
Liposomal SLA co-incorporated with PO CpG ODNs or PS CpG ODNs induce the same protection against the...
8
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorporated with PO CpG ODNs or PS CpG ODNs induce the same protection against the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.vaccine.2012.03.040 ARTICLE IN PRESS G Model JVAC 13016 1–8 Vaccine xxx (2012) xxx–xxx Contents lists available at SciVerse ScienceDirect Vaccine jou rn al h om epa ge: www.elsevier.com/locate/vaccine Liposomal SLA co-incorporated with PO CpG ODNs or PS CpG ODNs induce the same protection against the murine model of leishmaniasis 1 2 Vahid Heravi Shargh a , Mahmoud Reza Jaafari a,b,∗,1 , Ali Khamesipour c , Iman Jaafari a , Seyed Amir Jalali a , Q1 Azam Abbasi a , Ali Badiee a,∗,1 3 4 a Nanotechnology Research Center, School of Pharmacy, Mashhad, University of Medical Sciences (MUMS), Mashhad, Iran 5 b Biotechnology Research Center, School of Pharmacy, MUMS, Mashhad, Iran 6 c Center for Research and Training in Skin Diseases and Leprosy, Tehran University of Medical Sciences, Tehran, Iran 7 8 a r t i c l e i n f o 9 10 Article history: 11 Received 2 September 2011 12 Received in revised form 16 February 2012 13 Accepted 16 March 2012 14 Available online xxx 15 Keywords: 16 Vaccine 17 Liposome 18 SLA 19 CpG ODNs 20 Leishmaniasis 21 a b s t r a c t First generation Leishmania vaccines consisting of whole killed parasites with or without adjuvants have reached phase 3 trial and failed to show enough efficacy mainly due to the lack of an appropriate adjuvant. In this study, the nuclease-resistant phosphorothioate CpG oligodeoxynucleotides (PS CpG) or nuclease- sensitive phosphodiester CpG ODNs (PO CpG) were used as adjuvants to enhance immunogenicity and rate of protection against leishmaniasis. Due to the susceptibility of PO CpG to nuclease degradation, an efficient liposomal delivery system was developed to protect them from degradation. 1, 2-dioleoyl-3- trimethylammonium-propane (DOTAP) as a cationic lipid was used because of its unique adjuvanticity and electrostatic interaction with negatively charged CpG ODNs. To evaluate the role of liposomal formu- lation in protection rate and enhanced immune response, BALB/c mice were immunized subcutaneously with liposomal soluble Leishmania antigens (SLA) co-incorporated with PO CpG (Lip-SLA–PO CpG), Lip- SLA–PS CpG, SLA + PO CpG, SLA + PS CpG, SLA or buffer. As criteria for protection, footpad swelling at the site of challenge, parasite loads, the levels of IFN- and IL-4, and the IgG subtypes were evaluated. The groups of mice receiving Lip-SLA–PO CpG or Lip-SLA–PS CpG showed a high protection rate compared with the control groups. In addition, there was no significant difference in immune response generation between mice immunized with PS CpG and the group receiving PO CpG when incorporated into the lipo- somes. The results suggested that liposomal form of PO CpG might be used instead of PS CpG in future vaccine formulations as an efficient adjuvant. © 2012 Published by Elsevier Ltd. 1. Introduction 22 Leishmania species are dimorphic homoflagellate intracellu- 23 lar protozoan parasites of macrophage-dendritic cell lineage that 24 cause a spectrum of diseases ranging from a self-healing cutaneous 25 lesion to potentially fatal visceral form of disease, known as leish- 26 maniasis [1,2]. The current control strategies are either ineffective 27 or hard to maintain in many foci, available drugs are expensive, 28 need multiple injections, accompany with side effects and are not 29 always effective [3–5]. 30 It is well known for centuries that long lasting protection 31 induces upon recovery from cutaneous leishmaniasis (CL) or leish- 32 manization [4,5]. Leishmania subunit vaccines has not reached 33 ∗ Corresponding authors at: Nanotechnology Research Center, School of Phar- macy, Mashhad, University of Medical Sciences (MUMS), Mashhad, Iran. Tel.: +98 511 8823255; fax: +98 511 8823251. E-mail addresses: [email protected] (M.R. Jaafari), [email protected] (A. Badiee). 1 Both authors equally designed research. Q2 phase 3 while whole killed Leishmania with or without adjuvant 34 reached to phase 3 clinical trials. However, prophylactic studies 35 showed a limited efficacy mainly due to the limited Th1 inducer 36 adjuvant for use in human [4–8]. 37 Synthetic oligodeoxynucleotides, containing unmethylated CpG 38 motifs, are extremely efficient inducer of Th1 immune response and 39 generation of cytotoxic T lymphocyte (CTL) which have shown to 40 induce protection against an extensive range of viral, bacterial and 41 some parasitic pathogens in animal models [9]. While human tri- 42 als have yielded promising results [10–12], clinical use of free CpG 43 ODNs still faces several challenges which limit their effectiveness. 44 One of the limiting factors in the success of oligonucleotide-based 45 immunotherapeutics is rapid degradation of unmodified ODNs (PO 46 CpG) within the body. This problem is diminished by modifica- 47 tions such as the replacement of non-bridging oxygen with sulfur 48 in phosphate linkages to prepare nuclease-resistant phosphoroth- 49 ioate analogs (PS CpG) [13]. Despite backbone stabilization of CpG 50 ODNs, PS-modified ODNs are still susceptible to nuclease degrada- 51 tion, albeit at a lower rate [14]. More critically, phosphorothioate 52 modification is associated with inherent disadvantages including 53 0264-410X/$ – see front matter © 2012 Published by Elsevier Ltd. doi:10.1016/j.vaccine.2012.03.040
Transcript of Liposomal SLA co-incorporated with PO CpG ODNs or PS CpG ODNs induce the same protection against the...

G
J
Ls
1
2
VQ1
A3
4
a5b6c7
8
a9
10
A11
R12
R13
A14
A
15
K16
V17
L18
S19
C20
L21
122
23
l24
c25
l26
m27
o28
n29
a30
31
i32
m33
m5
(Q2
0d
ARTICLE IN PRESS Model
VAC 13016 1–8
Vaccine xxx (2012) xxx– xxx
Contents lists available at SciVerse ScienceDirect
Vaccine
jou rn al h om epa ge: www.elsev ier .com/ locate /vacc ine
iposomal SLA co-incorporated with PO CpG ODNs or PS CpG ODNs induce theame protection against the murine model of leishmaniasis
ahid Heravi Shargha, Mahmoud Reza Jaafari a,b,∗,1, Ali Khamesipourc, Iman Jaafari a, Seyed Amir Jalali a,zam Abbasia, Ali Badieea,∗,1
Nanotechnology Research Center, School of Pharmacy, Mashhad, University of Medical Sciences (MUMS), Mashhad, IranBiotechnology Research Center, School of Pharmacy, MUMS, Mashhad, IranCenter for Research and Training in Skin Diseases and Leprosy, Tehran University of Medical Sciences, Tehran, Iran
r t i c l e i n f o
rticle history:eceived 2 September 2011eceived in revised form 16 February 2012ccepted 16 March 2012vailable online xxx
eywords:accineiposomeLApG ODNseishmaniasis
a b s t r a c t
First generation Leishmania vaccines consisting of whole killed parasites with or without adjuvants havereached phase 3 trial and failed to show enough efficacy mainly due to the lack of an appropriate adjuvant.In this study, the nuclease-resistant phosphorothioate CpG oligodeoxynucleotides (PS CpG) or nuclease-sensitive phosphodiester CpG ODNs (PO CpG) were used as adjuvants to enhance immunogenicity andrate of protection against leishmaniasis. Due to the susceptibility of PO CpG to nuclease degradation, anefficient liposomal delivery system was developed to protect them from degradation. 1, 2-dioleoyl-3-trimethylammonium-propane (DOTAP) as a cationic lipid was used because of its unique adjuvanticityand electrostatic interaction with negatively charged CpG ODNs. To evaluate the role of liposomal formu-lation in protection rate and enhanced immune response, BALB/c mice were immunized subcutaneouslywith liposomal soluble Leishmania antigens (SLA) co-incorporated with PO CpG (Lip-SLA–PO CpG), Lip-SLA–PS CpG, SLA + PO CpG, SLA + PS CpG, SLA or buffer. As criteria for protection, footpad swelling at the
site of challenge, parasite loads, the levels of IFN-� and IL-4, and the IgG subtypes were evaluated. Thegroups of mice receiving Lip-SLA–PO CpG or Lip-SLA–PS CpG showed a high protection rate comparedwith the control groups. In addition, there was no significant difference in immune response generationbetween mice immunized with PS CpG and the group receiving PO CpG when incorporated into the lipo-somes. The results suggested that liposomal form of PO CpG might be used instead of PS CpG in futuren effi
34
35
36
37
38
39
40
41
42
43
vaccine formulations as a
. Introduction
Leishmania species are dimorphic homoflagellate intracellu-ar protozoan parasites of macrophage-dendritic cell lineage thatause a spectrum of diseases ranging from a self-healing cutaneousesion to potentially fatal visceral form of disease, known as leish-
aniasis [1,2]. The current control strategies are either ineffectiver hard to maintain in many foci, available drugs are expensive,eed multiple injections, accompany with side effects and are notlways effective [3–5].
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorpagainst the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.v
It is well known for centuries that long lasting protectionnduces upon recovery from cutaneous leishmaniasis (CL) or leish-
anization [4,5]. Leishmania subunit vaccines has not reached
∗ Corresponding authors at: Nanotechnology Research Center, School of Phar-acy, Mashhad, University of Medical Sciences (MUMS), Mashhad, Iran. Tel.: +98
11 8823255; fax: +98 511 8823251.E-mail addresses: [email protected] (M.R. Jaafari), [email protected]
A. Badiee).1 Both authors equally designed research.
44
45
46
47
48
49
264-410X/$ – see front matter © 2012 Published by Elsevier Ltd.oi:10.1016/j.vaccine.2012.03.040
cient adjuvant.© 2012 Published by Elsevier Ltd.
phase 3 while whole killed Leishmania with or without adjuvant
reached to phase 3 clinical trials. However, prophylactic studies
showed a limited efficacy mainly due to the limited Th1 inducer
adjuvant for use in human [4–8].
Synthetic oligodeoxynucleotides, containing unmethylated CpG
motifs, are extremely efficient inducer of Th1 immune response and
generation of cytotoxic T lymphocyte (CTL) which have shown to
induce protection against an extensive range of viral, bacterial and
some parasitic pathogens in animal models [9]. While human tri-
als have yielded promising results [10–12], clinical use of free CpG
ODNs still faces several challenges which limit their effectiveness.
One of the limiting factors in the success of oligonucleotide-based
immunotherapeutics is rapid degradation of unmodified ODNs (PO
CpG) within the body. This problem is diminished by modifica-
tions such as the replacement of non-bridging oxygen with sulfur
in phosphate linkages to prepare nuclease-resistant phosphoroth-
orated with PO CpG ODNs or PS CpG ODNs induce the same protectionaccine.2012.03.040
ioate analogs (PS CpG) [13]. Despite backbone stabilization of CpG 50
ODNs, PS-modified ODNs are still susceptible to nuclease degrada- 51
tion, albeit at a lower rate [14]. More critically, phosphorothioate 52
modification is associated with inherent disadvantages including 53

ING Model
J
2 accine
n54
a55
fi56
C57
i58
h59
d60
t61
t62
P63
a64
O65
t66
s67
c68
L69
70
e71
h72
o73
[74
t75
t76
u77
i78
l79
80
m81
(82
C83
v84
e85
c86
l87
288
289
90
m91
o92
p93
f94
D95
I96
f97
298
99
t100
h101
a102
A103
2104
o105
C106
107
i108
L109
[110
111
b112
s113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
ARTICLEVAC 13016 1–8
V.H. Shargh et al. / V
on-sequence specific toxicity, unfavorable pharmacokinetic (PK)nd bio-distribution (BD), poor cellular uptake and lack of speci-city for target cells [15–18]. Administration of high doses of PSpG has been demonstrated to result in a significant acute toxic-
ty in primates due to transient complement activation and otheremodynamic changes which, in extreme cases, may result in car-iovascular collapse and death [14]. Moreover, PS CpG causes longerm severe side effects in mice such as induction of arthritis [19],ransient splenomegaly [20], lymphoid follicle destruction [21], andS CpG-specific IgM production [22] depending on the CG sequencend backbone modification. There are evidences that PS-modifiedDNs do not closely mimic the interaction of natural PO CpG with
oll like receptor 9 (TLR9) [17,23–25]. Based on the mentioned rea-ons and the lower price of PO CpG in comparison with PS CpG, wehose PO CpG as the main adjuvant in combination with solubleeishmania antigens (SLA) into the cationic liposomes.
One strategy to protect and extend the activity of PO CpG isncapsulation of CpG ODNs into the liposomes [26,27]. Liposomesave been used as a delivery vehicle to provide a close associationf CpG ODNs with antigens, and enhance the immune responses28]. Lipid based delivery systems are also developed to protecthe CpG ODNs payload, alter their pharmacokinetic characteris-ics and enhance immune cell targeting and facilitate intracellularptake [9]. Induction of cell-mediated immune response to poorly
mmunogenic Ags is possible through encapsulation of Ags into theiposomes, particularly cationic ones [29,30].
We have previously assessed the role of CpG ODNs in enhance-ent of immune response against two recombinant antigens
rgp63 or rLmSTI1), when entrapped into the liposomes [31,32].onsidering the promising results obtained from Leishmania crudeaccine [8], in this study, SLA was used as a model of first gen-ration vaccine with PS or PO CpG either in free forms or wheno-incorporated in liposomes to immunize BALB/c mice againsteishmaniasis.
. Materials and methods
.1. Ethics statement
The protocol was approved by the Institutional Ethical Com-ittee and Research Advisory Committee of Mashhad University
f Medical Sciences (Education Office dated March 31, 2010; pro-osal code 88527), based on the Specific National Ethical Guidelinesor Biomedical Research issued by the Research and Technology,eputy of Ministry Of Health and Medical Education (MOHME) of
ran, issued in 2005. Animals were kept in cages and provided withood and water ad libitum.
.2. Animals, parasites, SLA and CpG ODNs
Female BALB/c mice 6–8 weeks old were purchased from Pas-eur Institute (Tehran, Iran). The mice were maintained in animalouse of Pharmaceutical Research Center and fed with tap waternd laboratory pellet chow (Khorassan Javane Co., Mashhad, Iran).nimals were housed in a colony room 12/12 h light/dark cycle at1 ◦C with free access to water and food. Experiments were carriedut according to Mashhad University of Medical Sciences, Ethicalommittee Acts.
Leishmania major strain (MRHO/IR/75/ER) used in this exper-ment is the one which was used for preparation of experimentaleishmania vaccine, leishmanin, and Leishmania for leishmanization
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorpagainst the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.v
33–35].The preparation of SLA was carried out using protocol developed
y Scott et al. [36] with some modifications. Briefly, the para-ites were harvested at stationary phase and washed 4 times using
PRESS xxx (2012) xxx– xxx
HEPES buffer (10 mM, pH 7.5). Then the number of promastigotes
was adjusted to 1.2 × 109/ml in buffer solution containing enzyme
inhibitor cocktail, 50 �l/ml (Sigma, St. Louis, MO, USA) and the
preparation was incubated in ice-water bath for 10 min and then
the parasites were lysed using freeze–thaw method followed by
probe sonication in an ice bath. The supernatant of centrifuged
lysate parasites was collected, dialyzed against buffer solution, ster-
ilized using a 0.22 �m membrane and stored at −70 ◦C until use. The
protein concentration of the SLA was determined using BCA protein
assay kit (Thermo Scientific, USA).
The PS and PO CpG (Microsynth, Balgach, Switzerland) used in
this study were a 20-mer (5′-TCCATGACGTTCCTGACGTT-3′) with
fully nuclease-resistant phosphorothioate or nuclease-susceptible
phosphodiester backbones, respectively, which contains two CpG
motifs with known Th1 type immunostimulatory effects in murine
model [37,38].
2.3. Liposomes preparation and characterization
Liposomes containing SLA and CpG ODNs (Lip-SLA–PO CpG or
Lip-SLA–PS CpG) were prepared using lipid film method. Briefly,
the lipid phase consisting of 1,2-dioleoyl-3-trimethylammonium-
propane (chloride salt) (DOTAP) (4 �mol/ml; Avanti Polar lipids,
USA) and cholesterol (Avanti Polar lipids, USA) (1:1 molar ratio)
was dissolved in chloroform in a sterile tube. The solvent was then
removed by rotary evaporation (Hettich, Germany) resulting in
deposition of a thin lipid film on the tube’s wall. The lipid film was
freeze-dried (Taitec, Japan) overnight to ensure complete removal
of the solvent. The lipid film was then hydrated and dispersed in
sterile HEPES buffer containing SLA (0.5 mg/ml) at 45 ◦C. The result-
ing multilamellar vesicles (MLVs) were converted to 200–400 nm
vesicles using the bath sonicator (Branson 5510, USA). The PS or PO
CpG (200 �g/ml) was then added slowly to liposomes containing
SLA using mild shaking tube. The sonication of liposomal formula-
tions was done before injection into the mice to reduce the size of
liposomes.
Particle size analyzer (Nano-ZS, Malvern, UK) was used to deter-
mine the mean diameter and zeta potential of the liposomes. The
concentration of SLA encapsulated in liposomes was determined
using BCA protein assay kit (Thermo Scientific, USA).
2.4. SDS-PAGE analysis of SLA and liposomal SLA
Analytical SDS-PAGE was carried out to characterize and esti-
mate qualitatively the concentration of SLA encapsulated in the
prepared Lip-SLA after purification by Vivaspin6® centrifugal con-
centrators (300 kDa) (Sartorius Stedim Biotech S.A., France). The gel
was consisted of running gel (10.22%, w/v, acrylamide) and stack-
ing gel (4.78%, w/v, acrylamide). The gel thickness was 1 mm. The
electrophoresis buffer was 25 mM Tris, 192 mM glycine, 0.1% SDS,
pH 8.3. Electrophoresis was carried out at 140 V constant voltage
for 45 min. After electrophoresis the gels were stained with silver
for protein detection.
2.5. Immunization of BALB/c mice
Different groups of mice, 8 mice per group, were subcutaneously
(SC) immunized in their left hind footpad 3 times at 2 weeks
intervals with one of the following formulations: Lip-SLA–PO
CpG (25 �g SLA-10 �g PO CpG/50 �l liposome/mouse), Lip-SLA–PS
CpG (25 �g SLA–10 �g PS CpG/50 �l liposome/mouse), SLA (25 �g
orated with PO CpG ODNs or PS CpG ODNs induce the same protectionaccine.2012.03.040
SLA/50 �l buffer/mouse), SLA (25 �g SLA/50 �l buffer/mouse) plus 169
PO CpG (10 �g PO CpG/50 �l buffer/mouse), SLA (25 �g SLA/50 �l 170
buffer/mouse) plus PS CpG (10 �g PS CpG/50 �l buffer/mouse) or 171
HEPES buffer (HEPES 10 mM, sucrose 10%, pH 7.5) alone. 172

IN PRESSG Model
J
accine xxx (2012) xxx– xxx 3
2173
174
c175
w176
i177
b178
M179
s180
t181
2182
183
g184
a185
w186
h187
t188
o189
h190
a191
p192
i193
v194
s195
2196
197
a198
b199
E200
p201
o202
2203
i204
1205
T206
m207
(208
m209
2210
211
t212
r213
s214
w215
2216
c217
(218
f219
I220
m221
2222
223
n224
s225
c226
o227
s228
Table 1Particle size, polydispersity index (PDI), and surface charge of various liposomalformulations. Results denote mean ± SD (n = 3).
Z-average (nm) PDI Zeta-potential (mV)
Empty Lipa 224.5 ± 82.1 0.4 ± 0.2 64.0 ± 13.7Lip-SLA 361.0 ± 84.1 0.4 ± 0.1 45.6 ± 11.4Lip-PS CpG 529.3 ± 52.3 0.6 ± 0.2 72.5 ± 4.8Lip-PO CpG 501.2 ± 48.9 0.5 ± 0.1 76.6 ± 2.3Lip-SLA–PS CpG 359.4 ± 85.7 0.3 ± 0.1 63.9 ± 2.7Lip-SLA–PO CpG 351.5 ± 86.8 0.4 ± 0.1 63.8 ± 6.0
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
PAGE (Fig. 1). SDS-PAGE analysis of soluble SLA revealed several 252
protein bands with a range of molecular weight between 10 and 253
70 kDa (Fig. 1). The SDS-PAGE analysis of liposomal SLA after 254
ARTICLEVAC 13016 1–8
V.H. Shargh et al. / V
.6. Challenge with L. major promastigotes
The immunized mice and control group (five per group) werehallenged SC, at week 2 after the last booster, in the left footpadith 1 × 106 L. major promastigotes harvested at stationary phase
n 50 �l volume. Lesion development was recorded in each mousey measuring the footpad swelling using a metric caliper (Mitutoyoeasuring Instruments, Japan). Grading of lesion size was done by
ubtracting the thickness of uninfected contralateral footpad fromhat of the infected one.
.7. Quantitative parasite burden after challenge
The number of viable L. major parasites in the spleen of differentroup of mice were estimated using limiting dilution assay methods described previously [39]. Briefly, the mice were sacrificed ateek 9 post-challenge; the spleens were aseptically removed andomogenized in RPMI 1640 supplemented with 10% (v/v) heat inac-ivated FCS (Eurobio, Les Ulys, France), 2 mM glutamine, 100 U/mlf penicillin and 100 �g/ml of streptomycin sulfate (RPMI-FCS). Theomogenate was diluted with the media in 8 serial 10-fold dilutionsnd then was placed in each well of flat-bottom 96-well microtiterlates (Nunc, Denmark) containing solid layer of rabbit blood agar
n tetraplicate and kept at 25 ± 1 ◦C for 7–10 days. The number ofiable parasite per spleen was determined using ELIDA software, atatistical method for limiting dilution assay [40].
.8. Antibody isotype assay
Blood samples were collected from mice before and at week 9fter the challenge and the sera were isolated and kept frozen untileing used to assess anti-SLA IgG total, IgG1 and IgG2a antibodies byLISA method as described before [39]. Briefly, 96-well micro titerlates (Nunc, Denmark) were coated with 50 �l of 10 �g/ml of SLAvernight at 4 ◦C. The plates were washed and blocked by adding00 �l per well of 1% of bovine serum albumin in PBS-Tween and
ncubated for 1 h at 37 ◦C. Serum samples were diluted to 1:200,:2000 or 1:20,000 with PBS-Tween and 50 �l applied to the plates.he plates were then washed and treated with HRP-rabbit anti-ouse IgG isotype according to the manufacturer’s instructions
Zymed Laboratories Inc., USA). Optical density (OD) was deter-ined at 450 nm using 630 nm as the reference wavelength.
.9. In vitro spleen cells responses
Three mice from each group were sacrificed at week 2 afterhe last booster injection and their spleens were asepticallyemoved. Mononuclear cells were isolated by Ficoll–Hypaque den-ity centrifugation method from spleen cell suspension. The cellsere washed and resuspended in RPMI 1640-FCS and seeded at
× 106/ml in 96-well flat-bottom plates (Nunc, Denmark). Theells were stimulated in vitro with either SLA (5 �g/ml), Con A2.5 �g/ml), or medium alone, and incubated at 37 ◦C in 5% CO2or 72 h. The culture supernatants were collected and the levels ofL-4 and IFN-� were checked using ELISA method according to the
anufacturer’s instructions (MabTech, Sweden).
.10. Statistical analysis
One-way ANOVA statistical test was used to assess the sig-ificance of the differences among various groups. In the case of
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorpagainst the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.v
ignificant F value, Tukey–Kramer multiple comparisons test wasarried out as a post-test to compare the means in different groupsf mice. Results with P < 0.05 were considered to be statisticallyignificant.
a Liposomes (Lip) were composed of DOTAP and cholesterol in a molar ratio of1:1.
3. Results
3.1. Liposome characterization
Liposomes used for in vivo study were 265–445 nm vesi-
cles with mean diameters 359.4 ± 85.7 nm (PDI: 0.3 ± 0.1)
and 351.5 ± 86.8 nm (PDI: 0.4 ± 0.1) and the zeta potentials
of + 63.9 ± 2.7 and + 63.8 ± 6.0 mV as calculated by particle size ana-
lyzer for Lip-SLA–PS CpG and Lip-SLA–PO CpG, respectively. In
terms of particle size and surface charge, there was no significant
difference between PS and PO CpG in liposomal form. The parti-
cle size results for all the formulations showed a slightly high PDI
values due to the used preparation method.
As shown in Table 1, the presence of SLA in formulations
decreased the surface charge of empty liposomes, Lip-PO CpG, and
Lip-PS CpG. However, the presence of CpG ODNs in the formula-
tions did not considerably affect the surface charge compared with
empty liposomes.
The entrapment of SLA in Lip-SLA–PS CpG or Lip-SLA–PO CpG
formulations was 45% and 50%, respectively. The entrapment of CpG
ODNs in liposomes was more than 90% based on the absorption at
260 nm because of their electrostatic interaction with DOTAP. The
concentration of SLA and CpG ODNs in liposomes was adjusted to
25 �g and 10 �g per 50 �l just before injection, respectively.
Identification of SLA and liposomal SLA was performed by SDS-
orated with PO CpG ODNs or PS CpG ODNs induce the same protectionaccine.2012.03.040
Fig. 1. SDS-PAGE analysis of SLA alone and liposomal SLA. Lane 1, low-range proteinstandard (Sigma, USA); lane 2, SLA (2.5 �g); lane 3, SLA (5 �g); lane 4, liposomal SLA(2.5 �g); lane 5, liposomal SLA (5 �g).
ARTICLE IN PRESSG Model
JVAC 13016 1–8
4 V.H. Shargh et al. / Vaccine xxx (2012) xxx– xxx
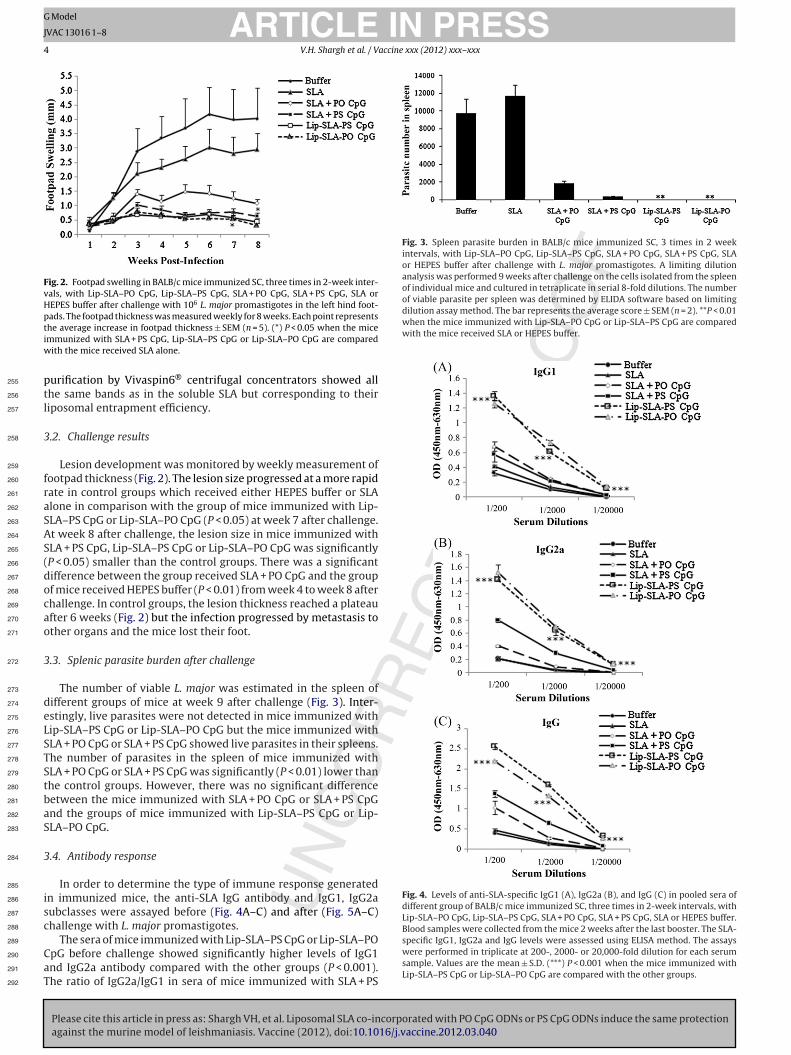
Fig. 2. Footpad swelling in BALB/c mice immunized SC, three times in 2-week inter-vals, with Lip-SLA–PO CpG, Lip-SLA–PS CpG, SLA + PO CpG, SLA + PS CpG, SLA orHEPES buffer after challenge with 106 L. major promastigotes in the left hind foot-pads. The footpad thickness was measured weekly for 8 weeks. Each point representstiw
p255
t256
l257
3258
259
f260
r261
a262
S263
A264
S265
(266
d267
o268
c269
a270
o271
3272
273
d274
e275
L276
S277
T278
S279
t280
b281
a282
S283
3284
285
i286
s287
c288
289
C290
a291
T292
Fig. 3. Spleen parasite burden in BALB/c mice immunized SC, 3 times in 2 weekintervals, with Lip-SLA–PO CpG, Lip-SLA–PS CpG, SLA + PO CpG, SLA + PS CpG, SLAor HEPES buffer after challenge with L. major promastigotes. A limiting dilutionanalysis was performed 9 weeks after challenge on the cells isolated from the spleenof individual mice and cultured in tetraplicate in serial 8-fold dilutions. The numberof viable parasite per spleen was determined by ELIDA software based on limitingdilution assay method. The bar represents the average score ± SEM (n = 2). **P < 0.01when the mice immunized with Lip-SLA–PO CpG or Lip-SLA–PS CpG are comparedwith the mice received SLA or HEPES buffer.
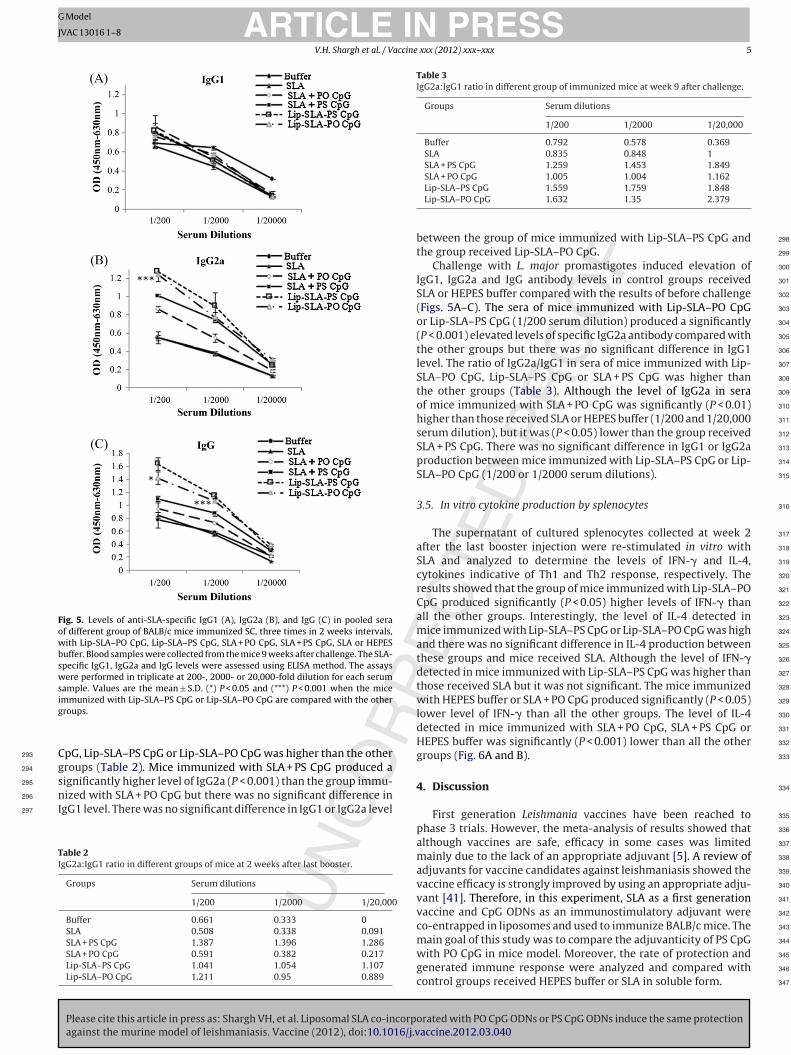
Fig. 4. Levels of anti-SLA-specific IgG1 (A), IgG2a (B), and IgG (C) in pooled sera ofdifferent group of BALB/c mice immunized SC, three times in 2-week intervals, withLip-SLA–PO CpG, Lip-SLA–PS CpG, SLA + PO CpG, SLA + PS CpG, SLA or HEPES buffer.Blood samples were collected from the mice 2 weeks after the last booster. The SLA-
he average increase in footpad thickness ± SEM (n = 5). (*) P < 0.05 when the micemmunized with SLA + PS CpG, Lip-SLA–PS CpG or Lip-SLA–PO CpG are compared
ith the mice received SLA alone.
urification by Vivaspin6® centrifugal concentrators showed allhe same bands as in the soluble SLA but corresponding to theiriposomal entrapment efficiency.
.2. Challenge results
Lesion development was monitored by weekly measurement ofootpad thickness (Fig. 2). The lesion size progressed at a more rapidate in control groups which received either HEPES buffer or SLAlone in comparison with the group of mice immunized with Lip-LA–PS CpG or Lip-SLA–PO CpG (P < 0.05) at week 7 after challenge.t week 8 after challenge, the lesion size in mice immunized withLA + PS CpG, Lip-SLA–PS CpG or Lip-SLA–PO CpG was significantlyP < 0.05) smaller than the control groups. There was a significantifference between the group received SLA + PO CpG and the groupf mice received HEPES buffer (P < 0.01) from week 4 to week 8 afterhallenge. In control groups, the lesion thickness reached a plateaufter 6 weeks (Fig. 2) but the infection progressed by metastasis tother organs and the mice lost their foot.
.3. Splenic parasite burden after challenge
The number of viable L. major was estimated in the spleen ofifferent groups of mice at week 9 after challenge (Fig. 3). Inter-stingly, live parasites were not detected in mice immunized withip-SLA–PS CpG or Lip-SLA–PO CpG but the mice immunized withLA + PO CpG or SLA + PS CpG showed live parasites in their spleens.he number of parasites in the spleen of mice immunized withLA + PO CpG or SLA + PS CpG was significantly (P < 0.01) lower thanhe control groups. However, there was no significant differenceetween the mice immunized with SLA + PO CpG or SLA + PS CpGnd the groups of mice immunized with Lip-SLA–PS CpG or Lip-LA–PO CpG.
.4. Antibody response
In order to determine the type of immune response generatedn immunized mice, the anti-SLA IgG antibody and IgG1, IgG2aubclasses were assayed before (Fig. 4A–C) and after (Fig. 5A–C)hallenge with L. major promastigotes.
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorporated with PO CpG ODNs or PS CpG ODNs induce the same protectionagainst the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.vaccine.2012.03.040
The sera of mice immunized with Lip-SLA–PS CpG or Lip-SLA–POpG before challenge showed significantly higher levels of IgG1nd IgG2a antibody compared with the other groups (P < 0.001).he ratio of IgG2a/IgG1 in sera of mice immunized with SLA + PS
specific IgG1, IgG2a and IgG levels were assessed using ELISA method. The assayswere performed in triplicate at 200-, 2000- or 20,000-fold dilution for each serumsample. Values are the mean ± S.D. (***) P < 0.001 when the mice immunized withLip-SLA–PS CpG or Lip-SLA–PO CpG are compared with the other groups.
ARTICLE IN PRESSG Model
JVAC 13016 1–8
V.H. Shargh et al. / Vaccine xxx (2012) xxx– xxx 5
Fig. 5. Levels of anti-SLA-specific IgG1 (A), IgG2a (B), and IgG (C) in pooled seraof different group of BALB/c mice immunized SC, three times in 2 weeks intervals,with Lip-SLA–PO CpG, Lip-SLA–PS CpG, SLA + PO CpG, SLA + PS CpG, SLA or HEPESbuffer. Blood samples were collected from the mice 9 weeks after challenge. The SLA-specific IgG1, IgG2a and IgG levels were assessed using ELISA method. The assayswere performed in triplicate at 200-, 2000- or 20,000-fold dilution for each serumsample. Values are the mean ± S.D. (*) P < 0.05 and (***) P < 0.001 when the miceig
C293
g294
s295
n296
I297
TI
Table 3IgG2a:IgG1 ratio in different group of immunized mice at week 9 after challenge.
Groups Serum dilutions
1/200 1/2000 1/20,000
Buffer 0.792 0.578 0.369SLA 0.835 0.848 1SLA + PS CpG 1.259 1.453 1.849SLA + PO CpG 1.005 1.004 1.162
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
mmunized with Lip-SLA–PS CpG or Lip-SLA–PO CpG are compared with the otherroups.
pG, Lip-SLA–PS CpG or Lip-SLA–PO CpG was higher than the otherroups (Table 2). Mice immunized with SLA + PS CpG produced aignificantly higher level of IgG2a (P < 0.001) than the group immu-
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorpagainst the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.v
ized with SLA + PO CpG but there was no significant difference ingG1 level. There was no significant difference in IgG1 or IgG2a level
able 2gG2a:IgG1 ratio in different groups of mice at 2 weeks after last booster.
Groups Serum dilutions
1/200 1/2000 1/20,000
Buffer 0.661 0.333 0SLA 0.508 0.338 0.091SLA + PS CpG 1.387 1.396 1.286SLA + PO CpG 0.591 0.382 0.217Lip-SLA–PS CpG 1.041 1.054 1.107Lip-SLA–PO CpG 1.211 0.95 0.889
335
336
337
338
339
340
341
342
343
Lip-SLA–PS CpG 1.559 1.759 1.848Lip-SLA–PO CpG 1.632 1.35 2.379
between the group of mice immunized with Lip-SLA–PS CpG and
the group received Lip-SLA–PO CpG.
Challenge with L. major promastigotes induced elevation ofIgG1, IgG2a and IgG antibody levels in control groups received
SLA or HEPES buffer compared with the results of before challenge
(Figs. 5A–C). The sera of mice immunized with Lip-SLA–PO CpG
or Lip-SLA–PS CpG (1/200 serum dilution) produced a significantly
(P < 0.001) elevated levels of specific IgG2a antibody compared with
the other groups but there was no significant difference in IgG1
level. The ratio of IgG2a/IgG1 in sera of mice immunized with Lip-
SLA–PO CpG, Lip-SLA–PS CpG or SLA + PS CpG was higher than
the other groups (Table 3). Although the level of IgG2a in sera
of mice immunized with SLA + PO CpG was significantly (P < 0.01)
higher than those received SLA or HEPES buffer (1/200 and 1/20,000
serum dilution), but it was (P < 0.05) lower than the group received
SLA + PS CpG. There was no significant difference in IgG1 or IgG2a
production between mice immunized with Lip-SLA–PS CpG or Lip-
SLA–PO CpG (1/200 or 1/2000 serum dilutions).
3.5. In vitro cytokine production by splenocytes
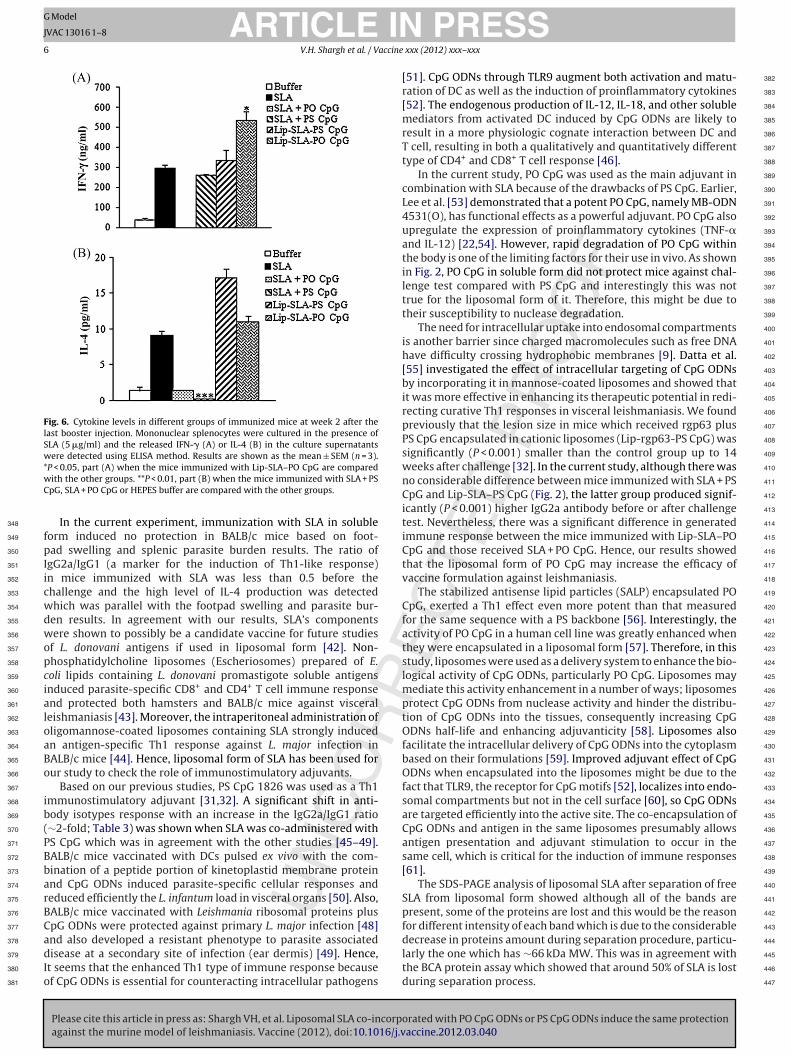
The supernatant of cultured splenocytes collected at week 2
after the last booster injection were re-stimulated in vitro withSLA and analyzed to determine the levels of IFN-� and IL-4,
cytokines indicative of Th1 and Th2 response, respectively. Theresults showed that the group of mice immunized with Lip-SLA–PO
CpG produced significantly (P < 0.05) higher levels of IFN-� than
all the other groups. Interestingly, the level of IL-4 detected in
mice immunized with Lip-SLA–PS CpG or Lip-SLA–PO CpG was high
and there was no significant difference in IL-4 production between
these groups and mice received SLA. Although the level of IFN-�
detected in mice immunized with Lip-SLA–PS CpG was higher than
those received SLA but it was not significant. The mice immunized
with HEPES buffer or SLA + PO CpG produced significantly (P < 0.05)
lower level of IFN-� than all the other groups. The level of IL-4
detected in mice immunized with SLA + PO CpG, SLA + PS CpG or
HEPES buffer was significantly (P < 0.001) lower than all the other
groups (Fig. 6A and B).
4. Discussion
First generation Leishmania vaccines have been reached to
phase 3 trials. However, the meta-analysis of results showed that
although vaccines are safe, efficacy in some cases was limited
mainly due to the lack of an appropriate adjuvant [5]. A review of
adjuvants for vaccine candidates against leishmaniasis showed the
vaccine efficacy is strongly improved by using an appropriate adju-
vant [41]. Therefore, in this experiment, SLA as a first generation
vaccine and CpG ODNs as an immunostimulatory adjuvant were
co-entrapped in liposomes and used to immunize BALB/c mice. The
orated with PO CpG ODNs or PS CpG ODNs induce the same protectionaccine.2012.03.040
main goal of this study was to compare the adjuvanticity of PS CpG 344
with PO CpG in mice model. Moreover, the rate of protection and 345
generated immune response were analyzed and compared with 346
control groups received HEPES buffer or SLA in soluble form. 347
ARTICLE ING Model
JVAC 13016 1–8
6 V.H. Shargh et al. / Vaccine
Fig. 6. Cytokine levels in different groups of immunized mice at week 2 after thelast booster injection. Mononuclear splenocytes were cultured in the presence ofSLA (5 �g/ml) and the released IFN-� (A) or IL-4 (B) in the culture supernatantswere detected using ELISA method. Results are shown as the mean ± SEM (n = 3).*wC
348
f349
p350
I351
i352
c353
w354
d355
w356
o357
p358
c359
i360
a361
l362
o363
a364
B365
o366
367
i368
b369
(370
P371
B372
b373
a374
r375
B376
C377
a378
d379
I380
o381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
P < 0.05, part (A) when the mice immunized with Lip-SLA–PO CpG are comparedith the other groups. **P < 0.01, part (B) when the mice immunized with SLA + PSpG, SLA + PO CpG or HEPES buffer are compared with the other groups.
In the current experiment, immunization with SLA in solubleorm induced no protection in BALB/c mice based on foot-ad swelling and splenic parasite burden results. The ratio of
gG2a/IgG1 (a marker for the induction of Th1-like response)n mice immunized with SLA was less than 0.5 before thehallenge and the high level of IL-4 production was detectedhich was parallel with the footpad swelling and parasite bur-en results. In agreement with our results, SLA’s componentsere shown to possibly be a candidate vaccine for future studies
f L. donovani antigens if used in liposomal form [42]. Non-hosphatidylcholine liposomes (Escheriosomes) prepared of E.oli lipids containing L. donovani promastigote soluble antigensnduced parasite-specific CD8+ and CD4+ T cell immune responsend protected both hamsters and BALB/c mice against visceraleishmaniasis [43]. Moreover, the intraperitoneal administration ofligomannose-coated liposomes containing SLA strongly inducedn antigen-specific Th1 response against L. major infection inALB/c mice [44]. Hence, liposomal form of SLA has been used forur study to check the role of immunostimulatory adjuvants.
Based on our previous studies, PS CpG 1826 was used as a Th1mmunostimulatory adjuvant [31,32]. A significant shift in anti-ody isotypes response with an increase in the IgG2a/IgG1 ratio∼2-fold; Table 3) was shown when SLA was co-administered withS CpG which was in agreement with the other studies [45–49].ALB/c mice vaccinated with DCs pulsed ex vivo with the com-ination of a peptide portion of kinetoplastid membrane proteinnd CpG ODNs induced parasite-specific cellular responses andeduced efficiently the L. infantum load in visceral organs [50]. Also,ALB/c mice vaccinated with Leishmania ribosomal proteins pluspG ODNs were protected against primary L. major infection [48]
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorpagainst the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.v
nd also developed a resistant phenotype to parasite associatedisease at a secondary site of infection (ear dermis) [49]. Hence,
t seems that the enhanced Th1 type of immune response becausef CpG ODNs is essential for counteracting intracellular pathogens
PRESS xxx (2012) xxx– xxx
[51]. CpG ODNs through TLR9 augment both activation and matu-
ration of DC as well as the induction of proinflammatory cytokines
[52]. The endogenous production of IL-12, IL-18, and other soluble
mediators from activated DC induced by CpG ODNs are likely to
result in a more physiologic cognate interaction between DC and
T cell, resulting in both a qualitatively and quantitatively different
type of CD4+ and CD8+ T cell response [46].
In the current study, PO CpG was used as the main adjuvant in
combination with SLA because of the drawbacks of PS CpG. Earlier,
Lee et al. [53] demonstrated that a potent PO CpG, namely MB-ODN
4531(O), has functional effects as a powerful adjuvant. PO CpG also
upregulate the expression of proinflammatory cytokines (TNF-�
and IL-12) [22,54]. However, rapid degradation of PO CpG within
the body is one of the limiting factors for their use in vivo. As shown
in Fig. 2, PO CpG in soluble form did not protect mice against chal-
lenge test compared with PS CpG and interestingly this was nottrue for the liposomal form of it. Therefore, this might be due totheir susceptibility to nuclease degradation.
The need for intracellular uptake into endosomal compartments
is another barrier since charged macromolecules such as free DNAhave difficulty crossing hydrophobic membranes [9]. Datta et al.
[55] investigated the effect of intracellular targeting of CpG ODNs
by incorporating it in mannose-coated liposomes and showed that
it was more effective in enhancing its therapeutic potential in redi-
recting curative Th1 responses in visceral leishmaniasis. We found
previously that the lesion size in mice which received rgp63 plus
PS CpG encapsulated in cationic liposomes (Lip-rgp63-PS CpG) was
significantly (P < 0.001) smaller than the control group up to 14
weeks after challenge [32]. In the current study, although there was
no considerable difference between mice immunized with SLA + PS
CpG and Lip-SLA–PS CpG (Fig. 2), the latter group produced signif-
icantly (P < 0.001) higher IgG2a antibody before or after challenge
test. Nevertheless, there was a significant difference in generated
immune response between the mice immunized with Lip-SLA–PO
CpG and those received SLA + PO CpG. Hence, our results showed
that the liposomal form of PO CpG may increase the efficacy of
vaccine formulation against leishmaniasis.
The stabilized antisense lipid particles (SALP) encapsulated PO
CpG, exerted a Th1 effect even more potent than that measured
for the same sequence with a PS backbone [56]. Interestingly, the
activity of PO CpG in a human cell line was greatly enhanced when
they were encapsulated in a liposomal form [57]. Therefore, in this
study, liposomes were used as a delivery system to enhance the bio-
logical activity of CpG ODNs, particularly PO CpG. Liposomes may
mediate this activity enhancement in a number of ways; liposomes
protect CpG ODNs from nuclease activity and hinder the distribu-
tion of CpG ODNs into the tissues, consequently increasing CpG
ODNs half-life and enhancing adjuvanticity [58]. Liposomes also
facilitate the intracellular delivery of CpG ODNs into the cytoplasm
based on their formulations [59]. Improved adjuvant effect of CpG
ODNs when encapsulated into the liposomes might be due to the
fact that TLR9, the receptor for CpG motifs [52], localizes into endo-
somal compartments but not in the cell surface [60], so CpG ODNs
are targeted efficiently into the active site. The co-encapsulation of
CpG ODNs and antigen in the same liposomes presumably allows
antigen presentation and adjuvant stimulation to occur in the
same cell, which is critical for the induction of immune responses
[61].
The SDS-PAGE analysis of liposomal SLA after separation of free
SLA from liposomal form showed although all of the bands are
present, some of the proteins are lost and this would be the reason
for different intensity of each band which is due to the considerable
orated with PO CpG ODNs or PS CpG ODNs induce the same protectionaccine.2012.03.040
decrease in proteins amount during separation procedure, particu- 444
larly the one which has ∼66 kDa MW. This was in agreement with 445
the BCA protein assay which showed that around 50% of SLA is lost 446
during separation process. 447

ING Model
J
accine
448
f449
w450
c451
f452
o453
o454
l455
a456
l457
t458
459
s460
H461
∼462
c463
l464
w465
o466
l467
l468
e469
b470
l471
c472
s473
o474
475
m476
o477
s478
r479
p480
[481
c482
a483
484
r485
(486
9487
T488
t489
f490
n491
r492
c493
s494
s495
r496
497
s498
a499
w500
t501
t502
i503
f504
f505
A506
507
a508
S509
e510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534
535
[ 536
537
538
539
[ 540
541
542
[ 543
544
545
546
547
[ 548
549
550
[ 551
552
553
[ 554
555
[ 556
557
558
559
[ 560
561
562
563
[ 564
565
[ 566
567
568
[ 569
570
571
[ 572
573
574
[ 575
576
577
[ 578
579
580
581
[ 582
583
584
[ 585
586
587
ARTICLEVAC 13016 1–8
V.H. Shargh et al. / V
DOTAP as a cationic lipid was used in the liposome formulationsor two reasons; firstly, to prepare positively charged liposomeshich interact more efficiently with CpG ODNs and the negatively
harged molecules on the APCs’ cell surface and so target antigensor endocytosis, and secondly, to utilize the intrinsic adjuvanticityf the synthetic quaternary ammonium compound as shown by thethers [62]. Yan et al. [62] showed that DOTAP is an active stimu-ator of DC resulting in extracellular-signal-regulated kinase (ERK)ctivation and CC chemokine (or �-chemokine) induction. Cationicipids with a quaternary ammonium group are more immunopo-ent than those with a tertiary ammonium group [63].
Due to the presence of DOTAP in the formulations, the neturface charge of empty liposomes was highly positive (+64 mV).owever, liposomes containing SLA had a zeta potential of+46 mV (Table 1). It means that SLA’s proteins have a negative
harge at pH 7.5 and interact electrostatically with the cationicipid in liposome bilayers. Hence, the zeta potential of liposomes
ill decrease. The electrostatic interaction could be the reasonf high entrapment efficiency of SLA in the positively chargediposomes (∼50%) compared with neutral ones (∼30%; unpub-ished data). Moreover, due to the fact that the zeta potential ofmpty liposomes was greater than those loaded with SLA, it cane concluded that some proteins in SLA were not entrapped within
iposomes and these fractions might be responsible for the surfaceharge decreasing. However, the zeta potential of empty liposomeshowed no significant change in the presence of CpG ODNs becausef very low molar ratio of CpG ODNs/DOTAP (7.5 × 10−3).
To assess the protection rate, footpad swelling induced by L.ajor was measured in immunized mice and compared with that
f the control groups. The results indicated the superiority of lipo-omal form of CpG ODNs in order to stimulate a Th1 immuneesponse. An improvement of adjuvanticity of CpG ODNs wasreviously reported by encapsulation of CpG ODNs in liposomes55,64–66]. However, PS CpG in their free forms showed signifi-antly higher protection rate than PO CpG as shown in antibodynd cytokine assay results.
Interestingly, IL-4 was produced in the spleen cells of miceeceived Lip-SLA–PS CpG or Lip-SLA–PO CpG after immunizationFig. 6B). However, the level of IL-4 decreased significantly at week
post-infection compared to the control groups (data not shown).he evidences that early IL-4 is needed to drive Th1 differentia-ion [67], to enhance IFN-� secretion [68,69] and to prime PBMCor cytokine production [70] imply that early IL-4 production doesot hinder the Th1 response. It might be in line with observationsevealing that IL-4 is crucial for the priming of long-term CD8+ T-ell memory responses [71–73]. As proposed by Sacks et al. [74], iteems that early IL-4 response might not be required to promoteusceptibility and there are more complexities in the mechanismsesponsible for acquired immunity.
The current data generated in murine model of L. major infectionhowed promising role of CpG ODNs and DOTAP cationic liposomess adjuvants to enhance stronger immune response against SLAhich is a crude Leishmania antigen. It is interesting to note that
here was no significant difference between PS CpG and PO CpG inerms of their ability to induce Th1 response when encapsulatednto the liposomes. Therefore, our results suggested that liposomalorm of PO CpG might be used instead of PS CpG in future vaccineormulations as an efficient adjuvant.
cknowledgment
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorpagainst the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.v
The financial support of the Nanotechnology Research Centernd Biotechnology Research Center, Mashhad University of Medicalciences (MUMS), and Center for Research and Training in Skin Dis-ases and Leprosy, Tehran University of Medical Sciences (TUMS)
[
[
PRESS xxx (2012) xxx– xxx 7
are gratefully acknowledged. This study was part of Pharm.D. thesis
of VHS that was done in Nanotechnology Research Center, MUMS,
Iran.
References
[1] Alexander J, Satoskar AR, Russell DG. Leishmania species: models of intracel-
lular parasitism. J Cell Sci 1999;112(18):2993–3002.
[2] Modabber F. Leishmaniasis vaccines: past, present and future. Int J Antimicrob
Agents 2010;36(Suppl. 1):S58–61.
[3] Croft SL, Sundar S, Fairlamb AH. Drug resistance in leishmaniasis. Clin Microbiol
Rev 2006;19(1):111–26.
[4] Modabber F. Vaccines against leishmaniasis. Ann Trop Med Parasitol
1995;89(Suppl. 1):83–8.
[5] Khamesipour A, Rafati S, Davoudi N, Maboudi F, Modabber F. Leishmania-
sis vaccine candidates for development: a global overview. Indian J Med Res
2006;123(3):423–38.
[6] Melby PC. Vaccination against cutaneous leishmaniasis: current status. Am J
Clin Dermatol 2002;3(8):557–70.
[7] Noazin S, Khamesipour A, Moulton LH, Tanner M, Nasseri K, Modabber F, et al.
Efficacy of killed whole-parasite vaccines in the prevention of leishmaniasis—a
meta-analysis. Vaccine 2009;27(35):4747–53.
[8] Noazin S, Modabber F, Khamesipour A, Smith PG, Moulton LH, Nasseri K, et al.
First generation leishmaniasis vaccines: a review of field efficacy trials. Vaccine2008;26(52):6759–67.
[9] Wilson KD, de Jong SD, Tam YK. Lipid-based delivery of CpG oligonucleotides
enhances immunotherapeutic efficacy. Adv Drug Deliv Rev 2009;61(3):233–42.
10] Krieg AM, Efler SM, Wittpoth M, Al Adhami MJ, Davis HL. Induction of sys-
temic TH1-like innate immunity in normal volunteers following subcutaneous
but not intravenous administration of CPG 7909, a synthetic B-class CpG
oligodeoxynucleotide TLR9 agonist. J Immunother 2004;27(6):460–71.
11] Cooper CL, Davis HL, Angel JB, Morris ML, Elfer SM, Seguin I, et al. CPG 7909
adjuvant improves hepatitis B virus vaccine seroprotection in antiretroviral-
treated HIV-infected adults. AIDS 2005;19(14):1473–9.
12] Friedberg JW, Kim H, McCauley M, Hessel EM, Sims P, Fisher DC, et al.
Combination immunotherapy with a CpG oligonucleotide (1018 ISS) andrituximab in patients with non-Hodgkin lymphoma: increased interferon-
alpha/beta-inducible gene expression, without significant toxicity. Blood
2005;105(2):489–95.
13] Brown DA, Kang SH, Gryaznov SM, DeDionisio L, Heidenreich O, Sullivan S, et al.
Effect of phosphorothioate modification of oligodeoxynucleotides on specific
protein binding. J Biol Chem 1994;269(43):26801–5.
14] Levin AA. A review of the issues in the pharmacokinetics and toxicol-
ogy of phosphorothioate antisense oligonucleotides. Biochim Biophys Acta1999;1489(1):69–84.
15] Agrawal S, Temsamani J, Galbraith W, Tang J. Pharmacokinetics of antisense
oligonucleotides. Clin Pharmacokinet 1995;28(1):7–16.
16] Sands H, Gorey-Feret LJ, Cocuzza AJ, Hobbs FW, Chidester D, Trainor GL.
Biodistribution and metabolism of internally 3H-labeled oligonucleotides. I.
Comparison of a phosphodiester and a phosphorothioate. Mol Pharmacol
1994;45(5):932–43.
17] Zhao Q, Matson S, Herrera CJ, Fisher E, Yu H, Krieg AM. Comparison of cel-
lular binding and uptake of antisense phosphodiester, phosphorothioate, and
mixed phosphorothioate and methylphosphonate oligonucleotides. Antisense
Res Dev 1993;3(1):53–66.
18] Krieg AM, Stein CA. Phosphorothioate oligodeoxynucleotides: antisense or
anti-protein? Antisense Res Dev 1995;5(4):241.
19] Deng GM, Nilsson IM, Verdrengh M, Collins LV, Tarkowski A. Intra-articularly
localized bacterial DNA containing CpG motifs induces arthritis. Nat Med
1999;5(6):702–5.
20] Sparwasser T, Hultner L, Koch ES, Luz A, Lipford GB, Wagner H. Immunostimu-
latory CpG-oligodeoxynucleotides cause extramedullary murine hemopoiesis.
J Immunol 1999;162(4):2368–74.
21] Heikenwalder M, Polymenidou M, Junt T, Sigurdson C, Wagner H, Akira S,
et al. Lymphoid follicle destruction and immunosuppression after repeated CpG
oligodeoxynucleotide administration. Nat Med 2004;10(2):187–92.
22] Kim D, Rhee JW, Kwon S, Sohn WJ, Lee Y, Kim DW, et al. Immunostimulation
and anti-DNA antibody production by backbone modified CpG-DNA. BiochemBiophys Res Commun 2009;379(2):362–7.
23] Kindrachuk J, Potter JE, Brownlie R, Ficzycz AD, Griebel PJ, Mookherjee
N, et al. Nucleic acids exert a sequence-independent cooperative effect
on sequence-dependent activation of Toll-like receptor 9. J Biol Chem
2007;282(19):13944–53.
24] Haas T, Metzger J, Schmitz F, Heit A, Muller T, Latz E, et al. The DNA sugar
backbone 2′ deoxyribose determines toll-like receptor 9 activation. Immunity
2008;28(3):315–23.
25] Yasuda K, Rutz M, Schlatter B, Metzger J, Luppa PB, Schmitz F, et al. CpG
motif-independent activation of TLR9 upon endosomal translocation of natural
phosphodiester DNA. Eur J Immunol 2006;36(2):431–6.
orated with PO CpG ODNs or PS CpG ODNs induce the same protectionaccine.2012.03.040
26] Klinman DM. Use of CpG oligodeoxynucleotides as immunoprotective agents. 588
Expert Opin Biol Ther 2004;4(6):937–46. 589
27] Semple SC, Klimuk SK, Harasym TO, Hope MJ. Lipid-based formulations of anti- 590
sense oligonucleotides for systemic delivery applications. Methods Enzymol 591
2000;313:322–41. 592

ING Model
J
8 accine
[593
594
[595
596
597
598
[599
600
601
602
[603
604
605
606
607
[608
609
610
611
[612
613
614
615
[616
617
618
619
[620
621
622
623
[624
625
626
[627
628
629
[630
631
632
[633
634
635
[636
637
[638
639
[640
641
642
[643
644
645
646
[647
648
649
650
651
[652
653
654
655
[656
657
658
659
660
[661
662
663
[664
665
666
667
[668
669
670
[ 671
672
673
674
[ 675
676
[ 677
678
[ 679
680
681
[ 682
683
684
[ 685
686
687
[ 688
689
690
[ 691
692
693
[ 694
695
696
[ 697
698
699
[ 700
701
[ 702
703
704
[ 705
706
707
[ 708
709
710
[ 711
712
713
[ 714
715
716
[ 717
718
719
[ 720
721
722
[ 723
724
725
[ 726
727
728
[ 729
730
731
732
[ 733
734
735
[ 736
737
738
739
[73] Stager S, Alexander J, Kirby AC, Botto M, Rooijen NV, Smith DF, et al. Natu- 740
ARTICLEVAC 13016 1–8
V.H. Shargh et al. / V
28] Klinman DM, Currie D, Gursel I, Verthelyi D. Use of CpG oligodeoxynucleotidesas immune adjuvants. Immunol Rev 2004;199(1):201–16.
29] Chikh GG, Kong S, Bally MB, Meunier J-C, Schutze-Redelmeier M-PM. Efficientdelivery of antennapedia homeodomain fused to CTL epitope with liposomesinto dendritic cells results in the activation of CD8+ T Cells. J Immunol2001;167(11):6462–70.
30] Nakanishi T, Kunisawa J, Hayashi A, Tsutsumi Y, Kubo K, Nakagawa S, et al.Positively charged liposome functions as an efficient immunoadjuvant ininducing cell-mediated immune response to soluble proteins. J Control Release1999;61(1–2):233–40.
31] Badiee A, Jaafari MR, Samiei A, Soroush D, Khamesipour A. Coencapsulationof CpG oligodeoxynucleotides with recombinant Leishmania major stress-inducible protein 1 in liposome enhances immune response and protectionagainst leishmaniasis in immunized BALB/c mice. Clin Vaccine Immunol2008;15(4):668–74.
32] Jaafari MR, Badiee A, Khamesipour A, Samiei A, Soroush D, Kheiri MT, et al. Therole of CpG ODN in enhancement of immune response and protection in BALB/cmice immunized with recombinant major surface glycoprotein of Leishmania(rgp63) encapsulated in cationic liposome. Vaccine 2007;25(32):6107–17.
33] Javadian E, Nadim A, Tahvildare-Bidruni G, Assefi V. Epidemiology of cuta-neous leishmaniasis in Iran: B. Khorassan part V: report on a focus ofzoonotic cutaneous leishmaniasis in Esferayen. Bull Soc Pathol Exot Filiales1976;69(2):140–3.
34] Momeni AZ, Jalayer T, Emamjomeh M, Khamesipour A, Zicker F, GhassemiRL, et al. A randomised, double-blind, controlled trial of a killed L. majorvaccine plus BCG against zoonotic cutaneous leishmaniasis in Iran. Vaccine1999;17(5):466–72.
35] Sharifi I, FeKri AR, Aflatonian MR, Khamesipour A, Nadim A, Mousavi MR,et al. Randomised vaccine trial of single dose of killed Leishmania majorplus BCG against anthroponotic cutaneous leishmaniasis in Bam, Iran. Lancet1998;351(9115):1540–3.
36] Scott P, Pearce E, Natovitz P, Sher A. Vaccination against cutaneous leishma-niasis in a murine model. I. Induction of protective immunity with a solubleextract of promastigotes. J Immunol 1987;139(1):221–7.
37] Chu RS, Targoni OS, Krieg AM, Lehmann PV, Harding CV. CpG Oligodeoxynu-cleotides act as adjuvants that Switch on T helper 1 (Th1) immunity. J Exp Med1997;186(10):1623–31.
38] Weeratna RD, Brazolot Millan CL, McCluskie MJ, Davis HL. CpG ODN can re-direct the Th bias of established Th2 immune responses in adult and youngmice. FEMS Immunol Med Microbiol 2001;32(1):65–71.
39] Badiee A, Jaafari MR, Khamesipour A. Leishmania major: immune response inBALB/c mice immunized with stress-inducible protein 1 encapsulated in lipo-somes. Exp Parasitol 2007;115(2):127–34.
40] Taswell C. Limiting dilution assays for the determination of immunocompetentcell frequencies. I. Data analysis. J Immunol 1981;126(4):1614–9.
41] Mutiso JM, Macharia JC, Gicheru MM. A review of adjuvants for Leishmaniavaccine candidates. J Biomed Res 2010;24(1):16–25.
42] Afrin F, Rajesh R, Anam K, Gopinath M, Pal S, Ali N. Characterization of Leish-mania donovani antigens encapsulated in liposomes that induce protectiveimmunity in BALB/c mice. Infect Immun 2002;70(12):6697–706.
43] Sharma SK, Dube A, Nadeem A, Khan S, Saleem I, Garg R, et al. Non PC liposomeentrapped promastigote antigens elicit parasite specific CD8+ and CD4+ T-cellimmune response and protect hamsters against visceral leishmaniasis. Vaccine2006;24(11):1800–10.
44] Shimizu Y, Takagi H, Nakayama T, Yamakami K, Tadakuma T, Yokoyama N, et al.Intraperitoneal immunization with oligomannose-coated liposome-entrappedsoluble leishmanial antigen induces antigen-specific T-helper type immuneresponse in BALB/c mice through uptake by peritoneal macrophages. ParasiteImmunol 2007;29(5):229–39.
45] Walker PS, Scharton-Kersten T, Krieg AM, Love-Homan L, Rowton ED, UdeyMC, et al. Immunostimulatory oligodeoxynucleotides promote protectiveimmunity and provide systemic therapy for leishmaniasis via IL-12- and IFN-gamma-dependent mechanisms. Proc Natl Acad Sci USA 1999;96(12):6970–5.
46] Rhee EG, Mendez S, Shah JA, Wu C-y Kirman JR, Turon TN, et al. Vaccina-tion with heat-killed leishmania antigen or recombinant leishmanial proteinand CpG Oligodeoxynucleotides induces long-term memory CD4+ and CD8+T cell responses and protection against Leishmania major infection. J Exp Med2002;195(12):1565–73.
47] Tafaghodi M, Khamesipour A, Jaafari MR. Immunization against leishmaniasisby PLGA nanospheres encapsulated with autoclaved Leishmania major (ALM)and CpG-ODN. Parasitol Res 2010;108(5):1265–73.
48] Iborra S, Parody N, Abanades DR, Bonay P, Prates D, Novais FO, et al. Vaccination
Please cite this article in press as: Shargh VH, et al. Liposomal SLA co-incorpagainst the murine model of leishmaniasis. Vaccine (2012), doi:10.1016/j.v
with the Leishmania major ribosomal proteins plus CpG oligodeoxynucleotidesinduces protection against experimental cutaneous leishmaniasis in mice.Microbes Infect 2008;10(10–11):1133–41.
49] Ramirez L, Iborra S, Cortes J, Bonay P, Alonso C, Barral-Netto M, et al. BALB/c micevaccinated with Leishmania major ribosomal proteins extracts combined with
[
PRESS xxx (2012) xxx– xxx
CpG oligodeoxynucleotides become resistant to disease caused by a secondary
parasite challenge. J Biomed Biotechnol 2009;2010:181690.
50] Agallou M, Margaroni M, Karagouni E. Cellular vaccination with bone marrow-
derived dendritic cells pulsed with a peptide of Leishmania infantum KMP-11
and CpG oligonucleotides induces protection in a murine model of visceral
leishmaniasis. Vaccine 2011;29(31):5053–64.
51] Dumont FJ. Modulation of Th1 and Th2 responses for immunotherapy. Exp Opin
Ther Patents 2002;12(3):341–67.
52] Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H, et al. A Toll-like
receptor recognizes bacterial DNA. Nature 2000;408(6813):740–5.
53] Lee KW, Jung J, Lee Y, Kim TY, Choi SY, Park J, et al. Immunostimulatory
oligodeoxynucleotide isolated from genome wide screening of Mycobacteriumbovis chromosomal DNA. Mol Immunol 2006;43(13):2107–18.
54] Kwon HJ, Lee KW, Yu SH, Han JH, Kim DS. NF-kappaB-dependent regulation of
tumor necrosis factor-alpha gene expression by CpG-oligodeoxynucleotides.
Biochem Biophys Res Commun 2003;311(1):129–38.
55] Datta N, Mukherjee S, Das L, Das PK. Targeting of immunostimulatory DNA
cures experimental visceral leishmaniasis through nitric oxide up-regulationand T cell activation. Eur J Immunol 2003;33(6):1508–18.
56] Mui B, Raney SG, Semple SC, Hope MJ. Immune stimulation by a CpG-containing
oligodeoxynucleotide is enhanced when encapsulated and delivered in lipidparticles. J Pharmacol Exp Ther 2001;298(3):1185–92.
57] Kim D, Rhee JW, Kwon S, Kim YE, Choi SY, Park J, et al. Enhancement of
immunomodulatory activity by liposome-encapsulated natural phosphodi-
ester bond CpG-DNA in a human B cell line. BMB Rep 2010;43(4):250–6.
58] De Oliveira MC, Boutet V, Fattal E, Boquet D, Grognet J-M, Couvreur P, et al.
Improvement of in vivo stability of phosphodiester oligonucleotide usinganionic liposomes in mice. Life Sci 2000;67(13):1625–37.
59] Zhang YM, Rusckowski M, Liu N, Liu C, Hnatowich DJ. Cationic liposomes
enhance cellular/nuclear localization of 99mTc-antisense oligonucleotides in
target tumor cells. Cancer Biother Radiopharm 2001;16(5):411–9.
60] Krieg AM. CpG motifs in bacterial DNA and their immune effects. Ann Rev
Immunol 2002;20(1):709–60.
61] Mutwiri GK, Nichani AK, Babiuk S, Babiuk LA. Strategies for enhancing the
immunostimulatory effects of CpG oligodeoxynucleotides. J Control Release2004;97(1):1–17.
62] Yan W, Chen W, Huang L. Mechanism of adjuvant activity of cationic lipo-
some: Phosphorylation of a MAP kinase, ERK and induction of chemokines.
Mol Immunol 2007;44(15):3672–81.
63] Vangasseri DP, Cui Z, Chen W, Hokey DA, Falo LD, Huang L. Immuno-
stimulation of dendritic cells by cationic liposomes. Mol Membrane Biol
2006;23(5):385–95.
64] Jiao X, Wang RY-H, Qiu Q, Alter HJ, Shih JW-K. Enhanced hepatitis C virus NS3
specific Th1 immune responses induced by co-delivery of protein antigen and
CpG with cationic liposomes. J Gen Virol 2004;85(6):1545–53.
65] Li WM, Dragowska WH, Bally MB, Schutze-Redelmeier M-P. Effective induction
of CD8+ T-cell response using CpG oligodeoxynucleotides and HER-2/neu-derived peptide co-encapsulated in liposomes. Vaccine 2003;21(23):3319–29.
66] Suzuki Y, Wakita D, Chamoto K, Narita Y, Tsuji T, Takeshima T, et al. Liposome-
encapsulated CpG oligodeoxynucleotides as a potent adjuvant for inducing type
1 innate immunity. Cancer Res 2004;64(23):8754–60.
67] Kamogawa Y, Minasi LA, Carding SR, Bottomly K, Flavell RA. The relationship
of IL-4- and IFN gamma-producing T cells studied by lineage ablation of IL-4-
producing cells. Cell 1993;75(5):985–95.
68] Platzer C, Richter G, Uberla K, Muller W, Blocker H, Diamantstein T, et al. Anal-
ysis of cytokine mRNA levels in interleukin-4-transgenic mice by quantitative
polymerase chain reaction. Eur J Immunol 1992;22(5):1179–84.
69] Noble A, Kemeny DM. Interleukin-4 enhances interferon-gamma synthesis
but inhibits development of interferon-gamma-producing cells. Immunology
1995;85(3):357–63.
70] D’Andrea A, Ma X, Aste-Amezaga M, Paganin C, Trinchieri G. Stimulatory and
inhibitory effects of interleukin (IL)-4 and IL-13 on the production of cytokines
by human peripheral blood mononuclear cells: priming for IL-12 and tumor
necrosis factor alpha production. J Exp Med 1995;181(2):537–46.
71] Huang LR, Chen FL, Chen YT, Lin YM, Kung JT. Potent induction of long-term
CD8+ T cell memory by short-term IL-4 exposure during T cell receptor stimu-
lation. Proc Natl Acad Sci USA 2000;97(7):3406–11.
72] Morrot A, Hafalla JC, Cockburn IA, Carvalho LH, Zavala F. IL-4 receptor
expression on CD8+ T cells is required for the development of protec-
tive memory responses against liver stages of malaria parasites. J Exp Med
2005;202(4):551–60.
orated with PO CpG ODNs or PS CpG ODNs induce the same protectionaccine.2012.03.040
ral antibodies and complement are endogenous adjuvants for vaccine-induced 741
CD8+ T-cell responses. Nat Med 2003;9(10):1287–92. 742
74] Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to 743
Leishmania major in mice. Nat Rev Immunol 2002;2(11):845–58. 744




















