
The Sgs1 Helicase of Saccharomyces cerevisiae Inhibits Retrotransposition of Ty1 Multimeric Arrays
16
10.1128/MCB.21.16.5374-5388.2001. 2001, 21(16):5374. DOI: Mol. Cell. Biol. Curcio Mary Bryk, Mukti Banerjee, Darryl Conte Jr. and M. Joan Ty1 Multimeric Arrays Inhibits Retrotransposition of cerevisiae Saccharomyces The Sgs1 Helicase of http://mcb.asm.org/content/21/16/5374 Updated information and services can be found at: These include: REFERENCES http://mcb.asm.org/content/21/16/5374#ref-list-1 at: This article cites 66 articles, 44 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on November 17, 2014 by guest http://mcb.asm.org/ Downloaded from on November 17, 2014 by guest http://mcb.asm.org/ Downloaded from
Transcript of The Sgs1 Helicase of Saccharomyces cerevisiae Inhibits Retrotransposition of Ty1 Multimeric Arrays

10.1128/MCB.21.16.5374-5388.2001.
2001, 21(16):5374. DOI:Mol. Cell. Biol. CurcioMary Bryk, Mukti Banerjee, Darryl Conte Jr. and M. Joan Ty1 Multimeric Arrays
Inhibits Retrotransposition ofcerevisiaeSaccharomycesThe Sgs1 Helicase of
http://mcb.asm.org/content/21/16/5374Updated information and services can be found at:
These include:
REFERENCEShttp://mcb.asm.org/content/21/16/5374#ref-list-1at:
This article cites 66 articles, 44 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/01/$04.0010 DOI: 10.1128/MCB.21.16.5374–5388.2001
Aug. 2001, p. 5374–5388 Vol. 21, No. 16
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
The Sgs1 Helicase of Saccharomyces cerevisiae InhibitsRetrotransposition of Ty1 Multimeric Arrays
MARY BRYK,1, 2 MUKTI BANERJEE,1 DARRYL CONTE, JR.,1† AND M. JOAN CURCIO1*
Molecular Genetics Program, Wadsworth Center and School of Public Health, State University of New York at Albany,Albany, New York 12208,1 and Department of Genetics, Harvard Medical School, Boston, Massachusetts 021152
Received 24 April 2001/Returned for modification 9 May 2001/Accepted 17 May 2001
Ty1 retrotransposons in the yeast Saccharomyces cerevisiae are maintained in a genetically competent buttranspositionally dormant state. When located in the ribosomal DNA (rDNA) locus, Ty1 elements are tran-scriptionally silenced by the specialized heterochromatin that inhibits rDNA repeat recombination. In addi-tion, transposition of all Ty1 elements is repressed at multiple posttranscriptional levels. Here, we demonstratethat Sgs1, a RecQ helicase required for genome stability, inhibits the mobility of Ty1 elements by a posttrans-lational mechanism. Using an assay for the mobility of Ty1 cDNA via integration or homologous recombina-tion, we found that the mobility of both euchromatic and rDNA-Ty1 elements was increased 32- to 79-fold insgs1D mutants. Increased Ty1 mobility was not due to derepression of silent rDNA-Ty1 elements, since deletionof SGS1 reduced the mitotic stability of rDNA-Ty1 elements but did not stimulate their transcription. Fur-thermore, deletion of SGS1 did not significantly increase the levels of total Ty1 RNA, protein, or cDNA and didnot alter the level or specificity of Ty1 integration. Instead, Ty1 cDNA molecules recombined at a highfrequency in sgs1D mutants, resulting in transposition of heterogeneous Ty1 multimers. Formation of Ty1multimers required the homologous recombination protein Rad52 but did not involve recombination betweenTy1 cDNA and genomic Ty1 elements. Therefore, Ty1 multimers that transpose at a high frequency in sgs1Dmutants are formed by intermolecular recombination between extrachromosomal Ty1 cDNA molecules beforeor during integration. Our data provide the first evidence that the host cell promotes retrotransposition ofmonomeric Ty1 elements by repressing cDNA recombination.
DNA helicases catalyze the unwinding of duplex DNA intoindividual DNA strands (42). A plethora of DNA helicaseswithin cells is involved in DNA replication, repair, recombina-tion, and transcription. Members of the RecQ family of DNAhelicases are involved in the maintenance of genome stabilityin all organisms characterized, from bacteria to humans (7).Mutations in the SGS1 gene, which encodes the only RecQhomologue in Saccharomyces cerevisiae, result in elevated lev-els of mitotic homologous and illegitimate recombination, in-creased rates of chromosomal nondisjunction, and acceleratedaging (21, 56, 61, 62, 65). Similarly, mutations in human genesencoding the RecQ homologues RecQL4 (35, 49), WRN (67),and BLM (18) give rise to rare hereditary disorders that arecharacterized by genome instability and a pronounced predis-position to cancer. Notably, expression of either WRN or BLMin yeast complements the hyperrecombination phenotypes ofan sgs1 mutant (65). These findings suggest that the mecha-nisms by which Sgs1 preserves genetic stability in yeast willserve as a paradigm for the role of RecQ homologues in hu-man disease.
The SGS1 gene was originally isolated in a screen for geneticinteraction with DNA topoisomerase III (21). Both topoisom-erase III and Sgs1 repress recombination of DNA repeats, andinactivation of either Sgs1 or topoisomerase III results in S-phase-specific defects. Therefore, it was proposed that the
combined activity of Sgs1 and topoisomerase III is required toinitiate DNA repair events during replication (7). In support ofthis model, Sgs1 was shown to colocalize with the cell cyclecheckpoint kinase, Rad53, in S-phase-specific foci and to par-ticipate in activation of the intra-S checkpoint (19). In theabsence of Sgs1, DNA lesions that arise during DNA replica-tion may be diverted into a homologous recombination path-way. This view is supported by recent reports that inactivationof Sgs1 and Srs2, another DNA helicase with a partially re-dundant role in genome maintenance, results in a severegrowth defect involving a high incidence of mitotic arrest (22,40, 46). The growth defect is suppressed by inactivation ofhomologous recombination proteins, including Rad51, Rad52,Rad55, and Rad57, suggesting that mitotic arrest in sgs1 srs2mutants results from DNA lesions that arise from abortiverecombination events (22, 46). Taken together, these findingssuggest that DNA lesions that block the replication fork act astriggers for homologous recombination in sgs1D mutants.
Sgs1 is critical for the stability of repeated DNA sequencessuch as those in the RDN1 locus. RDN1 contains 100 to 200tandem repeats of a 9.1-kb region encoding the RNA polymer-ase I (Pol I)-transcribed 35S rRNA gene and the Pol III-transcribed 5S rRNA gene. Both recombination between ribo-somal DNA (rDNA) repeats and Pol II-mediated transcriptionare inhibited in the RDN1 locus (6, 25, 57, 58). Recombinationin the rDNA can give rise to extrachromosomal rDNA circles(ERCs), which accumulate in aging cells and may trigger se-nescence (55). Many trans-acting factors that repress recombi-nation in rDNA have been identified. Where analyzed, genesthat inhibit rDNA recombination, including SIR2, TOP1,UBC2, and ZDS2, have been found to also be required for
* Corresponding author. Mailing address: Wadsworth Center, P.O.Box 22002, Albany, NY 12201-2002. Phone: (518) 473-6078. Fax: (518)474-3181. E-mail: [email protected].
† Present address: Program in Molecular Medicine, University ofMassachusetts Cancer Center, Worcester, MA 01605.
5374
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

transcriptional silencing in rDNA (6, 8, 20, 25, 52, 57). Fur-thermore, both SIR2 and ZDS2 forestall aging (31, 52). Theseand other findings have led to models in which a specific typeof heterochromatin mediates the repression of recombinationand transcription in the rDNA, and the integrity of this het-erochromatin is a primary determinant of longevity (26, 29). Insgs1 mutants, mitotic recombination in the rDNA is increasedsevenfold (21) and longevity is reduced by 60% (55, 56). Par-adoxically, ERCs accumulate to the same level in the presenceand absence of Sgs1 (28, 46). The role of Sgs1 in rDNA silenc-ing has not been investigated previously, despite its potential toprovide insight into the mechanism by which Sgs1 repressesrDNA recombination and aging.
In this study, we determined the effect of Sgs1 on the ex-pression and transposition of Ty1 elements in rDNA and eu-chromatic DNA. Ty1 elements constitute one of five families ofretrovirus-like transposons (Ty1 to Ty5) in yeast. Approxi-mately 30 Ty1 elements reside at dispersed sites in the haploidgenome, and the majority are transpositionally competent (12,34). Ty1 elements consist of a central coding domain flankedby long terminal repeats (LTRs). The coding domain containstwo overlapping open reading frames: TYA1, which encodes acapsid protein, and TYB1, which encodes protease, integrase(IN), and reverse transcriptase (RT). Ty1 elements are tran-scribed by Pol II to form a terminally redundant transcript. Ty1RNA is translated into two proteins, TyA1 and the TyA1-TyB1fusion protein, which assemble into virus-like particles (VLPs)that encapsulate Ty1 RNA during assembly. Maturation ofVLPs occurs by protease-mediated processing of Ty1 precur-sor proteins, and it is required for synthesis of a full-lengthlinear cDNA by RT, using Ty1 RNA as a template. Subse-quently, the Ty1 cDNA and IN protein are transported to thenucleus, presumably as components of a preintegration com-plex (33, 48). IN mediates transposition of Ty1 cDNA at anonhomologous target in the genome. Alternatively, Ty1cDNA can recombine with genomic Ty1 elements, which oc-curs by IN-independent and Rad52-dependent mechanisms(53). The introduction of Ty1 cDNA into the genome by trans-position or recombination is referred to as Ty1 cDNA-medi-ated mobility.
Several host mechanisms limit the potentially mutagenic ef-fects of Ty1 transposition. First, Ty1 elements are preferen-tially targeted to genomic regions that do not encode proteins,including the upstream regions of Pol III-transcribed genes(16, 30). Second, Ty1 elements that are located in the rDNAarray are subject to transcriptional silencing (6). Third, trans-position is inhibited by host factors that block posttranscrip-tional steps in Ty1 replication. For example, Fus3, a mitogen-activated protein kinase, destabilizes VLP-associated proteinsvia its negative regulation of the invasive growth pathway (10).In addition, Ssl2 and Rad3, two helicase components of tran-scription factor TFIIH, promote degradation of Ty1 cDNA,thereby repressing transposition 100-fold or more (39). Finally,members of the Rad52 epistasis group, including Rad50,Rad51, Rad52, Rad54, and Rad57, reduce Ty1 cDNA levelsand transposition by inhibiting cDNA synthesis or stability(50).
In this work, we demonstrate that Sgs1 inhibits the mobilityof Ty1 elements by a novel mechanism. Initially, we proposedthat Sgs1 might specifically repress transcription (and there-
fore transposition) of rDNA-Ty1 elements, since Sgs1 is re-quired for the repression of rDNA recombination. To oursurprise, we found that Sgs1 is not involved in the transcrip-tional silencing of Ty1 elements in the rDNA, although it isrequired for their mitotic stability. Furthermore, Sgs1 is aglobal inhibitor of Ty1 mobility that acts primarily at a stepfollowing cDNA synthesis. In the absence of Sgs1, extrachro-mosomal Ty1 cDNA molecules recombine at a high frequency,forming multimeric cDNA arrays that are subsequently inte-grated into the genome. We demonstrate that transposition ofmultimeric cDNA is the major cause of increased Ty1 mobilityin sgs1D mutants.
MATERIALS AND METHODS
Yeast strains, media, and genetic procedures. Standard yeast culture mediumwas prepared as described previously (51). The yeast strains used in this study arelisted in Table 1. The mapping of Ty1his3AI-236r and Ty1his3AI-816r to therDNA and Ty1his3AI-242 to a locus outside of rDNA on chromosome XII wasdescribed previously (6). Strain JC1109 is a haploid spore derived from a crossbetween strain JC1078 [MATa ura3(-52 or -167) trp1(-289 or :hisG) his3D200ade2D:hisG Ty1ade2AI-515] and strain JC816 (6). The leu2:hisG allele was in-troduced into strains JC236, JC242, and JC1109 by two-step transplacementusing plasmid pNK58, as described by Alani et al. (1). Subsequently, sgs1DLEU2derivatives were constructed by one-step transplacement using a DNA fragmentof plasmid pPWDSGS1 (62). The sgs1DLEU2 strain JC3161 was constructed bythe same method of transformation in strain YH8 (64). The sgs1DLEU2 disrup-tion alleles were verified by Southern analysis. Strains containing the rad52:hisGallele were constructed by two-step transplacement using plasmid pBDG542(12). To construct strains containing a HIS3-marked Ty1 element, the his3AImarker was replaced in strains JC236, JC242, and JC1109 and isogenic leu2:hisGsgs1DLEU2 derivatives by integrative transformation of a 0.8-kb ClaI fragment ofplasmid pGEM-HIS3 containing the HIS3 allele (6).
Plasmid construction. Plasmid pJC573, a URA3-based integrating vector con-taining '1.2 kb of yeast genomic DNA sequences from the BIK1-HIS4 intergenicregion (chromosome III, nucleotides 68462 to 69636) adjacent to aTy1his3AI[D1] element, was constructed in two steps. First, an 8.9-kb XhoI-EagIfragment of plasmid pOY1 (38) including the BIK1-HIS4 intergenic region ad-jacent to a Ty1-H3/912 element marked with his3AI was subcloned into pRS406(54). Second, a 0.9-kb ClaI fragment containing his3AI was deleted from thisplasmid and replaced with a 0.9-kb ClaI fragment containing the modified ret-rotranscript indicator gene (RIG), his3AI[D1]. The his3AI[D1] RIG contains thesame 104-bp artificial intron (AI) that is present in his3AI, but the intron hasbeen relocated to position 1440 in the HIS3 open reading frame. Consequently,the AI in his3AI[D1] is included within sequences that are deleted in the his3D1allele in yeast strain BY4742 and derivatives. Use of the his3AI[D1] RIG instrains carrying the his3D1 allele prevents formation of His1 prototrophs byectopic DNA-mediated recombination. The his3AI[D1] RIG was generouslyprovided by D. Garfinkel (Frederick Cancer Research and Development Center,National Cancer Institute), and its construction will be described elsewhere.
Plasmid pGEM-URA3-HIS3 contains a URA3-HIS3 cassette in the vectorpSP70 (Promega). It was constructed by subcloning a 1.3-kb BamHI-BglII frag-ment containing the URA3 allele into the unique BamHI site of plasmid pGEM-HIS3 (24). Plasmid pGEM-TYB1 contains the 934-bp HindIII-BglII fragment ofTy1-H3 (4) cloned into plasmid vector pSP70 (Promega).
Northern blot analysis. Total RNA was isolated from yeast strains JC236,JC280, JC2359, JC242, JC544, and JC2360 as described previously (6). Northernblot analysis was performed as described previously (59). 32P-labeled RNAprobes were used to detect Ty1his3AI, total Ty1, and PYK1 transcripts (13).Quantification was performed on a Storm 8600 phosphorimager using Image-Quant software.
Mitotic stability of Ty1HIS3 elements. The mitotic stability of Ty1HIS3 ele-ments was assayed as described previously (6). Briefly, single His1 colonies wereinoculated into 10 ml of yeast extract-peptone-dextrose (YPD) medium andgrown overnight at 30°C. Cultures were diluted 1:10,000 in fresh YPD and grownto saturation. A dilution of the ninth serial culture was plated on YPD mediumand replicated to synthetic complete (SC) medium lacking histidine to determinethe fraction of His2 auxotrophs.
Phenotypic assay for expression of mURA3-LEU2. SGS1 and congenic sgs1Dstrains containing the mURA3-LEU2 marker in rDNA or at the leu2D1 locuswere constructed using standard genetic techniques (51). Strain BY4742-10775
VOL. 21, 2001 Sgs1 INHIBITS TRANSPOSITION OF MULTIMERIC Ty1 ELEMENTS 5375
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

(sgs1DkanMX4) was crossed to strain JS210-1 (mURA3-LEU2 at leu2D1) andstrain JS215-10 (mURA3-LEU2 in rDNA) (57) to obtain SGS1 andsgs1DkanMX4 spores from the same tetrad. Individual spores were used to seed10-ml cultures of YPD medium, which were grown to saturation at 30°C. Tenfoldserial dilutions of each culture were made in sterile distilled water, and 5 ml ofeach dilution was spotted onto SC, SC-Ura, and 5-fluoro-orotic acid (5-FOA)agar.
Ty1 cDNA-mediated mobility assay. The rate of His1 prototroph formation instrains containing a Ty1his3AI element was determined by the maximum-likeli-hood method (37). Strains JC236, JC2359, JC1109, JC2378, JC242, JC2360,JC2712, and JC2698 were grown to saturation in YPD medium at 30°C. For eachstrain, 9 or 11 tubes containing 2 ml of YPD medium were inoculated with '500cells and grown to saturation at 20 or 23°C. The cell count in three cultures wasdetermined by determining the titers on YPD plates. All cultures were subse-quently plated to SC-His medium and grown at 30°C to quantify His1 pro-totrophs. SGS1 and sgs1DLEU2 congenic pairs were tested in the same experi-ment.
Protein analysis. Western blot analyses were performed on whole-cell yeastextracts prepared from exponential-phase cultures as described previously (2),except that 50 mg of protein was analyzed per lane on a sodium dodecyl sulfate–10% polyacrylamide gel. Proteins were transferred to a polyvinylidene difluoridemembrane and then incubated overnight with TyA1 antiserum (9) diluted1:5,000. Horseradish peroxidase (HRP)-conjugated anti-rabbit antibody (Amer-sham) diluted 1:2,500 was used as a secondary antibody. Subsequently, themembrane was incubated with goat antisera raised against N- and C-terminalpeptides of Fus3 (Santa Cruz Biotechnology, Inc.), which were combined andused at a 1:2,000 dilution for 1 to 2 h. HRP-conjugated anti-goat antibody (SantaCruz Biotechnology, Inc.) diluted 1:5,000 was used as a secondary antibody.
Subcellular fractions enriched for Ty1 VLPs were obtained by fractionating105 A260 units of cell lysate from each strain on 20 to 75% sucrose step gradients,as described previously (9). Approximately 50 mg of protein from each VLP-enriched fraction was analyzed on a sodium dodecyl sulfate–8% polyacrylamidegel and then transferred to a polyvinylidene difluoride membrane. The mem-brane was incubated sequentially with TYA1 antiserum diluted 1:5,000, B2
antiserum (66) diluted 1:5,000, and B8 antiserum (23) diluted 1:5,000. HRP-conjugated anti-rabbit antibody diluted 1:2,500 was used as a secondary antibody.
Immune complexes were detected by incubating membranes with 1.25 mMLuminol (Sigma) in 0.1 M Tris-HCl (pH 8.5)–0.68 mM para-coumaric acid–0.09% hydrogen peroxide for 1 min, followed by autoradiography. Quantitationof specific bands detected by autoradiography was performed by scanning den-sitometry using a Howtek Scanmaster31 and Scanalytics software.
Quantitation of Ty1 cDNA. Two independent colonies of each yeast strainwere grown on YPD medium at 20°C and then inoculated into a 10-ml culture ofYPD medium and grown to saturation at 20°C. Genomic DNA was preparedfrom each culture as described previously (9). DNA samples were digested withPvuII and subjected to electrophoresis on a 1% Seakem-GTG agarose (FMC)gel, followed by transfer of the DNA to a Hybond N1 membrane (Amersham).A 32P-labeled riboprobe containing sense-strand TYB1 sequences was synthe-sized using plasmid pGEM-TYB1. Bands were quantified by phosphorimageanalysis using a Molecular Dynamics Phosphorimager and ImageQuant soft-ware.
Ty1 integration into the CAN1 locus. Strains JC242 and JC2360 were grownovernight in YPD medium at 30°C. Twenty 2-ml cultures of YPD were inocu-lated with '500 cells and grown at 20°C to saturation. The titers of four cultureswere determined by plating dilutions on YPD medium, and the number ofcanavanine-resistant (Canr) cells in each culture was determined by plating onSC-Arg1Can medium. The rate of resistance to canavanine was calculated bythe maximum-likelihood method (37). To determine the fraction of can1 muta-tions caused by insertion of a Ty1 element, one Canr colony was obtained from40 independent cultures. Genomic DNA was prepared from each strain, and'250 ng of DNA was used as a template in PCR analysis with AX016 (GAAAATTTCGAGGAAGACGATAAGG) and AX019 (CAAATGCTTCTACTCCGTCTGC) to amplify a 2,265-bp fragment of the CAN1 gene (9). DNA samplesthat yielded no CAN1-specific PCR product were subjected to two additionalPCR amplifications with the CAN1-specific primer AX016 or AX019 and a Ty1LTR-specific oligomer, AX015 (GCCTTTATCAACAATGGAATCCC), to am-plify a Ty1:can1 junction fragment, if present. The rate of Ty1 transposition intoCAN1 in strain JC2360 was calculated by multiplying the rate of resistance to
TABLE 1. Yeast strains
Name Relevant genotype Reference or source
GRF167 MATa his3D200 ura3-167 5DG789 GRF167 spt3-101 14JC236 GRF167 Ty1his3AI-236r 14JC280 GRF167 Ty1his3AI-236r ubc2D:hisG 12JC1427 GRF167 Ty1his3AI-236r leu2:hisG This workJC2359 GRF167 Ty1his3AI-236r leu2:hisG sgs1DLEU2 This workMBY1487 GRF167 Ty1HIS3-236r leu2:hisG This workMBY1049 GRF167 Ty1HIS3-236r leu2:hisG sgs1DLEU2 This workJC242 GRF167 Ty1his3AI-242 14JC544 GRF167 Ty1his3AI-242 ubc2D:hisG 12JC1430 GRF167 Ty1his3AI-242 leu2:hisG 9JC2360 GRF167 Ty1his3AI-242 leu2:hisG sgs1DLEU2 This workJC2698 GRF167 Ty1his3AI-242 leu2:hisG sgs1DLEU2 rad52:URA3 This workJC2712 GRF167 Ty1his3AI-242 leu2:hisG rad52:URA3 9JC2148 GRF167 Ty1his3AI-242 leu2:hisG tec1:URA3 10JC959 GRF167 Ty1HIS3-242 6MBY1050 GRF167 Ty1HIS3-242 leu2:hisG sgs1DLEU2 This workJC1109 MATa his3D200 ura3(252 or 2167) trp1(2289 or :hisG) ade2D:hisG Ty1ade2AI-515
Ty1his3AI-816rThis work
JC2102 MATa his3D200 ura3(252 or 2167) trp1(2289 or :hisG) ade2D:hisG Ty1ade2AI-515Ty1his3AI-816r leu2:hisG
This work
JC2378 MATa his3D200 trp1-289 ade2D:hisG ura32 Ty1ade2AI-515 Ty1his3AI-816r leu2:hisG sgs1DLEU2
This work
MBY1008 MATa his3D200 trp1-289 ade2D:hisG ura32 Ty1ade2AI-515 Ty1HIS3-816r This workMBY1047 MATa his3D200 trp1-289 ade2D:hisG ura32 Ty1ade2AI-515 Ty1HIS3-816r leu2:hisG sgs1
DLEU2This work
BY4742 MATa his3D1 leu2D0 lys2D0 ura3D0 Research GeneticsBY4742-10775 MATa his3D1 leu2D0 lys2D0 ura3D0 sgs1DkanMX4 Research GeneticsJS210-1 MATa his3D200 ura3-167 trp1D63 leu2D1:mURA3-LEU2 57JS215-10 MATa his3D200 ura3-167 trp1D63 RDN1:mURA3-LEU2 57YH8 MATa his3D200 ura3-167 leu2D1 trp1D1 64JC3161 MATa his3D200 ura3-167 leu2D1 trp1D1 sgs1DLEU2 This work
5376 BRYK ET AL. MOL. CELL. BIOL.
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

canavanine by the fraction of Canr strains that contained a Ty1 element withinCAN1.
Ty1HIS3 cDNA-genomic Ty1his3AI element recombination assay. PlasmidpJC573 was linearized at the PacI site in BIK1-HIS4 intergenic DNA and used totransform strains BY4742, BY4742-10775, YH8, and JC3161. Ura1 transfor-mants were grown as large patches on SC-Ura medium at 30°C and then repli-cated to two YPD plates, one of which was grown at 20°C and the other of whichwas grown at 30°C. Following growth for 3 days, patches of cells were replicatedto SC-Ura-His medium to select for His1 Ura1 colonies that sustained aTy1HIS3 insertion and retained the Ty1his3AI[D1]-URA3 cassette. No His1
Ura1 prototrophs were detected following growth on YPD at 30°C, indicatingthat His1 Ura1 colonies that arose at 20°C were independent. His1 Ura1
colonies were transferred as small patches to YPD medium. Following growth at30°C for 2 to 3 days, patches of cells were replicated to 5-FOA–His medium andgrown for 3 days at 30°C. The fraction of His1 colonies that failed to generateHis1 Ura2 derivatives was determined.
PCR-based detection of multimeric Ty1HIS3 transposition events. Indepen-dent His1 colonies that sustained an insertion of Ty1HIS3 by retrotranspositionwere obtained as follows. Yeast strain JC236 and isogenic sgs1D, rad52, and sgs1Drad52 derivatives were single-colony purified and then spread onto YPD platesand grown at 30°C. After 2 days, the cells were replicated to SC-His to check thatno preexisting His1 colonies were present. Cells were also replicated to a freshYPD plate and grown at 20°C for three days to induce transposition. The cellswere subsequently replicated to SC-His medium and grown at 30°C. His1 pa-pillae were single-colony purified, and cells in a medium-sized colony were lysedusing Lyse-N-Go PCR Reagent (Pierce Chemical Co.), according to the manu-facturer’s specifications. Lysates were used in PCR amplification reactions witholigomers HISOUT-2 (GTACTAGAGGAGGCCAAGAG) and TYA1OUT-2(TCTCTGGAACAGCTGATGAAG). Oligomers TEL1-F (CGGATTTCTGACGATATGGAC) and TEL1-R (ACCAACGTACTGAAGGTATCC), which am-plify a 475-bp fragment of the TEL1 locus, were included in each PCR amplifi-cation to ensure that the genomic DNA was PCR competent. PCR mixtures wereincubated at 94°C for 30 s, 63°C for 30 s, and 72°C for 30 s, followed by 29 cyclesin which the annealing temperature (63°C) was successively lowered by 0.3°C. Aportion of each reaction mixture was analyzed on a 2% agarose gel.
Sequence analysis of 3* Ty1HIS3:genomic DNA junctions. Plasmid pGTy1-H3his3AI (14) was introduced into sgs1D strain JC3161. Ura1 transformantswere grown on SC-Ura–2% galactose medium at 20°C for 4 days to induceexpression of the pGTy1-H3his3AI element and subsequently plated on SC-His–2% glucose to isolate colonies containing Ty1HIS3 transpositions. Ura2
segregants were identified following single-colony purification of His1 isolates onYPD plates. Independent His1 Ura2 colonies containing a single .3.0-kb PvuIIfragment that hybridized to a 32P-labeled HIS3 riboprobe in Southern blotanalysis were identified. The 39 Ty1HIS3:genomic DNA junction was cloned byintegration and eviction of plasmid pGEM-URA3-HIS3 DNA linearized withNheI. The Ty1HIS3:genomic DNA junction fragments were evicted from pGEM-URA3-HIS3 transformants by digestion of genomic DNA that was prepared withAatII and ligation at ,0.1 mg of DNA/ml. Ligation reactions were used totransform Escherichia coli to ampicillin resistance. DNA sequencing was per-formed with the HIS3-specific primer AX009 (CTTTATCAACAATGGAATCCC), and sequencing data were analyzed by BLASTN homology search with theSaccharomyces Genome Database (http://genome-www2.stanford.edu/cgi-bin/SGD/).
RESULTS
Mitotic stability and transcriptional silencing of Ty1 ele-ments in rDNA. We demonstrated previously that Ty1 ele-ments located in the rDNA are subject to transcriptional si-lencing (6). To monitor the effect of deleting SGS1 ontranscriptional silencing of Ty1 elements in the rDNA, wequantified RNA from individual Ty1 elements marked with thehis3AI gene and from genomic Ty1 elements collectively inwild-type, sgs1D, and ubc2D cells (Fig. 1). RNA of the rDNAelement Ty1his3AI-236r was found at very low abundance inthe SGS1 strain JC236 because of rDNA silencing, and it wasincreased only twofold in an isogenic sgs1D strain (Fig. 1, lanes1 and 3). In contrast, deletion of UBC2, which is required forrDNA-specific transcriptional silencing (6), increased Ty1his3AI-236r RNA 13.3-fold (lanes 1 and 2). RNA of the euchromatic
element Ty1his3AI-242 was not significantly altered by deletionof SGS1 (lanes 4 and 6) or UBC2 in strain JC242 (lanes 4 and5). Deletion of SGS1 caused a minor increase in total Ty1RNA levels in both strains (lanes 1, 3, 4, and 6), as did deletionof UBC2 (lanes 1, 2, 4, and 5). In summary, the data show thatSGS1 is not involved in transcriptional silencing of rDNA Ty1elements or in the regulation of total Ty1 RNA levels.
Because the mitotic stability of marker genes in rDNA isdependent on Sgs1 (21, 61), we expected that recombination ofTy1 elements in rDNA would also be repressed by Sgs1. Toconfirm this, the mitotic stability of Ty1 insertions was analyzedby replacing the his3AI marker in different Ty1 elements witha HIS3 allele. We then determined the frequency of HIS3marker loss in isogenic SGS1 and sgs1D strains (Table 2). Therate of loss per generation of the rDNA element Ty1HIS3-236rwas increased 2.9-fold, and that of the rDNA elementTy1HIS3-816r was increased 7.0-fold, in sgs1D mutants. In con-trast, the rate of loss of Ty1HIS3-242, located outside therDNA, was decreased fivefold in an sgs1D derivative. Thesedata demonstrate that unlike previously analyzed regulators ofrDNA silencing and recombination, Sgs1 represses recombi-
FIG. 1. SGS1 is not required for transcriptional silencing ofTy1his3AI-236r in rDNA. Total RNA samples from strains JC236 andJC242 and sgs1D and ubc2D derivatives were analyzed by Northernblotting. The blot was hybridized to a 32P-labeled sense-strand HIS3riboprobe to detect Ty1his3AI RNA (top), an antisense Ty1 riboprobeto detect total Ty1 RNA (middle), and an antisense PYK1 riboprobe asa loading control (bottom). The ratios of Ty1his3AI-236 to PYK1 RNA,normalized to the SGS1 strain (lane 1), were 13.3 (ubc2D; lane 2) and2.0 (sgs1D; lane 3); the Ty1his3AI-242/PYK1 RNA ratios, normalizedto SGS1 (lane 4), were 1.1 (ubc2D; lane 5) and 1.5 (sgs1D; lane 6); theTy1/PYK1 RNA ratios, normalized to SGS1 (lane 1), were 1.5 (ubc2D;lane 2) and 1.7 (sgs1D; lane 3); and the Ty1/PYK1 RNA ratios, nor-malized to SGS1 (lane 4), were 1.5 (ubc2D; lane 5) and 1.7 (sgs1D; lane6).
VOL. 21, 2001 Sgs1 INHIBITS TRANSPOSITION OF MULTIMERIC Ty1 ELEMENTS 5377
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

nation without affecting transcriptional silencing at the rDNA.Therefore, silencing of Pol II transcription and repression ofrDNA repeat recombination are separable functions.
Silencing of the mURA3 marker in rDNA. To corroboratethe finding that SGS1 does not regulate transcriptional silenc-ing of Ty1 elements in rDNA, we analyzed the effect of thesgs1D mutation on silencing of the mURA3 gene in rDNAusing quantitative growth assays (Fig. 2). An mURA3-LEU2cassette was integrated in rDNA or at the leu2D1 locus, andserial dilutions of cells from SGS1 and sgs1D spores containingthe mURA3-LEU2 cassettes were plated on different media. InSGS1 strains, expression of mURA3 in rDNA was reducedrelative to the same marker at leu2D1, resulting in relativelyweaker growth on SC-Ura and stronger growth on 5-FOAmedium. This result reflects the fact that mURA3 is subject totranscriptional silencing in the rDNA (57). Deletion of SGS1did not relieve silencing of mURA3 in the rDNA (comparegrowth on SC, SC-Ura, and 5-FOA plates in columns 3 and 4or 5 and 6 of Fig. 2). As expected, deletion of SGS1 also did notaffect expression of mURA3 at leu2D1 (compare growth on SC,SC-Ura, and 5-FOA plates in columns 1 and 2). In summary,our data demonstrate that Sgs1 is not required for transcrip-tional silencing of the mURA3-LEU2 cassette or Ty1his3AIelements in rDNA.
Global increase in the cDNA-mediated mobility ofTy1his3AI elements in sgs1D mutants. To determine if Sgs1 isinvolved in maintaining transpositional dormancy, we mea-sured the effect of deleting SGS1 on the mobility of individualTy1 elements marked with his3AI. The his3AI RIG allows thecDNA-mediated mobility of Ty1 elements to be detected in aquantitative phenotypic assay for His1 prototroph formation(Fig. 3). Because the formation of His1 prototrophs is abso-lutely dependent on splicing of the intron from the Ty1his3AItranscript (14), this assay detects only cDNA-mediated trans-position or recombination events. The rate of His1 prototrophformation in a strain containing the rDNA element Ty1his3AI-236r was increased 79-fold as a result of deleting SGS1 (Table3). Similarly, deletion of SGS1 resulted in a 43-fold increase inthe cDNA-mediated mobility of a second element in therDNA, Ty1his3AI-816r, and a 32-fold increase in the mobilityof the euchromatic element, Ty1his3AI-242. These data dem-onstrate that Sgs1 is a global repressor of Ty1 mobility. Fur-thermore, the data are consistent with the finding that Sgs1does not specifically affect expression of Ty1 elements in therDNA. The increased Ty1 mobility in the absence of signifi-cantly elevated Ty1 RNA levels in sgs1D mutants (Fig. 1) sug-
gests that Sgs1 inhibits a posttranscriptional step in Ty1 retro-transposition.
Sgs1 affects a posttranslational step in retrotransposition.To determine if the sgs1D mutation affects the level of Ty1proteins, total cell proteins isolated from two different SGS1strains and isogenic sgs1D mutants were analyzed on a Westernblot. The blot was probed with antiserum against TyA1 proteinand subsequently with antiserum against Fus3, which served asa loading control (Fig. 4A). The levels of TyA1 protein relativeto Fus3 in the sgs1D mutants were 1.0- and 2.0-fold that ofTyA1 in the isogenic SGS1 strains. As a control, we showedthat an spt3 mutant had undetectable levels of TyA1 protein.TyB1 proteins are present in very low levels in whole-cellextracts and are therefore difficult to detect. For that reason,we obtained subcellular fractions enriched for Ty1 VLPs fromthe same SGS1 and sgs1D strain pairs. Equal amounts of pro-tein from each VLP-enriched fraction were analyzed with an-tisera directed against TyA1, IN, and RT (Fig. 4B). Only veryminor increases in the levels of TyA1 (1.6- and 1.3-fold),p60-RT (1.5- and 1.6-fold), and p90-IN (2.3- and 2.0-fold) wereobserved in the sgs1D mutants. These data suggest that Sgs1does not directly regulate the synthesis or stability of Ty1proteins.
To determine if Sgs1 inhibits Ty1 transposition by promotingthe degradation of Ty1 cDNA, we quantified unintegratedlinear Ty1 cDNA in an sgs1D strain relative to that in anisogenic SGS1 strain by Southern analysis. A TYB1 probe hy-bridized to total cellular DNA digested with PvuII detected a2.0-kb fragment of Ty1 linear extrachromosomal cDNA as wellas multiple .2.0-kb PvuII fragments consisting of the junctionbetween the 39 end of a genomic Ty1 element and flankingDNA (Fig. 5A). Ty1 cDNA levels were determined by quan-tifying the intensity of the 2.0-kb Ty1 cDNA band (band C inFig. 5B) relative to the intensities of two Ty1:genomic DNAjunction bands (bands G1 and G2 in Fig. 5B) in two DNAsamples from each strain. Deletion of SGS1 increased the levelof Ty1 cDNA 2.6-fold. As a control, we showed that Ty1 cDNAwas decreased 4.0-fold in a tec1 mutant, which has significantlyreduced levels of Ty1 RNA (36). The simplest explanation forthe 2.6-fold increase in cDNA is that it results from the minorincrease in Ty1 proteins in the sgs1D mutant. Therefore, ourresults suggest that Sgs1 does not significantly inhibit the syn-thesis or stability of Ty1 cDNA.
Can the minor increase in Ty1 cDNA levels in the sgs1Dmutant explain the 32- to 79-fold increases in transposition?We addressed this question by comparing the increase in Ty1
TABLE 2. Mitotic stability of Ty1HIS3 elements
Strain Ty1HIS3element
Relevantgenotype
No. of His2 auxotrophs/no. of cellsanalyzed (fraction)a HIS3 loss/generation Loss relative
to SGS1 strain
MBY1487 -236r SGS1 105/1,124 (0.093) 7.8 3 1024
MBY1049 -236r sgs1D 297/1,093 (0.272) 2.3 3 1023 2.9MBY1008 -816r SGS1 19/380 (0.050) 4.2 3 1024
MBY1047 -816r sgs1D 367/1,053 (0.348) 2.9 3 1023 7.0JC959 -242 SGS1 137/3,079 (0.044)b 3.7 3 1024
MBY1050 -242 sgs1D 10/972 (0.010) 8.6 3 1025 0.2
a Determined after 120 generations of nonselective growth in YPD liquid medium. Fractions are averages from two or three independent cultures.b Includes data from reference 6.
5378 BRYK ET AL. MOL. CELL. BIOL.
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

cDNA resulting from deletion of SGS1 to that resulting fromloss of function of another characterized regulator of Ty1transposition. Rad52 is an inhibitor of Ty1 transposition thatreduces the level of Ty1 cDNA (50). Deletion of RAD52 in a
strain harboring Ty1his3AI-242 resulted in a 25-fold increase inthe rate of His1 prototroph formation, which was similar inmagnitude to the 32-fold increase resulting from deletion ofSGS1 in the same strain (Table 3). However, the level of Ty1cDNA was increased 9.6-fold by deletion of RAD52, which wasalmost four times higher than the 2.6-fold increase resultingfrom deletion of SGS1 (Fig. 5B). Similar to a rad52 mutant, anisogenic sgs1D rad52 strain displayed an 11.3-fold increase inTy1 cDNA, suggesting that deletion of SGS1 resulted in littleor no increase in Ty1 cDNA in the absence of Rad52. Insummary, even though the repression of Ty1 mobility by Rad52was similar in magnitude to the repression caused by Sgs1,Rad52 mediated a significant reduction in Ty1 cDNA levels,whereas Sgs1 mediated a relatively minor reduction. The datasuggest that Sgs1 inhibits the mobility of Ty1 elements at a stepfollowing cDNA accumulation. Therefore, Sgs1 may regulatethe insertion of Ty1 cDNA into the genome.
Integration of Ty1 in sgs1D mutants. The hypothesis thatSgs1 represses integration of Ty1 cDNA was explored usingtwo different assays. First, a PCR-based assay was employed todetect de novo Ty1 integration events upstream of Pol IIItranscription units (38; M. Bryk and M. J. Curcio, unpublishedresults). Unselected integration events upstream of two typesof targets, the 16 glycyl-tRNA genes or the 100 to 200 5S rRNAgenes, were detected in DNA samples prepared from cellsgrown at 20°C, the permissive temperature for transposition.No significant change in the pattern or intensity of integrationevents was observed at either target set in an sgs1D mutantcompared to an isogenic SGS1 strain (data not shown). Thesame conclusion was drawn from quantitation of spontaneousTy1 insertions into the selectable target gene, CAN1, in SGS1and isogenic sgs1D strains. Loss-of-function mutations inCAN1 cause resistance to canavanine. Deletion of SGS1 re-sulted in a 2.4-fold increase in the rate of resistance to cana-vanine; however, the fraction of canavanine-resistant mutantswith a Ty1 insertion in CAN1 was not significantly increased inthe sgs1D strain (1/40) relative to the isogenic SGS1 strain(0/40). The rate of Ty1 transposition into CAN1 in the sgs1Dstrain JC2360 (1.15 3 1028) was equivalent to that previouslymeasured in a wild-type strain and was significantly lower thanthat in a fus3D mutant, despite the fact that the mobility ofTy1his3AI elements is elevated to similar levels in sgs1D andfus3D mutants (9). In summary, these data indicate that dele-tion of SGS1 does not significantly increase the frequency ofTy1 integration at preferred or selected target sites.
Recombination between Ty1 cDNA and genomic Ty1 ele-ments. In the absence of an increase in Ty1 integration, en-hanced recombination of Ty1 cDNA might explain how Ty1cDNA-mediated mobility events are increased in sgs1D mu-tants. Rad52 is required for homologous recombination ofcDNA (15, 53). Therefore, we determined whether Rad52 isrequired for the increase in Ty1HIS3 cDNA-mediated mobilityevents in an sgs1D mutant. Deletion of SGS1 in the rad52 strainJC2712, which contains Ty1his3AI-242, increased the rate ofHis1 formation 1.7-fold (Table 3). This is a minor increasecompared to the 32-fold increase resulting from deletion ofSGS1 in the isogenic RAD52 strain JC242. Hence, the datasuggest that Rad52 is required for most of the stimulation ofTy1HIS3 cDNA-mediated mobility in sgs1D mutants. There-fore, Sgs1 may repress recombination of Ty1 cDNA with
FIG. 2. Quantitative growth assay measuring the effect of sgs1D onsilencing of the mURA3 gene in rDNA. Tenfold serial dilutions ofsaturated cultures of strains containing the mURA3-LEU2 cassette atleu2D1 (columns 1 and 2) or in rDNA (columns 3 to 6) were spottedonto SC, SC-Ura, and 5-FOA plates. The plates were incubated for 3days at 30°C before being photographed. The strain pairs in columns 1(SGS1) and 2 (sgs1D), columns 3 (sgs1D) and 4 (SGS1), and columns5 (SGS1) and 6 (sgs1D) are progeny from the same tetrad.
VOL. 21, 2001 Sgs1 INHIBITS TRANSPOSITION OF MULTIMERIC Ty1 ELEMENTS 5379
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

genomic Ty1 elements or recombination between Ty1 cDNAmolecules prior to integration.
To differentiate between these models, we developed anassay to measure recombination between Ty1 cDNA and agenomic Ty1 element (Fig. 6). A cassette containing Ty1his3AIand URA3 between 1.2-kb direct repeats of BIK1-HIS4 inter-genic DNA was introduced into yeast by integration of plasmidpJC573. The cassette was used to determine how frequentlyTy1HIS3 cDNA recombines with the genomic Ty1his3AI ele-ment in SGS1 and sgs1D strains. Recombination of Ty1HIS3cDNA with the Ty1his3AI element would give rise to a His1
cell that contained Ty1HIS3 adjacent to URA3 and flanked bythe 1.2-kb direct repeats. Therefore, selection for loss of URA3by recombination between the flanking direct repeats wouldresult in concomitant loss of the functional HIS3 gene.
Strains containing the Ty1his3AI-URA3 cassette were grownat 20°C on rich medium, and then independent His1 Ura1
colonies that sustained a Ty1HIS3 cDNA-mediated mobilityevent were selected. To determine if the His1 Ura1 strainscould give rise to His1 Ura2 derivatives that retain a genomicTy1HIS3 element but have lost the Ty1his3AI-URA3 cassette,each isolate was transferred to 5-FOA–His medium. Inabilityof strains to grow on 5-FOA–His medium indicated that HIS3was cosegregating with URA3, which was the phenotype ex-pected if Ty1HIS3 cDNA had recombined with the Ty1his3AIelement next to URA3. Out of 305 His1 Ura1 coloniesscreened from the SGS1 strain BY4742, 3 (1%) were His2
Ura2 cosegregants. Similarly, 3 out of 286 (1%) independentHis1 Ura1 colonies of the isogenic sgs1D strain were His2
Ura2 cosegregants. When the Ty1his3AI-URA3 cassette wasintegrated in a second strain background, 4 out of 101 (4%)His1 Ura1 colonies of the SGS1 strain YH8 were His2 Ura2
cosegregants, whereas 5 out of 103 (5%) His1 Ura1 coloniesof the isogenic sgs1D strain were His2 Ura2 cosegregants. In
TABLE 3. cDNA-mediated mobility of genomic Ty1his3AI elements
Strain Relevant genotypeHis1 prototroph formation/cell/generation (mean 6 SE) Ratio of rates
(sgs1D/SGS1)SGS1 sgs1D
JC236 Ty1his3AI-236r (2.4 6 1.6) 3 1028 (1.9 6 0.8) 3 1026 79JC1109 Ty1his3AI-816r (1.2 6 0.88) 3 1028 (5.1 6 2.16) 3 1027 43JC242 Ty1his3AI-242 (2.4 6 1.0) 3 1027 (7.7 6 3.0) 3 1026 32JC2712 Ty1his3AI-242 rad52 5.9 3 1026 1.0 3 1025 1.7
FIG. 3. A phenotypic assay for Ty1 cDNA-mediated mobility events, which utilizes a genomic Ty1 element (tripartite rectangles flanking a blackrectangle) marked with the his3AI RIG. his3AI consists of an AI interrupting the HIS3 coding sequence (broken rectangle). The AI is in theantisense orientation relative to HIS3 transcription; consequently, his3AI is nonfunctional and cells containing it, which also harbor a deletion ofthe chromosomal HIS3 locus, are phenotypically His2. The his3AI gene is placed within the Ty1 element in the opposing transcriptionalorientation. Therefore, the AI is in the sense orientation in the Ty1his3AI RNA (wavy line) and can be removed by splicing. When the splicedtranscript is used as a template for reverse transcription, a linear double-stranded Ty1 cDNA containing a functional copy of the HIS3 gene isformed. Insertion of Ty1HIS3 cDNA into the genome by IN-mediated transposition (left) or Rad52-mediated recombination with preexistinggenomic Ty1 elements (right) results in formation of a His1 prototroph.
5380 BRYK ET AL. MOL. CELL. BIOL.
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

summary, these findings demonstrate that deletion of SGS1does not stimulate recombination between Ty1 cDNA andgenomic Ty1 elements.
Formation of multimeric Ty1 arrays during transposition insgs1D mutants. Our results suggested that Sgs1 inhibits a Ty1cDNA-mediated mobility process that is Rad52 dependent butdoes not involve recombination of Ty1 cDNA with genomicTy1 elements. Therefore, we hypothesized that Ty1 cDNAmolecules undergo intermolecular recombination in sgs1D mu-tants, forming tandem Ty1 arrays that transpose into the ge-nome. This process would increase the number of Ty1 cDNAmolecules per transposition event in sgs1D mutants.
To test this hypothesis, we isolated spontaneous His1 pro-totrophs from strains containing the chromosomal Ty1his3AI-236r element and determined if the transposed Ty1HIS3 in-sertion was present in a Ty1 tandem array. His1 prototrophsgenerated from a genomic Ty1his3AI element typically containonly one HIS3-marked Ty1 at a dispersed genomic location(14). DNA derived from independent His1 strains was sub-jected to PCR analysis using a HIS3-specific primer and aTYA1-specific primer. These primers allowed detection of Ty1arrays containing the HIS3 marker in the upstream Ty1 ele-ment (Fig. 7A). In the SGS1 strain, 12.5% of independentHis1 isolates contained a Ty1HIS3:Ty1 multimer (Fig. 7B).The fraction of Ty1HIS3:Ty1 arrays was increased more thanfourfold in an isogenic sgs1D derivative, to 53.3% of His1
isolates. The majority of PCR products derived from Ty1HIS3:Ty1 multimers were of a size consistent with the presence ofone LTR at the junction between Ty1 elements. These mul-timers can be explained as arising by homologous recombina-tion between LTRs of different Ty1 cDNA molecules. How-ever, one His1 isolate from the SGS1 strain and three from theisogenic sgs1D strain gave rise to a PCR product indicative oftwo LTRs at the junction between Ty1 elements (Fig. 7). Two-LTR Ty1 arrays probably arose by end joining of LTR se-quences.
Rad52 is required for increased mobility of Ty1 elements insgs1D mutants (Table 3). If transposition of Ty1HIS3:Ty1 mul-timers is responsible for the increased mobility, then we wouldexpect multimer formation in an sgs1D mutant to be Rad52dependent as well. In fact, formation of Ty1HIS3:Ty1 multim-ers was reduced 30-fold in the sgs1D strain when RAD52 wasdisrupted. Moreover, formation of Ty1 multimers was abol-ished in an SGS1 rad52 strain (Fig. 7B). Taken together, ourresults demonstrate that Ty1 cDNA forms multimers by aRad52-dependent process that is stimulated in sgs1D mutants.
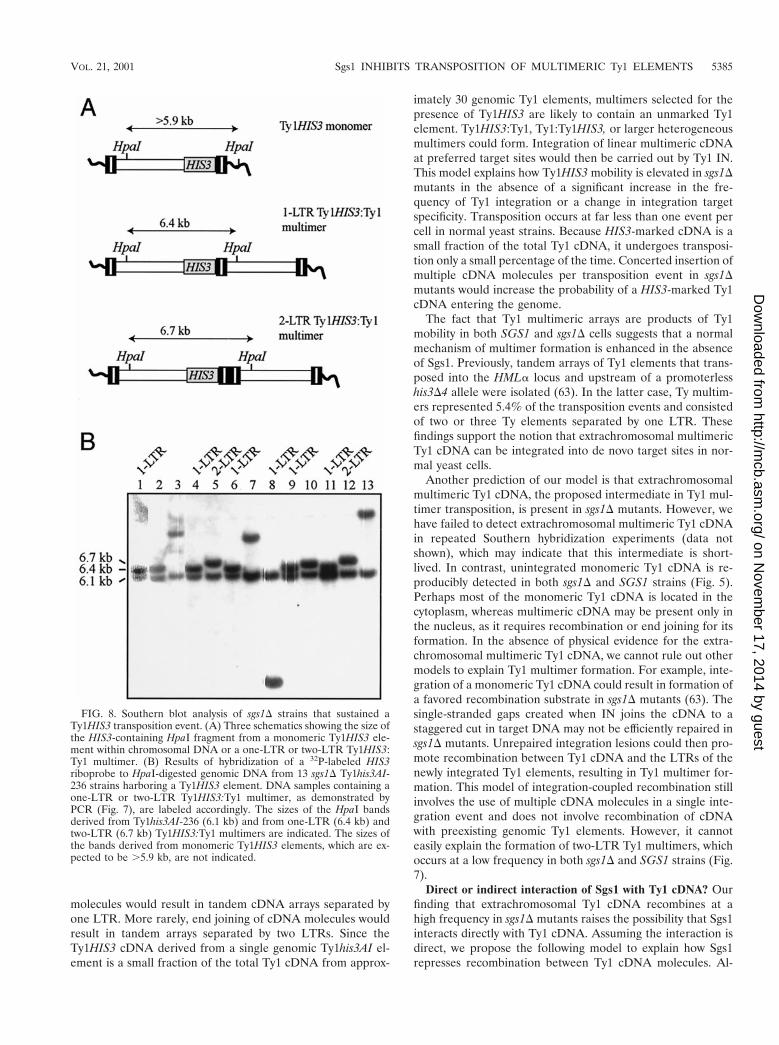
To extend our understanding of the fate of Ty1 cDNA in ansgs1D mutant, 13 of the His1 prototrophs that were analyzedby PCR were also subjected to Southern analysis. GenomicDNA was digested with HpaI, which cleaves once in Ty1 DNA.Hybridization to a HIS3 probe detected a 6.1-kb band derivedfrom Ty1his3AI-236r and a second, variably sized band derivedfrom the transposed Ty1HIS3 element in all 13 DNA samples(Fig. 8). The presence of two HIS3 bands in each DNA sampleindicated that none of the 13 genomic Ty1HIS3 elements hadreplaced the Ty1his3AI-236r element. This finding was consis-tent with data presented above demonstrating that deletion ofSGS1 does not stimulate gene conversion of genomic Ty1elements. DNA from each of the five His1 prototrophs thatcontained a monomeric Ty1HIS3 element yielded a 6.5- to.12-kb band representing the 39 junction of Ty1HIS3 withgenomic DNA at different target sites (Fig. 8B, lanes 1, 2, 7, 10,and 13). DNAs from seven of the eight remaining strainsyielded the predicted HpaI fragment for one-LTR Ty1HIS3:Ty1 multimers (6.4 kb) and two-LTR Ty1HIS3:Ty1 multimers(6.7 kb). The eighth strain had a 3.3-kb HpaI band, indicativeof a deletion or rearrangement within the multimeric array(Fig. 8B, lane 8). The presence of only one Ty1HIS3:genomicDNA junction band in addition to the Ty1his3AI-236r band inall eight strains demonstrates that Ty1 multimers are hetero-geneous arrays containing a Ty1HIS3 element and one or moreunmarked Ty1 elements. The same conclusion was drawn from
FIG. 4. Ty1 protein levels in sgs1D mutants. Western blot analyses of Ty1 proteins in total cell lysates (A) and VLP-enriched subcellularfractions (B) from SGS1 strain JC1430, the isogenic sgs1D derivative JC2360, strain JC2102, and the isogenic sgs1D derivative JC2378 are shown.The spt3 strain DG789 was used as a negative control. (A) The Western blot of total cell lysates was probed sequentially with antisera specific forTyA1 and Fus3. (B) The Western blot of VLP fractions was probed sequentially with antisera against TyA1, RT, and IN.
VOL. 21, 2001 Sgs1 INHIBITS TRANSPOSITION OF MULTIMERIC Ty1 ELEMENTS 5381
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

analysis of six isolates of the isogenic SGS1 strain containingTy1HIS3:Ty1 multimers (data not shown). These data confirmthe validity of the PCR assay for detecting Ty1HIS3 multimers.Furthermore, they demonstrate that Ty1HIS3:Ty1 multimersform by intermolecular recombination between a Ty1HIS3cDNA molecule and unmarked Ty1 cDNA molecules prior toor during integration into the genome.
Preferred target sites for Ty1 transposition in sgs1D mu-tants. We have observed a stimulation of Ty1 mobility in sgs1Dmutants that is not associated with elevated Ty1 integrationinto preferred or selected target sites. However, it is possiblethat transposition into a novel target, which would not bedetected by assays for Ty1 integration at glycyl-tRNA, 5SrRNA, or CAN1 genes, occurs at a high frequency in sgs1Dmutants. To test this hypothesis, unselected genomic DNAtargets of Ty1 transposition were identified in sgs1D mutants.Independent His1 colonies that harbored Ty1HIS3 transposi-tions were obtained following galactose induction of a plasmid-borne PGAL1Ty1-H3his3AI fusion element in the sgs1D strain
JC3161. Plasmid segregants were analyzed by Southern blot-ting with a HIS3 probe to identify strains containing oneTy1HIS3 element in genomic DNA. Of 56 PvuII fragmentsderived from Ty1HIS3 elements that were detected, 21(37.5%) were 3.0 kb, which is the length expected for Ty1HIS3elements within multimeric Ty1 arrays. Five strains were iden-tified that contained one .3.0-kb PvuII fragment, indicatingthe presence of a single 39 Ty1HIS3:genomic DNA junctionfragment. The junction fragment was cloned from each of thefive strains by plasmid integration and eviction, and the site ofTy1HIS3 integration into genomic DNA was determined bysequencing and comparison to the Saccharomyces GenomeDatabase. Four of the five Ty1HIS3 elements analyzed werelocated upstream of Pol III-transcribed genes (Table 4), whichare preferred target sites for Ty1 transposition in SGS1 strains.The fifth Ty1HIS3 element analyzed was adjacent to the be-ginning of the TYA2 open reading frame of a Ty2 element. Ty2elements are highly related to Ty1 elements and have nearlyidentical LTRs (34). Thus, this Ty1HIS3 element is a compo-
FIG. 5. Levels of unintegrated Ty1 cDNA in sgs1D, rad52, and sgs1D rad52 mutants. (A) The structures of unintegrated Ty1 cDNA and agenomic Ty1 element are shown. The hatched area within the Ty1 elements (tripartite rectangles bordering an open box) indicates the locationof the TYB1 probe. The locations of relevant PvuII sites are indicated. The TYB1 riboprobe detects a 2.0-kb fragment from the PvuII site in Ty1to the 39 end of the unintegrated Ty1 cDNA and .2.0-kb PvuII fragments from genomic Ty1 elements. (B) Southern blot analysis of PvuII-digestedgenomic DNAs from two independent colonies of each strain grown at 20°C. The strains analyzed are wild-type strain JC1430 (lanes 1 and 2), anisogenic sgs1D derivative (JC2360; lanes 3 and 4), an sgs1D rad52 derivative (JC2698; lanes 5 and 6), a rad52 derivative (JC2712; lanes 7 and 8),and a tec1 derivative (JC2148; lanes 9 and 10). Two Ty1:genomic DNA junction fragments (G1 and G2) detected by the TYB1 riboprobe were usedas controls in the quantification of the Ty1 cDNA band (C) by phosphorimage analysis.
5382 BRYK ET AL. MOL. CELL. BIOL.
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

nent of a heterogeneous Ty multimer containing an upstreamTy1HIS3 element and a downstream Ty2 element separated bya single LTR. In summary, this analysis failed to reveal a noveltarget site preference for Ty1 transposition in sgs1D mutants.As predicted by the effect of Sgs1 on spontaneous transposi-tion, expression of the inducible PGAL1Ty1-H3his3AI elementin an sgs1D mutant resulted in a high frequency of Ty1 multi-mer transposition. We conclude that transposition of Ty1cDNA multimers into typically preferred transposition targetsis primarily responsible for the increase in Ty1 cDNA-medi-ated mobility in sgs1D mutants.
DISCUSSION
Although host-mediated regulation of Ty1 transposition hasbeen evoked for some time, the identity and role of the host
regulators have only recently begun to be elucidated. The find-ings presented here demonstrate that Sgs1 is involved in main-taining transpositional dormancy by a novel mechanism. Sgs1does not regulate transcriptional silencing of Ty1 elementslocated in the rDNA, even though it represses rDNA recom-bination. This finding indicates that Sgs1 inhibits rDNA recom-bination by a mechanism that is independent of maintainingthe rDNA heterochromatin. Moreover, Sgs1 does not have amajor effect of the levels of Ty1 RNA, proteins, cDNA, orintegration. However, Sgs1 was shown to inhibit recombinationof extrachromosomal Ty1 cDNA molecules during transposi-tion. Our data suggest that repression of cDNA recombinationby Sgs1 limits the number of individual Ty1 cDNA moleculesthat are involved in each transposition event.
Sgs1 represses mitotic recombination without affecting genesilencing in the rDNA. Our findings demonstrate that themechanism by which Sgs1 represses recombination in therDNA is independent from the mechanism of maintainingrDNA silencing. The three- to sevenfold increase in the loss ofrDNA-Ty1HIS3 elements in sgs1D mutants (Table 2) is com-parable to the sevenfold increase in loss of marker genes in therDNA that was observed previously (21). Variation in the rateof loss may be due to the different locations of individualTy1HIS3 elements within the rDNA gene array. In contrast tothe requirement for Sgs1 in the mitotic stability of rDNA-Ty1elements, Sgs1 was not required for and may inhibit the sta-bility of euchromatic Ty1 elements.
Despite the role of Sgs1 in repressing rDNA recombination,deletion of SGS1 did not derepress transcription of Ty1 ele-ments (Fig. 1) or the mURA3 marker gene (Fig. 2) in rDNA.Transcriptional silencing in rDNA is dependent on a uniquetype of heterochromatin (6, 57). Several modulators of chro-matin structure have been shown to promote rDNA silencingand increase longevity (6, 20, 52, 57, 58). For example, thehistone deacetylase Sir2 is a dosage-dependent regulator ofrDNA silencing and longevity that also functions at sites ofDNA damage (20, 31, 44, 47, 60). Arguably, loss of rDNAsilencing promotes aging by allowing an increase in rDNArecombination and ERC formation (26, 29). However, ourfindings demonstrate that aging and rDNA recombination canbe stimulated without perturbing rDNA silencing. Therefore,the role of Sgs1 in repressing rDNA recombination may berelated to its function in initiating the repair of DNA lesionsduring replication (7). One model previously proposed to ex-plain the stimulation of rDNA recombination in sgs1 mutantsis that an accumulation of DNA damage outside the rDNAresults in sequestration of Sir2 away from the nucleolus. Ac-cordingly, rDNA silencing is perturbed and rDNA recombina-tion is stimulated (26). Because this model predicts that rDNAsilencing would be disrupted in sgs1 mutants, it is not consis-tent with our observations. An alternative model is that ele-vated rDNA recombination is a consequence of DNA lesionswithin the rDNA in sgs1 mutants. When Sgs1 cannot initiaterepair of these lesions during DNA replication, recombinationbetween rDNA repeats may provide an alternative mechanismto purge the lesions. A recent report suggests that much of therDNA recombination in sgs1D mutants occurs via the single-strand annealing pathway, which could lead to enhancedrDNA recombination without stimulating ERC formation(46). Hence, this model provides an explanation for the stim-
FIG. 6. Assay for recombination between Ty1HIS3 cDNA and agenomic Ty1his3AI element. Yeast strains containing direct repeats ofa 1.2-kb region of yeast DNA (horizontal black arrows) flanking aTy1his3AI element and the URA3 gene were constructed as describedin Materials and Methods. Splicing of AI from Ty1his3AI RNA andsubsequent reverse transcription result in formation of Ty1HIS3cDNA. The Ty1HIS3 cDNA can transpose into nonhomologous targetsites (not illustrated) or recombine with genomic Ty1 elements, includ-ing the Ty1his3AI element that is linked to URA3. One possible mech-anism of recombination in which Ty1HIS3 cDNA initiates gene con-version of the Ty1his3AI element is illustrated in brackets. Geneconversion would result in replacement of Ty1his3AI with a Ty1HIS3element adjacent to URA3. Subsequent selection for the loss of URA3by recombination between the flanking direct repeats would result inconcomitant loss of the Ty1HIS3 element. Therefore, cosegregation ofUra2 and His2 phenotypes would be observed. Wavy line, plasmidvector sequences; double line with open circle, chromosome III.
VOL. 21, 2001 Sgs1 INHIBITS TRANSPOSITION OF MULTIMERIC Ty1 ELEMENTS 5383
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

ulation of rDNA recombination in the absence of an effect onERC formation and rDNA silencing in sgs1D mutants. It doesnot readily provide an explanation for the shortened life spanof sgs1 mutants, which probably involves a high incidence ofmitotic arrest (46).
Rad52 is required for increased Ty1 cDNA mobility in sgs1Dmutants. In strains containing Ty1his3AI elements at differentlocations, deletion of SGS1 caused a marked increase in thecDNA-mediated mobility of Ty1 elements (Table 3) accompa-nied by a twofold or lower increase in Ty1 RNA (Fig. 1).Hence, Sgs1 may be a weak inhibitor of the transcription orstability of Ty1 RNA. Ty1 protein levels (Fig. 4) and cDNAlevels (Fig. 5) are also slightly elevated in sgs1D mutants, prob-ably as a direct result of the small increase in Ty1 RNA,although this cannot be determined with certainty. Severalfindings indicate that modestly elevated Ty1 cDNA levels arenot the major reason for the increase in Ty1 mobility in sgs1Dmutants. First, no significant stimulation of Ty1 integrationupstream of glycyl-tRNA genes or 5S rRNA genes or into theCAN1 locus was observed in sgs1 mutants. These findings donot support a simple model in which elevated levels of Ty1cDNA result in an increase in the number of integration eventsin sgs1D mutants. Second, the level of Ty1 cDNA in an sgs1Dmutant was about one-quarter of that in a rad52 mutant (Fig.5), even though both mutations stimulated the mobility ofTy1his3AI-242 to an equivalent level (Table 3). This findingsuggests that there is a second mechanism of increasing thecDNA-mediated mobility of Ty1 elements in sgs1D mutantsaside from the small increase in Ty1 cDNA. Third, deletion ofSGS1 in a rad52 strain did not cause a significant increase inthe mobility of Ty1his3AI-242 or the level of Ty1 cDNA, indi-cating that Sgs1 inhibits a Rad52-dependent mechanism of Ty1mobility.
Sgs1 inhibits transposition of multimeric Ty1 cDNA. Weenvisaged two Rad52-dependent pathways for entry of Ty1cDNA into the genome that might be stimulated in the absenceof Sgs1. First, recombination of Ty1 cDNA with genomic Ty1
elements might be enhanced, resulting in introduction ofTy1HIS3 cDNA into the genome at an elevated rate. Notably,genomic multimers of Ty5 elements are commonly formed by“ends-out” recombination between Ty5 cDNA and the up-stream LTR of a genomic Ty5 element (32). However, thispossibility was excluded for Ty1 multimers, since recombina-tion between Ty1HIS3 cDNA and a genomic Ty1his3AI ele-ment was not affected by deletion of SGS1. Second, intermo-lecular recombination between extrachromosomal Ty1 cDNAmolecules including Ty1HIS3 cDNA might be stimulated insgs1 mutants, leading to transposition of tandem Ty cDNAarrays. In fact, the fraction of His1 strains that containTy1HIS3 as a component of a multimeric array was fourfoldhigher in sgs1D mutants (Fig. 7). The PCR assay we useddetected multimers minimally containing a Ty1HIS3:Ty1 array.However, the total number of Ty1 elements within each mul-timer could be higher, especially in sgs1D strains. Hence, ele-vated Ty1 mobility in sgs1D mutants could result from in-creases both in the fraction of multimeric transposition eventsand in the average number of Ty1 cDNA molecules per mul-timer. Formation of Ty1 multimers required Rad52 in bothSGS1 and sgs1D strains. Since the increase in Ty1 mobility insgs1D mutants is also dependent on Rad52, our data suggestthat transposition of Ty1 multimers is the primary cause of theelevated Ty1 mobility in sgs1D mutants.
Independent Ty1 multimers in eight sgs1D strains and sixSGS1 strains all contained one Ty1HIS3 element and one ormore unmarked Ty1 elements. Therefore, Ty1 multimer for-mation involves two or more different cDNA molecules. Thisconclusion is also supported by the isolation of a Ty1HIS3:Ty2multimer in an sgs1D mutant. The evidence for heterogeneousTy arrays eliminates models in which multimeric cDNA isformed by rolling-circle replication of a monomeric Ty1HIS3cDNA. Moreover, it supports a model in which intermolecularrecombination between different Ty1 cDNA molecules resultsin the formation of linear cDNA arrays. Crossover recombi-nation between the 39 and 59 LTRs of different Ty1 cDNA
FIG. 7. Deletion of SGS1 increases genomic Ty1HIS3:Ty1 multimers. (A) Tandem arrays of Ty1 elements, consisting of LTRs (tripartiterectangles) bordering a central domain that carries TYA1 and TYB1 (open box), within chromosomal DNA (curved line). The left schematicrepresents a dimeric Ty1 array with a single shared LTR between each coding domain and the HIS3 marker adjacent to the upstream TYB1 domain(one-LTR Ty1HIS3:Ty1 multimer). The right schematic represents a Ty1 dimer with two joined LTRs between coding domains (two-LTRTy1HIS3:Ty1 multimer). PCR primers that hybridize to the HIS3 marker gene and to the TYA1 domain amplify a 570-bp DNA fragment of aone-LTR Ty1HIS3:Ty1 element and a 905-bp DNA fragment of a two-LTR Ty1HIS3:Ty1 element. (B) Independent His1 revertants of the SGS1RAD52 strain JC236 and isogenic mutant derivatives were analyzed by PCR to determine the fraction of His1 revertants that contained a one-LTRTy1HIS3:Ty1 multimer, a two-LTR Ty1HIS3:Ty1 multimer, or either.
5384 BRYK ET AL. MOL. CELL. BIOL.
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
molecules would result in tandem cDNA arrays separated byone LTR. More rarely, end joining of cDNA molecules wouldresult in tandem arrays separated by two LTRs. Since theTy1HIS3 cDNA derived from a single genomic Ty1his3AI el-ement is a small fraction of the total Ty1 cDNA from approx-
imately 30 genomic Ty1 elements, multimers selected for thepresence of Ty1HIS3 are likely to contain an unmarked Ty1element. Ty1HIS3:Ty1, Ty1:Ty1HIS3, or larger heterogeneousmultimers could form. Integration of linear multimeric cDNAat preferred target sites would then be carried out by Ty1 IN.This model explains how Ty1HIS3 mobility is elevated in sgs1Dmutants in the absence of a significant increase in the fre-quency of Ty1 integration or a change in integration targetspecificity. Transposition occurs at far less than one event percell in normal yeast strains. Because HIS3-marked cDNA is asmall fraction of the total Ty1 cDNA, it undergoes transposi-tion only a small percentage of the time. Concerted insertion ofmultiple cDNA molecules per transposition event in sgs1Dmutants would increase the probability of a HIS3-marked Ty1cDNA entering the genome.
The fact that Ty1 multimeric arrays are products of Ty1mobility in both SGS1 and sgs1D cells suggests that a normalmechanism of multimer formation is enhanced in the absenceof Sgs1. Previously, tandem arrays of Ty1 elements that trans-posed into the HMLa locus and upstream of a promoterlesshis3D4 allele were isolated (63). In the latter case, Ty multim-ers represented 5.4% of the transposition events and consistedof two or three Ty elements separated by one LTR. Thesefindings support the notion that extrachromosomal multimericTy1 cDNA can be integrated into de novo target sites in nor-mal yeast cells.
Another prediction of our model is that extrachromosomalmultimeric Ty1 cDNA, the proposed intermediate in Ty1 mul-timer transposition, is present in sgs1D mutants. However, wehave failed to detect extrachromosomal multimeric Ty1 cDNAin repeated Southern hybridization experiments (data notshown), which may indicate that this intermediate is short-lived. In contrast, unintegrated monomeric Ty1 cDNA is re-producibly detected in both sgs1D and SGS1 strains (Fig. 5).Perhaps most of the monomeric Ty1 cDNA is located in thecytoplasm, whereas multimeric cDNA may be present only inthe nucleus, as it requires recombination or end joining for itsformation. In the absence of physical evidence for the extra-chromosomal multimeric Ty1 cDNA, we cannot rule out othermodels to explain Ty1 multimer formation. For example, inte-gration of a monomeric Ty1 cDNA could result in formation ofa favored recombination substrate in sgs1D mutants (63). Thesingle-stranded gaps created when IN joins the cDNA to astaggered cut in target DNA may not be efficiently repaired insgs1D mutants. Unrepaired integration lesions could then pro-mote recombination between Ty1 cDNA and the LTRs of thenewly integrated Ty1 elements, resulting in Ty1 multimer for-mation. This model of integration-coupled recombination stillinvolves the use of multiple cDNA molecules in a single inte-gration event and does not involve recombination of cDNAwith preexisting genomic Ty1 elements. However, it cannoteasily explain the formation of two-LTR Ty1 multimers, whichoccurs at a low frequency in both sgs1D and SGS1 strains (Fig.7).
Direct or indirect interaction of Sgs1 with Ty1 cDNA? Ourfinding that extrachromosomal Ty1 cDNA recombines at ahigh frequency in sgs1D mutants raises the possibility that Sgs1interacts directly with Ty1 cDNA. Assuming the interaction isdirect, we propose the following model to explain how Sgs1represses recombination between Ty1 cDNA molecules. Al-
FIG. 8. Southern blot analysis of sgs1D strains that sustained aTy1HIS3 transposition event. (A) Three schematics showing the size ofthe HIS3-containing HpaI fragment from a monomeric Ty1HIS3 ele-ment within chromosomal DNA or a one-LTR or two-LTR Ty1HIS3:Ty1 multimer. (B) Results of hybridization of a 32P-labeled HIS3riboprobe to HpaI-digested genomic DNA from 13 sgs1D Ty1his3AI-236 strains harboring a Ty1HIS3 element. DNA samples containing aone-LTR or two-LTR Ty1HIS3:Ty1 multimer, as demonstrated byPCR (Fig. 7), are labeled accordingly. The sizes of the HpaI bandsderived from Ty1his3AI-236 (6.1 kb) and from one-LTR (6.4 kb) andtwo-LTR (6.7 kb) Ty1HIS3:Ty1 multimers are indicated. The sizes ofthe bands derived from monomeric Ty1HIS3 elements, which are ex-pected to be .5.9 kb, are not indicated.
VOL. 21, 2001 Sgs1 INHIBITS TRANSPOSITION OF MULTIMERIC Ty1 ELEMENTS 5385
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

though a fraction of Ty1 cDNA in normal cells enters thegenome by integration or recombination, most of it is degraded(39). Hence, some of the nuclear cDNA may not be protectedby Ty1 IN and instead may be recognized as a recombinationsubstrate. By analogy to a chromosomal double-strand break,free cDNA ends are likely to be potent initiators of recombi-nation. Since 59-end resection is the initial step in all pathwaysof double-strand break repair (27), the cDNA ends would beresected by a 59-to-39 exonuclease activity, leaving 39 single-stranded DNA tails. The single-stranded 39 ends would be ableto invade homologous Ty1 cDNA molecules and initiate re-combination, thereby forming structures in which the DNAstrands from different molecules are annealed. Sgs1 may bindthese branched structures and unwind the annealed strands toprevent homologous recombination. In fact, purified Sgs1 hasbeen shown to bind preferentially to forked DNA substrates(3). Following unwinding by Sgs1, partially degraded Ty1cDNA molecules may lack the terminal nucleotides requiredfor recognition by Ty1 IN and be completely degraded. In theabsence of Sgs1, homologous recombination between the LTRsequences of extrachromosomal Ty1 cDNAs results in the for-mation of multimeric Ty1 arrays with double-stranded DNAends restored by gap filling. Hence, these multimers would besuitable substrates for Ty IN to carry out integration.
Because Ty1 cDNA is extrachromosomal and does not con-tain autonomously replicating sequences, it is not expected toundergo DNA replication. Therefore, interaction of Sgs1 withTy1 cDNA would constitute a novel role for Sgs1 outside of itscharacterized role as a component of the pathway that moni-tors DNA replication fork progression (19). The identificationof factors that inhibit Ty1 mobility along with Sgs1 would helpto determine how Sgs1 represses Ty1 cDNA recombination.One interesting candidate is the yeast Ku70 protein, a compo-nent of the Ku heterodimer that binds to double-strand breaksand is required for nonhomologous end joining (43). Ku70 hasbeen shown to bind to Ty1 cDNA and to be required for thehigh levels of transposition that result from induction of aPGAL1Ty1 element (17). Notably, the human Ku heterodimerassociates with the human RecQ homologue WRN and spe-cifically stimulates its exonuclease activity (11, 41). PerhapsSgs1 and Ku associate and bind the ends of Ty1 cDNA toprevent recombination and/or promote integration.
Alternatively, the effect of deleting SGS1 on Ty1 cDNArecombination may be indirect. An elevated level of chromo-somal lesions that cannot be repaired in sgs1D mutants mightsignal the induction of recombination proteins that enhanceintermolecular cDNA recombination. In fact, stochastic cellcycle arrest characteristic of sgs1D mutants is mediatedthrough the RAD9-dependent DNA damage checkpoint path-
way (46). Perhaps specific recombination functions are in-duced through this pathway in the absence of Sgs1.
Implications for the host cell and Ty1 retrotransposons. Theresults presented here implicate a host mechanism in the re-pression of Ty1 cDNA recombination and the transposition ofmonomeric Ty1 elements. This is the first evidence that thehost cell actively represses Ty1 cDNA recombination, and itsuggests that transposition of Ty1 multimers may be disadvan-tageous to the host. Previous results have indicated the impor-tance of the Ty1 IN protein in sequestering cDNA from ho-mologous recombination pathways (53). When IN-mediatedintegration is blocked, Ty1 cDNA forms complex multimericarrays by homologous recombination. Notably, strains contain-ing genomic insertions of complex Ty1 arrays at independentlocations had a growth defect, suggesting that large multimerscan perturb some feature of chromosome organization. In ad-dition, it has been demonstrated that multimeric Ty1 insertionscan disrupt the silent chromatin domain of the HMLa locus,whereas single Ty1 or solo-LTR derivatives of the multimersrestore silencing (45).
Transposition of monomeric cDNA is likely to be importantfor the maintenance of retrotransposons as well. Although ourdata indicate that multimeric Ty1 insertions arise spontane-ously, they are not a stable component of the S. cerevisiaegenome (34). Hence, they may be reduced to monomeric Ty1elements or LTR derivatives by recombination at a relativelyhigh frequency. Therefore, transposition of Ty1 multimericarrays is not likely to be an efficient mechanism for Ty1 tospread throughout the genome.
ACKNOWLEDGMENTS
We thank Ed Louis, Jeff Smith, and David Garfinkel for supplyingplasmids and strains used in this study, the Wadsworth Center Molec-ular Genetics Core Facility for oligonucleotide synthesis and DNAsequencing, and Abram Gabriel, John Mueller, and Keith Derbyshirefor critical reading of the manuscript. Moreover, we are grateful toFred Winston and Winston lab members for support and encourage-ment.
M. Bryk is the recipient of a Special Fellowship from the Leukemia& Lymphoma Society. M. Bryk’s research in Fred Winston’s lab wasfunded by grant GM32967 from the National Institutes of Health. Thiswork was funded by National Institutes of Health grant GM52072 toM. J. Curcio.
REFERENCES
1. Alani, E., L. Cao, and N. Kleckner. 1987. A method for gene disruption thatallows repeated use of URA3 selection in the construction of multiply dis-rupted yeast strains. Genetics 116:541–545.
2. Atkin, A. L., N. Altamura, P. Leeds, and M. R. Culbertson. 1995. Themajority of yeast UPF1 co-localizes with polyribosomes in the cytoplasm.Mol. Biol. Cell 6:611–625.
3. Bennett, R. J., J. A. Sharp, and J. C. Wang. 1998. Purification and charac-terization of the Sgs1 DNA helicase activity of Saccharomyces cerevisiae.J. Biol. Chem. 273:9644–9650.
TABLE 4. Locations of Ty1HIS3 elements in an sgs1D strain
Ty1HIS3insertion Chromosome Nucleotide coordinate 39 of
Ty1 insertionaNeighboring Pol III-transcribed
gene (encoded RNA)Distance (bp) upstream of Pol
III-transcribed gene
-3 V 99793 tM(CAU)E (tRNAMet) 341-22 XII rDNA RDN5 (5S rRNA) 125-56 VIII 475922 tV(CAC)H (tRNAVal) 148-59 XII 1052246 tI(AUU)L2 (tRNAIle) 106
a Chromosomal coordinates are those provided in the Saccharomyces Genome Database (http://genome-www2.stanford.edu/cgi-bin/SGD/).
5386 BRYK ET AL. MOL. CELL. BIOL.
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

4. Boeke, J. D., D. Eichinger, D. Castrillon, and G. R. Fink. 1988. The Sac-charomyces cerevisiae genome contains functional and nonfunctional copiesof transposon Ty1. Mol. Cell. Biol. 8:1432–1442.
5. Boeke, J. D., D. J. Garfinkel, C. A. Styles, and G. R. Fink. 1985. Ty elementstranspose through an RNA intermediate. Cell 40:491–500.
6. Bryk, M., M. Banerjee, M. Murphy, K. E. Knudsen, D. J. Garfinkel, andM. J. Curcio. 1997. Transcriptional silencing of Ty1 elements in the RDN1locus of yeast. Genes Dev. 11:255–269.
7. Chakraverty, R. K., and I. D. Hickson. 1999. Defending genome integrityduring DNA replication: a proposed role for RecQ family helicases. Bioes-says 21:286–294.
8. Christman, M. F., F. S. Dietrich, and G. R. Fink. 1988. Mitotic recombina-tion in the rDNA of S. cerevisiae is suppressed by the combined action ofDNA topoisomerases I and II. Cell 55:413–425.
9. Conte, D., E. Barber, M. Banerjee, D. J. Garfinkel, and M. J. Curcio. 1998.Posttranslational regulation of Ty1 retrotransposition by mitogen-activatedprotein kinase Fus3. Mol. Cell. Biol. 18:2502–2513.
10. Conte, D., and M. J. Curcio. 2000. Fus3 controls Ty1 transpositional dor-mancy through the invasive growth MAPK pathway. Mol. Microbiol. 35:415–427.
11. Cooper, M. P., A. Machwe, D. K. Orren, R. M. Brosh, D. Ramsden, and V. A.Bohr. 2000. Ku complex interacts with and stimulates the Werner protein.Genes Dev. 14:907–912.
12. Curcio, M. J., and D. J. Garfinkel. 1994. Heterogeneous functional Ty1elements are abundant in the Saccharomyces cerevisiae genome. Genetics136:1245–1259.
13. Curcio, M. J., and D. J. Garfinkel. 1992. Posttranslational control of Ty1retrotransposition occurs at the level of protein processing. Mol. Cell. Biol.12:2813–2825.
14. Curcio, M. J., and D. J. Garfinkel. 1991. Single-step selection for Ty1 ele-ment retrotransposition. Proc. Natl. Acad. Sci. USA 88:936–940.
15. Derr, L. K., and J. N. Strathern. 1993. A role for reverse transcripts in geneconversion. Nature 361:170–173.
16. Devine, S. E., and J. D. Boeke. 1996. Integration of the yeast retrotransposonTy1 is targeted to regions upstream of genes transcribed by RNA polymeraseIII. Genes Dev. 10:620–633.
17. Downs, J. A., and S. P. Jackson. 1999. Involvement of DNA end-bindingprotein Ku in Ty element retrotransposition. Mol. Cell. Biol. 19:6260–6268.
18. Ellis, N. A., J. Groden, T. Z. Ye, J. Straughen, D. J. Lennon, S. Ciocci, M.Proytcheva, and J. German. 1995. The Bloom’s syndrome gene product ishomologous to RecQ helicases. Cell 83:655–666.
19. Frei, C., and S. M. Gasser. 2000. The yeast Sgs1p helicase acts upstream ofRad53p in the DNA replication checkpoint and colocalizes with Rad53p inS-phase-specific foci. Genes Dev. 14:81–96.
20. Fritze, C. E., K. Verschueren, R. Strich, and R. E. Esposito. 1997. Directevidence for SIR2 modulation of chromatin structure in yeast rDNA. EMBOJ. 16:6495–6509.
21. Gangloff, S., J. P. McDonald, C. Bendixen, L. Arthur, and R. Rothstein.1994. The yeast type I topoisomerase Top3 interacts with Sgs1, a DNAhelicase homolog: a potential eukaryotic reverse gyrase. Mol. Cell. Biol.14:8391–8398.
22. Gangloff, S., C. Soustelle, and F. Fabre. 2000. Homologous recombination isresponsible for cell death in the absence of the Sgs1 and Srs2 helicases. Nat.Genet. 25:192–194.
23. Garfinkel, D. J., A. M. Hedge, S. D. Youngren, and T. D. Copeland. 1991.Proteolytic processing of pol-TYB proteins from the yeast retrotransposonTy1. J. Virol. 65:4573–4581.
24. Garfinkel, D. J., M. F. Mastrangelo, N. J. Sanders, B. K. Shafer, and J. N.Strathern. 1988. Transposon tagging using Ty elements in yeast. Genetics120:95–108.
25. Gottlieb, S., and R. E. Esposito. 1989. A new role for a yeast transcriptionalsilencer gene, SIR2, in regulation of recombination in ribosomal DNA. Cell56:771–776.
26. Guarente, L. 2000. Sir2 links chromatin silencing, metabolism, and aging.Genes Dev. 14:1021–1026.
27. Haber, J. E. 2000. Partners and pathways: repairing a double-strand break.Trends Genet. 16:259–264.
28. Heo, S.-J., K. Tatebayashi, I. Ohsugi, A. Shimamoto, Y. Furuichi, and H.Ikeda. 1999. Bloom’s syndrome gene suppresses premature ageing caused bySgs1 deficiency in yeast. Genes Cells 4:619–625.
29. Jazwinski, S. M. 2000. Metabolic control and ageing. Trends Genet. 16:506–511.30. Ji, H., D. P. Moore, M. A. Blomberg, L. T. Braiterman, D. F. Voytas, G.
Natsoulis, and J. D. Boeke. 1993. Hotspots for unselected Ty1 transpositionevents on yeast chromosome III are near tRNA genes and LTR sequences.Cell 73:1007–1018.
31. Kaeberlein, M., M. McVey, and L. Guarente. 1999. The SIR2/3/4 complexand SIR2 alone promote longevity in Saccharomyces cerevisiae by two differ-ent mechanisms. Genes Dev. 13:2570–2580.
32. Ke, N., and D. F. Voytas. 1999. cDNA of the yeast retrotransposon Ty5preferentially recombines with substrates in silent chromatin. Mol. Cell. Biol.19:484–494.
33. Kenna, M. A., C. B. Brachmann, S. E. Devine, and J. D. Boeke. 1998.
Invading the yeast nucleus: a nuclear localization signal at the C terminus ofTy1 integrase is required for transposition in vivo. Mol. Cell. Biol. 18:1115–1124.
34. Kim, J. M., S. Vanguri, J. D. Boeke, A. Gabriel, and D. F. Voytas. 1998.Transposable elements and genome organization: a comprehensive survey ofretrotransposons revealed by the complete Saccharomyces cerevisiae genomesequence. Genome Res. 8:464–478.
35. Kitao, S., N. M. Lindor, M. Shiratori, Y. Furuichi, and A. Shimamoto. 1999.Rothmund-thomson syndrome responsible gene, RECQL4: genomic struc-ture and products. Genomics 61:268–276.
36. Laloux, I., E. Dubois, M. Dewerchin, and E. Jacobs. 1990. TEC1, a geneinvolved in the activation of Ty1 and Ty1-mediated gene expression inSaccharomyces cerevisiae: cloning and molecular analysis. Mol. Cell. Biol.10:3541–3550.
37. Lea, D. E., and C. A. Coulson. 1949. The distribution of the numbers ofmutants in bacterial populations. J. Genet. 49:264–285.
38. Lee, B.-S., C. P. Lichtenstein, B. Faiola, L. A. Rinckel, W. Wysock, M. J.Curcio, and D. J. Garfinkel. 1998. Posttranslational inhibition of Ty1 retro-transposition by nucleotide excision repair/transcription factor TFIIH sub-units Ssl2p and Rad3p. Genetics 148:1743–1761.
39. Lee, B. S., L. Bi, D. J. Garfinkel, and A. M. Bailis. 2000. Nucleotide excisionrepair/TFIIH helicases RAD3 and SSL2 inhibit short-sequence recombina-tion and Ty1 retrotransposition by similar mechanisms. Mol. Cell. Biol.20:2436–2445.
40. Lee, S.-K., R. E. Johnson, S.-L. Yu, L. Prakash, and S. Prakash. 1999.Requirement of yeast SGS1 and SRS2 genes for replication and transcrip-tion. Science 286:2339–2342.
41. Li, B., and L. Comai. 2000. Functional interaction between Ku and theWerner syndrome protein in DNA end processing. J. Biol. Chem. 275:28349–28352.
42. Lohman, T. M., and K. P. Bjornson. 1996. Mechanisms of helicase-catalyzedDNA unwinding. Annu. Rev. Biochem. 65:169–214.
43. Lustig, A. J. 1999. The Kudos of non-homologous end-joining. Nat. Genet.23:130–131.
44. Martin, S. G., T. Laroche, N. Suka, M. Grunstein, and S. M. Gasser. 1999.Relocalization of telomeric Ku and SIR proteins in response to DNA strandbreaks in yeast. Cell 97:621–633.
45. Mastrangelo, M. F., K. G. Weinstock, B. K. Shafer, A. M. Hedge, D. J.Garfinkel, and J. N. Strathern. 1992. Disruption of a silencer domain by aretrotransposon. Genetics 131:519–529.
46. McVey, M., M. Kaeberlein, H. A. Tissenbaum, and L. Guarente. 2001. Theshort life span of Saccharomyces cerevisiae sgs1 and srs2 mutants is a com-posite of normal aging processes and mitotic arrest due to defective recom-bination. Genetics 157:1531–1542.
47. Mills, K. D., D. A. Sinclair, and L. Guarente. 1999. MEC1-dependent redis-tribution of the Sir3 silencing protein from telomeres to DNA double-strandbreaks. Cell 97:609–620.
48. Moore, S. P., L. A. Rinckel, and D. J. Garfinkel. 1998. A Ty1 integrasenuclear localization signal required for retrotransposition. Mol. Cell. Biol.18:1105–1114.
49. Puranam, K. L., and P. J. Blackshear. 1994. Cloning and characterization ofRecQL, a potential human homologue of the Escherichia coli DNA helicaseRecQ. J. Biol. Chem. 269:29838–29845.
50. Rattray, A. J., B. K. Shafer, and D. J. Garfinkel. 2000. The Saccharomycescerevisiae DNA recombination and repair functions of the RAD52 epistasisgroup inhibit Ty1 transposition. Genetics 154:543–556.
51. Rose, M. D., F. Winston, and P. Hieter. 1990. Methods in yeast genetics: alaboratory course manual. Cold Spring Harbor Laboratory Press, Plainview,N.Y.
52. Roy, N., and K. W. Runge. 2000. Two paralogs involved in transcriptionalsilencing that antagonistically control yeast life span. Curr. Biol. 10:111–114.
53. Sharon, G., T. J. Burkett, and D. J. Garfinkel. 1994. Efficient homologousrecombination of Ty1 element cDNA when integration is blocked. Mol. Cell.Biol. 14:6540–6551.
54. Sikorski, R. S., and P. Hieter. 1989. A system of shuttle vectors and yeasthost strains designed for efficient manipulation of DNA in Saccharomycescerevisiae. Genetics 122:19–27.
55. Sinclair, D. A., and L. Guarente. 1997. Extrachromosomal rDNA circles—acause of aging in yeast. Cell 91:1033–1042.
56. Sinclair, D. A., K. Mills, and L. Guarente. 1997. Accelerated aging andnucleolar fragmentation in yeast sgs1 mutants. Science 277:1313–1316.
57. Smith, J. S., and J. D. Boeke. 1997. An unusual form of transcriptionalsilencing in the yeast ribosomal DNA. Genes Dev. 11:241–254.
58. Smith, J. S., E. Caputo, and J. D. Boeke. 1999. A genetic screen for ribo-somal DNA silencing defects identifies multiple DNA replication and chro-matin-modulating factors. Mol. Cell. Biol. 19:3184–3197.
59. Swanson, M. S., E. A. Malone, and F. Winston. 1991. SPT5, an essential geneimportant for normal transcription in Saccharomyces cerevisiae, encodes anacidic nuclear protein with a carboxy-terminal repeat. Mol. Cell. Biol. 11:3009–3019.
60. Tsukamoto, Y., J. Kato, and H. Ikeda. 1997. Silencing factors participate in
VOL. 21, 2001 Sgs1 INHIBITS TRANSPOSITION OF MULTIMERIC Ty1 ELEMENTS 5387
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

DNA repair and recombination in Saccharomyces cerevisiae. Nature 388:900–903.
61. Watt, P. M., I. D. Hickson, R. H. Borts, and E. J. Louis. 1996. SGS1, ahomologue of the Bloom’s and Werner’s syndrome genes, is required formaintenance of genome stability in Saccharomyces cerevisiae. Genetics 144:935–945.
62. Watt, P. M., E. J. Louis, R. H. Borts, and I. D. Hickson. 1995. Sgs1: aeukaryotic homolog of E. coli RecQ that interacts with topoisomerase II invivo and is required for faithful chromosome segregation. Cell 81:253–260.
63. Weinstock, K. G., M. F. Mastrangelo, T. J. Burkett, D. J. Garfinkel, and J. N.Strathern. 1990. Multimeric arrays of the yeast retrotransposon Ty. Mol.Cell. Biol. 10:2882–2892.
64. Xu, H., and J. D. Boeke. 1990. Host genes that influence transposition inyeast: the abundance of a rare tRNA regulates Ty1 transposition frequency.
Proc. Natl. Acad. Sci. USA 87:8360–8364.65. Yamagata, K., J.-I. Kato, A. Shimamoto, M. Goto, Y. Furuichi, and H. Ikeda.
1998. Bloom’s and Werner’s syndrome genes suppress hyperrecombinationin yeast sgs1 mutant: implication for genomic instability in human diseases.Proc. Natl. Acad. Sci. USA 95:8733–8738.
66. Youngren, S. D., J. D. Boeke, N. J. Sanders, and D. J. Garfinkel. 1988.Functional organization of the retrotransposon Ty from Saccharomyces cer-evisiae: Ty protease is required for transposition. Mol. Cell. Biol. 8:1421–1431.
67. Yu, C. E., J. Oshima, Y. H. Fu, E. M. Wijsman, F. Hisama, R. Alisch, S.Matthews, J. Nakura, T. Miki, S. Ouais, G. M. Martin, J. Mulligan, andG. D. Schellenberg. 1996. Positional cloning of the Werner’s syndrome gene.Science 272:258–262.
5388 BRYK ET AL. MOL. CELL. BIOL.
on Novem
ber 17, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from




















