
Evolution of bamboo mosaic virus in a nonsystemic host results in mutations in the helicase-like...
10
Virus Research 58 (1998) 127–136 Evolution of bamboo mosaic virus in a nonsystemic host results in mutations in the helicase-like domain that cause reduced RNA accumulation Cheng-Yen Huang a , Ching-Hsiu Tsai a,b, * a Graduate Institute of Agricultural Biotechnology, National Chung Hsing Uni6ersity, Taichung 402, Taiwan b Agricultural Biotechnology Laboratories, National Chung Hsing Uni6ersity, Taichung 402, Taiwan Received 1 May 1998; received in revised form 10 September 1998; accepted 14 September 1998 Abstract A mutant population of bamboo mosaic potexvirus (BaMV) was isolated after serial passage using Chenopodium quinoa plants. While the wild type inoculum induced indistinct chlorotic lesions, the mutant produced obvious lesions on C. quinoa although RNA accumulation of the mutant in Nicotiana benthamiana protoplasts was significantly reduced compared to wild type. Mutations were identified in the helicase-like domain. One RT-PCR-generated cDNA clone (designated pL1-33) representing the helicase-like region showed four nucleotide mutations encoding three amino acid changes that were shown to result in dramatically decreased viral accumulation. Independent analyses of the effects of these substitutions showed that nucleotide changes at position 1722 resulting in a leucine to proline switch and position 2129 resulting in a histidine to tyrosine switch had the greatest effect on viral accumulation. Combination of these two mutations resulted in a undetectable viral accumulation. We have identified that amino acids within the helicase domain but outside the universally conserved helicase-like motifs that play an important role in viral amplification. © 1998 Elsevier Science B.V. All rights reserved. Keywords: Bamboo mosaic potexvirus; Helicase; Mutations; Viral accumulation 1. Introduction Bamboo mosaic potexvirus (BaMV) was dis- covered infecting plantations of common bamboo species in Taiwan in 1979 (Lin et al., 1979). In two major cultivated species, Ma Chu (Dendro - calamus latiflorus Munro) and green bamboo (Bambusa oldhamii Munro), more than 70–80% of the plants are commonly infected (Lin et al., 1993). BaMV has flexuous rod-shaped particles about 480–500 nm in length (Lin et al., 1977, * Corresponding author. Tel.: +886 4 2874754; fax: +886 4 2861905. 0168-1702/98/$ - see front matter © 1998 Elsevier Science B.V. All rights reserved. PII S0168-1702(98)00109-9
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Evolution of bamboo mosaic virus in a nonsystemic host results in mutations in the helicase-like...

Virus Research 58 (1998) 127–136
Evolution of bamboo mosaic virus in a nonsystemic host resultsin mutations in the helicase-like domain that cause reduced
RNA accumulation
Cheng-Yen Huang a, Ching-Hsiu Tsai a,b,*a Graduate Institute of Agricultural Biotechnology, National Chung Hsing Uni6ersity, Taichung 402, Taiwan
b Agricultural Biotechnology Laboratories, National Chung Hsing Uni6ersity, Taichung 402, Taiwan
Received 1 May 1998; received in revised form 10 September 1998; accepted 14 September 1998
Abstract
A mutant population of bamboo mosaic potexvirus (BaMV) was isolated after serial passage using Chenopodiumquinoa plants. While the wild type inoculum induced indistinct chlorotic lesions, the mutant produced obvious lesionson C. quinoa although RNA accumulation of the mutant in Nicotiana benthamiana protoplasts was significantlyreduced compared to wild type. Mutations were identified in the helicase-like domain. One RT-PCR-generated cDNAclone (designated pL1-33) representing the helicase-like region showed four nucleotide mutations encoding threeamino acid changes that were shown to result in dramatically decreased viral accumulation. Independent analyses ofthe effects of these substitutions showed that nucleotide changes at position 1722 resulting in a leucine to prolineswitch and position 2129 resulting in a histidine to tyrosine switch had the greatest effect on viral accumulation.Combination of these two mutations resulted in a undetectable viral accumulation. We have identified that aminoacids within the helicase domain but outside the universally conserved helicase-like motifs that play an important rolein viral amplification. © 1998 Elsevier Science B.V. All rights reserved.
Keywords: Bamboo mosaic potexvirus; Helicase; Mutations; Viral accumulation
1. Introduction
Bamboo mosaic potexvirus (BaMV) was dis-covered infecting plantations of common bamboo
species in Taiwan in 1979 (Lin et al., 1979). Intwo major cultivated species, Ma Chu (Dendro-calamus latiflorus Munro) and green bamboo(Bambusa oldhamii Munro), more than 70–80%of the plants are commonly infected (Lin et al.,1993). BaMV has flexuous rod-shaped particlesabout 480–500 nm in length (Lin et al., 1977,
* Corresponding author. Tel.: +886 4 2874754; fax: +8864 2861905.
0168-1702/98/$ - see front matter © 1998 Elsevier Science B.V. All rights reserved.
PII S0168-1702(98)00109-9

C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136128
1979) and contains a single-stranded positivesense RNA genome 6366 nucleotides long, exclud-ing the 3% poly(A) tail, with five open readingframes (ORF1–5). ORF1 encodes a 155-kDaprotein, comprising a methyltransferase-like do-main, an RNA helicase-like domain, and a RNA-dependent RNA polymerase (RdRp)-like domain,that is involved in viral RNA replication (Lin etal., 1994).
The most conserved motif among the viral heli-case-like proteins is the NTP-binding motif(Kadare and Haenni, 1997). This element is di-rectly involved in binding the b and g phosphatesof the NTP, as well as in chelating the Mg2+ ofthe Mg-NTP complex and in hydrolysis of theNTP g-phosphate (Gorbalenya et al., 1988; Tol-skaya et al., 1994; Kadare and Haenni, 1997).Mutations have been introduced into this regionto identify the function of helicase-like proteins ofseveral RNA viruses. Poliovirus with changes inconserved amino acids within the NTP-bindingmotif or in nonconserved regions of poliovirus 2Chelicase-like protein was no longer capable ofreplication in host cells (Mirzayan and Wimmer,1994; Teterina et al., 1992). A series of mutationsartificially introduced into the NTP-binding motif[glycine–lysine–serine (GKS)] of potato virus X(PVX) to yield GNS or GES rendered the virusunable to accumulate in tobacco plants or proto-plasts, while mutants with the mutation GKS toAKS had an accumulation level of 30–50% com-pared to the wild-type PVX RNA (Davenportand Baulcombe, 1997). Mutations introduced intothe helicase-like domain of BMV by linker-inser-tion resulted in lethal or temperature-sensitive (ts)mutants (Kroner et al., 1990).
In an error-prone viral replication system, vari-ation can be introduced by serial passaging thehomogeneous viral transcripts onto host plants(Kurath and Palukaitis, 1990; Kurath and Dodds,1995). Fitness gains in one environment may leadto fitness losses in another environment (Domingoand Holland, 1997). Therefore, transcripts withless infectivity in certain hosts or harboring muta-tions causing a poor replication rate in one hostcan have the potential to produce fitter variants inanother host, these variants will replace the origi-nal inoculum. This strategy has been successfully
explored to identify mutations that outgrow theinoculum of mutants of turnip yellow mosaicvirus (Tsai and Dreher, 1992) and brome mosaicvirus (Smirnyagina et al., 1994). In thismanuscript, we focused on isolating a necroticlesion formating mutant by inoculating the clonedBaMV RNAs onto the nonsystemic host C.quinoa, in which BaMV-O produces light chloro-sis symptoms. A variant (L1) that gives rise tonecrotic local lesions in C. quinoa was isolated,and shown to possess decreased replication activ-ity in N. benthamiana protoplasts.
2. Materials and methods
2.1. Materials
Chenopodium quinoa and N. benthamiana weregrown in a greenhouse under natural light or in agrowth chamber under 16-h day length at 27°C.T7 RNA polymerase and reverse transcriptase(Superscript II) were purchased from GIBCO-BRL, m7GpppG cap analog from New EnglandBiolabs, Inhibit-Ace RNase inhibitor from 5Prime-3 Prime Inc., T7 DNA polymerase (Seque-nase) from United States Biochemical, Thermusaquaticus DNA polymerase from Promega, andrestriction enzymes were purchased from GIBCO-BRL, New England Biolabs, and Stratagene.Macerase and cellulysin were purchased from Cal-biochem. Synthetic deoxyoligonucleotides weremade by automated phosphoramidite synthesis.
2.2. Virion RNA extraction and characterization
BaMV was isolated from infected plants andprecipitated by polyethylene glycol according toLin and Chen (1991) except that centrifugationwith CaCl2 was omitted. Virion RNA was pre-pared from BaMV by double phenol–chloroformextraction and ethanol precipitation (Lin et al.,1992), then quantitated spectrophotometricallyand analyzed by gel electrophoresis. RNase pro-tection experiments were performed in whichvirion RNA was mixed with an antisense RNAprobe and denatured at 85°C for 5 min in a 30-m lreaction mixture containing 40 mM piperazine-

C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136 129
N,N %-bis (2-ethanesulfonic acid) (PIPES; pH 6.7),0.4 M NaCl, 1 mM EDTA, and 80% deionizedformamide. After hybridization overnight at55°C, 300 m l of RNase protection buffer contain-ing 30 mM Tris–HCl (pH 7.5), 200 mM NaCl,100 mM LiCl, 5 mM EDTA, RNase A (32 mg/ml), and RNase T1 (1.6 mg/ml) was added to thesample, then incubated at 37°C for 40 min.RNase was removed by treatment with 50 mg/mlproteinase K in the presence of 0.5% SDS at 37°Cfor 30 min (Tsai and Dreher, 1992). RNA wasprobed with minus-strand RNA representingBaMV sequences between the following restric-tion sites: PA0.6, P6uII1002–ApaI1576; AS2.1,ApaI1576–SphI3735; BS1.1, BsrGI2593–SphI3735;SB2.6, SphI3735–BamHI3%.
In order to obtain the sequence of the regionscontaining the mutation(s) identified by RNaseprotection with probe AS2.1, two primers, (− )2347 d(AGAGGGGCTGAGATCGTT) primingat nucleotide position 2347 and (+ )1449 d(GAC-CAAAACCCTACACCAACC) priming at posi-tion 1449 were used for reverse transcription andpolymerase chain reaction (RT-PCR). The am-plified fragments were then cloned into T-vector(Invitrogen) and sequenced.
2.3. Cloning and transcription
Fragments between the unique ApaI1576 andBstBI2305 sites within the infectious clone pBaMV-O (Lin et al., 1994; Tsai et al., unpublished data)were replaced with mutant fragments. In prepar-ing four single-nucleotide substitution mutants,six primers were designed: (+ )1449, (− )2347both described above, (+ )2066T d(CAAG-GAGGACTCCATCGTCACCAT), (+ )2129Td(AAACTGCCCGCTTATGACCCTGA), (+)2215G d(TACACTAAGCTGCCACGCGGGT),and (− )1722C d(CCAACTGTTCAGGGTCGC-CTT) with mutating nucleotides underlined. Increating mutants 2066, 2129 and 2215, pBaMV-Owas used as template for two-step PCR. Primers(+ )2066T, (+ )2129T and (+ )2215G were firstused individually with the primer (− )2347 toamplify short fragments, which were then used asmegaprimers with the upstream primer (+ )1449to generate the mutant fragments for subcloning
(Sarkar and Sommer, 1990). For generating dou-ble or triple mutants, the same primers and strat-egy were used except that the pBaMV-O orsingle-nucleotide substitution mutants were usedas templates as appropriate.
Plasmid DNAs were prepared from 50 ml bac-terial cultures and confirmed by double-strandedDNA sequencing (Chen and Seeburg, 1985).Capped genomic transcripts were prepared withT7 RNA polymerase from DNA templates lin-earized with BamHI3% in the condition adoptedfrom Tsai and Dreher (1991). The transcriptswere analyzed on agarose gel electrophoresis andthen quantified by Bioimage (IntelligentQuantifier™).
2.4. Plant and protoplast inoculations
Leaves were removed from 7-week-old C.quinoa plants except five to seven leaves used forviral inoculations. Five-week-old N. benthamianaplants with two leaves were used for inoculation.Each leaf was mechanically inoculated with 10 m lof RNA transcript (0.5 mg/ml) or virion RNA (2mg/ml) in 50 mM glycine–30 mM K2HPO4 (pH9.2)–1% bentonite–1% celite, and plants werethen held in a growth chamber until furtheranalysis.
For protoplast isolation, about 4 g of freshhealthy leaves were taken from 7-week-old growthchamber grown N. benthamiana plants. Proto-plasts were prepared and inoculated (5 mg oftranscript RNA per 4×105 cells) as describedpreviously (Tsai and Dreher, 1991).
2.5. Analysis of 6iral products by Westernimmunoblotting and Northern blotting
The levels of coat protein in inoculated plantsor protoplasts were analyzed by Western blots asdescribed in Weiland and Dreher (1989) with theanti-BaMV antiserum. Detection of immuno-reac-tive protein bands was performed with 5-bromo-4-chloro-3-indolyl phosphate (Bio-Rad) orchemiluminescent development (ECL, AmershamInternational). Protein levels were quantified byBioimage (Intelligent Quantifier™). RNA was ex-tracted and analyzed as described previously
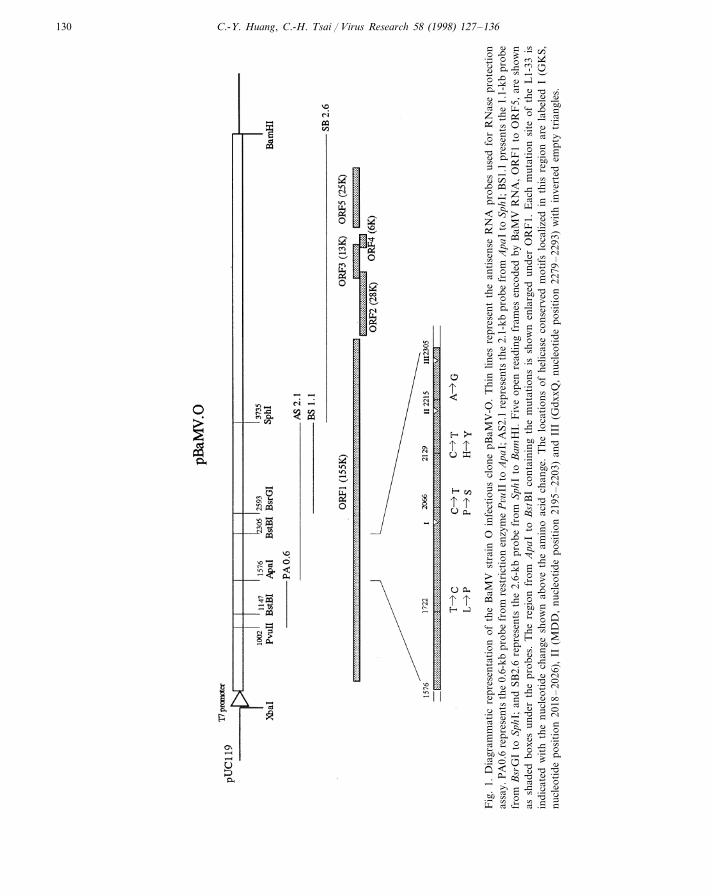
C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136130
Fig
.1.
Dia
gram
mat
icre
pres
enta
tion
ofth
eB
aMV
stra
inO
infe
ctio
uscl
one
pBaM
V-O
.T
hin
lines
repr
esen
tth
ean
tise
nse
RN
Apr
obes
used
for
RN
ase
prot
ecti
onas
say.
PA
0.6
repr
esen
tsth
e0.
6-kb
prob
efr
omre
stri
ctio
nen
zym
eP6u
IIto
Apa
I;A
S2.1
repr
esen
tsth
e2.
1-kb
prob
efr
omA
paI
toS
phI;
BS1
.1pr
esen
tsth
e1.
1-kb
prob
efr
omB
srG
Ito
Sph
I;an
dSB
2.6
repr
esen
tsth
e2.
6-kb
prob
efr
omS
phI
toB
amH
I.F
ive
open
read
ing
fram
esen
code
dby
BaM
VR
NA
,O
RF
1to
OR
F5,
are
show
nas
shad
edbo
xes
unde
rth
epr
obes
.T
here
gion
from
Apa
Ito
Bst
BI
cont
aini
ngth
em
utat
ions
issh
own
enla
rged
unde
rO
RF
1.E
ach
mut
atio
nsi
teof
the
L1-
33is
indi
cate
dw
ith
the
nucl
eoti
dech
ange
show
nab
ove
the
amin
oac
idch
ange
.T
helo
cati
ons
ofhe
licas
eco
nser
ved
mot
ifs
loca
lized
inth
isre
gion
are
labe
led
I(G
KS,
nucl
eoti
depo
siti
on20
18–
2026
),II
(MD
D,
nucl
eoti
depo
siti
on21
95–
2203
)an
dII
I(G
dxxQ
,nu
cleo
tide
posi
tion
2279
–22
93)
wit
hin
vert
edem
pty
tria
ngle
s.
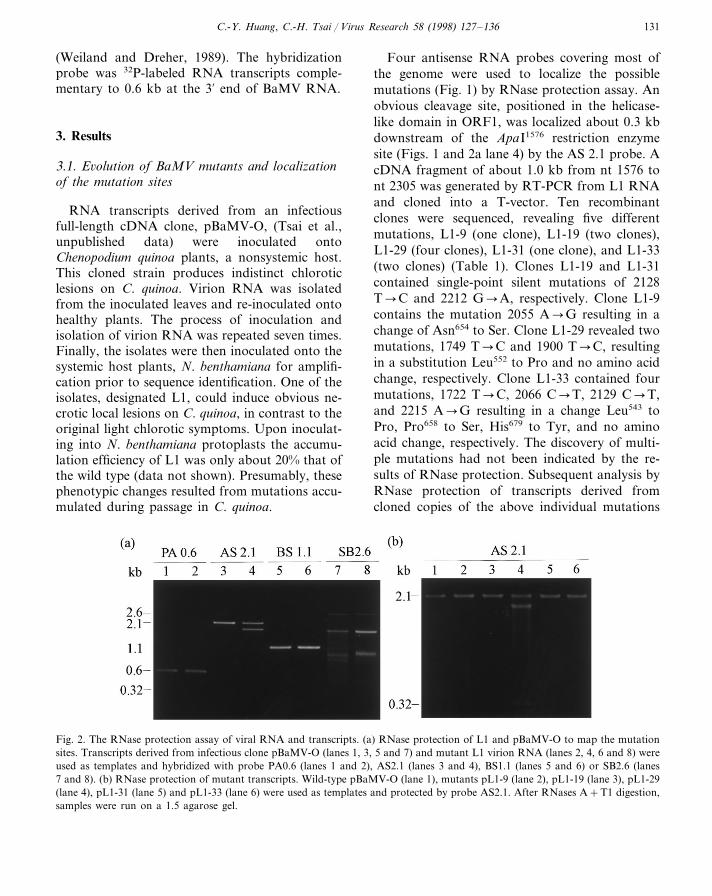
C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136 131
(Weiland and Dreher, 1989). The hybridizationprobe was 32P-labeled RNA transcripts comple-mentary to 0.6 kb at the 3% end of BaMV RNA.
3. Results
3.1. E6olution of BaMV mutants and localizationof the mutation sites
RNA transcripts derived from an infectiousfull-length cDNA clone, pBaMV-O, (Tsai et al.,unpublished data) were inoculated ontoChenopodium quinoa plants, a nonsystemic host.This cloned strain produces indistinct chloroticlesions on C. quinoa. Virion RNA was isolatedfrom the inoculated leaves and re-inoculated ontohealthy plants. The process of inoculation andisolation of virion RNA was repeated seven times.Finally, the isolates were then inoculated onto thesystemic host plants, N. benthamiana for amplifi-cation prior to sequence identification. One of theisolates, designated L1, could induce obvious ne-crotic local lesions on C. quinoa, in contrast to theoriginal light chlorotic symptoms. Upon inoculat-ing into N. benthamiana protoplasts the accumu-lation efficiency of L1 was only about 20% that ofthe wild type (data not shown). Presumably, thesephenotypic changes resulted from mutations accu-mulated during passage in C. quinoa.
Four antisense RNA probes covering most ofthe genome were used to localize the possiblemutations (Fig. 1) by RNase protection assay. Anobvious cleavage site, positioned in the helicase-like domain in ORF1, was localized about 0.3 kbdownstream of the ApaI1576 restriction enzymesite (Figs. 1 and 2a lane 4) by the AS 2.1 probe. AcDNA fragment of about 1.0 kb from nt 1576 tont 2305 was generated by RT-PCR from L1 RNAand cloned into a T-vector. Ten recombinantclones were sequenced, revealing five differentmutations, L1-9 (one clone), L1-19 (two clones),L1-29 (four clones), L1-31 (one clone), and L1-33(two clones) (Table 1). Clones L1-19 and L1-31contained single-point silent mutations of 2128T�C and 2212 G�A, respectively. Clone L1-9contains the mutation 2055 A�G resulting in achange of Asn654 to Ser. Clone L1-29 revealed twomutations, 1749 T�C and 1900 T�C, resultingin a substitution Leu552 to Pro and no amino acidchange, respectively. Clone L1-33 contained fourmutations, 1722 T�C, 2066 C�T, 2129 C�T,and 2215 A�G resulting in a change Leu543 toPro, Pro658 to Ser, His679 to Tyr, and no aminoacid change, respectively. The discovery of multi-ple mutations had not been indicated by the re-sults of RNase protection. Subsequent analysis byRNase protection of transcripts derived fromcloned copies of the above individual mutations
Fig. 2. The RNase protection assay of viral RNA and transcripts. (a) RNase protection of L1 and pBaMV-O to map the mutationsites. Transcripts derived from infectious clone pBaMV-O (lanes 1, 3, 5 and 7) and mutant L1 virion RNA (lanes 2, 4, 6 and 8) wereused as templates and hybridized with probe PA0.6 (lanes 1 and 2), AS2.1 (lanes 3 and 4), BS1.1 (lanes 5 and 6) or SB2.6 (lanes7 and 8). (b) RNase protection of mutant transcripts. Wild-type pBaMV-O (lane 1), mutants pL1-9 (lane 2), pL1-19 (lane 3), pL1-29(lane 4), pL1-31 (lane 5) and pL1-33 (lane 6) were used as templates and protected by probe AS2.1. After RNases A+T1 digestion,samples were run on a 1.5 agarose gel.

C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136132
Table 1The nucleotide changes from wild-type pBaMV-O to mutants
Amino acid change Viral accumulationa (%)Clones Numbers of clone Mutation site Lesions
100pBaMV-O No111 NoAsn654�Ser2055 A�GpL1-9 1
No 89pL1-19 2 No2128 T�CNo115Leu552�Pro1749 T�CpL1-29 4
1900b T�C No94 NopL1-31 1 2212 G�A No
Leu543�Pro B0.1cpL1-33 2 1722 T�C NoPro658�Ser2066 C�T
2129 C�T His679�TyrNo2215 A�G
a Coat protein accumulation in protoplasts, detected by Western blotting with anti-BaMV coat protein serum. All data representaverages of at least three or more experiments.b The only mutation site mapped by RNase protection.c Undetectable signal, with accumulation below the 0.1% detection limit.
indicated that only the transcript derived frompL1-29 showed the same cleavage as that of L1virion RNA in RNase protection assay (Fig. 2blane 4). Of the two mutations present in pL1-29,1900 (T�C) and 1749 (T�C), the mutation atnucleotide position 1900 located 324 bp down-stream of the ApaI1576 is the most likely site of thecleavage assayed by RNase protection.
3.2. Mutations in the helicase-like domain ofBaMV ORF1 that can interfere with efficientaccumulation
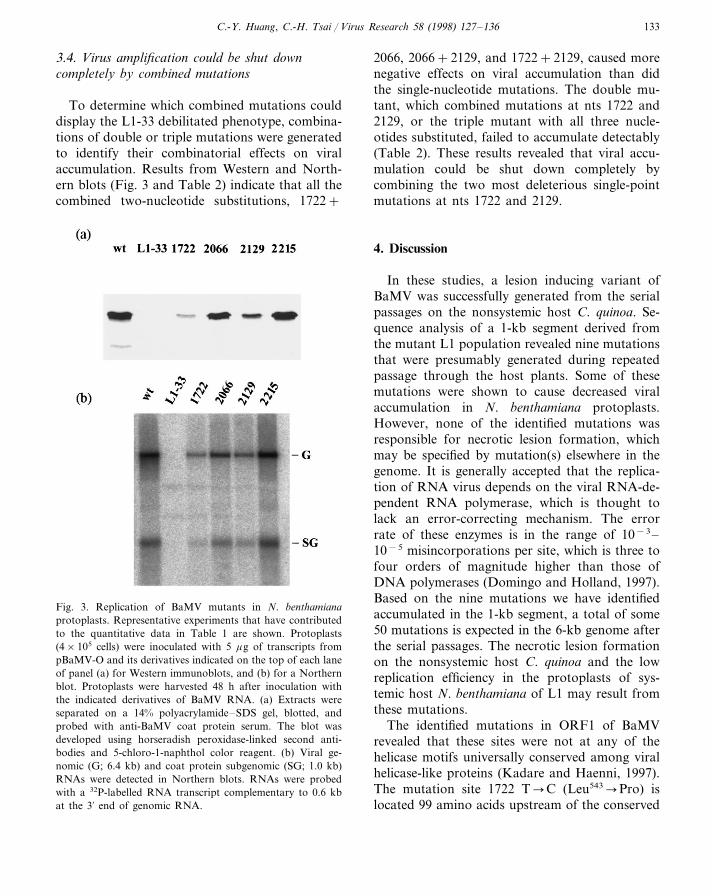
Fragments bearing mutations were subclonedinto pBaMV-O. Full length capped transcriptsderived from pL1-9, -19, -29, -31, and -33 weregenerated in vitro and inoculated into protoplastsof N. benthamiana. The inoculated protoplastswere held and incubated at 25°C with constantlight for 2 days. The accumulation efficiency ofeach mutant was measured by comparing therelative amount of coat protein accumulation onimmunoblots with at least three independent inoc-ulations. All the mutants, except L1-33, couldreplicate efficiently, accumulating to levels close towild type (Table 1). Mutant L1-33 containing fourmutations with three amino acid changes showedno detectable signal of coat protein accumulationon the immunoblots.
3.3. Indi6idual amino acid changes encoded by thefour mutations of L1-33 differentially affectaccumulation efficiency
Since L1-33 contained four mutations aroundthe helicase-like domain and showed no de-tectable signal of replication, we were interestedto identify the contribution of the individual mu-tations to the reduction in viral RNA accumula-tion. We used two-step PCR (see Section 2) tocreate four single-nucleotide substitution mutantsand introduced these into pBaMV-O. Full-lengthtranscripts derived from pBaMV-O and mutantspL1-33/1722, pL1-33/2066, pL1-33/2129, andpL1-33/2215 were inoculated into protoplasts.Western blotting was used to detect and quanti-tate the accumulation of coat protein and North-ern blotting was used to detect the multiplicationof genomic and subgenomic RNAs (Fig. 3). Mu-tant L1-33/2215, encoding no amino acid change,accumulated to 116% of the wild type level,whereas the other three mutants with alteredamino acid residues displayed different degrees ofdefective accumulation. The strongest effect re-sulted from the mutation at nucleotide 1722 withonly 10% viral accumulation relative to wild type(Table 2). However, none of the individual single-nucleotide substitutions could cause the lethal ef-fect shown for L1-33 transcripts.
C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136 133
3.4. Virus amplification could be shut downcompletely by combined mutations
To determine which combined mutations coulddisplay the L1-33 debilitated phenotype, combina-tions of double or triple mutations were generatedto identify their combinatorial effects on viralaccumulation. Results from Western and North-ern blots (Fig. 3 and Table 2) indicate that all thecombined two-nucleotide substitutions, 1722+
2066, 2066+2129, and 1722+2129, caused morenegative effects on viral accumulation than didthe single-nucleotide mutations. The double mu-tant, which combined mutations at nts 1722 and2129, or the triple mutant with all three nucle-otides substituted, failed to accumulate detectably(Table 2). These results revealed that viral accu-mulation could be shut down completely bycombining the two most deleterious single-pointmutations at nts 1722 and 2129.
4. Discussion
In these studies, a lesion inducing variant ofBaMV was successfully generated from the serialpassages on the nonsystemic host C. quinoa. Se-quence analysis of a 1-kb segment derived fromthe mutant L1 population revealed nine mutationsthat were presumably generated during repeatedpassage through the host plants. Some of thesemutations were shown to cause decreased viralaccumulation in N. benthamiana protoplasts.However, none of the identified mutations wasresponsible for necrotic lesion formation, whichmay be specified by mutation(s) elsewhere in thegenome. It is generally accepted that the replica-tion of RNA virus depends on the viral RNA-de-pendent RNA polymerase, which is thought tolack an error-correcting mechanism. The errorrate of these enzymes is in the range of 10−3–10−5 misincorporations per site, which is three tofour orders of magnitude higher than those ofDNA polymerases (Domingo and Holland, 1997).Based on the nine mutations we have identifiedaccumulated in the 1-kb segment, a total of some50 mutations is expected in the 6-kb genome afterthe serial passages. The necrotic lesion formationon the nonsystemic host C. quinoa and the lowreplication efficiency in the protoplasts of sys-temic host N. benthamiana of L1 may result fromthese mutations.
The identified mutations in ORF1 of BaMVrevealed that these sites were not at any of thehelicase motifs universally conserved among viralhelicase-like proteins (Kadare and Haenni, 1997).The mutation site 1722 T�C (Leu543�Pro) islocated 99 amino acids upstream of the conserved
Fig. 3. Replication of BaMV mutants in N. benthamianaprotoplasts. Representative experiments that have contributedto the quantitative data in Table 1 are shown. Protoplasts(4×105 cells) were inoculated with 5 mg of transcripts frompBaMV-O and its derivatives indicated on the top of each laneof panel (a) for Western immunoblots, and (b) for a Northernblot. Protoplasts were harvested 48 h after inoculation withthe indicated derivatives of BaMV RNA. (a) Extracts wereseparated on a 14% polyacrylamide–SDS gel, blotted, andprobed with anti-BaMV coat protein serum. The blot wasdeveloped using horseradish peroxidase-linked second anti-bodies and 5-chloro-1-naphthol color reagent. (b) Viral ge-nomic (G; 6.4 kb) and coat protein subgenomic (SG; 1.0 kb)RNAs were detected in Northern blots. RNAs were probedwith a 32P-labelled RNA transcript complementary to 0.6 kbat the 3% end of genomic RNA.
C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136134
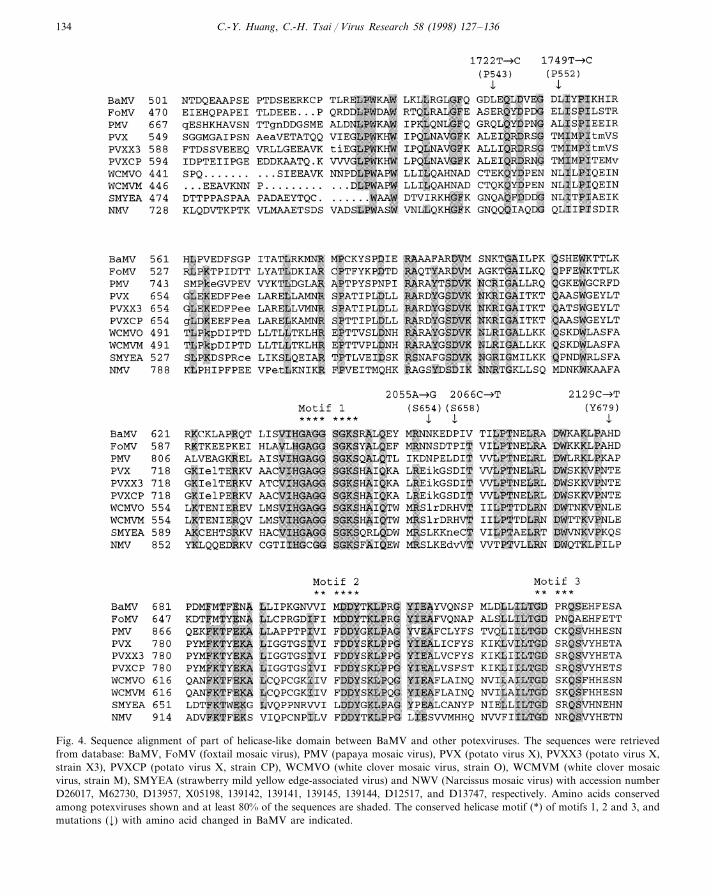
Fig. 4. Sequence alignment of part of helicase-like domain between BaMV and other potexviruses. The sequences were retrievedfrom database: BaMV, FoMV (foxtail mosaic virus), PMV (papaya mosaic virus), PVX (potato virus X), PVXX3 (potato virus X,strain X3), PVXCP (potato virus X, strain CP), WCMVO (white clover mosaic virus, strain O), WCMVM (white clover mosaicvirus, strain M), SMYEA (strawberry mild yellow edge-associated virus) and NWV (Narcissus mosaic virus) with accession numberD26017, M62730, D13957, X05198, 139142, 139141, 139145, 139144, D12517, and D13747, respectively. Amino acids conservedamong potexviruses shown and at least 80% of the sequences are shaded. The conserved helicase motif (*) of motifs 1, 2 and 3, andmutations (¡) with amino acid changed in BaMV are indicated.

C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136 135
Table 2The accumulation rates of mutants with single or multiple mutations
Single-point mutations Combined mutations
Accumulation levela (%)ClonesClones Accumulation level (%)
pL1-33/1722+2066 0.4pL1-33/1722 1052 pL1-33/2066+2129pL1-33/2066 3.6
pL1-33/1722+2129 B0.1b25pL1-33/2129116 pL1-33/1722+2066+2129 B0.1pL1-33/2215
a Coat protein accumulation in protoplasts, detected by Western blotting with anti-BaMV coat protein serum. All data representaverages of at least three or more experiments.b Undetectable signal, with accumulation below the 0.1% detection limit.
nucleotide binding fold GxxGxGKS/T (GKS)motif (Fig. 1), whereas the mutations 2066 C�T(Pro658�Ser) and 2129 C�T (His679�Tyr) arelocated between the conserved motifs I (GKS)and II (DDY) (Figs. 1 and 4). Each substitutionreduced the viral accumulation level. Amino acidalignment (PredictProtein server at http://www.embl-heidelberg.de/predictprotein/ppDo-PredDef.html) of proteins encoded bypotexviruses indicated that the sites of these muta-tions were surrounded by highly conserved se-quences (Fig. 4). The conserved sequencesextended upstream of the GKS motif of BaMVORF1 by 117 amino acids to include the site ofthe 1722 mutation. This suggested that nucleotide1722 was most likely to be functionally associatedwith the helicase domain. It is not known whetherthese changes diminished protein stability, inter-fered with protein folding or protein–protein in-teraction, or directly affected protein function.However, the fact that these mutations lie inamino acid sequence strongly conserved amongthe potexviruses suggest that the influence on viralamplification is due to the encoded amino acidchanges rather than changes at the RNA level.
In summary, a mutant population was obtainedby serial passage from infected to healthy plants.This mutant, though capable of lesion formationin C. quinoa, showed a greatly reduced accumula-tion in N. benthamiana. Results of sequencinganalysis showed that the reduced accumulationmight be due to the mutations in the noncon-served region of the helicase-like domain. Intro-duction of the defined mutations into the
helicase-like coding domain of wild-type viralRNA showed that these mutations do have dra-matic effects on viral accumulation. Interestingly,none of the mutants we have constructed showsany necrotic lesion formation. Therefore, genomicsequences responsible for lesion formation mustexist on other regions of the L1 genome.
Acknowledgements
We thank Drs Theo Dreher and Tony H.H.Chen at Oregon State University for helpful ad-vice and English editing. We acknowledge theassistance of Dr Menghsiou Meng in sequencealignment. This work was supported in part byNational Science Council grant NSC 88-2311-B-005-040.
References
Chen, E.Y., Seeburg, P., 1985. Supercoil sequencing: a fastand simple method for sequencing plasmid DNA. DNA 4,165–170.
Davenport, G.F., Baulcombe, D.C., 1997. Mutation of theGKS motif of the RNA-dependent RNA polymerase frompotato virus X disables or eliminates virus replication. J.Gen. Virol. 78, 1247–1251.
Domingo, E., Holland, J.-J., 1997. RNA virus mutations andfitness for survival. Annu. Rev. Microbiol. 51, 151–178.
Gorbalenya, A.E., Koonin, E.V., Donchenko, A.P., Blinov,V.M., 1988. A novel superfamily of nucleoside triphos-phate-binding motif containing proteins which are proba-bly involved in duplex unwinding in DNA and RNAreplication and recombination. FEBS Lett. 235, 16–24.

C.-Y. Huang, C.-H. Tsai / Virus Research 58 (1998) 127–136136
Kadare, G., Haenni, A.-L., 1997. Virus-encoded RNA heli-cases. J. Virol. 71, 2583–2590.
Kroner, P.A., Young, B.M., Ahlquist, P., 1990. Analysis ofthe role of brome mosaic virus 1a protein domains in RNAreplication, using linker insertion mutagenesis. J. Virol. 64,6110–6120.
Kurath, G., Palukaitis, P., 1990. Serial passage of infectioustranscripts of a cucumber mosaic virus satellite RNA cloneresults in sequence heterogeneity. Virology 176, 8–15.
Kurath, G., Dodds, J.A., 1995. Mutations analyses of molecu-larly cloned satellite tobacco mosaic virus during serialpassage in plants: evidence for hotspots of genetic change.RNA 1, 491–500.
Lin, N.S., Chen, C.C., 1991. Association of bamboo mosaicvirus (BoMV) and BoMV specific electron-dense crystallinebodies with chloroplast. Phytopathology 81, 1551–1555.
Lin, M.T., Kitajima, E.W., Cupertino, F.P., Costa, C.L., 1977.Partial purification and some properties of bamboo mosaicvirus. Phytopathology 67, 1439–1443.
Lin, N.S., Chen, M.J., Kiang, T., Lin, W.C., 1979. Preliminarystudies on bamboo mosaic virus disease in Taiwan. TaiwanForest Res. Inst. 317, 1–10.
Lin, N.S., Lin, F.Z., Huang, T.Y., Hsu, Y.H., 1992. Genomeproperties of bamboo mosaic virus. Phytopathology 82,731–734.
Lin, N.S., Chai, Y.J., Huang, T.Y., Hsu, Y.H., 1993. Inci-dence of bamboo mosaic potexvirus in Taiwan. Plant Dis.77, 448–450.
Lin, N.S., Lin, B.Y., Lo, N.W., Hu, C.C., Chow, T.Y., Hsu,Y.H., 1994. Nucleotide sequence of the genomic RNA ofbamboo mosaic potexvirus. J. Gen. Virol. 75, 2513–2518.
Mirzayan, C., Wimmer, E., 1994. Biochemical studies onpoliovirus polypeptide 2C: evidence for ATPase activity.Virology 199, 176–187.
Sarkar, G., Sommer, S.S., 1990. The ‘megaprimer’ method ofsite-directed mutagenesis. Biotechnology 8, 404–407.
Smirnyagina, E., Hsu, Y.-H., Chua, N., Ahlquist, P., 1994.Second-site mutations in the brome mosaic virus RNA3intercistronic region partially suppress a defect in coatprotein mRNA transcription. Virology 198, 427–436.
Teterina, N.L., Kean, K.M., Gorbalenya, A.E., Agol, V.I.,Girard, M., 1992. Analysis of the functional significance ofamino acid residues in the putative NTP-binding pattern ofthe poliovirus 2C protein. J. Gen. Virol. 73, 1977–1986.
Tolskaya, E.A., Romanova, L.I., Kolesnikova, M.S., Gmyl,A.P., Gorbalenya, A.E., Agol, V.I., 1994. Genetic studieson the poliovirus 2C protein, an NTPase a plausible mech-anism of guanidine effect on the 2C function and evidencefor the importance of 2C oligomerization. J. Mol. Biol.236, 1310–1323.
Tsai, C.-H., Dreher, T.W., 1991. Turnip yellow mosaic virusRNAs with anticodon loop substitutions that result indecreased valylation fail to replicate efficiently. J. Virol. 65,3060–3067.
Tsai, C.-H., Dreher, T.W., 1992. Second-site suppressor muta-tions assist in studying the function of the 3% noncodingregion of turnip yellow mosaic virus RNA. J. Virol. 66,5190–5199.
Weiland, J.J., Dreher, T.W., 1989. Infectious TYMV RNAfrom cloned cDNA: effects in vitro and in vivo of pointsubstitutions in the initiation codons of two extensivelyoverlapping ORFs. Nucleic Acids Res. 17, 4675–4687.
.
.




















