
Malaria circumsporozoite protein inhibits the respiratory burst in Kupffer cells
19
Malaria circumsporozoite protein inhibits the respiratory burst in Kupffer cells Ivan Usynin, Christian Klotz and Ute Frevert* Department of Medical Parasitology, New York University School of Medicine, 341 E 25 St, New York, NY 10010, USA. Summary After transmission by infected mosquitoes, malaria sporozoites rapidly travel to the liver. To infect hepa- tocytes, sporozoites traverse Kupffer cells, but sur- prisingly, the parasites are not killed by these resident macrophages of the liver. Here we show that Plasmo- dium sporozoites and recombinant circumsporozoite protein (CSP) suppress the respiratory burst in Kupffer cells. Sporozoites and CSP increased the intracellular concentration of cyclic adenosyl mono- phosphate (cAMP) and inositol 1,4,5-triphosphate in Kupffer cells, but not in hepatocytes or liver endothelia. Preincubation with cAMP analogues or inhibition of phosphodiesterase also inhibited the respiratory burst. By contrast, adenylyl cyclase inhibition abrogated the suppressive effect of sporozoites. Selective protein kinase A (PKA) inhibi- tors failed to reverse the CSP-mediated blockage and stimulation of the exchange protein directly activ- ated by cAMP (EPAC), but not PKA inhibited the respiratory burst. Both blockage of the low-density lipoprotein receptor-related protein (LRP-1) with receptor-associated protein and elimination of cell surface proteoglycans inhibited the cAMP increase in Kupffer cells. We propose that by binding of CSP to LRP-1 and cell surface proteoglycans, malaria sporozoites induce a cAMP/EPAC-dependent, but PKA-independent signal transduction pathway that suppresses defence mechanisms in Kupffer cells. This allows the sporozoites to safely pass through these professional phagocytes and to develop inside neighbouring hepatocytes. Introduction Minutes after entering a cutaneous blood capillary at the mosquito bite site (Vanderberg and Frevert, 2004; Amino et al., 2006), malaria sporozoites are arrested in the liver (Shin et al., 1982; Hollingdale, 1985). To infect hepato- cytes, their initial target cells in the mammalian host, sporozoites must cross the layer of sinusoidal cells composed of specialized, highly fenestrated sinusoidal endothelia interspersed with Kupffer cells, the resident macrophages of the liver (Wisse et al., 1985; Bouwens and Wisse, 1992). Various in vivo, in vitro and, more recently, intravital studies have provided evidence that sporozoites cross the sinusoidal barrier by passing through Kupffer cells. According to our current model, sporozoites are initially arrested in the liver by binding with their two major surface proteins, the circumsporozoite protein (CSP) and the thrombospondin-related adhesive protein (TRAP), to unique extracellular matrix proteogly- cans of the liver (Robson et al., 1995; Pradel et al., 2002; 2004). The parasites glide along the sinusoid and even- tually recognize selected syndecans, small cell surface proteoglycans, on Kupffer cells, which they actively invade and traverse unharmed (Meis et al., 1983; 1985; Pradel and Frevert, 2001; Pradel et al., 2002; Ishino et al., 2004; Frevert et al., 2005; 2006a,b; Baer et al., 2007). After exiting Kupffer cells towards the space of Disse, sporozoites traverse several hepatocytes before even- tually settling down for multiplication to thousands of erythrocyte-infective merozoites (Mota et al., 2001; Frevert et al., 2005; Sturm et al., 2006; Tarun et al., 2006). Circumsporozoite protein binding to the surface of mammalian cells (Shakibaei and Frevert, 1996; Ying et al., 1997; Ancsin and Kisilevsky, 2004) is mediated by a dual interaction with (i) syndecans, a family of small trans- membrane proteoglycans that are expressed on almost all cell types (Bernfield et al., 1992) and (ii) the low- density lipoprotein receptor-related protein (LRP-1), a multifunctional scavenger receptor that is predominantly found in the liver (Herz, 1993; Strickland et al., 1995). LRP-1, also known as the alpha-2-macroglobulin receptor (a 2MR) or CD91, is responsible for the clearance from the blood of a large number of molecules including activated alpha-2-macroglobulin (a2M*), proteases and their com- plexes with inhibitors, matrix proteins, growth factors, as well as small particles such as lipoprotein remnants (Herz, 1993; Strickland et al., 1995). Both syndecans and LRP-1 induce intracellular signalling cascades. Depending on the cytoplasmic domain of their various core proteins, syndecans are involved in distinct, but overlapping signal Received 26 February, 2007; revised 17 April, 2007; accepted 14 May, 2007. *For correspondence. E-mail [email protected]; Tel. (+1) 212 263 6755; Fax (+1) 212 263 8116. Cellular Microbiology (2007) 9(11), 2610–2628 doi:10.1111/j.1462-5822.2007.00982.x First published online 15 June 2007 © 2007 The Authors Journal compilation © 2007 Blackwell Publishing Ltd
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Malaria circumsporozoite protein inhibits the respiratory burst in Kupffer cells

Malaria circumsporozoite protein inhibits therespiratory burst in Kupffer cells
Ivan Usynin, Christian Klotz and Ute Frevert*Department of Medical Parasitology, New YorkUniversity School of Medicine, 341 E 25 St, New York,NY 10010, USA.
Summary
After transmission by infected mosquitoes, malariasporozoites rapidly travel to the liver. To infect hepa-tocytes, sporozoites traverse Kupffer cells, but sur-prisingly, the parasites are not killed by these residentmacrophages of the liver. Here we show that Plasmo-dium sporozoites and recombinant circumsporozoiteprotein (CSP) suppress the respiratory burst inKupffer cells. Sporozoites and CSP increased theintracellular concentration of cyclic adenosyl mono-phosphate (cAMP) and inositol 1,4,5-triphosphatein Kupffer cells, but not in hepatocytes or liverendothelia. Preincubation with cAMP analogues orinhibition of phosphodiesterase also inhibited therespiratory burst. By contrast, adenylyl cyclaseinhibition abrogated the suppressive effect ofsporozoites. Selective protein kinase A (PKA) inhibi-tors failed to reverse the CSP-mediated blockage andstimulation of the exchange protein directly activ-ated by cAMP (EPAC), but not PKA inhibited therespiratory burst. Both blockage of the low-densitylipoprotein receptor-related protein (LRP-1) withreceptor-associated protein and elimination of cellsurface proteoglycans inhibited the cAMP increase inKupffer cells. We propose that by binding of CSPto LRP-1 and cell surface proteoglycans, malariasporozoites induce a cAMP/EPAC-dependent, butPKA-independent signal transduction pathway thatsuppresses defence mechanisms in Kupffer cells.This allows the sporozoites to safely pass throughthese professional phagocytes and to develop insideneighbouring hepatocytes.
Introduction
Minutes after entering a cutaneous blood capillary at themosquito bite site (Vanderberg and Frevert, 2004; Amino
et al., 2006), malaria sporozoites are arrested in the liver(Shin et al., 1982; Hollingdale, 1985). To infect hepato-cytes, their initial target cells in the mammalian host,sporozoites must cross the layer of sinusoidal cellscomposed of specialized, highly fenestrated sinusoidalendothelia interspersed with Kupffer cells, the residentmacrophages of the liver (Wisse et al., 1985; Bouwensand Wisse, 1992). Various in vivo, in vitro and, morerecently, intravital studies have provided evidence thatsporozoites cross the sinusoidal barrier by passingthrough Kupffer cells. According to our current model,sporozoites are initially arrested in the liver by binding withtheir two major surface proteins, the circumsporozoiteprotein (CSP) and the thrombospondin-related adhesiveprotein (TRAP), to unique extracellular matrix proteogly-cans of the liver (Robson et al., 1995; Pradel et al., 2002;2004). The parasites glide along the sinusoid and even-tually recognize selected syndecans, small cell surfaceproteoglycans, on Kupffer cells, which they activelyinvade and traverse unharmed (Meis et al., 1983; 1985;Pradel and Frevert, 2001; Pradel et al., 2002; Ishino et al.,2004; Frevert et al., 2005; 2006a,b; Baer et al., 2007).After exiting Kupffer cells towards the space of Disse,sporozoites traverse several hepatocytes before even-tually settling down for multiplication to thousands oferythrocyte-infective merozoites (Mota et al., 2001;Frevert et al., 2005; Sturm et al., 2006; Tarun et al., 2006).
Circumsporozoite protein binding to the surface ofmammalian cells (Shakibaei and Frevert, 1996; Yinget al., 1997; Ancsin and Kisilevsky, 2004) is mediated by adual interaction with (i) syndecans, a family of small trans-membrane proteoglycans that are expressed on almostall cell types (Bernfield et al., 1992) and (ii) the low-density lipoprotein receptor-related protein (LRP-1), amultifunctional scavenger receptor that is predominantlyfound in the liver (Herz, 1993; Strickland et al., 1995).LRP-1, also known as the alpha-2-macroglobulin receptor(a2MR) or CD91, is responsible for the clearance from theblood of a large number of molecules including activatedalpha-2-macroglobulin (a2M*), proteases and their com-plexes with inhibitors, matrix proteins, growth factors, aswell as small particles such as lipoprotein remnants (Herz,1993; Strickland et al., 1995). Both syndecans and LRP-1induce intracellular signalling cascades. Depending onthe cytoplasmic domain of their various core proteins,syndecans are involved in distinct, but overlapping signal
Received 26 February, 2007; revised 17 April, 2007; accepted 14May, 2007. *For correspondence. E-mail [email protected];Tel. (+1) 212 263 6755; Fax (+1) 212 263 8116.
Cellular Microbiology (2007) 9(11), 2610–2628 doi:10.1111/j.1462-5822.2007.00982.xFirst published online 15 June 2007
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd

transduction cascades (Carey, 1997; Woods and Couch-man, 1998; Zimmermann and David, 1999; Rapraeger,2000). LRP-1 is directly or indirectly responsible for alarge variety of signal transduction events. A direct role ofLRP-1 in signal transduction is supported by the findingthat receptor ligation leads to tyrosine and serine phos-phorylation of its cytoplasmic domain (Li et al., 2001; vander Geer, 2002) and that signalling adapter proteins asso-ciate with the NPXY motifs in the cytoplasmic domain(Gotthardt et al., 2000). Direct LRP-1 signalling can beinhibited by the receptor-associated protein (RAP) andoccurs via the MEKK/JNK/cJun pathway (Lutz et al.,2002; Schneider and Nimpf, 2003).
A well-documented example for an indirect role ofLRP-1 in signal transduction is its cooperation withthe a2M signalling receptor (a2MSR) on peritonealmacrophages. Upon ligation with activated a2M (a2M*),the a2MSR activates phospholipase C (PLC), whichgenerates the two second messengers inositol 1,4,5-triphosphate (IP3) and diacylglycerol (DAG) (Misra andPizzo, 1998a,b; Misra et al., 2002a). a2M* binding toa2MSR also raises the intracellular concentration ofcyclic adenosyl mono-phosphate (cAMP) (Misra et al.,2002a,b). The binding of other LRP-1 ligands such asapolipoprotein E (apoE), lactoferrin, lipoprotein lipase andPseudomonas exotoxin A to macrophages increases theintracellular concentration of calcium, cAMP and IP3, andactivates protein kinase A (PKA) (Misra and Pizzo, 1998b;2002; Misra et al., 2002a,b; 2005). These LRP-1 ligands,however, signal via a pertussis-sensitive G protein andthe exact mechanism of signal induction is unknown.
Phagocytosis of microorganisms usually results in acti-vation of the NADPH oxidase and the generation of reac-tive oxygen species (ROS), a potent defence mechanismof macrophages against pathogenic intruders (Mauël,1996). The induction of the formation of ROS is a complexevent that requires the assembly of the heterohexamericNADPH oxidase (Lambeth, 2004; Quinn and Gauss,2004). This enzyme complex consists of two membrane-bound subunits, gp91phox and p22phox, which together formthe flavocytochrome b558 complex (Fig. S1).
Like many other microorganisms, malaria parasites arehighly susceptible to oxidative damage (Miller and Briti-gan, 1997; Becker et al., 2004; Müller, 2004). To combatthe oxidative stress imposed by the host, obligate intrac-ellular parasites have acquired a wide range of strategies.One common microbial defence mechanism againstphagocyte-derived ROS is avoidance of exposure to oxi-dants, which can be accomplished by killing the macro-phage before the onset of the respiratory burst, byprevention of phagocytosis, phagolysosomal fusion andphagosomal acidification, or by interference with signal-ling pathways of the phagocyte (Mauël, 1996; Miller andBritigan, 1997). Other strategies aim to prevent oxidative
damage, either by non-enzymatic blockage of the assem-bly of the NADPH oxidase or its toxic oxidative products,or by decomposition of ROS with antioxidant enzymessuch as superoxide dismutase (SOD), catalase, variousglutathione- and thioredoxin-dependent peroxidase/reductase systems, and peroxiredoxins (Miller andBritigan, 1997; Becker et al., 2004; Müller, 2004). Thus,survival of obligate intracellular microbes depends onmaintaining a delicate redox balance, which requires con-stant reestablishment of oxidant homeostasis.
Kupffer cells represent more than 80% of the totalpopulation of tissue macrophages of the body (Kuiperet al., 1994). With their strategic position in the sinusoidallumen, they play an important role in the removal ofaltered self or foreign substances as well as pathogenicmicroorganisms from the blood (Kuiper et al., 1994).Phagocytosis results in the generation of ROS, which arelethal for many microorganisms (Mauël, 1996). Surpris-ingly, however, Kupffer cells (Pradel and Frevert, 2001)and other macrophages (Danforth et al., 1980; Smith andAlexander, 1986; Vanderberg et al., 1990) do not killviable malaria sporozoites, even after prolonged coincu-bation in vitro. In contrast to normal salivary gland sporo-zoites, altered or dead parasites elicit a respiratory burstin peritoneal macrophages and are phagocytozed(Danforth et al., 1980; Smith and Alexander, 1986). Theseobservations led us to hypothesize that the parasites areable to manipulate Kupffer cell function by inducing intra-cellular signalling pathways that suppress macrophageactivation. Here we show that by raising the intracellularconcentration of cAMP and IP3, malaria sporozoites sup-press the respiratory burst in rat Kupffer cells.
Results
Effect of respiratory burst induction in Kupffer cells onPlasmodium sporozoites
Using a nitro-blue tetrazolium (NBT) assay, Smith andAlexander (Smith and Alexander, 1986) demonstratedthat heat- or trypsin-inactivated Plasmodium bergheisporozoites, but not viable salivary gland sporozoites,induce a local oxidative burst in peritoneal macrophages.In agreement with this report, we found that freeze/thaw-inactivated, but not viable Plasmodium yoelii sporozoitestrigger a strong localized NBT reduction in murine Kupffercells (Table 1). In addition, inactivated sporozoites werealso surrounded by a strong NBT reaction in Kupffer cellsthat had been activated with phorbol-12-myristate-13-acetate (PMA). As expected, PMA treatment, which trig-gers a respiratory burst in Kupffer cells (see below),caused a light diffuse NBT reduction in the majority of theKupffer cells. Interestingly, PMA treatment increased thenumber of NBT-positive parasites only roughly twofold,
Malaria sporozoites deactivate Kupffer cells 2611
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

while the majority of the Kupffer cell-associated sporozoi-tes remained NBT negative (Table 1), suggesting that livePlasmodium sporozoites either do not elicit sufficientburst activity to be killed or, alternatively, are able toactively suppress ROS formation (see below).
Sporozoites induce the generation of intracellular cAMPin Kupffer cells
Previous work has shown that malaria sporozoites passthrough Kupffer cells before invading hepatocytes (Pradeland Frevert, 2001) and that CSP binds to proteoglycansand LRP-1 on the surface of mammalian cells (Shakibaeiand Frevert, 1996). Because ligand binding to LRP-1causes a cAMP increase in macrophages (Goretzki andMueller, 1997; Misra et al., 2002b; Misra and Pizzo, 2002;Zhu and Hui, 2003), we asked whether sporozoites areable to induce the generation of this second messenger inKupffer cells. Sporozoites were incubated with Kupffercells at a ratio of two sporozoites per Kupffer cell andintracellular cAMP levels were analysed after coincuba-tion for 30 or 60 min. Exposure to P. berghei (Fig. 1A) orP. yoelii sporozoites (not shown) increased the cAMPconcentration in Kupffer cells significantly, while controlextracts prepared from the same number of uninfectedAnopheles stephensi salivary glands had no effect. Thus,malaria sporozoites elicit the generation of cAMP inKupffer cells.
Circumsporozoite protein raises the intracellular cAMPconcentration in Kupffer cells
To elucidate the mechanism of the sporozoite-inducedelevation of cAMP in Kupffer cells, we performed similarexperiments with recombinant Plasmodium falciparumCSP. Incubation with CSP induced a concentration-dependent elevation of the cAMP level in Kupffer cells withmaximum (threefold) increase at 50 nM (Fig. 1B). Theeffect of CSP on the intracellular cAMP level in Kupffer cells
was also time-dependent; immediately upon CSP binding,the cAMP concentration began to rise and reached amaximum between 30 and 60 min (Fig. 1C). The intracel-lular cAMP level depends on the balance betweensynthesis by adenylyl cyclase and degradation byphosphodiesterase (Sunahara and Taussig, 2002).Preincubation of Kupffer cells with IBMX, a specificphosphodiesterase inhibitor, did not change the CSP-induced generation of cAMP suggesting that the elevationof cAMP was due to activation of adenylyl cyclase ratherthan inhibition of phosphodiesterase (Fig. 1D). Interest-ingly, CSP had no effect on the cAMP concentration inother types of liver cells (hepatocytes and endothelial cells)or in the human hepatoma cell line HepG2 (Table 2) sug-gesting that the CSP-induced signalling is specific forKupffer cells.
LRP-1 expression on the surface of liver cells
To determine whether this selective responsiveness ismirrored by a cell type-specific expression of LRP-1 in therat liver, frozen sections were labelled with mAb 8B8, amonoclonal antibody specific for the 85 kDa light chain ofLRP-1 (Strickland et al., 1990) in combination with proteinA-FITC or protein G Alexa 488. In agreement with earlierwork (Moestrup et al., 1992), LRP-1 was found in highconcentrations on the surface of Kupffer cells as well aslining the sinusoids (Fig. 2A). However, the close associa-tion between hepatocytes and sinusoidal endothelia alongthe space of Disse made it difficult to assign the LRP-1label to one or the other cell type. We therefore deter-mined the contribution of the various liver cell types to thissinusoidal labelling pattern using cultures of purifiedprimary rat hepatocytes, Kupffer cells and sinusoidalendothelia. Confocal microscopy revealed high expres-sion levels of LRP-1 on the surface of hepatocytes(Fig. 2B) and Kupffer cells (Fig. 2C), while sinusoidalendothelia were only faintly labelled (Fig. 2D). HumanHepG2 hepatoma cells, which had previously been
Table 1. NBT reduction in Kupffer cells cocultivated with Plasmodium sporozoites.
Experiment Treatment
Per cent SPZ with positive NBT reaction
Viable SPZ Inactivated SPZ
1 –PMA 7.8 � 1.4 65.2 � 1.7+PMA 13.9 � 4.5 74.3 � 2.9
2 –PMA 13.6 � 3.8 75.6 � 4.4+PMA 28.2 � 2.6 79.8 � 2.0
Plasmodium yoelii sporozoites (SPZ) were incubated with Kupffer cells at a ratio of 1:1 for 45 min in RPMI containing 10% FCS and 0.1% nitro-bluetetrazolium (NBT). After washing with PBS, the cultures were fixed in 0.1% glutaraldehyde in PBS. Only close parasite host cell events wereanalysed for formazan formation surrounding the sporozoites. The data are represented as average measurements from triplicate wells � STD.Kupffer cells were activated with 1 mm PMA 5 min before sporozoite addition. Sporozoites were inactivated by four cycles of freeze in liquid nitrogenand thawing at 37°C.
2612 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628
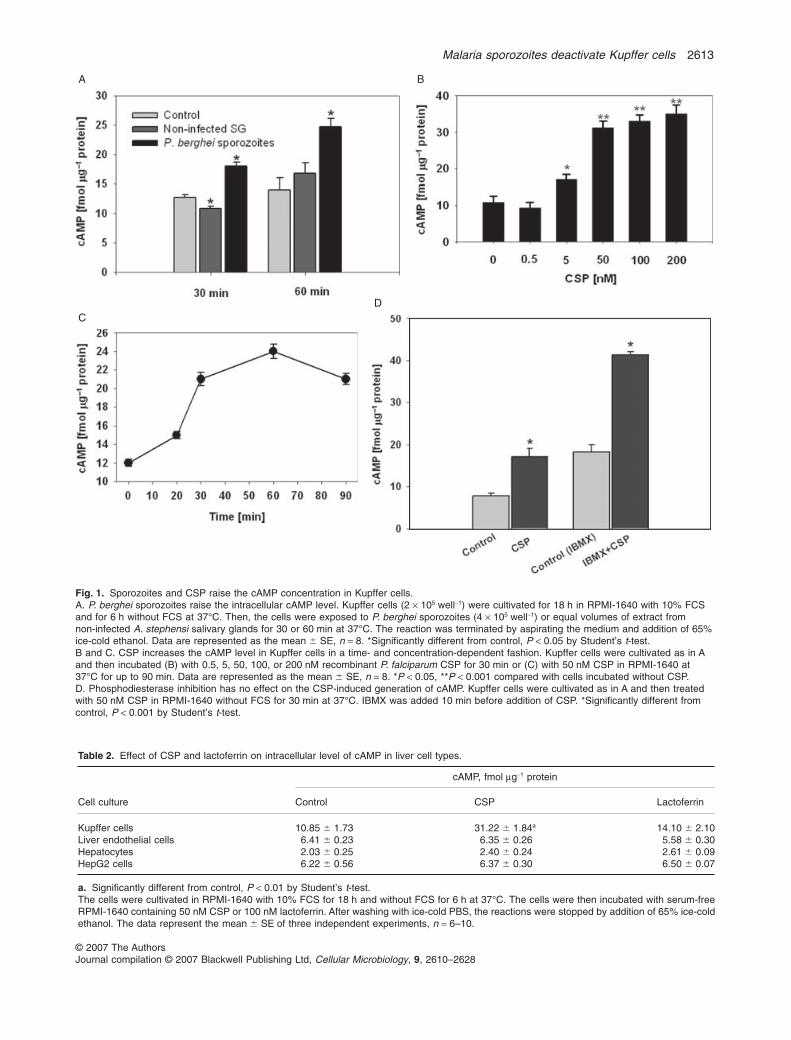
Table 2. Effect of CSP and lactoferrin on intracellular level of cAMP in liver cell types.
Cell culture
cAMP, fmol mg-1 protein
Control CSP Lactoferrin
Kupffer cells 10.85 � 1.73 31.22 � 1.84a 14.10 � 2.10Liver endothelial cells 6.41 � 0.23 6.35 � 0.26 5.58 � 0.30Hepatocytes 2.03 � 0.25 2.40 � 0.24 2.61 � 0.09HepG2 cells 6.22 � 0.56 6.37 � 0.30 6.50 � 0.07
a. Significantly different from control, P < 0.01 by Student’s t-test.The cells were cultivated in RPMI-1640 with 10% FCS for 18 h and without FCS for 6 h at 37°C. The cells were then incubated with serum-freeRPMI-1640 containing 50 nM CSP or 100 nM lactoferrin. After washing with ice-cold PBS, the reactions were stopped by addition of 65% ice-coldethanol. The data represent the mean � SE of three independent experiments, n = 6–10.
A B
CD
Fig. 1. Sporozoites and CSP raise the cAMP concentration in Kupffer cells.A. P. berghei sporozoites raise the intracellular cAMP level. Kupffer cells (2 ¥ 105 well-1) were cultivated for 18 h in RPMI-1640 with 10% FCSand for 6 h without FCS at 37°C. Then, the cells were exposed to P. berghei sporozoites (4 ¥ 105 well-1) or equal volumes of extract fromnon-infected A. stephensi salivary glands for 30 or 60 min at 37°C. The reaction was terminated by aspirating the medium and addition of 65%ice-cold ethanol. Data are represented as the mean � SE, n = 8. *Significantly different from control, P < 0.05 by Student’s t-test.B and C. CSP increases the cAMP level in Kupffer cells in a time- and concentration-dependent fashion. Kupffer cells were cultivated as in Aand then incubated (B) with 0.5, 5, 50, 100, or 200 nM recombinant P. falciparum CSP for 30 min or (C) with 50 nM CSP in RPMI-1640 at37°C for up to 90 min. Data are represented as the mean � SE, n = 8. *P < 0.05, **P < 0.001 compared with cells incubated without CSP.D. Phosphodiesterase inhibition has no effect on the CSP-induced generation of cAMP. Kupffer cells were cultivated as in A and then treatedwith 50 nM CSP in RPMI-1640 without FCS for 30 min at 37°C. IBMX was added 10 min before addition of CSP. *Significantly different fromcontrol, P < 0.001 by Student’s t-test.
Malaria sporozoites deactivate Kupffer cells 2613
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

shown to support LRP-1-mediated endocytosis of CSP(Shakibaei and Frevert, 1996), expressed a high LRP-1concentration similar to hepatocytes (data not shown).These data explain the absence of CSP-mediated eleva-tion of cAMP in sinusoidal endothelia and also suggestthat components crucial for the signalling cascade in mac-rophages are missing in hepatocytes.
Blockage of LRP-1 prevents CSP-induced signalling inKupffer cells
To evaluate the contribution of LRP-1 to the CSP-mediated generation of cAMP, we used the universalinhibitor RAP, which blocks the binding of CSP and allother ligands to LRP-1 (Herz and Strickland, 2001; Strick-land and Ranganathan, 2003). Preincubation with a10-fold excess of RAP (500 nM) blocked the CSP-inducedgeneration of cAMP in Kupffer cells, while 500 nM RAPalone had no effect (Fig. 3A). Lactoferrin, another LRP-1ligand, had no significant effect on the cAMP level in anyof the liver cell types studied (Table 2). These datasuggest that LRP-1 is crucial for the cAMP elevation inKupffer cells.
Because in addition to cAMP, a2M* also increases theconcentration of IP3 in peritoneal macrophages (Misraand Pizzo, 1998a,b), we analysed the effect of CSP
binding to Kupffer cells and hepatocytes on the genera-tion of this second messenger. Upon addition of 50 nMCSP, Kupffer cells responded with a time-dependentincrease in the intracellular IP3 concentration, while hepa-tocytes showed no effect (Fig. S2). Preincubation ofKupffer cells for 10 min with 500 nM RAP before additionof CSP prevented this increase, and RAP alone had noeffect (data not shown). Taken together, these datasupport the concept that the CSP-induced signalling isselective for Kupffer cells and that LRP is cruciallyinvolved in these events.
Cell surface glycosaminoglycans (GAGs) contribute tothe CSP-mediated signalling in Kupffer cells
Because syndecans are involved in the binding of CSP toKupffer cells (Pradel et al., 2002), we asked whether theelevation of cAMP in Kupffer cells can be prevented byelimination of GAGs as CSP binding sites by digestionwith specific GAG lyases (Frevert et al., 1993) in combi-nation with sodium chlorate treatment (Baeuerle andHuttner, 1986; Pinzon-Ortiz et al., 2001). Chlorate treat-ment alone inhibited the CSP-induced generation ofcAMP in Kupffer cells by 20% (Fig. 3B). In combinationwith GAG lyase digestion using a cocktail of heparinase,
Fig. 2. LRP-1 expression is liver celltype-specific.A. LRP-1 is associated with the sinusoids andthe surface of Kupffer cells (arrow) on frozenliver sections.B–D. In cultures of purified liver cells, LRP-1is detectable in large amounts on hepatocytes(B) and Kupffer cells (C), but not onsinusoidal endothelia (D). The cells werecultivated for 18 h and then fixed. LRP-1 wasdetected with mAb 8B8 directed against thelight chain of the receptor in combination withFITC-conjugated protein A (green). Evansblue (shown in red) was used as acounter-stain.
2614 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

heparitinase and chondroitinase ABC, chlorate treatmentreduced the cAMP increase by 50%, indicating that syn-decans contribute to the CSP-induced cAMP productionin Kupffer cells.
Sporozoites use a cAMP-dependent mechanism toinhibit the respiratory burst in Kupffer cells
Agents that stimulate the adenylyl cyclase generallyinhibit the generation of ROS (Nielson et al., 1992). Basedon the finding that sporozoites and CSP elevate the cAMPlevel in Kupffer cells, we hypothesized that malaria sporo-zoites are able to suppress the respiratory burst in thesemacrophages. To examine the effect of live parasites onthe respiratory burst that is normally elicited by phagocy-tosis of zymosan, we exposed Kupffer cells to P. yoeliisporozoites and measured ROS production by luminol-enhanced chemiluminescence (Wang et al., 1993). Iden-tically prepared extracts from uninfected salivary glandswere used as a control to determine the effect of mosquitotissue debris and bacteria, which contaminate routinesporozoite preparations in large quantities (Mack et al.,1978; U. Frevert, unpublished). Neither P. yoelii sporozoi-tes nor uninfected salivary gland extracts had any effecton the chemiluminescence within the 60 min incubationperiod (Fig. 4A) suggesting that neither sporozoites norbacteria or salivary gland debris induce a respiratoryburst in resting Kupffer cells. Subsequent exposure tozymosan, however, triggered burst activity with amaximum chemiluminescence response at 10 min(Fig. 4A). In relation to control cells incubated with HBSS,Kupffer cells that had been exposed to extract from unin-fected control glands showed an enhanced ROS genera-tion in response to zymosan (Fig. 4A) suggesting that thesalivary gland debris and/or the bacteria had a primingeffect on the Kupffer cells. Sporozoites inhibited thisenhanced burst activity by 78%. The specific adenylylcyclase inhibitor SQ 22 536 abrogated the inhibitory effectof sporozoites on the zymosan-induced respiratory burst(Fig. 4A), suggesting that the increased cAMP concentra-tion was responsible for the inhibition of ROS production.Indeed, the zymosan-induced respiratory burst wasinhibited by 22% and 67%, respectively, when the intra-cellular cAMP concentration was raised by db-cAMP, amembrane-permeable cAMP analogue, or with the phos-phodiesterase inhibitor IBMX (Fig. 4B). Taken together,the data indicate that the increased cAMP levels lead to ablock in ROS formation.
Circumsporozoite protein is responsible for thesporozoite-mediated suppression of ROS formation
Preincubation of Kupffer cells with recombinantP. falciparum CSP inhibited the zymosan-induced ROSproduction a dose-dependent fashion, ranging from 16%at 5 nM to 43% at 100 nM (Fig. 5A), confirming the roleof CSP in the inhibition of the respiratory burst bysporozoites. To exclude that Escherichia coli-derived(heat-resistant) lipopolysaccharide (LPS) contributed to
A
B
Fig. 3. Blockage of LRP-1 and syndecans prevents the CSP-induced generation of cAMP in Kupffer cells.A. RAP inhibits the CSP-mediated cAMP elevation. Kupffer cellswere cultivated in RPMI-1640 with 10% FCS for 18 h and withoutFCS for 6 h at 37°C. The cells were then incubated with 50 nM CSPin RPMI-1640 without FCS for 30 min. RAP (500 nM) was added tosome wells 10 min before addition of CSP. The reaction was termi-nated by aspirating the medium and adding 65% ice-cold ethanol.*P < 0.001 compared with cells treated with CSP.B. Combined inhibition of GAG sulphation and digestion of surfaceGAGs inhibits the CSP-induced generation of cAMP. Kupffer cellswere incubated for 18 h at 37°C with RPMI-1640 with 10% FCS orwith a 4:1 mixture of Ham’s F-12 and RPMI-1640 medium containing20 mM sodium chlorate and 2% desalted FCS. The cells were thendigested for 3 h with 20 mU ml-1 heparinase, 20 mU ml-1 hepariti-nase and 500 mU ml-1 chondroitinase ABC in the same mediumwithout FCS. Subsequently, 50 nM CSP was added for another30 min. The data represent the CSP-induced cAMP elevation in percent. Results are means � SE of three independent experiments.*P < 0.05, **P < 0.001 compared with control.
Malaria sporozoites deactivate Kupffer cells 2615
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

the observed effect of CSP on ROS formation, we studiedthe effect of heat-inactivation on the activity of our recom-binant CSP preparation. Boiling for 10 min at 95° com-pletely prevented the CSP-mediated suppression of therespiratory burst in Kupffer cells indicating that LPS wasnot responsible for this activity. This is supported by thefinding that elimination of LPS from the incubationmedium by polymyxin B did not change the effect of CSPon the respiratory burst, but completely prevented theeffect of E. coli 055:B5 LPS (data not shown). We alsocompared the time-dependent effect of CSP and LPS on
A
B
Fig. 4. Sporozoites suppress the respiratory burst in Kupffer cells.Kupffer cells (1.5–2.5 ¥ 105 well-1) were cultivated for 18 h at 37°Cin 96-well plates in RPMI-1640 containing 10% FCS and foranother 6 h in the same medium without FCS. After washing, thecells were incubated for 30–60 min with HBSS in the presence of(A) P. yoelii sporozoites (4 ¥ 105 well-1) with or without 100 mM ofthe adenylyl cyclase inhibitor SQ 22 536 or extract fromnon-infected A. stephensi salivary glands, or (B) 1 mM IBMX or1 mM db-cAMP. The production of ROS was determined byluminol-enhanced chemiluminescence. The kinetics of lightemission was analysed before and after stimulation of the cells withnon-opsonized zymosan. The results are expressed as relative lightunits (RLU) and represent the mean of four to six wells from one ofthree representative experiments.
A
B
C
Fig. 5. CSP blocks the generation of ROS in Kupffer cells. Kupffercells (1.5–2.5 ¥ 105 well-1) were cultivated for 48 h at 37°C in96-well plates in phenol red-free RPMI-1640 containing 10% FCSand for another 6 h in the same medium without FCS. Afterwashing, the cells were incubated for 0–90 min with HBSScontaining (A) 5, 50, or 100 nM CSP; (B) 100 nM CSP or 1 mg ml-1
LPS; (C) 100 nM CSP.A and B. The kinetics of luminol-enhanced light emission wasanalysed after stimulation of the cells with non-opsonized zymosan.C. Superoxide production was monitored for 60 min bylucigenin-enhanced chemiluminescence after stimulation with200 nM PMA and expressed as per cent compared with the control,to which equal volumes of vehicle were added. *P < 0.05;**P < 0.01 versus control (0 min).
2616 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

the production of superoxide anions and found that thesuppressive effect of CSP peaked 30 min after addition ofthe recombinant protein and slowly decreased thereafter,while LPS began to show a small inhibitory effect onlyafter 90 min (Fig. 5B). Because LPS can induce cyclooxy-genase in Kupffer cells, thereby triggering the productionof prostaglandins (Victorov and Hoek, 1995), we assumethat the observed delayed LPS effect is a result of aprostanoid-mediated cAMP elevation. Taken together, thedata suggest that sporozoites use CSP to block the res-piratory burst.
Circumsporozoite protein blocks the respiratory burstdownstream from PKC
By phosphorylating p47phox, PKC is involved in the assem-bly of the NADPH oxidase and functions as an importantregulator of the respiratory burst (Reeves et al., 1999). Todefine the action of CSP on the NADPH oxidase activationcascade in relation to PKC, Kupffer cells were stimulatedwith PMA, a synthetic analogue of DAG and activator ofPKC. PMA induced a rapid generation of ROS in Kupffercells, which was inhibited by 47% by a 30 min preincuba-tion of the cells with 100 nM CSP (Fig. 5C). CSP treat-ment alone did not induce the production of ROS inKupffer cells (Fig. 5A and C). These results suggest thatthe CSP-mediated block of the NADPH oxidase activationcascade occurs downstream from PKC.
The inhibition of the respiratory burst iscAMP-dependent, but PKA-independent
To obtain further insight into the ROS signalling cascadein Kupffer cells, we used lucigenin-enhanced chemilumi-nescence to investigate the effect of adenylyl cyclaseinhibitors on the CSP-induced suppression of the respira-tory burst. Figure 6 demonstrates that the adenylylcyclase inhibitor SQ 22 536 reversed the suppressiveeffect of CSP on superoxide generation, suggesting thatadenylyl cyclase activation led to the inhibition of theNADPH oxidase. Because PKA activation has beenreported to interfere with the assembly of the NADPHoxidase in human neutrophils by preventing the PKC-mediated phosphorylation of p47phox (Bengis-Garber andGruener, 1996), we also tested the effect of PKAinhibitors. Preincubation of Kupffer cells with the specificPKA inhibitors H-89, PKI14-22 and Rp-cAMP (Gjertsenet al., 1995) failed to prevent the CSP-mediated inhibitionof the respiratory burst (Fig. 6A). These data indicate thatthe intracellular cAMP increase is responsible for theCSP-mediated inhibition of the respiratory burst, but thatan alternative, PKA-independent pathway prevents theactivation of the NADPH oxidase in Kupffer cells.
Exchange protein directly activated by cAMP regulatesROS formation in Kupffer cells
Because activation of the ‘exchange protein directly acti-vated by cAMP’ (EPAC) is known to suppress ROS for-mation in human alveolar macrophages by regulating theassembly of ROS-producing NADPH oxidase (Aronoffet al., 2005), we considered the possibility that EPACdoes this in Kupffer cells as well. To test the ability of bothPKA and EPAC to regulate the respiratory burst in Kupffer
A
B
Fig. 6. Suppression of the zymosan-induced respiratory burst inKupffer cells by CSP is cAMP- and EPAC-dependent, butPKA-independent.A. Kupffer cells (1.5–2.5 ¥ 105 well-1) were cultivated for 48 h at37°C in 96-well plates in phenol red-free RPMI-1640 containing10% FCS and for another 6 h in the same medium without FCS.After washing, the cells were pretreated for 30 min with or withoutthe adenylyl cyclase inhibitor SQ 22 536 or the protein kinase Ainhibitors PKI14-22, H-89 and Rp-cAMP) followed by incubation with100 nM CSP for 30 min. The kinetics of light emission wasanalysed after stimulation of the cells with non-opsonized zymosan.Superoxide production was monitored for 60 min bylucigenin-enhanced chemiluminescence and expressed as per centof the control, to which equal volumes of vehicle or inhibitors wereadded. *P < 0.001.B. Kupffer cells were pretreated for 40 min with 3 mM of the EPACactivator 8-CPT-2-Me-cAMP, the PKA activator 6-MB-cAMP, or bothactivators. Superoxide production was monitored byluminol-enhanced chemiluminescence. The data are expressed asrelative light units (RLU) and the values represent the mean of fourwells from one representative of three experiments.
Malaria sporozoites deactivate Kupffer cells 2617
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

cells, we used selective PKA and EPAC activators(Rehmann et al., 2003; Seino and Shibasaki, 2005).Figure 6B shows that when incubated with 3 mM concen-trations, the selective EPAC activator 8-CPT-2-Me-cAMPinhibited the zymosan-induced ROS formation by Kupffercells, while the PKA activator 6-MB-cAMP had no effect.Simultaneous incubation with both 8-CPT-2-Me-cAMPand 6-MB-cAMP did not exceed the effect of the EPACactivator alone. Thus, EPAC activation can inhibit ROSproduction by rat Kupffer cells.
Discussion
Malaria sporozoites pass through Kupffer cells to reachhepatocytes, their initial site of multiplication in the mam-malian host (Frevert et al., 2005; 2006a,b; Baer et al.,2007). Here we show that sporozoite contact not only failsto induce a respiratory burst in Kupffer cells, but alsosuppresses the induction of ROS formation in response tosecondary stimuli. By using the high-affinity interactionbetween CSP and the scavenger receptor LRP-1 (Shak-ibaei and Frevert, 1996), sporozoites raise the intracellu-lar cAMP concentration in Kupffer cells, thus preventingthe fatal consequences of the assembly of the NADPHoxidase (see model in Fig. 7). Syndecans contribute tothe induction of the signalling cascade by providing alarge number of multivalent CSP-binding GAG chains(Pradel et al., 2002), thereby facilitating the binding ofCSP to LRP-1. These findings explain why malaria sporo-zoites survive the Kupffer cell passage unharmed. In addi-
tion, suppression of respiratory burst activity in responseto subsequent stimuli avoids the establishment of a pro-inflammatory microenvironment and tissue injury thusenabling Plasmodium liver stages to develop in hepato-cytes in the vicinity of the traversed Kupffer cells. Thus,Plasmodium sporozoites exploit the overall macrophagedeactivating and anti-inflammatory properties of cAMP tosuppress ROS production in Kupffer cells, thus facilitatingthe establishment of a malaria infection.
The archetypal second messenger cAMP, which isresponsible for the conversion of many intercellularsignals to intracellular events, has been implicated in themodulation of numerous immunologic reactions. In mac-rophages including Kupffer cells, increased cAMP levelsinhibit phagocytosis, phagolysosomal fusion, the LPS-mediated induction of macrophage activation, and killingof carcinoma cells (Lowrie et al., 1979; Sionov and Gallily,1990; Newman et al., 1991; Chaves et al., 1996; Rossiet al., 1998; Nogueira-Machado et al., 2003; Aronoffet al., 2005). There is also solid evidence that cAMPelevating agents suppress the respiratory burst in varioustypes of phagocytic cells in a dose-dependent fashion(Hoffman et al., 1983; Mitsuyama et al., 1993; Wanget al., 1998; Martins Chaves et al., 2002; Nogueira-Machado et al., 2003; O’Dowd et al., 2004). However,information on the exact time sequence by which thevarious subunits assemble to form the functional NADPHoxidase and the molecular events that control the onset ofthe assembly cascade is scarce, and data on the NADPHoxidase from Kupffer cells are lacking. In human neutro-
Fig. 7. Model for the CSP-mediated inhibition of ROS formation in Kupffer cells. Plasmodium sporozoite adhesion to Kupffer cells is mediated bya multivalent, high-avidity interaction between the major sporozoite surface protein CSP and chondroitin and heparan sulphate chains from syn-decans. This interaction facilitates engagement of some CSP molecules in a specific, high affinity interaction with LRP-1. The ensuing activationof a Gas protein stimulates adenylyl cyclase leading to the formation of a local pool of cAMP. LRP-1 ligation also rapidly increases the IP3 level,which triggers the release of calcium from intracellular stores thus contributing to adenylyl cyclase activation. Phosphodiesterases control thecAMP level by converting it to AMP. The increase in the intracellular cAMP concentration induces EPAC activation, which prevents ROS forma-tion. Because CSP inhibits the respiratory burst also after direct stimulation of PKC with PMA, EPAC may prevent the PKC-mediated phosphory-lation of p47phox. The resulting block in the assembly of the NADPH oxidase prevents the formation of ROS.
2618 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

phils, elevation of cAMP by prostaglandins, histamine,adenosine and b-adrenergic agonists inhibit superoxidegeneration in response to stimulation with the chemotacticpeptide formyl-methionyl-leucyl-phenylalanine (fMLP)(Cronstein et al., 1985; Sedgwick et al., 1985; Nielson,1987). The mechanism of suppression has been sug-gested to involve inhibition of phospholipase D (Tyagiet al., 1991) and phospholipase A2 signalling pathways(Nielson et al., 1992), resequestration of cytosolic calciumby upregulation of the endomembrane calcium-ATPase(Anderson et al., 1998), and PKA-dependent downregu-lation of p47phox phosphorylation (Bengis-Garber andGruener, 1996). CSP inhibited the respiratory burst inKupffer cells both after exposure to zymosan and afterdirect stimulation of PKC with PMA suggesting that thecAMP-mediated block of ROS formation occurred at ordownstream from PKC. However, unlike human neutro-phils, in which PKA downregulates the phosphorylation ofp47phox and specific PKA inhibitors consequently restorethe cAMP-mediated inhibition of the respiratory burst (Mit-suyama et al., 1993; Bengis-Garber and Gruener, 1996),PKA does not appear to play a role in the CSP-mediatedsuppression of the respiratory burst in Kupffer cells. Analternative pathway known to link cAMP increase to block-age of ROS formation in some cells involves members ofthe EPAC family of cAMP binding proteins (Kawasakiet al., 1998; de Rooij et al., 1998). Activation of either PKAor EPAC-1 in alveolar macrophages inhibits H2O2 produc-tion in a dose-dependent fashion (Aronoff et al., 2005).We used the selective EPAC and PKA activators 8-CPT-2-Me-cAMP and 6-MB-cAMP (Rehmann et al., 2003;Seino and Shibasaki, 2005), respectively, and found thatEPAC activation inhibits zymosan-induced ROS formationby Kupffer cells, but PKA activation does not. Although wecould not test the role of EPAC using selective inhibitorsbecause none is available, we did use the specific PKAinhibitors H-89, PKI14-22 and Rp-cAMP and found theyhad no effect on the CSP-mediated inhibition of ROSformation. Rac activation blocks ROS production in somecells, but there is no evidence that Rac mediates cAMP-mediated inhibition of ROS in macrophages (Bokoch andDiebold, 2002; Diebold and Bokoch, 2005; Hordijk, 2006).Based on these data and the lack of any other knownmechanism linking increased cAMP and inhibition of ROSproduction, we consider it likely that Plasmodium CSPactivates a cAMP/EPAC-dependent pathway in Kupffercells that interferes with the complex assembly cascade ofthe NADPH oxidase.
Signalling through syndecans is not associated with anincrease in the intracellular cAMP level (Carey, 1997;Woods and Couchman, 1998; Zimmermann and David,1999; Rapraeger, 2000). We assume therefore that thecontribution of syndecans to the observed signalling isto provide a multivalent, high-avidity platform on the
macrophage surface that facilitates the transfer of CSP toLRP-1. On the other hand, LRP-1 has been reported tostimulate adenylyl cyclase directly by interacting with aGás protein (Goretzki and Mueller, 1998). Despite itsnature as a universal scavenger receptor and its fairlywide tissue distribution (Moestrup et al., 1992), LRP caninduce selective, cell type-specific signalling cascades bycooperating with a variety of signalling receptors (Herzand Strickland, 2001; Boucher and Gotthardt, 2004).Taken together, the data support a model in which LRP-1ligation leads to Gás-mediated adenylyl cyclase stimula-tion as well as to PLC activation; the resulting IP3 increasetriggers the release of calcium from intracellular stores,which in turn further stimulates cAMP formation. Weobserved by intravital microscopy that after adhering tothe surface of a Kupffer cell, sporozoites pause for a fewminutes before starting the entry process (Frevert et al.,2005). This time may be required to accumulate sufficientcAMP to ensure protection against the fatal conse-quences of the respiratory burst.
Other microbial products are also known to inhibit therespiratory burst in phagocytes. For example, Leishmaniaspp., which replicate in macrophages, use surface lipo-phosphoglycan (LPG) to block oxidative damage by non-enzymatic mechanisms: LPG, besides being a potentROS scavenger, can also prevent the respiratory burstby directly inhibiting PKC, analogous to bacterial LPS(Buchmuller-Rouiller and Mauel, 1987; McNeely et al.,1989; Frankenburg et al., 1990; Brandonisio et al., 1994;Miller and Britigan, 1997). Legionella pneumophila, a bac-terial pathogen that infects various macrophages, uses ametalloproteinase to inhibit the fMLP- or zymosan-induced ROS formation in monocytes and polymorph-nuclear blood macrophages (Sahney et al., 2001). Theapicomplexan Toxoplasma gondii, a distant cousin ofPlasmodium that replicates in many cell types includingmacrophages, possesses a well-developed enzymaticantioxidant system as a defence mechanism against oxi-dative stress (Ding et al., 2004). A broad range of antioxi-dant enzymes including SODs, a catalase, glutathioneperoxidases, and peroxiredoxins enable T. gondii to copewith the pro-oxidative intracellular environment of theirphagocytic host cells (Ding et al., 2004). P. falciparumblood stages are highly adapted to the oxidative microen-vironment of erythrocytes. Lacking a catalase and glu-tathione peroxidases, P. falciparum blood stages rely on athioredoxin redox cycle, peroxiredoxins and plasmore-doxin for efficient detoxification of ROS (Müller, 2004;Nickel et al., 2006). Thus, a broad variety of differentantioxidant systems have evolved in many parasitic bac-teria and protozoa; these potent virulence factors havebeen recognized as potential drug targets (Miller and Briti-gan, 1997; Steenkamp, 2002; Becker et al., 2004; Dinget al., 2004; Müller, 2004).
Malaria sporozoites deactivate Kupffer cells 2619
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

Plasmodium faces enormous environmental changes inthe course of its complex life cycle. Because parasitesurvival depends on a delicate balance between pro-andantioxidant activities, disturbances in oxidant homeosta-sis are likely to occur. For example, despite their entirelyfermentative lifestyle, P. falciparum blood stages are nev-ertheless highly susceptible to alterations of the redoxequilibrium (Wozencraft et al., 1984; Müller, 2004). In fact,the parasites may have adopted a strictly anaerobicmetabolism, because they are unable to cope with redoxstress in addition to the oxidative environment of theerythrocyte (Oliveira and Oliveira, 2002). While attemptsto establish a role of NADPH oxidase-derived ROS in thecontrol of the parasitaemia failed (Sanni et al., 1999;Gillman et al., 2004), blood stage parasites are clearlyvulnerable to oxidative damage via the xanthine oxidasepathway (Dockrell and Playfair, 1984; Ockenhouse et al.,1984; Wozencraft et al., 1984; Greve et al., 1999) and topro-oxidant drugs such as artimisinins (Dong and Venner-strom, 2003; Tang et al., 2004). In addition, increasedmelanotic encapsulation of P. berghei ookinetes in refrac-tory mosquitoes has been correlated with elevated levelsof oxidative stress (Dimopoulos et al., 2002; Kumar et al.,2003). Consequently, impaired parasite development inand transmission by these unsuitable vectors demon-strates the susceptibility of Plasmodium ookinetes toROS-inflicted damage (Kumar et al., 2003). Previouswork showing that opsonization renders P. berghei sporo-zoites susceptible to killing by Kupffer cells or peritonealmacrophages (Danforth et al., 1980; Seguin et al., 1989),further supports the notion that P. yoelii sporozoites aresensitive to respiratory burst activity. In addition, a searchof P. yoelii EST databases indicated that, compared withblood stages, sporozoites and liver stage parasites under-express or lack most of the proteins with antioxidativefunction that are expressed in blood stages. Except forSOD and thioredoxin, no EST hit for other detoxificationproteins was found. The nature of the detoxificationsystems and their exact roles during the various life cyclestages of Plasmodium remain await experimental confir-mation, but all available evidence suggests that Plasmo-dium sporozoites depend on manipulation of Kupffer cellfunction to prevent oxidative damage during their passageinto the liver parenchyma. Kupffer cell stimulation withPMA prior to sporozoite addition caused only a twofoldincrease in the number of killed parasites (Table 1). Weattribute this relatively modest effect to the fact thatlive Plasmodium sporozoites continuously release CSPfrom their surface (Stewart and Vanderberg, 1991). Wepropose that by binding to Kupffer cells, the shed CSPprevented the strong local accumulation of ROS, whichwas typically observed in association with deadsporozoites. Future studies with p47 phox–/– or gp91phox–/– mice, both of which lack a functional NADPH
oxidase (Jackson et al., 1995; Pollock et al., 1995), maysupport the notion that Plasmodium sporozoite infectionof the liver depends on suppression of the respiratoryburst in Kupffer cells.
Mosquito salivary gland extract had a priming effect onKupffer cells as shown by the considerably enhancedrespiratory burst activity upon subsequent exposure tozymosan. We interpret this phenomenon as a responseof the phagocytes to the bacteria and/or mosquitotissue debris, major contaminants of routine sporozoitepreparations. Importantly, sporozoites suppressed thisenhanced burst activity by 78% suggesting that in additionto the CSP-mediated block of ROS formation, the para-sites are also able to prevent macrophage priming. Neu-trophil priming can occur within seconds to minutes(Condliffe et al., 1998; Binder et al., 1999; Dewas et al.,2003) thus explaining the rapid reaction of the Kupffercells to salivary gland extract. While Kupffer cells arenot exposed to mosquito contaminants under naturaltransmission conditions, sporozoites cause substantialdamage to hepatocytes while migrating to their final des-tination in the liver (Mota et al., 2001; Frevert et al., 2005),and this may be one reason why Plasmodium hasacquired a mechanism to avoid macrophage priming inresponse to tissue injury. Another important considerationin this context is that many in vivo studies involving Plas-modium infection of the liver are performed by intrave-nous injection of isolated sporozoites. Because themajority of the contaminants contained in such sporozoitepreparations must be expected to be removed from theblood by Kupffer cells, suppressive or otherwise modulat-ing effects on Plasmodium liver stage development due topriming of the entire Kupffer cell population cannot beexcluded. Sporozoite purification by affinity chromatogra-phy (Mack et al., 1978; Kappe et al., 2001) should helpminimize such unpredictable effects.
To define the role of LRP-1 in sporozoite infection of theliver, transgenic LRP-1flox/flox Cre+ mice with a functionaldefect of LRP-1 in hepatocytes were generated by condi-tional gene targeting (Rohlmann et al., 1996; 1998). Onelikely explanation for the unexpected finding that infectionof these LRP-1-deficient animals with P. yoelii sporozoiteswas not diminished (Marshall et al., 2000) is that the Creadenovirus Ad5 used in these studies infects predomi-nantly hepatocytes and to a minor degree also sinusoidalendothelia (Herz and Gerard, 1993), but not Kupffer cells(Shayakhmetov et al., 2004). Thus, LRP-1-mediatedsignal transduction was likely unaffected in the Kupffercells of the LRP-1flox/flox Cre+ mice, explaining the unalteredrate of P. yoelii sporozoite entry into the liver.
The host’s adenylyl cyclase/phosphodiesterase systemrepresents a key target for many microorganisms to sabo-tage macrophage function. For instance, pathogenicbacteria such as Bordetella pertussis, Bacillus anthracis,
2620 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

Pseudomonas aeruginosa and Yersinia pestis secreteadenylate cyclase toxins that become activated byeukaryotic cofactors and either modulate or completelyshut down the normal cell function (Ahuja et al., 2004). Byraising the cAMP concentration in neutrophils, forexample, the B. anthracis oedema toxin blocks phagocy-tosis and inhibits both particulate and PMA-inducedsuperoxide production. In lymphocytes, this bacterialenzyme alters the expression of immuno-regulatorygenes and inhibits proliferation. Parasites such asT. gondii, Giardia lamblia, Entamoeba histolytica, Schis-tosoma mansoni and Trypanosoma cruzi manipulate thecAMP level in various cells of the host to down-modulateNO production, inhibit ROS formation, induce IL-6 synthe-sis, and inhibit contractility and locomotion (Rico et al.,1995; Rodriguez et al., 1999; Angeli et al., 2001; Joensenet al., 2003; Rozenfeld et al., 2003; Rawal et al., 2004;Shant et al., 2004). Primary targets are immune effectorcells, whose deactivation naturally facilitates survival ofthe microorganisms in the host. In addition to suppressingROS formation in vitro, Plasmodium sporozoites are alsoable to disrupt the normal Kupffer cell function in vivo:consistent with induction of a cAMP increase (Feng et al.,2002), infectious P. berghei sporozoites inhibit the expres-sion of MHC class I molecules and the production ofIL-12p40 (Steers et al., 2005). Because any alteration inthe concentration of the ubiquitous second messengercAMP has a profound effect on many cellular processes,it must be expected that Plasmodium sporozoites are ableto manipulate other macrophage functions as well.
Experimental procedures
Materials
Heparinase (EC 4.2.2.7), heparitinase (EC 4.2.2.8) and chon-droitinaseABC (EC 4.2.2.4) were from Seikagaku (Falmouth, MA),collagenase H (EC 3.4.24.3) was from Boehringer (Indianapolis,IN). Recombinant human RAP was purchased from InnovativeResearch (Southfield, MI). The mAb 8B8 against the 85 kDa lightchain of LRP-1 was from Maine Biotechnology Services (Portland,ME). Goat anti-mouse IgG conjugated to fluorescein isothio-cyanate (FITC) was from Chemicon (Temecula, CA), proteinA-FITC and protein G-Alexa 488 were from Molecular Probes(Eugene, OR). Zymosan A (Saccharomyces cerevisiae), phorbol12-myristate 13-acetate (PMA), type I collagen, E. coli 055:B5LPS, lactoferrin (LF; EC 3.4.21.-; from bovine colostrum), luminol(5-amino-2,3-dihydro-1,4-phthalazinedione), lucigenin (N,N ′-Dimethyl-9,9′-biacridinium dinitrate), NBT, dibutyryl cAMP (db-cAMP), 9-(Tetrahydro-2-furanyl)-9H-purin-6-amine (SQ 22 536),N-[2-(p-Bromocinnamylamino)ethyl]-5-isoquinolinesulfonamidedihydrochloride (H89), Rp-cAMPS triethylammonium salt (Rp-cAMP) and 3-isobutyl-1 methylxanthine (IBMX) were purchasedfrom Sigma (St. Louis, MO). Myristoylated PKA inhibitor amide14–22 (PKI14-22) was from Calbiochem (San Diego, CA). HeparinSepharose 6 Fast Flow and Percoll were from Amersham (Piscat-away, NJ), Ni-NTA Superflow resin was from Qiagen (Valencia,
CA). Detoxi-Gel AffinityPak prepacked columns were fromPierce (Rockford, IL). The PKA and EPAC activators 8-[4-chlorophenylthio]-2′-O-methyladenosine-3′,5′-cAMP (8-CPT-2-Me-cAMP) and N6-monobutyryladenosine-3′,5′-cAMP (6-MB-cAMP) were from BioLog (Life Science, Arlington Heights, IL).
Purification of recombinant CSP
The P. falciparum CS protein CS27IVC-His6 (AA 27–123[NANPNVDP]3[NANP]21300–411) was expressed in E. coliand purified by two step affinity chromatography using Ni-NTASuperflow and Heparin Sepharose 6 Fast Flow resins (Stüberet al., 1990; Takacs and Girard, 1991). The protein comprisesamino acid sequences representing region I and II-plus as well asthe complete repeat region from the T4 isolate (Cerami et al.,1992). The plasmid encoding the construct was kindly providedby Drs Bela Takacs and Dietrich Stüber (F. Hoffmann-La Roche,Basel, Switzerland). To exclude a potential effect of LPS on ROSformation, we tested our recombinant CSP preparations for con-tamination with LPS using a Limulus amebocyte lysate assay(E-TOXATE Kit, Sigma) according to the instructions of thesupplier. The final concentration of LPS in some of the experi-ments with recombinant CSP was less than 0.06 EU ml-1. LPSwas removed using polymyxin B-agarose prepacked columns(Detoxi-Gel, Pierce) according to manufacturer’s instructions.
Plasmodium sporozoite purification
Plasmodium berghei or P. yoelii sporozoites were isolated frominfected A. stephensi salivary glands. Control extracts from unin-fected salivary glands were prepared in an identical fashion.
Liver cell isolation
Liver cells were isolated from female Sprague Dawley rats(230–250 g). Livers were perfused with collagenase as describedby Neufeld (Neufeld, 1997). Subsequently, liver cells were puri-fied by a combined differential and Percoll gradient centrifugationusing a Sorvall HS4 swinging bucket rotor (Smedsrod andPertoft, 1985). After filtration through a 100-mm mesh polypropy-lene filter (Spectra/Mesh; Spectrum, Laguna Hills, CA), the cellsuspension was centrifuged for 5 min at 50 g resulting in ahepatocyte-enriched pellet and a non-parenchymal cell-enrichedsupernatant. The pellet was resuspended in ice-cold phosphate-buffered saline (PBS) and mixed in an equal volume of PBS-buffered isotonic Percoll. The gradient was centrifuged for 10 minat 50 g. The pellet was resuspended in PBS, centrifuged again,and the final pellet was resuspended in RPMI 1640 mediumcontaining 10% fetal calf serum (FCS). The non-parenchymalcell-enriched supernatants were centrifuged for 15 min at 900 g.The pellet was resuspended in ice-cold PBS, layered over a2-cushion Percoll gradient (Smedsrod and Pertoft, 1985) andcentrifuged for 30 min at 900 g. The intermediate zone, contain-ing mostly liver endothelial cells, and the bottom cushion, con-taining mostly Kupffer cells, were saved, mixed with PBS andcentrifuged for 15 min at 900 g. The pellet was resuspended inPBS with 1% FCS and the cells were further separated intovarious fractions by centrifugal counterflow elutriation (Knook andSleyster, 1976) using the Beckman Coulter JE-5.0 elutriator rotorsupplied with a Sanderson chamber (Beckman Coulter, Palo Alto,
Malaria sporozoites deactivate Kupffer cells 2621
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

CA. USA). The cell suspension was introduced into the elutriatorat a flow rate of 12 ml min-1, and the elutriator speed set to3250 r.p.m. Endothelial cells were obtained at a 32 ml min-1 flowrate, whereas Kupffer cells were collected at 70 ml min-1. Thecells were centrifuged at 200 g and resuspended in RPMI-1640medium containing 10% FCS. The cell purity and viability wereabove 95% as determined by peroxidase staining and Trypanblue exclusion.
Cell culture
Kupffer cells were seeded into 96-well plates (0.25 ¥ 106 cells/well) or 24-well plates (1.0 ¥ 106 cells/well). Endothelial cellswere plated on 24-well plates (1.5 ¥ 106 cells/well), which hadbeen previously coated with type I collagen (Sigma, St. Louis,MO). Rat hepatocytes and HepG2 cells (#HB-8065, ATCC,Rockville, MD) were seeded at a density of 1 ¥ 105 cells cm-2
into Matrigel-coated (BD Biosciences, Bedford, MA) 12-well or24-well plates (Nunc, Napierville, IL). All types of cells werecultivated in RPMI 1640 medium supplemented with 10% FCS,25 mM HEPES and antibiotics (100 U ml-1 penicillin G and100 mg ml-1 streptomycin sulphate) at 37°C in 5% CO2. Non-adherent cells were removed after 1 h by replacing medium, andthe cells were cultured in the same medium for 18 h and then inRPMI-1640 without FCS for 6 h. HepG2 cells were used on day3 after plating.
Cell treatment
To study the effect of CSP on cell signalling, the cells wereincubated with different concentration of CSP for 0–90 min at37°C in 5% CO2. Before use, CSP was dialysed extensively with150 mM NaCl, 10 mM HEPES, pH 7.4 and diluted in culturemedium. To inhibit GAG sulphation, the cells were cultivated for18 h in a mixture (4:1) of low sulphate Ham’s F-12 medium andRPMI-1640, containing 20 mM sodium chlorate and 2% desaltedFCS. In wells with 20 mM chlorate, an appropriate amount ofmedium was replaced with water in order to maintain normalosmolarity (Pinzon-Ortiz et al., 2001). For digestion of cellsurface GAGs, the cells were treated with 20 mU ml-1 hepari-nase, 20 mU ml-1 heparitinase and 500 mU ml-1 chondroitinaseABC in the medium without FCS for 3 h (Shakibaei and Frevert,1996). To prevent binding of CSP to LRP-1, the cells were incu-bated with 500 nM of recombinant RAP for 10 min before additionof CSP. To inhibit adenylyl cyclase, the cells were incubated10 min with 100 mM SQ 22 536 before addition of sporozoites orCSP. To inhibit PKA, the cells were incubated for 30 min with10 mM H-89, 10 mM PKI14-22, or 200 mM Rp-cAMP (Sozzani et al.,1995; Orlic et al., 2002; Nogueira-Machado et al., 2003; Aronoffet al., 2005; Makranz et al., 2006). The selective activators8-CPT-2-Me-cAMP and 6-MB-cAMP were used to stimulate PKAand EPAC, respectively (Christensen et al., 2003; Kang et al.,2003). In some experiments, 3-isobutyl-1 methylxanthine (IBMX)or dibutyryl cAMP (db-cAMP) were added to the cells 30 minbefore measuring. IBMX and db-cAMP were used at a finalconcentration of 1 mM. IBMX was dissolved in dimethyl sulphox-ide (DMSO). The final concentration of DMSO in the culturemedium did not exceed 0.1%. All other additives were dissolvedin culture medium and were added as indicated in the figurelegends. To study the effect of sporozoites on signalling, 2 freshly
isolated sporozoites were added per Kupffer cell. As a control,Kupffer cells were incubated with extracts from uninfected sali-vary glands.
NBT reduction assay
Murine Kupffer cells were incubated for 45 min with P. yoeliisporozoites at a ratio of 1:1 in RPMI containing 10% FCS and0.1% NBT. After washing with PBS, the cells were fixed with 0.1%glutaraldehyde in PBS. Parasite Kupffer cell interactions wereanalysed microscopically for local NBT reduction as indicated bythe formation of insoluble purple formazan. In some experiments,Kupffer cells were activated with 1 mm PMA 5 min before sporo-zoite addition. Sporozoite inactivation was by four cycles of freez-ing in liquid nitrogen and thawing at 37°C.
Cyclic adenosyl mono-phosphate radioimmunoassay
For cAMP determination, liver cell cultures were incubated inculture medium without FCS at 37°C for 0–90 min. The cultureswere then placed on ice, and the reaction was stopped by aspi-ration of the incubation medium followed by the addition of 1 mlof ice cold 65% (v/v) ethanol. The cells were scraped into Eppen-dorf tubes and centrifuged at 10 000 g for 10 min at 4°C. Thesupernatants were dried in SC110 Speedvac Concentrator(Savant, Farmingdale, NY) at room temperature, and the driedextracts were dissolved in assay buffer for cAMP determinationwith cAMP [125I] Direct Biotrak Assay from Amersham Bio-sciences (Piscataway, NJ), according to the instructions of themanufacturer. Samples were analysed with a Beckman LS 6000scintillation counter. The cAMP level in each sample was normal-ized to the protein concentration and expressed as fmol mg-1 ofprotein. Ethanol-precipitated cell pellets were diluted with 0.15 MNaOH and the protein concentration was measured with theMicro BCA Protein Assay Kit (Pierce Chemical, Rockford, IL)according to manufacturer’s instructions.
Inositol-1,4,5-trisphosphate (IP3) assay
Intracellular IP3 levels were measured with a IP3 [3H] BiotrakAssay (Amersham Biosciences, Piscataway, NJ). Liver cells(3.0 ¥ 106 well-1) were incubated for 0, 30, 60, 120 s with 100 nMCSP in RPMI-1640 containing 10 mM LiCI. The plates were thenplaced on ice, and the reaction was stopped by aspiration of theincubation medium followed by the addition of 20% ice-cold per-chloric acid. After incubation on ice for 20 min, the cells werescraped into Eppendorf tubes. The precipitated protein wasremoved by centrifugation at 10 000 g for 15 min at 4°C. Thesupernatant was neutralized to pH 7.5 by titrating with ice-cold1.5 M KOH containing 60 mM HEPES buffer. The precipitatedKClO4 was removed by centrifugation at 5000 g for 15 min at4°C. Radioactivity in the supernatant was measured with aBeckman LS 6000 scintillation counter.
Chemiluminescence
The production of ROS by Kupffer cells was measured usingluminol- or lucigenin-enhanced chemiluminescence. Lucigeninreacts predominantly with superoxide, generated by the NADPH
2622 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

oxidase complex (Gyllenhammar, 1987), while luminol detectsmainly hydrogen peroxide plus an additional spectrum of ROS(hydrochlorous acid, nitric oxide, peroxyl radicals and peroxyni-trite anions) (Allen and Loose, 1976). Kupffer cells were culti-vated in white 96-well plates (Isoplate, PerkinElmer Life andAnalytical Sciences, Boston, MA) in RPMI-1640 without phenol.Before starting the experiments, the plates were washed threetimes with Hanks’ balanced salt solution (HBSS), pH 7.4 and thenincubated in HBSS containing 50 mM luminol or lucigenin. Thelight emission kinetics were analysed at 37°C using a GENios-Plus multifunctional microplate reader (Tecan Austria GmbH,Salzburg, Austria) using the following conditions: each well wasmeasured 60–90 times at 120 s intervals with a integration timeof 1000 ms per measurement. Data were acquired with Magellanstandard software and exported into XFluor4 software (Tecan).The results are expressed as relative light units (RLU) and rep-resent the mean of four to six wells. To stimulate Kupffer cells, thecells were treated with 200 nM PMA (Brehm et al., 1996) or0.25 mg ml-1 non-opsonized zymosan (Wang et al., 1993).Luminol and PMA were dissolved in DMSO. The final concentra-tion of DMSO in medium did not exceed 0.1%. All other additiveswere dissolved in HBSS and were added as indicated in thefigure legends. Zymosan A from S. cerevisiae was suspended(20 mg ml-1) and boiled in distilled water for 30 min followed bythree washes in HBSS.
Immunolocalization of LRP-1
Frozen sections of paraformaldehyde-fixed normal rat liver werecut with a Reichert-Jung 2800 Frigocut E cryostat. Hepatocytes,Kupffer cells and sinusoidal endothelia were purified from ratliver, cultivated overnight, and fixed with 4% paraformaldehyde inPBS. Frozen sections or fixed liver cell cultures were labelled withmAb 8B8 against the 85 kDa light chain of human LRP-1 followedby goat anti-mouse IgG conjugated to FITC (GAM-FITC) orprotein A-FITC (PA-FITC) and counterstained with 0.1% Evansblue in PBS.
Microscopy and image analysis
Immunofluorescence specimens were analysed using a ZeissLSM 510 confocal laser scanning microscope. Digital imageswere processed with ImagePro-Plus (Media Cybernetics, SilverSpring, MD) and Microsoft PowerPoint software.
Database analysis
To determine the EST expression level of Plasmodium parasitesgenes involved in ROS detoxification, we analysed EST data-bases at the ‘Gene Index Databases’ for P. yoelii from the Com-putational Biology and Functional Genomics Laboratory atthe Dana Faber Cancer Institute and Harvard School of PublicHealth (http://compbio.dfci.harvard.edu/tgi/cgi-bin/tgi/gireport.pl?gudb=p_yoelii). At the time of analysis, the libraries contained12471 ESTs from blood stage cDNA and 5523 ESTs from sporo-zoite and liver stage cDNA. Key word search was conductedfor ‘superoxide dismutase’, ‘thioredoxin reductase’, ‘glutathionereductase’, ‘thioredoxin’, ‘glutaredoxin’, ‘thioredoxin peroxidase’,‘glutathion peroxidase’ and ‘peroxiredoxin’.
Statistical analysis
All experimental data were presented as the mean � STD from 4to 8 wells from one of three representative experiments. Resultswere normalized per well or mg of cell protein. Statistical signifi-cance between groups was determined by Student’s paired t-test.
Acknowledgements
We thank Rita Altszuler for expert help with Plasmodium sporo-zoite purification and Dr Lakshmi Kumaratilake for performinginitial chemiluminescence experiments. We thank Dr DudleyStrickland, American Red Cross, for a generous gift of RAP. Weare indebted to Drs Mauricio Calvo and Jerome Vanderberg forcritically reading the manuscript. The work was supported by NIHGrants RO1 AI51656 and S10 RR019288 to U.F.
References
Ahuja, N., Kumar, P., and Bhatnagar, R. (2004) The adeny-late cyclase toxins. Crit Rev Microbiol 30: 187–196.
Allen, R.C., and Loose, L.D. (1976) Phagocytic activation of aluminol-dependent chemiluminescence in rabbit alveolarand peritoneal macrophages. Biochem Biophys ResCommun 69: 245–252.
Amino, R., Thiberge, S., Martin, B., Celli, S., Shorte, S.,Frischknecht, F., and Menard, R. (2006) Quantitativeimaging of Plasmodium transmission from mosquito tomammal. Nat Med 12: 220–224.
Ancsin, J.B., and Kisilevsky, R. (2004) A binding site forhighly sulfated heparan sulfate is identified in the amino-terminus of the circumsporozoite protein: significance formalarial sporozoite attachment to hepatocytes. J BiolChem 279: 21824–21832.
Anderson, R., Goolam Mahomed, A., Theron, A.J., Ramafi,G., and Feldman, C. (1998) Effect of rolipram and dibutyrylcyclic AMP on resequestration of cytosolic calcium inFMLP-activated human neutrophils. Br J Pharmacol 124:547–555.
Angeli, V., Faveeuw, C., Delerive, P., Fontaine, J., Barriera,Y., Franchimont, N., et al. (2001) Schistosoma mansoniinduces the synthesis of IL-6 in pulmonary microvascularendothelial cells: role of IL-6 in the control of lung eosino-philia during infection. Eur J Immunol 31: 2751–2761.
Aronoff, D.M., Canetti, C., Serezani, C.H., Luo, M., andPeters-Golden, M. (2005) Cutting edge: macrophage inhi-bition by cyclic AMP (cAMP): differential roles of proteinkinase A and exchange protein directly activated bycAMP-1. J Immunol 174: 595–599.
Baer, K., Roosevelt, M., Van Rooijen, N., Clarkson, A.B., Jr,and Frevert, U. (2007) Kupffer cells are obligatory for Plas-modium sporozoite infection of the liver. Cell Microbiol 9:397–412.
Baeuerle, P.A., and Huttner, W.B. (1986) Chlorate – a potentinhibitor of protein sulfation in intact cells. Biochem BiophysRes Comm 141: 870–877.
Becker, K., Tilley, L., Vennerstrom, J.L., Roberts, D.,Rogerson, S., and Ginsburg, H. (2004) Oxidative stressin malaria parasite-infected erythrocytes: host–parasiteinteractions. Int J Parasitol 34: 163–189.
Malaria sporozoites deactivate Kupffer cells 2623
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

Bengis-Garber, C., and Gruener, N. (1996) Protein kinase Adownregulates the phosphorylation of p47 phox in humanneutrophils: a possible pathway for inhibition of the respi-ratory burst. Cell Signal 8: 291–296.
Bernfield, M., Kokenyesi, R., Kato, M., Hinkes, M.T., Spring,J., Gallo, R.L., and Lose, E.J. (1992) Biology of thesyndecans: a family of transmembrane heparan sulfateproteoglycans. Ann Rev Cell Biol 8: 365–393.
Binder, R., Kress, A., Kan, G., Herrmann, K., and Kirschfink,M. (1999) Neutrophil priming by cytokines and vitamin Dbinding protein (Gc-globulin): impact on C5a-mediatedchemotaxis, degranulation and respiratory burst. MolImmunol 36: 885–892.
Bokoch, G.M., and Diebold, B.A. (2002) Current molecularmodels for NADPH oxidase regulation by Rac GTPase.Blood 100: 2692–2696.
Boucher, P., and Gotthardt, M. (2004) LRP and PDGF sig-naling: a pathway to atherosclerosis. Trends CardiovascMed 14: 55–60.
Bouwens, L., and Wisse, E. (1992) The origin of Kupffer cellsand their relationship to hepatocytes. In Hepatocyte andKupffer Cell Interactions. Billiar, T.R. and Curran, R.D.(eds). Boca Raton, FL: CRC Press, pp. 3–21.
Brandonisio, O., Panaro, M.A., Marzio, R., Marangi, A.,Faliero, S.M., and Jirillo, E. (1994) Impairment of thehuman phagocyte oxidative responses caused by Leish-mania lipophosphoglycan (LPG): in vitro studies. FEMSImmunol Med Microbiol 8: 57–62.
Brehm, M., Schiller, E., and Zeller, W.J. (1996) Quantificationof reactive oxygen species generated by alveolar macro-phages using lucigenin-enhanced chemiluminescence –methodical aspects. Toxicol Lett 87: 131–138.
Buchmuller-Rouiller, Y., and Mauel, J. (1987) Impairment ofthe oxidative metabolism of mouse peritoneal macro-phages by intracellular Leishmania spp. Infect Immun 55:587–593.
Carey, D.J. (1997) Syndecans: multifunctional cell-surfaceco-receptors. Biochem J 327: 1–16.
Cerami, C., Frevert, U., Sinnis, P., Takacs, B., Clavijo, P.,Santos, M.J., and Nussenzweig, V. (1992) The basolateraldomain of the hepatocyte plasma membrane bears recep-tors for the circumsporozoite protein of Plasmodium falci-parum sporozoites. Cell 70: 1021–1033.
Chaves, M.M., Silvestrini, A.A., Silva-Teixeira, D.N., andNogueira-Machado, J.A. (1996) Effect in vitro of gammainterferon and interleukin-10 on generation of oxidizingspecies by human granulocytes. Inflamm Res 45: 313–315.
Christensen, A.E., Selheim, F., de Rooij, J., Dremier, S.,Schwede, F., Dao, K.K., et al. (2003) cAMP analogmapping of Epac1 and cAMP kinase. Discriminatinganalogs demonstrate that Epac and cAMP kinase act syn-ergistically to promote PC-12 cell neurite extension. J BiolChem 278: 35394–35402.
Condliffe, A.M., Kitchen, E., and Chilvers, E.R. (1998)Neutrophil priming: pathophysiological consequences andunderlying mechanisms. Clin Sci (Lond) 94: 461–471.
Cronstein, B.N., Rosenstein, E.D., Kramer, S.B., Weissmann,G., and Hirschhorn, R. (1985) Adenosine; a physiologicmodulator of superoxide anion generation by humanneutrophils. Adenosine acts via an A2 receptor on humanneutrophils. J Immunol 135: 1366–1371.
Danforth, H.D., Aikawa, M., Cochrane, A.H., and Nussenz-weig, R.S. (1980) Sporozoites of mammalian malaria:attachment to, interiorization and fate within macrophages.J Protozool 27: 193–202.
Dewas, C., Dang, P.M., Gougerot-Pocidalo, M.A., andEl-Benna, J. (2003) TNF-alpha induces phosphorylation ofp47 (phox) in human neutrophils: partial phosphorylation ofp47phox is a common event of priming of human neutro-phils by TNF-alpha and granulocyte-macrophage colony-stimulating factor. J Immunol 171: 4392–4398.
Diebold, B.A., and Bokoch, G.M. (2005) Rho GTPases andthe control of the oxidative burst in polymorphonuclearleukocytes. Curr Top Microbiol Immunol 291: 91–111.
Dimopoulos, G., Christophides, G.K., Meister, S., Schultz, J.,White, K.P., Barillas-Mury, C., and Kafatos, F.C. (2002)Genome expression analysis of Anopheles gambiae:responses to injury, bacterial challenge, and malariainfection. Proc Natl Acad Sci USA 99: 8814–8819.
Ding, M., Kwok, L.Y., Schluter, D., Clayton, C., and Soldati,D. (2004) The antioxidant systems in Toxoplasma gondiiand the role of cytosolic catalase in defence against oxi-dative injury. Mol Microbiol 51: 47–61.
Dockrell, H.M., and Playfair, J.H. (1984) Killing of Plasmo-dium yoelii by enzyme-induced products of the oxidativeburst. Infect Immun 43: 451–456.
Dong, Y., and Vennerstrom, J.L. (2003) Mechanisms of in situactivation for peroxidic antimalarials. Redox Rep 8: 284–288.
Feng, W.G., Wang, Y.B., Zhang, J.S., Wang, X.Y., Li, C.L.,and Chang, Z.L. (2002) cAMP elevators inhibit LPS-induced IL-12 p40 expression by interfering with phospho-rylation of p38 MAPK in murine peritoneal macrophages.Cell Res 12: 331–337.
Frankenburg, S., Leibovici, V., Mansbach, N., Turco, S.J.,and Rosen, G. (1990) Effect of glycolipids of Leishmaniaparasites on human monocyte activity. Inhibition bylipophosphoglycan. J Immunol 145: 4284–4289.
Frevert, U., Sinnis, P., Cerami, C., Shreffler, W., Takacs, B.,and Nussenzweig, V. (1993) Malaria circumsporozoiteprotein binds to heparan sulfate proteoglycans associatedwith the surface membrane of hepatocytes. J Exp Med177: 1287–1298.
Frevert, U., Engelmann, S., Zougbédé, S., Stange, J., Ng, B.,Matuschewski, K., et al. (2005) Intravital observation ofPlasmodium berghei sporozoite infection of the liver. PLoSBiol 3: e192.
Frevert, U., Usynin, I., Baer, K., and Klotz, C. (2006a) Pen-etrating biological barriers. Liver: Plasmodium sporozoitepassage across the sinusoidal cell layer. In MolecularMechanisms of Parasite Invasion. Soldati, D., and Bur-leigh, B.A. (eds). Landes Bioscience, Austin, TX, pp. 1–16.
Frevert, U., Usynin, I., Baer, K., and Klotz, C. (2006b)Nomadic or sessile: can Kupffer cells function as portals formalaria sporozoites to the liver? Cell Microbiol 8: 1537–1546.
van der Geer, P. (2002) Phosphorylation of LRP1: regulationof transport and signal transduction. Trends CardiovascMed 12: 160–165.
Gillman, B.M., Batchelder, J., Flaherty, P., and Weidanz,W.P. (2004) Suppression of Plasmodium chabaudi para-sitemia is independent of the action of reactive oxygen
2624 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

intermediates and/or nitric oxide. Infect Immun 72: 6359–6366.
Gjertsen, B.T., Mellgren, G., Otten, A., Maronde, E.,Genieser, H.G., Jastorff, B., et al. (1995) Novel (Rp)-cAMPS analogs as tools for inhibition of cAMP-kinasein cell culture. Basal cAMP-kinase activity modulatesinterleukin-1 beta action. J Biol Chem 270: 20599–20607.
Goretzki, L., and Mueller, B.M. (1997) Receptor-mediatedendocytosis of urokinase-type plasminogen activator isregulated by cAMP-dependent protein kinase. J Cell Sci110 (Pt 12): 1395–1402.
Goretzki, L., and Mueller, B.M. (1998) Low-density-lipoprotein-receptor-related protein (LRP) interacts with aGTP-binding protein. Biochem J 336 (Pt 2): 381–386.
Gotthardt, M., Trommsdorff, M., Nevitt, M.F., Shelton, J.,Richardson, J.A., Stockinger, W., et al. (2000) Interactionsof the low density lipoprotein receptor gene family withcytosolic adaptor and scaffold proteins suggest diversebiological functions in cellular communication and signaltransduction. J Biol Chem 275: 25616–25624.
Greve, B., Lehman, L.G., Lell, B., Luckner, D., Schmidt-Ott,R., and Kremsner, P.G. (1999) High oxygen radical pro-duction is associated with fast parasite clearance in chil-dren with Plasmodium falciparum malaria. J Infect Dis 179:1584–1586.
Gyllenhammar, H. (1987) Lucigenin chemiluminescence inthe assessment of neutrophil superoxide production.J Immunol Methods 97: 209–213.
Herz, J. (1993) The LDL-receptor-related protein – portrait ofa multifunctional receptor. Curr Opin Lipidol 4: 107–113.
Herz, J., and Gerard, R.D. (1993) Adenovirus-mediatedtransfer of low density lipoprotein receptor gene acutelyaccelerates cholesterol clearance in normal mice. ProcNatl Acad Sci USA 90: 2812–2816.
Herz, J., and Strickland, D.K. (2001) LRP: a multifunctionalscavenger and signaling receptor. J Clin Invest 108: 779–784.
Hoffman, M., Feldman, S.R., and Pizzo, S.V. (1983) Alpha2-macroglobulin ‘fast’ forms inhibit superoxide productionby activated macrophages. Biochim Biophys Acta 760:421–423.
Hollingdale, M.R. (1985) Malaria and the liver. Hepatology 5:327–335.
Hordijk, P.L. (2006) Regulation of NADPH oxidases: the roleof Rac proteins. Circ Res 98: 453–462.
Ishino, T., Yano, K., Chinzei, Y., and Yuda, M. (2004) Cell-passage activity is required for the malarial parasite tocross the liver sinusoidal cell layer. Plos Biol 2: E4.
Jackson, S.H., Gallin, J.I., and Holland, S.M. (1995) Thep47phox mouse knock-out model of chronic granulomatousdisease. J Exp Med 182: 751–758.
Joensen, L., Borda, E., Kohout, T., Perry, S., Garcia, G., andSterin-Borda, L. (2003) Trypanosoma cruzi antigen thatinteracts with the beta1-adrenergic receptor and modifiesmyocardial contractile activity. Mol Biochem Parasitol 127:169–177.
Kang, G., Joseph, J.W., Chepurny, O.G., Monaco, M.,Wheeler, M.B., Bos, J.L., et al. (2003) Epac-selectivecAMP analog 8-pCPT-2′-O-Me-cAMP as a stimulus forCa2+-induced Ca2+ release and exocytosis in pancreaticbeta-cells. J Biol Chem 278: 8279–8285.
Kappe, S.H.I., Gardner, M.J., Brown, S.M., Ross, J., Matus-chewski, K., Ribeiro, J.M., et al. (2001) Exploring the tran-scriptome of the malaria sporozoite stage. Proc Natl AcadSci USA 98: 9895–9900.
Kawasaki, H., Springett, G.M., Mochizuki, N., Toki, S.,Nakaya, M., Matsuda, M., et al. (1998) A family of cAMP-binding proteins that directly activate Rap1. Science 282:2275–2279.
Knook, D.L., and Sleyster, E.C. (1976) Separation of Kupfferand endothelial cells of the rat liver by centrifugalelutriation. Exp Cell Res 99: 444–449.
Kuiper, J., Brouwer, A., Knook, D.L., and Berkel, T.J.C.V.(1994) Kupffer and sinusoidal endothelial cells. In TheLiver: Biology and Pathobiology. Arias, I.M., Boyer, J.L.,Fausto, N., Jakoby, W.B., Schachter, D.A., and Shafritz,D.A. (eds). New York: Raven Press, pp. 791–818.
Kumar, S., Christophides, G.K., Cantera, R., Charles, B., Han,Y.S., Meister, S., et al. (2003) The role of reactive oxygenspecies on Plasmodium melanotic encapsulation in Anoph-eles gambiae. Proc Natl Acad Sci USA 100: 14139–14144.
Lambeth, J.D. (2004) NOX enzymes and the biology of reac-tive oxygen. Nat Rev Immunol 4: 181–189.
Li, Y., van Kerkhof, P., Marzolo, M.P., Strous, G.J., and Bu,G. (2001) Identification of a major cyclic AMP-dependentprotein kinase A phosphorylation site within the cytoplas-mic tail of the low-density lipoprotein receptor-relatedprotein: implication for receptor-mediated endocytosis. MolCell Biol 21: 1185–1195.
Lowrie, D.B., Aber, V.R., and Jackett, P.S. (1979)Phagosome-lysosome fusion and cyclic adenosine 3′:5′-monophosphate in macrophages infected with Myco-bacterium microti, Mycobacterium bovis BCG or Mycobac-terium lepraemurium. J Gen Microbiol 110: 431–441.
Lutz, C., Nimpf, J., Jenny, M., Boecklinger, K., Enzinger, C.,Utermann, G., et al. (2002) Evidence of functional modu-lation of the MEKK/JNK/cJun signaling cascade by the lowdensity lipoprotein receptor-related protein (LRP). J BiolChem 277: 43143–43151.
Mack, S.R., Vanderberg, J.P., and Nawrot, R. (1978) Columnseparation of Plasmodium berghei sporozoites. J Parasitol64: 166–168.
McNeely, T.B., Rosen, G., Londner, M.V., and Turco, S.J.(1989) Inhibitory effects on protein kinase C activity bylipophosphoglycan fragments and glycosylphosphatidyli-nositol antigens of the protozoan parasite Leishmania.Biochem J 259: 601–604.
Makranz, C., Cohen, G., Reichert, F., Kodama, T., and Rot-shenker, S. (2006) cAMP cascade (PKA, Epac, adenylylcyclase, Gi, and phosphodiesterases) regulates myelinphagocytosis mediated by complement receptor-3 andscavenger receptor-AI/II in microglia and macrophages.Glia 53: 441–448.
Marshall, P., Rohlmann, A., Nussenzweig, V., Herz, J., andSinnis, P. (2000) Plasmodium sporozoites invade cells withtargeted deletions in the LDL receptor related protein. MolBiochem Parasitol 106: 293–298.
Martins Chaves, M., Prates Rodrigues, A.L., Pereira dosReis, A., Gerzstein, N.C., and Nogueira-Machado, J.A.(2002) Correlation between NADPH oxidase and proteinkinase C in the ROS production by human granulocytesrelated to age. Gerontology 48: 354–359.
Malaria sporozoites deactivate Kupffer cells 2625
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

Mauël, J. (1996) Intracellular survival of protozoan parasiteswith special reference to Leishmania spp., Toxoplasmagondii and Trypanosoma cruzi. Adv Parasitol 38: 1–51.
Meis, J.F.G.M., Verhave, J.P., Jap, P.H.K., and Meuwissen,J.H.E.T. (1983) An ultrastructural study on the role ofKupffer cells in the process of infection by Plasmodiumberghei sporozoites in rats. Parasitology 86: 231–242.
Meis, J.F.G.M., Verhave, J.P., Brouwer, A., and Meuwissen,J.H.E.T. (1985) Electron microscopic studies on the inter-action of rat Kupffer cells and Plasmodium bergheisporozoites. Z Parasitenkd 71: 473–483.
Miller, R.A., and Britigan, B.E. (1997) Role of oxidants inmicrobial pathophysiology. Clin Microbiol Rev 10: 1–18.
Misra, U.K., and Pizzo, S.V. (1998a) Ligation of the a2Msignalling receptor elevates the levels of p21Ras-GTP inmacrophages. Cell Signal 10: 441–445.
Misra, U.K., and Pizzo, S.V. (1998b) Ligation of the a2Msignaling receptor with receptor-recognized forms ofa2-macroglobulin initiates protein and DNA synthesis inmacrophages: the effect of intracellular calcium. BiochimBiophys Acta 1401: 121–128.
Misra, U.K., and Pizzo, S.V. (2002) Regulation of cytosolicphospholipase A2 activity in macrophages stimulated withreceptor-recognized forms of alpha 2-macroglobulin: rolein mitogenesis and cell proliferation. J Biol Chem 277:4069–4078.
Misra, U.K., Akabani, G., and Pizzo, S.V. (2002a) The role ofcAMP-dependent signaling in receptor-recognized forms ofalpha 2-macroglobulin-induced cellular proliferation. J BiolChem 277: 36509–36520.
Misra, U.K., Gonzalez-Gronow, M., Gawdi, G., Hart, J.P.,Johnson, C.E., and Pizzo, S.V. (2002b) The role of Grp 78in alpha 2-macroglobulin-induced signal transduction. Evi-dence from RNA interference that the low density lipopro-tein receptor-related protein is associated with, but notnecessary for, GRP 78-mediated signal transduction. J BiolChem 277: 42082–42087.
Misra, U.K., Gonzalez-Gronow, M., Gawdi, G., and Pizzo,S.V. (2005) The role of MTJ-1 in cell surface translocationof GRP78, a receptor for alpha 2-macroglobulin-dependentsignaling. J Immunol 174: 2092–2097.
Mitsuyama, T., Takeshige, K., and Minakami, S. (1993) CyclicAMP inhibits the respiratory burst of electropermeabilizedhuman neutrophils at a downstream site of protein kinaseC. Biochim Biophys Acta 1177: 167–173.
Moestrup, S.K., Gliemann, J., and Pallesen, G. (1992) Distri-bution of the a2-macroglobulin receptor/low density lipopro-tein receptor-related protein in human tissues. Cell TissueRes 269: 375–382.
Mota, M.M., Pradel, G., Vanderberg, J.P., Hafalla, J.C.R.,Frevert, U., Nussenzweig, R.S., et al. (2001) Migration ofPlasmodium sporozoites through cells before infection.Science 291: 141–144.
Müller, S. (2004) Redox and antioxidant systems of themalaria parasite Plasmodium falciparum. Mol Microbiol 53:1291–1305.
Neufeld, D.S. (1997) Isolation of rat liver hepatocytes. InMethods in Molecular Biology, Vol 75: Basic Cell CultureProtocols. Pollard, J.W., and Walker, J.M. (eds). HumanaPress Inc., Totawa, NJ, pp. 145–151.
Newman, S.L., Mikus, L.K., and Tucci, M.A. (1991) Differen-
tial requirements for cellular cytoskeleton in humanmacrophage complement receptor- and Fc receptor-mediated phagocytosis. J Immunol 146: 967–974.
Nickel, C., Rahlfs, S., Deponte, M., Koncarevic, S., andBecker, K. (2006) Thioredoxin networks in the malarialparasite Plasmodium falciparum. Antioxid Redox Signal 8:1227–1239.
Nielson, C.P. (1987) Beta-adrenergic modulation of the poly-morphonuclear leukocyte respiratory burst is dependentupon the mechanism of cell activation. J Immunol 139:2392–2397.
Nielson, C.P., Bayer, C., Hodson, S., and Hadjokas, N.(1992) Regulation of the respiratory burst by cyclic-3′,5′–AMP, an association with inhibition of arachidonic acidrelease. J Immunol 149: 4036–4040.
Nogueira-Machado, J.A., Lima e Silva, F.C., Medina, L.O.,Costa, D.C., and Chaves, M.M. (2003) Modulation of thereactive oxygen species (ROS) generation mediated bycyclic AMP-elevating agents or Interleukin 10 in granulo-cytes from type 2 diabetic patients (NIDDM): a PKA-independent phenomenon. Diabetes Metab 29: 533–537.
O’Dowd, Y.M., El-Benna, J., Perianin, A., and Newsholme, P.(2004) Inhibition of formyl-methionyl-leucyl-phenylalanine-stimulated respiratory burst in human neutrophils byadrenaline: inhibition of Phospholipase A2 activity but notp47phox phosphorylation and translocation. BiochemPharmacol 67: 183–190.
Ockenhouse, C.F., Schulman, S., and Shear, H.L. (1984)Oxidative killing of malaria parasites by mononuclearphagocytes. Prog Clin Biol Res 155: 93–108.
Oliveira, P.L., and Oliveira, M.F. (2002) Vampires, Pasteurand reactive oxygen species. Is the switch from aerobic toanaerobic metabolism a preventive antioxidant defence inblood-feeding parasites? FEBS Lett 525: 3–6.
Orlic, T., Loomis, W.H., Shreve, A., Namiki, S., and Junger,W.G. (2002) Hypertonicity increases cAMP in PMNand blocks oxidative burst by PKA-dependent and-independent mechanisms. Am J Physiol Cell Physiol 282:C1261–C1269.
Pinzon-Ortiz, C., Friedman, J., Esko, J., and Sinnis, P. (2001)The binding of the circumsporozoite protein to cell surfaceheparan sulfate proteoglycans is required for Plasmodiumsporozoite attachment to target cells. J Biol Chem 276:26784–26791.
Pollock, J.D., Williams, D.A., Gifford, M.A., Li, L.L., Du, X.,Fisherman, J., et al. (1995) Mouse model of X-linkedchronic granulomatous disease, an inherited defect inphagocyte superoxide production. Nat Genet 9: 202–209.
Pradel, G., and Frevert, U. (2001) Plasmodium sporozoitesactively enter and pass through Kupffer cells prior to hepa-tocyte invasion. Hepatology 33: 1154–1165.
Pradel, G., Garapaty, S., and Frevert, U. (2002) Proteogly-cans mediate malaria sporozoite targeting to the liver. MolMicrobiol 45: 637–651.
Pradel, G., Garapaty, S., and Frevert, U. (2004) Kupffer andstellate cell proteoglycans mediate malaria sporozoite tar-geting to the liver. Comp Hepatol 3 (Suppl. 1): S47.
Quinn, M.T., and Gauss, K.A. (2004) Structure and regulationof the neutrophil respiratory burst oxidase: comparison withnonphagocyte oxidases. J Leukoc Biol 76: 760–781.
2626 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

Rapraeger, A.C. (2000) Syndecan-regulated receptorsignaling. J Cell Biol 149: 995–997.
Rawal, S., Majumdar, S., Dhawan, V., and Vohra, H. (2004)Entamoeba histolytica Gal/GalNAc lectin depletes antioxi-dant defences of target epithelial cells. Parasitology 128:617–624.
Reeves, E.P., Dekker, L.V., Forbes, L.V., Wientjes, F.B.,Grogan, A., Pappin, D.J., and Segal, A.W. (1999) Directinteraction between p47phox and protein kinase C: evi-dence for targeting of protein kinase C by p47phox inneutrophils. Biochem J 344 (Pt 3): 859–866.
Rehmann, H., Prakash, B., Wolf, E., Rueppel, A., De Rooij,J., Bos, J.L., and Wittinghofer, A. (2003) Structure andregulation of the cAMP-binding domains of Epac2. NatStruct Biol 10: 26–32.
Rico, G., Diaz-Guerra, O., and Kretschmer, R.R. (1995)Cyclic nucleotide changes induced in human leukocytes bya product of axenically grown Entamoeba histolytica thatinhibits human monocyte locomotion. Parasitol Res 81:158–162.
Robson, K.J.H., Frevert, U., Reckmann, I., Cowan, G., Beier,J., Scragg, I.G., et al. (1995) Thrombospondin relatedadhesive protein (TRAP) of Plasmodium falciparum:expression during sporozoite ontogeny and binding tohuman hepatocytes. EMBO J 14: 3883–3894.
Rodriguez, A., Martinez, I., Chung, A., Berlot, C.H., andAndrew, N.W. (1999) cAMP Regulates Ca2+-dependentExocytosis of Lysosomes and Lysosome-mediated cellInvasion by Trypanosomes. J Biol Chem 274: 16754–16759.
Rohlmann, A., Gotthardt, M., Willnow, T.E., Hammer, R.E.,and Herz, J. (1996) Sustained somatic gene inactivation byviral transfer of Cre recombinase. Nat Biotechnol 14:1562–1565.
Rohlmann, A., Gotthardt, M., Hammer, R.E., and Herz, J.(1998) Inducible inactivation of hepatic LRP gene by cre-mediated recombination confirms role of LRP in clearanceof chylomicron remnants. J Clin Invest 101: 689–695.
de Rooij, J., Zwartkruis, F.J., Verheijen, M.H., Cool, R.H.,Nijman, S.M., Wittinghofer, A., and Bos, J.L. (1998) Epac isa Rap1 guanine-nucleotide-exchange factor directly acti-vated by cyclic AMP. Nature 396: 474–477.
Rossi, A.G., McCutcheon, J.C., Roy, N., Chilvers, E.R.,Haslett, C., and Dransfield, I. (1998) Regulation ofmacrophage phagocytosis of apoptotic cells by cAMP.J Immunol 160: 3562–3568.
Rozenfeld, C., Martinez, R., Figueiredo, R.T., Bozza, M.T.,Lima, F.R., Pires, A.L., et al. (2003) Soluble factorsreleased by Toxoplasma gondii-infected astrocytes down-modulate nitric oxide production by gamma interferon-activated microglia and prevent neuronal degeneration.Infect Immun 71: 2047–2057.
Sahney, N.N., Summersgill, J.T., Ramirez, J.A., and Miller,R.D. (2001) Inhibition of oxidative burst and chemotaxis inhuman phagocytes by Legionella pneumophila zincmetalloprotease. J Med Microbiol 50: 517–525.
Sanni, L.A., Fu, S., Dean, R.T., Bloomfield, G., Stocker, R.,Chaudhri, G., et al. (1999) Are reactive oxygen speciesinvolved in the pathogenesis of murine cerebral malaria?J Infect Dis 179: 217–222.
Schneider, W.J., and Nimpf, J. (2003) LDL receptor relatives
at the crossroad of endocytosis and signaling. Cell Mol LifeSci 60: 892–903.
Sedgwick, J.B., Berube, M.L., and Zurier, R.B. (1985)Stimulus-dependent inhibition of superoxide generation byprostaglandins. Clin Immunol Immunopathol 34: 205–215.
Seguin, M.C., Ballou, W.R., and Nacy, C.A. (1989) Interac-tions of Plasmodium berghei sporozoites and murineKupffer cells in vitro. J Immunol 143: 1716–1722.
Seino, S., and Shibasaki, T. (2005) PKA-dependentand PKA-independent pathways for cAMP-regulatedexocytosis. Physiol Rev 85: 1303–1342.
Shakibaei, M., and Frevert, U. (1996) Dual interaction of themalaria circumsporozoite protein with the low density lipo-protein receptor-related protein (LRP) and cell surfaceheparan sulfate. J Exp Med 184: 1699–1711.
Shant, J., Ghosh, S., Bhattacharyya, S., Ganguly, N.K., andMajumdar, S. (2004) The alteration in signal transductionparameters induced by the excretory-secretory productfrom Giardia lamblia. Parasitology 129: 421–430.
Shayakhmetov, D.M., Li, Z.Y., Ni, S., and Lieber, A. (2004)Analysis of adenovirus sequestration in the liver, transduc-tion of hepatic cells, and innate toxicity after injection offiber-modified vectors. J Virol 78: 5368–5381.
Shin, S.C.J., Vanderberg, J.P., and Terzakis, J.A. (1982)Direct infection of hepatocytes by sporozoites of Plasmo-dium berghei. J Protozool 29: 448–454.
Sionov, R.V., and Gallily, R. (1990) Engulfment and intracel-lular killing of F9 teratocarcinoma cells by non-activatedmurine macrophages. Int Immunol 2: 291–301.
Smedsrod, B., and Pertoft, H. (1985) Preparation of purehepatocytes and reticuloendothelial cells in high yield froma single rat liver by means of Percoll centrifugation andselective adherence. J Leukocyte Biol 38: 213–230.
Smith, J.E., and Alexander, J. (1986) Evasion of macrophagemicrobicidal mechanisms by mature sporozoites of Plas-modium yoelii yoelii. Parasitology 93: 33–38.
Sozzani, P., Cambon, C., Vita, N., Seguelas, M.H., Caput, D.,Ferrara, P., and Pipy, B. (1995) Interleukin-13 inhibitsprotein kinase C-triggered respiratory burst in humanmonocytes. Role of calcium and cyclic AMP. J Biol Chem270: 5084–5088.
Steenkamp, D.J. (2002) Trypanosomal antioxidants andemerging aspects of redox regulation in the trypano-somatids. Antioxid Redox Signal 4: 105–121.
Steers, N., Schwenk, R., Bacon, D.J., Berenzon, D., Will-iams, J., and Krzych, U. (2005) The immune status ofKupffer cells profoundly influences their responses to infec-tious Plasmodium berghei sporozoites. Eur J Immunol 35:2335–2346.
Stewart, M.J., and Vanderberg, J.P. (1991) Malaria sporozoi-tes release circumsporozoite protein from their apical endand translocate it along their surface. J Protozool 38: 411–421.
Strickland, D.K., and Ranganathan, S. (2003) Diverse role ofLDL receptor-related protein in the clearance of proteasesand in signaling. J Thromb Haemost 1: 1663–1670.
Strickland, D.K., Ashcom, J.D., Williams, S., Burgess, W.H.,Migliorini, M., and Argraves, W.S. (1990) Sequence iden-tity between the a2-macroglobulin receptor and low densitylipoprotein receptor-related protein suggests that this
Malaria sporozoites deactivate Kupffer cells 2627
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628

molecule is a multifunctional receptor. J Biol Chem 265:17401–17404.
Strickland, D.K., Kounnas, M.Z., and Argraves, W.S. (1995)LDL receptor-related protein: a multiligand receptor for lipo-protein and proteinase catabolism. FASEB J 9: 890–897.
Stüber, D., Matile, H., and Garotta, G. (1990) System forhigh-level production in Escherichia coli and rapid purifica-tion of recombinant proteins: application to epitopemapping, preparation of antibodies, and structure – func-tion analysis. Immunol Methods IV: 121–152.
Sturm, A., Amino, R., van de Sand, C., Regen, T., Retzlaff,S., Rennenberg, A., et al. (2006) Manipulation of hosthepatocytes by the malaria parasite for delivery into liversinusoids. Science 313: 1287–1290.
Sunahara, R.K., and Taussig, R. (2002) Isoforms of mamma-lian adenylyl cyclase: multiplicities of signaling. Mol Interv2: 168–184.
Takacs, B.J., and Girard, M.-F. (1991) Preparation of clinicalgrade proteins produced by recombinant DNA tech-nologies. J Immunol Methods 143: 231–240.
Tang, Y., Dong, Y., and Vennerstrom, J.L. (2004) Syntheticperoxides as antimalarials. Med Res Rev 24: 425–448.
Tarun, A.S., Baer, K., Dumpit, R.F., Gray, S., Lejarcegui, N.,Frevert, U., and Kappe, S. (2006) Quantitative isolationand in vivo imaging of malaria parasite liver stages. Int JParasitol 36: 1283–1293.
Tyagi, S.R., Olson, S.C., Burnham, D.N., and Lambeth, J.D.(1991) Cyclic AMP-elevating agents block chemoattractantactivation of diradylglycerol generation by inhibiting phos-pholipase D activation. J Biol Chem 266: 3498–3504.
Vanderberg, J.P., and Frevert, U. (2004) Intravital microscopydemonstrating antibody-mediated immobilization ofPlasmodium berghei sporozoites injected into skin bymosquitoes. Int J Parasitol 34: 991–996.
Vanderberg, J.P., Chew, S., and Stewart, M.J. (1990) Plas-modium sporozoite interactions with macrophages in vitro:a videomicroscopic analysis. J Protozool 37: 528–536.
Victorov, A.V., and Hoek, J.B. (1995) Secretion of prostag-landins elicited by lipopolysaccharide and ethanol in cul-tured rat Kupffer cells. Biochem Biophys Res Commun215: 691–697.
Wang, J.F., Komarov, P., and de Groot, H. (1993) Luminolchemiluminescence in rat macrophages and granulocytes:the role of NO, O2
-/H2O2, and HOCl. Arch Biochem Biophys304: 189–196.
Wang, J.P., Raung, S.L., Huang, L.J., and Kuo, S.C. (1998)Involvement of cyclic AMP generation in the inhibition ofrespiratory burst by 2-phenyl-4-quinolone (YT-1) in ratneutrophils. Biochem Pharmacol 56: 1505–1514.
Wisse, E., Zanger, R.B., Charels, K., Smissen, P., andMcCuskey, R.S. (1985) The liver sieve: considerationsconcerning the structure and function of endothelial fenes-trae, the sinusoidal wall and the space of Disse. Hepatol-ogy 5: 683–692.
Woods, A., and Couchman, J.R. (1998) Syndecans: synergis-tic activators of cell adhesion. Trends Cell Biol 8: 189–193.
Wozencraft, A.O., Dockrell, H.M., Taverne, J., Targett, G.A.,and Playfair, J.H. (1984) Killing of human malaria parasitesby macrophage secretory products. Infect Immun 43: 664–669.
Ying, P., Shakibaei, M., Patankar, M.S., Clavijo, P., Beavis,
R.C., Clark, G.F., and Frevert, U. (1997) The malariacircumsporozoite protein: interaction of the conservedregions I and II-plus with heparin-like oligosaccharides inheparan sulfate. Exp Parasitol 85: 168–182.
Zhu, Y., and Hui, D.Y. (2003) Apolipoprotein E binding to lowdensity lipoprotein receptor-related protein-1 inhibits cellmigration via activation of cAMP-dependent protein kinaseA. J Biol Chem 278: 36257–36263.
Zimmermann, P., and David, G. (1999) The syndecans,tuners of transmembrane signaling. FASEB J 13 (Suppl.):S91–S100.
Supplementary material
The following supplementary material is available for this articleonline:Fig. S1. Induction of the respiratory burst. The formation of super-oxide is a complex process that requires the assemblyof the heterohexameric NADPH oxidase (Bengis-Garber andGruener, 1996; Bokoch and Diebold, 2002; Lambeth, 2004; Quinnand Gauss, 2004; Diebold and Bokoch, 2005; Hordijk, 2006). Thetwo membrane-bound subunits gp91phox and p22phox form theflavocytochrome b558 complex. In the resting state, the four regu-latory subunits of the enzyme, p47phox, p67phox, p40phox, and a smallmonomeric G protein (Rac1 or Rac2) are located as an auto-inhibited complex in the cytosol. Upon cell stimulation withmicrobes or chemical agents, initiation of ROS formation caninvolve activation of phospholipase C (PLC) leading to generationof DAG and IP3 from PIP2. DAG and Ca++, released by IP3 fromintracellular stores, activate various isoforms of PKC leading tomultiple phosphorylation of p47phox (Reeves et al., 1999). Alterna-tively, ROS formation can be triggered by phosphoinositide3-kinase (PI-3K) activation causing phosphorylation of p47phox bykinases such as ERK,AKT, or various isoforms of PKC. p47phox canalso be phosphorylated by direct activation of PKC with phorbolesters such as PMA. Phosphorylation enables p47phox to targetp22phox, resulting in translocation of the cytosolic triad p47phox/p67phox/p40phox to the membrane. Another crucial event in NADPHoxidase assembly is activation of the small GTPase Rac, whichanchors to the membrane and associates with p67phox, thus posi-tioning it correctly for interaction with the cytochrome. Conforma-tional changes in flavocytochrome b558 then activate its gp91phox
subunit to allow flow of electrons from NADPH via FAD and haemredox centres to oxygen thus producing superoxide anions.Fig. S2. CSP increases the intracellular IP3 level in Kupffercells, but not in hepatocytes. Liver cells (3.0 ¥ 106 well-1) werecultivated for 18 h at 37°C in RPMI-1640 with 10% FCS and foranother 6 h in the same medium without FCS. The cells werethen incubated with 50 nM CSP in RPMI-1640 containing 10 mMlithium chloride for the times indicated. The reactions werestopped by addition of 6.25% ice-cold perchloric acid, and IP3
was extracted as described in Experimental procedures.*P < 0.001 compared with basal level.
This material is available as part of the online article from:http://www.blackwell-synergy.com/doi/abs/10.1111/j.1462-5822.2007.00982.x
Please note: Blackwell Publishing is not responsible for thecontent or functionality of any supplementary materials suppliedby the authors. Any queries (other than missing material) shouldbe directed to the corresponding author for the article.
2628 I. Usynin, C. Klotz and U. Frevert
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 2610–2628




















