
Vermetid gastropods from Sao Miguel, Azores: Comparative anatomy, systematic position and...
20
Af;OREANA, SUPPLEMENT 1995: 173-192 VERMETID GASTROPODS FROM SAO MIGUEL, AZORES: COMPARATIVE ANATOMY, SYSTEMATIC POSITION AND BIOGEOGRAPHIC AFFILIATION Rildiger Bieler Center for Evolutionary and Environmental Biology, Field Museum of Natural History, Chicago, Illinois 60605, U.S.A. ABSTRACT Two vermetid species inhabit the intertidill and upper subtidal rocky shores of Sao Miguel, Azores. The larger-shelled species, identified as Vermetus cf. triquetrus Bivona-Bernardi, 1832, has highly variable shell characters, spanning previously assumed diagnostic features of several nominal genera. The smaller form is recognized as Thylaeodus cf. rugu- losus Monterosato, 1878, a species previously deemed restricted to the Mediterranean. Females of both species brood their eggs in stalked capsules attached to the interior shell wall. After feeding on nurse eggs, juveniles are released at the crawling stage and attach to suitable sub- stratum. Neither form is known to date from the western Atlantic, and colonization of the mid-Atlantic Azores by these vermetids is hypothe- sized as passive long-distance dispersal by "rafting" from an eastern Atlantic/Mediterranean source. A lectotype is selected for nominal spe- cies Vermetus rugulosus Monterosato, 1878 (Museo Civico di Zoologia, Rome). The commonly used name "Vermetus triqueter" is dismissed as an incorrect subsequent spelling of Vermetus triquetrus Bivona-Bernar- di, 1832. RESUMO Duas especies de vermetfdeos habitam o intertidal e subtidal superior das costas rochosas de Sao Miguel, Ayores. A especie de concha maior, identificada como Vermetus cf. triquetrus Bivona-Bernardi, 1832 possui caracteres conquiliol6gicos altamente variaveis, abrangendo varios gene- ros nominais caracterfsticas antes tidas como diagnosticantes. A forma menor e reconhecida como Thylaeodus cf. rugulosus Monterosato, 1878, especie previamente tida como restricta ao Mediterraneo. As femeas de ambas as especies incubam os ovos em capsulas pedunculadas fixas a pa- rede interior da concha. Depois de se alimentarem de ovos de criayao, os juvenis sao largados na fase rastejante e fixam-se a um substrato adequa- do. Ate a data nenhuma das formas e conhecida do Atlantico oeste, e avanr;a-se a hip6tese que a colonizayao dos Ar;ores no meio do Atlantico por estes vermetfdeos fez-se por dispersao passiva a longa distancia por meio de "jangada" a partir de uma origem do Atlantico leste/Mediterra- neo. E seleccionado um lectotipo para a especie nominal Vermetus rugu- losus Monterosato, 1878 (Museo Civico di Zoologia, Rome). 0 nome co- mummente utilizado "Vermetus triqueter" e abandonado por ser uma soletrar;ao incorrecta subsequente de Vermetus triquetrus Bivona- Bernardi, 1832.
-
Upload
fieldmuseum -
Category
Documents
-
view
7 -
download
0
Transcript of Vermetid gastropods from Sao Miguel, Azores: Comparative anatomy, systematic position and...

Af;OREANA, SUPPLEMENT 1995: 173-192
VERMETID GASTROPODS FROM SAO MIGUEL, AZORES: COMPARATIVE ANATOMY, SYSTEMATIC POSITION AND BIOGEOGRAPHIC AFFILIATION
Rildiger Bieler
Center for Evolutionary and Environmental Biology, Field Museum of Natural History, Chicago, Illinois 60605, U.S.A.
ABSTRACT Two vermetid species inhabit the intertidill and upper subtidal rocky
shores of Sao Miguel, Azores. The larger-shelled species, identified as Vermetus cf. triquetrus Bivona-Bernardi, 1832, has highly variable shell characters, spanning previously assumed diagnostic features of several nominal genera. The smaller form is recognized as Thylaeodus cf. rugulosus Monterosato, 1878, a species previously deemed restricted to the Mediterranean. Females of both species brood their eggs in stalked capsules attached to the interior shell wall. After feeding on nurse eggs, juveniles are released at the crawling stage and attach to suitable substratum. Neither form is known to date from the western Atlantic, and colonization of the mid-Atlantic Azores by these vermetids is hypothesized as passive long-distance dispersal by "rafting" from an eastern Atlantic/Mediterranean source. A lectotype is selected for nominal species Vermetus rugulosus Monterosato, 1878 (Museo Civico di Zoologia, Rome). The commonly used name "Vermetus triqueter" is dismissed as an incorrect subsequent spelling of Vermetus triquetrus Bivona-Bernardi, 1832.
RESUMO Duas especies de vermetfdeos habitam o intertidal e subtidal superior
das costas rochosas de Sao Miguel, Ayores. A especie de concha maior, identificada como Vermetus cf. triquetrus Bivona-Bernardi, 1832 possui caracteres conquiliol6gicos altamente variaveis, abrangendo varios generos nominais caracterfsticas antes tidas como diagnosticantes. A forma menor e reconhecida como Thylaeodus cf. rugulosus Monterosato, 1878, especie previamente tida como restricta ao Mediterraneo. As femeas de ambas as especies incubam os ovos em capsulas pedunculadas fixas a parede interior da concha. Depois de se alimentarem de ovos de criayao, os juvenis sao largados na fase rastejante e fixam-se a um substrato adequado. Ate a data nenhuma das formas e conhecida do Atlantico oeste, e avanr;a-se a hip6tese que a colonizayao dos Ar;ores no meio do Atlantico por estes vermetfdeos fez-se por dispersao passiva a longa distancia por meio de "jangada" a partir de uma origem do Atlantico leste/Mediterraneo. E seleccionado um lectotipo para a especie nominal Vermetus rugulosus Monterosato, 1878 (Museo Civico di Zoologia, Rome). 0 nome comummente utilizado "Vermetus triqueter" e abandonado por ser uma soletrar;ao incorrecta subsequente de Vermetus triquetrus BivonaBernardi, 1832.

174 A<;OREANA SUPPLEMENT 1995: 173-192
INTRODUCTION
Vermetidae (Caenogastropoda: Vermetoidea incertae sedis) is a gastropod family whose members are highly modified snails, adapted to a sessile life mode. Their irregularly coiled and frequently corroded shells, which in the postlarval stage are firmly cemented to rock or other hard substrate, resemble those of serpulid polychaete worm tubes. The animals collect food from the surrounding seawater by ciliary action or by producing particletrapping mucus strands which are then ingested (e.g., Boettger, 1930). Generic or specific identification from empty shell tube fragments (which is the usual condition of historic vermetid collection material) is often impossible. Although it contains common members of many temperate to tropical rocky shore communities, the group has remained poorly studied. The high variability of shell features defeats attempts to apply "standard" gastropod shell characters for descriptive and identification purposes. This, and the difficulty in collecting (preferably by hammer and chisel; often from the under-sides of rocks and boulders in the surf zone and in deeper water) have excluded them from many studies of molluscan communities.
Field work for the present study concentrated on the intertidal and upper subtidal (SCUBA depth) rocky shore of Sao Miguel, mostly in the vicinity of Vila Franca do Campo and on Ilh6u de Vila Franca, a small
island formed by a drowned volcanic crater (B. Morton, 1990).
Although now known to be common, the vermetids of Sao Miguel are relatively small and inconspicuous, and were not mentioned in the more general descriptions of local intertidal habitats and ecology (Arruda & Gordo·, 1984; B. Morton, 1990; Hawkins et al., 1990).
Several morphotypes ultimately determined to represent two vermetid species were found in the study area. Comparative sampling of the rocky shore at the far north-eastern end of Sao Miguel, near Mosteiros, resulted in the collection of the same two species. The larger-shelled one is the form previously referred in the Azorean literature to Vermetus t riq u e te r (V. cf. t riq ue t rus in this work). The smaller one is a new record for the Azores, and here is called Thylaeodus cf. rugulosus Monterosato, 1878. Morphological and anatomical characters of the two species, as well as aspects of their reproductive biology will be compared.
The known range of reproductive strategies in the family Vermetidae includes brooding of very small clutches resulting in crawl-away larvae as well as brooding of egg masses releasing numerous planktotrophic larvae that facilitate wide geographic dispersal (Hadfield et al .• 1972). The isolated position of the mid-Atlantic Azores poses interesting questions of the geographic origin of its fauna and flora (see, e.g., Gofas, 1990; Prud'homme van Reine,

BIELER: VERMETIDAE OF THE AZORES 175
1988). For the Vermetidae, the question is here whether the local species can be recognized as part of the eastern Atlantic/Mediterranean fauna, whether they are part of the western Atlantic vermetid fauna (currently under rev1s1on by the present author). or whether they represent endemic forms of the Azores.
MATERIALS AND METHODS
During two weeks in July and August of 1991, living specimens were collected on Sao Miguel by wading, snorkling and SCUBA diving. The snails were removed from overturned rocks and boulders in 0.1-10 m depth of water, using a knife blade or hammer and chisel. Some specimens were collected with shell hash in 20 m depth, employing a suction device attached to a SCUBA air tank. Animals were transferred to the workshop laboratory in Vila Franca do Campo on Sao Miguel, where they were measured, observed and sketched under a dissecting microscope and prepared for further study. Measurements were taken with calipers or by using calibrated eyepieces on the dissecting microscope. Photographs were taken with standard 35 mm equipment and macrolenses. For selected listings of material examined, see individual species descriptions, below.
Animals were anestethized by placing menthol crystals on the water surface of finger bowls housing the specimens. Soft-bodies and brooded egg masses were removed
from the shell after cracking with a small vice. Soft-parts were measured in the "relaxed" state and their gross anatomy studied using a dissecting microscope. For histological studies, specimens were relaxed in MgCl2 solution isotonic with seawater and chemically fixed in either 5% buffered formalin or Bouin's solution. On return to the home laboratory at FMNH, specimens were embedded in paraplast, sectioned at 7-8 µ m and stained with Alcian B 1 ue/Periodic-Acid-Schiff (PAS) and/or Hematoxylin/Eosin. Radular ribbons and jaws were extracted by dissolving the surrounding tissue in 10% KOH solution at room temperature for 24 hours, with subsequent washing in distilled water. Air-dried shells, radulae, jaws, and opercula were coated with gold, and observed and photographed using an AMRA Y 1810 scanning electron microscope at Field Museum. Nomenclature for radular characters follows the outline given by Bandel (1984: Figs. 1-3).
Preserved Azorean specimens were compared with freshly collected western Atlantic vermetids at the Smithsonian Marine Station at Link Port, Ft. Pierce, Florida, U.S.A., in March of 1992. For comparison with Mediterranean material, specimens in the Monterosato Collection, Museo Civico di Zoologia, Rome, Italy, were studied in August of 1992. Previously reported Azorean vermetid specimens (Dautzenberg, 1889; 'Hirondelle' cruise of 1887; Mu see Oceanographique de Monaco) were

176 A<;OREANA SUPPLEMENT 1995: 173-192
examined on loan. Abbreviations used in text ('*'
denotes institutions where voucher material was deposited):
*FMNH Field Museun of Natural History, Chicago, Illinois, U.S.A.
*HBOM Harbor Branch Oceanographic Museum, Ft. Pierce, Florida, U.S.A.
*MCZ Museum of Comparative Zoology, Harvard University, Cam-bridge, Massachusetts, U.S.A.
MC7R. Museo Civico. di Zoologia, Rome, Italy.
*MNHNP Museum National d'Histoire Naturelle, Paris, France.
Additional voucher material will be deposited with the Museu Carlos Machado, Ponta Delgada, Sao Miguel, Azores (awaiting upgrade of museum facilities; Frias Martins, pers. comm.).
DESCRIPTIONS
Ve rm e tu s cf. tr i q u et r us Bivona-Bernardi, 1832
Figs. 1-14
?Vermetus triquetrus Bivona-Bernardi, 1832: 6 (Palermo, Italy; Recent)
Material studied (selected listing; all Sao Miguel, Azores; collected during 2nd International Workshop, 21 VII - 3 VIII 1991): Ponta da Piramide (37°42.?'N, 25°28.0'W), 0.5-1 m, in strong currents on sides and under-surfaces of rocks and boulders (FMNH 224921-23, 224927, 224933-37. 224939-41, 224944, 224946 [incl. sectioned specimen], 224925-26, 224930-31, 224945; MCZ 302758; MNHNP unnumbered); off Ponta da Piramide (37°42'N, 25°28'W), 5-13 m (SCUBA), on rocks (FMNH 224924, 224943, 224947); 300 m off Ponta da Piramide, 20 m (SCUBA suction device), in shell hash with red algae Antithamnion cruciatum (C. Agardh) Nligeli (FMNH 224928; HBOM 065:03700); W of Rosto do Cao, Sao Roque, E of Ponta Delgada (FMNH 224932); Ilheu de Vila Franca (37°42.3'N, 25°26.6'W), eastern part of inside lagoon, intertidal, among coralline algal turf (FMNH 224929, 224942); Mosteiros (37°54.0'N, 25°49.1 'W), intertidal, on sides and under-surface of rocks and boulders (FMNH 224938).
Teleoconch (descriptions based on live-taken material; for measurements, n=30): Coiling pattern, shell thickness, sculpture and coloration very variable. Maximum dimensions of individual adult shell mass ranging from 10 x 8 mm to 19 x 13 mm; maximum outside whorl diameter
FIGS. 1-7. Vermetus cf. triquetrus (Bivona-Bernardi). Sao Miguel, Azores. l, living animal, with final part of shell removed to expose head-foot area (note long pedal tentacles), bar=2 mm, FMNH 224930. 2, strongly-keeled morph, live-collected, greatest dimension 11.4 mm, FMNH 224930. 3, multi-ribbed morph, live-collected, greatest dimension 13.6 mm, FMNH 224932. 4, smooth (corroded) morph, live-collected, early whorls opened to expose transverse septa; greatest dimension 16 mm, FMNH 224935. 5, intertwined cluster, greatest dimension 45 mm, FMNH 224944. 6-7, surface shell sculpture (SEM): 6, early teleoconch whorl (aperture facing toward right), bar=l mm, FMNH 224928; 7, first teleoconch whorl, bar=lOO µm, FMNH 224929.
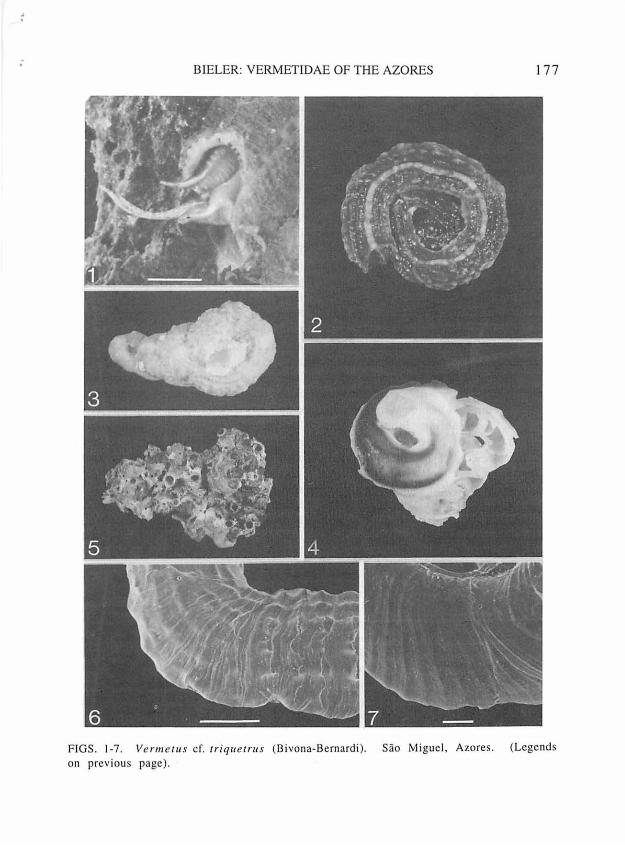
; BIELER: VERMETIDAE OF THE AZORES 177
F IGS. 1-7. Ve n11e t11 s cf. 1riq11etrus (Bivona-B ernardi ). Sao Miguel, Azores. (Legends on previous page).

178 Ac;OREANA SUPPLEMENT 1995: 173-192
from 3.8 to 5.1 mm (x= 4.7). Coiling rarely deviates from counter-clockwise direction, but ranges from a regular, tight coil (Fig. 2), or a "Turritella-squashed-sideways" growth form (Fig. 3), to very loosely coiled, irregular morphs, the latter especially when gregarious (Fig. 5). Shape of cross sections through whorls variable, ranging from circular to depressed-oval or almost triangular, influenced by substrate and shell sculpture. Usually with short erect feeding tube (raising aperture 4-10 mm above surface), narrower than those whorls appressed to the substrate, with inner apertural diameter ranging 2.6 to 3.6 mm (x= 3.2). Some specimens with remnants of earlier feeding tubes. Without longitudinal columellar lamellae, but occasionally with one or several internal septa (Fig. 4). Early postlarval sculpture of axial growth lines (Fig. 7), subsequent sculpture of 8-10 nodose longitudinal (spiral) ribs of varying thickness (Fig. 6), the outermost ones drawn out into flanges appressed to substrate. One rib
usually considerably stronger, forming an often prominent shell keel on inner "dorsum" of shell. Rib tubercles often scaly, coordinated into irregular transverse (axial) bulges. In weakly sculptured or somewhat eroded shells, these bulges form the only discernible sculpture. Shell wall thickest at dorsal keel and lateral flanges; attached ("bottom") part of tube very thin-walled, especially on smooth surfaces. Inside of tube lined with smooth, glossy layer; brown in the adult. Juvenile shells translucent, off-white, tan or dark brown. Adult coloration also variable, dark brown with dorsal keel lighter, or mottled light-tan/brown with tubercles lighter; occasionally only dorsal keel with discernible color pattern, or all ribs with alternating brown/off-white pattern (Fig. 1). Smooth shells often appearing gray, with inner brown shell layer shining through outer whitish layer. Shells of living snails can be strongly eroded, smooth and partly trans-1 ucen t, resulting in a purplishbrown (inner layer) and white (cor-
FIGS. 8-14. Vermetus cf. triquetrus (Bivona-Bernardi). Sao Miguel, Azores; SEM. 8, protoconch (725 µm in length) and early teleoconch whorls (normally attached to rock surface), with first teleoconch whorl partially opened to illustrate tight wrapping of whorls, FMNH 224941. 9, operculum, external side, 870 µm maximum diameter, FMNH 224946. 10, shell of intracapsular juvenile, after metamorphosis; apical aspect (note wavy growth marks on caplike embryonic shell), greatest diameter 640 µm, FMNH 224946. 11, shell of freshly hatched juvenile (note beginning teleoconch growth), maximum dimension 760 µm, FMNH 224946. 12, shell of settling juvenile (note protoconch umbilicus, fine spiral striation, and part of first teleoconch whorl); protoconch dimensions 815 x 680 µm, FMNH 224946. 13, radula, approximately mid-ribbon; arrow pointing to single inner flanking cusp of outer marginal tooth; bar=lOO µm, FMNH 224938. 14, fine detail of jaw surface; bar=lO µm, FMNH 224943.
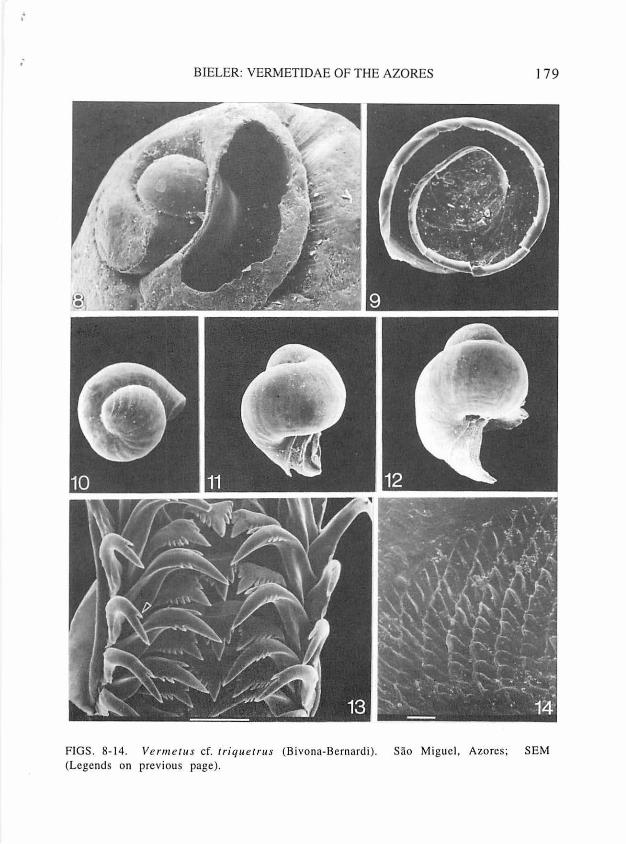
; BIELER: VERMETIDAE OF THE AZORES 179
FIGS. 8-1 4. V e n11e111s cf. rr iqu errus (B ivona-Bernardi). Sao Miguel, Azores; SEM (Legends on previous page).

180 A<;OREANA SUPPLEMENT 1995: 173-192
roded outer layer) shell (Fig. 4). Shells often fouled by numerous organisms, especially coralline algae, sponges and barnacles.
Protoconch: Globular shell, with a little over 1 2/3 whorls; maximum shell length 720-815 µ m, diameter 600-680 µ m; embryonic part capshaped, with wavy growth lines (see Fig. 10); larval shell (comprising approximately final full whorl) inflated, with faint spiral riblets (especially · n~ar protoconch umbilicus). No noticeable growth between hatching of crawl-away larva (see below) and settlement (compare Figs. 8, 10-12). Translucent off-white to light tan, with suture darker.
Operculum: Round, slightly concave, 700-870 µ m in adult specimens, about 1/5 to 1/3 of relaxed pedal disk diameter; exposed side with edge of earlier whorls rising as free-standing lamella along suture of final 1.5 whorls (edge rolling inward in dry specimens; see Fig. 9); horny, transparent.
Radula (Fig. 13): Regularly taenioglossate; transparent, largest specimens with some horn-colored staining; consisting of 25-30 rows of teeth (for counts and dimensions: n= 15, from specimens with maximum shell mass dimensions of 12-19 mm; no difference noted between sexes or between specimens from different localities on Sao Miguel).
Total length of (artificially straightened) radular ribbon 1.8-2.4 mm, width (mid-ribbon, marginal teeth not folded outward) 340-440 µ m. Radular configuration agreeing
with general description of vermetid characters given by Bandel (1984: 44-46) and the sketch for "Bivonia triquetra" (see Bandel, 1984: fig. 51 ). Trapezoidal rachidian tooth with strong main cusp and 4-5 flanking cusps on either side (reducing in size outwardly); basal denticles strongly developed. Inner margin of lateral tooth fitting into deep marginal depressions of rachidian tooth; cusp arrangement on triangular cutting edge as in central tooth. Marginal teeth long, slender, with long and strong main cusp. Inner marginal with 1 flanking cusp on inside, 2-3 on outside. Outer marginal with single inner flanking cusp, smooth on outside. Indicated variation in numbers of flanking cusps may occur between left and right sides of same radula.
Jaws: Elongate bean-shaped; attached to transparent cuticula; lightly tan-colored cutting area consisting of small (5 x 10 µm) obliquely quadrangular subunits (Fig. 14); maximum jaw dimensions 800x300 µm
Anatomy: Animal (removed from shell) serpentine, large specimens to 30 mm in (relaxed) length, 3.5 mm in width; long posterior lobe of digestive gland and gonad occupying at least half its length. Columellar muscle a long ·narrow strip with a deep insertion in the shell _(animal capable of retreating deeply into shell). Pedal tentacles long, extremely slender and tapered (Fig. 1 ), in fully extended position with right tentacle longer than left (e.g., 5.0 vs.
-'

...
BIELER: VERMETIDAE OF THE AZORES 181
4.3 mm); ciliated groove running along mesial side as far as tip, diverging from the median opening of very large pedal gland. Sole of foot small and ovoid, between and anterior to pedal tentacles. Large and bulbous head with short tentacles (ca. 0.80 mm in living adults). Head with no trace of deepened or flattened food tract. Pallial skirt of brooding females deeply incised in the dorsal midline. Gill extending full length of mantle cavity, approximately 14 fifaments per mm in large specimens; anteriormost filaments very short, quickly increasing to average adult length of 800 µ m ; filaments forming small triangles, somewhat taller than wide. Extensive area of hypobranchial gland left entirely uncovered by small gill. Osphradium vermiform (about 2.4x0.24 mm in large specimens), beginning somewhat anterior to gill filaments. Nervous and alimentary systems not studied in detail; fecal pellets ovoid, about 900x320 µ m . Female capsule gland not running the length of mantle cavity, but forming ovoid, ventrally slit pouch (as described by J.E. Morton, 1965: 609-611).
Soft-body coloration: Adult coloration: head-foot including pedal disk usually appearing brown-black; darker-shelled animals also with darker body pigmentation; pedal disk yellowish, mottled orange, black and white, with some orange granules more deeply embedded in tissue; upper rim (toward snout) of pedal disk with four evenly spaced
black dots; forehead with fine black and orange coloration, appearing brown; forehead and lightly-orange lateral concave regions of head with white "tiger-stripe" pattern; snout with orange, black and white pigment; cephalic tentacles whitish, with some fine black surface pigment and orange granules embedded within; rims of cephalic tentacles and sole of foot white; pedal tentacles translucent, with embedded yellowwhite and/or orange bodies and with yellow tips, in some specimens also with sparse black pigment; lower part of foot orange-yellow to bright orange with greenish iridescence, and some black pigment; outside of mantle appearing brightly orange, pigments consisting of yellow, granular orange, and white; mantle edge with alternating pattern of black and whitish-orange; inside of mantle yellow and orange. Young post-larva without dark body pigment, only some white at mantle margin and on anterior foot; after reaching about 1/2 teleoconch "whorl," with fine black pigment on head-foot and sparse orange spots on mantle.
Eggs and development: 5-9 egg capsules brooded in mantle cavity, arranged in single, occasionally double, row (second row started after completion of first); capsules attached by stalks passing through mantle slit to interior lining of shell; terminal capsule stalk 400-500 µ m in length, attaching at one smaller end of oval egg capsule; capsule dimension approximately 11OOx1600 µ m to 1500x1600 µm; in early develop-

182 A<;OREANA SUPPLEMENT 1995: 173-192
mental stages often more rounded. Egg capsules apparently deposited over a period of time, oldest (most developed) closest to shell aperture [a single clutch contained· 5 capsules with early cleavage stages, 2 with developing embryos, and with veligers and crawling stages]. Yolk yellowish-white; developing embryos/larvae feeding on nurse eggs, with 5-14 developing larvae per capsule; velar lobes resorbed during intracapsular metamorphosis to crawling stage; crawling stage with 2 black eye spots, long cephalic tentacles, agile foot (anterior foot and cephalic tentacles · heavily . ciliated); anterior part and sole of foot of prehatching animal with fine black pigment; shell transparent, light-tan, with suture and columella somewhat darker brown.
Habits/habitat: Attaching to more-or-less smooth patches of volcanic rock; usually singly or in pairs, occasionally in clusters attaching to each other. Juveniles often settling on mother shell. Not actively corroding into substratum. Feeding by mucus-net trapping.
Specific identity: The name "Vermetus triqueter" has been used for Azorean records (Dautzenberg, 1889; .Drouet, 1858: 26, who erroneously credited it to Linne). Bivona-Bernardi (1832: , 6) introduced the name as Vermetus triquetrus, using the correctly formed Latin suffix -q u et r us meaning angled, cornered, or sided. Philippi ( 1836: 170) changed this name to Vermetus triq ue te r, an incorrect subsequent
spelling without nomenclatural status (ICZN Art. 33[c]) in use ever since.
Vermetus triquetrus Bivona-Bernardi, 1832, was originally described from Palermo in the Mediterranean, and has subsequently been cited for the Atlantic and Mediterranean coasts of the Portuguese mainland (e.g., Nobre, 1936; Saldanha, 1979). The taxonomy of Mediterranean Vermetidae is still in a state of confusion, and the systematics of the Mediterranean members of the triquetrus-complex poses questions beyond the scope of the present study. The last monograph on the subject was published by Monterosato ( 1892) who expanded on the earlier works of Bivona-Bernardi ( 1832) and Morch (papers spanning 1859-1871), but like most of his predecessors, Monterosato concentrated on naming numerous nominal species and "varieties" based on teleoconch characters alone. Some authors (e.g., Morch, 1871) suggested conspecificity of the nominal species Vermetus triquetrus and V. semisurrectus Bivona-Bernardi, 1832, while Keen (1961) placed them in separate subgenera of Vermetus. The best work available on members of the triquetrus-complex is that by Lacaze-Duthiers ( 1860), who documented the anatomy and embryology of specimens identified as Vermetus .triquete r and V. semisurrectus "Philippi" (err. pro Bivona-Bernardi, 1832). The very detailed, study has an unfortunate flaw. Vermetus triquetrus was said (1860: 210) to have

; BIELER: VERMETIDAE OF THE AZORES 183
been used mainly for dissections and observations, but shells and anatomical material had become separated during the course of that study. Lacaze-Outhier, who studied these nominal species first on Corsica and then continued his work on the Spanish island of Menorca, was uncertain about the species identity for some of his data. In addition to striking similarities of the anatomical descriptions and illustrations, the developmental data for the Mediterranean forms are close to those reported here for the Azorean "cf. triquetrus." Mediterranean Vermetus triquetrus was described as having a series of 4-6 (sometimes more) egg capsules, containing 10-30 eggs each (Lacaze-Duthiers, 1860: 266; see also partial anatomical redescription by J. E. Morton [1965: 608-611] based on material from Monaco, and Bandel's [1984: fig. 51] sketch of radula). The Azorean species has 5-9 capsules containing 5-14 eggs each.
Previous Azorean records: Several previously published records mention the occurrence of vermetids in the Azores. Martens in Simroth (1888: 214) reported a "Vermetus sp." on the shell of a Patella species in the current study area ("Villafranca"). In his contribution on the molluscan fauna of the Azores, Dautzenberg (1889: 43) listed two species. One, cited as Ver met us triqueter Bivona- Bernardi, 1832, is based on Drouet's ( 1858) record from Sao Miguel. Drouet (1858: 26-27) described two forms, "1 'une d'un
jaune rose, tres c6telee; 1 'autre blanchatre, moins rugueuse (peutetre une autre espece?)," but it remains unclear whether he referred to two of many morphs of cf. triquetrus, or to the two species recognized in the present study.
The second Azorean vermetid species mentioned by Dautzenberg (1889: 43; from Sao Miguel and from shallow water off the island of Pico ("Lagens de Pico" [Lages do Pico], 'Hirondelle' station 108) is given as Vermetus cristatus Biondi, 1859. This nominal species was also originally described from Lhe Mediterranean. It was subsequently (e.g., Monterosato, 1892) interpreted as a species of "Spiroglyphus" [ Dendropoma Morch, 1861], a group not encountered during the present study. The original material on which Dautzenberg's (1889) "cristatus" record was based (Collections du Musee Oceanographique de Monaco, unnumbered; vidi) consist of somewhat corroded shells that are here identified as V. cf. triquetrus ('Hirondelle' material: 16.9 x 7.9 mm and 15.5 x }.5 mm greatest shell dimensions).
Thy la e o du s cf. rug u lo s u s (Monterosato, 1878)
Figs. 15-20
?Vermetus rugulosus Monterosato, 1878: 29 (Palermo, Italy).
? Vermetus ( Petaloconchus?) rugulosus, Monterosato, 1892: 2, pl. I, fig. 8.
Material studied (selected listing; all Sao Miguel, Azores; collected during 2nd

184 A<;OREANA SUPPLEMENT 1995: 173-192
International Workshop. 21 VII - 3 VIII 1991): Ponta da Pirimide (37°42.7'N. 25°28.0'W). 0.5-1 m. in strong currents on sides and under-surface of rocks and boulders (FMNH 224951-53); . 300 m off Ponta da Piramide (37°42'N. 25°28'W). 20 m (SCUBA suction device). in shell hash with red algae Antithamnion cruciatum (C. Agardh) Niigeli (FMNH 224949); Ilheu de Vila Franca (37°42.3'N. 25°26.6'W). eastern part of inside lagoon. intertidal. among coralline algal turf (FMNH 224950); Ilbeu de Vila Franca, inside lagoon. 1.5 m, under-surfaces of rocks (FMNH 224954-57; HBOM 065: 03701; MCZ 302759; MNHNP unnumbered); Mosteiros (37°54.0'N. 25°49. l 'W). intertidal, on sides and under-surface of rocks and boulders (FMNH 224948).
Teleoconch (descriptions based on live-taken material; for measurements. n=15): Maximum dimensions of individual adult shell masses ranging from 4.0 x 2.5 mm to 7 .8 x 5.3 mm; maximum outside whorl diameter ranging from 1.2 to 1.9 mm (x= 1.6). Coiling counter-clockwise, usually very irregular, but rarely forming open loops; spreading sideways (Fig. 15). Shape of cross sections through whorls round to oval, influenced ' by subStrate and development of shell flanges. Usual-
ly with short erect feeding tube (raising aperture 1.5-2.2, rarely to 5 mm above surface), narrower than whorls appressed to the substrate, with adult inner apertural diameter normally ranging 0. 74-1.0 mm (x=0.81 ), sometimes narrowed to as small as 0.40 mm. Remnants of earlier feeding tubes usually present (Fig. 17). Without longitudinal columellar lamellae, and without longitudinal (spiral) sculpture. With distinctive, Caecum-like, transverse (axial) sculpture, consisting of weak axial growth lines at first, later forming smooth, more-orless regular, closely spaced, somewhat wavy axial ribs; the latter strongest in appressed drawn-out lateral shell flanges (Figs. 15, 17). Outer shell layer translucent white, inner layer in most specimens brown (in late whorls especially dark in the unattached, "roof"-forming part of the shell tube); outer layer thin in juveniles, shell thus appearing tan to dark brown, later especially ribs and flanges whitish; apertural rim often darker (showing inner layer). Inner layer in about 40% of the animals not. brown, shells thus overall translucent milk-white; early whorls of these morphs
FIGS. 15-20. Thylaeodus cf. rugulosus (Monterosato), Sao Miguel, Azores; SEM. 15, entire shell. taken from living animal (Q ), bar=l mm, FMNH 224956. 16, operculum. external side. diameter 800 µm. FMNH 224953. 17, early teleoconch whorls, with part of whorl, removed (dashed line) to expose protoconch; arrow pointing to remnant of early feeding tube; same specimen. as in Fig. 15. 18, protoconch (730x390 µm), same
specimen as in Fig. 15. 19, radula (Q ). approximately mid-ribbon; ribbon-width 150 µm, FMNH 224953. 20. as in previous figure.· with outer marginal tooth folded outward to expose inner flanking cusps (arrow).
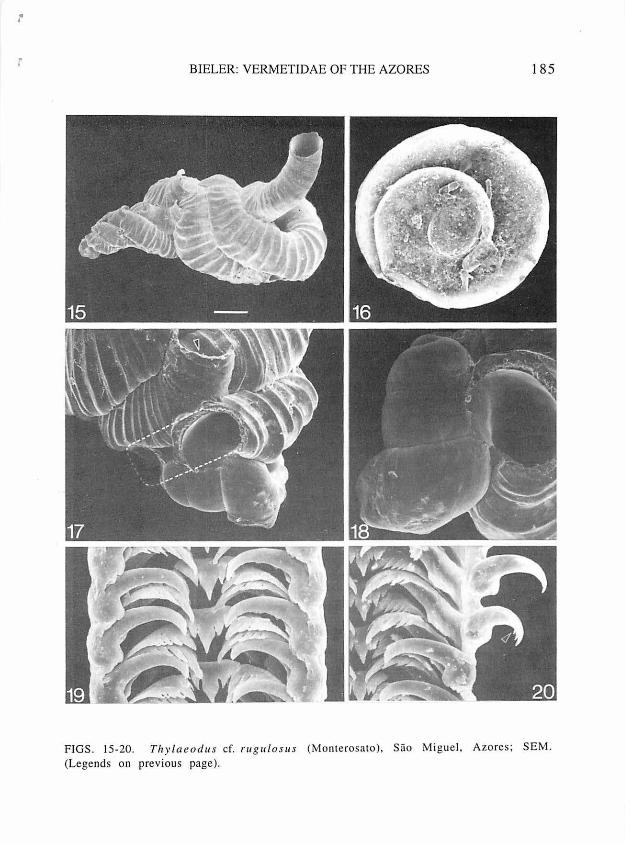
l
~ I
BIELER: YERMETIDAE OF THE AZORES 185
FIGS. 15-20. Thylaeod 11 s cf. ru gulorns (Monterosato), Sao Miguel, Azores; SEM. (Legends on previous page).

186
appearing tan with darker digestive gland shining through. Brooding females : appearing greenish with coloration of eggs visible through thin shell. Shells often fouled by several different organisms, especially foraminiferans and serpulid worm tubes.
Protoconch (Fig. 18): Relatively slender, pupa-shaped, with sinuous outer lip; with 2 1/2 whorls; maximum shell length 700-730 µm, diameter 390-420 µ m. Without noticeable growth between hatching of crawl-away larva (see below) and settlement. Suture deeply impressed around embryonic apex. Overall tan with lighter central part of body whorl, suture brown.
0 p e r cu l um : As large as pedal disk (in living animal}, 0.74-1.0 mm, filling aperture; exposed side with edge of earlier whorl rising as freestanding lamella along suture of final 1.5 to 2 whorls (see Fig. 16); transparent.
Radula (Figs. 19-20): Regularly taenioglossate; transparent; consisting of 28-32 rows of teeth (for counts and dimensions: n=7, from specimens with shell mass dimensions of over 5 mm; no difference noted between sexes or between specimens from different localities on Sao Miguel). Total length of (artificially straightened) radular ribbon 660-700 µ m, width (mid-ribbon, marginal teeth not folded outward) 140-150 µm. Radular configuration agreeing with general description of vermetid characters given by Bandel ( 1984: 44-46). Trapezoidal rachidian tooth with strong main cusp and
SUPPLEMENT 1995: 173-192
4-5 flanking cusps on either side (decreasing in size ot~!wardly); marginal ridges strongly developed. Inner margin of lateral tooth fitting into deep marginal depressions of rachidian tooth: cusp arrangement on triangular cutting edge as in central tooth. Marginal teeth long, slender, with long and strong main cusp. Inner marginal with 4-5 flanking cusps on either side. Outer marginal (about 64 µ m in length) with 5-6 flanking cusps on inside (largest toward main cusp}, smooth on outside (Fig. 20).
Jaws: Crescent-shaped opaque cutting area; maximum length 280 µ m, attached to transparent cuticle.
Anatomy: Animal (removed from shell) long and very narrow, coiled into several loops; large specimens to 14 mm in (relaxed) length, 0.8 mm in width; long posterior lobe of digestive gland and gonad occupying about 2/3 of its length. Columellar muscle a very long narrow strip with deep insertion in shell (animal capable of retreating very deeply into shell). Pedal tentacles short, even when in extended position rarely exceeding 400 µm. Sole of foot a small and bulging protrusion between and anterior to pedal tentacles. Pallial skirt of brooding females with deep incision, running obliquely over dorsum to back of mantle cavity. Gill extending full length of mantle cavity, approximately 20 filaments per mm in large specimens; anteriormost filaments very short, quickly increasing to average adult length of 260 µm; filaments
II 'c

BIELER: VERMETIDAE OF THE AZORES 187
forming small triangles, taller than wide. Osphradium vermiform, arched with back to gill (about 800 x 160 µm in large specimens), beginning at level of anteriormost gill filaments.
Soft-body coloration: Overall whitish or yellowish; sparse pink granules embedded in outer mantle area, in the 4 tentacles and around snout; fine white granules in headfoot, coarser in outer mantle; remainder of body translucent with more deeply embedded white pigment in tissue of foot and head; coloration of the dark-shelled morph additionally with purplish-pink granules embedded in head-foot, tentacles and mantle.
Egg and larval development: 4-8 egg capsules brooded in mantle cavity, arranged in single row); capsules attached by stalks passing through deep mantle slit to interior lining of shell; capsule stalk 200 µ m in length, subterminal, never centered at pole; egg capsule dimensions approximately 750 x 950 µm to 800 x 1200 µm. Capsules apparently deposited over a period of time, oldest (most developed) closest to shell aperture. Egg diameter 100 µm, yolk light-green [green color lost upon preservation in alcohol]; capsules filled with dozens of light-green eggs, only 3-5 of which developing into embryos, the rest being consumed as nurse yolk; larval shell pupa-shaped, transparent, with brown suture; larval digestive gland filled with light green yolk; no velar lobes observed; crawl-away larvae
(approximately 700 x 400 µm shell size) developing in capsule, with tentacles and black eyes; at hatching, shell with columella (shining through), suture, and lower outer shell rib browm.
Habits/habitat: Found attached to the under-surfaces of loose lava rocks and concrete blocks in shallow water, together with coralline algae, sponge, barnacles and tube-building polychaetes; singly or in small clusters of 2-5 per rock. Small mucous feeding nets observed. Animals not actively corroding into substratum.
Specific identity: Monterosato did not illustrate his new species "Vermetus rugulosus" in the original description (1878: 29); the diagnosis only read "Specie minuta, spirale, con le rugositA regolari simile a quelle del Caecum trachea." A subsequent, more detailed description accompanied by a figure (Monterosato, 1892: 22, pl. I, fig. 8) suggests identity with this Azorean form. The Monterosato collection in Rome contains a specimen lot of 11 syntypes (MCZR no. 21280, vidi); the largest and best-preserved . of which is the shell subsequently illustrated by Monterosato as "Forma, typica" (1892: fig. 8). This shell is here selected as lectotype of nominal species Vermetus rugulosus (greatest overall length 4.5, apertural height above attachment plane 3.5, apertural diameter 0.7 mm). No anatomical or biological data from Mediterranean specimens were available for comparison, hence the tentativeness of the assignment to this nominal

188 A~OREANA SUPPLEMENT 1995: 173-192
species. DISCUSSION
Generic placement The present study confirms the
presence of enormous morphological variability within a single vermetid species.
Azorean Vermetus cf. triquetrus was found to have smooth, multiribbed or sharply keeled shells, and growth forms ranging from tight and regular coils to elo~gated and irregularly· twisted individual shell masses. Shell , coloration of this species is similarly variable, and animals occur singly; in small aggregations, or in intertwined clusters. Figures 1-5 illustrate the range of encountered morphs, compnsmg coiling patterns·· .previously regarded as · diagriost:ic of several nominal genus-group taxa ·(e.g., Cladopa, Petalocorichus, Novastoq,). Taxonomic decisions based on such teleoconch characters· alone obviously lead to a.rtificial groupings. · · ' ,J ,
As is· the case with species ' defi:.. nitions,' ~our understanding : of monophyletic' groups within the Vermetidae is·: still . p:oor. The . excellent monographs by Keen ( 1961) and J.E. Morton (1965) cleared up much of the historic confusion. However, most of the genus-level assignments remain based on features of relative shell coiling patterns, relative size of operculum, and presence or absence of internal shell lamellae, and not on well-defined suites of synapomorphies.
The two shallow-water Azorean
species ar/ here placed tentatively in the nominal genera Ver'metus Daudin, 1800, and Thylaeodus Morch, 1860, respectively. The placement of tr i q u et r us in Ve rm e tu s agrees with Keen's (1961: 193) classifica.; tion, although that . author considered the general absence of feeding-tube scars a generic character (present in some Azorean specimens). The name Bivonia Gray, 1847, used by some authors for this nominal species (e.g., Bandel, 1984), is not available because preoccupied by Bivonia Cocco, 1832 (Crustacea). Thylaeodus is here employed at the generic level; it was treated as a subgenus of Vermetus by Keen (1961), based on the shared absence of internal lamellae~ and as a subgenus of.. Pe talooon ch us, by Morton (1965: 600), based on overall anatomical similarity. ·. ]ts relationships> to (or possi-: hie synonymy with). nominal .genera Vermetus or Petaloconchits need further study ..
Origin of 'the Azorean vermetid fauna Despite the ·eastward .direction of
the North Atlantic Drift, the Azorean molluscan fauna is known to have close· affinities to those of the European/North African mainland and the other Macaronesian islands, especially the Canaries and Madeira (for overview of surface current patterns around the Azores and some theories on colonization pathways-,. see Gofas, 1990). As was proposed for other marine organisms such as seaweeds (Prud'homme van Reine, 1988), the gastropod fauna of the

BIELER: VERMETIDAE OF THE AZORES 189
Azores seems to consist of chance survivors of chance invasions. "Island hopping" by forms with lecithotrophic development, either via passive larval transport (e.g., Leal & Bouchet, 1991 ), or drifting of postlarval stages (Martel & Chia, 1991 ), appears possible within the archipelago. However, for the long-distance dispersal over several thousand kilometers to the mid-Atlantic Azores, species with planktotrophic larvae and those adapted to "rafting" are obviously advantaged.
Azorean species with intracapsular development leading to "crawlaway larvae" would have to be descendants of such planktotrophic forms, or have arrived by "rafting" or similar means of transport. As obligatorily sessile and (in the broadest sense) filter-feeding organisms, vermetids are especially well-equipped for rafting. Although members of this family do · not normally settle on wood, they attach to organisms that do (such as barnacles and oysters), and have been found as members of the fouling community on ship hulls (pers. observ.). Passive long-distance dispersal, and thus specific identity with vermetids previously described from elsewhere in the Atlantic, is thus likely. Neither of the two Azorean species is known to date from the western Atlantic vermetid fauna, currently under review by the present author. Two species originally described from the Mediterranean appear to be very similar, and may prove identical after additional and
much needed study of that fauna.
ACKNOWLEDGMENTS
I thank Ant6nio M. de Frias Martins (Universidade dos A~ores)
for the invitation to participate in the 1991 Workshop, and the logistic support provided by him and the other staff members and research students of the Departamento de Biologia. Special thanks are due to Robert C. Bullock, who shared personal laboratory equipment and his knowledge of the local collecting sites with me. Joao Manuel Brum, Robert C. Bullock, Paula M. Mikkelsen and Brian Morton helped with the field collecting during the workshop in the Azores. Italo Nofroni kindly assisted with access to the Monterosato Collection at the Museo Civico di Zoo logia in Rome,, Italy. The staff of the Musee Oceanographique de Monaco provided a loan of Dautzenberg's ( 1889) vermetid material from the Azores. I thank FMNH associate Katherine Wiegmann for producing some of the histological sections, Alan Kabat (USNM) for providing photocopies of publications otherwise inaccessible, and Don Cameron (University of Michigan) for answering my etymological questions.
Travel funding to the Portuguese Azores was provided in part by the Foster Fund of the Department of Mollusks, MCZ, and the FMNH Marshall Field Fund. Work on western Atlantic Vermetidae was supported, in part, by Smithsonian Marine Sta-

190
tion and NATO postdoctoral research fellowships.
Additional comparative studies in Florida were made possible through the Visiting Scientist Program of the Smithsonian Marine Station at Link Port; the local support by Mary E. Rice and her staff is greatly appreciated. This is Smithsonian Marine Station Contribution No. 324.
LITERATURE CITED
ARRUDA, L.M. & L.S. GORDO, 1984. On the study of a sample of gastropods (Prosobranchia) from the imperial [sic, for "intertidal"] range of Azores. Arquipelago, 5: 25-32.
BANDEL, K., 1984. The radulae of Caribbean and other Mesogastropoda and Neogastropoda. Zoologischf! Verhandelingen, 214: 1-188, pis. 1-22.
BIONDI, S., 1859. Descrizione di alcune specie malacologiche nuove che vivono nel nostro littorale. Memoria 2a. Atti della Accademia Gioenia di Scienze Naturali di Catania, ser. 2, 14: 113-123, 1 pl.
BIVONA-BERNARDI, A., 1832. Continuazione dell'articolo sui vermeti, estratto dalle Collettanee di Storia naturale del barone Bivona. Effemeridi scientifiche e letterarie per la Sicilia, 2: 3-13, 1 pl.
BOETTGER, C.R., 1930. Studien zur Physiologie der Nahrungsaufnahme festgewachsener Schnecken. Die Ernahrung der Wurmschnecke Vermetus. Biologisches Zentralblatt, 50(10): 581-598.
SUPPLEMENT 1995: 173-192
DAUTZENBERG, P., 1889. Revision des mollusques marins des A~o
res. Contribution a la faune malacologique des Iles A~ores. Resultats des dragages effectues par le yacht 1 'Hirondelle pendant sa campaigne scientifique de 1887. Resultats des campagnes scientifiques, accomplies sur son yacht par Albert 1e r Prince Souverain de Monaco, 1: 1-120.
DROUET, H., 1858. Mollusques marins des Iles A~ores. Memoires de la Societe Academique de l 'Aube, 22: 1-53, pis. 1-2, Paris.
GOFAS, S., 1990. The littoral Rissoidae and Anabathridae of Sao Miguel, Azores. In: MARTINS, A.M.F. (Ed.), The Marine Fauna and Flora of the Azores (Proceedings of the First International Workshop of Malacology, Sao Miguel, 1988). Aforeana, Supplement 1990: 97-134.
HADFIELD, M.G., E.A. KAY, M.U. GILLETTE & M.C. LLOYD, 1972. The Vermetidae (Mollusca: Gastropoda) of the Hawaiian Islands. Marine Biology, 12(1): 81-98, 20 figs.
HAWKINS, S.J., L.P. BURNAY, A.I. NETO, R. TRIST AO DA CUNHA & A.M. FRIAS MARTINS, 1990. A description of the zonation patterns of molluscs and other important biota on the south coast of Sao Miguel, Azores. In: MARTINS, A.M.F. (Ed.), The Marine Fauna and Flora of the Azores (Proceedings of the First International Workshop of Malacology, Sao Miguel, 1988). Aforeana, Supple-

BIELER: VERMETIDAE OF THE AZORES 191
ment 1990: 21-38. [ICZN] International Commission on
Zoological Nomenclature. International Code of Zoological Nomenclature. Third Edition. International Trust for Zoological Nomenclature and British Museum (Nat. Hist.), xx + 338 pp. London.
KEEN, A.M., 1961. A proposed reclassification of the gastropod family Vermetidae. Bulletin of the British Museum (Natural History), Zoology, 7 (3): 181-213, pis. 54-55.
LACAZE-DUTHIERS, H., 1860. Memoire sur 1 'anatomie et l 'embryogenie des vermets (Vermetus triquete r [sic] et V. semisurrectus Phil.). Anna/es des Sciences na-turelles, 4e Serie (Zoologie), 13: 209-296, pis. 4-9.
LEAL, J.H., & P. BOUCHET, 1991. Distribution patterns and dispersal of prosobranch gastropods along a seamount chain in the Atlantic Ocean. Journal of the Marine Biological Association of the United Kingdom, 71 (1): 11-25.
MARTEL, A., & FU-SHIANG CHIA, 1991. Drifting and dispersal of small bivalves and gastropods with direct development. Journal of Experimental Marine Biology and Ecology, 150: 131-147.
MARTINS, H.R., 1990. A selected bibliography of the marine fauna and flora of the Azores. A rquipelago, Life and Earth Sciences, 8: 35-44.
MORCH, O.A.L., 1859. Etudes sur la famille des vermets. Journal de
Conchyliologie, 7 (1858): 342-360.
MORCH, 0.A.L., 1860. Etudes sur la famille des vermets (suite). Jou rnal de Conchyliologie, 8 (1): 27-48.
MORCH, O.A.L., 1861-1862. Review of the Vermetidae. Part I. Pro -ceedings of the Zoological Society of London, 1861: 145-181, pl. 25; Part II: 1861: 326-365; Part III: 1862: 54-83.
MORCH, O.A.L., 1865. Supplementary notes to the review of Vermetidae. Proceedings of the Zoological Society of London, 1865 ( 1 ): 96-99.
MORCH, O.A.L., 1871. Uebersicht der europaischen Malakozaologische 128-131.
Vermetiden. Blatter, 18:
MONTEROSATO, T., 1878. Enumerazione e sinonimia delle conchiglie mediterranee. Giornale Scienze Naturali ed Economiche, Palermo, 13: 61-115.
MONTEROSATO, T ., 1892. Monografia dei Vermeti del Mediterraneo. Bullettino della Societa Malacologica Italiana, 17: 7-48, pis. 1-7.
MORTON, B., 1990. The intertidal ecology of Ilheu de Vila Franca - a drowned volcanic crater in the Azores. In: MARTINS, A.M.F. (Ed.), The Marine Fauna and Flora of the Azares (Proceedings of the First International Workshop of Malacology, Siio Miguel, 1988). A~oreana, Supplement 1990: 3-20.
MORTON, J.E., 1965. Form and function in the evolution of the Vermetidae. Bulletin of the Bri-

192 A<;OREANA SUPPLEMENT 1995: 173-192
tish Museum (Natural History), Zoology, 11(9): 583-630.
NOBRE, A., 1936. Moluscos marinhos de Portugal. 2° Volume, 378 pp., pis. 81-86. Barcelos.
PHILIPPI, R.A., 1836. Enumeratio molluscorum Siciliae cum viventium tum in tellure tertiaria fossilium, quae in itinere suo observavit, 261 pp., 12 pis. S. Schroppii & Soc., Berlin.
PRUD'HOMME VAN REINE, W.F., 1988. Phytogeography of seaweeds of the Azores. Helgolander
Meeresuntersuchungen, 42: 165-185.
SALDANHA, L., [1979]. Fauna Submarina Atltintica. Portugal Continental, Afores, Madeira, 1 7 9 ( +22) pp. Publicarroes Europa-America. [date teste Martins, 1990).
SIMROTH, H., 1888. Zur Kenntniss der Azorenfauna. Mit Beitragen von Prof. Dr. von Martens, Dr. F. Hilgendorf und S. Clessin. Archiv fiir Naturgeschichte, (54) 1 (3): 179-234, pis. 14-15.




















