
THE CAMBRIAN OF THE CADENAS IBÉRICAS (NE SPAIN) AND ITS TRILOBITES

Geol. Mag. 144 (4 ), 2007, pp. 661–686. c© 2007 Cambridge University Press 661doi:10.1017/S0016756807003366 First published online 16 April 2007 Printed in the United Kingdom
Cambrian trilobites from Brzechow, Holy Cross Mountains(Poland) and their significance in stratigraphic correlation
and biogeographic reconstructions
ANNA ZYLINSKA∗ & MONIKA MASIAK†∗Faculty of Geology, University of Warsaw, Al. Zwirki i Wigury 93, PL-02-089 Warsaw, Poland
†Institute of Geological Sciences, Polish Academy of Sciences, ul. Twarda 51/55, PL-00-818 Warsaw, Poland
(Received 23 February 2006; accepted 1 August 2006)
Abstracts – A revision of the trilobite fauna from Brzechow in the Holy Cross Mountains (Poland)shows that the assemblage consists of Palaeolenus medius (Bednarczyk), Issafeniella orlowinensis(Samsonowicz), Kingaspidoides sanctacrucensis (Czarnocki), Paradoxides (Acadoparadoxides)oelandicus Sjogren, P. (A.) cf. mureroensis Sdzuy and P. (A.) sp. It is earliest Middle Cambrianin age according to the West Gondwana standard of Geyer & Landing (2004) and can be correlatedwith the Cephalopyge notabilis Zone of Morocco. Moreover, the co-occurrence of the ranges of P.(A.) oelandicus with P. (A.) cf. mureroensis might suggest that P. (A.) oelandicus appears earlierin the Brzechow sandstones than in Scandinavia, thus the strata exposed here do not find theircorrelative in the Scandinavian succession. As a result, the Banian/Agdzian boundary (Lower–MiddleCambrian boundary according to the West Gondwana standard) probably lies within the KamieniecFormation, and also below the Brzechow sandstones, in the upper part of the Ocieseki Formation. Closesimilarity of the discussed assemblage with contemporaneous assemblages from Morocco, Jordan orNew Brunswick indicates that the fauna is generally of peri-Gondwanan and Gondwanan aspect.
Keywords: Lower Cambrian, Middle Cambrian, stratigraphy, Holy Cross Mountains, Trilobita, Poland.
1. Introduction
The inconspicuous exposure of Cambrian sandstonesin Brzechow yields an assemblage of fossils uniquefor the Holy Cross Mountains, Poland. The trilobitesand accompanying fauna were discovered by Czarnocki(1927), however, palaeontological descriptions andphotographs of the fauna were not presented at thattime. Czarnocki’s collection of the alleged medusoidBrzechowia brzechowiensis (now: Velumbrella czar-nockii Stasinska) from Brzechow was redescribed byStasinska (1960). The trilobite collections from thislocality housed in the Polish Geological Survey inWarsaw were entirely destroyed during the SecondWorld War. Therefore, formal palaeontological de-scriptions of the trilobite fauna from Brzechow werelater based on new specimens collected by Bednarczyk(1970).
A new approach to Velumbrella czarnockii andRotadiscus sp. has recently been discussed in a numberof papers (e.g. Masiak & Zylinska, 1994; Zhu, Zhao &Chen, 2002; Conway Morris & Grazhdankin, 2005;Ivantsov et al. 2005); a modern revision of the trilobiteshas not yet been offered. The redescription of thetrilobite assemblage is particularly important withregard to the new approach to the definition of theLower–Middle Cambrian boundary (Geyer & Landing,
∗Author for correspondence: [email protected]
2004; Zhao et al. 2004; Babcock et al. 2005; Geyer,2005).
The material studied comprises over 250 trilobitespecimens from five collections, including almostthe entire type collection of Bednarczyk (1970).Unpublished field data of Wiesław Bednarczyk havebeen applied to evaluate the thickness of the studiedstrata.
2. Geological setting
The study area is located in the Holy Cross Mountains,which are part of one of the main geotectonicdomains in Europe, the Trans-European Suture Zone(Berthelsen, 1992). On the basis of different faciesdevelopment, stratigraphy and tectonic evolution, theHoly Cross Mountains are divided into two tectono-stratigraphic regions (recognized first by Czarnocki,1919): the southern Kielce Region and the northernŁysogory Region, separated by the WNW–ESE-trending Holy Cross Fault. The Kielce Region isconsidered to be the northern part of the MałopolskaMassif, whereas the Łysogory Region is generally theequivalent of the Łysogory block (see Pozaryski, 1990).
The village of Brzechow lies in the western partof the Kielce Region, about 10 km ESE of Kielce(Fig. 1; see Masiak & Zylinska, 1994, fig. 3 for adetailed map of the area). The WNW–ESE-orientedvalley where Brzechow lies is bounded from the

662 A. ZYLINSKA & M. MASIAK
Figure 1. Geological sketch map of the Holy Cross Mountains showing the distribution of Cambrian deposits (compiled fromSamsonowicz, 1962 and Orłowski, 1975b, 1992, with modifications), with location of the studied exposure. Lower and MiddleCambrian deposits as indicated on the map follow the traditional subdivisions. HCM – Holy Cross Mountains, TTL – Teisseyre–Tornquist Line, EEC – East European Craton.
north by low hills generally built of N-dipping LowerDevonian sandstones forming the northern limb ofthe Brzechow anticline. Cambrian strata build thecore part of the anticline, which currently is a flat-bottom valley. At present, only coarse- and medium-grained sandstones can be observed as heaps of stoneson fields and field balks in the area, particularlyafter spring ploughing. The large number and size ofthe sandstone fragments on the fields suggests closevicinity of the bedrock. According to the unpublisheddata of W. Bednarczyk, who dug trenches across thevalley in the mid-1960s, apart from the yellow, reddishor white-coloured coarse-grained sandstones, as wellas olive-coloured medium-grained sandstones with avariable degree of quartzitization, the Cambrian stratacomprise also olive-coloured siltstones and clayey, siltyand sandy shales. The presence of strata susceptibleto erosion is responsible for the inversion of theanticline. The beds observed by W. Bednarczyk inthe trenches often showed changing dip directions,suggesting folding and tectonic repetition of thesuccession. The lithological similarities between thestrata observed by W. Bednarczyk have been usedherein to distinguish several lithological associationswithin the succession. Based on the measured dipvalues (Wiesław Bednarczyk, unpub. data), we havereconstructed the orientation of these associations, androughly estimated the thickness of the succession asnot exceeding 90 m.
The presently accessible coarse-grained sandstonescomprise well-rounded quartz grains with syntaxialrims and sutured grains. Metamorphic quartz is alsopresent, and the sediment is petrologically mature.
Numerous large-sized clay clasts occur and thedeposit displays poor sorting, thus pointing to current-dominated sedimentation (Masiak & Zylinska, 1994).The lithology and lack of trace fossils, as well as thehigh degree of quartzitization, point to the SłowiecFormation rather than to the Ocieseki Formation(Fig. 2). The latter is characterized by fine-grained,
Figure 2. Position of the Brzechow sandstones (black spot)in relation to the Lower/Middle Cambrian biostratigraphicand lithostratigraphic scheme in the Kielce Region (HolyCross Mountains) (after Orłowski, 1975b, 1988, 1992, withmodifications of Masiak (M. Masiak, unpub. M.Sc. thesis, Univ.Warsaw, 1992) and this paper).

Lower–Middle Cambrian trilobites, Poland 663
well-sorted sandstones and siltstones, with a lowdegree of quartzitization, and numerous trace fossils(Orłowski, 1975b, 1989, 1992). It should, however, bepointed out that this comparison can be made onlyfor the sandstone, upper part of the 90 m successionfrom Brzechow. Taking into account the fact thatthe sandstones are intercalated and underlain by finesiliciclastic sediments, it is possible that the Brzechowsuccession may represent a transition between theOcieseki and Słowiec formations, reflecting a changeof the source area. The fine siliciclastics underlyingthe Brzechow sandstones might in this case representan unknown member within the uppermost part of theOcieseki Formation, or even the Kamieniec Formation,although the latter possibility is rather improbabledue to the fact that the westernmost occurrence ofthe Kamieniec Formation lies almost 20 km eastwardsof Brzechow, in the Zareby 2 borehole (Bednarczyk,Jurkiewicz & Orłowski, 1965; Jurkiewicz, 1971). Thus,in the lithostratigraphic scheme of Orłowski (1975b),the Brzechow sandstones are assigned to the SłowiecFormation (Fig. 2), cropping out within the OrłowinyRange (with Słowiec Hill), and further eastwards inthe Wygiełzow Range (Fig. 1) (Samsonowicz, 1962;Orłowski, 1971, 1975b, 1988, 1992).
3. Biostratigraphy
3.a. Review of earlier concepts
The Brzechow exposures were mentioned for the firsttime in literature by Czarnocki (1927). The trilobitesreported then from clayey shales with intercalationsof sandstones included Protolenus (Bergeronia) Rade-gasti sp.n., Protolenus Bodzanti sp.n. and Protolenus(?) Percuni sp.n. (original spelling after Czarnocki,1927). Unfortunately, as is the case with almost allspecies created by Jan Czarnocki, these fossils wereneither described nor illustrated, and the collectionsgenerally do not exist, although Czarnocki (1927)pointed out the close resemblance of the latter twoforms to Paradoxides, considering them the oldestrepresentatives of this genus. At that time bedswith these fossils were believed to represent LowerCambrian strata. This view was also later expressed byCzarnocki (1933) and Stasinska (1960).
The first after Jan Czarnocki to collect new trilobitespecimens in Brzechow was Bednarczyk (1970).Because the original collections were lost and couldnot be described, the trilobite species mentioned byCzarnocki (1927, 1933) should have been treated asnomina nuda. However, some specimens of trilobitesfrom the Holy Cross Mountains were sent out before theSecond World War to foreign geological institutions,either by Jan Czarnocki himself or by the PolishGeological Survey, of which he was the head. Forexample, such specimens were sent in 1932 to theSmithsonian Institution in Washington, D.C. Latex
casts of these specimens were made available toStanisław Orłowski in 1963 by A. R. (Pete) Palmer.Bednarczyk was also able to examine the latex casts ofthe Brzechow specimens when writing his paper (seeBednarczyk, 1970, p. 31). This, in turn, must have ledto improper determination of the author of Protolenusbodzanti as Czarnocki, 1927 (Bednarczyk, 1970). IfBednarczyk wanted to retain Protolenus bodzanti, heshould have given his name as the author in theoriginal paper. Eventually, this was corrected in laterpublications (Orłowski, 1988; Bednarczyk & Orłowskiin Pajchlowa, 1990).
Specimens of Paradoxides oelandicus Sjogren havebeen noted to co-occur with the alleged protolenids;therefore, following the traditional concept of Brøgger(1886), the beds from Brzechow were consideredMiddle Cambrian in age (Bednarczyk, 1970). This viewhas also been followed by Orłowski (1988), Masiak (M.Masiak, unpub. M.Sc. thesis, Univ. Warsaw, 1992) andMasiak & Zylinska (1994). A contradictory, althoughunsupported, conclusion about the Early Cambrian ageof the strata has been recently expressed by Lendzion(2001).
3.b. Lower–Middle Cambrian boundary problem
Traditionally, the Middle Cambrian deposits of theHoly Cross Mountains were compared to the successionin Scandinavia (Orłowski, 1964a, b), and thus thescheme of Westergard (1946) was applied. In this view,the definition of the Lower–Middle Cambrian boundaryfollows the concept of Brøgger (1886), with the base ofthe Middle Cambrian marked by the first appearance ofParadoxides sensu lato. Besides Scandinavia (Brøgger,1886; Westergard, 1946), this concept has also beenapplied in Poland (e.g. Orłowski, 1964a, b; Lendzion,1976), Bohemia (e.g. Snajdr, 1958; Havlıcek, 1971),Spain (e.g. Sdzuy, 1971) and Morocco (e.g. Hupe,1953; Geyer, 1990a; Geyer & Palmer, 1995). On theother hand, the extinction of olenelloids was chosento mark the Lower–Middle Cambrian boundary inNorth America (Walcott, 1891). The ‘type area’ ofBrøgger’s concept is Scandinavia, where the Lower–Middle Cambrian transition is marked by a sharp faunalcontrast, which, as has been eventually proved, is aresult of a stratigraphical gap at the Lower–MiddleCambrian boundary (Bergstrom & Gee, 1985) linkedwith late Early Cambrian circum-Iapetus regressionsreferred to the ‘Hawke Bay Event’ (Palmer & James,1980). Therefore, it took a time to discover that notonly do the olenelloids and paradoxidids overlap intheir ranges (e.g. in Morocco: Geyer, 1993), but alsothat the disappearance of olenelloids and appearance ofparadoxidids are diachronous events in different partsof the world (Geyer & Palmer, 1995). As a result,it has been shown that the Lower–Middle Cambrianboundary cannot be further based on either concept.

664 A. ZYLINSKA & M. MASIAK
Practically since 1999, the aim of the InternationalSubcommission on Cambrian Stratigraphy (Geyer &Shergold, 2000) has been to select potential correlationdata, on which the Lower–Middle Cambrian boundarycould be based (see Babcock et al. 2005; Geyer,2005, for details). Although it has been asserted thattrilobites should act as such correlation tools, in thesaid interval in particular they display fairly highendemicity; moreover, in different palaeogeographicareas this endemicity is ‘overprinted’ by biofacies ofdifferent bathymetry, each characterized by their ownsuite of trilobites. Therefore it is in many cases not clearwhether differences in the composition of particularassemblages result from bathymetric or biogeographiccauses, or both.
One of the most widely accepted horizons fordefining the base of the Middle Cambrian stage isthe first appearance datum (FAD) of Oryctocephalusindicus (Reed) (e.g. Sundberg & McCollum, 1997;Zhao et al. 2004; Geyer, 2005). Oryctocephalidshave a fairly wide global distribution (Sundberg &McCollum, 1997). However, they are confined to open-shelf to slope environments of the intertropical regions.Furthermore, apart from the problems linked with thecorrect taxonomic assignment of the oryctocephalidspecies (large degree of intraspecific variability andlarge susceptibility to tectonic distortion of smallindividuals) (Sundberg & McCollum, 1997; Geyer,2005), this group of trilobites has so far not beenrecognized in West Gondwana and Baltica (but seeGeyer, 2006).
Other possible correlation horizons in the Lower–Middle Cambrian boundary interval, which arediscussed with regard to the global Lower–MiddleCambrian boundary standard, include: (1) FADof Ovatoryctocara granulata Chernysheva and/orKiskinella cristata Romanenko & Romanenko; (2)FAD of Arthricocephalus chauveaui Bergeron; (3) baseof the Acidiscus–Cephalopyge assemblage zone, and(4) base of the Serrodiscus–Triangulaspis–Hebediscusband (Geyer & Shergold, 2000; Geyer, 2005). Of thetaxa defining these levels, none has a distribution wideenough to cover all the Cambrian continents. When,however, auxiliary taxa are included for correlationbetween regions and/or palaeocontinents, the baseof the Acidiscus–Cephalopyge assemblage zone andthe base of the Serrodiscus–Triangulaspis–Hebediscusband can serve as correlation levels on almost allCambrian continents (see Geyer, 2005, fig. 7).
Recently, a redefinition of the Lower–MiddleCambrian boundary based on the Moroccan stages hasbeen suggested for West Gondwana (Geyer & Landing,2004, fig. 2; Fig. 4). Following this, the Banian Stage(Atlasian Series), defined by an antatlasiine–strenuelline–saukiandine-dominated trilobite assem-blage, passes into the Agdzian Stage (CeltiberianSeries) defined by the dominance of protolenine–ellipsocephaline faunas (e.g. Geyer, 1990b; Geyer &
Palmer, 1995; Geyer & Landing, 2004; Fig. 4). Thus,the newly defined Lower–Middle Cambrian boundarycorresponding to the Banian/Agdzian boundarywould be marked by a facies-independent faunalturnover marking a stratigraphic interval evidently ofbetter resolution than the diachronous appearance ofparadoxidids (Geyer & Landing, 2004). It should bestressed, however, that this approach can be appliedonly in the West Gondwanan and Avalonian succes-sions. The Lower–Middle Cambrian boundary forWest Gondwana lies below the base of the Acidiscus–Cephalopyge assemblage zone (Geyer & Landing,2004; Geyer, 2005, fig. 8). The FAD of Oryctocephalusindicus correlates to the uppermost part of theMoroccan Middle Cambrian Cephalopyge notabilisZone (Geyer & Shergold, 2000; Geyer, 2005, fig. 8).
3.c. Biostratigraphic analysis of the Brzechow trilobites
Based on the taxonomic revision of the Brzechowtrilobites (see systematic descriptions, Section 6.b), theassemblage is dominated by Palaeolenus medius (Bed-narczyk) and Issafeniella orlowinensis (Samsonowicz),which in almost equal proportions (40 % and 35 %,respectively) make up 75 % of the collection (Fig. 3).The remaining trilobites include Kingaspidoides sanc-tacrucensis (Czarnocki) (15 %), representatives ofParadoxides (Acadoparadoxides) spp. (6 %) (P. (A.)oelandicus Sjogren, P. (A.) cf. mureroensis Sdzuy andP. (A.) sp.) and Myopsolenites kielcensis (Bednarczyk)(4 %). It is quite probable, however, that this proportiondoes not reflect the original abundance of the trilobitesin the environment, and is biased by sedimentaryconditions, which results in the higher proportion ofsmaller individuals and lower numbers of generallylarger individuals.
Palaeolenids are small trilobites numerously knownfrom China (East Gondwana) and Siberia (Mansuy,1912; Chernysheva et al. 1956; Sivov, 1960; Zhang,
Figure 3. Pie-chart showing the content of particular taxa in theBrzechow assemblage.

Lower–Middle Cambrian trilobites, Poland 665
Figure 4. Chrono- and biostratigraphic correlation of the Lower–Middle Cambrain boundary interval between Morocco, Iberia andthe Holy Cross Mountains against the West Gondwanan standard. Modified from Geyer & Landing (2004).
1966; Repina et al. 1999), where they occur aroundthe Lower–Middle Cambrian boundary interval (baseof the Amgan in Siberia or the upper Tsanglangpuanin North China) (Geyer & Shergold, 2000). Thereare, however, several occurrences of these trilobitesrecently noted also in West Gondwana successions.Representatives of the Palaeoleninae (Palaeolenusantiquus (Chernysheva)) have been observed in theBurj Formation in Jordan (Rushton & Powell, 1998),which correlates to the Hupeolenus Zone of Morocco(see Fig. 4). Palaeoleninae (Schistocephalus? ornatusGeyer) and Ichangiinae (Gigoutella atlasensis Hupe)have been described from the Cephalopyge notabilisZone of Morocco (Geyer, 1998; see Fig. 4). Otheroccurrences of the Palaeoleninae include: Ferralsiablayaci Cobbold from the Montagne Noire (e.g. Alvaro,Linan & Vizcaıno, 1998), and Ferralsia saxonica Geyer& Elicki from the Gorlitz Synclinorium in Germanyand the Ossa–Morena zone in Spain (Geyer & Elicki,1995; Alvaro, Linan & Vizcaıno, 1998). A middleto late ‘Marianian’ age has been determined for F.saxonica and a middle ‘Marianian’ to early ‘Bilbilian’age for F. blayaci (Alvaro, Linan & Vizcaıno, 1998),thus they correlate to the upper Banian of Morocco(Geyer & Shergold, 2000; see Fig. 4). Concluding, thepalaeolenines span the Banian/Agdzian boundary inWest Gondwana (Geyer & Landing, 2004), with theearliest occurrences in France, Spain and Germanyand the latest in Jordan and Morocco. This appearsto conflict with the migration of the biodiversitypeak across the Lower–Middle Cambrian boundaryinterval in West Gondwana from Morocco to theIberian Peninsula and the Montagne Noire as indicated
by Alvaro et al. (2003) and Alvaro & Clausen(2005).
In Morocco, the antatlasiine Issafeniella Geyer isrepresented by three species (I. turgida Geyer, I.modesta Geyer and I. ? marocana (Hupe)), whichcome from the middle and upper Banian Antatlasiaguttapluviae and Sectigena zones in the Issafen Form-ation (Geyer, 1990b). Additionally, Geyer (1990b) alsoassigned to this genus I. sampelayoi (Richter & Richter)from the uppermost Lower Cambrian deposits in Spain(Richter & Richter, 1940; Henningsmoen, 1958; Sdzuy,1962), I. melendezi (Gil Cid) from the ‘Marianian’ ofAlanıs in the Sierra Morena (Gil Cid, 1972) and twotaxa from the Protolenus–Strenuaeva Zone of the HolyCross Mountains (Samsonowicz, 1959a; Orłowski,1985a): I. trifida (Orłowski) and I. orlowinensis(Samsonowicz), the latter discussed herein. Thus, thegenus is characteristic of the interval encompassingthe middle and upper Banian, and possibly alsothe lower Agdzian of West Gondwana (Geyer &Landing, 2004; see Fig. 4).
Kingaspidoides sanctacrucensis (Czarnocki) has amuch longer age range in the Holy Cross Mountains.This very characteristic endemic ellipsocephaline hasbeen observed in exposures near Widełki (Łapigrosz)(Fig. 1) in strata assigned to the Protolenus–StrenuaevaZone (corresponding to the upper Banian and lower Ag-dzian of Morocco; see Fig. 4), where it is most abund-ant, and in slightly younger deposits in Brzechow;additionally, Orłowski (1964a) lists K. sanctacrucensisfrom the Paradoxides insularis Zone in Helenow.Moreover, Orłowski’s collection yields a single cran-idium of this species from exposures on Słowiec

666 A. ZYLINSKA & M. MASIAK
Hill from younger strata assigned to the Paradoxidespolonicus Zone, which in turn can be correlatedwith the lower part of the Caesaraugustian (Geyer &Landing, 2004). An occurrence from the much youngerstrata (corresponding to the Caesaraugustian or evenLanguedocian) in the Pieprzowe Mountains has alsobeen noted (see Orłowski, 1964a, table 1, p. 42),although the specimen has not been illustrated ordiscussed in the text. In Morocco, the largest diversityof Kingaspidoides spp. is observed in the Cephalopygenotabilis Zone (Agdzian series) with representativesof at least seven species; of these, only Kingaspidoidesborjensis Geyer ranges from the Hupeolenus Zone up tothe Kymataspis arenosa Zone (Geyer, 1990b), that is, itcovers the entire Agdzian of West Gondwana (Geyer &Landing, 2004).
The Myopsolenites issue has been recently extens-ively discussed in Geyer & Landing (2004). Followingthis, the Onaraspis clade (including Myopsolenites)occurs in a fairly narrow stratigraphic range generallyspanning the co-occurrence of typical ‘Lower’ Cam-brian olenellids and ‘Middle’ Cambrian paradoxidids.Moreover, the geographic range of this clade of asyet unknown affinities has been proved to be of largeextent (Geyer & Landing, 2004), with occurrencesin the Australian Ordian (Onaraspis somniurna Opik,O. adusta Opik and O. rubra Jell: Opik, 1968; Jellin Bengston et al. 1990), in the terminal LowerCambrian Megapalaeolenus Zone of the YangtzePlatform (Chengkouaspis longioculus Zhang & Lin:Zhang et al. 1980), in the possibly terminal LowerCambrian deposits in Israel (Myopsolenites palmeri(Parnes): Parnes, 1971; see also comments in Geyer &Landing, 2004), in the early Agdzian (‘Bilbilian’) ofSpain (Onaraspis altus (Linan & Gozalo): Linan &Gozalo, 1986; Gozalo & Linan, 1997) and theAgdzian Cephalopyge notabilis Zone of Morocco(Myopsolenites boutiouiti Geyer & Landing: Geyer &Landing, 2004), thus within a narrow interval in theEast and West Gondwana successions (see Fig. 4).Other possible members of the Onaraspis clade mayinclude Conomicmacca plana (Matthew) from theProtolenus elegans Zone of southern New Brunswick(Westrop & Landing, 2000), Bathynotellus yermolaeviLerm. (MS) from the Middle Cambrian succession ofNovaya Zemlya (Lermontova, 1940), as well as Rec-tifrontinella olhae Konstantinenko from the supposedFurongian (upper Cambrian) strata of the CarpathianForedeep in the Ukraine (Drygant & Konstantinenko,2001). The Furongian age of these strata is based onold geophysical data, and the presence of a possibleelement of the Onaraspis clade within them mightsuggest a much earlier age of these deposits. Thetwo latter occurrences may extend the geographicrange of the Onaraspis clade onto the margin of thepalaeocontinent of Baltica.
Paradoxides (Acadoparadoxides) oelandicusSjogren is a typical Scandinavian taxon and has
been described from the basal Middle CambrianAcadoparadoxides oelandicus Superzone (or stage),both from the Eccaparadoxides insularis andPtychagnostus praecurrens zones in Sweden (Sjogren,1872; Linnarsson, 1877; Westergard, 1928, 1936) andin the upper part of the Acadoparadoxides oelandicusStage in Norway (Henningsmoen, 1952). Theoccurrences of P. (A.) oelandicus outside Scandinavia(apart from Brzechow) seem to be doubtful (Cobbold &Pocock, 1934; Lake, 1935; cf. Henningsmoen, 1952,p. 25; A. Rushton, pers. comm. 2005).
The FAD of Paradoxides (Acadoparadoxides)mureroensis Sdzuy, the eponymous species for thelowest Middle Cambrian zone in Spain, is consideredto be the base of the upper Agdzian (‘Leonian’)(Fig. 4) and thus defines the Lower–Middle Cambrianboundary therein (e.g. Sdzuy, 1958; Gozalo et al. 1993;Geyer, 1998). The species has also been observedin the Cal Tepe Formation of approximately similarage in the western Taurids, Turkey (Dean & Ozgul,1994; Dean, 2005) and in the Cephalopyge notabilisZone of Morocco (Geyer, 1998). P. (A.) mureroensisis very closely related to P. (A.) harlani (Green) fromthe lowermost Acadian series of Massachusetts andNewfoundland (Geyer & Landing, 2001; Fletcher et al.2005), to the ‘giant Moroccan Paradoxides’, P. (A.)briareus Geyer from the Cephalopyge notabilis Zoneand the lowermost part of the Ornamentaspis frequensZone in the Agdzian of Morocco (Geyer, 1993; Geyer &Landing, 2001), as well as to P. (A.) nobilis Geyer fromthe uppermost Hupeolenus Zone, the Cephalopygenotabilis Zone and the basal Ornamentaspis frequensZone in the Agdzian of Morocco (Geyer, 1998).Particularly, it has been claimed that pygidia of thesefour species form a morphocline that extends from theearliest P. (A.) nobilis, through the stratigraphicallyyounger P. (A.) mureroensis and P. (A.) briareus, toP. (A.) harlani (Geyer & Landing, 2001), although thishas been contested by Fletcher et al. (2005).
Concluding, most elements of the Brzechow as-semblage point to its correspondence to the Cephalo-pyge notabilis Zone of Morocco (see also Geyer &Landing, 2004, fig. 2; Fig. 4). Significant, however, isthe large abundance of Issafeniella which in Moroccoappears in slightly older strata (Antatlasia guttapluviaeand Sectigena zones). Moreover, the co-occurrenceof P. (A.) oelandicus with P. (A.) cf. mureroensismight suggest an earlier appearance of the first speciesthan in Scandinavia, which would mean that theBrzechow sandstones do not find their correlative inthe Cambrian succession of the epicontinental sea inBaltica, where generally there is a gap at the Lower–Middle Cambrian boundary (e.g. Lendzion, 1976;Bergstrom & Gee, 1985), approximately equivalentto the circum-Iapetus regressive ‘Hawke Bay Event’(Palmer & James, 1980). As a result, the FAD of P.(A.) oelandicus probably pre-dates its appearance inBaltica.
Lower–Middle Cambrian trilobites, Poland 667
3.d. Lower–Middle Cambrian boundary intervalin the Holy Cross Mountains
Deposits from the Lower–Middle Cambrian boundaryinterval in the Holy Cross Mountains are referred to theProtolenus–Strenuaeva and the Paradoxides insulariszones. They belong to four lithostratigraphic units, thatis, the Lower Cambrian Kamieniec Formation, theLower–Middle Cambrian Ocieseki Formation andthe Middle Cambrian Słowiec and Usarzow forma-tions (Fig. 2). Distribution of trilobites within theseformations is random, and boundaries between thelithostratigraphic units are typically not exposed.
The Kamieniec Formation from the eastern partof the Holy Cross Mountains is represented by clayand clay-siltstone shales with intercalations of finesandstones. The upper part of this unit (Protolenus–Strenuaeva Zone) is characterized by the antatlasiinesIssafeniella trifida (Orłowski) and Berabichia kiaeri(Czarnocki), protolenines Protolenus (Protolenus) ex-pectans Orłowski (Fig. 5a), Protolenus (Hupeolenus)czarnockii Orłowski & Bednarczyk, Hamatolenus(Hamatolenus) glabellosus (Orłowski), as well asthe corynexochid Strettonia cobboldi Orłowski &Bednarczyk and eodiscids Cobboldites comleyen-sis (Cobbold) and Serrodiscus primarius Orłowski(Samsonowicz, 1962; Bednarczyk, Jurkiewicz &Orłowski, 1965; Orłowski, 1985a; Geyer, 1990b).The strata were deposited in low-energy deep-waterenvironments, referred to the outer shelf (Studencki,1988; Mizerski, Orłowski & Waksmundzki, 1991).The protolenines most probably point to an intervalcorresponding to the Hupeolenus–Cephalopyge notab-ilis zones of Morocco, with, for example, Protolenus(P.) expectans falling into the definition of species ofProtolenus (P.) such as Protolenus (P.) densigranulatusGeyer in Geyer (1990b, text-fig. 63, pl. 45, figs 7–11) and in Geyer & Landing (2004, text-fig. 7.1,2,4). Correspondence with the Cephalopyge notabilisZone is also likely through Strettonia cobboldi, asStrettonia sp. has been noted from the Cephalopygenotabilis Zone in the Siskawn region (Geyer, 1994;
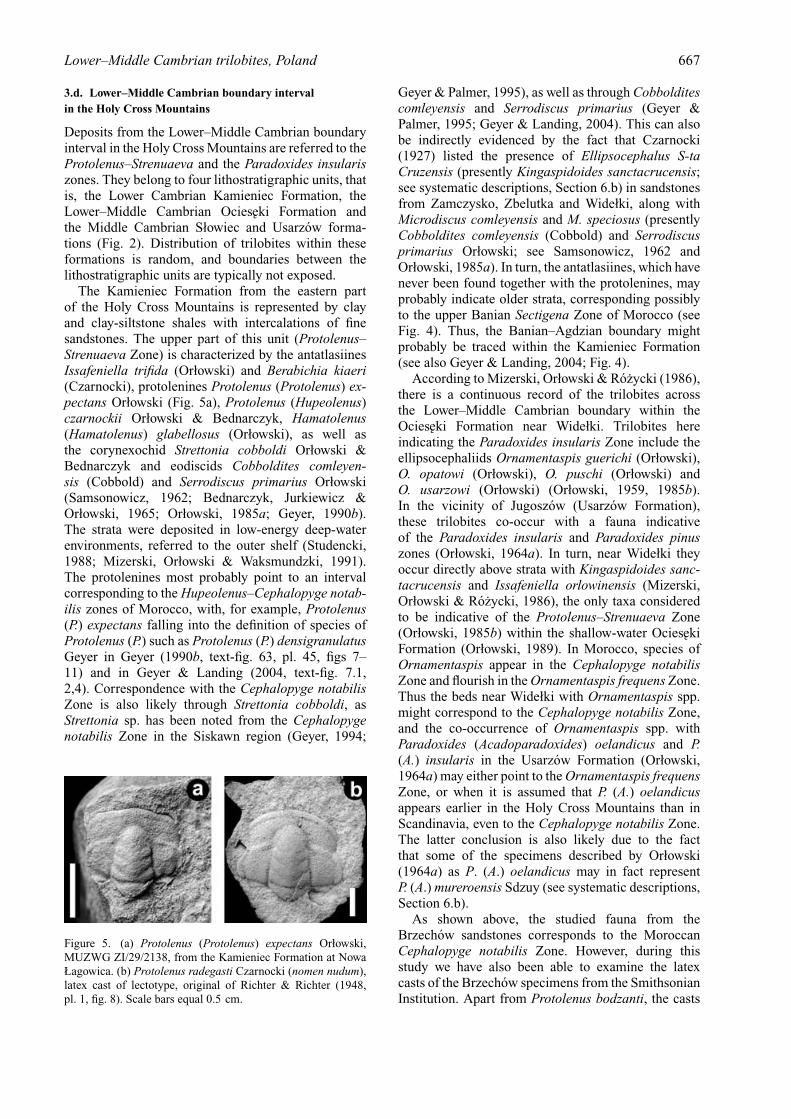
Figure 5. (a) Protolenus (Protolenus) expectans Orłowski,MUZWG ZI/29/2138, from the Kamieniec Formation at NowaŁagowica. (b) Protolenus radegasti Czarnocki (nomen nudum),latex cast of lectotype, original of Richter & Richter (1948,pl. 1, fig. 8). Scale bars equal 0.5 cm.
Geyer & Palmer, 1995), as well as through Cobbolditescomleyensis and Serrodiscus primarius (Geyer &Palmer, 1995; Geyer & Landing, 2004). This can alsobe indirectly evidenced by the fact that Czarnocki(1927) listed the presence of Ellipsocephalus S-taCruzensis (presently Kingaspidoides sanctacrucensis;see systematic descriptions, Section 6.b) in sandstonesfrom Zamczysko, Zbelutka and Widełki, along withMicrodiscus comleyensis and M. speciosus (presentlyCobboldites comleyensis (Cobbold) and Serrodiscusprimarius Orłowski; see Samsonowicz, 1962 andOrłowski, 1985a). In turn, the antatlasiines, which havenever been found together with the protolenines, mayprobably indicate older strata, corresponding possiblyto the upper Banian Sectigena Zone of Morocco (seeFig. 4). Thus, the Banian–Agdzian boundary mightprobably be traced within the Kamieniec Formation(see also Geyer & Landing, 2004; Fig. 4).
According to Mizerski, Orłowski & Rozycki (1986),there is a continuous record of the trilobites acrossthe Lower–Middle Cambrian boundary within theOcieseki Formation near Widełki. Trilobites hereindicating the Paradoxides insularis Zone include theellipsocephaliids Ornamentaspis guerichi (Orłowski),O. opatowi (Orłowski), O. puschi (Orłowski) andO. usarzowi (Orłowski) (Orłowski, 1959, 1985b).In the vicinity of Jugoszow (Usarzow Formation),these trilobites co-occur with a fauna indicativeof the Paradoxides insularis and Paradoxides pinuszones (Orłowski, 1964a). In turn, near Widełki theyoccur directly above strata with Kingaspidoides sanc-tacrucensis and Issafeniella orlowinensis (Mizerski,Orłowski & Rozycki, 1986), the only taxa consideredto be indicative of the Protolenus–Strenuaeva Zone(Orłowski, 1985b) within the shallow-water OciesekiFormation (Orłowski, 1989). In Morocco, species ofOrnamentaspis appear in the Cephalopyge notabilisZone and flourish in the Ornamentaspis frequens Zone.Thus the beds near Widełki with Ornamentaspis spp.might correspond to the Cephalopyge notabilis Zone,and the co-occurrence of Ornamentaspis spp. withParadoxides (Acadoparadoxides) oelandicus and P.(A.) insularis in the Usarzow Formation (Orłowski,1964a) may either point to the Ornamentaspis frequensZone, or when it is assumed that P. (A.) oelandicusappears earlier in the Holy Cross Mountains than inScandinavia, even to the Cephalopyge notabilis Zone.The latter conclusion is also likely due to the factthat some of the specimens described by Orłowski(1964a) as P. (A.) oelandicus may in fact representP. (A.) mureroensis Sdzuy (see systematic descriptions,Section 6.b).
As shown above, the studied fauna from theBrzechow sandstones corresponds to the MoroccanCephalopyge notabilis Zone. However, during thisstudy we have also been able to examine the latexcasts of the Brzechow specimens from the SmithsonianInstitution. Apart from Protolenus bodzanti, the casts

668 A. ZYLINSKA & M. MASIAK
include Protolenus radegasti Czarnocki nomen nudum(Fig. 5b). The gypsum cast of the lectotype has beenillustrated in Richter & Richter (1948, pl. 1, fig. 8).Analysis of the photograph of the gypsum cast andthe latex cast at hand indicates that the casts weremade from the same specimen. Czarnocki’s originalspecimens of this species are considered missing.Nevertheless, from the available illustrations and latexcast it seems that P. radegasti (Fig. 5b) might be closelyrelated to Protolenus (Protolenus) expectans Orłowski(Fig. 5a), at present noted only from the KamieniecFormation (Orłowski, 1985a). The existing collectionsof the Brzechow specimens lack forms comparablewith Protolenus radegasti, but if we assume thatsuch fossils were once noted in the study area, theycould possibly indicate the Hupeolenus Zone, thus thelowermost part of the Middle Cambrian stage sensuGeyer & Landing (2004).
4. Palaeogeographic remarks
The unique character of the Brzechow exposure lies inthe fact that, although the area now lies in the directvicinity of the East European Craton, considered asthe palaeocontinent Baltica, the strata yield trilobitesfrom an interval unknown from Scandinavia. As shownby the biostratigraphic analysis, although continuoussections are not exposed, it is highly probable thatthe Holy Cross Mountains succession does not bearsignificant gaps across the Lower–Middle Cambriantransition, linked with a regressive phase in a roughlysimilar position to the Hawke Bay event (Palmer &James, 1980), although the Słowiec and Usarzow form-ations above the Ocieseki and Kamieniec formationsindirectly seem to evidence this fact because they wereboth deposited in shallower marine environments. Theinfluence of a local tectonic phase at the Lower–MiddleCambrian boundary, which caused shallowing and insome cases also emersion has also been postulatedfor the area (Kowalczewski, 1990; Gagała, 2005).The succession of trilobites from the Lower–MiddleCambrian boundary transition on the genus levelclosely resembles that from West Gondwana, and itis possible to correlate the assemblages from differentparts of the succession with the assemblages fromMorocco. Correlatives are also present in the faunasfrom the Burj Formation in Jordan (Rushton & Powell,1998) and in the Hanford Brook Formation fromsouthern New Brunswick (Westrop & Landing, 2000).The only link to Baltica is via P. (A.) oelandicus, but, asshown above, the Brzechow assemblage may reflect theoverlap of the latter species with P. (A.) mureroensis,pre-dating its appearance in Baltica. The Gondwananaspect of the trilobite fauna is additionally indicatedby the co-occurring soft-bodied Velumbrella czarnockiiand Rotadiscus sp. (Stasinska, 1960; Masiak &Zylinska, 1994), organisms similar to disc-like or-ganisms from the Chengjiang and Kaili Lagerstatten
in southwest China (e.g. Zhu, Zhao & Chen, 2002)of lophophorate or echinoderm nature. Apart fromthese, the sandstones also contain remains of brachi-opods, aglaspid-like chelicerates, monoplacophorans(?helcionellids), sclerites of polyplacophorans, as wellas other as yet unidentified fossils (A. Zylinska, unpub.data). The equivalent Agdzian Stage in Morocco isa widespread interval from the foothills of the HighAtlas and much of the Anti-Atlas, characterized by richassemblages with trilobites, brachiopods, helcionellids,bivalves, hyoliths, SSFs and echinoderms (Geyer,Landing & Heldmeier, 1995; Geyer & Landing, 2004).
5. Conclusions
(1) Redescription of the trilobites from Brzechowin the Holy Cross Mountains, Poland, showsthat the assemblage comprises Palaeolenusmedius (Bednarczyk), Issafeniella orlowinensis(Samsonowicz), Kingaspidoides sanctacrucen-sis (Czarnocki), Paradoxides (Acadoparadox-ides) oelandicus Sjogren, P. (A.) cf. mureroensisSdzuy, P. (A.) sp. and Myopsolenites kielcensis(Bednarczyk).
(2) Based on the unpublished data of WiesławBednarczyk, the thickness of the upper part ofthe succession, partly exposed as the Brzechowsandstones, is estimated as not exceeding 90 m.The Brzechow sandstones are included in theSłowiec Formation, whereas the underlying finesiliciclastics might represent the uppermost partof the Ocieseki Formation.
(3) Biostratigraphic analysis of the assemblagespoints to correlation with the Moroccan Ceph-alopyge notabilis Zone. Moreover, the co-occurrence of P. (A.) oelandicus with P. (A.)cf. mureroensis suggests an appearance of thefirst taxon earlier than in Scandinavia, and thus,that the Brzechow sandstones do not find theircorrelatives in the Scandinavian succession.
(4) Biogeographically, the trilobite assemblage fromBrzechow, as well as the Lower–Middle Cam-brian boundary interval in the Kielce regionof the Holy Cross Mountains, bears strongsimilarities to the West Gondwanan successionas recognized in Morocco and Spain. Thesimilarities are observed on the genus level,whereas on species level, trilobites from all theseareas display a distinct endemicity.
6. Palaeontological notes (AZ)
6.a. Material and methods
All specimens studied are preserved as internal orexternal moulds of detached parts of the shield.

Lower–Middle Cambrian trilobites, Poland 669
Cranidia prevail in the collections, whereas pygidia andlibrigenae are much rarer.
The studied trilobites (254 specimens) come fromfive different collections:
(1) collection INGPAN WB (64 specimens; nos 1–47, 49–56, 62, 73–6), housed in the Institute of GeologicalSciences, Polish Academy of Sciences. This is a largepart of the original collection of Wiesław Bednarczyk(1970);
(2) collection MUZWG/ZI/29 (62 specimens; nos 0937–0990), housed in the Museum of the Faculty of Geology,University of Warsaw, collected by Stanisław Orłowskibetween 1992 and 1996 and Monika Masiak between1989 and 1991 (the latter were described in ‘M. Masiak,unpub. M.Sc. thesis, Univ. Warsaw, 1992’);
(3) collection WK (93 specimens; nos 1–76), belonging toWojciech Kozłowski, temporarily housed in the Instituteof Geology at the Faculty of Geology, University ofWarsaw, collected between 1992 and 2002;
(4) collection IGPUW B/II (16 specimens; nos 33, 40, 41,43, 55, 60–67) housed in the Institute of Geology atthe Faculty of Geology, University of Warsaw, collectedbetween 1992 and 2004 by Anna Zylinska and MonikaMasiak;
(5) collection AK (19 specimens, nos 2/TA1–2, 6/TA,7/TA, 8/TA2-1, 30/TA1-2, 41/TA1-2, 42/TA1, TA2, TA3,TA3’, TA4, TA5, TA6, 56/TA, 300/CD2-1, 300/CD2-2, 300/CD3), belonging to Adrian Kin, housed inthe Museum of the Geoscience Friends Association‘Phacops’ in Łodz and collected between 1996 and 1998.
Terms applied to the trilobite exoskeleton are usedaccording to the recommendations of the TrilobiteTreatise (Kaesler, 1997). Measurements of the analysedspecimens have been made with callipers, with anaccuracy of 0.1 mm. Character lengths were measuredeither sagittally (sag.) or exsagittally (exs.) and widthswere measured transversely (tr.). For each specimen themeasurements were taken in one plane. Symbols usedfor sclerite dimensions are taken from Shaw (1956,1957) and Temple (1975). The particular parameterswere calculated with the help of Microsoft Excelsoftware. The values of parameters are preceded bythe term ‘approximately’ when only one specimenis measured; a range of values is given for two tofour specimens, whereas for five specimens or morethe mean value along with the standard deviationis given (after Sundberg & McCollum, 1997, withmodifications). The number of specimens for whichthe length of the cranidium has been measured isgiven in parentheses after the mean value (e.g. n = 6).Specimens were photographed after coating withammonium chloride.
6.b. Systematic descriptions
Superfamily ELLIPSOCEPHALACEA Matthew, 1887Family PALAEOLENIDAE Hupe, 1953
Remarks. The family is closely related to Ellipso-cephalidae Matthew (Geyer, 1990b, 1998), however, itincludes forms which did not give rise to other lineages.
Geyer (1998) suggested a phylogenetic lineage fromPalaeolenus Mansuy and related genera via Megapa-laeolenus Chang to Schistocephalus Chernysheva withan expanding glabella in progressively younger species.Palaeolenidae are generally characteristic of North andEast Gondwana, although numerous genera have alsobeen recognized in Siberia (Chernysheva et al. 1956;Sivov, 1960), as well as in West Gondwana. The lattergenera include representatives of the PalaeoleninaeHupe, as well as the Ichangiinae Zhu (Geyer & Elicki,1995; Alvaro, Linan & Vizcaıno, 1998; Geyer, 1998;Rushton & Powell, 1998). The most significant featuresof the palaeolenid cranidium include a club-shapedglabella, strong and thick eye ridges and a characteristicpattern of lateral glabellar furrows: S2–S4 typicallyequidistant, rarely S3 and S4 more closely spacedcompared to the posterior pairs, S4 slightly obliqueinward and forward, S1 usually bifid, with posteriorbranch curving backwards and often transglabellar(Geyer, 1998; Rushton & Powell, 1998).
Genus Palaeolenus Mansuy, 1912
Type species. Palaeolenus douvillei Mansuy, 1912, bysubsequent designation of Vogdes (1925).
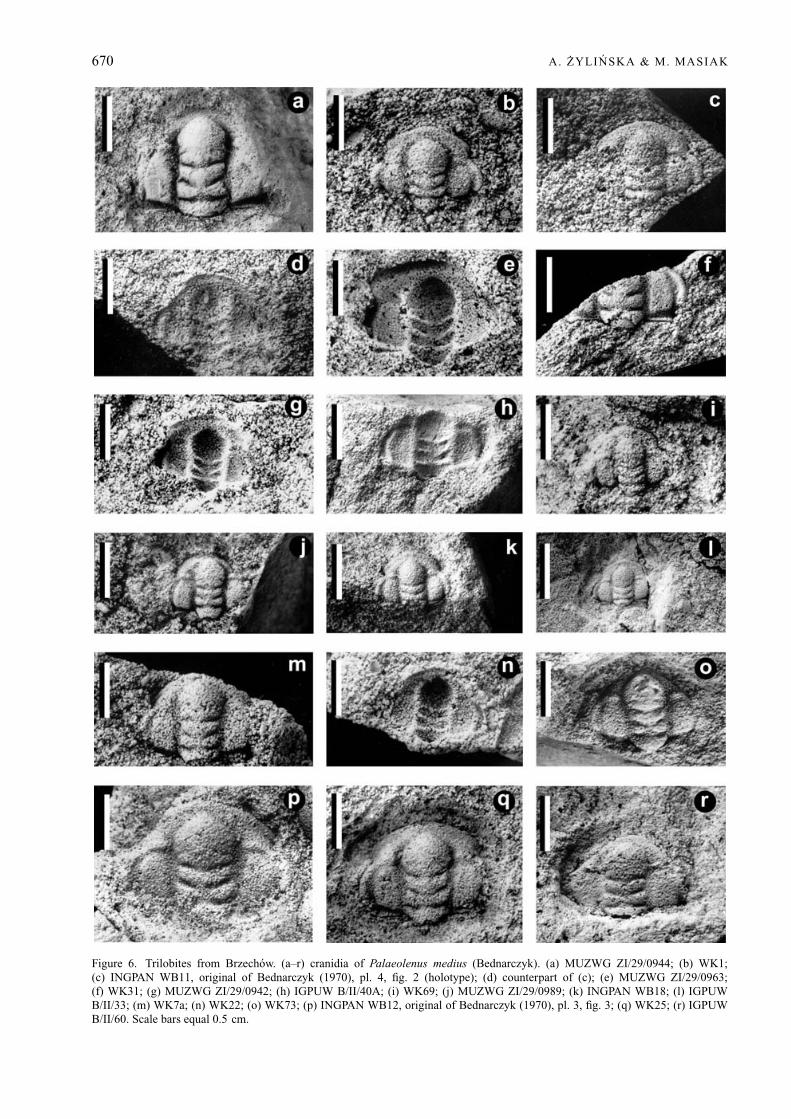
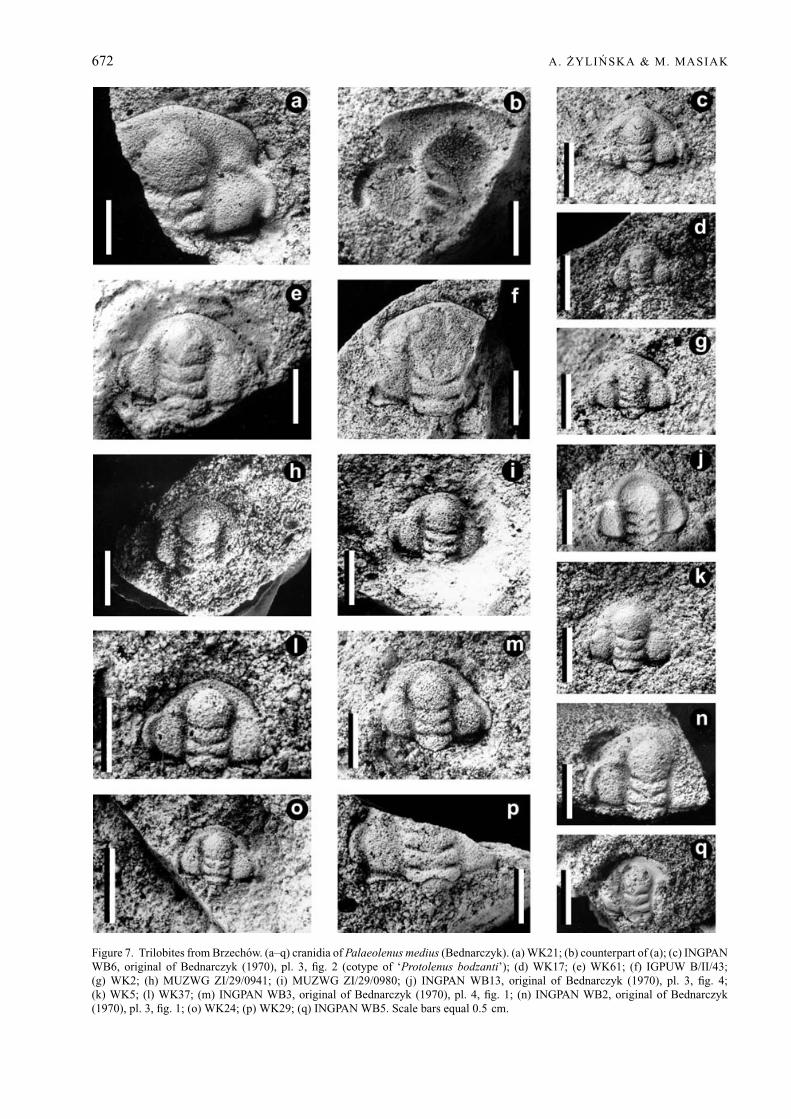
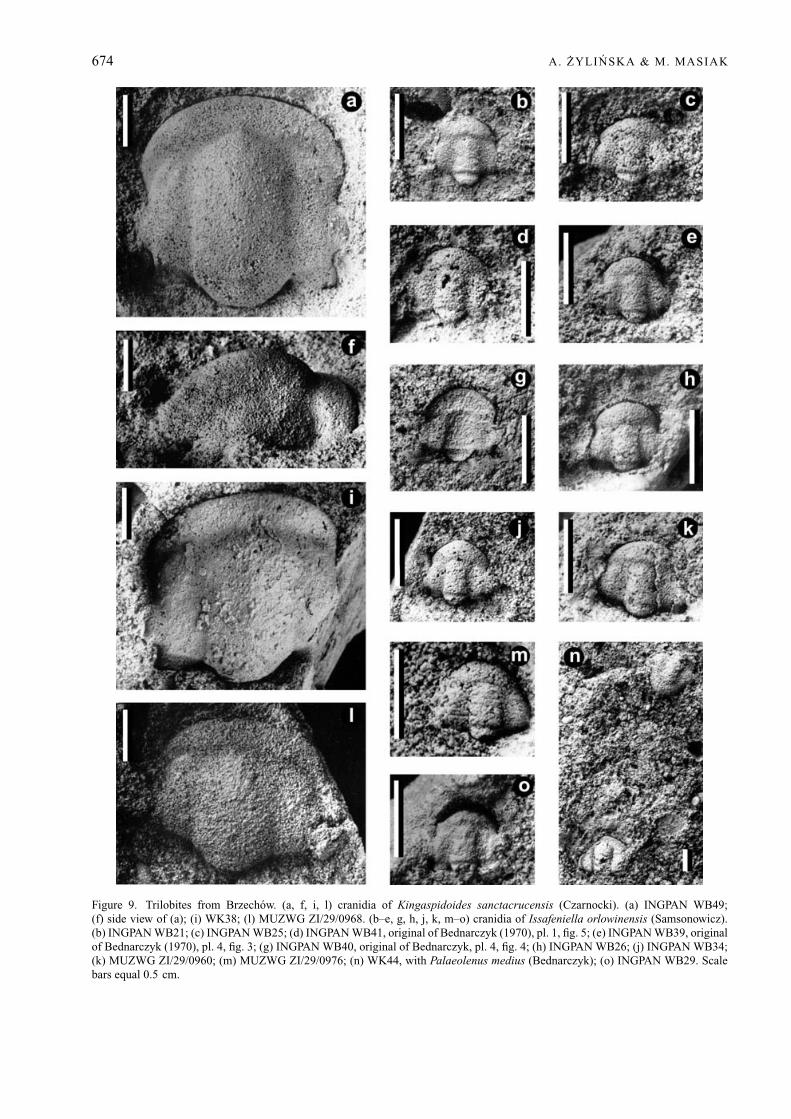
Palaeolenus medius (Bednarczyk, 1970)Figures 6a–r, 7a–q, 8, 9n
1927 Protolenus Bodzanti; Czarnocki, p. 194 (nomennudum).
?1927 Protolenus (?) Percuni; Czarnocki, p. 194 (nomennudum).
v1970 Protolenus bodzanti Czarnocki, 1927 [sic]; Bednar-czyk, pp. 30–1, pl. 3, figs 1–5, pl. 4, fig. 1.
v1970 Protolenus medius n.sp.; Bednarczyk, pp. 31–2, pl.3, fig. 6, pl. 4, fig. 2.
v1990 Protolenus bodzanti Bednarczyk; Bednarczyk &Orłowski in Pajchlowa, p. 57, pl. 16, fig. 2.
v1990 Protolenus medius Bednarczyk; Bednarczyk &Orłowski in Pajchlowa, p. 57, pl. 16, fig. 3.
Types. The holotype of Palaeolenus medius(Bednarczyk) is cranidium INGPAN WB11 fromBrzechow, illustrated in Bednarczyk (1970), pl. 4,fig. 2 as Brz11 (Fig. 6c, d herein, and pl. 16, fig. 3in Bednarczyk & Orłowski in Pajchlowa, 1990).Bednarczyk (1970) selected the ‘cotypus’ of theconspecific Protolenus bodzanti Czarnocki (cranidiumINGPAN WB6, illustrated in Bednarczyk 1970, pl. 3,fig. 2 as Brz5: Fig. 7c herein, and pl. 16, fig. 2in Bednarczyk & Orłowski in Pajchlowa, 1990),however, not from Czarnocki’s original collection,which had been destroyed, but from his own topotypecollection. In papers published after 1970, Bednarczykis considered the author of both Protolenus medius andProtolenus bodzanti (see Orłowski, 1988; Bednarczyk& Orłowski in Pajchlowa, 1990).
Emended diagnosis. A species of Palaeolenus withglabella slightly expanded forward, and with sides
670 A. ZYLINSKA & M. MASIAK
Figure 6. Trilobites from Brzechow. (a–r) cranidia of Palaeolenus medius (Bednarczyk). (a) MUZWG ZI/29/0944; (b) WK1;(c) INGPAN WB11, original of Bednarczyk (1970), pl. 4, fig. 2 (holotype); (d) counterpart of (c); (e) MUZWG ZI/29/0963;(f) WK31; (g) MUZWG ZI/29/0942; (h) IGPUW B/II/40A; (i) WK69; (j) MUZWG ZI/29/0989; (k) INGPAN WB18; (l) IGPUWB/II/33; (m) WK7a; (n) WK22; (o) WK73; (p) INGPAN WB12, original of Bednarczyk (1970), pl. 3, fig. 3; (q) WK25; (r) IGPUWB/II/60. Scale bars equal 0.5 cm.

Lower–Middle Cambrian trilobites, Poland 671
slightly concave inwards at level of S2; four pairs oflateral glabellar furrows, S1 and S2 wide, directedobliquely posteriorly, S1 slightly more curved thanS2 and occasionally transglabellar, S3 and S4 ratherfaint and short, S3 almost straight or slightly curvedanteriorly, S4 much shorter than S3, directed anteriorly;S3 and S4 more closely spaced compared to the pos-terior pairs. Occipital furrow almost straight, occipitalring with strongly convex posterior margin and smallmedian tubercle; anterior part of facial suture notextending laterally beyond palpebral lobe; preglabellarfield sagittally slightly wider than anterior border.
Material. 99 cranidia, variably preserved, in somecases effaced: INGPAN WB1–13, 16–19, 46, 51, 52,54, 73, 74, 76; MUZWG ZI/29/0937–0945, 0947,0949, 0963–0965, 0978, 0980, 0984, 0988–0990;WK1, 2, 4, 5, 7, 9, 13, 17, 21–26, 28–31, 33, 37, 44–46,56, 61, 62, 65, 67, 69, 71, 73; IGPUW B/II/33, 40, 43,60, 61, 63, 65; AK 6/TA, 7/TA, 30/TA1-2; 42/TA1–4, 6,56/TA. Thorax and pygidium unknown. All specimensare from the Paradoxides insularis Zone at Brzechow,Holy Cross Mountains, Poland.
Description. Cranidium subquadrangular, overall con-vexity modest, length 78 ± 7 % of maximum widthacross centre of palpebral lobes (n = 35). Glabellastrongly convex, 86 ± 5 % of cephalic length (n = 46)and 43 ± 4 % of cephalic width across occipital ring(n = 29); middle part of glabella slightly concave in-wards, anterior part (anterior to L2) widening forwards.Frontal lobe rounded or sub-triangular, giving theglabella an overall club-shaped outline. Four pairs oflateral glabellar furrows, S1 and S2 wide, directedposteriorly, S1 slightly more strongly curved than S2and occasionally transglabellar, S3 and S4 obsolete andshort, visible only on some specimens (see Figs 6a–d,j, o–q; 7a, b, j, m, q), S3 almost straight or slightlycurved anteriorly, S4 much shorter than S3, directedanteriorly. Occipital furrow moderately wide in sagittalsection, deeper and slightly backward-directed later-ally, medially with slight forward bend. Occipital ring15 ± 5 % cephalic length (n = 46), sagittally slightlywider than at axial furrows, posterior rim curved.Traces of small tubercle visible on some specimens(see Figs 6b, g; 7c, j, m). Axial furrows rather deepand wide, deeper opposite L2 and almost obsolescentadjacent to frontal lobe of the glabella. Fixigenae evenlyconvex, 73 ± 10 % width of occipital ring (n = 44)and 32 ± 6 % cephalic length (n = 38). Palpebral lobecrescent-shaped, with slightly narrower anterior andposterior ends, length 42 ± 5 % of cephalic length(n = 37). Palpebral furrow rather wide and not verydeep, widest in posterior part, narrows anteriorly.Palpebral lobe separated from eye ridge by shallow,indistinct groove. Eye ridges exsagittally as wide aspalpebral lobe, reaching glabella at level of S2, notconnected to glabella. Anterior branch of facial sutureshort and slightly divergent from palpebral lobes tofrontal margin, but not extending exsagittally beyond
palpebral lobe, then curved sharply adaxially, and insome cases giving the anterior border a sub-triangularshape. Posterior branch strongly diverging, curved,not extending exsagittally beyond palpebral lobes.Preglabellar field in front of glabella slightly widerthan anterior border. Anterior border faintly convex,rather flat, not continuing the general convexity of thecephalon, slightly wider exsagittally. Posterior borderslightly widens distally. Posterior border furrow distallydeeper and slightly wider, running slightly obliquely toaxis.
Measurements. The holotype cranidium (INGPANWB11) has a length of 7 mm and a width acrossthe mid-length of the palpebral lobes of 10 mm. Thecranidial length varies between 2.7 and 15 mm, and thecranidial width across the mid-length of the palpebrallobes varies between 3 and 19 mm (n = 50).
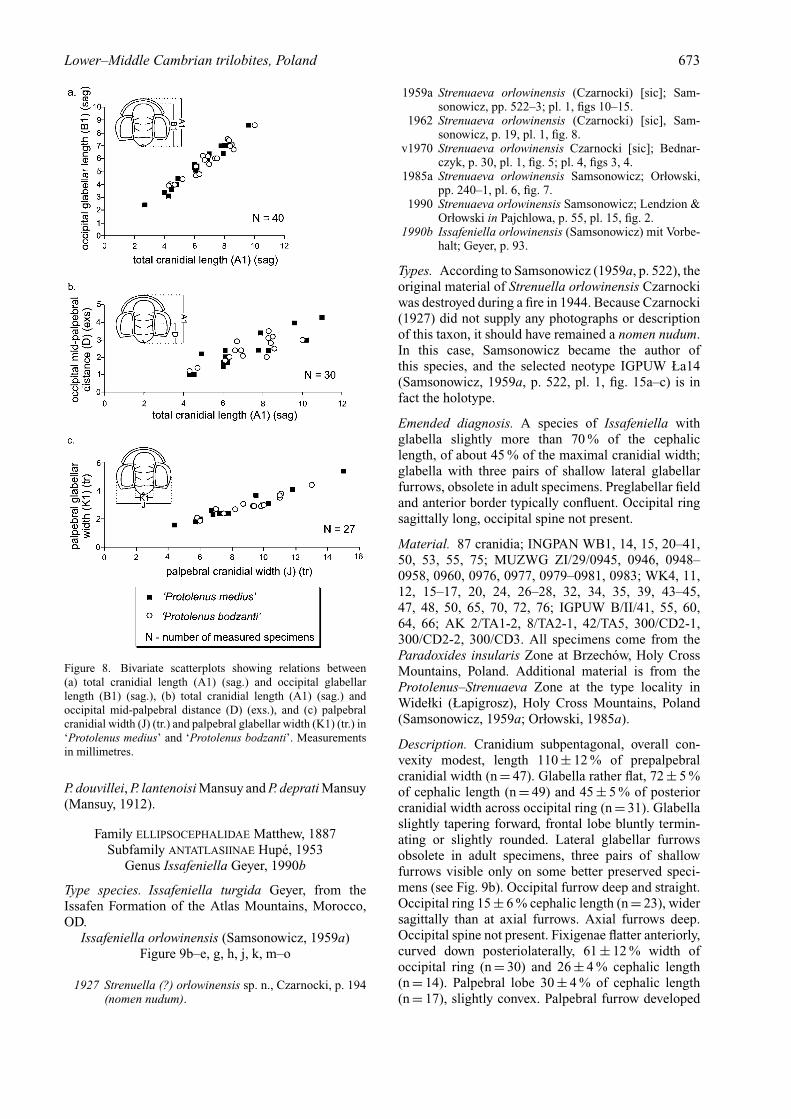
Remarks. The specimens assigned here to Palaeolenusmedius have been considered by Bednarczyk (1970) torepresent two separate species: Protolenus medius andProtolenus bodzanti. Geyer & Landing (2004) com-mented on the Brzechow assemblage and suggestedthe palaeolenid affinity of bodzanti and medius as wellas their assignment to a single species. Analysis ofa larger collection of specimens has confirmed thesesuggestions, and shown that the differences pointedout by Bednarczyk (1970) are intraspecific in character,with forms transitional between medius and bodzantipresent in the studied collection. Moreover, bivariatescatterplots performed for specimens of the two allegedspecies (Fig. 8) indicate that they are conspecific.
Protolenus bodzanti as applied by Czarnocki (1927)is a nomen nudum. The confusion was caused whenBednarczyk (1970) decided to use the same specificname for specimens from his topotype collection withCzarnocki as the species creator. Since 1970 (Orłowski,1988; Bednarczyk & Orłowski in Pajchlowa, 1990),Bednarczyk is considered the author of both species,P. bodzanti and P. medius. In order to resolve thisconfusion, we have decided to abandon bodzanti andretain medius as the specific name, with Bednarczyk asthe creator. In this case, specimens formerly assigned tobodzanti fall into the intraspecific variability of medius,and bodzanti becomes a junior synonym of medius.
Palaeolenus medius differs from other species of thisgenus (Palaeolenus douvillei Mansuy: Mansuy, 1912or Palaeolenus antiquus (Chernysheva): Rushton &Powell, 1998) in its almost parallel-sided posterior partof the glabella and well-rounded frontal lobe expandingforwards from S2, whereas in P. antiquus the frontallobe expands already from S1 (e.g. Chernysheva, 1961,pl. 3, figs 2, 6–10; Rushton & Powell, 1998, figs 29a,34b, 37), and in P. douvillei the widening of the frontallobe is not as distinct (e.g. Mansuy, 1912, pl. 4, fig. 1).P. medius also has a slightly wider (tr.) cranidium atlevel of palpebral lobes in comparison to P. antiquus,and in this respect approaches the Chinese palaeolenids
672 A. ZYLINSKA & M. MASIAK
Figure 7. Trilobites from Brzechow. (a–q) cranidia of Palaeolenus medius (Bednarczyk). (a) WK21; (b) counterpart of (a); (c) INGPANWB6, original of Bednarczyk (1970), pl. 3, fig. 2 (cotype of ‘Protolenus bodzanti’); (d) WK17; (e) WK61; (f) IGPUW B/II/43;(g) WK2; (h) MUZWG ZI/29/0941; (i) MUZWG ZI/29/0980; (j) INGPAN WB13, original of Bednarczyk (1970), pl. 3, fig. 4;(k) WK5; (l) WK37; (m) INGPAN WB3, original of Bednarczyk (1970), pl. 4, fig. 1; (n) INGPAN WB2, original of Bednarczyk(1970), pl. 3, fig. 1; (o) WK24; (p) WK29; (q) INGPAN WB5. Scale bars equal 0.5 cm.
Lower–Middle Cambrian trilobites, Poland 673
Figure 8. Bivariate scatterplots showing relations between(a) total cranidial length (A1) (sag.) and occipital glabellarlength (B1) (sag.), (b) total cranidial length (A1) (sag.) andoccipital mid-palpebral distance (D) (exs.), and (c) palpebralcranidial width (J) (tr.) and palpebral glabellar width (K1) (tr.) in‘Protolenus medius’ and ‘Protolenus bodzanti’. Measurementsin millimetres.
P. douvillei, P. lantenoisi Mansuy and P. deprati Mansuy(Mansuy, 1912).
Family ELLIPSOCEPHALIDAE Matthew, 1887Subfamily ANTATLASIINAE Hupe, 1953
Genus Issafeniella Geyer, 1990b
Type species. Issafeniella turgida Geyer, from theIssafen Formation of the Atlas Mountains, Morocco,OD.
Issafeniella orlowinensis (Samsonowicz, 1959a)Figure 9b–e, g, h, j, k, m–o
1927 Strenuella (?) orlowinensis sp. n., Czarnocki, p. 194(nomen nudum).
1959a Strenuaeva orlowinensis (Czarnocki) [sic]; Sam-sonowicz, pp. 522–3; pl. 1, figs 10–15.
1962 Strenuaeva orlowinensis (Czarnocki) [sic], Sam-sonowicz, p. 19, pl. 1, fig. 8.
v1970 Strenuaeva orlowinensis Czarnocki [sic]; Bednar-czyk, p. 30, pl. 1, fig. 5; pl. 4, figs 3, 4.
1985a Strenuaeva orlowinensis Samsonowicz; Orłowski,pp. 240–1, pl. 6, fig. 7.
1990 Strenuaeva orlowinensis Samsonowicz; Lendzion &Orłowski in Pajchlowa, p. 55, pl. 15, fig. 2.
1990b Issafeniella orlowinensis (Samsonowicz) mit Vorbe-halt; Geyer, p. 93.
Types. According to Samsonowicz (1959a, p. 522), theoriginal material of Strenuella orlowinensis Czarnockiwas destroyed during a fire in 1944. Because Czarnocki(1927) did not supply any photographs or descriptionof this taxon, it should have remained a nomen nudum.In this case, Samsonowicz became the author ofthis species, and the selected neotype IGPUW Ła14(Samsonowicz, 1959a, p. 522, pl. 1, fig. 15a–c) is infact the holotype.
Emended diagnosis. A species of Issafeniella withglabella slightly more than 70 % of the cephaliclength, of about 45 % of the maximal cranidial width;glabella with three pairs of shallow lateral glabellarfurrows, obsolete in adult specimens. Preglabellar fieldand anterior border typically confluent. Occipital ringsagittally long, occipital spine not present.
Material. 87 cranidia; INGPAN WB1, 14, 15, 20–41,50, 53, 55, 75; MUZWG ZI/29/0945, 0946, 0948–0958, 0960, 0976, 0977, 0979–0981, 0983; WK4, 11,12, 15–17, 20, 24, 26–28, 32, 34, 35, 39, 43–45,47, 48, 50, 65, 70, 72, 76; IGPUW B/II/41, 55, 60,64, 66; AK 2/TA1-2, 8/TA2-1, 42/TA5, 300/CD2-1,300/CD2-2, 300/CD3. All specimens come from theParadoxides insularis Zone at Brzechow, Holy CrossMountains, Poland. Additional material is from theProtolenus–Strenuaeva Zone at the type locality inWidełki (Łapigrosz), Holy Cross Mountains, Poland(Samsonowicz, 1959a; Orłowski, 1985a).
Description. Cranidium subpentagonal, overall con-vexity modest, length 110 ± 12 % of prepalpebralcranidial width (n = 47). Glabella rather flat, 72 ± 5 %of cephalic length (n = 49) and 45 ± 5 % of posteriorcranidial width across occipital ring (n = 31). Glabellaslightly tapering forward, frontal lobe bluntly termin-ating or slightly rounded. Lateral glabellar furrowsobsolete in adult specimens, three pairs of shallowfurrows visible only on some better preserved speci-mens (see Fig. 9b). Occipital furrow deep and straight.Occipital ring 15 ± 6 % cephalic length (n = 23), widersagittally than at axial furrows. Axial furrows deep.Occipital spine not present. Fixigenae flatter anteriorly,curved down posteriolaterally, 61 ± 12 % width ofoccipital ring (n = 30) and 26 ± 4 % cephalic length(n = 14). Palpebral lobe 30 ± 4 % of cephalic length(n = 17), slightly convex. Palpebral furrow developed
674 A. ZYLINSKA & M. MASIAK
Figure 9. Trilobites from Brzechow. (a, f, i, l) cranidia of Kingaspidoides sanctacrucensis (Czarnocki). (a) INGPAN WB49;(f) side view of (a); (i) WK38; (l) MUZWG ZI/29/0968. (b–e, g, h, j, k, m–o) cranidia of Issafeniella orlowinensis (Samsonowicz).(b) INGPAN WB21; (c) INGPAN WB25; (d) INGPAN WB41, original of Bednarczyk (1970), pl. 1, fig. 5; (e) INGPAN WB39, originalof Bednarczyk (1970), pl. 4, fig. 3; (g) INGPAN WB40, original of Bednarczyk, pl. 4, fig. 4; (h) INGPAN WB26; (j) INGPAN WB34;(k) MUZWG ZI/29/0960; (m) MUZWG ZI/29/0976; (n) WK44, with Palaeolenus medius (Bednarczyk); (o) INGPAN WB29. Scalebars equal 0.5 cm.

Lower–Middle Cambrian trilobites, Poland 675
as a shallow, indistinct depression (see Fig. 9j).Anterior branch of facial suture short, only slightlydivergent from palpebral lobes to frontal margin, thencurving sharply adaxially to give the anterior bordera sub-triangular shape. Posterior branch diverging,short, extending slightly distally to palpebral lobes.Frontal area convex, 28 ± 6 % of cephalic length(n = 49). Preglabellar furrow almost straight, ratherdeep. Posterior border furrow distally deeper.
Measurements. The cranidial length varies between2.8 and 6.5 mm, and the pre-palpebral cranidial widthvaries between 2.3 and 5.9 mm (n = 49). The largestillustrated cranidium of this species (Samsonowicz,1959a, p. 522, pl. 1, fig. 15) is also the holotype (seeabove) and is 10.5 mm long and 12.5 mm wide.
Remarks. Although the species is insufficiently known(only isolated cranidia have been found so far), itrepresents Issafeniella Geyer rather than StrenuaevaRichter & Richter in cranidial proportions. Thespecimens at hand are not very well preserved, whichis the reason why some cranidial characters are notclear. For example, the preglabellar field is apparentlyconfluent with the anterior border in the specimens athand, whereas in the type collection from Widełki, theborder furrow is very shallow but distinct (see e.g. pl. 1,figs 10, 14 in Samsonowicz, 1959a). Likewise thelateral glabellar furrows are more evident in thespecimens from Widełki than in the specimens at hand.Geyer & Landing (2004) have pointed out the possiblesimilarity of Strenuaeva orlowinensis to Comluellasp. from the southern Negev in Israel (Parnes, 1971);the original paper, however, illustrates only Strenuella(Strenuella) sp. and Strenuella (Comluella) nevoin.sp. In both cases the specimens differ from thespecimens in hand in having a much more taperingglabella, distinct eye-ridges and less pronounced andtransversely narrower preglabellar field; moreover, theyseem to be distinct from Issafeniella.
An evident feature in Issafeniella orlowinensis isthe lack of an occipital spine. In this respect it isdistinct from other species of Issafeniella Geyer andrecalls Strenuaeva Richter & Richter, although worthmentioning is the fact that the occipital spines are notvisible on internal moulds of, for example, I. turgidaGeyer (Geyer, 1990b, pl. 12, figs 3, 4), I. sampelayoi(Richter & Richter) (Richter & Richter, 1940, pl. 3,figs 51–53) and I. melendezi (Gil Cid) (Gil Cid, 1972,pl. 1, figs 1–6).
Subfamily ELLIPSOCEPHALINAE Matthew, 1887Genus Kingaspidoides Hupe, 1953
Type species. Kingaspis (Kingaspidoides) armatus[sic] Hupe, a subjective synonym of Kingaspis (Kin-gaspis) brevifrons Hupe from the Breche a Micmacca
facies of the lower Jbel Wawrmast Formation ofMorocco (cf. Geyer, 1990b).
Kingaspidoides sanctacrucensis (Czarnocki, 1927)Figure 9a, f, i, l
1927 Ellipsocephalus S-ta Crucensis sp.n., Czarnocki,pp. 193–4.
v1959b Germaropyge sancta-crucensis (Czarnocki), Sam-sonowicz, pp. 527, 529, pl. 2, figs 4–12.
v1970 Germaropyge sancta-crucensis Czarnocki, Bednar-czyk, p. 30, pl. 1, figs 2, 4–6.
1973 Germaropyge cf. sanctacrucensis (Czarnocki),Orłowski, p. 369, pl. 1, fig. 3.
v1975a Ellipsocephalus sanctacrucensis (Samsonowicz)[sic], Orłowski, pp. 371–4, pl. 1, figs 1–7, pl. 2,figs 1–10, pl. 3, figs 1–8, pl. 4, fig. 1.
v1985a Ellipsocephalus sanctacrucensis (Samsonowicz)[sic], Orłowski, p. 238, pl. 7, fig. 4a, b.
v1990 Ellipsocephalus sanctacrucensis (Samsonowicz)[sic], Lendzion & Orłowski in Pajchlowa, p. 54,pl. 14, fig. 9.
Types. The neotype is cranidium IGPUW Ła 1,selected and illustrated by Samsonowicz (1959b, pl. 2,fig. 4a–c) from Łapigrosz near Widełki, Holy CrossMountains. This cranidium has also been illustrated byOrłowski (1975a, pl. 3, fig. 4), however, without anyannotation about its identity. Additionally, Orłowskiselected a dorsal skeleton as a lectotype (Orłowski,1975a, p. 371, pl. 1, fig. 1a, b and Lendzion & Orłowskiin Pajchlowa, 1990, p. 54, pl. 14, fig. 9). This selectionis considered erroneous.
Emended diagnosis. Species of Kingaspidoides withglabella clearly elevated with respect to the preglabellarfield and fixigenae; glabella 80–82 % of cranidiallength and about 50 % of cranidial width acrosspalpebral lobes, with faint median ridge; occipitalring without spine; fixigenae about 60 % of maximumglabellar width; exsaggital length of palpebral lobesapproximately 24 % of the cranidial length; anteriorborder and preglabellar field obsolescent.
Material. 33 cranidia, INGPAN WB49, 56, 62;MUZWG ZI/29/0968–0970, 0982, 0983, 0986; WK4–6, 8, 10, 11, 14, 17–19, 22, 23, 38, 41, 42, 49,52–56, 64; IGPUW B/II/62, 67; in some caseswith counterparts; 3 librigenae, MUZWG ZI/29/0971,0987; WK68. The studied specimens come fromthe Paradoxides insularis Zone at Brzechow, HolyCross Mountains, Poland. Most specimens of thistaxon illustrated by Bednarczyk (1970, pl. 1, figs 3,4, 6) are missing and were not available for study.Additional material from the Holy Cross Mountains,including complete specimens, has been noted in theProtolenus–Strenuaeva Zone in Widełki (Łapigrosz)(Samsonowicz, 1959b; Orłowski, 1975a, 1985a), theParadoxides insularis Zone in Helenow (Orłowski,1964a), and the much younger strata (correspondingto the Caesaraugustian or even Languedocian) in the

676 A. ZYLINSKA & M. MASIAK
Pieprzowe Mountains (see Orłowski, 1964a, table 1,p. 42), although the specimen has not been illustratedor discussed in the text. A single cranidium representingthis species from the coarse sandstones representing theParadoxides polonicus Zone on Słowiec Hill (SłowiecFormation) has been found in Orłowski’s collection.
Description. Cranidium subrectangular, overall con-vexity distinct; length 85–99 % of palpebral cranidialwidth. Glabella distinctly elevated above the fixigenae,82 ± 2 % of cephalic length (n = 8) and 46–53 % ofposterior cranidial width across occipital ring. Glabellaalmost parallel-sided, frontal lobe bluntly pointed orslightly rounded anteriorly. Lateral glabellar furrowseffaced. Occipital furrow shallow, straight, not reachingthe axial furrows. Occipital ring 11 ± 2 % of cephaliclength (n = 8), sagittally distinctly wider than at axialfurrows. Axial furrows rather shallow. Occipital spinenot developed. Fixigenae flatter anteriorly, 55 ± 6 %width of occipital ring (n = 8) and 25–31 % cephaliclength. Palpebral lobes 24 ± 3 % of cephalic length(n = 5), slightly convex. Palpebral furrow developedas a shallow and broad, poorly defined depression.Anterior branch of facial suture slightly divergent frompalpebral lobes to frontal margin, then curving sharplyadaxially. Posterior branch only slightly divergent, veryshort. Frontal area 18–20 % of cephalic length, entire,without border furrow. Preglabellar furrow crescent-shaped, shallow. Posterior furrow deeper distally.
Librigena flat, with stout genal spine only slightlydivergent from lateral margin. Inner spine angle closeto a right angle. Border wide, flat, only barely distinctfrom genal field. Ocular area slightly elevated.
Measurements. The cranidial length varies between12.8 and 20.3 mm (n = 8) and the pre-palpebral cran-idial width varies between 14.8 and 17.8 mm (n = 3).The holotype cranidium (Samsonowicz, 1959b, pl. 2,fig. 4a) is 26 mm long and 25 mm wide. The largestillustrated cranidium of this species (Orłowski, 1975a,pl. 3, fig. 8) is 43 mm long and 35 mm wide.
Remarks. The discussed species is known from isol-ated parts of the exoskeleton as well as completespecimens (see Orłowski, 1975a, pl. 4, fig. 1).Ontogenic analysis on specimens from the type localityin Widełki (Łapigrosz) has shown that in youngerindividuals the glabella is more convex and bettermarked, whereas in the older specimens it is lessconvex, and the glabellar outline is effaced (Orłowski,1975a). Moreover, younger individuals seem to havea narrower (sag.) cranidial frontal area in comparisonwith the older ones (cf. Orłowski, 1975a, pl. 3). Possibletectonic compression has been suggested by Sam-sonowicz (1959b) to explain the fact that some of thecranidia are more swollen than others. On the contrary,the studied specimens do not display effacement ofcranidial outline in larger specimens, as is particularly
well visible in specimen INGPAN WB49 (Fig. 9a, f).The reason for this may lie in a different susceptibilityto compaction of the fine-grained sandstones fromWidełki (Łapigrosz) compared with the coarse-grainedBrzechow sandstones.
Assignment of this species to Kingaspidoides Hupeis justified by cranidial features which include: distinctextensions of the anterolateral corners of the glabellaconnected with eye ridges, the bar-like elevation ofglabella above the fixigenae and the clearly convex andrather wide (sag.) preglabellar area. K. sanctacrucensisshows features characteristic of many other membersof this genus, that is, rather small palpebral lobesresembling those of K. obliquoculatus Geyer (Geyer,1990b, pl. 18, figs 1, 2, 4, 5, 7, 8, 10, 11), in which,however, the cranidium at level of palpebral lobes iswider (tr.); sub-triangular anterior part of frontal lobein some specimens as in K. neglectus Geyer (Geyer,1990b, pl. 22, figs 1, 2, 6), from which it differs innarrower (tr.) frontal area, smaller palpebral lobes andless distinct eye-ridges; and librigenae (see Orłowski,1975a, pl. 2, figs 5a, 6) passing into stout genal spine,similarly to Kingaspidoides borjensis Geyer (Geyer,1990b, pl. 23, fig. 5), in which, however, the cran-idium is wider (tr.) and the preglabellar furrow lesspronounced.
Family uncertainGenus Myopsolenites Geyer & Landing, 2004
Type species. Myopsolenus palmeri Parnes, 1971 fromthe Timna Formation, Timna area, southern Israel, OD.
Remarks. For family assignment of this genus seediscussion in Geyer & Landing (2004).
The availability of this generic name is notstraightforward. The name appeared for the first timein Opik (1975) in the combination Myopsolenitespalmeri in a footnote to Figure 3, and apparentlywas a misprint as shown by a corrigendum printedon the page facing the title page in that paper. Assuch it was considered by Rushton & Powell (1998),who also assigned Myopsolenus palmeri Parnes toOnaraspis Opik, and showed that the differencesbetween the two taxa are rather of specific significance.Geyer (1990b) pointed that Myopsolenus palmeri isclearly distinct from other myopsolenines and possiblybelongs to Bathynotidae; moreover, he recalled thatthe specimens assigned to this species have beendescribed by Opik (1975) as representing a separategenus Myopsolenites. Accordingly, Geyer & Landing(2004) treated Myopsolenites as a valid taxon created byOpik (1975), although they did not supply a diagnosisseparating this genus from Onaraspis. In the view ofthe International Code of Zoological Nomenclature(Ride et al. 1999), neither Opik (1975) nor Geyer(1990b) have made Myopsolenites an available name. It

Lower–Middle Cambrian trilobites, Poland 677
seems that the thorough discussion of the type materialof Myopsolenus palmeri in Geyer & Landing (2004)is sufficient to make the Myopsolenites available (A.Rushton, pers. comm. 2006), although, as suggestedabove, the diagnosis lacks clear distinction from On-araspis. Following this, the authors of Myopsolenitesare Geyer & Landing, 2004 and not Opik, 1975, andthis view is presented herein.
Myopsolenites kielcensis (Bednarczyk, 1970)Figure 10a–h
v1970 ?Jakutus kielcensis n.sp.; Bednarczyk, pp. 32–4, pl.2, figs 1–3.
v1990 ?Jakutus kielcensis Bednarczyk; Bednarczyk inPajchlowa, p. 62, pl. 18, figs 5, 6.
2004 Myopsolenites kielciensis [sic] (Bednarczyk); Geyer& Landing, pp. 200–1.
Types. The holotype is cranidium INGPAN WB44,illustrated in Bednarczyk (1970), pl. 2, fig. 3 asBrz51 (Fig. 10c herein) and pygidium INGPAN WB42,illustrated in Bednarczyk (1970), pl. 2, fig. 2 as Brz49(Fig. 10e herein) (also illustrated in Bednarczyk inPajchlowa, 1990, pl. 18, figs 5, 6); from Brzechow,Holy Cross Mountains, Poland.
Emended diagnosis. Species of Myopsolenites withmoderately tapering glabella and slightly curved axialfurrows; well-developed, transglabellar S1 and truncatefront; relatively short palpebral lobes; pygidium sub-triangular (heart-shaped) in outline.
Material. 8 cranidia: INGPAN WB43–45; MUZWGZI/29/0966, 0974, 0985; WK66; AK 41/TA1-2; 3pygidia: INGPAN WB42; MUZWG ZI/29/0967, 0973.All specimens are from the Paradoxides insularis Zoneat Brzechow, Holy Cross Mountains, Poland.
Description. Medium- to large-sized trilobites. Onlycranidia and pygidia are known. Cranidium trans-versely trapezoidal, rather flat, length 71–76 % widthacross centre of palpebral lobes. Glabella slightlyconvex, length in adult specimens approximately 87 %cephalic length and about half the width of thecranidium across occipital ring; continuously taperingforward. Frontal lobe sagittally short, subtruncate.Three pairs of shallow lateral glabellar furrowsdeveloped, all commencing at axial furrows. S1transglabellar, both on adult and juvenile specimens,directed backwards. S2 slightly directed backwards,separated medially on glabella in adult specimensand transglabellar in the juvenile specimen (INGPANWB45, Fig. 10f). S3 much shorter, slightly directedbackwards, not visible on juvenile specimen. Occipitalring defined only in juvenile specimen, approximately22 % of cephalic length, posterior rim gently curved,with subterminal occipital node. Occipital furrowdistinct with deeper lateral sections. Axial furrows
wide and rather deep. Fixigenae moderately convex,subtriangular, the exsagittal occipital mid-palpebraldistance is 45–46 % maximum cephalic length andthe transverse width of fixigenae is 27–29 % ofmaximum cephalic width across centre of palpebrallobes. Posterolateral projections of fixigenae smalland narrow. Palpebral lobes slightly convex and withslight curvature, 46–53 % maximum cephalic length,oblique to exsagittal axis, posterior tips distant fromposterior border furrow. Palpebral furrow narrower(tr.) than palpebral lobes, shallow, well defined. Eyeridges oblique to axes, directed approximately 45◦
anteriorly from anterior tips of palpebral lobes, lessdistinct in larger individuals. Eye ridges slightly curvedforward adaxially, reach glabella slightly behind itsanterolateral corners. Anterior branch of facial sutureshort, slightly diverging towards anterior margin, thencurved sharply adaxially towards anterolateral margin.Posterior branches strongly divergent, curving gentlyfrom posterior tips of palpebral lobes. Frontal area 12–15 % of total cranidial length, approximately 7 % injuvenile specimen. Preglabellar field narrow, forminga distinct depression between glabella and anteriorborder furrow. Anterior border more convex in smallerindividuals, much lower in large individuals. Anteriorborder furrow indistinct. Posterior border moderatelywide and convex, adaxial section slightly obliqueto axis, with small socket approximately midway offixigenal width. Posterior border furrow wide close toaxial furrows, shallows outwards and does not reachlateral margin.
Thorax unknown.Pygidium large, sub-triangular in outline; length
from articulating furrow to posterior margin 62–77 % maximum width (tr.). Lateral margins smooth.Articulating half-ring narrow (tr.). Axis 92–95 % ofpygidial length, with four relatively distinct rings inthe adult specimens, followed by a large terminal axialpiece. Pleural areas fused, with two or three weaklydefined interpleural furrows recognizable. Border lowand narrow, border furrow indistinct and defined assubtle change in convexity.
Surface of test covered with low, densely spacedgranules (e.g. MUZWG ZI/29/0974).
Measurements. The holotype cranidium (INGPANWB44) has a length of about 26 mm and a width acrossthe mid-length of the palpebral lobes of about 42 mm.The smallest cranidium (INGPAN WB45), a juvenilespecimen, is 6.8 mm long and its width across the mid-length of the palpebral lobes is 9 mm. The holotypepygidium (INGPAN WB42) is about 12 mm longand about 15.5 mm wide. The second well-preservedpygidium (MUZWG ZI/29/0967) is about 13 mm longand almost 21 mm wide.
Remarks. The genus, its species and their stratigraphicposition have been recently described in detail by
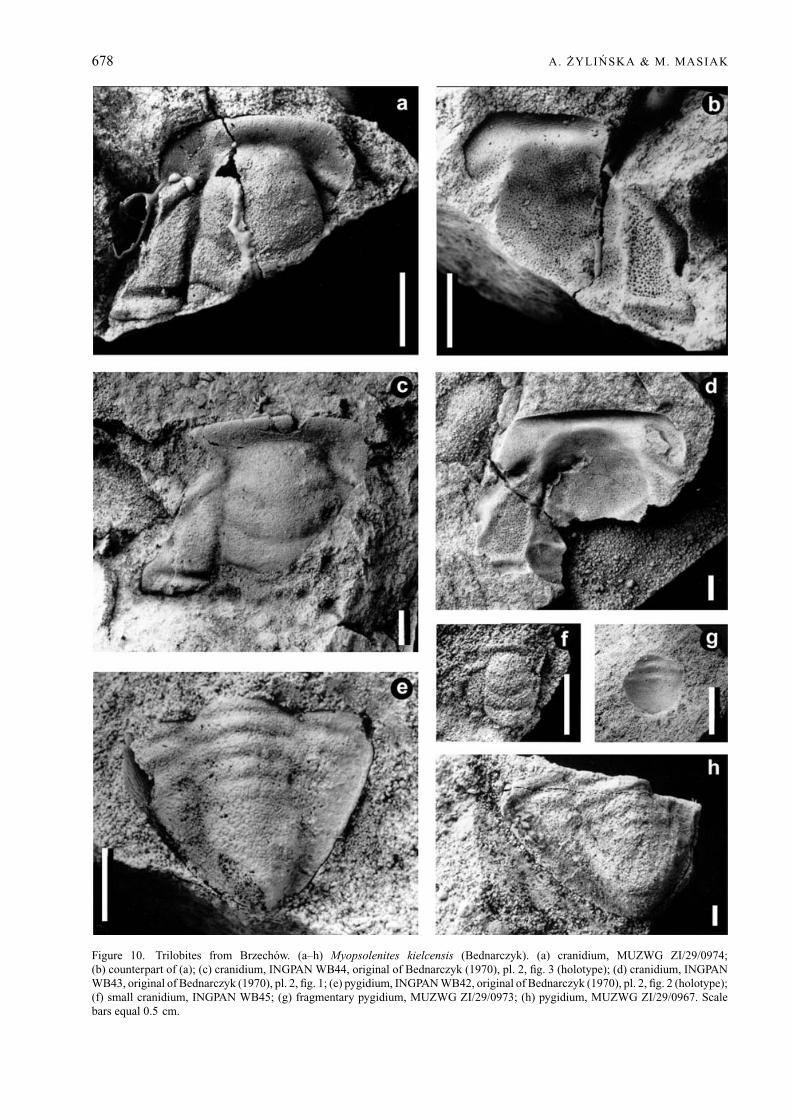
678 A. ZYLINSKA & M. MASIAK
Figure 10. Trilobites from Brzechow. (a–h) Myopsolenites kielcensis (Bednarczyk). (a) cranidium, MUZWG ZI/29/0974;(b) counterpart of (a); (c) cranidium, INGPAN WB44, original of Bednarczyk (1970), pl. 2, fig. 3 (holotype); (d) cranidium, INGPANWB43, original of Bednarczyk (1970), pl. 2, fig. 1; (e) pygidium, INGPAN WB42, original of Bednarczyk (1970), pl. 2, fig. 2 (holotype);(f) small cranidium, INGPAN WB45; (g) fragmentary pygidium, MUZWG ZI/29/0973; (h) pygidium, MUZWG ZI/29/0967. Scalebars equal 0.5 cm.

Lower–Middle Cambrian trilobites, Poland 679
Geyer & Landing (2004). Despite large variabilitythe species can be clearly distinguished from eachother (Geyer & Landing, 2004, table 1). Other possiblemembers of this or a closely related genus may includeConomicmacca plana (Matthew) from the Protolenuselegans Zone of southern New Brunswick (Westrop &Landing, 2000), Bathynotellus yermolaevi Lerm. (MS)from the Middle Cambrian succession of NovayaZemlya (Lermontova, 1940), as well as Rectifrontinellaolhae Konstantinenko from the Carpathian Foredeep inthe Ukraine (Drygant & Konstantinenko, 2001).
Superfamily PARADOXIDOIDEA Hawle & Corda, 1847Family PARADOXIDIDAE Hawle & Corda, 1847
Genus Paradoxides Brongniart, 1822
Type species. Entomostracites paradoxissimus Wah-lenberg from the Middle Cambrian Triplagnostusgibbus Zone at Oltorp, Vastergotland, Sweden, bysubsequent designation (I.C.Z.N. 1957).
Remarks. The need for a comprehensive revision ofthe Paradoxides clade has been expressed by manyauthors. The main reason for this is that the differentideas for subdividing the members of this genus (e.g.Snajdr, 1957, 1958; Solov’ev, 1969; Dean & Rushtonin Kaesler, 1997) have been based on the ontogenicsimilarities of the protaspid and meraspid forms, dueto the fact that in holaspids the distinctions become lesspronounced because of morphological convergence.In reality, however, a great many species have beenbased on forms preserved only as holaspids, withoutknowledge of earlier ontogeny, and, moreover, thediagnostic features proposed for genera and subgenerahave been differently applied by various authors.This has led to establishing more than 140 speciesand subspecies of Paradoxides sensu lato (Geyer &Landing, 2001).
Generally, a pragmatic view persists (followingSdzuy, 1968, 1970 and Solov’ev, 1969, 1980), torecognize and apply Snajdr’s (1957, 1958) genera:Paradoxides Brongniart, Hydrocephalus Barrande, Ec-caparadoxides Snajdr and Acadoparadoxides Snajdr.This approach has been recently expressed by, forexample, Dean & Ozgul (1994), Dean & Rushtonin Kaesler (1997), Kim, Westrop & Landing (2002)and Dean (2005), and in modified form (the generaare considered as subgenera of Paradoxides s. l.)by, for example, Linan & Gozalo (1986), Geyer(1998) or Geyer & Landing (2001). An attempt tosort out problems linked with paradoxidid classific-ation has been recently presented by Fletcher et al.(2005), who emphasized the importance of cephalicmorphology in this classification, and following this,recognized the following subgenera of Paradoxides s.l.: Paradoxides Brongniart, Eccaparadoxides Snajdr,Hydrocephalus Barrande and Plutonides Hicks. Wefollow Geyer (1998) and Geyer & Landing (2001) inaccepting Snajdr’s (1957, 1958) genera as subgenera of
Paradoxides s. l. and do not try to resolve the problemswith the subdivision of the clade in this paper.
Subgenus Acadoparadoxides Snajdr, 1957
Type species. Paradoxides sacheri Barrande, 1852, OD.
Remarks. The paradoxidids studied herein belong ingeneral to the ‘oelandicus group’, assigned since Sdzuy(1957, 1968) to Acadoparadoxides. This approach isfollowed here. An emended diagnosis of Acadopara-doxides Snajdr has been recently presented in Geyer(1993), although accordingly the group is not supposedto include forms with spines on the pygidial posteriormargin, thus excluding P. (A.) oelandicus itself. P.(A.) oelandicus has been referred to Acadoparadoxides(Baltoparadoxides) by Dean & Rushton in Kaesler(1997). In Fletcher et al. (2005), AcadoparadoxidesSnajdr and Baltoparadoxides Snajdr are considered asjunior synonyms of Plutonides Hicks.
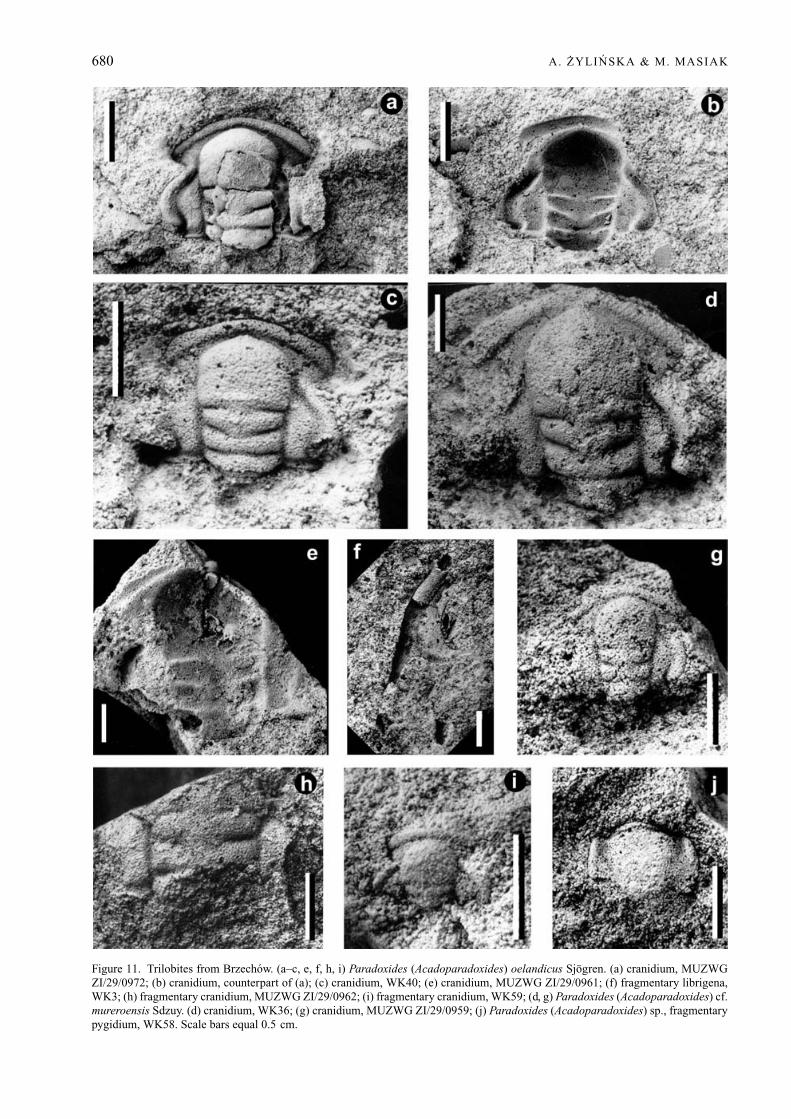
Paradoxides (Acadoparadoxides) cf. mureroensis(Sdzuy, 1958)Figure 11d, g
?1959 Paradoxides oelandicus Sjogren, Orłowski, pp. 441–2, pl. 1, figs 1–4.
Types of Paradoxides (Acadoparadoxides) mureroensisSdzuy. The holotype is cranidium X 1370c housed inthe Senckenberg Museum, Frankfurt am Main, fromBarranco de Valdemiedes, Northern Murero, Spain,illustrated in Sdzuy (1958, pl. 1, fig. 12). An imageof the holotype, after retrodeformation in Photoshop,has been recently presented in Fletcher et al. (2005,fig. 12).
Material. Two cranidia: MUZWG ZI/29/0959 andWK36 from the Paradoxides insularis Zone atBrzechow, Holy Cross Mountains, Poland. Additionalcranidia are probably also present in the UsarzowFormation (Orłowski, 1959).
Description. Cranidium with evenly curved anteriorborder. Glabella pyriform, expanding forward to about1.3 times width across occipital ring. Posterior sectionwith faintly diverging sides. Frontal lobe stronglyconvex in transverse profile, glabellar front slightlyarcuate. S1 and S2 well impressed, connected althoughmuch shallower over the axis. S1 more stronglyindented, clearly anteriorly directed at the axialfurrows; S2 less strongly indented, slightly curvedbackwards medially. S3 very poorly marked, short,visible only in specimen WK36 (Fig. 11d), directedobliquely forward. Occipital furrow rather broad andwell impressed, slightly shallower medially. Occipitalring slightly convex. Posterior margin of occipital ringmoderately curved, projecting well beyond posteriorborder. Axial furrows shallow, narrow, deepest close toL2. Fixigenae narrow, about 18 % width of cranidium
680 A. ZYLINSKA & M. MASIAK
Figure 11. Trilobites from Brzechow. (a–c, e, f, h, i) Paradoxides (Acadoparadoxides) oelandicus Sjogren. (a) cranidium, MUZWGZI/29/0972; (b) cranidium, counterpart of (a); (c) cranidium, WK40; (e) cranidium, MUZWG ZI/29/0961; (f) fragmentary librigena,WK3; (h) fragmentary cranidium, MUZWG ZI/29/0962; (i) fragmentary cranidium, WK59; (d, g) Paradoxides (Acadoparadoxides) cf.mureroensis Sdzuy. (d) cranidium, WK36; (g) cranidium, MUZWG ZI/29/0959; (j) Paradoxides (Acadoparadoxides) sp., fragmentarypygidium, WK58. Scale bars equal 0.5 cm.

Lower–Middle Cambrian trilobites, Poland 681
across palpebral lobes. Palpebral lobe relatively long,exsagittally almost 50 % cephalic length, posterior endalmost reaching posterior border, moderately wide(about 10 % of cranidial width), sausage-shaped andrather flat. Palpebral furrow not discernible. Anteriorborder well developed, distinctly convex, sagittallyrather narrow in front of the glabella (less than 10 %of cephalic width), expanding to almost double itswidth exsagittally. Anterior border furrow merged withpreglabellar area forming a narrow depression, whichis only slightly wider than axial furrows. Posteriorborder rather narrow and convex, otherwise verypoorly preserved in the analysed specimens. Facialsuture with strongly diverging anterior branch, stronglycurved toward axis when reaching anterior border.Anterolateral corner of cranidium projecting wellbeyond distal margin of palpebral lobe.
Measurements. The largest cranidium in the collection(WK36) has a length of 16.7 mm and a width acrossthe mid-length of the palpebral lobes of 17.3 mm. Thesecond, smaller cranidium (ZI/29/0959) is 7.8 mm longand its width across the mid-length of the palpebrallobes is 8.6 mm. The holotype cranidium of P. (A.)mureroensis is 36 mm long (Sdzuy, 1958).
Remarks. The two specimens assigned herein to P.(A.) mureroensis can be distinguished from the otherparadoxidid cranidia in the collection, which areassigned to P. (A.) oelandicus Sjogren (see below).The differences, however, are quite small, but include:the angulate outline of the anterior margin, slightlynarrower fixigenae at level of palpebral lobes, narrowerglabella at mid-length of palpebral lobes, more roundedanterior lobe, and lateral glabellar furrows bentbackwards medially. With these features the cranidia athand fall into the characteristics of P. (A.) mureroensisas illustrated by Sdzuy (1961, pl. 16, figs 2–11), Gozaloet al. (1993, pl. 1, figs 3, 5, 7) or Dean (2005, pl. 5,figs a, b, e, g, j, k, n, o). It is worth noting that mostknown specimens of this species are strongly distorted,and the observed large intraspecific variability might bein reality a tectonic effect. As shown by, for example,Sdzuy (1961), Geyer (1998), Geyer & Landing (2001)and Fletcher et al. (2005), the cranidia and hypostomataof this species are barely distinguishable from otherParadoxides species, such as P. (A.) pinus Holm MS,P. (A.) nobilis Geyer, P. (A.) harlani Green sensuGeyer & Landing (2001) or P. (Plutonides) haywardi(Raymond) sensu Fletcher et al. (2005). The lack ofcomplete specimens in the material from Brzechowdoes not allow a confident taxonomic assignment.
Paradoxides (Acadoparadoxides) oelandicusSjogren, 1872
Figure 11a–c, e, f, h, i
1872 Paradoxides Oelandicus sp. n., Sjogren, p. 72,pl. 5, fig. 1.
1877 Paradoxides oelandicus Sjogren, Linnarsson, pp.354–6, pl. 14, figs 1–6 (fig. 1 cop. Sjogren(1872)).
1936 Paradoxides oelandicus Sjogren, Westergard, pp.33–5, 51–2, text-fig. 8, pl. 2, figs 1–11, pl. 3, figs1–6.
1952 Paradoxides oelandicus Sjogren, Hennings-moen, p. 16, pl. 1, fig. 1.
1957 Eccaparadoxides oelandicus (Sjogren), Snajdr,p. 238.
1957 Paradoxides of the olandicus group, Orłowski,pp. 770–1, figs 1–6.
1958 Eccaparadoxides ? oelandicus (Sjogren), Snajdr,p. 115.
non 1959 Paradoxides oelandicus Sjogren, Orłowski, pp.441–2, pl. 1, figs 1–4 (?P. (Acadoparadoxides)cf. mureroensis Sdzuy).
pars 1959 Paradoxides pinus Holm, Orłowski, p. 442, pl. 1,figs 6, 7.
1964a Paradoxides oelandicus Sjogren, Orłowski, pl. 1,figs 1–11.
1970 Eccaparadoxides oelandicus (Sjogren), Tom-czykowa in Dumicz, Tomczykowa & Wojcik, pp.295–7, pl. 12, figs 1–5, pl. 13, figs 1–12.
1970 Paradoxides oelandicus Sjorgen [sic], Bednar-czyk, p. 34, pl. 1, fig. 1.
1973 Paradoxides (Acadoparadoxides) ex gr. oelandi-cus (Sjogren), Schrank, p. 88, pl. 2, figs 1, 2.
1986 Acadoparadoxides (Baltoparadoxides) oelandi-cus Sjogren, Snajdr, p. 174.
1989 Eccaparadoxides oelandicus (Sjogren), Ahlberg,p. 146, figs 8, ?9B.
1990 Eccaparadoxides oelandicus (Sjogren), Bednar-czyk, Lendzion & Orłowski in Pajchlowa, p. 61,pl. 16, fig. 10.
1994 Eccaparadoxides oelandicus (Sjogren),Rudolph, pp. 169–70, pl. 16, figs 1–9.
1997 Acadoparadoxides (Baltoparadoxides) oelandi-cus Sjogren, Dean & Rushton in Kaesler, pp.474–5, fig. 305, 1a, b.
Types. The holotype is a cephalon and thorax, Ar.1449, Palaeontological Department, State Museum inStockholm, illustrated by Sjogren (1872), pl. 5, fig. 1;re-illustrated by Linnarsson (1877, pl. 14, fig. 1) andWestergard (1936, pl. 2, fig. 10); either from Borgholmor from Stora Fro, Oland (see discussion in Westergard,1936, p. 35).
Material. 5 cranidia: MUZWG ZI/29/0961, 0962,0972; WK40, WK59; 5 librigenae: WK3, 24, 51,63; AK 42/TA3’. Specimens of this taxon from thecollection of Wiesław Bednarczyk are missing andwere not available for study. All studied specimens arefrom the Paradoxides insularis Zone at Brzechow, HolyCross Mountains, Poland. Other material from the HolyCross Mountains has been collected in the Paradoxidesinsularis and Paradoxides pinus zones at Jugoszow,Usarzow and Sternalice (Orłowski, 1964a).
Description. Cranidium with evenly rounded frontaloutline. Glabella pyriform, expanding forward toonly about 1.1 times width across occipital ring.Posterior section with sub-parallel sides. Frontal lobe

682 A. ZYLINSKA & M. MASIAK
strongly convex in transverse profile, glabellar frontblunt, sub-triangular. S1 and S2 well impressed,connected although much shallower over the mid-line.S1 oblique backwards; S2 almost transverse. S3 verypoorly marked, short (visible in specimens ZI/29/0972,Fig. 11a, b and WK40, Fig. 11c), directed obliquely for-ward. Occipital furrow rather broad and well impressed,slightly shallower medially. Occipital ring slightlyconvex. Posterior margin of occipital ring moderatelycurved, projecting well behind posterior border. Axialfurrows shallow, narrow, deepest close to L2. Fixigenaemoderately wide, over 20 % width of cranidium acrosspalpebral lobes. Palpebral lobe relatively long, exsagit-tally over 50 % cephalic length, posterior end almostreaching posterior border, moderately wide (about20 % of cranidial width), crescent shaped and slightlyelevated above fixigenae. Palpebral furrow shallowbut distinct. Anterior border well developed, distinctlyconvex, sagittally rather narrow in front of the glabella(slightly more than 10 % of cephalic width), expandingto about 1.5 its width exsagittally. Anterior borderfurrow distinct and shallow; preglabellar area forminga narrow depression. Posterior border very narrow andconvex. Facial suture with strongly diverging anteriorbranch, strongly curved toward axis when reachinganterior border. Anterolateral corner of cranidiumexsagittally almost reaches distal margin of palpebrallobe.
Librigenae with moderately curved lateral border.Genal spine long, slightly curved, longer than anteriorpart of librigena. Base of genal spine rather broad.Border furrow shallow and poorly defined. Posteriorborder straight. Genal angle close to a right angle.Librigenal field relatively wide (tr.).
Measurements. The largest cranidium in the collection(ZI/29/0961) is at least 26 mm long and its width acrossthe mid-length of the palpebral lobes is about 30 mm.The smallest cranidium (WK59) is 4 mm long and itswidth across the mid-length of the palpebral lobes is5.9 mm. The holotype cranidium of P. (A.) oelandicusis 18.4 mm long and 20 mm wide (Westergard, 1936).
Remarks. Specimens of P. (A.) oelandicus from Oland(Westergard, 1936, pls 2, 3) include cranidia which arecharacterized by an evenly rounded anterior glabellarlobe (e.g. Westergard, 1936, pl. 2, figs 2–5, 9, 10, pl. 3,fig. 2) and some in which the anterior lobe is slightlyangular (Westergard, 1936, pl. 2, fig. 6 and pl. 3, fig. 1).Also, the palpebral lobes seem to be less oblique insome of the specimens, both in the morphotypes withthe anterior lobe rounded and subangular. Most ofthe specimens in hand, along with the other specimenfrom Brzechow illustrated by Bednarczyk (1970, pl. 1,fig. 1) seem to represent the morphotype with thesubangular anterior lobe (see Fig. 11a–c), although theother morphotype with the rounded anterior lobe is alsopresent (Fig. 11i). The librigenae assigned herein to thisspecies are not very well preserved, but are similar to
the librigenae assigned to this species by Westergard(1936, pl. 2, figs 4, 7, 9–11) and can be distinguishedfrom those of P. (A.) mureroensis (Sdzuy, 1961, pl. 16,figs 12, 13) in the genal angle close to a right angleand not passing into acute as in P. (A.) mureroensis,narrower lateral border and slightly curved genal spine.
Paradoxides (Acadoparadoxides) sp.Fig. 11j
Material. One pygidium, WK58, from the Paradoxidesinsularis Zone at Brzechow, Holy Cross Mountains,Poland.
Description. Pygidium with posterior part incom-pletely preserved; subsemicircular, with slightly di-verging convex lateral margins, curving gently into amost probably rounded posterior margin. Axis broad,subtriangular, subdivision within axis not visible.Articulating half-ring sagittally narrow, separated fromaxis by shallow furrow.
Remarks. The taxonomic assignment of this specimenis not clear. On the one hand it resembles pygidia ofP. (A.) mureroensis Sdzuy (Sdzuy, 1961, pl. 17, figs 1–5), but its posterior part is not preserved, and thus theassignment to the other paradoxidid in the collection,P. (A.) oelandicus, which has pygidia with four distinctspines on posterior margin (see Westergard, 1936,pl. 2, figs 4, 8, 9, 11, pl. 3, figs 4–6) is also plausible.
Acknowledgements. We are greatly indebted to Prof.Wiesław Bednarczyk (Polish Academy of Sciences) formaking possible the study of his Brzechow collection andmanuscripts, which inspired us to take up this research.Warmest gratitude is expressed to Prof. Stanisław Orłowskiand to Wojciech Kozłowski (University of Warsaw) foraccess to their Brzechow collections. Adrian Kin fromthe ‘Phacops’ Geoscience Friends Association in Łodzkindly supplied his trilobite specimens from Brzechow forthis study. The critical remarks, helpful comments andsuggestions from Gerd Geyer (Wurzburg University) andAdrian Rushton (The Natural History Museum in London),the journal referees, substantially improving the manuscript,are cordially acknowledged. The photographs were madeby Stanisław Kolanowski. We are also grateful to EwaFalkowska (University of Warsaw) for aid during fieldwork.The research was supported by the Institute of Geology,Faculty of Geology, University of Warsaw (individual BWgrant to AZ).
References
AHLBERG, P. 1989. Cambrian stratigraphy of the Nar 1 deepwell, Gotland. Geologiska Foreningens i StockholmForhandlingar 111, 137–48.
ALVARO, J. J. & CLAUSEN, S. 2005. Major geodynamicand sedimentary constraints on the chronostratigraphiccorrelation of the lower–middle Cambrian transition inthe western Mediterranean region. Geosciences Journal9, 145–60.
ALVARO, J. J., ELICKI, O., GEYER, G., RUSHTON, A. W. A.& SHERGOLD, J. H. 2003. Palaeogeographical controls

Lower–Middle Cambrian trilobites, Poland 683
on the Cambrian trilobite immigration and evolutionarypatterns reported in the western Gondwana margin. Pa-laeogeography, Palaeoclimatology, Palaeoecology 195,5–35.
ALVARO, J. J., LINAN, E. & VIZCAINO, D. 1998. Biostrati-graphical significance of the genus Ferralsia (LowerCambrian, Trilobita). Geobios 31, 499–504.
BABCOCK, L. E., PENG, S. C., GEYER, G. & SHERGOLD, J. H.2005. Changing perspectives on Cambrian chronostrati-graphy and progress toward subdivision of the CambrianSystem. Geosciences Journal 9, 101–6.
BARRANDE, J. 1852. Systeme Silurien du Centre dela Boheme. I. Recherches Paleontologiques, vol. 1(Crustaces: Trilobites). Prague & Paris, 935 pp.
BEDNARCZYK, W. 1970. Trilobites Fauna [sic] of the LowerParadoxides oelandicus Stage from the BrzechowArea in the Western Part of the Swietokrzyskie Mts.Bulletin de l’Academie Polonaise des Sciences, Seriedes sciences geologiques et geographiques 18, 29–35.
BEDNARCZYK, W., JURKIEWICZ, H. & ORŁOWSKI, S. 1965.Lower Cambrian and its fauna from the boring of Zarebynear Łagow (Holy Cross Mts.). Bulletin de l’AcademiePolonaise des Sciences, Serie des sciences geologiqueet geographiques 13, 231–6.
BENGSTON, S., CONWAY MORRIS, S., COOPER, B. J., JELL, P.A. & RUNNEGAR, B. N. 1990. Early Cambrian fossilsfrom South Australia. Memoir of the Association ofAustralasian Palaeontologists 9, 1–364.
BERGSTROM, J. & GEE, D. G. 1985. The Cambrian inScandinavia. In The Caledonide Orogen – Scandinaviaand Related Areas (eds D. E. Gee & B. A. Sturt),pp. 247–71. John Wiley & Sons Ltd., London.
BERTHELSEN, A. 1992. From Precambrian to VariscanEurope. In A Continent Revealed: The EuropeanGeotraverse (eds D. Blundell, R. Freeman & S. Muller),pp. 153–64. Cambridge University Press.
BRØGGER, W. C. 1886. Om alderen af Olenelluszoneni Nordamerika. Geologiska Foreningens i StockholmForhandlingar 101, VIII, 182–213.
BRONGNIART, A. 1822. Histoire naturelle des Crustacesfossiles sous les rapports zoologique et geologique,savoir le Trilobites. Paris & Strasbourg, 65 pp.
CHERNYSHEVA, N. E. 1961. (Stratigraphy of the Cambrianof the Aldan Uplift and palaeontologic description ofthe species of the Amga stage.) Trudy VSEGEI, NovayaSerya 49, 1–282 (in Russian).
CHERNYSHEVA, N. E., EGOROVA, L. I., OGIENKO, L. V.,POLETAEVA, O. K. & REPINA, L. N. 1956. (New familiesand genera of trilobites.) In Novye semejstva i rody (edsO. K. Poletaeva & N. E. Chernysheva), pp. 145–82, 324–35. Materialy po paleontologii. Trudy VSEGEI, NovayaSerya 12, paleontologiya, (in Russian).
COBBOLD, E. S. & POCOCK, R. W. 1934. The Cambrian areaof Rushton (Shropshire). Philosophical Transactions ofthe Royal Society of London ser. B 223, 305–409.
CONWAY MORRIS, S. & GRAZHDANKIN, D. 2005. Enigmaticworm-like organisms from the Upper Devonian ofNew York: an apparent example of Ediacaran-likepreservation. Palaeontology 48, 395–410.
CZARNOCKI, J. 1919. Stratygrafja i tektonika GorSwietokrzyskich (Stratigraphy and tectonics ofthe Swiety Krzyz Mountains.) Prace TowarzystwaNaukowego Warszawskiego 28, 1–172.
CZARNOCKI, J. 1927. Kambr i jego fauna w srodkowejczesci Gor Swietokrzyskich. (Cambrian and its faunain the central part of the Holy Cross Mountains.)
Sprawozdania Polskiego Instytutu Geologicznego 4,189–207.
CZARNOCKI, J. 1933. Odsłoniecia kambru okolic Ocieseki Orłowin jako zabytek w znaczeniu naukowem. (Thescientific significance of Cambrian exposures in thevicinity of Ocieseki and Orłowiny.) Zabytki PrzyrodyNieozywionej 2, 78–84.
DEAN, W. T. 2005. Trilobites from the Cal Tepe Formation(Cambrian), near Seydisehir, Central Taurides, south-western Turkey. Turkish Journal of Earth Sciences 14,1–71.
DEAN, W. T. & OZGUL, N. 1994. Cambrian rocks and faunas,Hudai area, Taurus Mountains, southwestern Turkey.Bulletin de l’Institut Royal des Sciences Naturelles deBelgique, Sciences de la Terre 64, 5–20.
DRYGANT, D. & KONSTANTINENKO, L. 2001. (A new trilobitegenus from the Cambrian of the Carpathian Foredeep.)Paleontological Collection 33, 28–31 (in Ukrainian).
DUMICZ, M., TOMCZYKOWA, E. & WOJCIK, L. 1970. Onthe occurrence of Middle Cambrian trilobites in theKhobdo Region (Western Mongolia). Annales de laSociete Geologique de Pologne 40, 287–310.
FLETCHER, T. P., THEOKRITOFF, G., LORD, G. S. & ZEOLI,G. 2005. The early paradoxidid harlani trilobite faunaof Massachusetts and its correlatives in Newfoundland,Morocco, and Spain. Journal of Paleontology 79, 312–36.
GAGAŁA, Ł. 2005. Pre-Ordovician polyphase tectonics of theCambrian sequences in the Kielce Unit, Holy Cross Mts.(Central Poland). Geological Quarterly 49, 53–66.
GEYER, G. 1990a. Revised Lower to lower Middle Cambrianbiostratigraphy of Morocco. Newsletters on Stratigraphy22, 53–70.
GEYER, G. 1990b. Die marokkanischen Ellipsocephalidae(Trilobita: Redlichiida). Beringeria 3, 3–363.
GEYER, G. 1993. The giant Cambrian trilobites of Morocco.Beringeria 8, 71–107.
GEYER, G. 1994. Cambrian corynexochid trilobites fromMorocco. Journal of Paleontology 68, 1306–20.
GEYER, G. 1998. Intercontinental, trilobite-based correlationof the Moroccan early Middle Cambrian. CanadianJournal of Earth Sciences 35, 374–401.
GEYER, G. 2005. The base of a revised Middle Cambrian:are suitable concepts for a series boundary in reach?Geosciences Journal 9, 81–99.
GEYER, G. 2006. First African oryctocephalid trilobitesfrom the Lower–Middle Combrian boundary interval.Palaeoworld 15, 348–59.
GEYER, G. & ELICKI, O. 1995. The Lower Cambrian trilobitesfrom the Gorlitz Synclinorium (Germany) – review andnew results. Palaontologische Zeitschrift 69, 87–119.
GEYER, G. & LANDING, E. 2001. Middle Cambrian ofAvalonian Massachusetts: stratigraphy and correlationof the Braintree trilobites. Journal of Paleontology 75,116–35.
GEYER, G. & LANDING, E. 2004. A unified Lower–MiddleCambrian chronostratigraphy for West Gondwana. ActaGeologica Polonica 54, 179–218.
GEYER, G., LANDING, E. & HELDMEIER, W. 1995. Faunasand depositional environments of the Cambrian of theMoroccan Atlas regions. In Morocco ‘95 – The Lower–Middle Cambrian Standard of western Gondwana (edsG. Geyer & E. Landing), pp. 47–119. Beringeria SpecialIssue 2.
GEYER, G. & PALMER, A. R. 1995. Neltneriidae andHolmiidae (Trilobita) from Morocco and the problem

684 A. ZYLINSKA & M. MASIAK
of Early Cambrian intercontinental correlation. Journalof Paleontology 69, 459–74.
GEYER, G. & SHERGOLD, J. 2000. The quest for internation-ally recognised divisions of Cambrian time. Episodes23, 188–95.
GIL CID, D. 1972. Strenuaeva melendezi nov. sp. delCambrico inferior de Alanıs (Sevilla). EstudiosGeologicos 28, 463–7.
GOZALO, R., ALVARO, J., LINAN, E., SDZUY, K. & TRUYOLS,J. 1993. La distribucion de Paradoxides (Acado-paradoxides) mureroensis SDZUY, 1958 (CambricoMedio basal) y sus implicaciones paleobiogeograficas.Cuaderno Laboratorio Xeoloxico de Laxe 18, 217–30.
GOZALO, R. & LINAN, E. 1997. Revision of the genusOnaraspis Opik, 1968 and its biostratigraphic andbiogeographic significance. In Abstracts with Program,Second International Trilobite Conference, Brock Uni-versity, St. Catharines, Ontario, August 22–25, 1997,24–5.
HAVLICEK, V. 1971. Stratigraphy of the Cambrian of centralBohemia. Sbornık geologickych Ved, Geologie 20, 7–52.
HAWLE, I. & CORDA, A. J. C. 1847. Prodrom einerMonographie der bohmischen Trilobiten. KoniglicheBohmische Gesellschaft der Wissenschaften, Abhand-lungen, 5. Prague: J. G. Calve, 176 pp.
HENNINGSMOEN, G. 1952. Early Middle Cambrian faunafrom Rogaland, SW Norway. Norsk Geologisk Tidsskrift30, 13–32.
HENNINGSMOEN, G. 1958. Los trilobitos de las capas deSaukianda, Cambrico Inferior, en Andalucıa. EstudiosGeologicos 14, 251–71.
HUPE, P. 1953. Contribution a l’etude du Cambrien inferieuret du Precambrien III de l’Anti-Atlas marocain. Notes etMemoirs de la Service geologique du Maroc 103, 1–402(‘1952’).
IVANTSOV, A. YU., ZHURAVLEV, A. YU., LEGUTA, A. V.,KRASSILOV, V. A., MELNIKOVA, L. M. & USHATINSKAYA,G. T. 2005. Palaeoecology of the Early Cambrian Sinskbiota from the Siberian Platform. Palaeogeography,Palaeoclimatology, Palaeoecology 220, 69–88.
JURKIEWICZ, H. 1971. Wgłebna budowa geologiczna okolicŁagowa. (The deep geological structure of the Łagowarea.) Biuletyn Instytutu Geologicznego 242, 5–27.
KAESLER, R. L. (ed.) 1997. Treatise on InvertebratePaleontology, Part O, Arthropoda 1, Trilobita, Revised.Geological Society of America, Boulder, and Universityof Kansas Press, Lawrence, i–xxiv, 1–530.
KIM, D. H., WESTROP, S. R. & LANDING, E. 2002.Middle Cambrian (Acadian Series) conocoryphid andparadoxidid trilobites from the upper Chamberlain’sBrook Formation, Newfoundland and New Brunswick.Journal of Paleontology 76, 822–42.
KOWALCZEWSKI, Z. 1990. Grubookruchowe skały kambru nasrodkowym południu Polski (litostratygrafia, tektonika,paleogeografia). (Coarse-grained Cambrian rocks incentral south Poland (lithostratigraphy, tectonics, pa-laeogeography)). Prace Panstwowego Instytutu Geolo-gicznego 131, 5–82.
LAKE, P. 1935. A monograph of the British Cambriantrilobites. Palaeontographical Society Monographs 88,197–224 (‘1934’).
LENDZION, K. 1976. Stratygrafia kambru zachodniej czescisyneklizy perybałtyckiej. (Stratigraphy of the Cambrianin the western part of the Peri-Baltic Syneclise.) BiuletynInstytutu Geologicznego 270, 59–76.
LENDZION, K. 2001. Some biostratigraphic problems of theCambrian in the Holy Cross and Nida areas of Poland.Geological Quarterly 45, 107–15.
LERMONTOVA, E. V. 1940. Arthropoda. In (Atlas of theleading forms of the fossil faunas of the USSR, vol. I,Cambrian) (eds A. Vologdin, E. Lermontova, V. Yavor-sky & M. Janischevsky.), pp. 112–57 (in Russian).
LINAN, E. & GOZALO, R. 1986. Trilobites del Cambricoinferior y medio de Murero (Cordillera Iberica).Memorias del Museo Paleontologico de la Universidadde Zaragoza 2, 1–104.
LINNARSSON, J. G. O. 1877. Om faunan i lagren med Para-doxides olandicus. Geologiska Foreningens i StockholmForhandlingar 3, 352–75.
MANSUY, H. 1912. IIe Partie Paleontologie. In EtudeGeologique du Yun-nan Oriental (eds J. Deprat & H.Mansuy.), pp. 1–147. Memoirs du Service Geologiquede l’Indochine 1.
MASIAK, M. & ZYLINSKA, A. 1994. Burgess Shale-typefossils in Cambrian Sandstones of the Holy CrossMountains. Acta Palaeontologica Polonica 39, 329–40.
MATTHEW, G. F. 1887. Illustrations of the fauna of theSt. John Group. No. IV. On the smaller-eyed trilobitesof Division I, with a few remarks on the species ofthe higher divisions of the group. Canadian Record ofScience 2, 357–63.
MIZERSKI, W., ORŁOWSKI, S. & ROZYCKI, A. 1986. Tectonicsof the Ocieseki and Zamczysko ranges in the Holy CrossMts. Geological Quarterly 30, 187–200.
MIZERSKI, W., ORŁOWSKI, S. & WAKSMUNDZKI, B. 1991.New data on geology of the Kamieniec Shale Form-ation (Lower Cambrian, Holy Cross Mts). GeologicalQuarterly 35, 149–62.
OPIK, A. A. 1968. The Ordian Stage of the Cambrian and itsAustralian Metadoxididae. Bureau of Mineral Resourcesof Australia, Bulletin 92, 133–66.
OPIK, A. A. 1975. Cymbric Vale Fauna of New South Walesand Early Cambrian Biostratigraphy. Bureau of MineralResources, Geology and Geophysics, Bulletin 159, 1–78.
ORŁOWSKI, S. 1957. On the presence of Paradoxidesolandicus beds in the Holy Cross Mountains. Bulletinde l’Academie Polonaise des sciences, Classe troisieme5, 769–72.
ORŁOWSKI, S. 1959. Paradoxidae from lower MiddleCambrian strata in the vicinity of Sandomierz (centralPoland). Bulletin de l’Academie Polonaise des sci-ences, Serie des sciences chimiques, geologiques etgeographiques 7, 441–6.
ORŁOWSKI, S. 1964a. Kambr srodkowy i jego faunawe wschodniej czesci Gor Swietokrzyskich. (MiddleCambrian and its fauna in the eastern part of the HolyCross Mts.) Studia Geologica Polonica 16, 5–94.
ORŁOWSKI, S. 1964b. Kambr srodkowy w GorachSwietokrzyskich. (The Middle Cambrian in the HolyCross Mts.) Acta Geologica Polonica 14, 547–60.
ORŁOWSKI, S. 1971. The Middle Cambrian of the Klimontowanticlinorium, Holy Cross Mts. Acta Geologica Polonica21, 349–58.
ORŁOWSKI, S. 1973. Cambrian deposits of the Podlasiedepression. Acta Geologica Polonica 23, 365–74.
ORŁOWSKI, S. 1975a. The systematic position and ontogenyof the Lower Cambrian trilobite species Ellipsocephalussanctacrucensis (Samsonowicz, 1959). Acta GeologicaPolonica 25, 369–75.

Lower–Middle Cambrian trilobites, Poland 685
ORŁOWSKI, S. 1975b. Jednostki stratygraficzne kambru igornego prekambru Gor Swietokrzyskich. (Cambrianand Upper Precambrian lithostratigraphic units inthe Holy Cross Mts.) Acta Geologica Polonica 25,431–48.
ORŁOWSKI, S. 1985a. Lower Cambrian and its trilobites in theHoly Cross Mts. Acta Geologica Polonica 35, 231–50.
ORŁOWSKI, S. 1985b. New data on the Middle Cambriantrilobites and stratigraphy in the Holy Cross Mts. ActaGeologica Polonica 35, 251–63.
ORŁOWSKI, S. 1988. Stratigraphy of the Cambrian systemin the Holy Cross Mts. Kwartalnik Geologiczny 32,525–32.
ORŁOWSKI, S. 1989. Trace fossils in the Lower Cambriansequence in Swietokrzyskie Mountains, Central Poland.Acta Palaeontologica Polonica 34, 211–31.
ORŁOWSKI, S. 1992. Cambrian stratigraphy and stagesubdivision in the Holy Cross Mountains, Poland.Geological Magazine 129, 471–4.
PAJCHLOWA, M. (ed.) 1990. Atlas skamieniałosci przewod-nich i charakterystycznych, III, 1a, paleozoik starszy (zproterozoikiem gornym) (Atlas of Guide and Character-istic Fossils, Geology of Poland, III, 1a, Older Palaeozoic(with Upper Proterozoic)), 493 pp. (English versionpublished in 1991.) Polish Geological Institute.
PALMER, A. R. & JAMES, N. P. 1980. The Hawke Bay event:a circum-Iapetus regression near the Lower MiddleCambrian boundary. In The Caledonides in the USA.Proceedings, 1979 Meeting, Blacksburg, Virginia (ed.D. R. Wones), pp. 15–18. Virginia Polytechnic Instituteand State University.
PARNES, A. 1971. Late Lower Cambrian trilobites from theTimna area and Har’Amram (Southern Negev, Israel).Israel Journal of Earth-Sciences 20, 179–205.
POZARYSKI, W. 1990. Kaledonidy srodkowej Europy – oro-genem przesuwczym złozonym z terranow (The MiddleEurope Caledonides – wrenching orogen composed ofterranes.) Przeglad Geologiczny 38, 1–9.
REPINA, L. N., ROMANENKO, E. V., FEDJANINA, E. S. &PEGEL, T.V. 1999. Trilobites from the Lower and lower-most Middle Cambrian of the Kiya River Reference Sec-tion (Kuznetsk Alatau). Annales de Paleontologie 85,3–56.
RICHTER, R. & RICHTER, E. 1940. Studien im Palaozoikumder Mittelmeer-Lander, 5. Die Saukianda-Stufe vonAndalusien, eine fremde Fauna im europaischenOber-Kambrium. Abhandlungen der SenckenbergischenNaturforschenden Gesellschaft 450, 1–88.
RICHTER, R. & RICHTER, E. 1948. Studien im Palaozoikumder Mittelmeer-Lander, 8. Zur Frage des Unter-Kambriums in Nordost-Spanien. Senckenbergiana 29,23–39.
RIDE, W. D. L., COGGER, H. G., DUPUIS, C., KRAUS,O., MINELLI, A., THOMPSON, F. C. & TUBBS, P. K. (eds)1999. International Code of Zoological Nomenclature.Fourth Edition. International Trust for ZoologicalNomenclature, London, 306 pp.
RUDOLPH, F. 1994. Die Trilobiten der mittelkambrischenGeschiebe. Systematik, Morphologie und Okologie.Wankendorf: Verlag Frank Rudolph, 309 pp.
RUSHTON, A. W. A. & POWELL, J. H. 1998. A review ofthe stratigraphy and trilobite faunas from the CambrianBurj Formation in Jordan. Bulletin of the Natural HistoryMuseum of London (Geology) 54, 131–46.
SAMSONOWICZ, J. 1959a. On Strenuaeva from LowerCambrian in Klimontow Anticlinorium. Bulletin de
l’Academie Polonaise des Sciences, Serie des scienceschimiques, geologiques et geographiques 7, 521–4.
SAMSONOWICZ, J. 1959b. On Strenuella and Germaro-pyge from the Lower Cambrian in the KlimontowAnticlinorium. Bulletin de l’Academie Polonaise desSciences, Serie des sciences chimiques, geologiques etgeographiques 7, 525–9.
SAMSONOWICZ, J. 1962. Lower Cambrian fossils from theKlimontow anticlinorium of the Holy Cross Mts.,Poland. Prepared for print from the deceased author’sMS noted by K. Korejwo and L. Teller. In KsiegaPamiatkowa ku czci Profesora Jana Samsonowicza (ed.E. Passendorfer), pp. 9–29. Wydawnictwa Geologiczne,Warszawa.
SCHRANK, E. 1973. Fauna und Kontakt Mittelkam-brium/Oberkambrium in einem Geschiebe. Zeitschriftfur geologische Wissenschaften 22, 85–99.
SDZUY, K. 1957. Revision der mittelkambrischen Trilobitenvon Doberlug. Senckenbergiana lethaea 38, 7–28.
SDZUY, K. 1958. Neue Trilobiten aus dem Mittelkambriumvon Spanien. Senckenbergiana lethaea 39, 235–53.
SDZUY, K. 1961. Das Kambrium Spaniens. Teil II:Trilobiten. 2. Abschnitt. Akademie der Wissenschaftenund Literatur, Abhandlungen der matematisch-naturwissenschaftlichen Klasse 1961(8), 597–690.
SDZUY, K. 1962. Trilobiten aus dem Unter-Kambrium derSierra Morena (S-Spanien). Senckenbergiana lethaea43, 181–229.
SDZUY, K. 1968. Trilobites del Cambrico Medio deAsturias. Trabajos de Geologıa, Faculdad de Ciencias,Universidad de Oviedo 1, 77–133 (‘1967’).
SDZUY, K. 1970. Mittelkambrische Fossilien aus neuenBohrungen bei Doberlug und bei Dielitzsch. Geologie19, 1066–91.
SDZUY, K. 1971. La subdivision bioestratigrafica y lacorrelacion del Cambrico Medio en Espana. I CongresoHispano-Luso-Americano de Geologıa Economica,Seccion 1, Geologıa 2, 769–82.
SHAW, A. B. 1956. Quantitative trilobite studies I. The stat-istical description of trilobites. Journal of Paleontology30, 1209–24.
SHAW, A. B. 1957. Quantitative trilobite studies II. Measure-ment of the dorsal shell of non-agnostidean trilobites.Journal of Paleontology 31, 193–207.
SIVOV, A. G. 1960. (Descriptions of trilobites.) In Biostrati-grafiya paleozoya Sayano-Altajskoj gornoj oblasti,vol. 1. Nizhnij paleozoj (ed. L. L. Khalfina), pp. 152–253. Trudy SNIIGGiMS Novosibirsk 19.
SJOGREN, A. 1872. Om nagra forsteningar i Olandskambriska lager. Geologiska Foreningens i StockholmForhandlingar 1, 67–80.
SNAJDR, M. 1957. O novych trilobitech z ceskeho kambria.(On new trilobites of the Cambrian of Bohemia.) VestnikUstrednıho Ustavu Geologickeho 32, 235–44.
SNAJDR, M. 1958. Trilobiti ceskeho strednıho kambria.(The trilobites of the Middle Cambrian of Bohemia.)Rozpravy Ustrednıho Ustavu Geologickeho 24, 1–280.
SNAJDR, M. 1986. Two new paradoxid trilobites from theJince Formation (Middle Cambrian, Czechoslovakia).Vestnik Ustrednıho Ustavu Geologickeho 61, 169–74.
SOLOV’EV, I. A. 1969. (New species of Paradoxides (trilob-ites) from the oil shales of the Amga Stage of northernYakutia.) Uchenie zapiski NIIGA, paleontologiya ibiostratigrafiya 25, 9–20 (in Russian).

686 Lower–Middle Cambrian trilobites, Poland
SOLOV’EV, I. A. 1980. (Biostratigraphy of deposits in theAmga Stage in the north of the Siberian Platform.)Geologiya i Geofizika 22, 26–38 (in Russian).
STASINSKA, A. 1960. Velumbrella czarnockii n. gen., n.sp. – Meduse du Cambrien inferieur des Monts deSainte-Croix. Acta Palaeontologica Polonica 10, 337–46.
STUDENCKI, M. 1988. Warunki sedymentacji formacjidolnokambryjskich piaskowcow z Ociesek i łupkowz Kamienca w Gorach Swietokrzyskich. (Sedimentaryconditions of the Ocieseki Sandstone and KamieniecShale Formations (Lower Cambrian) in the Holy CrossMts.) Kwartalnik Geologiczny 32, 533–40.
SUNDBERG, F. A. & MCCOLLUM, L. B. 1997. Orycto-cephalids (Corynexochida: Trilobita) of the Lower–Middle Cambrian boundary interval from California andNevada. Journal of Paleontology 71, 1065–90.
TEMPLE, J. T. 1975. Standardisation of trilobite orientationand measurement. Fossils and Strata 4, 461–7.
VOGDES, A. W. 1925. Palaeozoic Crustacea. Part II. A list ofthe genera and subgenera of the Trilobita. Transactionsof the San Diego Society of Natural History 4, 89–115.
WALCOTT, C. D. 1891. The Fauna of the Lower Cambrianor Olenellus Zone. Tenth Annual Report of the U.S.Geological Survey 1, 509–774.
WESTERGARD, A. H. 1928. A deep boring through Middleand Lower Cambrian strata at Borgholm, Isle of Oland.Sveriges Geologiska Undersokning Ser. C 355, 3–19.
WESTERGARD, A. H. 1936. Paradoxides œlandicus beds ofOland with the account of a diamond boring throughthe Cambrian at Mossberga. Sveriges Geologiska Un-dersokning Ser. C 394, 5–66.
WESTERGARD, A. H. 1946. Agnostidea of the Middle Cam-brian of Sweden. Sveriges Geologiska Undersokning 40,1–141.
WESTROP, S. R. & LANDING, E. 2000. Lower Cambrian(Branchian) trilobites and biostratigraphy of the HanfordBrook Formation, southern New Brunswick. Journal ofPaleontology 74, 858–78.
ZHANG (CHANG), W. 1966. (On the classification ofRedlichiacea, with description of new families and newgenera.) Acta Palaeontologica Sinica 14, 135–84 (inChinese).
ZHANG (CHANG), W., LU, Y., ZHU, Z., QIAN, Y., LIN, H.,ZHOU, Z., ZHANG, S. & YUAN, J. 1980. (Cambriantrilobite faunas of southwestern China.) Palaeonto-logia Sinica 159, New Series B, 16, pp. 497 (inChinese).
ZHAO, Y., YUAN, J., PENG, S., GUO, Q., ZHU, L., PENG, J. &WANG, P. 2004. Proposal and prospects for the globalLower–Middle Cambrian boundary. Progress in NaturalScience 14, 1033–8.
ZHU, M., ZHAO, Y. & CHEN, J. 2002. Revision of theCambrian discoidal animals Stellostomites eumorphusand Pararotadiscus guizhouensis from South China.Geobios 35, 165–85.
Copyright © 2022 FDOKUMEN




















