
High density DNA methylation array with single CpG site resolution
Upload
independentCategory
view
2download
0
Oligodeoxynucleotides lacking CpG dinucleotides mediate Toll-like receptor 9
dependent T helper type 2 biased immune stimulation
JORG VOLLMER,* RISINI D. WEERATNA,� MARION JURK,* ULRIKE SAMULOWITZ,*
MICHAEL J. MCCLUSKIE,� PAUL PAYETTE,� HEATHER L. DAVIS,� CHRISTIAN SCHETTER* &
ARTHUR M. KRIEG� Coley Pharmaceutical, *Langenfeld, Germany, �Ottawa, Canada and �Wellesley, MA, USA
SUMMARY
Oligodeoxynucleotides (ODN) with unmethylated CpG dinucleotides mimic the immune stimu-
latory activity of bacterial DNA in vertebrates and are recognized by Toll-like receptor 9 (TLR9).
It is also possible to detect immune activation with certain phosphorothioate sequences that lack
CpG motifs. These ODN are less potent than CpG ODN and the mechanism by which they
stimulate mammalian leucocytes is not understood. We here provide several lines of evidence
demonstrating that the effects induced by non-CpGODN are mediated by TLR9. First, non-CpG
ODN could not stimulate cytokine secretion from the splenocytes of TLR9-deficient (TLR9–/––)
mice. Second, immunization of TLR9+/++ but not TLR9–/–– mice with non-CpG ODN enhanced
antigen-specific antibody responses, although these were T helper type 2 (Th2)-biased. Third,
reactivity to non-CpG ODN could be reconstituted by transfection of human TLR9 into non-
responsive cells. In addition, we define a new efficient immune stimulatory motif aside from the
CpG dinucleotide that consists of a 5¢-TC dinucleotide in a thymidine-rich background. Non-CpG
ODNcontaining this motif induced activation of humanB cells, but lacked stimulation of Th1-like
cytokines and chemokines. Our study indicates that TLR9 can mediate either efficient Th1- or
Th2-dominated effects depending on whether it is stimulated by CpG or certain non-CpG ODN.
Keywords B cells, dendritic cells, CpG DNA, Toll-like receptors
INTRODUCTION
Small synthetic oligodeoxynucleotides (ODN) containingunmethylated deoxycytidine-deoxyguanosine (CpG) dinu-
cleotides are able to mimic the immune stimulatoryactivity of bacterial DNA.1 Certain sequence motifs withcentral CpG dinucleotides are responsible for this stimu-lation. We have recently shown that the optimal CpG
motif for activating human cells is 5¢-GTCGTT-3¢,whereas the optimal motif for activating mouse cells is5¢-GACGTT-3¢.2,3 The strength of the immune response is
determined by the unmethylated CpG motif itself, as wellas the number and position of CpG dinucleotides in theODN and the spacing between such motifs.2 Modifi-
cations at the 5-position of cytosine are usually not verywell tolerated.1 Phosphorothioate modification of CpGODN enhances their in vitro activity usually by about10–100-fold.2–6
Studies using mice deficient in Toll-like receptor 9(TLR9) demonstrated that this TLR subtype is essential forthe effects mediated by bacterial or viral DNA and CpG
ODN.7 The responsiveness to CpG ODN stimulationin vitro could be reconstituted by transfection ofunresponsive cells with human TLR9.8,9 In addition, recent
publications strongly suggest that CpG DNA can directlybind to TLR9.10,11 TLR9 is a member of a family of at least12 different receptors recognizing certain molecular struc-
tures that are present in pathogens.12–14 Recently, TLRswere also shown to be responsible for the recognition ofdouble-stranded RNA (TLR3), small antiviral compounds,guanosine nucleoside analogues, or viral single-stranded
RNA (TLR7 and TLR8).15–19
Received 28 April 2004; revised 22 June 2004; accepted 8 July
2004
Abbreviations: CpG, unmethylated deoxycytidine-deoxyguan-
osine dinucleotide; HBsAg, hepatitis B surface antigen; ODN,
oligodeoxynucleotide; pDC, plasmacytoid dendritic cell; TLR,
Toll-like receptor; TT, tetanus toxoid antigen.
Correspondence: J. Vollmer, Coley Pharmaceutical GmbH,
Elisabeth-Selbert-Strasse 9, 40764 Langenfeld, Germany. E-mail:
Immunology 2004 113 212–223 doi:10.1111/j.1365-2567.2004.01962.x
212 � 2004 Blackwell Publishing Ltd

So far only a few types of human immune cells havebeen demonstrated to express TLR9. B cells and plasma-cytoid dendritic cells (pDC) express intracellular TLR9 andcan be directly stimulated by CpG ODN.10,20 The pDC are
the main producers of interferon-a (IFN-a), and these cellsare unique in their ability to respond to CpG ODN byproducing type I interferons.20–22 In contrast, mouse cells of
the myeloid lineage, such as mDC, monocytes, or macro-phages, all express TLR9.23 Other immune cells, such ashuman ab- or cd-T cells, or natural killer (NK) cells, appear
to be negative for TLR9 and seem to be only indirectlystimulated by CpG ODN.12
Aside from the CpG-specific effects, there are also CpG-
independent immune stimulatory effects of phosphoro-thioate ODN.24 Many of these non-sequence-specific effectsmay be the result of the increased affinity of phosphoro-thioate ODN for binding a wide variety of proteins.25
Phosphorothioate thymidine homopolymers, in contrast toother homopolymers, can induce the stimulation of humanB cells or murine splenocytes.26,27 We previously demon-
strated that the number of thymidine nucleobases in non-CpG phosphorothioate ODN, as well as the length of suchODN, directly relate to the magnitude of their stimulatory
effects, especially on human B cells.28 Nevertheless, themechanism of immune activation by non-CpG ODN andthe existence of specific stimulatory motifs besides the CpGmotifs still remain to be elucidated. TLR9 was reported to
be responsive to CpG but not to non-CpG ODN stimula-tion.8,29,30 Therefore, it was possible that non-CpG ODNacted via another pathway, which might also explain the
relatively small effects observed with non-CpG ODN whencompared to CpG ODN.
Here, we extended our previous studies and investigated
the dependency of non-CpG ODN-mediated immune sti-mulation on the expression of TLR9 in vivo as well asin vitro. Surprisingly, TLR9 is required for mediating im-
mune modulatory effects, not only by CpG ODN, but alsoby non-CpG ODN. In addition, efficient TLR9-mediatedactivation by such ODN depended strongly on a stablephosphorothioate backbone. Non-CpG ODN in vitro
induced efficient B-cell stimulation but lacked the stimula-tion of T helper type 1 (Th1) -like chemokines or cytokinesfrom human peripheral blood mononuclear cells (PBMC)
or pDC. In addition, we describe a non-CpG motif, a TCdinucleotide placed at the 5¢ end of thymidine-containingODN, that tremendously enhances the stimulatory capacity
of non-CpG ODN without mediating Th1-like effects. Ourdata suggest that the TLR9 receptor can induce differentprofiles of immune activation in response to different lig-ands, and provide a mechanistic explanation for previous
observations that a Th2-biased immune stimulatory effect isassociated with certain non-CpG ODN.31–33
MATERIALS AND METHODS
OligodeoxynucleotidesAll ODN were purchased from Biospring (Frankfurt,
Germany), controlled for identity (MALDI-TOF MS) andpurity (capillary gel electrophoresis) by Coley Pharmaceu-
tical Group (Langenfeld, Germany) and had undetectableendotoxin levels (< 0Æ1 EU ⁄ml), as measured by theLimulus assay (BioWhittaker, Verviers, Belgium). ODNwere suspended in sterile, endotoxin-free Tris–ethylenedi-
aminetetraacetic acid (Sigma, Deisenhofen, Germany), andstored and handled under aseptic conditions to preventboth microbial and endotoxin contamination. All dilutions
were carried out using pyrogen-free phosphate-bufferedsaline (Life Technologies, Eggenstein, Germany).
TLR assaysStably transfected HEK293 cells expressing human TLR9have been described previously.8,17 Briefly, HEK293 cells
were transfected by electroporation with vectors expressinghuman TLR9 and a 6xNFjB-luciferase reporter plasmid.Stable transfectants (3 · 104 cells ⁄well) were incubated
with ODN for 16 hr at 37� in a humidified incubator. Eachdata point was performed in triplicate. Cells were lysed andassayed for luciferase gene activity (using the BriteLite
kit from Perkin-Elmer, Zaventem, Belgium). Stimulationindices were calculated in reference to the reporter geneactivity of medium without addition of stimulus.
Cell purificationPeripheral blood buffy coat preparations from healthy
human donors were obtained from the Blood Bank of theUniversity of Dusseldorf (Germany) and PBMC were puri-fied by centrifugation over Ficoll–Hypaque (Sigma). Cellswere cultured in a humidified incubator at 37� in RPMI-1640
medium supplemented with 5% (v ⁄v) heat-inactivated hu-manAB serum (BioWhittaker) or 10% (v ⁄v) heat inactivatedfetal calf serum, 2 mm l-glutamine, 100 U ⁄ml penicillin and
100 lg ⁄ml streptomycin (all from Sigma).
Cytokine detection and flow cytometric analysisPBMC were resuspended at a concentration of 3 ·106 cells ⁄ml and added to 48-well flat-bottomed plates(1 ml ⁄well) or 96-well round-bottomed plates (250 ll ⁄well).PBMC were incubated with various ODN concentrationsand culture supernatants were collected after the indicatedtime points. If not used immediately, supernatants were
stored at )20� until required.Amounts of cytokines in the supernatants were assessed
using commercially available enzyme-linked immunosorb-
ent assay (ELISA) Kits [interleukin-6 (IL-6), IFN-inducibleprotein 10 (IP-10), IFN-c or IL-10; from Diaclone, Bes-ancon, France] or an in-house ELISA for IFN-a developedusing commercially available antibodies (from PBL, New
Brunswick, NJ).All monoclonal antibodies (mAb) for cell surface
staining were purchased from Becton-Dickinson (Heidel-
berg, Germany), except mAb to CD11c which wereobtained from DAKO (Hamburg, Germany). Flow cyto-metric data for CD86-expression on CD11c–, HLA-DRhigh
and CD123+ pDC were acquired on a FACSCalibur andwere analysed using the computer program cellquest
(both from Becton-Dickinson).
For B-cell proliferation assays, CD19+ B cells wereidentified after culturing 5-[and6-]carboxyfluorescein
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
TLR9 mediates non-CpG stimulation 213

diacitate succinmidyl ester (CFSE)-labelled PBMC withCpG ODN for 5 days by flow cytometry as describedbefore.9 Decreased CFSE content indicated proliferatingB cells.
Isolation of human B cells or pDC
Human B cells were isolated from whole PBMC with theCD19 B-cell isolation kit as described by the manufacturer(Miltenyi, Bergisch-Gladbach, Germany). To determinepurity, cells were stained with mAb to CD20 and CD14
and identified by flow cytometry. In all the experimentsB cells were more than 95% pure. Purified B cells(2 · 105)5 · 105 cells ⁄ml) were incubated with increasing
concentrations of ODN for 24 hr and IL-6 was measured asdescribed above. Purity was also confirmed using lipo-polysaccharide as a stimulus that should not induce IL-6
secretion from human B cells.The pDC were enriched with the BDCA-4 pDC isola-
tion kit following the manufacturer’s (Miltenyi) instruc-
tions. This enrichment was confirmed by staining with mAbto CD123, CD11c and HLA-DR and cells (5 · 105
cells ⁄ml) were cultured in the presence of IL-3 (10 ng ⁄ml;R & D Systems GmbH, Wiesbaden, Germany) for 48 hr
with 0Æ5 lm ODN. IFN-a in the supernatant was measuredas described above.
AnimalsFemale BALB ⁄c mice (6–8 weeks of age) were used for allexperiments. They were purchased from Charles River
Canada (Quebec, Canada) and housed in micro-isolators atthe animal care facility of the Ottawa Hospital ResearchInstitute. TLR9-deficient mice have been described before7
and were obtained from Dr Shizuo Akira (Osaka, Japan).The animals were back-crossed to BALB ⁄c for severalgenerations.
In vitro murine assaysBALB ⁄c or TLR9– ⁄– mouse splenocytes were used for all
in vitro assays. Spleens were removed from anaesthetizedanimals under aseptic conditions and were homogenized.Splenocytes were re-suspended in RPMI-1640 medium
(Life Technologies) supplemented with 2% normal mouseserum (Cedarlane Laboratories, Ontario, Canada), 2 mm
l-glutamine, penicillin–streptomycin solution (final con-
centration of 1000 U ⁄ml and 1 mg ⁄ml, respectively), and5 · 10)5 m b-mercaptoethanol (all from Sigma). Spleno-cytes were plated in 96-well, round-bottomed plates(5 · 106 cells ⁄ml). Each splenocyte sample was plated in
quadruplicate and the cells were incubated in a humidified5% CO2 incubator at 37� for 48 hr. Supernatants wereharvested and a commercially available assay kit for IL-6
(mouse OptEIA kit; PharMingen, Mississauga, ON,Canada) was used, according to the manufacturer’sinstructions, to assay cytokine levels.
Immunization of miceFor systemic immunization naıve BALB ⁄c or TLR9– ⁄–
mice (n ¼ 5 ⁄group) were immunized at 0 and 4 weeks byintramuscular injection with 1 lg hepatitis B surface anti-
gen (HBsAg) subtype ad (International Enzymes, CA,USA) alone or in combination with 50 lg CpG or non-CpG ODN in saline, 25 lg Al3+ (alum; Alhydrogel‘85¢-Superfos Biosector, Vedbaek, Denmark) or complete
Freund’s adjuvant (CFA; Sigma, 1 : 1 v ⁄v). Animals werebled at 4 and 8 weeks, and plasma was recovered and storedat )20� until assay.
For mucosal vaccine delivery, mice (n ¼ 5 to n ¼10 ⁄group) were immunized on days 0, 1 and 2 with 10 lgformalin-inactivated tetanus toxoid (TT, Aventis Pasteur,
Swiftwater, PA), alone or combined with 10 lg ODN insaline. Mucosal formulations were administered using a1-cm3 tuberculin syringe (Becton-Dickinson, Franklin
Lakes, NJ) attached to a 20-gauge olive tip steel feedingtube (Fine Science Tools Inc., North Vancouver, BC,Canada) which was passed into the stomach. The titre ofTT-specific antibodies in plasma was taken at 8 weeks,
which was 4 weeks post-boost.Antibodies against HBsAg (anti-HBs) or anti-TT anti-
bodies [total immunoglobulin G (IgG), IgG1 or IgG2a]
were detected and quantified by end-point dilution ELISAassay as described previously31 which was performed induplicate on samples from individual animals.
Statistical analysisStatistical analysis was performed using the instat program
(Graph PAD Software, San Diego, CA). The statisticaldifference between groups was determined by Student’st-test (for two groups) or by one-factor analysis of variance
(anova) followed by Tukey’s test (for three or more groups)on raw data or transformed data (log10, for heteroscedasticpopulations).
RESULTS
Induction of B-cell proliferation and B-cell IL-6
or IL-10 secretion by non-CpG ODN
B-cell proliferation was measured by decreasing CFSE
staining of freshly isolated PBMC. Cells staining low withCFSE were found to be mainly CD19-positive B cells forCpG or non-CpG ODN (for non-CpG ODN 1982 see
Fig. 1a). The commonly used control non-CpG ODN, 1982(for sequences see Table 1), induced human B-cell prolifer-ation, although higher concentrations were needed to yield
activation comparable to CpG ODN 2006 (Fig. 1b). Inaddition, proliferating B cells stimulated with ODN 1982showed a larger cell size and higher granularity as describedpreviously for CpG ODN (data not shown).34 Furthermore,
ODN 1982 induced B cells to secrete IL-6 and IL-10(Figs 1c,d), as has been reportedwithCpGODN.1However,higher concentrations were required and levels of cytokine
production were usually higher with CpG ODN 2006. Othernon-CpGODN such as the GC control (ODN 2137) of CpGODN 2006 behaved similarly, as shown for the secretion of
IL-10 from human PBMC in Fig. 1(e). Similar results forIL-6 secretion were obtained with non-CpG ODN 1982compared to CpG ODN 2006 when stimulation was carriedout on purified human B cells (Fig. 1f).
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
214 J. Vollmer et al.
Non-CpG ODN do not stimulate the production of type I
and type II interferons or CXCL-10
Given the ability of phosphorothioate non-CpG ODN suchas 1982 to activate human B cells, albeit with weaker effects
and at higher concentrations than required for CpG ODN,we hypothesized that non-CpG ODN might have the samefull range of immune effects as induced by CpG ODN. One
of the most striking attributes of CpG ODN is their abilityto induce strong Th1 immune responses, mediated in partby the induction of IFN-a production from pDC. However,
the results of the present study indicate that CpG ODN2006, but not ODN 1982, can stimulate significant pro-duction of IFN-a from human PBMC (Fig. 2a), even at
concentrations of up to 16 lm or at various culture periods(6 hr, 16 hr, 24 hr, or 48 hr) (data not shown). Similarresults were obtained using pDC-enriched cell cultures(Fig. 2b). The Th1 cytokine IFN-c, which was efficiently
induced by CpG ODN 2006, appeared not to be consider-ably enhanced by non-CpG ODN 1982 (Fig. 2c). Further-more, the Th1-related chemokine CXCL-10 (IP-10) was notproduced above background in the presence of ODN 1982
(Fig. 2d; and up to 2 lm, not shown). Similar results wereobtained with ODN 2137 (data not shown). To investigatefurther whether pDC respond at all to ODN lacking CpG
dinucleotides, human PBMC were cultured with CpGODN 2006 or ODN 1982 and up-regulation of CD86 wasmeasured on CD123+ CD11c– pDC by flow cytometry
(Fig. 2e). Although not as efficient as CpG ODN 2006,ODN 1982 did induce enhanced expression of CD86.
Requirement for TLR9 to mediate immune stimulation
by non-CpG ODN
It is possible that the immune stimulatory effects of non-CpG ODN could be mediated through TLR9, which is
(a) (b)
(c) (d)
(f)(e)
12
6
0
2006
12
6
0
1982
0.015 µM
20
10
0
0.031 µM
2006
0.125 µM
Cou
nts
CFSE
20
10
0
1982
20
10
0
2006
20
10
0
1982
100 101 102 103 104
19820.015 µM
CD
19CFSE
CD
19
IL-1
0 (p
g/m
l)
2006 1982 Med
0100200300400500600
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
2006 1982 Med
IL-6
(pg
/ml)
0
100
200
300
400
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
ODN (µM)
0
100
200
300
400
IL-1
0 (p
g/m
l)
2006 2137 Med
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
ODN (µM)
0
50
100
150
200
250
0·01
60·
063
0·25
01·
000
0·01
60·
063
0·25
01·
000
2006 1982 LPS Med
IL-6
(pg
/ml)
19820.125 µM
100 101 102 103 104100 10
1 10210
3 104
10010
1 10
210
310
4
100 101 102 103 104 100 101 102 103 104100 101 102 103 104
100 101 102 103 104100 101 102 103 104 100 101 102 103 104
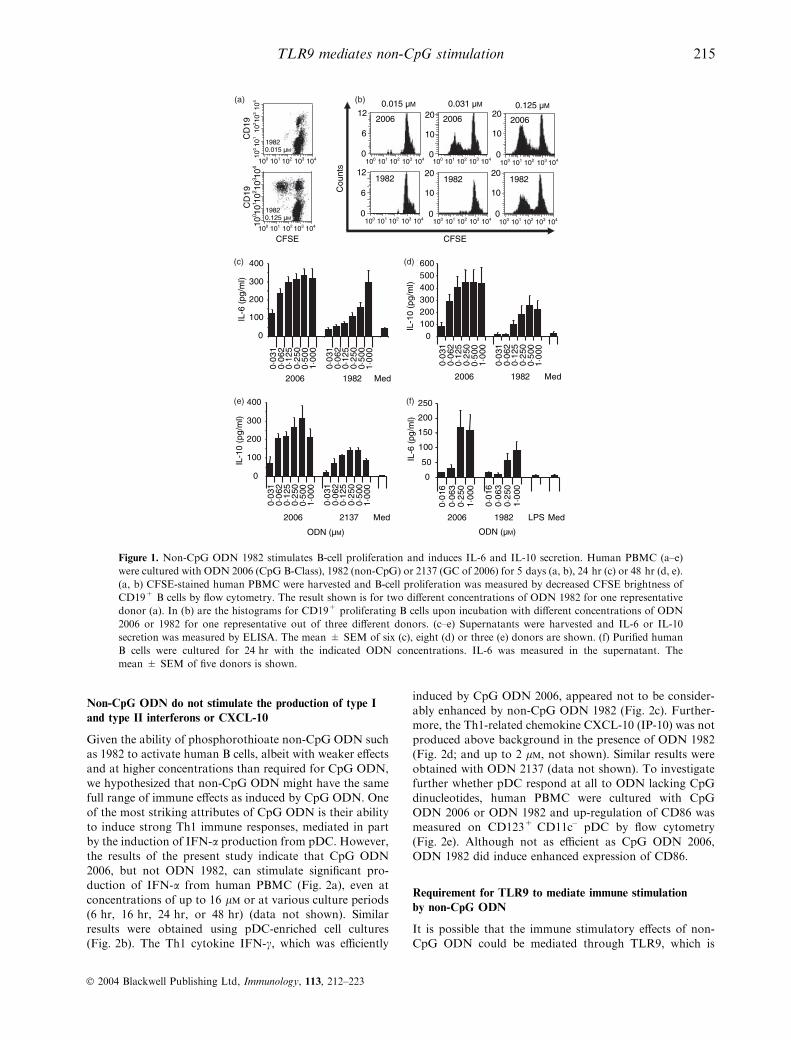
Figure 1. Non-CpG ODN 1982 stimulates B-cell proliferation and induces IL-6 and IL-10 secretion. Human PBMC (a–e)
were cultured with ODN 2006 (CpG B-Class), 1982 (non-CpG) or 2137 (GC of 2006) for 5 days (a, b), 24 hr (c) or 48 hr (d, e).
(a, b) CFSE-stained human PBMC were harvested and B-cell proliferation was measured by decreased CFSE brightness of
CD19+ B cells by flow cytometry. The result shown is for two different concentrations of ODN 1982 for one representative
donor (a). In (b) are the histograms for CD19+ proliferating B cells upon incubation with different concentrations of ODN
2006 or 1982 for one representative out of three different donors. (c–e) Supernatants were harvested and IL-6 or IL-10
secretion was measured by ELISA. The mean ± SEM of six (c), eight (d) or three (e) donors are shown. (f) Purified human
B cells were cultured for 24 hr with the indicated ODN concentrations. IL-6 was measured in the supernatant. The
mean ± SEM of five donors is shown.
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
TLR9 mediates non-CpG stimulation 215

known to be the receptor for CpG ODN, or through adifferent immune pathway. To determine the TLR9-dependency of immune stimulation induced by non-CpGODN, we first examined the ability of these ODN to
activate IL-6 production in vitro from the spleen cells ofwild-type and TLR9– ⁄– mice. Non-CpG ODN 2137 in-duced low levels of IL-6 secretion from wild-type cells
(Table 2), that were approximately one-fifth to one-tenthof those induced by CpG ODN 2006. However, neitherCpG ODN 2006 nor non-CpG ODN 2137 induced
detectable IL-6 secretion in spleen cells from TLR9– ⁄–
mice (Table 2).The optimized human CpG ODN 2006 cross-reacts with
the murine TLR9, although maximal murine TLR9 sti-mulation is only achieved with the optimized mouse CpGODN 1826.8,9 Therefore, we further evaluated in vivo CpGODN 1826 and its GC control ODN 2138 (for sequences
see Table 1) as vaccine adjuvants to HBsAg in TLR9+ ⁄+
or TLR9– ⁄– mice, and compared them to alum and CFA(Fig. 3). Mice immunized with HBsAg plus ODN 1826 had
higher antibody titres compared to animals immunized withantigen alone (Fig. 3a). The anti-HBs titres were alsohigher in mice immunized with HBsAg plus non-CpG
ODN 2138 compared to animals immunized with antigenalone at 4 weeks post-boost (P ¼ 0Æ0056) (Fig. 3a). How-ever, there was no difference in anti-HBs titres in TLR9– ⁄–
mice immunized with HBsAg alone and antigen plus CpG
ODN 1826 or non-CpG ODN 2138 (Fig. 3b). Antibodytitres in wild-type mice with non-CpG ODN 2138 werehigher than those with alum (P ¼ 0Æ01) (Fig. 3a). CFA,
which presumably can also activate immunity through
other TLRs, had an equal adjuvant effect in TLR9– ⁄– andTLR9+ ⁄+ mice (P ¼ 0Æ03 for both BALB ⁄c and TLR9– ⁄–
mice) (Fig. 3a,b).In mice, IgG isotype distribution can be used as an
indication of the Th bias of the immune response, with highIgG2a ⁄IgG1 ratios indicating a Th1-biased immuneresponse. According to the results of this study, CpG ODN
1826 induced a Th1-biased antibody response with three-and four-fold increases in IgG2a over IgG1 titres at 4 weeks(IgG2a ⁄IgG1 ratio, 2Æ6) and 8 weeks (IgG2a ⁄IgG1 ratio,
4Æ4) post primary immunization, respectively (Fig. 3c).Animals immunized with HBsAg using alum, CFA, or non-CpG ODN as adjuvant showed more Th2-biased antibody
responses with higher IgG1 over IgG2a titres (IgG2a ⁄IgG1ratios with non-CpG ODN 2138, 0Æ1 and 0Æ2 at 4 and8 weeks post-immunization, respectively) (Fig. 3c). CFAinduced a Th2-biased immune response even in TLR9– ⁄–
mice (Fig. 3d). There was no adjuvant effect of CpG ornon-CpG ODN in TLR9– ⁄– mice and they had similar titresand ratios of IgG subtypes with CpG ODN, non-CpG
ODN and HBsAg alone (Fig. 3d).
TLR9 is sufficient to restore NFjB activation by ODN
lacking CpG dinucleotides
Even though the results in the TLR9-deficient mice stronglysuggested that TLR9 was required for non-CpG ODN-mediated in vitro cytokine secretion or in vivo adjuvant
effects, it remained possible that TLR9 alone may not besufficient to mediate non-CpG ODN stimulation. To dis-tinguish between these possibilities, we tested the ability of
Table 1. ODN sequences used in this study
ODN Sequence 5¢ fi 3¢
1826 T*C*C*A*T*G*A*C*G*T*T*C*C*T*G*A*C*G*T*T
1982 T*C*C*A*G*G*A*C*T*T*C*T*C*T*C*A*G*G*T*T
2006 T*C*G*T*C*G*T*T*T*T*G*T*C*G*T*T*T*T*G*T*C*G*T*T
2137 T*G*C*T*G*C*T*T*T*T*G*T*G*C*T*T*T*T*G*T*G*C*T*T
2138 T*C*C*A*T*G*A*G*C*T*T*C*C*T*G*A*G*C*T*T
2178 C*C*C*C*C*C*C*C*C*C*C*C*C*C*C*C*C*C*C*C*C*C*C
2183 T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T
2351 T-C-C-A-T-G-A-G-C-T-T-C-C-T-G-A-G-C-T-T
2352 T*C*C-A-T-G-A-G-C-T-T-C-C-T-G*A*G*C*T*T
5314 T-C-C-A-G-G-A-C-T-T-C-T-C-T-C-A-G-G-T-T
5529 T*C*G*T*T*T*T*T*T*T*T*T*T*T*T*T*T
6056 T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T
20201 T*C*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T
21113 A*A*A*A*A*A*A*A*A*A*A*A*A*A*A*A*A*A*A*A
21114 C*G*C*G*C*G*C*G*C*G*C*G*C*G*C*G*C*G*C*G*C*G*C*G
21115 G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G*G
21570 T*C*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T
21571 T*A*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T
21572 T*G*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T
21573 T*Z*T*T*T*T*T*T*T*T*T*T*T*T*T*T*T
21698 A*C*C*A*G*G*A*C*A*A*C*A*C*A*C*A*G*G*A*A
21699 T**C*C*A*G*G*A*C*A*A*C*A*C*A*C*A*G*G*A*A
*: phosphorothioate linkage; -: phosphodiester linkage; Z: 5-methyl-cytosine.
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
216 J. Vollmer et al.

non-CpG ODN 1982 and 2137 to induce transcription froma human TLR9-transfected HEK293 NFjB reporter cellline.9 Non-CpG ODN 2138 (GC control of CpG ODN1826) induced similar TLR9-dependent NFjB activation in
experiments not shown. Incubation with increasing ODNconcentrations (Fig. 4a) resulted in considerable NFjBsignalling, although the non-CpG ODN were stronglyinferior to CpG ODN 2006 (Fig. 4b).
As with other immune stimulatory effects of non-CpGODN, the thymidine content was critical for TLR9-dependent signalling. Phosphorothioate poly (C) (ODN
2178), poly (A) (ODN 21113), poly (CG) (ODN 21114) orpoly (G) (ODN 21115) ODN did not induce considerableTLR9-mediated NFjB signalling (Fig. 4c). In contrast, a
poly (T) ODN appeared to stimulate TLR9-dependentsignalling (Fig. 4c).
Influence of the non-CpG phosphorothioate backbone
on TLR9-mediated immune stimulation
To determine the effect of the phosphorothioate backboneon the TLR9-dependent NFjB activation by ODN such as
ODN 1982 or ODN 2138, we incubated human TLR9transfectants with increasing concentrations of phos-phorothioate ODN 1982 and the same sequence except
containing only phosphodiester linkages (ODN 5314)
IFN
-α (
pg/m
l)
2006 1982 Med
0
40
80
120
160
200IF
N-γ
(pg/
ml)
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
2006 1982 Med
0
200
400
600
800
IP-1
0 (n
g/m
l)
2006 1982 Med
0
1
2
3
4
5
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0
0·50
00·
250
0·12
50·
062
0·03
1
1·00
0 0200400600800
10001200
2006 1982 Med
IFN
-α (
pg/m
l)
0·50
0
0·50
0
MF
I CD
86 p
DC
(×
102 )
2006 1982 Med
0·50
00·
250
0·12
50·
062
0·03
10·
015
0
2
4
6
8
100·
500
0·25
00·
125
0·06
20·
031
0·01
5
ODN (µM)
ODN (µM)
(a) (b)
(d)(c)
(e)
Figure 2. Non-CpG ODN do not stimulate Th1-like effects in human PBMC. Human PBMC (a, c, d) were incubated with the
indicated concentrations of ODN 2006 or 1982 for 48 hr. Supernatants were harvested and secretion of IFN-a, IFN-c, orIP-10 was measured by ELISA. Mean ± SEM of six (a, c) or nine (d) donors is shown. (b) PDC-enriched cell cultures were
incubated with 0Æ5 lm ODN 2006 or 1982 for 48 hr. Cells were harvested and IFN-a in the supernatant was measured by
ELISA. The mean ± SEM of two donors is shown. (e) Up-regulation of CD86 on CD11c–, HLA-DRhigh and CD123+ pDC
was measured upon culture of human PBMC with ODN 1982 or 2006 for 48 hr by flow cytometry and the mean ± SEM of
the mean fluorescence intensity of three donors is shown.
Table 2. IL-6 secretion induced by CpG and non-CpG ODN in
murine TLR9– ⁄– splenocytes
ODN
Conc.
(lg ⁄ml)
IL-6 secretion (pg ⁄ml)
TLR9+ ⁄+ TLR9– ⁄–
None 98Æ3 ± 4Æ9 54Æ4 ± 36Æ42006 1 923Æ8 ± 20Æ2 79Æ5 ± 73Æ9
3 1977Æ6 ± 191Æ5 80Æ8 ± 50Æ410 3449Æ2 ± 225Æ7 113Æ1 ± 30
2137 1 190 ± 5Æ6 80 ± 54Æ13 258Æ3 ± 17Æ6 97Æ1 ± 39Æ910 335 ± 21Æ7 95Æ4 ± 25Æ5
TLR9+ ⁄+ or TLR9– ⁄ – BALB ⁄c splenocytes were cultured with the
indicated ODN for 48 hr. Supernatants were harvested and IL-6 was
measured by ELISA. The mean ± SD of two independent ELISA of one out
of two experiments is shown.
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
TLR9 mediates non-CpG stimulation 217
(Fig. 5a). ODN 5314 gave little NFjB stimulation, andonly at concentrations greater than 25 lm, whereas ODN1982 showed stimulation at lower concentrations.
To determine the role of the phosphorothioate back-
bone on the adjuvant effect by non-CpG ODN 2138, weevaluated responses to oral immunization with TT usingthe same sequence with a chimeric phosphorothioate ⁄phosphodiester backbone (ODN 2352; for sequences seeTable 1) or with a phosphodiester backbone (ODN 2351).TT alone did not induce measurable plasma anti-TT anti-
body titres, and only with the addition of CpG ODN 1826or non-CpG ODN 2138 were significant levels of IgGdetected (P < 0Æ001 and P < 0Æ05 for CpG ODN 1826 and
non-CpG ODN 2138, respectively), although those withODN 2138 were inferior to those with the same dose ofCpG ODN 1826 (P < 0Æ01) (Fig. 5b). Neither the chimericnor the phosphodiester versions of ODN 2138 had a con-
siderable adjuvant effect at the dose used. Ratios of IgGsubtypes with CpG ODN, non-CpG ODN, or TT aloneindicate that only the CpG ODN induced a strong IgG2a
response. The non-CpG ODN induced a Th2-biasedresponse (Fig. 5c).
Sequence-dependence of immune stimulation mediated
by non-CpG ODN
Recently, we demonstrated that the thymidine content aswell as the length of an ODN lacking CpG dinucleotides
strongly determines its immune stimulatory potential.28 Arecent publication suggested that the presence of a sequencesuch as 5¢-CATTTTGT-3¢ in a non-CpG ODN would be
sufficient to stimulate human B-cell activation.35 Indeed,non-CpG ODN 2137 is thymidine-rich and contains at leastin part the described potential non-CpG motif.35 Never-
theless, other non-CpG ODN, such as ODN 1982 or 2138used in this study, have a lower thymidine content (35%
BALB/c (TLR9+/+)
BALB/c (TLR9+/+)
TLR9−/−4 week8 week
(a) (b)
102
103
104
105
106
Ant
i-HB
s (G
MT
)
None 1826 2138 Alum CFAAdjuvant None 1826 2138 Alum CFA
0
Ant
i-HB
s (G
MT
)
IgG1IgG2a
TLR9−/−
(c) (d)
2·5x105
5·0x105
7·5x105
Adjuvant None 1826 2138 Alum CFA None 1826 2138 Alum CFA
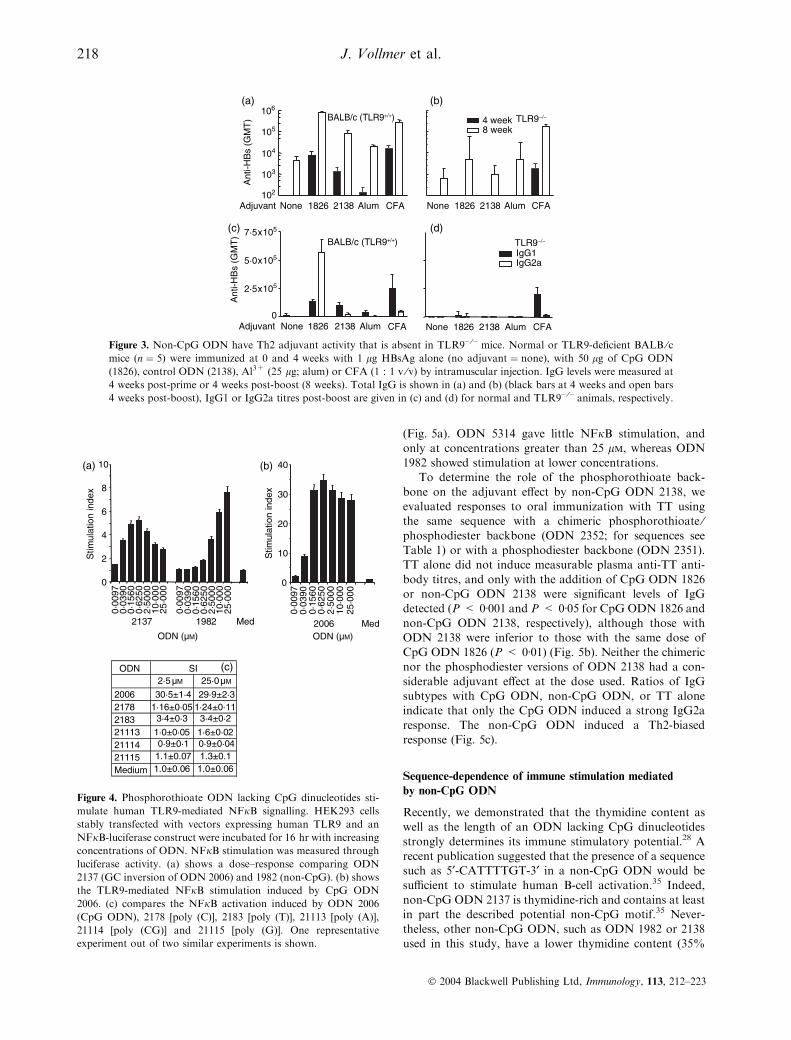
Figure 3. Non-CpG ODN have Th2 adjuvant activity that is absent in TLR9– ⁄– mice. Normal or TLR9-deficient BALB ⁄cmice (n ¼ 5) were immunized at 0 and 4 weeks with 1 lg HBsAg alone (no adjuvant ¼ none), with 50 lg of CpG ODN
(1826), control ODN (2138), Al3+ (25 lg; alum) or CFA (1 : 1 v ⁄v) by intramuscular injection. IgG levels were measured at
4 weeks post-prime or 4 weeks post-boost (8 weeks). Total IgG is shown in (a) and (b) (black bars at 4 weeks and open bars
4 weeks post-boost), IgG1 or IgG2a titres post-boost are given in (c) and (d) for normal and TLR9– ⁄– animals, respectively.
ODN (µM)
0
10
20
30
40
2006 Med
Stim
ulat
ion
inde
x
0·00
970·
0390
0·15
600·
6250
2·50
0010
·000
25·0
00
(b)
0
2
4
6
8
10
0·00
970·
0390
0·15
600·
6250
2·50
0010
·000
25·0
00
2137 1982 Med
ODN (µM)
Stim
ulat
ion
inde
x
0·00
970·
0390
0·15
600·
6250
2·50
0010
·000
25·0
00
(a)
(c)ODN2·5 µM 25·0 µM
2006 30·5±1·4 29·9±2·32178 1·16±0·05 1·24±0·112183 3·4±0·3 3·4±0·2
21113 1·0±0·05 1·6±0·0221114 0·9±0·1 0·9±0·04
21115 1.1±0.07 1.3±0.1
Medium 1.0±0.06 1.0±0.06
SI
Figure 4. Phosphorothioate ODN lacking CpG dinucleotides sti-
mulate human TLR9-mediated NFjB signalling. HEK293 cells
stably transfected with vectors expressing human TLR9 and an
NFjB-luciferase construct were incubated for 16 hr with increasing
concentrations of ODN. NFjB stimulation was measured through
luciferase activity. (a) shows a dose–response comparing ODN
2137 (GC inversion of ODN 2006) and 1982 (non-CpG). (b) shows
the TLR9-mediated NFjB stimulation induced by CpG ODN
2006. (c) compares the NFjB activation induced by ODN 2006
(CpG ODN), 2178 [poly (C)], 2183 [poly (T)], 21113 [poly (A)],
21114 [poly (CG)] and 21115 [poly (G)]. One representative
experiment out of two similar experiments is shown.
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
218 J. Vollmer et al.

compared to 58% for ODN 2137) and do not have longthymidine stretches as well as having no similarities to theabove proposed non-CpG motif. ODN 2138 and 1982 havesequence similarities and both contain a 5¢-TC. Therefore,we asked the question if the 5¢-TC would be sufficient toinduce enhanced TLR9-dependent stimulation. We com-pared the effects of a phosphorothioate 17-mer poly (T)
ODN (ODN 6056 with 5¢-TTT) with the same 17-mer se-quence containing a 5¢-TCT (ODN 20201) or a 5¢-TCG(ODN 5529) (for sequences see Table 1). Surprisingly, the
secretion of IL-10 from human PBMC was increased atleast 300% by modifying only one nucleotide from athymidine to a cytosine in ODN 20201 (Fig. 6a). A further
increase was achieved by introducing a 5¢-TCG (ODN5529). Similar results were obtained for IL-6 production(data not shown). ODN 20201 also increased TLR9-medi-ated NFjB signalling compared to poly (T) ODN 6056
after incubation of TLR9-transfected HEK293 cells
[stimulation index (SI) at 25 lm ODN 6056: 1Æ4 ± 0Æ2;ODN 20201: 2Æ9 ± 0Æ2]. Nevertheless, only ODN 5529 witha 5¢-TCG was able to induce IFN-a production fromhuman PBMC (Fig. 6b). Although stimulating consider-
able IL-10 secretion, both non-CpG ODN, 6056 and 20201,failed completely to produce IFN-a above backgroundlevels (Fig. 6b). This is in agreement with our results
obtained with ODN 1982.
5¢-TC and thymidine content determine the immune
stimulatory activity of non-CpG ODN
We wanted to investigate further the role of the 5¢-TC forhuman PBMC stimulation. It was possible that nucleotides
Stim
ulat
ion
Inde
x
5314 1982 Med
(a)
(b)
(c)
0
1
2
3
4
5
2·50
00·
625
0·15
60·
039
10·0
025
·00
50·0
0
2·50
00·
625
0·15
60·
039
10·0
0
10·0
0
SI 17±1·3
2006
None 1826 2138 2352 2351
Ant
i-TT
IgG
(G
MT
, 4 w
eeks
)
None 1826 2138 2352 2351101
102
103
104
105
IgG1IgG2a
Adjuvant
Ant
i-TT
IgG
ODN (µM)
101
102
103
104
105
Figure 5. TLR9-mediated immune stimulation by non-CpG ODN
in vitro or in vivo is dependent on the phosphorothioate backbone.
(a) TLR9 transfectants were incubated as described in Figure 4 with
non-CpG phosphorothioate ODN 1982, phosphodiester ODN 5314
or CpG ODN 2006 (10 lm). NFjB stimulation was measured
through luciferase activity. (b) BALB ⁄c mice (n ¼ 5–10) were
immunized by oral delivery of 10 lg TT alone (no adjuvant ¼ none),
or combined with 10 lg CpG ODN 1826, non-CpG control ODN
2138, the same sequence as chimeric phosphodiester ⁄phosphoro-thioate ODN (ODN 2352) or phosphodiester ODN (ODN 2351).
TT-specific IgG titres in plasma were measured 4 weeks after the
final immunization. (c) shows IgG1 and IgG2a titres.
0
100
200
300
400
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
6056 (5'-TTT) 20201 (5'-TCT) 5529 (5'-TCG) Med
ODN (µM)
IL-1
0 (p
g/m
l)
0
100
200
300
ODN (µM)
IFN
-α (
pg/m
l)
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
6056 (5'-TTT) 20201 (5'-TCT) 5529 (5'-TCG) Med
(a)
(b)
Figure 6. 5¢-TC stimulates efficient IL-10 production from human
PBMC but lacks induction of IFN-a secretion. Human PBMC
were stimulated with increasing concentrations of poly (T) ODN
6056 (5¢-TTT), 5¢-TCT ODN 20201 or 5¢-TCG ODN 5529 for
48 hr. Supernatants were harvested and IL-10 (a) or IFN-a (b)
secretion was measured by ELISA. The mean ± SEM of three
donors are shown.
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
TLR9 mediates non-CpG stimulation 219
other than cytosine would have a similar effect. Therefore,we stimulated human PBMC with phosphorothioate ODNwith a 5¢-TT (ODN 6056), 5¢-TC (ODN 20201), 5¢-TA(ODN 21571) or 5¢-TG (ODN 21572) (Fig. 7). All ODN
induced at least some IL-10 secretion from human PBMC.Nevertheless, ODN 20201 with a 5¢-TC was clearly superiorin its activity to the other non-CpG ODN. In addition,
methylation of the cytosine as in ODN 21573 (5¢-TZ, Z:5-methylated cytosine) reduced IL-10 to the same levels asthose obtained with the poly (T) ODN 6056. Elongation of
ODN 20201 (to 24 nucleotides) further increased IL-10production, as observed previously for CpG ODN.28,36
None of these described ODN stimulated the production of
IFN-a from human PBMC (data not shown).To determine if the thymidine content and the 5¢-TC
indeed contribute to the stimulatory activity of ODN 1982we generated ODN with the same sequence that lacked all
except the 5¢-T (ODN 21699) or all thymidines (ODN21698) after exchange of thymidine to adenosine. Indeed,reducing the thymidine content, as in ODN 21699, con-
siderably decreased IL-10 production from human PBMCand modification of the 5¢-TC appeared to decrease IL-10secretion further (Fig. 8).
DISCUSSION
In this study, we have extended our previous observationson the immune modulatory activity of phosphorothioate
ODN that lack unmethylated CpG dinucleotides, and wehave revealed a surprising and hitherto unsuspected diver-sity in the immune effects mediated by TLR9 stimulation.
The mechanism of immune modulation induced by such
ODN was heretofore not understood. Previous work sug-gested that CpG ODN, but not non-CpG ODN, couldmediate TLR9-mediated signalling.8,29,30 Phosphorothioate
ODN can bind to a variety of different cellular proteins25
and therefore it was possible that proteins or receptorsother than TLR9 might be involved in the recognition ofsuch ODN.
Non-CpG ODN as a vaccine adjuvant with either HBsor TT as an antigen in mice induced enhanced Th2-biasedantigen-specific antibody responses compared to the strong
Th1-dominated responses observed with CpG ODN. Boththese responses appeared to be TLR9-dependent becausethey were abrogated in TLR9– ⁄– mice using HBsAg. In
contrast, CFA which can presumably also activate throughother TLRs had equal adjuvant effects in both TLR9+ ⁄+
and TLR9– ⁄– mice. In addition, splenocytes of TLR9– ⁄–
mice did not respond by IL-6 production to in vitro culture
with non-CpG ODN in contrast to wild-type cells. TLR9was further demonstrated to be sufficient to mediate sig-nalling by non-CpG ODN, although the magnitude of
TLR9-mediated stimulation was always inferior to thatwith CpG ODN. Interestingly, the thymidine content ofphosphorothioate non-CpG ODN determined the extent of
NFjB stimulation observed with TLR9-transfected cells asreported for human immune cells.26,28,37 Only a phos-phorothioate thymidine homopolymer was able to induce
TLR9-dependent stimulation. Collectively, these in vitroand in vivo results show that TLR9 is required and sufficientfor the NFjB-mediated stimulation induced by ODNlacking CpG dinucleotides. Nevertheless, with these data
we cannot completely rule out the involvement of otherreceptors or adaptor proteins in generating additionalco-stimulatory or co-mitogenic signals beside those medi-
ated by NFjB required for immune modulation.
ODN (µM)
0·01 0·1 1
IL-1
0 (p
g/m
l)
0
50
100
150
200
250
300
350
6056 (5'-TT) 20201 (5'-TC) 20570 (24mer) 20571 (5'-TA) 20572 (5'-TG) 20573 (5'-TZ) Medium
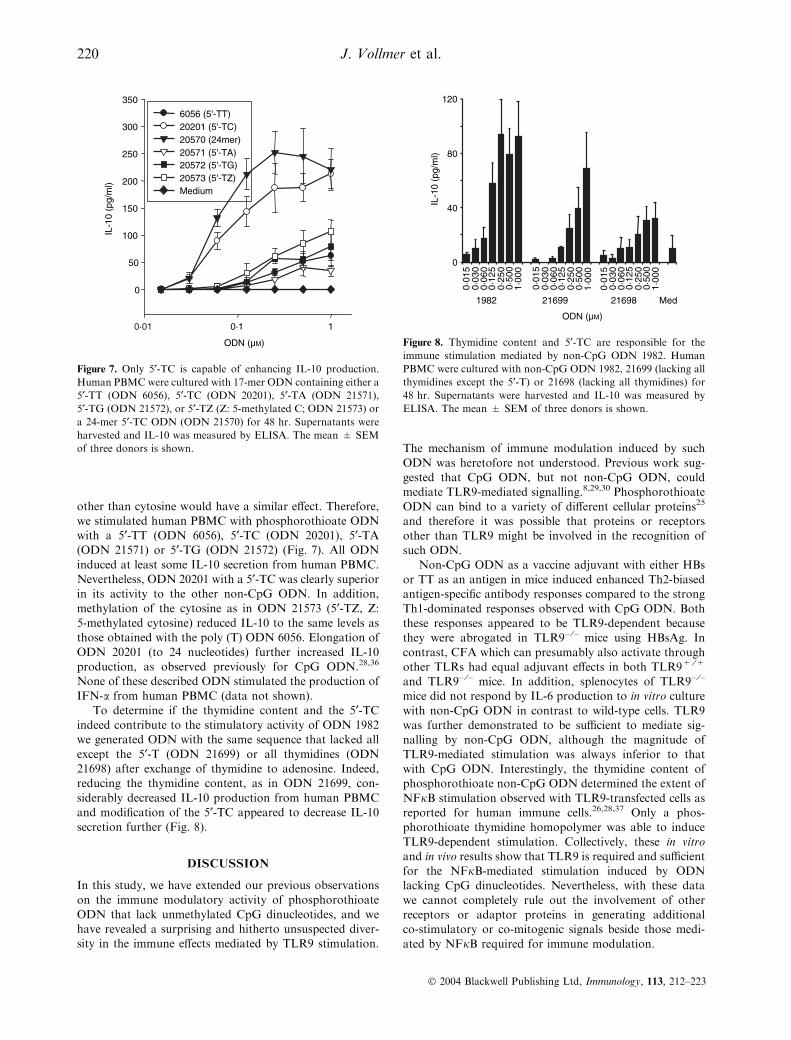
Figure 7. Only 5¢-TC is capable of enhancing IL-10 production.
Human PBMC were cultured with 17-mer ODN containing either a
5¢-TT (ODN 6056), 5¢-TC (ODN 20201), 5¢-TA (ODN 21571),
5¢-TG (ODN 21572), or 5¢-TZ (Z: 5-methylated C; ODN 21573) or
a 24-mer 5¢-TC ODN (ODN 21570) for 48 hr. Supernatants were
harvested and IL-10 was measured by ELISA. The mean ± SEM
of three donors is shown.
0
40
80
120
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
1982 21699 21698 Med
ODN (µM)
IL-1
0 (p
g/m
l)
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
0·01
50·
030
0·06
00·
125
0·25
00·
500
1·00
0
Figure 8. Thymidine content and 5¢-TC are responsible for the
immune stimulation mediated by non-CpG ODN 1982. Human
PBMC were cultured with non-CpG ODN 1982, 21699 (lacking all
thymidines except the 5¢-T) or 21698 (lacking all thymidines) for
48 hr. Supernatants were harvested and IL-10 was measured by
ELISA. The mean ± SEM of three donors is shown.
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
220 J. Vollmer et al.

Without a phosphorothioate backbone, the presence ofCpG dinucleotides becomes more critical for immune sti-mulation.28,34 Only a few reports describe immune stimu-lation mediated by phosphodiester non-CpG ODN, and
they had usually to be added at extremely high concentra-tions and on several occasions.28,35,38 PhosphorothioateODN, in contrast to phosphodiester ODN, are not only
more stable but also show better cellular uptake.4–6,23 Whenwe added a phosphodiester non-CpG ODN to cultures ofhuman TLR9-transfected cells, NFjB activation to a sim-
ilar level as that with the same phosphorothioate sequencewas observed, but only with 10–20 times higher concen-trations. In addition, Th2-dominated anti-TT responses
observed in mice using non-CpG ODN as an adjuvant toTT required the entire molecule to contain stabilizingphosphorothioate linkages.
The stimulatory capacities of phosphorothioate non-
CpG ODN on human immune cells had not previouslybeen investigated in much detail. We observed efficientstimulation of human B cells by non-CpG ODN to pro-
liferate and secrete IL-6, even when purified, although aCpG ODN was superior. Non-CpG ODN lacked com-pletely stimulation of Th1-like cytokines or chemokines
such as IFN-a, IFN-c, or IP-10. This was also true whenusing cultures enriched for pDC, the main producers ofIFN-a upon CpG-mediated stimulation.20 In contrast,reproducible up-regulation of pDC cell surface co-stimu-
latory molecules by non-CpG ODN was observed. As avaccine adjuvant in mice, non-CpG ODN induce Th2-dominated immune responses (refs 31–33 and this study),
in contrast to the Th1-biased effects seen with CpG ODN.Our in vitro results might provide an explanation forthese in vivo observations as non-CpG ODN appear to
lack one of the most important features of CpG ODN,the efficient stimulation of Th1-like cytokines, includingtype I interferons. The pDC type I interferons have been
demonstrated to be responsible for the indirect activationof a variety of immune cells such as monocytes, Th1 cells,or NK cells that do not express TLR9.1,23 Antigen-presenting cells that had become competent to present
antigen to T cells in the presence of non-CpG ODN (byup-regulation of co-stimulatory molecules) might skewT-cell responses towards Th2 in the absence of these Th1
influences. Therefore, the lack of a Th1-like pattern ofcytokine production would result in the default Th2-biased antigen-specific responses.
Recently, it was suggested that thymidine content orthe presence of a sequence such as 5¢-CATTTTGT-3¢ in anon-CpG ODN would be sufficient to stimulate humanB-cell activation.28,35 Nevertheless, this does not explain
the relatively strong stimulation observed with ODN 1982or 2138 with relatively low thymidine content and noputative non-CpG motif. We therefore investigated in
more detail the possible sequence requirements, beside thethymidine content, to induce immune stimulation by non-CpG ODN. In a previous study we described that the
addition of a 5¢-TC to a CpG ODN enhances stimulatoryactivities.2 As both non-CpG ODN contain a 5¢-TC butlack a CpG motif, we asked whether the 5¢-TC itself
would be sufficient to increase immune stimulation.Indeed, the addition of a 5¢-TC to a poly (T) ODNgreatly enhanced its capability to induce cytokine secre-tion from human PBMC, although no production of type
I interferon above background levels could be observed.In addition, none of the other tested nucleotides, adeno-sine or guanosine (as a 5¢-TA or 5¢-TG) or a 5-methylated
cytosine (as a 5¢-TZ), were able to replace cytosine toenhance cytokine secretion. Increasing the length of a5¢-TC ODN resulted in enhanced IL-10 production,
comparable to our previous results with CpG ODN.28,36
The observation that modifying the sequence of ODN1982 by deleting all thymidines including the 5¢-T adja-
cent to the cytosine considerably decreases immune sti-mulation confirms that both thymidine content as well as5¢-TC appear to be responsible for the stimulationmediated by this class of ODN. The remaining low sti-
mulation upon deletion of these motifs may be attributedto the guanosines in the sequence or the phosphorothio-ate backbone itself.1
It was previously described that a 5¢-TCG can enhancethe stimulatory activity of different CpG ODN clas-ses.2,9,36,39 In addition, we demonstrated that the CpG
guanosine position is more flexible to induce human TLR9-dependent immune stimulatory effects than the cytosineposition.40 Therefore, the 5¢-TCT could also be seen as amodification of a 5¢-TCG with the guanosine substituted by
a thymidine. Although thymine and guanine have differentstructures, TLR9 may be more forgiving at the guanosineposition and accept more substantial modifications than
can be tolerated at the cytosine position. Nevertheless,although such 5¢-TCT ODN lacking a guanosine inducesome immune stimulation, such as B-cell proliferation, they
completely lack the stimulation of Th1-like effects in vitroand in vivo. This may give rise to a completely new class ofimmune stimulatory ODN that may be useful for promo-
ting Th2 immune responses and demonstrates that anunmodified CpG dinucleotide is essential for the strong Th1stimulation induced by immune modulatory ODN.
One of the surprising aspects of our work is the
demonstration that different TLR9 stimuli can induce Th1or Th2 patterns of immune activation, depending on thepresence or absence of CpG motifs. ODN induction of type
I and type II interferon expression, and the secretion ofthe IFN-inducible protein, IP-10, shows a near absoluterequirement for the presence of CpG motifs. The explan-
ation for this finding is unlikely to be a simple matter ofaffinity, because for every readout except IFN secretion,increased concentrations of the non-CpG ODN could atleast partially restore TLR9-dependent responses, including
B-cell proliferation, co-stimulatory molecule expression, IL-10 and IL-6 secretion, and in vivo adjuvant activity. To date,only a single signalling pathway downstream from TLR9
has been identified, and this is mediated by MyD88.23 Ourdata lead us to hypothesize that CpG ODN interact with theTLR9 receptor in a qualitatively different way from non-
CpG ODN, and that although either interaction can inducemost of the immune effects of TLR9 activation, the induc-tion of the IFN promoter uniquely may be dependent on the
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
TLR9 mediates non-CpG stimulation 221

presence of a CpG motif. Further studies will be required toelucidate the molecular signalling pathways responsible forthis differential signalling through TLR9.
In summary, this is the first study to show that TLR9 is
responsible for the recognition of immune stimulatory non-CpG ODN. Such ODN in vitro and in vivo induce a Th2-like pattern of immune modulatory effects and these effects
are dependent on the thymidine content as well as thepresence of a 5¢-TC sequence. Th2 responses may havetherapeutic applications for a number of diseases such as
Crohn’s disease or organ-specific autoimmune disorders.Thus, there may be a use for such Th2-biased ODNas vaccine adjuvants or in the development of effective
Th2-dominated prophylactic or therapeutic strategies.
ACKNOWLEDGMENTS
We are grateful to Sybille Tluk, Hanna Hartmann, Tanja Wader,
Bettina Schulte and Ming Liu for excellent technical assistance. We
thank Dr S. Bauer for providing TLR9 transfected HEK293 cells
and Dr S. Akira for providing TLR9– ⁄– mice.
REFERENCES
1 Uhlmann E, Vollmer J. Recent advances in the development of
immunostimulatory oligonucleotides. Curr Opin Drug Discov
Devel 2003; 6:204–17.
2 Hartmann G, Weeratna RD, Ballas ZK et al. Delineation of a
CpG phosphorothioate oligodeoxynucleotide for activating
primate immune responses in vitro and in vivo. J Immunol 2000;
164:1617–24.
3 Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA,
Teasdale R, Koretzky GA, Klinman DM. CpG motifs in bac-
terial DNA trigger direct B-cell activation. Nature 1995;
374:546–9.
4 Zhao Q, Matson S, Herrera CJ, Fisher E, Yu H, Krieg AM.
Comparison of cellular binding and uptake of antisense phos-
phodiester, phosphorothioate, and mixed phosphorothioate and
methylphosphonate oligonucleotides. Antisense Res Dev 1993;
3:53–66.
5 Krieg AM, Matson S, Fisher E. Oligodeoxynucleotide modifi-
cations determine the magnitude of B cell stimulation by CpG
motifs. Antisense Nucl Acid Drug Dev 1996; 6:133–9.
6 Sester DP, Naik S, Beasley SJ, Hume DA, Stacey KJ. Phos-
phorothioate backbone modification modulates macrophage
activation by CpG DNA. J Immunol 2000; 165:4165–73.
7 Hemmi H, Takeuchi O, Kawai T et al. A Toll-like receptor
recognizes bacterial DNA. Nature 2000; 408:740–5.
8 Bauer S, Kirschning CJ, Hacker H, Redecke V, Hausmann S,
Akira S, Wagner H, Lipford GB. Human TLR9 confers
responsiveness to bacterial DNA via species-specific CpG motif
recognition. Proc Natl Acad Sci USA 2001; 98:9237–42.
9 Vollmer J, Weeratna R, Payette P et al. Characterization of
three CpG oligodeoxynucleotide classes with distinct immuno-
stimulatory activities. Eur J Immunol 2004; 34:251–62.
10 Latz E, Schoenemeyer A, Visintin A et al. TLR9 signals after
translocating from the ER to CpG DNA in the lysosome. Nat
Immunol 2004; 5:190–8.
11 Cornelie S, Hoebeke J, Schacht AM, Bertin B, Vicogne J,
Capron M, Riveau G. Direct evidence that toll-like receptor 9
(TLR9) functionally binds plasmid DNA by specific CpG
motifs recognition. J Biol Chem 2004; e-published.
12 Schetter C, Vollmer J. Toll-like receptors involved in the
response to microbial pathogens: development of agonists for
Toll-like receptor 9. Current Opinion Drug Discovery Devel-
opment 2004; 7:204–10.
13 Zhang D, Zhang G, Hayden MS, Greenblatt MB, Bussey C,
Flavell RA, Ghosh S. A toll-like receptor that prevents infection
by uropathogenic bacteria. Science 2004; 303:1522–6.
14 Tabeta K, Georgel P, Janssen E et al. Toll-like receptors 9 and 3
as essential components of innate immune defense against
mouse cytomegalovirus infection. Proc Natl Acad Sci USA
2004; 101:3516–21.
15 Heil F, Ahmad-Nejad P, Hemmi H et al. The Toll-like receptor
7 (TLR7) -specific stimulus loxoribine uncovers a strong rela-
tionship within the TLR7, 8 and 9 subfamily. Eur J Immunol
2003; 33:2987–97.
16 Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. Recogni-
tion of double-stranded RNA and activation of NF-kappaB by
Toll-like receptor 3. Nature 2001; 413:732–8.
17 Jurk M, Heil F, Vollmer J, Schetter C, Krieg AM, Wagner H,
Lipford G, Bauer S. Human TLR7 or TLR8 independently
confer responsiveness to the antiviral compound R-848. Nat
Immunol 2002; 3:499.
18 Heil F, Hemmi H, Hochrein H et al. Species-specific recognition
of single-stranded RNA via Toll-like receptor 7 and 8. Science
2004; 8:1526–9.
19 Diebold SS, Kaisho T, Hemmi H, Akira S, Reis e Sousa. Innate
antiviral responses by means of TLR7-mediated recognition of
single-stranded RNA. Science 2004; 303:1529–31.
20 Krug A, Towarowski A, Britsch S et al. Toll-like receptor
expression reveals CpG DNA as a unique microbial stimulus
for plasmacytoid dendritic cells which synergizes with CD40
ligand to induce high amounts of IL-12. Eur J Immunol
2001; 31:3026–37.
21 Kadowaki N, Antonenko S, Liu YJ. Distinct CpG DNA and
polyinosinic-polycytidylic acid double-stranded RNA, respect-
ively, stimulate CD11c- type 2 dendritic cell precursors and
CD11c+ dendritic cells to produce type I IFN. J Immunol
2001; 166:2291–5.
22 Jarrossay D, Napolitani G, Colonna M, Sallusto F, Lanzavec-
chia A. Specialization and complementarity in microbial mole-
cule recognition by human myeloid and plasmacytoid dendritic
cells. Eur J Immunol 2001; 31:3388–93.
23 Krieg AM. CpG motifs in bacterial DNA and their immune
effects. Annu Rev Immunol 2002; 20:709–60.
24 Monteith DK, Henry SP, Howard RB, Flournoy S, Levin AA,
Bennett CF, Crooke ST. Immune stimulation – a class effect of
phosphorothioate oligodeoxynucleotides in rodents. Anticancer
Drug Des 1997; 12:421–32.
25 Stein CA, Cheng YC. Antisense oligonucleotides as therapeutic
agents – is the bullet really magical? Science 1993; 261:1004–12.
26 Liang H, Nishioka Y, Reich CF, Pisetsky DS, Lipsky PE.
Activation of human B cells by phosphorothioate oligodeoxy-
nucleotides. J Clin Invest 1996; 98:1119–29.
27 Pisetsky DS, Reich CF III Influence of backbone chemistry
on immune activation by synthetic oligonucleotides. Biochem
Pharmacol 1999; 58:1981–8.
28 Vollmer J, Janosch A, Laucht M, Ballas ZK, Schetter C, Krieg
AM. Highly immunostimulatory CpG-free oligodeoxynucleo-
tides for activation of human leukocytes. Antisense Nucl Acid
Drug Dev 2002; 12:165–75.
29 Takeshita F, Leifer CA, Gursel I, Ishii KJ, Takeshita S, Gursel
M, Klinman DM. Cutting edge. Role of Toll-like receptor 9 in
CpG DNA-induced activation of human cells. J Immunol 2001;
167:3555–8.
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
222 J. Vollmer et al.

30 Chuang TH, Lee J, Kline L, Mathison JC, Ulevitch RJ. Toll-
like receptor 9 mediates CpG-DNA signaling. J Leukoc Biol
2002; 71:538–44.
31 McCluskie MJ, Weeratna RD, Davis HL. The potential of
oligodeoxynucleotides as mucosal and parenteral adjuvants.
Vaccine 2001; 19:2657–60.
32 McCluskie MJ, Davis HL. Oral, intrarectal and intranasal
immunizations using CpG and non-CpG oligodeoxynucleotides
as adjuvants. Vaccine 2000; 19:413–22.
33 Sano K, Shirota H, Terui T, Hattori T, Tamura G. Oligode-
oxynucleotides without CpG motifs work as adjuvant for the
induction of th2 differentiation in a sequence-independent
manner. J Immunol 2003; 170:2367–73.
34 Hartmann G, Krieg AM. Mechanism and function of a
newly identified CpG DNA motif in human primary B cells.
J Immunol 2000; 164:944–53.
35 Elias F, Flo J, Lopez RA, Zorzopulos J, Montaner A, Rodri-
guez JM. Strong cytosine-guanosine-independent immuno-
stimulation in humans and other primates by synthetic
oligodeoxynucleotides with PyNTTTTGT motifs. J Immunol
2003; 171:3697–704.
36 Jurk M, Schulte B, Kritzler A et al. C-Class CpG ODN.
Sequence requirements and characterization of immunostimu-
latory activities on mRNA level. Immunobiology 2004; in press.
37 Mannon RB, Nataraj C, Pisetsky DS. Stimulation of thymocyte
proliferation by phosphorothioate DNA oligonucleotides. Cell
Immunol 2000; 201:14–21.
38 Pisetsky DS, Reich CF III The influence of base sequence on the
immunological properties of defined oligonucleotides. Immu-
nopharmacology 1998; 40:199–208.
39 Verthelyi D, Ishii KJ, Gursel M, Takeshita F, Klinman DM.
Human peripheral blood cells differentially recognize and
respond to two distinct CPG motifs. J Immunol 2001;
166:2372–7.
40 Vollmer J, Jepsen JS, Uhlmann E, Schetter C, Jurk M, Wader
T, Wullner M, Krieg AM. Modulation of CpG oligodeoxy-
nucleotide-mediated immune stimulation by locked nucleic acid
(LNA). Oligonucleotides 2004; 14:23–31.
� 2004 Blackwell Publishing Ltd, Immunology, 113, 212–223
TLR9 mediates non-CpG stimulation 223
Copyright © 2022 FDOKUMEN




















