
Monteiro's Storm-petrel Oceanodroma monteiroi: a new species from the Azores
Upload
independentCategory
view
0download
0
- Primary succession on lava domes on Terceira (Azores) - 331
Journal of Vegetation Science 15: 331-338, 2004© IAVS; Opulus Press Uppsala.
Abstract.Questions: Are the vegetation attributes significantly differ-ent among lava domes and among geomorphologic units as aresult of age and soil features? Are the successional rates equalin all the geomorphologic units of the domes? Are the colonizerspecies of lava domes totally replaced by other species in thelate successional stages?Location: Terceira Island, Azores (Portugal).Methods: Three comparable domes of 240, 370 and 2080 yrold were selected. Data on floristic composition, vegetationbioarea (area occupied by plant species in transects), structure,demography and soil nutrients were collected. Quantitativeand qualitative changes along the succession gradient werealso analyzed.Results: Vegetation attributes were consistent with the suc-cessional stage of each dome in the primary sere; however, thegeomorphologic units did not follow the same pattern. Theinfluence of the rates of plant colonization and soil formationare responsible for the decrease of the successional rates fromfootslopes, to summits, to slopes. The vegetation successionallychanges from Juniperus scrub, to Juniperus wood and forest,and there is little species replacement since the similarity inspecies composition is high between the 3 domes.Conclusions: This is a special type of direct succession thattakes place mainly through an increase in biomass and struc-tural complexity. We observed small wooded areas in thefissures of very young domes that are samples of later succes-sional forests – a phenomenon that we call ‘zoom effect’.
Keywords: Bio-area; Direct succession; Oceanic island; Soilnutrient; Succession rate; Vegetation structure; Zoom effect.
Nomenclature: Hansen & Sunding (1993); Schäfer (2002).
Abbreviations: TIR = Terceira Três Irmãs; TLD = TerceiraLomba do Dias; TMN = Terceira Mistérios Negros.
Introduction
Primary succession can be defined as the develop-ment of vegetation on newly formed or exposedsubstrates with no biological legacy and low fertility,especially a low nitrogen level (e.g. Miles 1979;Glenn-Lewin & van der Maarel 1992). Successionimplies changes in species composition, density,cover, species richness and diversity (Bornkamm1981), and in normal succession there is a gradual andsuccessive replacement of species, including canopyspecies (Tagawa 1992). This may be true for mostmainland ecosystems, but in the case of island succes-sion, processes can have unique characteristics thatresult from the disharmony and paucity of the flora(Bush et al. 1995; Dias 1996; Mueller-Dombois 1987).In fact, island effects, such as low rate of successfulcolonization and large ecological amplitude, can beresponsible for a very limited indigenous establish-ment of forest building tree taxa and for the persistenceof species, from early to late succession stages, as inthe Hawaiian islands (Drake & Mueller-Dombois 1993;Kitayama et al. 1995; Mueller-Dombois 1987).
Patterns of succession can be studied through directmonitoring using permanent plots (del Moral & Wood1993; Tsuyuzaki 1991) or can be inferred from a se-quence of closely located sites which differ only in ageor time after disturbances (Drake & Mueller-Dombois1993; Kitayama et al. 1995; Mueller-Dombois &Ellenberg 1974). These two, potentially complementaryapproaches focus on succession at different levels ofspatial resolution and ecological organization, but thechronosequence approach is best to assess long-termsuccessional trajectories (Foster & Tilman 2000).
Using the chronosequence approach we will ad-dress three questions regarding primary succession onlava domes: 1. Are the vegetation attributes (such asdemography, bio-area and structure) significantly dif-ferent among lava domes and among the differentareas of the domes (geomorphologic units) as a resultof age and soil features? 2. Are the successional ratesequal in all geomorphologic units of the domes? 3. Arethe colonizer species of lava domes completely re-placed by other species in the late successional stages?
Primary succession on lava domes on Terceira (Azores)
Elias, Rui B.* & Dias, Eduardo
Universidade dos Açores, Departamento de Ciências Agrárias, Terra Chã,9701-851 Angra do Heroísmo, Azores, Portugal; *Corresponding author: E-mail [email protected]

332 Elias, R.B. & Dias, E.
Methods
Study area
The study area is located in the western part of theisland Terceira (Azores), on the eastern slope of theSanta Bárbara volcano (38º44'N; 27º17'W). After theformation of the caldera this volcano has experienced,in the last 15 000 years, the emission of several lava(trachyte) domes and coulées both inside and on thewestern and eastern slopes of the caldera. Subsequentdeposits of pumice down-fall from eruptions in thisvolcano covered some of these domes (Elias 2001;Self 1976; Zbyszewski et al. 1971).
Lava domes result from the accumulation of vis-cous peralkaline salic lavas around an emission con-duct. As a result, these geologic formations have around or oval contour, no crater and very steep slopes(Self 1976). The most recent dome was formed ca. 240yr ago (Drummond 1836; Neves 1826). Our study waslimited to those domes that are, at present, not coveredby layers of pumice. Radiocarbon analysis indicatesthat the oldest dome without pumice deposit is ca.2080 yr old (Elias 2001; Self 1976). The study area ismainly occupied by natural vegetation and is part ofone of the largest concentrations of natural areas of theAzores islands (Community Interest Site of SantaBárbara-Pico Alto).
Dome selection
The chronosequence approach (Miles 1979; Glenn-Lewin & van der Maarel 1992; Foster & Tilman 2000)was used to study the primary succession on lavadomes. According to Miles (1979) the validity of thechronosequence method depends on all sites havingeffectively identical soils, climatic and microclimaticconditions. Thus, for the purpose of dome selection,the geology, geomorphology, climate, soils and veg-etation were studied (Elias 2001). After this prelimi-nary study was completed, the selection was limited tothree domes which were 240, 370 and 2080 yr old. Thedomes were named Terceira Mistérios Negros (TMN),Terceira Três Irmãs (TIR) and Terceira Lomba do Dias(TLD), respectively.
These domes have the same geologic substrate(trachyte material without pumice fall deposits), thesame geomorphology and the same climatic conditions(the differences of altitude are less than 100 m betweenthe three and they all occur on the eastern slope ofSanta Bárbara volcano). TLD is located 1 km south ofTIR and TMN which avoids possible spatial auto-correlations between the older and the younger domes.The surrounding vegetation is the same for all three
domes, which allows the assumption that the seed rainis similar and the probability of a seed reaching thedome is also similar for all three domes.
Field data collection
One transect was established from the footslope tothe summit of each dome (Fig. 1). Transects were 84,67 and 73 m long for TMN, TIR and TLD respectively.The method was adapted from the transect methoddescribed by Mueller-Dombois & Ellenberg (1974). Ineach transect we measured every plant that interceptedit. Algae, lichens and bryophytes were not measuredwith the exception of Sphagnum species and Poly-trichum commune. Maximum height, medium canopyheight, canopy length and distance to the beginning ofthe transect were measured for each individual. In thecase of grasses, small ferns and macro bryophytes onlythe height and length were measured.
A profile of each dome, with each individual plantin its exact position, was made. Individuals were re-corded as seedlings when the cotyledons were presentand the height was < 10 cm, saplings when the maximumheight was between 10 and 30 cm and as adults whenthe maximum height was > 30 cm and/or reproductiveorgans were present. Three soil samples were collectedin each 10 m segment of each transect.
Data analysis
Previous studies showed that the best approach forthe study of Azorean natural vegetation was the use ofthe biovolume (Dias 1989, 1996). The suitability ofthis floristic-structural parameter for the study of is-land plant communities was also confirmed for theCanary Islands by Fernández-Palacios (1987). In thepresent study the transect method was applied, thus wedid not have information for all three axes of the three-dimensional space and it was not possible to usebiovolume. Instead, we applied an adaptation of thebiovolume which we defined as the ‘bio-area’. Assum-ing that the shape of the canopies (in lateral view) issimilar to an ellipse, it is possible to calculate the areaoccupied by the canopies along the transect (giving the
Fig. 1. Geomorphology of a lava dome showing the threegeomorphologic units identified: foot slope, slope and summit.

- Primary succession on lava domes on Terceira (Azores) - 333
trunk a symbolic bio-area value).Thus the absolute bio-area (BAa) (m
2) of a givenindividual plant is given by:
BAa = p · (CL/2) · (CH/2) (1)
where, BAa is the absolute bio-area, CL is the canopylength and CH is the canopy height. For grasses, smallferns and macro-bryophytes the absolute bio-area isgiven by:
BAa = L · H (2)
where BAa is the absolute bio-area occupied by theindividual (or set of individual plants), L is its lengthand H is its height. In this case the shape of theindividual plants in lateral view was considered simi-lar to a rectangle.
The bio-area of a given species is the result of thesum of the bio-areas of each individual plant. Therelative bio-area (BAr) (m2.m–1) of a given speciesresults from the division of its absolute bio-area (BAa)by the number of metres of the transect (M):
BAr = BAa/M (3)
The vertical structure of the vegetation was ana-lysed through the height distribution of the bio-area bycomputing the area occupied by each species in heightclasses (using 0.5 m intervals), similarly to Dias (1996).
The soil samples were analysed by the Soil Labora-tory of the Department of Agrarian Sciences of theAzores University, to determine its organic matter(%), total nitrogen (%), phosphorus (ppm) and potas-sium (ppm) as well as pH and carbon-nitrogen ratio(C:N). To analyse the correlation between dome ageand soil depth, nitrogen content, phosphorus contentand relative bio-area, simple linear regression wasapplied, assuming dome age as the independent vari-able (Zar 1996). The significance of the differencesbetween mean nitrogen, phosphorus, potassium, C:Nand relative bio-area values of the three domes wasevaluated by Kruskal-Wallis and Mann-Whitney’s Unon-parametric tests (Fernández-Palacios & Santos1996; Zar 1996).
Species diversity was evaluated through Shannon’sindex (H') (Magurran 1983) – using the relative bio-area of a species instead of its number of individuals.Qualitative changes in the vegetation were approachedthrough the Community Coefficient (CC) as definedby Sørensen and modified by Bornkamm (1981):
CC = 2a / (2a+b+c) (4)
where a is the number of species common to bothsuccession stages (for example TMN and TIR), b thenumber of species occurring in one of the stages (TMN,for example) and c the number of species occurring in
the other stage (e.g. TIR). Quantitative changes weredetermined by the Percent Similarity (PS) (Bornkamm1981) but using relative bio-area instead of percent cover:
PS = 2 S min (xi, yi) / S (xi + yi) · 100 (6)
where xi and yi are the relative bio-areas of species i inyears x and y, and min (xi, yi) is the lower relative bio-area value of species i occurring in both years. Clearly,CC will be sensitive to changes in species presence/absence, whereas PS will indicate changes in relativebio-area relations.
Analysis between soil features and vegetation data(using the five most abundant species in each geo-morphologic unit of the domes) was conducted throughCanonical Correspondence Analysis (CCA) to depictrelationships among domes and among geomorphologicunits within domes. The CANOCO program (ter Braak &Smilauer 1998) was used to perform the analysis. Scalingwas focused on inter-sample distances and downweightingof rare species was used. In the other steps of the compu-tation the default options were chosen.
Results
Geochronology and soil analysis
Vegetation data, soil depth, soil nitrogen and phos-phorus contents confirmed the geochronology datasince there is a clear increase of these parameters withdome age. Regression analysis revealed a good corre-lation between these parameters and dome age (Table 1).The soil data indicated significant differences be-tween the three domes with respect to nitrogen, phos-phorus, potassium and carbon-nitrogen ratio but nodifferences were found between the younger domes(TMN and TIR), with the exception of nitrogen con-tent. The percentage of organic matter and all nutrientsincreased from the younger to the older domes. Incontrast, the carbon-nitrogen ratio and pH decreasedwith increasing dome age.
Based on the soil features the domes were dividedinto geomorphologic units: footslope (near the undis-turbed vegetation), slope (the steep lateral slopes of thedome) and summit (the upper part of the dome). In the
Table 1. Relationship between dome age and soil and vegeta-tion data. r = Correlation coefficient.
Parameters TMN TIR TLD r
Age (yr BP) 239 370 2080 -Relative bio-area (m2.m–1) 0.28 0.40 1.73 1.0Mean soil depth (cm) 4 7 23 0.997Nitrogen (%) 0.69 0.99 1.36 0.924Phosphorus (ppm) 12.70 14.40 25.52 0.998
‹
334 Elias, R.B. & Dias, E.
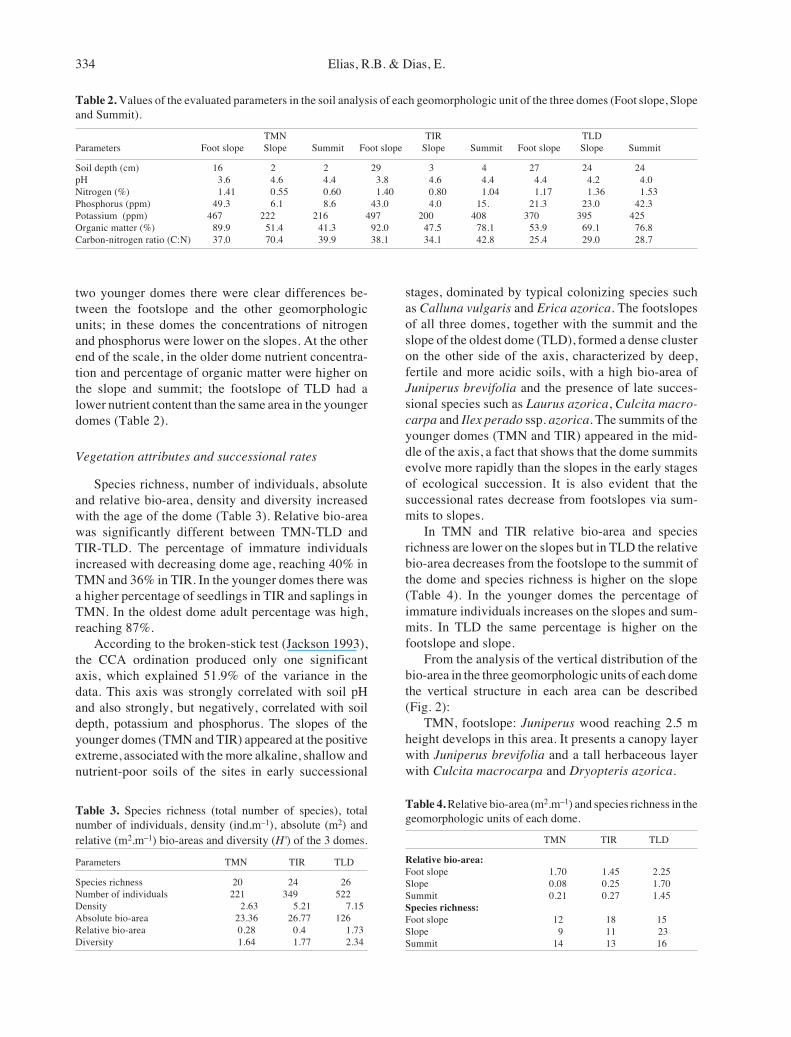
two younger domes there were clear differences be-tween the footslope and the other geomorphologicunits; in these domes the concentrations of nitrogenand phosphorus were lower on the slopes. At the otherend of the scale, in the older dome nutrient concentra-tion and percentage of organic matter were higher onthe slope and summit; the footslope of TLD had alower nutrient content than the same area in the youngerdomes (Table 2).
Vegetation attributes and successional rates
Species richness, number of individuals, absoluteand relative bio-area, density and diversity increasedwith the age of the dome (Table 3). Relative bio-areawas significantly different between TMN-TLD andTIR-TLD. The percentage of immature individualsincreased with decreasing dome age, reaching 40% inTMN and 36% in TIR. In the younger domes there wasa higher percentage of seedlings in TIR and saplings inTMN. In the oldest dome adult percentage was high,reaching 87%.
According to the broken-stick test (Jackson 1993),the CCA ordination produced only one significantaxis, which explained 51.9% of the variance in thedata. This axis was strongly correlated with soil pHand also strongly, but negatively, correlated with soildepth, potassium and phosphorus. The slopes of theyounger domes (TMN and TIR) appeared at the positiveextreme, associated with the more alkaline, shallow andnutrient-poor soils of the sites in early successional
stages, dominated by typical colonizing species suchas Calluna vulgaris and Erica azorica. The footslopesof all three domes, together with the summit and theslope of the oldest dome (TLD), formed a dense clusteron the other side of the axis, characterized by deep,fertile and more acidic soils, with a high bio-area ofJuniperus brevifolia and the presence of late succes-sional species such as Laurus azorica, Culcita macro-carpa and Ilex perado ssp. azorica. The summits of theyounger domes (TMN and TIR) appeared in the mid-dle of the axis, a fact that shows that the dome summitsevolve more rapidly than the slopes in the early stagesof ecological succession. It is also evident that thesuccessional rates decrease from footslopes via sum-mits to slopes.
In TMN and TIR relative bio-area and speciesrichness are lower on the slopes but in TLD the relativebio-area decreases from the footslope to the summit ofthe dome and species richness is higher on the slope(Table 4). In the younger domes the percentage ofimmature individuals increases on the slopes and sum-mits. In TLD the same percentage is higher on thefootslope and slope.
From the analysis of the vertical distribution of thebio-area in the three geomorphologic units of each domethe vertical structure in each area can be described(Fig. 2):
TMN, footslope: Juniperus wood reaching 2.5 mheight develops in this area. It presents a canopy layerwith Juniperus brevifolia and a tall herbaceous layerwith Culcita macrocarpa and Dryopteris azorica.
Table 2. Values of the evaluated parameters in the soil analysis of each geomorphologic unit of the three domes (Foot slope, Slopeand Summit).
TMN TIR TLDParameters Foot slope Slope Summit Foot slope Slope Summit Foot slope Slope Summit
Soil depth (cm) 16 2 2 29 3 4 27 24 24pH 3.6 4.6 4.4 3.8 4.6 4.4 4.4 4.2 4.0Nitrogen (%) 1.41 0.55 0.60 1.40 0.80 1.04 1.17 1.36 1.53Phosphorus (ppm) 49.3 6.1 8.6 43.0 4.0 15. 21.3 23.0 42.3Potassium (ppm) 467 222 216 497 200 408 370 395 425Organic matter (%) 89.9 51.4 41.3 92.0 47.5 78.1 53.9 69.1 76.8Carbon-nitrogen ratio (C:N) 37.0 70.4 39.9 38.1 34.1 42.8 25.4 29.0 28.7
Table 3. Species richness (total number of species), totalnumber of individuals, density (ind.m–1), absolute (m2) andrelative (m2.m–1) bio-areas and diversity (H') of the 3 domes.
Parameters TMN TIR TLD
Species richness 20 24 26Number of individuals 221 349 522Density 2.63 5.21 7.15Absolute bio-area 23.36 26.77 126Relative bio-area 0.28 0.4 1.73Diversity 1.64 1.77 2.34
Table 4. Relative bio-area (m2.m–1) and species richness in thegeomorphologic units of each dome.
TMN TIR TLD
Relative bio-area:Foot slope 1.70 1.45 2.25Slope 0.08 0.25 1.70Summit 0.21 0.27 1.45Species richness:Foot slope 12 18 15Slope 9 11 23Summit 14 13 16
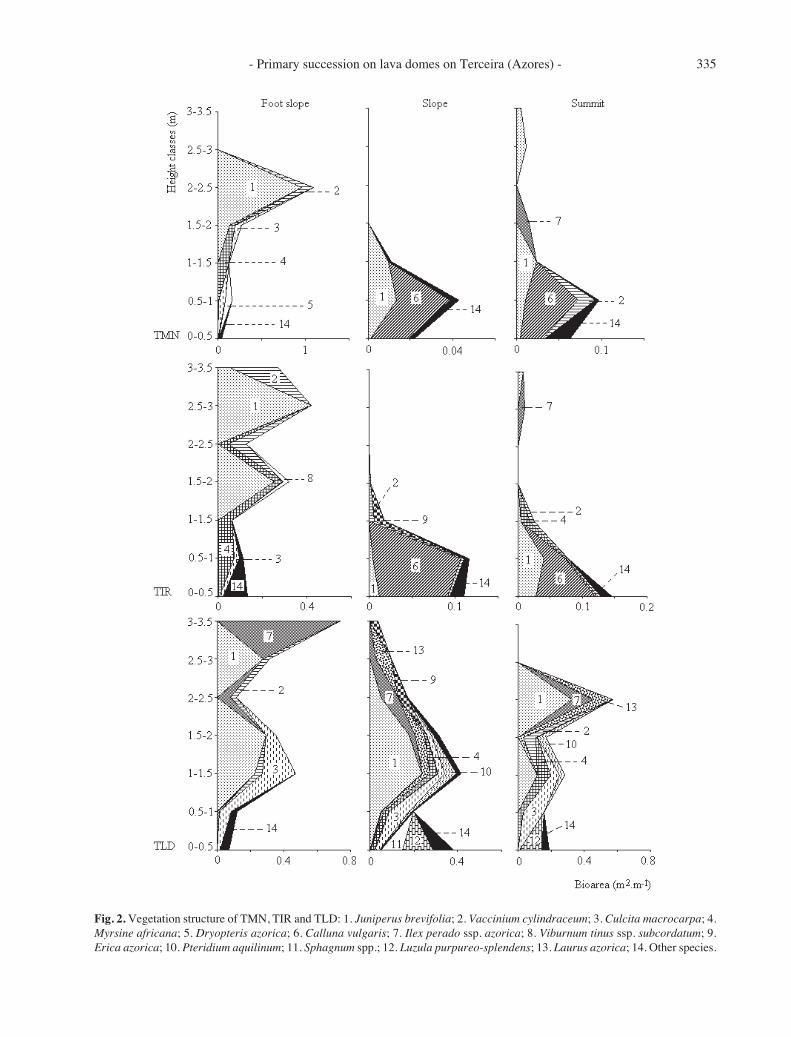
- Primary succession on lava domes on Terceira (Azores) - 335
Fig. 2. Vegetation structure of TMN, TIR and TLD: 1. Juniperus brevifolia; 2. Vaccinium cylindraceum; 3. Culcita macrocarpa; 4.Myrsine africana; 5. Dryopteris azorica; 6. Calluna vulgaris; 7. Ilex perado ssp. azorica; 8. Viburnum tinus ssp. subcordatum; 9.Erica azorica; 10. Pteridium aquilinum; 11. Sphagnum spp.; 12. Luzula purpureo-splendens; 13. Laurus azorica; 14. Other species.

336 Elias, R.B. & Dias, E.
TMN, slope: Calluna-Juniperus pioneer scrubreaching 1.5 m height develops here. The vegetationcover values are very low (with only 20% of thesurface covered) and the community presents a clearpatchy horizontal structure with ‘spots’ of vegetationin rock surface depressions and with the more exposedsurfaces almost clear of vegetation.
TMN, summit: This area harbours vegetation com-posed of three different formations according to themicro- and mesohabitats. The depressions on the rocksurface are occupied by a Calluna-Juniperus scrubbut a Juniperus scrub develops in the micro-fissures.Small Juniperus-Ilex woods, which (in spite of theyoung age of this dome) can reach a height of 3.5 m,are present in the macro-fissures. The more exposedrock surfaces are almost clear of vegetation. Thisspatial heterogeneity is reflected in Fig. 2 with theapparent existence of two layers: one in the 0.5 - 1 minterval and another at 2.5 - 3 m.
TIR, foot slope: Juniperus forest reaching 3.5 mheight develops in this area. It presents a canopy layerwith J. brevifolia, a shrub layer also dominated by J.brevifolia and a low herbaceous/bryophyte layer withseveral species.
TIR, slope: With 75% of the rock surface covered,this area presents a Calluna-Juniperus scrub with amaximum height of 2 m.
TIR, summit: In this area the vegetation is similarto that of the top of TMN. Here, however the vegeta-tion cover reaches 80% of the rock surface.
TLD, footslope: This area is covered by a Juniperus-Ilex forest composed of three layers: a canopy layer ofJ. brevifolia and Ilex perado ssp. azorica with amaximum height of 3.5 m; a shrub / tall herbaceouslayer mainly with J. brevifolia and Culcita macrocarpaand a low herbaceous/bryophyte layer mainly withLuzula purpureo-splendens and Sphagnum spp.
TLD, slope: A Juniperus wet scrub with two layersdevelops in this area: a shrub/tall herbaceous layerwith high species richness and dominated by J. brevi-folia and a well developed low herbaceous/ bryophytelayer dominated by Sphagnum spp. and L. purpureo-splendens.
TLD, summit: This area harbours a Juniperus-Ilex-Laurus forest composed of a canopy layer with J.brevifolia, Ilex perado ssp. azorica and Laurus azorica,a shrub/tall herbaceous layer dominated by J. brevifolia,Myrsine africana and Culcita macrocarpa and a lowherbaceous/bryophyte layer dominated by L. purpureo-splendens.
An epiphytic layer of Elaphoglossum semicylindri-cum and Hymenophyllum tunbrigense is present in allgeomorphologic units of TLD and in the footslopes ofTMN and TIR.
Species replacement
In Table 5 we present the relative bio-area of theten most abundant species in each dome. In TMN andTIR there is a dominance of J. brevifolia and Callunavulgaris but J. brevifolia dominates in TLD followedby Ilex perado ssp. azorica and Culcita macrocarpa.From the 33 species identified in the three domes, 15occur in all of them, two are common to TMN and TIR,four occur in TIR and TLD and one is common toTMN and TLD. Five species (J. brevifolia, Ilex peradossp. azorica, Culcita macrocarpa, Myrsine africanaand Vaccinium cylindraceum) are among the ten mostabundant species in all the domes.
The Community Coefficient is similar between thethree domes, with values between 0.70 and 0.77, whichindicates that a significant percentage of the speciespresent in the younger domes are also present through-out the succession. This fact is reinforced by the highsimilarity (0.70) between the younger domes and thesurrounding vegetation (undisturbed by the eruption).However, the Percent Similarity is low between theyounger domes (TMN and TIR) and TLD (23.6%between TMN and TLD and 25.7% between TIR andTLD), which shows that in spite of the similarity inspecies composition there are important changes invegetation bio-area.
Table 5. Relative bio-area (m2.m–1) of the ten most abundantspecies in each dome.
Species TMN TIR TLD
Juniperus brevifolia 0.136 0.132 0.54Calluna vulgaris 0.055 0.134 -Vaccinium cylindraceum 0.023 0.05 0.07Myrsine africana 0.022 0.039 0.08Culcita macrocarpa 0.016 0.007 0.19Dryopteris azorica 0.007 - -Ilex perado ssp. azorica 0.006 0.007 0.21Blechnum spicant 0.005 0.006 -Sphagnum spp. 0.004 - 0.08Hymenophyllum tunbrigense 0.001 - -Erica azorica - 0.005 0.06Polytrichum commune - 0.004 -Rubia agostinhoi - 0.004 -Laurus azorica - - 0.14Luzula purpureo-splendens - - 0.13Pteridium aquilinum - - 0.07

- Primary succession on lava domes on Terceira (Azores) - 337
Discussion
Vegetation attributes and successional rates
Vegetation attributes were consistent with the suc-cessional stage of each dome in the primary sere. Theage increase is followed by an increase in the numberof individuals, density, bio-area, percentage of adultsand structural complexity. However, in the geomorpho-logic units the vegetation did not follow this pattern.Clearly the successional sequence of geomorphologicunits is from earlier to later successional stages: slopesof TMN and TIR, summits of TMN and TIR, TLDslope, footslopes of TMN and TIR, TLD summit andTLD footslope. In this gradient the vegetation suc-cessionally changes from Calluna-Juniperus scrubs(slopes and summits of TMN and TIR), to Juniperuswet scrubs (TLD slope), to Juniperus woods (TMNfootslope), to Juniperus forests (TIR footslope), toJuniperus-Ilex forests (TLD footslope).
According to Marchese & Grillo (2000) the dy-namic vegetation stages cannot always be related tothe age of the lava as other factors such as surfacemorphology and soil depth become relevant. The suc-cessional pattern of the geomorphologic units is prob-ably the consequence of: (1) different rates of plantcolonization and secondary dispersion and (2) differ-ent rates of soil formation. The influence of these twofactors results in different successional rates: wherethe colonization and secondary dispersion rates arehigher and the conditions for soil formation are better(as on the footslopes, due to the proximity of vegeta-tion undisturbed by the eruption, and in the summits,due to the presence of fissures) the successional ratesare higher; where the colonization and secondary dis-persion rates are lower and the conditions for soilformation are worse (as on the slopes) the successionalrates are lower. The decrease of the successional ratesis reflected in the decrease in structural complexityfrom foot slopes via summits to slopes.
Species replacement
Unlike mainland ecosystems and non-oceanic is-lands in which the successional processes involvechanges in diversity and floristic composition (e.g.Frenot et al. 1998; Kamijo et al. 2002; Marchese &Grillo 2000), in the lava domes of Terceira most changesare found in biomass and structure. There is littlespecies replacement since the similarity in speciescomposition is high between the three domes. Quanti-tative changes are more important due to the cleardifferences in bio-area. Calluna vulgaris is a typicalcolonizing species that decreases progressively in
biomass as shading by tree species increases. The otherimportant colonizing species is J. brevifolia, which isat the same time the dominant canopy species in thelater succession stages. Typical successions are oftencharacterized by a sequence of lichen/moss, herba-ceous, shrub and forest communities of different speciescomposition. In our volcanic domes we found no evi-dence of herbaceous communities, and the vegetationsuccessionally changes from Juniperus scrubs, toJuniperus woods and forests.
Other oceanic islands have somewhat similar suc-cessional processes. In the Hawaii islands Metrosiderospolymorpha is the dominant canopy species through-out the succession (Drake & Mueller-Dombois 1993;Kitayama et al. 1995) but in the case of the Azoresseveral other species, besides J. brevifolia, from thecanopy and under canopy layers, are also present fromthe beginning (e.g. Ilex perado ssp. azorica, Culcitamacrocarpa, Myrsine africana and Vaccinium cylindra-ceum). According to Aplet et al. (1998) in Mauna Loa(Hawaii) there is an increase in species richness andbiomass during the primary sere. In our case, in spite ofthe clear increase of biomass, species richness remainslargely unaffected by time.
The primary succession on lava domes can be con-sidered a special type of ‘direct succession’ describedby Miles (1987). The succession process takes placemainly through an increase in biomass and structuralcomplexity, and it is possible to see small and structur-ally simpler Juniperus-Ilex woods in the macro-fis-sures of young domes that are small samples of theforests we find in late succession stages. We havenamed this process ‘zoom effect’. This type of succes-sion is probably due to the relatively poor native spe-cies richness of the Azores islands. Isolated islandsoften have disharmonic and poor floras (Bush et al.1995; Dias 1996; Mueller-Dombois 1987, 2000) andin the case of the Azores this is even more evidentsince they have only ca. 300 native species.
Acknowledgements. We would like to thank all those whocontributed to this work, in particular José Madeira, FernandoPereira and Lurdes Matos. This study was supported by theProject LIFE96NAT-P-3022 ‘Study and conservation of theAzorean natural patrimony’, of the Departamento de CiênciasAgrárias da Universidade dos Açores / Direcção Regionaldos Recursos Florestais.

338 Elias, R.B. & Dias, E.
References
Aplet, G.H., Hughes, R.F. & Vitousek, P.M. 1998. Ecosys-tem development on Hawaiin lava flows: biomass andspecies composition. J. Veg. Sci. 9: 17-26.
Bornkamm, R. 1981. Rates of change in vegetation duringsecondary succession. Vegetatio 47: 213-220.
Bush, M.B., Whittaker, R.J. & Partomihardjo, T. 1995. Colo-nization and succession on Krakatau: an analysis of theguild of vining plants. Biotropica 27: 355-372.
del Moral, R. & Wood, D.M. 1993. Early primary successionon the volcano Mount St. Helens. J. Veg. Sci. 4: 223-234.
Dias, E. 1989. Flora e vegetação endémica na ilha Terceira,Açores. M.Sc. Thesis, Universidade dos Açores, Angrado Heroísmo, Azores, PT.
Dias, E. 1996. Vegetação natural dos Açores. Ecologia esintaxonomia das florestas naturais. Ph.D. Thesis, Univer-sidade dos Açores, Angra do Heroísmo, Azores, PT.
Drake, D.R. & Mueller-Dombois, D. 1993. Population de-velopment of rain forest trees on a chronosequence ofHawaiian lava flows. Ecology 74: 1012-1019.
Drummond, F.F. 1836. Annaes da ilha Terceira. CâmaraMunicipal de Angra do Heroísmo. Angra do Heroísmo,Azores, PT.
Elias, R.B. 2001. Sucessão primária em domas traquíticos.M.Sc. Dissertation, Universidade dos Açores, Departa-mento de Ciências Agrárias, Angra do Heroísmo, Azores,PT.
Fernández-Palacios, J.M. 1987. Analisis de la variaciónespacial de la vegetación de Tenerife e interpretación enterminos ambientales. Ph.D. Thesis, Universidade de LaLaguna. Tenerife, Canarias, ES.
Fernández-Palacios, J.M. & Santos, A. 1996. Ecologia de lasislas Canarias – Muestreo y análisis de poblaciones ycomunidades. Sociedad La Cosmológica, Santa Cruz deLa Palma, Canarias, ES.
Foster, B.L. & Tilman, D. 2000. Dynamic and static views ofsuccession: testing the descriptive power of the chrono-sequence approach. Plant Ecol. 146: 1-10.
Frenot, Y., Gloaguen, J.C., Cannavacciulo, M. & Bellido, A.1998. Primary succession on glacier forelands in thesubantartic Kerguelen Islands. J. Veg. Sci. 9: 75-84.
Glenn-Lewin, D. & van der Maarel, E. 1992. Patterns andprocesses of vegetation dynamics. In: Glenn-Lewin, D.,Peet, R.K. & Veblen, T.T. (eds.) Plant succession –theory and prediction, pp. 11-59. Chapman & Hall, Lon-don, UK.
Hansen, A. & Sunding, P. 1993. Flora of Macaronesia.Checklist of vascular plants. 4. rev. ed. Sommerfeltia 17.
Jackson, D.A. 1993. Stopping rules in principal componentsanalysis: a comparison of heuristical and statistical ap-proaches. Ecology 74: 2201-2214.
Kamijo, T., Kitayama, K., Sugawara, A., Urushimichi, S. &Sasai, K. 2002. Primary succession on the warm-temper-ate broad leave forest on a volcanic island, Miyake-Jima,Japan. Folia Geobot. 37: 71-91.
Kitayama, K., Mueller-Dombois, D. & Vitousek, P.M. 1995.Primary succession of Hawaiian montane rain foreston a chronosequence of eight lava flows. J. Veg. Sci. 6:
211-222.Magurran, A.E. 1983. Ecological diversity and its measure-
ment. Chapman & Hall, London, UK.Marchese, E.P. & Grillo, M. 2000. Vegetation dynamics on
lava flows on Mt. Etna. Proceedings IAVS Symposium,pp. 210-213. Opulus Press, Uppsala, SE.
Miles, J. 1979. Vegetation dynamics. Chapman & Hall, Lon-don, UK.
Miles, J. 1987. Vegetation succession: past and present per-ceptions. In: Gray, A.J., Crawley, M.J. & Edwards, P.J.(eds.) Colonization, succession and stability, pp. 1-29.Blackwell Science, Oxford, UK.
Mueller-Dombois, D. 1987. Forest dynamics in Hawaii.Trends Ecol. Evol. 2: 216-219.
Mueller-Dombois, D. 2000. Succession and zonation in theHawaiian Islands. Proceedings IAVS Symposium, pp.203-206. Opulus Press, Uppsala, SE.
Mueller-Dombois, D. & Ellenberg, H. 1974. Aims and meth-ods of vegetation ecology. Wiley, London, UK.
Neves, J. A. 1826. Entretenimentos cosmológicos, geográficose históricos, Tomo I. Impressão Regia, Lisbon, PT.
Schäfer, H. 2002. Flora of the Azores. Margraf Verlag,Weikersheim, DE.
Self, S. 1976. The recent volcanology of Terceira, Azores. J.Geol. Soc. Lond. 132: 645-666.
Tagawa, H. 1992. Primary succession and the effect of firstarrivals on subsequent development of forest types.GeoJournal 28.2: 175-183.
ter Braak, C.J.F. & Smilauer, P. 1998. CANOCO Referencemanual and user’s guide to Canoco for Windows. Soft-ware for canonical community ordination (version 4.0).Centre for Biometry, Wageningen, NL.
Tsuyuzaki, S. 1991. Species turnover and diversity duringearly stages of vegetation recovery on the volcano Usu,northern Japan. J. Veg. Sci. 2: 301-306.
Zar, J., 1996. Biostatiscal analysis, 3rd. ed. Prentice Hall,Englewood Cliffs, NJ, US.
Zbyszewski, G., Medeiros, A.C., Ferreira, O.V. & Assunção,C.T. 1971. Carta geológica de Portugal. Notícia explica-tiva da folha ilha Terceira. Serviços Geológicos de Por-tugal, Lisbon, PT.
Received 3 March 2003;Accepted 19 November 2003.
Co-ordinating Editor: E. Ezcurra.
‹
Copyright © 2022 FDOKUMEN




















