
Essential function of HIPK2 in TGFβ-dependent survival of midbrain dopamine neurons

Ž .Biochimica et Biophysica Acta 1353 1997 241–252
Antisense S-oligodeoxynucleotides down-regulate TGFb-production byKupffer cells from CCl -injured rat livers4
Juan Armendariz-Borunda a,), Leighton LeGros Jr. b, Octavio Campollo a, Arturo Panduro a,´Ana Rosa Rincon a´
a Institute of Molecular Biology in Medicine, CUCS, UniÕersity of Guadalajara, Apdo. Postal 2-500, Guadalajara, Jal, Mexico 44281b Veterans Administration Medical Center, Memphis, TN, USA
Received 3 March 1997; accepted 21 March 1997
Abstract
TGFb is a pleiotropic cytokine involved in multiple physiological and pathophysiological regulatory mechanisms. SinceTGFb is a disparate modulator of cell recruitment, proliferation and extracellular matrix phenotype for mesenchymal andnonmesenchymal cells, we have been investigating the role of this cytokine in the pathophysiology of liver. In the presentpaper we investigate which hepatic cell types from CCl -injured rat livers express TGFb mRNA and produce TGFb in4
culture, with the aim of further obliterating its biological activity by means of antisense technology. We performed a seriesof comprehensive molecular studies of in situ hybridization, northern blots, and RT-PCR and we found that onlynon-parenchymal cells produce TGFb while its expression in hepatocytes was absent. Consistent with the in situhybridization findings, we observed that Kupffer cells expressed high steady-state levels of TGFb mRNA, while circulatingmonocytes expressed a smaller amount of TGFb transcripts. We did not detect TGFb gene expression in endothelial cells.
w3 xThese findings were further confirmed by RT-PCR analyses. TGFb activity, as measured by inhibition of H thymidineincorporation by Mv 1 Lu mink lung epithelial cells, was down-regulated in culture by antisense phosphorothioateoligonucleotides. These effects of antisense oligomers were dose-dependent and the sense oligonucleotides had no effect atthe same concentration. q 1997 Elsevier Science B.V.
Keywords: Kupffer cell; Transforming growth factor b ; In situ hybridization; Gene expression regulation; Antisense oligomer; RT-PCR
1. Introduction
TGFb is a member of a steadily growing family ofgrowth factors whose biological functions are multi-ple and disparate depending on the target cell onwhich they act. TGFb 1 is the best characterized andis considered the prototype of this family of cy-
) Ž .Corresponding author. Fax: q52 3 617 4159; E-mail:[email protected]
w xtokines 1 . TGFb receptors are present in virtuallyall cells of the body and TGFb 1 can bind to itsreceptor only as a biologically active molecule, i.e. asa 25-kDa homodimer after the latency associated
w xpeptide has been clipped off 2–4 . TGFb plays acentral role in initiating, modulating, and terminating
Ž .the process of normal repair wound healing follow-ing all forms of tissue injury. TGFb also acts to limitor control inflammation through its immunoregula-
w xtory properties 5,6 . Platelets contain high concentra-
0167-4781r97r$17.00 q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0167-4781 97 00066-3

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´242
tions of TGFb and PDGF and, upon degranulation,release these cytokines into the tissue at the site of
w xinjury 7 . TGFb autoinduces its own production bycells and thus strongly amplifies its biological effects.It then promotes and stimulates the production anddeposition of extracellular matrix proteins, chemoat-tracts inflammatory cells and fibroblasts to the site ofinjury. Here, as these cells encounter higher concen-trations of TGFb , they become activated by mecha-nisms which remain to be elucidated. It is believedthat the strong chemotactic properties displayed byTGFb are probably due to up-regulation of variousadhesion receptors, such as the integrins, which thesecells use following a chemoattractant gradient to
w xmigrate to the site of injury 7 . In this regard, wehave used an experimental model to understand thepivotal role of TGFb in mediating inflammation and
Ž .overproduction of extracellular matrix proteins ECMafter the rat liver has been injured either acutely orchronically by the hepatotoxin CCl . We have re-4
Žcently shown that nonparenchymal cells specifically.Kupffer cells from CCl -treated rat livers accumu-4
late increased levels of TGFb transcripts during thew xinflammatory phase of liver damage 5,8 . Accord-
ingly with other investigators, we have also shownthat TGFb strongly up-regulates the transcription ofthe type I collagen in hepatic stellate cells obtainedfrom cirrhotic animals and that this up-regulationdepends on the previous induction of transcriptionalfactors like c-Fos and c-Jun that bind to a cognateAP-1 binding site situated within the enhancer of the
w xhuman type I collagen gene 9 . We and others havealso shown that during rat liver fibrosis induced byCCl , TGFb gene expression takes place early and is4
sustained throughout the process, always correlatingwith the gene expression for several collagens includ-
w xing collagen types I, III and IV 8,10 . The concentra-tion of TGFb 1 is also increased in other models ofexperimental liver fibrosis, such as murine schistoso-miasis and following hepatic irradiation, and also in
w xcirrhotic human livers 11–13 . It is clear, therefore,that an experimental strategy to obliterate the in vitroandror in vivo gene expression of an excess ofTGFb becomes attractive. A number of studies haveused phosphorothioate antisense oligodeoxynu-cleotides specifically targeted against coding se-
wquences of TGFb 1 mRNA and TGFb 2 mRNA 14–x17 . In this case, TGFb 1 antisense oligomer can be
directed to be complementary to the translationalinitiation site, the first seven codons for the N-termi-nal region of active TGFb 1 or 3X end of the mRNAw x14–17 . These various strategies to prevent the se-cretion or activity of TGFb 1 rendered different de-grees of inhibition. Using a different experimentalapproach, investigators have employed, as an anti-sense oligomer, a complete TGFb 1 cDNA that isspecific for TGFb 1 genomic DNA. This TGFb 1antisense oligomer, complementary to the nucleotidesflanking the first transcription start site of the humanTGFb 1 gene and phosphorothioate modified, wasefficacious in constraining TGFb 1 promoter activity
w xand reducing TGFb 1 secretion 18 . In the light ofthese findings, we reasoned that inactivation or neu-tralization of TGFb 1 production at the mRNA levelin an experimental model of hepatic cirrhosis, wouldrender a promising approach to modulate the exacer-bated in situ actions of TGFb 1 when this cytokine isproduced in uncontrolled fashion. This excess ofTGFb would be produced by the activated cellspresent at the site of the injury in the liver. Usingsuch an experimental model, we demonstrate herethat TGFb 1 production in liver cells derived fromCCl -treated animals, is mainly, if not only, sup-4
p o r te d b y r e s id e n t a n d in c o m in gKupfferrmonocytic-macrophagic cells. Furthermore,we could abrogate the biological activity of TGFb 1Ž .determined by a mink lung cells inhibition assay inactivated cells by using antisense technology.
2. Materials and methods
2.1. Fractionation of liÕer cells and in Õitro cultiÕa-tion
Detailed protocols for the isolation and cultivationw xof liver cells have been described earlier 8 . Hepato-
cytes and nonparenchymal cells were isolated by atwo-step sequential perfusion procedure by using col-
w xlagenase and pronase 19 . Cell suspensions werewashed and enriched cell populations were obtained
Ž .virtually free of cross-contamination see Fig. 1 . Inorder to fractionate the cell components of NPC, cellswere layered over a three-step gradient consisting of
Ž .8, 13 and 18% vrv metrizamide and centrifuged at

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´ 243
700=g for 17 min. Fraction enriched in Kupffercells was collected from the bottom layer while theendothelial cell-enriched fraction sedimented in themiddle layer. Metrizamide was removed from cellsrepresenting different fractions by washing twice with
´Hanks balanced salt solution. Cells were then smearedonto slides for morphological and histochemical anal-ysis. Peripheral blood monocytes were purified fromheparinized blood of CCl -treated rats by density4
gradient centrifugation on Ficoll–Hypaque.
2.2. Histochemistry
After fixation of the cells in cold acetone, stainingprocedures were performed. Staining for nonspecificesterase was done according to a previously de-
w xscribed method 20 ; cell-associated factor VIII wasdetected by immunostaining with rabbit anti-human
Žfactor VIII polyclonal antibody Dako Corporation,.St. Barbara, CA, USA by the peroxidase–antiperoxi-
dase staining technique according to the instructionsof the supplier. Intracellular vimentin was detected bythe indirect immunofluorescence staining method us-ing mouse anti-vimentin monoclonal antibodiesŽ .Labsystems, Helsinki, Finland .
2.3. In situ localization of TGFb transcripts in liÕercells
Radiolabeled riboprobes in sense and antisenseorientations were used for in situ hybridization. We
w xcloned a full length chicken b-actin cDNA 5,21Žinto a unique Pst I site of pSP6rT7–19 Bethesda
.Research Laboratories . A different cDNA encodingw xcomplete human TGFb 19 was inserted into a unique
Eco RI site of pSP6rT7–19 multiple cloning site.Orientation of both cDNAs with respect to T7 or SP6promoters was established by diagnostic enzymaticdigestions and by annealing of in vitro synthesizedradiolabeled RNA to cellular mRNA. To synthesizeb-actin RNA in antisense orientation, plasmid b-actin-pSP6rT7–19 was linearized with Hind III andradiolabeled RNA was synthesized by SP6 poly-merase; the same plasmid was restricted with Bam HIand sense RNA was synthesized with the polymerase.
ŽPlasmid containing TGFb specific cDNA TGFb-.pSP6rT7–19 was linearized by either Hind III or
Nhe I in order to synthesize TGFb-specific RNA insense or antisense orientations, respectively.
w35 xSynthesis of riboprobes labeled with S uridine 5Ž . Žw35 x .a-thio triphosphate S UTP was essentially as
w xdescribed 22 and was done according to the instruc-tions of the supplier of pSP6rT7–19 system. Thereaction mixture was prepared essentially as we de-
w xscribed in previous communications 5 and eachradiolabeled riboprobe ranging in specific activity1–10=108 cpmrmg, was purified by phenolrchlo-roform extraction and ethanol precipitation. Isolatedcells were cultivated on sterilized glass coverslipsand subjected to in situ hybridization according to
w xpreviously described protocols 5,23 . Cells were fixedwith 4% paraformaldehyde and DNA was depuri-nated by a 20-min exposure to 0.2 M HCl. Coverslipswere then sequentially incubated in 2X SSC at 708Cfor 30 min, in 20 mM Tris–HCl, pH 7.2, 2 mM
ŽCaCl containing 1 mgrml proteinase K 30 min at2.378C , washed in distilled water and dehydrated in a
graded ethanol series. Cells were then hybridizedw35 xwith 15 000 cpm of S UTP-radiolabeled riboprobe
in either sense and antisense orientation for 24 h atw xroom temperature 5 . After hybridization, coverslips
were washed three times with 4X SSC for 15 min atroom temperature; 30 min at 378C with 0.5 M NaClin 10 mM Tris–HCl, pH 7.5 containing 20 mgrmlRNase A; and for 24 h at room temperature with 2XSSC, changing the buffer every 8 h. After two finalwashes with 0.1X SSC for 2 h each, coverslips weredehydrated in the graded ethanol series. Dry cover-slips were mounted on slides and coated with KodakNTB-3 nuclear track emulsion which was melted at458C and diluted 1:1 with 600 mM ammonium ac-etate. Slides were stored in dessicated light-tight boxesat 48C for 10–15 days and developed in Kodak D-19developer for 4 min. This was followed by washings,
w xfixation and staining using standard procedures 23 .
2.4. Biological assay for quantification of TGFb
TGFb 1 was measured according to Danielpourw x24 . This method is based on the ability of TGFb 1 toinhibit proliferation of Mv 1 Lu mink lung epithelialcells. Pure TGFb 1 standards were obtained from Rand D Systems, Minneapolis and utilized to obtain astandard curve. Typical dose-response curves were inthe range of 10–250 pgrml TGFb 1, with an ED50
Ž .of approximately 60 pgrml TGFb 1 see Fig. 6 .Fresh serum-free medium had no effect on the stan-

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´244
dard curve generated by pure TGFb 1. For evaluatingTGFb 1 secretion in culture, the various hepatic cellsŽ .Kupffer cells and transfected Hep G2 cells wereplated and maintained for 1–2 days under standardconditions, washed and incubated further in serum-free medium. This serum-free supernatant was har-vested, cleared of cells by centrifugation, and assayedfor TGFb 1. BSA, 1 mgrml, and and protease in-
Ž .hibitors Leupeptin, and Pepstatin A , 1 mgrml, wereadded and activation of latent TGFb 1 was accom-plished by either heating at 808C for 10 min or
Žacidification of the medium 0.05 N HCl, final con-.centration . After 10 min incubation at 258C, the
.medium was neutralized with 0.1 N NaOH . AnŽ . Žaliquot 80 ml was diluted with assay medium 600
.ml, total volume and used to perform each experi-mental assay.
2.5. Inhibition of TGFb biological actiÕity by anti-sense oligonucleotides
Kupffer cells obtained from CCl -intoxicated ani-4
mals were incubated overnight in DMEM supple-mented with 10% FCS after which they were washedand cultured in serum-free DMEM supplemented with2% lactalbumin hydrolizate. A 23-base phosphoroth-
Ž .iote-DNA oligonucleotide Antisense S-Oligo DNAdesigned to hybridize to the AUG translation initia-
Ž Xtion codon of murine TGFb 1 mRNA 5 -CCC CGAX.GGG CGG CAT GGG GGA GG-3 was added at 0,
1 and 5 mM. Cells were incubated for an additional24 h and then the serum-free supernatants were col-lected for TGFb 1 biological assays. Parallel cultureswere treated with a sense TGFb oligonucleotide andprocessed in the same way.
HepG2 cells were transfected with a plasmid con-taining the cDNA for an active and mature TGFb
Ž . w xpRcrCMV-mTGFb 1 S223r225 25 using 10 mgDNAr60 mm dish. Cells were transfected at 30%confluency and left overnight incubated in DMEMsupplemented with 10% FCS. The morning after,cells were carefully washed and further replenishedwith fresh 10% FCS-DMEM for another 36 h untilthey reached nearly 100% confluency. Then, themedia was removed, cells were washed to completelyremove any traces of serum protein and incubated inserum-free DMEM for an additional 24 h in the
Ž .presence of antisense S-oligo DNA 1 and 5 mM forthe experimental HepG2 cells, while sense oligo-nucleotide was incorporated into the control HepG2cells.
2.6. Determination of gene expression by RT-PCR
In order to increase the sensitivity of detection ofTGFb 1 gene expression by the different cell fractionsfrom the CCl -injured livers, we standardized a semi-4
quantitative PCR method based in the co-amplifica-Žtion of the target gene and a constitutive gene b-
.actin in the same reaction tube. Briefly, RNA fromcells was isolated with TRIZOLe and 2 mg of totalRNA were reverse transcribed in 0.05 M Tris–HClpH 8.3, 40 mM KCl, 7 mM MgCl2 buffer containing0.05 mgrml of random hexamers, 1 mM dNTPs mix,0.05 Urml RNase inhibitor and 200 Urml murineleukemia virus reverse transcriptase M-MLV. Sam-ples were incubated for 10 min at 708C and then 60min at 37.58C. Inactivation of the reverse transcrip-tase was achieved by heating the samples at 958C for10 min. cDNAs obtained this way, were used imme-diately or stored at y208C until use. PCR reactionswere performed according to our previous observa-tions where we determine the optimal conditions topick up TGFb 1 gene expression always in the con-text of the co-determination of the expression of a
wconstitutive gene Armendariz-Borunda, unpublishedxresults . Briefly, amplification was performed in a
PCR buffer of 50 mM Tris–HCl, pH 9.0, and 50 mMNaCl containing a mix of 100 mM dNTPs and 1 U ofTaq DNA polymerase. Final primer concentrationswere as follows: 0.6 mM for TGFb sense and anti-sense primers and 0.3 mM for b-actin sense andantisense oligonucleotides. Amplification reactionswere set up and run in an automated thermal cycler at948C for 1 min, 608C for 1 min and 728C for 1.5 minfor a total of 30 cycles.
3. Results
3.1. Non-parenchymal cells from CCL4-injured liÕersexpress TGFb in culture
We previously observed that the levels of TGFb
mRNA in non-parenchymal cells isolated 48 h after a

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´ 245
single intragastric dose of CCl were significantly4
higher than controls. With an aim to identify whetherŽ .the cell s responsible for elevated TGFb gene tran-
scription in CCl -treated livers continue to express4
this cytokine mRNA in vitro, we separated hepato-cytes and non-parenchymal cells. We further subfrac-tionate the non-parenchymal cells in Kupffer and
endothelial cells by using metrizamide gradients. Fig.1 depicts the immunohistochemical characterizationand identification of both types of cells which thenwere subjected to in situ hybridization using TGFb
and b-actin-specific riboprobes. In the normal liver,endothelial and Kupffer cells make up the majorityŽ .85% of the non-parenchymal cells, the remaining
Fig. 1. Immunohistochemical characterization of nonparenchymal cells subfractionated on metrizamide gradients. Cells obtained from ratliver 48 h after acute hepatic injury with CCl were centrifuged through a 3-step metrizamide gradient as described in Section 2. Fraction4
Ž . Ž .enriched in endothelial a,b,c and Kupffer cells d,e,f are shown. Endothelial cells were either stained with rabbit antihuman Factor VIIIŽ .polyclonal antibody by using the peroxidase-antiperoxidase staining technique a or were reacted with nonimmune rabbit IgG instead of
Ž . Ž .the primary antibody b . Fractions enriched in Kupffer cells readily exhibited nonspecific esterase staining d , while no similar stainingŽ .was seen for the endothelial cell fraction e . Both endothelial and Kupffer cell-enriched fractions reacted positively to indirect
Ž .immunofluorescence staining of vimentin with a monoclonal antivimentin antibody c,f .
( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´246
fraction is composed of fibroblasts, hepatic stellatecells and Pit cells. However, in the CCl -treated liver4
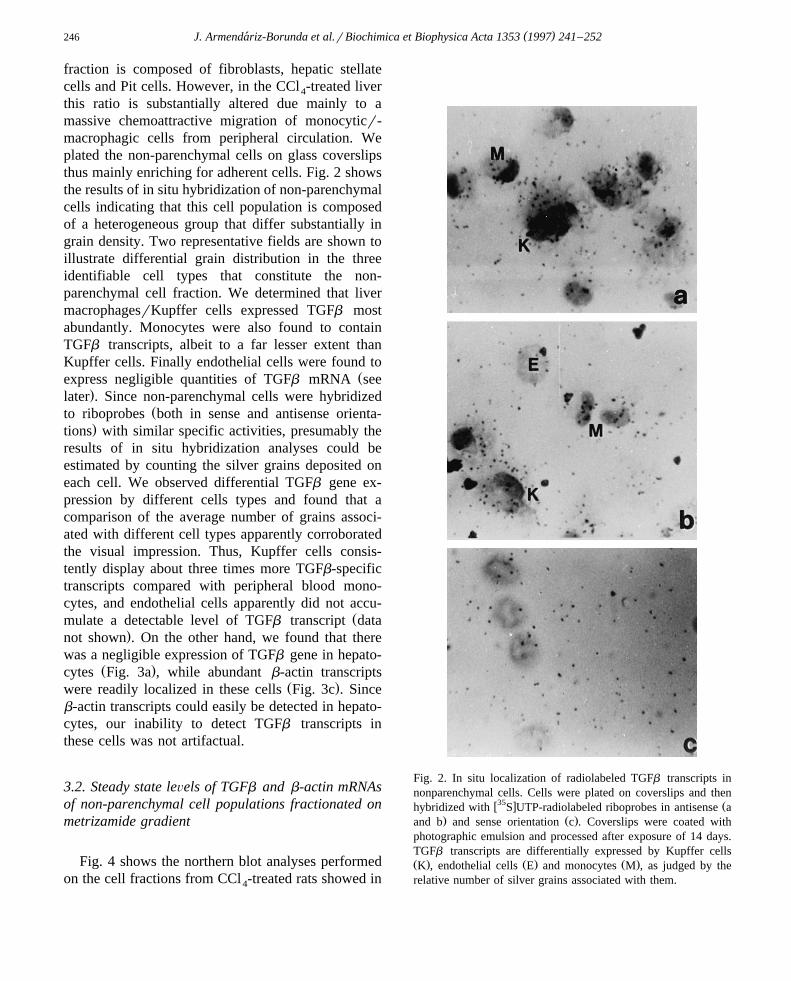
this ratio is substantially altered due mainly to amassive chemoattractive migration of monocyticr-macrophagic cells from peripheral circulation. Weplated the non-parenchymal cells on glass coverslipsthus mainly enriching for adherent cells. Fig. 2 showsthe results of in situ hybridization of non-parenchymalcells indicating that this cell population is composedof a heterogeneous group that differ substantially ingrain density. Two representative fields are shown toillustrate differential grain distribution in the threeidentifiable cell types that constitute the non-parenchymal cell fraction. We determined that livermacrophagesrKupffer cells expressed TGFb mostabundantly. Monocytes were also found to containTGFb transcripts, albeit to a far lesser extent thanKupffer cells. Finally endothelial cells were found to
Žexpress negligible quantities of TGFb mRNA see.later . Since non-parenchymal cells were hybridized
Žto riboprobes both in sense and antisense orienta-.tions with similar specific activities, presumably the
results of in situ hybridization analyses could beestimated by counting the silver grains deposited oneach cell. We observed differential TGFb gene ex-pression by different cells types and found that acomparison of the average number of grains associ-ated with different cell types apparently corroboratedthe visual impression. Thus, Kupffer cells consis-tently display about three times more TGFb-specifictranscripts compared with peripheral blood mono-cytes, and endothelial cells apparently did not accu-
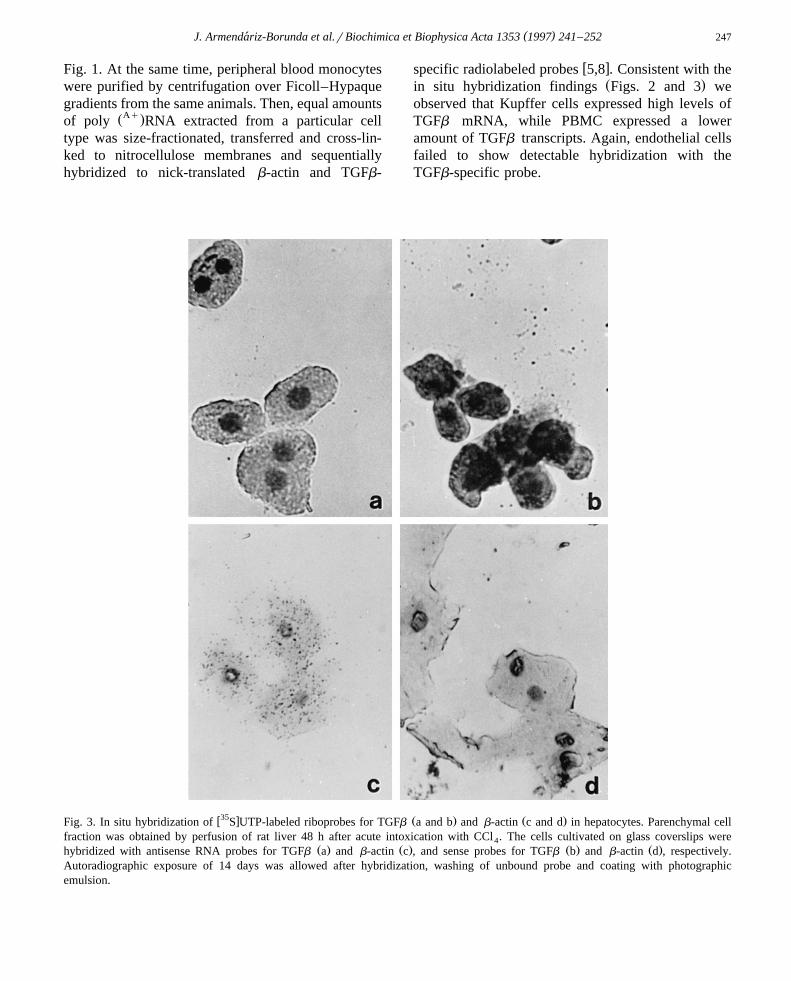
Žmulate a detectable level of TGFb transcript data.not shown . On the other hand, we found that there
was a negligible expression of TGFb gene in hepato-Ž .cytes Fig. 3a , while abundant b-actin transcripts
Ž .were readily localized in these cells Fig. 3c . Sinceb-actin transcripts could easily be detected in hepato-cytes, our inability to detect TGFb transcripts inthese cells was not artifactual.
3.2. Steady state leÕels of TGFb and b-actin mRNAsof non-parenchymal cell populations fractionated onmetrizamide gradient
Fig. 4 shows the northern blot analyses performedon the cell fractions from CCl -treated rats showed in4
Fig. 2. In situ localization of radiolabeled TGFb transcripts innonparenchymal cells. Cells were plated on coverslips and then
w35 x Žhybridized with S UTP-radiolabeled riboprobes in antisense a. Ž .and b and sense orientation c . Coverslips were coated with
photographic emulsion and processed after exposure of 14 days.TGFb transcripts are differentially expressed by Kupffer cellsŽ . Ž . Ž .K , endothelial cells E and monocytes M , as judged by therelative number of silver grains associated with them.
( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´ 247
Fig. 1. At the same time, peripheral blood monocyteswere purified by centrifugation over Ficoll–Hypaquegradients from the same animals. Then, equal amounts
ŽAq.of poly RNA extracted from a particular celltype was size-fractionated, transferred and cross-lin-ked to nitrocellulose membranes and sequentiallyhybridized to nick-translated b-actin and TGFb-
w xspecific radiolabeled probes 5,8 . Consistent with theŽ .in situ hybridization findings Figs. 2 and 3 we
observed that Kupffer cells expressed high levels ofTGFb mRNA, while PBMC expressed a loweramount of TGFb transcripts. Again, endothelial cellsfailed to show detectable hybridization with theTGFb-specific probe.
w35 x Ž . Ž .Fig. 3. In situ hybridization of S UTP-labeled riboprobes for TGFb a and b and b-actin c and d in hepatocytes. Parenchymal cellfraction was obtained by perfusion of rat liver 48 h after acute intoxication with CCl . The cells cultivated on glass coverslips were4
Ž . Ž . Ž . Ž .hybridized with antisense RNA probes for TGFb a and b-actin c , and sense probes for TGFb b and b-actin d , respectively.Autoradiographic exposure of 14 days was allowed after hybridization, washing of unbound probe and coating with photographicemulsion.

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´248
3.3. Determination of TGFb gene expression by RT-PCR
In order to make sure that the differential expres-sion of TGFb by the several liver cell componentswas accurate and not underestimated, we standard-ized a semiquantitative method using reverse tran-
Ž .scriptase-polymerase chain reaction RT-PCR to de-termine gene expression. This method has proven tobe highly sensitive and efficient. To accomplish this,we assayed several PCR conditions varying cDNAamount, primer concentrations, temperature and num-
Ž .ber of cycles data not shown . We finally met ouroptimal PCR conditions to detect and co-amplify inthe same reaction tube two different PCR products:one band of 878 bp corresponding to TGFb 1 geneand another band of 314 bp corresponding to theb-actin gene. Since we co-amplified the two PCRproducts in the same reaction mixture, we were cer-tain that no artifacts were included in the experiment.Furthermore, we were able to use the b-actin PCRproduct as an internal control in order to specificallyquantitate and normalize the levels of TGFb 1 geneexpression by each cell type. Fig. 5 shows an EtBr-stained agarose gel where the PCR products areseparated according to their molecular weight. Theexpression of the two gene products is perfectly clear,demonstrating that while the band of 314 bp corre-sponding to b-actin gene is present in all the lanes,the band of 878 bp corresponding to the TGFb 1amplified gene product is distinctly present only inthe whole non-parenchymal fraction and in the subse-quent purified Kupffer cell subfraction. A band repre-senting an non-specific PCR product was detected inthe endothelial cell fraction.
3.4. Production of TGFb1 is inhibited by phosphoro-thioate antisense oligodeoxynucleotides
To evaluate this concept, we tested the effects ofantisense S-oligodeoxynucleotides targeted to thetranslational initiation site of murine TGFb 1 mRNAŽ .23 mer by introducing them into the medium ofcultured Kupffer cells from CCl -injured rat livers4
and HepG2 cells transfected with the TGFb 1cDNA-containing plasmid pRcrCMV-mTGFb 1Ž . w xS223r225 25 . Phosphorothioation of theoligomers was employed to retard their intracellular
degradation. In order to achieve this, we previouslystandardized a dose-response curve using pure TGFb 1to determine the ED-50. Fig. 6 shows that approxi-mately 60 pgrml of pure TGFb 1 induced a 50%
w3 xinhibition in the H thymidine incorporation by Mv1 Lu mink lung epithelial cells. As Fig. 7 shows,there is a dose-dependent effect of antisense S-
Fig. 4. Steady-state levels of b-actin and TGFb mRNAs inŽ .nonparenchymal cell fractions. Peripheral blood monocytes MŽ .were purified by Ficoll–Hypaque gradient while endothelial E
Ž .and Kupffer cell K fractions were obtained from CCl -treated4
rat liver by metrizamide gradient centrifugation as described inSection 2, Methods. Equal amounts of total RNA were fraction-
ŽAy . ŽAq .ated into poly or poly fractions and electrophoresed inŽAq . Ž .1.2% agarose gel. Poly RNA 3 mgrlane electrophoresed
in agarose gel was immobilized on nitrocellulose and sequentiallyhybridized with nick-translated cDNA plasmids containing either
Ž . Ž .TGFb middle panel or b-actin bottom panel sequences. Ethid-ŽAy . Ž .ium bromide stained pattern of the poly RNA 7.5 mgrlane
Ž .is depicted with ribosomal sizes 28S and 18S in the top panel.

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´ 249
Fig. 5. TGFb gene expression determination by RT-PCR. Agarose gel showing PCR amplified products. Lane 1: f X 174rHae III; lane2: pmTGFb cDNA, lane 3: non-parenchymal cell fraction; lane 4: hepatocytes; lane 5: Kupffer cell subfraction; lane 6: endothelial cellsubfraction. Hepatocytes and non-parenchymal cells from CCl -treated animals were sub-fractionated and RNA from each cell type was4
isolated with TRIZOLe. Two mg of total RNA were reverse transcribed in 0.05 M Tris–HCl, pH 8.3, 40 mM KCl, 7 mM MgCl buffer2
containing 0.05 mgrml of random hexamers, 1 mM dNTPs mix, 0.05 Urml RNase inhibitor and 200 Urml murine leukemia virusreverse transcriptase M-MLV. Samples were incubated for 10 min at 708C and then 60 min at 37.58C. Inactivation of the reversetranscriptase was achieved by heating the samples at 958C for 10 min. cDNAs obtained this way, were used for PCR amplification in aPCR buffer of 50 mM Tris–HCl, pH 9.0, and 50 mM NaCl containing a mix of 100 mM dNTPs and 1 U of Taq DNA polymerase. Finalprimer concentrations were 0.6 mM for TGFb sense and antisense primers and 0.3 mM for b-actin sense and antisense oligonucleotides.Amplification reactions were set up and run in an automated thermal cycler at 948C for 1 min, 608C for 1 min and 728C for 1.5 min for atotal of 30 cycles.
oligodeoxynucleotide complementary to TGFb 1mRNA on the biological effects of this cytokine asmeasured by its ability to inhibit proliferation ofmink lung cells. Kupffer cells were obtained fromCCl -treated animals and incubated for 24 h in4
serum-free medium that contained various concentra-
Fig. 6. Standard curve for TGFb biological activity. Pure TGFb 1standards were obtained from R and D Systems and utilized toobtain a standard curve. Mv 1 Lu mink lung epithelial cells were
w3 xgrown as described in Section 2 pulsed with H thymidine andTGFb was added at the concentrations indicated. TGFb inhibitsproliferation of mink lung cells in a dose-dependent manner.Typical dose-response curves were in range of 10–250 pgrmlTGFb 1, with an ED50 of approximately 60 pgrml TGFb 1.
tions of antisense S-oligodeoxynucleotides. Thisdose-dependent effect was maintained at 5 mM anti-sense oligomer. On the other hand, sense S-oligomerat 5 mM final concentration had no effect on
Fig. 7. Antisense oligonucleotides inhibit endogenous productionof TGFb by Kupffer cells from CCl -injured rats. Kupffer cells4
were plated in D-MEM supplemented with 10% FCS overnight.After this period, cells were washed and cultured in serum-freeD-MEM plus 2% lactalbumin hydrolyzate. TGFb-antisense andsense oligomers were added at the concentrations indicated andcells were further incubated at 378C for 24 h and then serum-freesupernatants were collected for TGFb-biological activity determi-nation.

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´250
Fig. 8. Antisense oligonucleotides down-regulate the inducibleproduction of TGFb by Hep G2 cells. Subconfluent cultures ofHepG2 cells were transfected with a plasmid containing thecDNA for an active and mature TGFb pRcrCMV-
Ž .mTGFb 1 S223r225 as described in Section 2. After the trans-fection period was complete, the media was removed, cells werewashed to completely remove any trace of serum protein andincubated in serum-free D-MEM for an additional 24 h in the
Ž .presence of antisense S-oligo DNA 1 and 5 mM for theexperimental HepG2 cells, while sense oligonucleotide was in-corporated to the control HepG2 cells. Supernatants were col-lected in the presence of protease inhibitors and frozen until used.Immediately before use, an aliquot was heated at 808C for 10 minandror acidified, diluted with medium and the inhibition assaywas performed.
w3 xH thymidine incorporation by Mv 1 Lu mink lungcells.
Fig. 8 depicts a similar behavior and dose-depen-dent effect of antisense S-oligodeoxynucleotide onthe physiological activity of TGFb 1. Again, the con-trol serum-free supernatants that were incubated withthe sense S-oligodeoxynucleotides showed no down-regulation of TGFb 1 activity in vitro.
4. Discussion
For several years the primary focus of our labora-tory has been the molecular mechanisms that mediatetissue fibrosis and the role of cytokines in this pro-cess. Using experimental models of hepatic fibrosis,we have demonstrated that TGFb was transiently
w xstimulated in the injured organs 5,8 . In the liver wenoted that the steady-state levels of TGFb mRNAwere maximally increased around 48 h after CCl4
administration. Non-parenchymal cells almost exclu-
sively accounted for increased TGFb gene expres-sion, while hepatocytes accumulated negligible levelsof TGFb transcripts. In the present communicationwe perform a comprehensive study using in situhybridization, RT-PCR and cell fractionation meth-ods to examine TGFb gene expression in differentcell types that comprise the nonparenchymal cell
Ž .fraction in order to investigate the following: a wewanted to know whether isolated and cultured livercells consistently express in vitro TGFb gene as they
Ž .do in vivo; b if they do so, we intended to block theexpression of TGFb gene at the mRNA level in vitroby using phosphorotioate antisense S-oligonucleo-tides directed specifically against the TGFb mRNA
Ž .translational initiation site; and c by using the sameapproach we wanted to block the inducible TGFb
gene expression in Hep G2 cells.Consistent with our previously published data, we
found that hepatocytes contained undetectable levelsof TGFb transcripts. This was true even when we use
Žreverse transcriptase-polymerase chain reaction RT-.PCR to study TGFb gene expression. This fact
acquires capital relevance in the light of recent find-w xings 26 by other investigators where they showed
TGFb 1 to be produced by hepatocytes and hepaticŽ .stellate cells Ito cells or lipocytes during liver re-
generation induced by a 70% partial hepatectomy.Those investigators used RNase protection assay todetect TGFb 1 mRNA expression by hepatocytes. Itis noteworthy to emphasize that even when this lattertechnique is highly sensitive it is far less accurate andsensitive compared with RT-PCR. Bissell and collab-orators invoke the participation of several other fac-tors like in situ hybridization or Northern blot analy-sis as the major caveat in the inability to detectTGFb 1 mRNA in hepatocytes as shown by previousstudies. Regardless of the intrinsic physiological dif-ferences between the partial hepatectomy- and CCl -4
induced liver regeneration accounting for the differ-ential cell-gene expression, we have shown here acomprehensive and comparative study, using variousmolecular biology techniques, that hepatocytes do notexpress TGFb 1 in the CCl -induced liver injury. We4
also postulate that enhanced expression of TGFb
gene by liver-derived Kupffer cellsrmacrophagesrepresents a process of activation of these cells, themolecular mechanism of which remain elusive. Nev-ertheless, such pronounced activation of TGFb 1 gene

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´ 251
expression might be a critical early response of cer-tain nonparenchymal cells to toxic liver injury. It isimportant to note that we also failed to detect signifi-cant TGFb 1 gene expression in endothelial cellseither by in situ, northern hybridization and RT-PCRmethods. This is in contrast to what has been reportedfor regenerative liver where endothelial cells were
w xfound to be the main producers of TGFb 1 19 . Itappears, therefore, that there is a fundamental differ-ence between the processes of tissue regenerationinitiated as a result of partial hepatectomy and CCl4
administration. The differences between our experi-mental model and others reported elsewhere remainto be reconciled.
Antisense RNA technology is emerging as an ef-fective means of blocking or decreasing the intra-cellular or secreted amounts of a given gene productw x27,28 . It is based on the fact that these antisensesequences hybridize to specific RNA transcripts, dis-rupting normal RNA processing, stability and transla-tion, thereby preventing the expression of a targetedgene. Administration of antisense oligonucleotides ortransfer of expression constructs capable of produc-ing intracellular antisense sequences complementaryto the mRNA of interest, have been shown to blockthe translation of specific genes in vitro and in vivo.Therefore, we employed antisense RNA technologyto block the TGFb gene expression in cultured livercells which play a major role in inducing and perpet-uating the molecular mechanisms regulating the onsetof hepatic fibrosis.
The biological activities of TGFb 1 antisenseoligomer were sequence-specific since the TGFb 1sense oligomer did not prevent TGFb protein produc-tion. Our findings, in addition to demostrating theefficacy and specificity of the TGFb 1 antisense S-oligonucleotide, suggest that the oligomer might beof value for the treatment of diseases in which TGFb 1
Žoverexpression might play a pathogenic role e.g.liver cirrhosis, diabetic renal disease, glomerulo-
.nephritis, etc. .
Acknowledgements
We acknowledge the competent assistance of Ro-gelio Troyo in the preparation of the graphs of this
manuscript. These studies were supported in part by aŽ .grant from CONACyT 4947-M9409 .
References
w x1 N. Sanderson, V. Factor, P. Nagy, J. Kopp, P. Kondaiah, L.Wakefield, A.B. Roberts, M.B. Sporn, S.S. Thorgeirsson,
Ž .Proc. Natl. Acad. Sci. USA 92 1995 2572–2576.w x Ž .2 J. Massague, Annu. Rev. Cell Biol. 6 1990 597–641.´w x Ž .3 H.L. Moses, E.Y. Yang, J.A. Pietenpol, Cell 63 1990
245–247.w x4 H.Y. Lin, X.F. Wang, E.E. Ng, R.A. Weinberg, H.F. Lodish,
Ž .Cell 68 1992 775–785.w x5 J. Armendariz-Borunda, H. Katai, C.M. Jones, J.M. Seyer,
Ž .A.H. Kang, R. Raghow, Lab. Investigation 69 1993 283–294.
w x6 B. Waters, M.J. Malecha, J.M. Seyer, D. Martinez-Fong, A.Martinez-Hernandez, J. Armendariz-Borunda, Hepatology
Ž . Ž .20 4 1994 227A.w x Ž .7 Border, W.A. and Noble, N.A. 1995 TGF-b. Scientific
American SCI. and MED. January 68-77.w x8 J. Armendariz-Borunda, J.M. Seyer, A.H. Kang, R. Raghow,
Ž .FASEB J. 4 1990 215–221.w x9 J. Armendariz-Borunda, N. Roy, C. Simkevich, R. Raghow,
Ž .J.M. Seyer, A.H. Kang, Bichemical Journal 304 1994817–824.
w x10 H. Nakatsukasa, P. Nagy, R.P. Evarts, C.-C. Hsia, E. Mars-Ž .den, S.S. Thorgeirsson, J. Clin. Invest. 85 1990 1833–1843.
w x11 M.S. Anscher, I.R. Crocker, R.L. Jirtle, Radiat. Res. 122Ž .1990 77–85.
w x12 M.J. Czaja, F.R. Weiner, K.C. Flanders, M.A. Giambrone,Ž .R. Wind, L. Biempica, M.A. Zern, J. Cell Biol. 108 1989
2477–2482.w x Ž .13 A. Castilla, J. Prieto, N. Fausto, N. Engl. J. Med. 324 1991
933–940.w x14 M. Spearman, W.R. Taylor, A.H. Greenberg, J.A. Wright,
Ž .Gene 149 1994 25–29.w x15 M.J. Merrilees, L. Scott, Journal of Vascular Research 31
Ž .1994 322–329.w x16 P. Jachimczak, B. Hessdorfer, K. Fabel-Schulte, W. Brysch,
K.H. Schlingensiepen, A. Blesch, U. Bogdahn, CellularŽ .Immunology 165 1995 125–133.
w x17 P. Jachimczak, B. Hessdorfer, K. Fabel-Schulte, C. Wis-meth, W. Brysch, K.H. Schlingensiepen, A. Bauer, A.
Ž .Blesch, U. Bogdahn, Int. J. Cancer 65 1996 332–337.w x18 A. Khanna, B. Li, P. Li, M. Suthanthiran, Bioch. Biophys.
Ž .Res. Comm. 204 1994 1061–1066.w x19 L. Braun, J.E. Mead, M. Pamzica, R. Mikumo, G.I. Bell, N.
Ž .Fausto, Proc. Natl. Acad. Sci. USA 85 1988 1539–1543.w x20 L.T. Yam, C.Y. Li, W.H. Crosby, Am. J. Clin. Pathol. 55
Ž .1971 283–290.w x21 D.W. Cleveland, M.A. Lopata, R.J. MacDonald, N.J. Gowan,
Ž .W.H. Rutter, M.W. Kirschner, Cell 20 1980 95–105.

( )J. Armendariz-Borunda et al.rBiochimica et Biophysica Acta 1353 1997 241–252´252
w x22 D.A. Melton, P.A. Krieg, M.R. Rebagliati, T. Maniatis, K.Ž .Zinn, M.R. Green, Nucleic Acids Res. 12 1984 7035–7056.
w x23 L. Angerer, D. DeLeou, K. Cox, R. Maxson, L. Kedes, J.Ž .Kaumeyer, E. Weinberg, R. Angerer, Dev. Biol. 112 1985
157–166.w x24 D. Danielpour, L.L. Dart, K.C. Flanders, A.B. Roberts,
Ž .M.B. Sporn, J. Cell. Physiol. 138 1989 79–86.w x25 D. Martınez-Fong, J. Mullersman, A.F. Purchio, J. Armen-´
dariz-Borunda, A. Martinez-Hernandez, Hepatology 20Ž .1994 1602–1608.
w x26 D.M. Bissell, S.-S. Wang, W.R. Jarnagin, F.J. Roll, J. Clin.Ž .Invest. 96 1995 447–455.
w x27 A. Harel-Bellan, D.R. Ferris, M. Vinocour, J.T. Holt, W.L.Ž .Farrar, J. Immunol. 140 1988 2431.
w x Ž .28 D.A. Melton, Proc. Natl. Acad. Sci. USA 82 1985 144.
Copyright © 2022 FDOKUMEN




















