de 3 L contendo 2 L de amostra cada;
E1- experimento com proporção de nitrogênio maior do que o fósforo: 6 L de
amostra mantidos em água do ambiente natural com adição de nitrogênio e fósforo na
proporção de 75:25, divididos em três erlenmeyers de 3 L contendo 2 L de amostra cada;
E2- experimento com proporção de nitrogênio igual ao fósforo: 6 L de amostra
mantidos em água do ambiente natural com adição de nitrogênio e fósforo na proporção
de 50:50, divididos em três erlenmeyers de 3 L contendo 2 L de amostra cada;
E3- experimento com proporção de nitrogênio menor que o fósforo: 6 L de
amostra mantidos em água do ambiente natural com adição de nitrogênio e fósforo na
proporção de 25:75, divididos em três erlenmeyers de 3 L contendo 2 L de amostra cada.
As concentrações de nitrogênio e fósforo foram inseridas em cada experimento a
partir da determinação das concentrações prévias destes nutrientes nas amostras. Os sais
adicionados às soluções (experimento) foram nitrato de sódio (NaNO3), cloreto de amônio
(NH4Cl3) e potássio de fosfato (K2HPO4.3H2O). Para a montagem e adição dos nutrientes
foram realizados cálculos estequiométricos para determinar a concentração de N e P nos
compostos adicionais.
Após a montagem do experimento, os erlenmeyers foram acondicionados em
câmara de crescimento de microalgas sob contínua aeração, temperatura 28 ± 2 °C,
luminosidade de 100 µmol.fótons.m2s-1 e fotoperíodo de 12 horas com duração de 40 dias.
A cada cinco dias foram retirados 250 mL de subamostra de cada experimento,
incluindo suas réplicas, 50 mL para análise quali-quantitativa do fitoplâncton
(UTERMÖHL, 1958) e 200 mL para análise físico-química e de nutrientes (nitrogênio e
fósforo total).
2.3 ANÁLISE DO FITOPLÂNCTON
As amostras quali-quantitativas provenientes de cada bloco experimental foram
fixadas com lugol acético. O método de sedimentação de Utermöhl (1958) foi empregado

329
Belém (PA), 11 a 13 de dezembro de 2019
para a identificação dos táxons e a determinação do número de indivíduos por litro. O
método de contagem foi por transectos. A identificação, a nomenclatura e o
enquadramento taxonômico foram realizados de acordo com a literatura especializada.
2.4 ANÁLISE DE NUTRIENTES E FATORES FÍSICO-QUÍMICOS
Para as amostras do ambiente e dos experimentos foram realizadas as medições
das variáveis: temperatura, pH, condutividade elétrica, sólidos totais dissolvidos (STD),
salinidade e oxigênio dissolvido utilizando a Sonda multiparamétrica HI 9828 HANNA®.
Em laboratório foram analisados: Nitrogênio Total e Fósforo Total. Todos os métodos
analíticos empregados obedeceram às recomendações do Standard Methods for the
Examination of Waterand Wastewater (2012).
2.5 ANÁLISE DE DADOS
As análises estatísticas descritivas e analíticas dos dados foram realizadas no
programa Statistica 7.0. A análise de variância foi realizada para testar se havia diferença
de densidades do fitoplâncton entre os diferentes experimentos.
3. RESULTADOS E DISCUSSÃO
Com relação à temperatura não se pode atribuir interferências na composição do
fitoplâncton, pois se trata de uma variável sensível e que se modificou em função do
processo de análise no laboratório durante as retiradas das amostras, neste sentido não
houve variação significativa (p<0,05) deste parâmetro. O pH se manteve alcalino durante
todo o experimento. A Condutividade elétrica variou de 110,46 µS.cm (controle) a
3739,29 µS.cm (E1). Oxigênio dissolvido se manteve em níveis superiores a 5 mg.L-1
considerando as águas oxigenadas. STD foi mais elevado no experimento E1 com
1955,19 mg.L-1. A salinidade variou de 0,05 (controle) a 2,03 (E1) (Tabela 1).
Não houve variação temporal significativa do fósforo total ao longo dos 40 dias
de observação do microcosmo. As concentrações de fósforo total foram
significativamente menores (F= 6,5; p< 0,05) no controle (8,95 ± 10,38 mg.L-1) e maiores
no experimento E2 (325,33 ± 85,18 mg.L-1).

330
Belém (PA), 11 a 13 de dezembro de 2019
Também não houve diferença significativa do nitrogênio total ao longo do tempo
do experimento. Entretanto, se observou significativamente (F= 12,62; p< 0,05) menor
concentração de nitrogênio no controle (1,35 ± 1,19 mg.L-1) e maior no experimento E1
(233,42 ± 45,85 mg.L-1). A relação nitrogênio e fósforo não variou significativamente ao
longo do tempo, apresentando uma relação pequena nos tempos iniciais T1 e T2. Entre
os experimentos, a relação nitrogênio e fósforo foi significativamente (F= 4,8; p< 0,05)
maior no controle (19,19 ± 30,72) e menor no E2 (0,31 ± 0,09) (Tabela 1).
Tabela 1. Estatística descritiva das variáveis físico-químicas nos experimentos realizados
em novembro/2018 com águas provenientes do Lago Bolonha (Belém, Pará).
Controle E1 E2 E3
Média ± Dp Média ± Dp Média ± Dp Média ± Dp
Temperatura (° C) 20,14 ± 0,26 19,84 ± 0,19 20,44 ± 0,18 20,63 ± 0,48
pH 7,41 ± 023 8,01 ± 0,05 8,96 ± 0,36 8,10 ± 0,73
Condutividade Elétrica 110,46 ± 14,55 3739,29 ± 23,38 2981,79 ± 43,31 3260,23 ± 466,25
Oxigênio Dissolvido 5,36 ± 0,14 6,10 ± 0,21 6,1 3 ± 0,45 6,35 ± 0,73
Sólidos totais dissolvidos 61,04 ± 8,37 1955,19 ± 345,13 1616,42 ± 22,04 1723,98 ± 426,18
Salinidade 0,05 ± 0,01 2,03 ± 0,02 1,59 ± 0,02 1,74 ± 0,32
F. Total 8,95 ± 10,38 95,60 ± 15,83 325,3 3 ± 85,18 161,32 ± 76,96
N. Total 1,35 ± 1,19 233,42 ± 45,85 89,54 ±33,99 166,31 ± 14,35
NT/PT 19,19 ± 30,72 2,69 ± 0,54 0,31 ± 0,09 0,96 ± 0,16
Fonte: Pinto et al. (2019)
Os experimentos apresentaram características de eutrofização, porém as baixas
razões de N/P (<10) não promoveram o crescimento de cianobactérias.
Ao longo do experimento foram encontradas 42 espécies. A densidade média foi
aumentando significativamente (F= 2,7; p< 0,05) ao longo do tempo do experimento
variando de 11,57 x 103 ind.L-1 a 1862,6 x 103 ind.L-1 nos tempos T1 e T8,
respectivamente (Figura 2A). A densidade foi significativamente menor no experimento
controle (100,37 ± 15,65 x 103 ind. L-1) e maior no experimento E3 (1651 ± 785 x 103
ind. L-1) (Figura 2B).

331
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2. Variação da densidade do fitoplâncton nos experimentos realizados em
novembro/2018 com águas provenientes do Lago Bolonha (Belém, Pará). A- variação temporal
e B- variação entre os experimentos.
Pinto et al. (2019)
Em termos de estrutura da comunidade fitoplanctônica, sua constituição foi
incorporada por organismos das classes Euglenophyceae, Cyanophyceae,
Bacillariophyceae e Chlamydophyceae, sendo este último dominante a partir do 20° dia,
compondo mais de 70% da densidade (Figura 3A). Os experimentos foram dominados
pela classe Chlamydophyceae havendo um elevado aumento de Bacillariophyceae nos
experimentos E2 e E3 (Figura 3B).
As classes que foram mais representativas durante o estudo podem ser encontradas
em ambientes rasos, ricos em matéria orgânica (nitrogênio e fósforo) (FRANCESCHINI
et al., 2010).
A classe Chlamydophyceae foi muito representativa durante o experimento, com
a espécie Chlamydomonas sp. sendo a mais dominante. Tais espécies são mais comuns e
abundantes em ambientes de águas mais transparentes com maior quantidade de
nutrientes (LACHI; SIPAÚBA-TAVARES, 2008). Este grupo esteve dominando em
todos os tratamentos, onde apresentou variação clara na densidade com 40% no controle
e 84% no experimento E1.

332
Belém (PA), 11 a 13 de dezembro de 2019
Figura 3. Variação da densidade relativa (%) das classes do fitoplâncton nos experimentos
realizados em novembro/2018 com águas provenientes do Lago Bolonha (Belém, Pará). A-
variação temporal e B- variação entre os experimentos.
Pinto et al. (2019)
As macrófitas aquáticas limitam o crescimento de Chamydophyceaes, pois em
condições naturais esta classe é pouco comum no reservatório Bolonha como observado
por Martins-da-Silva (1997), Costa et al., (2010) e Pires (2018). Essa limitação pode ser
por competição pelo consumo de nutrientes, pelo sombreamento fornecido pelas
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
T1 (5°) T2 (10°) T3 (15°) T4 (20°) T5 (25°) T6 (30°) T7 (35°) T8 (40°)
Den
sid
ad
e
Tempo (dias)
ACyanophyceae ChlorophyceaeEuglenophyceae TrebouxiophyceaeZygnematophyceae ChlamydophyceaeBacillariophyceae
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Controle E1 E2 E3
Den
sid
ad
e
Tipo de experimento
BCyanophyceae Chlorophyceae
Trebouxiophyceae Zygnematophyceae
Euglenophyceae Chlamydophyceae
Coscinodiscophyceae Bacillariophyceae

333
Belém (PA), 11 a 13 de dezembro de 2019
macrófitas ou pela condição de assoreamento do reservatório causado pelo
aprisionamento de sedimento nas suas raízes.
4. CONSIDERAÇÕES FINAIS
O aumento da densidade média do fitoplâncton durante o período do experimento
ocorreu concomitante ao decréscimo de nitrogênio e fósforo totais e da relação nitrogênio
e fósforo, sendo esta relação menor que 10, beneficiando os grupos de microalgas
Euglenophyceae e Chlamidophyceae, mostrando ainda que quanto maior a relação N/P
menor foi à densidade do fitoplâncton.
Além disso, este estudo mostrou a importância de entender as dinâmicas das
comunidades fitoplanctônicas em relação à descarga de nutrientes nos ambientes
aquáticos, visto que esta é uma grande problemática da região, onde os sistemas de
abastecimento de Belém são impactados pelo avanço da urbanização e a falta de
saneamento básico. Este experimento é pioneiro e precisa de repetições e ajustes para se
aproximar às condições do ambiente que se pretende simular.

334
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
AMERICAN PUBLIC HEALTH ASSOCIATION (APHA). Standard methods for the
examination of water and wastewater. American Public Health Association;
Washington: Unity State American, 2012. 1496 p.
COMPANHIA DE SANEAMENTO DO PARÁ (COSANPA). Plano Diretor do
Sistema de Abastecimento de Água da Região Metropolitana de Belém. Belém:
COSANPA, 2004.
FRANCESCHINI, I. M., BURLIGA, A. L., REVIERS, B., PRADO, J. F. & RÉZIG, S.
H., 2010. Algas – Uma abordagem filogenética, taxonômica e ecológica. 1ª Ed., Porto
Alegre, Artmed, 332p.
KLUMPP, A. et al. Variation of nutrient and metal concentrations in aquatic
macrophytes along the Rio Cachoeira in Bahia (Brazil). Environment International. v.
28, p. 165-171. 2002.
LACHI, G. B.; SIPAÚBA-TAVARES, L. H. Qualidade da água e composição
fitoplanctônica de um viveiro de piscicultura utilizado para fins de pesca esportiva e
irrigação. B. Inst. Pesca, São Paulo, v. 34, n. 1, p. 29 - 38, 2008.
MARTINS-DA-SILVA, R. C. V. Chlorellaceae (Chlorophyceae, Chlorococcales) do
Lago Água Preta, município de Belém, Estado do Pará. Boletim do Museu Paraense
Emílio Goeldi, Série Botânica, v.13, n.2, p.113-138, 1997.
MEES, J. B. R. et al. Removal of organic matter and nutrients from slaughterhouse
wastewater by using Eichhornia crassipes and evaluation of the generated biomass
composting. Revista Engenharia Agrícola. v. 29, n. 3, p. 466-473. 2009.
PIRES, P. V. B. O uso de grupos funcionais do fitoplâncton na determinação da
qualidade das águas e tróficas do reservatório Bolonha, Belém, Pará. 2018. Trabalho de
Conclusão de Curso (graduação em Ciências Biológicas), Instituto de Ciências
Biológicas, Universidade Federal do Pará. Belém, 2018.
UTERMÖHL H. Zur vervolkommung der quantitativen phytoplankton-Methodik.
Mitteilungen Internationale Vereiningung fuer Theoretische und Angewandte
Limnologie, v. 9, p. 1-38. 1958.

335
Belém (PA), 11 a 13 de dezembro de 2019
ANÁLISE DA EFICIÊNCIA DA COLETA DE MATERIAL BOTÂNICO PARA
A MELHORA DA IDENTIFICAÇÃO CIENTÍFICA DAS ESPÉCIES
ARBÓREAS NA FLONA DO TAPAJÓS
Jefferson Matheus Tavares de Oliveira1; Ademir Roberto Ruschel2.
1 Graduando em Engenharia Florestal. Universidade do Estado do Pará.
2 Orientador: Pesquisador-A. Embrapa Amazônia Oriental. [email protected].
RESUMO
Considerando a importância do auxílio dos herbários na identificação taxonômica, este
trabalho visa analisar o nível de eficácia das informações para a identificação de exsicatas.
Vários projetos de âmbito acadêmico ou empresarial necessitam de inventários florestais
confiáveis, com informações corretas acerca dos nomes científicos das espécies e, para
isso, dependem da identificação botânica. Estes produtos da identificação são de extrema
importância, pois subsidiam a construção de planos de manejo e auxiliam nas
informações sobre diversidade florística. Os dados foram coletados na Floresta Nacional
do Tapajós. Um universo amostral de 2.192 exsicatas foram coletadas, herborizadas e
analisadas pelo herbário IAN – Embrapa Amazônia Oriental. As informações dos laudos
da identificação foram tabuladas em planilhas eletrônicas Excel e as análises das
identificações foram comparadas em nível de família, gênero e espécie. O método de
coleta e identificação em herbário foi considerado eficaz, pois, mesmo com material
estéril, a identificação ao nível de espécie foi obtida para a maioria para as coletas.
Contudo, os menores ganhos foram obtidos para os espécimes da família Sapotaceae e
Lauraceae.
Palavras-chave: Botânica. Exsicatas. Sapotaceae.
Área de Interesse do Simpósio: Botânica

336
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Identificação botânica é a utilização de órgãos vegetativos e reprodutivos de
plantas, previamente determinada nos acervos dos herbários e literaturas especializadas,
para determinação da identidade de espécies coletadas no campo (MARTINS-DA-
SILVA et al, 2014). A identificação do material botânico é de extrema importância, pois
subsidia estudos taxonômicos e auxilia na construção de informações sobre diversidade
florística de áreas e planos de manejo adequados para espécies (FERREIRA, 2006).
Apesar de todo acervo taxonômico disponível nos herbários, ainda temos
dificuldades em avançar no que tange a identificação precisa de algumas espécies,
principalmente pela semelhança de seus órgãos vegetativos (BARBOSA, 2016), onde, na
maioria dos casos, é necessária a presença do órgão reprodutivo do vegetal (material
fértil), o que nem sempre é possível, pois o período de floração e frutificação varia entre
as espécies (BARBOSA, 2016; WOODSON, 1939).
Os métodos mais utilizados para o de identificação são a chave dicotômica e a
comparação de exsicatas. O primeiro é complexo e requer conhecimento em botânica para
a construção da chave (BRAZ; MOURA; ROSA, 2004), o segundo é utilizado nos
herbários, comparando as características das amostras coletadas com exsicatas
previamente identificadas (WIGGERS; STANGE, 2008).
Nos inventários florestais percebemos a adoção do nome vernacular ou popular
para identificação de espécies, geralmente obtido com o conhecimento empírico de
nativos da área (SECCO; PROCÓPIO, 2008). O uso do nome popular prejudica a
autenticidade do inventário, pois dificulta a identificação da ocorrência geográfica da
espécie, por existir grande quantidade de nomes populares associados a diferentes
espécies botânicas. Os nomes vernaculares mudam de acordo com região, uso na
comercialização e cultura (CAMARGOS et al, 2001). O presente estudo objetiva avaliar
a eficiência da coleta de material botânico para a melhoria da identificação à nível de
espécie no monitoramento de parcelas permanentes.

337
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
As exsicatas utilizadas no presente estudo foram coletadas nas parcelas
permanentes de inventário contínuo, situadas nas áreas experimentais do km-67 e km-114
da rodovia Santarém-Cuiabá. A Floresta Nacional do Tapajós é uma Unidade de
Conservação localizada no estado do Pará, abrange os municípios de Aveiro, Belterra,
Placas e Rurópolis, no Oeste do estado. O clima é tropical úmido, com temperatura média
anual de 25,5 ºC e variação térmica anual inferior a 5 ºC, classificado como Ami no
sistema Köppen. Segundo Silva et al. (1995), a precipitação média anual está em torno de
1.900-2.110 mm, apresentando grande variação no regime de chuvas durante o ano, com
as maiores precipitações ocorrendo nos meses de janeiro a maio.
A pesquisa foi baseada em materiais coletados pelo Laboratório de Manejo e
Conservação Florestal – Projeto Bom Manejo e identificados por parataxônomos do
Herbário IAN, ambos localizados na Empresa Brasileira de Pesquisa Agropecuária -
Embrapa Amazônia Oriental, Belém - PA. Foram analisadas 2.192 exsicatas coletadas
entre os anos de 2008 e 2017. As análises gráficas e estatísticas foram feitas em tabelas
eletrônicas (Microsoft Office – Excel), onde foram diferenciadas em famílias
predominantes e pelo nível de identificação, sendo família, gênero e espécie. Foram
analisados, também, quais erros eram predominantes associados a identificação das
exsicatas.
3. RESULTADOS E DISCUSSÃO
A Tabela 1 mostra a identificação das 2.192 exsicatas coletadas. Dessas 2.192
exsicatas, apenas três exsicatas (0,14%) ficaram indeterminadas em sua família botânica,
o que mostra a precisão da identificação ao nível de família, como mostra o Gráfico 1. As
famílias mais coletadas foram respectivamente: Fabaceae, Burceraceae, Lauraceae,
Sapotaceae, Moraceae, Myrtaceae, Lecythidaceae e Annonaceae. Tais oito famílias
acumularam 54% de todas as coletas, indicando representarem as famílias com maior
dificuldade de identificação das espécies em campo.

338
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1 – Famílias botânicas e número de espécies identificadas (Nº spp).
Família Nº spp Família Nº spp
Acanthaceae 1 Lamiaceae 6
Achariaceae 2 Lauraceae 199
Anacardiaceae 10 Lecythidaceae 106
Annonaceae 105 Lomariopsidaceae 1
Apocynaceae 41 Malpighiaceae 5
Aquifoliaceae 1 Malvaceae 32
Araceae 1 Melastomataceae 48
Araliaceae 4 Meliaceae 47
Arecaceae 3 Monimiaceae 1
Bignoniaceae 5 Moraceae 112
Bixaceae 2 Myristicaceae 31
Boraginaceae 29 Myrtaceae 112
Burseraceae 203 Nyctaginaceae 30
Calophyllaceae 2 Ochnnaceae 10
Capparaceae 1 Olacaceae 31
Cardiopteridaceae 1 Opiliaceae 4
Caryocaraceae 5 Passifloraceae 1
Cecropiaceae 1 Peraceae 3
Celastraceae 18 Phyllanthaceae 5
Chrysobalanaceae 36 Picramniaceae 1
Clusiaceae 10 Poaceae 1
Combretaceae 6 Polygonaceae 1
Connaraceae 9 Primulaceae 4
Dichapetalaceae 8 Putranjivaceae 7
Ebenaceae 13 Quiinaceae 9
Elaeocarpaceae 20 Rhizophoraceae 3
Erythroxylaceae 2 Rosaceae 7
Euphorbiaceae 43 Rubiaceae 87
Fabaceae 268 Rutaceae 10
Flacourtiaceae 6 Salicaceae 24
Gesneriaceae 1 Sapindaceae 56
Goupiaceae 3 Sapotaceae 183
Humiriaceae 11 Simaroubaceae 3
Hypericaceae 9 Siparunaceae 18
Icacinaceae 1 Solanaceae 5
Indetermidado 3 Sterculiaceae 1
Lacistemataceae 14 Styracaceae 3

339
Belém (PA), 11 a 13 de dezembro de 2019
Symplocaceae 6 Verbenaceae 2
Tiliaceae 3 Violaceae 55
Ulmaceae 4 Vochysiaceae 11
Urticaceae 6
Total Geral 2192
Fonte: Jefferson Oliveira, 2019
Um total de 526 exsicatas foram identificadas em nível de gênero,
correspondendo a 24% das amostras. A identificação do gênero, seja em campo ou no
herbário, depende das características morfológicas das plantas. Um estudo de Barroso
(1976) diferenciou o gênero Baccharis de acordo com a presença de cladódios
substituindo suas folhas, sua característica marcante.
Figura 1 - Nível de identificação à nível de espécie, gênero e família.
Fonte: Jefferson Oliveira, 2019.
A dificuldade de identificação botânica se deu pelos seguintes erros de coleta
identificados no estudo: coleta de material estéril e erro de codificação da coleta em
campo. Durante a análise, 35 exsicatas apresentaram erro de codificação de coleta em
campo, correspondendo a 1,6% das amostras. O equivalente 99,8% das exsicatas eram
estéreis, ou seja, não possuíam caracteres reprodutivos em sua coleta, o que dependeu do
acervo botânico do herbário para determinar sua espécie.

340
Belém (PA), 11 a 13 de dezembro de 2019
Segundo Martins-da-silva (2002), o sucesso da identificação botânica depende
dos seguintes fatores: modo como as amostras são coletadas, preparadas, transportadas
até o local de estudo e das anotações realizadas no campo.
4. CONCLUSÃO
O estudo mostrou a eficácia do método de coleta de material botânico
(exsicatas) para melhoria da determinação botânica em nível de espécie, mesmo quando
um quarto de todas as exsicatas ficou indefinida ao nível de espécie, ressaltando a
complexidade principalmente das espécies de um pequeno grupo de famílias, além da
inacessibilidade de material fértil durante a coleta de exsicatas em campo.

341
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BARBOSA, C. V. O. Revisão taxonômica de Geissospermum Allemão
(Apocynaceae). Dissertação de Mestrado. Universidade Federal Rural da Amazônica,
Belém. 58p, 2016.
BARROSO, G.M. Compositae - Subtribo Baccharidinae Hoffmann - Estudo das
espécies ocorrentes no Brasil. Rodriguésia 28: 1-273. 1976.
BRAZ, D.M.; MOURA, M.V.L.P.; ROSA, M.M.T da. Chave de identificação para as
espécies de Dicotiledôneas arbóreas da Reserva Biológica do Tinguá, RJ, com base
em caracteres vegetativos. Acta bot. bras. 18(2): 225-240. 2004.
CAMARGOS, J. A.; CZARNESKI, C.M.; MEGUERDITCHIAN, I.; OLIVEIRA, D. de.
2001. Catálogo de árvores do Brasil. Brasília: IBAMA/ Laboratório de Produtos
Florestais. Brasília. 643-644p.
FERREIRA, G. C. Diretrizes para coleta e identificação de material botânico. Belém-
PA: Embrapa, 2006.
MARTINS-DA-SILVA, R. C. V. Coleta e identificação de espécimes botânicos -
Belém: Embrapa Amazônia Oriental, 2002. 40p. 21cm. - (Embrapa Amazônia
Oriental. Documentos, 143).
MARTINS-DA-SILVA, R. C. V.; SILVA, A. S. L.; FERNANDES, M. M.;
MARGALHO, L. F. Noções morfológicas e taxonômicas para identificação botânica.
– Brasília, DF: Embrapa, 2014.
PROCÓPIO, L. C.; SECCO, R. S. 2008. A importância da identificação botânica nos
inventários florestais: o exemplo do "tauari" (Couratari spp. e Cariniana spp. -
Lecythidaceae) em duas áreas manejadas no Estado do Pará. Acta Amazonica 38 (1): 31-
44.
SILVA, J. N. M.; CARVALHO, J. O. P.; LOPES, J. do C. A. Inventário florestal de
uma área experimental na Floresta Nacional de Tapajós. Boletim de Pesquisa
Florestal, Colombo, n. 10-11, p. 38-110, jun./dez. 1985.
SILVA, J. N. M.; CARVALHO, J. O. P. de; LOPES, J. do C. A.; ALMEIDA, B. F. de;
COSTA, D. H. M.; OLIVEIRA, L. C. de; VANCLAY, J. K.; SKOVSGAARD, J. P.
Growth and yield of a tropical rain forest in the Brazilian Amazon 13 years after logging.
Forest Ecology and Management, v. 71, p. 267-274, 1995.
WIGGERS, I. STANGE, C.E.B. Manual de instruções para coleta, identificação e
herborização de material botânico. Programa de Desenvolvimento Educacional –
SEED – PR UNICENTRO. 2008.
WOODSON JR, R.; E. Apocynaceae. In: Smith AC (ed.). Lloydia 2: 207-209, 1939.

342
Belém (PA), 11 a 13 de dezembro de 2019
LEVANTAMENTO ETNOBOTÂNICO DE PLANTAS MEDICINAIS
UTILIZADAS NA PASTORAL DA SAÚDE NO MUNICÍPIO DE MOJU-PA
Mayanna Igreja dos Santos1; Raianne Ferreira Farias2; Luiz Alberto Cavalcante
Guimarães3, Sebastião da Cunha Lopes4
1 Pós-graduanda do curso de Especialização em Teorias e Metodologias de Ensino para
a Educação Básica. [email protected] 2 Licenciada em Ciências Naturais habilitação Biologia, UEPA.
[email protected] 3 Mestre em Botânica Tropical. UFRA. [email protected] 4Doutor em Ciências Agrárias, UEPA. [email protected]
RESUMO
A utilização das plantas como medicamento é tão antiga quanto o próprio homem.
Partindo desse contexto, o presente trabalho teve como objetivo realizar o levantamento
etnobotânico de plantas medicinais utilizadas no tratamento de enfermidades na Pastoral
da Saúde do município de Moju/PA. A pesquisa do tipo exploratória, teve como técnica
o estudo de caso e foi realizada na Pastoral da Saúde, localizada em Moju. Foram
entrevistadas as agentes voluntárias, cujas perguntas foram sobre: plantas medicinais
(planta utilizada, preparo e origem) e teste bioenergético (diagnóstico). Essas entrevistas
foram realizadas através de gravações. As plantas foram coletadas nos quintais da pastoral
e vizinhanças e identificadas no herbário MFS Profa. Dra. Marlene Freitas da Silva,
Universidade do Estado do Pará. Foram listadas 63 espécies de plantas medicinais
utilizadas, destas, 37 foram coletadas e identificadas no herbário MFS, e 26 espécies não puderam ser coletadas, por isso não foram identificadas cientificamente. As espécies
identificadas foram distribuídas em 26 famílias sendo as mais representativas: Lamiaceae,
Asteraceae, Amarantaceae, Myrtaceae, Malvaceae, Piperaceae, Rutaceae e Verbenacea.
Considera-se a inflamação como doença de maior ocorrência, verificou se que são
utilizadas para confecção de remédios folhas (45%), casca (20%), raízes (11%) e flor
(10%) para o preparo são utilizados chás (42%), Xaropes (20%), tinturas (18%, pomadas
(5%) sucos e banhos (4%), óleos (3%), lavagens e xampu (1%). Os resultados obtidos
nesta pesquisa enfatizam a importância significativa do trabalho desenvolvido pela
Pastoral da Saúde com plantas medicinais no município de Moju.
Palavras-chave: Botânica. Medicamentos Caseiros. Pastoral da Saúde.
Área de Interesse do Simpósio: Botânica

343
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A utilização das plantas como medicamento seja tão antiga quanto o próprio
homem. Nos antigos livros de medicina encontram-se uma imensa variedade de
medicamentos simples e eficazes que o reino vegetal oferecia aos nossos antepassados.
O uso de plantas medicinais esteve muito tempo associado a práticas mágicas, místicas e
ritualísticas, daí a dificuldade de delimitar com exatidão a evolução da arte de curar
(OLIVEIRA et al. 2009).
Segundo Ustulin et al. (2009), a utilização de plantas medicinais para tratamento,
cura e prevenção de doenças é uma das mais antigas formas de prática medicinal da
humanidade. Estudos de Jorge (2009) mostram que desde o ano 3000 a.C., a China
dedicava-se ao cultivo de plantas medicinais quando o Imperador Sheng-Nung utilizou
uma série de plantas em seu próprio corpo e formulou o "Cânone das Ervas" com cerca
de 252 espécies mencionadas.
Conforme evidencia Ford (1978) a forma como o homem vem utilizando, ao logo do
tempo, os vegetais para obtenção de cura das doenças, é que desperta o interesse pela
etnobotânica que por sua vez, vem sendo estudada e definida como o estudo da relação
sociedade-natureza, ou homem-plantas. Segundo Amorozo e Gély (1988), a etnobotânica
ainda consiste em compreender quanto ao uso e aplicação tradicional dos vegetais pela
sociedade.
Para Farnsworth (1988), a importância dada pela comunidade acadêmica a
respeito do conhecimento do uso de plantas medicinais como um método fitoterápico
surge a partir da constatação de que a base empírica utilizada por povos tradicionais ao
longo do tempo pode, em muitos casos, ter uma comprovação científica, que possibilita
a extensão destes usos à sociedade industrializada, produzindo assim maior diversidade
terapêutica de uso farmacológico.
No Brasil, o uso das plantas como medicamento tem enorme influência para o
tratamento de doenças. E muitos são os estudiosos sobre o assunto no país, entre tantos
se pode citar Scoles (2006), Jorge (2009), Pasa (2011) e Vásquez et al. (2014).
A Amazônia, tendo uma flora tão diversificada e abundante possui um expressivo
número de trabalhos publicados sobre plantas medicinais e suas utilidades, Di Stasi e
Hiruma-lima (2002), Medina et al. (2010), Castro, Souza e Fraxe (2010), Souza e

344
Belém (PA), 11 a 13 de dezembro de 2019
Scudeller (2010), são alguns dos estudiosos que dedicam suas pesquisas à região
Amazônica.
A região do Baixo Tocantins apresenta alguns trabalhos na área da etnobotânica
podem ser citados, entre eles destacam-se as pesquisas de Pinto (2008), e Lobato et al.
(2014). Em relação ao município de Moju, o número de publicações sobre plantas
medicinais vem sendo relativamente menor, onde se destacam os trabalhos de, Machado
e Braga (2011) e Cunha e Neto (2011).
Partindo desse contexto, o objetivo desse trabalho foi realizar um levantamento
etnobotânico de plantas medicinais utilizadas no tratamento de enfermidades na Pastoral
da Saúde do município de Moju/PA, tendo em vista a grande procura, por parte da
população mojuense, em relação aos remédios caseiros produzidos por esta instituição.
2. MATERIAL E MÉTODOS
2.1. ÁREA DE ESTUDO
A pesquisa foi realizada no município de Moju distante a 61 km da capital, Belém.
Para a coleta de dados foi utilizada a Pastoral da Saúde localizada no centro da cidade.
A Pastoral da Saúde é uma das pastorais sociais da Conferência Nacional dos
Bispos do Brasil (CNBB) e com organização cívico-religiosa, sem fins lucrativos, de
atuação em âmbito nacional e de referência internacional, comprometida em defender,
preservar, cuidar, promover e celebrar a vida (ou seja, saúde plena) de todo o povo de
Deus, independente de quaisquer fatores de exclusão social, inclusive do credo. Com
vários agentes espalhados por todo território nacional, esta pastoral atua em três
dimensões: solidária, comunitária e político-institucional (PASTORAL DA SAÚDE
NACIONAL, 2015).
A estrutura da Pastoral se baseia na formação de agentes de pastoral, os quais se
dispõem a prestar serviços voluntários à entidade. Essas pessoas têm a missão de cuidar
dos enfermos nos hospitais ou em suas residências, também se interessam por todos
aqueles que o assistem: profissionais de saúde, familiares, agentes pastorais, comunidade
e voluntariado, em favor dos pobres.
Conforme Baldessini (2010), uma das características da Pastoral da Saúde é a
utilização de fitoterápicos como alternativa no tratamento às doenças, além do trabalho

345
Belém (PA), 11 a 13 de dezembro de 2019
de prevenção. O uso das plantas medicinais retrata a importância que a instituição dá à
experiência da população, já que um dos objetivos da Pastoral é recuperar e revalorizar a
sabedoria popular, sua fé e sua religiosidade.
A Pastoral da Saúde trabalha com plantas medicinais em dioceses espalhadas por
todo o país, principalmente na região sul do Brasil foi encontrado relatos de que os
agentes de pastoral são treinados para o serviço voluntário participando de palestras e
formações referentes à utilização de plantas medicinais para a produção de remédio
caseiro (COMUNICAÇÃO SOCIAL, 2011).
A Pastoral da Saúde foi criada no município de Moju no ano de 1997 com a vinda
da Congregação das Irmãs Franciscanas de Ingolstad para a região. O trabalho da pastoral
começou a ser realizado no município, com iniciativa da Irmã (Ir) Matilde Picagevicz que
reuniu outras pessoas da comunidade detentoras de conhecimento empírico acerca de
plantas medicinais. Foi fundada no mesmo prédio da Pastoral da Criança devido não
possuir condições financeiras de se instalar em um local com prédio próprio e funciona
até hoje neste mesmo local.
2.2. MÉTODO DE PESQUISA
Foi utilizada a pesquisa exploratória e tendo como técnica o estudo de caso,
fornecendo dados de origem quantitativa (CERVO e BERVIAN, 2002).
2.3. A ENTREVISTA
Foram realizadas duas entrevistas com duas agentes da pastoral, a primeira com
perguntas para informar sobre a espécie utilizada, parte usada, preparo e origem, e a
segunda entrevista foi sobre a realização do teste bioenergético, método utilizado para o
diagnóstico de doenças.
2.4. COLETA E IDENTIFICAÇÃO DAS PLANTAS
As plantas foram coletadas em três locais diferentes, sendo eles: quintal da
pastoral, quintal da casa das irmãs e quintal de residências vizinhas à pastoral. O material
botânico passou pelo processo de herborização e suas exsicatas foram montadas e
incorporadas ao Herbário Prof.ª Drª. Marlene Freitas da Silva (MFS) da UEPA.

346
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
3.1. ESPÉCIES LEVANTADAS
Foram listadas 63 espécies de plantas medicinais, no entanto foram
identificadas e incorporadas no herbário (MFS) apenas 37 espécies, distribuídas em 26
famílias, 26 espécies não puderam ser coletadas, por isso não possuem nome cientifico
confirmado.
As 37 espécies coletadas e identificadas foram distribuídas em 26 famílias, as mais
frequentes foram: Lamiaceae contendo 4 espécies e Asteraceae com 3 espécies. As
famílias Amaranthaceae, Malvaceae, Myrtaceae, Piperaceae, Rutaceae e Verbenaceae
apresentaram 2 espécies cada, totalizando 12 representantes. As demais famílias foram
citadas uma vez cada e juntas totalizam 18 representantes.
Estes dados coincidem com os das pesquisas de Santos et al. (2018), em que as
famílias mais representativas também foram Lamiaceae e Asteraceae. Segundo eles, as
espécies pertencentes a estas famílias podem ser encontradas em diferentes regiões do
Brasil, e ainda dominam o grupo de plantas medicinais, por isso geralmente são as que
apresentam maior número de espécies.
Corroborando os dados obtidos nessa pesquisa, Vasquéz et al. (2014) também
observaram as famílias Asteraceae e Lamiaceae como as mais representativas em seu
trabalho realizado no município de Manacapuru-AM. Entretanto, Amorozo (2002) aponta
em sua pesquisa realizada no município de Santo Antônio de Leverger-MT que a família
mais representativa foi a Euphorbiaceae, tendo em vista que não houve registro de
nenhuma espécie desta família neste trabalho.
3.2. DOENÇAS TRATADAS COM PLANTAS MEDICINAIS
Considerando todas as plantas, foram citadas 40 indicações de uso, dentre as quais
se destacam como mais representativas no uso contra inflamações com 25 citações, gripe
com 12, diarreia com 10, verminose com 9, reumatismo 8, cólica intestinal com 8, tosse
com 7, cicatrizante 6, colesterol 5, alergia 5 e anemia, infecções, calmante, diabetes,
pressão alta com 4 citações cada, as demais indicações apresentaram uma ou duas
citações.
De acordo com os dados obtidos, a inflamação é a doença que pode ser tratada por
um maior número de plantas, já que apareceu com maior frequência no levantamento,

347
Belém (PA), 11 a 13 de dezembro de 2019
seguida pela gripe, que além de ter diversas espécies de plantas medicinais para seu
tratamento, é uma doença que ocorre com muita frequência na região, a diarreia também
uma doença muito recorrente e pode ser tratada com a utilização de diversas plantas.
3.3. PARTES DAS PLANTAS MAIS UTILIZADAS
As partes das plantas mais utilizadas no tratamento das enfermidades são as folhas
com 35 citações (45%), seguida da casca com 15 (20%), raiz 8 (11%), flor com 7 (10%),
planta toda 2 (3%), fruto 2 (3%), semente, bulbo, óleo, talo, fava, broto, cipó e fibras
todos com 1 citação cada (1%).
Pesquisas realizadas por Rodrigues e Andrade (2014), apresentam semelhanças
com estes resultados, onde também mostram as folhas como as partes das plantas mais
utilizadas, seguidas de cascas de troncos e raízes.
Na instituição da pesquisa conclui-se que o uso das folhas é bem mais frequente,
se for comparado com as demais partes da planta, isto se deve provavelmente pelo fato
desta região apresentar plantas que mantém suas folhas sempre em boas condições de uso
durante quase todo o ano, permitindo assim, uma disponibilidade de medicamentos
sempre que necessário.
3.4. FORMA DE PREPARO DE REMÉDIOS
Os trabalhos de Pinto et al. (2006), Pasa (2011), Oliveira e Menini Neto (2012),
Zucchi et al. (2013), Vásquez et al. (2014), mostram como resultados o chá como forma
de preparo mais utilizada para produção de remédios caseiros. Nesses trabalhos, decocção
e infusão são os modos de chás mais representativos, o que coincidem com os dados desta
pesquisa.
De acordo com o levantamento realizado na Pastoral observou-se que a em forma
de chá é a mais frequente com 42 citações. Em seguida apresentam-se: xaropes com 20,
tinturas com 18, pomadas com 05, sucos, banhos, óleos com 04 citações cada e por fim
lavagens e xampu com 01 citação cada.
Xarope é também um preparo bastante usado na pastoral da saúde, sendo
igualmente mencionado na pesquisa de Amorozo (2002). Além disso, deve-se considerar
o uso acentuado de tinturas para produção dos remédios caseiros na pastoral, como
também mencionado na pesquisa, de Tomazzoni et al. (2006) sobre a utilização de plantas
medicinais pela comunidade do município de Cascavel – Paraná (PR).

348
Belém (PA), 11 a 13 de dezembro de 2019
Quanto às formas de preparo e utilização de remédios caseiros, percebe-se que
podem ser usados de diferentes maneiras, seja para uso interno ou para uso externo.
4. CONCLUSÃO
Os resultados obtidos neste trabalho ressaltam a importância que a Pastoral da
Saúde possui, pois é considerada uma instituição de referência no município, em se
tratando da aquisição de remédios caseiros para a cura de determinadas enfermidades.
Uma característica que a diferencia das demais instituições que também trabalham com
plantas medicinais é a forma exclusiva de diagnóstico de doenças, através do teste
bioenergético.
Ressaltamos que a busca por plantas medicinais para a cura de doenças através da
medicina tradicional não está restrita a comunidades tradicionais e comunidades
interioranas, pessoas da cidade buscam este método de tratamento, enfatizando assim a
importância significativa do trabalho desenvolvido pela Pastoral da Saúde com plantas
medicinais no município de Moju.

349
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
AMOROZO, M. C. de M. Uso e diversidade de plantas medicinais em Santo Antônio do
Leverger, MT, Brasil. Acta botânica brasílica. v.16, n.2, p.189-203, 2002. Disponível
em: < http://www.scielo.br/pdf/abb/v16n2/a06v16n2.pdf> Acesso em: 15 jul. 2015.
AMOROZO e GÉLY, A. Uso de plantas medicinais por caboclos do Baixo Amazonas.
Barcarena, PA, Brasil. Boletim do Museu Paraense Emilio Goeldi, Belém, v.4, n.1, p.
47-131, 1988.CASTRO, A. P. da; SOUZA, H. H. de; FRAXE, T. de J. P. Estudos
etnobotânicos em uma comunidade de várzea na Amazônia Ocidental. Anais do
Congresso Nacional de Botânica. Manaus, v. 1, n.3, p.1, Set., 2010.
BALDESSIN, A. Como fazer pastoral da saúde? 1. ed. São Paulo: Loyola, 2010.
CERVO, A. L.; BERVIAN, P. A. Metodologia Científica. 5. ed. São Paulo: Prentice
Hall, 2002.
COMUNICAÇÃO SOCIAL. Encontro do GEPPLAM com a Pastoral da Saúde.
UNESC. 2015. Disponível em: <http://www.unesc.net/portal/blog/index/314> Acesso
em: 20 out. 2015.
CUNHA, D. B.; NETO, P. G. de M. Etnobotânica de Plantas Medicinais na
Comunidade Quilombola São Bernadino, munícipio de Moju – PA. 2011. 61
f.Trabalho de Conclusão de Curso (Graduação em Licenciatura em Biologia)
Universidade do Estado do Pará, Moju, 2011.
DI STASI, L. C.; HIRUMA-LIMA, C. A. Plantas medicinais na Amazônia e na Mata
Atlântica. - 2. ed. rev. e ampl. São Paulo: UNESP, 2002.
FARNSWORTH, N.R. Screening plants for new medicines. In: Wilson, E.O. (ed)
Biodiversity .Washington DC: Nac. Acad. Press, 521p. 1988. Disponível em:
<http://www.aba-
agroecologia.org.br/revistas/index.php/cad/article/viewFile/2898/2469> Acesso em: 11
nov. 2015.
FORD, Irving Richard. Ethnobotany: historical diversity and synthesis. In: R.I. Ford; M.
Hodge & W.L. Merril (eds.). The nature and status of ethnobotany. Annals of Arnold
Arboretum.Michigan: Museum of Anthropology, University of
Michigan.Anthropological Papers 67: 33-49. PDF. 1978.Disponível em: <https://ay14-
15.moodle.wisc.edu/prod/pluginfile.php/59438/mod_resource/content/0/Ford1978Ethno
botanyHistoricalDiversity.pdf>. Acesso em: 05 mai. 2015.
JORGE, S. da S. A. Plantas Medicinais: Coletânea de Saberes. 2009. PDF. Disponível
Em : <http://www.fazendadocerrado.com.br/fotos_noticias/1280/Livro.pdf.> Acesso em:
28 mar. 2015.
LOBATO, G. de J. M. et al. Biodiversidade Urbana: um estudo nos quintais do bairro
Mutirão, Abaetetuba, PA, Brasil. In: XIV Safety, Health and Environment Word
Congress. Cubatão, Brasil, V.1, n.1, p. 5-20, 2014.

350
Belém (PA), 11 a 13 de dezembro de 2019
MACHADO, A. R.; BRAGA, M. de F. da S. Estudo Etnobotânico das Plantas
Medicinais da comunidade Congregação do Rio Ubá, município de Moju, Pará.
Belém, 2011, 68 f. Trabalho de Conclusão de Curso (Graduação em Licenciatura em
Biologia) Universidade do Estado do Pará, Belém, 2011.
MEDINA, G., et al. Frutíferas e plantas medicinais úteis na vida amazônica. 2. ed.
Brasil: Cifor, 2010.
OLIVEIRA, E.R. e MENINI NETO, L. Levantamento etnobotânico de plantas
medicinais utilizadas pelos moradores do povoado de Manejo, Lima Duarte – MG.
Revista Brasileira de Plantas Medicinais, Botucatu, v.14, n.2, p.311-320, 2012.
OLIVEIRA, F. C. de, et al. Avanços nas pesquisas etnobotânicas no Brasil. Acta
botânica brasílica. v.23, n.2, p. 590-605, 2009.
PASA, M. C. Saber local e medicina popular: a etnobotânica em Cuiabá, Mato Grosso,
Brasil. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, Belém, v. 6, n.
1, p. 179-196, 2011.
PASTORAL DA SAÚDE NACIONAL. Pastoral da saúde. Disponível em:
<http://pastoraldasaudenacional.com.br/pastoral-da-sa%C3%BAde.php> Acesso em: 06
de ago. 2015.
PINTO, E. P. P; AMOROZO, M. C. M. & FURLAN, A. Conhecimento popular sobre
plantas medicinais em comunidades rurais de mata atlântica - Itacaré, BA, Brasil. Acta
Botanica Brasilica. v. 20, n. 4, p. 751-762. 2006.
PINTO, L. do N. Plantas medicinais utilizadas em comunidades do município de
Igarapé-Miri, Pará: Etnofarmácia do município de Igarapé-Miri-PA. 2008. 103 f.
Dissertação (Mestrado em Ciências Farmacêuticas) – Universidade do Federal do Pará,
Belém, 2008.
RODRIGUES, A.P.; ANDRADE, L.H.C. Levantamento etnobotânico das plantas
medicinais utilizadas pela comunidade de Inhamã, Pernambuco, Nordeste do Brasil.
Revista Brasileira de Plantas Medicinais, Campinas, v.16, n.3, p.721-730, 2014.
SANTOS, et al. Etnobotânica de plantas medicinais em mercados públicos da Região
Metropolitana de Belém do Pará, Brasil. Biota Amazônia ISSN 2179-5746 Macapá, v.
8, n. 1, p. 1-9, 2018. Disponível em http://periodicos.unifap.br/index.php/biota. Acesso
em: 22/11/2019.
SCOLES, R. Sabiduria popular y plantas medicinales: el ejemplo de la comunidad negra
de Itacoâ, acará, Pará. Boletim do Museu Paraense Emílio Goeldi, Ciência Naturais.
Belém, v. 1. p. 79-102, 2006.
SOUZA, C. C.; SCUDELLER, V. V. V. Etnobotânica de quintais em três comunidades
ribeirinhas na Amazônia Central – Manaus (AM). Anais do Congresso Nacional de
Botânica. Manaus, v.1, n.3, p.1, Set.,2010.

351
Belém (PA), 11 a 13 de dezembro de 2019
TOMAZZONI, M. I. et al. Fitoterapia popular: a busca instrumental enquanto prática
terapêutica. Texto Contexto Enferm. Florianópolis, v.15, n.1, p. 115-21, 2006.
USTULIN, M. et al. Plantas medicinais comercializadas no Mercado Municipal de
Campo Grande - MS. Revista Brasileira de Farmacognosia, João Pessoa, v. 19, n. 3,
p. 805-813, 2009.
VÁSQUEZ, S. P. F. et al. Etnobotânica de plantas medicinais em comunidades
ribeirinhas do Município de Manacapuru, Amazonas, Brasil. Acta Amazônica. v. 44, n.
4 p. 457 – 472, 2014.
ZUCCHI, M.R. Levantamento etnobotânico de plantas medicinais na cidade de Ipameri
– GO. Revista Brasileira de Plantas Medicinais, Campinas, v.15, n.2, p.273-279, 2013.

352
Belém (PA), 11 a 13 de dezembro de 2019
LEVANTAMENTO E IDENTIFICAÇÃO DE ESPÉCIES COMESTÍVEIS E
ORNAMENTAIS COM POTENCIAL TÓXICO NO MUNICÍPIO DE
SALVATERRA-PARÁ
Keyla Thaise Carvalho de Sousa1; Raimme Paola do Nascimento Pinto1; Munírih Rosa
Amaro1; Igor dos Santos Soares2; Marlyson Jeremias Rodrigues Costa3.
1Graduandas em Ciências Biológicas. Universidade do Estado do Pará - UEPA. Emails:
[email protected]; [email protected];
munirí[email protected];
2 Mestrando em Ciências Ambientais. Universidade do Estado do Pará - UEPA/PPGCA.
Email: [email protected]
3 Drº. em Biodiversidade e Biotecnologia. Professor da Universidade do Estado do Pará
- UEPA. Email: [email protected]
RESUMO
O desconhecimento de características toxicológicas de espécies botânicas cultivadas com
fins ornamentais e alimentares acaba dificultando a tomada de precauções quanto ao
caráter de toxicidade das mesmas. Nesse sentido, o trabalho objetivou levantar e
identificar espécies de plantas tóxicas utilizadas com fins ornamentais e comestíveis
presentes no município de Salvaterra-PA, na perspectiva de evidenciar os riscos que o
manuseio inadequado e a ingestão destes tipos de plantas podem oferecer ao bem-estar
de pessoas e animais. Com o auxílio de aplicativos de identificação e consulta a diretórios
botânicos, foram levantadas e identificadas 40 espécies. Dentre estas, nove espécies
distribuídas em seis famílias botânicas foram categorizadas como ornamentais e alimentíciais: Zephyranthes rosea Lindl. (Amaryllidaceae); Dieffenbachia seguine Jacq.
Schott. e Aglaonema rotundum N.E.Br. (Araceae); Dracaena fragrans (L.) Ker Gawl.
(Asparagaceae); Plumeria rubra L., Catharanthus roseus L. e Pentalinon luteum L.
(Apocynaceae); Ipomoea batatas (L.) Lam. (Convolvulaceae) e Manihot esculenta
Crantz. (Euphorbiaceae). Por fim, estas foram utilizadas para pesquisa de status de
toxicidade através das plataformas Scientific Eletronic Library Online (SCIELO),
Semantic Scholar, National Center for Biotechnology Information (NCBI),
ResearchGate, Flora do Brasil 2020 (Reflora), World Flora Online (WFO) e Sistema
Nacional de Informações Tóxico-Farmacológicas (Sintox). De acordo com a literatura
constatou-se que, em sua maioria, o cultivo destas plantas é baseado em conhecimento
informal, o que repercute em acidentes devido ao desconhecimento das espécies. Todas
as espécies identificadas são responsáveis por provocar acidentes em diversas localidades
no Brasil. Assim, verifica-se a necessidade de ações que disseminem informações com
vistas à prevenção e ao cuidado no uso e manuseio adequado das espécies identificadas.
Palavras-chave: Ilha de Marajó. Botânica. Espécies tóxicas.
Área de Interesse do Simpósio: Botânica

353
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A utilização dos vegetais pelos seres humanos ocorre desde os primórdios da
humanidade onde, a partir da agricultura eram utilizados para fins alimentícios, sendo
também usados pelos seus efeitos curativos na medicina popular (CORRÊA;
MEDEIROS, 1991; MAEZUMY et al., 2018).
A despeito de benefícios que estas plantas possuem, muitas espécies apresentam
substâncias tóxicas, oriundas da soma de características biológicas, físicas e químicas,
que são capazes de produzir reações biológicas adversas se manipuladas indevidamente
por organismos humanos e animais (VASCONCELOS et al., 2009).
Espécies de plantas de uso ornamental e alimentícias também podem apresentar
princípios tóxicos quando ingeridos e/ou tocados, cujos componentes são responsáveis
por irritações cutâneas, fotofobia, sérias manifestações toxicológicas gastrointestinais,
entre outros (ALVES et al., 2016). Entre estes princípios ativos estão os alcaloides,
glicosídeos cardioativos ou cardiotônicos, glicosídeos cianogênicos ou cianogenético,
taninos, saponinas, oxalato de cálcio dentre outros (COSTA SILVA, 2009).
Ao visar à prevenção de eventualidades e acidentes decorrentes da manipulação
de plantas tóxicas, são importantes medidas que visem à precaução no contato repetido e
ingestão, em sua grande maioria devido à proximidade e facilidade de acesso a essas
espécies. Essas condições aumentam a exposição a riscos de pessoas (principalmente
crianças) e animais (SCHVARTSMAN, 1992; ZONETTI et al., 2019). O objetivo deste
trabalho foi levantar e identificar espécies de plantas tóxicas utilizadas com fins
ornamentais e comestíveis presentes no município de Salvaterra-PA, na perspectiva de
evidenciar os riscos que o manuseio inadequado e a ingestão destes tipos de plantas
podem oferecer ao bem-estar de pessoas e animais.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
A pesquisa foi desenvolvida em Salvaterra, Ilha de Marajó, no estado do Pará. O
município possui uma extensão territorial de aproximadamente 1.039,072 km², com

354
Belém (PA), 11 a 13 de dezembro de 2019
população estimada em 23.424 habitantes, distribuídos nos espaços urbano e rural (IBGE,
2018).
2.2 COLETA E ANÁLISE DE DADOS
A coleta de dados ocorreu em jardins e quintais do espaço urbano de Salvaterra,
onde foram registradas e identificadas 40 espécies de plantas, das quais foram
selecionadas apenas as nove mais conhecidas pelo status de toxicidade, de acordo com a
literatura. As visitas aos locais de coleta de dados foram consentidas pelos proprietários
dos imóveis. Em campo, as espécies foram identificadas com auxílio dos aplicativos
PlantNet e PlantSnap, disponíveis na plataforma Android. Posteriormente, para a revisão
de nomenclatura e alocação nas famílias botânicas, se consultou os diretórios botânicos
Flora do Brasil 2020 (Reflora), World Flora Online (WFO), para levantamento de dados
toxicológicos utilizou-se o Sistema Nacional de Informações Tóxico-Farmacológicas
(Sintox), além de artigos retirados das plataformas Scientific Eletronic Library Online
(SCIELO), Semantic Scholar, National Center for Biotechnology Information (NCBI) e
ResearchGate.
Os dados coletados serão dispostos no decorrer da discussão e inseridos em uma
tabela, sendo abordados de maneira que compreenda o status de toxidade e em qual parte
das espécies identificadas está presente.
3. RESULTADOS E DISCUSSÃO
O levantamento resultou um quantitativo de 40 espécies, com maioria sendo
introduzidas vindas de outras partes do mundo, mas bem distribuídas nas Américas. As
nove plantas (Figura 01: A, B, C, D, E, F, G, H, I) referidas como tóxicas estão
distribuídas em seis famílias botânicas, e suas respectivas descrições estão listadas no
quadro 1.

355
Belém (PA), 11 a 13 de dezembro de 2019
Figura 1-registro fotográfico da identificação botânica das espécies tóxicas: A) Zephyranthes
rosea Lindl; B) Dieffenbachia seguine (Jacq.) Schott; C) Aglaonema rotundum N.E.Br.; D)
Dracaena fragrans (L.) Ker Gawl.; E) Plumeria rubra L.; F) Catharanthus roseus (L.) G.; G)
Pentalinon luteum L. BFHansen & Wunderlin; H) Ipomoea batatas (L.) Lam.; I) Manihot
esculenta Crantz.
Quadro 1- espécies botânicas tóxicas de jardins e quintais da cidade de Salvaterra, PA.
Plantas tóxicas
Família/Espéc
ie
Princípio (s)
Ativo (s)
Parte (s)
tóxica(s) Quadro toxicológico
Proce-
dência
Amaryllidaceae
Zephyranthes
rosea Lindl Alcaloides Todas
Vômitos, convulsões e
morte de humanos,
gado e aves.
Ja, Qu*
Araceae
Dieffenbachia
seguine (Jacq.)
Schott
Oxalato de
cálcio e
saponinas
Todas
Irritação por ingestão e
contato, dor em
queimação, inchaço de
lábios língua, palato e
faringe, cólicas
abdominais, náuseas e
lacrimejamento.
Ja, Qu

356
Belém (PA), 11 a 13 de dezembro de 2019
Aglaonema
rotundum
N.E.Br.
Oxalato de
cálcio --
Irritação das
membranas mucosas,
irritação da pele e
erupção cutânea
dolorosa.
Ja, Qu
Asparagaceae
Dracaena
fragrans (L.)
Ker Gawl. Saponinas
Bulbos
germinativos
Anorexia,
agressividade, arritmias
cardíacas, problemas na
mucosa ocular e lesões
de pele tipo
fotossensibilização.
Ja, Qu
Apocynaceae
Plumeria
rubra L.
Alcaloides
agoniadina,
plumária e
ácido
plumeritânico.
Caule e
folhas
Náuseas, alucinações,
redução dos reflexos,
diarreia e hipotensão.
Ja, Qu
Catharanthus
roseus (L.) G.
Alcaloide da
vinca
Todas Alucinações Ja, Qu
Pentalinon
luteum L.
BFHansen &
Wunderlin
-- Folhas Intoxicação animal Ja, Qu
Convolvulaceae
Ipomoea
batatas (L.)
Lam.
Alcaloides,
antraquinonas,
taninos e
saponinas.
Ramos, flores Sintomas
pneumotóxicos Qu
Euphorbiaceae
Manihot
esculenta
Crantz.
Glicosídeos
cianogênicos
Folhas e
raízes
Cansaço e falta de ar,
fraqueza, taquicardia,
taquipneia, agitação,
confusão mental,
convulsão, coma e
morte.
Qu
Fonte: Presente estudo. *Lê-se: Ja – jardim; Qu - Quintal
Geralmente, a maioria das plantas que foram identificadas como tóxicas em
Salvaterra estão cultivadas em jardins e/ou quintais, com livre acesso de pessoas. No
entanto, além destes locais domiciliares, Dantas (2007a) e Costa Silva (2009), registram

357
Belém (PA), 11 a 13 de dezembro de 2019
que estas plantas também são encontradas em espaços ornamentais de parques, pátios,
grandes jardins e shoppings, aumentando o risco de acidentes. É o caso da Plumeria rubra
L. e Catharanthus roseus (L.) G., (COSTA SILVA, 2009), que também foram
identificadas nos locais de coleta do trabalho. Dantas (2007a) reitera que são as
caracteristicas chamativas das flores, frutos ou sementes coloridas, e ainda a presença do
látex, que favorecem o contato com estas espécies.
Por outro lado, para Carvalho e Silveira (2010) a propagação das plantas só é
difundido como recursos que trazem somente beneficios, devido as propriedades que as
permitem serem utilizadas como fonte de alimentação e em muitos casos com própositos
terapêuticos para o tratamento de diversas doenças na medicina tradicional. Isso contribui
com as concepções errôneas sobre essas plantas, e, acabam por representar riscos
potenciais à saúde humana.
Durante a coleta de dados do estudo, foi possível observar a ocorrência acentuada
das espécies Dieffenbachia seguinte (Jacq.) Schott, Pentalinon luteum e Catharanthus
roseus (L.) G., nas decorações de jardins caseiros, e que as pessoas as cultivam sem
informações prévias das mesmas. A proximidade e a exposição destas espécies as
pessoas, principalmente, crianças e animais (considerados grupos de risco), que
costumam manuseá-las sem conhecimento e os cuidados necessários, faz deles as
principais vítimas de acidentes por intoxicação com plantas ornamentais (ZONETTI et
al., 2019).
A vulnerabilidade a grupos de riscos supracitados, demonstra a necessidade no
(re)conhecimento do potencial de toxicidade dessas espécies antes de utiliza-las, do
contrário tornam improprias pra fins ornamentais. Segundo Amador et al. (2000), através
de uma análise de fichas de ocorrências toxicológicas em Maringá, identificou-se 20 casos
de intoxicação tendo a planta ‘comigo-ninguém-pode’ (Dieffenbachia seguine) como a
principal causadora em grupo de criança menores de 14 anos, no ano de 1995. Dantas
(2007b), também refere intoxicação de caprinos, em Recife/PE, pela mesma planta.
Quanto às espécies Ipomoea batatas e Manihot esculenta Crantz, que são espécies
alimentícias, observa-se um quadro mais preocupante, devido ao elevado risco de
intoxicação se não preparadas de forma adequada. Canela et al. (1998) argumenta que a
riqueza em ácido cianídrico dos tubérculos da Mandioca (HCN), tem como consequência
a distribuição de altas taxas de toxicidade na raiz e folhas, que podem causar até a morte

358
Belém (PA), 11 a 13 de dezembro de 2019
se consumidas cruas ou mal cozidas. Estes registros indicam a necessidade de educação
a respeito plantas tóxicas.
Colombo et al. (2010) recomenda, portanto, que utilização de plantas como
remédio e alimento deve se limitar a plantas conhecidas e/ou corretamente identificadas,
para que não ocorram graves acidentes, devido aos princípios ativos que fazem parte da
composição química dos vegetais. Jesus; Suchara (2013), reiteram ainda, que estes
metabólitos secundários ao serem tocados, inalados ou ingeridos, são capazes de
promoverem alterações patológicas em pessoas e animais, assim como as descritas para
as espécies do estudo (Quadro 1).
Com vistas à prevenção, as contribuições Alves et al. (2016) e Zonetti et al.
(2019), demonstram ser possível obter resultados satisfatórios, a partir da socialização de
informações para públicos distintos, sobre os cuidados com a toxicidade de plantas. Estes
estudos sinalizam que ações que partem de temas gerais e mais conhecidos, como o de
plantas medicinais, podem ser estratégicos para a introdução ao assunto de toxicidade dos
vegetais.
O trabalho com a temática em diferentes esferas da sociedade, se torna mais
necessário, ao se considerar as dificuldades de diagnóstico e a identificação da espécie
vegetal que provocou a intoxicação (CAMPOS, et al., 2016). Neste contexto, estão entre
os fatores de dificuldade: o não relato, pelo acidentado, do consumo ou contato com a
planta; a barreira de poucas informações sobre o potencial tóxico das espécies e a ausência
de profissional que possa fazer a correta identificação da espécie nos pontos de
atendimento médico (PEACOK et al., 2009; MONSENY et al., 2015).
4. CONSIDERAÇÕES FINAIS
A presença de plantas com potencial tóxico nos jardins e quintais da cidade de
Salvaterra como a Manihot esculenta Crantz., que tem fins alimentícios e é muito
utilizada pelos residentes, principalmente para a produção de farinha d’água, ou a
Dieffenbachia seguine (Jacq.) Schott., que tem fins ornamentais e está presente em grande
quantidade nos jardins dos moradores, sinaliza o risco de eventuais casos de intoxicação
local. Os problemas acometidos pelas plantas identificadas se dão pela falta de
conhecimento da população em relação aos riscos que estas plantas podem trazer. Para
isso, são necessários estudos mais aprofundados na temática que possam identificar e

359
Belém (PA), 11 a 13 de dezembro de 2019
divulgar todas as espécies que oferecem perigo à população humana e animais. Para tanto,
campanhas promovidas pelo poder público, ações extensivas em escolas, hospitais, entre
outros espaços, podem favorecer o reconhecimento do risco, a prevenção de acidentes e
o combate a eventuais intoxicações, ações importantes à manutenção da saúde pública.

360
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALVES, R. B. S. et al. Plantas ornamentais x plantas tóxicas: prevenção de acidentes
com crianças. Revista Ciência em Extensão, v.12. n. 3, p.79-87, 2016.
AMADOR, J. C. et al. Perfil das intoxicações agudas exógenas infantis na cidade de
Maringá (PR) e região, sugestões de como se pode enfrentar o problema. Pediatria
(São Paulo). 2000.
BARG, D. G. Plantas tóxicas. Trabalho acadêmico: Faculdade de Ciências da Saúde de
São Paulo, 2004.
CAMPOS, S. C., et al. Toxicidade de espécies vegetais. Revista Brasileira de Plantas
Medicinais, v.18. n.1 p. 373-382, 2016.
CANELLA, C. F. C. et al. Intoxicação experimental pela maniçoba (Manihot glaziovi
Muell. Arg.) em bovinos. Pesquisa Agropecuária Brasileira, v. 3, p. 22-28, 1968.
COLOMBO, M. L. et al. Most commonly plant exposures and intoxications from
outdoor toxic plants. Journal of Pharmaceutical Sciences and Research, v.2, n.7, p.417-
25, 2010.
CORREA, F. R.; MEDEIROS, R. M. T. Intoxicações por plantas em ruminantes no
Brasil e no Uruguai: importância econômica, controle e riscos para a saúde pública.
Pesq. Vet. Bras, 2001.
COSTA, R. L. D. et al. Um caso de intoxicação de bovinos por Enterolobium
contortisiliquum (timboril) no Brasil. Archivos de Zootecnia. V. 58, n. 222, p. 313-
316, 2009.
COSTA SILVA, L. Plantas ornamentais tóxicas presentes no shopping Riverside
Walk em Teresina–PI. Revista da Sociedade Brasileira de Arborização Urbana, v.4, n.3,
p.69-85, 2019.
CUMPSTON, K. L. et al. Acute airway compromise after brief exposure to a
Dieffenbachia plant. J. Emerg Med. Novembro de 2003; 25 (4): 391-7.
DANTAS, I. C. O Raizeiro. Campina Grande: EDUEPB. 2007a.
DANTAS, A. C. et al. Intoxicação natural por comigo-ninguém-pode (Dieffenbachia
sp.) em caprino. Recife-PE, Ciênc. vet. trop. maio/dezembro, 2007b.
FERREIRA, M. M. Avaliação das características de crescimento e desenvolvimento
da Vinca (Catharanthus roseus) em relação à adubação em época de colheita. Minais
Gerais. LAVRAS, 2013.
FOCHTMAN, F. W. et al. A. Toxicity of the genus Dieffenbachia. Toxicol Appl
Pharmacol. 1969, 25 Jan, 2014.

361
Belém (PA), 11 a 13 de dezembro de 2019
FURTADO F. M. V. et al Intoxicações causadas pela ingestão de espécies vegetais em
ruminantes. Ciência Animal, 2012.
HSUEH, K. F. et al. Ocular injuries from plant sap of genera Euphorbia and
Dieffenbachia. J Chin Med Assoc. 25 Jan, 2014.
IBGE. Cidades, 2018. Disponível em:
https://cidades.ibge.gov.br/brasil/pa/salvaterra/panorama. Acesso 20 de out., 2019.
JESUS, N. A.; SUCHARA, E. A. Cultivo de plantas tóxicas e a ocorrência de
intoxicações em domicílios no município de Barra do Graças. Revista Eletrônica da
UNIVAR, v.2; n.10, p.89-95, 2013.
KNOX, G. W. (2005). “Rainlily, Zephyranthes e Habranthus spp: Bulbos de Baixa
Manutenção para Jardins da Flórida” (PDF). ENH1151. Instituto de Extensão de Ciências
Alimentares e Agrícolas, universidade da Flórida.
MAEZUMI, S. Y. et al. The legacy of 4,500 years of polyculture agroforestry in the
eastern Amazon. Nature plants, v. 4, n. 8, p. 540, 2018.
MARINHO, S. R. M. Função dos Alcalóides Indóiicos Monoterpenóides de
Catharanthus roseus Catharanthus roseus (L.) G. Don (PDF). Faculdade do Porto,
2011.
MARTINS, T. D; GERON, V. L. M. G. Plantas ornamentais tóxicas: conhecer para
previnir acidentes domésticos. Revista Científica da Faculdade de Educação e Meio
Ambiente, Jan-Jun, 2014.
MATOS, F. J. A. et al. Plantas tóxicas – Estudo de Fitotoxicologia Química de Plantas
Brasileiras. São Paulo, Instituto Plantarum de Estudos da Flora, 247 p. 2011.
MONSENY, A. M. et al. Poisonous plants: an ongoing problem. Anales de Pediatria,
v.85, n.2, p.347-353, 2015.
MORTON, J. F. (1962). “Plantas ornamentais com propriedades tóxicas e/ ou
irritantes”. Proceedings of the Florida Estado Horticultural Society. Sociedade.
Sociedade Horticultural do Estado da Flórida.75: 484-491.
PEACOK, B. M. et al. Intoxicaciones por plantas tóxicas atendidas desde un servicio
de información toxicológica. Revista Cubana de Plantas Medicinales, v.14, n.2, p.1-8,
2009.
PlantNet. Disponível em: https:/www.plantsnap.com. Acessado em 17 maio 2019.
PlantSnap. Disponível em: https:/www.plantsnap.com. Acessado em 17 maio 2019.
Plumeria rubra information from NPGS/GRIN. Germplasm Resources Information
Network- (GRIN). Departamento de Agricultura dos Estados Unidos da América.

362
Belém (PA), 11 a 13 de dezembro de 2019
RIET-CORREA, F. et al. Intoxicações por plantas e micotoxicoses em animais
domésticos. Montevideo. Editorial Hemisfério Sul, 340 p. 1993
ROCHA, L. D. et al. Organização estrutural e localização das estruturas tóxicas em
comigo-ninguém-pode (Dieffenbachia picta (L.) Schott) e copo-de- leite
(Zantedeschia aethiopica (L.) Spreng). RUBS, Universidade Positivo. 25 Jan, 2014.
ROSSETTI, A. C. P. A.; CORSI, M. Plantas tóxicas de interesse pecuário. Revisão
bibliográfica 01 – projeto capim – pesquisa e extensão; departamento de zootecnia; esalq-
usp, abril de 2009.
SCHVARTSMAN, S. Plantas venenosas e animais peçonhentos. 2. ed. São Paulo:
Sarvier. 288p., 1992.
SILVA, L.R.R. et al. Plantas Tóxicas: Conhecimento de populares para a prevenção de
acidentes. RevInter Revista Intertox de Toxicologia, Risco Ambiental e Sociedade, v. 7,
n. 2, p. 17-36, jun, 2014.
SNAJDAUF, J. et al. Aortoesophageal fistula – an unusual complication of
esophagitis caused by Dieffenbachia ingestion. Philadelphia, Journal of pediatric
surgery, p. 29 – 31, 2005.
TEIXEIRA, J. B. P; LIMA, A. A. Plantas ornamentais tóxicas: prevenção de acidentes,
2013.
TOKARNIA, C. H. et al. Plantas tóxicas do Brasil. RevInter (Revista Intertox de
Toxicologia, Risco Ambiental e Sociedade), 2014.
VASCONCELOS, J. et al. Plantas tóxicas: conhecer para prevenir. Revista Científica da
UFPA, v.7, n.1, p.1-10, 2009.
ZONETTI, P. C. et al. Crianças e plantas medicinais: o conhecimento por meio de
atividades lúdicas. Expressa Extensão, v.24, n.1, p. 63-76, 2019.

363
Belém (PA), 11 a 13 de dezembro de 2019
PERFIL QUÍMICO E ATIVIDADE ANTIOXIDANTE DA PLANTA VERÔNICA
(Dalbergia monetaria L.) COLETADA NO ARQUIPÉLAGO DO MARAJÓ - PA
Diogo Gabriel Barbosa Pimentel1; Abraão de Jesus Barbosa Muribeca2; Rayana Silva
Craveiro3; Endril Pablo Brasil de Freitas4; Ronilson Freitas de Souza5
1Cursando o Ensino Médio. Escola Estadual de Ensino Médio Salomão Matos /
Programa Institucional de Bolsas de Iniciação Científica – Junior.
[email protected] 2Doutorando em Química. Universidade Federal do Pará.
[email protected] 3Graduanda em Licenciatura Plena em Ciências Naturais com Habilitação em Química.
Universidade do Estado do Pará. [email protected] 4Graduando em Licenciatura Plena em Ciências Naturais com Habilitação em Química.
Universidade do Estado do Pará. [email protected] 5Professor no Departamento de Ciências Naturais e Tecnologias – CCSE, Campus XIX.
Universidade do Estado do Pará. [email protected]
RESUMO
A Amazônia é um local onde existe uma cultura fitoterápica, decorrente de vários fatores:
pelo fácil acesso as plantas medicinais, pelo conhecimento popular, a questão financeira,
os benefícios terapêuticos e entre outros. Até mesmo nas antigas civilizações se usavam
extratos de plantas como fontes de remédios naturais, para tratamento, cura e prevenção
de diversos males de diversas etiologias causadas ao homem, sem qualquer conhecimento
da composição química destes extratos. Grande parte desta população atualmente ainda
faz uso destes vegetais baseando-se apenas no saber empírico, o que torna essencial a
realização de levantamentos científicos para verificar e validar a possibilidade de tornar-
los em fármacos com potencial curativos. Na região Amazônica existe uma grande
diversidade de espécimes de plantas medicinais, dessas muitas ainda são desconhecidas
e outras não são muito estudadas como é o caso Verônica (Dalbergia monetaria L.). Neste
contexto o objetivo deste trabalho foi determinar o perfil químico e avaliar o potencial
antioxidante de planta medicinal Dalbergia monetaria L., utilizando o CCDC para
determinar o perfil químico e por meio do método de DPPH, averiguar a atividade
antioxidante deste vegetal. Com os resultados obtidos foi possível verificar a presença
considerável de compostos antioxidantes, as quais pertencentes à classe dos flavonoides,
no entanto, há uma necessidade de isolamento e elucidação de possíveis estruturas com
princípios ativos.
Palavras-chave: Verônica. Plantas Medicinais. Potencial Antioxidante
Área de Interesse do Simpósio: Botânica

364
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
As plantas medicinais são utilizadas há muito tempo no tratamento de
enfermidades e alteração no curso de doenças, muitas vezes sendo o único recurso
terapêutico disponível, como no caso de populações que não possuem fácil acesso a
serviços de saúde ou medicamentos industrializados. Segundo a Organização Mundial da
Saúde (OMS) o consumo de medicamentos à base de ervas e plantas medicinais é hábito
comum para aproximadamente 80% da população mundial (VITAL, 2014).
As propriedades medicinais das plantas têm um papel muito importante na questão
socioeconômica, tanto para as populações que vivem no meio rural, como as que vivem
no meio urbano. Na maioria das vezes nativas da região, ou cultivadas em quintais,
reduzindo os gastos com medicamentos sintéticos. Este é um aspecto que também pode
estar ocasionando a prática de cultivos de plantas com atividades biológicas na região
amazônica (MERA et al.,2018). Como grande parte da população faz uso destes vegetais
baseando-se apenas no saber empírico, torna-se essencial realizar de levantamentos
científicos para possibilidade de tornar estas plantas como fitoterápicos.
2. METODOLOGIA
A coleta da espécie foi realizada no município de Soure, localizado no
Arquipélago de Marajó - PA. Foi realizada em dois momentos: uma coleta no período de
agosto a outubro de 2018, e em março a abril de 2019, posteriormente as amostras foram
encaminhadas para o Laboratório de Ciências Naturais na Universidade do Estado do
Pará, Campus XIX, em Salvaterra - PA.
2.1 OBTENÇÕES DOS EXTRATOS SECOS
Para obtenção dos extratos secos foram utilizados às partes aéreas e sistemas
radiculares da planta Verônica (Dalbergia monetaria L) que foi submetido à secagem em
estufa à temperatura de 40ºC por um período de três dias, e triturados em um
microprocessador Wallita®, a fim de favorecer a solubilização com o solvente no
processo de maceração. O material foi macerado com álcool etílico 92,8° INPM em
temperatura ambiente e a extração realizada em três ciclos de 72 h, totalizando um período

365
Belém (PA), 11 a 13 de dezembro de 2019
de nove dias, a cada 72 h o material vegetal passou por filtração a vácuo sendo adicionado
um novo solvente. Ao final, os extratos foram armazenados em frascos âmbar.
2.2 CCDC (CROMATOGRAFIA EM CAMADA DELGADA COMPOSTA)
Segundo a indicação de Collins (2006) a caracterização química das amostras foi
realizada por CCDC, que foi possibilitado pelo uso dos marcadores químicos específicos.
As amostras e os marcadores químicos foram aplicados, com volumes iguais (10 μL), em
placas cromatográficas de sílica gel, empregando o revelado metanol / ácido fórmico
(5:5), diclorometano / metanol / acetona / ácido (79:10:10:01), diclorometano / metanol /
ácido fórmico (97:02:01) e diclorometano / metanol (5:5), acetona / diclorometano (5:5).
2.2.1 Sistemas de reveladores
Os sistemas reveladores foram embasados nos trabalhos de Barros e Pedrosa
(2018) e Reis (2018). VAS: a utilização da solução de VAS foi realizada nas seguintes
etapas, aplicação da solução na placa de sílica gel, posteriormente com o auxílio de uma
placa térmica aquecida em temperatura constante de 100ºC, durante 10 minutos para a
visualização dos resultados. NP/PEG: a solução de NP foi aplicada primeiro, seguida pela
solução de PEG. Após aplicação dos reveladores foi possível observar a placa sob a luz
ultravioleta (366 nm). DRAGENDORFF: Revelação com Dragendorff: após a aplicação
do reagente Dragendorff, foi aplicado o revelador ácido sulfúrico 10%, para melhor
visualização dos resultados. A solução aquosa faz com que a evaporação do revelador
seja muito lenta e por vezes o tempo de evaporação faz com que as manchas desapareçam.
Para acelerar o processo de evaporação, foi utilizado um secador de cabelo na temperatura
mínima.
2.2.2 Atividade antioxidante dos extratos
As amostras foram tratadas seguindo os parâmetros de aplicação em CCD. Os
sistemas de eluição foram os mesmos empregados para o perfil químico. A solução
0,005% de DPPH, ou seja, 25 mg em 50 mL de metanol, para derivatizar a placa após a
eluição. Em seguida, a placa foi coberta por papel alumínio durante 20 minutos e os
resultados fotografados.
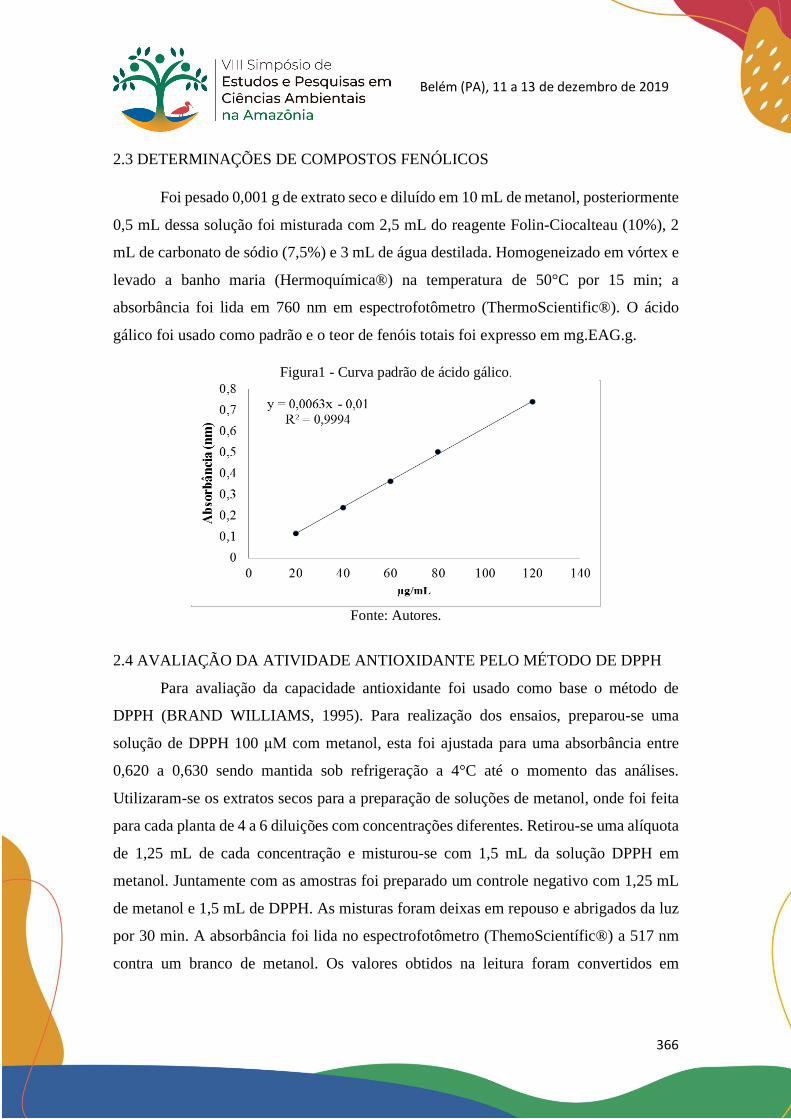
366
Belém (PA), 11 a 13 de dezembro de 2019
2.3 DETERMINAÇÕES DE COMPOSTOS FENÓLICOS
Foi pesado 0,001 g de extrato seco e diluído em 10 mL de metanol, posteriormente
0,5 mL dessa solução foi misturada com 2,5 mL do reagente Folin-Ciocalteau (10%), 2
mL de carbonato de sódio (7,5%) e 3 mL de água destilada. Homogeneizado em vórtex e
levado a banho maria (Hermoquímica®) na temperatura de 50°C por 15 min; a
absorbância foi lida em 760 nm em espectrofotômetro (ThermoScientific®). O ácido
gálico foi usado como padrão e o teor de fenóis totais foi expresso em mg.EAG.g.
Figura1 - Curva padrão de ácido gálico.
Fonte: Autores.
2.4 AVALIAÇÃO DA ATIVIDADE ANTIOXIDANTE PELO MÉTODO DE DPPH
Para avaliação da capacidade antioxidante foi usado como base o método de
DPPH (BRAND WILLIAMS, 1995). Para realização dos ensaios, preparou-se uma
solução de DPPH 100 μM com metanol, esta foi ajustada para uma absorbância entre
0,620 a 0,630 sendo mantida sob refrigeração a 4°C até o momento das análises.
Utilizaram-se os extratos secos para a preparação de soluções de metanol, onde foi feita
para cada planta de 4 a 6 diluições com concentrações diferentes. Retirou-se uma alíquota
de 1,25 mL de cada concentração e misturou-se com 1,5 mL da solução DPPH em
metanol. Juntamente com as amostras foi preparado um controle negativo com 1,25 mL
de metanol e 1,5 mL de DPPH. As misturas foram deixas em repouso e abrigados da luz
por 30 min. A absorbância foi lida no espectrofotômetro (ThemoScientífic®) a 517 nm
contra um branco de metanol. Os valores obtidos na leitura foram convertidos em

367
Belém (PA), 11 a 13 de dezembro de 2019
porcentagem da inibição por meio da equação: % de Inibição do DPPH = (Abs. controle
negativo – Abs. amostra / Abs. controle negativo) x 100.
Por meio do programa Microsoft Office Excel 2016, construiu-se um gráfico de
inibição (eixo x) versus as respectivas concentrações dos extratos (eixo y). A curva gerada
foi interpretada segundo uma linha de tendência reta, por meio desta, obteve-se equações
que permitiram estimar a concentração de cada extrato eficiente para reduzir a quantidade
inicial de DPPH pela metade (EC50).
3. RESULTADOS E DISCUSSÃO
3.1 CCDC
Foram utilizadas placas de sílica em gel 20x20 cm, em seguida foram cortadas no
tamanho 10x10 cm e lavadas com metanol. A amostra de Verônica foi disposta em
frascos, acrescida de metanol, colocada no sonicador para solubilizar, em seguida diluída
em metanol até ficarem na concentração de 5.000 miligramas. Foi encaminhada para
camag automatic tlc sampler 4. A amostra depositada nas cubetas para a realização das
corridas; que foram reveladas e fotografadas, em seguida foi estabelecido um padrão do
sistema de diclorometano / metanol / ácido fórmico (9,7:0,2:0,1), sistema um pouco leve
a fim de arrastar substâncias menos polares e diclorometano / metanol / acetona / ácido
(7,9:11: 0,1) sistema mais forte, para arrastar substâncias polares que não foram arrastadas
pelo anterior. Durante as análises os resultados foram fotografados na máquina Camag
tlc visualizer usando os aspectos luz visível; luz 254 nm e a luz 366 nm para visualização
de um número maior de substâncias.
3.1.1 Sistemas reveladores
Os sistemas reveladores utilizados foram o reagente vanilina – ácido sulfúrico
(VAS). Após a aplicação da solução VAS, a placa com as amostras foi aquecida com o
auxílio de uma placa térmica em temperatura constante de 100ºC, durante 10 minutos
para a visualização dos resultados e depois foi fotografada na luz visível.

368
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2 – Sistema revelador VAS.
Fonte: Autores.
Legenda: Coluna 1 - Amostra padrão (Rutina). Coluna 2 - Amostra de Verônica.
Na coluna 1 (Rutina) foi encontrada Flavonoides. Na coluna 2 (Verônica) foi
encontrado as substâncias: flavonoides; Irioides glicosilados e secoirioides.
Reagente NP/PEG: a solução de NP foi aplicada primeira, em seguida foi aplicada
a solução do PEG. Após aplicação dos reveladores observou-se a placa sob a luz
ultravioleta (366 nm).
Figura 3 – Sistema revelador NP/PEG.
Fonte: Autores.
Legenda: Coluna 1 - Amostra padrão (Rutina). Coluna 2 - Amostra de Verônica.
Na coluna 1 (Rutina) foram encontrados antronas e antronoise, enquanto que na
coluna 2 (Verônica) foram encontrados ácidos carboxílicos, fenólicos, cumarinas e
flavonoides.
DRAGENDORFF: Após a aplicação do reagente Dragendorff, foi adicionada a
solução do revelado ácido sulfúrico 10% que melhorou visualização dos resultados. Para

369
Belém (PA), 11 a 13 de dezembro de 2019
acelerar o processo de evaporação, a placa foi aquecida com auxílio de um secador de
cabelo.
Figura 4 – Sistema revelador Dragendorff.
Fonte: Autores.
Legenda: Coluna 1 - Amostra padrão (Brucina). Coluna 2 - Amostra de Verônica.
Nas colunas 1 (Brucina) e 2 (Verônica) foram detectadas a presença de
alcaloides.
Figura 5 – Sistema revelador DPPH.
Fonte: Autores.
Legenda: Coluna 1 - Amostra padrão (ácido ascórbico). Coluna 2 - Amostra de Verônica.
Na coluna 1, possui a presença de antioxidantes e na coluna 2 também foi possível
identificar metabólitos com potencial antioxidantes.
3.2 COMPOSTOS FENÓLICOS

370
Belém (PA), 11 a 13 de dezembro de 2019
O resultado obtido para a quantificação do teor total de compostos fenólicos da
amostra Verônica, coletada no município de Soure, na Ilha do Marajó - Pará está
apresentado na Tabela 1.
Tabela 1 - Resultados da quantificação de fenóis totais
das amostras de Verônica.
Nome da planta (nome popular) Fenóis Totais1
Dalbergia monetaria L. (Verônica) 725,40 ± 1,22
1teor total de fenóis expressos em miligramas equivalentes de ácido gálico por grama.
Fonte: Autores.
3.3 CAPACIDADE ANTIOXIDANTE
Tabela 2 – Inibição do DPPH para cada concentração de extrato da Verônica.
Nome da planta (Nome popular)
Concentração
(µg.mL-1) Inibição do
DPPH(%)
EC50
(µg.mL-1)
Dalbergia monetaria L.
(Verônica)
15 12 9 6 3
1,5
91,90 73,45 71,92 54,38 31,57 23,61
6,17
Fonte: Autores.
4. CONSIDERAÇÕES FINAIS
A região amazônica tem uma biodiversidade riquíssima que pode guardar avanços
no ramo farmacológico, por isso, é valido investigar de forma científica os conhecimentos
empíricos disseminados através da tradição oral, para averiguar os possíveis benefícios
da utilização desses conhecimentos na região. A partir das análises realizadas, ressalta-se
que a verônica tem propriedades fitoterápicas e uma boa atividade antioxidante,
comprovando dessa forma que ela pode ser considerada uma planta medicinal.

371
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
Barros, D. P. D. Avaliação do perfil químico e físico-químico de plantas medicinais
de Salvaterra – Ilha de Marajó, Pará. 2018. 58 f. Trabalho de Conclusão de Curso
(Graduação em Ciências Naturais com Habilitação em Química) - Universidade do
Estado do Pará. Salvaterra, 2018.
BRANDO-WILLIAMS, W; CUVELIER, M. E.; BERST, C. Use f free radical method
to evaluate antioxidant activity. LebensmittelWissenschatUndTechnologie, v. 28, p.
25-30, 1995.
COLLINS, C.H.; BRAGA, G.L.; BONATO,P.S. Fundamentos de Cromatografia.
São Paulo: Unicamp, 2006.
REIS, J. D. E .Triagem fitoquímica e avaliação do potencial antioxidante das folhas
da espécie Chamaecristadiphylla (L.) Greene (Fabaceae). 2018. 83f. Trabalho de
Conclusão de Curso (Licenciatura em Ciências Naturais com Habilitação em Química) -
Universidade do Estado do Pará, Salvaterra, 2018.
MERA. R. L et al. Conhecimento, percepção e ensino sobre plantas medicinais em duas
escolas públicas no município de Benjamin Constant – Am. Experiências em Ensino
de Ciências, v.13, n. 62, 2018.
VITAL, S. Plantas medicinais com ação antifúngica. 2014. 30f. Trabalho de
conclusão de Curso (Farmácia) - Universidade Católica de Brasília, Distrito Federal,
2014.

372
Belém (PA), 11 a 13 de dezembro de 2019
PROBLEMÁTICA NA ASSOCIAÇÃO DE NOMES VERNACULARES E
CIENTÍFICOS: O CASO DO CAJU
Stefany de Andrade Silva1; Gracialda Costa Ferreira²; Ingrid Raphaela Cromwell
Pereira³; Daniel Pereira da Silva Filho4; Alessandra de Cassia Nascimento Vinagre5.
1Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia. E-mail:
²Doutora em Botânica Tropical, Universidade Federal Rural da Amazônia. E-mail:
3Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia. E-mail:
4Graduando em Engenharia Florestal. Universidade Federal Rural da Amazônia. E-mail:
5Graduanda em Engenharia Florestal. Universidade federal Rural da Amazônia. E-mail:
RESUMO
O processo de manejo florestal exige uma correta identificação das espécies para seu uso
de acordo com suas propriedades físicas, químicas e mecânicas. No entanto, existe uma
variabilidade de nomes vernaculares e científicos na família Anacardiaceae, o que pode
acarretar um diagnóstico equivocado e comprometer a comercialização e biodiversidade.
O objetivo do trabalho é relacionar nomes vernaculares com científicos para verificar
família, ocorrência no estado do Pará, ameaça de extinção, valor e volume de madeira.
Para isso, foram obtidos dados por meio da base do SISFLORA do relatório de Extração
Comércio de Toras de Madeira por Essência, pela SEMAS (Secretaria de Meio Ambiente
e Sustentabilidade) e pela plataforma do Flora do Brasil. Com isso, verificou-se a
existência de 8 nomes vernaculares na base de dados do SISFLORA, que contabilizam o
total de 147.696,28 m³ e R$16.941.638,63. Para esses 8 nomes vernaculares foram
listados 11 nomes científicos, tendo entre eles apenas 5 nomes aceitos de acordo com o
Flora do Brasil. As características macromorfológicas são de extrema importância para a
diferenciação das espécies em campo por possuírem estrutura especificas. Desse modo, a
correção e divulgação da nomenclatura corrigida de espécies dentro do gênero são
imprescindíveis para auxiliar em estudos científicos confiáveis e para o bom manejo das
espécies.
Palavras-chave: Manejo Florestal. Identificação. Características Macromorfológicas.
Área de Interesse do Simpósio: Botânica

373
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A identificação botânica é de grande importância no setor madeireiro, pois com a
identificação correta é possível acessar informações sobre o uso da madeira para cada
espécie, de acordo com suas características físicas, químicas e mecânicas (LIMA, 2015).
No entanto, o agrupamento de várias espécies no mesmo vernacular inviabiliza o
diagnóstico correto para o uso da madeira, portanto, é necessário avaliar a relação na
identificação das madeiras inventariadas do “caju” e variantes. Para, além disso, a
importância ecológica e para o uso sustentável de recursos florestais, que não venha
prejudicar o setor econômico, comercial e a biodiversidade por um diagnóstico incorreto.
(ARAUJO, 2016).
É prática na realização de inventários florestais se atribui a determinação as
espécies a profissionais que detém o conhecimento empírico sobre as plantas, por serem
nativos desta, os chamados “mateiros” que utilizam nome vernaculares para nomear as
espécies. Posteriormente, esses nomes vernaculares serão associados a nomes científicos,
obtidos em literatura, consulta em sites ou disponível em listas de outros inventários
comerciais. É uma pratica que não considera qualquer informação mais consistente sobre
a verdadeira identidade das árvores nomeadas. Para Kanashiro (2002), este procedimento
leva a uma confusão entre os nomes vernaculares e os científicos acarretando exploração
desordenada e não sustentável das florestas, trazendo prejuízos para a empresa e para a
biodiversidade.
O Ministério do Meio Ambiente (MMA, 2002), por meio da Instrução Normativa
Nº 04, estabeleceu critérios para a exploração sustentável da floresta e, destaca sobre que
a identificação e levantamento de todas as árvores a serem exploradas nos inventários,
devem apresentar dados consistentes e autênticos. Em vista disso, a legislação determina
esses os parâmetros para que os atores manejadores possam trabalhar, causando menor
impacto.
Anacardium giganteum, no Pará vulgarmente chamado de “Caju”, pertence à
família botânica, ocorrente em algumas partes do Brasil, presente também na Índia,
Vietnã, Moçambique e Nigéria. A amêndoa do caju é o produto comercial de maior
importância, seu mercado é centrado na exportação, que é o destino de cerca de 90% da
produção nacional, além de apresentar importância madeireira por possuir alto brilho e
atrativamente marcada com desenhos formados pelos veios. A madeira é leve, possuindo

374
Belém (PA), 11 a 13 de dezembro de 2019
propriedades mecânicas e retratilidade baixa. A espécie em questão foi escolhida devida
a importância econômica e por ter poucas literaturas vigentes a respeito das espécies. O
objetivo do trabalho é relacionar nomes vernacular com científicos para verificar família,
ocorrência no estado do Pará, ameaça de extinção, valor e volume por meio do
SISFLORA (Sistema de Comercialização e transportes de Produtos Florestais).
2. METODOLOGIA
Os dados utilizados relacionados a nomes científicos, nomes vernacular, gênero,
família, volume e valor da madeira do caju foram adquiridos pelo portal SISFLORA
(Sistema de Comercialização e transportes de Produtos Florestais) e pelos relatórios de
Extração e Comercio de Toras de Madeiras Nativas por Essência da SEMAS (Secretaria
do Meio Ambiente e Sustentabilidade).
Utilizando o Excel os dados referentes aos nomes científicos e sua ocorrência no
Estado do Pará e no Brasil, foram organizados e corrigidos visando sua verídica ortografia
e ocorrências, adquiridas por meio do sistema da Flora do Brasil. Por meio dos dados
obtidos pelo Flora do Brasil (Reflora), foi realizada uma verificação referente às
características macromorfológicas que são importantes para a identificação da família e
da espécie em campo.
3. RESULTADOS E DISCUSSÃO
No presente estudo, verificou-se a existência de 8 nomes vernaculares na base de
dados do SISFLORA, que contabilizam o total de 147.696,28 m³ e R$16.941.638,63.
(Quadro 1).
Foram listados 11 nomes científicos, sendo que, após a correção na plataforma do
Reflora, apenas 5 nomes são considerados válidos - Anacardium spruceanum Benth. ex
Engl., Anacardium sp., Anacardium parvifolium Ducke, Anacardium occidentale L.,
Anacardium microsepalum Loes-. Além disso, verificou-se na mesma plataforma que a
espécie Anacardium excelsum não ocorre no Brasil e a Anacardium parvifolium não
ocorre o Pará.

375
Belém (PA), 11 a 13 de dezembro de 2019
Quadro 1- Volumes e valores associados aos nomes vernaculares da base de dados do SISFLORA.
Nome vernacular Volume (m³) Valor (R$)
Caju 34.529,99 4.369.902,32
Cajuaçu 88.664,37 9.919.026,91
Caju-da-mata 1.640,31 187.836,43
Caju-de-janeiro 1.745,24 218.041,98
Caju-do-mato 4.862,34 460.887,20
Cajueiro 1.326,47 106.497,31
Cajuí 14.868,59 1.674.573,67
Cajuí-da-mata 59,07 4.902,81
Total: 147.696,38 16.941.638,63
Fonte: SISFLORA. Quadro 2- Volumes e valores associados aos nomes científicos e quantidade de nomes
vernaculares (Qv).
Nome científico correto Nome científico Qv Volume
(m³) Valor (R$)
Anacardium occidentale L. Anacardium
curatellaefolium A.St.-
Hil.
1 329 51.268,11
Anacardium excelsum (Bertero
& Balb. ex Kunth) Skeels Anacardium excelsum
Skeels. 2 2.313,56 284.564,37
Anacardium giganteum
W.Hancock ex Engl. Anacardium giganteum
Hanck ex Engl. 6 71.264,76 8.042.938,35
Anacardium occidentale L. Anacardium microcarpum
Ducke 2 2.883.30 367.831,44
Anacardium microsepalum
Loes. Anacardium
microsepalum Loes 1 1.609,07 181.327,51
Anacardium microsepalum
Loes. Anacardium negrense
Pires & Fróes 1 243,01 18.525,39
Anacardium occidentale L. Anacardium occidentale
L. 1 4.200,73 582.982,78
Anacardium parvifolium
Ducke Anacardium parvifolium
Ducke 3 20.030,67 2.356.127,03
Anacardium sp. Anacardium sp. 1 1.745,24 218.041,98
Anacardium spruceanum
Benth. ex Engl. Anacardium spruceanum
Benth. ex Engl. 4 42.654,34 4.791.437,1
Anacardium parvifolium
Ducke Anacardium tenuifolium
Ducke 2 422,7 46.624,57
Fonte: Reflora e SISFLORA.

376
Belém (PA), 11 a 13 de dezembro de 2019
A espécie Anacardium giganteum. possui a maior quantidade de nomes
vernaculares relacionados - caju, cajuaçu, caju-do-mato, cajueiro, cajuí e cajuí-da-mata -
e consequentemente, o maior volume e valor, respectivamente, 71.264,76 m³ e R$
8.042.938,35. Enquanto a espécie Anacardium microsepalum Loes. (nome corrigido da
espécie Anacardium negrense Pires & Fróes), apresenta apenas um nome vernacular,
cajú, e o menor volume e valor, respectivamente, 243,01 m³ e R$18.525,39.
A diversidade de nomes vernaculares associados a um único nome científico
ocasiona uma identificação incerta das espécies, ocasionando indevidas explorações que
afetam a biodiversidade e a comercialização. Segundo a publicação de SANTOS, S R.
M., O. R. KATO & M. M. TOURINHO, 2019, a espécie Anacardium giganteum
W.Hancock ex Engl. é classificada como cajuí. E Gonçalves & Lucas, 2017 classificam
a mesma espécie com o nome vernacular de caju-do-mato.
Para a identificação correta das espécies, é necessário o reconhecimento das
características macromorfológicas referentes a tipos foliares, tronco, casca, entre outros
(Quadro 3).
Quadro 3- Características macromorfológicas correspondente ao nome científico correto.
Espécies Folhas Casca Tronco
Anacardium spruceanum
Benth. ex Engl. Alternas compostas, ou
imparipinadas, ou simples Lisa com
lenticelas Base reta
Anacardium parvifolium
Ducke Simples, agregadas em
direção ao ápice Lisa Base reta
Anacardium occidentale L. Alternas, pecioladas, ovadas
e obtudas Escamosa ou
lisa Tortuoso e
ramificado
Anacardium excelsum
Skeels. Simples e sem estípulas,
relativamente grandes Escamosa ou
lisa Base reta
Anacardium giganteum
Hanck ex Engl Simples, alternas e
espiraladas Lisa com
lenticelas Aéreo
Anacardium microsepalum
Loes Pecioladas, com a base
obtusa Lisa com
lenticelas Aéreo
Fonte: Flora do Brasil e Flora da Reserva Ducke (1999).

377
Belém (PA), 11 a 13 de dezembro de 2019
4. CONSIDERAÇÕES FINAIS
Pela análise da pesquisa, conclui-se que 11 nomes científicos correspondem a 8
nomes vernaculares do caju e suas variantes, por conta da identificação incorreta ou
desconhecida feita comumente feita por mateiros.
A identificação equivocada das espécies da família Anacadiaceae, podem
acarretar uma exploração desordenada e uma comercialização imprópria prejudicando a
finalidade para o mercado madeireiro devido as suas particularidades.
Consequentemente, afeta a biodiversidade e a manutenção ecológica da região explorada.
O reconhecimento das espécies pode ser facilitado por meio das características
macromorfológicas como folha, casca e tronco que podem se diferenciar entre espécies
distintas.
Desse modo, a correção e divulgação da nomenclatura corrigida de espécies
dentro do gênero Anacardium são imprescindíveis para auxiliar estudos científicos
visando o manejo adequado das espécies.

378
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ARAÚJO, J. A.; NASCIMENTO, C. C. da.; CALDERON, C. M. A.; CALDERON, R. A
IDENTIFICAÇÃO ANATÔMICA DE MADEIRA COMO FERRAMENTA AUXILIAR
DA SUSTENTABILIDADE DOS RECURSOS FLORESTAIS MADEREIROS. Seminário
internacional de ciências do ambiente e sustentabilidade na Amazônia, IV.; ENCONTRO
AMAZÔNICO DA ASSOCIAÇÃO NACINAL DE PÓS-GRADUAÇÃO E PESQUISA EM
MEIO AMBIENTE E SOCIEDADE, 1°,2016.
KANASHIRO, M. 2002.AS DIFERENÇAS ENTRE AS ÁRVORES. Pesquisa FAPESP, nº 82.
São Paulo.
LIMA, P. P.; ILKIU-BORGES, F.; FARO, B. L. S. de O.; COSTA, P. A. da. IMPORTÂNCIA
DA IDENTIFICAÇÃO BOTÂNICA NA COMERCIALIZAÇÃO DE ESPEÉCIES
MADEREIRAS. SEMINÁRIO DE INICIAÇÃO CIENTÍFICA, 19.; SEMINÁRIO DE PÓS-
GRADUAÇÃO DA EMBRAPA AMAZÔNIA ORIENTAL, 3., 2015, Belém, PA.
SANTOS, S. R. M., O. R. KATO & M. M. TOURINHO, 2019. DIVERSIDADE
FLORÍSTICA E ESTOQUE DE CARBONO DE SISTEMAS AGROFLORESTAIS EM
DOIS MUNICÍPIOS DO NORDESTE PARAENSE, BRASIL. Boletim do Museu Paraense
Emílio Goeldi. Ciências Naturais, 14(1): 31-4. Disponível em:
<https://ww.bb.com.br/docs/pub/inst/dwn/Vol4FrutCaju.pdf>. Acesso em: 6.nov.2019

379
Belém (PA), 11 a 13 de dezembro de 2019
MAPEAMENTO E LEVANTAMENTO DAS COLEÇÕES DE ESPÉCIES
ORNAMENTAIS DO HERBÁRIO FELISBERTO CAMARGO REGISTRADAS
PELO BRAHMS
Thalisson Johann Michelon de Oliveira1; Anderson dos Santos Monteiro2; Matheus
Lopes Nascimento3; Roberta Marselle dos Santos4; Manoel Euclides do Nascimento5.
1 Graduando em Agronomia. Universidade Federal Rural da Amazônia.
[email protected]. 2 Graduando em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 3 Graduando em Engenharia Florestal. Universidade Federal Rural da Amazônia.
[email protected]. 4 Graduanda em Agronomia. Universidade Federal Rural da Amazônia.
[email protected]. 5 Doutor em Fitotecnia e curador do Herbário Felisberto Camargo. Universidade Federal
Rural da Amazônia. [email protected].
RESUMO
O projeto Herbário Virtual de Coleções Temáticas com Espécies da Flora Amazônica
com Interesse Agronômico e Florestal discrimina as espécies conforme seu potencial,
incluindo utilização madeireira, ornamental, invasoras, medicinais e frutíferas. Este
trabalho teve como objetivo quantificar as espécies das coleções ornamentais que já foram
incorporadas ao banco de dados do Herbário. Foram realizadas coletas de material
botânico no Parque Estadual do Utinga (PEUT) e na Universidade Federal Rural da
Amazônia (UFRA - Belém). O método de coleta foi o usual em Taxonomia Vegetal. As
amostras foram prensadas em campo, levadas para secar em estufa e montadas em
cartolina no tamanho padrão para exsicatas. O programa Brahms (Botanical Research
And Herbarium Management System) – específico para fins de gerenciamento de dados
de herbário – auxiliou na organização das diferentes espécies coletadas e na junção das
imagens das exsicatas com dados de campo. O mapa de localização foi feito através de
software de sistema de informação geográfica (QGIS – versão 3.4). Foram coletadas 262
amostras, com 39 famílias distintas, sendo a Fabaceae a que mais teve espécies,
totalizando 18% do total, e dentro desta família foram encontrados 15 gêneros distintos.
Esta primeira fase do projeto contribuiu enormemente para a área de Taxonomia Vegetal
e áreas correlatas da UFRA, pois restaurou o herbário da instituição e já possui espécies
ornamentais depositadas e organizadas conforme a perspectiva de coleções temáticas,
com vistas a criação de um site para disponibilização destas informações, assim a
disseminação das informações das espécies coletadas se tornará mais abrangente e mais
prática para muitos interessados, otimizando o tempo que levariam para realizar a mesma
consulta em herbários físicos.
Palavras-chave: Flora amazônica. BRAHMS. QGIS. Fabaceae.
Área de Interesse do Simpósio: Sistemática e Biogeografia.

380
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Desde que o ser humano começou a se relacionar mais com o ambiente através da
fixação em um determinado lugar, o mesmo começou a fazer atividades com plantas com
o viés estético e paisagístico, sendo que gerações futuras começaram a cultivá-la com
outras finalidades, como na cultura e religiosidade (HEIDEN et al, 2006). Além de que o
cultivo de plantas pelo prazer estético está intrinsicamente ligado a relevância ambiental
e socioeconômica.
Muitas plantas são comercializadas para cultivo por sua beleza, podendo utilizá-
las em paisagismo, arquitetura e em diversas áreas, denomina-se à essas plantas sendo
como ornamentais. O mercado brasileiro de flores e plantas ornamentais está em grande
crescimento atualmente no Brasil (SEBRAE, 2015), visto que muitas pessoas começaram
a utilizar produtos naturais para a decoração em suas casas e locais públicos, logo é uma
ótima fonte de renda para pequenos e médios produtores (LINS, 2004). Além do que
conforme Lilian Anefalos et al (2003), este setor está se expandindo para o mercado
externo.
Consoante Araújo et al (2013), herbários em si são ótimos projetos de ensino,
extensão e pesquisa. Tendo em vista o ensino, muitos herbários aceitam visitantes para
exposição de seus trabalhos, o que seria uma ótima forma de assimilação taxonômica e
morfológica para serem utilizadas em práticas desenvolvidas pelos professores, visto que
são ótimos lugares para ministração de aulas botânicas. Na extensão, herbários ofertam a
toda a comunidade conhecimentos sobre a flora local, o que se feito de maneira correta,
geraria maior interesse em cuidar do ambiente. E em pesquisas, herbários abrigam muito
conhecimento científico, estimulando a criação de artigos para a disseminação desse
conhecimento no meio científico.
O Herbário Felisberto Camargo, da Universidade Federal Rural da Amazônia, foi
criado na segunda metade do século XX. O objetivo de sua criação, foi, sobretudo,
fornecer subsídios às disciplinas Botânica e Sistemática Vegetal, bem como contribuir ao
desenvolvimento de trabalhos de cunho científico na graduação e pós-graduação,
naquelas disciplinas e em áreas afins. Desde sua criação, várias coletas foram realizadas
por sucessivas turmas de alunos dos cursos de Agronomia e Engenharia Florestal, em que
cerca de 3.600 amostras encontram-se registradas.

381
Belém (PA), 11 a 13 de dezembro de 2019
A região amazônica, especificamente a cidade de Belém, conta com dois grandes
herbários, o da EMBRAPA e do Museu Paraense Emilio Goeldi, porém, nesses herbários,
o objetivo principal é fazer coletas generalizadas e amplos levantamentos botânicos para
formar coleções clássicas de toda flora amazônica. Entretanto, os referidos herbários não
trabalham em contato direto com o aluno, ou destinadas a atender áreas específicas de
estudo nas Ciências Agrárias, como prevê a presente proposta. Assim, é necessária a
formação de coleções temáticas com espécies de interesse agroflorestais para atender aos
anseios dos professores e estudantes desta universidade, nos cursos de Graduação e Pós-
Graduação, outros profissionais de áreas afins e a própria comunidade.
O projeto Herbário Virtual de Coleções Temáticas com Espécies da Flora
Amazônica com Interesse Agroflorestais discrimina as espécies conforme seu uso,
incluindo utilização madeireira, ornamental, invasoras, medicinais e frutíferas. Este
trabalho teve como objetivo quantificar as espécies das coleções ornamentais que já foram
incorporadas ao banco de dados do programa Brahms sobre o Parque Estadual do Utinga
(1°25'33.6"S 48°26'40.6"W) e Universidade Federal Rural da Amazônia (1°27'17.5"S
48°26'46.0"W) e levantar mais dados a respeito.
2. METODOLOGIA
Foram realizadas coletas de material botânico na Universidade Federal Rural da
Amazônia (UFRA) – Campus Belém, e no Parque Estadual do Utinga (PEUT) foi através
de registros feitos pelo BRAHMS em diversos períodos a partir do mês de março de 2018
– mês em que foi inaugurado o PEUT após sua revitalização.
O método de coleta foi a utilização de podão e tesoura de poda, sendo que as
amostras foram prensadas em campo para que não ocorra a degradação do material até
que o mesmo chegue ao laboratório para a prensagem final evidenciando as flores e
características peculiares da amostra. Para a proteção dos coletores foi feita a utilização
dos Equipamentos de Proteção Individual (EPI), como o uso de capacete, luva, calça
comprida e perneira para evitar picadas de animais peçonhentos.

382
Belém (PA), 11 a 13 de dezembro de 2019
Após a coleta as amostras foram levadas para secar em estufa a 60°C por um
período de 72h para retirar a umidade e evitar que o material botânico crie fungos e
degrade o material fazendo com que o mesmo perca características importantes.
A montagem foi feita em cartolina no tamanho padrão para exsicatas, as colagens
foram com cola de isopor para a fixação do material, os envelopes foram feitos com papel
manteiga e colada com cola branca.
Após finalizadas, as amostras ficam armazenadas em locais com temperatura e
umidade controlada, a temperatura deve estar em torno de 25° C e a umidade em 50%,
pois assim evita a proliferação de fungos que estão naturalmente pelo ar nas espécies
coletadas, fazendo com que assim o material fique conservado por mais tempo (ROTTA
et al, 2008).
O programa Brahms (Botanical Research And Herbarium Management System)
– específico para fins de gerenciamento de dados de herbário – auxiliou na organização
das diferentes espécies coletadas e na junção das imagens das exsicatas com dados de
campo. O programa também auxiliou na criação da etiqueta com todos os dados
necessários para ser colada no material como pode ser observado na figura 1.

383
Belém (PA), 11 a 13 de dezembro de 2019
O mapa de localização das espécies foi feito através de softwares de sistema de
informação geográfica (QGIS – Versão 3.4), com coordenadas geográficas coletadas em
campo. Ademais, os shapefiles utilizados foram adquiridos no site do IBGE e Ideflor.
3. RESULTADOS E DISCUSSÃO
O número de espécies coletadas soma 262 amostras em 39 famílias distintas:
Apocynaceae, Bignoniaceae, Fabaceae, Malvaceae, Melastomataceae, Nyctaginaceae e
Rubiaceae, estas são as que apresentaram maior quantitativo de espécies. Como pode-se
observar na tabela 1 a família Fabaceae apresenta 15% de todo o quantitativo coletado,
Apocynaceae 10,8% e Rubiaceae com 9,6%. Conforme o Jardim Botânico do Rio de
Janeiro (2019), a flora brasileira conta com 33.300 espécies de angiospermas, com a
família Fabaceae representando 8,4% deste todo, Rubiaceae com 4,2% e Apocynaceae
com 2,4%. No Pará registraram-se 6537 espécies de plantas angiospermas diferentes.
Fabaceae Lindl. se encontrou como a família em destaque dentro da coleta, e os
principais gêneros encontrados foram: a Acacia Mill., Albizia Durazz., Arachis L.,
Bauhinia L., Caesalpinia L., Calliandra Benth., Cassia L., Cenostigma Tul., Centrosema
(DC.) Benth., Clitoria L., Cynometra L., Delonix Raf., Mimosa L. e Senna Mill.. Essa
grande variedade de gêneros e espécies de plantas se dá pelo fato desta família ter a forma
de vida como arbusto, árvore, liana e erva nos ambientes aquáticos, epífitos, rupícolas e
terrícolas, tanto é que sua distribuição se dá por todas as regiões do Brasil com 222
gêneros e 2807 espécies (REFLORA, 2019).
Consoante Cardoso et al (2005) que também realizou um levantamento de
espécies botânicas, afirma que estudos com este objetivo são importantes pois subsidiam
outros trabalhos interligados a botânica, preservação, conservação e repovoamento das
espécies da flora local.
Na tabela 1 pode-se observar a quantidade de amostras, gêneros e espécies de cada
família. Apesar da Fabaceae apresentar-se em destaque nas coleções, a espécie em maior
quantidade foi a Hibiscus rosa-sinensis L., um arbusto da família Malvaceae, a espécie
possui flores de coloração vermelha muito atrativa para agentes polinizadores, com folhas
simples de filotaxia alterna, além do mais, as folhas tem formato ovalado pontiagudo e
com a margem serrilhada.

384
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1. Dados de amostras, gêneros e famílias coletadas do BRAHMS no ano de 2019, Belém
(PA).
Famílias Quantitativo
Amostras Gêneros Espécies
Fabaceae Lindl. 48 14 20
Apocynaceae Juss. 29 5 8
Rubiaceae Juss. 25 5 7
Bignoniaceae Juss. 22 3 3
Malvaceae Juss. 20 3 3
Nyctaginaceae Juss. 20 1 3
Melastomataceae A. Juss 13 4 4
Acanthaceae Juss. 9 4 6
Outros 84 42 56
Total 272 81 110
Fonte: Os autores, 2019.
Pode-se notar que nas coletas realizadas, Malvaceae Juss. apresentou 20 amostras
distribuído em 3 gêneros e 3 espécies, quando comparado a flora brasileira a Malvaceae
tem 791 espécies e 73 gêneros.
Como a Fabaceae teve mais amostras, foi realizado um mapa de localização de
todas essas espécies ornamentais (figura 3), ao todo encontram-se 49 amostras com 21
espécies, para os pontos de coleta visíveis, sendo selecionada a área em que mais se
concentravam esses pontos. A Senna multijuga (Rich.) H.S.Irwin & Barneby foi a espécie
dessa família com maior número de coletas, nas coleções registradas esta espécie foi
coletada 6 vezes, sendo cinco amostras coletadas na Universidade Federal Rural da
Amazônia e uma coletada no Parque Estadual do Utinga.
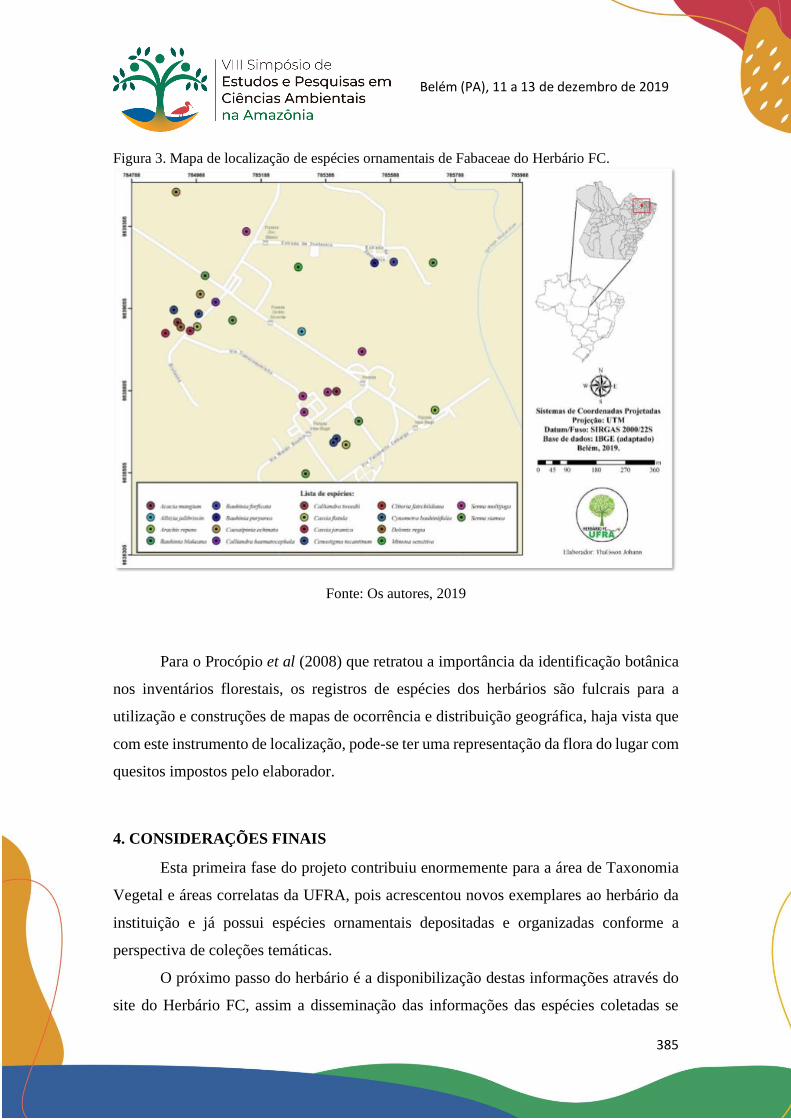
385
Belém (PA), 11 a 13 de dezembro de 2019
Figura 3. Mapa de localização de espécies ornamentais de Fabaceae do Herbário FC.
Fonte: Os autores, 2019
Para o Procópio et al (2008) que retratou a importância da identificação botânica
nos inventários florestais, os registros de espécies dos herbários são fulcrais para a
utilização e construções de mapas de ocorrência e distribuição geográfica, haja vista que
com este instrumento de localização, pode-se ter uma representação da flora do lugar com
quesitos impostos pelo elaborador.
4. CONSIDERAÇÕES FINAIS
Esta primeira fase do projeto contribuiu enormemente para a área de Taxonomia
Vegetal e áreas correlatas da UFRA, pois acrescentou novos exemplares ao herbário da
instituição e já possui espécies ornamentais depositadas e organizadas conforme a
perspectiva de coleções temáticas.
O próximo passo do herbário é a disponibilização destas informações através do
site do Herbário FC, assim a disseminação das informações das espécies coletadas se

386
Belém (PA), 11 a 13 de dezembro de 2019
tornará mais abrangente e trará mais praticidade a pesquisadores da área, comunidade
científica em si e a quem interessar, otimizando o tempo que levariam para realizar a
mesma consulta em herbários físicos.

387
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ANEFALOS, Lilian C., GUILHOTO, Joaquim J. M. Estrutura do mercado brasileiro
de flores e plantas ornamentais. Agric. São Paulo, SP. v. 50, n. 2, pág. 41-63, 2003.
Disponível em: <https://mpra.ub.uni-
muenchen.de/38318/1/MPRA_paper_38318.pdf>.Acesso em: 06/04/2019.
ARAÚJO, Miria S., MIGUEL, João R. Herbário Didático no ensino da Botânica. In:
Anais do I Encontro de Pesquisa em Ensino de Ciência e Matemática: questões atuais da
Universidade Unigranrio. v. 1 n. 1 p. 1-3, 2013. Disponível em:
<http://publicacoes.unigranrio.edu.br/index.php/pecm/article/view/2216/1035>. Acesso
em: 06/04/2019.
CARDOSO, Jean Carlos et al. Levantamento de espécies da família Orchidaceae em
Águas de Sta. Bárbara (SP) e seu cultivo. Hortic. bras. v. 23, n. 2, 2005. Disponível em:
<http://www.scielo.br/pdf/%0D/hb/v23n2/25046.pdf>. Acesso em: 16/09/2019.
PROCÓPIO, Lílian Costa; SECCO, Ricardo de Souza. A importância da identificação
botânica nos inventários florestais: o exemplo do “tauari” (Couratari spp. e Cariniana spp.
- Lecythidaceae) em duas áreas manejadas no estado do Pará. Acta Amazonica. v. 38, n.
1, p. 31-44, 2008. Disponível em: <http://www.scielo.br/pdf/aa/v38n1/v38n1a05.pdf>.
Acesso em: 17/09/2019.
REFLORA. Fabaceae in Flora do Brasil 2020 em construção. Jardim Botânico do Rio de
Janeiro. Disponível em: <http://reflora.jbrj.gov.br/reflora/floradobrasil/FB115>. Acesso
em: 16/09/2019.
HEIDEN, Gustavo et al. Considerações sobre o uso de plantas ornamentais nativas.
Revista Brasileira de Horticultura Ornamental. v. 12, n.1, p. 2-7, 2006. Acesso em:
07/04/2019.
LINS, Servina R. O. & COELHO, Rildo S. B. Ocorrência de Doenças em Plantas
Ornamentais Tropicais no Estado de Pernambuco. Fitopatol. bras. v. 29, n. 3, pág. 332-
335, 2004. Disponível em: <http://www.scielo.br/pdf/%0D/fb/v29n3/20374.pdf>.
Acesso em: 06/04/2019.
ROTTA, Emilio et al. Manual de Prática de Coleta e Herborização de Material Botânico.
EMBRAPA. Colombo, PR. Disponível em:
<https://www.infoteca.cnptia.embrapa.br/bitstream/doc/315636/1/Doc173.pdf>. Acesso
em: 07/11/2019.
SEBRAE. Serviço Brasileiro de Apoio às Micro e Pequenas Empresas. Flores e plantas
ornamentais do Brasil. v. 1. 2015. Disponível em:
<http://www.bibliotecas.sebrae.com.br/chronus/ARQUIVOS_CHRONUS/bds/bds.nsf/7
ed114f4eace9ea970dadf63bc8baa29/$File/5518.pdf>. Acesso em: 07/11/2019.

388
Belém (PA), 11 a 13 de dezembro de 2019
APÊNDICE A: Local de fotografia das exsicatas

389
Belém (PA), 11 a 13 de dezembro de 2019
COMPORTAMENTO ALIMENTAR DE ESTRELAS-DO-MAR EM AMBIENTE
RECIFAL CONTROLADO
Lucas Garcia Martins1; Ana Carla de Araújo Gomes2; Rafael Anaisce das
Chagas3; Vicente Savonitti Miranda4
1 Graduando em Engenharia de Pesca na Universidade Federal Rural da Amazônia.
Laboratório de Pesca e Biodiversidade Aquática. E
mail: [email protected] 2 Graduanda em Engenharia de Pesca na Universidade Federal Rural da Amazônia.
Museu de Conservação em Ciências Naturais. Email: [email protected] 3Doutorando em Ecologia (PPGEco/UFPA/Embrapa), pesquisador no Museu de
Zoologia (MZUFRA). Universidade Federal do Pará Email: [email protected] 4Doutor em Biotecnologia, Professor da Universidade Federal Rural da Amazônia.
Museu de Conservação em Ciências Naturais. Email: [email protected]
RESUMO
Equinodermos são animais bentônicos encontrados em todas as latitudes, desde a região
entre marés até a zona abissal e mar profundo. Por sua beleza, são comumente utilizadas
como animais ornamentais. No entanto, ainda são escassos os estudos acerca das
adaptações e variações alimentares de estrelas do mar em aquários marinhos. O presente
trabalho teve como objetivo analisar a preferência alimentar, entre ração industrial e
alimento vivo de quatro espécies de estrelas-do-mar confinadas em ambiente controlado.
Os animais foram mantidos em três aquários: 800L (Turbinella sp.), 500L (Protoreaster
linckii e P. nodosus) e 100L (Echinaster (othilia) spinulosus) na Sala de Conservação de
Ciências Naturais localizada na Universidade Federal Rural da Amazônia. A alimentação
dos animais foi feita utilizando a ração industrializada (Tetra Veggie) e o alimento vivo
(Asterina sp.) durante período de julho a setembro de 2019 com rotina de coleta de três
dias semanais com 6 a 8 horas de esforço de observação em cada dia. A análise da
preferência alimentar das estrelas foi feita utilizando o teste do Qui Quadrado no
programa Statistic (versão 7.0) a um nível de significância de 5%. Para o teste da hipótese
de que mesmo em condições experimentais e alimentadas anteriormente com ração
industrializada, as estrelas naturalmente preferirão alimento vivo, a análise foi realizada
a nível de espécie separadamente. A partir das análises demonstrou-se que,
estatisticamente, não houve preferência alimentar entre os dois tipos de alimentos,
contudo pode-se perceber a manutenção do comportamento alimentar das estrelas em
relação a sua fonte alimentar, a ração industrial representou apenas mais um recurso
alimentar aos animais. Conclui-se que as estrelas não apresentaram preferência alimentar,
mas mantém seus hábitos alimentares naturais, além disso mostram-se como
controladores paliativos de Asterinas sp., trazendo uma nova solução aos aquaristas que
possuem corais vivos.
Palavras-chave: Aquarismo. Alimentação. Controle biológico
Área de Interesse do Simpósio: Zoologia

390
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O ambiente marinho é habitado por diversos animais, nos quais os invertebrados
representam uma grande parcela. Destes, um dos grupos mais comuns e conhecidos são
os pertencentes ao filo Echinodermata (Grego: echinos=espinhoso, derma= pela). O filo
é constituído por cerca de 7.000 espécies distribuídas em seis classes: Asteroidea
(estrelas-do-mar), Ophiuroidea (ofíuros ou serpentes-do-mar), Crinoidea (lírios-do-mar),
Echinoidea (ouriços-do-mar e bolachas-do-mar), Holothuroidea (pepinos-do-mar) e
Concentricloidea (margaridas-do-mar) (HICKMAN, 2016; BRUSCA e BRUSCA, 2007).
Equinodermos são animais bentônicos encontrados em todas as latitudes, desde a
região entre marés até a zona abissal e mar profundo, além disso podem habitar tanto
substratos consolidados como inconsolados ou até vivendo sobre outros organismos
(STORER et al., 2005; BRUSCA e BRUSCA, 2007).
Os organismos que compõem a classe Asteroidae são vulgarmente conhecidos
como “estrelas do mar”, são membros conspícuos das áreas litorâneas, habitando
sobretudo em costões rochosos. Possuem simetria pentaradial, com membros chamados
braços fundidos na região central do corpo (disco central), sendo achatados e flexíveis,
cobertos por endoesqueleto calcário onde se insere uma epiderme coberta de espinhos e
de diferentes colorações (BOOLOOTIAN, 1966; RIBEIRO-COSTA e ROCHA, 2006).
Devido sua beleza são bastante representativas como organismos ornamentais
(CARRACA, 2016). Normalmente são utilizados em conjunto com corais na criação de
aquários que ilustram ambientes de recifes (BACELAR, 2007). Portanto, a elaboração
dos ambientes marinhos artificiais engloba espécies de diferentes regiões e que, segundo
estudos de MORISHITA et al. 2010, podem apresentar mudanças nos hábitos de cada
espécie e suas interações tróficas, além das mudanças em relação aos processos físico-
químicos da água do aquário.
Estudos acerca de reprodução e distribuição têm sido amplamente aplicados
(SAMPAIO, 2010; MOURA, 1999; LIMA, 2006). Além disso, tem sido utilizado
modelos de dinâmica populacional e ecologia alimentar para auxiliar em estudos de
história de vida e modelos morfométricos (MASCARENHAS; 1996; TAVARES, 2007).
Adicionalmente, são muito fortes os estudos de diversidade de equinodermos (XAVIER,
2010; LIMA, 2009; MANSO, 2008). No entanto, ainda existe uma lacuna acerca dos
estudos sobre preferência alimentar e comportamento em cativeiro.

391
Belém (PA), 11 a 13 de dezembro de 2019
Neste sentido, o presente trabalho teve como objetivo analisar a preferência
alimentar, entre ração industrial e alimento vivo de quatro espécies de estrelas-do-mar
confinadas em ambiente controlado. Considerando que as estrelas-do-mar são animais
carnívoros, nossa hipótese é que, mesmo em condições experimentais e alimentadas
anteriormente com ração industrializada, as estrelas naturalmente preferirão alimento
vivo.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
Os animais foram mantidos em três aquários: 800L (Turbinella sp.), 500L
(Protoreaster linckii e P. nodosus) e 100L (Echinaster (othilia) spinulosus) na Sala de
Conservação de Ciências Naturais localizada na Universidade Federal Rural da
Amazônia.
Figura 1: Fotos dos aquários do experimento, em A aquário de 800L, em B 300L e C 100 L
Fonte: Os autores
2.2 TIPO DE PESQUISA

392
Belém (PA), 11 a 13 de dezembro de 2019
As asterinas (Asterina sp.) são pequenos equinodermos que por vezes representam
uma ameaça aos aquários marinhos por viverem em associação aos corais para se
alimentarem de sua gastroderme (ROWE e GATES, 1995). No entanto, o controle dela
ainda é difícil de ser feito pela ausência de estudos de sua biologia e pela intensa
reprodução por fissão (EMSON e CRUMP, 1979), por tal fato foi escolhida como
alimento vivo deste experimento.
2.3 COLETA DE DADOS
A alimentação dos animais foi feita utilizando a ração industrializada (Tetra
Veggie) e o alimento vivo (Asterina sp.) (Fig. 2) durante período de julho a setembro de
2019 com rotina de coleta de três dias semanais com 6 a 8 horas de esforço de observação
em cada dia. As estrelas eram alimentadas simultaneamente com a ração e alimento vivo.
Ao final das observações eram planilhados os números de Asterinas sp. consumidas e a
ração industrializada.
Figura 2: Em A ração industrializada utilizada e em B o alimento vivo, a Asterina sp.
Fonte: A: modificado de produto.mercadolivre.com.br/ B: modificado de instarix.org
2.4 ANÁLISE DE DADOS
A análise da preferência alimentar das estrelas foi feita utilizando o teste do Qui
Quadrado no programa Statistic (versão 7.0) a um nível de significância de 5%. Para o
teste da hipótese de que mesmo em condições experimentais e alimentadas anteriormente
com ração industrializada, as estrelas naturalmente preferirão alimento vivo, a análise foi
realizada a nível de espécie separadamente.

393
Belém (PA), 11 a 13 de dezembro de 2019
3. RESULTADOS E DISCUSSÃO
Figura 3: Em A E. spinulosus, em B P. nodosus e em C Turbinella sp se alimentando de
asterinas
Fonte: Os autores
Ao analisar a preferência alimentar de Turbinella sp., verifica-se que 72,2% (26)
das vezes alimentou-se apenas Asterina sp., 2,8% (1) de apenas ração industrial e 13,9%
(5) dos dois alimentos, sendo 11,1% (4) não se alimentando. Tais diferenças foram
consideradas como devidas ao acaso (χ2=0,464; gl=1; p=0,829) e deste modo, por mais
que numericamente seja diferente, estatisticamente refutou-se a hipótese de preferência
alimentar.
Ao analisar a preferência alimentar de Protoreaster linckii, verifica-se que 36,0%
(13) das vezes alimentou-se apenas Asterina sp., 2,8% (1) de apenas ração industrial e
61,1% (22) dos dois alimentos. Assim como no exemplo anterior, tais diferenças foram
consideradas como devidas ao acaso (χ2=0,581; gl=1; p=0,445) e deste modo, por mais
que numericamente seja diferente, estatisticamente refutou-se a hipótese de preferência
alimentar.
Do mesmo modo, ao analisar a preferência alimentar de Protoreaster nodosus,
verifica-se que 38,90% (14) das vezes alimentou-se apenas Asterina sp., 2,8% (1) de
apenas ração industrial e 58,3% (21) dos dois alimentos. Tais diferenças foram
consideradas como devidas ao acaso (χ2=0,654; gl=1; p=0,418) e deste modo, por mais
que numericamente seja diferente, estatisticamente refutou-se a hipótese de preferência
alimentar.
Por último, ao analisar a preferência alimentar de Echinaster (othilia) spinulosus,
verifica-se que 41,7% (15) das vezes alimentou-se apenas Asterina sp., 16,7% (6) de
apenas ração industrial e 2,8% (1) dos dois alimentos, sendo 38,9% (15) não se
alimentando. Tais diferenças foram consideradas como devidas ao acaso (χ2=3,2; gl=1;

394
Belém (PA), 11 a 13 de dezembro de 2019
p=0,073) e deste modo, por mais que numericamente seja diferente, estatisticamente
refutou-se a hipótese de preferência alimentar.
Turbinella sp. embora, estatisticamente, não tenha apresentado preferência
alimentar em relação as Asterinas e ração, demonstrou que mantém seu hábito alimentar
natural na qual é predominantemente carnívora (BARNES, 1996), mas podendo se
alimentar da ração industrial que possui as proteínas de origem animal.
A estrela P. linckii que é bastante explorada em aquários do mundo devido sua
beleza (INDIVIGLIO, 2006), no entanto é considerada um animal incompatível com
outros invertebrados devido ser uma predadora voraz de corais mole, poliquetas
tubulares, esponjas, ameijoas e outras estrelas (SCHEIBLING, METAXAS, 2008).
Portanto, embora não tenha mostrado preferência, mantém o comportamento natural de
predador oportunista alimentando-se tanto das asterinas como da ração industrial.
Vulgarmente conhecido como estrela-do-mar com chifres ou estrela-do-mar com
pedaços de chocolate, P. nodosus é uma espécie com preferência por substratos
inconsolidados fundos abrigados, arenosos ou levemente enlameados (WABNITZ,
GREEN E RAZAK, 2003). Ela é uma espécie oportunista, quando adultos atacam
principalmente invertebrados sésseis como corais duros e esponjas, em aquários acaba
sendo uma ameaça aos corais, além disso também pode se alimentar de outros
invertebrados como gastrópodes e estrelas (BOS, 2008). A partir do visto, ela mantém-se
no aquário como predador oportunista que se alimentou das duas fontes alimentares,
percebeu-se que apesar de se alimentar muitas vezes apenas das asterinas, o percentual
de vezes que se alimentou de ambos alimentos foi maior que caracterizou a não
preferência.
A estrela E. spinulosus também conhecida como estrela vermelha é uma espécie
bastante comum no litoral brasileiro que se alimenta sobretudo de esponjas do mar,
anêmonas, ofiuróides, caranguejos e moluscos. No entanto, estudos de QUEIROZ et al.
(2012) e AMARAL et al. (1996), observaram que esta espécie apresenta preferência
alimentar por poríferos, especialmente as esponjas moles. No ambiente em que era
mantido o espécime, apesar da ração e asterinas via-se que esta espécie em questão se
alimentava principalmente dos poríferos da classe demospongie que se mantinham
incrustados nas rochas. Embora, aparentemente, tenha mostrado interesse maior nas

395
Belém (PA), 11 a 13 de dezembro de 2019
esponjas, percebemos que podia se alimentar tanto das asterinas como da ração industrial
devido sua plasticidade alimentar.
As análises (Figura 4) mostraram que para Turbinella sp., P. linckii e P. nodosus
se alimentaram com maior frequência das asterinas e de asterinas e ração no mesmo dia
que demostra que, mesmo mantidas confinadas em ambiente artificial, mantém seus
hábitos alimentares naturais e evidencia a clara adaptação a alimentação por ração
industrial como nova fonte energética. Apesar de não ser tão significativa se alimentando
da ração, E. spinulosus mostrou-se uma controladora paliativa em potencial.
Figura 4: Gráfico de comparação da dieta alimentar das estrelas
Fonte: Os autores
As estrelas do mar são, no ambiente bentônico, animais que normalmente ocupam
o topo da cadeia alimentar (HONEY-ESCANDÓN, SOLÍS-MARÍN e LAGUARDA-
FIGUERAS, 2008). No entanto, estudos acerca da alimentação das espécies em questão
ainda são escassos, especialmente em ambientes controlados, apesar de serem organismos
bastante populares na aquariofilia e aquários públicos. Adicionalmente, percebeu-se a
possibilidade de utilização das estrelas como controladores paliativos em aquários
marinhos, sobretudo Turbinella sp. e E. spinulosus por serem espécies “Reef safe” e não
apresentam perigo aos corais.

396
Belém (PA), 11 a 13 de dezembro de 2019
4. CONCLUSÃO
Conclui-se que, apesar de serem alimentadas com ração industrializada, as estrelas
do mar não perderam seu hábito alimentar natural de predadores de topo de cadeia.
Portanto, tanto a ração como o alimento vivo demonstraram ser alimentos em potencial
para os espécimes e não houve, estatisticamente, preferência alimentar por nenhuma das
duas fontes. Adicionalmente, este trabalho mostrou que estas estrelas são controladores
paliativos de Asterinas sp. o que torna viável aos aquaristas seu uso para manutenção de
aquários com recifes artificiais.

397
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BACELAR, Alberto José de Freitas. Aquários marinhos de recifes de corais. São
Paulo: Nbl Editora. 173 p. 1997.
RUPPERT, E. E.; BARNÉS, R. D. Zoologia dos Invertebrados. 6a. Edição. Roca, São Paulo, 1029p, 1996.
BOOLOTIAN, R.A. (In.) Physiology of Echinodermata. Nova Iorque: Interscience
Publishers. 822p. 1966.
BOS, Arthur R. et al. Population dynamics, reproduction and growth of the Indo-Pacific
horned sea star, Protoreaster nodosus (Echinodermata; Asteroidea). Marine Biology, v.
156, n. 1, p. 55-63, 2008.
BRUSCA, R. C. & BRUSCA, J. G. Invertebrados. 2a. Ed. Guanabara Koogan. 1098 p.
2007.
CARRACA, Susana Isabel Soares. Estágio no Oceanário de Lisboa Técnicas de
aquariofilia em aquários públicos. 2016.
DE BARROS LIMA, Eduardo José; FERNANDES, Múcio Luiz Banja. Diversidade de
equinodermos (Echinodermata) no Estado de Pernambuco (Brasil). Revista Brasileira
de Zoociências, v. 11, n. 1, 2009.
EMSON, R. H.; CRUMP, R. G. Description of a new species of Asterina (Asteroidea),
with an account of its ecology. Journal of the Marine Biological Association of the
United Kingdom, v. 59, n. 1, p. 77-94, 1979.
HICKMAN, Cleveland P.; ROBERTS, Larry S.; KEEN, Susan L. Princípios integrados
de zoologia. Grupo Gen-Guanabara Koogan, 2016.
HONEY-ESCANDÓN, M.; SOLÍS-MARÍN, F. A.; LAGUARDA-FIGUERAS, A.
Equinodermos (Echinodermata) del Pacífico Mexicano. Revista de Biología Tropical,
v. 56, n. 3, p. 57-73, 2008.
INDIVIGLIO, Frank. The Everything Aquarium Book: All You Need to Build the
Acquarium of Your Dreams. Simon and Schuster, 2006.
LIMA, Eduardo José de Barros. Dinâmica populacional de Echinometra lucunter
(Linnaeus, 1758) (Echinodermata: Echinoidea) na praia de Muro Alto, Ipojuca,
Pernambuco. 2006. Dissertação de Mestrado. Universidade Federal de Pernambuco.
MANSO, Cynthia Lara de Castro; ALVES, Orane Falcão de Souza; MARTINS, Luciana
Ribeiro. Echinoderms from Todos os Santos Bay and Aratu Bay (Bahia, Brazil). Biota
Neotropica, v. 8, n. 3, p. 0-0, 2008.
MORISHITA, Vanessa Rimoli. Resposta comportamentais do ouriço-do-mar preto a
pistas químicas que indicam risco de predação. 2010.
MOURA, Ronny Scheffler. Dinâmica da reproduçao e curva de crescimento de
Astropectem marginatus Gray, 1840 (Echinodermata: Asteroidea) da fauna
acompanhante dos arrastos de camaroes provenientes do Balneário de Shangri-lá, PR.
1999.

398
Belém (PA), 11 a 13 de dezembro de 2019
QUEIROZ, Camila Helena de Souza. Estratégias e fisiologia do consumo e digestão de
esponjas (Porifera) por Echinaster brasiliensis (Echinodermata: Asteroidea). 2012.
Dissertação (Mestrado em Fisiologia Geral) - Instituto de Biociências, University of São
Paulo, São Paulo, 2012. doi:10.11606/D.41.2012.tde-24102012-085504. Acesso em:
2019-10-28.
RIBEIRO-COSTA, C. S. (Org.); ROCHA, R.M. (Org.). Invertebrados: Manual de Aulas
Práticas. 2a. ed. Ribeirão Preto: Holos, Editora Ltda., v. 1. 271 p. 2006.
ROWE, F. W. E.; GATES, J. Echinodermata. Vol. 33 in A. Wells, ed. Zoological
catalogue of Australia. CSIRO Australia, Melbourne, 1995.
SAMPAIO, FLM. Distribuição espaço-temporal de mega equinodermos e reprodução
assexuada de Linckia guildingii (Echinodermata: Asteroidea) no platô do recife de Coroa
Vermelha, Bahia, Brasil. Ilhéus, BA, Dissertação de mestrado, Universidade Estadual
de Santa Cruz, 30p, 2010.
SCHEIBLING, Robert E.; METAXAS, Anna. Abundância, distribuição espacial e
estrutura de tamanho da estrela do mar Protoreaster nodosus em Palau, com notas sobre
alimentação e reprodução. Boletim de Ciência Marinha, v. 82, n. 2, p. 221-235, 2008.
STORER, T.I., USIN KUKENTHAL GER, R.L., STEBBINS, R.C., NYBAKKEN, J.W.
Zoologia geral. São Paulo: Companhia Editora Nacional, 816 p. 2005.
TAVARES, Y. A. G. et al. DINÂMICA POPULACIONAL DE Astropecten marginatus
(ECHINODERMATA: ASTEROIDEA) NO SUL DO BRASIL 2007.
WABNITZ C, TAYLOR M, GREEN E, RAZAK T From ocean to aquarium. UNEP-
WCMC, Cambridge, UK. 2003.
XAVIER, Luís Augusto Ramos. Inventário dos equinodermos do estado de Santa
Catarina, Brasil. Brazilian Journal of Aquatic Science and Technology, v. 14, n. 2, p.
73-78, 2010.

399
Belém (PA), 11 a 13 de dezembro de 2019
LEVANTAMENTO BIBLIOGRÁFICO SOBRE A MARRECA-CABOCLA
Dendrocygna autumnalis (ANSERIFORMES: ANATIDAE): TEMAS
ESTUDADOS E HISTÓRICO DOS TRABALHOS
Francisco Donnadone Reis Araújo 1; Sanae Nogueira Hayashi 2.
1 Acadêmico do Curso de Licenciatura em Biologia. Universidade Federal Rural da
Amazônia – UFRA. [email protected]
2 Dra. Eng. Florestal, Orientadora, Docente. Universidade Federal Rural da Amazônia–
UFRA. [email protected]
RESUMO
O objetivo desse trabalho foi analisar as publicações sobre marrecas-cabocla
(Dendrocygna autumnalis), através de um levantamento bibliográfico. Para isso foi
realizado buscas dos artigos produzidos em diversas partes do mundo referentes a
marrecas, extraídos de diversos bancos de dados do tipo: Google Acadêmico, Scielo, e
Portal CAPES. Com isso foi possível analisar o panorama dos temas estudados sobre a
ave em nível nacional e internacional. Foram encontrados 70 (setenta) artigos com
relevância a pesquisas feitas sobre D. autumnalis, realizados no período de 1964 a 2018.
Todos os trabalhos foram analisados e classificados de forma sistemática através de uma
abordagem quantitativa. Os resultados adquiridos mostram que a maior parte dos estudos
estão concentradas na área de ecologia. No entanto, também foram encontrados trabalhos
desenvolvidos na área da saúde, além de um único trabalho relacionado à história natural,
trazendo informações pertinentes sobre a espécie. Por fim, foram detectadas poucas
pesquisas sobre a ave aquática, ou seja, com uma quantidade baixa de artigos, haja vista
o fato de ser um levantamento de abrangência mundial.
Palavras-chave: Ave. Mundial. Dendrocygna autumnalis.
Área de Interesse do Simpósio: Zoologia

400
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A riqueza e diversidade de espécies de ecossistemas tropicais são bem evidentes,
e para realização de estudos nesse quesito, se faz necessária uma delimitação clara dos
objetivos e métodos a serem utilizados nessa produção, principalmente se forem usados
para preservação da biodiversidade em uma ampla escala (BISPO et al., 2016).
A ave aquática denominada Dendrocygna autumnalis tem distribuição ampla em
diversas partes do continente, estendendo-se desde o centro oeste dos EUA até o norte
argentino, a mesma reside em regiões baixas e úmidas, mas também podem ser
encontradas em locais com características diferentes (PETERS, 1979). No Brasil pela rica
biodiversidade e por sua variedade de espécies, a marreca cabocla (Dendrocygna
autumnalis) pode ser encontrada em todos os estados que componham o país, tendo uma
ocorrência bem ampla (PINTO, 1976).
De acordo com PIZZANI et al. (2012) a pesquisa bibliográfica, também chamada
de levantamento ou revisão bibliográfica está situada como a revisão de literaturas com
intuito de nortear o trabalho científico, uma revisão geral em vários tipos de materiais,
sendo eles: livros, periódicos, artigos, entre outras fontes. Dentro dos objetivos desse
estudo é citado o aprendizado em determinada área do conhecimento, facilitador de
métodos e técnicas que o pesquisador utilizará, e vários outros subsídios de revisão de
literatura e discussão de determinado trabalho de cunho científico.
Estando em um patamar de trabalho investigativo e fundamental facilitador de
pesquisa, justifica-se então em primeiro lugar como facilitador de trabalhos que venham
trazer dificuldades na localização, identificação e manejo de informações e dados
pertinentes de determinados trabalhos estando eles em inúmeras colocações (PIZZANI et
al., 2012).
Com relação aos resumos de artigos em bases de dados eletrônicas nacionais e
internacionais corroboram para obtenção de informações pertinentes dessa pesquisa para
elaboração do estudo. Visando os tipos de produção feitas na área e a quantidade de
trabalhos desenvolvidos, visando o estudo da arte como uma estratégia sistemática e de
grande importância na comunidade acadêmica (FERREIRA, 2002).
Para obter dados relevantes sobre trabalhos relacionados a marrecas
(Dendrocygna autumnalis), se faz extremamente necessária essa metodologia, levando

401
Belém (PA), 11 a 13 de dezembro de 2019
em conta toda exploração e revisões necessárias no quesito de pesquisas de matérias e um
levantamento organizado de todos os dados pertinentes ao estudo dessa determinada
espécie. Dessa forma o objetivo geral desse trabalho se estabeleceu em analisar os
diferentes trabalhos científicos sobre D. autumnalis através de um levantamento
bibliográfico ao redor do mundo utilizando os bancos de dados nacionais e internacionais
disponíveis para pesquisa.
2. METODOLOGIA
A justificativa relacionada com a necessidade da produção desse trabalho é a
ausência de levantamentos bibliográficos em trabalhos de cunho científico referentes a
marrecas (Dendrocygna autumnalis). Dessa forma, foi necessário um estudo relevante
com obtenção ampla de dados dos artigos sobre a ave aquática, tendo o intuito de
conhecer a demanda de trabalhos, locais de estudos e os tipos de pesquisas realizadas,
sendo uma ferramenta que possibilite um embasamento teórico vindo nortear facilitando
o desenvolver de futuros trabalhos científico.
A problemática se dá pela ausência de dados para compor registros de mais
informações sobre D. autumnalis, ou seja, com a ausência de registros acaba-se
dificultando pesquisas relacionadas à espécie, limitando determinadas investigações. Para
cobrir essa lacuna, foi realizada uma pesquisa em sites acadêmicos do tipo: Google
Acadêmico, Scielo, e Portal CAPES. Nesse levantamento foram extraídos e lidos diversos
artigos para a análise quantitativa sobre os diferentes tipos de trabalhos relacionados à
espécie, sistemáticamente com a busca escrita como “Dendrocygna autumnalis”, além
de avaliar a importância dos trabalhos sobre a espécie.
A abordagem metodológica utilizada foi a quantitativa, visando o resultado
numérico e também as considerações analisadas diante das perspectivas que
caracterizaram as áreas e os locais de onde os estudos foram realizados e notificados em
escritas, tabela e gráficos para melhor ilustrar as atribuições.
O método de investigação foi o levantamento bibliográfico visando uma coleta de
informações para compor um banco de dados sobre D. autumnalis, delimitando tempo e
sistematização para realização da pesquisa e geração dos resultados citados no presente
trabalho.

402
Belém (PA), 11 a 13 de dezembro de 2019
O instrumento de coleta para o trabalho foi o levantamento e busca de informações
do tipo: títulos dos trabalhos, autores e co-autores, revista onde o trabalho foi publicado
(periódicos), ano de publicação, local de estudo (País/Continente), área teórica, subtemas
e observações gerais de acordo com a apreciação e o entendimento da leitura dos trabalhos
analisados. Todas essas informações foram retiradas de banco de dados eletrônicos em
diversas plataformas digitais, corroborando para a base teórica e fundamental resultante
da pesquisa.
3. RESULTADOS E DISCUSSÃO
Na amostra foi identificado um total de 70 publicações de artigos referentes a
Dendrocygna autumnalis, das quais 41% eram produções nacionais (desenvolvidas no
Brasil) e 59% internacionais, desenvolvidas em diversos países espalhados pelo mundo.
Entre os diversos periódicos de publicação foram pesquisados e encontrados em um total
de 47, na qual alguns tinham mais de uma publicação. (Tabela 1).
Tabela 1 – Distribuição quantitativa de artigos sobre Dendrocygna autumnalis de acordo com o
periódico de publicação.
Periódicos Número de
artigos (n)
Amazôn., Rev. Antropol. (Online) 1
Atualidades Ornitológicas 4
AvianDiseases 1
BiologicalConservation 1
Biological Reviews 1
BioTemas 1
Bird ConservationInternational 1
Birds of North America, Incorporated 1
BrazilianJournal of Biology 2
CanadianJournal of Zoology 1
Ciencia Animal 2
Conservatyon e Society 2
Ecotoxicology 1
Florida Field Naturalist 1
Food and agriculture orgazatio of the united nations 1

403
Belém (PA), 11 a 13 de dezembro de 2019
Huitzil 2
Ibis 1
Instituto de Pesquisas Cientificas e Tecnológicas do Estado do
Amapá-IEPA 1
Instituto Nacional de Pesquisas da Amazônia 1
Interciencia 1
Jornal Zoológico do Brasil 1
JournalArticle 5
Journal of Comparative and Physiological Psychology 2
Journal of Ornithology 1
Journal of WildlifeDiseases 3
O TexmexNaturalist 1
Open Juornal System 1
Ornitologia 1
ParasitologyResearch 1
Regional Environmental Change 1
Revista Brasileira de Ornitologia 3
Revista Brasileira de Zoologia 1
Revista Científica do CEMAVE 1
Revista de Biología Tropical 3
Revista de Ornitologia Neotropical 2
Revista Mexicana de Biodiversidad 1
Revista Mexicana de Ornitologia 2
ScienceDirect 1
Springer InternationalPublishingSwitzerland 1
Texas A & I University 1
The AukOrnithologicalAdvances 1
The Cooper Ornithological Society 1
The Journal of Parasitology 1
The Royal Society 1
The Society of Environmental Toxicology and Chemistry 1
The SouthwesternNaturalist 3
Wildfowl 2
TOTAL 70
Fonte: ARAÚJO, F. D. R. (2019)

404
Belém (PA), 11 a 13 de dezembro de 2019
Entre as áreas de estudo foi possível encontrar trabalhos sobre a espécie em
ecologia, saúde e história natural, distribuídos por várias partes do mundo na condição
que a ave está presente em diferentes continentes. A maioria dos trabalhos estão
concentrados na área de ecologia, apresentando a maior porcentagem de produção, ou
seja, 84% do total 59 artigos. Em seguida, estão os trabalhos das áreas referentes à saúde
do organismo com 14%, totalizando 10 trabalhos. E por fim, foi encontrado somente um
trabalho na área de história natural.
Na área da ecologia, foram encontrados artigos sobre migração, reprodução,
monitoramento, adaptação da alimentação, desenvolvimento, comportamento, conflito
ambiental, preservação, rastreamento, registro de ocorrência, comparação de grupos,
relações ecológicas, estruturas físicas das aves, adaptação, monitoramento populacional,
análise sistemática, biogeografia, marcação da espécie, fatores socioeconômicos,
diversidade, monitoramento de caça, aves como bioindicadores, habitat de aves, ecologia
de aves, ocorrência de ninho e estudos estruturais sobre a ave aquática. Em relação a área
da saúde os artigos abordaram temos sobre implicações dos patos como possíveis
reservatórios de bactérias, biomonitoramento, bioindicadores de atividade viral, análise
laboratorial sobre vetores de doenças, estudo patológico, análise de conservação da
espécie, estudo imunológico, análise parasitológica e patológica. E, o único artigo na área
de história natural tratava de informações descrevendo as principais características do
organismo D. autumnalis.
Em relação ao local onde os trabalhos foram realizados, o levantando bibliográfico
identificou 10 países como área de estudo, porém em algumas situações os artigos
apresentaram somente a localização do continente e, em outros casos, a localização estava
ausente. Entre esses países, o Brasil foi responsável pela maioria das publicações (42%),
equivalente a 29 produções científicas, sendo a maioria representada por artigos recentes
entre os anos de 2016-2018. Em seguida, estão os EUA (21%) com 15 produções, e o
México (20%) com 14. O restante dos artigos analisados está representado pela Venezuela
(4%), África (2%), Argentina (2%), Austrália (2%), Colômbia (1%), Continente Europeu
(1%), Israel (1%), e Peru (1%) (Figura 01).

405
Belém (PA), 11 a 13 de dezembro de 2019
Figura 01 – Quantidade bruta de publicações por localidades sobre a ave Dendrocygna
autumnalis.
Fonte: ARAÚJO, F. D. R. (2019)
Em uma escala temporal de produção dos trabalhos analisados, foi possível
constatar publicações de 1964 a 2018. Em um gradiente bem equilibrado se destaca o ano
de 2010 com o maior número de publicação, totalizando 08 artigos dos 70 coletados. Os
demais anos oscilaram entre 07, 05, 04, 03, 02, e a maioria com 01 artigo no ano referente
a publicação (Figura 02).
Entre os trabalhos produzidos no ano de 2010, a maioria são do Brasil. Entretanto,
alguns desses artigos mostraram similaridades, ou seja, são produções complementares
uma da outra, abordando subtemas diferentes com temáticas semelhantes. Além disso, no
mesmo ano, tiveram produções do México e da Venezuela, e coincidentemente, todas as
08 produções situaram-se na área da ecologia.

406
Belém (PA), 11 a 13 de dezembro de 2019
Figura 02 – Ano da publicação e quantidade de artigos publicados anualmente referentes à ave
Dendrocygna autumnalis.
Fonte: ARAÚJO, F. D. R. (2019)
Um ponto positivo foi a predominância do grande número de produções recentes
de pesquisas sobre a espécie, trazendo informações pertinentes sobre a espécie e deixando
o banco de dados atualizado trazendo benefícios a nível de conhecimento e de produção
de futuras pesquisas.
4. CONSIDERAÇÕES FINAIS
Através deste levantamento bibliográfico referente aos trabalhos produzidos sobre
a ave aquática marreca cabocla (Dendrocygna autumnalis), foi possível identificar que a
maioria dos artigos estão concentrados na área de ecologia, envolvendo vários subtemas.
Também foi possível verificar que o fator de investigação da pesquisa forneceu
informações importantes e inéditas sobre os trabalhos analisados, mostrando a
importância da D. autumnalis e, através das publicações no decorrer dos anos.
A pesquisa foi um levantamento de abrangência mundial para uma maior
apreciação das informações adquiridas, na qual o Brasil foi responsável pelo maior
número de produções e contou com os artigos com anos de publicações mais recentes.
Apesar disso, dentro de uma perspectiva evocada deste trabalho, é notório que ainda se
faz necessário o desenvolvimento de mais estudos referentes a espécie D. autumnalis,
sejam nas áreas de ecologia, saúde ou de história natural.

407
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
PINTO, Olivério M. De Oliveira. NOVO CATÁLOGO DAS AVES DO BRASIL -
PRIMEIRA PARTE. Revista dos Tribunais: São Paulo. v. 1, p. 446. São Paulo, 1978.
PETERS, J. L. Check-Iist of birds of the world. Museum of Comparative Zoology
Cambridge. v. 1, n. 2. Cambridge, 1979.
PIZZANI L. et al. A ARTE DA PESQUISA BIBLIOGRÁFICA NA BUSCA DO
CONHECIMENTO. Revista digital de biblioteconomia na ciência da informação. v.10,
n.1, p.53-66, jul-dez. Campinas, 2012.
FERREIRA, N. S. A. (2002). As pesquisas denominadas “Estado da Arte”. Educação
&Sociedade. v. 79, p. 257-272. Campinas, 2002.
BISPO A. A. et al. Protocolo para Monitoramento de Comunidades de Aves em Unidades
de Conservação Federais: Instituto Chico Mendes de Conservação da Biodiversidade,
Parque Nacional Viruá, Boa Vista, 2016.

408
Belém (PA), 11 a 13 de dezembro de 2019
ANEXO I - Referências dos trabalhos originários das buscas feitas
PINTO, Olivério M. De Oliveira. NOVO CATÁLOGO DAS AVES DO BRASIL - PRIMEIRA PARTE. Revista dos
Tribunais: São Paulo. v. 1, p. 446, 1978., jan, 1974. PETERS, J. L. Check-Iist of birds of the world. Museum of Comparative
Zoology Cambridge. v. 1, n. 2. 1979. BISPO A. A. et al. Protocolo para Monitoramento de Comunidades de Aves em Unidades de Conservação Federais: Instituto Chico Mendes de Conservação da Biodiversidade, Parque Nacional Viruá, Boa Vista.
2016. RYLANDER, Michael Kent; BOLEN, Eric G. Feeding Adaptations in Whistling Ducks (Dendrocygna). JOURNAL
ARTICLE, v. 91, n. 1, p. 86-94 . AGUIRRE. A.A.; QUAN., T.J.; COOK, R.S. MCLEAN RG. Cloacal flora isolated from
wild black-bellied whistling ducks (Dendrocygna autumnalis) in Laguna La Nacha, Mexico. Avian Diseases, v. 36, n. 2, p.
62-459, apr-jun, 1992. OSTEN, J.R.; SOARES, A.M.; GUILHERMINO, L. Black-bellied whistling duck (Dendrocygna
autumnalis) brain cholinesterase characterization and diagnosis of anticholinesterase pesticide exposure in wild populations
from Mexico. Environ Toxicol Chem. v. 24, n. 2, p. 7-313, feb, 2005. BOLEN E. G.; MCDANIEL B.; C. COTTAM. Natural
History of the Black-Bellied Tree Duck (Dendrocygna autumnalis) in Southern Texas. JOURNAL ARTICLE. v. 9, n. 2, p. 78-88. may, 1964. MATTA N. E. et al. Description and molecular characterization of Haemoproteus macrovacuolatus n. sp.
(Haemosporida, Haemoproteidae), a morphologically unique blood parasite of black-bellied whistling duck (Dendrocygna
autumnalis) from South America. Parasitol Res. v. 8, doi: 10.1007/s00436-014-3961-2, p. 2991-3000. aug, 2014. AGUIRRE,
A. Alonso; MCLEAN, Robert; COOK, Robert S. Experimental inoculation of three arboviruses in black-bellied whistling
ducks (Dendrocygna autumnalis). Journal of wildlife diseases. v. 4, doi: 10.7589/0090-3558-28.4.521, p. 5-521. nov, 1992.
RODRIGUES, Monique Rocha; SCHMITZ, Heribert. Conflito Social: O Caso da “Pesca de Marrecas” na Reserva Extrativista
Marinha de Tracuateua (PA). Amazôn., Rev. Antropol. v. 9, n. 1. P. 132-194, 2017. ESTABROOKS S.R. Ingested lead shot
in northern red-billed whistling ducks (Dendrocygna autumnalis) and northern pintails (Anas acuta) in Sinaloa, Mexico. J Wildl Dis. v. 23. n. 1. p. 169. jan, 1987. HERSLOFF L.; LEHNER P.N.; BOLEN E.G.; RYLANDER M.K. Visual sensitivity
in the black-bellied tree duck (Dendrocygna autumnalis), a crepuscular species. J Comp Physiol Psychol. v. 86, n. 3, p. 92-
486. mar 1974. QUEZADA, David Colón. Presencia de patos pijije, alablanca (Dendrocygna autumnalis) y canelo
(Dendrocygna bicolor), en Toluca, Estado de México. Huitzil. Revista Mexicana de Ornitología. v. 5. n. 1. p. 1-2. jun, 2004.
CASTRO, Gonzalo; STOYAN, Nina; MYERS, J.P. Assimilation efficiency in birds: A function of taxon or food type?
ScienceDirect. v. 92, n. 3, p. 271-278. 1989. RYLANDER M. Kent; BOLEN Eric G. Analysis and Comparison of Gaits in
Whistling Ducks (Dendrocygna). The Wilson Journal of Ornithology. v. 86, n. 3, p. 237-245. sep, 1974. GUADAGNIN, D.;
DOTTO, J. C.; BURGER, M. I. Ocorrencia da Marreca Cabocla Dendrocygna autumnalis no noroeste do Rio Grande do Sul, Brasil. Hornero. Revista de Ornitología Neotropical. v. 14, n. 01y02, p. 74-75. jul, 1995. JAMES, J. D.; THOMPSON, J. E.
Black-bellied Whistling-Duck (Dendrocygna autumnalis). In The Birds of North America (A. F. Poole and F. B. Gill, Editors).
v. 2, n. 9, p. 78-88. jan, 2001. CAIN, Brian W. Energetics of Growth for Black-Bellied Tree Ducks. The Condor. v. 78, n. 1,
p. 124-128. spr, 1976. DECHELLE. M. Black-bellied Whistling Duck - Dendrocygna autumnalis. Geographic range by
countries - Department of Guiana. v. 43, n. 1. mar, 2006. ERIC, G. BOLEN; KENTRYLANDER, M. Foot adaptations in four
species of Whistling Duck Dendrocygna. Wildfowl. p. 81-83. 1974. SIEGFRIED, W. Roy. Morphology and Ecology of the
Southern African Whistling Ducks (Dendrocygna). The Auk. v. 90, n. 1, p. 198-201 jan, 1973. AGUIRRE A. A.; MCLEAN R.G.; COOK R.S.; QUAN T. J. Serologic survey for selected arboviruses and other potential pathogens in wildlife from
Mexico. J Wildl Dis. v. 28, n. 3. p. 42-435. jul. 1992. EDMONDS, Samuel T.; STOLLEY, Dorie S. Population Decline of
Ground-Nesting Black-Bellied Whistling Ducks (Dendrocygna autumnalis) on Islands in Southern Texas. The Southwestern
Naturalist. v. 53, n. 2, p. 185-189. jun, 2008. WELLS, M. C.; LEHNER, P. N.; BOLEN, E.G.; RYLANDER, M. K.
Comparison of scotopic sensitivity in diurnal (Anas platyrhynchos) and crepuscular (Dendrocygna autumnalis) ducks. v. 88,
n. 2, p. 4-940. feb, 1975. STOLLEY, Dorie S.; EDMONDS, Samuel T.; METEYER, Carol U. Mortality of Ducklings of the
Black-Bellied Whistling Duck (Dendrocygna autumnalis) During Their Premier Swim in a Hypersaline Lake in South Texas.
The Southwestern Naturalist. v. 53, n. 2, p. 230-235. jun, 2008. FEDYNICH, Alan M.; PENCE, Danny B.; BERGAN, James F. Helminth community structure and pattern in sympatric populations of black-bellied and fulvous whistling-ducks. Canadian
Journal of Zoology. v. 74, n. 12, p. 2219-2225. 1996. FERREYRA, H.; BELDOMENICO, P. M.; MARCHESE, K.;
ROMANO, M.; CASELLI, A.; CORREA, A. I.; UHART, M. Lead exposure affects health indices in free-ranging ducks in
Argentina. Ecotoxicology. v. 24, n. 4, p. 45-735. may, 2015. LÓPEZ, Rubén Pineda; SANAPHRE, Alejandro Arellano.
Noteworthy records of aquatic birds in the state of Querétaro, Mexico. Huitzil Revista de Ornitología Mexicana. v. 11, n. 2,
p. 49-59. dec, 2010. COOKE, Richard G.; OLSON, Storrs L. An Archaeological Record for the White-Faced Whistling-Duck
(Dendrocygna viduata) in Central Panama. The Condor. v. 86, n. 4, p. 493-494. nov, 1984. CARMONA, Roberto; AYALA-
PÉREZ, Víctor; AGUILAR, Antonio Gutiérrez. Registros nuevos y sobresalientes de anátidos en humedales artificiales de Baja California Sur, México. REVISTA MEXICANA DE BIODIVERSIDAD. v. 82. n. 2, p. 727-730. june, 2011. BANKS,
Richard C. NOMENCLATURE OF THE BLACK-BELLIED WHISTLING-DUCK. The Auk. v. 95, p. 348-352. april, 1978.
OLSON, Storrs L. An Evaluation of the Supposed Anhinga of Mauritius. The Auk. v. 92, n. 2, p. 374-376. apr, 1975. YOM-
TOV, Yoram. Intraspecific nest parasitism in birds. Biological Reviews. v. 55, n. 1, p. 93-108. jan, 2008. BRUZUAL, Jose;
BRUZUAL, Iris. Feeding habits of whistling ducks in the Calabozo Ricefields, Venezuela, during the non-reproductive period.
Wildfowl. v. 34, p. 20-26. 1983. SEBAIO, Fabiane. Et al. Blood parasites in passerine birds from the Brazilian Atlantic Forest.
Rev. Bras. Parasitol. Vet. v. 21, n.1. mar, 2012. MATSON K. D.; COHEN A. A.; KLASING K.C.; RICKLEFS R.E.;
SCHEUERLEIN A. No simple answers for ecological immunology: relationships among immune indices at the individual level break down at the species level in waterfowl. Proc Biol Sci. v. 273, n. 7, p. 22-815. apr, 2006.YOM‐TOV, Yoram. An
updated list and some comments on the occurrence of intraspecific nest parasitism in birds. International Journal of Avion
Science. v. 143, n. 1, p. 133-143. june. 2008. PEREIRA, Washington Luiz Assunção. Et al. LESÕES CUTÂNEAS TIPO
TUMORAIS ASSOCIADAS À INFEÇÃO POR AVIPOXVIRUS EM UMA MARRECA-CABOCLA (Dendrocygna
autumnalis). Cienc. anim. v.15, n.2, p. 234-238, abr./jun, 2014. FEEKES, Francisca. The black-bellied whistling duck in
Mexico—from traditional use to sustainable management? ScienseDirect. v. 56, n. 2, p. 123-131. 1991. BERGSTROM,
Bradley J. FIRST REPORTED BREEDING OF BLACK-BELLIED WHISTLING-DUCK IN NORTHERN FLORIDA.

409
Belém (PA), 11 a 13 de dezembro de 2019
Florida Field Naturalist. v. 27, n. 4, p. 177-179. 1999. MELLINK, Eric; RIOJAS-LÓPEZ, Mónica E. Waterbirds (other than
Laridae) nesting in the middle section of Laguna Cuyutlán, Colima, México. Revista de Biología Tropical. v. 56, n. 1, p. 391-
397. mar, 2008. MELLINK, Eric; RIOJAS-LÓPEZ, Mónica. Waterbirds and human-related threats to their conservation in Laguna Cuyutlán, Colima, México. Rev. biol. trop. v. 57 n. 1-2, p. 1-12. mar./jun. 2009. VILELLA, Francisco J. Abundance
and Distribution of Waterbirds in the Llanos of Venezuela. The Wilson Journal of Ornithology. v. 122, DOI: 10.1676/09-
070.1, p. 102-115. feb, 2010. HARVEY, Michael G. The avian biogeography of an Amazonian headwater: the Upper Ucayali
River, Peru. The Wilson J. of Ornithology. v. 126, n. 2, p. 179-191. jun, 2014. PIÑA-ORTIZ, Alberto; CASTILLO-
GUERRERO, José Alfredo; ROCHA-ARMENTA, Jesús Alfonso; GUEVARA-MEDINA, Miguel Ángel. Presencia y
abundancia de aves que se reproducen en islas de la bahía de Mazatlán, Sinaloa, México. Huitzil. v. 18 n .1, p. 87-101. jun,
2017. GONZÁLEZ-MARÍN, Rosa María; MORENO-CASASOLA, Patricia; CASTRO-LUNA, Alejandro Antonio;
CASTILLO, Alicia. Regaining the traditional use of wildlife in wetlands on the coastal plain of Veracruz, Mexico: ensuring food security in the face of global climate change. Regional Environmental Change. v. 17, n. 5, p. 1343-1354. jun, 2017.
ANTAS, Paulo de Tarso Zuquim. Anilhamento de Dendrocygna spp. no Brasil entre 1973 e 1994 (Anseriformes: Anatidae).
Revista Brasileira de Ornitologia. v. 3, n. 62 , p. 62-65. abr, 1995. MELO-JÚNIOR, Tadeu A.; VASCONCELOS, Marcelo F.
de; FERNANDES, Geraldo W.; MARINI, Miguel. Bird species distribution and conservation in Serra do Cipó, Minas Gerais,
Brazil. Bird Conservation International, v. 11, n. 3, p. 189-204. nov, 2001. RODRIGUES, Marcos; MICHELIN, Vania B.
Riqueza e diversidade de aves aquáticas de uma lagoa natural no sudeste do Brasil. Rev. Bras. Zool. v. 22, n .4, p. 928-935.
dez, 2005.PIVATTO, Maria Antonietta Castro. et all. AVES DO PLANALTO DA BODOQUENA, ESTADO DO MATO
GROSSO DO SUL (BRASIL). Atualidades Ornitológicas. v. 1, n. 129, p. 1-26. fev, 2006. OLIVEIRA, Dalci Mauricio Miranda De. Efeitos bióticos e abióticos de ambientes alagáveis nas assembléias de aves aquáticas e piscívoras no Pantanal,
Brasil. Instituto Nacional de Pesquisas da Amazônia. v. 1, n. s/n, p. 1-198. jan, 2006. LOPES, Leonardo; BRAZ, Vivian.
Aves da região de Pedro Afonso, Tocantins, Brasil. Braz Revista Brasileira de Ornitologia. v. 15, n. 4, p. 530-537. dez, 2007.
AGUIAR, Kurazo Mateus Okada; NAIFF, Rafael Homobono; XAVIER, Bruno. Aves da Reserva Biológica do Lago Piratuba,
Amapá, Brasil. Revista Científica do CEMAVE. v. 4, n. 1, p. 1-14. dez, 2010. AGUIAR, Kurazo Mateus Okada; NAIFF,
Rafael Homobono. Composição da avifauna da Área de Proteção Ambiental do Rio Curiaú, Macapá, Amapá, Brasil. Revista
Científica do CEMAVE. v. 4, n. 1, p. 1-13. dez, 2010. NUNES, Alessandro Pacheco. et all. Aves da Estrada Parque Pantanal,
Corumbá, Mato Grosso do Sul, Brasil. Atualidades Ornitológicas. v. 156, n. s/n, p. 33-47. ago, 2010. GOMES, Carla Raphaela Gonzaga; EPIFÂNIO, Ariana Dias; VASCONCELOS, Marcelo Ferreira de. Estudo etnoornitológico no município de
Corumbá, Mato Grosso do Sul, Brasil. Atualidades Ornitológicas. v. 158, n. s-n, p. 49-54. dez, 2010. ALHO, C. J. R.
Biodiversity of the Pantanal: its magnitude, human occupation, environmental threats and challenges for conservation. Braz.
J. Biol. v.71 n.1, p. 1-4. apr, 2011.SANJAD, Nelson et al. Documentos para a história do mais antigo jardim zoológico do
Brasil: o parque zoobotânico do Museu Goeldi. Bol. Mus. Para. Emílio Goeldi. Ciênc. hum. v.7, n.1, p.197-258. abr, 2012.
REGISTROS FOTOGRÁFICOS, (divérsos fotográfos). Registros reprodutivos da asa-branca, Dendrocygna autumnalis, no
estado do Rio de Janeiro, Brasil. Atualidades Ornitológicas. v. 1, n. 169 , p. 1. oct, 2012. JUNIOR, Miguel Benedito Ferreira Dias; CUNHA, Helenilza Ferreira Albuquerque; DIAS, Teresa Cristina Albuquerque de Castro. CARACTERIZAÇÃO DAS
APREENSÕES DE FAUNA SILVESTRE NO ESTADO DO AMAPÁ, AMAZÔNIA ORIENTAL, BRASIL. Biota
Amazônia. v. 4, n. 1, p. 65-73. mar, 2014. TAVARES, Davi Castro; SICILIANO, Salvatore. Variação temporal na abundância
de espécies de aves aquáticas em uma lagoa costeira do Norte Fluminense, sudeste do Brasil. Biotemas. v. 27, n. 1, p. 121-
132. mar, 2014. DONATELLI, R. J.; POSSO, S. R.; TOLEDO, M. C. B. Distribution, composition and seasonality of aquatic
birds in the Nhecolândia sub-region of South Pantanal, Brazil. Braz. J. Biol. v. 74, n. 4, p. 844-853. nov, 2014. CINTRA,
Renato. Spatial distribution and composition of waterbirds in relation to limnological conditions in the Amazon basin.
Hydrobiologia. v. 747, n. 1, p. 235-252. mar, 2015. PIACENTINI, Vítor de Q. et al. Annotated checklist of the birds of Brazil by the Brazilian Ornithological Records Committee. Revista Brasileira de Ornitologia. v. 23, n. 2, p. 91-298. jun, 2015.
VIEIRA, Marina Albuquerque Regina de Mattos; MUHLEN, Eduardo Matheus von, SHEPARD, Glenn H. Participatory
Monitoring and Management of Subsistence Hunting in the Piagaçu-Purus Reserve, Brazil. Conservat Soc. v. 12, n. 3, p. 254-
264. nov, 2015. LOPES, Leonardo Esteves; MARÇAL, Bráulio de Freitas. Avifauna do Campus Florestal da Universidade
Federal de Viçosa, Minas Gerais, Brasil. Atualidades Ornitológicas. v. 193, n. 41, p. 41-56. out, 2016. DONATELLI,
Reginaldo José et al. Temporal and spatial variation of richness and abundance of the community of birds in the Pantanal
wetlands of Nhecolândia(Mato Grosso do Sul, Brazil). Rev. biol. Trop. v.65, n.4, p.1358-1380. dec, 2017. SOARES, Vanessa
Moura dos Santos; SOARES, Hyago Keslley de Lucena ; LUCENA, Reinaldo Farias Paiva de; BARBOZA, Raynner Rilke Duarte. CONHECIMENTO, USO ALIMENTAR E CONSERVAÇÃO DA AVIFAUNA CINEGÉTICA: ESTUDO DE
CASO NO MUNICÍPIO DE PATOS, PARAÍBA, BRASIL. Interciencia. v. 43, n. 7, p. 491-497. jun, 2018. AGUIAR, Kurazo
Mateus Okada; NAIFF, Rafael Homobono; XAVIER, Bruno. Aves da Reserva Biológica do Lago Piratuba, Amapá, Brasil.
Ornithologia. v. 4, n. 1, p. 1-14. dez, 2010. JOHNSGARD, Paul A. The North American Whistling-Ducks, Pochards, and
Stifftails. Zca Books Lincoln, Nebrasca. v. 54, n. 1, p. 1-184. mar, 2017. JOHNSGARD, Paul. "The North American Perching
and Dabbling Ducks: Their Biology and Behavior". Zca Books Lincoln, Nebrasca. v. 53, n. 1, p. 1-228. jan, 2017.

410
Belém (PA), 11 a 13 de dezembro de 2019
PRIMEIRO REGISTRO DO MOLUSCO Polymesoda caroliniana (BOSC, 1801)
(BIVALVIA: CYRENIDAE) NO LITORAL BRASILEIRO
Josiney Farias de Araújo1; Weverton John Pinheiro dos Santos2; Rafael Anaisce das
Chagas3; Mara Rubia Ferreira Barros4; Andrea Magalhães Bezerra5
1 Mestrando em Ecologia (PPGEco). Universidade Federal do Pará. E-mail:
[email protected] 2 Mestre em Aquicultura e Recursos Aquáticos Tropicais. Universidade Federal Rural
da Amazônia. E-mail: [email protected] 3 Doutorando em Ecologia (PPGEco). Universidade Federal do Pará. E-mail:
[email protected] 4 Mestra em Aquicultura e Recursos Aquáticos Tropicais. Universidade Federal Rural
da Amazônia. E-mail: [email protected] 5 Doutora em Saúde e Produção Animal na Amazônia. Universidade Federal Rural da
Amazônia. E-mail: [email protected]
RESUMO
O bivalve Polymesoda caroliniana é um molusco que habita águas costeiras, sendo
encontrado sobre sedimentos lamosos ou arenosos, possuindo distribuição da América do
Norte à América Central. Porém, comumente organismos são transportados de um
continente para o outro através da água de lastro e bioincrustações em navios e, dentre
estes, os moluscos são os que apresentam maiores índices de introdução em outros
ambientes. O presente estudo objetivou relatar a ocorrência de P. caroliniana em área de
manguezal no município de Primavera – PA. Em setembro de 2019, realizou-se coletas
de bivalves em quatro mangues localizados em Primavera, nordeste paraense, onde
delimitou-se um quadrante de 1m2 para retirada do substrato a uma profundidade de 5cm,
as amostras foram peneiradas em malha de 500µm, armazenados em recipientes plásticos
e fixadas em etanol 70 %. Posteriormente, foram transportadas ao Museu de Zoologia da
UFRA para análise. Após identificação morfológica, constatou-se a presença do bivalve
P. caroliniana nas coletas, o qual compunha 65,8% da diversidade local de moluscos
bivalves. Tendo em visto o que se sabe sobre a biogeografia de P. caroliniana, este
achado contribui com conhecimento sobre a sua distribuição, além de ser o primeiro
registro de ocorrência desta espécie no litoral brasileiro.
Palavras-chave: Bivalve. Amêijoa da Carolina. Mangue.
Área de Interesse do Simpósio: Zoologia.

411
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Espécies de animais que ocorrem em áreas fora do seu limite natural e que
possuem potencial de distribuição nessa nova área são denominadas de exóticas,
introduzidas ou não-nativas. Quando ocasionam algum dano socioeconômico e
ambiental, ameaçando a diversidade biológica nativa, ou para a saúde humana, são
consideradas invasoras (IUCN, 2000; MEDEIROS; NAHUZ, 2006). De acordo com
Souza et al. (2003), a introdução destas espécies em distintas áreas é algo que ocorre
desde a expansão europeia, por meio da água de lastro e bioincrustantes (FERREIRA et
al., 2004, HEWITT; CAMPBELL, 2007). Neste contexto, em razão da enorme área
costeira, o Brasil é considerado um dos países potencialmente mais afetados (ZANELLA,
2015).
Dentre os organismos invasores, os moluscos são os que mais são introduzidos
mundialmente, com destaque para os bivalves marinhos que possuem uma alta dispersão
pelos oceanos, sendo transportados de forma intencional (e.g. aquariofilia) e não-
intencional (e.g. água de lastro) (CARLTON & GELLER, 1993; STEFFANI; BRANCH,
2005; ZANELLA, 2015)
O bivalve Polymesoda caroliniana (Bosc, 1801) é um molusco pertencente a
família Cyrenidae. Habita águas costeiras, encontrado sobre sedimentos lamosos ou
arenosos. As características das conchas de P. caroliniana são, presença de superfície
lisa, contorno subtriangular arredondada, dobradiça com três dentes cardinais situados
sob umbo, coloração externa da concha pode ser branca e a interna, raramente, branca
manchada de roxo e perióstraco marrom brilhante, alcançando o tamanho máximo de
40mm do comprimento anteroposterior (ABBOTT, DANCE, 2000; ABBOTT, MORRIS,
1995; CERVIGÓN et al., 1992; LEAL, 2002; ROSENBERG, 2009).
P. caroliniana apresenta distribuição delimitada ao longo da costa atlântica da
Virgínia, norte da Carolina do Norte e da Flórida (nos EUA), com relatos de ocorrência
em Carolina do Sul e em Georgia, passando pelo Texas até a região do extremo sul do
México, aparentemente, estando restrita as regiões ao longo da costa norte do Golfo do
México (ANDREWS, 1951; LEAL, 2002; MORTON, 1983; SCHALIE, 1933;
WAKIDA-KUSUNOKI; MACKENZIE JR., 2004; WILLIAMS et al., 2014).

412
Belém (PA), 11 a 13 de dezembro de 2019
O presente trabalho teve como objetivo relatar a primeira ocorrência do molusco
bivalve P. caroliniana em área de manguezal situada no município de Primavera – Pará,
Brasil, e desta forma contribuir com as informações à cerca de extensão de sua
biogeografia.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
O município de Primavera, pertence à Mesorregião Nordeste Paraense e à
Microrregião Bragantina está distante cerca de 190 km da capital do estado, Belém (Fig.
1). Apresenta um clima tropical quente e úmido, com altos índices pluviométricos com
variações de precipitação ao longo do ano (MORAES et al., 2005). Este município e
formado por diversas comunidades, como Vila da Telha, do Laranjal, do Bacabal (Boa
Vistinha), da Doca, Galpão e Paca, entre outras, onde a principal atividade exercida por
seus moradores consiste na pesca artesanal, na agricultura, pequenos cultivos e no
extrativismo de caranguejos e mariscos (FADESP, 2016).
Figura 1 – Localização da área de estudo: Município de Primavera-PA.
Fonte: Elaborado pelos autores.

413
Belém (PA), 11 a 13 de dezembro de 2019
2.2 COLETA DE DADOS
P. caroliniana foi coletado durante amostragens em quatro manguezais, em marés
de quadratura, entre os dias 18 e 21 de setembro de 2019. Para a amostragem utilizou-se
quadrados de um 1m2 de área. Em cada quadrado, realizou-se a raspagem da 5cm
superfície do sedimento. Posteriormente, o sedimento coletado foi peneirado em malha
de 500µm, armazenado/codificando/fixando os moluscos encontrados em recipientes
plásticos contendo etanol a 70 %, transportando-os ao Museu de Zoologia da
Universidade Federal Rural da Amazônia para análises posteriores.
Em laboratório, identificou-se P. caroliniana de acordo com os caracteres
morfológicos segundo descrição da concha por Leal (2002) e bibliografias específicas
para o gênero (ABBOTT, DANCE, 2000; ABBOTT, MORRIS, 1995; CERVIGÓN et al., 1992).
Após isso, os espécimes foram depositados no Museu de Zoologia da UFRA (MZUFRA),
sob os códigos: Moll930; Moll936; Moll946 e Moll948.
2.3 ANÁLISE DE DADOS
Para uma análise preliminar, estimou-se a densidade de P. caroliniana em cada
local amostrado, determinando a morfometria externa (comprimento antero-posterior,
largura e altura) dos indivíduos encontrados.
3. RESULTADOS E DISCUSSÃO
Ao total, encontrou-se 142 P. caroliniana (Fig. 2A), com comprimento médio da
concha de 5,5±3,3mm, variando de 1,3 a 21,2mm. Ressalta-se que P. caroliniana foi
65,8% da diversidade local de moluscos bivalves.
Encontrou-se P. caroliniana nos quatro locais de amostragens (Fig. 2B), sendo
51% encontrado no Mangue A, seguido de 28% no Mangue B, 11% no Mangue D e 10%
no Mangue C (Fig. 3).
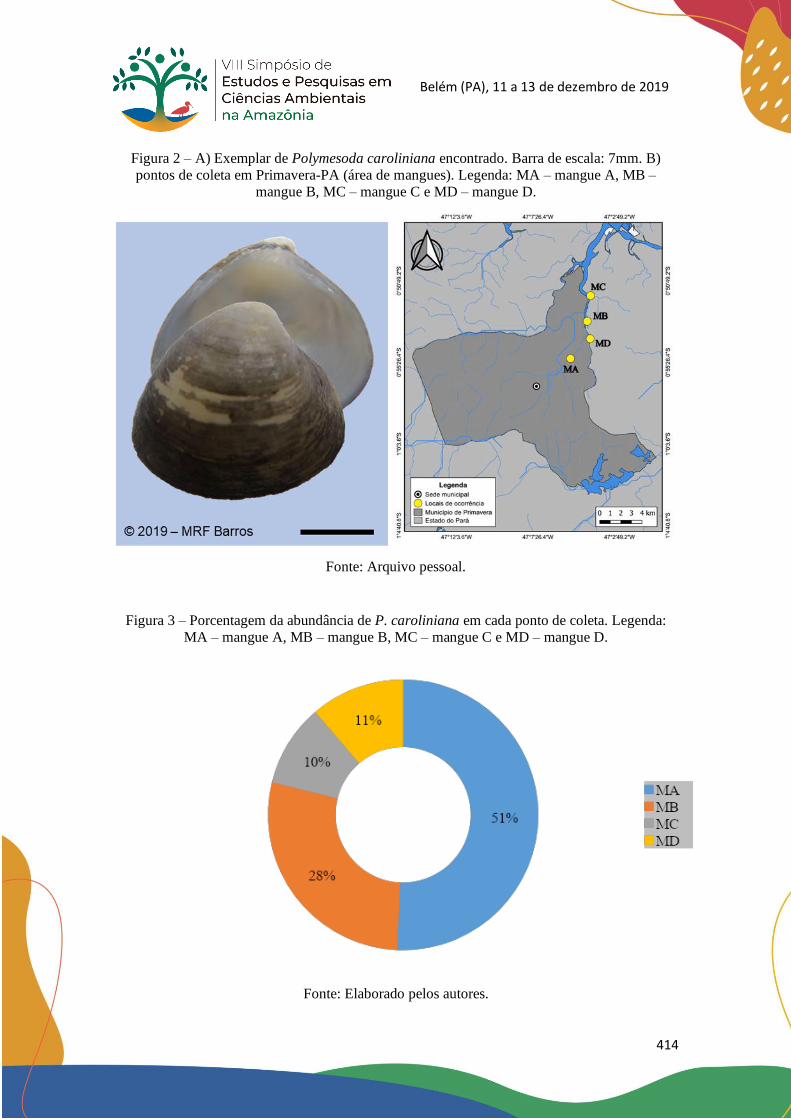
414
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2 – A) Exemplar de Polymesoda caroliniana encontrado. Barra de escala: 7mm. B)
pontos de coleta em Primavera-PA (área de mangues). Legenda: MA – mangue A, MB –
mangue B, MC – mangue C e MD – mangue D.
Fonte: Arquivo pessoal.
Figura 3 – Porcentagem da abundância de P. caroliniana em cada ponto de coleta. Legenda:
MA – mangue A, MB – mangue B, MC – mangue C e MD – mangue D.
Fonte: Elaborado pelos autores.

415
Belém (PA), 11 a 13 de dezembro de 2019
Na América do Sul, a família Cyrenidae é representada pelos gêneros
Polymesoda, Cyanocyclas e Corbicula, sendo este último uma espécie límnica
considerada invasora e não apresenta sinus palial (PEREIRA et al., 2014). As espécies
do gênero Polymesoda ocorrem em águas salobras no norte do continente, como
Polymesoda artata (Deshayes, 1854) (Sul do caribe ao Norte da América do Sul) e
Polymesoda triangulata a (Philippi, 1849) (Caribe mexicano ao Panamá) (LEAL, 2002).
Apesar destas espécies possuírem características em comum as evidenciadas para
P. caroliniana, há certas características que as diferenciam, como apesenta escultura de
fios concêntricos bem definidos, dentes laterais lisos e perióstraco com escalas
minúsculas em P. artata; concha com contorno triangular e perióstraco suave em P.
triangulata (LEAL, 2002).
No Brasil, há poucos relatos de Polymesoda spp., Noelli et al. (2011), que relatou
sua ocorrência após estudo zooarqueológico do lastro de embarcação naufragada na Praia
dos Ingleses, Ilha de Santa Catarina. Schroeder et al. (2014), que constatou a presença de
P. triangulata após coleta em pesca com emalhe de fundo nas regiões sudeste/sul.
Visto que não há nenhum relato da presença da espécie P. caroliniana em estados
brasileiros e que sua biogeografia se restringe à algumas regiões do norte da América
Central e sul da América do Norte, este é o primeiro registro de ocorrência desta espécie
no Brasil e diante disto, possivelmente, há uma extensão na distribuição da espécie no
Atlântico.
4. CONSIDERAÇÕES FINAIS
O registro do bivalve P. caroliniana nos mangues do município de Primavera-PA,
contribui ao conhecimento acerca de sua biogeografia, estendendo-se atualmente até o
norte do litoral brasileiro. Ressalta que, sendo este registro de P. caroliniana é de grande
relevância, uma vez que esta espécie apresenta importância comercial em regiões do
México, necessitando-se de estudos acerca da dinâmica população sob influência
latitudinal, já que sua distribuição geográfica foi ampliada com este estudo.

416
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ABBOTT, R. T.; DANCE, S. P. Compendium of Seashells. California: Odyssey Pub,
411p., 2000.
ABBOTT, R.T.; MORRIS, P.A. A field guide to shells: Atlantic and Gulf Coasts and
the West Indies. Houghton Mifflin Harcourt, 424p., 1995.
https://www.researchgate.net/publication/48377205_A_Field_Guide_to_Shells_Atlantic
_and_Gulf_Coasts_and_the_West_Indies
ANDREWS, J. Range and Habitat of the Clam Polymesoda Caroliniana (Bosc) in
Virginia (Family Cycladidae). Ecology, v. 32, n. 4, p. 758-760, 1951.
doi:10.2307/1932745
CARLTON, J. T.; GELLER, J. B. Ecological Roulette: The global transport of
nonindigenous organisms. Science, v. 261, p. 78-82, 1993.
CERVIGÓN, F.; CIPRIANI, R.; FISCHER, W.; GARIBALDI, L.; HENDRICKX, M.;
LEMUS, A. J.; MÁRQUEZ, R.; POUTIERS, J. M.; ROBAINA, G.; RODRIGUEZ, B.
Guia de campo de las especies comerciales marinas y de agua salobres de la costa
septentrional de sur america. Roma: FAO, 577p., 1992.
FADESP (Fundação ao Amparo e Desenvolvimento de Pesquisa). Diagnóstico e
caracterização socioambiental das áreas propostas para criação e ampliação de Reservas
Extrativistas na Mesorregião do Nordeste Paraense no Estado do Pará - PRODUTO 4:
Relatório com diagnóstico socioambiental referente a proposta de criação da Resex
Rio Primavera. Manguezais do Brasil, Belém, 2016. 99p.
FERREIRA, C. E. L.; GONÇALVES, J. E. A.; COUTINHO, R. Cascos de navios e
plataformas como vetores na introdução de espécies exóticas. p. 143-156. In: SILVA, J.
S. V.; SOUZA, R. C. C. L. (orgs.). Água de lastro e bioinvasão, Interciência, Rio de
Janeiro, 2004. 224p.
HEWITT, C. L.; CAMPBELL, M.L. Mechanisms for the prevention of marine
bioinvasions for better biosecurity. Marine Pollution Bulletin, v. 55, p. 395-401, 2007.
IUCN (The World Conservation Union). IUCN Guidelines for the Prevention of
Biodiversity Loss Caused by Alien Invasive Species. 2000. 21p.
https://portals.iucn.org/library/efiles/documents/Rep-2000-052.pdf
LEAL, J. H. Bivalves. In: CARPENTER, K. E. The Living Marine Resources of the
Western Central Atlantic. FAO: Rome, 2002. p. 25-98. http://www.fao.org/3/a-
i5712e.pdf
MEDEIROS, D. S.; NAHUZ, M. A. R. Avaliação de risco da introdução de espécies
marinhas exóticas por meio de água de lastro no terminal portuário de Ponta Ubu (ES).
Interfacehs. Revista de gestão integrada em saúde do trabalho e meio ambiente.

417
Belém (PA), 11 a 13 de dezembro de 2019
SENAC: Santo Amaro. 2006. http://www3.sp.senac.br/hotsites/blogs/InterfacEHS/wp-
content/uploads/2013/07/2006-v2-Inte-1.pdf
NOELLI, F.S.; MONTEIRO, P.C.; VIANA, A. Praia dos Ingleses 1: Arqueologia
subaquática na Ilha de Santa Catarina, Brasil (Parte 2). Revista Do Museu De
Arqueologia E Etnologia, São Paulo, n. 21, p. 293-314, 2011.
MORAES, B. C. D.; COSTA, J. M. N. D.; COSTA, A. C. L. D.; COSTA, M. H. Variação
espacial e temporal da precipitação no estado do Pará. Acta Amazonica. v. 35, n. 2,
p. 207-214, 2005.
MORTON, B. Mangrove Bivalves, In: RUSSEL-HUNTER, W. D. The Mollusca, Vol.
6. Ecology: Academic Press,1983. p. 77-138.
PEREIRA, D.; MANSUR, M. C. D.; PIMPÃO, D. M. Identificação e diferenciação dos
bivalves límnicos invasores dos demais bivalves nativos do Brasil. Cap.5, p 75-94. In:
MANSUR, M. C. D.; SANTOS, C. P.; PEREIRA, D.; PAZ, I. C. P; ZURITA, M.L.L.;
RODRIGUEZ, M. T. R. NEHRKE, M. V.; BERGONCI, P. E. A. Moluscos límnicos
invasores no Brasil: biologia, prevenção e controle. Porto Alegre: Editora Redes, Cap.
5, 2014. 412p.
ROSENBERG, G. Malacolog 4.1.1: A Database of Western Atlantic Marine
Mollusca. [WWW database (version 4.1.1)]. 2009. http://www.malacolog.org/
SCHALIE, H. V. Notes on the brackish water bivalve Polymesoda caroliniana (Bosc).
Occasional Papers of the Museum of Zoology, v. 258, p. 1-8, 1933.
https://deepblue.lib.umich.edu/bitstream/handle/2027.42/56697/OP258.pdf?sequence=1
&isAllowed=y
SCHROEDER, R.; PIO, V. M.; BAIL, G. C.; LOPES, F. R. A; WAHRLICH, R. Análise
Espaço-Temporal da Composição da Captura da Pesca com Emalhe de Fundo no
Sudeste/Sul do Brasil. Boletim do Instituto de Pesca, São Paulo, v. 40, n. 3, p. 323 –
353, 2014.
SOUZA, R. C. C. L.; FERNANDES, F. C.; SILVA, E. P. A study on the occurrence of
the brown mussel Perna perna on the sambaquis of the Brazilian coast. Revista do
Museu de Arqueologia e Etnologia, v. 13, p. 3-24, 2003.
STEFFANI, C. N.; BRANCH, G. M. Mechanisms and consequences of competition
between an alien mussel, Mytilus galloprovincialis, and an indigenous limpet,
Scutellastra argenvillei. Journal of Experimental Marine Biology and Ecology, v. 317,
p.127-142, 2005.
WAKIDA-KUSUNOKI A. T.; MACKENZIE JR, C. L. Rangia and marsh clams, Rangia
cuneata, R. flexuosa and Polymesoda caroliniana, in Eastern Mexico: distribution,
biology and ecology, and historical fisheries. Marine Fisheries Review, v. 66, n. 3, p.
13-20, 2004.

418
Belém (PA), 11 a 13 de dezembro de 2019
WILLIAMS, J. D.; BUTLER, R. S.; WARREN, G. L.; JOHNSON, N. A. Freshwater
Mussels of Florida. University Of Alabama Press: Alabama, 2014. 528p.
ZANELLA, T. V. Água de lastro e bioinvasão no Brasil: Uma análise do posicionamento
do Brasil frente ao risco de bioinvasão de espécies exóticas via água de lastro dos navios.
Revista Jurídica Luso Brasileira. v. Ano 1, n. 6, p. 1639-1671, 2015.

419
Belém (PA), 11 a 13 de dezembro de 2019
CARACTERIZAÇÃO MORFOLÓGICA DAS CEPAS DE Microcystis LBARP-1 e
Microcystis LBARP-2 ISOLADAS DO RIO PARÁ (BARCARENA, PARÁ).
Eliane Brabo de Sousa1, Gabriel dos Santos Moura2; Aline Lemos Gomes3; Vanessa
Bandeira da Costa Tavares4; Francisco de Arimatéia dos Santos Alves5
1Doutorado em Saúde Coletiva. Instituto Evandro Chagas. [email protected]
2 Graduação em Farmácia. Faculdade Cosmopolita. [email protected]. 3Mestrado em Ecologia Aquática e Pesca. Instituto Evandro Chagas.
[email protected]. 4 Mestrado em Biologia Ambiental. Instituto Evandro Chagas. [email protected].
5 Mestrado em Biologia Ambiental. Instituto Evandro Chagas
RESUMO
A cianobactéria Microcystis possui o maior número de relatos de florações no mundo. O
objetivo deste estudo foi avaliar o crescimento das cepas Microcystis aeruginosa e M.
wesenbergii isoladas do rio Pará (Barcarena, Pará). As amostras foram obtidas com rede
de plâncton de 45 µm em arrasto horizontal na superfície da água do rio Pará em março
de 2018. O material foi resfriado, analisado, isolado e cultivado em meio líquido BG-11
e ASM-1. A curva de crescimento de M. aeruginosa foi determinada por um experimento
adicionando-se 10 ml de meio de cultura BG-11, para M. aeruginosa, e 10 ml de meio
ASM-1, para M. wesenbergii em tubos de ensaios contendo 3 ml de cada cepa. Os tubos
foram acondicionados em câmara de crescimento durante 30 dias (M. aeruginosa) e 44
dias (M. wesenbergii). A cada três dias foram retirados três amostras para a análise de
clorofila- a. O crescimento de M. aeruginosa apresentou três fases (latência, exponencial
e declínio). A cepa apresentou cor verde oliva e as colônias se distribuíram por toda a
coluna d’água, sugerindo que as florações podem ocupar toda a zona eufótica. A curva
M. wesenbergii apresentou duas fases (latência e exponencial) e o aspecto da cepa era
verde claro, onde as colônias se concentravam na superfície da água, sugerindo que as
florações ocorram na subsuperfície da água formando escumas. Em M. aeruginosa as
células se arranjaram de forma densa sem mucilagem e os diâmetros das colônias e das
células foram de 142 µm e 5,1 µm, respectivamente. Em M. wesenbergii, as colônias
tiveram arranjo denso com mucilagem e o diâmetro de 180,2 µm, em seu eixo menor, e
245 µm, em seu eixo maior e diâmetro celular de 3,4 µm. Sugere-se o monitoramento
mensal das águas da região e coletas em diferentes profundidades da zona eufótica.
Palavras-chave: Amazônia. Água. Eutrofização.
Área de Interesse do Simpósio: Ecologia e Biodiversidade

420
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Muitas espécies de cianobactérias são formadoras de florações e produtoras de
toxinas, sendo prejudiciais para o meio ambiente e a saúde publica. Entre essas espécies
estão às pertencentes ao gênero Microcystis, o qual possui maior número de relatos de
florações. O gênero se caracteriza por apresentar colônias esféricas, ovais, ou irregulares,
com arranjo celular denso ou esparso em volto por mucilagem. Apresenta uma mucilagem
incolor, células esféricas ou hemisféricas com aerótopos. Divisão principalmente em três
planos em gerações sucessivas (KOMÁREK; ANAGNOSTIDIS, 2008). Trata-se de
organismos exclusivamente planctônicos que ocorrem em água doce e salobra, com
ampla distribuição pelo mundo.
Já foram descritas 25 espécies de Microcystis ao redor do mundo e 12 no Brasil.
Na revisão sobre algas do estado do Pará, Costa et al. (2014) reconheceu 11 espécies de
Microcystis.
No que se refere à produção de toxinas, espécies do gênero Microcystis produzem
as microcistinas, uma hepatotoxina constituída por heptapeptídeos cíclicos
hepatocarcinogênico, sendo considerada a cianotoxina com maior ocorrência e
conhecimento em todo mundo, onde são reconhecidas mais de 80 variantes. Nos
mamíferos os efeitos tóxicos podem ser desde agudos (dermatites e parada respiratória) a
crônicos (tumores), podendo causar morte num intervalo de poucas horas a poucos dias
(MOLICA; AZEVEDO, 2009).
Florações de espécies do gênero Microcystis já foram registradas em diversas
regiões do Brasil e no estado do Pará, o estudo realizado por Vieira et al. (2005) apontou
a presença de cepas tóxicas de Radiocystis fernandoi e Microcystis viridis e Sá et al.
(2010) apontaram a presença de uma floração tóxica no Rio Tapajós ocasionada por
espécies dos gêneros Anabaena e Microcystis.
A dinâmica destes organismos em ensaios de microcosmo e a determinação de
produção de microcistinas em cepas de duas espécies de Microcystis isoladas nas águas
do rio Pará próximas a região portuária de Vila do Conde (Barcarena, Pará) podem prever
um possível cenário de Bloom algal e riscos de intoxicação da população ribeirinha que
mantém práticas de contatos primários e secundários neste rio.
Este estudo avaliou o crescimento populacional e a produção de microcistinas nas
cepas de cianobactérias Microcyistis aeruginosa LBARP- 1 e M. wesenbergii LBARP-2

421
Belém (PA), 11 a 13 de dezembro de 2019
(in microcosmo) isoladas das águas do rio Pará, próximo ao Porto de Vila do Conde
(Barcarena, Pará).
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
O rio Pará possui aproximadamente 500 m de largura e 170 Km de extensão, é
um estuário sem nascente definida, pois recebe a contribuição de outros rios menores
como Anapu- Pacajá, Jacundá, Araticu, Cupijó, Tocantins, Moju, Acará e Guamá, seu
início é na baía das Bocas (delta de Boiuçu/Breves) e segue até o oceano Atlântico. Por
receber forte influência oceânica é uma região muito dinâmica e apresenta intenso fluxo
de marés que podem atingir picos de 300.000 m3/s durante a vazante e 200.00 m3/s em
maré enchente (ROSÁRIO, 2016).
2.2 COLETA E CULTIVO DE CIANOBACTÉRIAS
As coletas foram realizadas em março de 2018 (período chuvoso), em um ponto
do rio Pará, próximo à zona portuária de Vila do Conde, município de Barcarena (Figura
1).
Figura 1 – Mapa da localização da área de estudo, rio Pará defronte do porto de Vila do
Conde, município de Barcarena (Pará).
Fonte: Sousa et al. (2019).

422
Belém (PA), 11 a 13 de dezembro de 2019
As amostras foram obtidas com rede de plâncton de 45 µm de abertura de malha
em arrasto horizontal na sub-superfície da água (durante 3 minutos). O material foi
resfriado e analisado em laboratório através de lâminas temporárias em microscópio
biocular (Axiostarplus, Carl Zeiss, Germany) com oculares de medição acoplado a
câmera fotográfica (AxiocamMRc, Carl Zeiss), isolado por capilaridade e mantido em
grande volume (2L) em erlenmeyer com meio líquido BG-11, acrescido de
ciclohexamida para interromper o crescimento de eucariotos. Este material serviu para
análise da curva de crescimento e análise de microcistinas.
As culturas foram não contínuas e se renovavam por meio de repicagem a cada 30
dias. A temperatura de incubação foi de 22°C±1°, luminosidade de aproximadamente 20
µmol photon m.s-1 e fotoperíodo de 14 h de luz.
2.3 CURVA DE CRESCIMENTO
O inóculo das cianobactérias foi estabelecido a partir de 60 dias de cultivo em
meio BG-11 para Microcystis aeruginosa e para Microcystis wesenbergii em meio ASM-
1.
A curva de crescimento de M. aeruginosa foi determinada por meio de um
experimento utilizando tubos de ensaios de 15 ml contendo 10 ml de meio de cultura BG-
11. Em cada tubo foi adicionado 3 ml do inóculo de M. aeruginosa. As amostras (tubos
de ensaios) foram acondicionadas em câmara de crescimento e cultivadas durante 30 dias,
sendo que a cada três dias foram retirados três amostras (tréplica), escolhidas através de
sorteios aleatórios utilizando o Office Excell®. A curva de Microcystis wesenbergii foi
determinada por meio de um experimento utilizando tubos de ensaios de 15 ml contendo
8 ml do meio ASM-1 mais 2 ml do inóculo. As amostras também foram acondicionadas
em câmaras de cultivo com condições ambientais simuladas às condições de Barcarena.
Todos os dias foram retirados amostras (tréplica) para análise de cianobactérias durante
44° dia. As curvas de crescimento foram medidas pela concentração de clorofila- a
(PARSONS; STRICKLAND, 1963).
2.4 ANÁLISE DOS DADOS
Foi construída a curva de crescimento para ambas a cepas e realizada a análise de
variância para testar se houve diferença entre as fases/tempos considerados (teste F de
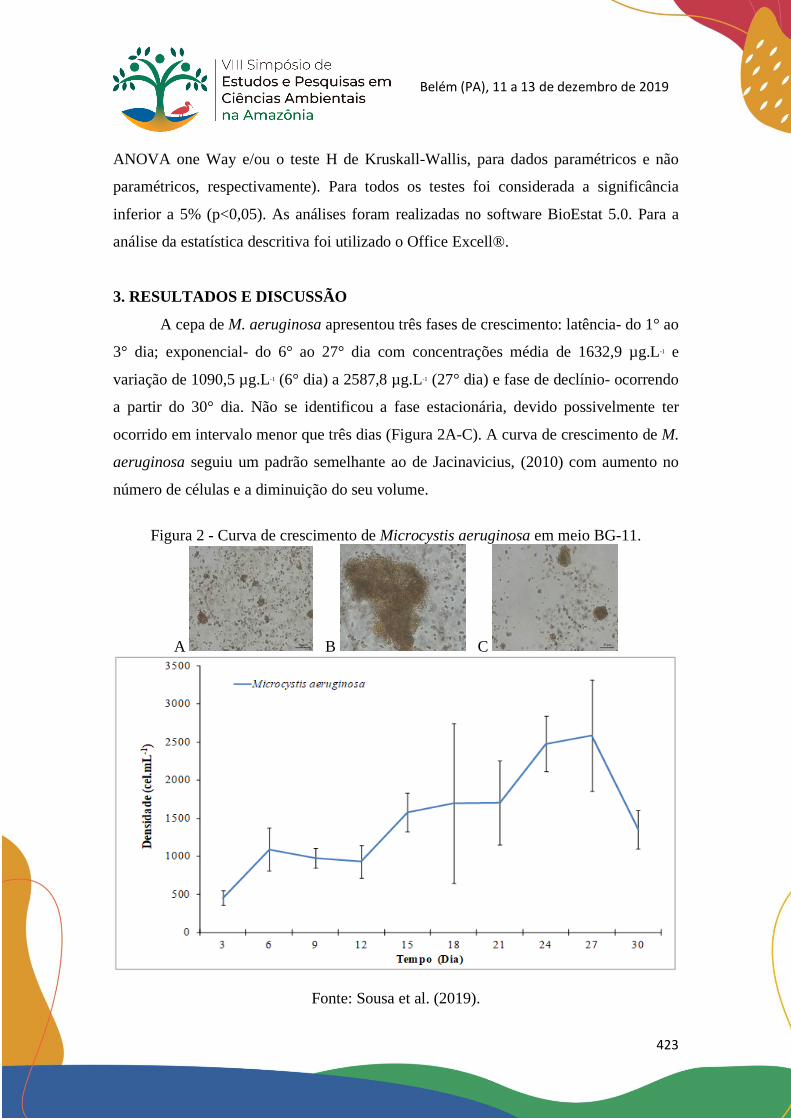
423
Belém (PA), 11 a 13 de dezembro de 2019
ANOVA one Way e/ou o teste H de Kruskall-Wallis, para dados paramétricos e não
paramétricos, respectivamente). Para todos os testes foi considerada a significância
inferior a 5% (p<0,05). As análises foram realizadas no software BioEstat 5.0. Para a
análise da estatística descritiva foi utilizado o Office Excell®.
3. RESULTADOS E DISCUSSÃO
A cepa de M. aeruginosa apresentou três fases de crescimento: latência- do 1° ao
3° dia; exponencial- do 6° ao 27° dia com concentrações média de 1632,9 µg.L-1 e
variação de 1090,5 µg.L-1 (6° dia) a 2587,8 µg.L-1 (27° dia) e fase de declínio- ocorrendo
a partir do 30° dia. Não se identificou a fase estacionária, devido possivelmente ter
ocorrido em intervalo menor que três dias (Figura 2A-C). A curva de crescimento de M.
aeruginosa seguiu um padrão semelhante ao de Jacinavicius, (2010) com aumento no
número de células e a diminuição do seu volume.
Figura 2 - Curva de crescimento de Microcystis aeruginosa em meio BG-11.
A B C
Fonte: Sousa et al. (2019).

424
Belém (PA), 11 a 13 de dezembro de 2019
A aparência da cepa de M. aeruginosa era verde oliva e se distribuía por toda a
coluna d’água do inóculo. O que sugere que as florações envolvendo esta espécie pode
ocupar toda a zona eufótica dos corpos d’água em função dos vários aerótopos nas células.
Apresentou mucilagem hialina fina e muitas colônias pequenas. Observaram-se muitas
células espalhadas e com diâmetro reduzido (~3,5 µm), poucos aerótopos e colônias com
tamanho de 55,5 µm. Estas colônias eram pouco densas e assumiram cor azul esverdeado
claro (Figura 2A).
Na fase exponencial, as células se arranjaram de forma densa sem a formação de
mucilagem (Figura 2B). Os diâmetros das colônias e das células foram de 142 µm e 5,1
µm, respectivamente. No início da fase de declínio as colônias diminuíram o seu
diâmetro, mas não apresentaram qualquer alteração quando comparado ao início do
experimento (Figura 2C).
A curva Microcystis wesenbergii apresentou duas fases: fase de latência que
variou de 125,3 a 888,1 µg.L-1, respectivamente do 1° ao 24° dia, que foi
significativamente diferente (F=72,3; p=0,0011) que a fase exponencial que variou
1265,2 a 4511,0 µg.L-1 do 25° ao 44° dia, respectivamente. A média para as fases de
latência e exponencial foram 427, 0 µg.L-1 e 2153,0 µg.L-1, respectivamente.
O aspecto da cepa Microcystis wesenbergii era verde claro onde a floração se
concentrava na superfície da água desde a fase inicial do cultivo até as fases madura e
senescente, sugerindo que no ambiente natural as florações com esta espécie ocorra na
subsuperfície da zona eufótica.
Na fase de latência as células encontravam- se muito agregadas em colônias
grandes e sem forma definida, envolvidas por mucilagem densa e firme. O diâmetro
médio das células foi de 3,4 µm e as colônias mediram 180,2 µm de comprimento, em
seu eixo menor, e 245 µm, em seu eixo maior. Não foram visualizados aerótopos nas
células o que sugerem que as colônias no estágio inicial mantiveram-se suspensa na água
pela mucilagem (Figura3A-C).
Na fase exponencial o diâmetro variou de 4-7 µm com poucos ou sem aerótopos.
Colônias variando de 724 a 802 µm no seu eixo maior. A partir do 41° as colônias
começam a perder a coloração e ocorreu a formação da clatra, estrutura circular que se
localiza no interior da colônia marcando o início da senescência (Figuras 3D e E).

425
Belém (PA), 11 a 13 de dezembro de 2019
Microcystis wesenbergii é uma espécie dominante no rio Pará na zona portuária
de vila do Conde (Barcarena, Pará), onde a espécie foi isolada, e apresenta maior
densidade no período seco, principalmente de setembro a dezembro (OLIVEIRA, 2018).
Figura 4. Curva de crescimento de Microcystis wesenbergii em meio ASM-1.
Fonte: Sousa et al. (2019).
4. CONCLUSÃO
- O meio BG-11 foi mais eficaz ao crescimento de M. aeruginosa e o meio ASM-
1 foi mais eficaz para M. wesenbergii;
- M. wesenbergii apresenta um crescimento mais lento que M. aeruginosa;
- As florações de M. aeruginosa ocorrem em toda zona fótica da coluna d’água,
de cor verde escura, enquanto que M. wesenbergii na subsuperfície da água, de cor verde
claro e verde pálido com formação de massas agregadas (escumas);
- As florações de M. wesenbergii produz forte mucilagem o que a associa a outras
espécies de cianobactérias, logo sua floração pode estar associada a com outras espécies,
sobretudo filamentosa;
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
5500
6000
1 2 3 4 5 6 7 8 9 1011121314151617181920212223242526272829303132333435363738394041424344
Clo
rofi
la-a
(µg
.L-1
)
Tempo (Dia)
Microcystis wesenbergii

426
Belém (PA), 11 a 13 de dezembro de 2019
- Para a vigilancia se pode considerar que o monitoramento das águas da região
de Barcarena, porto de Vila do Conde, pode ocorrer com frequência mensal.

427
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
COSTA, S. D. et al. Algas e cianobactérias continentais no estado do Pará, Brasil.
Belém: Embrapa Amazônia Oriental, 2014. 351 p.
JACINAVICIUS, Fernanda Rios. Efeitos da temperatura, irradiância e competição
no crescimento e na produção de cianotoxinas da cepa SPC777- Microcystis
aeruginosa (Kützing) Kützing (Cyanobacteria). 2010. Dissertação (Biodiversidade
Vegetal e Meio Ambiente)- Instituto de Botânica da Secretaria de Estado do Meio
Ambiente, São Paulo, 2010.
KOMÁREK, J.; ANAGNOSTIDIS, K. Cyanoprocaryota 1. Teil: Chroococcales. In:
MOESTRUP, Ø.; CALADO, A. Süßwasserflora von Mitteleuropa freshwater flora of
central Europa. Berlim: Spektrum Akademischer Verlag, 2008. 548 p.
MOLICA, R.; AZEVEDO, S. Ecofisiologia de cianobactérias produtoras de cianotoxinas.
Oecologia Brasiliensis, Rio de Janeiro, v. 13, n. 2, p. 229-246, 2009.
OLIVEIRA, Graziela Jones. Estrutura espaço-temporal das cianobactérias
(cyanophyceae) nos portos de Outeiro e Vila do Conde. Pará -Amazônia – Brasil.
2018. Dissertação (Aquicultura e Recursos Aquáticos Tropicais)- Universidade Rural da
Amazônia, Belém, 2018.
PARSONS, T. R.; STRICKLAND, J. D. H. Discussion of spectrophotometric
determination of marine planckton pigments with revised equations of ascertaining
clorophyll a and carotenoids. Journal of Marine Research, New Haven, v. 21, n. 3, p.
155-163, 1963.
ROSÁRIO, R. P. Análise de processos oceanográficos no estuário do Rio Pará. Tese
(Doutorado em Geofísica)- Universidade Federal do Pará, Belém, 2016.
SÁ, L. L. C. et al. Ocorrência de uma Floração de Cianobactérias Tóxicas na Margem
direita do Rio Tapajós, no Município de Santarém (Pará, Brasil). Revista Pan-
Amazônica de Saúde, Belém, v. 1, p. 159-166, 2010.
VIEIRA, J. M. S. et al. Toxic Cyanobacteria and Microcystin Concentrations in a Public
Water Supply Reservoir in the Brazilian Amazonia region. Toxicon, Australia, v. 45, n.
7, p. 901-909, 2005.

428
Belém (PA), 11 a 13 de dezembro de 2019
CIANOBACTÉRIAS COMO INDICADORAS DA QUALIDADE DAS ÁGUAS
USADAS PARA O ABASTECIMENTO DE COMUNIDADES TRADICIONAIS
NA RESERVA BIOLÓGICA RIO TROMBETAS (PARÁ)
Paola Vitoria Brito Pires1; Celly Jenniffer da Silva Cunha2; Aline Lemos Gomes3;
Eliane Brabo de Sousa4; Vanessa Bandeira da Costa Tavares5
1Mestranda em Aquicultura e Recursos Aquáticos Tropicais. Universidade Federal
Rural da Amazônia. [email protected] 2Mestre em Ecologia Aquática e Pesca. Instituto Evandro Chagas.
[email protected] 3Mestre em Ecologia Aquática e Pesca. Instituto Evandro Chagas.
[email protected] 4Doutora em Saúde Coletiva. Instituto Evandro Chagas. [email protected].
5Mestre em Biologia Ambiental. Instituto Evandro Chagas- IEC.
RESUMO
A Reserva Biológica do rio Trombetas está localizada na Calha Norte do Pará, é uma
unidade de conservação de proteção integral, a qual vem sofrendo fortes impactos pela
exploração de minérios. As cianobactérias respondem as mudanças dos fatores físico
químicos da água, podendo ajudar a compreender a respostas dos organismos aos
impactos causados no ambiente aquático. O estudo tem como objetivo caracterizar a
estrutura da comunidade de cianobactérias do rio Trombetas e sua variação espaço-
temporal, destacando o emprego desta comunidade como bioindicadora da qualidade da
água, ao longo do rio e seus afluentes. As coletas foram realizadas no rio Trombetas e
alguns tributários nos meses de maior e menor precipitação pluviométrica em 8 pontos
amostrais. Foram realizadas coletas das variáveis físico químicas, as amostras qualitativas
de cianobactérias obtidas com rede de plâncton (45μm), amostras quantitativas e de
clorofila-a coletadas com garrafas de Van Dorn. Foi realizada analise das espécies
indicadoras (IndVal) e correlação de Spearman. Foram identificados 59 taxóns de
cianobactérias. Em media a riqueza e densidade foram maiores no período chuvoso. A
biomassa foi maior no período menos chuvoso. A espécie Pseudanabaena limnetica foi
indicadora para o lago do Batata. Clorofila-a apresentou correlação positiva com
temperatura e nitrato. As cianobactérias no rio Trombetas e seus afluentes apresentam
alta diversidade, entretanto não foi possível aferir relação entre as variáveis ambientais e
a ocorrência das espécies registradas no período estudado.
Palavras-chave: Bioindicadores. Cianobactérias. Trombetas.
Área de Interesse do Simpósio: Ecologia e Biodiversidade

429
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A Reserva Biológica (REBIO) do rio Trombetas está localizada na Calha Norte
do Pará, pertencente ao grupo de unidades de conservação de proteção integral. A
Mineração Rio do Norte (MRN) está presente na região como a maior empresa produtora
de bauxita do Brasil, matéria prima utilizada na produção do alumínio (MRN, 2014).
Sabe-se que os principais impactos gerados pela extração minério são o total
desmatamento da floresta, através da escavação do solo para extrair o minério, e a
contaminação dos recursos hídricos através das águas utilizadas nos diversos processos
da cadeia produtiva (ÁVILA e MONTE-MÓR, 2007).
Nesta REBIO são encontradas dezenas de populações tradicionais (povos
indígenas e comunidades quilombolas), as quais dependem diretamente dos recursos
naturais desta região para o abastecimento, recreação e pesca de subsistência. Além disso,
enquanto unidade de conservação (UC) de proteção integral é extremamente importante
para a proteção de tabuleiros de desova de tartarugas, com boa conectividade, e dos
demais atributos para aves, mamíferos, biota aquática, macroinvertebrados e vegetação,
apresentando alta diversidade e com grande endemismo (ICMBIO, 2004).
A Mineração Rio do Norte (MRN) já foi responsável pelo assoreamento do lago
Batata, afluente do rio Trombetas, fato denominado por alguns como “o maior desastre
industrial da Amazônia”, onde 20% da superfície líquida tornou-se sólida, aterrada pelos
rejeitos da lavagem do minério que eram depositados no lago (WANDERLEY, 2008). O
alto nível de assoreamento do lago colocou em perigo de contaminação o rio Trombetas.
Esta contaminação pode alterar a qualidade da água, através de mudanças nas suas
características físico-químicas, da entrada de nutrientes e substâncias alóctonas no
sistema hídrico, desiquilibrando as comunidades biológicas.
A legislação brasileira que rege a qualidade das águas, a resolução CONAMA
357/2005. Determina diversos padrões de qualidade, desde físicos, químicos a biológicos.
No Art 8º § 3º da resolução indica que a qualidade dos ambientes aquáticos poderá ser
avaliada por indicadores biológicos, utilizando-se organismos e/ou comunidades
aquáticas. Dentre estas comunidades a fitoplanctônica tem sido um importante parâmetro
para a determinação desta qualidade.
As cianobactérias fazem parte do fitoplâncton, os quais são organismos
fotossintetizantes microscópicos que respondem às alterações dos fatores ambientais

430
Belém (PA), 11 a 13 de dezembro de 2019
(REYNOLDS, 2006). Algumas espécies de cianobactérias são produtoras de metabólitos
tóxicos, principalmente quando estão em desequilíbrio, isto é, quando possuem
crescimentos acentuados (Blooms) no ambiente, além de causarem a alteração das
características organolépticas da água e desequilíbrio no ecossistema aquático.
O conhecimento das espécies de cianobactérias da região da REBIO pode fornecer
informações para o entendimento da resposta desses organismos às alterações ambientais
e sobre os possíveis riscos de intoxicação de populações tradicionais (quilombolas) na
que consomem estas águas. Portanto o estudo teve como objetivo caracterizar a estrutura
da comunidade de cianobactérias do rio Trombetas e sua variação espaço-temporal,
destacando o emprego desta comunidade como bioindicadora da qualidade da água, ao
longo do rio e seus afluentes.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO E ESTRATEGIA AMOSTRAL
A Reserva Biológica do rio Trombetas tem uma área de 385.000 km2, tendo sido
criada pelo Decreto- 84.018 em 21/09/1979. Está localizada no noroeste do estado do
Pará, no município de Oriximiná, na margem esquerda do rio Trombetas e caracteriza-se
por ser um clima típico de região continental, quente e úmido. A área apresenta uma das
precipitações mais baixas da região Amazônica, sendo a média anual de 2.152 mm. A
maior parte das chuvas se concentra nos meses de janeiro a maio.
As coletas foram realizadas no rio Trombetas e alguns tributários nos meses de
maior e menor precipitação pluviométrica (Figura 1). Foram estabelecidos 08 pontos de
amostragem: 2 pontos no igarapé Água Fria, 2 pontos no igarapé Moura, 2 pontos no rio
Trombetas e 2 pontos no lago Batata. Em cada ponto de amostragem foram coletadas
amostras para análise de cianobactérias, clorofila-a (biomassa indireta) e os fatores físico-
químicos da água.

431
Belém (PA), 11 a 13 de dezembro de 2019
Figura 1. Localização das estações de coleta ao longo do rio Trombetas e seus tributários.
Fonte: autores
2.2 COLETA E ANÁLISE DOS FATORES FÍSICO-QUÍMICOS
A transparência da água foi estimada com o uso de um disco de Secchi. As análises
físico-químicas: temperatura (T°C) e pH utilizando a Sonda multiparamétrica. Já as
variáveis turbidez, demanda bioquímica de oxigênio (DBO), N-nitrito (N-NO2-), N-nitrato
(N-NO3-), nitrogênio amoniacal (N-NH4) e fosforo total foram coletadas de forma direta
na sub-superfície da água. Em laboratório, estes parâmetros foram determinados por
espectrometria de UV-VIS e pelo sistema de cromatografia de íons obedecendo aos
procedimentos e as recomendações da American Public Health Association (APHA,
2005).
2.3 COLETA E ANÁLISE DAS CIANOBACTÉRIAS
As amostras qualitativas foram obtidas com rede de plâncton (45μm) e fixadas em
formol 4%. No laboratório foram analisadas por meio da montagem de lâminas
temporárias e observadas sob um microscópio acoplado a um sistema de captura de
imagem. A identificação, nomenclatura e o enquadramento taxonômico foram realizados
de acordo com a literatura especializada.
Já as amostras quantitativas e de clorofila-a foram coletadas com garrafas de Van
Dorn na camada de água anterior ao desaparecimento do disco de secchi, sendo
armazenadas em frascos de polipropileno (250 ml) e preservadas em solução de lugol
10% e refrigeradas, respectivamente. Em laboratório as amostras quantitativas foram

432
Belém (PA), 11 a 13 de dezembro de 2019
analisadas através do método de sedimentação de Utermöhl (1958). Já a clorofila-a foi
analisada por espectrofotometria (STRICKLAND e PARSONS, 1972).
2.4 TRATAMENTO DOS DADOS
Para as análises estatísticas descritivas e analíticas foram utilizados os softwares
PAST10 (HAMMER et al., 2001), PCORD 511 (McMUNE e MEFFORD, 2011) e
CANOCO 4.512 (TER BRAAK e SMILAUER, 2002). Também foi feita análise de
variância para testar as variações dos fatores ambientais por meio do teste F (ANOVA
One Way), para dados normais, e o teste H de Kruskall-Wallis para dados não normais.
Foi realizada uma Análise de Agrupamento (Cluster), a partir da qual foi avaliada
a relação entre as amostras (Modo Q) e as associações de espécies de cianobactérias
(Modo R). Os grupos de amostras formados no Cluster foram utilizados na determinação
das espécies indicadoras do ambiente estudado (IndVal – Valor Indicativo das espécies)
(DUFRENE e LEGENDRE, 1997). Para relacionar as variáveis ambientais com as
variáveis bióticas foi realizada uma Correlação de Spearman.
3. RESULTADOS E DISCUSSÃO
Através da análise da variância foi possível observar que as variáveis pH, turbidez
e salinidade não apresentaram diferenças significativas entre os diversos fatores testados
(períodos sazonais e pontos de amostragem).
A variável temperatura apresentou diferença entre os períodos sazonais (H= 6,35;
p < 0,05), sendo o período menos chuvoso (33,05 ± 3,28 °C o de maior temperatura em
relação ao período mais chuvoso (29,92 ± 1,05 °C), oscilação provavelmente ocasionada
pelo aumento das chuvas no período. O nitrato (F= 0,51; p < 0,05) também variou
somente entre os períodos sazonais. No entanto, este parâmetro esteve maior no período
menos chuvoso (1,6 ± 0,42 mg/L) em relação ao mais chuvoso (0,76 ± 0,25 mg/L).
Foram identificados 59 espécies de cianobactérias, distribuídas em três ordens, 10
famílias e 23 gêneros. A ordem mais representativa foi Oscillatoriales com 43 spp. as
demais ordens (Chroococales e Nostocales) estiveram apresentando 9 spp. e 7 spp.,
respectivamente. As maiores representatividades de espécies foram das famílias
Oscillatoriaceae (9 spp.), Phormidiaceae (11 spp.), e Pseudanabaenaceae (15 spp.).

433
Belém (PA), 11 a 13 de dezembro de 2019
Em relação à riqueza de espécies foi possível observar que a maior riqueza ocorreu
no mês chuvoso (23 spp.) e a menor no mês menos chuvoso (9 spp.), e de maneira geral
a riqueza foi maior no período chuvoso. A maior riqueza taxonômica foi constituída pelos
indivíduos da ordem Oscillatoriales, a qual é uma ordem que inclui todos os gêneros cujas
células possuem talos exclusivamente filamentosos e homocitados (KOMÁREK e
ANAGNOSTIDIS, 1989, 2005) apresentando grande diversidade de habitats, desde
sistemas aquáticos até terrestres, podendo ser planctônicas ou bentônicas (TATON et al.,
2006).
A maioria das espécies foi classificada como esporádicas (35 spp), seguida de
pouco frequente (14 spp), muito frequente (7 spp) e freqüente (3 spp). As espécies com
maior frequência foram Aphanocapsa elachista, Merismopedia tenuissima,
Planktolyngbya brevicellularis, Planktolyngbya limnetica, Pseudanabaenoideae,
Pseudanabaena limnetica e Pseudanabaena sp. 1. Dentre os gêneros das espécies citadas
mais frequentes, podemos destaca-los como possíveis produtores de toxinas e formadores
de florações (SANT’ANNA et al., 2006).
Os valores de densidade variaram entre 3924,48 cel/mL (no periodo menos
chuvoso; ponto 08) a 68499,2 cel/mL (no periodo mais chuvoso; ponto 06). Quando
testados os valores de densidade não diferiram significativamente em relação aos
períodos sazonais e pontos. No entanto, o período chuvoso, em média, apresentou maior
densidade (29703,52 ± 25425,7 cel/mL) (Figura 2).
As espécies que mais contribuíram para a densidade foram Aphanocapsa
elachista, Merismopedia tenuissima, Pseudanabaena limnetica e Pseudanabaenoideae. A
densidade de espécies estando maior no período chuvoso pode estar relacionada à maior
entrada de nutrientes na água. Além disso, as espécies de maior densidade são de gêneros
que podem estar relacionados a habitas metafiticos, organismos que vivem frouxamente
relacionados à macrófitas ou plantas aquáticas, dando maior sustentabilidade no
ambiente, já que o período chuvoso acarreta águas mais agitadas (FRANCESCHINI et
al., 2010).

434
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2. Densidade de células de cianobactérias registrada para região e período de estudo.
Fonte: autores
A alta densidade de cianobactérias pode alterar a qualidade da água (odor, cor e
sabor) e o equilíbrio de vida da biota aquática. Podem ser organismos prejudiciais à saúde
publica por serem potenciais produtores de toxinas (dermatotoxinas, hepatotoxinas e
neurotoxinas) e formadores de florações. Modificam a transparência da água e
consequentemente a penetração de luz solar na coluna d’água, prejudicando
principalmente outros organismos fotossintetizantes (CARMICHAEL et al., 1986;
BEYRUTH, 1991; CETESB, 2009).
A biomassa em termos de clorofila-a oscilou de 0,00 μg.L-1 (P12, chuvoso) a 13,06
μg.L-1 (P12, menos chuvoso). A clorofila-a variou significativamente somente entre os
períodos sazonais (F= 46,91; p< 0,05), sendo o período menos chuvoso com maior
concentração deste pigmento (9,14 ± 2,90 μg.L-1). Todos os valores estiveram abaixo do
valor preconizado pelo Brasil (2005) (Figura 3).

435
Belém (PA), 11 a 13 de dezembro de 2019
Figura 3. Biomassa (Clorofila-a) registrada para a região e período de estudo.
Fonte: autores
O período menos chuvoso foi o que apresentou temperaturas mais elevadas, e com
maiores concentrações de compostos orgânicos na água, estes fatores se relacionam
diretamente com a taxa de atividade fotossintética das comunidades fitoplanctônicas, as
quais fazem parte as cianobactérias, por conta disso podemos relacionar a maior
concentração de clorofila-a em relação a estes fatores (BUZELI e CUNHA-SANTINO,
2013).
A diversidade (H’) variou de um mínimo de 3,17 bits.log2 (período menos
chuvoso) e máximo de 4,5 bits.log2 (período chuvoso). Já a equitabilidade (J’), obteve
valor 1 para todos os pontos e períodos. Para cianobactérias o ambiente foi considerado
de certa forma diverso e homogêneo temporal e espacialmente.
Apenas uma espécie indicadora foi identificada, em relação ao local dos pontos
de coleta, sendo esta Pseudanabaena limnetica (IndVal= 51,5%; p < 0,05), para o lago
Batata. A análise de espécies indicadoras leva em consideração a exclusividade e a
fidelidade das espécies nas amostras (ambientes/habitat), possibilitando assim inferir se
uma ou mais espécies são boas indicadoras ambientais.
O lago Batata já foi alvo de assoreamento por rejeitos de lavagem de minério e
teve 20% da sua superfície sólida (WANDERLEY, 2008). Atualmente a situação do lago
encontra-se de certa forma revertida, contudo, podemos dizer que o lago ainda sofre

436
Belém (PA), 11 a 13 de dezembro de 2019
influencia do assoreamento sofrido, pois a espécie indicadora deste ambiente é capaz de
viver em habitats com camadas turvas e mescladas, apresentando resistência a condições
de alta deficiência de luz (FRANCESCHINI et al., 2010), demonstrando que ainda há
ocorrência de impacto sobre o ambiente.
A descrição dos resultados para a correlação de Spearman encontra-se listados na
Tabela 1. Foi observada correlação direta e positiva entre clorofila-a, nitrato e
temperatura, quando estes parâmetros aumentam o nível deste pigmento também
aumenta. Devido ao aumento da temperatura, os organismos presentes no ambiente
podem ter seu metabolismo alterado, de forma que essas temperaturas elevadas
promovam um crescimento da taxa metabólica. Para organismos que realizam
fotossíntese, como o fitoplâncton em geral, o aumento dessa taxa pode acarretar em uma
maior atividade fotossintética, produzindo assim, mais pigmento fotossintetizante, a
clorofila (BUZELI e CUNHA-SANTINO, 2013).
Já a presença de nitrato no ambiente proporciona maior crescimento das
populações de cianobactérias, que são beneficiadas pela presença de compostos orgânicos
na água, uma maior população gera maior produção fotossintética, o que pode ser
explicado pela correlação positiva da presença de pigmento fotossintetizante e nitrato na
água (BUZELI e CUNHA-SANTINO, 2013).
Tabela 1. Correlação de Spearman entre os parâmetros bióticos e abióticos da região e período
de estudo.
Fonte: autores
4. CONCLUSÃO
A ordem Oscillatoriales e a família Pseudanabaenaceae, foram as mais
representativas por terem apresentado maior riqueza de espécies no presente estudo.

437
Belém (PA), 11 a 13 de dezembro de 2019
Apenas uma espécie indicadora foi identificada, sendo esta para o lago Batata, onde a
espécie aponta características que indicam que o ambiente ainda continua afetado pelas
consequências dos impactos sofridos. As cianobactérias no rio Trombetas e seus afluentes
apresentam alta diversidade, entretanto não foi possível encontrar relação significativa
entre as variáveis ambientais e a ocorrência da maioria das espécies registradas no período
estudado.

438
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ÁVILA, J. L. T.; MONTE-MÓR, R. L. M. Urbanização e impactos ambientais: uma
análise da relação entre as características dos espaços urbanos e a poluição hídrica na
região do médio Rio Doce (MG). In: VII Encontro da Sociedade Brasileira de Economia
Ecológica, 2007, Fortaleza Anais do VII Encontro da Sociedade Brasileira de Economia
Ecológica. 2007.
APHA- American Public Health Association. Standard Methods For The Examination
Of Water And Wasterwater. 19th. Washington: Apha, 2005.
BRASIL. [Resolução Conama 357/2005] Conselho Nacional do Meio Ambiente -
CONAMA. Resolução Nº 357, de 17 de março de 2005. Dispõe sobre a classificação dos
corpos de água e diretrizes ambientais para o seu enquadramento, bem como estabelece
as condições e padrões de lançamento de efluentes, e dá outras providências. Disponível
em: http://www2.mma.gov.br/port/conama/legiabre.cfm?codlegi=459. Acesso em: 12
out. 2019.
BEYRUTH, Z. Guarapiranga Também Pede Água. In: Comunidade fitoplanctônica da
represa de Guarapiranga: Aspectos ecológicos, sanitários e subsídios para
reabilitação ambiental. 1996. 191f. Tese (Doutorado em Saúde Coletiva) –
Universidade de São Paulo, São Paulo, 1996.
BUZELLI, G. M.; CUNHA-SANTINO, M. B. Análise e diagnóstico da qualidade da água
e estado trófico do reservatório de Barra Bonita (SP). Revista Ambiente & Água, v. 8,
n. 1, p. 186-205. 2013.
CARMICHAEL, L. W. W.; PINNOTI, M. H.;FRALEIGH, P. C. Toxicity of a clonal
isolate os the cyanobacterium (blue-green alga) microcystis aeruginosa from Lake Erie.
In: Annual Meeting Ohio Academy Of Science, 95, 1986, Proceddings... Toledo: (s.n).
1986.
CETESB - Companhia De Tecnologia de Saneamento Ambiental. Qualidade das Águas
Interiores no Estado de São Paulo. Série Relatórios. Apêndice A - Significado
Ambiental e Sanitário das Variáveis de Qualidade das Águas e dos Sedimentos e
Metodologias Analíticas e de Amostragem. 2009
DUFRENE, M.; LEGENDRE, P. Species assemblages and indicator species: the need for
a flexible asymmetrical approach. Ecological Monographs, v. 67, n. 3, p. 345-366, 1997.
FRANCESCHINI, I. M.; BURLIGA, A. L.; REVIERS, B.; PRADO, J. F.; HAMLAOUI,
S. Algas: Uma abordagem filogenética, taxonômica e ecológica. Porto Alegre: Artmed,
2010. 332 p.
HAMMER, Ø.; HARPER, D. A. T.;RYAN, P. D. Past. Paleontological statistics software
package for education and data analysis. Palaeontologia Electronica. 2001.
ICMBIO. Plano de Manejo da Reserva Biológica do Rio Trombetas. Instituto Chico
Mendes de Conservação da Biodiversidade. 2004. Disponível em:
[http://www.icmbio.gov.br/portal/images/stories/imgsunidadescoservacao/rebio_trombe
tas.pdf]. Acesso em: 30 nov. 2017.

439
Belém (PA), 11 a 13 de dezembro de 2019
KOMÁREK, J.; ANAGNOSTIDIS, K. Modern approach to the classification system of
cyanophytes. In: 4: Nostocales: Algological Studies, 1989.
KOMÁREK, J.; ANAGNOSTIDIS, K. Cyanoprokaryota 2. Teil/ 2nd Part: Oscillatoriales.
In: BÜDEL, B.; KRIENITZ, L., et al. (Ed.). Sußwasserflora Von Mitteleuropa 19/2.
Heidelberg: Elsevier/Spektrum, Heidelberg, v.19/2, p.1-759. 2005.
MCCUNE, B.; MEFFORD, M. J. PC-ORD: Multivariate Analysis Of Ecological Data.
Gleneden Beach, Oregon: MjM Software, 2011.
MRN - Mineração Rio Do Norte – Relatório Da Administração, 2014.
REYNOLDS, C. S. Ecology of phytoplankton. Cambrigde University Press, 2006.
552p.
SANT’ANNA, C. L.; AZEVEDO, M. T. P.; AGUJARO, L.; CARVALHO, M. C. et al.
Manual ilustrado para: identificação e contagem de cianobactérias planctônicas de
águas continentais brasileiras. Rio de Janeiro: Editora Interciência, 2006. 60p.
STRICKLAND, J. D. H.; PARSONS, T. R. A practical handbook of sea water
analysis. 2 ed. Ottawa: Fisheries Research Board Of Canadá, 1972. 310p.
TATON, A.; GRUBISIC, S.; ERTZ, D.; HODGSON, D. A. et al. Polyphasic study of
antarctic cyanobacterial strains. Journal Of Phycology, v. 42, n. 6, p. 1257 - 1270. 2006.
TER BRAAK, C. J. F.; SMILAUER, P. CANOCO Reference Manual and CanoDraw
for Windows User's Guide: Software for Canonical Community Ordination.
Version 4.5. Ithaca, New York: www.canoco.com (Microcomputer Power), 2002.
UTERMÖHL, H. Zur vervollkommung der quantitativen phytoplankton-methodic.
Mitteilungen Internationale Vereiningung Fur Teoretische Und Angewandle
Limnologie, v. 19, p. 1-38, 1958.
WANDERLEY, L. J. M. Conflitos e movimentos sociais populares em área de
mineração na Amazônia brasileira. 2008. 152f. Dissertação (Mestrado em Geografia)
- Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2008.

440
Belém (PA), 11 a 13 de dezembro de 2019
COMPOSIÇÃO FLORÍSTICA E FITOSSOCIOLOGIA ESPACIAL DE
COMUNIDADES VEGETAIS EM UM AMBIENTE RIPÁRIO ANTROPIZADO
NO NORDESTE DO ESTADO DO PARÁ, BRASIL
Luna Gabriela Santiago Monteiro1; João Victor de Moraes Souza Pinheiro²; Lucélia
Thaís da Silva Cruz³.
1Mestranda em Biologia Ambiental. UFPA Campus Bragança.
2 Mestrando em Biologia Ambiental. UFPA Campus Bragança.
³Mestranda em Estudos Antrópicos na Amazônia. UFPA Campus Castanhal.
RESUMO
As matas ciliares possuem grande importância aos sistemas hidrográficos e ecossistemas
terrestres, sendo consideradas pela legislação brasileira como Áreas de Preservação
Permanente (APP), no entanto têm sido continuamente suprimida devido vários fatores
antrópicos. Dessa maneira este trabalho teve como objetivo avaliar a composição
florística e distribuição de comunidades vegetais na mata associada ao rio Cereja,
localizada no município de Bragança, NE-PA, bem como determinar os índices de
densidade e equitabilidade dos espécimes registrados, dentre outras variáveis
fitossociológicas. A análise florística foi realizada em duas parcelas (P1 e P2) paralelas
ao rio, onde cada parcela foi dividida em dois lotes de 5X5 m. Dentro de cada lote foi
feito uma subdivisão em sub-lotes de 1X1 m para a obtenção de dados quantitativos utilizáveis ao estudo fitossociológico. As principais famílias registradas foram:
Asteraceae (5 espécies), Fabaceae (5 espécies) e Poaceae (4 espécies). Os maiores valores
de densidade, diversidade e equitabilidade ocorreram na P1, dessa forma estima-se que
esses valores estejam relacionados às condições ambientais menos perturbadas do local
em comparação a P2, favorecendo uma heterogeneidade de táxons. A vegetação ripária
do rio Cereja se apresentou bastante diversa e heterogeneamente distribuída, sendo
constituída principalmente por plantas rasteiras e de pequeno porte. A presença de
Spermacoce verticillata Meye em ambas parcelas analisadas é devido esta espécie ser
altamente tolerante a solos ácidos e pobres em nutrientes, sendo comuns em áreas
degradadas ou em fase de degradação, além de serem grandes competidoras por nutrientes
e água, inibindo em parte a presença de outras espécies.
Palavras-chave: Antropização. Parâmetros Fitossociológicos. Vegetação Ripária.
Área de Interesse do Simpósio: Ecologia e Biodiversidade.

441
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Os cursos d’água, assim como os reservatórios e as nascentes dos rios, atuam
como limitantes ao desenvolvimento e sobrevivência de grande parte dos organismos que
vivem nestes ambientes e em suas adjacências (Metzger, 2010). A preservação de corpos
hídricos está diretamente associada às condições em que se encontra a vegetação
proximal, estando esta vegetação composta principalmente por florestas ripárias que
estabilizam e protegem os solos das margens dos rios e represas (Faria, 2001). Essas
importantes formações vegetais se encontram associadas aos corpos d’água, ao longo dos
quais podem se estender por dezenas de metros a partir das margens e apresentar
marcantes variações na composição florística e na estrutura comunitária, dependendo das
interações que se estabelecem entre ecossistema aquático e o ambiente terrestre adjacente
(Chaves, 2005). As faixas de vegetação que cercam os corpos hídricos são dependentes
de uma série de fatores, dentre eles, a dinâmica de equilíbrio entre fauna, flora e o meio
abiótico existente, devendo a faixa de mata pré-estabelecida ser capaz de desemprenhar
satisfatoriamente todas as funções vitais deste ecossistema (Metzger, 2010).
As florestas ripárias, ripícolas ou ainda matas ciliares, como são mais conhecidas,
são consideradas pela legislação brasileira como Áreas de Preservação Permanente
(APP), no entanto o alto nível de degradação desses ecossistemas é resultado direto da
ação indiscriminada do homem verificado pelo mau uso do solo, desmatamento,
queimadas e cujas consequências são danosas para a geração atual e futura (Santos, et al.,
2014). As matas ciliares agem dificultando o escoamento da água de enxurradas,
diminuindo sua velocidade e promovendo maior infiltração no solo, devendo-se este
fenômeno às arvores, suas raízes e a camada de matéria orgânica depositada sobre o solo
(serapilheira) (Vogel et al., 2009), além do controle de erosão em áreas fluviais (Durlo e
Sutili, 2005). Desta maneira, em virtude da grande importância das vegetações ciliares
aos sistemas hidrográficos e ecossistemas terrestres, este trabalho teve como objetivo
avaliar a composição florística e distribuição de comunidades vegetais ao longo do Rio
Cereja, localizado no nordeste do estado do Pará, bem como determinar os índices de
densidade e equitabilidade dos espécimes registrados, dentre outras variáveis
fitossociológicas.

442
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
O município de Bragança está situado sobre uma extensa superfície de planície
costeira, entre dois grandes estuários, o do Taperaçu e o do rio Caeté (Pereira et al., 2009)
e apresentando características inerentes, tais como rios extensos e hidrodinâmicos, amplas
áreas de manguezais e planaltos rebaixados (Carvalho, 2002). A hidrografia do município
é intensamente recortada por igarapés e apresenta dois rios principais, o rio Caeté, que
desagua no oceano Atlântico, e o rio Cereja, que é um dos afluentes do rio Caeté (Figura
1) (IBGE, 2003). O rio atravessa sete bairros de Bragança: Taíra, Alegre, Cereja, Vila
Sinhá, Padre Luís, Centro e Aldeia, e apresenta aproximadamente 1,5 metros de
profundidade, 6 metros de largura e 5 km de comprimento (Santos et al., 2014). O
crescimento desordenado da população de Bragança culminou em um processo crescente
de degradação ambiental e na prática de constantes agressões contra a climatização local,
a boa e correta drenagem, as áreas verdes e matas ciliares, aos cursos d’água e a topografia
original (Mello, 2002), tendo como reflexo disso as condições atuais do rio Cereja.
Figura 1 - Localização geográfica da área de amostragem: (A) América do Sul, (B)
Salgado Bragantino; (C) os dois pontos de coleta ao longo do Rio Cereja.
Fonte: autores.

443
Belém (PA), 11 a 13 de dezembro de 2019
2.2 COLETAS E ANÁLISES DE DADOS
Foram determinadas duas parcelas de amostragem ao longo do Rio Cereja, Parcela
1 (P1 – S1°3’15,80616 e W46°46’19,16112) e Parcela 2 (P2 – S1°3’4,464 e
W46°45’56,88792) (Figura 1). Cada parcela foi subdividida em dois lotes, tendo cada
lote 5X5 m, os quais foram medidos com o auxílio de fita métrica. Para cada lote foram
coletadas amostras de todas as espécies vegetais detectadas, a fim de se fazer uma análise
qualitativa da composição florística do ambiente. Como método de preservação dos
espécimes vegetais coletados, foram confeccionadas exsicatas (Mori et al., 1989), as
quais foram armazenadas em estufa por 48 horas e posteriormente levadas ao herbário da
Universidade Federal do Pará (UFPA – Bragança) para serem classificadas quanto à
taxonomia.
A classificação taxonômica foi realizada com a ajuda de especialista em
fanerógamas da própria UFPA. Sub-lotes de 1X1 m foram delimitados dentro dos lotes
maiores de 5X5 m. Todos os organismos vegetais dentro dos sub-lotes de 1X1 m foram
coletados, sendo os de pequeno porte coletados desde suas raízes, e contabilizados a fim
de se obter dados quantitativos para o cálculo da diversidade específica, equitabilidade,
densidade das espécies, abundância relativa e da frequência de ocorrência.
3. RESULTADOS E DISCUSSÃO
3.1 COMPOSIÇÃO FLORÍSTICA
No levantamento florístico foram registradas 34 espécies, pertencentes a 12
famílias, sendo que seis organismos não foram identificados em nenhum nível
taxonômico. (Tabela 1). As famílias com maior número de espécies foram: Asteraceae
(5) e Fabaceae (5), seguido por Poaceae (4), Malvaceae (3), Rubiaceae (2), Acanthaceae
(2), Amaranthaceae (2), Commelinaceae (2), Plantaginaceae (1), Onagraceae (1),
Passifloraceae (1), Myrtaceae (1), Combretaceae (1), Convolvulaceae (1), Solanaceae (1),
Arecaceae (1), Cleomaceae (1). Considerando o número de espécies por gênero, três
organismos não foram classificados a esse nível taxonômico e seis em nenhum nível
taxonômico, no total, os gêneros que apresentaram maior número de espécies foram:

444
Belém (PA), 11 a 13 de dezembro de 2019
Paspalum (4), Alternanthera (2), Commelina (2), Spermacoce (2), Mimosa (2), Sida (2)
Senna (1), Macrolobium (1), Sphagneticola (1), Synedrella (1), Eclipta (1), Ruellia (1),
Hygrophila (1), Scoparia (1), Triumfetta (1), Ludwigia (1), Turnera (1), Psidium (1),
Terminalia (1), Ipomoea (1), Solanum (1), Euterpe (1), Cleone (1) (Tabela 1). Foi
registrado para o Rio Cereja amplo espectro de variação quanto à sua preservação, desde
a nascente até a foz (Silva-Filho, 2015), e assim como no trabalho deste autor, a vegetação
ripária do Rio Cereja foi constituída predominantemente por espécies rasteiras, de
pequeno porte, com maior diversidade e maior conservação de espécies nativas,
registrando-se maiores índices de densidade e diversidade nas porções à jusante do rio, o
que se deve em parte a menor ação antrópica neste local.
Tabela 1 - Composição das espécies de vegetais presentes às margens do Rio Cereja. FAMÍLIAS/TÁXONS
FABACEAE POACEAE Senna sp. Paspalum urvillei Steud Macrolobium sp. Paspalum sp.1 Mimosa sp. Paspalum sp.2 Mimosa hostilis Benth Paspalum sp.3 RUBIACEAE MALVACEAE Spermacoce verticillata Meye Triumfetta sp. Spermacoce sp. Sida sp.1 ASTERACEAE Sida sp.2 Sphagneticola triloba Pruski ONAGRACEAE Asteraceae sp.1 Ludwigia sp. Asteraceae sp.2 PASSIFLORACEAE Synedrela sp. Turnera sp. Eclipta sp. MYRTACEAE ACANTHACEAE Psidium guajava Burman. Ruellia blechum Browne COMBRETACEAE Hygrophila costata Nees Terminalia catappa Linn PLANTAGINACEAE CONVOLVULACEAE Scoparia dulcis Valke Ipomoea asarifolia Schult COMMELINACEAE SOLANACEAE Commelina sp1 Solanum sp. Commelina sp2 ARECACEAE AMARANTHACEAE Euterpe oleracea Mart Alternanthera brasiliana Kunt. CLEOMACEAE Alternanthera sp. Cleone sp.
3.2 DIVERSIDADE ESPECÍFICA, EQUITABILIDADE E DENSIDADES
Os valores de diversidade específica de Shannon oscilaram entre 0,89 ± 0,28 na
Parcela 2 (porção mais externa do rio) a 2,05 ± 0,02 na Parcela 1 (porção mais interna).

445
Belém (PA), 11 a 13 de dezembro de 2019
Os índices de equitabilidade (Pielou, 1977) seguiram o mesmo padrão da diversidade,
com valores mais baixos ao longo da P2 (0,64 ± 0,20) e mais altos ao longo da P1 (0,75
± 0,02). Por outro lado, a densidade total de espécies variou de 59 ± 35,35 organismos/m2
na P2 a 119,5 ± 64,34 organismos/m2 na P1. Os táxons que apresentaram maior valor de
densidade por m2 na P1 foram: Asteraceae sp 2 com 39,5 organismos/m2, Senna sp com
24 organismos/m2 e Sphagneticola triloba, com 17 organismos/m2; e os que apresentaram
maior valor de densidade na P2 foram: Poaceae sp 3 com 35 organismos/m2, Spermacoce
verticillata 13 organismos/m2 e Ruellia blechum com 9 organismos/m2 (Figura 2).
Figura 2 - Médias e ± DP das variáveis fitossociológicas: Diversidade Específica (H’),
Equitabilidade (J’) e Densidade total dos organismos/m2, considerando-se as duas parcelas.
Fonte: autores.
Os valores de diversidade específica (0,89 ± 0,28) e equitabilidade (0,64 ± 0,20)
mais baixos para a parcela 2 (porção mais externa do rio) pode ser elucidado pelos
impactos e as alterações decorrentes da ocupação desordenada desta área, visto que essa
condição insalubre contribui expressivamente para a degradação ambiental, como já visto
por Silva-Filho & Nunes (2017), onde verificaram que a invasão da população às

446
Belém (PA), 11 a 13 de dezembro de 2019
proximidades do Rio Cereja causa a diminuição nos índices de integridade do rio,
contribuindo para o processo de eutrofização.
3.3 ABUNDÂNCIA RELATIVA
Em ambas as parcelas, três espécies apresentaram abundância relativa
significante. Na parcela 1, Asteraceae sp 2, Sphagneticola triloba Pruski e Ruellia
blechum Browne apresentaram abundância relativa de 33,05%, 14,22% e 12,55%
respectivamente. Na parcela 2, Poaceae sp 3, Spermacoce verticillata Meye e R. blechum
apresentaram 59,32%, 22,03 e 7,62 respectivamente. R. blechum esteve presente tanto na
parcela 1 quanto na parcela 2, apresentando valores consideráveis de abundância de
12,55% e 7,62 respectivamente.
A riqueza de espécies relativamente elevada é característica comum em matas
ciliares devido à heterogeneidade ambiental comumente superior à de florestas de terra
firme próximas (Oliveira Filho et al., 1990). Segundo Rodrigues e Nave (2000), esse fato
é condicionado pela natureza ecotonal da faixa ciliar, que é ocupada por mais de um tipo
vegetacional ou mesmo por formações fitofisionômicas distintas, que diferem fortemente
entre si em termos de composição florística. Sendo assim, no presente estudo se constatou
ampla riqueza de espécies, com índices relativamente mais altos na parcela 1, a qual
demonstrou elevada heterogeneidade.
Figura 3 - Abundância Relativa (%) dos principais táxons que compuseram a vegetação
ripária nas duas parcelas estudadas ao longo do Rio Cereja.
Fonte: autores.

447
Belém (PA), 11 a 13 de dezembro de 2019
3.4 FREQUÊNCIA DE OCORRÊNCIA
Quanto à frequência de ocorrência das espécies vegetais registradas, apenas
Spermacoce verticillata apresentou 100% de frequência, estando presente em todos os
sub-lotes amostrados (ou seja, nas duas parcelas). A Tabela 2 mostra outros dados sobre
a frequência de ocorrência dos demais táxons registrados em suas respectivas parcelas ao
longo do Rio Cereja. Dentre as espécies vegetais coletadas, apenas Spermacoce
verticillata Meye foi comum a todas as parcelas e sub-lotes. Outros táxons com maior
distribuição foram Ruellia blechum Browne e Poaceae sp 3. A ampla distribuição e
relativa dominância de S. verticillata, comumente conhecida como vassourinha-de-botão,
podem ser atribuídas ao fato desta ser altamente tolerante a solos ácidos e pobres em
nutrientes (Kissmann e Groth, 2000).
Na Amazônia é uma das espécies mais comuns em pastagens nativas ou cultivadas
em terra firme, sobre tudo em áreas degradantes ou em fase de degradação (Dutra et al.,
2004; Fontes et al., 2011). Outro importante ponto a ser destacado é que a espécie S.
verticillata é uma grande competidora por nutrientes, capaz de interferir na presença de
outras espécies, como já foi verificado por Fontes e Tonato (2016), em que registraram
forte extração de nutrientes pela espécie como, por exemplo, nitrogênio, cálcio, potássio,
magnésio, enxofre, fosforo, ferro, manganês, zinco e boro, que ficaram indisponíveis para
outras espécies, causando redução no crescimento e aparecimento destas. Este pode ser
um fato explicativo do menor número de espécies na parcela 2, uma vez que, S.
verticillata, embora não tenha constituído a espécie dominante para esta parcela,
representou o segundo maior numero (26) de indivíduos por m2, levando à redução da
diversidade neste ambiente.

448
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 2 - Frequência relativa de ocorrência dos espécimes vegetais registrados nas respectivas
parcelas de amostragem. * Valores em percentagem (%)
Espécie P1 P2 Espécie P1 P2
Alternanthera brasiliana 100 - Eclipta sp 50 -
Alternanthera sp 100 - Euterpe oleracea 50 -
Asteraceae sp 2 100 - Fabaceae sp 1 50 -
Ruellia blechum 100 50 Macrolobium sp 50 -
Sp1 desconhecida 100 - Poaceae Sp 2 50 -
Sp2 desconhecida 100 - Poaceae Sp 3 50 100
Sp4 desconhecida 100 - Senna sp 50 50
Spermacoce vertillata 100 100 Sp3 desconhecida 50 -
Sphagneticola triloba 100 - Sp5 desconhecida 50 -
Poaceae sp 1 50 50 Sp6 desconhecida 50 -
Commelina sp 1 50 - Mimosa hostilis - 50
Commelina sp 2 50 -
4. CONSIDERAÇÕES FINAIS
A vegetação ripária do rio Cereja se apresentou bastante diversa e
heterogeneamente distribuída, sendo constituída principalmente por plantas rasteiras e de
pequeno porte. As condições abióticas e o nível de degradação do ambiente aparentam
interferir diretamente nos índices de densidade por metro quadrado de vegetação, bem
como nos valores de diversidade de espécies, dentre outras variáveis fitossociológicas.
Com isso, pode-se inferir que quanto mais interno for o trecho analisado ao longo do rio
Cereja, aparentemente menor será o nível de perturbação do ambiente, tendendo a certa
preservação do ecossistema em questão, o que garante melhor condição de acolhimento
a uma maior gama de espécies vegetais, que colaboram para o aumento da diversidade
local. Além disso, a presença de Spermacoce verticillata Meye em ambos os trechos
analisados constitui outro fator determinante ao estabelecimento e desenvolvimento de
outras espécies no ambiente, já que segundo a literatura esta espécie é caracterizada como
grande competidora, sendo capaz de influenciar nos índices de diversidade vegetacional
do ambiente em questão.

449
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
CARVALHO, M. L. Aspectos da produtividade primária dos bosques de mangue
do Furo Grande, Bragança-Pará. Dissertação (Mestrado) - Universidade Federal do
Pará, Belém-Pará, 2002.
CHAVES, M. M. F. Reflorestamentos mistos com essências nativas para recomposição
de Matas Ciliares. In: Andrade, J.; Sanquetta, C. R.; Ugaya, C. Identificação de áreas
prioritárias para recuperação da Mata Ciliar na UHE Salto Caxias. Revista Espaço
Energia. n. 3. 2005.
DURLO, M. A.; SUTILI, F. J. Bioengenharia: Manejo biotécnico de cursos de água.
Porto Alegre: EST Edições. p. 189, 2005.
DUTRA, S.; SOUZA FILHO, A. P.; MASCARENHAS, R. E. B.; GONÇALVES, C.A.
Controle integrado de plantas invasoras em pastagens cultivadas no município de Terra
Alta, Nordeste Paraense. Belém-PA: Embrapa Amazônia Oriental, (Embrapa Amazônia
Oriental. Boletim de Pesquisa e Desenvolvimento, 25, p. 30, 2004.
FARIA, H. H.; SÉRIO, F. C.; GARRIDO, M. A. O. Reposição da vegetação ciliar
integrada à conservação de microbacia. São Paulo: Série Registros, vol. 21, p. 1-13,
2001.
FONTES, J. R. A.; PERIN, R., SOUSA, J. N.; MARTINS, G. C; SANTOS, A. M. B.
Manejo integrado de plantas daninhas em pastagens na integração lavoura-pecuária-
floresta. Manaus-AM: Embrapa Amazônia Ocidental (Embrapa Amazônia Ocidental.
Circular Técnica, 37), p. 10, 2011.
FONTES, J. R. A.; TONATO, F. Acúmulo de nutrientes por vassourinha-de-botão
(Spermacoce verticillata), planta daninha de pastagens na Amazônia. Embrapa
Amazônia Ocidental-Circular Técnica (INFOTECA-E), 2016.
IBGE – Instituto Brasileiro de Geografia e Estatística, Cidades/População, 2003.
KISSMANN, K. G.; GROTH, D. Plantas infestantes e nocivas. 2. Ed. São Paulo:
BASF, T., p. 722, 2000.
MELLO, F. A. O. Análise do processo de formação da paisagem urbana no
município de Viçosa, Minas Gerais. Universidade Federal de Viçosa: Dissertação de
mestrado, p. 122, 2002.
METZGER, J. P. O Código Florestal tem base científica? Associação Brasileira de
Ciência Ecológica e Conservação: editora cubo, vol. 8, n. 1, p. 1-5, 2010.
MORI, S. A.; SILVA, L. A. M.; LISBOA, G.; CORADIN, L. Manual de manejo do
herbário fanerogâmico. Ilhéus: Centro de Pesquisas do Cacau, p. 103, 1989.
OLIVEIRA FILHO, A.T., RATTER, J.A. & SHEPHERD, G.J. Floristic composition
and community structure of a central Brazilian gallery forest. Flora, vol. 184, 103-117,
1990.

450
Belém (PA), 11 a 13 de dezembro de 2019
PEREIRA, L. C. C. et al. Morphological and sedimentological changes in a macrotidal
sand beach in the Amazon Littoral (Vila dos Pescadores, Pará, Brazil). Portugal: Jornal
of Coastal Research, p. 113-117, 2009.
PIELOU, E. C. Mathematical Ecology. Wiley, New York, p. 385, 1977.
RODRIGUES, R.R.; NAVE, A.G. Heterogeneidade florística das matas ciliares. In
Matas Ciliares: conservação e recuperação (R.R. Rodrigues & H.F. Leitão-Filho, eds.).
Edusp, São Paulo, p.45-71, 2000.
SANTOS, M. R. S.; MOREIRA, A. M.; SANTOS, M. N. S. Análise socioambiental
dos moradores da APP urbana do rio Cereja, Bragança-Pará. APP Urbana, UFPA –
Belém, 2014.
SILVA-FILHO, C. P.; NUNES, Z. M. P. Índice de integridade ambiental do rio Cereja-
Bragança-PA. In: FÓRUM INTERNACIONAL DE RESÍDUOS SÓLIDOS-Anais,
2017.
VOGEL, H. F.; ZAWADZKI, C. H.; METRI, R. Florestas ripárias: importância e
principais ameaças. Campo Mourão: SaBios: Rev., Saúde e Biol., vol. 4, n. 1, p. 24-
30, 2009.

451
Belém (PA), 11 a 13 de dezembro de 2019
DESEMPENHO DE MACRÓFITAS AQUÁTICAS SOBRE INTERFERÊNCIA
DE ESPÉCIES NATIVAS E INVASORAS
Rayssa Silva do Carmo¹; Nayara Louback Franco²; Thaisa Sala Michelan³.
¹Graduanda de Ciências Biológicas, curso Bacharelado. Instituto de Ciências
Biológicas, Universidade Federal do Pará. [email protected]
²Mestre em Ciências Ambientais. Instituto de Ciências Biológicas, Universidade
Federal do Pará. [email protected]
³Doutora em Ecologia de Ambientes Aquáticos. Instituto de Ciências Biológicas,
Universidade Federal do Pará. [email protected]
RESUMO
O presente estudo objetivou verificar o desempenho de macrófitas aquáticas sobre
interferência de espécies nativas e invasoras. O experimento foi realizado na
Universidade Federal do Pará e teve duração de 13 dias no mês de maio de 2019. As
amostras das plantas foram coletadas na Universidade Federal Rural da Amazônia,
Belém, Brasil. Visto em ambientes aquáticos continentais, macrófitas aquáticas
frequentemente se tornam invasoras e tendem a causar fortes impactos ao criar
estruturação física e controlar aspectos físicos e químicos da água e dos sedimentos. As
plantas invasoras podem deter características que as auxiliam a se estabelecer em um
ambiente, dentre elas, destacamos os efeitos alelopáticos, que são um tipo de interações
bioquímicas entre diversas espécies de vegetais, considerada uma forma de adaptação
química defensiva, além de ser um fator de estresse ambiental para muitas espécies. Com
isso, selecionadas, as plantas Urochloa arrecta, Pistia stratiotes e, Hydrocleys
nymphoides, foram lavadas e separadas dos resíduos do lago. Após mensuradas, as
plantas foram colocadas em 3 diferentes tratamentos com P. stratiotes, com 10 réplicas
para cada tratamento, totalizando 30 unidades experimentais. Porém, não houve diferença
de crescimento entre os tratamentos, mas sim, um crescimento entre os tratamentos
controle (água) e nativa (Hydrocleys nymphoides). Nesse sentido, a hipótese de que
haveria ações alelopáticas da espécie de macrófita invasora Urochloa arrecta foi rejeitada,
pois a espécie nativa Pistia stratiotes não apresentou diferença significativa de
crescimento.
Palavras-chave: Efeitos alelopáticos, Urochloa arrecta, capacidade competitiva
Área de Interesse do Simpósio: Ecologia

452
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O aumento das atividades antrópicas nas últimas décadas tem afetado
negativamente os recursos naturais, como por exemplo, contaminação dos ambientes
aquáticos, desmatamentos, contaminação de lençol freático e introdução de espécies
exóticas, resultando na perda de hábitats e redução da biodiversidade (Goulart & Callisto,
2003). A conservação da biodiversidade tornou-se uma questão central na ecologia,
especialmente nas últimas três décadas, quando as extinções em massa começaram a ser
reconhecidas como uma das mais severas mudanças globais (Clavero & Garcıa-Berthou
2005).
A introdução de espécies não-nativas é considerada uma das principais causas de
perda de biodiversidade (Clavero & Garcıa-Berthou 2005), embora as invasões de plantas
possam afetar as comunidades nativas sem causar extinções (p.e. Jäger et al. 2009).
Entender como as espécies de plantas exóticas se tornam invasivas é uma questão
fundamental na ecologia de invasão. As invasões biológicas ocorrem quando uma espécie
é introduzida em uma nova região, coloniza, se estabelece e se propaga em um ambiente
além da sua extensão original (Richardson et al. 2000).
Fatores abióticos e bióticos podem explicar o sucesso das invasões. Entre os
bióticos, o grau de parentesco filogenético de uma espécie com os outros na comunidade
a ser invadida é um importante preditor da capacidade de invasão de espécies exóticas
(Darwin, 1859). O processo de invasão, promovido por ações humanas que eliminam
barreiras naturais, transportam espécies acidentalmente e até mesmo as introduzem
propositalmente (Rejmánek et al. 2005). Espécies exóticas são comumente introduzidas
em ecossistemas de água doce, onde ameaçam vários nativos. Esse fenômeno tornou-se
uma preocupação pois estes ecossistemas são frequentemente mais diversificados em
comparação aos ecossistemas terrestres e marinhos (Balian et al. 2008) e estão passando
por altas taxas de extinção (Jenkins 2003).
Em ambientes aquáticos continentais, macrófitas aquáticas frequentemente se
tornam invasoras e tendem a causar fortes impactos ao criar estruturação física e controlar
aspectos físicos e químicos da água e dos sedimentos (e.g. Caraco e Cole 2002; Perna e
Burrows 2005; Urban et al. 2009; Tall et al. 2011). O termo macrófitas aquáticas constitui

453
Belém (PA), 11 a 13 de dezembro de 2019
uma designação geral para os vegetais que habitam desde brejos até ambientes totalmente
submersos, sendo esta terminologia baseada no contexto ecológico, independentemente,
em primeira instância, de aspectos taxonômicos. As macrófitas aquáticas constituem, em
sua grande maioria, vegetais superiores que retornaram ao ambiente aquático. Dessa
forma, apresentam algumas características de vegetais terrestres e grande capacidade de
adaptação a diferentes tipos de ambientes (Esteves, 1998). Alguns dos impactos
atribuídos a macrófitas invasoras são alterações na disponibilidade de habitat e refúgio
(Wright et al. 2014), dinâmica do oxigênio dissolvido (Caraco e Cole 2002; Perna e
Burrows 2005), ciclagem de nutrientes (Urban et al. 2009; Tall et al. 2011).
As plantas invasoras podem deter características que as auxiliam a se estabelecer
em um ambiente, dentre elas, destacamos os efeitos alelopáticos, que são um tipo de
interações bioquímicas entre diversas espécies de vegetais, considerada uma forma de
adaptação química defensiva, além de ser um fator de estresse ambiental para muitas
espécies (Lovett e Ryuntyu, 1992). O conceito de alelopatia descreve a influência de um
indivíduo sobre o outro, seja prejudicando ou favorecendo o segundo, e sugere que o
efeito é realizado por biomoléculas (denominadas aleloquímicos) produzidas por uma
planta e lançadas no ambiente, seja na fase aquosa do solo ou substrato (Rizvi et al.,
1992). Alterações nestes aspectos geralmente causam impactos diretos e indiretos na
fauna associada às macrófitas (Kornijów et al. 2010; Wright et al. 2014).
Dentre as macrófitas invasoras, a Poaceae Urochloa arrecta (Hack. ex T. Durand
e Schinz) Morrone e Zuloaga (capim-braquiária), é uma planta perene e estolonífera, com
reprodução seminífera pouco significativa, nativa do continente africano, é um exemplo
de macrófita aquática altamente invasora e com ampla abrangência no Brasil. Sua
adaptabilidade a solos úmidos e alagados permitiu a invasão de áreas cultivadas
(Kissmann & Groth, 2007). Há evidências de que, no geral, o enriquecimento por
nutrientes facilita os processos de invasão pelo aumento da disponibilidade de recursos
(Byers 2002; Engelhardt 2011).
Nesse sentido, nossa hipótese é que há interferência pela liberação de substâncias
alelopáticas de U. arrecta no meio em que a Pistia stratiotes, uma macrófita nativa,
cresce. Essa interferência pode ser positiva se a absorção de nutrientes disponíveis pela

454
Belém (PA), 11 a 13 de dezembro de 2019
U. arrecta for eficiente para o desenvolvimento e reprodução de P. stratiotes, porém
podemos encontrar um efeito negativo caso os aleloquímicos de U. arrecta forem
potencialmente ativos agindo na inibição do crescimento de P. stratiotes.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
O experimento foi realizado na Universidade Federal do Pará e teve duração de
13 dias no mês de maio de 2019. As amostras das plantas foram coletadas na Universidade
Federal Rural da Amazônia, Belém, Brasil.
2.2 AMOSTRA
Depois de selecionadas, as plantas Urochloa arrecta, Pistia stratiotes e,
Hydrocleys nymphoides, foram lavadas e separadas dos resíduos do lago. A princípio,
verificou-se o peso inicial (g) de cada indivíduo. Após mensuradas, as plantas foram
colocadas em 3 diferentes tratamentos com P. stratiotes, com 10 réplicas para cada
tratamento, totalizando 30 unidades experimentais. As plantas que constituíram os
tratamentos (Urochloa arrecta e Hydrocleys nymphoides) passaram por um processo de
fragmentação no liquidificador, no qual colocamos 1.515g de planta (tanto invasora,
quanto nativa) para cada 1 litro de água. Depois de processada, 30ml da solução foi
adicionada em recipientes com capacidade de 500ml e completados com água. No
tratamento controle foi colocado somente água no recipiente com a espécie nativa P.
stratiotes.
O primeiro tratamento consistiu em recipientes com solução de Urochloa arrecta,
o segundo com solução de Hydrocleys nymphoides, e o terceiro apenas com água
(controle). Utilizou-se 10 réplicas de cada tratamento para analisar a interferência de
ambos os experimentos com as plantas invasora e nativa, no qual as amostras ficaram
distribuídas aleatoriamente (figura 1). A cada dois dias foram trocadas as soluções por
uma nova solução nas amostras. Após os 13 dias, os indivíduos de P. stratiotes foram
retirados dos recipientes e lavados, e em seguida foi feita uma nova mensuração da
biomassa. Após esse processo, as amostras foram colocadas dentro da estufa em

455
Belém (PA), 11 a 13 de dezembro de 2019
temperatura de 65° graus até atingir o peso constante (peso seco), e foi aferido o diâmetro
posteriormente com paquímetro de cada indivíduo.
Figura 1 – Distribuição das amostras
Análise de dados
Para testar a hipótese de que haveria efeitos alelopáticos no crescimento de Pistia
stratiotes sobre influência da espécie invasora U. arrecta utilizamos a análise de variância
(ANOVA). Foram feitas três ANOVAS utilizado o crescimento (biomassa/peso), o
diâmetro e peso seco como variáveis resposta para cada análise, e os tratamentos como
variáveis preditoras em todas as análises. Para isso, utilizou-se o pacote car (Fox &
Weisberg, 2011) do programa R versão 3.6.1 (R Core Team, 2019), foram testados os
pressupostos através da homogeneidade e normalidade para verificar a distribuição das
amostras. A posteriori foi aplicado o teste de Tukey para evidenciar diferença entre os
tratamentos.
3. RESULTADOS E DISCUSSÃO
A ANOVA considerando a taxa de crescimento de P. stratiotes, apresentou diferença
significativa entre os tratamentos. A hipótese de que haveria um maior efeito alelopático
da espécie invasora Urochloa arrecta sobre o crescimento de Pistia stratiotes não foi
corroborada, pois não houve diferença significativa nos atributos de crescimento e
ausência de efeito alelopáticos. Entretanto, o teste evidenciou diferença significativa entre
o tratamento controle (água) e o tratamento da espécie nativa Hydrocleys. (Figura 2). Já
entre os tratamentos invasora (Urochloa arrecta) e nativa (Hydrocleys) não há diferença
significativa de crescimento (Tukey I-N 0.101). A tabela abaixo (Tabela 1) mostra o

456
Belém (PA), 11 a 13 de dezembro de 2019
desvio padrão, a média dos quadrados e o valor de p dos resultados da análise de variância
(ANOVA), da taxa de crescimento relativa, diâmetro e da biomassa seca.
Tabela 1: Resultados da análise de variância (ANOVA) referente ao crescimento de Pistia
stratiotes.
Taxa de crescimento relativo (g) Diâmetro (mm) Biomassa seca (g)
Desvio padrão 2 2 2
Média 19,549 361,2 1,234
Valor de p 0,020 0,031 0,392
Apesar de não ter corroborado a nossa hipótese, as análises de variância mostraram
que existem diferenças entre os tratamentos e pode-se observa-las nas variáveis respostas
que obtiveram valores significativos na taxa de crescimento e diâmetro dos indivíduos de
P. stratiotes, diferente do resultado da biomassa seca que não apresentou diferença entre
os tratamentos.
Figura 2 - O desenvolvimento de P. stratiotes considerando os atributos aferidos como taxa de
crescimento relativa (a), diâmetro em milímetros (b), biomassa seca dos divíduos (c), sobre
interferência de espécies nativas e invasoras.
Fonte: Rayssa Carmo, 2019

457
Belém (PA), 11 a 13 de dezembro de 2019
De acordo com o resultado, a hipótese de que haveria ações alelopáticas da espécie de
macrófita invasora Urochloa arrecta foi rejeitada, pois a espécie nativa Pistia stratiotes
não apresentou diferença significativa de crescimento no tratamento com a espécie
invasora (U. arrecta), mostrando somente diferença nos tratamentos com nativa
(Hydrocleys) e controle (água) observados na tabela 2 abaixo.
Tabela 2. Crescimento de Pistia stratiotes nos diferentes tratamentos.
Valores de Tukey
Tratamentos Taxa de crescimento Diâmetro Biomassa seca
I-C 0,723 0,916 0,606
N-C 0,019* 0,038* 0,920
N-I 0,101 0,088 0,379
I. Espécie invasora; C – Controle; N – Espécie nativa
Tal fato, mostra que as macrófitas aqui estudadas podem ser ou não
potencialmente causadoras de danos em relação as espécies nativas nos ambientes
aquáticos, devido a mudanças na luz e temperatura que causam aclimatação significativa
nas plantas aquáticas, que têm demonstrado em vários estudos (por exemplo, Sculthorpe,
1967; Barko e Inteligente, 1981; Barko et al., 1982; Middelboe e Markager, 1997;
Hussner et al., 2010, 2011). Mesmo pequenas diferenças, por exemplo, na temperatura da
água, podem afetar a força competitiva das espécies, que tem implicações para avaliar e
prever a invasão por plantas exóticas (Sorte et al., 2013). Devido a isto, infere-se que a
espécie de macrófita aquática U. arrecta não apresentou ações alelopáticas sobre a nativa
P. stratiotes, mas talvez o sucesso dessa invasora nos ambientes aquáticas se deva a sua
capacidade competitiva mais forte por recursos (luz e nutriente) que espécies nativas,
tornando-se mais eficientes na assimilação de nutrientes do ambiente e limitando o
crescimento das nativas.
Sabe-se que espécies invasoras tem um sucesso competitivo em ambientes
aquáticos, isso se deve ao rápido crescimento, alta absorção de nutrientes disponíveis e
elevada produção de biomassa, porém, esse estudo mostrou que a espécie nativa P.
stratiotes relevou resistência biótica quando em ambientes com espécies invasoras. Por
outro lado, podemos sugerir que exista uma relação positiva entre as nativas de um
ambiente (seja por questões adaptativas ou filogenéticas), onde o crescimento de ambas
sejam maior quando estiverem próximas ou que a P.stratiotes tenha vantagem

458
Belém (PA), 11 a 13 de dezembro de 2019
adaptativa/competitiva sobre a Hydroclyes. Dessa forma, sugere-se um aprofundamento
da abordagem para se entender o processo de invasão de macrófitas aquáticas que podem
causar a perda da biodiversidade de ambientes aquáticos.
Sendo assim, atentamos que não se observou a inibição do crescimento de Pistia
stratiotes por substâncias alelopáticas da espécie invasora U. arrecta no experimento, que
a espécie nativa P. stratiotes tem um bom desempenho na presença de outra nativa
Hydrocleys. Logo, a espécie invasora não limitaria o crescimento da nativa por meio de
substâncias alelopáticas que possivelmente liberasse no ambiente, mas sim pela invasora
ter mais sucesso em assimilar os recursos do ambiente assim deixando a nativa sem tanto
recurso para crescer ou sobreviver.
4. CONCLUSÃO
Neste estudo, foi possível verificar que a espécie de macrofita invasora Urochloa arrecta
não possui efeitos alopáticos sobre a espécie nativa Pista stratiotes, mas houve um
crescimento significativa entre a P. stratiotes com a espécie Hydrocleys, uma possível
explicação é que a espécie nativa teria disponibilizado uma maior concentração de
nutrientes em comparação com espécies invasora, porém vale ressaltar que são
necessários experimentos a posteriori para comprovação desses resultados. Porém, apesar
de não ter apresentado inibição no crescimento da nativa por meio de substâncias
alelopáticas, a U.arrecta, faz parte do grupo de macrófita potencialmente causadora de
danos nos ambientes aquáticos, de acordo com outros estudos, portanto é necessário
realizar pesquisas que busquem entender como esta espécie se comporta nestes ambientes
e o que promove o seu sucesso invasivo, para que sejam desenvolvidas técnicas de
controle e manejo nos ambientes aquáticos invadidos.

459
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
CARACO, N.F., J.J. COLE. Contrasting impacts of a native and alien macrophyte on diss
olved oxygen in a large river. Ecological Applications 12.5 (2002): 1496-1509.
CLAVERO, M., E. GARCÍA-BERTHOU. Invasive species are a leading cause of animal
extinctions. Trends in ecology & evolution 20.3 (2005): 110.
CORDANI, U.G., B.B. BRITO NEVES, A. THOMAZ FILHO. Estudo preliminar de
integração do Pré-Cambriano com os eventos tectônicos das bacias sedimentares
brasileiras (Atualização). Bol. Geociênc. Petrobras, Rio de Janeiro 17.1 (2009): 205-219.
DARWIN, C.. On the origin of species, 1859. Routledge, 2004.
ESTEVES, A.J., J. AZEVEDO. Metodologias qualitativas para as ciências sociais. 1998.
GOULART, M.D., M. CALLISTO. Bioindicadores de qualidade de água como
ferramenta em estudos de impacto ambiental. Revista da FAPAM 2.1 (2003): 156-164.
HUSSNER, A. et al. Response capacity to CO2 depletion rather than temperature and
light effects explain the growth success of three alien Hydrocharitaceae compared with
native Myriophyllum triphyllum in New Zealand. Aquatic Botany 120 (2015): 205-211.
JAMES, R. et al. Tendon: biology, biomechanics, repair, growth factors, and evolving
treatment options. The Journal of hand surgery 33.1 (2008): 102-112.
JENKINS, J.C. et al. Estimadores de biomassa em escala nacional para espécies de
árvores dos Estados Unidos. Ciências Florestais 49.1 (2003): 12-35
LOVETT, J., M. RYUNTYU. Allelopathy: broadening the context. Allelopathy.
Springer, Dordrecht, 1992. 11-19.
MICHELAN, T.S. et al. Regeneration and colonization of an invasive macrophyte grass
in response to desiccation. Nat & Conserv 8 (2010): 133-139.
POWELL, K.I., J.M. CHASE, T.M. KNIGHT. A synthesis of plant invasion effects on
biodiversity across spatial scales. American Journal of Botany 98.3 (2011): 539-548.
REJMÁNEK, M., D.M. RICHARDSON, P. PYŠEK. Plant invasions and invasibility of
plant communities. Vegetation ecology 20 (2005): 332-355.
RIIS, T., T.V. MADSEN, R.S.H. SENNELS. Regeneration, colonisation and growth rates
of allofragments in four common stream plants. Aquatic Botany 90.2 (2009): 209-212.
RIZVI, S. J. H. et al. A discipline called allelopathy. Allelopathy. Springer, Dordrecht,
1992. 1-10.
SANTAMARÍA, L. Why are most aquatic plants widely distributed? Dispersal, clonal
growth and small-scale heterogeneity in a stressful environment. Acta oecologica23.3
(2002): 137-154.

460
Belém (PA), 11 a 13 de dezembro de 2019
EFEITOS DO MANEJO DE IMPACTO REDUZIDO SOBRE A ESTRUTURA
HORIZONTAL E DIVERSIDADE FLORÍSTICA DA ÁREA DE CONCESSÃO
FLORESTAL MAMURU-ARAPIUNS, OESTE PARAENSE
Tárita Rafaela Zahluth Ferreira¹, Emil Hernandez², Anderson Borges Serra³
1Graduanda de Engenharia Florestal – UFPA¹. [email protected];
2Professor Doutor, na Universidade Federal do Pará – UFPA². [email protected];
3Professor Mestre, na Universidade Federal do Pará – UFPA³. [email protected].
RESUMO
O objetivo deste trabalho é avaliar a estrutura horizontal e diversidade florística de uma
floresta de fitofisionomia ombrófila densa após a exploração numa área de concessão
florestal. Para isso, foram inventariadas 12 parcelas permanentes de 50X50 na UPA 1,
localizada na UMF 1. Foram medidos todos os indivíduos com DAP (Diâmetro Altura do
Peito - 1,30 m, DAP > 32 cm) no período de pré-exploração (2012) e remedidos no
período pós exploratório (2013 e 2015). A espécie mais frequente nas parcelas da UPA 1
da área de concessão florestal Estadual Mamuru Arapiuns à família Fabaceae; As espécies
que possuem em média os maiores Índice de Valor de Importância são aquela que
obtiveram os resultados de Fr(%), D(%) e Do(%). Os índices de Shannon obtidos em cada
parcela amostrada foi abaixo do valor encontrado para florestas tropicais, indicando que
a área possui uma baixa diversidade florística, já o índice de Equabilidade também foi
considerado baixo, uma vez que o valor varia de 0 a 1, evidenciando a má distribuição
das espécies na área de estudo.
Palavras-chave: Exploração Florestal, Sustentável, Pós-exploração.
Área de Interesse do Simpósio: Ecologia e Biodiversidade.

461
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O manejo florestal vem sendo experimentado na Amazônia em larga escala em
áreas de concessão florestal como forma de viabilizar a sustentabilidade da atividade e
manutenção da qualidade da floresta (REMOR, 2009), favorecendo a estabilização do
setor madeireiro na região e tendo em vista o potencial de diminuição do desmatamento
ilegal e degradação florestal (MORAES, 2014).
Contudo, existem dúvidas sobre a capacidade das técnicas de manejo florestal de
impacto reduzido vem sendo feita sendo realizada de forma que não haja grandes
modificação em sua estrutura florestal e grandes alterações na sua capacidade de
recuperação (resiliência). Uma das formas de avaliar a qualidade/eficiência do manejo é
por meio da diversidade florística, tendo em vista ser imprescindível para o entendimento
sobre o comportamento da floresta após o seu manejo.
As pesquisas sobre a composição florística e a estrutura fitossociológica das
formações florestais são de imprescindível importância, pois oferecem subsídios para a
compreensão da estrutura e da dinâmica destas formações, parâmetros imprescindíveis
para o manejo e regeneração das diferentes comunidades vegetais (CHAVES et al., 2013).
Neste sentido, o objetivo deste trabalho é avaliar a estrutura horizontal e diversidade
florística de uma floresta de fitofisionomia ombrófila densa após a exploração na área de
concessão florestal Mamuru – Arapiuns (PA) - oeste paraense.
2. MATERIAL E METODOLOGIA
2.1 CARACTERIZAÇÃO DA ÁREA DE ESTUDO
O estudo foi realizado a partir do banco de dados coletados pela empresa LN
GUERRA Indústria e Comércio de Madeiras LTDA. A área da propriedade é de
45.657,5373 hectares, sendo 43.371,4675 hectares destinados ao manejo. De acordo com
a Lei 11.284 de 02 março de 2006, a área Reserva Absoluta corresponde a 5% da área de
manejo. A UMF I possui uma tipologia florestal caracterizada por formações de floresta
ombrófila densa e ombrófila densa + aberta (IBGE, 1992; UFRA, 2010), sendo ainda
identificada algumas áreas que já foram exploradas anteriormente e/ou tiveram outro tipo
de uso (MORAES, 2014).

462
Belém (PA), 11 a 13 de dezembro de 2019
2.2 COLETA DE DADOS
Foram coletadas informações da UPA I, de 12 parcelas permanentes de 50X50m
(2.500 m² ou 0,25 ha) totalizando 30.000 m² (3 ha). Dentro de cada parcela foram medidos
todos os indivíduos com DAP (Diâmetro Altura do Peito - 1,30 m, DAP > 32 cm) no
período de pré-exploração (2012) e remedidos no período pós exploratório (2013 e 2015).
2.3 ANÁLISE DE DADOS
Na análise de estrutura horizontal foram calculados os seguintes parâmetros
populacionais por espécie, parcela e tipo de fitofisionomia com auxílio do software Excel
(2016) sugeridos por Souza e Soares (2013): densidade, que é o número de indivíduos;
dominância, que expressa o grau de ocupação da área da parcela por cada espécie;
frequência, que mede a distribuição de cada espécie em termos percentuais, sobre a área;
e índice de valor de importância, o qual é a combinação, em uma única expressão dos
valores relativos de densidade, dominância e frequência. Também foram calculados os
índices de Shannon-Winner (H’) para determinação da diversidade do local e índice de
Equabilidade de Pielou (E’), visando obter o grau de uniformidade da distribuição dos
indivíduos entre as espécies existentes.
3. RESULTADOS E DISCUSSÃO
A área amostrada inicialmente possuía mensurações de DAP (Diâmetro Altura do
Peito - 1,30 m, DAP > 32 cm) de pelo menos 1345 indivíduos no total, porém devido a
diversos erros de anotação e mensuração de dados, foram utilizados sistemas de filtragens
para realização da exclusão dos dados inconsistentes da tabela do inventário. Após a
filtragem, foram restados 765 indivíduos distribuídos em 132 espécies (gráfico 1) e 41
famílias.
A espécie que obteve maior abundância na área foi Pouteria neoxythece sp (31),
seguida respectivamente por Rinorea guianensis (27), Coussarea paniculata (26), Zygia
saman (24), Eschweilera odora (22), Miconia sp (21), Aniba sp (21), Abiu-casca-fina sp
(19), Chrysophyllum pachycarpa (19), Eschweilera subglandulosa (18), Protium
sagotianum (18) (figura 01) correspondendo a 27% de toda a abundância da área.

463
Belém (PA), 11 a 13 de dezembro de 2019
Figura 1 - Gráfico das espécies encontradas na área de manejo
.
Fonte: Ferreira et al., (2019).
A espécie Pouteria neoxythece sp, Aniba sp, Eschweilera florinbunda, foram as
mais frequentes dentre as 12 parcelas amostradas na área. Para melhor ilustração optou-
se por ranquear as dez espécies mais frequentes, seguidos de outros parâmetros de
estrutura da floresta descritos na tabela 4.
Figura 2 - tabela de parâmetros fitossociológicos das dez espécies mais importantes (IVI) na área
de floresta de terra firme no município de Anapú, PA. Onde, N = o nº total de indivíduo
mensurado da espécie; Fai = Frequência absoluta Fr (%) = Frequência relativa da espécie; Doi =
Dominância absoluta; DoR (%) = Dominância Relativa; Abund. = Abundância da espécie
(ind/ha); DR (%) Densidade relativa da espécie; IVI = Índice de Valor de importância.
Espécie N Fai Fr(%) Da Dr(%) Doi Dor(%) IVI Pouteria neoxythece sp 31 0,91 2,47 10,3 4,05 5,22 3,36 9,89
Aniba sp 20 0,83 2,24 6,66 2,61 2,35 1,52 6,38 Eschweilera floribunda 14 0,75 2,02 4,66 1,83 2,79 1,80 5,65
Protium sagotianum 18 0,75 2,02 6 2,35 1,29 0,83 5,21 Abiu-casca-seca sp 12 0,66 1,79 4 1,56 1,41 0,91 4,28
Bagassa sp 16 0,66 1,79 5,33 2,09 2,21 1,42 5,31 Eschweilera odora 22 0,66 1,79 7,33 2,87 2,10 1,35 6,02
Eschweilera subglandulosa 18 0,66 1,79 6 2,35 4,18 2,69 6,84 Neea constricta 16 0,66 1,79 5,33 2,09 2,61 1,68 5,57
Abiu-casca-fina sp 19 0,58 1,57 6,33 2,48 1,06 0,68 4,74 Total parcial 186 7,16 19,32 62 24,31 25,2 16,28 59,9
TOTAL 765 37,0 100 255 100 155 100 300 Fonte: Ferreira et al. (2019).

464
Belém (PA), 11 a 13 de dezembro de 2019
Na Figura 03 são demonstradas as 10 espécies de maior valor de importância antes
da intervenção (2012) e após a intervenção (2013 e 2015) na unidade de produção anual
1. As dez espécies mais importantes na comunidade florestal estudada, nos dois períodos
analisados foram Zygia saman, Pouteria neoxythece sp, Eschweilera subglandulosa
Eschweilera odora, Abiu-casca-fina sp, Aniba sp, Rinorea guianensis, Geissospermum
sericeum, Tachigali myrmecophylla e Bertholletia excelsa.
Figura 3 - Gráfico das espécies de maior IVI.
Fonte: Ferreira et al., (2019)
A manutenção da estrutura da floresta, antes e após a exploração, está relacionada
à boa distribuição horizontal das espécies, principalmente daquelas mais frequentes e
dominantes, que são fundamentais nos processos básicos de equilíbrio da floresta
(OLIVEIRA; AMARAL, 2004; SILVA et al., 2008).
A família Fabaceae apresentou o maior número de espécies na área de estudo (31),
seguida da Sapotaceae (13), Moraceae (10), Apocynaceae (7) e Lecythidaceae (6).
Conjuntamente, essas 5 espécies, somam mais de 50% da riqueza local (figura 2)
corroborando com resultados encontrados por Pinheiro et al. (2007) e Castro et al. (2017),
os quais são estudos também realizados em área de manejo florestal.
De acordo com Escobar (2016) a família Fabaceae é a terceira maior família
botânica existente com cerca de 19.325 espécies, neste estudo ela possuiu a maior
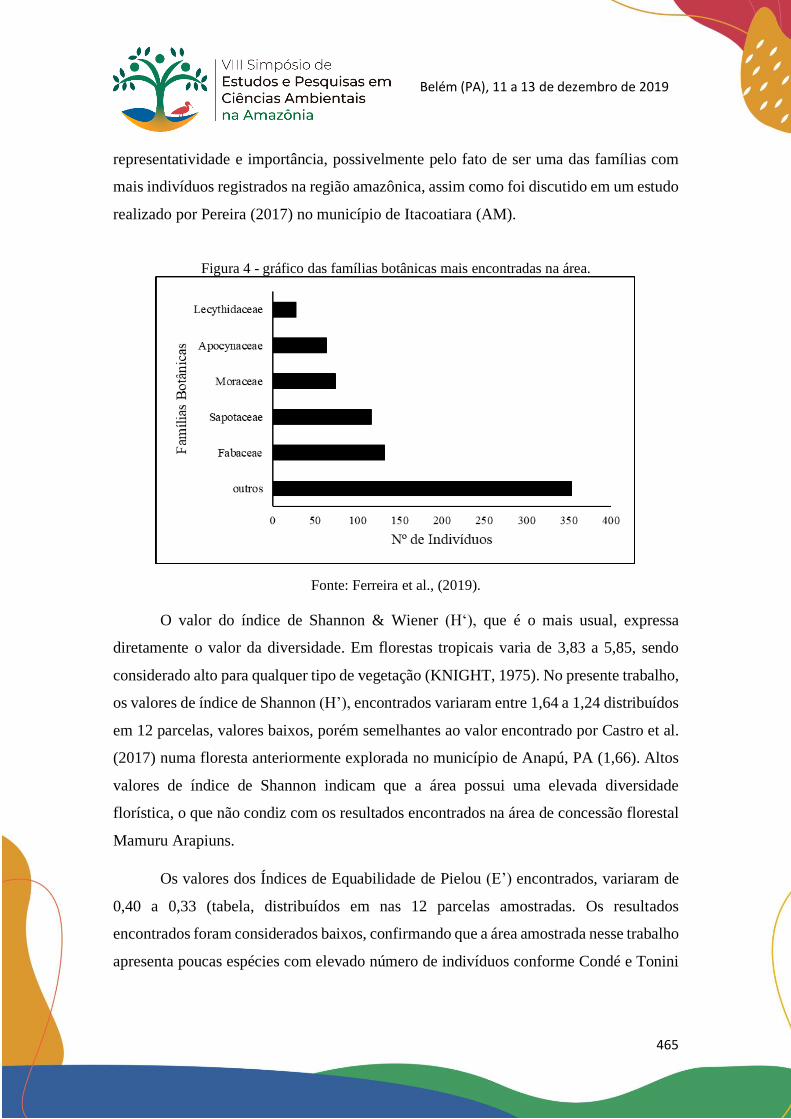
465
Belém (PA), 11 a 13 de dezembro de 2019
representatividade e importância, possivelmente pelo fato de ser uma das famílias com
mais indivíduos registrados na região amazônica, assim como foi discutido em um estudo
realizado por Pereira (2017) no município de Itacoatiara (AM).
Figura 4 - gráfico das famílias botânicas mais encontradas na área.
Fonte: Ferreira et al., (2019).
O valor do índice de Shannon & Wiener (H‘), que é o mais usual, expressa
diretamente o valor da diversidade. Em florestas tropicais varia de 3,83 a 5,85, sendo
considerado alto para qualquer tipo de vegetação (KNIGHT, 1975). No presente trabalho,
os valores de índice de Shannon (H’), encontrados variaram entre 1,64 a 1,24 distribuídos
em 12 parcelas, valores baixos, porém semelhantes ao valor encontrado por Castro et al.
(2017) numa floresta anteriormente explorada no município de Anapú, PA (1,66). Altos
valores de índice de Shannon indicam que a área possui uma elevada diversidade
florística, o que não condiz com os resultados encontrados na área de concessão florestal
Mamuru Arapiuns.
Os valores dos Índices de Equabilidade de Pielou (E’) encontrados, variaram de
0,40 a 0,33 (tabela, distribuídos em nas 12 parcelas amostradas. Os resultados
encontrados foram considerados baixos, confirmando que a área amostrada nesse trabalho
apresenta poucas espécies com elevado número de indivíduos conforme Condé e Tonini

466
Belém (PA), 11 a 13 de dezembro de 2019
(2013). Quando comparado a outras realizados na Amazônia onde os valores encontrados
ficaram entre 0,64 a 0,89 (DIONISIO et al. 2016; CONDÉ E TONINI, 2013)
Um dos fatores agravantes que podem ter corroborado para baixos índices de
diversidade, pode ser o fato de área já ter tido outros tipos de usos anteriormente, tal como
a exploração ilegal madeireira.
Figura 5 - tabela sobre a Diversidade florística da regeneração natural de espécies arbóreas e
arbustivas de uma floresta ombrófila densa de terra firme da Amazônia Central, Amazonas,
Brasil. Onde: H’ e E’ = Índices de Diversidade e Equabilidade de Shannon, respectivamente, Nf
= número de famílias, Ne = número de espécies, Ni = número de indivíduos amostrados.
PARCELA H’ E’ NF NE NI 1 1,39 0,38 15 28 38 2 1,36 0,36 16 28 42 3 1,24 0,36 15 21 31 4 1,51 0,37 19 36 55 5 1,39 0,37 14 28 39 6 1,48 0,34 19 38 74 7 1,30 0,34 11 23 42 8 1,64 0,40 25 53 80 9 1,57 0,35 24 48 81
10 1,55 0,35 25 43 75 11 1,55 0,33 20 49 109 12 1,62 0,35 27 50 99
TOTAL
765 Fonte: Ferreira et al., (2019).
4. CONCLUSÃO
Sabe-se que a área de concessão Mamuru-Arapiuns, além de ter sido explorada
comercialmente, também teve outros tipos de exploração anteriormente, contribuindo
para que a área obtivesse resultados pouco satisfatórios nos parâmetros fitossociológicos,
além de também ter se tornado uma área de baixa diversidade e distribuição desigual das
espécies encontradas, mostrando que o poder de resiliência do território tem sido
grandemente afetado pelas perturbações frequentes na floresta.

467
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
AMARAL, I. L.; MATOS, F. D. A.; LIMA, J. Composição florística e estrutural de
um hectare de floresta densa de terra firme no Rio Uatumã, Amazônia, Brasil. Acta
Amazonica, Manaus, v.30, n. 3, p. 377-392, 2000.
CASTRO, A. C. J.; RODRIGUES, A. R. M.; COELHO, F. A.; SOUSA, J. B. S.;
NASCIMENTO, A. M.; RUSCHEL, A. R. Caracterização florística e análise
fitossociológica da regeneração natural em uma área de manejo florestal no
município de anapú – pa. Anais Trabalhos Completos – Repositório Alice. Belém. 2017
CONDÉ, T. M.; TONINI, H. Fitossociologia de uma Floresta Ombrófila Densa na
Amazônia Setentrional, Roraima, Brasil. Univerisdade Estadual de Roraima – UERR,
Revista Acta Amaônica, Vol. 43 (3) 247 – 260, 2013.
CHAVES, A. D. C. G.; SANTOS, R. M. S.; SANTOS, J. O.; FERNANDES, A. A.;
MARACAJÁ, P. B. A importância dos levantamentos florísticos e fitossociológicos
para a conservação e preservação das florestas. R. Agropecuária Científica do
Semiárido, Campina Grande – Paraíba, v 09, n.2, 2013.
DIONISIO, L. F.; BONFIM FILHO, O. S.; CRIVELLI, B. R. S.; GOMES, J. P.;
OLIVEIRA, M. H. S.; CARVALHO, J. O. P. Importância de um Fragmento de
Floresta Ombrófila Densa no Estado de Roraima, Brasil. Revista Agro@mbiente, v.
10, n. 3, p. 243-252. Centro de Ciências Agrárias – Universidade Federal de Roraima,
Boa Vista, RR. 2016.
ESCOBAR, F. B. PADRÕES ESTRUTURAIS FITOSSOCIOLÓGICOS E ÁREAS
POTENCIAIS PARA CORREDORES ECOLÓGICOS NA BACIA DO
PURAQUEQUARA MANAUS – AM. Mestrado (Mestrado em Biodiversidade e
Conservação). Universidade Federal do Amazonas. Manaus, AM. 2016.
KNIGHT, D. H. A phytosociological analysis of species-rich tropical forest on Barro
Colorado Island, Panamá. Ecological Monographs, 45: 259-284, 1975.
MORAES, I. S. Quantificação e avaliação de abertura no dossel em áreas de
concessões florestais: Mamuru-Arapiuns-PA. (Tese de Doutorado). Universidade
Federal do Pará. Belém, PA. 2014.
OLIVEIRA, M. V. N.; BRAZ, E. M. Estudo da dinâmica da floresta manejada no
projeto de manejo florestal comunitário do PC Pedro Peixoto na Amazônia
Ocidental. Acta Amazônica, Manaus, v. 36, n. 2, p. 177-182, 2006.
PEREIRA, L. F, B. ANÁLISE DA FITOSSOCIOLOGIA DE UMA ÁREA DE
TERRA FIRME NO RIO MAUÉS MIRIM, MUNICÍPIO DE MAUÉS – AM.
Monografia (Bacharelado em Engenharia florestal). Universidade do Estado do
Amazônas. Itacoatiara, AM. 2017.
PINHEIRO, K. A. O.; CARVALHO, J. O. P.; QUANZ, B.; FRANCEZ, L. M. B.;
SCHWARTZ, G. Fitossociologia de uma área de preservação permanente no leste da
Amazônia: indicação de espécies para recuperação de áreas alteradas. FLORESTA,
Curitiba, PR, v. 37, n. 2, mai./ago. 2007.

468
Belém (PA), 11 a 13 de dezembro de 2019
SILVA, K. E.; MATOS, F. D. A.; FERREIRA, M. M. Composição florística e
fitossociologia de espécies arbóreas do Parque Fenológico da Embrapa Amazônia
Ocidental. Acta Amazônica, Manaus, vol. 38, n. 2, p. 213-222, 2008.
SOUZA, A. L.; SOARES, C. P. B. Florestas nativas: estrutura, dinâmica e manejo.
Universidade Federal de Viçosa. Viçosa – Minas Gerais, v. 1, p. 38-39, 2013.
UFRA. Relatório Ambiental preliminar das áreas de concessão florestal do Conjunto
de Glebas Mamuru-Arapiuns. Belém: UFRA, 2010. 123 p.

469
Belém (PA), 11 a 13 de dezembro de 2019
FENODINÂMICA DA POPULAÇÃO DE Euxylophora paraensis Huber EM
FRAGMENTO FLORESTAL NO MUNICÍPIO DE PARAGOMINAS-PARÁ
Joyce Ananda Paixão Duarte¹; Gracialda Costa Ferreira²; Jakeline Prata de Assis Pires³;
José Natalino Macedo Silva4; Raphael Lobato Prado Neves5
1 Doutoranda, Programa de Pós Graduação em Ciências Florestais, Universidade
Federal Rural da Amazônia. [email protected] 2 Professora Dra. do Instituto de Ciências Agrárias; Universidade Federal Rural da
Amazônia, [email protected] 3 Professora Assistente I, Departamento de Biologia, Pontifícia Universidade católica do
Rio de Janeiro, [email protected] 4 Professor Dr. do Programa de Pós Graduação em Ciências Florestais, Universidade
Federal Rural da Amazônia. [email protected] 4 Doutorando, Programa de Pós Graduação em Ciências Florestais, Universidade
Federal Rural da Amazônia. [email protected]
RESUMO
Na Amazônia brasileira, os fragmentos florestais têm aumentado de forma alarmante,
levando muitas espécies à extinção local. A Euxylophora paraensis Huber é endêmica do
norte do Brasil, e apresenta ocorrência natural restrita aos estados do Amazonas, Pará,
Tocantins e Maranhão. A limitação demográfica, a intensa exploração madeireira e o
aumento do desmatamento fizeram com que a espécie fosse incluída na lista de espécies
ameaçadas em extinção na categoria “Criticamente em Perigo”. O avanço da
fragmentação e a ameaça de extinção de E. paraensis, motivou este trabalho, cujo
objetivo é avaliar a fenologia em um fragmento florestal no município de Paragominas -
Pará, tendo em vista sua resiliência e manutenção de populações em florestas próximas
às áreas de mineração. Foram feitas observações das fenofases vegetativas e reprodutivas
mensalmente durante o período de cinco anos (2013 – 2017) em 37 indivíduos, sendo: 25
matrizes em área de borda, cinco matrizes em área intermediária e sete matrizes no
interior do fragmento. Foram calculados mensalmente os índices de atividade e de
intensidade das fenofases vegetativas e reprodutivas. Para relacionar os fatores climáticos
com as fenofases vegetativas e reprodutivas foi utilizada a correlação de Spearman. As
fenofases reprodutivas são anuais. A população apresentou botão floral, flor e fruto uma
vez ao ano em todo período observado. A correlação das fenofases reprodutivas com as
variáveis climáticas foi significativa, sendo a precipitação a única variável correlacionada
negativamente, onde a emissão de botões florais, a abertura das flores e a presença de
frutos iniciam com o final da estação chuvosa.
Palavras-chave: Pau-amarelo. Fragmentação florestal. Amazônia.
Área de Interesse do Simpósio: Ecologia e Biodiversidade

470
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
As florestas amazônicas brasileiras estão sendo fragmentadas em grande escala e
velocidade (HADDAD et al., 2015). Em apenas dois anos (2000 a 2002), os fragmentos
florestais aumentaram 64% na Amazônia brasileira (BROADBENT et al., 2008). Apesar
disso, os fragmentos florestais dispersos pela paisagem, tem se tornado uma das formas
de conservação disponíveis atualmente, mesmo sendo áreas sensíveis aos efeitos da
fragmentação e mais suscetíveis à degradação (MACHADO et al., 2010).
Um dos principais efeitos causados pela fragmentação de áreas naturais é a
formação de bordas (LAURANCE et al., 2018). Próximo das bordas, os fragmentos
florestais apresentam características peculiares, como maiores temperaturas, menor
umidade no solo, maior luminosidade e maior incidência de vento (LAURANCE et al.,
2018). As árvores que permanecem às margens de habitats fragmentados podem ter
diferentes respostas fenológicas que variam com as espécies, dependendo da sua ecologia,
condições bióticas e abióticas e histórico de vida (LIMA et al., 2018).
Além do efeito de borda causado pela fragmentação, mudanças nos fatores
climáticos como temperatura, radiação e precipitação, também têm forte influência na
fenologia das espécies lenhosas (KHAROUBA et al., 2017). A mudança na resposta
reprodutiva das espécies florestais, frente às alterações climáticas e ambientais, é um dos
fatores de grande relevância a ser observado, pois compromete diversos serviços
ambientais como polinização, dispersão de sementes e disponibilidade de frutos para a
fauna, ocasionando um efeito cascata que pode levar diversas espécies à extinção
(ALBERTI; MORELLATO, 2010) como é o caso da Euxylophora paraenses Huber.
Conhecida popularmente como pau-amarelo, a E. paraensis é a única espécie do
gênero Euxylophora da família Rutaceae (PIRANI, 2016), sendo endêmica do Brasil com
ocorrência restrita apenas em quatro estados (Amazonas, Pará, Tocantins e Maranhão)
(PIRANI, 2016). Devido ter sido intensamente explorada, sem qualquer planejamento, do
século XIX até o final da década de 1990, teve suas populações naturais drasticamente
reduzidas nos últimos anos (BRANDÃO et al., 2018), levando a sua inclusão na lista de
espécies ameaçadas de extinção, na categoria “Vulnerável” em 1998 e, em 2014 foi
reclassificada como “Criticamente em Perigo” (BRASIL, 2014).

471
Belém (PA), 11 a 13 de dezembro de 2019
Identificar as consequências dos efeitos de borda e a influência dos fatores
climáticos sobre a fenodinâmica de espécies arbóreas é fundamental para compreender as
respostas da espécie e vegetação em relação às mudanças climáticas. Além disso, pode-
se promover o manejo destas espécies de acordo com as características ambientais na qual
estão inseridas, assim como serem utilizadas como espécies bioindicadoras para estudos
de impactos climáticos e auxiliar na perpetuação das espécies em florestas fragmentadas
principalmente àquelas que já estão em risco de extinção como a Euxylophora paraensis
Huber (MAUÉS; OLIVEIRA, 2010; FREITAS, et al., 2013).
Desta forma o presente estudo buscou avaliar a influência de borda e as variações
climáticas na fenodinâmica vegetativa e reprodutiva de Euxylophora paraensis em
matrizes existentes em um fragmento florestal localizado em área de mineração de
bauxita no município de Paragominas-Pará.
2. MATERIAL E MÉTODOS
2.1. ÁREA DE ESTUDO
O estudo foi conduzido em um fragmento florestal, localizado em uma área com
atividade minerária, denominada Platô Miltônia III, pertencente à empresa Mineração
Paragominas S.A. que, atualmente, está sob a administração do grupo Hydro. A área está
localizada no município de Paragominas na mesorregião do Sudeste Paraense, nas
coordenadas 3°15'50"S e 47°44'19"W (Figura 2). A mineração de bauxita em
Paragominas fica a 64 quilômetros da área urbana e 350 km da capital Belém.

472
Belém (PA), 11 a 13 de dezembro de 2019
Figura 1- Mapa de localização da área e de estudo e das matrizes de Euxylophora
paraensis Huber no fragmento florestal no município de Paragominas - Pará.
Fonte: Joyce Duarte, 2019
A região apresenta clima equatorial do tipo quente e úmido, temperatura média
anual de 26,3ºC, umidade relativa do ar média de 81% (BASTOS et al. 2006). A
pluviosidade média anual é de 1.800 milímetros, com o período mais chuvoso, de
dezembro a maio e o mais seco de junho a novembro evidenciando o período de estiagem
(BASTOS et al., 2006). O relevo é plano a suavemente ondulado, com altitude média de
200 metros em relação ao nível do mar. O solo de Paragominas é do tipo latossolo amarelo
distrófico, com alto grau de intemperismo, profundos, ácidos e ricos em alumínio
(RODRIGUES et. al. 2003).
2.2 COLETA DE DADOS
Os dados utilizados foram obtidos por meio do projeto “Estudos para o
monitoramento de áreas degradadas e, de eventos fenológicos de espécies ameaçadas de
extinção nas áreas da Mineração Paragominas S.A., Paragominas-PA” que teve início em
2012 pela Universidade Federal Rural da Amazônia em convênio com a empresa
Mineração Paragominas S.A.

473
Belém (PA), 11 a 13 de dezembro de 2019
As observações fenológicas foram realizadas mensalmente durante cinco anos
(2013 – 2017) em 37 matrizes de E. paraensis, sendo: 25 matrizes em área de borda, cinco
matrizes em área intermediária e sete matrizes no interior do fragmento.
Para avaliar a fenologia de E. paraensis em diferentes ambientes do fragmento,
foram obtidas as distâncias a partir das estradas da mineração, sendo: áreas de bordas o
perímetro de 0 – 99 metros do fragmento, as áreas intermediárias de 100 – 199 metros e
o interior do fragmento áreas ≥ 200 metros da borda da floresta. Cada indivíduo foi
marcado com placa de alumínio e, obtida sua coordenada geográfica. Respeitou-se a regra
da distância mínima de 100m entre os indivíduos recomendado por Higa & Duque Silva
(2006). A distância de cada indivíduo de E. paraensis em relação a borda do fragmento
foi obtida utilizando a ferramenta measure do programa Programa ArcGis.
Foram registradas as fenofases vegetativas e reprodutivas da população de E.
paraensis. A fenofase vegetativa (foliação) foi caracterizada por meio da presença de
folhas novas e maduras. A fenofase reprodutiva (floração) foi subdividida em botões
florais e flores abertas e para frutificação foi considerada os frutos imaturos, frutos
maduros até o final da dispersão das sementes. As observações foram realizadas sempre
pela mesma equipe com o auxílio de binóculo.
Foram coletados dados de atividade (presença e ausência) e intensidade de
Fournier (1974). A intensidade de Fournier (1974) para cada fenofase atribui valor 0 para
ausência da fenofase e valores de 1 a 4 em intervalos de 25% (1 = 1-25%; 2 = 26-50%; 3
= 51-75%; 4 = 76-100%) para presença da fenofase.
A determinação dos padrões de produção e queda de folhas seguiu a classificação
proposta por Rivera et al., (2002): decídua, semidecídua e perenifólia (sempre-verde). A
periodicidade dos eventos reprodutivos foi baseada segundo Newstrom et al., (1994) que
considerou quatro classes: contínua, sub anual, anual e supra anual.
Dados climáticos como: precipitação (total, maior precipitação do mês, número
de dias com chuva), temperatura (média, máxima e mínima absoluta), e radiação foram
obtidos juntos ao Instituto Nacional de Meteorologia (INMET) para estação
meteorológica automática localizada no município de Paragominas (Latitude 03°00'37''
S/ Longitude 047°20'35'' W) com 113m de altitude. Dados de fotoperíodo foram
disponibilizados pelo Observatório Nacional do Ministério da Ciência, Tecnologia,
Inovações e Comunicações.

474
Belém (PA), 11 a 13 de dezembro de 2019
2.3 ANÁLISE DE DADOS
Para caracterizar a fenodinâmica da população de E. paraensis, foram calculados
mensalmente os índices de atividade (BENCKE; MORELLATO 2002) e intensidade
(FOURNIER, 1974) de cada fenofase vegetativa e reprodutiva. Foram construídos
fenogramas para caracterizar graficamente as fenofases vegetativas e reprodutivas por
ambiente.
Para relacionar os fatores abióticos (precipitação, temperatura média, horas com
sol e radiação) com as fenofases (foliação, botão floral, flor e frutificação) foi utilizada a
correlação de Spearman que é recomendada para dados que não apresentam distribuição
normal (ZAR, 1999). Foi utilizado o programa estatístico Past 2.16.
3. RESULTADOS E DISCUSSÃO
O padrão vegetativo é sempre verde ou perenifólio, em que a queda e o brotamento
de folhas ocorreram simultaneamente. Porém, houve grande perda da proporção de folhas
nos períodos mais chuvosos, mas em nenhum momento as copas perderam totalmente
suas folhas independente do ambiente que esteja (Figura 2A; Figura 2E). Houve uma
intensa perda foliar (75%) da população, em fevereiro de 2015, porém no mês seguinte
as matrizes apresentaram alta produção foliar. A perda foliar foi mais intensa nas matrizes
localizadas na borda e no interior do fragmento em relação às do ambiente intermediário.
A atividade para produção de folhas foi correlacionada com fatores climáticos, a
intensidade para a foliação apresentou correlação baixa e positiva com a temperatura
média, radiação, horas de sol e correlação baixa e negativa com a precipitação (Tabela 1).
O padrão fenológico reprodutivo (floração e frutificação) de E. paraensis é anual,
observou-se que população apresentou botão floral, flor e fruto uma vez ao ano não
ultrapassando 12 meses de duração.
Os maiores picos de atividade e intensidade ocorreram nos períodos menos
chuvosos e nas matrizes em áreas intermediárias (Figura 2B; Figura 2F). A presença de
botões florais durou de três a seis meses na maioria dos indivíduos observados. Nos meses
de junho e julho de 2015 houve emissão dos botões florais em 100% dos indivíduos
observados, ocorrendo nos três ambientes observados e em julho a maior intensidade de
emissões de botões florais. A atividade e intensidade da emissão de botão floral teve

475
Belém (PA), 11 a 13 de dezembro de 2019
correlação significativa com todas as variáveis climáticas, com exceção apenas da
temperatura. A precipitação e as horas com sol foram as variáveis que apresentaram maior
correlação, porém negativa para esta fenofase (Tabela 1).
Figura 2: Intensidade e atividades fenológicas de Euxylophora paraensis Huber em
áreas de borda, intermediária e interior de fragmento florestal localizado no município
de Paragominas-PA. Foliação (A; E), Botão floral (B; F), Flor (C; G) e frutos (D; H)
Fonte: Joyce Duarte, 2019.
Na Amazônia, estudos mostraram que algumas espécies podem ser fortemente
afetadas por alterações nas condições abióticas (e.g. LEPSCH-CUNHA; MORI, 1999;

476
Belém (PA), 11 a 13 de dezembro de 2019
PEREIRA; TONINI, 2012; FREITAS et al., 2013). Por exemplo, a dispersão dos frutos
de Carapa guianensis Aubl. (Meliaceae) correlaciona-se fortemente com a precipitação,
de forma que, quanto maior a precipitação, maior o número de árvores que dispersam
frutos (PEREIRA & TONINI, 2012). Já para Couratari multiflora (Sm.) Eyma
(Lecythidaceae) foi verificado que a espécie tem como estratégia florescer e frutificar
durante a estação seca por conta da facilidade de dispersar suas sementes aladas
(LEPSCH-CUNHA; MORI, 1999).
A abertura das flores iniciou sempre no final da estação mais chuvosa. Nos
períodos com maior precipitação, a presença de flores foi reduzida. Em junho de 2015
ocorreu a floração em todas as matrizes, independente do ambiente, com alta intensidade
no mês de agosto em todos os ambientes (Figura 2C; Figura 2G). O pico de atividade e
intensidade, durante os cinco anos de observação (2013 a 2017), ocorreu em junho de
2015 em todas as matrizes, independente do ambiente com alta intensidade em 2015. A
correlação da atividade e intensidade da floração foi significativa com todas as variáveis
climáticas avaliadas. A precipitação teve alta correlação negativa na emissão de flores,
enquanto que a radiação foi a única variável que se correlacionou positivamente (Tabela
1).
Todos os indivíduos frutificaram pelo menos uma vez em todos os anos
observados, porém de forma irregular, variando entre os meses. O pico de atividade
ocorreu, na maioria dos anos, de julho a setembro, porém o período de frutificação
estendeu-se por até 11 meses, mesmo com baixa produção. A precipitação foi a única
variável que apresentou correlação negativa com a produção de frutos (Tabela 1).

477
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1: Valores para a correlação de Spearman obtidos entre os fatores climáticos (temperatura
média, radiação, horas com sol e precipitação) e as fenofases de foliação, botão floral, flor e fruto
da população de Euxylophora paraensis Huber em áreas sob mineração de bauxita, Paragominas,
Pará.
Atividade Fenofase Temperatura Radiação Horas com sol Precipitação Foliação 0,31*** 0,41*** 0,53*** -0,23*** Botão floral -0,01ns 0,46*** -0,65*** -0,56*** Flor -0,23*** 0,36*** -0,56*** -0,64*** Fruto 0,22*** 0,25*** 0,21*** -0,16***
Intensidade Foliação 0,26*** 0,25*** 0,07*** -0,29*** Botão floral -0,09** 0,47*** -0,68*** -0,63*** Flor -0,19*** 0,43*** -0,46*** -0,66*** Fruto 0,45*** 0,14*** 0,46*** -0,05ns
Nota: ns: não significativo; *p<0,05; **p<0,01; ***p<0,001. Fonte: Joyce Duarte, 2019
A mudança na resposta reprodutiva das espécies florestais, frente às mudanças
ambientais e climatológicas, é um dos fatores de grande relevância a ser observado, pois
este compromete diversos outros serviços ambientais, como polinização, dispersão de
sementes e disponibilidade de frutos para a fauna, o que pode interferir na distribuição
espacial e na dinâmica populacional das plantas, ocasionando um efeito cascata até levar
diversas espécies à extinção (ALBERTI & MORELLATO 2010; MAUÉS & OLIVEIRA,
2010; PIRES et al., 2018).
4. CONCLUSÃO
A população de Euxylophora paraensis teve seu padrão reprodutivo
influenciado pelas variáveis climáticas nas diferentes áreas de borda. A emissão de botões
florais, a abertura das flores e a presença de frutos iniciam com o final da estação chuvosa
e o maior pico apresentado ocorreu em áreas intermediárias. Não houve influência da área
de borda e das variáveis climáticas no padrão vegetativo de Euxylophora paraensis.

478
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALBERTI, L.F.; MORELLATO, P.C. Variation on fruit production of Nectandra
megapotamica (Lauraceae) trees on the edge and interior of a semideciduous forest - a
case study. Naturalia 33: 57-68, 2010.
BASTOS, T.; SILVA, G.; PACHECO, N.; & FIGUEIREDO, R. (2006). (E. A. Oriental,
Editor). Disponível em: <https://www.embrapa.br/busca-de-publicacoes/-
/publicacao/579613/informacoes-agroclimaticas-do-municipio-de-paragominas-para-o-
planejamento-agricola> . Acesso em 31 de fevereiro de 2018.
BENCKE, C. S., & MORELLATO, P. C. Comparação de dois métodos de avaliação da
fenologia de plantas, sua interpretação e representação. Brasileira Botânica, v.25, n.3,
p.269-275, 2002.
BROADBENT, E. N., ASNER, G. P., KELLER, M., KNAPP, D. E., OLIVEIRA, P. J.,
& SILVA, J. N. Forest fragmentation and edge effects from deforestation and selective
logging in the Brazilian Amazon. Biological Conservation, 141(7), 1745-1757, 2008.
FORTUNATO, M. M.; QUIRINO, Z. M. Efeitos da fragmentação na fenologia
reprodutiva de espécies arbóreas presentes em borda e interior de Mata Atlântica
Paraibana. Rodriguésia, v.67, n.3, p.603-614. 2016.
FOURNIER, L.A. Un método cuantitativo para la medición de características fenológicas
en árboles. Turrialba, 24, p.422–423, 1974.
FREITAS, J. L.; SANTOS, A. C.; SILVA, R. B.; RABELO, F. G.; SANTOS, E. S.;
SILVA, T. L. Fenologia reprodutiva da espécie Carapa guianensis Aubl. (Andirobeira)
em ecossistemas de terra firme e várzea, Amapá, Brasil. Biota Amazônia, p.31-38, 2013.
HIGA, A. R.; DUQUE SILVA, L. 2006. Certificação da Produção de Sementes e Mudas
de Espécies Florestais Nativas. In: SILVA, L.D. & HIGA, A.R. Pomar de espécies
florestais nativas. Curitiba: FUPEF. p.65-77.
HADDAD, N. M.; BRUDVIG, L. A.; CLOBERT, J.; DAVIES, K. F.; GONZALEZ, A.;
HOLT, R. D.; LOVEJOY, T. E.; SEXTON, J. O.; AUSTIN, M. P.; COLLINS, C. D.;
COOK, W. M.; DAMSCHEN, E. I.; EWERS, R. M.; FOSTER, B. L.; JENKINS, C.N.;
KING, A. J.; LAURANCE, W. F.; LEVEY, D. J.; MARGULES, C. R.; MELBOURNE,
B. A.; NICHOLLS, A. O.; ORROCK, J. L.; SONG, D.; TOWNSHEND, J.R. Habitat
fragmentation and its lasting impact on Earth’s ecosystems, Science Advances, v.1, n.2,
p.1-9, 2015.
KHAROUBA, H. M.; EHRLÉN, J.; GELMAN, A.; BOLMGREN, K.; ALLEN, J. M.;,
TRAVERS, S. E.; WOLKOVICH, E. M. Global shifts in the phenological synchrony of
species interactions over recent decades. Proceedings of the National Academy of
Sciences of the United States of America, v.115, n.20, p. 5211-5216, 2018.
LAURANCE, W. F.; CAMARGO, J. L. C.; FEARNSIDE, P. M.; LOVEJOY, T. E.;
WILLIAMSON, G. B.; MESQUITA, R. C. G.; MEYER, C. F. J.; BOBROWIEC, P.
E. D.; LAURANCE, S. G. W. An Amazonian rainforest and its fragments as a laboratory
of global change. Biological Reviews, v.93, p. 223 – 247, 2018.

479
Belém (PA), 11 a 13 de dezembro de 2019
LEPSCH-CUNHA, N.; MORI, S. A. Reproductive phenology and mating potential in a
low density tree population of Couratari multiflora (Lecythidaceae) in central Amazonia.
Journal of Tropical Ecology, v.15, p.97–121, 1999.
LIMA, J. M. T.; CARUSO, N. M.; CLUGSTON, J.; KAINER, K. A. Landscape change
alters reproductive phenology and sex expression in Attalea palms (Arecaceae) of
southwestern Amazonia. Plant Ecology, v.219, n.10, p.1225–1245, 2018.
MACHADO, E. L. M.; GONZAGA, A. P. D.; CARVALHO, W. A. C.; SOUZA, J. S.;
HIGUCHI, P.; SANTOS, R. M.; SILVA, A. C.; OLIVEIRA FILHO, A. T. Flutuações
temporais nos padrões de distribuição Diamétrica da comunidade arbóreo-arbustivo e de
15 populações em um fragmento florestal. Revista Árvore, v.34, n.4, p.723-732, 2010.
MAUÉS, M. M; OLIVEIRA, P. E. A. M. Consequências da fragmentação do habitat na
ecologia reprodutiva de espécies arbóreas em florestas tropicais, com ênfase na
Amazônia. Oecologia Australis, 14(1), 238-250, 2010.
NEWSTROM L. E.; FRANKIE G. W.; BAKER G. A new classification for plant
phenology based on flowering patterns in lowland tropical rain forest trees at La Selva,
Costa Rica. Biotropica, v.26, p.141–159, 1994.
PEREIRA, M. R.; TONINI, H. Fenologia da andiroba (Carapa guianensis, Aubl.,
(Meliaceae) no sul. Ciência Florestal, v.22, n.1, p.47-58, 2012.
PIRANI, J. R. Euxylophora. Lista de Espécies da Flora do Brasil, 2016. Jardim Botânico
do Rio de Janeiro. Online. Available from: <http://
floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB621>. Acesso em: Agosto de 2019.
PIRES, J. P.; MARINO, N. A.; SILVA, A. G.; RODRIGUES, P. J.; FREITAS, L. Tree
Community Phenodynamics and Its Relationship with Climatic Conditions in a Lowland
Tropical Rainforest. Forests, v.9, n.3, p.1-18. 2018.
VOGADO, N. O.; CAMARGO, M. G. G.; LOCOSSELLI, G. M.; MORELLATO, L. P.
C. Edge effects on the phenology of the guamirim, Myrcia guianensis (Myrtaceae), a
cerrado tree, Brazil. Tropical Conservation Science, v.9, n.1, p.291-312, 2016.

480
Belém (PA), 11 a 13 de dezembro de 2019
FITOSSOCIOLOGIA E CARACTERIZAÇÃO DE ESPÉCIES EM ÁREA DE
VEGETAÇÃO SECUNDÁRIA NO ECOSSISTEMA DE CAPOEIRA DO
MUNICÍPIO DE BRAGANÇA (PA)
Leonardo Correa Cordeiro1; Bruno Mateus Santiago da Silva2; Mônica Trindade Abreu
de Gusmão³
1 Graduando em Engenharia ambiental e Energias Renováveis. Universidade Federal
Rural da Amazônia. [email protected].
2 Graduando em Engenharia Ambiental e Energias Renováveis. Universidade Federal
Rural da Amazônia. [email protected].
3Professora Dra. da Universidade Federal Rural da Amazônia.
RESUMO
O ecossistema de vegetação secundária, conhecido por capoeira, pode ser caracterizado
como área impactada por atividades antrópicas formadas por florestas secundárias
importantes para a manutenção desse ecossistema, visto que é comum ser relacionada à
regeneração de áreas impactadas. O estudo da fitossociologia das espécies vegetais do
ecossistema capoeira auxilia a compreensão do comportamento ecofitossociológico dessa
área. Neste contexto, objetivou-se caracterizar uma área de capoeira para correlacionar as
espécies com a ecologia do ambiente no município de Bragança (PA). Dentre os
parâmetros fitossociológicos foram avaliadas as densidades absoluta e relativa, a
frequência relativa, a área basal e o diâmetro à altura do peito (DAP). Para a identificação
das espécies foram utilizados como base a lista de espécies ameaçadas do website “The IUCN Red List of Threatened Species”. Foram avaliadas cinco áreas de capoeira para
serem utilizadas pesquisa, sendo cada área delimitada em 10 m x 10 m. As espécies
consideradas como raras obtiveram frequências muito baixas em relação as demais
espécies, suas frequências juntas somam 17,65%. As espécies Astrocaryum
aculeatissimum Schott. e Sansevieria trifasciata Wild. com 35,3% e 47,1% de frequências
foram classificadas como Ocasional e Comum. Os impactos causados pelo homem na
área de coleta de dados não foram tão significativos a ponto de inibir o desenvolvimento
das espécies vegetais descritas e foram suficientes para a caracterização fitossociologica
da região. Mesmo com seis tipos de espécies vegetais no ambiente podem-se compreender
suas importâncias para a preservação do ambiente como seus usos para o ser humano.
Palavras-chave: Florística. Importância Ambiental. Nordeste Paraense.
Área de Interesse do Simpósio: Ecologia e Biodiversidade.

481
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O levantamento de espécies em uma região pode ser feito através de estudos
fitossociologicos e florísticos, visto que essas ferramentas possibilitam a compreensão da
variabilidade de determinadas espécies na ecologia do ambiente (SILVA, 2017). O meio
ambiente está sujeito a impactos antrópicos, sendo que esses não só impactam o meio
físico, como as espécies vegetais presentes. Como áreas impactadas têm-se as capoeiras,
que podem ser caracterizadas como florestas secundárias com importância ecológica para
o crescimento das espécies vegetais, para o acúmulo de biomassa e para a contribuição
da manutenção do ecossistema.
O fato de ser uma vegetação secundária nos permite inferir que é normal encontrar
uma área de capoeira relacionada a diversos estágios de regeneração de área degradados
(THOMAZ, 2005). Nesse tipo de área as espécies locais estão se regenerando
naturalmente nos agroecossistemas (VIEIRA; PROCTOR, 2007).
Segundo Pereira et al. (2009), verificou-se que por meio dos parâmetros
fitossociológicos, dentre eles a frequência de indivíduos, o diâmetro à altura do peito
(DAP), a densidade relativa e a área basal, diferenciar áreas naturais e antropizadas. Este
resultado ressalta a importância da avaliação dos parâmetros de análise fitossociológicos
para os estudos de ecossistemas amazônicos.
A existência de degradação vegetal em áreas de capoeiras pode ser estudada por
meio dos estudos fitossociológicos e os resultados são explicados através de variáveis
quantitativas e qualitativas. As variáveis qualitativas podem ser compreendidas como as
responsáveis por descrever a estrutura das espécies por meio do número de indivíduos e
sua densidade por unidade de área; e a variável qualitativa permite a compreensão da
relação das espécies ocorrentes no meio de estudo, informando se há alguma relação entre
elas (DE OLIVEIRA et al., 2008).
O objetivo deste trabalho foi caracterizar uma área de capoeira para correlacionar
as espécies com a ecologia do ambiente no município de Bragança (PA). Como as áreas
de capoeiras são caracterizadas por vegetação secundaria resultado de degradação
antrópica, fez-se uso de ferramentas fitossociológicas para verificar os impactos
antrópicos na região.

482
Belém (PA), 11 a 13 de dezembro de 2019
2. METODOLOGIA
2.1. CARACTERIZAÇÃO DA ÁREA DE ESTUDO
O estudo foi desenvolvido em duas áreas, uma de capoeira às margens da PA-112
(1°06'38.3"S; 46°48'13.2"W) Figura 1. A média anual de precipitação de 2.329,43 mm,
com a temperatura média compensada do ar variando de 25,7 a 27,2°C e a umidade
relativa do ar variando de 77,9 a 91,4% ao longo do ano. De acordo com a classificação
climática de Köppen-Geiger, descrita por Alvares et al. (2013), a região de Bragança
localiza-se em uma zona “Am”, classificada como clima tropical monçônico, com
precipitação média anual maior que 1.500 mm e com a precipitação do mês mais seco
menor que 60 mm.
Figura 1 - Localização da área de estudo.
Fonte: Autores, 2019.
2.2 AMOSTRAGEM
Foram avaliadas 5 áreas de capoeira para a pesquisa, sendo cada área delimitada
em 10 m x 10 m. Para demarcar a área foi utilizada uma trena de 20 metros e barbante,
os dados referentes às espécies do estudo foram anotados em tabelas que posteriormente
foram utilizadas para estabelecer os parâmetros do estudo.

483
Belém (PA), 11 a 13 de dezembro de 2019
Foi realizada a contagem das espécies para a elaboração da frequência e
determinação dos parâmetros de densidade relativa e absoluta, área basal e diâmetro à
altura do peito (DAP). Para a identificação das espécies foi utilizado como base o site
www.iucnredlist.org.
2.3 ANÁLISE FITOSSOCIOLÓGICA E ESTATÍSTICA
Para a análise da frequência e dos parâmetros foram utilizadas as seguintes
equações:
FR= FA de determinada espécieTotal de FA (1)
Sendo, FA = Frequência absoluta, é o número de indivíduos de determinada
espécie e FR = Frequência relativa.
DA= Nº de indíviduos de uma espécie100m² (2)
Sendo, DA = Densidade absoluta.
DR = DA de cada espécieDA total (3)
Sendo, DR = Densidade relativa.
AB = DAP²4π (4)
Sendo, DAP médio = Diâmetro à altura do peito, o DAP de cada espécie é o
somatório do diâmetro de cada indivíduo dentro da mesma espécie e dividido pelo total
de indivíduos dessa espécie e AB = Área basal em cm².
3. RESULTADOS E DISCUSSÃO
O levantamento do número de indivíduos na área de capoeira e a identificação de
suas espécies podem ser observados na Tabela 1. Foram encontrados na área mais de 60
indivíduos de Sansevieria trifasciata De Wild, seguido por 48 indivíduos de Astrocaryum
aculeatissimum Schott, mas também a área apresentou poucos indivíduos de outras espécies como
Protium heptaphyllum Aubl, Mangifera indica L, Goupia glabra Aubl e Siparuna decipiens Tul.

484
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1 - Espécies e número de indivíduos encontrados na área de capoeira do município de
Bragança-PA.
Espécie Nº de indivíduos
Mangifera indica L (Mi) 4
Protium heptaphyllum Aubl (Ph) 12
Goupia glabra Aubl (Gg) 4
Siparuna decipiens Tul (Sd) 4
Astrocaryum aculeatissimum Schott (Aa) 48
Sansevieria trifasciata De Wild (St) 64
Total 136 Fonte: Autores, 2019.
Em áreas de capoeira é comum a diversidade de espécies, pois trata-se de uma
área que está se recuperando ecologicamente para a realização de suas funções. Gentry
(1979) em trabalho realizado, mostrou a importância da identificação pelas espécies, pois
cada uma pode apresentar peculiaridades que possibilitam o entendimento dos processos
ecológicos que ocorrem em seu local. Baseando-se nessa premissa foi elaborada a Tabela
2, apresentando dados fitossociológicos com as características qualitativas e quantitativas
das espécies coletadas.
Tabela 2 - Dados fitossociológicos de espécies observadas em área de capoeira do município de
Bragança-PA.
Nome da espécie FR (%) DA DR (%) DAP (cm) AB (cm^2)
Mangifera indica L. (Mi) 2,9 0,04 2,94 186,73 14,860
Protium heptaphyllum Aubl. (Ph) 8,8 0,12 8,82 250 19,894
Goupia glabra Aubl. (Gg) 2,9 0,04 2,94 270,2 21,502
Siparuna decipiens Tul. (Sd) 2,9 0,04 2,94 54,5 4,337
Astrocaryum aculeatissimum Schott. (Aa) 35,3 0,48 35,29 479,5 38,157
Sansevieria trifasciata De Wild. (St) 47,1 0,64 47,06 260 20,690
Total 100,00% 1,36 100,00% 1500,93 119,440 Fonte: Autores, 2019.
FR = Frequência relativa, DA= Dominância Absoluta, DR=Densidade Relativa, DAP=
Diâmetro à altura do peito, AB= Área basal.
A frequência relativa das espécies coletadas na área de capoeira no município de
Bragança – PA podem ser observadas na Figura 2. Percebe-se a maior frequência das
espécies Sansevieria trifasciata De Wild. seguido da Astrocaryum aculeatum Schott
podendo serem classificadas com ocorrência Comum e Ocasional. EM JUPI et al. (2013),

485
Belém (PA), 11 a 13 de dezembro de 2019
recomenda a espécie Sansevieria trifasciata Wild. para ser utilizada em quintais
florestais, pois possibilita a contribuição para o equilíbrio ecológico natural sem a
utilização de insumos químicos. No presente trabalho observou-se que a distribuição de
Sansevieria trifasciata De Wild. foi uniforme, permitindo pressupor a influência do efeito
borda nesta espécie. Tal efeito borda, de acordo com Alves Jr. et al. (2006), pode ser
entendido como fragmentação florestal que provoca o isolamento de trechos da floresta
deixando uma parte mais exposta à insolação e outras variáveis que possam interferir na
ecologia do ambiente.
Figura 2 – Frequência relativa das espécies coletadas na área de capoeira no município de
Bragança – PA.
Fonte: Autores, 2019.
Em relação à espécie Astrocaryum aculeatum Schott. encontrada na área, segundo
CAMPÊLO et al. (2018), sua casca pode ser utilizada para a combustão de fornos
destinados ao processo de beneficiamento de oleaginosas. De acordo com a literatura,
pode-se inferir a importância dessas espécies tanto para o equilíbrio do ambiente quanto
para a questão econômica, como o reaproveito dos restos orgânicos.
As demais espécies Siparuna decipiens Tul., Goupia glabra Aubl., Protium
heptaphyllum Aubl. e Mangifera indica L. totalizaram 17,65 % de frequência relativa
indicando sua baixa presença na área de estudo, contudo não se deve ignorar sua
importância para a ecologia do local. A Mangifera indica L., por exemplo, possui uso
medicinal para doenças como a diabetes e analgésicos, conforme afirmação de Canuto

486
Belém (PA), 11 a 13 de dezembro de 2019
(2009), bem como a utilização dos seus frutos para consumo in natura ou confecção de
sucos e geleias; a Protium heptaphyllum Aubl. e Siparuna decipiens Tul. também
possuem uso medicinal e a Goupia glabra Aubl. pode ser utilizada como cosmético e
seus resíduos destinados para a exploração de carvão (FREITAS et al., 2006; GURGEL
et al., 2015; ROCHA et al., 2017). Isso permite refletir sobre a importância ecológica e
econômica que cada planta possui independente da sua frequência no ambiente.
Outro parâmetro analisado foi o diâmetro à altura do peito (DAP). Na Figura 3
percebe-se a diferença entre as espécies. Para as espécies analisadas, verificou-se as
maiores médias de DAP para Goupia glabra Aubl. com 67,55 % e Mangifera indica L.
com 46,68 %. As demais espécies totalizaram 48,51 %. Os altos valores médios de DAP
foram para as espécies de menor frequência, ambas com 2,9 %, isso permite inferir que o
parâmetro de frequência relativa indica a ocorrência da espécie vegetal no ambiente, mas
não que ela terá alto média de DAP, visto que tal parâmetro varia entre os tipos de
espécies.
Figura 3 – DAP (cm) das espécies presentes na capoeira no município de Bragança – PA.
Fonte: Autores, 2019.
Na figura 4 apresenta a área basal das espécies observadas na capoeira no
município de Bragança – PA, onde a área basal de todas as espécies foi de 119,44 cm2.
Três espécies somam 51,98 % da área basal total, sendo elas a Sanseviriana trifasciata

487
Belém (PA), 11 a 13 de dezembro de 2019
De Wild, Goupia glabra Aubl e Protium heptaphyllum Aubl. As outras duas espécies
participam com aproximadamente 48 %. A espécie que apresentou a maior área basal foi
a Astrocaryum aculeatissimim Schott que é responsável por 31,95 %. Percebe-se que o
fato de uma espécie possuir alta frequência relativa, como a Sanseviriana trifasciato De
Wild. 47,1% não equivale a afirmar que essa mesma terá maior área basal, visto que esta
ficou entre as espécies que possuíram o menor valor nesse índice.
Figura 4 – Área basal das espécies na capoeira no município de Bragança – PA
Fonte: Autores, 2019.
4. CONCLUSÃO
O estudo fitossociológico auxilia a caracterização de uma área de capoeira em
relação a sua ecologia, indicando a importância das espécies observadas na área para a
preservação do ambiente em que se encontram além de indicar seus usos para o ser
humano. Verificou-se que o alto índice de frequência relativa de uma espécie não
ocasiona um alto índice de DAP, visto que esse está relacionado ao diâmetro basal
variando entre as espécies.

488
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALVES JR, F. T. et al. Efeito de borda na estrutura de espécies arbóreas em um fragmento
de floresta ombrófila densa, Recife, PE. Revista Brasileira de Ciências Agrárias, v. 1,
n. 1, p. 49-56, 2006.
Botanic Gardens Conservation International (BGCI) & IUCN SSC Global Tree Specialist
Group 2019. Goupia glabra. The IUCN Red List of Threatened Species 2019:
e.T62018945A150003422. Disponível em: < http://dx.doi.org/10.2305/IUCN.UK.2019-
2.RLTS.T62018945A150003422.en.>. Acesso em: 29.jul. 2019.
BRASIL, AMBIENTE. Classificação climática de Köppen-Geiger. 2008.
BRAUN-BLANQUET, V. Fitosociología, bases para el estudio de las comunidades
vegetales. Madrid: H. Blume, 1979. 820 p.
Flora do Brasil 2020 em construção. Jardim Botânico do Rio de Janeiro.Disponível em:
<http://reflora.jbrj.gov.br/reflora/floradobrasil/FB6593>. Acesso em: 29 Jul. 2019.
CAMPÊLO, José Euber Almeida et al. BENEFÍCIOS E DESAFIOS DA
COMERCIALIZAÇÃO DE PRODUTOS DE SEMENTES OLEAGINOSAS NO
MUNICÍPIO DE BRAGANÇA-PA. 2018.
CANUTO, K. M. Propriedades químicas e farmacológicas de mangiferina: um composto
bioativo de manga (Mangifera indica L.). Embrapa Semiárido-Documentos
(INFOTECA-E), 2009. 27 p.
DE OLIVEIRA, A. R.; FREITAS, S. P. Levantamento fitossociológico de plantas
daninhas em áreas de produção de cana-de-açucar. Embrapa Semiárido-Artigo em
periódico indexado (ALICE), p 33-46, 2008.
DE SOUSA, L. A. S.; JARDIM, M. A. G. Sobrevivência e Mortalidade de Plantas de
Açaizeiro (Euterpe oleracea Mart.) Cultivadas em Capoeira no Nordeste Paraense.
Revista Brasileira de Biociências, v. 5, n. S1, p. 255-257, 2007.
EM JUPI, M. et al. GT 11–Ação coletiva e mediadores no espaço rural brasileiro. 2013.
FLORES-LOPES, F.; CETRA, M.; MALABARBA, L. R.. Utilização de índices
ecológicos em assembléias de peixes como instrumento de avaliação da degradação
ambiental em programas de monitoramento. Biota Neotropica, v. 10, n. 4, p. 183-193,
2010.
FREITAS, J. C.; FERNANDES, M. E. B.. Uso de plantas medicinais pela comunidade
de Enfarrusca, Bragança, Pará. Boletim do Museu Paraense Emílio Goeldi Ciências
Naturais, v. 1, n. 3, p. 11-26, 2006.
GENTRY, A. H.. Diversidade e regeneração da capoeira do INPA, com referência
especial às Bignoniaceae. Acta Amaz., Manaus, v. 8, n. 1, p. 67-70, Mar. 1978 .
GURGEL, E. S. et al. Conhecendo Espécie de Plantas da Amazônia: Cupiúba (Goupia
glabra Aubl.–Goupiaceae). Embrapa Amazônia Oriental-Comunicado Técnico
(INFOTECA-E), 2015. 7 p.

489
Belém (PA), 11 a 13 de dezembro de 2019
NOBLICK, L. 1998. Astrocaryum aculeatissimum. The IUCN Red List of Threatened
Species
1998:e.T38755A10147973.Disponívelem:<http://dx.doi.org/10.2305/IUCN.UK.1998.R
LTS.T38755A10147973.en>. Acesso em: 29.jul. 2019.
PEREIRA, Fabiano Volponi et al. Estrutura da vegetação em duas áreas com diferentes
históricos de antropização no manguezal de Anchieta, ES. Boletim do Laboratório de
Hidrobiologia, v. 22, n. 1, 2009.
ROCHA, T. T.; TAVARES-MARTINS, A. C. C.; LUCAS, F. C. A. Traditional
populations in environmentally protected areas: an ethnobotanical study in the Soure
Marine Extractive Reserve of Brazil. Boletín Latinoamericano y del Caribe de Plantas
Medicinales y Aromáticas, v. 16, n. 4, p. 410-427, 2017.
RODAL, M. J. N.; SAMPAIO, E. V. de S. B.; FIGUEIREDO, M. A. Manual sobre
métodos de estudo florístico e fitossociológico: Ecossistema Caatinga. Brasília:
Sociedade Botânica do Brasil, 1992. 24 p.
SILVA, Dodlânia Araújo. Composição florística de plantas daninhas em lavoura de soja
no cerrado maranhense. 2017.
THOMAZ, E. L. Avaliação de interceptação e precipitação interna em capoeira e floresta
secundária em Guarapuava–PR. GEOGRAFIA (Londrina), v. 14, n. 1, p. 47-60, 2005.
VIEIRA, I. C. G.; PROCTOR, J.. Mechanisms of plant regeneration during succession
after shifting cultivation in eastern Amazonia. Plant Ecology, v. 192, n. 2, p. 303-315,
2007.
World Conservation Monitoring Centre 1998. Mangifera indica. The IUCN Red List of
Threatened Species1998: e.T31389A9624842. Disponível
em:<http://dx.doi.org/10.2305/IUCN.UK.1998.RLTS.T31389A9624842.en>. Acesso
em: 29. Jul. 2019.

490
Belém (PA), 11 a 13 de dezembro de 2019
IMPACTOS NA BIODIVERSIDADE COM OS FOCOS DE QUEIMADAS NA
AMAZÔNIA PARAENSE
Rafael Dias Bicalho 1; Kevin Oliveira Moura²; Ana Júlia da Silva Moura³; Letícia
Picanço da Silva4; Gleidson Marques Pereira5
1 Graduando em engenharia ambiental e sanitária. Universidade do estado do Pará -
UEPA. [email protected]. 2 Graduando em engenharia ambiental e sanitária. Universidade do estado do Pará -
UEPA. [email protected]. 3 Graduando em engenharia ambiental e sanitária. Universidade do estado do Pará -
UEPA. [email protected]. 4 Graduando em engenharia ambiental e sanitária. Universidade do estado do Pará -
UEPA. [email protected] 5 Mestre em Agronomia (Solos e nutrição de plantas). Universidade do estado do Pará -
UEPA. [email protected].
RESUMO
A Amazônia, maior floresta tropical do mundo é possuidora de uma biodiversidade
inigualável e admirada, tem sofrido com ações antrópicas como pecuária, a mineração,
comércio madeireiro e demais atividades cujos desmatamentos têm sido provocados em
larga escala. Desta forma, torna-se imprescindível a realização de pesquisas e estudos
com intuito de analisar espacial e temporalmente os focos de calor juntamente com dados
de queimadas e desmatamento de áreas amazônicas, o que enriquece a relevância dessa
pesquisa cujo objetivo consiste em fazer correlação entre esses fatores somada ainda a
impactos decorrentes de tais ações sobre a biodiversidade da fauna e da flora. Para isto,
foram utilizadas informações referentes aos focos de incêndio e desmatamento na região
amazônica contida no estado do Pará, disponibilizadas pela plataforma do Instituto
Nacional de Pesquisas Espaciais-INPE e bases cartográficas do IBGE. A análise dos
dados permitiu identificar o quantitativo de áreas desmatadas em cada estado, os
principais tipos de desmatamento e os municípios com as maiores taxas de focos de
incêndio no Pará. Conclui-se, desta forma, que o estado do Pará é o estado que apresenta
maiores taxas de desmatamento e focos de incêndio na Amazônia brasileira. Em
decorrência disso, tem-se consequências negativas diretas sobre espécies, solo, ciclo
hidrológico e clima.
Palavras-chave: Fogo. Exploração. Solo.
Área de Interesse do Simpósio: Ecologia e Biodiversidade

491
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A diversidade da vida é de suma importância para o equilíbrio ambiental do
planeta Terra, qualificando os ecossistemas para responderem da melhor maneira às
alterações no meio ambiente resultantes de fatores naturais e sociais.Além disso, a
biodiversidade dispõe possibilidades para que o homem se adapte às modificações que
ocorrem em seu meio tanto físico quanto social, e disponibiliza recursos para atender suas
necessidades (ALBAGLI, 2010). Todavia, devido às ações antrópicas essa tem sido cada
vez mais ameaçada, uma vez que o homem vem utilizando os recursos naturais em parte
de forma errônea e desordenada.
Nessa conjuntura, as causas mais predominantes que tem colocado a
biodiversidade brasileira em risco são a perda e a degradação de habitat, ressaltando que
o desmatamento e a expansão agrícola têm contribuído consideravelmente para esse
cenário, especialmente quando somados com outros fatores que também influenciam
nessa problemática, sendo esses a inserção voluntária e involuntária e a disseminação de
espécies exóticas invasoras; a utilização do fogo para limpar terrenos; e a poluição e
contaminação da água e do solo (MMA, 2011).
Nos últimos anos, a região do norte do país em especifico no estado do Pará, vem
sofrendo um aumento expressivo no seu contingente populacional, tal quadro evidencia
o aumento da complexidade urbana da região paraense, pois ocorre um processo de
desmembração e a criação de novos dos municípios. Atestando uma característica de
dinâmica de crescimento de forma endógena (ALVES, 2010).
Os estados da região amazônica tendem a expandir sua fronteira agrícola em busca
do desenvolvimento econômico. Esta área é um espaço estratégico em que visa a
expansão dos plantios de grãos como arroz, milho e soja, a criação de animais para o
abate, exploração madeireira, e a exploração de jazidas de minérios em que a Amazônia
possui em grandes quantidades em sua área. Portanto a Amazônia se tornou influente para
o mercado nacional e de exportação de produtos e insumos (CASTRO, 2012).
Em relação ao quantitativo de biodiversidade, a Amazônia brasileira se destaca,
além de ser a maior extensão de floresta tropical do planeta. A diversidade dos recursos
naturais ali presentes decorre devido às múltiplas associações vegetais que se
desenvolvem devido a fatores ambientais específicos de cada ecossistema que constitui
esse bioma (GAMA et al., 2005). Em razão disso, essa sofre bastante com a exploração,

492
Belém (PA), 11 a 13 de dezembro de 2019
pois devido a sua riqueza, muitos têm se beneficiado de seus recursos sendo esses em
alguns casos extraídos de maneira ilegal e não sustentável resultando em diversos
impactos no meio.
Todavia, a fim de fortalecer o planejamento e promover o desenvolvimento social
e econômico dos estados da região amazônica, o governo implementou o que hoje é
conhecido como Amazônia Legal, composta por Acre, Pará, Amazonas, Roraima,
Rondônia, Amapá e Mato Grosso e as regiões situadas ao norte do paralelo 13º S, dos
Estados de Tocantins e Goiás, e ao oeste do Meridiano de 44º W, do estado do Maranhão
(PRATES; BACHA, 2011).
Toda política pública para a Amazônia deve estar voltada para a utilização de mais
de 67 milhões de hectares (2004) que já foram desmatados e que constituem a "segunda
natureza". Essa área é três vezes a do estado do Paraná ou mais do que a soma dos estados
do Rio Grande do Sul, Santa Catarina e Paraná (HOMMA, 2005), assim como citado por
Álcali (2001) a perca da biodiversidade já era instrumento de pesquisa.
Contudo, ainda assim, a Amazônia legal conta com o maior índice de queimadas
dos últimos anos no Brasil, essas sendo consequência da grande concentração do
desmatamento atual. A maior parte do desmatamento encontra-se principalmente ao
longo de um "arco" que abrange desde o sudeste do Maranhão, incluindo o norte do
Tocantins, o sul do Pará, norte de Mato Grosso, Rondônia, sul do Amazonas, até o sudeste
do Acre. Concentra cerca de 524 municípios (IGNOTTI, et al., 2007).
Tendo em vista todo o exposto, a relevância desta pesquisa se dá pela correlação
realizada entre queimadas e os focos de calor analisados em áreas amazônicas e as
atividades desenvolvidas no período em questão, a levar em consideração também os
prejuízos expressivamente negativos que incêndios e desmatamentos ocasionam à
biodiversidade, ao ciclo hidrológico e ao clima.
O presente estudo visa analisar e associar a ocorrência de focos de calor com dados
de desmatamento, queimadas e manchas urbanas sobre a biodiversidade da Amazônia
paraense durante o período compreendido entre 1998 e 2018.

493
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
A Floresta Amazônica analisada está contida no estado do Pará (Figura 1),
fazendo parte de uma das 27 unidades federativas do Brasil, localiza-se na região norte,
e é o segundo maior território do país com aproximadamente 1.247.950km², com 8,5
milhões de habitantes segundo o IBGE.
Figura 1 – Mapa de localização da Amazônia e do Pará
Fonte: Autores (2019)
2.2 MÉTODO
O presente trabalho se trata de uma revisão bibliográfica que foi desenvolvido a
partir de pesquisas integrativas e exploratórias. O estudo foi feito através de um
levantamento de dados obtido pelo Instituto Nacional de Pesquisas Espaciais (INPE)
entre os anos de 2008 a 2018 e com um banco de informações adquirido de outros artigos
como periódicos nacionais e internacionais realizadas no portal Google Acadêmico,
IMPE, IBGE, IMET e PRODES. Com um recorte temporal de dezessete anos (2002 –
2019).

494
Belém (PA), 11 a 13 de dezembro de 2019
2.3 PROCESSAMENTO DE DADOS
Através dos dados dos satélites e de monitoramento da vegetação do PRODES,
em especifico no Pará, foi possível realizar um processamento de dados com os softwares
Qgis que foi para tratar as camadas vetoriais utilizadas para a confecção da sobreposição
dos mapas de localização e identificação, e o Excel que foi utilizado para confeccionar os
gráficos. E a comparação de dados através de artigos de periódicos.
3. RESULTADOS E DISCUSSÃO
O INPE disponibiliza dados anuais de 2008 a 2018 relacionados ao desmatamento
da Floresta Amazônica no estado do Pará e em unidades de conservação- UC (Figura 2)
e dados de alerta de desmatamento a partir de 2015.
Figura 2 – Mapa de desmatamento da Amazônia e do Pará
Fonte: INPE (2019) e editado por Autores (2019).
De acordo com os dados fornecidos, o Pará é o estado que apresenta maior taxa
de desmatamento da Amazônia. Desde o ano de 2008 a 2018 permaneceu como o líder
desse grupo, mesmo havendo uma queda considerável nessa taxa. Todavia, altos valores
de desmatamento têm sido observados na Amazônia a começar da década de 70, sendo
que em 1995 esses valores atingiram o seu maior nível, e posteriormente a esse ano, os
mesmos vem mostrando oscilações dissemelhantes advindas de várias causas, como
incêndios, comercialização madeireira, crescimento da atividade agropecuária, expansão

495
Belém (PA), 11 a 13 de dezembro de 2019
da densidade populacional e incentivos fiscais (ARRAES, MARIANO, SIMONASSI,
2012).
No que se refere às taxas anuais de desmatamento da Amazônia Legal, de acordo
com os dados disponibilizados pelo INPE, é possível fazer comparações entre o
quantitativo de áreas desmatadas de cada estado entre 2008 e 2018 (Figura 3).
Figura 3 – Grafico da taxa de desmatamento anual na Amazônia por estados.
Fonte: INPE (2019)
Segundo Carvalho (2012) a expansão agropecuária é a grande causadora do
desmatamento, pois os empreendedores precisam de mais áreas para aumentar o lucro e
a floresta se torna um empecilho para o desenvolvimento econômico dessas pessoas.
Dentre os impactos do desmatamento, está a perda de possibilidades para a utilização de
maneira sustentável da floresta, além da fabricação de produtos tradicionais seja através
do manejo florestal para madeira ou até mesmo pela retirada de produtos não advindos
dessa. Ademais, tal ato impossibilita que o valor dos serviços ambientais advindos da
floresta seja capturado (FEARNSIDE, 2006).
Quanto aos tipos de desmatamento presentes no estado do Pará, o INPE fornece
informações relativas ao tema entre os anos de 2017 e 2018 (Figura 4) e permite afirmar
que a principal razão de desmatamento do local refere-se a incêndio florestal, com
8.929,1km².

496
Belém (PA), 11 a 13 de dezembro de 2019
Figura 4 – Gráfico dos tipos de desmatamento no Pará nos anos de 2017 a 2018
Fonte: INPE (2019) e editado por Autores (2019)
Segundo Soares e Santos (2002), entre os diversos agentes que impactam os
recursos florestais, o fogo aparentemente é o maior desastre. Tanto as florestas quanto
demais variedades de vegetação estão incessantemente suscetíveis a incêndios de
intensidades dissemelhantes. Assim, também, Cabral, Filho e Borges (2013), afirmam
que o fogo é um instrumento usado pelos agropecuaristas para limpar o pasto e preparar
a área para o plantio, em condições laboratoriais onde é controlado os fatores climáticos,
essa prática é excelente, porém, o vento e o tempo seco podem alastrar causando um
incêndio nas florestas próximas.
O fogo no solo pode gerar várias modificações de diferentes aspectos, podendo
ser de natureza física, química ou biológica, o fator determinante da sua intensidade e o
grau de alteração e seu tipo de ação que se configura nas formas diretas e indiretas,
levando em consideração fatores de influência mais específicos como tipo de solo,
cobertura vegetal, duração, intensidade e frequência de uso (REDIN, 2011).
Em função da influência das queimadas, sendo elas ocasionadas por meio natural
ou antrópico, o solo é sujeito a impactos significativos, e por consequências ele pode
degradar a biota decompositora existente, acarretando a diminuição da infiltração de
água, interferindo negativamente nos ciclos biogeoquímicos, ocasionando a redução de
nutrientes do solo (SILVA, 2018).

497
Belém (PA), 11 a 13 de dezembro de 2019
Com isso, a região que está sujeita à prática de queimadas tem seu clima afetado
negativamente, pois a cobertura vegetal que é importante para a regulação do ciclo
hidrológico da região que está sendo suprimida, estendendo os períodos de seca da região
em questão, além de intensificar a emissão de gás carbônico (CO2) para a atmosfera, faz
com que a temperatura da região aumente a níveis alarmantes (ARTAXO, 2014). Assim,
com a queda da pluviosidade, a ocorrência de incêndios nas florestas aumenta, o que fará
com que haja a produção de mais seca e também fumaça e outras florestas fiquem
propícias também a sofrerem incêndios e desmatamentos (MOUTINHO, 2009).
Essas mudanças ocasionadas em decorrências das queimadas que influenciam o
solo e na atmosfera se propagam de maneira rápida e proporciona mudanças ambientais
mais profundas com efeitos no ecossistema amazônico e possivelmente no clima de outras
regiões do Brasil (LARA, 2005). Ademais, nas queimadas são liberados poluentes como
NOx, CO, HC, material particulado e entre outros, resultando em efeitos negativos à
saúde da população onde as mais afetadas são as pessoas mais próximas à área da
queimada, destacando-se as que estejam atuando no seu combate. As consequências
podem ir de intoxicação até a morte por asfixia (RIBEIRO; ASSUNÇÃO, 2002).
Como citado anteriormente, o Pará lidera a lista de estados com maior índice de
desmatamento, fator esse que pode influenciar na ocorrência de incêndios florestais de
forma natural. Todavia, ressalta-se que tal acontecimento também pode ocorrer através
do alastramento de incêndios causados pela ação humana.
Já em relação ao quantitativo de focos de incêndios relacionado com alguns
municípios pertencentes a estados da Amazônia Legal, os dados fornecidos pelo INPE
permitem analisar do mesmo modo quais locais apresentam o total mais alarmante no que
se refere ao estado do Pará (Figura 5).

498
Belém (PA), 11 a 13 de dezembro de 2019
Figura 5 – Municípios com maior taxa de focos de incêndio no Brasil.
Fonte: INPE (2019)
A análise dos dados permite inferir que os municípios do estado do Pará que
apresentam a maior taxa de focos de incêndio no Brasil são: Altamira e São Félix do
Xingu, seguidos de Novo Progresso e, por último, Itaituba. No que tange aos dois
municípios cujos dados apresentam maior expressividade, um estudo realizado por
Martinez, Fiedler e Lucatelli (2007) sobre focos de calor, desflorestamentos nesses
municípios e o elo entre esses, mostra que a relação é positiva e que a maior fração das
ocorrências de focos de calor correspondia à conversão de áreas de floresta em pastagens.
Já em relação à estimativa de focos de calor registrados no município de Novo
Progresso, um outro estudo efetuado por Lopes et al. (2017) sobre a distribuição de focos
de calor nesse município, os autores abordam que, além da influência de madeireiras,
principais responsáveis pelos focos de incêndio na região, a tendência de variação
temporal desses focos apresenta relação direta com o período de estiagem do município.
Da mesma forma, Alves et al. (2019) apresenta alguns dados: em 2016 os autores
afirmam que houve mais eventualidades de queimadas e incêndios nos meses de agosto
com 2.066 e novembro com 1.512, e março e abril apresentaram os menores registros

499
Belém (PA), 11 a 13 de dezembro de 2019
sendo esses 13 e 5 subsequentemente. Já no ano posterior, o mês de agosto teve 4.440 e
setembro 6.222, havendo assim maiores detecções de focos de calor. Tais dados acabam
por sustentar o apresentado por Lopes et al. no que tange à relação entre focos e períodos
de estiagem.
No que diz respeito aos focos de queimadas no estado do Pará, de forma mais
abrangente, a partir de uma análise espacial, Santos et al. (2015) chamam atenção para a
maior concentração presente no nordeste e sudeste do estado, bem como aumento em
períodos menos chuvosos e a relação entre maior incidência dos casos em municípios de
grandes extensões territoriais como São Félix do Xingu, Altamira Itaituba.
4. CONCLUSÃO
A análise da relação entre focos de calor e desmatamento, atividades
desenvolvidas e impactos diretos na biodiversidade amazônica, permitiu inferir alguns
pontos:
- O Pará é o estado que apresenta maiores índices relativos a focos de incêndio e
desmatamento, sendo que, quanto a este último, os municípios de São Félix do Xingu e
Altamira denotam valores maiores expressivos;
- O aumento do desmatamento e focos de incêndios possivelmente relacionam-se
às práticas agropecuárias e madeireiras, cujo crescimento tem se mostrado evidente nos
últimos anos;
- As queimadas na atualidade impactam de forma expressiva o meio ambiente,
tanto natural quanto urbano, desta forma prejudicando as espécies que vivem nesse meio,
inclusive o ser humano. Com o aumento das queimadas, a floresta tem seus serviços
ecossistêmicos afetados, constituindo esses como de profunda importância para a
manutenção do ciclo hidrológico, clima, biodiversidade e saúde humana.
- O processo de queimadas na Amazônia tem se intensificado devido a intensa
expansão econômica formado pela demanda do mercado interno e externo, fomentando
o grande desafio da sociedade brasileira atual que é crescer economicamente sem deixar
de preservar o meio – ambiente. Portanto, faz-se necessário o incentivo para estudos e
pesquisas na região para compreender e aliar as necessidades do meio social à
sustentabilidade, de forma a manter o crescimento econômico da região sem afetar as
gerações futuras em termos de recursos e bem-estar.

500
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALBAGLI, S. Amazônia: fronteira geopolítica da biodiversidade. Parcerias
estratégicas, v. 6, n. 12, p. 05-19, 2010.
ALVES, P. A. et al. Explorando as relações entre a dinâmica demográfica, estrutura
econômica e mudanças no uso e cobertura da terra no sul do Pará: lições para o distrito
florestal sustentável da BR-163. Geografia, v. 35, n. 1, p. 165-182, 2010.
ALVES, R. J. M. et al. Análise do monitoramento de focos de calor e propostas para a
redução de queimadas e incêndios em áreas protegidas no estado do Pará. Contribuições
a las Ciencias Sociales, 2019.
ARRAES, R. de A.; MARIANO, F. Z.; SIMONASSI, A. G. Causas do desmatamento no
Brasil e seu ordenamento no contexto mundial. Revista de Economia e Sociologia
Rural, v. 50, n. 1, p. 119-140, 2012.
ARTAXO, P. et al. Perspectivas de pesquisas na relação entre clima e o funcionamento
da floresta Amazônica. Ciência e Cultura, v. 66, n. 3, p. 41-46, 2014.
CABRAL, A. L.; FILHO, L. O. M.; BORGES, L. A. C. Uso do fogo na agricultura:
legislação, impactos ambientais e realidade na Amazônia. Periódico Eletrônico Fórum
Ambiental da Alta Paulista, v. 9, n. 5, 2013. Disponível
em:<http://www.amigosdanatureza.org.br/publicacoes/index.php/forum_ambiental/artic
le/view/577/601 >. Acesso em: 06 set. 2019.
CARVALHO, A. C. Expansão da fronteira agropecuária e a dinâmica do desmatamento
florestal na Amazônia Paraense. 2012. 349 p. Tese (doutorado) - Universidade
Estadual de Campinas, Instituto de Economia, Campinas, SP. Disponível
em:<http://www.repositorio.unicamp.br/handle/REPOSIP/285926>. Acesso em: 06 set.
2019.
CASTRO, E. Expansão da fronteira, megaprojetos de infraestrutura e integração sul-
americanas. Caderno CRH, v. 25, n. 64, 01-04, 2012, pp. 45-61 Universidade Federal
da Bahia Salvador, Brasil.
FEARNSIDE, P. M. Desmatamento na Amazônia: dinâmica, impactos e controle.
2006. GAMA, J. R. V. et al. Comparação entre florestas de várzea e de terra firme do
Estado do Pará. Revista Árvore, v. 29, n. 4, p. 607-616, 2005.
FLORESTAL, Novo Código. Lei 12.651 de 25 de maio de 2012. Presidência da
República. Casa Civil. Subchefia para Assuntos Jurídicos, 2016.
HOMMA, A. O. Amazônia: como aproveitar os benefícios da destruição? Estudos
avançados, v. 19, n. 54, p. 115-135, 2005.
IGNOTTI, E. et al. Efeitos das queimadas na Amazônia: método de seleção dos
municípios segundo indicadores de saúde. Revista Brasileira de Epidemiologia, v. 10,
p. 453-464,2007.
INSTITUTO NACIONAL DE PESQUISAS ESPACIAIS (INPE). COORDENAÇÃO
GERAL DE OBSERVAÇÃO DA TERRA. PROGRAMA DE MONITORAMENTO DA

501
Belém (PA), 11 a 13 de dezembro de 2019
AMAZÔNIA E DEMAIS BIOMAS. Desmatamento – Amazônia – Disponível em:
http://terrabrasilis.dpi.inpe.br/. Acesso em: 04 set. 2019.
LARA, L. et al. Química atmosférica na Amazônia: A floresta e as emissões de queimadas
controlando a composição da atmosfera amazônica. SciELO: São Paulo, Brasil, 2005.
LOPES, A. C. L. et al. Análise da distribuição de focos de calor no município de Novo
Progresso, Pará. Revista Verde de Agroecologia e Desenvolvimento Sustentável, v. 12,
n. 2, p. 298-303, 2017.
MARTINEZ, L. L.; FIEDLER, N. C.; LUCATELLI, G. J. Análise das relações entre
desflorestamentos e focos de calor. Estudo de caso nos municípios de Altamira e São
Félix do Xingu, no estado do Pará. Revista Árvore, v. 31, n. 4, p. 695-702, 2007.
MEC. Dados estatísticos [http://sinaes.inep.gov.br/sinaes – Acesso em 03/09/2019].
BRASIL – PRESIDÊNCIA DA REPÚBLICA. Lei nº 1.806/53 de 06.01.1953 (criou a
SPVEA). www.presidencia.gov.br/legislacao,. Brasília, 2007.
MINISTÉRIO DO MEIO AMBIENTE. Quarto relatório nacional para a convenção
sobre diversidade biológica: Brasil. Brasília: MMA, 2011. 248 p.
MOUTINHO, P. Desmatamento na Amazônia: desafios para reduzir as emissões de
gases de efeito estufa do Brasil. http://www. ipam. org. br/biblioteca>. Acesso em, v. 5,
n. 2009, p. 2-3, 2009.
PRATES, R. C.; BACHA, C. J. C. Os processos de desenvolvimento e desmatamento da
Amazônia. Economia e Sociedade. Campinas. v. 20, n. 3, p. 601-636, 2011.
REDIN, M. et al. Impactos da queima sobre atributos químicos, físicos e biológicos do
solo. Ciência Florestal, v. 21, n. 2, p. 381-392, 2011.
RIBEIRO, H.; ASSUNÇÃO, J. V. Efeitos das queimadas na saúde humana. Estudos
avançados, v. 16, n. 44, p. 125-148, 2002.
SANTOS, K. S. et al. Análise espacial das queimadas nos municipios do estado do Pará,
no período de 2015. In: XVII Simpósio Luso Brasileiro de Engenharia Sanitária e
Ambiental – SILUBES. Anais. Florianópolis-SC, 2015.
SILVA, P. et al. Proposta de recuperação de área degradada por queimadas no município
de Marabá-PA. Revista Brasileira de Gestão Ambiental, v. 12, n. 3, p. 08-17, 2018.
SOARES, R. V.; SANTOS, J. F. Perfil dos incêndios florestais no Brasil de 1994 a 1997.
Floresta, v. 32, n. 2, 2002.

502
Belém (PA), 11 a 13 de dezembro de 2019
ÍNDICES DE EMERGÊNCIA DE PLÂNTULAS DE ACMELLA OLERACEA (L.)
R.K. JANSEN E ACMELLA CILIATA (KUNTH) CASS
Camila Monteiro Salgado1; Mônica Trindade Abreu de Gusmão2; Gabriel Rodrigues
Santos3; Joyce da Costa Dias4.
1 Graduanda em agronomia. Universidade Federal Rural da Amazônia.
2 Doutora em Produção Vegetal. Universidade Federal Rural da Amazônia.
3 Mestrando em aquicultura e recursos aquáticos. Universidade Federal Rural da
Amazônia. [email protected]
4 Graduanda em agronomia. Universidade Federal Rural da Amazônia.
RESUMO
As plantas de jambu (Acmella oleracea (l.) R.K. Jansen e Acmella ciliata (Kunth) Cass)
são nativas da região amazônica, pertencentes à família Asteraceae (Compositae), muito
utilizadas para fins medicinais e alimentícios, entretanto há carência de informações
relacionadas aos aspectos agronômicos, principalmente em relação à tecnologia de
sementes. Nesse contexto, o objetivo deste trabalho foi estimar os parâmetros genéticos
em relações aos caracteres biométricos de sementes de duas espécies de jambu. Foi
utilizado delineamento inteiramente casualizado (DIC), com três tratamentos e quatro
repetições. Os tratamentos foram constituídos por três acessos: acesso 1 (T1),
representante de Tapanã; acesso 2 (T2), representante de Salvaterra e acesso 3 (T3),
representante de Bragança. Inicialmente as sementes foram debulhadas e separadas das
impurezas. Foram colocadas 100 sementes em cada caixa plástica gerbox em substrato de
papel filme umedecido com água destilada. Em seguida, foram levadas para uma câmara
incubadora do tipo BOD em temperatura de 29°C. Foram analisados os dados de
porcentagem de emergência (%E); índice de velocidade de emergência (IVE) e tempo
médio de emergência (TME). Para a análise da dissimilaridade genética utilizou-se o
software Rbio com suprimento do R 3.4.3. Com isso, na análise da %E houve maior
influência genotípica no fenótipo do material, onde a relação cgv/cve ficou próxima de
1, ademais, no IVE foi possível observar que a herdabilidade apresentou ótimo resultado
com altas perpesctivas para a transferência do material às gerações futuras, já em TME
os dados apresentaram alta relação intraclasse denotando a confiabilidade nos demais
dados analisados. Portanto, para as três variáveis analisadas com o teste de germinação,
o valor genotípico demonstrou maior importância na expressão das características, além
disso, a dissimiliridade evidenciou que os acessos de tapanã e bragança são mais distantes,
geneticamente, entre sí, representando maior heterogeneidade entres os materiais.
Palavras-chave: Jambu. Dissimilaridade. Teste de germinação.
Área de Interesse do Simpósio: Biodiversidade

503
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
As plantas de jambu são nativas da região amazônica, pertencentes à família
Asteraceae (Compositae) e tem seu cultivo difundido entre os pequenos produtores de
alguns municípios do nordeste do Estado do Pará (COSTA, 2010). As espécies Acmella
oleracea (l.) R.K. Jansen e Acmella ciliata (Kunth) Cass são espécies herbáceas não
convencionais, utilizadas para fins medicinais e alimentícios com poucas informações
relacionadas aos aspectos agronômicos, principalmente em relação à tecnologia de
sementes (SOUSA, 2018).
Além de fazer parte da culinária típica do estado do Pará, como o pato no tucupi
e o tacacá, o jambu é bastante utilizado em saladas ou compondo outros pratos. Sua
composição química, incluindo o espilantol, tem potencial para o uso industrial, sendo
ainda citado como indicativo para diversas doenças. É uma hortaliça rica do ponto de
vista nutricional possuindo diversos elementos como ferro, e ainda possui as vitaminas
B1, B2, niacina, vitamina C, vitamina A e cálcio (GUSMÃO; GUSMÃO, 2013).
De modo geral, o estudo da morfologia das sementes, o conhecimento da
emergência e o desenvolvimento da plântula são importantes para distinguir as famílias
botânicas e diferenciá-las em subfamílias, gêneros e espécies, contribuindo nos trabalhos
sobre regeneração e de tecnologia de sementes, além de colaborar para as análises de
dispersão, sucessão e regeneração das espécies (SANTOS et al., 2014; SOUZA, 2018).
O conhecimento da emergência de plântulas é um fator importante para
caracterização das espécies, pois é uma maneira importante para detectar a variabilidade
genética dentro de populações de uma mesma espécie, assim como as variações
individuais da mesma e entre diferentes localidades devido às condições ambientais.
Nesse contexto, o objetivo deste trabalho foi estimar os parâmetros genéticos em relações
aos caracteres de emergência de plântulas de duas espécies de jambu oriundas de três
localidades.
2. MATERIAL E MÉTODOS
Os dados foram coletados a partir do plantio de plantas de jambu que
permaneceram em campo durante três meses, período no qual os aquênios ficaram
maduros. Posteriormente à colheita, as sementes foram direcionadas ao laboratório de

504
Belém (PA), 11 a 13 de dezembro de 2019
sementes do Instituto de Ciências Agrárias localizado no Campus da Universidade
Federal Rural da Amazônia, em Belém (PA) para a condução do ensaio.
O experimento foi conduzido em Delineamento Inteiramente Casualizado (DIC),
utilizando-se três tratamentos e quatro repetições, sendo que os tratamentos foram
constituídos por duas espécies de plantas de jambu, a Acmella oleracea (l.) R.K. Jansen
e a Acmella ciliata (Kunth).
Inicialmente, as sementes foram debulhadas e separadas das impurezas. Em
seguida foram colocadas 100 sementes em cada caixa de poliestireno Gerbox com
dimensões de 11mm x 11mm x 3,5mm. Cada Gerbox foi disposto em câmara incubadora
do tipo BOD (Figura 1), onde as sementes foram mantidas em temperatura de 29°C, com
fotoperíodo de 12 horas e umidade relativa do ar acima de 90, durante oito dias.
Figura1 – Estufa incubadora do tipo BOD contendo plantas de jambu.
Fonte: Autores, 2019.
Em cada Gerbox foi utilizado como substrato uma folha de papel filtro umedecida
com água destilada durante o intervalo de 24 horas, sendo translocado 2mL de água
destilada com auxílio de pipeta, em cada repetição.
Foram realizadas diariamente até o oitavo dia após a semeadura, as avaliações
quanto ao número de sementes emergidas; fungadas; moles e duras, onde foram
consideradas mortas, as sementes que estavam em outra condição adversa da emergida.
Também foram avaliadas a porcentagem de emergência (E), o índice de velocidade de

505
Belém (PA), 11 a 13 de dezembro de 2019
emergência (IVE) e tempo médio e emergência (TME). Sendo utilizado para sua
estimativa o software Excel de domínio da Microsoft.
Para a porcentagem de emergência, considerou-se a expressão: E = (Σn/ss)*100
Para o índice de velocidade de emergência, considerou-se a expressão: IVE =
(E1/N1)+(E2/N2)+...(En/Nn).
Para o tempo médio e emergência, considerou-se a expressão: TME =
Σ(E1*N1+E2*N2+En*Nn)/(Σn).
Posteriormente avaliaram-se as estimativas dos parâmetros genéticos, conforme
abaixo:
Para a variância fenotípica considerou-se a expressão: σ2f = (QMP/r)
Para a variância genotípica considerou-se a expressão: σ2g = (QMP/QMR/r)
Para a variância ambiental considerou-se a expressão: σ2e = (QMR/r)
Para a herdabilidade considerou-se a expressão: H2 = σ2g/σ2f
Para a correlação intraclasse considerou-se a expressão: C.I = σ2entre
grupos/(σ2entre grupos + σ2dentro do grupo).
Para o coeficiente de variação genético considerou-se a expressão: cvG =
100*((√Φg)/x)
Para a Razão de variância genotípica e ambiental considerou-se a expressão=
cvG/cvE
Para a acurácia considerou-se a expressão: Rgg = (r2mm * r2
mq)1/2
3. RESULTADOS E DISCUSSÃO
Os resultados indicam maior atribuição à variância genotípica na definição das
características quando comparada à variância ambiental (Tabela 1), desta forma, os
efeitos dos genes são mais relevantes na expressão do fenótipo o que representa algo
positivo tendo em vista que é buscado pelo melhorista o potencial gênico da espécie.

506
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1 – Porcentagem de emergência (E), quanto à variância fenotípica (σ2f); variância
genotípica (σ2g); variância ambiental (σ2e); herdabilidade (H2); correlação intraclasse (C.I);
coeficiente de variação genético (cvG); Razão de variância genotípica e ambiental) e Rgg
(acurácia).
σ2f σ2g σ2e H2(%) C.I(%) cvG(%) cvG/cvE Rgg
0,078 0,06 0,018 77,09 45,7 21,50 0,91 0,87
Fonte: Autores, 2019.
O coeficiente de herdabilidade fomenta a influência genotípica desses materiais,
tendo em vista que os resultados apontam 77,09% de possibilidade das características de
germinação serem repassadas à próxima geração. Sendo assim, esse percentual indica alta
herdabilidade, onde há maior probabilidade da característica ser fixada (Tabela 1).
A relação cvG/cvE foi tendente a 1 com valor de 0,91 na análise de porcentagem
de emergência (Tabela 1) e ultrapassou essa medida nas variáveis de IVG e TMG, sendo
respectivamente, 1,12 (Tabela 2) e 1,83 (Tabela 3). Com valores acima de 1 ou tendentes
a 1 infere-se que há situação favorável ao ganho de seleção em decorrência da variância
genotípica ser superior à variância do ambiente.
Tabela 2 – Índice de Velocidade de Emergência (IVE), quanto à variância fenotípica (σ2f);
variância genotípica (σ2g); variância ambiental (σ2e); herdabilidade (H2); correlação intraclasse
(C.I); coeficiente de variação genético (cvG); Razão de variância genotípica e ambiental) e Rgg
(acurácia).
σ2f σ2g σ2ge H2(%) C.I(%) cvG(%) cvG/cv Rgg
4,706 3,933 0,773 83,57 55,98 28,28 1,12 0,91
Fonte: Autores, 2019.
Conforme Resende (2015) a herdabilidade de forma singular detém a seguinte
classificação, baixa (0,01 ≤ h2 ≤ 0,15); média (0,15 < h2 ≤ 0,50); alta (h2 ≥ 0,50). Com
herdabilidade a 83,57% é possível obter confiabilidade no valor genotípico.
As variações genotípica e ambiental foram respectivamente de 3,933 e 0,773
(Tabela 2) indicando maior influência do gene no fenótipo apresentado para velocidade

507
Belém (PA), 11 a 13 de dezembro de 2019
de emergência. Este fato é ratificado pela relação cvG/cvE, a qual manteve-se acima de
1 demonstrando alta possibilidade ao ganho de seleção ( Tabela 2).
A estimativa da correlação intraclasse dos materiais analisados para variável
Tempo Médio de Emergência mostrou-se satisfatória quanto à confiabilidade das medidas
para definição da relação entre os indivíduos dentro da mesma classe, em torno de 77,l5%
(Tabela 3).
Tabela3 – Tempo Médio de Emergência (TME), quanto à variância fenotípica (σ2f); variância
genotípica (σ2g); variância ambiental (σ2e); herdabilidade (H2); correlação intraclasse (C.I);
coeficiente de variação genético (cvG); Razão de variância genotípica e ambiental) e Rgg
(acurácia).
σ2f σ2g σ2e H2(%) C.I(%) cvG(%) cvG/cvE rgg
0,1217 0,1133 0,0083 93,10 77,15 6,14 1,83 0,96
Fonte: Autores, 2019.
O teste foi significativo a 5 rejeitando-se, portanto, H0 e adotando-se H1 para os
tratamentos, indicando que há diferença significativa entre os materiais genéticos
analisados, logo, as médias entre os acessos não são iguais havendo variabilidade genética
(Tabela 4).
Tabela 4 – ANOVA resultante da porcentagem de germinação do cultivo de plantas de jambú A.
oleraceae e A. ciliata. Teste Turkey significativo a 5.
GL SQ QM F p-valor
Tratamento 2 6,30 3,15 4,367 0,0463
Resídual 9 6,49 0,72
Fonte: Autores, 2019.
Com a dissimilaridade é possível estimar a variabilidade entre os genótipos,
dividindo-os em grupos, os quais demonstram os materiais mais próximos e mais
distantes geneticamente, obtendo-se com os resultados mais distantes os melhores
resultados para o melhoramento genético. Sendo estes ambientes passíveis de serem

508
Belém (PA), 11 a 13 de dezembro de 2019
agrupados por conglomeração, com a utilização de medida de dissimilaridade do fator
ambiental e, por conseguinte, com método de agrupamento (Cruz, 2006)
O par formado entre os acessos dois e três são vizinhos próximos, o que demonstra
sua proximidade genética, ou seja, compartilham de genes semelhantes (Figura 2).
Figura 2 – Dendograma de Cluster representativo da dissimilaridade genética entre genótipos de
três diferentes cultivares de plantas de jambú, utilizando o método do vizinho mais próximo por
meio da matriz de distância euclidiana.
Fonte: Autores, 2019.
Com genes em comum é possível dizer que esses materiais genéticos apresentam
baixa heterogeneidade. No entanto, observa-se que entre esses dois grupos há ampla
dissimilaridade, indicando a diferença e distanciamento entre o acesso um e o grupo
formado pelos acessos dois e três, o que representa genes distintos e com alta
heterogeneidade (Figura 2). Com isso, esses acessos podem ser explorados para fins de
melhoramento, já que apresentam alta variabilidade genética, podendo ser indicadas para
cruzamento.

509
Belém (PA), 11 a 13 de dezembro de 2019
Os acessos com maiores dissimilaridades entre os três avaliados foram o um (A.
oleracea advinda de Tapanã) e o três (A. oleracea advinda de bragança) formando grupos
distanciados uns dos outros (Figura 2).
4. CONCLUSÃO
Os acessos apresentaram alta herdabilidade para as características de Porcentagem
de emergência, Velocidade de Emergência e Tempo Médio de Emergência evidenciando a
importância do potencial gênico na expressão desses fenótipos e a agregação das
características em gerações futuras.
Por conseguinte, para as três variáveis analisadas houve ótima relação cvG/cvE
indicando materiais genéticos propícios ao ganho de seleção sendo, portanto, indicados
para participarem de programa de melhoramento genético.
Por intermédio da dissimilaridade, o acesso um (A. oleracea advinda de Tapanã)
e o acesso três (A. oleracea advinda de bragança) analisados demonstram alta
variabilidade genética, indicando sua heterogeneidade.

510
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
Bhering, L.L. Rbio: A Tool For Biometric And Statistical Analysis Using The R
Platform. Crop Breeding and Applied Biotechnology, v.17: 187-190p, 2017
COSTA, C. M. L. Caracterização e análise experimental do recobrimento de
sementes de jambu (Spilanthes oleracea) em leito fluidizado. 2010. 203 p. Trabalho de
Dissertação de Doutorado. Universidade Estadual de Campinas, Campinas, SP, 2010.
CRUZ, Cosme Damião, 1958 – Programa GENES : biometria / Cosme Damião Cruz. –
Viçosa : Ed.UFV, 54p.2006.
GUSMÃO, M. T. A.; GUSMÃO, S. A. L. Jambu da amazônia (Acmella oleracea [(L.) R.
K. Jansen]: características gerais, cultivo convencional, orgânico e hidropônico. Editora
Universidade Federal Rural da Amazônia, UFRA, 2013. 135 p.
SANTOS, H. H. D.; MATOS, V. P.; ALBUQUERQUE, A. P. C.; SENA, L. H. M.;
FERREIRA, E. G. B. S. Morfologia de frutos, sementes e plântulas de Averrhoa bilimbi
L. oriundas de dois estágios de maturação. Ciência Rural. Santa Maria, v. 44, n. 11, p.
1995-2002, 2014.
SOUSA, E. C. Morfologia e germinação de sementes de jambu (Acmella oleracea L.
R. K. Jansen). 2018. 82 p. Trabalho de Dissertação de Mestrado. Universidade Rural do
Semi árido, Mossoró, RN, 2018.
RESENDE, M.D.V. Genética quantitativa e de populações.Visconde do Rio Branco:
Ed. Suprema, 2015.

511
Belém (PA), 11 a 13 de dezembro de 2019
REGISTROS REPRODUTIVOS DA MARRECA, DENDROCYGNA
AUTUMNALIS, NO ENTORNO DA RESEX MARINHA DE TRACUATEUA-PA
Euller Pimentel Teixeira1; Sanae Nogueira Hayashi2; Neuma Teixeira dos Santos³;
Jaime Viana de Sousa4.
1Graduando de Engenharia Ambiental e Energias Renováveis. LabGEFA - Universidade
Federal Rural da Amazônia. [email protected]
2 Doutora em Biologia Ambiental. LabGEFA - Universidade Federal Rural da
Amazônia. [email protected]
3Mestre em Engenharia Elétrica. LabGEFA - Universidade Federal Rural da Amazônia.
4Doutor em Bioinformática e Genética Humana Molecular. LabGEFA - Universidade
Federal Rural da Amazônia. [email protected]
RESUMO
Conhecida popularmente como marreca, Dendrocygna autumnalis, são aves de grande
interesse tanto para a caça de subsistência, amadorista ou comercial de sua carne, além
de muitas vezes ser criada como mascote. A captura dessa fauna pode ser uma prática
recorrente em áreas de reservas e seus arredores, apresentando aspectos conflitivos como
o que ocorre na Reserva Extrativista Marinha de Tracuateua (PA) e na sua área
circundante, nesse caso, a captura constante dessa ave pode causar desequilíbrio
ecológico para o ecossistema da região. Nesse sentido, este trabalho tem como objetivo
divulgar registros de reprodução de D. autumnalis, em cativeiro, assim como a
documentação e descrição das características do ninho e do habitat de nidificação, no
entorno da Reserva Extrativista Marinha de Tracuateua (PA). O trabalho seguiu uma
pesquisa exploratória, pois não havia evidências da ocorrência de reprodução da avifauna
estudada na região. Os métodos utilizados seguiram Simon e Pacheco (2005), em
consonância com Camargo e Pimentel (2012). No ponto de coleta foram encontrados
apenas ninhos artificiais de marreca, onde são criados pelos próprios marrequeiros. Para
acontecer a reprodução, o ambiente em que as D. autumnnalis estão inseridas, precisa
apresentar alguma cobertura vegetal como exemplo, as gramíneas, que tem como
finalidade diminuir a temperatura do local. Os ninhos artificiais da marreca, D.
autumnalis, são feitos utilizando uma parte de madeira de buriti coberta, muitas das vezes,
por areia ou vegetação e em apenas um ninho foi possível observar ovos e,
consequentemente, mensurar seu tamanho e diâmetro. Dessa forma, foi possível
descrever a estrutura dos ninhos construídos artificialmente, o ambiente de nidificação e
as características dos ovos da marreca.
Palavras-chave: Conservação. Ninho. Cativeiro.
Área de Interesse do Simpósio: Ecologia e Biodiversidade

512
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A rica diversidade da fauna e flora silvestre, desde os tempos de colonização no
Brasil, sempre foi alvo de grandes interesses, seja por motivos que envolvem admiração
por esses animais, seja pela própria cobiça do homem, ou até mesmo para outas variadas
finalidades. Nesse sentindo, o uso dessas espécies sendo animais ou vegetais silvestres
interfere nas suas especificas funções no ambiente natural e, principalmente, na sua
existência (LIMA, 2007).
A marreca, Dendrocygna autumnalis (LINNAEUS, 1758), é considerada como
ave silvestre e um Anseriforme das Américas que ocorre nos Estados Unidos à Bolívia,
Argentina e com maior frequência no Brasil, sendo comum na região da Amazônia e rara
ou de pouca ocorrência nas regiões sudeste e sul do Brasil (BERGSTRON, 1999; SICK,
1997).
A Reserva Extrativista (RESEX) Marinha de Tracuateua, no Estado do Pará, é
uma Unidade de Conservação (UC) formada por regiões de campos naturais, além dos
tradicionais manguezais, onde os respectivos moradores que encontram-se ao seu
entorno, vivem da pesca artesanal, da extração de outros recursos naturais, como a
“tiração de caranguejo” (COSTA, 2014), e de uma prática denominada como “pesca de
marrecas” (RODRIGUES, SCHMITZ, 2017).
Segundo Nascimento e Antas (1995), aves da ordem Anseriformes, como a
marreca, são de grande interesse tanto para a caça de subsistência, amadorista ou
comercial de sua carne, além de muitas vezes ser criada como mascote. Para Rodrigues e
Schmitz (2017), a captura dessa fauna, também conhecida como “pesca de marrecas”,
pode ser uma prática recorrente em áreas de reservas e seus arredores, apresentando
aspectos conflitivos como o que ocorre na Reserva Extrativista Marinha de Tracuateua
(PA) e na sua área circundante. Nesse caso, a captura constante dessa ave pode causar
desequilíbrio ecológico para o ecossistema da região (SILVA, 2018).
Diante desse cenário, a análise dos aspectos ecológicos da espécie D. autumnalis
são importantes para o desenvolvimento de estratégia de conservação e manejo das
marrecas na região amazônica, tendo em vista que no Brasil, dados sobre a reprodução
de D. Autumnalis ainda é bastante escasso (CAMACHO, PIMENTEL, 2012). Assim, a
identificação dos pontos e períodos de desasagem e reprodução de D. autumnalis, são

513
Belém (PA), 11 a 13 de dezembro de 2019
essenciais para nortear a conservação dessas avifauna, pois são esses locais que os
anatídeos ficam mais vulneráveis, tornando-se impossibilitados de voar e escapar de
predadores naturais e ações antrópicas (NASCIMENTO, ANTAS, 1995).
Nesse sentido, este trabalho tem como objetivo divulgar registros de reprodução
de Dendrocygna autumnalis, em cativeiro, assim como a documentação e descrição das
características do seu ninho e do habitat de nidificação, no entorno da Reserva Extrativista
Marinha de Tracuateua (PA).
2. METODOLOGIA
2.1. ÁREA DE ESTUDO
A área de estudo está localizada no entorno da Reserva Extrativista Marinha de
Tracuateua, entre as coordenadas 0º57'18.1"S e 46º53'43.6"W, as margens dos campos
alagados no município de Tracuateua, PA (Figura 1). Dentro desse contexto, a RESEX
Marinha de Tracuateua foi criada através do Decreto Presidencial em 20 de maio de 2005,
totalizando uma área de aproximadamente 278,64 km2 (ICMbio, 2019), da qual, 168,8 km2
são áreas compostas por florestas de mangue (HAYASHI, 2018), 81,2 km2 por espelho
d`água, e os demais por restingas, praias, campos alagados e ilhas (MAIA et al., 2016).
Figura 1 – Localização da área de estudo e do ponto de coleta dos dados.
Fonte: Alexandre Reis, 2019.

514
Belém (PA), 11 a 13 de dezembro de 2019
2.2. COLETA DOS DADOS
O trabalho seguiu uma pesquisa exploratória, pois não havia evidências da
ocorrência de reprodução da avifauna estudada na região. Foi utilizada uma abordagem
quali-quantitativa, tendo em vista que foi fundamental a realização de uma entrevista
aberta, com uma família de marrequeiro, bem como, quantificar os dados obtidos da
mensuração dos ovos de D. autumnalis. Os métodos utilizados seguiram Simon e Pacheco
(2005), em consonância com Camargo e Pimentel (2012), como estudos primordiais para
auxiliar no trabalho. Utilizou-se uma câmera para os registros fotográficos e um
paquímetro foi essencial para mensurar as medidas dos ovos da marreca em estudo.
3. RESULTADOS E DISCUSSÃO
No ponto de coleta foi encontrado apenas ninhos artificiais (2 ninhos) de marreca,
onde são criados pelos próprios marrequeiros. Os mesmos informaram que os ninhos são
construídos apenas para a criação e reprodução doméstica (Figura 2). Quando as
marrecas, D. autumnalis, estão no período de chocarem os ovos não é recomendável
mexer com elas e nem no seu ninho, pois elas podem simplesmente deixar a ninhada ou
até mesmo quebrar seus ovos.
O estresse acaba sendo um estado comum entre as aves encontradas em cativeiro,
tendo em vista à falta de interações sociais com indivíduos coespecíficos, além da
privação de suas reais atividades cotidianas (COSTA, 2012). O comportamento das
marrecas de destruir ou deixar sua ninhada, no período de reprodução, pode estar
relacionado com o estado de estresse nesses ambientes, uma vez que não estão inseridas
nos seus habitats naturais.
Simon e Pacheco (2005), definem um ninho como determinado lugar onde um
pássaro escolhe para colocar seus ovos, independentemente da escavação, limpeza, forro,
ou a construção que executa. No presente estudo, apesar de ocorrer interferência do
homem na formação do ambiente de criação do ninho das marrecas, ainda assim, elas que
preparam o lugar para colocarem seus ovos.

515
Belém (PA), 11 a 13 de dezembro de 2019
Figura 2 - Ninho artificial da marreca, Dendrocygna autumnalis.
Fonte: Felipe Lima, 2019.
De acordo com o marrequeiro, ninhos naturais da marreca, D. autumnnalis, não
são encontrados nos campos naturais, dessa forma, nunca foi avistado nenhum momento
de reprodução nesses ambientes, porque elas já migram na fase adulta para os campos
(Figura 3). Apenas ninhos naturais da marreca Oureré, Dendrocygna viduata, são
encontrados nos campos naturais. Em relação aos ninhos naturais da marreca, D.
autumnalis, o entrevistado afirmou que já foram avistados em troncos de siribeiras, no
manguezal da RESEX Marinha de Tracuateua. A vista disso, ninhos naturais da marreca,
D. autumnnalis, foram encontrados, no estado de Rio de Janeiro, no interior de caules de
palmeiras, Acrocomia sp. (CAMACHO, PIMENTEL, 2012). Dessa maneira, essa espécie
prefere construir seus ninhos naturais em troncos de arvores ou no solo (SICK, 1997), e
não nos campos alagados.
Figura 3 – Campos alagados, no entorno da RESEX Marinha de Tracuateua, PA.
Fonte: Felipe Lima, 2019.

516
Belém (PA), 11 a 13 de dezembro de 2019
Atrelado a isso, segundo o marrequeiro, o momento de reprodução dessa ave
ocorre durante os meses de fevereiro a maio, período chuvoso. Sobretudo, para acontecer
a reprodução o ambiente em que as D. autumnnalis estão inseridas, precisa apresentar
alguma cobertura vegetal como exemplo, as gramíneas, que tem como finalidade diminuir
a temperatura do local.
Nesse período, após completarem 10 ovos ou quando termina o período de choca,
a marreca fêmea começa a revezar o choco com a marreca macho, demonstrando assim,
um comportamento em casal (Figura 4). O revezamento dura próximo de 24 horas,
quando chega o momento de trocarem de turno, elas cantam anunciando a outra marreca
que o seu turno terminou. Adicionalmente, para poder diferenciar os gêneros dessas
marrecas, é necessário observar seus tamanhos e seus cantos. No trabalho de Camacho e
Pimentel (2012), foram vistos dois indivíduos adultos junto com nove filhotes nadando
em um lago, no entanto, não foi possível saber se era um comportamento em casal.
Através das informações dos marrequeiros, a marreca, D. autumnnalis, apresenta
comportamento em casal, no ambiente de cativeiro.
Figura 4 - Marreca, Dendrocygna sutumnalis, no período de reprodução.
Fonte: Felipe Lima, 2019.
Conforme informado, as marrecas podem chegar até 20 ovos por ninhada (Figura
5), porém nem todos esses ovos chegam a gerar filhotes e, atrelado a isso, muitos desses
filhotes não conseguem chegar a vida adulta. A média de ovos encontrados por Bolen,
McDaniel e Cottam (1964), foi de 14 ovos e cerca de 10 ovos por Camacho e Pimentel
(2012), em ninhos naturais. No presente estudo, foram encontrados apenas sete ovos em

517
Belém (PA), 11 a 13 de dezembro de 2019
um ninho artificial, dessa forma, o resultado se difere da média dos outros estudos. Além
disso, como informação complementar, os filhotes nascem amarelos como patos
domésticos, além disso, se tornam independentes quando as marrecas mães entram
novamente em período de reprodução.
Figura 5 – ninho com ovos da marreca, Dendrocygna Autumnalis.
Fonte: Felipe Lima, 2019.
3.1. DESCRIÇÃO DOS NINHOS ARTIFICIAIS
Os ninhos artificiais da marreca, Dendrocygna autumnalis, são feitos utilizando
uma parte de madeira de buriti coberta por areia ou vegetação, como foi possível notar na
Figura 2. Entretanto, às vezes são utilizadas folhas como base para as aves fazerem seus
ninhos. Não foi possível realizar a mensuração da largura, altura e comprimento dos
ninhos artificiais, assim como o tamanho das marrecas, porque elas se encontravam no
período de reprodução. Por esse motivo, o recomendável pelos marrequeiros foi que se
tivesse o mínimo contato possível com as aves.
O ninho de uma das marrecas se encontrava as margens dos campos alagados,
apresentando um ambiente mais agradável, devido a presença abundante de vegetação e
a temperatura do habitat ser mais amena. Por outro lado, o segundo ninho se encontrava
mais distante das margens dos campos, localizando-se dentro do quintal do entrevistado,
onde o ambiente disponibilizava pouca cobertura vegetal, mas ainda assim era possível
encontrar gramíneas no local. Ambos os ninhos eram delimitados por um cercado,
limitando assim o ambiente em que elas vivem.

518
Belém (PA), 11 a 13 de dezembro de 2019
Em apenas um ninho foi possível observar ovos e, consequentemente, mensurar
seu tamanho e diâmetro. Com a ajuda do marrequeiro, somente um ovo foi mensurado,
pois as marrecas estavam no período reprodutivo, como foi citado anteriormente.
Entretanto, procedeu-se a medida com os devidos cuidado para preservar as
características e a formação do ovo. Nesse sentido, como resultados da medição o ovo
apresentou: 4,2 cm de altura e 3,1 cm de diâmetro.
4. CONSIDERAÇÕES FINAIS
Foram encontrados somente ninhos artificiais da marreca, Dendrocygna
autumnalis, nos campos alagados, na área circundante da RESEX Marinha de Tracuateua,
PA. Dessa forma, foi possível descrever a estrutura dos ninhos construídos, o ambiente
de nidificação e as características dos ovos.
Embora não tenha sido possível encontrar ninhos naturais de D. autumnalis nos
campos alagados, os moradores relataram que já avistaram ninhos naturais da marreca no
mangue da RESEX Marinha de Tracuateua, PA. Nesse caso, é possível que ninhos
naturais de D. autumnalis, possam ser encontrados no mangue. Dentro dessa perspectiva,
esta pesquisa poderá servir como norte para futuros trabalhos envolvendo registro de
reprodução da marreca, D. autumnalis, na região.

519
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
BERGSTRON, B. First reported breeding of black-bellied wistling-duck in Northern
Florida. Florida Field Naturalist 27 (4): 177-179, 1999.
BOLEN, E.G.; MCDANIEL, B.; COTTAM, C. Natural History of the Black--Bellied Tree
Duck (Dendrocygna autumnalis) in Southern Texas. TheSouthwestern Naturalist 9 (2):
78-88, 1964.
CAMACHO, I.; PIMENTEL, L.M.S. Registro reprodutivo da asa-branca, Dendrocygna
autumnalis, no estado de Rio de Janeiro, Brasil. Atualidades Ornitológicas, Nº 169, set.
/out. 2012.
COSTA, J. N. As novas regras do jogo para o acesso aos recursos naturais na Reserva
Extrativista Marinha de Tracuateu, no Pará. Pós-graduação (Ciências Ambientais),
Instituto de Filosofia e Ciências Humana, Universidade Federal do Pará, Belém-PA, 2014
COSTA, V. A. Aves silvestres criadas em cativeiro em Santa Bárbara do Pará:
aspectos sócio-culturais e etológicos. 2012. 68 P. Dissertação (Mestrado em Ecoetologia) -
Programa de Teoria e Pequisa do Comportamento, Universidade Federal do Pará, Belém,
2012.
HAYASHI, S.N. Uso e Conservação dos manguezais na costa Amazônia brasileira:
uma abordagem a partir de sensoriamento remoto. 147f. Tese (Doutorado em Biologia
Ambiental) – Instituto de Estudos Costeiros, Universidade Federal do Pará, Bragança-Pará,
2018.
INSTITUTO CHICO MENDES DE CONSERVAÇÃO DA BIODIVERSIDADE
(ICMBio). 2019. Disponível em:
<http://www.icmbio.gov.br/portal/unidadesdeconservacao/biomas-
brasileiros/marinho/unidades-de-conservacao-marinho/2293-resex-marinha-de-
tracuateua>. Acesso em: 13 de jun. de 2019.
LIMA, G. G. B. Conservação da fauna e da flora silvestres no Brasil: a questão do tráfico
ilegal de plantas e animais silvestres e o desenvolvimento sustentável. Revista Jurídica,
Brasília, DF, v. 9, n. 86, p.134-150, ago. /set. 2007.
MAIA, C.; LIMA, M.C.P.; SILVA, M.M.T.; PAIVA, S.P. Densidade e estrutura
populacional do caranguejo-uçá, Ucides cordatus (Linnaeus, 1763) na Reserva Extrativista
Marinha de Tracuateua, Pará, Brasil. Biota Amazônia, v. 6, n.2, p.86-92, 2016.
NASCIMENTO, J.L.X.; ANTAS P.T.Z. Anilhamento de Dendrocygna spp. No Brasil
entre 1973 e 1994 (Anseriformes: Anatidae). Ararajuba 3: 62-65,1995.
RODRIGUES, M. R.; SCHMITZ, H. Conflito social: o caso da “pesca de marrecas” na
reserva extrativista marinha de Tracuateua (Pa). Amazon.; Ver. Antropol. (Online),
v.9, n.1, p. 132 – 154, 2017.
SICK, H. Ornitologia Brasileira. 2. ed. Rio de Janeiro: Nova Fronteira,1997.
SILVA, S. M. Hipermapa como Ferramenta de Sensibilização Ambiental para a
Resex Marinha de Tracuateua – PA. 2018. pág.39. Trabalho de Conclusão de Curso –
Universidade Federal Rural da Amazônia, Campus de Capanema, Capanema-PA. 2018.

520
Belém (PA), 11 a 13 de dezembro de 2019
SIMON, J.E.; S. PACHECO. On the standardization of nest description of neotropical
birds. Revista Brasileira de Ornitologia 13 (2): 143-154, 2005.

521
Belém (PA), 11 a 13 de dezembro de 2019
INFLUÊNCIA CLIMÁTICA NA FLUTUAÇÃO SAZONAL DE INSETOS
CERATOPOGONÍDEOS CAPTURADOS NA ÁREA DE PROTEÇÃO
AMBIENTAL ILHA DO COMBU, BELÉM, PARÁ
Marcelo Alves Farias1; Keissy Karoline Pinheiro Miranda2; Glennda Juscely Galvão
Pereira3; Derek Chrystian Monteiro Leitão4; Valéria Lima Carvalho5
1 Mestre em Virologia. Instituto Evandro Chagas. E-mail: [email protected] 2 Mestre em Virologia. Instituto Evandro Chagas. E-mail: [email protected]
3 Especialista em Microbiologia. Instituto Evandro Chagas. E-mail:
[email protected] 4 Graduando em Nutrição. Universidade da Amazônia. E-mail:
[email protected] 5 Doutora em Biologia de Agentes Infecciosos e Parasitários. Instituto Evandro Chagas.
E-mail: [email protected]
RESUMO
Levantamentos entomofaunísticos podem fornecer informações sobre o ciclo biológico,
picos de ocorrência e densidade populacional de insetos, além de disponibilizar
informações sobre o grau de integridade dos ambientes em que se encontram, dos
processos biológicos dos ecossistemas naturais e da sazonalidade de doenças. Este
trabalho teve como objetivo inicial a captura de insetos hematófagos na APACOMBU,
um fragmento de floresta amazônica pertencente à região insular do município de Belém,
Pará, e a partir da abundância desses animais de importância médica em diferentes épocas
do ano, considerar que condições climáticas específicas diferenciadas podem interferir na
biologia de espécimes pertencentes a família Ceratopogonidae. Foram realizadas duas
excursões científicas para a área de estudo no ano de 2018, sendo utilizadas armadilhas
luminosas modelo CDC e realizadas capturas concomitantes por atração humana com
aspiradores manuais na obtenção dos insetos. Os lotes foram identificados e agrupados
em pools de acordo com critérios taxonômicos, data, quantidade, ponto e método de
captura, e a partir da riqueza da entomofauna obtida, estimou-se o parâmetro de
abundância para os espécimes de ceratopogonídeos. Foram capturados 12.646
exemplares de dípteros hematófagos totais na área de estudo, sendo 973 destes agrupados
na família Ceratopogonidae. Na primeira excursão, durante a estação chuvosa, foram
capturados 873 espécimes de insetos ceratopogonídeos, e na segunda excursão, realizada
na estação seca, foram capturados 100 representantes. Esses dados convergiram com a
biologia desses animais, bem como com as características climáticas detectadas na ilha
no período de captura, caracterizando a flutuação sazonal desses insetos nas diferentes
estações. Dessa forma, os estudos entomofaunísticos permitem, em adição, analisar a
funcionalidade de ecossistemas naturais, da mesma forma que alterações na quantificação
observadas por meio de estudos como este torna-se complementar para o entendimento
preliminar acerca das influências climáticas no estabelecimento e na circulação de agentes
etiológicos vetorizados por estes animais.
Palavras-chave: Ilha do Combu. Insetos Vetores. Ceratopogonidae.
Área de Interesse do Simpósio: Ecologia e Biodiversidade.

522
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O conhecimento ecológico e taxonômico de várias das principais ordens da
classe Insecta é geralmente restrito ou focalizado em espécies de importância econômica
ou médica. A ordem Diptera é uma das maiores entre esses artrópodes, com grande
diversidade de espécies, onde são conhecidas aproximadamente 153 mil espécies
representantes, distribuídas em 160 famílias em todo o mundo. No Brasil, ocorrem cerca
de 8.700 espécies e estão presentes na maioria dos habitats, ocupando diversos nichos
alimentares, podendo ser parasitas, predadores e hematófagos, quando, neste último,
passam a ter importância médica ao se tornarem vetores de doenças (PAPAVERO;
GUIMARÃES, 2000; CARVALHO et al., 2012).
No que diz respeito a família Ceratopogonidae, esta compreende um táxon de
insetos de pequenas dimensões, que variam entre 1-6 mm de comprimento, conhecidos
popularmente como maruins, mosquitos-do-mangue e mosquitos-pólvora (COSTA et al.,
2013). Este grupo é bastante diversificado, contendo mais de 5.900 espécies descritas no
mundo, com espécimes possuindo as asas em geral escuras e manchadas. Os indivíduos
adultos de ambos os sexos podem visitar flores em busca de substâncias açucaradas, mas
as fêmeas usualmente buscam uma alimentação rica em proteína animal para maturação
dos seus ovos (TRINDADE; GORAYEB, 2010).
Dentro desta família, o gênero Culicoides destaca-se por ser um grupo
cosmopolita cujas espécies são amplamente distribuídas no mundo, com destaque em
países na região neotropical, como o Brasil, onde são conhecidas pelo menos 103
espécies, das quais 82 ocorrem na Região Amazônica, onde estudos levantamentos
entomológicos voltados para o conhecimento da diversidade desse gênero foram feitos
nos Estados do Amazonas e Pará (TRINDADE; GORAYEB, 2010). Ao praticarem a
hematofagia, esses animais podem funcionar como vetores de várias espécies de
parasitas, mas é principalmente como vetores de viroses humanas e de animais que as
espécies de culicoides têm sua principal importância (SAKKAS et al., 2018).
Na Amazônia brasileira, local onde doenças como a malária e diversas
arboviroses apresentam grande importância epidemiológica, faz-se importante realizar
estudos de avaliação entomofaunística como este, a fim de conhecer as espécies de
dípteros hematófagos transmissores de agentes etiológicos circulantes nessa região e sua

523
Belém (PA), 11 a 13 de dezembro de 2019
correlação com fatores climáticos. Em particular, ressalta-se sua relevância em áreas
florestais como a Área de Proteção Ambiental Ilha do Combu (APACOMBU), que fica
muito próxima da capital do estado onde se localiza e que é considerado local turístico
muito visitado por brasileiros e estrangeiros, além de receber muitos praticantes de
atividades de ecoturismo, o que propicia maior contato de pessoas com esses possíveis
agentes circulantes na área (FARIAS, 2019).
Apesar desta proximidade da Ilha com a capital acabar por contribuir de algum
modo para a promoção de pressões antrópicas sobre a mesma, áreas amazônicas como a
APACOMBU se tornam bons focos de diversos estudos por apresentarem características
rurais-ribeirinhas com áreas cujas coberturas vegetais ainda se apresentam em grande
parte conservadas (NASCIMENTO et al., 2010; COSTA et al., 2015; FARIAS, 2019).
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
Segundo o Instituto de Desenvolvimento Florestal e da Biodiversidade do Estado
do Pará, a Ilha do Combu é uma das 39 ilhas que compõem o município de Belém, estando
situada a 1,5 km ao sul da cidade. É limitada ao norte pelas margens do rio Guamá, ao sul
circundada pelo furo São Benedito, à leste pelo furo da Paciência e à oeste pela Baía do
Guajará, entre as coordenadas geográficas de 01°29'20" (extremo norte), 01°31'11"
(extremo sul), 48°25'54" (extremo leste) e 48°29'34" (extremo oeste) (PARÁ, 1997).
Sua área de 15,972 km2 apresenta ecossistema típico de várzea, com paisagem
florestal exuberante em equilíbrio, formada por um mosaico peculiar de espécies
composto por árvores, arbustos, herbáceas, lianas e espécies de sub-bosque, abrigando
grande diversidade de avifauna, servindo como um belo exemplo de local com um dos
mais altos níveis da biodiversidade amazônica (MAUÉS et al., 2011).
O clima da Ilha do Combu é o equatorial, quente e úmido, com índice
pluviométrico anual médio de 2.500 mm e temperatura média anual de 27 °C. O solo da
ilha é típico de área de várzea, sofrendo influência direta das marés do estuário do Guamá,
com alta percentagem de siltes, sendo um pouco úmido e argiloso, com baixa infiltração
e relevo plano, pobre em oxigênio e nutrientes (SILVA; SAMPAIO, 1998).

524
Belém (PA), 11 a 13 de dezembro de 2019
Através da Lei Estadual nº 6.083 de 13/11/1997, a Ilha do Combu foi
reconhecida como área especialmente protegida, com o objetivo de proteção e restauração
da diversidade biológica, dos recursos genéticos, das espécies ameaçadas de extinção,
bem como para a promoção do desenvolvimento sustentável, através do ordenamento dos
recursos naturais e da melhoria da qualidade de vida da comunidade local (PARÁ, 1997).
2.2 EXCURSÕES CIENTÍFICAS PARA CAPTURA DE INSETOS
Durante o ano de 2018, foram realizadas duas excursões científicas para a
APACOMBU, com duração de aproximadamente 15 dias cada uma. A primeira delas,
correspondente a estação chuvosa amazônica, foi executada entre os meses
fevereiro/março (26/02 a 16/03) e a segunda ocorreu entre os meses de outubro/novembro
(15/10 a 02/11), e correspondeu a estação seca. Ambas excursões apresentaram as
mesmas características quanto à captura dos insetos e ocorreram no mesmo ponto da ilha,
sob as coordenadas geográficas S 01º30.114' e W 048º27.611' (Figura 1).
Figura 1– Ponto determinado para a realização das capturas dos insetos na área de estudo.
Fonte: Laboratório de Geoprocessamento – LABGEO/IEC.

525
Belém (PA), 11 a 13 de dezembro de 2019
Os insetos adultos capturados e destinados a realização deste estudo foram
coletados vivos, respeitando seus hábitos e biologia, utilizando-se de dois métodos: a
técnica de captura por atração humana, na qual esses animais são atraídos pelos odores
corporais naturais e se faz uso de instrumentos de captura manufaturados e de
equipamentos de proteção individual, sendo mantidos temporariamente em tubos de
ensaio. O outro método utilizado na obtenção dos insetos foi a captura por meio de
versões melhoradas de armadilhas luminosas desenvolvidas pelo Centro de Controle de
Doenças dos Estados Unidos (CDC), colocadas em funcionamento no período
compreendido entre às 12:00, quando se interrompia a captura por atração humana, até às
09:00 do dia seguinte. Ambas técnicas utilizadas na obtenção dos insetos deste estudo
foram aplicadas tanto em solo quanto em dossel de árvore.
Após capturados, os artrópodes foram armazenados em tubos KMA
identificados com data, ponto e método de obtenção, e conservados em nitrogênio líquido
(-196 ºC) até a chegada ao Laboratório de Entomologia da SAARB/IEC, onde os mesmos
foram armazenados em freezer a -70 ºC. Posteriormente, os insetos foram identificados
quanto ao grupo taxonômico a que pertenciam com o auxílio de microscópios
estereoscópicos (lupas) em mesa refrigerada a -20 ºC e embasadas com a utilização de
chaves dicotômicas propostas por Consoli e Oliveira (1994) e Forattini (2002), sendo
então, agrupados em lotes (pools) de acordo com os critérios de identificações
mencionados, conferindo a riqueza entomofaunística do fragmento da Ilha determinado
para captura.
Convém mencionar que foram realizadas medições da temperatura ambiente e
da umidade relativa do ar antes e após o início das capturas diárias, cujas médias dos
registros estão demonstrados no Quadro 1.

526
Belém (PA), 11 a 13 de dezembro de 2019
Quadro 1 – Intervalos das temperaturas ambientes e umidades relativas diárias registradas
nas duas excursões.
Estação Temperatura Ambiente Umidade Relativa
Menor Maior Média Menor Maior Média
Chuvosa 25 ºC
(15/03) 30.1 ºC (12/03)
27.8 ºC (26/02 a
16/03)
68 % (05/03)
91 % (Vários dias)
87.2 % (15/10 a
02/11)
Seca 26.9 ºC (26/10)
31 ºC (16, 22/10)
29.2 ºC (15/10 a
02/11)
48 % (16/10)
91 % (26/10)
64.8 % (15/10 a
02/11)
Fonte: Laboratório de Entomologia da Seção de Arbovírus e Febres Hemorrágicas (SAARB/IEC).
2.3 ANÁLISE ENTOMOFAUNÍSTICA
Com base nos dados disponibilizados com as identificações dos insetos
capturados, foi realizada uma análise descritiva a partir dos valores da riqueza de espécies
obtidos e que possibilitaram a estimativa do parâmetro da abundância dos insetos
pertencentes a família Ceratopogonidae, correspondendo ao número de espécimes
identificados agrupados neste grupo taxonômico, por refletir a influência climática e o
arranjo das condições ambientais, bem como dos recursos ecológicos disponíveis na área
de estudo para as população de insetos capturados.
3. RESULTADOS E DISCUSSÃO
Considerando os dois métodos de captura utilizados e as duas áreas determinadas
para a coleta, como resultados das amostragens dos insetos capturados nas duas excursões
realizadas durante o ano de 2018, foram obtidos 12.646 exemplares de dípteros
hematófagos totais na área de estudo. Após a identificação, foram agrupados 973
espécimes na família Ceratopogonidae: na primeira excursão, correspondente a estação
chuvosa amazônica, foram coletados 5.019 exemplares de insetos hematófagos, sendo
873 destes identificados como ceratopogonídeos. Já na segunda excursão, realizada na
estação seca amazônica, foram capturados 7.627 insetos, sendo 100 membros agrupados
na família-alvo deste estudo, conforme podem ser vistos nos Quadros 2 e 3.

527
Belém (PA), 11 a 13 de dezembro de 2019
Quadro 2 – Pools de insetos ceratopogonídeos obtidos na estação chuvosa.
Pool Quant. Datas de Captura Local Método
1 147 28/Fev. e 01/Mar.
Solo
Atração
Humana
2 157 02, 05/Mar.
3 153 27/Fev. e 06, 07/Mar.
4 142 08, 09, 12, 13, 14, 15, 16/Mar.
5 1 14/Mar. Dossel
6 1 28/Fev.
7 78 01, 02, 07, 09, 13, 14, 15, 16/Mar. Solo
CDC 8 147 01, 07, 08, 09, 16/Mar. Dossel
9 47 15/Mar.
10 3 17, 19/Out. Solo Atração
Humana 11 1 24/Out. Dossel
12 69 16, 17, 19, 23, 24, 25, 30, 31/Out. e
01/Nov. Solo
CDC
13 27 23, 24, 30/Out. e 01/Nov. Dossel
Ressalta-se a diferença observada na abundância dos insetos ceratopogonídeos
capturados por estação, o que sugere forte influência de fatores climáticos, como a
pluviosidade, na flutuação sazonal destes insetos durante as diferentes estações,
demonstrando-se a necessidade do desenvolvimento de estudos mais aprofundados
voltados para os representantes incluídos nessa família, reforçado pela possibilidade de
vetorização de agentes virais com potencial patogênico (SAKKAS et al., 2018).
Essa dinâmica sazonal de insetos culicoides, evidenciada com a avaliação de
abundância desses artrópodes por estação climática, está diretamente correlacionada com
a temperatura e a umidade típicos do microclima encontrado na APACOMBU. Esta
correlação foi apontada em um estudo de monitorização da dinâmica sazonal de
culicoides na Sérvia, nos períodos de 2006-2007, 2011-2012 e de 2015-2017. O estudo
apontou que esses fatores foram importantes influências nas flutuações a curto prazo
desses insetos em áreas epizoóticas de circulação de arbovírus naquele país (aumento

528
Belém (PA), 11 a 13 de dezembro de 2019
repentino no número) e depois na sua propagação a longo prazo (PAVLOVIĆ et al.,
2018).
Outras pesquisas pelo globo estabeleceram que a maioria dos insetos
ceratopogonídeos não é registrada quando a temperatura média atmosférica está abaixo
de 13 ºC e acima de 35 ºC, sendo numerosos dentro desse intervalo térmico tipicamente
registrado em áreas tropicais e, certamente, na Ilha do Combu (Ver Quadro 1), onde foram
realizadas as capturas. Além disso, climas mais quentes favorecem o desenvolvimento
larval o que reforça que estudos de levantamento entomofaunístico sejam realizados em
diferentes estações climáticas (PURSE et al., 2015; PAVLOVIĆ et al., 2016),.
Sabe-se que os maruins são relatados pela ampla capacidade de reprodução em
áreas periurbanas e por serem abundantes durante os meses quentes e chuvosos do ano
(TRINDADE; GORAYEB, 2010), o que está de acordo com o observado neste estudo.
As larvas de Culicoides paraensis, por exemplo, são mais ativas durante o dia, durante o
período de fortes chuvas e sob iluminação intensa, e podem se desenvolver em vários
habitats úmidos em períodos de seca, como florestas, margens de rios, solo úmido,
buracos de árvore em decomposição e contendo matéria orgânica de origem vegetal, tais
como bananas em apodrecimento, troncos e cascas de cacau (CASTELLÓN; VERAS,
2015), muito frequentes na vegetação da Ilha do Combu, nos arredores da área
determinada para a captura dos artrópodes que foram alvo deste estudo.
Esses dados entomofaunísticos podem ser associados com dados
epidemiológicos para melhorar a compreensão da sazonalidade de algumas enfermidades
virais, como é o caso da febre do Oropouche, transmitida por C. paraensis. Contudo,
ainda são escassos trabalhos desenvolvidos na Região Amazônica voltados para a
ecologia de insetos hematófagos silvestres de interesse médico, geradores de dados e
informações que contribuam para o entendimento de alguns importantes aspectos
ecoepidemiológicos de algumas arboviroses (GUEDES, 2012).
Convém mencionar, em adição, que esses dados preliminares da entomofauna
da APACOMBU obtidos com a captura e identificação dos insetos nortearam a aplicação
de testes investigativos de diagnóstico virológico nos insetos ceratopogonídeos obtidos
(FARIAS, 2019). A família Ceratopogonidae apresentou grande número de espécimes
capturados, o que, apesar de possuírem pequenas dimensões, permitiu-se agrupamentos
em um número de pools compatível com a proposta temporal da pesquisa adicional de

529
Belém (PA), 11 a 13 de dezembro de 2019
arbovírus aplicada nesse estudo (Dados não mostrados). Além disso, um número
considerável de diferentes vírus já foi isolado a partir de insetos Culicoides em todo o
mundo, muitos deles de grande interesse veterinário e médico (SAKKAS et al., 2018).
4. CONSIDERAÇÕES FINAIS
Estudos entomofaunísticos podem fornecer informações sobre ciclo biológico,
picos de ocorrência e densidade populacional de insetos, além de disponibilizar
informações sobre o grau de integridade dos ambientes em que se encontram, da sua
importância nos processos biológicos dos ecossistemas naturais e na transmissão de
doenças (CAJAIBA; SILVA, 2017). Inventariar e monitorar a variedade de insetos e as
alterações na quantificação por meio de estudos como este, torna-se essencial para o
entendimento preliminar da biodiversidade em ecossistemas como na área estudada, e de
que forma elas podem influenciar indiretamente no estabelecimento e na expansão de
agentes patogênicos.
Por conseguinte, é sabido que fatores microclimáticos e bióticos podem
influenciar a capacidade vetorial de populações adultas de insetos na transmissão de
doenças. Tais fatores, particularmente a temperatura e a umidade, influenciam a
proporção de culicoides adultos capazes de transmitir vírus por causar impacto na duração
do período larval, na sobrevivência adulta, na taxa de picadas e na competência de vetores
em sequência, por exemplo (PURSE et al., 2015).
Além disso, a diferença na quantidade de insetos capturados na estação chuvosa
quando comparada com a quantidade de insetos obtida na estação seca pode ser,
futuramente, associada a estudos epidemiológicos preditivos de surtos ou epidemias de
doenças que pudessem ocorrer nesta estação, uma vez que um maior número de insetos
circulantes aumenta as chances de contato com hospedeiros humanos em potencial que
visitam a Ilha com objetivos turísticos em tempos chuvosos, sendo sugestiva a promoção
de medidas de controle desses vetores nessas épocas.
Nesse contexto, os levantamentos populacionais de insetos tornam-se
imprescindíveis em áreas amazônicas protegidas, pois, além de amostrar a densidade
populacional das espécies de insetos em determinado local, permitem caracterizar sua
comunidade e podem contribuir na vigilância em saúde e na promoção de medidas

530
Belém (PA), 11 a 13 de dezembro de 2019
preventivas em áreas turísticas, como a APACOMBU, que apresenta aumento na
interferência ambiental de seus ecossistemas.

531
Belém (PA), 11 a 13 de dezembro de 2019
5.REFERÊNCIAS
CAJAIBA, R. L.; SILVA, W. B. Levantamento de entomofauna em arborização urbana
no município de Uruará, Pará, norte do Brasil. Biota Amazônia. v. 7, n. 1, p. 69-73, jan.
2017.
CASTELLÓN, E. G.; VERAS, R. S. (Org.). Maruins (Culicoides: Ceratopogonidae) na
Amazônia brasileira. Manaus: INPA, 2015. 144p., il. Color.
CARVALHO, et al. Diptera Linnaeus, 1758. In: RAFAEL, J. A.; MELO, G. A. R.;
CARVALHO, C. J. B.; CASARI, S. A.; CONSTANTINO, R. (Eds.). Insetos do Brasil:
diversidade e taxonomia. Ribeirão Preto: Holos Editora, p. 701-743, 2012.
CONSOLI, R. A. G. B.; OLIVEIRA, R. L. Principais mosquitos de importância
sanitária no Brasil. Rio de Janeiro: Fiocruz, 1994, 228 p., il.
COSTA, et al. Espécies de Culicoides (Diptera; Ceratopogonidae) e hospedeiros
potenciais em área de ecoturismo do Parque Nacional dos Lençóis Maranhenses, Brasil.
Revista Pan-Amazônica de Saúde. v. 4, n. 3, p. 11-18, set. 2013.
COSTA, et al. Ilha do Combu: realidades e desafios. Revista Interdisciplinar Saúde e
Meio Ambiente. v. 4, n. 2, p. 32-48, jul./dez. 2015.
FARIAS, M. A. Avaliação Entomofaunística e Pesquisa de Arbovírus na Área de
Proteção Ambiental Ilha do Combu, Belém, Pará. 90 f. Dissertação (Mestrado em
Virologia) – Instituto Evandro Chagas, Programa de Pós-Graduação em Virologia,
Ananindeua, 2019.
FORATTINI, O. P. Culicidologia Médica. São Paulo: EDUSP, v. 2, 2002. 864 p., il.
GUEDES, M. L. P. Culicidae (diptera) no Brasil: relações entre diversidade, distribuição
e enfermidades. Oecologia Australis. v. 16, n. 2, p. 283-296, jun. 2012.
MAUÉS, et al. Composição florística e estrutura do estrato inferior da floresta de várzea
na área de proteção ambiental Ilha do Combu, município de Belém, estado do Pará.
Revista Árvore. Viçosa, MG, v. 35, n. 3, p. 669-677, 2011. Edição especial.
NASCIMENTO, et al. Um estudo dos problemas ambientais da área de proteção
ambiental da ilha do Combu, Belém-Pa. In: I Congresso Brasileiro de Gestão Ambiental,
2010, Bauru/SP. Anais... – CONGRESSO BRASILEIRO DE GESTÃO AMBIENTAL.
2010.
PAPAVERO, N.; GUIMARÃES, J. H. The Taxonomy of Brazilian Insects Vectors of
Transmissible Diseases (1900-2000) - Then and Now. Memórias do Instituto Oswaldo
Cruz. Rio de Janeiro, v. 95, Supl. 1, p 109-118, 2000.
PARÁ. Lei Estadual 6.083, de 13 de novembro de 1997 – Dispõe sobre a criação da Área
de Proteção Ambiental da Ilha do Combu no Município de Belém. Diário Oficial do
Estado. Poder Executivo, PARÁ, 17 nov. 1997.
PAVLOVIĆ, et al. Impact of environmental factors on the biodiversity of Culicoides
(Insecta: Ceratopogonidae) in Serbia. Book of Abstracts of International Conference

532
Belém (PA), 11 a 13 de dezembro de 2019
on Ecological Crisis: Technogenesis and Climate Change. Belgrade, Serbia, p. 71-72,
2016.
______. et al. Study results of the presence of Culicoides spp. in Serbia during 2017.
Arhiv Veterinarske Medicine, v. 11, n. 1, p. 45-51, 2018.
PURSE, et al. Bionomics of temperate and tropical Culicoides midges: Knowledge gaps
and consequences for transmission of Culicoides-borne viruses. Annual Review of
Entomology. v. 60, p. 373-392, jan. 2015.
SAKKAS, et al. Oropouche Fever: A Review. Viruses. v. 10, n. 4, p. 175, abr. 2018.
SILVA, C. A. R.; SAMPAIO, L. S. Speciation of phosphorus in a tidal floodplain forest
in the Amazon estuary. Mangrove And Salt Marshes. v. 2, n. 1, p. 51-57, mar. 1998.
TRINDADE, R. L.; GORAYEB, I. S. Maruins (Diptera: Ceratopogonidae: Culicoides),
após a estação chuvosa, na Reserva de Desenvolvimento Sustentável Itatupã-Baquiá,
Gurupá, Pará, Brasil. Revista Pan-Amazônica Saúde. v. 1, n. 2, p. 121-130, 2010.

533
Belém (PA), 11 a 13 de dezembro de 2019
DEPOSIÇÃO DE SERAPILHEIRA E NUTRIENTES EM ÁREA DE
RESTAURAÇÃO APÓS MINERAÇÃO DE BAUXITA NA AMAZÔNIA
ORIENTAL
Julia Isabella de Matos Rodrigues1; Walmer Bruno Rocha Martins2; Gracialda Costa
Ferreira3; Mário Lima dos Santos4 Francisco de Assis Oliveira5
1 Graduanda em Engenharia Florestal. Universidade Federal Rural da Amazônia.
2 Mestre em Ciências Florestais. Universidade Federal Rural da Amazônia.
3 Dra. Profa. da Universidade Federal Rural da Amazônia.
4 Mestre em Ciências Florestais, Universidade de Brasília.
5 Prof. da Universidade Federal Rural da Amazônia. [email protected]
RESUMO
Apesar dos benefícios econômicos, o avanço da atividade minerária, como a exploração
da bauxita, provoca uma série de impactos ambientais. Para isso, faz-se necessário a
recuperação de áreas afetadas por tal prática, sendo que, o método mais utilizado é o
plantio de mudas de espécies arbóreas nativas. Neste sentido, o estudo busca responder a
seguinte questão: qual ecossistema em recuperação assemelha-se mais ao ecossistema de
floresta nativa? Com a hipótese de que se os ecossistemas em estágios mais avançados de
sucessão possuem maior estoque de serapilheira e nutrientes e por isso, estão próximos aos valores de um ecossistema de referência (floresta nativa ou testemunha), então o
ecossistema de 9 anos é o mais semelhante à floresta nativa. O objetivo do estudo foi
avaliar e quantificar o estoque de serapilheira e o conteúdo nutricional de quatro
ecossistemas em área de extração de bauxita. O trabalho foi conduzido na empresa
Mineração Paragominas S.A. (MPSA), do grupo Hydro e as coletas foram realizadas em
dois períodos: mais e menos chuvoso. O estudo evidenciou maior estoque de serapilheira
(7,12±0,34 Mg.ha-1) no ecossistema de floresta, sendo que, não houve diferença entre este
e o plantio de 9 anos nos dois períodos de coleta. Logo, a hipótese foi aceita (p<0,05),
mostrando que o plantio de 9 anos é o que apresenta maior semelhança ao ecossistema
testemunha.
Palavras-chave: Ciclagem de nutrientes. Recuperação de áreas degradadas. Plantio de
mudas.
Área de Interesse do Simpósio: Ecologia e Bidiversidade

534
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
Com o intuito de diminuir os impactos causados pela exploração da bauxita e pela
atividade minerária, busca-se, cada vez mais, restaurar o ambiente degradado por essas
práticas. Para isso, faz-se o plantio de mudas de diversas espécies arbóreas nativas da
região (FONSECA et al., 2017) e acompanha-se o desenvolvimento do ecossistema por
meio da avaliação de indicadores de características dos atributos da área em restauração
(PRACH; WALKER, 2019).
Um desses indicadores é a serapilheira, a qual corresponde à camada de resíduos
florestais depositados no solo, como folha, galhos e frutos (FERNANDES et al., 2018),
pois a decomposição é um processo essencial para manutenção dos ecossistemas, com o
retorno dos nutrientes ao solo e deste as plantas (VIVANCO; AUSTIN, 2019). Além
disso, a serapilheira é responsável pela proteção do solo contra os impactos causados pela
constante precipitação pluviométrica, principalmente na Amazônia (SCHUMACHER et
al., 2018). Logo, quanto maior a deposição e decomposição de serapilheira, maior será a
ciclagem de nutrientes e o desenvolvimento das plantas em ecossistemas em processo de
restauração (MACHADO et al., 2019).
Nesse sentido, avaliou-se quatro ecossistemas, sendo três em restauração (Plantios
de mudas com 9, 8 e 7 anos de idade) e uma floresta nativa adjacente para efeito de
comparação (Floresta Ombrófila Densa) a fim de responder a seguinte questão: qual
ecossistema em recuperação assemelha-se mais ao ecossistema de floresta nativa?
Partindo da hipótese de que se os ecossistemas em estágios mais avançados de sucessão
possuem maior estoque de serapilheira e nutrientes e por isso, estão próximos aos valores
de um ecossistema de referência (floresta nativa ou testemunha), então o ecossistema de
9 anos é o mais semelhante à floresta nativa. O objetivo desse estudo, portanto, foi avaliar
e quantificar o conteúdo nutricional de quatro ecossistemas em área de extração de
bauxita.
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO
A pesquisa foi desenvolvida em área da empresa Mineração Paragominas S.A.
(MPSA), do grupo Hydro, localizada a 70 km da sede municipal de Paragominas. O clima
da região é do tipo “Aw”, segundo a classificação de Köppen-Geiger. A temperatura

535
Belém (PA), 11 a 13 de dezembro de 2019
média é de 26,3 °C com índice pluviométrico anual de aproximadamente 1.800 mm, com
umidade relativa do ar em torno de 81% (ALVARES et al., 2013).
Foram selecionadas três áreas em processo de restauração pelo método de plantio
de mudas de espécies florestais nativas e uma área referência classificada como Floresta
Ombrófila Densa. O plantio de mudas de 9 anos recebeu subsolagem e adubação pré-
plantio com Fosfato natural: 33% P2O5 total, 10% solúvel em ácido cítrico e adubação
de cobertura com calcário dolomítico: CaO-28%, MgO-12%, sendo que esta última
também foi realizada para o ecossistema de 8 anos.
As espécies Croton matourensis Aubl. e Vismia guianensis (Aubl.) Choisy foram
encontradas em todos os ecossistemas e estão entre as espécies como maior Índice de
Valor de Importância (IVI), sendo que a primeira apresentou maior IVI (33,31%) no
ecossistema com 8 anos de restauração, enquanto a segunda, maior IVI (12,84%) no
ecossistema com 9 anos de restauração.
2.2 ESTOQUE DE SERAPILHEIRA E NUTRIENTES
A coleta foi realizada com auxílio de um gabarito metálico de 0,5 m x 0,5 m (0,25
m²) e 0,10 m de altura. Foram coletadas 6 amostras em cada parcela, totalizando 72 no
ecossistema com 8 anos de restauração e 48 amostras nos demais ecossistemas. As coletas
foram realizadas nos períodos mais e menos chuvoso (março e novembro,
respectivamente), ambos em 2018. Após a coleta, a serapilheira foi triada em duas
frações, lenhosa (ramos e material enrijecido) e não lenhosa (folhas e material não
enrijecido), e posteriormente, seca em estufa a 70 °C por 72 horas. Em seguida o material
foi pesado em balança analítica digital de precisão de 0,01 g e a porção não lenhosa foi
triturada em um moinho de lâminas do tipo Willey com peneira de 0,85 mm de abertura
para análise química de Nitrogênio (N), Fósforo (P), Potássio (K), Cálcio (Ca), Magnésio
(Mg) e Sódio (Na) no Laboratório da Embrapa Amazônia Oriental - PA, seguindo o
método descrito por Malavolta et al. (1997).
2.3 ANÁLISE DE DADOS
Conforme o delineamento inteiramente casualizado, os dados foram submetidos ao
teste de normalidade de Shapiro-Wilk. Posteriormente, para comparar os diferentes
ecossistemas no mesmo mês, a hipótese foi testada por meio do teste Tukey ao nível de
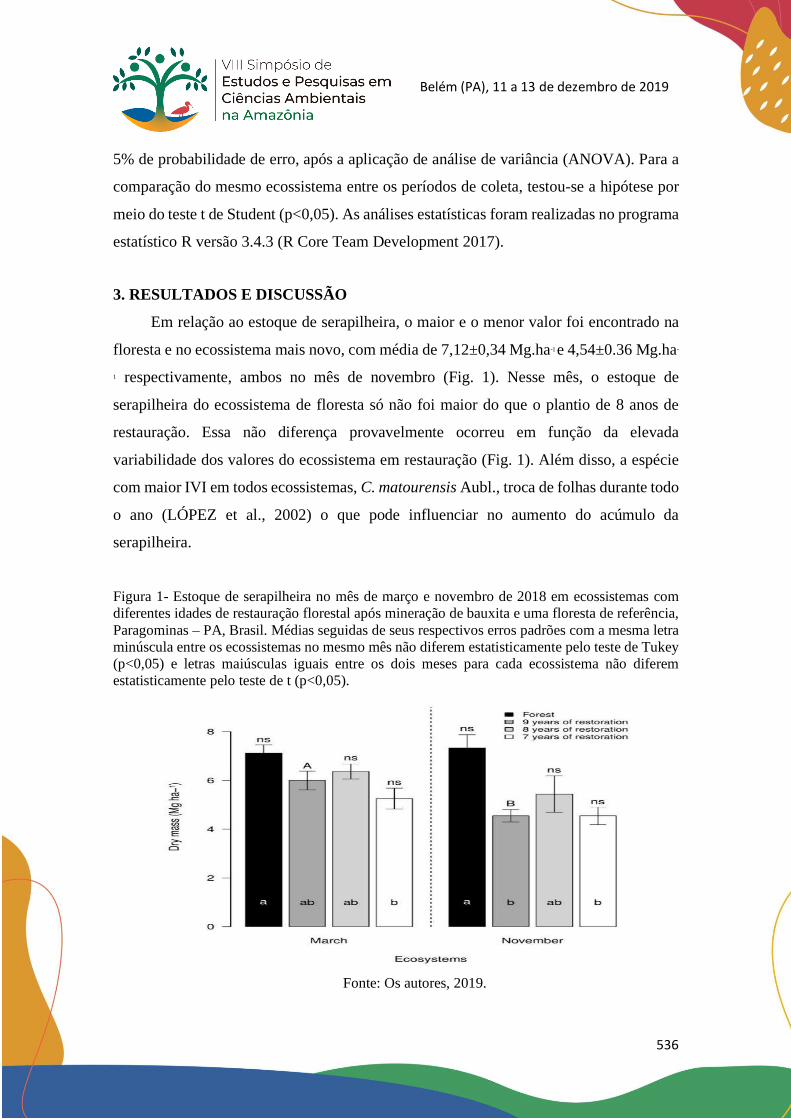
536
Belém (PA), 11 a 13 de dezembro de 2019
5% de probabilidade de erro, após a aplicação de análise de variância (ANOVA). Para a
comparação do mesmo ecossistema entre os períodos de coleta, testou-se a hipótese por
meio do teste t de Student (p<0,05). As análises estatísticas foram realizadas no programa
estatístico R versão 3.4.3 (R Core Team Development 2017).
3. RESULTADOS E DISCUSSÃO
Em relação ao estoque de serapilheira, o maior e o menor valor foi encontrado na
floresta e no ecossistema mais novo, com média de 7,12±0,34 Mg.ha-1 e 4,54±0.36 Mg.ha-
1 respectivamente, ambos no mês de novembro (Fig. 1). Nesse mês, o estoque de
serapilheira do ecossistema de floresta só não foi maior do que o plantio de 8 anos de
restauração. Essa não diferença provavelmente ocorreu em função da elevada
variabilidade dos valores do ecossistema em restauração (Fig. 1). Além disso, a espécie
com maior IVI em todos ecossistemas, C. matourensis Aubl., troca de folhas durante todo
o ano (LÓPEZ et al., 2002) o que pode influenciar no aumento do acúmulo da
serapilheira.
Figura 1- Estoque de serapilheira no mês de março e novembro de 2018 em ecossistemas com
diferentes idades de restauração florestal após mineração de bauxita e uma floresta de referência,
Paragominas – PA, Brasil. Médias seguidas de seus respectivos erros padrões com a mesma letra
minúscula entre os ecossistemas no mesmo mês não diferem estatisticamente pelo teste de Tukey
(p<0,05) e letras maiúsculas iguais entre os dois meses para cada ecossistema não diferem
estatisticamente pelo teste de t (p<0,05).
Fonte: Os autores, 2019.

537
Belém (PA), 11 a 13 de dezembro de 2019
A serapilheira acumulada no ecossistema de floresta diferiu significativamente do
mais jovem nos dois períodos de coleta. Esse resultado reflete o fato de que o segundo,
foi aquele onde ocorreu os impactos da mineração mais recentemente, apresenta maior
possibilidade de possuir um solo mais compactado, e consequentemente, apresenta menor
acúmulo de serapilheira (SILVA et al., 2018).
Em relação aos nutrientes, a concentração desses elementos é diretamente afetada
pelas condições climáticas, sobretudo pela precipitação pluviométrica (MARTINS et al.,
2018). Isto foi observado neste estudo, principalmente para K e Na (Fig. 2), já que ambos
apresentaram maior teor no período menos chuvoso. No mês mais chuvoso, o valor
superior de K foi encontrado na floresta, apesar de não haver diferença entre esse
ecossistema e o de 8 anos.
Figura 2- Concentração de macronutrientes na serapilheira estocada no solo nos diferentes
ecossistemas estudados. Médias e seus respectivos erros padrões. Mesma letra minúscula entre os
ecossistemas no mesmo mês não diferem estatisticamente pelo teste de Tukey (p<0,05) e letras
maiúsculas iguais entre os dois meses para cada ecossistema não diferem estatisticamente pelo
teste de t (p<0,05). ns = diferença não significativa.
Fonte: Os autores, 2019.

538
Belém (PA), 11 a 13 de dezembro de 2019
Os baixos valores encontrados para K são explicados pela alta mobilidade deste
elemento no tecido vegetal, o que facilita sua retranslocação antes da abscisão foliar
(GODINHO et al., 2013) somado ao fato de que a maior parte deste elemento é encontrada
na forma solúvel e na matéria orgânica, o K é liberado por lavagem (FAQUIN, 2005).
O elemento químico N apresentou maior teor no período chuvoso para o
ecossistema natural de floresta. Entretanto, não houve diferença entre este e o ecossistema
de 7 anos, pois apesar de haver uma tendência de aumento de N em ecossistemas mais
jovens (MACHADO et al., 2016), a espécie com segundo maior IVI no ecossistema de
floresta é da família fabaceae, a qual faz associação com bactérias fixadoras do elemento
em questão (CALDEIRA et al., 2019).
Para o Ca e Mg, o maior valor foi encontrado no plantio de 8 anos de restauração e
estes não diferiram do ecossistema controle e do plantio de 9 anos, o que provavelmente
ocorreu devido à adubação de cobertura realizada com calcário dolomítico. Os altos
valores de Ca ocorrem devido à função estrutural desempenhada por este nutriente na
célula vegetal, o qual é um dos principais componentes da parede celular (GODINHO et
al., 2014). Além disso, uma das características do Cálcio é possuir baixa mobilidade
dentro da planta, ou seja, este é lentamente distribuído antes da abscisão foliar
(CARVALHO et al., 2019).
Nesse sentido, por meio do teste Tukey (p<0,05) a hipótese de que o plantio de
mudas com 9 anos possui maior semelhança com o ecossistema referência foi aceita, pois
apesar de algumas não diferenças nutricionais, o plantio de mudas de 9 anos de
restauração foi o único que não diferiu estatisticamente em relação ao estoque de
serapilheira nos dois períodos de coleta.
4. CONSIDERAÇÕES FINAIS
O plantio de mudas de 9 anos apresenta melhores indicadores de restauração
quando comparado aos plantios com idades inferiores.
A menor precipitação influencia no maior aporte de serapilheira.
A presença de Croton matourensis Aubl. entre as espécies com maior IVI acarreta
no maior acúmulo de serapilheira.
O teor nutricional é afetado diretamente pela adubação de base.

539
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
ALVARES, C. A. et al. Köppen’s climate classification map for Brazil. Meteorologische
Zeitschrift, v. 22, n. 6, p. 711–728, 2013.
CALDEIRA, M. V. W. et al. Litter as an Ecological Indicator of Forest Restoration
Processes in a Dense Ombrophylous Lowland Forest. Floresta e Ambiente, v. 26, n.
spe1, p. 1–11, 2019.
CARVALHO, H. C. DE S. et al. Estoque de nutrientes na serapilheira acumulada em
quatro tipos de vegetação no Cerrado em Goiás, Brasil. Revista Ecologia e Nutrição
Florestal - ENFLO, v. 7, n. May, p. 06, 2019.
FAQUIN, V. Nutrição Mineral das Plantas. [s.l.] Universidade Federal de Lavras-
UFLA, 2005.
FERNANDES, M. M. et al. Aporte de serapilheira em reflorestamento misto. Acta
Biológica Catarinense, v. 5, n. 3, p. 90, 2018.
FONSECA, D. A. et al. Avaliação da regeneração natural em área de restauração
ecológica e mata ciliar de referência. Ciencia Florestal, v. 27, n. 2, p. 521–534, 2017.
GODINHO, T. DE O. et al. Biomassa, macronutrientes e carbono Orgânico na
serapilheira depositada em trecho de floresta Estacional Semidecidual Submontana, ES.
Scientia Forestalis/Forest Sciences, v. 41, n. 97, p. 131–144, 2013.
GODINHO, T. DE O. et al. Quantificação de biomassa e nutrientes na serapilheira
acumulada em trecho de Floresta Estacional Semidecidual Submontana, ES. Cerne, v.
20, n. 1, p. 11–20, 2014.
LÓPEZ, C. V. DA C. et al. Fenologia de espécies da floresta secundária potencialmente
acumuladoras de fósforo. Biomass, n. 74, 2002.
MACHADO, D. L. et al. Organic matter and soil fertility in different successional stages
of seasonal semidecidual forest. Revista Caatinga, v. 32, n. 1, p. 179–188, 2019.
MALAVOLTA, E.; VITTI, G. C.; OLIVEIRA, S. . Avaliação do Estado Nutricional
das Plantas. 2. ed. Piracicaba: Associação Brasileira para Pesquisa de Potassa e do
Fosfato, 1997.
MARTINS, W. B. R. et al. Deposição de serapilheira e nutrientes em áreas de mineração
submetidas a métodos de restauração florestal em paragominas, Pará. Floresta, v. 48, p.
67–76, 2018.
PRACH, K.; WALKER, L. R. Differences between primary and secondary plant
succession among biomes of the world. Journal of Ecology, v. 107, n. 2, p. 510–516,
2019.
SCHUMACHER, M. V. et al. Aporte De Serapilheira E Nutrientes Em Uma Floresta
Estacional Decidual Na Região Central Do Rio Grande Do Sul. Ciência Florestal, v. 28,
n. 2, p. 532, 2018.
SILVA, K. DE A. et al. Estoque de serapilheira em uma floresta em processo de
restauração após mineração de bauxita. Rodriguesia, v. 69, n. 2, p. 853–861, 2018.

540
Belém (PA), 11 a 13 de dezembro de 2019
TAPIA-CORAL, S. et al. Influencia Da Massa E Nutrientes Da Liteira Sobre a
Composição Dos Macro-Invertebrados Em Plantíos Florestais Na Amazônia Peruana.
Folia Amazónica, v. 23, n. 2, p. 171, 2014.
VIVANCO, L.; AUSTIN, A. T. The importance of macro- and micro-nutrients over
climate for leaf litter decomposition and nutrient release in Patagonian temperate forests.
Forest Ecology and Management, v. 441, n. March, p. 144–154, 2019.

541
Belém (PA), 11 a 13 de dezembro de 2019
PROMOÇÃO DE CRESCIMENTO DE MUDAS DE JAMBÚ INOCULADAS
COM TECNOLOGIA MICROBIANA
Alice de Paula de Sousa Cavalcante1; Gleiciane Rodrigues dos Santos2; Ricardo
Christin Lobato Machado3; Rafael Rodrigo da Silva Costa4; Gisele Barata da Silva5.
1 Graduanda em Agronomia. Universidade Federal Rural da Amazônia. E-mail:
[email protected]. 2 Doutoranda em Agronomia. Universidade Federal Rural da Amazônia. E-mail:
[email protected]. 3Mestrando em Agronomia. Universidade Federal Rural da Amazônia. E-mail:
[email protected]. 4Mestrando em Agronomia. Universidade Federal Rural da Amazônia. E-mail:
[email protected]. 5Professora de Microbiologia. Universidade Federal Rural da Amazônia. E-mail:
RESUMO
O jambú é uma hortaliça não convencional nativa da região norte, além de ser muito
consumida para fins medicinais e como condimento. É produzida principalmente por
agricultores familiares, de forma que, para muitos, o cultivo do Jambú é uma importante
fonte de renda. A demanda crescente por esse produto estimula pesquisas voltadas para
aprimorar tecnologias de produção e, principalmente, que causem menor impacto ao meio
ambiente. Nesse cenário o uso de biofertilizantes é uma alternativa ao uso insumos
químicos, possibilitando que o agricultor forneça um produto com menor acumulo de
resíduos químicos ao consumidor. Nesse trabalho foram utilizados 3 isolados de
rizobactérias, mix de rizobactérias (R46 e R92), e o controle (apenas água), consistindo
assim de 5 tratamentos, inoculados nas sementes de jambú. O experimento foi realizado
em blocos inteiramente casualizados, contendo 5 repetições cada. Os dados de biometria
e clorofila foram submetidos a análise estatística e as médias foram comparadas pelo teste
SNK (P<0,05). Os isolados R1 e Mix(R46 e R92) promoveram maior crescimento e
aumento do teor de clorofila em relação ao tratamento controle. Resultados indicam que
esses isolados de rizobactérias contribuem para o manejo sustentável da produção de
jambú na região Amazônia, além de garantir maior segurança alimentar e ganhos de
produtividade.
Palavras-chave: Jambú. Biofertilizantes. Manejo sustentável.
Área de Interesse do Simpósio: Agronomia

542
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
O Jambú (Acmella oleraceae) é uma hortaliça não convencional pertencente a
família das Asteraceaes, nativa da Amazônia serve como especiaria e para fins
medicinais. Trata-se de uma hortaliça amplamente utilizada na culinária paraense. Desta
forma, esta espécie é muito importante como fonte de renda para os agricultores
familiares dos municípios do Pará, pois é considerada uma planta de múltiplo uso
(medicinal, condimentar e ornamental), reunindo elementos essenciais para formação de
um sistema sustentável (GUSMÃO et al., 2009).
A planta apresenta propriedades químicas importantes, que desperta o interesse da
indústria farmacêutica, principalmente pela presença do metabólito especial mais
abundante, considerado mais bioativo, o espilantol (BARBOSA 2016, pág. 1). Além do
espilantol, o Jambu contém outros constituintes nutricionais e funcionais, como elevado
conteúdo de ferro, cálcio e ainda possui as vitaminas B1, B2, niacina, C e pró-vitamina
A. Apresenta também baixo valor calórico, assim como, a maioria das hortaliças. Outra
propriedade relevante desta planta é sua capacidade antioxidante (BARBOSA 2016, pág.
1).
“A demanda crescente por hortaliças estimula estudos direcionados à tecnologias
que possibilitem a maior produtividade da cultura, visando sempre uma produção mais
limpa e sustentável (Acmella oleraceae), utilizou-se 4 isolados de rizobactérias para
verificar a sua capacidade de promoção de crescimento em plantas de Jambú. A promoção
de crescimento, que pode ser detectada de várias formas, como o aumento em altura ou matéria
seca ou, ainda, em produtividade, é desejável sob a maioria dos pontos de vista. Inúmeros
trabalhos já foram publicados sobre o assunto, como os de Freitas (1989), Silveira et al. (1995),
Dashti et al. (1998), Nandakumar et al. (2001), além de outros, para diferentes espécies vegetais.”
Sendo assim o objetivo do trabalho foi empregar o uso de microrganismos
benéficos do solo na produção de mudas de Jambú e assim diminuir o uso de insumos
químicos, garantindo maior segurança alimentar aos consumidores dessa hortaliça.
2. MATERIAL E MÉTODOS
1. Isolados: Os isolados de rizobactérias (R46, R92, R1 e Mix de 46 e 92)
utilizados no experimento são provenientes da coleção de microrganismos do Laboratório
de Proteção de Plantas da Universidade Federal Rural da Amazônia campus Belém.
2. Inoculação: os isolados de rizobactérias foram cultivadas em meio sólido 523
(Kado and Heskett 1970) durante 48 h, a 28 ºC. As suspenções bacterianas foram

543
Belém (PA), 11 a 13 de dezembro de 2019
preparadas com água destilada e esterilizada, e a concentração foi ajustada em
espectrofotômetro para A540 = 0,5 (108 UFC). . Posteriormente, preparou-se uma
suspensão bacteriana utilizando água estéril e ajustada a 540 nm de absorbância usando
um espectrofotómetro (A550 = 0,1 correspondendo a 108 U.F.C. mL-1) para
microbiolização das sementes de Jambú. Em seguida, as sementes de Jambú foram
imersas na suspensão bacteriana e colocadas em uma mesa giratória por 4 horas, a uma
velocidade de 110 rpm.
3. Avaliação do crescimento: A promoção do crescimento foi avaliada 30 dias
após a germinação A altura das plantas e o diâmetro do coleto foi mensurado com
paquímetro digital (precisão de 0,02 mm). O índice de robustez foi calculado pela razão
entre a altura da planta e diâmetro do coleto. O número de folhas foi avaliado através da
contagem direta das folhas.
4. Analise estatística: O experimento foi realizado em delineamento inteiramente
casualizado com 5 tratamentos, 3 isolados de rizobactérias, Mix de rizobactérias e o
controle (só agua) e 5 repetições. Os dados de crescimento foram submetidos à análise
de agrupamento usando os valores médios de cada indivíduo e o grau de similaridade foi
obtido pela distância Euclidiana padrão. Os grupos foram submetidos à análise de
variância (ANOVA) e as médias foram comparadas pelo teste Student-Newman-Keuls
(SNK), P < 0,05, usando o software R versão 3.1.
3. RESULTADOS E DISCUSSÃO
Todos os isolados testados promoveram crescimento nas mudas de Jambú (tabela
1). Para as variáveis de crescimento o aumento variou de 43% a 80% para altura da planta,
em 3% a 23% para o diâmetro do coleto, aumento de 91% no número de folhas, e aumento
de 43% no índice de robustez para o isola, e aumento de 43% no índice de robustez para
o isolado R46. Os isolados que induziram maiores teores de clorofila foram
respectivamente o Mix (15%) e R1 (12%).
O crescimento da parte aérea, raiz e número de folhas podem ser observados na
figura 1 que pode ser atribuído a capacidade das rizobactérias de induzir a síntese de
giberelinas e citocininas que regulam a expansão foliar e síntese de clorofilas (DODD et
al. 2010; KANG et al. 2014). Com o aumento da área foliar e número de folhas, aumenta
também a área fotossintetizante, pois contribui para a maior captação de luz. A mesma

544
Belém (PA), 11 a 13 de dezembro de 2019
lógica se observa para a maior expansão do sistema radicular, visto que a raiz consegue
absorver mais nutriente e maior teor de água.
Figura 1 – controle e Mix de bactérias
Fonte: Alice Cavalcante, 2019.
Quando se trata de massa fresca da parte aérea todos os tratamentos obtiveram
sucesso, porem os melhores isolados foram, respectivamente, R92 com média de 103% e
o isolado de R1 com média de 101%. A massa fresca da raiz também demonstrou
aumentos significativos, os melhores tratamentos foram dos isolados Mix de rizobactérias
com média de 13%, e R1 com aumento de 6,6%. Todos os tratamentos obtiveram aumento
na massa seca da parte aérea, com as melhores médias pertencentes aos isolados R46 com
média de 66,6%, e o isolado R1 com média de 28,5%. A determinação da massa de
matéria seca (da planta e/ou de suas partes: frutos, caule, folhas e outros) é mais
apropriada para a análise de crescimento (TAIZ; ZEIGER, 2004) e, via de regra a área
foliar é também determinada (BENINCASA, 2003).

545
Belém (PA), 11 a 13 de dezembro de 2019
Tabela 1 – Parâmetros biométricos avaliados nas mudas de Jambú.
aGrupos bIsolados cALT (cm) cTR cDC (mm) cIR
1 Controle 15,21 c 13,9 b 2,79 a 5,76 ab
2 R46 25,1 ab 14,5 ab 3,05 a 8,26 a
3 Mix de rizobactérias 26,8 a 18,1 a 3,44 a 7,82 a
4 R92 23,7 ab 15,3 ab 3,41 a 6,96 a
5 R1 21,7 b 13,5 b 2,88 a 7,7 a
CV (%)
9,74 9,39 2,38 5,18
Fonte: Próprio autor. (2019). aGrupos de similaridade de acordo com a matriz Euclidiana;
bIsolados de rizobactérias ( R 46, R 1, R 92, e Mix de 46 e 92) e cAvaliações biométricas: Altura
em cm; Tamanho da raiz em cm; Diametro do coleto em mm; Indice de robustez. *Médias com
letras iguais não diferem significativamente de acordo com o teste de Student-Newman-Keuls
(P<0,05).
O índice SPAD avalia o teor de nitrogênio presente nas folhas. O incremento na
produção da cultura está sendo otimizado por meio da utilização de biofertilizantes a base
de microrganismos procariotos, capazes de fixar no solo o nitrogênio atmosférico e
disponibilizá-lo para as plantas (JORDÃO, et al., 2010). Muitos estudos comprovam que
as bactérias associadas a rizosfera da planta, contribuem com a adubação nitrogenada da
mesma. Com base nos dados obtidos através do índice SPAD, avaliações biométricas,
massa seca e massa fresca da parte aérea e da raiz, pode se dizer que as rizobactérias
foram responsáveis pelo maior acumulo de matéria seca. Elevadas dosagens de
nitrogênio, indicam relação com o aumento da atividade das enzimas fotossintéticas e de
assimilação de nitrogênio (DIDONET et al., 1996).
As figuras abaixo demonstram médias por agrupamento dos índices de SPAD
(Figura 2), massa seca da raiz, massa seca da folha e massa fresca da parte aérea, é visível
que os isolados Mix e R1 obtiveram as melhores médias, de modo que “folhas bem
nutridas de nitrogênio tem maior capacidade de assimilar CO2 e sintetizar carboidratos

546
Belém (PA), 11 a 13 de dezembro de 2019
durante a fotossíntese (FERREIRA et al.1997)”. Resultando do maior acumulo de
biomassa, o que pode ser visualizado nos Figuras 3, 4 e 5.
Figura 2 – índice SP AD
Fonte: próprio autor. (2019).
Figura 3 – Massa seca raiz
Fonte: próprio autor. (2019).
abb
aab a
0
5
10
15
20
25
30
35
Controle R-46 R-46 +
R-92
R-92 R-1
Ch
la+
b(µ
g c
m-2
)
a a
a
a
a
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
0.1
Controle R-46 R-46 +
R-92
R-92 R-1
Mass
a s
eca d
e ra
ízes
(g)

547
Belém (PA), 11 a 13 de dezembro de 2019
Figura 4 – Massa seca das folhas
Fonte: próprio autor. (2019).
Figura 5 – Massa fresca aerea
Fonte: próprio autor. (2019).
4. CONCLUSÃO
Os isolados de rizobactérias promoveram crescimento e induziram o
acumulo relativo de clorofila. Os isolados R1 e Mix de 46 e 92 obtiveram médias de
destaque em quase todas as analises, exceto pelo diâmetro do coleto, onde o isolado R1
não demonstrou tanto vigo. Conclui-se que o uso da tecnologia microbiana na produção
de Jambú, pode contribuir para o manejo sustentável, pois promovem crescimento,
aumentando assim a produtividade da cultura.
aa a
aa
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
Controle R-46 R-46 + R-92 R-92 R-1
Mas
sa s
eca
de
fo
lhas
(g)
b
ab
a a a
0
0.5
1
1.5
2
2.5
3
3.5
4
Controle R-46 R-46 + R-92 R-92 R-1
Mass
a f
resc
a d
e fo
lhas
(g)

548
Belém (PA), 11 a 13 de dezembro de 2019
5. REFERÊNCIAS
DIDONET, A. D.; RODRIGUES, O.; KENNER, M. H. Acúmulo de nitrogênio e de
massa seca em plantas de trigo inoculadas com Azospirillum brasiliense. Pesquisa
Agropecuária Brasileira, Brasília, v.31, p. 645-651, 1996.
BENINCASA, M. M. P. Análise de crescimento de plantas (noções básicas). 2. ed.
Jaboticabal: Funep, 2003.
BARBOSA, Alan Franco. Tese de Doutorado Avaliação química e biológica do Jambu
(Acmella oleraceae L.) seco com ar frio e liofilizado. Seropédica, RJ Dezembro de
2016.
DASHTI, N.; ZHANG, F.; HYNES R. & SMITH, D.L. Plant growth-promoting
rhizobacteria accelerate nodulation and increase nitrogen fixaton activity by field
grown soybean [Glycine max (L.) Merr.] under short season conditions. Plant Soil,
200:205-213,1998.
DIDONET, A. D.; RODRIGUES, O.; KENNER, M. H. Acúmulo de nitrogênio e de
massa seca em plantas de trigo inoculadas com Azospirillum brasiliense. Pesquisa
Agropecuária Brasileira, Brasília, v.31, p. 645-651, 1996.
DODD IC, ZINOVKINA NY, SAFRONOVA VI, BELIMOV AA (2010) Rhizobacterial
mediation of plant hormone status. Ann Appl Biol 157:361–379. doi:
10.1111/j.17447348.2010.00439.
FARIAS, Vivian Dielly da Silva; MOREIRA, Sheila Doria; GOMES, Rafaelle Fazzi ;
SILVA, Josiane Pereira da; SOUZA, Gisele Teixeira de; Sergio GUSMÃO, Antonio
Lopes. Rendimento de variedades de jambu (Acmella oleracea) conduzidas em solo
e hidroponia NFT. Hortic. bras., v. 28, n. 2 (Suplemento - CD Rom), julho 2010.
FERREIRA, A. C. B. Efeitos da adubação com N, Mo, Zn sobre a produção,
qualidade dos grãos e concentração de nutrientes no milho. 1997. Tese (Mestrado) –
Universidade Federal de Viçosa, Viçosa, 1997.
FREITAS, S.S. Desenvolvimento de plântulas de café pela inoculação
de Pseudomonas sp. R. Bras. Ci. Solo, 13:31-34,1989.
KANG SM, KHAN AL, YOU YH, et al. Gibberellin production by newly isolated strain
Leifsonia soli SE134 and Its potential to promote plant growth. J Microbiol Biotechnol
24:106–112. doi: 10.4014/jmb.1304.04015, 2014.
RATZ, Raquel Jacheline et al. Potencial biotecnológico de rizobactérias promotoras
de crescimento de plantas no cultivo de milho e soja. ENGEVISTA, V. 19, n.4, p. 890-
905, outubro 2017.
GUSMÃO, Sergio Antonio Lopes; HOMMA, Alfredo Kingo Oyama; SANCHES,
Ronaldo da Silva; MENEZES, Antonio Jose Elias. Etnocultivo do jambu para
abastecimento da cidade de belém, estado do pará. Amazônia: Ci. & Desenv., Belém, v. 6,
n. 12, jan./jun. 2011.
NEVES, Jucimar Ferreira, DIAS, Leonardo Diogo Ehle; JÚNIOR, Santino Seabra

549
Belém (PA), 11 a 13 de dezembro de 2019
BORGES, Luciana da Silva; LOURENÇÃO, Willian Augusto Peres. Cultivo de Jambu
em campo abertosobre telas de sombreamento e termo refletoras. Enciclopedia
Biosfera – Goiânia, v.9, n.17; p.926, 2013.
NANDAKUMAR, R.; BABU, S.; VISWANATHAN, R.; RAGUCHANDER, T. &
SAMIYAPPAN, R. Induction of systemic resistance in rice against sheath blight
disease by Pseudomonas fluorescens. Soil Biol. Biochem., 33:603-612,2001.
SILVEIRA, A.P.D.; FREITAS, S.S.; SILVA, L.R.C.; LOMBARDI, M.L.C.O. &
CARDOSO, E.J.B.N. Interações de micorrizas arbusculares e rizobactérias
promotoras do crescimento em plantas de feijão. R. Bras. Ci. Solo, 19:205-211,1995.
SPOLAOR Leandro Teodoski ; GONÇALVES, Leandro Simões Azeredo;
SANTOS,Odair José Andrade Pais dos; OLIVEIRA, André Luiz Martinez de; SCAPIM,
Carlos Alberto; BERTAGNA, Filipe Augusto Bengosi; KUKI, Mauricio Carlos.
Bactérias promotoras de crescimento associadas a adubação nitrogenada de
cobertura no desempenho agronômico de milho pipoca. Bragantia, Campinas v. 75, n.
1, p.33-40, 2016.
TAIZ, L.; ZEIGER, E. (Ed.). Fisiologia vegetal. 3. ed. Porto Alegre: Artmed, 2004. cap.
16, p. 365-400.

550
Belém (PA), 11 a 13 de dezembro de 2019
USO DE MICRORGANISMOS PROMOTORES DE CRESCIMENTO
ASSOCIADOS À ADUBAÇÂO MINERAL EM UROCHLOA BRIZANTHA Hélio Elias de Aguiar1; Sidney Daniel Araújo da Costa2; Aline Figueiredo Cardoso3;
Thiago da Silva Carvalho4; Gisele Barata da Silva5
1 Graduação em andamento. Universidade Federal Rural da Amazônia.
[email protected]. 2Doutorado em andamento. Universidade Federal Rural da Amazônia.
[email protected]. 3Doutorado em andamento. Universidade Federal Rural da Amazônia.
[email protected] 4Doutor. Universidade Federal Rural da Amazônia. [email protected]
5Doutora. Universidade Federal Rural da Amazônia. [email protected]
RESUMO A utilização de adubação mineral pode causar problemas ambientais por conta da
lixiviação. O uso de microrganismos promotores de crescimento pode contribuir para o
aumento da produtividade do capim-marandú. o objetivo foi avaliar as alterações
morfogênicas do capim Urochola brizantha cv. Marandú em função da aplicação de
biopromotores de crescimento associadas a diferentes doses de NPK. Os parâmetros
biométricos calculados foram: taxa de aparecimento de folhas (TApF), taxa de
alongamento das folhas (TAlF). Foi determinado também o comprimento da parte aérea
e raíz, massa fresca e massa seca da folha e da raíz. A dose ótima encontrada de NPK foi
equivalente a 50% do recomendado, correspondendo a 25-40-20 kg de ha-1, o que garantiu
um incremento de 40% em comprimento radicular em plantas inoculadas, incrementou
56% em altura de plantas comparadas a plantas com 100% de adubação mineral
recomendada. A massa fresca da parte aérea também teve incrementos significativos de
15% e 35% com os tratamentos R92 e MIX, respectivamente. A massa fresca da raiz
apresentou incrementos significativos de 338% e 263% nesses parâmetros com a
aplicação dos inoculantes R92 e MIX, respectivamente. A taxa de aparecimento foliar
com 50% da adubação apresentou valores iguais ao tratamento com 100% da adubação
recomendada, enquanto que a taxa de alongamento apresentou incrementos de 19% com
o uso de R92 e MIX.
Palavras-chave: Manejo sustentável, Forragem, Biopromotores.
Área de Interesse do Simpósio: Agronomia

551
Belém (PA), 11 a 13 de dezembro de 2019
1. INTRODUÇÃO
A Urochloa brizantha cv. Marandu é uma alternativa viável para os pecuaristas
na implantação das pastagens, por conta das suas características agronômicas e índices
zootécnicos ocupando extensas áreas de cultivo em todo Brasil (TEIXEIRA, 2011). Essa
forrageira apresenta como característica bom valor nutritivo, menor estacionalidade na
produção, melhor relação folha/haste, e resistência à cigarrinha das pastagens, quando
comparada a outras variedades do mesmo gênero (EMBRAPA, 1985).
Atualmente cerca de 60 % das áreas de pastagem apresentam algum grau de
degradação em função do manejo inadequado, principalmente devido ao uso de taxas de
lotação acima da capacidade de suporte e à falta de adubação de manutenção (BROCH et
al., 2008). FAGUNDES et al. (2006) relataram que a baixa disponibilidade de nutrientes
é, certamente, um dos fatores que mais prejudicam a produtividade e a qualidade da
forrageira. Deste modo, alternativas para o estabelecimento de plantas forrageiras vêm
sendo estudadas de forma que busquem aumentar os efeitos positivos no seu
desenvolvimento, e minimizar os riscos ambientais diante do uso de produtos químicos
no pasto (FONSECA et al., 2011).
Uma estratégia é o uso de rizobactérias promotoras de crescimento na formação
de pastagens que possibilitam melhor desenvolvimento das forrageiras, além de
auxiliarem no uso sustentável do sistema, visando reduzir o uso da adubação mineral e
promover uso eficiente desses fertilizantes (GAZOLA et al., 2017). O uso do fungo
Trichoderma sp., também é uma estratégia para reduzir o uso de fertilizantes minerais e
possibilitar maior desenvolvimento do sistema radicular das plantas, gerando efeitos
benéficos no desenvolvimento da cultura (CHAGAS et al., 2016).
Mecanismos de ação que envolve a produção de hormônios vegetais relacionados
ao maior crescimento do sistema radicular, produção de ácidos orgânicos responsáveis
pela solubilização de fósforo e potássio, assim como mineralização da matéria orgânica
do solo pelos microrganismos promotores de crescimento são relatados (HAEMAD e
KIBRET, 2014), tais condições favorecem o maior desenvolvimento do capim. Desse
modo, o objetivo foi avaliar as alterações morfogênicas do capim Urochola brizantha cv.
Marandú em função da aplicação de biopromotores de crescimento associadas a
diferentes doses de NPK.

552
Belém (PA), 11 a 13 de dezembro de 2019
2. MATERIAL E MÉTODOS
O experimento foi conduzido em ambiente semi-controlado, na unidade de
produção de mudas de viveiro da Universidade Federal Rural da Amazônia (UFRA) (01
° 27′25 ″ S, 48 ° 26′36 ″ O) em Belém (PA). O clima regional de acordo com a
classificação de Köppen é Af (equatorial). Dez sementes de Urochloa brizantha cv.
Marandu foram semeadas em sacos de polietileno (20×30×0,05 cm) preenchidos com
substrato e mantidos em casa de vegetação.
O delineamento experimental foi inteiramente casualizado em esquema fatorial
5×3 (U. brizantha cv. Marandú não inoculado, inoculado com a rizobactéria Bacillus
Subtilis, inoculado com B. Subtilis + Trichoderma asperellum (MIX), e 5 porcentagens
de redução das doses de NPK (100%, 75%, 50%, 25%, 0%) com 5 repetições, cada
repetição corresponde a uma planta. As doses recomendadas de NPK kg ha-1 (50-80-40)
foram calculadas conforme Embrapa Cerrados e as reduções ocorreram a partir dessa
recomendação (Santos, 2016).
O isolado de Rizobactéria Bacillus Subtilis e Trichoderma asperellum foram
provenientes da coleção de microrganismos do Laboratório de Proteção de Plantas da
Universidade Federal da Amazônia. A rizobactéria foi cultivada em placas de Petri com
meio de cultura sólido 523 (Kado & Heskett, 1970) e incubadas por 24 horas a 28 °
C. A suspensão bacteriana foi preparada em água estéril e ajustada no espectrofotômetro
a 550 nm (108 UFC).
O fungo foi multiplicado em placas de Petri contendo o meio de cultura BDA
(batata, dextrose e ágar) em seguida, as placas foram incubadas em uma câmara de
crescimento por 96 h, aproximadamente 28ºC. Após a obtenção do isolado foi preparado
suspensão aquosa de esporos com concentração de 108 esporos/mL. A inoculação foi feita
por meio de rega 14 dias após a emergência das plântulas, cada saco de polietileno
recebeu uma suspensão de 5 mL.
Os seguintes parâmetros biométricos foram calculados de acordo com Gomide e
Gomide (2000): taxa de aparecimento de folhas (TApF), taxa de alongamento das folhas
(TAlF). Foi determinado também o comprimento da parte aérea e raíz. Aos 35 dias após
a emergência das plântulas, cinco plantas por tratamento foram coletadas e separadas em
parte aérea (lâminas foliar) e raízes. O material vegetal foi seco em estufa (60 ° C) até
massa constante e pesado a massa fresca e massa seca da folha e da raíz.

553
Belém (PA), 11 a 13 de dezembro de 2019
Os dados foram submetidos à análise de variância e, quando significativos,
realizou-se teste de regressão para as doses de NPK. Foi utilizado o software Sisvar 5.6
para as análises estatísticas.
3. RESULTADOS E DISCUSSÃO O comprimento radicular apresentou efeito significativo (P<0,05) com
comportamento quadrático em todos os tratamentos à medida que se aumentou as doses
de NPK (Figura 1). A dose ótima que promoveu melhor incremento das características
avaliadas foi utilizando 50% da recomendação de NPK. O incremento do sistema
radicular foi de 40% em plantas inoculadas, quando comparadas a plantas controle com
100% da adubação recomendada. O maior desenvolvimento radicular a partir do uso da
tecnologia microbiana ocorre em função das melhorias proporcionadas com essa
associação. Os microrganismos atuam no crescimento radicular, pois promove o
desenvolvimento de raízes devido à produção de fitohormônios, o que possibilita um
aumento da massa radicular. Além de auxiliar no processo de solubilização e absorção de
nutrientes pelas plantas (HARMAN et al., 2004).
Figura 1 – Comprimento da Parte Aérea de Urochloa brizantha associadas a diferentes
porcentagens de NPK.
Fonte: Costa et al. (2019).
A parte aérea apresentou maior desenvolvimento em função das doses de NPK
quando associadas aos biopromotores, apresentando comportamento quadrático (Figura
y = -0,001x2 + 0,0752x + 40,929 R² = 0,9067
y = -0,0013x2 + 0,1695x + 45,592 R² = 0,5251
y = -0,0036x2 + 0,36x + 42,155 R² = 0,90770
10
20
30
40
50
60
0 25 50 75 100
Co
mpri
mento
de
P.A
(cm
)
Doses de NPK (%)
Controle
R92
R92+Tricho

554
Belém (PA), 11 a 13 de dezembro de 2019
2), apresentando incremento de 56% nessa variável, em comparação com plantas não
inoculadas e com 100% da adubação recomendada. O maior desenvolvimento radicular
com maior quantidade de pêlos radiculares em plantas inoculadas, foram responsáveis
por promover maior capacidade de exploração de água e nutrientes pelas plantas. A
melhor arquitetura das raízes associadas aos mecanismos diretos de ação, como a
mineralização e solubilização dos nutrientes (HUNGRIA et al., 2016), promoveram
maior desempenho das plantas com o uso dos biopromotores de crescimento. Tal
comportamento sugere que houve maior solubilização do fertilizante aplicado, e maior
capacidade na aquisição dos recursos essenciais para o seu desenvolvimento, ou seja, a
inoculação contribuiu para o aumento de oferta e consequente absorção pelas plantas
(CANELLAS et al., 2008)
Figura 2 – Comprimento de parte aérea de Urochloa brizantha associadas a diferentes
porcentagens de NPK.
Fonte: Costa et al. (2019).
Foi observado um comportamento linear em massa fresca da folha em todos os
tratamentos (Figura 3). Apresentando incremento de 15% e 35% nos tratamentos R92 e
Mix, respectivamente, quando comparados a plantas controle com 100% da adubação
recomendada. O resultado é decorrente do valor ótimo da adubação combinada ao uso do
microrganismo. Compreende-se que a adubação para a forragem, pode favorecer o
desenvolvimento do número de folhas vivas e a diminuição do tempo de aparecimento
foliar (SALES et al., 2014). Segundo, Lucon 2009, o crescimento de plantas estimulado
y = -0,001x2 + 0,0752x + 40,929 R² = 0,9067
y = -0,0013x2 + 0,1695x + 45,592 R² = 0,5251
y = -0,0036x2 + 0,36x + 42,155 R² = 0,9077
0
10
20
30
40
50
60
0 25 50 75 100
Co
mpri
mento
de
P.A
(cm
)
Doses de NPK (%)
Controle
R92
R92+Tricho
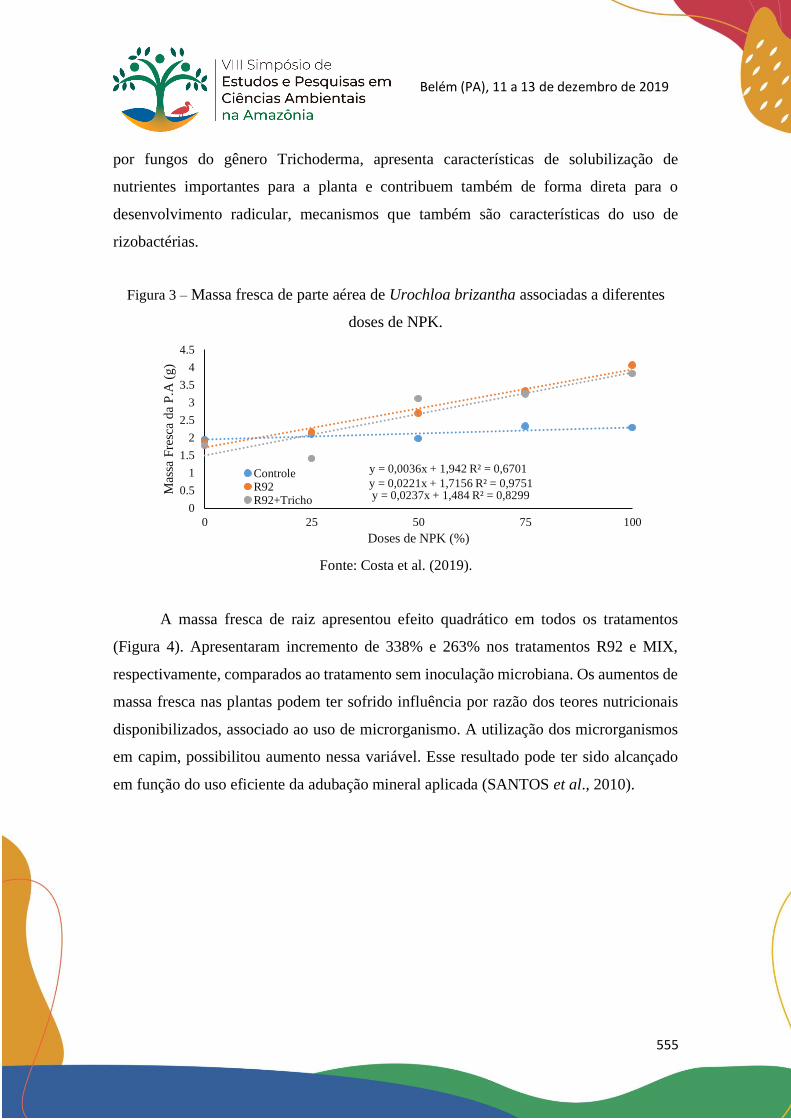
555
Belém (PA), 11 a 13 de dezembro de 2019
por fungos do gênero Trichoderma, apresenta características de solubilização de
nutrientes importantes para a planta e contribuem também de forma direta para o
desenvolvimento radicular, mecanismos que também são características do uso de
rizobactérias.
Figura 3 – Massa fresca de parte aérea de Urochloa brizantha associadas a diferentes
doses de NPK.
Fonte: Costa et al. (2019).
A massa fresca de raiz apresentou efeito quadrático em todos os tratamentos
(Figura 4). Apresentaram incremento de 338% e 263% nos tratamentos R92 e MIX,
respectivamente, comparados ao tratamento sem inoculação microbiana. Os aumentos de
massa fresca nas plantas podem ter sofrido influência por razão dos teores nutricionais
disponibilizados, associado ao uso de microrganismo. A utilização dos microrganismos
em capim, possibilitou aumento nessa variável. Esse resultado pode ter sido alcançado
em função do uso eficiente da adubação mineral aplicada (SANTOS et al., 2010).
y = 0,0036x + 1,942 R² = 0,6701
y = 0,0221x + 1,7156 R² = 0,9751y = 0,0237x + 1,484 R² = 0,8299
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
0 25 50 75 100
Mas
sa F
resc
a da
P.A
(g)
Doses de NPK (%)
ControleR92R92+Tricho

556
Belém (PA), 11 a 13 de dezembro de 2019
Figura 4 – Massa fresca da raiz de Urochloa brizantha associadas a diferentes
porcentagens de NPK.
Fonte: Costa et al. (2019).
Figura 5 – Massa seca da folha de Urochloa brizantha associadas a diferentes
porcentagens de NPK.
Fonte: Costa et al. (2019).
Foi observado que a produção de massa seca das raízes demostrou um
comportamento quadrático em todos os tratamentos (Figura 6). Considerando as doses de
NPK, observou-se que a dose reduzida de 50% somada ao uso de microrganismo
promoveu incremento de 242% e 200% nessa variável, nos tratamentos R92 e MIX,
respectivamente quando comparados a plantas controle com 100% da adubação
recomendada. O que afirma a eficiência do uso reduzido de NPK associado ao uso desses
y = -0,0025x2 + 0,2131x + 8,1925 R² = 0,7133
y = -0,0011x2 + 0,1861x + 8,2209 R² = 0,7573
y = -0,0022x2 + 0,2541x + 5,7523 R² = 0,8020
5
10
15
20
0 25 50 75 100
Mas
sa f
resc
a da
raiz
(g)
Doses de NPK (%)
Controle
R92
R92+tricho
y = 0,0012x + 0,214 R² = 0,9073
y = 0,0027x + 0,2643 R² = 0,940
y = 0,0028x + 0,252 R² = 0,7652
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0 25 50 75 100
Mas
sa s
eca
da
folh
a (g
)
Doses de NPK (%)
Controle
R92
R92+Tricho

557
Belém (PA), 11 a 13 de dezembro de 2019
biopromotores de crescimento. A utilização de rizobactérias promotoras de crescimento
promoveram incrementos na biomassa do capim Marandú com o uso de 50% da adubação
recomendada (PAUNGFOO-LONHIENNE et al., 2019). Segundo Chagas (2016), houve
um aumento de massa seca da parte aérea, massa seca da raiz em decorrência de maior
absorção de nutrientes.
Figura 6 – Massa seca da raiz de Urochloa brizantha associadas a diferentes
porcentagens de NPK.
Fonte: Costa et al. (2019).
A taxa de aparecimento foliar (Figura 7) e alongamento foliar (Figura 8)
demonstraram um comportamento linear em todos os tratamentos avaliados. Avaliando a
taxa de aparecimento foliar, o uso dos microrganismos apresentou valores iguais ao
tratamento controle com 100% de adubação. A taxa de alongamento foliar utilizando o
MIX proporcionou incremento de 19%, em relação ao tratamento controle com 100% da
adubação. De acordo com Oliveira et al. (2007), quando se encontra disponibilidade de
nitrogênio necessária, ocorre aumento do crescimento da planta, com alongamento dos
entrenós, ocorrendo a incidência de uma nova folha para fora da bainha da folha
precedente, o que pode causar aumento na taxa de aparecimento foliar. O teor nutricional
adequado disponível para a planta permitiu que ocorresse tal comportamento.
O nitrogênio possui essa característica responsável por estimular o alongamento
foliar, pois o seu acúmulo atua no aumento da expansão celular (YANG et al., 2009). O
aumento desta variável é crucial, pois, a partir do momento que ocorre o crescimento
y = -0,0003x2 + 0,0244x + 1,1895 R² = 0,9657
y = -0,0002x2 + 0,0286x + 1,0707 R² = 0,7009y = -0,0003x2 + 0,0391x + 0,9069 R² = 0,7466
0
0.5
1
1.5
2
2.5
3
0 25 50 75 100
Mas
sa s
eca
da
raiz
(g)
Doses de NPK (%)
Controle
R92
R92+Tricho
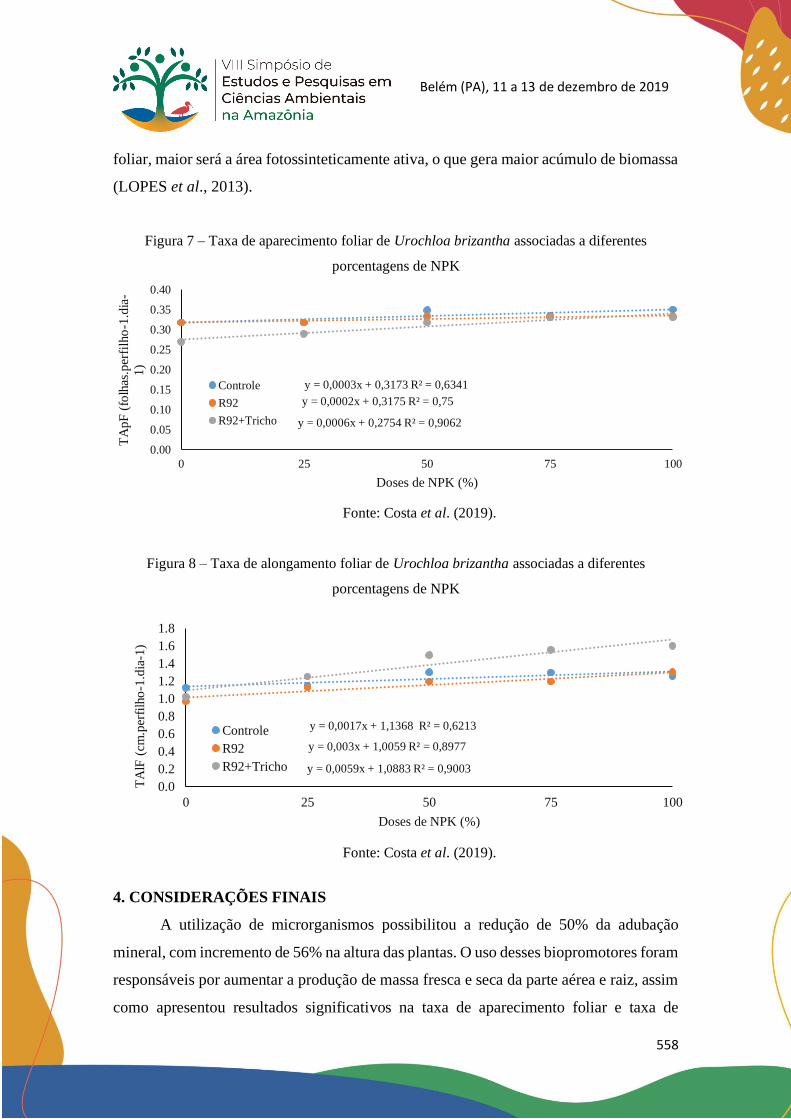
558
Belém (PA), 11 a 13 de dezembro de 2019
foliar, maior será a área fotossinteticamente ativa, o que gera maior acúmulo de biomassa
(LOPES et al., 2013).
Figura 7 – Taxa de aparecimento foliar de Urochloa brizantha associadas a diferentes
porcentagens de NPK
Fonte: Costa et al. (2019).
Figura 8 – Taxa de alongamento foliar de Urochloa brizantha associadas a diferentes
porcentagens de NPK
Fonte: Costa et al. (2019).
4. CONSIDERAÇÕES FINAIS
A utilização de microrganismos possibilitou a redução de 50% da adubação
mineral, com incremento de 56% na altura das plantas. O uso desses biopromotores foram
responsáveis por aumentar a produção de massa fresca e seca da parte aérea e raiz, assim
como apresentou resultados significativos na taxa de aparecimento foliar e taxa de
y = 0,0003x + 0,3173 R² = 0,6341
y = 0,0002x + 0,3175 R² = 0,75
y = 0,0006x + 0,2754 R² = 0,9062
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0 25 50 75 100
TA
pF
(fo
lhas
.per
filh
o-1
.dia
-
1)
Doses de NPK (%)
Controle
R92
R92+Tricho
y = 0,0017x + 1,1368 R² = 0,6213
y = 0,003x + 1,0059 R² = 0,8977
y = 0,0059x + 1,0883 R² = 0,9003
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
0 25 50 75 100
TA
lF (
cm.p
erfi
lho
-1.d
ia-1
)
Doses de NPK (%)
Controle
R92
R92+Tricho

559
Belém (PA), 11 a 13 de dezembro de 2019
alongamento das folhas em tratamentos com adubação reduzida e associados a
microrganismos promotores de crescimento.
5. AGRADECIMENTOS
A UFRA pelo apoio e disponibilização do espaço, a FAPESPA pela
disponibilização dos recursos financeiros destinados a bolsa de estudos.

560
Belém (PA), 11 a 13 de dezembro de 2019
6. REFERÊNCIAS
AHEMAD, M.; KIBRET, M. Mechanisms and applications of plant growth promoting
rhizobacteria: current perspective. Journal of King Saud University-Science, v. 26, n. 1,
p. 1-20, 2014.
ALCÂNTARA, P.B. Origem das Brachiarias e suas características morfológicas de
interesse forrageiro. In: ENCONTRO PARA DISCUSSÃO SOBRE CAPINS DO
GÊNERO BRACHIARIA, L, Nova Odessa, 1986. Anais... Nova Odessa: Instituto de
Zootecnia, 1987. p.1-18.
CANELLAS, L. P. et al Reações da matéria orgânica. In: SANTOS, G. A. et al. (Ed.).
Fundamentos da matéria orgânica do solo: ecossistemas tropicais subtropicais, 2. ed.
Porto Alegre: Metrópole, 2008. p. 45-63.
CHAGAS, L. F. B.; CASTRO, H. G.; COLONIA, B. S. O.; CARVALHO FILHO, M.
R.; MILLER, L. O.; CHAGAS JUNIOR, A. F. Efficiency of Trichoderma spp. as a
growth promoter of cowpea (Vigna unguiculata) and analysis of phosphate solubilization
and indole acetic acid synthesis. Brazilian Journal of Botany, São Paulo-SP, v. 38, n. 4,
p. 1- 11, 2016.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Centro Nacional de
Pesquisa de Gado de Corte. Brachiaria brizantha v. Marandu. Campo Grande, 1985. 31p.
(EMBRAPA/CNPGC. Documentos, 21).
HARMAN, G.E. et al. Trichoderma species – opportunistic, avirulent plant symbionts.
Nature, v. 2, p. 43-56, 2004.
LUCON, C.M.M. (2009) - Promoção de crescimento de plantas com o uso de
Trichoderma spp (em linha). Infobibos, Informações Tecnológicas. (Acesso em
2010.05.31).
SANTOS, A.R. & MONTEIRO, F.A. Produção e perfilhamento de Brachiaria decumbens
Stapf. em função de doses de enxofre. Sci. Agric., 56:689-692, 1999
SANTOS, H. A.; MELLO, S. C. M.; PEIXOTO, J. R. Associação de isolados de
Trichoderma spp. e ácido indol-3-butírico (AIB) na promoção de enraizamento de estacas
e crescimento de maracujazeiro. Bioscience Journal, Uberlândia-MG, v. 26, n. 6, p. 966-
972, 2010.
Teixeira, P. Quem sabe responde: Brachiaria e Panicum são o mesmo capim?
BlumenauSC: Rural Centro, 21 set. 2011..
Fonseca, D. M., Martuscello, J. A. & Santos, M. E. R. (2011). Adubação de pastagens:
inovações e perspectivas. In: Anais do XXI Congresso Nacional de Zootecnia.
Universidade Federal de Alagoas, Brasil, 1- 13.
OLIVEIRA, A.B.; PIRES, A.J.V.; MATOS NETO, U. et al. Morfogênese do capim-
tanzânia submetido a adubações e intensidades de corte. Revista Brasileira de Zootecnia,
v.36, n.4, p.1006-1013, 2007.

561
Belém (PA), 11 a 13 de dezembro de 2019
PAUNGFOO-LONHIENNE C., REDDING M., PRATT C. and WANG W. (2019) Plant
growth promoting rhizobacteria increase the efficiency of fertilisers while reducing
nitrogen loss. Journal of Environmental Management, 233, 337-341.




















