
Uranium extraction from purified wet process Jordanian phosphoric acid: a development study
Upload
pasaycitysciencehighCategory
view
3download
0
Vaccine 22 (2003) 77–86
Use of a purifiedTrypanosoma cruziantigen and CpGoligodeoxynucleotides for immunoprotection against
a lethal challenge with trypomastigotes
Fernanda M. Franka,b, Patricia B. Petrayc, Silvia I. Cazorlaa,b, Marina C. Muñoza,b,Ricardo S. Corralc, Emilio L. Malchiodia,b,∗
a Cátedra de Inmunolog´ıa, Instituto de Estudios de la Inmunidad Humoral (IDEHU), CONICET-UBA, Facultad de Farmacia y Bioqu´ımica,Universidad de Buenos Aires, Jun´ın 956 4o P, 1113 Buenos Aires, Argentina
b Departamento de Microbiolog´ıa, Parasitolog´ıa e Inmunolog´ıa, Facultad de Medicina, Universidad de Buenos Aires, Buenos Aires, Argentinac Laboratorio de Virolog´ıa, Hospital de Niños Ricardo Gutiérrez, Buenos Aires, Argentina
Received 8 May 2003; accepted 3 July 2003
Abstract
The crucial role played by Ag163B6/cruzipain, the major cystein proteinase ofTrypanosoma cruzi, in the process of parasite internal-ization into mammalian cells and IgG hydrolysis, signals this antigen as a potential target for raising a protective immune response againstChagas’ disease. On the other hand, synthetic oligodeoxynucleotides containing CpG-motifs (CpG-ODN) are capable of driving immunitytoward a Th1 bias. Considering the importance of Th1 mechanisms in resistance against this intracellular parasite, we analyzed the abilityof Ag163B6/cruzipain plus CpG-ODN to induce immunoprotection against a lethal challenge with trypomastigotes. Mice were immunizedwith Ag163B6+ CpG-ODN showing high specific antibody titers, mostly IgG2a. Spleen cells from these mice strongly proliferated andpresented significant increase of IL-2 and IFN-� concentrations in their supernatant upon antigen stimulation. Trypomastigote challengerendered elevated parasitemia and mortality in all control groups, meanwhile Ag163B6+ CpG-ODN mice displayed the lowest level ofblood parasites and 100% survival to acute infection. Besides, we demonstrated that other parasite antigens introduced into mice whenchallenged, and consequently never seen before by the immune system, also elicited a Th1 immune response. Taken together, these resultsplus others provide the basis for the design of a multicomponent anti-T. cruzivaccine which may ultimately be used not only to protecthumans at risk of infection, but also may alleviate or prevent the pathogenic responses characteristic of chronic Chagas’ disease by reducingor perhaps eliminating tissue parasites from infected patients.© 2003 Elsevier Ltd. All rights reserved.
Keywords: Trypanosoma cruzi; CpG-ODN; Ag163B6/cruzipain
1. Introduction
Chagas’ disease (American trypanosomiasis), caused byTrypanosoma cruzi, is a parasitic infection transmitted by areduviid insect, that affects 16–18 million people in Centraland South America and 70% of the population are at risk ofcontracting the disease[1] Chagas’ disease is characterizedby an acute phase with high parasitemia, followed by anindeterminate stage that can last for years without signs norsymptoms. Between 20 and 30% of patients progress to achronic phase along which different types of heart pathologymay appear, such as mild arrhythmia, right or left branch
∗ Corresponding author. Tel.:+54-11-4964-8260;fax: +54-11-4964-0024.
E-mail address:[email protected] (E.L. Malchiodi).
block and severe myocardiopathies which can cause death.Disorders of the esophagus and/or colon (megaviscera) mayalso be present in other chronic patients. Chemotherapy ofChagas’ disease has limited efficacy and is not innocuous,added to the fact that neither reliable nor safe vaccine forprevention or treatment of the disease is so far available.
Whole parasites and some purified antigenic constituents,as well as recombinant proteins andT. cruzi antigen-encoding DNA, have been tested as experimental immuno-gens with variable outcome[2–4]. Using a monoclonalantibody (mAb) againstT. cruzi, we have purified by affin-ity chromatography an antigen named Ag163B6[5], whichis identical to the major cystein proteinase ofT. cruzi,cruzipain[6], also known as cruzain or GP57/51[7]. Thisenzyme is a member of the papain superfamily and hasbeen reported by Bontempi and Cazzulo[8] to be able to
0264-410X/$ – see front matter © 2003 Elsevier Ltd. All rights reserved.doi:10.1016/S0264-410X(03)00541-3

78 F.M. Frank et al. / Vaccine 22 (2003) 77–86
hydrolyze human IgG, leading to complete degradation ofthe Fc fragment. In the parasite, cruzipain is accumulatedin lysosomes near the flagellar pocket and considering thatis not only secreted but it is also present at surface level,could be responsible for proteolysis of the IgG attached tomembrane and thus, involved in the parasite defense mech-anisms against the host immune response. Besides, it wasshown that cruzipain plays a role in the process of inter-nalization ofT. cruzi into mammalian cells[9] and specificenzyme inhibitors have been proved to interfere with cellinvasion and inhibitT. cruzi intracellular replication in vitro[10]. These characteristics signal cruzipain as a potentialtarget for the generation of an immune response able toblock the progression of the parasite inside the host.
The immunogenicity of defined protein antigens is poorunless aided by adjuvants. Oligodeoxynucleotides contain-ing CpG-motifs (CpG-ODN) act as adjuvant promoting aTh1 immune response in mice, associated with protectionagainst intracellular pathogens[11,12]. Recently, it hasbeen reported that synthetic ODNs with unmethylated se-quences induce: murine B cells to proliferate and secreteimmunoglobulin in vitro and in vivo[13]; secretion of Th1cytokines as IL-12, IL-18 and IFN-� [14,15]; expression ofco-stimulatory molecules and a general increase in antigenpresenting function, markedly in dendritic cells[16]. Sev-eral antigen-CpG-ODN candidates are under pre-clinicaland clinical evaluation against a broad range of infectiousdiseases and tumors (reviewed in[17]).
Particularly, CpG-ODNs combined with vaccine can-didates or alone were able to induce a protective and/orcurative response in parasitoses caused by intracellularprotozoa. Thus, freeze-thawedLeishmania majoror a com-plex Leishmaniaantigen co-administered with CpG-ODNs,provided protection against a lethal challenge of promastig-otes[18,19]; a DNA vaccine or recombinant protein com-bined with CpG-ODNs-induced protection against a lethalchallenge withPlasmodium falciparum[20] and Gramzin-ski et al. [21] showed that CpG-ODNs administered inthe absence of antigen, 1–2 days before challenge withPlasmodium yoelii, conferred sterile protection against in-fection. Furthermore, we recently have shown that wholehomogenate ofT. cruziepimastigotes co-administered withCpG-ODN-induced protection to a challenge with lethaldoses of trypomastigotes[22].
Different mechanisms have been proposed to explain theimmunopathology of the chronic stage of Chagas’ disease,e.g.T. cruzi shares antigens with the host and the effectorelements elicited as a response to infection recognize ownantigens (molecular mimicry[23,24]), whereas serum an-tibodies against homologous antigens betweenT. cruziandmammals have been described, such as some ribosomalproteins[25], laminin [26], as well as muscle antigens[27]and some antigens of the nervous system[28,29]. Theseand other reports[30] suggesting an autoimmune etiologyfor Chagas’ disease prompted us to investigate whethera purified antigen instead of a complex protein mixture
would confer protection against infection. Accordingly, weanalyzed the ability of Ag163B6/cruzipain co-administeredwith ODN-CpG to induce protection in a murine model ofacuteT. cruzi infection.
2. Material and methods
2.1. Parasites
T. cruziepimastigotes, República Argentina (RA) strain,were grown in biphasic medium as previously described[31]. Parasites were harvested during exponential growthphase by centrifugation at 5000× g for 15 min and washedthree times with 0.1 M phosphate buffered saline (PBS) pH7.2. Bloodstream trypomastigotes ofT. cruziRA strain wereisolated from acutely infected mice at the peak of para-sitemia.
2.2. Purification of Ag163B6/cruzipain
Epimastigotes were re-suspended in 0.25 M sucrose and5 mM KCl containing protease inhibitors (2�M PMSF,5�M leupeptin, 5�M pepstatin and 5�M E-64; Sigma,St. Louis, MO) and broken by three cycles of freezing andthawing. After centrifugation at 105,000×g the obtained su-pernatant, called F105, was conserved at−20◦C as a sourceof antigens. Purified monoclonal antibody 163B6 was fixedto Sepharose 4B-CNBr[32] and used as immunosorbentfor F105. Unbound fraction was used as source of antigensdepleted of Ag163B6. After extensive washing of the im-munosorbent with PBS, the retained Ag163B6 was elutedwith 0.1 M glycine–HCl buffer pH 3.0, dialyzed againstPBS and stored at−70◦C until use. Purity was >95%, asassessed by SDS-PAGE. Before inoculation into mice, orfor in vitro stimulations, Ag163B6 and F105 were filteredthroughout a 0.2�m-pore membrane.
2.3. Oligodeoxynucleotides
The sequence for the CpG-ODN 1826 was 5′-TCCATG ACG TTC CTG ACG TT-3′, with the sequence CpGunderlined for clarity, and the sequence for non CpG-ODN1745 was 5′-TCC ATG AGC TTC CTG AGC CT-3′. AllODNs were synthesized under GMP conditions, and witha nuclease-resistant phosphorothioate backbone, by OligosEtc. (Wilsonville, OR). Endotoxin levels in the ODNs wereundetectable (<1.0 EU/mg), as determined using aLimu-lus amebocytes lysate analysis kit (Whittaker Bioproducts,Walkersville, MD).
2.4. Immunizations and challenge
All studies were carried out with inbred female 6–8-weekold C3H/HeN mice from University of Buenos Aires,

F.M. Frank et al. / Vaccine 22 (2003) 77–86 79
Argentina, with 10 mice in each experimental group, andmaintained under standard conditions. Mice were immu-nized and boosted 1 week later with 10�g of Ag163B6with added 100�g of the indicated ODN or alum (Al2O3,Alhydrogel, Superfos Biosector, Vedbaek, Denmark) bybilateral injection in the quadriceps muscles. Additionalcontrol groups were injected similarly with PBS or 100�gof CpG-ODN 1826 alone. Blood was collected from thetail vein on days 0, 14 and 28 post-immunization and thesera analyzed for the presence of specific antibodies. Twoweeks after the last immunization, half of the mice in eachgroup were killed by cervical dislocation and their spleenswere removed aseptically. The remaining animals werechallenged i.p. 4 weeks after the last immunization with500 bloodstream trypomastigotes of RA strain. Parasitemiawas monitored by counting every 2 or 3 days, the numberof trypomastigotes per 5�l of fresh blood and mortalitywas recorded daily. Surviving animals were killed 100 dayslater by cervical dislocation.
2.5. Antibody titers and IgG isotype determination
An indirect ELISA for antibody detection was used asdescribed by Voller et al.[33]. Plates (Nunc, Roskilde, Den-mark) were sensitized with 1�g per well of the soluble frac-tion F105, or with 0.1�g per well of the Ag163B6/cruzipain.Peroxidase-conjugated goat immunoglobulins to mouseIgG (Sigma), or biotinylated rat monoclonal antibodiesto mouse IgG1 or IgG2a subclasses (Pharmingen, SanDiego, CA) were used as secondary antibody, followed bystreptavidin-peroxidase. Plates were developed by addingOPD/H2O2, dark incubated for 20 min and the reactionwas stopped using 4N H2SO4. Optical density was readby an ELISA reader (Bio-Rad Laboratories, Hercules, CA)at 490 nm. End point titers were defined as the dilutionthat resulted in OD value greater than 0.1. Negative con-trol serum samples included in the assay had values lowerthan 0.1.
2.6. SDS-PAGE and immunoblotting
Epimastigotes containing protease inhibitors were dilutedin sample buffer for SDS-PAGE and electrophoretically sep-arated according to Laemmli[34] on 3% stacking/7.5% re-solving gel. Samples were electrophoresed at 200 V during1 h using a Mini-Protean II apparatus (Bio-Rad, Richmond,CA) and blotted onto nitrocellulose at 300 mA for 1.5 h in aTrans Blot apparatus (Bio-Rad)[35]. After blocking in Trisbuffer solution (TBS: 50 mM Tris–HCl, 150 mM NaCl, pH7.4) with 1% skim milk, nitrocellulose was cut into 4 mmstrips and incubated for 1 h at 37◦C with 1/100 dilutionof mice sera. After three washes with TBS, strips were in-cubated with peroxidase-conjugated goat immunoglobulinsto mouse total immunoglobulins or biotinylated rat mono-clonal antibodies to mouse IgG1 or IgG2a subclasses anddeveloped with H2O2/4-Cl-1-naphtol.
2.7. Proliferation assay and cytokine quantification
Spleens from immunized, as well as immunized and in-fected surviving mice, were aseptically removed 2 weeksafter the boost immunization and cell suspensions were pre-pared. Erythrocytes were lysed by 2 min incubation in lysisbuffer (0.15 M NH4Cl, 1.0 M KHCO3, 0.1 mM EDTA, pH7.2). Cells were washed twice and re-suspended in completeRPMI medium containing 10% foetal bovine serum (Gibco,Gaithersburg, MD), 100 U/ml of penicillin and 100�g/mlstreptomycin. Cells were cultured by triplicate in the pres-ence of 10�g/ml Ag163B6, 10�g/ml F105, 5�g/ml Con-canavalin A (Con A) or medium alone. For proliferationassays 2× 105 cells were cultured in 96-well-round bottomplates (Sarstedt Inc., Newton, NC). For the last 18 h, cellswere pulsed with 1�Ci per well (3H) thymidine (Amer-sham, Little Chalfont, England). Cells stimulated with ConA and control with RPMI alone were harvested at 24 h. Cellsstimulated with Ag163B6 or F105 and the correspondingcontrols were harvested at 5 days. Protein concentration offraction F105 and Ag163B6 used for stimulation of spleencells were 10�g/ml, as determined by Lowry. Counts perminute (CPM) of triplicate cultures were measured by anautomated liquid scintillation counter (Packard, DownersGrove, IL). The stimulation index (SI) was calculated us-ing the following formula: SI = CPM stimulated/CPMnon-stimulated. For cytokine analyses, spleen cells (4×106)from each mouse or pooled lymph node cells (1×106) werecultured by triplicate in 24-wells plates (Sarstedt, Newton,USA). Supernatants were collected after 48 h and stored at−70◦C. IL-2, IL-4, IL-10, and IFN-� measurements weredone by sandwich ELISA (R&D Systems, Minneapolis,MN) according to manufacturer’s instructions. Suppliedstandards were used to generate a standard curve.
2.8. Statistical analysis
Statistical analyses were carried out with the Prisma3.0 Software (GraphPad, San Diego, CA) using one-wayanalyses of variance (ANOVA) for proliferation, cytokineand antibody assays, Mann–Whitney non-parametric testfor parasitemia data and Log rank test for survival curves.Values ofP < 0.05 were considered significant.
3. Results
3.1. Immunization with Ag163B6 plus CpG-ODN 1826elicited a vigorous humoral response
Mice immunized with Ag163B6/cruzipain combined withCpG-ODN 1826 presented ELISA antibody titers (6389±2173; mean± S.D.) between 12.5- and 48-fold higher (P <
0.001) than those observed in mice immunized with Ag+nonCpG-ODN 1745 (258± 80), and between 1- and 3.8-foldhigher (P < 0.05) than those immunized with Ag+ alum
80 F.M. Frank et al. / Vaccine 22 (2003) 77–86
PBS alum 1826 1745 1826
2
3
4
5
Ag163B6 - + + + -
Sp
eci
ficto
tal I
gG
tite
r (L
og
)
PBS alum 1826 1745 1826
2
3
4
5Ig G1
Ig G2a
Ag163B6 - + + + -
***
***
Spe
cific
IgG
tite
r (L
og)
(A)
(B)
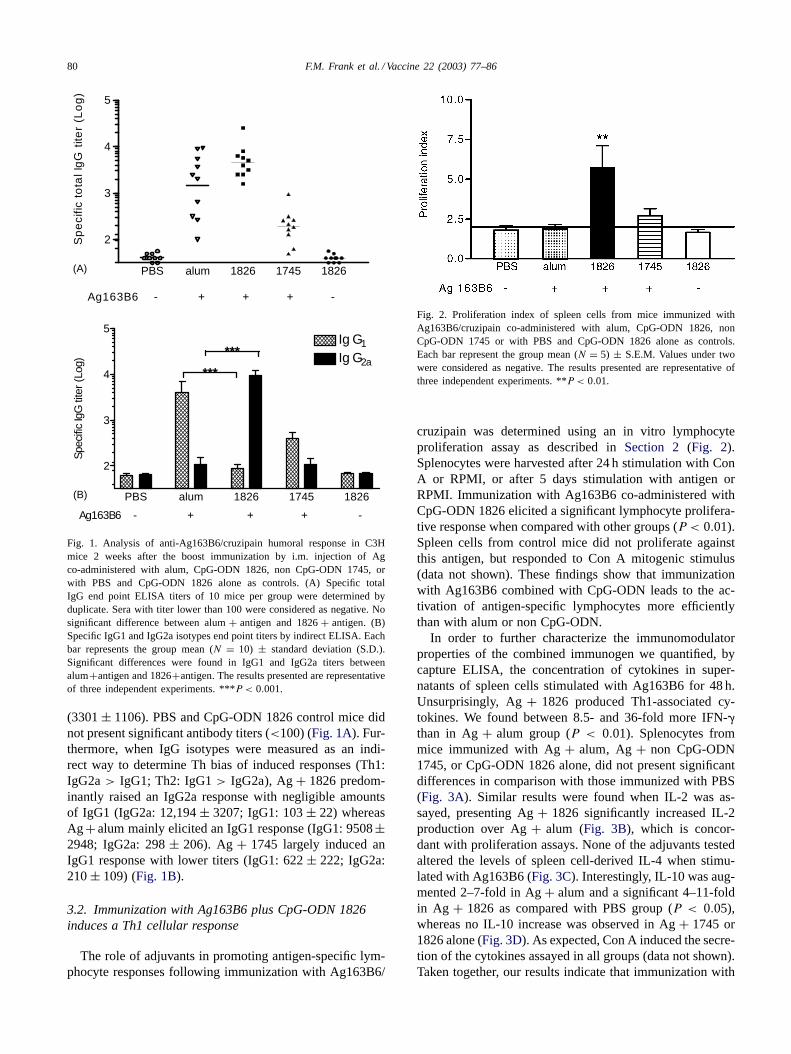
Fig. 1. Analysis of anti-Ag163B6/cruzipain humoral response in C3Hmice 2 weeks after the boost immunization by i.m. injection of Agco-administered with alum, CpG-ODN 1826, non CpG-ODN 1745, orwith PBS and CpG-ODN 1826 alone as controls. (A) Specific totalIgG end point ELISA titers of 10 mice per group were determined byduplicate. Sera with titer lower than 100 were considered as negative. Nosignificant difference between alum+ antigen and 1826+ antigen. (B)Specific IgG1 and IgG2a isotypes end point titers by indirect ELISA. Eachbar represents the group mean (N = 10) ± standard deviation (S.D.).Significant differences were found in IgG1 and IgG2a titers betweenalum+antigen and 1826+antigen. The results presented are representativeof three independent experiments. ***P < 0.001.
(3301± 1106). PBS and CpG-ODN 1826 control mice didnot present significant antibody titers (<100) (Fig. 1A). Fur-thermore, when IgG isotypes were measured as an indi-rect way to determine Th bias of induced responses (Th1:IgG2a> IgG1; Th2: IgG1> IgG2a), Ag+ 1826 predom-inantly raised an IgG2a response with negligible amountsof IgG1 (IgG2a: 12,194± 3207; IgG1: 103± 22) whereasAg + alum mainly elicited an IgG1 response (IgG1: 9508±2948; IgG2a: 298± 206). Ag+ 1745 largely induced anIgG1 response with lower titers (IgG1: 622± 222; IgG2a:210± 109) (Fig. 1B).
3.2. Immunization with Ag163B6 plus CpG-ODN 1826induces a Th1 cellular response
The role of adjuvants in promoting antigen-specific lym-phocyte responses following immunization with Ag163B6/
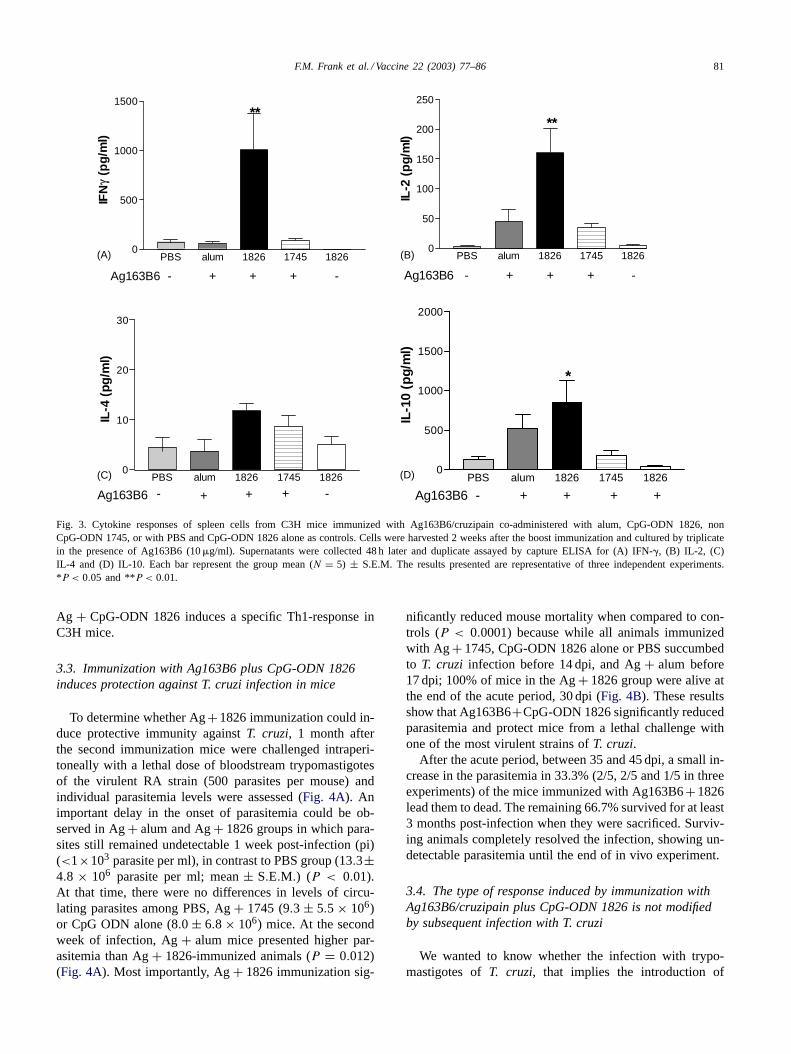
Fig. 2. Proliferation index of spleen cells from mice immunized withAg163B6/cruzipain co-administered with alum, CpG-ODN 1826, nonCpG-ODN 1745 or with PBS and CpG-ODN 1826 alone as controls.Each bar represent the group mean (N = 5) ± S.E.M. Values under twowere considered as negative. The results presented are representative ofthree independent experiments. **P < 0.01.
cruzipain was determined using an in vitro lymphocyteproliferation assay as described inSection 2 (Fig. 2).Splenocytes were harvested after 24 h stimulation with ConA or RPMI, or after 5 days stimulation with antigen orRPMI. Immunization with Ag163B6 co-administered withCpG-ODN 1826 elicited a significant lymphocyte prolifera-tive response when compared with other groups (P < 0.01).Spleen cells from control mice did not proliferate againstthis antigen, but responded to Con A mitogenic stimulus(data not shown). These findings show that immunizationwith Ag163B6 combined with CpG-ODN leads to the ac-tivation of antigen-specific lymphocytes more efficientlythan with alum or non CpG-ODN.
In order to further characterize the immunomodulatorproperties of the combined immunogen we quantified, bycapture ELISA, the concentration of cytokines in super-natants of spleen cells stimulated with Ag163B6 for 48 h.Unsurprisingly, Ag+ 1826 produced Th1-associated cy-tokines. We found between 8.5- and 36-fold more IFN-�than in Ag+ alum group (P < 0.01). Splenocytes frommice immunized with Ag+ alum, Ag+ non CpG-ODN1745, or CpG-ODN 1826 alone, did not present significantdifferences in comparison with those immunized with PBS(Fig. 3A). Similar results were found when IL-2 was as-sayed, presenting Ag+ 1826 significantly increased IL-2production over Ag+ alum (Fig. 3B), which is concor-dant with proliferation assays. None of the adjuvants testedaltered the levels of spleen cell-derived IL-4 when stimu-lated with Ag163B6 (Fig. 3C). Interestingly, IL-10 was aug-mented 2–7-fold in Ag+ alum and a significant 4–11-foldin Ag + 1826 as compared with PBS group (P < 0.05),whereas no IL-10 increase was observed in Ag+ 1745 or1826 alone (Fig. 3D). As expected, Con A induced the secre-tion of the cytokines assayed in all groups (data not shown).Taken together, our results indicate that immunization with
F.M. Frank et al. / Vaccine 22 (2003) 77–86 81
PBS alum 1826 1745 18260
500
1000
1500**
Ag163B6 - + + + -
IFN
(p
g/m
l)
PBS alum 1826 1745 18260
50
100
150
200
250
**
Ag163B6 - + + + -
IL-2
(p
g/m
l)
PBS alum 1826 1745 18260
10
20
30
- + + + -Ag163B6
IL-4
(pg
/ml)
PBS alum 1826 1745 18260
500
1000
1500
2000
*
Ag163B6 - + + + +
IL-1
0 (p
g/m
l)
(A) (B)
(C) (D)
Fig. 3. Cytokine responses of spleen cells from C3H mice immunized with Ag163B6/cruzipain co-administered with alum, CpG-ODN 1826, nonCpG-ODN 1745, or with PBS and CpG-ODN 1826 alone as controls. Cells were harvested 2 weeks after the boost immunization and cultured by triplicatein the presence of Ag163B6 (10�g/ml). Supernatants were collected 48 h later and duplicate assayed by capture ELISA for (A) IFN-�, (B) IL-2, (C)IL-4 and (D) IL-10. Each bar represent the group mean (N = 5) ± S.E.M. The results presented are representative of three independent experiments.*P < 0.05 and **P < 0.01.
Ag + CpG-ODN 1826 induces a specific Th1-response inC3H mice.
3.3. Immunization with Ag163B6 plus CpG-ODN 1826induces protection against T. cruzi infection in mice
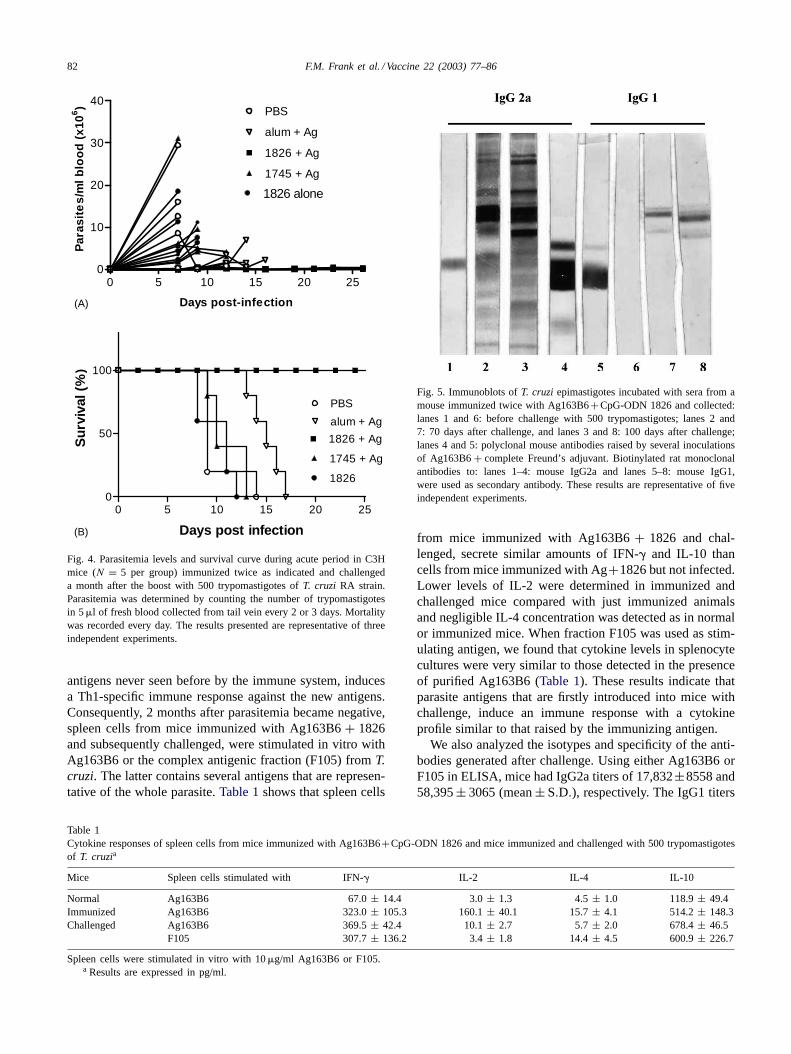
To determine whether Ag+1826 immunization could in-duce protective immunity againstT. cruzi, 1 month afterthe second immunization mice were challenged intraperi-toneally with a lethal dose of bloodstream trypomastigotesof the virulent RA strain (500 parasites per mouse) andindividual parasitemia levels were assessed (Fig. 4A). Animportant delay in the onset of parasitemia could be ob-served in Ag+ alum and Ag+ 1826 groups in which para-sites still remained undetectable 1 week post-infection (pi)(<1×103 parasite per ml), in contrast to PBS group (13.3±4.8 × 106 parasite per ml; mean± S.E.M.) (P < 0.01).At that time, there were no differences in levels of circu-lating parasites among PBS, Ag+ 1745 (9.3 ± 5.5 × 106)or CpG ODN alone (8.0 ± 6.8 × 106) mice. At the secondweek of infection, Ag+ alum mice presented higher par-asitemia than Ag+ 1826-immunized animals (P = 0.012)(Fig. 4A). Most importantly, Ag+ 1826 immunization sig-
nificantly reduced mouse mortality when compared to con-trols (P < 0.0001) because while all animals immunizedwith Ag + 1745, CpG-ODN 1826 alone or PBS succumbedto T. cruzi infection before 14 dpi, and Ag+ alum before17 dpi; 100% of mice in the Ag+ 1826 group were alive atthe end of the acute period, 30 dpi (Fig. 4B). These resultsshow that Ag163B6+CpG-ODN 1826 significantly reducedparasitemia and protect mice from a lethal challenge withone of the most virulent strains ofT. cruzi.
After the acute period, between 35 and 45 dpi, a small in-crease in the parasitemia in 33.3% (2/5, 2/5 and 1/5 in threeexperiments) of the mice immunized with Ag163B6+1826lead them to dead. The remaining 66.7% survived for at least3 months post-infection when they were sacrificed. Surviv-ing animals completely resolved the infection, showing un-detectable parasitemia until the end of in vivo experiment.
3.4. The type of response induced by immunization withAg163B6/cruzipain plus CpG-ODN 1826 is not modifiedby subsequent infection with T. cruzi
We wanted to know whether the infection with trypo-mastigotes ofT. cruzi, that implies the introduction of
82 F.M. Frank et al. / Vaccine 22 (2003) 77–86
0 5 10 15 20 250
10
20
30
40PBS
alum + Ag
1826 + Ag
1745 + Ag
1826 alone
Days post-infection
Par
asit
es/
ml b
loo
d (
x106
)
0 5 10 15 20 250
50
100
PBS
1745 + Ag
alum + Ag
1826
1826 + Ag
Days post infection
Sur
viva
l (%
)
(A)
(B)
Fig. 4. Parasitemia levels and survival curve during acute period in C3Hmice (N = 5 per group) immunized twice as indicated and challengeda month after the boost with 500 trypomastigotes ofT. cruzi RA strain.Parasitemia was determined by counting the number of trypomastigotesin 5�l of fresh blood collected from tail vein every 2 or 3 days. Mortalitywas recorded every day. The results presented are representative of threeindependent experiments.
antigens never seen before by the immune system, inducesa Th1-specific immune response against the new antigens.Consequently, 2 months after parasitemia became negative,spleen cells from mice immunized with Ag163B6+ 1826and subsequently challenged, were stimulated in vitro withAg163B6 or the complex antigenic fraction (F105) fromT.cruzi. The latter contains several antigens that are represen-tative of the whole parasite.Table 1shows that spleen cells
Table 1Cytokine responses of spleen cells from mice immunized with Ag163B6+CpG-ODN 1826 and mice immunized and challenged with 500 trypomastigotesof T. cruzia
Mice Spleen cells stimulated with IFN-� IL-2 IL-4 IL-10
Normal Ag163B6 67.0± 14.4 3.0± 1.3 4.5± 1.0 118.9± 49.4Immunized Ag163B6 323.0± 105.3 160.1± 40.1 15.7± 4.1 514.2± 148.3Challenged Ag163B6 369.5± 42.4 10.1± 2.7 5.7± 2.0 678.4± 46.5
F105 307.7± 136.2 3.4± 1.8 14.4± 4.5 600.9± 226.7
Spleen cells were stimulated in vitro with 10�g/ml Ag163B6 or F105.a Results are expressed in pg/ml.
Fig. 5. Immunoblots ofT. cruziepimastigotes incubated with sera from amouse immunized twice with Ag163B6+CpG-ODN 1826 and collected:lanes 1 and 6: before challenge with 500 trypomastigotes; lanes 2 and7: 70 days after challenge, and lanes 3 and 8: 100 days after challenge;lanes 4 and 5: polyclonal mouse antibodies raised by several inoculationsof Ag163B6+ complete Freund’s adjuvant. Biotinylated rat monoclonalantibodies to: lanes 1–4: mouse IgG2a and lanes 5–8: mouse IgG1,were used as secondary antibody. These results are representative of fiveindependent experiments.
from mice immunized with Ag163B6+ 1826 and chal-lenged, secrete similar amounts of IFN-� and IL-10 thancells from mice immunized with Ag+1826 but not infected.Lower levels of IL-2 were determined in immunized andchallenged mice compared with just immunized animalsand negligible IL-4 concentration was detected as in normalor immunized mice. When fraction F105 was used as stim-ulating antigen, we found that cytokine levels in splenocytecultures were very similar to those detected in the presenceof purified Ag163B6 (Table 1). These results indicate thatparasite antigens that are firstly introduced into mice withchallenge, induce an immune response with a cytokineprofile similar to that raised by the immunizing antigen.
We also analyzed the isotypes and specificity of the anti-bodies generated after challenge. Using either Ag163B6 orF105 in ELISA, mice had IgG2a titers of 17,832±8558 and58,395± 3065 (mean± S.D.), respectively. The IgG1 titers

F.M. Frank et al. / Vaccine 22 (2003) 77–86 83
were much lower than those, 1682±1229 and 1835±1534,respectively. In order to identify the antigens able to in-duce these antibody responses, we used fraction F105 inimmunoblotting assays. InFig. 5, it is shown that miceimmunized with Ag163B6+ CpG-ODN 1826, but not in-fected, produced IgG2a antibodies only against Ag163B6.Seventy and 100 days after challenge, mice also secretedIgG2a antibodies with specificity for several other antigenspresent in F105 and a very low amount of IgG1 againstfewer antigens. Notably, the IgG1-specific antibodies wereincreasing from the day of infection throughout day 100 pi.The production of specific IgG2a antibodies against otherantigens than Ag163B6, confirms that the induced Th1bias of the immune response remains after infection. An-tibody reactivity in sera collected post-immunization withAg163B6 indirectly reflects the purity of the immunogen,as this is the only component from a complex mixture ofT.cruzi antigens recognized by immunoblotting (Fig. 1, lane1). Pre-immune sera were used as negative control.
4. Discussion
The enzymatic properties of cruzipain, the major cys-tein proteinase ofT. cruzi, were firstly studied by Cazzuloet al. [36,37] and Murta et al.[7]. Its usefulness as anti-gen in serological test was determined by Martinez et al.[38] and Malchiodi et al.[39], who showed that most serafrom chronic Chagasic patients have specific antibodiesagainst Ag163B6/cruzipain. Later on, it was demonstratedthat Ag163B6 would be useful for serological differentialdiagnosis of Chagas’ disease and leishmaniasis[40,41].Moreover, these studies using Ag163B6 led to the firstdemonstration of human concomitant infection with bothparasites,T. cruziandLeishmaniaspp.[42].
The demonstrated immunogenic properties of Ag163B6/cruzipain, its enzymatic ability to effectively cleave im-munoglobulins and the strong suggestions of its participa-tion in parasite penetration into mammalian cells, promptedus to investigate whether this antigen would be able to in-duce protection againstT. cruziinfection. Earlier studies us-ing Ag163B6 plus complete Freund’s adjuvant (CFA) werevery disappointing because no difference in parasitemia andsurvival compared with control groups was seen (unpub-lished data). Sepulveda et al.[3] found similar results us-ing recombinant complement regulatory protein (CRP) ofT.cruzi plus CFA, although immunoprotection was achievedthrough DNA-based immunization.
A successful vaccine for induction of protection againstT. cruzi will likely to elicit strong humoral and cellularimmune responses and several publications indicate thata Th1, rather than a Th2, driven immune response willbe protective[43–46]. The search for effective and safeTh1-promoting adjuvant is an active field, as most estab-lished vaccines use alum adjuvant which results in a bias toTh2 responses, characterized by IgG1 antibodies in the ab-
sence of IgG2a and lack of specific cytotoxic T lymphocytes[47–49,50,51]. The knowledge about CpG-ODN propertiesto induce a Th1 bias to the immune response and the intra-cellular replication of this parasite, supported further studieson Ag163B6/cruzipain plus CpG-ODN as immunogen forprotection againstT. cruzi infection.
In this paper, antibody titers were determined before chal-lenge in all groups finding high total specific IgG in miceimmunized with both, Ag plus alum or Ag plus CpG-ODN1826, without significant difference between them. In con-trast, the IgG isotype distribution was dependent on the adju-vant used in each group. Accordingly, Ag+alum-immunizedmice displayed high titer of specific IgG1 and a low amountof specific IgG2a whereas the inverse situation was seen inmice immunized with Ag+ CpG-ODN 1826, firmly sug-gesting a bias to a Th1 driven immune response in the lattercase[50]. A confirmation was obtained when lymphocyteproliferation and cytokines in supernatant of spleen cellsfrom immunized mice were determined. Thus, spleen cellsfrom animals receiving Ag+ CpG-ODN 1826 strongly pro-liferated and secreted both IFN-� and IL-2 when Ag163B6was added to the culture, indicating that the immunizationhad generated antigen-specific IFN-�-producing cells. Sur-prisingly, we observed a significant increase in the releaseof IL-10, which is a LTh2 cytokine, in Ag+ CpG-ODN1826 compared with PBS group. Other authors communi-cated that CpG-ODN can induce secretion of this cytokineaccompanying others that characterize Th1 bias[52,53]. Al-though the mechanism is not very well understood, it hasbeen proposed that IL-10, probably B cell- or regulatory Tcell-derived, may play a role in vivo to keep under controltoxic shock caused by a self-amplifying loop of secretion ofIL-12, TNF-�, NO and IFN-� [54].
In order to analyze whether Ag+CpG-ODN 1826 wouldconfer protection, we decided to use a murine model ofacute infection with 500 trypomastigotes of one of the mostlethal strains ofT. cruzi. This amount of RA strain para-sites is able to kill all challenged C3H mice in less than 14days. To understand the virulence of RA strain it must betaken into account that other authors challenged mice with103–5×105 trypomastigotes, Brazil or Tulahuen strain, andthe mice began to die 20–30 dpi[55,4] whereas Sepulvedaet al. [3] needed 2× 106 Y-strain trypomastigotes, intra-venously administered, to observe lethal effect at 14 dpi. Inour experiment, Ag163B6+ CpG-ODN 1826-immunizedmice not only controlled parasitemia but also survived acutephase. Parasitemia in surviving mice became negative onday 48 pi, remaining undetectable until they were sacrificedon day 100 pi.
One of the pending questions about the use of CpG-ODNas adjuvant is whether any new antigen will render aTh1-driven immune response. To analyze whether the par-asite could be able to revert the Th1 bias raised by im-munization and what kind of response could be inducedagainst these new antigens, we determined specific IgGantibody titers using a complex antigenic fraction (F105),

84 F.M. Frank et al. / Vaccine 22 (2003) 77–86
representative of the whole parasite. At 70 and 100 dpi,we found a high titer of specific IgG2a against antigensincluded in fraction F105. Immunoblotting analysis clearlydemonstrated that these IgG2a antibodies recognizedT.cruzi antigens other than Ag163B6. In addition to this, bothF105- as well as Ag163B6-stimulated splenocytes showedenhanced IFN-� production, strongly suggesting the in-duction of a Th1-biased response to infection. The use ofF105 or Ag163B6 as stimulating antigen did not induce asignificant increase of IL-2, probably because T cells frominfected mice have reduced capacity to produce IL-2 anddisplay IL-2R[2]. To discard that the observed stimulationwhere due to the presence of Ag163B6 in F105 fraction,in spite is <0.5% of antigen content, we used the F105fraction after depletion with monoclonal antibody 163B6obtaining similar results (data not shown). Further studieswill be necessary to elucidate whether the Th1 bias-inducedagainst the new antigens, is dependent or not on the pres-ence of Ag163B6 in the infecting parasite. The possibilitythat Ag163B6+ CpG-ODN immunization may precludethe natural development of Th2 responses, beneficial forresistance to some infections, should be ruled out beforebeing evaluated in vaccine trials.
Thus far, the exploration of vaccines againstT. cruzihasbeen widely avoided due to the fear that such interventionmethods would exacerbate rather than prevent a disease thatmany still consider to have an autoimmune etiology[29].However, a growing body of evidence indicates that it is thepersistence ofT. cruzi in the diseased tissues and not theparasite-induced immune response to self molecules whichcorrelates best with the induction and maintenance of the in-flammatory disease process[56–61]. In that regard, publica-tions from Gea’s laboratory[62,63] reported that cruzipaincan induce cardiac conduction abnormalities, autoimmuneresponse and skeletal muscle damage in mice but later theyfound that the pathogenic immune response against cruzi-pain would depends on murine genetic background[64].Besides, in those experiments mice were immunized withCFA, which is not used in humans and it has been recentlydemonstrated to induce important damage to muscle[65].Moreover, we were not able to identify any abnormality inhearts, by microscopical examination of stained tissue sec-tions, from Ag163B6-immunized mice, either adjuvantedby alum or CpG-ODN (data not shown). Additional studiesshould be required to clarify whether the immune responseagainst own antigens induced with cruzipain mainly dependson the antigen itself, the associated adjuvant or the host’sgenetic background.
In the present work, we tested two adjuvants that couldbe used in humans to improve the immune response againsta single antigen. Alum is widespread licensed for humanvaccines whereas there are extensive data generated frompre-clinical trials with antisense oligonucleotides containingCpG motifs for viral infections and cancers[17] adminis-tered to humans even at doses up to 20 mg/kg without sig-nificant specific toxicity. Considering that CpG DNA elicits
adjuvant ability at�g/kg doses, it is less likely to provokeadverse effects[66].
Perhaps the most significant finding of our studies is thata protective immune response toT. cruzican be elicited byimmunization with a natural, parasite-derived antigen usingCpG-ODN as adjuvant. Meanwhile this manuscript was onpreparation, Schnapp et al.[55] communicated that a recom-binant cruzipain, plus IL-12 and a neutralizing IL-4 mAbs,induces protection againstT. cruzi. Taken together, these re-sults are important because Ag163B6/cruzipain is not onlyone of the major antigens but is expressed in all strains ofT.cruzi studied so far[67,32], indicating that it could protectagainst infection from different endemic regions. However,it is significant that, despite the control of parasitemia andincrease of survival achieved, Ag163B6+CpG-ODN failedto prevent recipient mice from getting infected: they devel-oped acute phase-limited parasitemias, albeit at lower levelthan controls, and survival after acute period was limited to67% of vaccinated animals. Protection from disease mightnot require the perhaps unachievable goal of sterilizing im-munity, but may only have to provide efficient control of theparasite load at a sub-pathogenic level[60].
The ability of Ag163B6/cruzipain+CpG-ODN to inducesignificant murine protective immunity toT. cruziprovidesthe basis for the design of multicomponent, including nat-ural, recombinant or/and DNA-based anti-T. cruzi vaccinewhich may ultimately be used not only to protect humansat risk of infection, but also may alleviate or prevent thepathogenic responses characteristic of chronic Chagas’ dis-ease by reducing or perhaps eliminating tissue parasites frominfected patients.
Acknowledgements
The authors are grateful to Dr. Mónica G. Chiaramontefor first ideas on this project. We also thank Dr. Mónica Es-teva, Instituto Fatala Chaben, Buenos Aires, Argentina, forproviding F105 and Lucas Bukata for helping in mice infec-tion. Financial support was received from: Universidad deBuenos Aires (Grant B802); Consejo Nacional de Investiga-ciones Cient́ıficas y Técnicas (CONICET); Swedish Inter-national Development Agency (SIDA) for the Network forResearch and Training in Parasitic Diseases in the South-ern Cone of Latin America (RTPD Network) and AgenciaNacional de Investigaciones Cientı́ficas y Técnicas (PICT4415), Argentina.
References
[1] Tropical disease research. In: Twelfth Programme Report of theUNDP/World Bank/WHO Special Programme for Research andTraining in Tropical Disease (TDR). Geneva, Switzerland: WorldHealth Organization; 1999.
[2] Rottemberg ME, Lindqvist C, Koman A, Segura EL, OrnA. Modulation of both interleukin-2 receptor expression and

F.M. Frank et al. / Vaccine 22 (2003) 77–86 85
interleukin-2 production during experimental murineTrypanosomacruzi infection. Scand J Immunol 1989;30:65–72.
[3] Sepulveda P, Hontebeyrie M, Liegeard P, Mascilli A, Norris KA.DNA-based immunization withTrypanosoma cruzicomplementregulatory protein elicits complement lytic antibodies and confersprotection againstTrypanosoma cruziinfection. Infect Immun2000;68:4986–91.
[4] Wizel B, Garg N, Tarleton RL. Vaccination with trypomastigotesurface antigen 1-encoding plasmid DNA confers protection againstlethal Trypanosoma cruziinfection. Infect Immun 1998;66:5073–81.
[5] Carbonetto CH, Malchiodi EL, Chiaramonte MG, Durante de IsolaEL, Fossati CA, Margni RA. Isolation of aTrypanosoma cruziantigenby affinity chromatography with a monoclonal antibody. Preliminaryevaluation of its possible applications in serological tests. Clin ExpImmunol 1990;82:93–6.
[6] Malchiodi EL, Chiaramonte MG, Martinez JA, Zwirner NW, MargniRA, Cazzulo JJ. Identity of the major cysteine proteinase (cruzipain)from Trypanosoma cruziand an antigen (Ag163B6) isolated with amonoclonal antibody. Immunol Lett 1993;35:59–62.
[7] Murta AC, Persechini PM, de Padrón ST, de Souza W, GuimaraesJA, Scharfstein J. Structural and functional identification of GP57/51antigen ofTrypanosoma cruzias a cysteine proteinase. Mol BiochemParasitol 1990;43:27–38.
[8] Bontempi E, Cazzulo JJ. Digestión of human immunoglobulin Gby the major cysteine proteinase (cruzipain) fromT. cruzi. FEMSMicrobiol Lett 1999;58:337–41.
[9] Souto-Padron T, Campetella O, Cazzulo JJ, De Souza W. Cysteineproteinases in T. cruzi immunocytochemical localization andinvolvement in parasite-host cell interaction. J Cell Sci 1990;96:485–90.
[10] Meirelles MN, Juliano L, Carmona E, Silva SG, Costa EM, MurtaAC, et al. Inhibitors of the major cysteinyl proteinase (GP57/51)impair host cell invasion and arrest the intracellular development ofTrypanosoma cruziin vitro. Mol Biochem Parasitol 1992;52:175–84.
[11] Chu RS, Targoni OS, Krieg AM, Lehmann PV, Harding CV. CpGoligodeoxynucleotides act as adjuvants that switch on T helper 1(Th1) immunity. J Exp Med 1997;186:1623–31.
[12] Roman M, Martin-Orozco E, Goodman JS, et al. ImmunostimulatoryDNA sequences function as T helper1-promoting adjuvants. Nat Med1997;3:849–54.
[13] Krieg AM, Yi AK, Matson S, et al. CpG motifs in bacterial DNAtrigger direct B-cell activation. Nature 1995;374:546–9.
[14] Bohle B, Jahn-Schmid B, Maurer D, Kraft D, Ebner C.Oligodeoxynucleotides containing CpG motifs induce IL-12, IL-18and IFN-� production in cells from allergic individuals and inhibitIgE synthesis in vitro. Eur J Immunol 1999;29:2344–53.
[15] Yamamoto S, Yamamoto T, Kataoka T, Kuramoto E, Yano O,Tokunaga T. Unique palindromic sequences in synthetic oligo-nucleotides are required to induce IFN and augment IFN-mediatednatural killer activity. J Immunol 1992;148:4072–6.
[16] Chu RS, Askew D, Noss EH, Tobian A, Krieg AM, Harding CV.CpG oligodeoxinucleotides down-regulate macrophage class II MHCantigen processing. J Immunol 1999;163:1188–94.
[17] Agrawal S, Kandimalla ER. Medicinal chemistry and therapeuticpotential of CpG DNA. Trends Mol Med 2002;8:114–21.
[18] Walker PS, Scharton-Kersten T, Krieg AM. Immunostimulatoryoligodeoxinucleotides promote protective immunity and providesystemic therapy for leishmaniasis via IL-12 and IFN-�- dependentmechanisms. Proc Natl Acad Sci USA 1999;96:6970–5.
[19] Zimmermann S, Egeter O, Hausmann S, et al. Cutting edge: CpGoligodeoxinucleotides trigger protective and curative Th1 responsesin lethal murine leishmaniasis. J Immunol 1998;160:3627–30.
[20] Jones TR, Narum DL, Gozalo AS, et al. Protection ofAotusmonkeysby Plasmodium falciparumEBA-175 region II DNA prime-proteinboost immunization regimen. J Infect Dis 2001;183:303–12.
[21] Gramzinski RA, Doolan DL, Sedegah M, Davis HL, KriegAM, Hoffman SL. Interleukin-12- and gamma interferon-dependent
protection against malaria conferred by CpG oligodeoxynucleotidein mice. Infect Immun 2001;69:1643–9.
[22] Corral RS, Petray PB. CpG DNA as a Th1-promoting adjuvant inimmunization againstTrypanosoma cruzi. Vaccine 2000;19:234–42.
[23] Brener Z. Immunity toTrypanosoma cruzi. Adv Parasitol 1980;18:247–9.
[24] Hudson L. Autoinmune phenomena in chronic chagasic cardiopathy.Parasitol Today 1985;1:6–9.
[25] Skeiky YAW, Benson DR, Parsons M, Elkon KB, Reed SG.Cloning and expression ofTrypanosoma cruziribosomal protein POand epitope analysis of anti-PO autoantibodies in Chagas’ diseasepatients. J Exp Med 1992;176:201–11.
[26] Szarfman A, Terranova VP, Rennard SI, et al. Antibodies to lamininin Chagas’ disease. J Exp Med 1982;155:1161–71.
[27] Zwirner NW, Malchiodi EL, Chiaramonte MG, Fossati CA. Alytic monoclonal antibody toTrypanosoma cruzibloodstreamtrypomastigotes which recognizes an epitope expressed in tissuesaffected in Chagas’ disease. Infect Immun 1994;62:2483–9.
[28] Van Voorhis WC, Eisen H. FI-160. A surface antigen ofTrypanosoma cruzithat mimics mammalian nervous tissue. J ExpMed 1989;169:641–52.
[29] Wood JN, Hudson L, Jessell TM, Yamamoto M. A monoclonalantibody defining antigenic determinants on subpopulations ofmammalian neurones andTrypanosoma cruziparasites. Nature1982;296:34–8.
[30] Kalil J, Cunha-Neto E. Autoimmunity in Chagas’ diseasecardiomiopathy: fulfilling the criteria at last? Parasitol Today1996;12:396–9.
[31] Chiari E, Camargo EP. Culturing and cloning ofT. cruzi. In: MorelCM, editor. Genes and antigens of parasites. Rio de Janeiro: FundaçãoOswaldo Cruz; 1984. p. 23–6.
[32] Carbonetto CH, Malchiodi EL, Chiaramonte MG, Durante de IsolaEL, Fossati CA, Margni RA. Isolation of aTrypanosoma cruziantigenby affinity chromatography with a monoclonal antibody. Preliminaryevaluation of its possible applications in serological tests. Clin ExpImmunol 1990;82:93–6.
[33] Voller A, Bidwell D, Bartlett A. Enzyme-linked immunoadsorbentassay. In: Rose NR, Friedman H, editors. Manual of clinicalimmunology. Washington (DC): American Society for Microbiology;1988. p. 359–71.
[34] Laemmli UK. Cleavage of structural proteins during the assemblyof the head of bacteriophage T4. Nature 1970;227:680–5.
[35] Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteinsfrom polyacrilamide gels to nitrocellulose sheets: procedure and someapplications. Proc Natl Acad Sci USA 1979;76:4350–4.
[36] Cazzulo JJ, Couso R, Raimondi A, Wernstedt C, Hellman U.Further characterization and partial amino acid sequence of acysteine proteinase fromTrypanosoma cruzi. Mol Biochem Parasitol1989;33:33–41.
[37] Cazzulo JJ, Stoka V, Turk V. Cruzipain, the major cysteineproteinase from the protozoan parasiteTrypanosoma cruzi. BiolChem 1997;378:1–10.
[38] Martinez J, Campetella O, Frasch ACC, Cazzulo JJ. The majorcysteine proteinase (cruzipain) fromT. cruzi is antigenic in humaninfections. Infect Immun 1991;59:4275–7.
[39] Malchiodi EL, Chiaramonte MG, Taranto NJ, Zwirner NW, MargniRA. Cross reactivity studies and differential serodiagnosis ofTrypanosoma cruziand Leishmaniaspp. human infection: Use ofimmunoblotting and ELISA with a purified antigen (Ag163B6). ClinExp Immunol 1994;97:417–23.
[40] Chiaramonte MG, Zwirner NW, Caropresi SL, Taranto NJ, MalchiodiEL. Trypanosoma cruziandLeishmaniaspp. human mixed infection.Am J Trop Med Hyg 1996;54:271–3.
[41] Frank FM, Fernández MM, Taranto NJ, et al. Characterization ofhuman infection byLeishmaniaspp. in the northwest of Argentina:humoral immune response double infection withT. cruziand speciesof Leishmaniainvolved. Parasitology 2003;129:1–9.

86 F.M. Frank et al. / Vaccine 22 (2003) 77–86
[42] Chiaramonte MG, Frank FM, Furer GM, Taranto NJ, MargniRA, Malchiodi EL. Polymerase chain reaction revealsT. cruziinfection suspected by serology in leishmaniasis patients. Acta Trop1999;72:295–308.
[43] Fujimura AE, Kinoshita, Pereira-Chioccola VL, Rodrı́guez MM.DNA sequences encoding CD4+ and CD8+ T-cell epitopes areimportant for efficient protective immunity induced by DNAvaccination withTrypanosoma cruzigene. Infect Immun 2001;69:5477–86.
[44] Hoft DF, Schnapp AR, Eickhoff CS, Roodman ST. Involvement ofCD4+ Th1 cells in systemic immunity protective against primaryand secondary challenges withTrypanosoma cruzi. Infect Immun2000;68:197–204.
[45] Kumar S, Tarleton RL. Antigen-specific Th1 but not Th2 cellsprovide protection from lethalTrypanosoma cruziinfection in mice.J Immunol 2001;166:4596–603.
[46] Rodrigues MM, Riberao M, Pereira-Chioccola V, Renia L, CostaF. Predominance of CD4 Th1 and CD8 Tc1 cells revealed bycharacterization of the cellular immune response generated byimmunization with a DNA vaccine containing aTrypanosoma cruzigene. Infect Immun 1999;67:3855–63.
[47] Brewer JM, Alexander J. Cytokines and the mechanisms ofaction of vaccine adjuvants. Cytokines Cell Mol Ther 1997;3:233–46.
[48] Brewer JM, Conacher M, Hunter CA, Mohrs M, BrombacherF, Alexander J. Aluminium hydroxide adjuvants initiates strongantigen-specific Th2 responses in the absence of IL-4- orIL-13-mediated signaling. J Immunol 1999;163:6448–54.
[49] Brewer JM, Conacher M, Satoskar A, Buethmann H, AlexanderJ. In interleukin-4 deficient mice, alum not only generates Thelper-1 responses equivalent to Freund’s complete adjuvant, butcontinues to induce T helper-2 cytokine production. Eur J Immunol1996;26:2062–6.
[50] Davis HL, Weeratna R, Waldschmidt TJ, Tygrett L, Schorr J, KriegAM. CpG DNA is a potent enhancer of specific immunity in miceimmunized with recombinant hepatitis B surface antigen. J Immunol1998;160:870–6.
[51] Grun JL, Maurer PH. Different T helper cell subsets elicitedin mice utilizing two different adjuvant vehicles: the role ofendogenous IL-1 in proliferative responses. Cell Immunol 1989;121:134–45.
[52] Anitescu M, Chace JH, Tuetken R, et al. Interleukin-10 functionsin vitro and in vivo to inhibit bacterial DNA-induced secretion ofinteleukin-12. J Interferon Cytokine Res 1997;17:781–8.
[53] Redford TW, Yi AK, Ward CT, Krieg AM. Cyclosporine Aenhances IL-12 production by CpG in bacterial DNA and syntheticoligodeoxinucleotides. J Immunol 1998;161:3930–5.
[54] Stacey KJ, Sester DP, Sweet MJ, Hume DA. Macrophage activationby immunostimulatory DNA. In: Wagner H, editor. Immunobiologyof bacterial CpG-DNA. Berlin: Springer; 2000. p. 41–58.
[55] Schnapp AR, Eickhoff CS, Sizemore D, Curtiss III R, Hoft DF.Cruzipain induces both mucosal and systemic protection againstTrypanosoma cruziin mice. Infect Immun 2002;70:5065–74.
[56] Bellotti G, Bocchi EA, de Moraes AV, et al. In vivo detectionof Trypanosoma cruziantigens in hearts of patients with chronicChagas’ hearth disease. Am Heart J 1996;131:301–7.
[57] Brandariz S, Schijman A, Vigliano C, et al. Detection of parasiteDNA in Chagas’ heart disease. Lancet 1997;346:1370–1.
[58] Jones EM, Colley DG, Tostes S, Lopes ER, Vnencak-Jones CL,McCurley TL. Amplification of aTrypanosoma cruziDNA sequencefrom inflammatory lesions in human chagasic cardiomyopathy. AmJ Trop Med Hyg 1993;48:348–57.
[59] Lane JE, Olivares-Villagomez D, Vnencak-Jones CL, McCurley TL,Carter CE. Detection ofTrypanosoma cruziwith the polymerasechain reaction and in situ hybridization in infected murine cardiactissue. Am J Trop Med Hyg 1997;56:588–95.
[60] Tarleton RL, Zhang L. Chagas disease etiology: autoimmunity orparasite persistence? Parasitol Today 1999;15:94–9.
[61] Tarleton RL, Zhang L, Downs MO. “Autoimmune rejection” ofneonatal heart transplants in experimental Chagas’ disease is aparasite-specific response to infected host tissue. Proc Natl Acad SciUSA 1997;94:3932–7.
[62] Giordanengo L, Fretes R, Dı́az H, et al. Cruzipain inducesautoimmune response against skeletal muscle and tissue damage inmice. Muscle Nerve 2000;23:1407–13.
[63] Giordanengo L, Maldonado C, Rivarola HW, et al. Induction ofantibodies reactive to cardiac myosin and development of heartalterations in cruzipain-immunized mice and their offspring. Eur JImmunol 2000;30:3181–9.
[64] Guiñazú AL, Aoki P, Giordanengo L, et al. The genetic profile isa key factor in the outcome of the cruzipain pathogenic immuneresponse, immunodominant antigen ofTrypanosoma cruzi. Medicina(B Aires) 2002;62(5):496.
[65] Weeratna RD, McCluskie MJ, Xu J, Davis HL. CpG DNA inducesstronger immune responses with less toxicity than other adjuvants.Vaccine 2000;18:1755–62.
[66] Klinman DM, Kamstrup S, Verthelyi D, et al. Activation of the innateimmune system by CpG oligodeoxynucleotides: immunoprotectiveactivity and safety. Springer Semin Immunopathol 2000;22:173–83.
[67] Campetella OE, Henriksson J, Aslund L, Frash ACC, PettersonU, Cazzulo JJ. The major cysteine proteinase (cruzipain) fromTrypanosoma cruziis encoded by multiple polymorphic tandemlyorganized genes located on different chromosomes. Mol BiochemParasitol 1992;50:225–34.
Copyright © 2022 FDOKUMEN




















