
TNFα enhances the motility and invasiveness of prostatic cancer cells by stimulating the expression...
6
TNFa enhances the motility and invasiveness of prostatic cancer cells by stimulating the expression of selective glycosyl- and sulfotransferase genes involved in the synthesis of selectin ligands Prakash Radhakrishnan a , Vishwanath Chachadi a , Ming-Fong Lin a,b , Rakesh Singh b,c , Reiji Kannagi d , Pi-Wan Cheng a,b,⇑ a Department of Biochemistry and Molecular Biology, College of Medicine, University of Nebraska Medical Center, Omaha, NE 68198, USA b Eppley Cancer Center for Research in Cancer and Allied Diseases, University of Nebraska Medical Center, Omaha, NE 68198, USA c Department of Pathology and Microbiology, University of Nebraska Medical Center, Omaha, NE 68198, USA d Aichi Cancer Center, Nagoya, Japan article info Article history: Received 26 April 2011 Available online 8 May 2011 Keywords: LNCaP cells sLe x TNFa Glycogenes Motility and invasion abstract Sialyl Lewis x (sLe x ) plays an important role in cancer metastasis. But, the mechanism for its production in metastatic cancers remains unclear. The objective of current study was to examine the effects of a pro- inflammatory cytokine on the expression of glycosyltransferase and sulfotransferase genes involved in the synthesis of selectin ligands in a prostate cancer cell line. Androgen-independent human lymph node-derived metastatic prostate cancer cells (C-81 LNCaP), which express functional androgen receptor and mimic the castration-resistant advanced prostate cancer, were used. TNFa treatment of these cells increased their binding to P-, E- and L-selectins, anti-sLe x antibody, and anti-6-sulfo-sialyl Lewis x anti- body by 12%, 240%, 43%, 248% and 21%, respectively. Also, the expression of C2GnT-1, B4GalT1, Glc- NAc6ST3, and ST3Gal3 genes was significantly upregulated. Further treatment of TNFa-treated cells with either anti-sLe x antibody or E-selectin significantly suppressed their in vitro migration (81% and 52%, respectively) and invasion (45% and 56%, respectively). Our data indicate that TNFa treatment enhances the motility and invasion properties of LNCaP C-81 cells by increasing the formation of selectin ligands through stimulation of the expression of selective glycosyl- and sulfotransferase genes. These results support the hypothesis that inflammation contributes to cancer metastasis. Ó 2011 Elsevier Inc. All rights reserved. 1. Introduction Prostate cancer is the second leading cause of cancer death of men in the United States [1]. At the time of initial diagnosis, more than 70% of prostate cancer patients have lymph node metastases [2–5]. Although metastasis is the primary cause of the fatalities of cancer patients, the mechanism is not completely understood. One key step in the cancer metastatic process is the adhesion of circu- lating cancer cells to the endothelium at distant sites. This step in- volves selectin-ligand interactions between host cells (selectins) and cancer cells (selectin ligands). Understanding how the produc- tion of these two key players is regulated during cancer progres- sion could help develop cancer therapeutic strategy. Selectins are a family of type II transmembrane proteins found on the surface of leukocytes, platelets and activated endothelial cells. They are calcium-dependent C-type lectins [6]. The three major types of selectins include E-, P-, and L-selectins, which are located on endothelium, platelets and activated endothelium, and leuko- cytes, respectively [6]. The naturally occurring vascular carbohy- drate ligands for these selectins include E-selectin ligand-1 (ESL-1), P-selectin glycoprotein ligand-1 (PSGL-1), CD34, cutaneous lymphocyte-associated antigen (CLA), glycoprotein GlyCAM-1, MAdCAM-1, Podocalyxin-like protein and Sgp200 [7]. These selec- tin-ligand interactions were originally shown to be involved in the trafficking of immune cells to inflamed tissues [8] and lymphoid or- gans, and recently in the metastasis of cancer cells [9]. The naturally occurring vascular ligands for these selectin molecules are mostly mucin-type glycoproteins which carry sialylated, fucosylated and sulfated glycans [10] although some N-linked glycans also can par- ticipate in this process [11]. Increased expression and altered glyco- sylation of mucins are prominent features of carcinoma progression [10]. For example, sialyl Lewis x (sLe x ) and sialyl Lewis a (sLe a ) are 0006-291X/$ - see front matter Ó 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2011.05.019 ⇑ Corresponding author. Address: Department of Biochemistry and Molecular Biology, College of Medicine, 985870 Nebraska Medical Center, University of Nebraska Medical Center, Omaha, NE 68198-5870, USA. Fax: +1 402 559 6650. E-mail address: [email protected] (P.-W. Cheng). Biochemical and Biophysical Research Communications 409 (2011) 436–441 Contents lists available at ScienceDirect Biochemical and Biophysical Research Communications journal homepage: www.elsevier.com/locate/ybbrc
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of TNFα enhances the motility and invasiveness of prostatic cancer cells by stimulating the expression...

Biochemical and Biophysical Research Communications 409 (2011) 436–441
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
TNFa enhances the motility and invasiveness of prostatic cancer cells bystimulating the expression of selective glycosyl- and sulfotransferase genesinvolved in the synthesis of selectin ligands
Prakash Radhakrishnan a, Vishwanath Chachadi a, Ming-Fong Lin a,b, Rakesh Singh b,c, Reiji Kannagi d,Pi-Wan Cheng a,b,⇑a Department of Biochemistry and Molecular Biology, College of Medicine, University of Nebraska Medical Center, Omaha, NE 68198, USAb Eppley Cancer Center for Research in Cancer and Allied Diseases, University of Nebraska Medical Center, Omaha, NE 68198, USAc Department of Pathology and Microbiology, University of Nebraska Medical Center, Omaha, NE 68198, USAd Aichi Cancer Center, Nagoya, Japan
a r t i c l e i n f o a b s t r a c t
Article history:Received 26 April 2011Available online 8 May 2011
Keywords:LNCaP cellssLex
TNFaGlycogenesMotility and invasion
0006-291X/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.bbrc.2011.05.019
⇑ Corresponding author. Address: Department ofBiology, College of Medicine, 985870 Nebraska MNebraska Medical Center, Omaha, NE 68198-5870, US
E-mail address: [email protected] (P.-W. Cheng).
Sialyl Lewis x (sLex) plays an important role in cancer metastasis. But, the mechanism for its productionin metastatic cancers remains unclear. The objective of current study was to examine the effects of a pro-inflammatory cytokine on the expression of glycosyltransferase and sulfotransferase genes involved inthe synthesis of selectin ligands in a prostate cancer cell line. Androgen-independent human lymphnode-derived metastatic prostate cancer cells (C-81 LNCaP), which express functional androgen receptorand mimic the castration-resistant advanced prostate cancer, were used. TNFa treatment of these cellsincreased their binding to P-, E- and L-selectins, anti-sLex antibody, and anti-6-sulfo-sialyl Lewis x anti-body by 12%, 240%, 43%, 248% and 21%, respectively. Also, the expression of C2GnT-1, B4GalT1, Glc-NAc6ST3, and ST3Gal3 genes was significantly upregulated. Further treatment of TNFa-treated cellswith either anti-sLex antibody or E-selectin significantly suppressed their in vitro migration (81% and52%, respectively) and invasion (45% and 56%, respectively). Our data indicate that TNFa treatmentenhances the motility and invasion properties of LNCaP C-81 cells by increasing the formation of selectinligands through stimulation of the expression of selective glycosyl- and sulfotransferase genes. Theseresults support the hypothesis that inflammation contributes to cancer metastasis.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
Prostate cancer is the second leading cause of cancer death ofmen in the United States [1]. At the time of initial diagnosis, morethan 70% of prostate cancer patients have lymph node metastases[2–5]. Although metastasis is the primary cause of the fatalities ofcancer patients, the mechanism is not completely understood. Onekey step in the cancer metastatic process is the adhesion of circu-lating cancer cells to the endothelium at distant sites. This step in-volves selectin-ligand interactions between host cells (selectins)and cancer cells (selectin ligands). Understanding how the produc-tion of these two key players is regulated during cancer progres-sion could help develop cancer therapeutic strategy.
ll rights reserved.
Biochemistry and Molecularedical Center, University ofA. Fax: +1 402 559 6650.
Selectins are a family of type II transmembrane proteins found onthe surface of leukocytes, platelets and activated endothelial cells.They are calcium-dependent C-type lectins [6]. The three majortypes of selectins include E-, P-, and L-selectins, which are locatedon endothelium, platelets and activated endothelium, and leuko-cytes, respectively [6]. The naturally occurring vascular carbohy-drate ligands for these selectins include E-selectin ligand-1(ESL-1), P-selectin glycoprotein ligand-1 (PSGL-1), CD34, cutaneouslymphocyte-associated antigen (CLA), glycoprotein GlyCAM-1,MAdCAM-1, Podocalyxin-like protein and Sgp200 [7]. These selec-tin-ligand interactions were originally shown to be involved in thetrafficking of immune cells to inflamed tissues [8] and lymphoid or-gans, and recently in the metastasis of cancer cells [9]. The naturallyoccurring vascular ligands for these selectin molecules are mostlymucin-type glycoproteins which carry sialylated, fucosylated andsulfated glycans [10] although some N-linked glycans also can par-ticipate in this process [11]. Increased expression and altered glyco-sylation of mucins are prominent features of carcinoma progression[10]. For example, sialyl Lewis x (sLex) and sialyl Lewis a (sLea) are

P. Radhakrishnan et al. / Biochemical and Biophysical Research Communications 409 (2011) 436–441 437
increased substantially in tumors [12], including prostate cancers[13,14]. Elevated expression of sLex and sLea on mucin type O-gly-cans is highly correlated with lymphatic and venous invasion of can-cerous cells [15,16]. SLex and 6-sulfo-sLex also were elevated underin vivo inflammatory condition [9] and in vitro induction with proin-flammatroy cytokine, tumor necrosis factor a (TNFa) [17,18]. Glyco-syl- and sulfotransferases responsible for the biosynthesis of theabove mentioned epitopes were selectively upregulated in variouscancers [19] and inflammatory conditions [9] and/or cells treatedwith TNFa [17,18]. However, a concerted effort to correlate the ef-fects of inflammatory cytokine on the expression of selectin ligands,glycosyl- and sulfotransferase genes responsible for the synthesis ofthese ligands, interactions of these ligands with selectins, and theacquisition of metastatic potential of the TNFa-treated cells waslacking.
Recently, serum TNFa level was found to be increased in prostatecancer patients with metastatic disease and elevated PSA [20]. Thisobservation prompted us to examine whether TNFa could enhancethe metastatic potential of prostate cancer cells via stimulating theproduction of sLex, an important determinant of cancer metastasis.To carry out a more comprehensive study on the effect of TNFa onthe acquisition of the sLex-related metastatic potential in prostatecancer, we employed LNCaP C-81 cells, which exhibit propertiesmimicking the clinical characteristics of advanced prostate cancers[21–23]. C-81 cells are androgen independent, grow faster and havehigh colony-forming property on soft agar as compared to LNCaPparental C-33 cells [22]. We found that treatment of the C-81 LNCaPcells with TNFa increased the motility and invasiveness, whichaccompanied an increase in selectin ligands resulted from elevatedexpression of selected glycosyl- and sulfotransferase genes.
2. Materials and methods
2.1. Cell lines
The human prostate cancer cell line LNCaP was originally pur-chased from the American Type Culture Collection ATCC (Rockville,MD). The LNCaP cell model including C-33 and C-81 cells used inthe present study was developed by Lin et al. [22] and further char-acterized by Igawa et al. [21]. The cell line was maintained in RPMI1640 medium supplemented with 5% Fetal bovine serum, 1% L-glu-tamine and 1% Penicillin–Streptomycin.
2.2. Flow cytometry analysis of selectin interactions with ligands onLNCaP cells
Flow cytometry analysis of selectin interactions with ligands oncancer cells was carried out as described previously [24]. Briefly,LNCaP cells of both control and TNFa (20 ng/ml, 36 h) treated weredetached from plates after incubation with PBS containing 2 mMEDTA for 5 min at 37 �C, washed three times with Hanks’ balancedsalt solution (HBSS), and then treated with 0.5% BSA in HBSS for30 min to block nonspecific binding. Chimeras of selectins P(50 lg/ml), E (50 lg/ml), and L (50 lg/ml) fused with Fc region ofhuman IgG were purified from human 293 cells (transformedembryonic kidney fibroblast cell line) of Psel IgG, ESIg B3293 andLSRg293 Cytel, respectively [24]. These selectins were pre-incu-bated with a goat-anti human IgG conjugated with FITC for 1 h atroom temperature. Similarly, sLex (KM93) (10 lg/ml) and 6-sulfo-sLex (G152) (1:10 diluted) [25] mouse monoclonal antibodies werepre-incubated with goat anti-mouse antibody conjugated with FITCbefore use. These complexes (Selectin, KM93 Ab, or G152 Ab com-plexed with respective FITC-conjugated secondary antibodies) wereincubated with tumor cells at 4 �C for 2 h and then washed succes-sively with HBSS/BSA and HBSS. Then cells were fixed with 2% (wt/
vol) paraformaldehyde in HBSS at room temperature for 15–30 min. The cells were then washed with HBSS and resuspended in1 ml of HBSS/BSA for flow cytometry analysis. Controls were cellsstained in a presence of 5 mM EDTA (calcium chelation) or 30 mMEDTA in the case of P-selectin. In some instances, tumor cell surfaceswere pretreated before probing for selectin ligands. Cells treatedwith FITC-conjugated secondary antibody alone served as an anti-body control.
2.3. Real time PCR analysis of the expression of glycosyl- andsulfotransferase genes
RT-PCR analyses on LNCaP C-81 cells were performed asdescribed previously [26]. Total cellular RNAs were isolated fromTNFa treated (20 ng/ml, 3 h) and control LNCaP cells using the TRI-REAGENT (Molecular Research Center, Inc, USA) according to themanufacturer protocol. cDNA was synthesized by using 2 lg of totalRNA as the template in a 20 ll RT reaction mixture by using Verso™cDNA kit (ABgene, UK) according to the manufacturer instruction.Quantitative real time PCR was performed on a Master cycler ep real-plex2 (Eppendorf AG, Hamburg) using SYBR Green PCR kit (TakaraBio Inc, USA). The forward and reverse primers used for amplifica-tion of selective glycosyl- and sulfotransferase genes are listed inTable 1. The data were analyzed using Eppendorf realplex software,version 1.5 (Eppendorf). The amounts of various glycogene tran-scripts were normalized to the amount of GAPDH transcript in samecDNA sample and expressed as % of GAPDH. Relative fold differencesin transcript expression were determined using the following com-parative CT method: 2�[DCt(TNFa)�DCt(Control)] = 2�DDCt, whereDCt = Ct(Target)�Ct(GAPDH) as described previously [26]. Theresults were expressed as the amount (%) relative to that (100%) ofGAPDH and plotted as mean fold changes ± SEM.
2.4. Motility and invasion assays
Motility and invasion assays were performed as previously de-scribed [23]. For motility assays, 5 � 104 cells/ml of control, TNFatreated (20 ng/ml, 36 h), and TNFa treated cells further incubatedwith sLex monoclonal antibody KM93 (10 lg/ml) or E-selectin(100 lg/ml) were placed on the top chamber of non-coated polyeth-ylene terapthalate membranes. After incubation for 24 h, the cellsthat did not migrate through the pores in the membrane were re-moved by scraping the membrane with a cotton swab. Cells thattransversed the membranes were stained with a Diff-Quick cellstaining kit (Dade Behring, Inc., Newark, DE). Invasion assay of pros-tate cancer cells was carried out as described above except that 24well BD BioCoat™ Matrigel™ invasion chambers (8 lm pore size;Becton Dickinson) were used and the incubation time was 22 h. Cellsat three random fields per insert were counted with 40�magnifica-tion and expressed as the average number of cells per field of view.Three independent experiments were performed in triplicates ineach condition. The data were presented as the mean of three inde-pendent experiments ± SD.
2.5. Statistical analysis
The Student’s t-test was used for analysis of the significance ofdifferences in means between control and tested samples. P < 0.05was considered significant.
3. Results
3.1. TNFa enhancement of selectin binding to LNCaP cells
LNCaP C-81 cells with and without TNFa treatment were as-sessed for their binding to P-, E-, and L-selectins by flow cytometry.
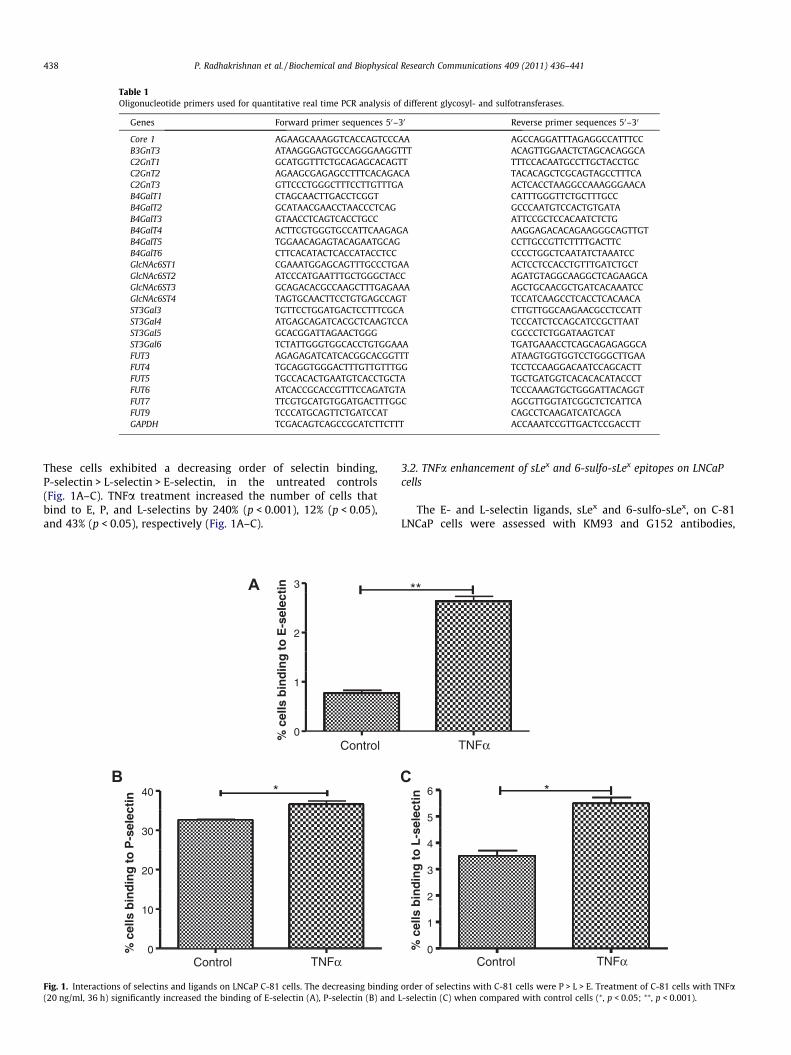
Table 1Oligonucleotide primers used for quantitative real time PCR analysis of different glycosyl- and sulfotransferases.
Genes Forward primer sequences 50–30 Reverse primer sequences 50–30
Core 1 AGAAGCAAAGGTCACCAGTCCCAA AGCCAGGATTTAGAGGCCATTTCCB3GnT3 ATAAGGGAGTGCCAGGGAAGGTTT ACAGTTGGAACTCTAGCACAGGCAC2GnT1 GCATGGTTTCTGCAGAGCACAGTT TTTCCACAATGCCTTGCTACCTGCC2GnT2 AGAAGCGAGAGCCTTTCACAGACA TACACAGCTCGCAGTAGCCTTTCAC2GnT3 GTTCCCTGGGCTTTCCTTGTTTGA ACTCACCTAAGGCCAAAGGGAACAB4GalT1 CTAGCAACTTGACCTCGGT CATTTGGGTTCTGCTTTGCCB4GalT2 GCATAACGAACCTAACCCTCAG GCCCAATGTCCACTGTGATAB4GalT3 GTAACCTCAGTCACCTGCC ATTCCGCTCCACAATCTCTGB4GalT4 ACTTCGTGGGTGCCATTCAAGAGA AAGGAGACACAGAAGGGCAGTTGTB4GalT5 TGGAACAGAGTACAGAATGCAG CCTTGCCGTTCTTTTGACTTCB4GalT6 CTTCACATACTCACCATACCTCC CCCCTGGCTCAATATCTAAATCCGlcNAc6ST1 CGAAATGGAGCAGTTTGCCCTGAA ACTCCTCCACCTGTTTGATCTGCTGlcNAc6ST2 ATCCCATGAATTTGCTGGGCTACC AGATGTAGGCAAGGCTCAGAAGCAGlcNAc6ST3 GCAGACACGCCAAGCTTTGAGAAA AGCTGCAACGCTGATCACAAATCCGlcNAc6ST4 TAGTGCAACTTCCTGTGAGCCAGT TCCATCAAGCCTCACCTCACAACAST3Gal3 TGTTCCTGGATGACTCCTTTCGCA CTTGTTGGCAAGAACGCCTCCATTST3Gal4 ATGAGCAGATCACGCTCAAGTCCA TCCCATCTCCAGCATCCGCTTAATST3Gal5 GCACGGATTAGAACTGGG CGCCCTCTGGATAAGTCATST3Gal6 TCTATTGGGTGGCACCTGTGGAAA TGATGAAACCTCAGCAGAGAGGCAFUT3 AGAGAGATCATCACGGCACGGTTT ATAAGTGGTGGTCCTGGGCTTGAAFUT4 TGCAGGTGGGACTTTGTTGTTTGG TCCTCCAAGGACAATCCAGCACTTFUT5 TGCCACACTGAATGTCACCTGCTA TGCTGATGGTCACACACATACCCTFUT6 ATCACCGCACCGTTTCCAGATGTA TCCCAAAGTGCTGGGATTACAGGTFUT7 TTCGTGCATGTGGATGACTTTGGC AGCGTTGGTATCGGCTCTCATTCAFUT9 TCCCATGCAGTTCTGATCCAT CAGCCTCAAGATCATCAGCAGAPDH TCGACAGTCAGCCGCATCTTCTTT ACCAAATCCGTTGACTCCGACCTT
438 P. Radhakrishnan et al. / Biochemical and Biophysical Research Communications 409 (2011) 436–441
These cells exhibited a decreasing order of selectin binding,P-selectin > L-selectin > E-selectin, in the untreated controls(Fig. 1A–C). TNFa treatment increased the number of cells thatbind to E, P, and L-selectins by 240% (p < 0.001), 12% (p < 0.05),and 43% (p < 0.05), respectively (Fig. 1A–C).
3A
2
1
tin
lec
E-s
e t
o
din
gb
inel
ls
% c
Control0
*B
40
30
20
10
Control TNFα0
in
lect
-se
to
Pin
gb
ind
lls
ce
%
Fig. 1. Interactions of selectins and ligands on LNCaP C-81 cells. The decreasing binding(20 ng/ml, 36 h) significantly increased the binding of E-selectin (A), P-selectin (B) and
3.2. TNFa enhancement of sLex and 6-sulfo-sLex epitopes on LNCaPcells
The E- and L-selectin ligands, sLex and 6-sulfo-sLex, on C-81LNCaP cells were assessed with KM93 and G152 antibodies,
**
TNFα
*C
6 *
tin
5elec
4 L-s
g t
o
3
din
2
bin
ells
1
% c
Control TNFα0
order of selectins with C-81 cells were P > L > E. Treatment of C-81 cells with TNFaL-selectin (C) when compared with control cells (⁄, p < 0.05; ⁄⁄, p < 0.001).
P. Radhakrishnan et al. / Biochemical and Biophysical Research Communications 409 (2011) 436–441 439
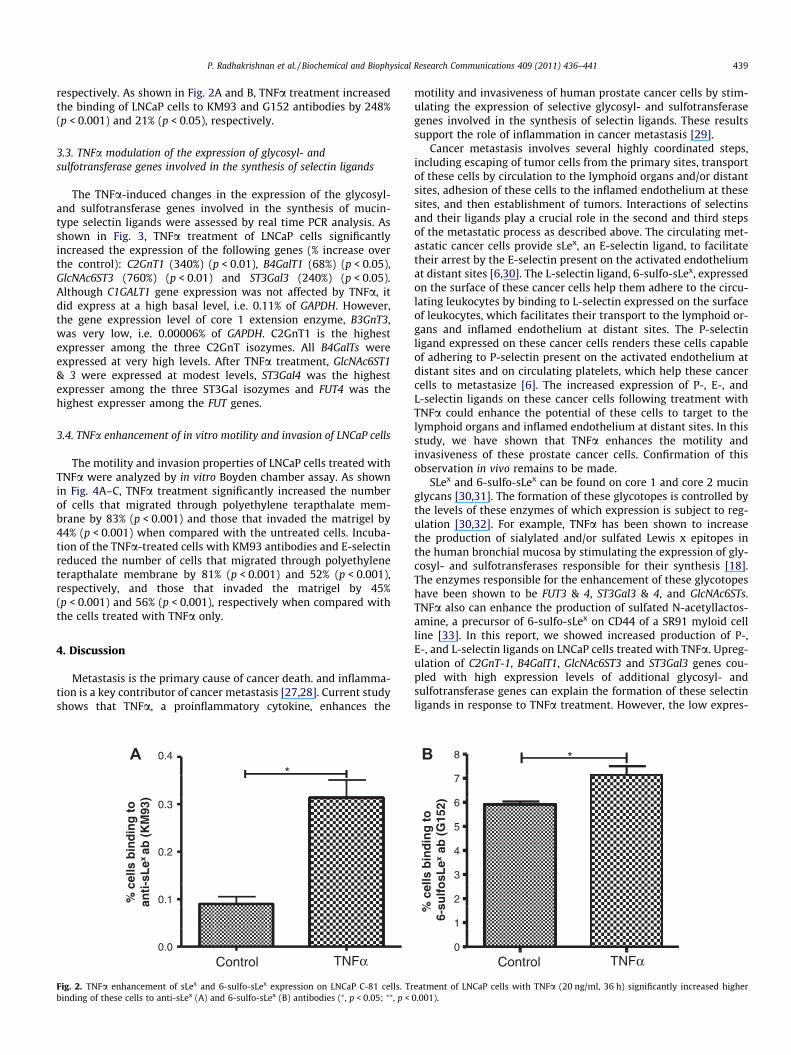
respectively. As shown in Fig. 2A and B, TNFa treatment increasedthe binding of LNCaP cells to KM93 and G152 antibodies by 248%(p < 0.001) and 21% (p < 0.05), respectively.
3.3. TNFa modulation of the expression of glycosyl- andsulfotransferase genes involved in the synthesis of selectin ligands
The TNFa-induced changes in the expression of the glycosyl-and sulfotransferase genes involved in the synthesis of mucin-type selectin ligands were assessed by real time PCR analysis. Asshown in Fig. 3, TNFa treatment of LNCaP cells significantlyincreased the expression of the following genes (% increase overthe control): C2GnT1 (340%) (p < 0.01), B4GalT1 (68%) (p < 0.05),GlcNAc6ST3 (760%) (p < 0.01) and ST3Gal3 (240%) (p < 0.05).Although C1GALT1 gene expression was not affected by TNFa, itdid express at a high basal level, i.e. 0.11% of GAPDH. However,the gene expression level of core 1 extension enzyme, B3GnT3,was very low, i.e. 0.00006% of GAPDH. C2GnT1 is the highestexpresser among the three C2GnT isozymes. All B4GalTs wereexpressed at very high levels. After TNFa treatment, GlcNAc6ST1& 3 were expressed at modest levels, ST3Gal4 was the highestexpresser among the three ST3Gal isozymes and FUT4 was thehighest expresser among the FUT genes.
3.4. TNFa enhancement of in vitro motility and invasion of LNCaP cells
The motility and invasion properties of LNCaP cells treated withTNFa were analyzed by in vitro Boyden chamber assay. As shownin Fig. 4A–C, TNFa treatment significantly increased the numberof cells that migrated through polyethylene terapthalate mem-brane by 83% (p < 0.001) and those that invaded the matrigel by44% (p < 0.001) when compared with the untreated cells. Incuba-tion of the TNFa-treated cells with KM93 antibodies and E-selectinreduced the number of cells that migrated through polyethyleneterapthalate membrane by 81% (p < 0.001) and 52% (p < 0.001),respectively, and those that invaded the matrigel by 45%(p < 0.001) and 56% (p < 0.001), respectively when compared withthe cells treated with TNFa only.
4. Discussion
Metastasis is the primary cause of cancer death. and inflamma-tion is a key contributor of cancer metastasis [27,28]. Current studyshows that TNFa, a proinflammatory cytokine, enhances the
A*
0.3
0.4
to
93
)d
ing
(KM
0.2 bin
x ab
ells
sLe
0.1% c
nti
-a
Control TNFα0.0
Fig. 2. TNFa enhancement of sLex and 6-sulfo-sLex expression on LNCaP C-81 cells. Trbinding of these cells to anti-sLex (A) and 6-sulfo-sLex (B) antibodies (⁄, p < 0.05; ⁄⁄, p <
motility and invasiveness of human prostate cancer cells by stim-ulating the expression of selective glycosyl- and sulfotransferasegenes involved in the synthesis of selectin ligands. These resultssupport the role of inflammation in cancer metastasis [29].
Cancer metastasis involves several highly coordinated steps,including escaping of tumor cells from the primary sites, transportof these cells by circulation to the lymphoid organs and/or distantsites, adhesion of these cells to the inflamed endothelium at thesesites, and then establishment of tumors. Interactions of selectinsand their ligands play a crucial role in the second and third stepsof the metastatic process as described above. The circulating met-astatic cancer cells provide sLex, an E-selectin ligand, to facilitatetheir arrest by the E-selectin present on the activated endotheliumat distant sites [6,30]. The L-selectin ligand, 6-sulfo-sLex, expressedon the surface of these cancer cells help them adhere to the circu-lating leukocytes by binding to L-selectin expressed on the surfaceof leukocytes, which facilitates their transport to the lymphoid or-gans and inflamed endothelium at distant sites. The P-selectinligand expressed on these cancer cells renders these cells capableof adhering to P-selectin present on the activated endothelium atdistant sites and on circulating platelets, which help these cancercells to metastasize [6]. The increased expression of P-, E-, andL-selectin ligands on these cancer cells following treatment withTNFa could enhance the potential of these cells to target to thelymphoid organs and inflamed endothelium at distant sites. In thisstudy, we have shown that TNFa enhances the motility andinvasiveness of these prostate cancer cells. Confirmation of thisobservation in vivo remains to be made.
SLex and 6-sulfo-sLex can be found on core 1 and core 2 mucinglycans [30,31]. The formation of these glycotopes is controlled bythe levels of these enzymes of which expression is subject to reg-ulation [30,32]. For example, TNFa has been shown to increasethe production of sialylated and/or sulfated Lewis x epitopes inthe human bronchial mucosa by stimulating the expression of gly-cosyl- and sulfotransferases responsible for their synthesis [18].The enzymes responsible for the enhancement of these glycotopeshave been shown to be FUT3 & 4, ST3Gal3 & 4, and GlcNAc6STs.TNFa also can enhance the production of sulfated N-acetyllactos-amine, a precursor of 6-sulfo-sLex on CD44 of a SR91 myloid cellline [33]. In this report, we showed increased production of P-,E-, and L-selectin ligands on LNCaP cells treated with TNFa. Upreg-ulation of C2GnT-1, B4GalT1, GlcNAc6ST3 and ST3Gal3 genes cou-pled with high expression levels of additional glycosyl- andsulfotransferase genes can explain the formation of these selectinligands in response to TNFa treatment. However, the low expres-
*B 8 *
7
) 6
to
15
2
5
din
gb
(G
4
bin ex
a
3
ells
osL
2
% c
sulf
16-
Control TNFα0
eatment of LNCaP cells with TNFa (20 ng/ml, 36 h) significantly increased higher0.001).
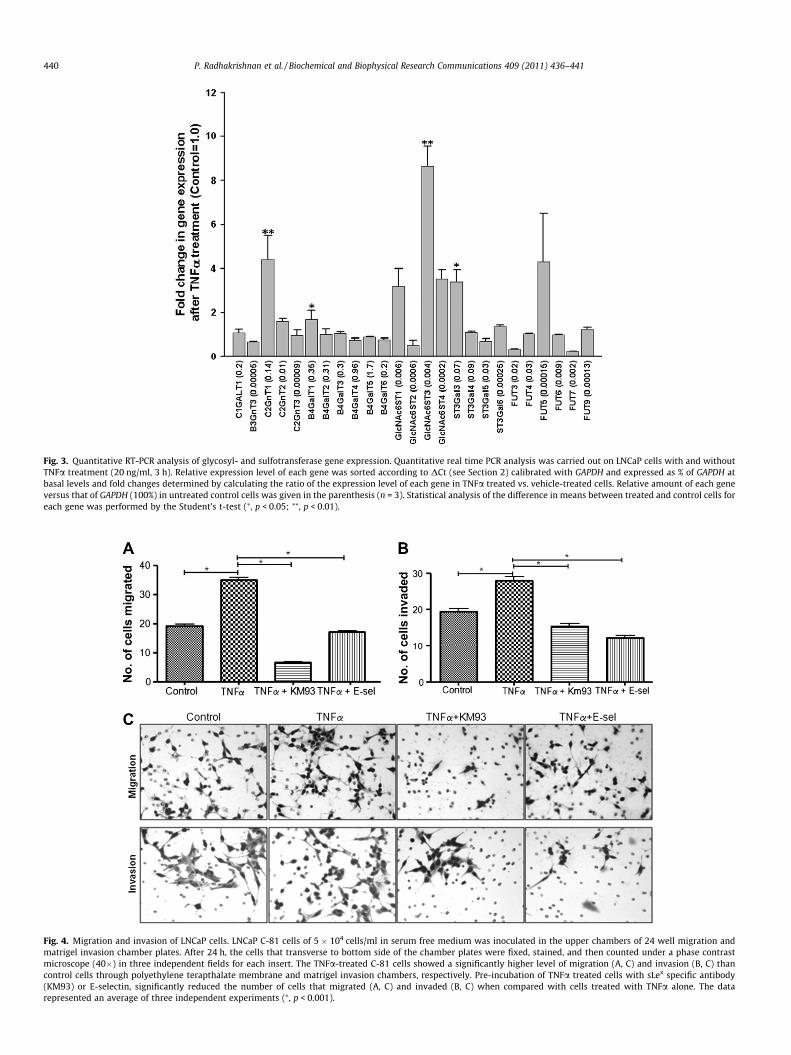
Fig. 3. Quantitative RT-PCR analysis of glycosyl- and sulfotransferase gene expression. Quantitative real time PCR analysis was carried out on LNCaP cells with and withoutTNFa treatment (20 ng/ml, 3 h). Relative expression level of each gene was sorted according to DCt (see Section 2) calibrated with GAPDH and expressed as % of GAPDH atbasal levels and fold changes determined by calculating the ratio of the expression level of each gene in TNFa treated vs. vehicle-treated cells. Relative amount of each geneversus that of GAPDH (100%) in untreated control cells was given in the parenthesis (n = 3). Statistical analysis of the difference in means between treated and control cells foreach gene was performed by the Student’s t-test (⁄, p < 0.05; ⁄⁄, p < 0.01).
Fig. 4. Migration and invasion of LNCaP cells. LNCaP C-81 cells of 5 � 104 cells/ml in serum free medium was inoculated in the upper chambers of 24 well migration andmatrigel invasion chamber plates. After 24 h, the cells that transverse to bottom side of the chamber plates were fixed, stained, and then counted under a phase contrastmicroscope (40�) in three independent fields for each insert. The TNFa-treated C-81 cells showed a significantly higher level of migration (A, C) and invasion (B, C) thancontrol cells through polyethylene terapthalate membrane and matrigel invasion chambers, respectively. Pre-incubation of TNFa treated cells with sLex specific antibody(KM93) or E-selectin, significantly reduced the number of cells that migrated (A, C) and invaded (B, C) when compared with cells treated with TNFa alone. The datarepresented an average of three independent experiments (⁄, p < 0.001).
440 P. Radhakrishnan et al. / Biochemical and Biophysical Research Communications 409 (2011) 436–441

P. Radhakrishnan et al. / Biochemical and Biophysical Research Communications 409 (2011) 436–441 441
sion level (0.00005% of GAPDH) of the core 1 extension enzyme(B3GnT3) suggests that the core 1 can not be extended to supportthe formation of these glycotopes on this core structure. Therefore,these selectin ligands are most likely localized to the core 2 branch.This prediction remains to be confirmed by glycan analysis.
We extended the biochemical and molecular biologic observa-tions by showing that TNFa-induced changes in glycogeneexpression profile led to increased binding of these cells toP-, E-, and L-selectins, and increased their adhesion, motility,and invasion properties. This observation confirmed a previousreport that TNFa increased migration and invasion of a humanmelanoma cell line [34]. Collectively, these results support theidea that inflammatory cytokines can enhance the metastaticproperties of cancer cells and further substantiate the linkbetween inflammation and cancer progression as previouslydescribed [27,28].
In conclusion, the present study shows the TNFa effect onenhancement of prostate cancer cell migration and invasionthrough activation of selective glycosyl and sulfotransferase genesinvolved in the biosynthesis of sialylated and/or sulfated sLex.These genes may serve as potential therapeutic targets.
Acknowledgments
We wish to acknowledge the research support from NIH RO1CA88184 (MFL), RO1 HL48282 and R21 HL97238, the State of Ne-braska-NRI cancer glycobiology program and LB506 (PWC), andthe Department of Defense Postdoctoral Fellowship (PR), and ex-pert technical assistance of Helen Cheng. We also thank Dr. AjitVarki at the University of California at San Diego for providing293 cells stably transfected with plasmids containing cDNAs thatencode secreted P-, E-, and L-selectins fused with Fc portion of hu-man IgG.
References
[1] A. Jemal, R. Siegel, J. Xu, E. Ward, Cancer statistics, CA-Cancer. J. Clin. 60 (2010)277–300.
[2] W.G. Nelson, A.M. De Marzo, W.B. Isaacs, Prostate cancer, New Engl. J. Med.349 (2003) 366–381.
[3] L. Borsig, R. Wong, J. Feramisco, D.R. Nadeau, N.M. Varki, A. Varki, Heparincancer revisited: mechanistic connections involving platelets, P-selectin,carcinoma mucins, and tumor metastasis, P. Natl. Acad. Sci. USA 98 (2001)3352–3357.
[4] Y. Imai, M.S. Singer, C. Fennie, L.A. Lasky, S.D. Rosen, Identification of acarbohydrate-based endothelial ligand for a lymphocyte homing receptor, J.Cell Biol. 113 (1991) 1213–1221.
[5] S. Hemmerich, E.C. Butcher, S.D. Rosen, Sulfation-dependent recognition ofhigh endothelial venules (HEV)-ligands by L-selectin and MECA 79, andadhesion-blocking monoclonal antibody, J. Exp. Med. 180 (1994) 2219–2226.
[6] A. Varki, Selectin ligands, P. Natl. Acad. Sci. USA 91 (1994) 7390–7397.[7] J.C. Yeh, N. Hiraoka, B. Petryniak, J. Nakayama, L.G. Ellies, D. Rabuka, O.
Hindsgaul, J.D. Marth, J.B. Lowe, M. Fukuda, Novel sulfated lymphocyte homingreceptors and their control by a Core 1 extension beta 1,3-N-acetylglucosaminyltransferase, Cell 105 (2001) 957–969.
[8] P.R. Streeter, B.T. Rouse, E.C. Butcher, Immunohistologic and functionalcharacterization of a vascular addressin involved in lymphocyte homing intoperipheral lymph nodes, J. Cell Biol. 107 (1988) 1853–1862.
[9] R.P. McEver, Selectin-carbohydrate interactions during inflammation andmetastasis, Glycoconjugate J. 14 (1997) 585–591.
[10] Y.J. Kim, L. Borsig, H.L. Han, N.M. Varki, A. Varki, Distinct selectin ligands oncolon carcinoma mucins can mediate pathological interactions amongplatelets, leukocytes, and endothelium, Am. J. Pathol. 155 (1999) 461–472.
[11] J. Mitoma, X. Bao, B. Petryanik, P. Schaerli, J.M. Gauguet, S.Y. Yu, H. Kawashima,H. Saito, K. Ohtsubo, J.D. Marth, K.H. Khoo, U.H. von Andrian, J.B. Lowe, M.Fukuda, Critical functions of N-glycans in L-selectin-mediated lymphocytehoming and recruitment, Nat. Immunol. 8 (2007) 409–418.
[12] C. Ohyama, S. Tsuboi, M. Fukuda, Dual roles of sialyl lewis X oligosaccharidesin tumor metastasis and rejection by natural killer cells, EMBO J. 18 (1999)1516–1525.
[13] S. Martensson, S.A. Bigler, M. Brown, P.H. Lange, M.K. Brawer, S. Hakomori,Sialyl-lewis(x) and related carbohydrate antigens in the prostate, Hum. Pathol.26 (1995) 735–739.
[14] J. Wei, L. Cui, F. Liu, Y. Fan, R. Lang, F. Gu, X. Guo, P. Tang, L. Fu, E-selectin andsialyl lewis X expression is associated with lymph node metastasis of invasivemicropapillary carcinoma of the breast, Int. J. Surg. Pathol. 18 (2010) 193–200.
[15] S. Sumikura, S. Ishigami, S. Natsugoe, F. Miyazono, K. Tokuda, A. Nakajo, H.Okumura, M. Matsumoto, S. Hokita, T. Aikou, Disseminated cancer cells in theblood and expression of sialylated antigen in gastric cancer, Cancer Lett. 200(2003) 77–83.
[16] U. Jeschke, I. Mylonas, N. Shabani, C. Kunert-Keil, C. Schindlbeck, B. Gerber, K.Friese, Expression of sialyl lewis X, sialyl lewis A, E-cadherin and cathepsin-Din human breast cancer: Immunohistochemical analysis in mammarycarcinoma in situ invasive carcinomas and their lymph node metastasis,Anticancer Res. 25 (2005) 1615–1622.
[17] Y. Ishibashi, Y. Inouye, T. Okano, A. Taniguchi, Regulation of sialyl-lewis xepitope expression by TNF-alpha and EGF in an airway carcinoma cell line,Glycoconjugate J. 22 (2005) 53–62.
[18] P. Delmotte, S. Degroote, J.J. Lafitte, G. Lamblin, J.M. Perini, P. Roussel, Tumornecrosis factor alpha increases the expression of glycosyltransferases andsulfotransferases responsible for the biosynthesis of sialylated and/or sulfatedlewis x epitopes in the human bronchial mucosa, J. Biol. Chem. 277 (2002)424–431.
[19] T. Kudo, Y. Ikehara, A. Togayachi, K. Morozumi, M. Watanabe, M. Nakamura, S.Nishihara, H. Narimatsu, Up-regulation of a set of glycosyltransferase genes inhuman colorectal cancer, Lab. Invest. 78 (1998) 797–811.
[20] V. Michalaki, K. Syrigos, P. Charles, J. Waxman, Serum levels of IL-6 and TNF-alpha correlate with clinicopathological features and patient survival inpatients with prostate cancer, Brit. J. Cancer 90 (2004) 2312–2316.
[21] T. Igawa, F.F. Lin, M.S. Lee, D. Karan, S.K. Batra, M.F. Lin, Establishment andcharacterization of androgen-independent human prostate cancer LNCaP cellmodel, Prostate 50 (2002) 222–235.
[22] M.F. Lin, M.S. Lee, X.W. Zhou, J.C. Andressen, T.C. Meng, S.L. Johansson, W.W.West, R.J. Taylor, J.R. Anderson, F.F. Lin, Decreased expression of cellularprostatic acid phosphatase increases tumorigenicity of human prostate cancercells, J. Urol. 166 (2001) 1943–1950.
[23] P. Radhakrishnan, M.F. Lin, P.W. Cheng, Elevated expression of L-selectinligand in lymph node-derived human prostate cancer cells correlates withincreased tumorigenicity, Glycoconjugate. J. 26 (2009) 75–81.
[24] L. Borsig, R. Wong, R.O. Hynes, N.M. Varki, A. Varki, Synergistic effects of L- andP-selectin in facilitating tumor metastasis can involve non-mucin ligands andimplicate leukocytes as enhancers of metastasis, P. Natl. Acad. Sci. USA 99(2002) 2193–2198.
[25] C. Mitsuoka, M. Sawada-Kasugai, K. Ando-Furui, M. Izawa, H. Nakanishi, S.Nakamura, H. Ishida, M. Kiso, R. Kannagi, Identification of a majorcarbohydrate capping group of the L-selectin ligand on high endothelialvenules in human lymph nodes as 6-sulfo sialyl lewis X, J. Biol. Chem. 273(1998) 11225–11233.
[26] F. Tassone, R.J. Hagerman, A.K. Taylor, L.W. Gane, T.E. Godfrey, P.J.Hagerman, Elevated levels of FMR1 mRNA in carrier males: a newmechanism of involvement in the fragile-X syndrome, Am. J. Hum. Genet.66 (2000) 6–15.
[27] F. Balkwill, C. Joffroy, TNF: a tumor-suppressing factor or a tumor-promotingfactor?, Future On 6 (2010) 1833–1836.
[28] F. Balkwill, Tumor necrosis factor or tumor promoting factor?, CytokineGrowth F R. 13 (2002) 135–141.
[29] H. Clevers, At the crossroads of inflammation and cancer, Cell 118 (2004) 671–674.
[30] P.-W. Cheng, P. Radhakrishnan, Mucin glycan branching enzymes: structure,function and gene regulation, in: Albert Wu (Ed.), Molecular Immunology ofComplex Carbohydrates-3: , Advances in Experimental Medicine and Biology,vol. 705, Plenum Press, NY, NY, 2011, pp. 465–492.
[31] H. Kawashima, B. Petryniak, N. Hiraoka, J. Mitoma, V. Huckabyet, J. Nakayama,K. Uchimura, K. Kadomatsu, J.B. Lowe, M. Fukuda, N-acetylglucosamine-6-O-sulfotransferases 1 and 2 cooperatively control lymphocyte homing through L-selectin ligand biosynthesis in high endothelial venues, Nat. Immunol. 6(2005) 1096–1104.
[32] I. Brockhausen, Mucin-type O-glycans in human colon and breast cancer:glycodynamics and functions, EMBO Rep. 7 (2006) 599–604.
[33] M. Delcommenne, R. Kannagi, P. Johnson, TNF-alpha increases thecarbohydrate sulfation of CD44: Induction of 6-sulfo N-acetyl lactosamineon N- and O-linked glycans, Glycobiology 12 (2002) 613–622.
[34] E. Katerinaki, G.S. Evans, P.C. Lorigan, S. MacNeil, TNF-alpha increases humanmelanoma cell invasion and migration in vitro: the role of proteolyticenzymes, Brit. J. Cancer 89 (2003) 1123–1129.




















