
TrkB-Mediated Protection against Circadian Sensitivity to Noise Trauma in the Murine Cochlea
16
Current Biology 24, 658–663, March 17, 2014 ª2014 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2014.01.047 Report TrkB-Mediated Protection against Circadian Sensitivity to Noise Trauma in the Murine Cochlea Inna Meltser, 1,3 Christopher R. Cederroth, 1,3 Vasiliki Basinou, 1 Sergey Savelyev, 2 Gabriella S. Lundkvist, 1,2 and Barbara Canlon 1, * 1 Department of Physiology and Pharmacology, Karolinska Institutet, 17177 Stockholm, Sweden 2 Department of Neuroscience, Karolinska Institutet, 17177 Stockholm, Sweden Summary Noise-induced hearing loss (NIHL) is a debilitating sensory impairment affecting 10%–15% of the population, caused primarily through damage to the sensory hair cells or to the auditory neurons. Once lost, these never regenerate [1], and no effective drugs are available [2, 3]. Emerging evidence points toward an important contribution of synap- tic ribbons in the long-term coupling of the inner hair cell and afferent neuron synapse to maintain hearing [4]. Here we show in nocturnal mice that night noise overexposure trig- gers permanent hearing loss, whereas mice overexposed during the day recover to normal auditory thresholds. In view of this time-dependent sensitivity, we identified a self- sustained circadian rhythm in the isolated cochlea, as evi- denced by circadian expression of clock genes and ample PERIOD2::LUCIFERASE oscillations, originating mainly from the primary auditory neurons and hair cells. The tran- scripts of the otoprotecting brain-derived neurotrophic factor (BDNF) showed higher levels in response to day noise versus night noise, suggesting that BDNF-mediated signaling regulates noise sensitivity throughout the day. Administration of a selective BDNF receptor, tropomyosin- related kinase type B (TrkB), in the night protected the inner hair cell’s synaptic ribbons and subsequent full recovery of hearing thresholds after night noise overexposure. The TrkB agonist shifted the phase and boosted the amplitude of circadian rhythms in the isolated cochlea. These findings highlight the coupling of circadian rhythmicity and the TrkB receptor for the successful prevention and treatment of NIHL. Results Diurnal Sensitivity in Noise Overexposure and Its Effects on the Auditory Clock To evaluate whether the auditory clock would play a role in hear- ing functions in nonanesthetized animals, CBA/CaJ male mice were exposed to an acoustic startle paradigm during the inac- tive phase (9 a.m., zeitgeber time [ZT] 3) or the active phase (9 p.m., ZT 15). Acoustic startle amplitude was lower (p = 0.044; Figure 1A) and latency longer (p = 0.001; Figure 1B) in animals tested during the active phase than during the inactive phase. We next challenged awake mice with a noise trauma (6–12 kHz broadband noise of 100 dB sound pressure level [SPL] for 1 hr) during the inactive or active phase. This noise trauma is known to cause temporary damage characterized by the synaptic swelling of the auditory neuron dendrites [5]. We estimated auditory brainstem response (ABR) threshold shifts at either 24 hr or 2 weeks postexposure. Similar threshold shifts were found 24 hr postexposure for both day and night groups. While the recovery was complete for the day group at 2 weeks posttrauma (Figure 1C), the night group showed perma- nent threshold shifts (higher sound levels required to elicit a response) (p < 0.001) in the absence of hair cell loss (data not shown). These findings illustrate differences in recovery rate de- pending on the time of the day the noise trauma is delivered. Biological rhythms that are controlled throughout the day, namely circadian rhythms, are important for many bodily functions (e.g., metabolism, cell cycle, sleep-wake cycle, reproduction, immune function, and hormonal regulation). The master clock, the suprachiasmatic nucleus (SCN) of the hypothalamus, synchronizes and coordinates rhythms to regulate these various physiological functions [6, 7]. The central and peripheral clockwork circuitry contains a self- regulated feedback loop consisting of four transcriptional repressor-encoding genes (Per1, Per2, Cry1, and Cry2), which are regulated by BMAL1-CLOCK complexes, which in turn will be repressed by PER-CRY complexes resulting in a 24 hr cycle. A secondary loop involves REVERB-a, which represses Bmal1 transcription in a circadian manner. These core clock elements are necessary for the generation and maintenance of circadian rhythms [8]. We examined the temporal expres- sion patterns of mRNAs encoding central clock components from CBA/CaJ mouse cochleae. As depicted in Figures S1A– S1D available online, SYBR Green RT-PCR assays show smaller amplitudes of Per1, Bmal1, and Reverb-a transcript amplitudes in the cochlea (2-fold) than in the well-character- ized liver (15- to 30-fold). However, Per2 mRNA transcripts were as ample in the cochlea as in the liver (6-fold), which prompted us to evaluate whether the clock is expressed autonomously in the cochlea. For this purpose, we evaluated by means of real-time bioluminescence the expression of PER2 in adult cochlear explants using PERIOD2::LUCIFERASE (PER2::LUC) mice, in which a Luc gene has been fused in frame to the 3 0 end of the endogenous mouse Per2 gene generating a PER2 protein fused with luciferase [9]. We found that iso- lated cochleae demonstrated robust self-sustained molecular rhythmicity of PER2::LUC expression (Figures S1E–S1G), which damping could be kick-started by the addition of the synchronizing agent dexamethasone (Figure S1H). PER2 was localized mainly in inner and outer hair cells and in spiral gan- glion neurons from the cochlea (Figures S1L–S1O). Immunore- activity was not expressed in the hair cells and spiral ganglion neurons in mPer2 Brdm1 mutants. We next found that night noise overexposure affected molecular cochlear rhythms more than day noise. Night noise (p = 0.001) but not day noise (p = 0.078) overexposure decreased the amplitude of Per2 mRNA transcripts by 30% (Figure 1D). Night noise overexposure suppressed Reverb-a mRNA transcript oscillations to a greater extent than day noise overexposure (p < 0.007, day noise versus night noise; Figure 1F). The known antiphase oscillations of Bmal1 were 3 These authors contributed equally to this work *Correspondence: [email protected]
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of TrkB-Mediated Protection against Circadian Sensitivity to Noise Trauma in the Murine Cochlea

TrkB-Mediated Protection
Current Biology 24, 658–663, March 17, 2014 ª2014 Elsevier Ltd All rights reserved http://dx.doi.org/10.1016/j.cub.2014.01.047
Report
against Circadian Sensitivityto Noise Trauma in the Murine Cochlea
Inna Meltser,1,3 Christopher R. Cederroth,1,3
Vasiliki Basinou,1 Sergey Savelyev,2
Gabriella S. Lundkvist,1,2 and Barbara Canlon1,*1Department of Physiology and Pharmacology,Karolinska Institutet, 17177 Stockholm, Sweden2Department of Neuroscience, Karolinska Institutet,17177 Stockholm, Sweden
Summary
Noise-induced hearing loss (NIHL) is a debilitating sensoryimpairment affecting 10%–15% of the population, caused
primarily through damage to the sensory hair cells or tothe auditory neurons. Once lost, these never regenerate
[1], and no effective drugs are available [2, 3]. Emergingevidence points toward an important contribution of synap-
tic ribbons in the long-term coupling of the inner hair cell andafferent neuron synapse to maintain hearing [4]. Here we
show in nocturnal mice that night noise overexposure trig-gers permanent hearing loss, whereas mice overexposed
during the day recover to normal auditory thresholds. Inview of this time-dependent sensitivity, we identified a self-
sustained circadian rhythm in the isolated cochlea, as evi-denced by circadian expression of clock genes and ample
PERIOD2::LUCIFERASE oscillations, originating mainlyfrom the primary auditory neurons and hair cells. The tran-
scripts of the otoprotecting brain-derived neurotrophic
factor (BDNF) showed higher levels in response to daynoise versus night noise, suggesting that BDNF-mediated
signaling regulates noise sensitivity throughout the day.Administration of a selective BDNF receptor, tropomyosin-
related kinase type B (TrkB), in the night protected the innerhair cell’s synaptic ribbons and subsequent full recovery of
hearing thresholds after night noise overexposure. The TrkBagonist shifted the phase and boosted the amplitude of
circadian rhythms in the isolated cochlea. These findingshighlight the coupling of circadian rhythmicity and the
TrkB receptor for the successful prevention and treatmentof NIHL.
Results
Diurnal Sensitivity in Noise Overexposure and Its Effects
on the Auditory ClockToevaluatewhether the auditory clockwouldplay a role in hear-ing functions in nonanesthetized animals, CBA/CaJ male micewere exposed to an acoustic startle paradigm during the inac-tive phase (9 a.m., zeitgeber time [ZT] 3) or the active phase(9 p.m., ZT 15). Acoustic startle amplitude was lower (p =0.044; Figure 1A) and latency longer (p = 0.001; Figure 1B) inanimals tested during the active phase than during the inactivephase. We next challenged awake mice with a noise trauma(6–12 kHz broadband noise of 100 dB sound pressure level
3These authors contributed equally to this work
*Correspondence: [email protected]
[SPL] for 1 hr) during the inactive or active phase. This noisetrauma is known to cause temporary damage characterizedby the synaptic swelling of the auditory neuron dendrites [5].We estimated auditory brainstem response (ABR) thresholdshifts at either 24 hr or 2 weeks postexposure. Similar thresholdshifts were found 24 hr postexposure for both day and nightgroups. While the recovery was complete for the day group at2weeksposttrauma (Figure1C), thenightgroupshowedperma-nent threshold shifts (higher sound levels required to elicit aresponse) (p < 0.001) in the absence of hair cell loss (data notshown). These findings illustratedifferences in recovery ratede-pending on the time of the day the noise trauma is delivered.Biological rhythms that are controlled throughout the day,
namely circadian rhythms, are important for many bodilyfunctions (e.g., metabolism, cell cycle, sleep-wake cycle,reproduction, immune function, and hormonal regulation).The master clock, the suprachiasmatic nucleus (SCN) of thehypothalamus, synchronizes and coordinates rhythms toregulate these various physiological functions [6, 7]. Thecentral and peripheral clockwork circuitry contains a self-regulated feedback loop consisting of four transcriptionalrepressor-encoding genes (Per1, Per2, Cry1, and Cry2), whichare regulated by BMAL1-CLOCK complexes, which in turn willbe repressed by PER-CRY complexes resulting in a 24 hrcycle. A secondary loop involves REVERB-a, which repressesBmal1 transcription in a circadian manner. These core clockelements are necessary for the generation and maintenanceof circadian rhythms [8]. We examined the temporal expres-sion patterns of mRNAs encoding central clock componentsfrom CBA/CaJ mouse cochleae. As depicted in Figures S1A–S1D available online, SYBR Green RT-PCR assays showsmaller amplitudes of Per1, Bmal1, and Reverb-a transcriptamplitudes in the cochlea (2-fold) than in the well-character-ized liver (15- to 30-fold). However, Per2 mRNA transcriptswere as ample in the cochlea as in the liver (6-fold), whichprompted us to evaluate whether the clock is expressedautonomously in the cochlea. For this purpose, we evaluatedby means of real-time bioluminescence the expression ofPER2 in adult cochlear explants using PERIOD2::LUCIFERASE(PER2::LUC)mice, inwhich a Luc gene has been fused in frameto the 30 end of the endogenous mouse Per2 gene generatinga PER2 protein fused with luciferase [9]. We found that iso-lated cochleae demonstrated robust self-sustained molecularrhythmicity of PER2::LUC expression (Figures S1E–S1G),which damping could be kick-started by the addition of thesynchronizing agent dexamethasone (Figure S1H). PER2 waslocalized mainly in inner and outer hair cells and in spiral gan-glion neurons from the cochlea (Figures S1L–S1O). Immunore-activity was not expressed in the hair cells and spiral ganglionneurons in mPer2Brdm1 mutants.We next found that night noise overexposure affected
molecular cochlear rhythms more than day noise. Night noise(p = 0.001) but not day noise (p = 0.078) overexposuredecreased the amplitude of Per2 mRNA transcripts by 30%(Figure 1D). Night noise overexposure suppressed Reverb-amRNA transcript oscillations to a greater extent than daynoise overexposure (p < 0.007, day noise versus night noise;Figure 1F). The known antiphase oscillations of Bmal1 were
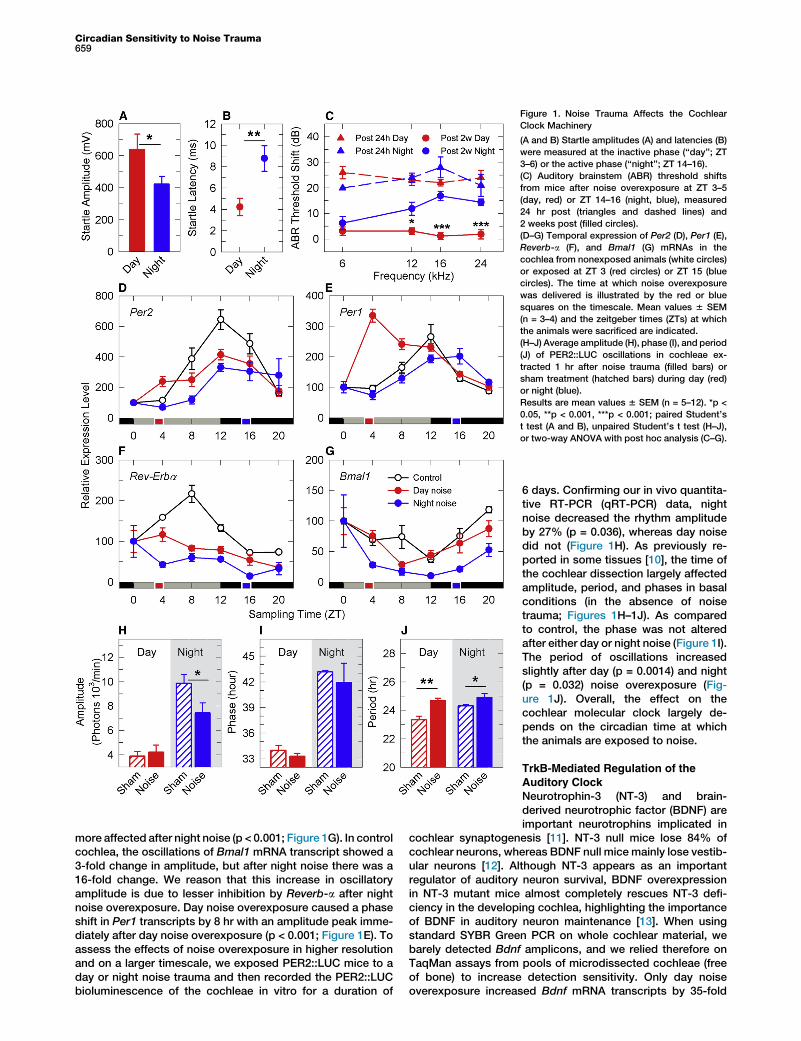
Figure 1. Noise Trauma Affects the Cochlear
Clock Machinery
(A and B) Startle amplitudes (A) and latencies (B)
were measured at the inactive phase (‘‘day’’; ZT
3–6) or the active phase (‘‘night’’; ZT 14–16).
(C) Auditory brainstem (ABR) threshold shifts
from mice after noise overexposure at ZT 3–5
(day, red) or ZT 14–16 (night, blue), measured
24 hr post (triangles and dashed lines) and
2 weeks post (filled circles).
(D–G) Temporal expression of Per2 (D), Per1 (E),
Reverb-a (F), and Bmal1 (G) mRNAs in the
cochlea from nonexposed animals (white circles)
or exposed at ZT 3 (red circles) or ZT 15 (blue
circles). The time at which noise overexposure
was delivered is illustrated by the red or blue
squares on the timescale. Mean values 6 SEM
(n = 3–4) and the zeitgeber times (ZTs) at which
the animals were sacrificed are indicated.
(H–J) Average amplitude (H), phase (I), and period
(J) of PER2::LUC oscillations in cochleae ex-
tracted 1 hr after noise trauma (filled bars) or
sham treatment (hatched bars) during day (red)
or night (blue).
Results are mean values 6 SEM (n = 5–12). *p <
0.05, **p < 0.001, ***p < 0.001; paired Student’s
t test (A and B), unpaired Student’s t test (H–J),
or two-way ANOVA with post hoc analysis (C–G).
Circadian Sensitivity to Noise Trauma659
more affected after night noise (p < 0.001; Figure 1G). In controlcochlea, the oscillations of Bmal1 mRNA transcript showed a3-fold change in amplitude, but after night noise there was a16-fold change. We reason that this increase in oscillatoryamplitude is due to lesser inhibition by Reverb-a after nightnoise overexposure. Day noise overexposure caused a phaseshift in Per1 transcripts by 8 hr with an amplitude peak imme-diately after day noise overexposure (p < 0.001; Figure 1E). Toassess the effects of noise overexposure in higher resolutionand on a larger timescale, we exposed PER2::LUC mice to aday or night noise trauma and then recorded the PER2::LUCbioluminescence of the cochleae in vitro for a duration of
6 days. Confirming our in vivo quantita-tive RT-PCR (qRT-PCR) data, nightnoise decreased the rhythm amplitudeby 27% (p = 0.036), whereas day noisedid not (Figure 1H). As previously re-ported in some tissues [10], the time ofthe cochlear dissection largely affectedamplitude, period, and phases in basalconditions (in the absence of noisetrauma; Figures 1H–1J). As comparedto control, the phase was not alteredafter either day or night noise (Figure 1I).The period of oscillations increasedslightly after day (p = 0.0014) and night(p = 0.032) noise overexposure (Fig-ure 1J). Overall, the effect on thecochlear molecular clock largely de-pends on the circadian time at whichthe animals are exposed to noise.
TrkB-Mediated Regulation of the
Auditory ClockNeurotrophin-3 (NT-3) and brain-derived neurotrophic factor (BDNF) areimportant neurotrophins implicated in
cochlear synaptogenesis [11]. NT-3 null mice lose 84% ofcochlear neurons, whereas BDNF null mice mainly lose vestib-ular neurons [12]. Although NT-3 appears as an importantregulator of auditory neuron survival, BDNF overexpressionin NT-3 mutant mice almost completely rescues NT-3 defi-ciency in the developing cochlea, highlighting the importanceof BDNF in auditory neuron maintenance [13]. When usingstandard SYBR Green PCR on whole cochlear material, webarely detected Bdnf amplicons, and we relied therefore onTaqMan assays from pools of microdissected cochleae (freeof bone) to increase detection sensitivity. Only day noiseoverexposure increased Bdnf mRNA transcripts by 35-fold
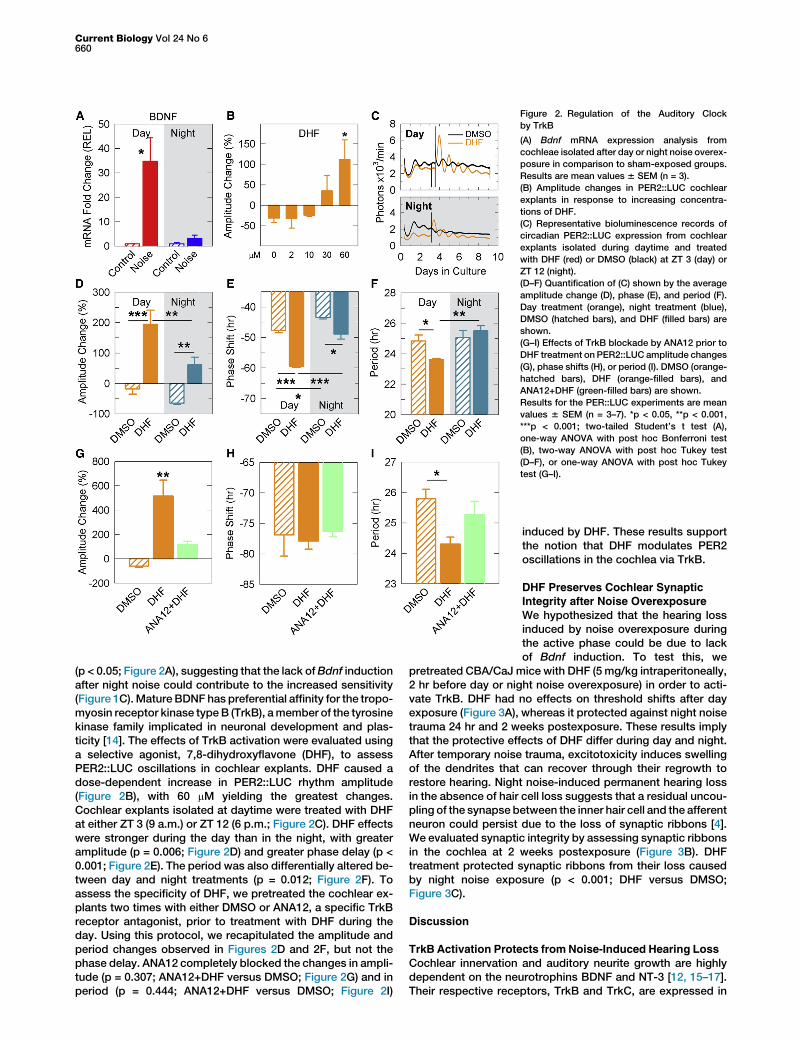
Figure 2. Regulation of the Auditory Clock
by TrkB
(A) Bdnf mRNA expression analysis from
cochleae isolated after day or night noise overex-
posure in comparison to sham-exposed groups.
Results are mean values 6 SEM (n = 3).
(B) Amplitude changes in PER2::LUC cochlear
explants in response to increasing concentra-
tions of DHF.
(C) Representative bioluminescence records of
circadian PER2::LUC expression from cochlear
explants isolated during daytime and treated
with DHF (red) or DMSO (black) at ZT 3 (day) or
ZT 12 (night).
(D–F) Quantification of (C) shown by the average
amplitude change (D), phase (E), and period (F).
Day treatment (orange), night treatment (blue),
DMSO (hatched bars), and DHF (filled bars) are
shown.
(G–I) Effects of TrkB blockade by ANA12 prior to
DHF treatment on PER2::LUC amplitude changes
(G), phase shifts (H), or period (I). DMSO (orange-
hatched bars), DHF (orange-filled bars), and
ANA12+DHF (green-filled bars) are shown.
Results for the PER::LUC experiments are mean
values 6 SEM (n = 3–7). *p < 0.05, **p < 0.001,
***p < 0.001; two-tailed Student’s t test (A),
one-way ANOVA with post hoc Bonferroni test
(B), two-way ANOVA with post hoc Tukey test
(D–F), or one-way ANOVA with post hoc Tukey
test (G–I).
Current Biology Vol 24 No 6660
(p < 0.05; Figure 2A), suggesting that the lack ofBdnf inductionafter night noise could contribute to the increased sensitivity(Figure 1C).Mature BDNF has preferential affinity for the tropo-myosin receptor kinase typeB (TrkB), amember of the tyrosinekinase family implicated in neuronal development and plas-ticity [14]. The effects of TrkB activation were evaluated usinga selective agonist, 7,8-dihydroxyflavone (DHF), to assessPER2::LUC oscillations in cochlear explants. DHF caused adose-dependent increase in PER2::LUC rhythm amplitude(Figure 2B), with 60 mM yielding the greatest changes.Cochlear explants isolated at daytime were treated with DHFat either ZT 3 (9 a.m.) or ZT 12 (6 p.m.; Figure 2C). DHF effectswere stronger during the day than in the night, with greateramplitude (p = 0.006; Figure 2D) and greater phase delay (p <0.001; Figure 2E). The period was also differentially altered be-tween day and night treatments (p = 0.012; Figure 2F). Toassess the specificity of DHF, we pretreated the cochlear ex-plants two times with either DMSO or ANA12, a specific TrkBreceptor antagonist, prior to treatment with DHF during theday. Using this protocol, we recapitulated the amplitude andperiod changes observed in Figures 2D and 2F, but not thephase delay. ANA12 completely blocked the changes in ampli-tude (p = 0.307; ANA12+DHF versus DMSO; Figure 2G) and inperiod (p = 0.444; ANA12+DHF versus DMSO; Figure 2I)
induced by DHF. These results supportthe notion that DHF modulates PER2oscillations in the cochlea via TrkB.
DHF Preserves Cochlear SynapticIntegrity after Noise Overexposure
We hypothesized that the hearing lossinduced by noise overexposure duringthe active phase could be due to lackof Bdnf induction. To test this, we
pretreated CBA/CaJ mice with DHF (5 mg/kg intraperitoneally,2 hr before day or night noise overexposure) in order to acti-vate TrkB. DHF had no effects on threshold shifts after dayexposure (Figure 3A), whereas it protected against night noisetrauma 24 hr and 2 weeks postexposure. These results implythat the protective effects of DHF differ during day and night.After temporary noise trauma, excitotoxicity induces swellingof the dendrites that can recover through their regrowth torestore hearing. Night noise-induced permanent hearing lossin the absence of hair cell loss suggests that a residual uncou-pling of the synapse between the inner hair cell and the afferentneuron could persist due to the loss of synaptic ribbons [4].We evaluated synaptic integrity by assessing synaptic ribbonsin the cochlea at 2 weeks postexposure (Figure 3B). DHFtreatment protected synaptic ribbons from their loss causedby night noise exposure (p < 0.001; DHF versus DMSO;Figure 3C).
Discussion
TrkB Activation Protects from Noise-Induced Hearing Loss
Cochlear innervation and auditory neurite growth are highlydependent on the neurotrophins BDNF and NT-3 [12, 15–17].Their respective receptors, TrkB and TrkC, are expressed in

Figure 3. Maintenance of Synaptic Integrity by TrkB Activation after Night Noise Injury
(A) ABR threshold shifts from mice exposed to noise at ZT 3–5 (day, red) or ZT 14–16 (night, blue), measured 24 hr post (left panel) and 2 weeks post (right
panel). DMSO-treated animals are indicated by white circles; DHF-treated animals are indicated by filled circles. Results are mean values6 SEM (n = 4–9);
two-way ANOVA post hoc Holm-Sidak.
(B) Representative images of synaptic ribbons (arrows) beneath inner hair cell nuclei from 0.5mm (low-frequency region) and 3.5mm (high-frequency region)
distance from the apex of controls (upper panel), Noise+DHF (middle panel), and Noise+DMSO (lower panel). Scale bar represents 10 mm. Corresponding
frequency map is provided at the upper right of (C).
(C) Quantification of the synaptic ribbons along the length of the cochlea from controls (empty gray circles) and 2 weeks post DMSO-treated night noise
(empty blue circles) and DHF-treated night noise (filled blue circles).
Results in (A) and (C) are mean values 6 SEM (n = 4). *p < 0.05, **p < 0.001, ***p < 0.001; two-way ANOVA with post hoc Tukey test.
Circadian Sensitivity to Noise Trauma661
auditory neurons [18–20]. Loss of TrkC function, but notTrkB function, results in a loss of 50% of auditory neurons;however, dual knockout of TrkB and TrkC causes a completeabsence of auditory neurons, highlighting the complementarysupport of both signaling pathways on auditory neuronal main-tenance [12]. In the cultured postnatal cochlea, TrkC, but notTrkB, played a role in axon growth and synaptogenesis,although both neurotrophins were equally efficient in regener-ating synapses after excitotoxicity [21]. Our study providesevidence that TrkB activationmay prove useful for maintainingsynaptic integrity after noise injury in the adult mouse cochleain vivo.
BDNF has been used by different means to protect fromhearing loss or to restore hearing [17, 22, 23], but its sizeand short lifespan preempt its ease of use. A selective TrkBagonist, DHF, has recently been identified in a neuronalcell-based apoptotic screen [24]. A structural activity rela-tionship study from the same group generated 7,8,30-trihydroxyflavone (7,8,30-THF) with three times greater po-tency on TrkB than DHF [25]. Local delivery of DHF andTHF in the ear has been shown to rescue auditory neuronalloss and function in a mouse model of auditory neuropathy(cCx26 null mice) [26, 27]. Our results are in line withthese findings and suggest that DHF may (1) protect immedi-ately, presumably by decreasing glutamate release duringnoise overexposure and subsequent excitotoxicity, (2) acton the recovery processes after noise overexposure byfacilitating synaptogenesis, or (3) a combination of the two.Although we do not exclude worse recovery or potentialdamage on outer hair cells after night noise overexposure,the decreased temporary threshold shifts observed 24 hrposttrauma in the DHF group (Figure 3A) suggest that theexcitotoxic damage does not occur to the same extent asin the control group.
A Circadian Machinery Responding to Noise
Although numerous molecular clocks have been found indifferent tissues [28, 29], this is the first demonstration thatthe adult cochlea contains rhythmically expressed clockgenes, with robust and self-sustained oscillations. We havefound, to our knowledge for the first time, that the auditorysensitivity to noise trauma depends on the time of day andthat noise overexposure alters circadian oscillations ofcochlear Per1, Per2, Reverb-a, and Bmal1 mRNA transcripts.Correlating with the increased sensitivity to noise overexpo-sure at night, greater effects on the circadian machinerywere observed after night noise overexposure (suppressionof Per2 and Reverb-a oscillations and greater oscillations ofBmal1; Figures 1D–1G). Since REVERB-a inhibits Bmal1transcription, we propose that the suppression of Reverb-aoscillations is linked to the increased oscillations of circa-dian Bmal1 mRNA transcripts occurring after night noiseoverexposure.TrkB may play a critical role in gating the circadian auditory
clock to sound. TrkB could modulate circadian rhythmsthrough upstream connections with the clockwork circuitry,including that of the SCN. Since TrkB mutant mice are embry-onic lethal [30], targeted deletion of TrkB specifically in theSCN or the use of SCN-specific dominant-negative TrkB-expressing mice should provide interesting insights into thecontribution of TrkB to the auditory clock machinery.
Conclusions
Sensitivity to noise trauma is greater at certain times of theday, both at the molecular and behavioral level, with conse-quences on hearing. It is likely that the mouse auditory clockdescribed here will have similar properties in humans, as theauditory and circadian systems in mammalian species arehighly homologous. For this reason, our findings could have

Current Biology Vol 24 No 6662
important implications with reference to the circadian variationin the potential for recovery from noise trauma that may play arole for people working in noisy environments, shift workers,flight crew that frequently travel across time zones, and thosewho recurrently visit nightclubs.
Experimental Procedures
Animal Handling
All experimental procedures involving animals were performed in accor-
dance with the guidelines and regulations set forth by Karolinska Institutet
and the Stockholm Norra Djurforsoksetiska Namnd. Recordings of circa-
dian oscillations of the PER2 protein were performed using tissues
obtained from knockin PER2::LUC transgenic mice with a C57BL/6 back-
ground [9], generously provided by J. Takahashi. CBA/CaJ and PER2::LUC
male mice aged between 2 and 4 months were used for audiological,
morphological, and molecular experiments. Temperature was maintained
between 19�C and 21�C. Because lights were on at 6 a.m. and off by
6 p.m., we set 6 a.m. as ZT 0. Handling at ZT 14–16 (darkness) was per-
formed in red light.
Organotypic Cultures
Adult cochleae, SCNs, and livers were dissected from PER2::LUC mice and
cultured organotypically on a membrane (Millipore; PICMORG50). The iso-
lated tissues were cultured as described [31, 32]. Cochleae, dissected
free of bone and stria vascularis, were kept in culture for a minimum of
6 days.
Drug Treatment
Mice were given a 4 ml/kg intraperitoneal injection of DHF (5 mg/kg) dis-
solved in vehicle (18% DMSO in PBS solution) 2 hr prior to noise trauma.
For in vitro treatments, two protocols were used for the treatment of
cochlear explants: (1) DHF day/night experiments (Figures 2D–2F), in which
cochlear explants were exposed to DMSO vehicle or 60 mM DHF at either
ZT3 or ZT12, and (2) DHF/ANA12 experiments (Figures 2G–2I), in which
cochlear explants were pre-exposed on day 0 and day 2 with DMSO
vehicle or ANA12 (Sigma-Aldrich; SML0209) at ZT 4 and the explants
were cotreated with DHF on day 3 (Tocris; 3826) and vehicle or ANA12
at ZT 4.
Acoustic Trauma and Auditory Brainstem Response
Awake and unanesthetized animals were treated with free-field broadband
noise at 6–12 kHz at an intensity of 100 dB SPL for 1 hr to generate hearing
loss, similar to a previously described noise trauma paradigm [33]. Post-
trauma measurements were performed either 24 hr after the day exposure
or 24 after the night exposure. However, the final measurements made at
2 weeks posttrauma were performed only during the day because (1) we
found no differences in basal ABR levels between day and night and (2) after
2 weeks posttrauma, ABR levels are considered stable.
Acoustic Startle Response
CBA/CaJ male mice were tested for the acoustic startle response (ASR) at
sleep (ZT 3–6) and awake (ZT 14–16) phases. The same animals were
used for the experiment, with 7-day intervals between day and night
sessions. The ASR paradigm was described previously [34]. The startle
magnitude (Vmax, where V is the mean maximal startle response to the
‘‘pulse-alone’’ trials) and startle latency (Tmax, where T is the mean latency
to the maximal response to the ‘‘pulse-alone’’ trials) were calculated.
Immunocytochemistry and Quantification of Synaptic Ribbons
For immunostaining of PER2, wild-type or mPer2Brdm1 mutants (having a
frame deletion that produces an unstable PER2 protein) [35] underwent
transcardiac perfusion (4% paraformaldehyde), and cochleae were decalci-
fied for 12 hr in EDTA 2% for 4 days and cryosectioned. Sections were
immunostained with a rabbit antibody directed against PER2 (PER21-A;
Alpha Diagnostic International, 1:100). For the quantification of synaptic rib-
bons, surface preparations were stained for C-terminal binding protein 2
(mouse anti-CtBP2; 612044 from BD Biosciences, 1:200) and secondary
fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit and tetrame-
thylrhodamine isothiocyanate (TRIC)-conjugated goat anti-mouse anti-
bodies (Jackson ImmunoResearch, 1:100). Confocal image stacks were
analyzed using ImageJ software. Quantification was performed using an
automated particle counter after converting the image to grayscale and
thresholding the image. This technique was manually validated before col-
lecting the data.
Quantitative Real-Time PCR
SYBR Green qRT-PCR assays were performed as described previously
[36, 37], with minor modifications. CBA/CaJ male mice were put into dark-
ness for 72 hr prior to their exposure to either silence or sound (100 dB,
6–12 kHz narrow-band white noise, for 1 hr). The cochleae (controls and
day and night noise exposure) were collected at fixed ZTs to allow compar-
isons. Total RNAs were extracted from whole cochleae using the Direct-zol
RNAMiniPrep Kit from Zymo Research (Nordic Biolabs AB) immediately fol-
lowed by DNase I treatment (Invitrogen). RNA integrity was assessed using
RNA 6000 Nanochips with an Agilent 2100 Bioanalyzer (Agilent Technolo-
gies), and quantity was evaluatedwith NanoDrop. RT-PCR assays were per-
formed with 0.5 mg total RNA from three or four adult cochleae or livers,
collected every 4 hr around the clock. Total RNAs were reverse transcribed
with the SuperScript II Reverse Transcriptase kit from Invitrogen (Life
Sciences). A mean quantity was calculated from triplicate PCR for each
sample, and this quantity was normalized with the geometric mean of two
to four most stable genes out of six reference genes (tubulin b [Tubb], glyc-
eraldehyde-3-phosphate dehydrogenase [Gapdh], transferrin receptor 1
[Trf1R], tubulin a2 [Tuba2], hypoxanthine phosphoribosyltransferase
[HPRT], and cyclophilin B) selected using the geNorm algorithm as
described previously [38]. Normalized quantities were averaged for three
technical replicates for each data point and are represented as mean 6
SD. The highest normalized relative quantity was arbitrarily designated as
a value of 1.0. Fold changes were calculated from the quotient of means
of these normalized quantities and reported as 6 SEM. The primers used
for qRT-PCR are listed in Table S1.
For the evaluation of BdnfmRNA transcript levels, we performed TaqMan
qRT-PCR assays (Applied Biosystems). Bdnf (Mm01334042_m1) gene
expression was quantified as described above using Cyclophilin E
(Mm00450929_m1) and Hprt (Mm00446968_m1) as the most stable normal-
ization genes.
Statistics
Data are presented as mean 6 SEM. Statistical analyses of PER2::LUC
rhythms and qRT-PCR data were performed with GraphPad Prism 5.04
(GraphPad Software). Analyses of other experiments were performed using
SigmaStat v3.5.
Supplemental Information
Supplemental Information includes one figure, one table, and Supplemental
Experimental Procedures and can be found with this article online at http://
dx.doi.org/10.1016/j.cub.2014.01.047.
Author Contributions
B.C., I.M., C.R.C., and G.S.L. designed the research. B.C., I.M., C.R.C.,
G.S.L., S.S., and V.B. performed research. B.C., I.M., C.R.C., G.S.L., S.S.,
and V.B. analyzed data. B.C., I.M., C.R.C., and G.S.L. wrote the manuscript.
Acknowledgments
We are most grateful to Urs Albrecht for generous help with sharing the
PER2 mutant mice (mPer2Brdm1) and wild-type mice samples. We thank
Didier Chollet from the Genomics facility of the NCCR program ‘‘Frontiers
in Genetics’’ at the University of Geneva for dedicated support. We also
thank Agneta Viberg and Anne-Sofie Johansson for technical support.
This work was supported by AFA Insurance Company (project and postdoc-
toral grant to I.M.), Swedish Medical Research Council grants K2010-62X-
09476-20-3 and 2008-3197 (B.C.), the National Institute on Deafness and
other Communication Disorders of the National Institutes of Health grant
R21DC013172 (B.C.), Swedish Society of Medicine grant SLS-95151
(G.S.L.), Knut and Alice Wallenberg Foundation grant KAW2008.0149
(B.C.), the Karolinska Institutet (B.C.), Stiftelsen Tysta Skolan (B.C., I.M.,
and C.R.C.), Stiftelsen Lars Hiertas Minne (C.R.C.), Magnus Bergvalls
Stiftelse (C.R.C. and G.S.L.), and Wenner-Gren Stiftelse (C.R.C.).
Received: August 30, 2013
Revised: December 20, 2013
Accepted: January 21, 2014
Published: February 27, 2014

Circadian Sensitivity to Noise Trauma663
References
1. Rubel, E.W., Furrer, S.A., and Stone, J.S. (2013). A brief history of hair
cell regeneration research and speculations on the future. Hear. Res.
297, 42–51.
2. Cederroth, C.R., Canlon, B., and Langguth, B. (2013). Hearing loss and
tinnitus—are funders and industry listening? Nat. Biotechnol. 31,
972–974.
3. Sheridan, C. (2013). Investors start backing hearing loss treatments.
Nat. Biotechnol. 31, 575–576.
4. Kujawa, S.G., and Liberman, M.C. (2009). Adding insult to injury:
cochlear nerve degeneration after ‘‘temporary’’ noise-induced hearing
loss. J. Neurosci. 29, 14077–14085.
5. Puel, J.L., Ruel, J., Gervais d’Aldin, C., and Pujol, R. (1998).
Excitotoxicity and repair of cochlear synapses after noise-trauma
induced hearing loss. Neuroreport 9, 2109–2114.
6. Bass, J., and Takahashi, J.S. (2010). Circadian integration of meta-
bolism and energetics. Science 330, 1349–1354.
7. Dibner, C., Schibler, U., and Albrecht, U. (2010). The mammalian circa-
dian timing system: organization and coordination of central and
peripheral clocks. Annu. Rev. Physiol. 72, 517–549.
8. Schibler, U. (2007). The daily timing of gene expression and physiology
in mammals. Dialogues Clin. Neurosci. 9, 257–272.
9. Yoo, S.H., Yamazaki, S., Lowrey, P.L., Shimomura, K., Ko, C.H., Buhr,
E.D., Siepka, S.M., Hong, H.K., Oh, W.J., Yoo, O.J., et al. (2004).
PERIOD2:LUCIFERASE real-time reporting of circadian dynamics re-
veals persistent circadian oscillations in mouse peripheral tissues.
Proc. Natl. Acad. Sci. USA 101, 5339–5346.
10. Yoshikawa, T., Yamazaki, S., and Menaker, M. (2005). Effects of prepa-
ration time on phase of cultured tissues reveal complexity of circadian
organization. J. Biol. Rhythms 20, 500–512.
11. Yang, T., Kersigo, J., Jahan, I., Pan, N., and Fritzsch, B. (2011). The
molecular basis of making spiral ganglion neurons and connecting
them to hair cells of the organ of Corti. Hear. Res. 278, 21–33.
12. Fritzsch, B., Tessarollo, L., Coppola, E., and Reichardt, L.F. (2004).
Neurotrophins in the ear: their roles in sensory neuron survival and fiber
guidance. Prog. Brain Res. 146, 265–278.
13. Farinas, I., Jones, K.R., Tessarollo, L., Vigers, A.J., Huang, E., Kirstein,
M., de Caprona, D.C., Coppola, V., Backus, C., Reichardt, L.F., et al.
(2001). Spatial shaping of cochlear innervation by temporally regulated
neurotrophin expression. J. Neurosci. 21, 6170–6180.
14. Shen, K., and Cowan, C.W. (2010). Guidance molecules in synapse for-
mation and plasticity. Cold Spring Harb. Perspect. Biol. 2, a001842.
15. Hegarty, J.L., Kay, A.R., and Green, S.H. (1997). Trophic support of
cultured spiral ganglion neurons by depolarization exceeds and is
additive with that by neurotrophins or cAMP and requires elevation of
[Ca2+]i within a set range. J. Neurosci. 17, 1959–1970.
16. Malgrange, B., Lefebvre, P., Van de Water, T.R., Staecker, H., and
Moonen, G. (1996). Effects of neurotrophins on early auditory neurones
in cell culture. Neuroreport 7, 913–917.
17. Wise, A.K., Richardson, R., Hardman, J., Clark, G., and O’leary, S.
(2005). Resprouting and survival of guinea pig cochlear neurons in
response to the administration of the neurotrophins brain-derived neu-
rotrophic factor and neurotrophin-3. J. Comp. Neurol. 487, 147–165.
18. Ylikoski, J., Pirvola, U., Moshnyakov, M., Palgi, J., Arumae, U., and
Saarma, M. (1993). Expression patterns of neurotrophin and their recep-
tor mRNAs in the rat inner ear. Hear. Res. 65, 69–78.
19. Pirvola, U., Arumae, U., Moshnyakov, M., Palgi, J., Saarma, M., and
Ylikoski, J. (1994). Coordinated expression and function of neurotro-
phins and their receptors in the rat inner ear during target innervation.
Hear. Res. 75, 131–144.
20. Pirvola, U., Hallbook, F., Xing-Qun, L., Virkkala, J., Saarma, M., and
Ylikoski, J. (1997). Expression of neurotrophins and Trk receptors in
the developing, adult, and regenerating avian cochlea. J. Neurobiol.
33, 1019–1033.
21. Wang, Q., and Green, S.H. (2011). Functional role of neurotrophin-3 in
synapse regeneration by spiral ganglion neurons on inner hair cells after
excitotoxic trauma in vitro. J. Neurosci. 31, 7938–7949.
22. Rejali, D., Lee, V.A., Abrashkin, K.A., Humayun, N., Swiderski, D.L., and
Raphael, Y. (2007). Cochlear implants and ex vivo BDNF gene therapy
protect spiral ganglion neurons. Hear. Res. 228, 180–187.
23. Pettingill, L.N., Wise, A.K., Geaney, M.S., and Shepherd, R.K. (2011).
Enhanced auditory neuron survival following cell-based BDNF treat-
ment in the deaf guinea pig. PLoS One 6, e18733.
24. Jang, S.W., Liu, X., Yepes, M., Shepherd, K.R., Miller, G.W., Liu, Y.,
Wilson, W.D., Xiao, G., Blanchi, B., Sun, Y.E., and Ye, K. (2010). A selec-
tive TrkB agonist with potent neurotrophic activities by 7,8-dihydroxy-
flavone. Proc. Natl. Acad. Sci. USA 107, 2687–2692.
25. Liu, X., Chan, C.-B., Jang, S.-W., Pradoldej, S., Huang, J., He, K., Phun,
L.H., France, S., Xiao, G., Jia, Y., et al. (2010). A synthetic 7,8-dihydrox-
yflavone derivative promotes neurogenesis and exhibits potent antide-
pressant effect. J. Med. Chem 53, 8274–8286.
26. Yu, Q., Chang, Q., Liu, X., Gong, S., Ye, K., and Lin, X. (2012). 7,8,30-Trihydroxyflavone, a potent small molecule TrkB receptor agonist, pro-
tects spiral ganglion neurons fromdegeneration both in vitro and in vivo.
Biochem. Biophys. Res. Commun. 422, 387–392.
27. Yu, Q., Chang, Q., Liu, X., Wang, Y., Li, H., Gong, S., Ye, K., and Lin, X.
(2013). Protection of spiral ganglion neurons from degeneration using
small-molecule TrkB receptor agonists. J. Neurosci. 33, 13042–13052.
28. Bell-Pedersen, D., Cassone, V.M., Earnest, D.J., Golden, S.S., Hardin,
P.E., Thomas, T.L., and Zoran, M.J. (2005). Circadian rhythms frommul-
tiple oscillators: lessons from diverse organisms. Nat. Rev. Genet. 6,
544–556.
29. Bass, J. (2012). Circadian topology of metabolism. Nature 491, 348–356.
30. Klein, R., Smeyne, R.J., Wurst, W., Long, L.K., Auerbach, B.A., Joyner,
A.L., and Barbacid, M. (1993). Targeted disruption of the trkB neurotro-
phin receptor gene results in nervous system lesions and neonatal
death. Cell 75, 113–122.
31. Savelyev, S.A., Larsson, K.C., Johansson, A.S., and Lundkvist, G.B.
(2011). Slice preparation, organotypic tissue culturing and luciferase
recording of clock gene activity in the suprachiasmatic nucleus. J. Vis.
Exp 48, 2439.
32. Yamazaki, S., Numano, R., Abe, M., Hida, A., Takahashi, R., Ueda, M.,
Block, G.D., Sakaki, Y., Menaker, M., and Tei, H. (2000). Resetting cen-
tral and peripheral circadian oscillators in transgenic rats. Science 288,
682–685.
33. Tahera, Y., Meltser, I., Johansson, P., Hansson, A.C., and Canlon, B.
(2006). Glucocorticoid receptor and nuclear factor-kappa B interactions
in restraint stress-mediated protection against acoustic trauma.
Endocrinology 147, 4430–4437.
34. Fisahn, A., Lavebratt, C., and Canlon, B. (2011). Acoustic startle hyper-
sensitivity inMcephmice and its effect on hippocampal excitability. Eur.
J. Neurosci. 34, 1121–1130.
35. Zheng, B., Larkin, D.W., Albrecht, U., Sun, Z.S., Sage, M., Eichele, G.,
Lee, C.C., and Bradley, A. (1999). The mPer2 gene encodes a functional
component of the mammalian circadian clock. Nature 400, 169–173.
36. Cederroth, C.R., Schaad, O., Descombes, P., Chambon, P., Vassalli,
J.D., and Nef, S. (2007). Estrogen receptor alpha is a major contributor
to estrogen-mediated fetal testis dysgenesis and cryptorchidism.
Endocrinology 148, 5507–5519.
37. Cederroth, C.R., Vinciguerra, M., Gjinovci, A., Kuhne, F., Klein, M.,
Cederroth, M., Caille, D., Suter, M., Neumann, D., James, R.W., et al.
(2008). Dietary phytoestrogens activate AMP-activated protein kinase
with improvement in lipid and glucose metabolism. Diabetes 57,
1176–1185.
38. Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De
Paepe, A., and Speleman, F. (2002). Accurate normalization of real-time
quantitative RT-PCR data by geometric averaging of multiple internal
control genes. Genome Biol 3, RESEARCH0034.

Current Biology, Volume 24
Supplemental Information
TrkB-Mediated Protection
against Circadian Sensitivity
to Noise Trauma in the Murine Cochlea
Inna Meltser, Christopher R. Cederroth, Vasiliki Basinou, Sergey Savelyev,
Gabriella S. Lundkvist, and Barbara Canlon

Figure S1. Circadian rhythmicity in the adult cochlea. Temporal expression of Reverb-a,
Per1 (b), Per2 (c), and Bmal1 (d) mRNAs in the cochlea (blue circles) and in the liver (gray
circles). Per1 mRNA levels oscillated with an amplitude of about 30-fold in the liver, whereas the
amplitude in the cochlea was only about two-fold. Per2 transcripts in the cochlea and liver
displayed similar circadian accumulation (6 fold amplitude). Transcript levels of Reverb- rose
more than 10-fold in the liver but with an amplitude of two-fold in the cochlea. The well-known
anti-phase circadian regulation of Bmal1 was found in both the liver and the cochlea. Results are

mean values ± SEM (n = 3-4) and the Zeitgeber times (ZT) at which the animals were sacrificed are
indicated. Without medium exchange or other pharmacological manipulations, isolated cochleae
demonstrated robust self-sustained molecular rhythmicity for at least 6 days as in the liver and the
SCN. Representative bioluminescence records of circadian PER2::LUC expression in cultured adult
cochleae (e), liver (f) and SCN (g) explants. Treatment with dexamethasone, a synchronizing agent,
restored rhythmicity when applied 4 days after damping (h), reflecting that cochlear cells became
asynchronous with time rather than dying off. Quantification of the rhythmic components are shown
by the average amplitude (i), phase (j) and period (k) for each organs. The average PER2::LUC
rhythm amplitude in photon/min was 3365 ± 648 in adult cochleae, 916 ± 551 in the liver and
40605 ± 2129 in the SCN (i). The phase of the oscillations was analyzed by measuring the time of
peak expression between 24 and 48 h in culture. PER2::LUC in the cochlea was maximally
expressed around Zeitgeber Time (ZT) 11-12 approximately at the same time as in the SCN
(cochlea at 36.7 ± 0.9; liver at 41.3 ± 0.5; SCN at 36 ± 0.2) (j). The period of the PER2::LUC
rhythm was 24 ± 0.2 h in the cochlea, 24.4 ± 0.5 in the liver, and 25.4 ± 0.1 h in the adult SCN (k).
Results are mean values ± SEM, n = 4-9. Immunostaining of PER2 in a cochlea of intact adult
CBA/CaJ mouse shows the localization of the protein in inner and outer hair and supporting cells of
the organ of Corti (l) and in the spiral ganglion neurons (m) of the cochlea. PER2 immunostaining
of a unilateral SCN (coronal section) is shown as positive control (n). As a negative control,
mPer2Brdm1
mutant cochleas (having a frame deletion that produces an unstable PER2 protein) were
used and immunoreactivity was not expressed in the hair cells or spiral ganglion neurons (data not
shown). Scale bar: 50 m.

Table S1. Sybr Green Primers
mPERIOD1 mPer1 CATTCCGCCTAACCCCATATG
CGGGGAGCTTCATAACCAGAG
mCLOCK mClock AGAGATGACAGTAGTATTTTTGATGGATTG
TCTCTACGTTTCTTTTCTGATTTGTTTCT
mBMAL1 mBmal1 CTCAGCTGCCTCGTTGCAATCGGG
GTACTCCATAGATTTCACCCGTATTTC
mREVERB-A mReverb-
ATG CCC ATG ACA AGT TAG GC
GGG CTA CCT GAT GCA TGA TT
mTUBB mTubb
GCAGTGCGGCAACCAGAT
AGTGGGATCAATGCCATGCT
mGAPDH mGapdh TCCATGACAACTTTGGCATTG
CAGTCTTCTGGGTGGCAGTGA
mTRF1R mTrf1R
GGAATCCCAGCAGTTTCTTTTTG
CAATGCCTCATAGGTATCCAATCTAG
mTUBA2 mTubA2 AGGAGCTGGCAAGCATGTG
CGGTGCGAACTTCATCGAT
mHPRT mHPRT
GCTCGAGATGTCATGAAGGAGAT
AAAGAACTTATAGCCCCCCTTGA
mCYCLO B mCycloB ATG TGG TTT TCG GCA AAG TT
TGA CAT CCT TCA GTG GCT TG

EXPERIMENTAL PROCEDURES
Ethics Statement and animal handling
All experimental procedures on animals were performed in accordance with the guidelines and
regulations set forth by Karolinska Institutet and “Stockholm’s Norra Djurförsöksetiska Nämnd”.
Recordings of circadian oscillations of the PER2 protein were performed using tissues obtained
from knock-in PERIOD2::LUCIFERASE (PER2::LUC) transgenic mice with a C57BL/6
background [S1], generously provided by Prof. J. Takahashi. CBA/J and PER2::LUC male mice
aged between 2 and 4 months mice were used for audiological, morphological and molecular
experiments. Animals had free access water and to food (Lactamin R34, Lantmännen). Food pellets
contained 43 mg/kg daidzin, 60 mg/kg genistin, 10 mg/kg glycitin, 2 mg/kg daidzein, 1.6 mg/kg
genistein (Lantmännen report). Temperature was maintained between 19° and 21°C. Because lights
were on at 6 a.m. and off by 6 p.m., we set 6 a.m. as the Zeitgeber time ZT 0. Handling at ZT 14-16
(darkness) was performed in red light.
Organotypic cultures
Adult cochleae, suprachiasmatic nucleus (SCN) and livers were dissected from PER2::LUC mice
and cultured organotypically on a membrane (Millipore, PICMORG50). The isolated tissues were
cultured as described [S2, S3]. Cochleae, dissected free of bone and stria vascularis, were kept in
culture for minimum 6 days. Real time luciferase reporter technology is described [S1-S3].
PER2::LUC rhythms (amplitude, phase, and period) were analyzed using Origin software 8.1 SR1
(Microcal Software, Northampton, MA, USA). Data from each recording trace was first de-trended
by subtraction of the 24 h baseline drift from the raw data. The period of one complete cycle was
defined as the time between two consecutive peaks (i.e. the highest photon count within one cycle)
and consecutive peaks were used for averaging periods. The amplitude was calculated as the
difference between highest (peak) and lowest (trough) photon count within one cycle. The

calculation was performed from trough-to-peak and from the peak-to-trough, thus giving two values
(half-cycles) within one cycle. In each recording, three half-cycles were used for amplitude
analyses. The very first peak after culture start was not used. The phase relationships were analyzed
by comparing the time of the peak (maximum bioluminescence) between 24 and 48 h after culture
start in each tissue. Phase shifts were calculated by comparing the time of the peak right before
treatment with the time of the peak right after the treatment.
Drug treatment
Mice were given a 4 ml/kg intraperitoneal injection of DHF (5 mg/kg) dissolved in the vehicle
(18 % DMSO in PBS solution) 2 h prior to noise trauma. For in vitro treatments, two protocols were
used for the treatment of cochlear explants:
1) DHF day-night experiments (Figure 3D-F): Cochlear explants were exposed to DMSO vehicle or
DHF 60 M either at ZT 3 or ZT12. Dexamethasone 21 phosphate disodium salt (Sigma Aldrich,
D1159) was applied on day 4. 2) DHF/ANA12 experiments (Figure 3G-I): Cochlear explants were
pre-exposed on day 0 and day 2 with DMSO vehicle or ANA12 (Sigma Aldrich, SML0209) at ZT
4. On day 3 the explants were co-treated with DHF (Tocris, 3826) and vehicle or ANA12 at ZT 4.
Acoustic trauma and auditory brainstem response (ABR)
To generate hearing loss, awake and unanesthetized animals were treated with free field broadband
noise at 6 - 12 kHz at intensity of 100 dB SPL for 1 h, similar to previously described noise trauma
paradigm [S4]. Auditory sensitivity was assessed with ABR thresholds for the frequency of 6, 12,16
and 24 kHz as described previously [S5]. Post-trauma measurements where performed either 24 h
after the day or night exposures. However, the final measurements made at 2 weeks post-trauma
were made only during the day because i) we found no differences in basal ABR levels between day
and night and ii) after 2 weeks post-trauma, the ABR levels are considered stable.

Acoustic startle response (ASR)
CBA male mice were tested for the ASR at sleep (ZT 3-6) and awake (ZT 14-16) phases. Same
animals were used for the experiment with 7 days interval between day and night sessions. The
ASR paradigm was previously described [S6]. The startle magnitude (Vmax, V - the mean maximal
startle response to the “pulse-alone” trials) and startle latency (Tmax, msec - the mean latency to the
maximal response to the “pulse-alone” trials) were calculated.
Immunocytochemistry and quantification of synaptic ribbons
For immunostaining of PER2, wild type or mPer2Brdm1
mutants (having a frame deletion that
produces an unstable PER2 protein) [S7] underwent transcardiac perfusion with 4%
paraformaldehyde and cochleae were decalcified in EDTA 2% for 4 days and cryosectioned
(14m). Sections were immunostained with a rabbit-antibody to PER2 (PER21-A, Alpha
Diagnostic, Texas USA; 1:100). For the quantification of synaptic ribbons, surface preparations
from fixed cochleae, decalcified for twelve hours in EDTA 2%, were stained for C-terminal binding
protein 2 (mouse anti-CtBP2, 612044 from BD-Biosciences, used at 1:200) and secondary FITC-
conjugated goat anti-rabbit and TRIC-conjugated goat anti-mouse antibodies (Jackson
ImmunoResearch; Pennsylvania USA; 1:100). Cochlear lengths were obtained for each sample and
a cochlear frequency map computed to precisely localize inner hair cells (IHCs) throughout the
length of the cochlea [S8]. Confocal z-stacks from every 0.3 mm distance along the basilar
membrane were made with a z-step-size of 642 nm. Averages were then made for every mm region.
Image stacks were then analyzed using Image J software. Synaptic ribbons were counted in 5-10
adjacent inner hair cells and divided the number of cells counted in order to obtain the number of
ribbons per cell. Quantification was performed using an automated particle counting after

converting the image to grayscale and thresholding the image. This technique was manually
validated before collecting the data.
Quantitative Real Time-PCR
SybrGreen qRT-PCR assays were performed as previously described [S9, S10] with minor
modifications. CBA male mice were put into darkness for 72 hours prior their exposure to either
silence or sound (100 dB, 6-12 kHz narrow band white noise, for 1 hour). To allow comparisons,
the cochleas (controls, day and night noise exposure) were collected at fixed ZT time points.
Animals were decapitated under isofluorane anesthesia and their cochleae and liver collected. Total
RNAs were extracted from whole cochleae using the Direct-zol™ RNA MiniPrep kit from
ZymoResearch (Nordic Biolabs AB, Sweden) according to the manufacturer’s protocol, and
immediately followed by DNAse I treatment (Invitrogen). RNA integrity was assessed using RNA
6000 nanochips with an Agilent 2100 bioanalyzer (Agilent Technologies, Inc., Palo Alto, CA) and
quantity was evaluated with Nanodrop. RT-PCR assays were performed with 0.5 g total RNA
from 3-4 adult cochleae, or livers, collected every 4 h around the clock. Total RNAs were reverse
transcribed with the Superscript II RT-kit from Invitrogen (Life Sciences, Sweden) according to
manufacturer’s instructions, and one-twentieth cDNA dilution was used as template for each PCR.
cDNA was PCR amplified in a 7900HT Sequence Detection Systems (Applied Biosystems, Foster
City, CA) using Power SYBR Green PCR master mix (Applied Biosystems). Raw threshold-cycle
(Ct) values were obtained from Sequence Detection Systems 2.0 software (Applied Biosystems).
Relative quantities (RQs) were calculated with the formula RQ = E - Ct, using efficiencies
calculated for each run with the Data Analysis for Real-Time PCR (DART-PCR) algorithm, as
described [S11]. A mean quantity was calculated from triplicate PCR for each sample, and this
quantity was normalized with the geometric mean of two to four most stable genes out of six
reference genes (tubulin , Tubb; glyceraldehyde-3-phosphate dehydrogenase, G3pdh; transferring

receptor 1, Trf1R; Tubulin 2, Tuba2; hypoxanthine phosphoribosyltransferase, HPRT; and
Cyclophilin B) selected using the geNorm algorithm as described [S12]. Normalized quantities
were averaged for three technical replicates for each data point and represented as the mean ± SD.
The highest normalized relative quantity was arbitrarily designated as a value of 1.0. Fold changes
were calculated from the quotient of means of these normalized quantities and reported as ± SEM.
The primers used for quantitative RT-PCR (qRT-PCR) are listed in Supplemental Table 1.
For the evaluation of Bdnf mRNA transcript levels, we performed Taqman qRT-PCR assays
(Applied Biosystems, CA, USA). Mice were decapitated under isofluorane anesthesia; cochleae
were extracted from the temporal bone and further dissected in RNAlater RNA stabilization reagent
(Qiagen). The outer bony shell of the cochlea and the vascular tissue (stria vascularis) were
removed, and cochleae were left in RNA stabilization reagent for 12-24 h at room temperature.
Bdnf (Mm01334042_m1) gene expression was quantified as described above using Cyclophilin E
(Mm00450929_m1) and Hprt (Mm00446968_m1) as the most stable normalization genes.
Statistics
Data are presented as a mean ± SEM. Statistical analysis of PER2::LUC rhythms and the qRT-PCR
was performed with GraphPad Prism 5.04 (GraphPad Software Inc., CA, USA). Analysis of other
experiments was performed using SigmaStat v 3.5. The tests used for each experiment are described
in the figure legends.
Supplemental references
S1. Yoo, S.H., Yamazaki, S., Lowrey, P.L., Shimomura, K., Ko, C.H., Buhr, E.D., Siepka, S.M., Hong,
H.K., Oh, W.J., Yoo, O.J., et al. (2004). PERIOD2::LUCIFERASE real-time reporting of circadian

dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proceedings of the
National Academy of Sciences of the United States of America 101, 5339-5346.
S2. Savelyev, S.A., Larsson, K.C., Johansson, A.S., and Lundkvist, G.B. (2011). Slice preparation,
organotypic tissue culturing and luciferase recording of clock gene activity in the suprachiasmatic
nucleus. Journal of visualized experiments : JoVE.
S3. Yamazaki, S., Numano, R., Abe, M., Hida, A., Takahashi, R., Ueda, M., Block, G.D., Sakaki, Y.,
Menaker, M., and Tei, H. (2000). Resetting central and peripheral circadian oscillators in transgenic
rats. Science 288, 682-685.
S4. Tahera, Y., Meltser, I., Johansson, P., Hansson, A.C., and Canlon, B. (2006). Glucocorticoid
receptor and nuclear factor-kappa B interactions in restraint stress-mediated protection against
acoustic trauma. Endocrinology 147, 4430-4437.
S5. Niu, X., Bogdanovic, N., and Canlon, B. (2004). The distribution and the modulation of tyrosine
hydroxylase immunoreactivity in the lateral olivocochlear system of the guinea-pig. Neuroscience
125, 725-733.
S6. Fisahn, A., Lavebratt, C., and Canlon, B. (2011). Acoustic startle hypersensitivity in Mceph mice
and its effect on hippocampal excitability. The European journal of neuroscience 34, 1121-1130.
S7. Zheng, B., Larkin, D.W., Albrecht, U., Sun, Z.S., Sage, M., Eichele, G., Lee, C.C., and Bradley, A.
(1999). The mPer2 gene encodes a functional component of the mammalian circadian clock. Nature
400, 169-173.
S8. Viberg, A., and Canlon, B. (2004). The guide to plotting a cochleogram. Hearing research 197, 1-10.
S9. Cederroth, C.R., Schaad, O., Descombes, P., Chambon, P., Vassalli, J.D., and Nef, S. (2007).
Estrogen receptor alpha is a major contributor to estrogen-mediated fetal testis dysgenesis and
cryptorchidism. Endocrinology 148, 5507-5519.
S10. Cederroth, C.R., Vinciguerra, M., Gjinovci, A., Kuhne, F., Klein, M., Cederroth, M., Caille, D.,
Suter, M., Neumann, D., James, R.W., et al. (2008). Dietary phytoestrogens activate AMP-activated
protein kinase with improvement in lipid and glucose metabolism. Diabetes 57, 1176-1185.
S11. Peirson, S.N., Butler, J.N., and Foster, R.G. (2003). Experimental validation of novel and
conventional approaches to quantitative real-time PCR data analysis. Nucleic Acids Res 31, e73.
S12. Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., and Speleman, F.
(2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of
multiple internal control genes. Genome Biol 3, RESEARCH0034.




















