
Vaccination with the Leishmania infantum Acidic Ribosomal P0 Protein plus CpG Oligodeoxynucleotides...
12
10.1128/IAI.73.9.5842-5852.2005. 2005, 73(9):5842. DOI: Infect. Immun. Alonso, David Sacks and Manuel Soto Salvador Iborra, Javier Carrión, Charles Anderson, Carlos Progressive Disease in BALB/c Mice C57BL/6 Mice but Does Not Prevent against Cutaneous Leishmaniasis in Oligodeoxynucleotides Induces Protection Acidic Ribosomal P0 Protein plus CpG Leishmania infantum Vaccination with the http://iai.asm.org/content/73/9/5842 Updated information and services can be found at: These include: REFERENCES http://iai.asm.org/content/73/9/5842#ref-list-1 at: This article cites 35 articles, 17 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on December 19, 2013 by guest http://iai.asm.org/ Downloaded from on December 19, 2013 by guest http://iai.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Vaccination with the Leishmania infantum Acidic Ribosomal P0 Protein plus CpG Oligodeoxynucleotides...

10.1128/IAI.73.9.5842-5852.2005.
2005, 73(9):5842. DOI:Infect. Immun. Alonso, David Sacks and Manuel SotoSalvador Iborra, Javier Carrión, Charles Anderson, Carlos Progressive Disease in BALB/c MiceC57BL/6 Mice but Does Not Preventagainst Cutaneous Leishmaniasis in Oligodeoxynucleotides Induces ProtectionAcidic Ribosomal P0 Protein plus CpG
Leishmania infantumVaccination with the
http://iai.asm.org/content/73/9/5842Updated information and services can be found at:
These include:
REFERENCEShttp://iai.asm.org/content/73/9/5842#ref-list-1at:
This article cites 35 articles, 17 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
on D
ecember 19, 2013 by guest
http://iai.asm.org/
Dow
nloaded from

INFECTION AND IMMUNITY, Sept. 2005, p. 5842–5852 Vol. 73, No. 90019-9567/05/$08.00�0 doi:10.1128/IAI.73.9.5842–5852.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Vaccination with the Leishmania infantum Acidic Ribosomal P0Protein plus CpG Oligodeoxynucleotides Induces Protection
against Cutaneous Leishmaniasis in C57BL/6 Mice butDoes Not Prevent Progressive Disease
in BALB/c MiceSalvador Iborra,1 Javier Carrion,1 Charles Anderson,2 Carlos Alonso,1
David Sacks,2 and Manuel Soto1*Centro de Biologıa Molecular “Severo Ochoa,” Universidad Autonoma de Madrid, 28049 Madrid, Spain,1
and Laboratory of Parasitic Diseases, National Institute of Allergy and Infectious Diseases,National Institutes of Health, Bethesda, Maryland 208922
Received 15 February 2005/Returned for modification 25 March 2005/Accepted 15 April 2005
We have examined the efficacy of the administration in mice of a molecularly defined vaccine based on theLeishmania infantum acidic ribosomal protein P0 (rLiP0). Two different challenge models of murine cutaneousleishmaniasis were used: (i) subcutaneous inoculation of L. major parasites in susceptible BALB/c mice (amodel widely used for vaccination analysis) and (ii) the intradermal inoculation of a low infective dose inresistant C57BL/6 mice (a model that more accurately reproduces the L. major infection in natural reservoirsand in human hosts). First, we demonstrated that C57BL/6 mice vaccinated with LiP0-DNA or rLiP0 proteinplus CpG oligodeoxynucleotides (ODN) were protected against the development of dermal pathology andshowed a reduction in the parasite load. This protection was associated with production of gamma interferon(IFN-�) in the dermal site. Secondly, we showed that immunization with rLiP0 plus CpG ODN is able to induceonly partial protection in BALB/c, since these mice finally developed a progressive disease. Further, wedemonstrated that LiP0 vaccination induces a Th1 immunological response in both strains of mice. In bothcases, the antibodies against LiP0 were predominantly of the immunoglobulin G2a isotype, which was corre-lated with an rLiP0-stimulated production of IFN-� in draining lymph nodes. Finally, we demonstrated thatLiP0 vaccination does not prevent the Th2 response induced by L. major infection in BALB/c mice. Takentogether, these data indicate that the BALB/c model of cutaneous leishmaniasis may undervalue the potentialefficacy of some vaccines based on defined proteins, making C57BL/6 a suitable alternative model to test vaccinecandidates.
Vaccines for a variety of diseases caused by intracellularinfections like tuberculosis, malaria and leishmaniasis requirethe induction and maintenance of cellular immune responses.Infection by Leishmania major, the main etiologic agent ofzoonotic cutaneous leishmaniasis in the Old World, has beenextensively used in mouse models to understand the require-ments for effective vaccination. Despite the evidence of ac-quired immunity and resistance to reinfection in natural Leish-mania hosts, suggesting that a vaccine is feasible, there is novaccine for human leishmaniasis. The chemotherapy availableat present for this disease is far from satisfactory and there isincreasing resistance against those drugs (6).
To date, leishmanization is the most effective induction ofprotective immunity to prevent disfiguring cutaneous leish-maniasis (9, 17, 23). However, this active infection with liveparasites has been restricted or abandoned completely due toseveral associated problems, such as the development of un-controlled skin lesions and immunosuppression (12). Recent
advances in the design of vaccines against leishmaniasis arebased on molecularly defined antigens, the so-called second-generation vaccines (24). One of these molecules is the L.infantum acidic ribosomal P0 protein (rLiP0), a structural com-ponent of the large ribosome subunit that has been describedas an immunodominant antigen recognized by sera from bothpatients and dogs infected with L. chagasi-L. infantum (31, 32).We have previously shown that DNA vaccination with thisantigen partially protects BALB/c mice against L. major infec-tion (16). In addition, the C-terminal region of the LiP0 ispresent in a multicomponent protein that when administeredto dogs, using live BCG as adjuvant, confers protection againstL. infantum infection (22).
Effective primary immunity against L. major in mouse isknown to require interleukin 12 (IL-12)-dependent productionof gamma interferon (IFN-�) from CD4� T cells (Th1 re-sponse), which mediates nitric oxide (NO)-dependent killingby infected macrophages (recently reviewed in references 10,27, and 29). Whereas C57BL/6 mice develop protective Th1responses and control infection, susceptible BALB/c mice ex-hibit an IL-4-driven Th2 response produced by a restrictedpopulation of V�8/V�4 CD4� T cells and are unable to con-trol infection. The genetic susceptibility of BALB/c mice to L.
* Corresponding author. Mailing address: Centro de Biologıa Mo-lecular “Severo Ochoa,” Universidad Autonoma de Madrid, 28049Madrid, Spain. Phone: (34-91) 4978454. Fax: (34-91) 4974799. E-mail:[email protected].
5842
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from

major may be prevented by treatment with IL-12 protein orneutralizing antibodies to IL-4 at the time of infection, whichhas been shown to shift the immune response to a Th1 profile(15, 28). The usefulness of this model for vaccine developmentrelies on the possibility to modify the Th responses and toassess the correlation of these responses with functional andbiological outcomes.
Although most studies involving vaccination against cutane-ous leishmaniasis have been conducted in BALB/c mice, thehealing lesions observed in C57BL/6 mice may provide a morerelevant model of L. major infection in natural reservoirs andin human hosts. This infection model takes into account twomain features of natural transmission: low dose (100 to 1,000metacyclic promastigotes) and inoculation into a dermal site(the ear dermis). The analysis of adaptive immunity in thismodel confirmed a role for Th1 cells, and in addition revealedan essential requirement for CD8� T cells (2).
Several vaccine studies have demonstrated that long-lastingcellular responses and protection against L. major may beachieved by genetic vaccination, or by coinoculation of Leish-mania antigen plus immunostimulatory CpG oligodeoxynucle-otides (ODN) (11, 25). It has been shown also that CpG ODNtrigger a therapeutic response in BALB/c mice infected with L.major (35) or L. donovani (7). Finally, it has been demon-strated that the pathogenicity of leishmanization may be re-duced by the coinjection of the parasite with CpG ODN (21).These effects probably rely on their ability to induce bothinnate and adaptive cellular immune responses. These ODNstimulate macrophages and dendritic cells (DC) to synthesizeseveral cytokines including IL-12, IL-18, tumor necrosis factoralpha (TNF-�), IFN-�, IFN-�, and IFN-� and to upregulatecostimulatory molecules such as CD40 and major histocom-patibility complex class II (MHC-II) (13, 18, 33). Moreover,CpG ODN may activate DC, leading to the presentation ofsoluble protein to class I-restricted T cells and the induction ofcytotoxic T-lymphocyte responses (4, 8, 34).
In the present paper, we have evaluated the vaccination withthe LiP0 protein using very different challenge systems: high-dose inocula in the footpad of BALB/c mice (widely used incutaneous leishmaniasis vaccine assays) and low dose in the earof C57BL/6 mice (a model that more closely mimics the humandisease in terms of route and dose infection). We demonstratethat C57BL/6 mice inoculated with LiP0 plus CpG ODN wereprotected against cutaneous leishmaniasis, showing a reduc-tion of the parasite number in the inoculation site, and amoderation of dermal pathology. In contrast, BALB/c micevaccinated in the same fashion were only partially protectedagainst L. major challenge, developing delayed but ultimatelyprogressive, nonhealing lesions. We show that although LiP0vaccine induces a specific Th1 response in both strains it doesnot abrogate the Th2 response induced by L. major challengein BALB/c mice, which may account for its different efficacy inthese two models.
MATERIALS AND METHODS
Mouse strains and parasites. Female BALB/c and C57BL/6 mice were 6 to 8weeks old at the onset of experiments and were purchased from Harlan Inter-fauna Iberica S.A. (Barcelona, Spain). L. major parasites (clone WHOM/IR/173)and clone V1 (MHOM/IL/80/Friedlin) were kept in a virulent state by passage inBALB/c mice. The L. major amastigotes were obtained from popliteal lymph
nodes, and after transformation from amastigote to promastigote form, parasiteswere cultured and harvested at late stationary phase. Promastigotes of bothclones were cultured at 26°C in Schneider’s medium (Gibco BRL) supplementedwith 20% fetal calf serum (FCS). Infective-stage promastigotes (metacyclics) ofL. major (clone V1) were isolated from stationary cultures by negative selectionusing peanut agglutinin (Vector Laboratories, Burlingame, CA).
Plasmids, ODN, and leishmanial antigens. The plasmids used in this studywere pcDNA3 (empty vector) and pcDNA3 encoding the Leishmania infantumP0 ribosomal protein (pcDNA3-LiP0) constructed as described previously (16).Endotoxin-free plasmid DNA was isolated using the EndoFree Plasmid Giga kit(QIAGEN, Hilden, Germany). Plasmids used in this study were all suspended insterile phosphate-buffered saline (PBS).
Phosphorothioate-modified ODN sequences containing CpG motifs were syn-thesized by Isogen (The Netherlands). The sequences of the immunostimulatoryODN (5� to 3�) were TCAACGTTGA and GCTAGCGTTAGCGT. The his-tagged rLiP0 was produced in Escherichia coli M15 transformed with pQE-LiP0and purified as previously described (16). The recombinant protein was furtherpassed through a polymyxin-agarose column (Sigma, St. Louis, Mo.) to eliminateendotoxins. The lipopolysacharide content was measured by the QuantitativeChromogenic Limulus Amebocyte Assay QCL-1000 (BioWhittaker, Walkers-ville, Md.), showing that preparations were essentially free of endotoxin (�12 pg�g�1 of rLiP0).
A library of overlapping peptides covering the whole LiP0 protein sequencewas synthesized by the simultaneous multiple-peptide solid phase syntheticmethod using a polyamide resin and FMOC chemistry as described before (32).ALM (autoclaved L. major) is prepared from freeze-thaw lysates of autoclavedL. major promastigotes bound to aluminum hydroxide, prepared as previouslydescribed (14).
Total proteins of L. major (soluble Leishmania antigen [SLA]) were preparedby three freezing and thawing cycles of stationary promastigotes of L. majorsuspended in PBS. After cell lysis, soluble antigens were separated from theinsoluble fraction by centrifugation for 15 min at 12,000 g using a microcen-trifuge.
Immunizations and parasite challenge. C57BL/6 mice were injected subcuta-neously (s.c.) in the footpad with either PBS, 100 �g of pcDNA3 (empty vector)or pcDNA3-LiP0. Each group was inoculated three times at 2-week intervals.Additional groups were immunized with either 10 �g of rLiP0 plus 50 �g of CpGODN (25 �g of each immunostimulatory ODN), 50 �g of ALM plus CpG ODN(50 �g), CpG ODN (50 �g) adjuvant alone, or PBS. These mice were boostedtwo weeks later with the same immunization regimen. The infection was per-formed 4 weeks after the last vaccination by intradermal (i.d.) inoculation of1,000 metacyclic promastigotes of L. major (clone V1) into the dermis of bothears of the mouse. The evolution of the infection was monitored by measuringthe diameter of the induration of the ear lesion with a metric caliper.
BALB/c mice were s.c. inoculated in the right footpad with either 0.5, 2, or 10�g of rLiP0 plus 50 �g of CpG ODN (25 �g of each immunostimulatory ODN),CpG ODN (50 �g) adjuvant alone or PBS. Each group was boosted 2 weeks laterusing the same regimen. Parasite challenge was carried out by s.c. inoculationwith 5 104 stationary-phase promastigotes of L. major (clone WHOM/IR/-173)into the left (untreated) footpad four weeks after the last inoculation. Theprogress of the infection was followed by measuring the thickness with a metriccaliper. The contralateral footpad of each animal represented the control value,and the swelling was calculated as follows: thickness of the left footpad minusthickness of the right footpad. The animals were euthanized when the lesionsbecame necrotic.
Parasite quantitation. The number of parasites was determined in ear andfootpad by limiting dilution assay (3). Briefly, footpad sections and ears wererecovered from infected BALB/c and C57BL/6 mice, respectively. The ventraland dorsal sheets of the infected ears were separated. The footpad sections andear sheets were deposited in Dulbecco’s modified Eagle medium containingLiberase CI enzyme blend (50 �g ml�1). After 2 h of incubation at 37°C, thetissues were cut into small pieces, homogenized and filtered using a cell strainer(70-�m pore size). The homogenized tissue was serially diluted in a 96-wellflat-bottomed microtiter plate containing Schneider’s medium plus 20% FCS orbiphasic medium, prepared using 50 �l of NNN medium containing 30% defi-brinated rabbit blood and overlaid with 50 �l of M199/S as previously described(21). The number of viable parasites was determined from the highest dilution atwhich promastigotes could be grown up to 7 days of incubation at 26°C. Thenumber of parasites was also determined in the local draining lymph nodes(DLN) of infected ears (retromaxillar) and footpad (popliteal). The lymph nodeswere recovered, mechanically dissociated and then serially diluted as above. Inthe ear and LN the parasite load is expressed as the number of parasites in the
VOL. 73, 2005 PROTECTION AGAINST L. MAJOR BY rLiP0 PLUS CpG ODN 5843
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
whole organ. In the footpad the parasite load is expressed as the number ofparasites per mg tissue.
Measurement of cytokines in supernatants. Spleens and lymph nodes wereremoved aseptically from mice after cervical dislocation. Splenocytes and drain-ing lymph node cell (LNC) suspensions were seeded in complete RPMI medium(RPMI 1640 supplemented with 10% FCS, 2 mM glutamine, and 10 mM 2-mer-captoethanol). Next, 3 106 cells were seeded in 48-well plates during 72 h at 37°Cin the presence of rLiP0 (3 �g ml�1) or SLA (12 �g ml�1). The release of IFN-� andIL-4 was measured in the supernatants of splenocytes and LNC cultures, and wasdetermined by commercial enzyme-linked immunosorbent assay (ELISA) kits (Dia-clone, Besancon, France). In parallel, LNCs stimulated with rLiP0 were incubated inthe presence of 10 �g ml�1 of monoclonal antibody (MAb) against either mouseCD4 (GK 1.5), mouse IL-12 (C17.8), or mouse CD8 (53-6.7). Appropriate isotype-matched controls were also analyzed in the assay. The antibodies (no azide/lowendotoxin) were purchased from BD (PharMingen).
When bone marrow-derived dendritic cells (BMDDC) were used as antigen-presenting cells they were pulsed for 24 h with L. major amastigotes, at a parasiteto cell ratio of 5:1. Afterwards, dermal cell suspensions from infected mice werestimulated with L. major-infected or uninfected BMDDC (2 105) in completeRPMI medium. Then IFN-� production was analyzed in 48-h supernatants asdescribed above. For T-cell epitope determination, BMDDC (2 105) werepulsed for 16 h with either rLiP0 (12 �g ml�1) or each peptide (15 �g ml�1)covering the whole LiP0 sequence in U-bottom plates. Aferwards, BMDDC wereharvested, washed and used to stimulate splenocytes from LiP0-vaccinated mice.Cytokine production was analyzed in supernatants after 72 h.
Intracytoplasmic staining of IFN-�. For the analysis of the frequency ofT-cell-producing IFN-�, single-cell suspensions from four ears were cultured ina 24-well tissue plate. Subsequently, the dermal cells were stimulated for 16 hwith L. major-infected BMDDC, as described before. Then, brefeldin A (10 �gml�1) reagent was added to the culture and further incubated for an additionalperiod of 6 h. Afterwards, cells were harvested, washed twice in PBS with 1%FCS and incubated with an anti-FC� III/II receptor (FcBlock) prior to staining.Then cells were stained for the surface markers TCR� (H57–597; Cy-chromelabeled) and either CD4 (GK1.5; fluorescein isothiocyanate-labeled) or CD8(53-6.7, fluorescein isothiocyanate-conjugated) for 30 min on ice. Cells were then
washed twice and fixed for 20 min in Cytofix/Cytoperm buffer. Next, cells werewashed twice and incubated with phycoerythrin (PE)-conjugated rat anti-mouseIFN-� (XGM.1.2) for 30 min at 4°C. Finally cells were washed twice and ana-lyzed on a FACSCalibur flow cytometer. The specificity of the anti-cytokine MAbwas tested by both negative staining of nonpermeabilized cells and a PE-conju-gated isotype-matched control (R3–34). For each sample, at least 100,000 cellswere analyzed. The data were collected and analyzed using FACScalibur andCELLQuest software. The frequency of CD4� and CD8� T cells was determinedby gating on TCR��cells. All reagents and conjugate monoclonal antibodieswere purchased from Becton Dickinson (PharMingen, San Diego, CA).
Determination of antibody titers and isotypes. Serum samples were analyzedfor specific anti-LiP0 antibodies. Briefly, standard ELISA plates were coatedovernight at room temperature with 100 �l of rLiP0 (2 �g ml�1 in PBS). The titerwas determined by serial dilution of the sera, and was defined as the inverse ofthe highest serum dilution factor giving an absorbance of 0.2.
The Falcon assay screening test-ELISA (FAST-ELISA; Becton Dickinson)was used instead of the standard ELISA for the determination of the reactivityagainst the synthetic peptides covering the LiP0 sequence. The lids were coatedwith 100 �l of each synthetic peptide (100 �g ml�1) as described before (32). Theisotype-specific analyses were done with the following horseradish peroxidase-conjugated anti-mouse immunoglobulins (Igs) (Nordic Immunological Labora-tories, Tilburg, The Netherlands): anti-IgG (1:2,000), anti-IgG1 (1:1,000) andanti-IgG2a (1:500). Orthophenylene diamine dihydrochloride (OPD; Dako A/S,Glostrup, Denmark) was used as peroxidase substrate. After 15 min, the reactionwas stopped with the addition of 100 �l of H2SO4 (1 M) and the absorbance wasread at 450 nm.
Statistical analysis. Statistical analysis was performed by Student’s t test.Differences were considered significant when P � 0.05.
RESULTS
Vaccination with LiP0-DNA or rLiP0 plus CpG ODN con-fers protection against dermal pathology due to L. major chal-lenge in C57BL/6 mice. We have previously described that
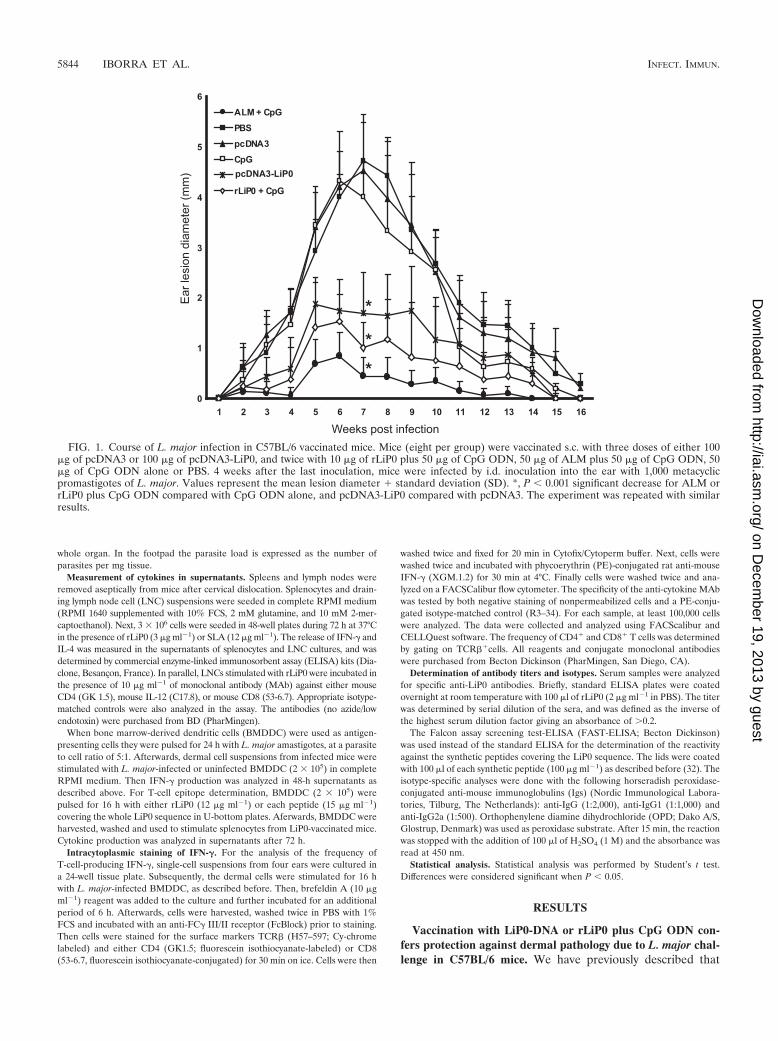
FIG. 1. Course of L. major infection in C57BL/6 vaccinated mice. Mice (eight per group) were vaccinated s.c. with three doses of either 100�g of pcDNA3 or 100 �g of pcDNA3-LiP0, and twice with 10 �g of rLiP0 plus 50 �g of CpG ODN, 50 �g of ALM plus 50 �g of CpG ODN, 50�g of CpG ODN alone or PBS. 4 weeks after the last inoculation, mice were infected by i.d. inoculation into the ear with 1,000 metacyclicpromastigotes of L. major. Values represent the mean lesion diameter � standard deviation (SD). *, P � 0.001 significant decrease for ALM orrLiP0 plus CpG ODN compared with CpG ODN alone, and pcDNA3-LiP0 compared with pcDNA3. The experiment was repeated with similarresults.
5844 IBORRA ET AL. INFECT. IMMUN.
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
vaccination with LiP0 DNA (pcDNA3-LiP0) partially protectsagainst L. major infection in BALB/c mice. Here, this antigenhas been tested, either as a DNA vaccine or as recombinantprotein (rLiP0) adjuvanted with CpG ODN, using low-doseintradermal challenge in C57BL/6 mice.
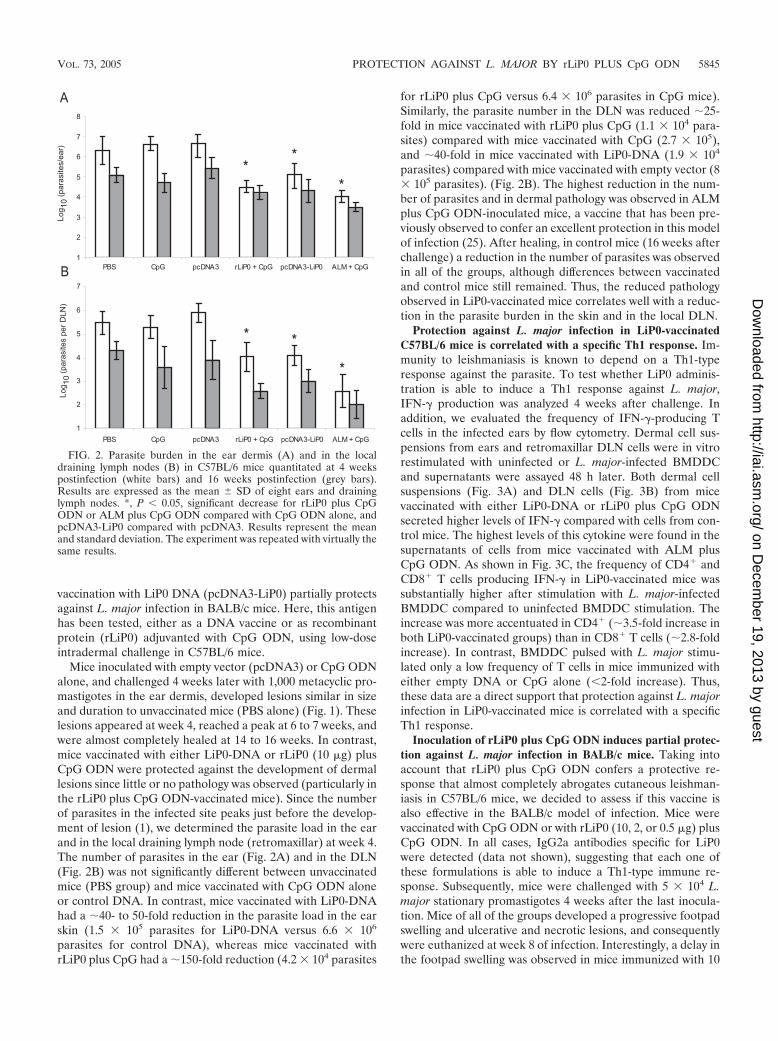
Mice inoculated with empty vector (pcDNA3) or CpG ODNalone, and challenged 4 weeks later with 1,000 metacyclic pro-mastigotes in the ear dermis, developed lesions similar in sizeand duration to unvaccinated mice (PBS alone) (Fig. 1). Theselesions appeared at week 4, reached a peak at 6 to 7 weeks, andwere almost completely healed at 14 to 16 weeks. In contrast,mice vaccinated with either LiP0-DNA or rLiP0 (10 �g) plusCpG ODN were protected against the development of dermallesions since little or no pathology was observed (particularly inthe rLiP0 plus CpG ODN-vaccinated mice). Since the numberof parasites in the infected site peaks just before the develop-ment of lesion (1), we determined the parasite load in the earand in the local draining lymph node (retromaxillar) at week 4.The number of parasites in the ear (Fig. 2A) and in the DLN(Fig. 2B) was not significantly different between unvaccinatedmice (PBS group) and mice vaccinated with CpG ODN aloneor control DNA. In contrast, mice vaccinated with LiP0-DNAhad a �40- to 50-fold reduction in the parasite load in the earskin (1.5 105 parasites for LiP0-DNA versus 6.6 106
parasites for control DNA), whereas mice vaccinated withrLiP0 plus CpG had a �150-fold reduction (4.2 104 parasites
for rLiP0 plus CpG versus 6.4 106 parasites in CpG mice).Similarly, the parasite number in the DLN was reduced �25-fold in mice vaccinated with rLiP0 plus CpG (1.1 104 para-sites) compared with mice vaccinated with CpG (2.7 105),and �40-fold in mice vaccinated with LiP0-DNA (1.9 104
parasites) compared with mice vaccinated with empty vector (8 105 parasites). (Fig. 2B). The highest reduction in the num-ber of parasites and in dermal pathology was observed in ALMplus CpG ODN-inoculated mice, a vaccine that has been pre-viously observed to confer an excellent protection in this modelof infection (25). After healing, in control mice (16 weeks afterchallenge) a reduction in the number of parasites was observedin all of the groups, although differences between vaccinatedand control mice still remained. Thus, the reduced pathologyobserved in LiP0-vaccinated mice correlates well with a reduc-tion in the parasite burden in the skin and in the local DLN.
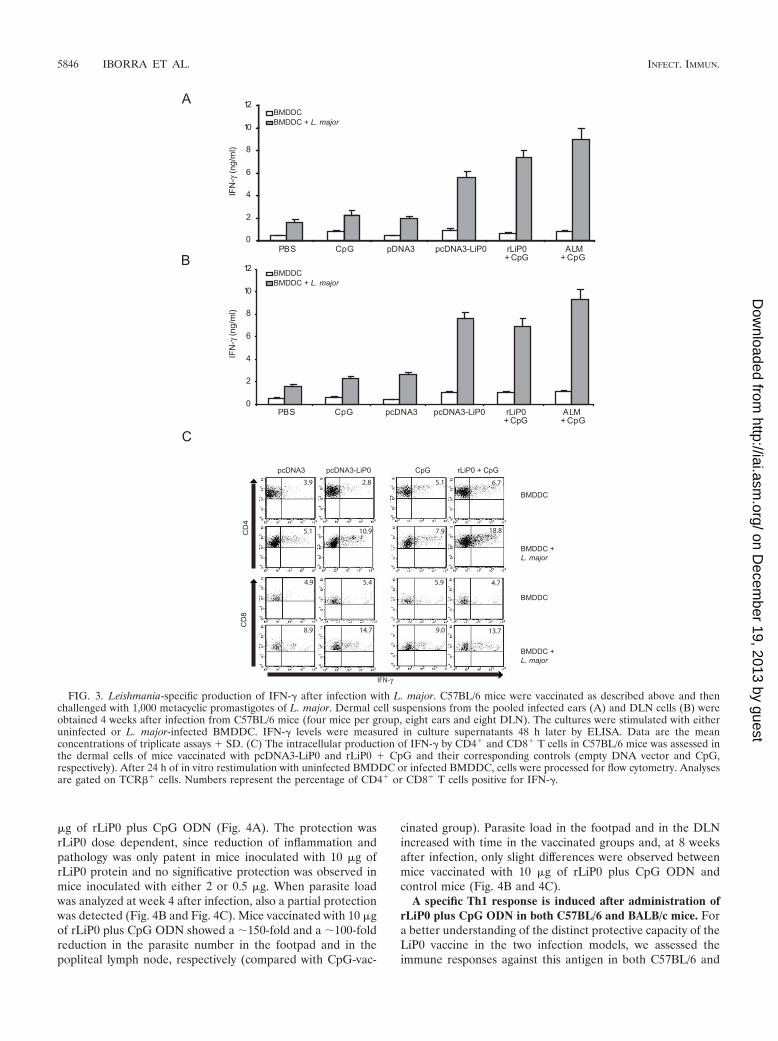
Protection against L. major infection in LiP0-vaccinatedC57BL/6 mice is correlated with a specific Th1 response. Im-munity to leishmaniasis is known to depend on a Th1-typeresponse against the parasite. To test whether LiP0 adminis-tration is able to induce a Th1 response against L. major,IFN-� production was analyzed 4 weeks after challenge. Inaddition, we evaluated the frequency of IFN-�-producing Tcells in the infected ears by flow cytometry. Dermal cell sus-pensions from ears and retromaxillar DLN cells were in vitrorestimulated with uninfected or L. major-infected BMDDCand supernatants were assayed 48 h later. Both dermal cellsuspensions (Fig. 3A) and DLN cells (Fig. 3B) from micevaccinated with either LiP0-DNA or rLiP0 plus CpG ODNsecreted higher levels of IFN-� compared with cells from con-trol mice. The highest levels of this cytokine were found in thesupernatants of cells from mice vaccinated with ALM plusCpG ODN. As shown in Fig. 3C, the frequency of CD4� andCD8� T cells producing IFN-� in LiP0-vaccinated mice wassubstantially higher after stimulation with L. major-infectedBMDDC compared to uninfected BMDDC stimulation. Theincrease was more accentuated in CD4� (�3.5-fold increase inboth LiP0-vaccinated groups) than in CD8� T cells (�2.8-foldincrease). In contrast, BMDDC pulsed with L. major stimu-lated only a low frequency of T cells in mice immunized witheither empty DNA or CpG alone (�2-fold increase). Thus,these data are a direct support that protection against L. majorinfection in LiP0-vaccinated mice is correlated with a specificTh1 response.
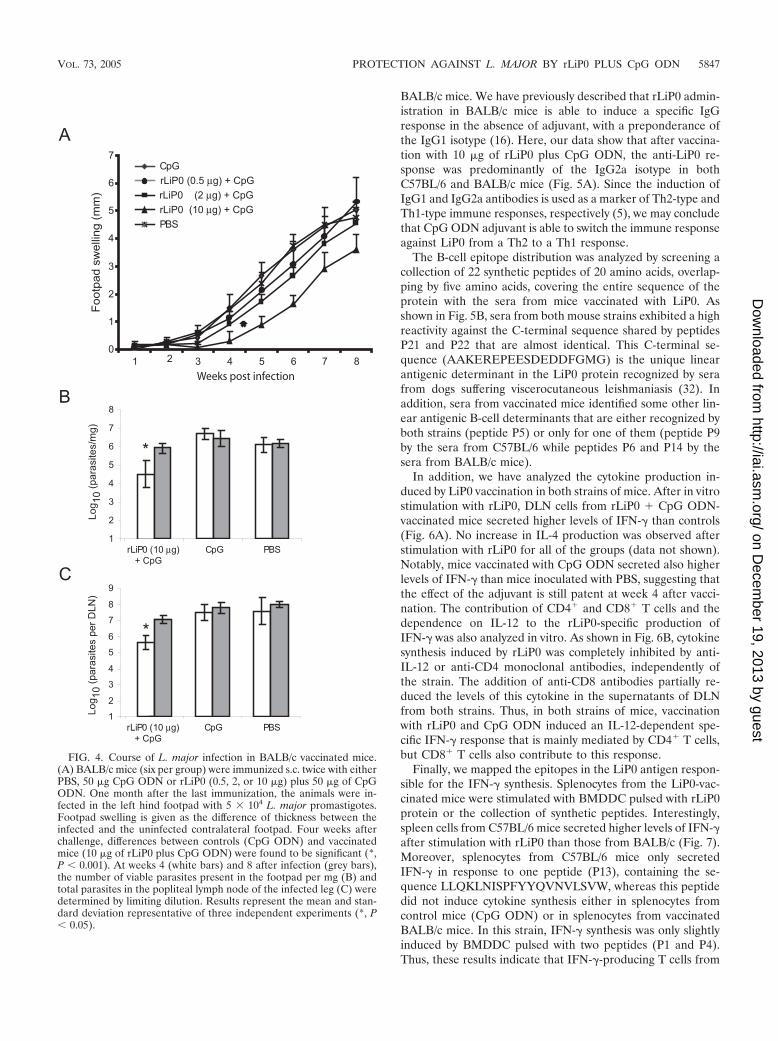
Inoculation of rLiP0 plus CpG ODN induces partial protec-tion against L. major infection in BALB/c mice. Taking intoaccount that rLiP0 plus CpG ODN confers a protective re-sponse that almost completely abrogates cutaneous leishman-iasis in C57BL/6 mice, we decided to assess if this vaccine isalso effective in the BALB/c model of infection. Mice werevaccinated with CpG ODN or with rLiP0 (10, 2, or 0.5 �g) plusCpG ODN. In all cases, IgG2a antibodies specific for LiP0were detected (data not shown), suggesting that each one ofthese formulations is able to induce a Th1-type immune re-sponse. Subsequently, mice were challenged with 5 104 L.major stationary promastigotes 4 weeks after the last inocula-tion. Mice of all of the groups developed a progressive footpadswelling and ulcerative and necrotic lesions, and consequentlywere euthanized at week 8 of infection. Interestingly, a delay inthe footpad swelling was observed in mice immunized with 10
FIG. 2. Parasite burden in the ear dermis (A) and in the localdraining lymph nodes (B) in C57BL/6 mice quantitated at 4 weekspostinfection (white bars) and 16 weeks postinfection (grey bars).Results are expressed as the mean � SD of eight ears and draininglymph nodes. *, P � 0.05, significant decrease for rLiP0 plus CpGODN or ALM plus CpG ODN compared with CpG ODN alone, andpcDNA3-LiP0 compared with pcDNA3. Results represent the meanand standard deviation. The experiment was repeated with virtually thesame results.
VOL. 73, 2005 PROTECTION AGAINST L. MAJOR BY rLiP0 PLUS CpG ODN 5845
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
�g of rLiP0 plus CpG ODN (Fig. 4A). The protection wasrLiP0 dose dependent, since reduction of inflammation andpathology was only patent in mice inoculated with 10 �g ofrLiP0 protein and no significative protection was observed inmice inoculated with either 2 or 0.5 �g. When parasite loadwas analyzed at week 4 after infection, also a partial protectionwas detected (Fig. 4B and Fig. 4C). Mice vaccinated with 10 �gof rLiP0 plus CpG ODN showed a �150-fold and a �100-foldreduction in the parasite number in the footpad and in thepopliteal lymph node, respectively (compared with CpG-vac-
cinated group). Parasite load in the footpad and in the DLNincreased with time in the vaccinated groups and, at 8 weeksafter infection, only slight differences were observed betweenmice vaccinated with 10 �g of rLiP0 plus CpG ODN andcontrol mice (Fig. 4B and 4C).
A specific Th1 response is induced after administration ofrLiP0 plus CpG ODN in both C57BL/6 and BALB/c mice. Fora better understanding of the distinct protective capacity of theLiP0 vaccine in the two infection models, we assessed theimmune responses against this antigen in both C57BL/6 and
FIG. 3. Leishmania-specific production of IFN-� after infection with L. major. C57BL/6 mice were vaccinated as described above and thenchallenged with 1,000 metacyclic promastigotes of L. major. Dermal cell suspensions from the pooled infected ears (A) and DLN cells (B) wereobtained 4 weeks after infection from C57BL/6 mice (four mice per group, eight ears and eight DLN). The cultures were stimulated with eitheruninfected or L. major-infected BMDDC. IFN-� levels were measured in culture supernatants 48 h later by ELISA. Data are the meanconcentrations of triplicate assays � SD. (C) The intracellular production of IFN-� by CD4� and CD8� T cells in C57BL/6 mice was assessed inthe dermal cells of mice vaccinated with pcDNA3-LiP0 and rLiP0 � CpG and their corresponding controls (empty DNA vector and CpG,respectively). After 24 h of in vitro restimulation with uninfected BMDDC or infected BMDDC, cells were processed for flow cytometry. Analysesare gated on TCR�� cells. Numbers represent the percentage of CD4� or CD8� T cells positive for IFN-�.
5846 IBORRA ET AL. INFECT. IMMUN.
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
BALB/c mice. We have previously described that rLiP0 admin-istration in BALB/c mice is able to induce a specific IgGresponse in the absence of adjuvant, with a preponderance ofthe IgG1 isotype (16). Here, our data show that after vaccina-tion with 10 �g of rLiP0 plus CpG ODN, the anti-LiP0 re-sponse was predominantly of the IgG2a isotype in bothC57BL/6 and BALB/c mice (Fig. 5A). Since the induction ofIgG1 and IgG2a antibodies is used as a marker of Th2-type andTh1-type immune responses, respectively (5), we may concludethat CpG ODN adjuvant is able to switch the immune responseagainst LiP0 from a Th2 to a Th1 response.
The B-cell epitope distribution was analyzed by screening acollection of 22 synthetic peptides of 20 amino acids, overlap-ping by five amino acids, covering the entire sequence of theprotein with the sera from mice vaccinated with LiP0. Asshown in Fig. 5B, sera from both mouse strains exhibited a highreactivity against the C-terminal sequence shared by peptidesP21 and P22 that are almost identical. This C-terminal se-quence (AAKEREPEESDEDDFGMG) is the unique linearantigenic determinant in the LiP0 protein recognized by serafrom dogs suffering viscerocutaneous leishmaniasis (32). Inaddition, sera from vaccinated mice identified some other lin-ear antigenic B-cell determinants that are either recognized byboth strains (peptide P5) or only for one of them (peptide P9by the sera from C57BL/6 while peptides P6 and P14 by thesera from BALB/c mice).
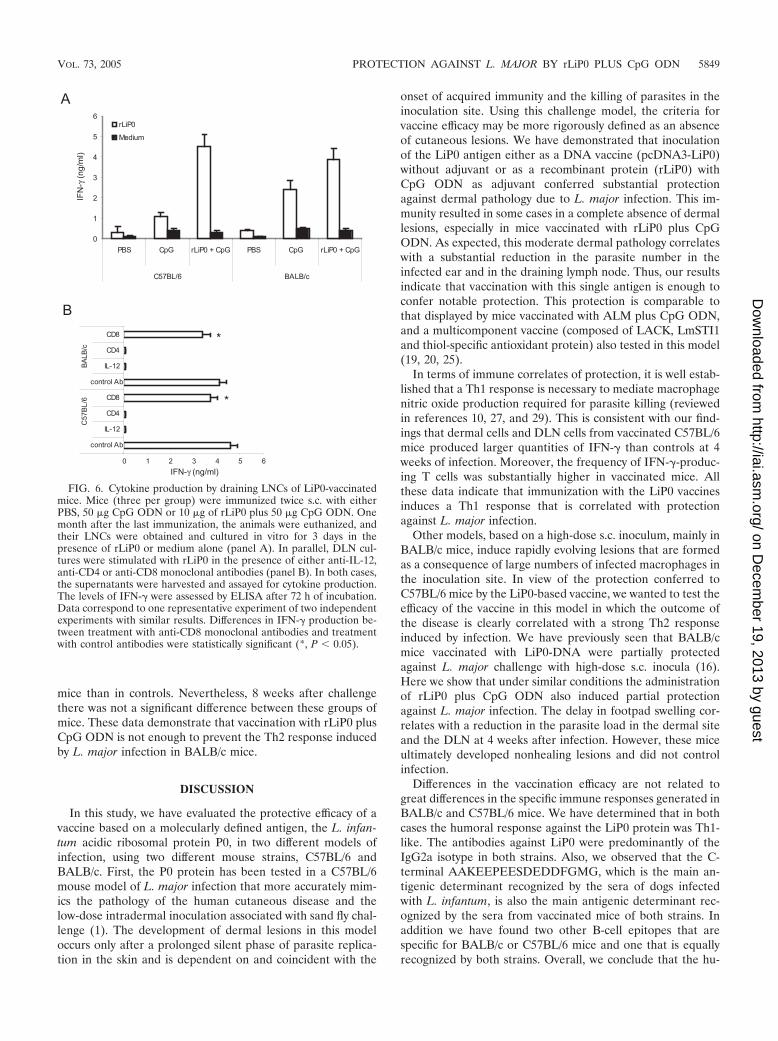
In addition, we have analyzed the cytokine production in-duced by LiP0 vaccination in both strains of mice. After in vitrostimulation with rLiP0, DLN cells from rLiP0 � CpG ODN-vaccinated mice secreted higher levels of IFN-� than controls(Fig. 6A). No increase in IL-4 production was observed afterstimulation with rLiP0 for all of the groups (data not shown).Notably, mice vaccinated with CpG ODN secreted also higherlevels of IFN-� than mice inoculated with PBS, suggesting thatthe effect of the adjuvant is still patent at week 4 after vacci-nation. The contribution of CD4� and CD8� T cells and thedependence on IL-12 to the rLiP0-specific production ofIFN-� was also analyzed in vitro. As shown in Fig. 6B, cytokinesynthesis induced by rLiP0 was completely inhibited by anti-IL-12 or anti-CD4 monoclonal antibodies, independently ofthe strain. The addition of anti-CD8 antibodies partially re-duced the levels of this cytokine in the supernatants of DLNfrom both strains. Thus, in both strains of mice, vaccinationwith rLiP0 and CpG ODN induced an IL-12-dependent spe-cific IFN-� response that is mainly mediated by CD4� T cells,but CD8� T cells also contribute to this response.
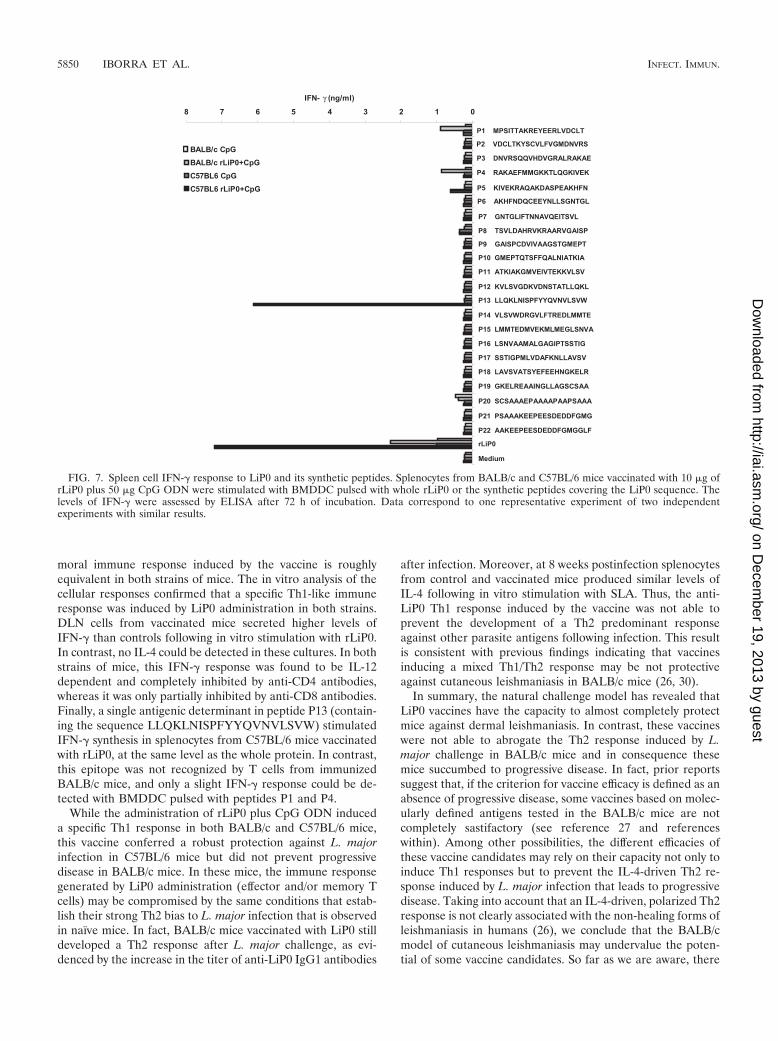
Finally, we mapped the epitopes in the LiP0 antigen respon-sible for the IFN-� synthesis. Splenocytes from the LiP0-vac-cinated mice were stimulated with BMDDC pulsed with rLiP0protein or the collection of synthetic peptides. Interestingly,spleen cells from C57BL/6 mice secreted higher levels of IFN-�after stimulation with rLiP0 than those from BALB/c (Fig. 7).Moreover, splenocytes from C57BL/6 mice only secretedIFN-� in response to one peptide (P13), containing the se-quence LLQKLNISPFYYQVNVLSVW, whereas this peptidedid not induce cytokine synthesis either in splenocytes fromcontrol mice (CpG ODN) or in splenocytes from vaccinatedBALB/c mice. In this strain, IFN-� synthesis was only slightlyinduced by BMDDC pulsed with two peptides (P1 and P4).Thus, these results indicate that IFN-�-producing T cells from
FIG. 4. Course of L. major infection in BALB/c vaccinated mice.(A) BALB/c mice (six per group) were immunized s.c. twice with eitherPBS, 50 �g CpG ODN or rLiP0 (0.5, 2, or 10 �g) plus 50 �g of CpGODN. One month after the last immunization, the animals were in-fected in the left hind footpad with 5 104 L. major promastigotes.Footpad swelling is given as the difference of thickness between theinfected and the uninfected contralateral footpad. Four weeks afterchallenge, differences between controls (CpG ODN) and vaccinatedmice (10 �g of rLiP0 plus CpG ODN) were found to be significant (*,P � 0.001). At weeks 4 (white bars) and 8 after infection (grey bars),the number of viable parasites present in the footpad per mg (B) andtotal parasites in the popliteal lymph node of the infected leg (C) weredetermined by limiting dilution. Results represent the mean and stan-dard deviation representative of three independent experiments (*, P� 0.05).
VOL. 73, 2005 PROTECTION AGAINST L. MAJOR BY rLiP0 PLUS CpG ODN 5847
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from

vaccinated mice recognized different antigenic determinants inthe LiP0 antigen, depending on the mouse strain.
Vaccination with rLiP0 plus CpG ODN does not prevent aTh2 response in BALB/c mice infected with L. major. SinceBALB/c mice vaccinated with LiP0 were partially protectedagainst L. major infection, we decided to analyze if this vaccinereversed the Th2 response induced by the parasite. First, weanalyzed the humoral response elicited against the LiP0 anti-gen in both strains of vaccinated mice 8 weeks after challenge.As shown in Fig. 8A, the antibodies against LiP0 were stillpredominantly IgG2a in C57BL/6 mice. In contrast, a mixed
IgG1/IgG2a anti-LiP0 response was observed in BALB/c mice,indicating that the L. major infection induced a Th2 responseagainst LiP0 in these mice. Second, we analyzed the IL-4 re-sponse 4 and 8 weeks after challenge. Splenocytes from controland vaccinated mice were in vitro re-stimulated with rLiP0 andSLA. No IL-4 was detected in C57BL/6 mice (data not shown).In addition, no IL-4 production could be detected in responseto rLiP0 stimulation in BALB/c mice, whereas SLA inducedIL-4 synthesis in both control and vaccinated mice (Fig. 8B).Concomitant with the early protection observed, at 4 weeksafter infection the levels of IL-4 were lower in the vaccinated
FIG. 5. Analysis of the specific humoral response induced in BALB/c and C57BL/6 mice immunized with 10 �g of rLiP0 plus CpG ODN. Onemonth after the second inoculation, mice (n 6) were bled and sera were tested by ELISA for specific anti-LiP0 antibody responses of both IgG1and IgG2a isotypes (A). Each sample was assayed separately, and the titer was determined as indicated in the Materials and Methods section. Thereactivity of these sera against the synthetic peptides covering the LiP0 sequence was analyzed by FAST-ELISA (B). The optical density at 450nm (OD450) values represent the reactivities obtained with the pooled sera diluted 1:500 using as second antibody a horseradish peroxidase-conjugated anti-IgG at 1:2,000 dilution. None of the preimmune, the PBS or the CpG ODN control sera showed reactivity against either therecombinant protein or the synthetic peptides (not shown).
5848 IBORRA ET AL. INFECT. IMMUN.
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
mice than in controls. Nevertheless, 8 weeks after challengethere was not a significant difference between these groups ofmice. These data demonstrate that vaccination with rLiP0 plusCpG ODN is not enough to prevent the Th2 response inducedby L. major infection in BALB/c mice.
DISCUSSION
In this study, we have evaluated the protective efficacy of avaccine based on a molecularly defined antigen, the L. infan-tum acidic ribosomal protein P0, in two different models ofinfection, using two different mouse strains, C57BL/6 andBALB/c. First, the P0 protein has been tested in a C57BL/6mouse model of L. major infection that more accurately mim-ics the pathology of the human cutaneous disease and thelow-dose intradermal inoculation associated with sand fly chal-lenge (1). The development of dermal lesions in this modeloccurs only after a prolonged silent phase of parasite replica-tion in the skin and is dependent on and coincident with the
onset of acquired immunity and the killing of parasites in theinoculation site. Using this challenge model, the criteria forvaccine efficacy may be more rigorously defined as an absenceof cutaneous lesions. We have demonstrated that inoculationof the LiP0 antigen either as a DNA vaccine (pcDNA3-LiP0)without adjuvant or as a recombinant protein (rLiP0) withCpG ODN as adjuvant conferred substantial protectionagainst dermal pathology due to L. major infection. This im-munity resulted in some cases in a complete absence of dermallesions, especially in mice vaccinated with rLiP0 plus CpGODN. As expected, this moderate dermal pathology correlateswith a substantial reduction in the parasite number in theinfected ear and in the draining lymph node. Thus, our resultsindicate that vaccination with this single antigen is enough toconfer notable protection. This protection is comparable tothat displayed by mice vaccinated with ALM plus CpG ODN,and a multicomponent vaccine (composed of LACK, LmSTI1and thiol-specific antioxidant protein) also tested in this model(19, 20, 25).
In terms of immune correlates of protection, it is well estab-lished that a Th1 response is necessary to mediate macrophagenitric oxide production required for parasite killing (reviewedin references 10, 27, and 29). This is consistent with our find-ings that dermal cells and DLN cells from vaccinated C57BL/6mice produced larger quantities of IFN-� than controls at 4weeks of infection. Moreover, the frequency of IFN-�-produc-ing T cells was substantially higher in vaccinated mice. Allthese data indicate that immunization with the LiP0 vaccinesinduces a Th1 response that is correlated with protectionagainst L. major infection.
Other models, based on a high-dose s.c. inoculum, mainly inBALB/c mice, induce rapidly evolving lesions that are formedas a consequence of large numbers of infected macrophages inthe inoculation site. In view of the protection conferred toC57BL/6 mice by the LiP0-based vaccine, we wanted to test theefficacy of the vaccine in this model in which the outcome ofthe disease is clearly correlated with a strong Th2 responseinduced by infection. We have previously seen that BALB/cmice vaccinated with LiP0-DNA were partially protectedagainst L. major challenge with high-dose s.c. inocula (16).Here we show that under similar conditions the administrationof rLiP0 plus CpG ODN also induced partial protectionagainst L. major infection. The delay in footpad swelling cor-relates with a reduction in the parasite load in the dermal siteand the DLN at 4 weeks after infection. However, these miceultimately developed nonhealing lesions and did not controlinfection.
Differences in the vaccination efficacy are not related togreat differences in the specific immune responses generated inBALB/c and C57BL/6 mice. We have determined that in bothcases the humoral response against the LiP0 protein was Th1-like. The antibodies against LiP0 were predominantly of theIgG2a isotype in both strains. Also, we observed that the C-terminal AAKEEPEESDEDDFGMG, which is the main an-tigenic determinant recognized by the sera of dogs infectedwith L. infantum, is also the main antigenic determinant rec-ognized by the sera from vaccinated mice of both strains. Inaddition we have found two other B-cell epitopes that arespecific for BALB/c or C57BL/6 mice and one that is equallyrecognized by both strains. Overall, we conclude that the hu-
FIG. 6. Cytokine production by draining LNCs of LiP0-vaccinatedmice. Mice (three per group) were immunized twice s.c. with eitherPBS, 50 �g CpG ODN or 10 �g of rLiP0 plus 50 �g CpG ODN. Onemonth after the last immunization, the animals were euthanized, andtheir LNCs were obtained and cultured in vitro for 3 days in thepresence of rLiP0 or medium alone (panel A). In parallel, DLN cul-tures were stimulated with rLiP0 in the presence of either anti-IL-12,anti-CD4 or anti-CD8 monoclonal antibodies (panel B). In both cases,the supernatants were harvested and assayed for cytokine production.The levels of IFN-� were assessed by ELISA after 72 h of incubation.Data correspond to one representative experiment of two independentexperiments with similar results. Differences in IFN-� production be-tween treatment with anti-CD8 monoclonal antibodies and treatmentwith control antibodies were statistically significant (*, P � 0.05).
VOL. 73, 2005 PROTECTION AGAINST L. MAJOR BY rLiP0 PLUS CpG ODN 5849
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
moral immune response induced by the vaccine is roughlyequivalent in both strains of mice. The in vitro analysis of thecellular responses confirmed that a specific Th1-like immuneresponse was induced by LiP0 administration in both strains.DLN cells from vaccinated mice secreted higher levels ofIFN-� than controls following in vitro stimulation with rLiP0.In contrast, no IL-4 could be detected in these cultures. In bothstrains of mice, this IFN-� response was found to be IL-12dependent and completely inhibited by anti-CD4 antibodies,whereas it was only partially inhibited by anti-CD8 antibodies.Finally, a single antigenic determinant in peptide P13 (contain-ing the sequence LLQKLNISPFYYQVNVLSVW) stimulatedIFN-� synthesis in splenocytes from C57BL/6 mice vaccinatedwith rLiP0, at the same level as the whole protein. In contrast,this epitope was not recognized by T cells from immunizedBALB/c mice, and only a slight IFN-� response could be de-tected with BMDDC pulsed with peptides P1 and P4.
While the administration of rLiP0 plus CpG ODN induceda specific Th1 response in both BALB/c and C57BL/6 mice,this vaccine conferred a robust protection against L. majorinfection in C57BL/6 mice but did not prevent progressivedisease in BALB/c mice. In these mice, the immune responsegenerated by LiP0 administration (effector and/or memory Tcells) may be compromised by the same conditions that estab-lish their strong Th2 bias to L. major infection that is observedin naıve mice. In fact, BALB/c mice vaccinated with LiP0 stilldeveloped a Th2 response after L. major challenge, as evi-denced by the increase in the titer of anti-LiP0 IgG1 antibodies
after infection. Moreover, at 8 weeks postinfection splenocytesfrom control and vaccinated mice produced similar levels ofIL-4 following in vitro stimulation with SLA. Thus, the anti-LiP0 Th1 response induced by the vaccine was not able toprevent the development of a Th2 predominant responseagainst other parasite antigens following infection. This resultis consistent with previous findings indicating that vaccinesinducing a mixed Th1/Th2 response may be not protectiveagainst cutaneous leishmaniasis in BALB/c mice (26, 30).
In summary, the natural challenge model has revealed thatLiP0 vaccines have the capacity to almost completely protectmice against dermal leishmaniasis. In contrast, these vaccineswere not able to abrogate the Th2 response induced by L.major challenge in BALB/c mice and in consequence thesemice succumbed to progressive disease. In fact, prior reportssuggest that, if the criterion for vaccine efficacy is defined as anabsence of progressive disease, some vaccines based on molec-ularly defined antigens tested in the BALB/c mice are notcompletely sastifactory (see reference 27 and referenceswithin). Among other possibilities, the different efficacies ofthese vaccine candidates may rely on their capacity not only toinduce Th1 responses but to prevent the IL-4-driven Th2 re-sponse induced by L. major infection that leads to progressivedisease. Taking into account that an IL-4-driven, polarized Th2response is not clearly associated with the non-healing forms ofleishmaniasis in humans (26), we conclude that the BALB/cmodel of cutaneous leishmaniasis may undervalue the poten-tial of some vaccine candidates. So far as we are aware, there
FIG. 7. Spleen cell IFN-� response to LiP0 and its synthetic peptides. Splenocytes from BALB/c and C57BL/6 mice vaccinated with 10 �g ofrLiP0 plus 50 �g CpG ODN were stimulated with BMDDC pulsed with whole rLiP0 or the synthetic peptides covering the LiP0 sequence. Thelevels of IFN-� were assessed by ELISA after 72 h of incubation. Data correspond to one representative experiment of two independentexperiments with similar results.
5850 IBORRA ET AL. INFECT. IMMUN.
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from

are only three studies that have tested vaccine candidates inthe ear model of infection in C57BL/6 mice (19, 20, 25). Thus,we think that further studies in this suitable model will con-tribute to better characterize the protective efficacy of poten-tial molecularly defined vaccines against this disease.
ACKNOWLEDGMENTS
This work was supported by grant BIO2002-04049-C02-C1 fromMinisterio de Ciencia y Tecnologıa from Spain. Also, an institutionalgrant from Fundacion Ramon Areces is acknowledged.
REFERENCES
1. Belkaid, Y., S. Kamhawi, G. Modi, J. Valenzuela, N. Noben-Trauth, E.Rowton, J. Ribeiro, and D. L. Sacks. 1998. Development of a natural modelof cutaneous leishmaniasis: powerful effects of vector saliva and saliva pre-exposure on the long-term outcome of Leishmania major infection in themouse ear dermis. J. Exp. Med. 188:1941–1953.
2. Belkaid, Y., E. Von Stebut, S. Mendez, R. Lira, E. Caler, S. Bertholet, M. C.Udey, and D. Sacks. 2002. CD8� T cells are required for primary immunityin C57BL/6 mice following low-dose, intradermal challenge with Leishmaniamajor. J. Immunol. 168:3992–4000.
3. Buffet, P. A., A. Sulahian, Y. J. Garin, N. Nassar, and F. Derouin. 1995.Culture microtitration: a sensitive method for quantifying Leishmania infan-tum in tissues of infected mice. Antimicrob. Agents Chemother. 39:2167–2168.
4. Cho, H. J., K. Takabayashi, P. M. Cheng, M. D. Nguyen, M. Corr, S. Tuck,and E. Raz. 2000. Immunostimulatory DNA-based vaccines induce cytotoxic
lymphocyte activity by a T-helper cell-independent mechanism. Nat. Bio-technol. 18:509–514.
5. Coffman, R. L. 1993. Mechanisms of helper T-cell regulation of B-cell ac-tivity. Ann. N. Y. Acad. Sci. 681:25–28.
6. Croft, S. L., and G. H. Coombs. 2003. Leishmaniasis, current chemotherapyand recent advances in the search for novel drugs. Trends Parasitol. 19:502–508.
7. Datta, N., S. Mukherjee, L. Das, and P. K. Das. 2003. Targeting of immu-nostimulatory DNA cures experimental visceral leishmaniasis through nitricoxide up-regulation and T cell activation. Eur. J. Immunol. 33:1508–1518.
8. Davila, E., and E. Celis. 2000. Repeated administration of cytosine-phos-phorothiolated guanine-containing oligonucleotides together with peptide/protein immunization results in enhanced CTL responses with anti-tumoractivity. J. Immunol. 165:539–547.
9. Greenblatt, C. L. 1980. The present and future of vaccination for cutaneousleishmaniasis. Prog. Clin. Biol. Res. 47:259–285.
10. Gumy, A., J. A. Louis, and P. Launois. 2004. The murine model of infectionwith Leishmania major and its importance for the deciphering of mechanismsunderlying differences in Th cell differentiation in mice from different ge-netic backgrounds. Int. J. Parasitol. 34:433–444.
11. Gurunathan, S., D. L. Sacks, D. R. Brown, S. L. Reiner, H. Charest, N.Glaichenhaus, and R. A. Seder. 1997. Vaccination with DNA encoding theimmunodominant LACK parasite antigen confers protective immunity tomice infected with Leishmania major. J. Exp. Med. 186:1137–1147.
12. Handman, E. 2001. Leishmaniasis: current status of vaccine development.Clin. Microbiol. Rev. 14:229–243.
13. Hartmann, G., G. J. Weiner, and A. M. Krieg. 1999. CpG DNA: a potentsignal for growth, activation, and maturation of human dendritic cells. Proc.Natl. Acad. Sci. USA 96:9305–9310.
14. Hashemi-Fesharki, R., S. Alle-Agha, P. Ahourai, E. Djavadian, A. Taghavi,A. Golabi, M. Kamail, K. Esmail-Nia, H. Ghafari, F. Mofasali, and R.Kheirol-Omour. 1992. Vaccine preparation and quality control of killedLeishmania major. Arch. Inst. Razi 43:39–50.
15. Heinzel, F. P., D. S. Schoenhaut, R. M. Rerko, L. E. Rosser, and M. K.Gately. 1993. Recombinant interleukin 12 cures mice infected with Leish-mania major. J. Exp. Med. 177:1505–1509.
16. Iborra, S., M. Soto, J. Carrion, A. Nieto, E. Fernandez, C. Alonso, and J. M.Requena. 2003. The Leishmania infantum acidic ribosomal protein P0 ad-ministered as a DNA vaccine confers protective immunity to Leishmaniamajor infection in BALB/c mice. Infect. Immun. 71:6562–6572.
17. Kellina, O. I. 1981. Problem and current lines in investigations on theepidemiology of leishmaniasis and its control in the U.S.S.R. Bull. Soc.Pathol. Exot. Filiales 74:306–318.
18. Klinman, D. M., A. K. Yi, S. L. Beaucage, J. Conover, and A. M. Krieg. 1996.CpG motifs present in bacteria DNA rapidly induce lymphocytes to secreteinterleukin 6, interleukin 12, and interferon gamma. Proc. Natl. Acad. Sci.USA 93:2879–2883.
19. Mendez, S., Y. Belkaid, R. A. Seder, and D. Sacks. 2002. Optimization ofDNA vaccination against cutaneous leishmaniasis. Vaccine 20:3702–3708.
20. Mendez, S., S. Gurunathan, S. Kamhawi, Y. Belkaid, M. A. Moga, Y. A.Skeiky, A. Campos-Neto, S. Reed, R. A. Seder, and D. Sacks. 2001. Thepotency and durability of DNA- and protein-based vaccines against Leish-mania major evaluated using low-dose, intradermal challenge. J. Immunol.166:5122–5128.
21. Mendez, S., K. Tabbara, Y. Belkaid, S. Bertholet, D. Verthelyi, D. Klinman,R. A. Seder, and D. L. Sacks. 2003. Coinjection with CpG-containing immu-nostimulatory oligodeoxynucleotides reduces the pathogenicity of a live vac-cine against cutaneous leishmaniasis but maintains its potency and durability.Infect. Immun. 71:5121–5129.
22. Molano, I., M. G. Alonso, C. Miron, E. Redondo, J. M. Requena, M. Soto,C. G. Nieto, and C. Alonso. 2003. A Leishmania infantum multi-componentantigenic protein mixed with live BCG confers protection to dogs experi-mentally infected with L. infantum. Vet. Immunol. Immunopathol. 92:1–13.
23. Nadim, A., E. Javadian, G. Tahvildar-Bidruni, and M. Ghorbani. 1983.Effectiveness of leishmanization in the control of cutaneous leishmaniasis.Bull. Soc. Pathol. Exot. Filiales 76:377–383.
24. Requena, J. M., S. Iborra, J. Carrion, C. Alonso, and M. Soto. 2004. Recentadvances in vaccines for leishmaniasis. Expert Opin. Biol. Ther. 4:1505–1517.
25. Rhee, E. G., S. Mendez, J. A. Shah, C. Y. Wu, J. R. Kirman, T. N. Turon,D. F. Davey, H. Davis, D. M. Klinman, R. N. Coler, D. L. Sacks, and R. A.Seder. 2002. Vaccination with heat-killed Leishmania antigen or recombi-nant leishmanial protein and CpG oligodeoxynucleotides induces long-termmemory CD4� and CD8� T cell responses and protection against Leishma-nia major infection. J. Exp. Med. 195:1565–1573.
26. Sacks, D., and C. Anderson. 2004. Re-examination of the immunosuppres-sive mechanisms mediating non-cure of Leishmania infection in mice. Im-munol. Rev. 201:225–238.
27. Sacks, D., and N. Noben-Trauth. 2002. The immunology of susceptibility andresistance to Leishmania major in mice. Nat. Rev. Immunol. 2:845–858.
28. Sadick, M. D., F. P. Heinzel, B. J. Holaday, R. T. Pu, R. S. Dawkins, and
FIG. 8. Analysis of the Th2 response after L. major infection.(A) Eight weeks after parasite challenge vaccinated BALB/c andC57BL/6 mice (n 5) were bled and sera were tested by ELISA forspecific anti-LiP0 antibody responses of both IgG1 and IgG2a isotypes.The titers of the anti-rLiP0 antibodies in the BALB/c CpG control miceat week 8 after challenge were 1/2,700 and 1/900 for IgG1 and IgG2a,respectively (not shown). (B) At weeks 4 and 8 after the L. majorchallenge, the animals were euthanized, and their splenocytes wereobtained and cultured in vitro for 3 days in the presence of rLiP0 orSLA. The supernatants were harvested and assayed for IL-4 by ELISA.
VOL. 73, 2005 PROTECTION AGAINST L. MAJOR BY rLiP0 PLUS CpG ODN 5851
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from

R. M. Locksley. 1990. Cure of murine leishmaniasis with anti-interleukin 4monoclonal antibody. Evidence for a T cell-dependent, interferon gamma-independent mechanism. J. Exp. Med. 171:115–127.
29. Scott, P., D. Artis, J. Uzonna, and C. Zaph. 2004. The development ofeffector and memory T cells in cutaneous leishmaniasis: the implications forvaccine development. Immunol. Rev. 201:318–338.
30. Sjolander, A., T. M. Baldwin, J. M. Curtis, and E. Handman. 1998. Inductionof a Th1 immune response and simultaneous lack of activation of a Th2response are required for generation of immunity to leishmaniasis. J. Im-munol. 160:3949–3957.
31. Skeiky, Y. A., D. R. Benson, M. Elwasila, R. Badaro, J. M. Burns, Jr., andS. G. Reed. 1994. Antigens shared by Leishmania species and Trypanosomacruzi: immunological comparison of the acidic ribosomal P0 proteins. Infect.Immun. 62:1643–1651.
32. Soto, M., J. M. Requena, L. Quijada, F. Guzman, M. E. Patarroyo, and C.Alonso. 1995. Identification of the Leishmania infantum P0 ribosomal pro-tein epitope in canine visceral leishmaniasis. Immunol. Lett. 48:23–28.
33. Sparwasser, T., and G. B. Lipford. 2000. Consequences of bacterial CpGDNA-driven activation of antigen-presenting cells. Curr. Top. Microbiol.Immunol. 247:59–75.
34. Sparwasser, T., R. M. Vabulas, B. Villmow, G. B. Lipford, and H. Wagner.2000. Bacterial CpG-DNA activates dendritic cells in vivo: T helper cell-independent cytotoxic T cell responses to soluble proteins. Eur. J. Immunol.30:3591–3597.
35. Zimmermann, S., O. Egeter, S. Hausmann, G. B. Lipford, M. Rocken, H.Wagner, and K. Heeg. 1998. CpG oligodeoxynucleotides trigger protectiveand curative Th1 responses in lethal murine leishmaniasis. J. Immunol.160:3627–3630.
Editor: W. A. Petri, Jr.
5852 IBORRA ET AL. INFECT. IMMUN.
on Decem
ber 19, 2013 by guesthttp://iai.asm
.org/D
ownloaded from




















