
Proteomic characterization of novel histone post-translational modifications
Upload
independentCategory
view
2download
0
234
Biochimica et Biophysica Acta, 425 (1976) 234--245 © Elsevier Scientific Publishing Company, Amsterdam -- Printed in The Netherlands
BBA 98531
TRANSLATIONAL CONTROL OF PROTEIN SYNTHESIS IN STIMULATED WI-38 FIBROBLASTS
SANDRA M. WHELLY and RENATO BASERGA
Department of Pathology and Fels Research Institute, Temple University School of Medicine, Philadelphia, Pa. 19140 (U.S.A.)
(Received August 14th, 1975)
Summary
A cell-free protein synthesis system employing ribosomes from WI-38 human diploid fibroblasts was developed and its opt imum MgC12 and KC1 levels and pH value found. The rate at which ribosomes are able to incorporate radioactive leucine into proteins ([14C]leucine incorporation/10 min/100 pg rRNA) and the number of growing peptide chains [ 3H] puromycinpeptides formed/100 pg rRNA) was determined. When confluent monolayers of WI-38 cells were stimu- lated to proliferate by serum, a transient increase in the rate of peptide elonga- tion by ribosomes was observed at 60 min after stimulation. This increase was not affected by the presence of act inomycin D (10 pg/ml) in the stimulating medium. A change in the relative amount of certain ribosome-associated pro- teins accompanied the increased elongation rate of peptide growth. The alter- ation in associated proteins could not be accounted for by an increased synthe- sis of protein. Finally, the early activation of ribosomes in stimulated WI-38 cells appears to result from the removal of an inhibitor(s) of ribosome function.
Introduction
Formation of confluent monolayers in cell culture by human diploid fibro- blasts results in virtual cessation of DNA synthesis and mitosis [1--4]. Station- ary WI-38 human diploid fibroblasts can be stimulated to proliferate by re- placement of conditioned medium with new medium containing 10% fetal calf serum [3]. Stimulation to proliferate results in a number of biochemical alter- ations which precede the onset of DNA synthesis. Two of the earliest biochemi- cal changes, occurring an hour after stimulation, are an increase in chromatin template activity [5,6] and an increased synthesis of non-histone nuclear pro- teins [4]. It has also been shown that stimulation of RNA synthesis in WI-38

235
human diploid fibroblasts is dependent on the simultaneous and/or prior syn- thesis of proteins [7] .
The present s tudy was undertaken to determine whether or not the efficien- cy of the ribosomal translational system of stationary WI°38 human diploid fibroblasts was altered by a nutritional change. The experimental approach was to measure both the number of growing peptide chains and the rate at which the r ibosomes are able to incorporate [ 14 C] leucine into protein in a standard- ized cell-free protein synthesis system. This permits the measurement of the relative rate of peptide chain elongation in the in vitro system by isolated WI-38 human diploid fibroblast ribosomes. It will be shown that 60 min after a nutritional change there is a transient increase in the rate of peptide elongation by isolated ribosomes. This activation coincides with a change in the relative amount of certain ribosome-associated proteins. Some evidence will be pre- sented which suggests that the increase in the elongation rate results from the removal of an inhibitor(s) from ribosomes of stationary WI-38 human diploid fibroblasts.
Experimental procedure
Materials Fetal calf serum was purchased from Flow Laboratories, Rockville, Md.,
Earle's balanced salt solution and trypsin (1 : 250) from Microbiological Asso- ciates. Sucrose, urea (ultrapure grade) and act inomycin D were obtained from Schwarz--Mann. [ 14 C] leucine {280 Ci/mol), [ 3 H] leucine (44.2 Ci/mol), and 3H-labeled methoxypuromycin (3.72 Ci/mmol) were purchased from New Eng- land Nuclear Corp., Boston, Mass. ATP, GTP, phosphoenol pyruvate and pyru- vate kinase were obtained from Sigma Chemicals, St. Louis, Mo. All other chemicals were supplied by Mallinckrodt Chemical Works, St. Louis, Mo., and the Fisher Scientific Co., Pittsburgh, Pa., analytical grades being used where available.
Methods Cell culture. WI-38 human diploid fibroblasts [8] were obtained from Dr. L.
Hayflick through Contract No. 1, HD42828. Cell stocks, between the 21st and 30th passages, were maintained in roller vessels with a growth area of 840 cm 2 and were grown in Basal Eagle's Medium supplemented with 10% fetal calf serum, 2 mM glutamine and 50/ag/ml s treptomycin [3] . Fo r experiments the cells were grown in 1-1 Blake bottles (surface area 210 cm 2 ) for 7 days until they reached confluence [9] . The confluent monolayers were stimulated to proliferate by replacing the old medium with medium containing 10% (v/v) fetal calf serum. The controls were cultures in which the old medium was not changed.
Preparation o f WI-38 fibroblast ribosomes. All operations were performed at 4°C unless otherwise noted. After the medium was removed the cells were washed twice with Earle's balanced salt solution. 5 ml of a 0.25% solution of trypsin (1 : 250, Microbiological Associates) in Earle's salt solution was added, and the cells were left in contact with the trypsin solution for 5 rain. The trypsin solution was then aspirated and the cells were scraped with a rubber

236
policeman into Earle's balanced salt solution. The cells were washed twice in Earle's balanced salt solution. Cells were pooled and homogenized with 10 strokes of a tight-fitting Dounce homogenizer in 1 ml/Blake bot t le of homoge- nizing medium (50 mM Tris/HC1 (pH 7.5) 25 mM KC1, 5 mM MgC12 and 7 mM /3-mercaptoethanol) containing 0.25 M sucrose. The homogenate was centri- fuged at 400 X g for 10 rain and the crude nuclear pellet was re-homogenized in 0.5 ml/Blake bott le of homogenizing medium and centrifuged at 400 X g for 10 min. The post-nuclear supernatants were pooled and centrifuged at 26 000 X g for 20 min. The postmitochondrial supernatant was adjusted to 1% sodium deoxychola te and aliquots (3 ml) were layered over 9 ml of homogenizing me- dium containing 1.5 M sucrose and centrifuged at 165 000 X g for 3 h. The ribosomes were suspended in 10 ml of suspension medium (50 mM Tris/HC1 (pH 7.5) 100 mM KC1, 5 mM MgC12 and 7 mM fl-mercaptoethanol, and 0.25 M sucrose, and centrifuged at 165 000 X g for 1.5 h. The washed ribosomes were resuspended in a small amount of suspension medium and the RNA content measured [10] . Ribosomes thus prepared have a A26o/A28onm ratio of 1.65--1.80 and an RNA/prote in ratio of 0.55. The yield was from 30 to 45 pg of rRNA/Blake bottle. Sucrose gradient analysis of the preparations revealed the presence of polysomes, monosomes and subunits (not shown).
WI-38 fibroblast ribosomes protein-synthesizing system. The protein-synthe- sizing capacity of WI-38 fibroblast r ibosomes was determined by measuring the rate of incorporation of [ 14 C] leucine into heated trichloroacetic acid-insoluble material. The optimal MgC12 and KC1 concentrations and pH value were found for the system. The reaction mixture (0.5 ml) had the following composit ion: Tris/HC1 (pH 7.5) 50 mM; KC1,100 mM; MgC12, 5 mM; ~-mercaptoethanol, 7.0 mM; sucrose, 250 mM; ATP, 1.0 mM; GTP, 0.4 mM; phosphoenolpyruvate, 10 mM; 19 amino acids (without leucine), 0.05 mM each; L-[14C]leucine, 0.5 t~Ci/ml (spec. act. 280 Ci/mol); pyruvate kinase, 10 pg/ml; "pH 5 enzyme , " 1.0 mg/ml; WI-38 fibroblast ribosomes, 50 pg rRNA/ml. The pH 5 enzyme was prepared by the method described by Moldave [11] . The reaction mixture (without ribosomes) was equilibrated at 37°C for 1 rain and the reaction was initiated by the addition of ribosomes. After 10 rain incubation with gentle agitation, a 0.1 ml al iquot was applied to filter paper disks and immediately placed in cold 10% trichloroacetic acid. The filters were transferred to 5% trichloroacetic acid and heated at 90°C for 30 rain. They were then washed three times in cold 5% trichloroacetic acid, once in 95% ethanol, once in ether, dried and counted. Control assays containing all components including ribo- somes, but wi thout incubation, were performed for each r ibosome preparation and the blank r0dioactivity (<30 cpm) has been subtracted from the reported values. All assays were performed in duplicate. The reaction rate was constant during the interval of incubation and was dependent on the amount of rRNA added to the reaction mixture. The system was dependent upon the presence of a high-energy-generating system (ATP, GTP, phosphoenolpyruvate and pyru- rate kinase) and on the presence of the pH 5 enzyme.
Determination of the amounts of nascent peptide (active ribosomes in WI-38 fibroblast ribosomes. The relative amount or number of r ibosomes in the vari- ous preparations of WI-38 fibroblast ribosomes containing a growing peptide capable of forming an pept idyl-puromycin complex in vitro was determined by a

237
method similar to that described by Wool and Kurihara [12] and by Suvatte and Hagerman [13]. The reaction mixture (0.25 ml) for the formation of a 3 H-labeled peptidyl-puromycin complex from [ 3 H] puromycin had the follow- ing composition: Tris/HC1 (pH 7.5), 50 raM; MgC12, 5 mM; KC1, 100 mM; fi-mercaptoethanol, 7 mM; sucrose, 0.25 M; ATP, 1.0 mM; GTP, 0.4 mM; phos- phoenolpyruvate, 10 mM; pyruvate kinase, 10 /~g/ml; 3H-labeled methoxy- puromycin, 10.0 pCi/ml (spec. act. 3.82 Ci/mmol; New England Nuclear Corp.); WI-38 fibroblast ribosomes; 100 pg of rRNA/ml. The mixture was incubated at 37°C for 45 rain after which the ribosomes and peptides were precipitated by addition of 5 ml of 5% trichloroacetic acid. The mixture was heated at 90°C for 15 min, cooled on ice for 30 min and then collected on glass fiber filters (Millipore Corp.). The filters were washed by passing 60 ml (three 20-ml aliquots) of cold 5% trichloroacetic acid through the filter followed by 60 ml (three 20-ml aliquots) of 8 M urea and 10 ml of 95% ethanol. The filters were then dried and counted. Control assays, prepared by the addition of ribosomes after the addition of cold trichloroacetic acid, were performed for each ribosome preparation and the radioactivity (<50 cpm) was subtracted from all values before calculation of radioactivity in the peptide fraction. All assays were performed in triplicate. The formation of a 3 H-labeled peptidyl- puromycin complex was ribosome-dependent but independent of both the incubation times over the range 30--50 min and the amount of [ 3 H] puromycin added over the range of 5--20/~Ci/ml.
Sodium dodecyl sulfate gel electrophoresis of ribosome-associated proteins. Ribosomes from WI-38 fibroblasts either pulse labeled for 45 min with [ 3HI- leucine, 1 #Ci/ml (spec. act. 44.2 Ci/mmol) or unlabeled, were incubated at 37°C for 40 rain to release all nascent peptides in a reaction mixture contain- ing; Tris/HC1 (pH 7.5), 50 mM; MgC12, 5.0 mM; KC1, 100 mM; fi-mercapto- ethanol, 7 mM; sucrose, 0.25 M; ATP, 10 mM; GTP, 0.4 mM; phosphoenol- pyruvate, 10 mM; pyruvate kinase, 10 /ag/ml; puromycin, 0.1 mM; and ribo- somes containing 100 pg ribosomal RNA/ml. The ribosomes were then sedi- mented by centrifugation at 165 000 × g for 2 h. The sedimented ribosomes were then solubilized by heating at 50°C for 12 h in 0.2 ml of a solution containing 1% sodium dodecyl sulfate, 6 M urea, 1% /3-mercaptoethanol and 0.1 M sodium phosphate (pH 7.0). The solubilized material was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis on 10% acrylamide gels as described by Palmiter et al. [14].
After the tracking dye had migrated 7 cm, the gels were removed from the tubes and either cut into 2-mm sections from the origin to the dye front (radiolabeled proteins) or stained in 1% fast green in 7% acetic acid overnight, followed by destaining in 7% acetic acid (unlabeled proteins). The 2-mm slices from gels of radiolabeled fibroblasts were placed in scintillation vials in 1.0 ml of NCS (Amersham/Searle Corp.) and incubated at 50°C for 18 h. Scintillation fluid (10 ml) was added and the radioactivity in each slice was determined. Recovery of total radioactivity applied to the gels was 90--95% and the results are expressed as cpm in each gel slice.
Determination of radioactivity. Dried filter papers containing'4C-labeled protein were placed in scintillation vials with 10 ml of a toluene-based scintilla- tion fluid (4 g of 2,5-diphenyloxazole and 0.1 g of 1,4-bis(2-(4-methyl-5-phenyl-
238
oxazole)) in 1-1 of toluene) and counted in a Packard Tri-Carb Liquict Scintilla- tion Spectrometer, Model 3375. Membrane filters containing the 3H-labeled peptidyl-puromycin complex were counted as above. The counting efficiency for 14 C and 3H was 80% and 3%, respectively.
Determination o f ribosome composition. Protein was determined by the method of Lowry et al. [15] and rRNA content was determined as suggested by Wool and Cavicchi [10].
Results
Effects o f serum stimulation o f confluent monolayers on the rate o f protein synthesis and on the nascent peptide content o f isolated WI-38 fibroblast ribosomes
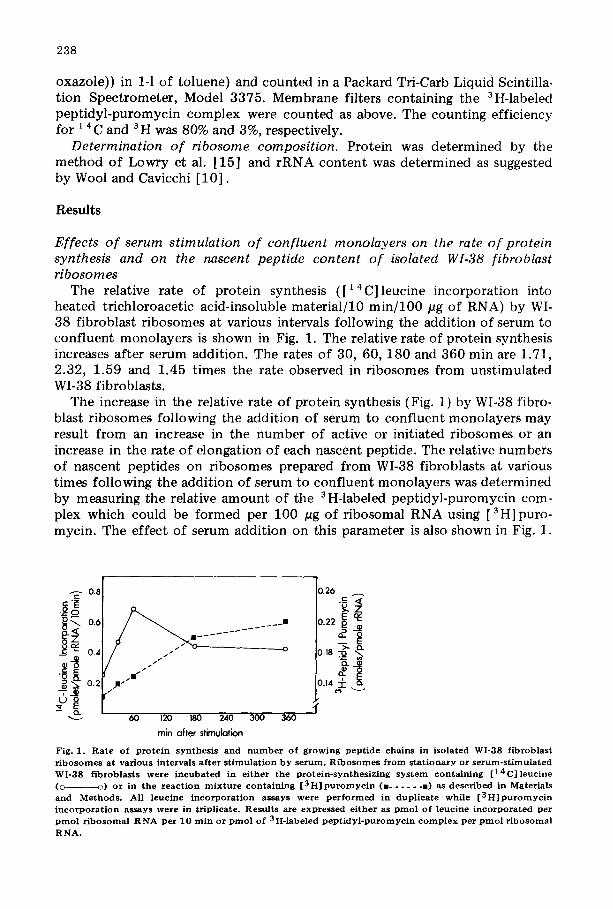
The relative rate of protein synthesis ([14C]leucine incorporation into heated trichloroacetic acid-insoluble material/10 min/100 pg of RNA) by WI- 38 fibroblast ribosomes at various intervals following the addition of serum to confluent monolayers is shown in Fig. 1. The relative rate of protein synthesis increases after serum addition. The rates of 30, 60, 180 and 360 min are 1.71, 2.32, 1.59 and 1.45 times the rate observed in ribosomes from unstimulated WI-38 fibroblasts.
The increase in the relative rate of protein synthesis (Fig. 1) by WI-38 fibro- blast ribosomes following the addition of serum to confluent monolayers may result from an increase in the number of active or initiated ribosomes or an increase in the rate of elongation of each nascent peptide. The relative numbers of nascent peptides on ribosomes prepared from WI-38 fibroblasts at various times following the addition of serum to confluent monolayers was determined by measuring the relative amount of the 3 H-labeled peptidyl-puromycin com- plex which could be formed per 100 pg of ribosomal RNA using [3H]puro- mycin. The effect of serum addition on this parameter is also shown in Fig. 1.
0.81
. n ~ ~PR ~
60 120 180 240 300 360
rnin after stimulation
0.26 .c_
/ °- ~
Fig. 1. R a t e of p r o t e i n s y n t h e s i s and n u m b e r o f g r o w i n g p e p t i d e chains in isola ted WI-38 f ibroblas t r i bosom es at var ious intervals a f ter s t i m u l a t i o n b y se rum. R i b o s o m e s f r o m s t a t i ona ry or s e r u m - s t i m u l a t e d WI-3S f ibrob las t s w e r e i n c u b a t e d in e i t h e r t h e p r o t e i n - s y n t h e s i z i n g s y s t e m con ta in ing [ 1 4 C ] l e u c i n e (o o) o r in t h e r e a c t i o n m i x t u r e con ta in ing [ 3 H ] p u r o m y c i n (a . . . . . . a) as d e s c r i b e d in Materia ls and Methods . All leucine i n c o r p o r a t i o n assays w e r e p e r f o r m e d in d u p l i c a t e w h i l e [ 3 H ] p u r o m y c i n i n c o r p o r a t i o n assays w e r e in t r ipl icate . Resul ts are e x p r e s s e d e i ther as p m o l of l e u c i n e i n c o r p o r a t e d per p m o l r i b o s o m a l R N A per 10 rain or p m o l of 3H-labeled p e p t i d y l - p u r o m y c i n c o m p l e x per p m o l r i b o so ma l RNA.

239
The number of growing peptides increases asymptotically. The formation of the labeled peptidyl-puromycin complex per pmol of ribosomal RNA increases from 0.125 pmol by control ribosomes to 0.223 pmol by 6-h serum-stimulated ribosomes. These results suggest that 12% of the r ibosomes from quiescent WI-38 fibroblasts are actively engaged in protein synthesis. Following serum stimulation for 6 h, the number of active ribosomes increases to 22%. It is possible that some nascent peptides are still located in the aminoacyl acceptor site of the ribosomes and unreactive to puromycin [16] . This would increase the percentage of active ribosomes, but probably would not affect the magni- tude of stimulation. Therefore, the increase in the relative rate of protein synthesis ([14 C] leucine incorporation into heated trichloroacetic acid-insolu- ble material/10 min/100 pg rRNA) is greater than can be accounted for by the increase in the number of active ribosomes {level of 3 H-labeled peptidyl-puro- mycin complex/100 pg rRNA) following 60 rain exposure to serum. Presum- ably at this time the rate of elongation of each nascent peptide is increased.
The effects of a 60-min serum exposure to confluent monolayers on these two parameters were reproducible with four different preparations of WI-38 fibroblasts. Compared to the control, the [14C]leucine incorporation was in- creased 1.71 + 0.0272-fold (mean + S.E.M.), while the [ 3 H] puromycin incor- poration was increased 1.10 + 0.037 fold {mean + S.E.M.). Therefore, at 60 min the relative increase in elongation rate (fold increase in [ ' 4 C] leucine incor- porat ion/fold increase in [ 3H] puromycin incorporation) of protein synthesis was increased 1.60 + 0.052-fold (mean + S.E.M.) over the control.
Effect o f inhibitor of the initiation reaction on the 60-min serum-induced increase in protein synthesis
The increase in the rate of protein synthesis observed in Fig. 1 could be explained if re-initiation of protein synthesis increases in the in vitro system containing isolated ribosomes from WI-38 fibroblasts exposed to serum for 60 min, relative to the rate of re-initiation by ribosomes from unstimulated WI-38 fibroblasts. The effect of 10 mM sodium fluoride or 0.02 mM aurintricarboxyl- ic acid on the cell-free protein synthesis system was investigated. These com- pounds have been shown to block specifically the initiation or re-initiation of protein synthesis in b o t h liver and reticulocyte cell-free systems [17--21] . In the absence of both inhibitors the ribosomes isolated from 60-min serum-stimu- lated cells had a leucine incorporation rate 1.88-fold higher than ribosomes isolated from control fibroblasts. In the presence of the inhibitors, NaF and aurintricarboxylic acid, the increases were 1.89- and 1.84-fold, respectively. The serum-induced increase was therefore not affected by the inhibitors in the incubation mixture, indicating that the increase in the rate of [14C]leucine incorporation into protein was not due to the preferential initiation or reinitia- t ion of protein synthesis in the in vitro protein-synthesizing system.
Effect of actinomycin D on the 60-min serum-stimulated increase in protein synthesis
RNA synthesis in WI-38 fibroblasts is stimulated l~y the addition of serum to confluent monolayers [5,6] . The question arises whether the increase in the rate of protein synthesis measured in the WI-38 fibroblast cell-free system is

240
dependent on previous RNA synthesis. The relationship between RNA synthe- sis and increased rate of protein synthesis by a 60-min exposure to serum was investigated by determining whether or not actinomycin D was capable o5 blocking the serum-induced increase in the rate of protein synthesis on isolated WI-38 fibroblast ribosomes.
Confluent monolayers of WI-38 fibroblasts were stimulated by the addition of fresh medium containing 10% fetal calf serum for 60 min, with or wi thout the addition of act inomycin D (10 pg/ml). This dose of act inomycin D inhibits RNA synthesis in WI-38 fibroblasts by more than 90% within 3 min of expo- sure. The antibiotic is wi thout effect on serum-induced stimulation of protein synthesis (data not shown).
Effect o f serum on WI-38 fibroblast ribosome-associated proteins Interactions between proteins and ribosomal nucleic acid are essential for the
process of protein synthesis. The effect of serum addition to the ribosome-asso- ciated proteins of confluent monolayers of WI-38 fibroblasts is shown on the sodium dodecyl sulfate-polyacrylamide gel electrophoresis profiles of Fig. 2. Nascent non-structural proteins were released and eliminated by prior incuba- tion of r ibosomes with puromycin. Serum addition for 60 min results in an alteration in the relative amounts of proteins associated with ribosomes in peaks 5 and 6. Whether or not these protein changes are responsible for the stimulation of elongation rate of protein synthesis is not known, but they do correlate with the increased rate of chain growth.
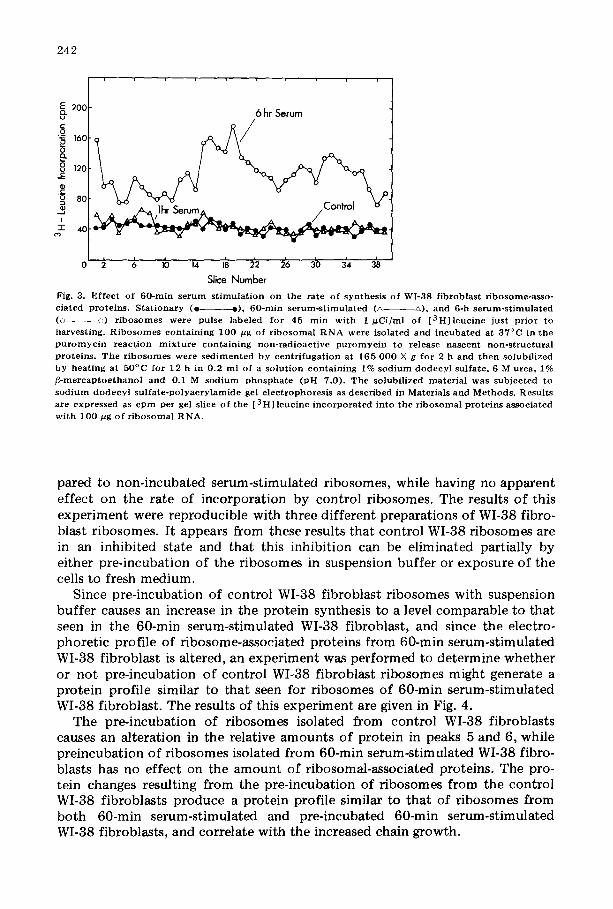
Changes in relative amounts of WI-38 fibroblast proteins following the addi- tion of serum to confluent monolayers may be due to an induced synthesis of these proteins. This question was investigated by studying the incorporation of [ 3 HI leucine into ribosome-associated proteins. The effect of serum addition on the rate of synthesis of WI-38 fibroblast ribosome-associated proteins (again nascent, nonstructural proteins were released by prior incubation of r ibosomes with puromycin) is shown in the sodium dodecyl sulfate-polyacrylamide gel electrophoretic profiles of Fig. 3. Serum addition to confluent monolayers for 60 min had no effect on the rate of incorporation of [~H]leucine into the proteins, but by 6 h after serum addition there was a significant increase in the rate of [ 3 H] leucine incorporation into ribosome-associated proteins. This t ime of synthesis o f proteins correlates well with the serum induction of ribosomal RNA synthesis [22] . These data indicate that the change in the relative amounts of ribosome-associated proteins after 60 min of serum addition is probably not due to a serum-induced increase in the rate of synthesis of specif- ic r ibosomal proteins.
Effects o f pre-incubation o f WI-38 fibroblast ribosomes with suspension buffer and conditioned medium
The increased rate of [14C]leucine by WI-38 fibroblast r ibosomes isolated from serum-stimulated cells may be due to the presence of a "s t imulator" or the removal of an inhibitor from fibroblast ribosomes. The in vitro effect of pre-incubation of WI-38 fibroblast r ibosomes at 37°C for 15 min with suspen- sion buffer (Tris/HC1 (pH 7.5), 50 mM; KC1, 100 mM; MgC12, 5 raM; ~-mer- captoethanol, 7 mM and sucrose, 250 mM) or with condit ioned medium (me-

241
, , , - - , , , ,
6 70 5/1 7 8
50
40
~ B . . . . 5 . . . .
.<~ 70t ~ 8 °°I t'l A
0.1 0.2 0.3 0,4 0.5 0.6 0.7 0.8 R F
Fig. 2 . E f f e c t o f s e r u m s t i m u l a t i o n o f t h e s o d i u m d o d e c y l s u l f a t e - p o l y a c r y l a m i d e gel e l e c t r o p h o r e t i c p ro f i l e o f WI-38 f i b r o b l a s t r i b o s o m e - a s s o c i a t e d p r o t e i n s . WI-38 f i b r o b l a s t r i b o s o m e s c o n t a i n i n g 1 0 0 #g r i b o s o m a l R N A i so la t ed f r o m s t a t i o n a r y (A) a n d 6 0 - r a i n - s t i m u l a t e d WI-38 cells (]3) w e r e i n c u b a t e d a t 3 7 ° C in t h e p u r o m y e i n r e a c t i o n m i x t u r e c o n t a i n i n g n o n - r a d i o a c t i v e p u r o m y c i n f o r 4 5 m i n to re lease n a s c e n t n o n - s t r u c t u r a l p r o t e i n s . T h e r i b o s o m e s were s e d i m e n t e d b y e e n t r i f u g a t i o n a t 1 6 5 0 0 0 X g f o r 2 h a n d t h e n so lub i l i zed b y h e a t i n g a t 5 0 ° C fo r 1 2 h in 0 .2 m l o f a s o l u t i o n c o n t a i n i n g 1% s o d i u m d o d e c y l su l f a t e , 6 M u r e a , 1% ~ - m e r c a p t o e t h a n o l a n d 0 .1 M s o d i u m p h o s p h a t e ( p H 7 .0 ) . T h e so lub i l i zed m a t e r i a l w a s s u b j e c t e d to s o d i u m d o d e c y l s u l f a t e - p o l y a c r y l a m i d e ~el e l e c t r o p h o r e s i s as d e s c r i b e d in Mate r i a l s a n d M e t h o d s .
dium removed from cells which had reached confluency) on the rate of protein synthesis by the subsequently re-isolated ribosomes (removed from the pre- incubation reaction by centrifugation at 165 000 × g for 2 h) was investigated. The pre-incubation of WI-38 fibroblast ribosomes (control and 60-min serum- stimulated) with suspension buffer eliminates the 60-min serum-induced in- crease in protein synthesis compared to the control rate of protein synthesis ([ ,4 C] leucine incorporation into heated trichloroacetic acid-insoluble materi- al/100 #g rRNA/10 min). The rate of protein synthesis by control ribosomes incubated in suspension buffer was 1.75 times that observed by non-incubated control ribosomes, while the pre-incubation has no effect on the rate of incor- poration by ribosomes isolated from WI-38 cells exposed to 10% serum for 1 h. Pre-incubation of WI-38 fibroblast ribosomes (control and 60-min serum-stimu- lated) in conditioned medium also eliminated the 60,min serum-induced in- crease in protein synthesis. The pre-incubation in conditioned medium caused a 65% decrease in the incorporation rate by serum-stimulated ribosomes corn-
242
200 6 hr Serum u
"~ 160
a
~ 40 -
t i i i i i i i i i i 0 2 6 10 14 18 22 26 30 34 38
Slice Number
Fig. 3. Ef fec t of 60-ra in s e r u m s t imu la t ion on the ra te of synthes is of WI-38 f ibroblas t r ibosome-asso- c ia ted prote ins . S t a t i o n a r y ( e e) , 60 -min s e r u m - s t i m u l a t e d (• •), and 6-h s e r u m - s t i m u l a t e d (o o) r i bosom es were pulse labeled for 45 min wi th 1 ~Ci /ml of [ 3 H ] l e u c i n e just p r io r to harves t ing. R i b o s o m e s con ta in ing 100 ~g of r i bos oma l R N A were isolated and i n cu b a t ed at 37°C in the p u r o m y e i n r e a c t i o n m i x t u r e con ta in ing non- rad ioac t ive p u r o m y c i n to release nascent non - s t ruc tu ra l p ro te ins . T he r i bosom es were s e d i m e n t e d b y een t r i fuga t i on at 165 0 0 0 × g for 2 h and t h e n solubi l ized by hea t ing at 50°C for 12 h in 0.2 ml of a so lu t ion con ta in ing 1% s o d i u m d o d e c y l sulfa te , 6 M urea, 1% j3-mercap toe thanol and 0.1 M s o d i u m p h o s p h a t e ( pH 7.0) . Th e solubi l ized ma te r i a l was sub jec ted to s o d i u m d o d e c y l s u l f a t e - p o l y a c r y l a m i d e gel e lec t rophores i s as descr ibed in Materials and Methods . Resul ts are expressed as c p m per gel slice of the [3 HI leucine i n c o r p o r a t e d in to the r i bosoma l p ro te ins associa ted wi th 100 pg o f r i b o s o m a l RNA.
pared to non-incubated serum-stimulated ribosomes, while having no apparent effect on the rate of incorporation by control ribosomes. The results of this experiment were reproducible with three different preparations of WI-38 fibro- blast ribosomes. It appears from these results that control WI-38 ribosomes are in an inhibited state and that this inhibition can be eliminated partially by either pre-incubation of the ribosomes in suspension buffer or exposure of the cells to fresh medium.
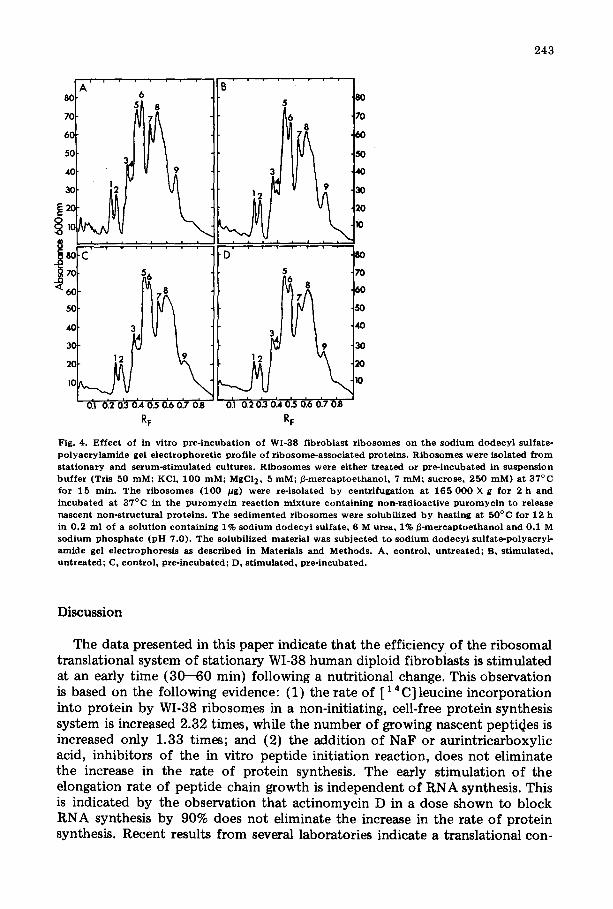
Since pre-incubation of control WI-38 fibroblast ribosomes with suspension buffer causes an increase in the protein synthesis to a level comparable to that seen in the 60-min serum-stimulated WI-38 fibroblast, and since the electro° phoretic profile of ribosome-associated proteins from 60-min serum-stimulated WI°38 fibroblast is altered, an experiment was performed to determine whether or not pre-incubation of control WI-38 fibroblast ribosomes might generate a protein profile similar to that seen for ribosomes of 60-min serum-stimulated WI-38 fibroblast. The results of this experiment are given in Fig. 4.
The pre-incubation of ribosomes isolated from control WI-38 fibroblasts causes an alteration in the relative amounts of protein in peaks 5 and 6, while preincubation of ribosomes isolated from 60-min serum-stimulated WI-38 fibro- blasts has no effect on the amount of ribosomal-associated proteins. The pro- tein changes resulting from the pre-incubation of ribosomes from the control WI-38 fibroblasts produce a protein profile similar to that of ribosomes from both 60-min serum-stimulated and pre-incubated 60-min serum-stimulated WI-38 fibroblasts, and correlate with the increased chain growth.
243
A B 80 6 $0
4¢ ~ J ~ ? 3 ~ 40 v ~ ]d
. ~ 8 o ' c . . . . . . . . . D . . . . . . . . .8o
~70 5, 5 . 70
• V
.50
, 0 I. O / :I,1 / 4 o
20 12 ~,9 . 12 v \ .20
oi~ o'.2 o~3 o'.4 o'.5 o'.~ o'.z o'.8 o.t o.2 o~ o~ o~ o.6 o.z o~ R F RF
Fig. 4. Ef fec t of in vi t ro p re - incuba t ion of WI-38 f ibroblas t r i b o so mes on t h e s o d i u m d o d e c y l sulfate- p o l y a c r y l a m i d e gel e l e c t ropho re t i c profi le of r ibosome-assoc ia t ed prote ins . R i b o s o m e s w e r e isolated f r o m s t a t i ona ry and s e rum-s t imu la t ed cul tures . R ibos ome s were e i ther t r ea t ed or p re - incuba ted in suspens ion b u f f e r (Tris 50 raM: KCI, 100 mM~ MgC12, 5 mMi ~ - m e r c a p t o e t h a n o l , 7 mM~ sucrose , 250 raM) a t 37°C for 15 rain. T he r i b o s o m e s (100 /~g) were re- isolated b y cen t r i fuga t i on at 165 0 0 0 × g for 2 h and i ncuba t ed a t 37°C in the p u r o m y c i n r eac t ion m i x t u r e con ta in ing non- rad ioac t ive p u r o m y c i n to release nascen t non-s t ruc t t t ra l pro te ins . T h e s e d i m e n t e d r i bos ome s were solubil ized b y hea t ing at 50°C for 12 h in 0 .2 ml of a so lu t ion con ta in ing 1% s o d i u m d o d c c y l sulfate , 6 M urea , 1% B-mexcap toe thano l and 0.1 M s o d i u m p h o s p h a t e ( p H 7.0). The solubi l ized ma te r i a l was sub jec ted to s o d i u m d o d e c y l su l fa te -po lyacry l - amide gel e lec t rophores i s as desc r ibed in Materials and Methods . A, con t ro l , unt rea ted~ B, s t imu la t ed , untzeated~ C, con t ro l , p re - incuba ted : D, s t imu la t ed , p re - incuba ted .
D i s c u s s i o n
The data presented in this paper indicate that the efficiency of the ribosomal translational system of stationary WI-38 human diploid fibroblasts is stimulated at an early time (30--60 min) following a nutritional change. This observation is based on the following evidence: (1) the rate of [14C]leucin e incorporation into protein by WI-38 ribosomes in a non-initiating, cell-free protein synthesis system is increased 2.32 times, while the number of growing nascent pepti~es is increased only 1.33 times; and (2) the addition of NaF or aurintricarboxylic acid, inhibitors of the in vitro peptide initiation reaction, does not eliminate the increase in the rate of protein synthesis. The early stimulation of the elongation rate of peptide chain growth is independent of RNA synthesis. This is indicated by the observation that act inomycin D in a dose shown to block RNA synthesis by 90% does not eliminate the increase in the rate of protein synthesis. Recent results from several laboratories indicate a translational con-

2 4 4
trol of protein synthesis either by an increase in the number of active ribo- somes or by increasing the rate of initiation and elongation of peptides [12,17--28].
Interactions between proteins and ribosomal nucleic acids are essential for the process of protein synthesis. Quantitative and qualitative changes in ribo- some-associated proteins have been reported in a number of mammalian sys- tems [29--35]. The biological significance of these alterations is not clear. This investigation showed that stimulation of WI-38 diploid fibroblasts by a nutri- tional change resulted in an alteration in the relative amounts of certain ribo- some-associated proteins. This alteration in the relative amount of these pro- teins can not be accounted for by an increased synthesis of fibroblast ribosome- associated proteins as measured by the rate of incorporation of [3H]leucine into the proteins of control and 60-min-stimulated WI-38 fibroblasts. Whether or not these alterations in the fibroblast ribosome-associated proteins are re- sponsible for the increased elongation rate of nascent peptide synthesis in 60-min-stimulated fibroblasts is not known, but they do correspond with the time of the response.
The experiments investigating the effects of preincubation of ribosomes from control and stimulated fibroblasts in conditioned medium (medium re- moved from stationary fibroblasts) suggest that ribosomes are maintained in a relatively inactive state in density-inhibited cultures. The catalytic activity (rate of elongation of nascent peptides) appears to be suppressed by a specific inhibi- tor(s), present both on the ribosome and in the culture medium. When station- ary cultures are stimulated by a nutritional change the inhibitor(s) may be removed, either by the direct action of a component(s) in the fresh medium, or by the establishment of a new concentration equilibrium between bound and free inhibitors. The existence of translational inhibitors of protein synthesis in other eukaryotic systems has been reported [17,35--43]. Whether or not these inhibitors regulate protein synthesis in similar ways remains to be established. In WI-38 fibroblasts there appears to be a correlation between increased elonga- tion rate, removal of an inhibitor and a change in the relative amount of certain ribosome-associated proteins. The actual role for the increased translational efficiency of protein synthesis remains uncertain, but the possibility exists that it represents synthesis of regulatory proteins or early completion of the pro- teins being synthesized so that pre-existing ribosomes are available for utiliza- tion of newly synthesized messenger RNA which is being released from the nucleus at this time.
Acknowledgments
This work was supported by U.S.P.H.S. Research Grant Ca-12923, and Training Grant (S.M.W.) CA-05222 from the National Cancer Institute.
References
1 Levine , E .M.Y. , Becke r , Y., B o o n e , C.W. a n d Eagle, H. ( 1 9 6 5 ) Proc , Nat l . A c a d . Sci. U.S. 53. 350--356
2 Rhode, III, S.L. and EIIem, K.A.O. (1968) Exp. Cell Res. 53, 184--204

245
3 Wiebel, F. and Baserga, R. (1969) J, Cell. Physiol. 74, 191--202 4 Rovera, G. and Baserga, R. (1971) J. Ceil. Physiol. 77 ,201 - -212 5 Father, J., Rovera, F. and Baserga, R. (1971) Bioehem. J. 122, 189--195 6 Bombik, B.M. and Baserga, R. (1974) Proc. Natl. Acad, Sci. U.S. 71, 2038--2042 7 Rovera, G., Farber, J, and Baserga, R. (1971) Proc. Natl. Acad. Sei. U.S. 68, 1725--1729 8 I-layflick, L. and Moorehead, P.S. (1961) Exp. Ceil Res. 25, 535---621 9 Rovera, G. and Baserga, R. (1973) Exp. Cell Res. 78, 118--126
10 Wool, I.G. and Cavicchi, P. (1967) Biochemistry 6, 1231--1242 11 Moldave, K. (1963) Methods Enzymol. 6, 757--761 12 Wool, I.G. and Kurihara, K. (1967) Proc. Natl. Acad. Sci. U.S. 58, 2401--2407 13 Suvatte, A. and I-Iagerman, D. (1970) Endocrinology 87 ,641- -645 14 Palmiter, R.D., Oka, T. and Sehimke, R.T. (1971) J. Biol. Chem. 246, 724--737 15 Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall, R.J. (1951) J. Biol. Chem. 193, 265--275 16 Baliga, B.S., Scheehtman, M.G., Nolan, R.D. and Munro, H.N. (1973) Biochem. Biophys. Aeta 312,
349--357 17 Lin, S.Y., Mosteller. R.D. and Hardesty, B. (1966) J. Mol. Biol. 21, 51--69 18 Shafritz, D.A., Laycock, D.G., Crystal, R.G. and Anderson, W,F. (1971) Proc. Natl. Acad. Sci. U.S.
68, 2246--2251 19 Stewart, M.L., Grollman, A.P. and Huang, M.T. (1971) Proc. Natl. Aead. Sci. U.S. 68, 97--101 20 Leader, D.P. (1972) FEBS Lett. 22, 245--247 21 Baglione, C., Jacobs-Lorena, M. and Meade, H. (1972) Biochim. Biophys. Acta 277 ,188- -197 22 Zardi, L. and Baserga, R. (1974) Exp. Mol. Pathol. 20, 69--77 23 Soeiro, R. and Amos, H. (1966) Science 154, 622--665 24 Becker, H., Stanners, P. and Kudlow, J.E. (1971) J. Ceil. Physiol. 77, 43--60 25 Kay, J.E., Ahem, T, and Atkins, M. (1971) Biochim. Biophys. Acta 331, 91--101 26 Comstock, J.P., Rosenfeld, G.C., O'Mailey, B.W. and Means, A.R. (1972) Proc. Natl. Acad. Sci. U.S.
69, 2377--2380 27 Palmiter, R.D. (1972) J. Biol. Chem. 247, 6670--6780 28 Wheily, S.M. and Barker, K.L. (1974) Biochemistry 13, 341--346 29 Hopkinson, J., Prichard, P.M. and Bresnick, E. (1974) Biochim. Biophys. Acta 374, 375--383 30 Engelhardt, D.L. and Sornaski, J. (1975) J. Cell. Physiol., in press 31 Rodgers, A. (1973) Bioehim. Biophys. Acta 294, 292--296 32 Kabat, D. (1970) Biochemistry 9, 4160--4175 33 Kabat, D. (1972) J. Biol. Chem. 247, 5338--5344 34 Liew, C.C. and Gbrnail, A.G. (1973) J. Biol. Chem. 248, 977--983 35 Eft, C. and Wool, I.G. (1973) J. Biol. Chem. 248, 5122--5129 36 Eft, C. and Wool, I.G. (1973) J. Biol. Chem. 248, 5130--5136 37 Anderson, W.M., Grundholm, A. and Sena, B.H. (1975) Biochem. Biophys. Res. Commun, 62,
669---676 38 Scornik, O.A., Hoagland, M.B., Pfefferkorn, L.C. and Bishop, E.A. (1967) J. Biol. Chem. 242,
131--139 39 Bechowsky-Slomnieki , L. (1969) Biochem. Biophys. Res. Commun. 37 , 750 - -756 40 Metafora, S., Feilcetti , L. and Gambeno, R. (1971) Proc. Natl. Acad. Sci. U.S. 68, 600--604 41 Engelhardt, D.L. (1971) J. Cell. Physiol. 78, 333--334 42 Hassell, J.A. and Engelhardt, D.L. (1973) Biochim. Biophys. Acta 325, 545--553 43 Adelman, T.G. and Lovett, J.S. (1974) Bioehim. Biophys. Acta 335, 236--245 44 Ulrich, F. (1974) Biochem. Biophys. Res. Commun. 60, 1453--1459
Copyright © 2022 FDOKUMEN




















