
Toolbox39b Final Report
29
Toolbox39b Project NADPH oxidases in symbiosis Students: Supervisors: Marjon Stel René Geurts Linda van Waalwijk van Doorn Alessandra Lillo Loan Ly Thi Phuong Anneke Hibbel Ruben Higler Max Friesen Nov-Dec 2009, Laboratory of Molecular Biology, Wageningen University
Transcript of Toolbox39b Final Report

Toolbox39b Project
NADPH oxidases in symbiosis
Students: Supervisors:
Marjon Stel René Geurts
Linda van Waalwijk van Doorn Alessandra Lillo
Loan Ly Thi Phuong
Anneke Hibbel
Ruben Higler
Max Friesen
Nov-Dec 2009, Laboratory of Molecular Biology, Wageningen University

2
Preface
In this report, Toolbox39b will discuss the results of seven weeks (plus one day) of experiments at
Molecular Biology in Wageningen, in November and December 2009, under supervision of René
Geurts and Alessandra Lillo.
The endosymbiosis between legumes and Rhizobium is being studied in research groups around
the world, and among them is Molecular Biology in Wageningen. Toolbox39b contributed during
the Toolbox course to this research, specifically in the role of NADPH oxidases in the symbiosis of
legumes and Rhizobium.
We gratefully thank René and Alessandra for their enthusiasm and encouragement.

3
Introduction
Rhizobium bacteria live in root nodules, organs formed by the plant specifically for the Rhizobium
to live in. These new organs are formed after a Rhizobium bacterium touches a root hair on the
root of the legume. The bacterium secretes oligosaccharides called Nod Factors. The plant
recognises these Nod Factors and a signal transduction pathway in the plant is initiated. The root
hair diverts its linear growth, growing around the bacterium.
When fully surrounded by the root hair, the bacterium forms an infection thread through the root
hair, surrounded by a plant cell wall. The infection thread will reach the cortical root cells, which
have already formed a nodule cells. The bacteria are released into the nodule cells, in a
mechanism that is not yet understood. In the nodule cells, they are encapsulated in a plant plasma
membrane, called the symbiosome. The bacterium, now inside the plant, differentiates and starts
to fix nitrogen gas from air. It transfers this fixed nitrogen to the plant in the form of ammonia. In
turn, the bacterium receives carbohydrates and other nutrients it needs from the plant (Giles et al.,
2008).
Emergence of legume-Rhizobium symbiosis
One of the questions raised about this symbiosis is, why legumes are able to form this symbiosis
with Rhizobium, while other plants can not? In evolution of plant species, when did the ability to
form root nodules for Rhizobium to live in emerge? What changes in genetics made it possible to
form nodules?
One hypothesis explains that in the evolution of legumes, a whole genome duplication (WGD)
occurred around the same time the legume family emerged. This WGD would have led to genes
gaining new functions, and a new organ (the root nodule) to be formed. These duplicated genes
would probably have functions specific to symbiosis with Rhizobium. This WGD would have taken
place approximately 60 million years ago. Many of the duplicated genes would have lost their
function, because of a lack of selection pressure to keep their function. Other duplicated genes
may have gained new functions compared to the original gene, and other gene pairs may have
specified their function, partially sharing it.
Our Toolbox started with this hypothesis. If gene pairs are retained after 60 million years, their
function may be important enough to keep. More importantly, they may fulfil a function in
symbiosis. We received data from a microarray analysis of Medicago truncatula from Benedito et
al. (2008) of 250 gene pairs thought to be leftovers from the WGD. We screened the expression
data of these gene pairs for genes that showed high expression in nodules, preferably with low
expression in other parts of the plant. We selected 18 of these genes. They have a variety of
functions, though for some no knowledge on function was available. Among these genes was one

4
NADPH oxidase. Since, at the Molecular Biology group, some research was already performed on
NADPH oxidases, we chose NADPH oxidases as the focus of this Toolbox.
NADPH oxidases
NADPH oxidases are membrane-bound proteins that produce reactive oxygen species (ROS).
Almost all eukaryotes have NADPH oxidases, in plants they are called respiratory burst oxidase
homologs (Rboh). The plant NADPH oxidase (see figure 1) consists of three major regions. First,
there is a trans-membrane part with six α-helices. Two of the helices contain histidine amino acids,
which bind the oxygen species. Second, the N-terminus part contains several regulatory sites,
among which are two EF-hands to which for example calcium can bind. Finally, there is a C-
terminus part with two binding sites, one for the co-factor FAD and one for NADPH. NADPH is the
reducing agent in this reaction and donates electrons to the oxygen compound, to produce ROS
(Sumimoto, 2008)
NADPH oxidases have several functions, most extensively studied in Arabidopsis thaliana and
Oryza sativa (rice). Their most well-known function is in the reaction to pathogens. The toxicity of
ROS comes to mind, a straightforward defence against pathogens. However, ROS may also play a
part in signal transduction pathways in defence mechanisms, and not just to kill pathogenic
bacteria or fungi. Pathogenic stress is not the only stress NADPH oxidases are involved in, abiotic
stress will evoke NADPH oxidase response as well. Arabidopsis thaliana RbohD and RbohF play
important roles in response to these biotic and abiotic stresses (Torres & Dangl, 2005).
Roles in developmental processes may be less obvious, but NADPH oxidases play important roles
in some of those processes. For example, AtRbohC is involved in root hair growth. The gene is
also called root hair deficient 2, or RHD2, which refers to the mutant rhd2. On the roots of this
mutant grow no root hairs. Another function of AtRbohC is the involvement in stomatal closure.
Mutants of AtRbohD, for example, were unable to close their stomata in case of drought (Torres &
Dangl, 2005; Takeda et al., 2008).
Figure 1: General structure of a plant NADPH oxidase. Three regions can be distinguished:
• Six trans-membrane α-helices. Two of the helices contain histidine amino acids, which bind the oxygen species.
• N-terminus regulatory region. Contains two EF-hands and other binding sites for regulation of the protein.
• C-terminus binding sites for FAD (cofactor) and NADPH (reducing agent).
Image from Sumimoto review 2008.

5
Regulation of NADPH oxidases occurs in a variety of ways. Their regulation depends on the
specific NADPH oxidase; we will concisely mention the most prevalent ways here. The EF-hands
on the regulatory part of the protein suggest that calcium ions can influence the activation of the
protein. But calcium dependent kinases also have an upstream function in regulation of NADPH
oxidases. Rac-GTPase can bind to the EF-hands as well, activating the NADPH oxidase.
Phospholipase Dα1 can regulate AtRbohD specifically through its product phosphatidic acid, which
binds to the regulatory region of the protein (Sumimoto, 2008). Interestingly, phospholipase Dα1
and phosphatidic acid play a role in Nod Factor signalling in legumes as well (Zhang et al, 2009).
Role of NADPH oxidases in symbiosis
The aim of this Toolbox was to investigate the role of NADPH oxidases in symbiosis of legumes
with Rhizobium. Thus, we return to Medicago truncatula, since Arabidopsis, some of which genes
we discussed earlier cannot undergo this symbiosis. The NADPH oxidase gene we selected from
the micro-array data, suggest that at least one gene from Medicago plays an important role in
nodulation. Research was already being done on NADPH oxidases at the Molecular Biology lab,
so we chose these genes to research in the Toolbox.
Medicago has six known NADPH oxidase genes, but not much is known about them yet. We
looked at more detailed expression analysis from the Gene Expression Atlas by Noble Foundation
[7] (see figure 2), though we could only find four of the six genes (these are called NADPH
oxidaseC, NADPH oxidaseM, NADPH oxidaseE and NADPH oxidaseF, we could not find NADPH
oxidaseI and NADPH oxidaseO). In the graph, we see the expression levels in certain processes
and plant tissues. We can see that NADPH oxidaseM has a low expression throughout the plant,
while NADPH oxidaseC has a very high expression throughout the plant. NADPH oxidaseF shows
an increased expression during nodule formation, and was also the gene we selected based on
the microarray from Benedito et al. (2008).

6
To investigate the role of NADPH oxidases in symbiosis of Medicago and Sinorhizobium meliloti
(the specific Rhizobium strain that can infect Medicago), we performed several experiments. We
will discuss these experiments in this report. Each experiment will be explained in its appropriate
section.
• Evolution of NADPH oxidases. We wanted to expand the phylogenetic tree that Stéphane de
Mita made of NADPH oxidases, to answer questions on the origin of nodulation and its relation to
other (non-)legumes.
• Localisation studies of NADPH oxidase E. Where is E expressed in the nodule?
• Expression studies of NADPH oxidase F. The specific expression pattern of NADPH oxidase F in
nodules.
• Functional analysis of NADPH oxidases F, C, M and O. Using RNAi on these genes, we tried to
see which function the NADPH oxidases have. We also tried to complement these genes with
Arabidopsis thaliana RbohC (or RHD2) in root hair growth.
Figure 2: Expression levels of four Medicago truncatula NADPH oxidases: C (in blue), E (in black), F (in pink) and M (in red). The NADPH oxidases I and O are missing from this graph, since there were no expression data available. A few points of note: M is not highly expressed throughout the plant and different processes, while C and E are. F shows a definite rise in expression during nodule formation.

7
1. NADPH oxidase-based evolution of legumes and nodulation
All flowering plants that have symbiotic Nitrogen-fixing symbioses fall in a part of Rosid 1 (Soltis et
al, 2000). In detail, nitrogen-fixing root nodule symbiosis occurs in two major forms: actinorhizal
plants and legumes (Fabales), which share a relatively close ancestor (Kistner & Parniske, 2002).
Legumes evolved about 60 millions years ago (Ma), while nodulation evolved 58 Ma ago (Sprent,
2006). Adopting the terminology of Lewis et al (2005), Leguminosae, rather than Fabaceae, is
used for the whole family, divided into three subfamilies: Caesalpinoideae, Mimosoideae and
Papilionoideae. It is believed that nodulation is uncommon in the caesalpinoideae, more common
in the mimosoideae and very common in the papilionoideae (Allen & Allen, 1981; Sprent, 2006)
(see Fig.3).
Figure 3. Phylogenetic tree showing the Rosid 1 clade.
It is known that many of the genes for nodulation have been acquired following duplication of
genes with related functions (Sprent, 2006). This statement is also supported according to the
phylogenetic tree made by Stephane de Mita (not published, Fig. 3). Following this, it is shown that
there are three important gene duplications of NADPH oxidase in legumes including soybean (Gm)
and Medicago (Mt), with the first duplication splitting the two branches of E/F group and C/M/I/O
NADPH oxidases. The duplication 2 and 3 were illustrated in the same time frame were maintained
in Mt and Gm. This observation is in line with the hypothesis in which the whole genome
duplications of the last common ancestor led to the speciation of Lotus, Medicago and Soybean
55-80 Ma. Nevertheless, there is no information on when these genes duplicated and whether
those events also took place in monocots and other sub-families of Leguminosae.
Rosales � Rosaceae
Malpighiales � Salecaceae � Poplar
Cucurbitales
Fabales
Oxalidales
Celastrales
Zygophyllales
Surianaceae
Quilajaceae
Polygalaceae
Leguminosae
Fagales
Rosid II � Brasicaceae � Arabidopsis
Vitis vinifera (wine grape)
Eurosid I
Cercidieae, Detarieae
Cassieae, Casealpiniieae
Mimosoideae
Papilionoideae
Casaelpinioideae � Chameacrista
� Medicago; Glycine

8
Additionally, there is no copy of RbohD (NADPH oxidase D) found in legumes. In Arabidopsis this
gene serves important roles in many biological activities including stomata closure regulation and
host defense against pathogen,. It is not known why RbohD was lost in legumes and how legumes
do their relevant functions without RbohD.
To gain insight into the roles of NADPH oxidase gene family in nodulation, our original phylogeny
was expanded, taking into account the NADPH oxidase homologs of Chameacrista, a symbiotic
nitrogen-fixing plant of Caeselpinioideae subfamily having the fixation threads in the active
indeterminate nodules, as well as, NADPH oxidase homologs of rice (Os), a well-studied species
of monocots (see Fig. 3). The resulting tree reveals that rice is supported to maintain several
copies of NADPH oxidases but no E/F and C/I/M/O duplicated copies are found in both
Chameacrista and rice. Moreover, the three duplications mentioned in Fig. 4 are shown to occur
after the rice speciation. The copies of F/E and C/I/M/O NADPH oxidases are not represented in
Chameacrista. Again, none Rboh was found in all three legumes (Chameacrista, Glycine and
Medicago). Following this, it is postulated that RBoh gene occurred before the last ancestor split
the non-legumes and legumes. From that point, the question is raised about whether the
duplicated genes found in Papillionoideae legumes (E/F and C/O/M/I NADPH oxidases) are
important for nitrogen fixing symbiosis. To tackle this problem, some functional analyses were
carried out to study the sub-cellular localization of NADPH oxidase E, followed by the experiment
of expression of NADPH oxidase F and RNAi knockdown of several NADPH oxidase genes.
Finally, the Arabidopsis rhd2 complementation study was conducted, with rhd2 as a very close
homologs of Medicago NADPH oxidases.

9
Figure 4. The original NADPH oxidase-based cladogram represented the evolution of some selected species
(soybean – Gm, Medicago – Mt, Poplar - Pt) involving in nitrogen-fixing symbiosis with the outgroup of Vitis
(Vv) and Arabidopsis (At) (Stephane de Mita, 2009). The three duplication events that led to duplicated
genes of C, E, F, I, M and O are marked as 1 to 3.
F
C
M
I O
1
2
3
RHD2
E

10
Figure 5. The NADPH oxidase-based dendogram extended from the original phylogeny with the rice (Os)
and Chameacrista (Cf) NADPH oxidase homologs. The studied genes are labeled as C, E, F, I, M and O.
I M
C O
RHD2
E
F

11
2. Sub-cellular localization studies of Mt NADPH oxidase E
In this localization study we used a GFP fusion construct consisting of a promoter region, GFP
gene and our NADPH oxidase of interest. Using confocal studies we found localization of the
protein in the plasma membrane of root hairs and uninfected nodule cells. Furthermore we found a
specific localization of gene E at the polar ends of the symbiosomes in infected nodule cells. This
specific localization has been confirmed using indirect immunostaining on the GFP part of the
fusion.
Sub-cellular localization
The Mt NADPH oxidase E gene is a member of a large family of NADPH oxidases in Medicago
Truncatula (Mt). We selected this gene for study based on its expression pattern as shown in
figure 61. Its profile is specifically interesting for its wide spectrum of expression in plant tissue, with
the exclusion of seed tissue, and its relative high levels of expression. Localization studies using
GFP fusions had already been tried with this protein2, however at the time only low GFP signals
had been observed. So low in fact, that it was very difficult to distinguish between real GFP signals
and artifact signals.
Therefore we decided to repeat the experiment to see whether something went wrong in the
previous experiment, or to confirm the low signals and possibly come up with a reason and
solution for the problem.
Figure 6. Expression pattern of NADPH oxidase E
1 http://bioinfo.noble.org/gene-atlas/v2/probeset.php?id=Mtr.32307.1.S1_s_at&submit=Go

12
To execute these studies we used a construct consisting of a 3kb region upstream of the NADPH
oxidase E gene which we labeled as its promoter region, the GFP gene and finally the NADPH
oxidase E gene. For the selection of transgenic roots we used DsRed expression.
The 3kb region we labeled as promoter consisted of the actual promoter region of the gene, the
transcriptional start site and the 5’-UTR. As control an empty vector carrying DsRed was used.
The vectors were constructed using Multisite Gateway® methods into a destination vector
specifically designed for transformation of Mt roots using Agrobacterium rhizogenes
After 7 days we tested for transgenic roots as can be seen in figures 7 and 8.
Sub-cellular localization in root hairs
Using confocal microscopy we tried to determine the sub-cellular localization of NADPH oxidase E
in root hairs. We found weak signals which localized the protein in the plasma membrane, at the
root hair tip and along its sides (figure 9). This observation was strengthened by the fact that one
of the root hairs we observed was in plasmolysis (figure 10). The GFP signal followed the structure
Fig. 7: Transgenic control roots Fig. 8: Transgenic GFP fusion roots
Fig. 9 Localization in root hairs Fig.10 Root hair in plasmolysis

13
of the plasma membrane, suggesting the E gene is indeed localized in the plasma membrane.
Although, it should be noted that just as in previous studies, our GFP signals were quite low.
Checking expression levels using qPCR
To make sure that our construct was actually being expressed and that we weren’t seeing artificial
signals in our localization studies we decided to compare expression levels of wild type NADPH
oxidase E and our GFP fusion gene. We designed 2 primer sets (see figure 11), one to detect our
fusion gene and one to detect the WT gene. The fusion set had a ~200b amplicon specifically set
on the GFP part of our fusion. The WT set had its forward primer on the 5‘UTR and it reverse
primer on the NADPH oxidase E gene part of our construct. This way it should only amplify when
the GFP gene is absent, aka the WT gene. It should work because the amplicon with the GFP
included would be too large for qPCR to amplify and thus detect.
The qPCR was performed on a cDNA sample from 10 transgenic roots from 10 different plants.
GADPH and Actin2 were used as reference genes. Unfortunately the melting curves of our primers
sets showed that the set we used to detect the WT gene had more than one amplicon rendering it
unusable for further analysis. Because of this we were only able to confirm that our construct is
indeed expressed in our transgenic roots. We were not able to compare this to WT expression.
We found a fifteen fold increased level of expression compared to our control plants, whose signal
is most likely just background noise. (Figure 12)
NADPHox E prom GFP NADPHox E gene
Fig.11: qPCR primers

14
Fig. 12. Q-PCR results on transgenic roots, confirming that our construct is expressed in our transgenic
roots.
Sub-cellular localization in nodule (9 dpi)
The second part of our localization studies consisted of determining the position of NADPH
oxidase E in nodules. For this we used nodules 9 dpi with RFP-Rhizobium, to make sure that there
is a contrast between bacteroid/symbiosomes signal and protein signal.
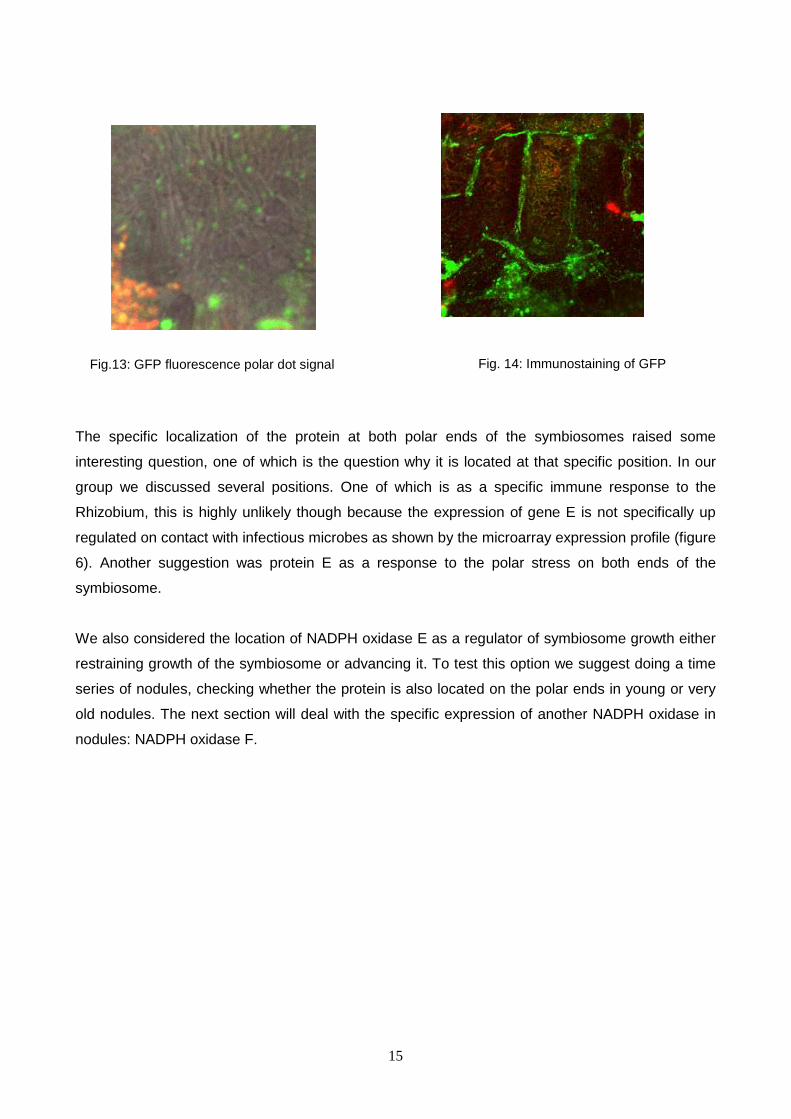
Using a confocal microscope we were able to locate the protein in a rather surprising position,
namely at both polar ends of the symbiosomes (Figure 13) This position had been seen in earlier
experiments which located ROP proteins, which interact with NADPH oxidases (Wong et al, 2007).
Thus that experiment indirectly also located NADPH oxidases.
To confirm that we were actually looking at a GFP signal and not some sort of artificial signal an
indirect immunostaining was performed3 on a different nodule from the same plant to check the
dot-localization still showed, (figure 14) which it did. Furthermore, apart from the amplified specific
signal due to immuno techniques, GFP immuno signal was also clearly visible in uninfected cells
where we couldn’t see a GFP fluorescence signal. This difference in signal strength made us ask
the question why the GFP signal of a highly expressed protein was so very weak. A few
possibilities were raised. A suggestion was raised that the FADH binding part of the protein might
quench the GFP fluorescence or that the fusion with GFP caused a misfolding which rendered the
protein unusable. As a third option we thought that the GPF could interfere with a locator peptide
signal in the WT NADPH ox E gene disrupting the proper localization of the protein.
3 By Alessandra Lillo
15
The specific localization of the protein at both polar ends of the symbiosomes raised some
interesting question, one of which is the question why it is located at that specific position. In our
group we discussed several positions. One of which is as a specific immune response to the
Rhizobium, this is highly unlikely though because the expression of gene E is not specifically up
regulated on contact with infectious microbes as shown by the microarray expression profile (figure
6). Another suggestion was protein E as a response to the polar stress on both ends of the
symbiosome.
We also considered the location of NADPH oxidase E as a regulator of symbiosome growth either
restraining growth of the symbiosome or advancing it. To test this option we suggest doing a time
series of nodules, checking whether the protein is also located on the polar ends in young or very
old nodules. The next section will deal with the specific expression of another NADPH oxidase in
nodules: NADPH oxidase F.
Fig.13: GFP fluorescence polar dot signal Fig. 14: Immunostaining of GFP
16
3. Expression of Mt NADPH oxidase F in nodules
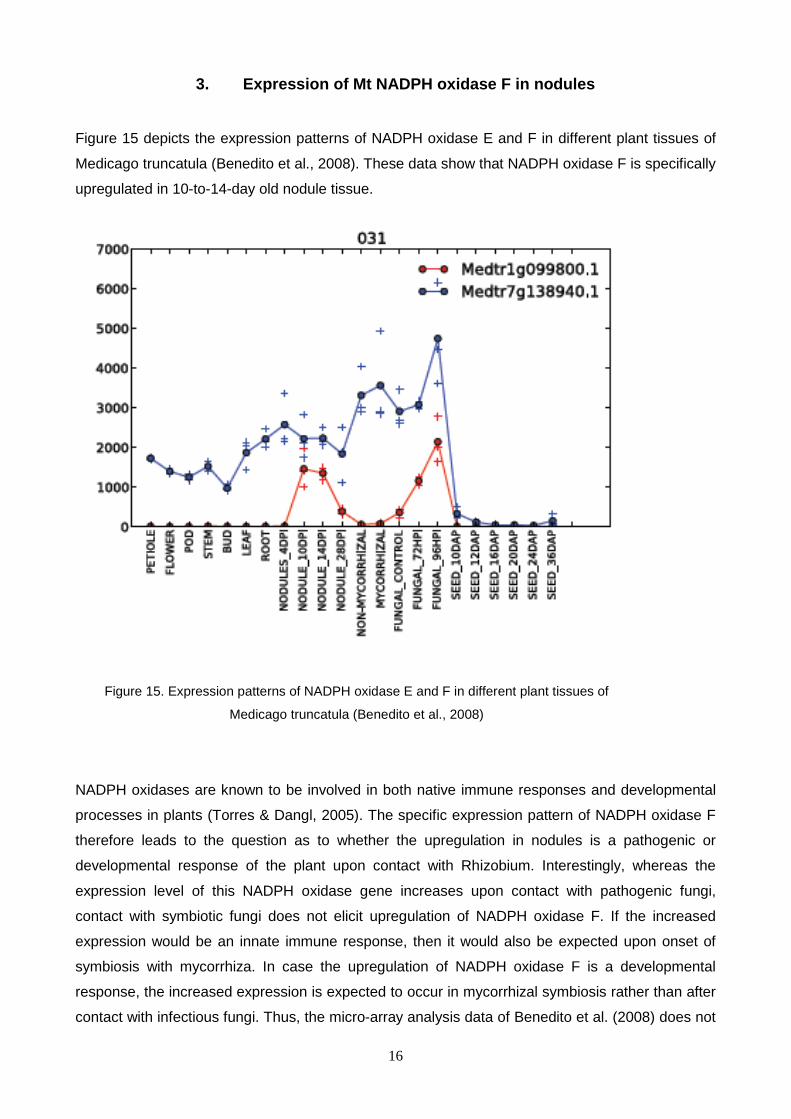
Figure 15 depicts the expression patterns of NADPH oxidase E and F in different plant tissues of
Medicago truncatula (Benedito et al., 2008). These data show that NADPH oxidase F is specifically
upregulated in 10-to-14-day old nodule tissue.
Figure 15. Expression patterns of NADPH oxidase E and F in different plant tissues of
Medicago truncatula (Benedito et al., 2008)
NADPH oxidases are known to be involved in both native immune responses and developmental
processes in plants (Torres & Dangl, 2005). The specific expression pattern of NADPH oxidase F
therefore leads to the question as to whether the upregulation in nodules is a pathogenic or
developmental response of the plant upon contact with Rhizobium. Interestingly, whereas the
expression level of this NADPH oxidase gene increases upon contact with pathogenic fungi,
contact with symbiotic fungi does not elicit upregulation of NADPH oxidase F. If the increased
expression would be an innate immune response, then it would also be expected upon onset of
symbiosis with mycorrhiza. In case the upregulation of NADPH oxidase F is a developmental
response, the increased expression is expected to occur in mycorrhizal symbiosis rather than after
contact with infectious fungi. Thus, the micro-array analysis data of Benedito et al. (2008) does not

17
uniformly support either of the hypotheses mentioned above. Insights in the functional role of the
upregulation of NADPH oxidase F between ten and fourteen days might be achieved by the
assessment of nodule-specific developmental processes in this time frame. Therefore, three
experiments were designed to study the expression of NADPH oxidase F in more detail.
GUS reporter study
A GUS reporter system study was conducted to localize promoter activity in WT Medicago
truncatula roots and nodules. We used A. rhizogenes to transiently transform the plants. Table 1
shows which constructs were made and the results obtained from this experiment. Due to time
limitations, this experiment could not be conducted to completion.
Constructs Results
NADPH oxidase_C::GUS Transgenic callus, no roots
NADPH oxidase_M::GUS Transgenic callus, no roots
NADPH oxidase_I::GUS Transgenic callus, no roots
NADPH oxidase_O::GUS Unable to amplify with PCR
NADPH oxidase_F::GUS No time to transform plants
NADPH oxidase_E::GUS Unable to amplify with PCR
Table 1. GUS reporter study on NADPH oxidases in Medicago truncatula
Time series experiment
A time series experiment was conducted to obtain a detailed assessment of the expression of
NADPH oxidase F in developing nodules. At the time of 0, 4, 6, 8, 10 and13-day- post-inoculation
with GFP-tagged Rhizobium, WT plants were scored for infection threads and nodule primordia,
using a stereomacroscope with a GFP filter. To determine the developmental stage of the nodules,
we intended to imbed nodules in a fixative wax for sectioning to create slides for microscopy. In
order to determine NADPH oxidase F expression levels in the roots and nodule primordia, the root
material was obtained at t=6 and t=8 for Q-PCR analysis. The images taken at t=4 and t=6 (Figure
16) show that GFP-tagged microbes are present on the root hairs of the Medicago plants.
However, in this experiment no symbiosis between Medicago truncatula and Rhizobium bacteria
did occur. Therefore, we were unable to obtain data on development and RNA expression levels in

18
nodules. Several reasons might explain this negative result. Firstly, the plants were too young at
the time of transplantation to perlite. However, since 17 days after inoculation still no nodulation
was observed, this hypothesis may be rejected. Secondly, it was speculated that the Rhizobium
strain used in the experiment was not efficiently infectious enough. The fact that in other
experiments nodulation was induced with the same bacterial strain counters this second
hypothesis. Finally, the nitrate concentration in the Färreaus medium might have been too high,
obstructing the symbiosis between Rhizobium and Medicago.
A. B.
C. D.
Figure 16. Stereomacroscopic images of Medicago truncatula roots inoculated with GFP-tagged Rhizobium.
A.) GFP image of tagged bacteria on root hairs. B.) Long-pass filter image of GFP-tagged Rhizobium. C.)
Possible infection thread in root hair. D.) Unidentified mass surrounding Medicago root.
NADPH oxidase F expression in mutants
To compare the NADPH oxidase F expression levels between WT and mutants, a study was set
up to perform Q-PCR on the nodule RNA obtained from WT plants in which bacteria do not leave
the infection thread and plants with empty nodules. WT plants transiently transformed with the HK*

19
construct (empty nodules) did not show transgenic callus, whereas dmi3 mutants transiently
transformed with poplar DMI3 showed transgenic roots without nodule primordia. It was therefore
not possible to obtain RNA for Q-PCR analysis.
4. Functional analysis of Mt NADPH oxidases
To find the function of our Medicago Truncatula genes, we designed two separate experiments to
test what functions they are actually responsible for in a plant. One of these is making an RNAi
knockdown for some of the genes and examining the phenotype of the plant to see if any of the
genes is responsible for a visible change in plant phenotype. The other experiment is determining
whether seeing if we can complement an Arabidopsis RHD2 (root hair deficient) mutant with one of
our Medicago genes. This is to find out if any of the Medicago genes has the function or part of the
function of the RbohC gene in Arabidopsis.
RNAi knockdown
For our RNAi knockdown experiment we received several constructs from Sjef Boeren, and we
made two of our own. The two we made ourselves were against NADPH oxidase gene F with a
constitutive 35S promoter, and we used two different RNAi fragments to try and silence the gene.
We transformed plants with these two constructs, and we have seen transgenic calluses forming,
but there was not enough time to let the plants grow enough to have clear transgenic roots visible.
The RNAi constructs we received from Sjef (Fig. 17) target three different genes, namely our
Medicago genes NADPH oxidase C, M and O. We also have a construct that targets all three
simultaneously, which is possible due to a perfectly homologous sequence in the three genes. All
of these have two different versions, one with the 35S promoter and the other with a nod-factor
induced Enod12 (from now on called E12) promoter.
Fig. 17. RNAi knockdown constructs against NADPH oxidase C and CMO
We transformed these into plants, grew them until we saw transgenic roots and applied nod-factors
to the roots. Next we scored for several things, including continued root growth, root hair growth, if
there was an absence of root hairs at the location of nod-factor appliance, primordia and if the

20
roots were red colored, probably due to flavonoids. We did not score every construct for these
attributes.
Root growth Root hair
growth
Root hair gap
at location of
nod-factor
appliance
Primordia Red
coloring
Control roots 15/17 16/17 7/17 3/17 15/17
35S:CMO RNAi 19/20
Impaired growth
17/19 7/19 3/19 11/19
E12:C RNAi 7/7 ? 4/7 2/7 6/7
35S:C RNAi 0/20 n.a. n.a. n.a. n.a.
Table 2: Scoring of different attributes of plants after appliance of nod-factors.
As had already been determined by others 4, the 35S:C RNAi construct was lethal. We confirmed
this, getting no transgenic roots at all for this line of plants.
Our E12:C RNAi line however did show transgenic roots, although even after application of nod-
factors which is supposed to activate the promoter, leading to an RNAi knockdown, we do not see
any difference from our control plants.
Our 35S:CMO construct does show an altered phenotype however, as it clearly shows impaired
growth after applying nod-factors. This must mean that one of the NADPH oxidase genes C, M or
O is involved in root growth. The interesting thing is that it does not seem to have any effect before
applying the nod-factors, so one of these genes, or a combination thereof only seems important for
root growth after an induced ‘stress’ situation of our treatment.
4 By Sjef Boeren

21
Figure 18: Transgenic roots of our RNAi fragments

22
Figure 19. Our plants after applying nod-factors, not showing a clear phenotype
Picture 20: A 35S:CMO transgenic root after appliance of nod-factor. It clearly shows impaired growth,
growing nearly nothing after treatment (at the black line).

23
Arabidopsis complementation
The Arabidopsis complementation experiment was meant to show if one of our Medicago genes
has the function that the RbohC gene has in Arabidopsis. RbohC, also called RHD2, is mostly
involved in root hair growth. We used a RHD2 mutant Arabidopsis line, leading to a root hair
deficient line of plants. It is however not an absolute mutant, so you still see some root hairs,
although heavily impaired in their development. We had to pick homozygote mutants from a
number of F1 seeds we received.
Figure 21: The difference between wild type (left) and RHD2 mutant (right) plants
Using the Multisite Gateway® technique, we constructed several vectors containing the RHD2
promoter from Arabidopsis itself driving a fusion between GFP and a gene of our interest. We used
our four NADPH oxidase genes from Medicago, these were E, F, M and O. We also made a
control construct with the RHD2 promoter and the normal RHD2 gene from Arabidopsis fused to
GFP. We transformed these into plants and examined the transgenic roots for root hairs. Sadly
enough none of our transgenic roots showed significant complementation, not even our control line
of plants. This might have several reasons, possibly our constructs including the control were not
correct, or the gene fusion with GFP interrupts its function.

24
Figure 22: Our Arabidopsis plants. This picture is representative of all our results, a transgenic root next to a
non-transgenic root showing no difference in root hairs. (This is the RHD2 control construct)

25
Conclusions and discussion
We come back to our initial question about the role NADPH oxidases play in symbiosis of
Medicago truncatula and Sinorhizobium meliloti. First, we discussed the possible evolution of
Medicago NADPH oxidases, compared to other plants, such as Chamaecrista and Arabidopsis.
Fig. 23. Phylogenetic tree with genome duplications indicated at the arrows.
In this simplified tree (Fig. 23), we see that there are three whole genome duplications. But which
duplication is important for the symbiosis between legumes and Rhizobium? If the emergence of
symbiosis was a single event, then none of the WGD is important for the symbiosis we studied. If it
is not a single event, but several events lead to the emergence of symbiosis, then the duplications
are important. From this tree we conclude that a whole genome duplication did occur in the
papillinoid lineage. Inspecting the tree more closely, we observe that in whole genome duplication
2, there is a splitting of two groups: legumes and non-legumes.
Unexpectedly, the RbohD gene is inherited only by non-legumes and the gene is lost in legumes.
RbohD has an important function in defense mechanism against biotic and abiotic stresses and in
a developmental role of stomatal closure (Torres & Dangl 2005), so this raises the question why it
would be lost after a gene duplication. A reason for this loss could be that there are other genes
that have a similar function or similar functions, making this gene redundant.
Another point of discussion is the moment where the RbohD gene was lost in legumes. There are
two possible explanations. The first one is that the loss of RbohD occurred prior to the split of
Chamaecrista and Soybean/Lotus/Medicago. The second explanation is that the RbohD was lost
after the split of Chamaecrista, Medicago and other legumes. Chamaecristae do not have the
NADPH oxidaseidase C/M/O/I gene, but uses the same initiation process for making nodules as

26
Medicago. We conclude that the duplication of NADPH oxidases was non-essential for initiation of
nodulation.
In the GFP localization studies, we observed the same symbiosomes in NADPH oxidaseE:GFP
visualization as Erik in his Rop2:GFP studies. The symbiosomes have only at the polar ends some
signal of GFP. From this we conclude that NADPH oxidaseE and Rop2 have the same localization,
so their functions are correlated, which is supported by the article “Regulation of rice NADPH
oxidaseidase by binding of Rac-GTPase to its N-terminal extension” (Rac is Rop2).
Figure 24: GFP localization of Rop2 done by Erik, symbiosomes with GFP-signal at the ends
Figure 25: GFP localization of NADPH oxidase E, symbiosomes with GFP-signal at the ends

27
The signal of GFP fused to NADPH oxidaseE only at the end of (older) symbiosomes is an
interesting occurrence. This unique observation of GFP-signals solely at the end of the
symbiosomes can be explained by the possible functions of NADPH oxidaseE: it might be an
immune response, a mechanical stress response, the blocking of elongated long symbiosomes or
(polar) growth.
Growth of the symbiosomes, or specifically polar growth, could be a very plausible explanation,
because a similar NADPH oxidase activity occurs when a root tip is growing. Further research
should be performed to clarify this.
Future experiments have to be performed to get a broader and more accurate view on the
localization and function on NADPH oxidases. Experiments which will be conducted by Anneke
Hibbel, one of the students in Toolbox39b:
1. Study the NADPH oxidase E localization in the DNF1 mutant. The symbiosomes are
formed in this mutant, but stay small.
2. Study the NADPH oxidase E localization in a mutant where bacteria can not be released.
3. Study the localization and regulation of NADPH oxidase F in new time series.
a) NADPH oxidase F is specifically expressed during symbiosis (10 day old nodules)
b) Study expression in different mutants (HK*, DNF1, Poplar DMI3 nodules)

28
References
Allen ON & Allen EK. (1981) The Leguminosae: a source book of characteristics, uses and
nodulation. Madison, WI, USA/London: University of Wisconsin Press/Macmillan.
Benedito V. et al. (2008). A gene expression atlas of the model legume Medicago truncatula.
Plant J. vol. 55 (3) pp. 504-13
Oldroyd GED. and Downie JA. (2008) Coordinating Nodule Morphogenesis with Rhizobial
Infection in Legumes, Annu. Rev. Plant Biol. 59:519–46
Kistner C & Parniske M. (2002) Evolution of signal transduction in intracellular symbiosis.
Trends in Plant Science. 7(11):511-518.
Lewis G, Schrire B, Mackinder B, Lock M, eds. (2005) Legumes of the world. London: Royal
Botanic Gardens Kew.
Soltis DE, Soltis PS, Chase ME, Mort ME, ALbach DC, Zannis M, Savolainen V, Hahn WH, Hoot
SB, Fay MF, Axtell M, Swensen SM, Prince LM, Kress WJ, Nixon KC, Farris JS. (2000)
Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Botanical Journal
of Linnaean Society. 133: 281-461.
Sprent JI. (2007) Evolving ideas of legume evolution and diversity: a taxonomy perspective
on the occurrence of nodulation. New Phytologist. 174: 11-25.
Sumimoto H. (2008) Structure, regulation and evolution of Nox-family NADPH oxidases that
produce reactive oxygen species, FEBS Journal 275: 3249–3277
Takeda S. et al (2008) Local Positive Feedback Regulation Determines Cell Shape in Root
Hair Cells, Science 319, 1241

29
Torres MA and Dangl JL. (2005) Functions of the respiratory burst oxidase in biotic
interactions, abiotic stress and development. Current Opinion in Plant Biology (2005) 8:397–
403
Wong HL, Pinontoan R, Hayashi K, Tabata R, Yaeno T, Hasegawa K, Kojima C, Yoshioka H, Iba
K,Kawasaki T et al. (2007) Regulation of rice NADPH oxidase by binding of Rac GTPase to its
N-terminal extension. Plant Cell 19, 4022–4034.
Zhang Y et al, (2009) Phospholipase Da1 and Phosphatidic Acid Regulate NADPH Oxidase
Activity and Production of Reactive Oxygen Species in ABA-Mediated Stomatal Closure in
Arabidopsis, The Plant Cell
Medicago truncatula Gene Expression Atlas by Noble Foundation, data extracted November
2009, http://bioinfo.noble.org/gene-atlas/v2/




















