
Nuclear Receptor Cofactors in PPARγ-Mediated Adipogenesis and Adipocyte Energy Metabolism
Upload
independentCategory
view
4download
0
TE
ÁIJ
*M
BittfepoagttuEdotraseCsawvwavimhct
TbomsS
GASTROENTEROLOGY 2007;132:1791–1803
he Role of PPAR� on Restoration of Colonic Homeostasis Afterxperimental Stress-Induced Inflammation and Dysfunction
NGEL PONFERRADA,* JAVIER R. CASO,‡ LUIS ALOU,§ ARTURO COLÓN,‡ DAVID SEVILLANO,§ MARÍA A. MORO,‡
GNACIO LIZASOAIN,‡ PEDRO MENCHÉN,* MARÍA L. GÓMEZ–LUS,§ PEDRO LORENZO,‡ ENRIQUE COS,*UAN C. LEZA,‡ and LUIS MENCHÉN*
Servicio de Aparato Digestivo, Hospital General Universitario “Gregorio Marañón,” Madrid; and Departamentos de ‡Farmacología and §Microbiología I, Facultad de
edicina, Universidad Complutense, Madrid, Spainsotttpwhtpattoaagtatnpc
itadsstopms
1ot
BA
SIC–
ALI
MEN
TARY
TRA
CT
ackground & Aims: Psychological stress has beenmplicated in the clinical course of several gastroin-estinal diseases, but the mechanisms implicated andhe effects of stress on the normal colon are not yetully understood. Methods: Male Wistar rats werexposed to various immobilization periods as a stressaradigm. Colon was processed to assess myeloper-xidase activity, nitric oxide synthase 2, cyclooxygen-se 2, and peroxisome proliferator-activated receptoramma (PPAR�) expression and production of pros-aglandins. Colonic permeability, bacterial transloca-ion, tight junctions ultrastructure, and immunoglob-lin (Ig) A levels were also evaluated. Results:xposure to acute (6 hours) immobilization stress pro-uced an increase in myeloperoxidase activity and nitricxide synthase 2 and cyclooxygenase 2 expression. Allhese parameters remained increased after 5 days ofepeated stress exposure, showing a trend to normalizefter 10 days. Levels of the anti-inflammatory eico-anoid 15-deoxy-�12,14-prostaglandin J2 (15d-PGJ2) andxpression of PPAR� run parallel with these changes.olonic epithelial barrier was altered after stress expo-
ure, and a significant decrease in colonic IgA levelsfter acute stress exposure was observed. Pretreatmentith PPAR� agonists 15d-PGJ2 and rosiglitazone pre-
ented colonic inflammation and barrier dysfunction asell as the decrease of IgA production induced aftercute stress; PPAR� specific antagonist T0070907 re-erted these effects. Conclusions: Activation of PPAR�n rat colon in vivo seems to counteract colonic inflam-
ation and dysfunction induced by stress. On the otherand, PPAR� ligands may be therapeutically useful inonditions in which inflammation and barrier dysfunc-ion takes place in colon after exposure to stress.
he relationship between psychosocial stress and sev-eral gastrointestinal disorders is widely accepted by
oth patients and physicians, although the rational dem-nstration of a casual nexus remains difficult because ofethodological aspects and interpretation of clinical
tudies and the shortage of experimental approaches.
tress influences subjective perception of gastrointestinalymptoms by patients1 and has been shown to trigger thenset and modify the clinical course of certain gastroin-estinal diseases.2 Irritable bowel syndrome and func-ional dyspepsia are the most prevalent among the gas-rointestinal disorders that have been related withsychological factors.3 It has been reported that patientsith irritable bowel syndrome and functional dyspepsiaad been exposed to stressful events more frequentlyhan control individuals and that life stress was highlyredictive of symptom intensity.4 Stress has been associ-ted also with inflammatory activity of ulcerative coli-is5,6 and with an increased risk of disease exacerba-ion.5,7,8 The high response to placebo frequentlybserved in inflammatory bowel disease clinical trials9,10
lso supports the concept that psychological factors playrole in the clinical course of these diseases. There is
rowing evidence from experimental studies supportinghe ability of psychosocial stress to (1) induce physiologicbnormalities in the gut such as increased motility, al-ered ion secretion, increased permeability, and entericeuron dysfunction,11–14 (2) increase the severity of ex-erimental colitis,15 and (3) even reactivate a quiescentolitis.16
Previous findings of our group showed that stressnduces macroscopic and histologic colonic inflamma-ion, increasing inducible nitric oxide synthase (NOS-2)ctivity and membrane lipid peroxidation, after a lowose of trinitrobenzene sulfonic acid unable to induceuch changes in nonstressed animals.17 Also, it has beenhown that stress increases colonic permeability, facili-ating the entry of luminal contents that activate previ-usly sensitized CD4� T cells in the colon,16 initiating orerpetuating an inflammatory response, but the exactechanisms of the association are still poorly under-
tood.
Abbreviations used in this paper: COX, cyclooxygenase; 15d-PGJ2,5-deoxy-�12,14-prostaglandin J2; NF-�B, nuclear factor �B; NOS, nitricxide synthase; MPO, myeloperoxidase; PPAR, peroxisome prolifera-or-activated receptor; TJ, tight junction.
© 2007 by the AGA Institute0016-5085/07/$32.00
doi:10.1053/j.gastro.2007.02.032

sntbimuep(taeaAv(igniopbrrsmNp
vihwtamraolscpddPdPtPPbi
iipfihp(furtS9
hradSctb
rwtdsswmt
iCiMpoitrwAagn1
i
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1792 PONFERRADA ET AL GASTROENTEROLOGY Vol. 132, No. 5
A question of special relevance is the possibility thatome of the changes observed in stress would be a featureot of the deleterious effects of stress but of some po-entially reversible type of adaptive plasticity, as proposedy McEwen.18 Most species of higher multicellular organ-
sms have evolved complex homeostatic and defenseechanisms that allow cells to overcome stressful stim-
li. One such mechanism that has received considerablexperimental attention as a possible anti-inflammatoryathway is peroxisome proliferator-activated receptors
PPARs), a group of 3 ligand-regulated transcription fac-ors originally implicated in adipocyte differentiationnd glucose homeostasis. One of its isoforms, PPAR�,xerts an anti-inflammatory effect by interfering with thectivity of inflammatory transcription factors, such asctivator Protein-1 (AP-1), Signal Transducers and Acti-ators of Transcription (STAT), and nuclear factor �BNF-�B).19 Three isoforms, as a result of alternative splic-ng and differential promoter usage within the sameene, are expressed in humans; PPAR�1, the predomi-ant isoform, is expressed in a broad range of tissues,
ncluding small bowel and colon.20 Therefore, activationf PPAR� by natural agonists, such as 15-deoxy-�12,14-rostaglandin J2 (15d-PGJ2),21 or synthetic agonists cane considered as a pathway to halt the inflammatoryesponse. Indeed, PPAR� ligands have been shown toeduce markedly colonic inflammation in both dextranodium sulfate and trinitrobenzene sulfonic acid colitis
odels in mice,22,23 probably through inhibition ofF-�B activity and decrease of proinflammatory cytokineroduction.To further investigate the possible mechanisms in-
olved in stress-induced inflammation and dysfunctionn the normal colon and the role of PPAR� as a possibleomeostatic mechanism in the colon of stressed animals,e decided to explore whether this may serve as a new
arget for protective pharmacologic maneuvers. We usedstress paradigm that has been reported to cause accu-ulation of oxidative mediators in the gut.17 Acute or
epeated immobilization stress is an easy, reproducible,nd widely used model to study pathophysiologic aspectsf the stress response and to study possible pharmaco-
ogic manipulations to decrease the negative effects oftress exposure on gastrointestinal, cardiovascular, orentral nervous systems.24 We used a pharmacologic ap-roach using natural and synthetic PPAR� agonists (theeoxy-prostaglandin 15d-PGJ2 and the thiazolidinedionerug rosiglitazone, respectively) as well as the specificPAR� antagonist T0070907. Rosiglitazone is a well-ocumented high-affinity and selective ligand forPAR�25 and has been shown to activate PPAR� in in-estinal epithelial cells both in vitro26 and in vivo.27 15d-GJ2 has also been shown to be a high-affinity ligand forPAR�.28,29 On the other hand, T0070907 has previouslyeen shown to inhibit PPAR� activity both in vitro30 and
n vivo.31 b
Materials and MethodsAnimals and Stress ProtocolYoung adult male Wistar rats (Animal Care Facil-
ty of the Complutense University, Madrid, Spain) weigh-ng 210 –230 g were used in this study. All experimentalrotocols adhered to the guidelines of the Animal Wel-are Committee of the Universidad Complutense follow-ng European legislation (2003/65/EC). The rats wereoused individually under standard conditions of tem-erature and humidity and a 12-hour light/dark cycle
lights on at 8 AM) with free access to standard pelletedormula and tap water. All animals were maintainednder constant conditions for 4 days before study. Theestraint was performed using a plastic rodent restrainerhat allowed for a close fit to rats in their home cages.32
tress consisted of 6 hours of immobilization starting atAM for 1, 5, or 10 consecutive days.Control animals were not subjected to stress but were
andled at 9 AM for a few seconds. Immediately afterestraint (still in the restrainer), animals were weighednd killed using intraperitoneally (IP) administered so-ium pentobarbital (320 mg/kg; Vetoquinol, Madrid,pain). Blood for plasma determinations was collected byardiac puncture and anticoagulated in the presence ofrisodium citrate (3.15% wt/vol, 1 vol citrate per 9 vollood).
The colon was removed, and after assessment of mac-oscopic damage, the distal 8-cm portion of the colonas removed and cut longitudinally for the determina-
ions described in the following text. For biochemicaleterminations, tissue was frozen at �80°C until as-ayed. For image studies, tissue was processed as de-cribed in the following text. A separate group of animalsas used for colonic paracellular permeability assay im-ediately after the stress sessions for the in vivo lumen-
o-blood ratio after 51Cr-EDTA instillation.33,34
Various groups of animals were IP injected with ros-glitazone maleate (3 mg/kg, dissolved in saline; Alexisorp, San Diego, CA) or 15d-PGJ2 (120 �g/kg, dissolved
n dimethyl sulfoxide 10%; Cayman Chemical, Ann Arbor,I). The doses of both drugs were chosen based on
revious findings showing maximum inhibitory activityn NOS-2 expression and tumor necrosis factor � release
n a model of stress-induced brain damage.35,36 None ofhe parameters studied were modified in saline-treatedats when compared with noninjected animals. The drugsere injected at the onset of the daily stress session.nimals receiving saline at the onset of stress (either incute or in repeated exposure) were used as the controlroup in Results and the figures. Finally, 2 groups ofonstressed animals received 3 mg/kg rosiglitazone IP or20 �g/kg 15d-PGJ2 IP at 9 AM.
On the other hand, various groups of rats were IPnjected with T0070907 (2-chloro-5-nitro-N-4-pyridinyl-
enzamide; Cayman Chemical), a specific PPAR� antag-
ot
dmmhTaipag
uSsucNazti
(a1�a(uRw
u(fwoBcC5r(PX
tAwge
Swmemn�Nmagbwpgmbof
fio3irn9ttlicma
(WEcs
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 PPAR� AND COLONIC DYSFUNCTION 1793
nist. The drug was injected IP 1 hour before the onset ofhe stress and the injection of PPAR� agonists.
Each experimental group contained at least 8 animals.
Myeloperoxidase ActivityImmediately after the animals were killed and
issected, colonic samples were minced on ice and ho-ogenized (glass/glass) in 0.5% hexadecyltrimethylam-onium bromide, 0.5% Nonidet P40 (Boehringer, Mann-
eim, Germany) in 20 mmol/L phosphate buffer, pH 6.0.he homogenates were then centrifuged for 20 minutest 12,000g. Tissue levels of myeloperoxidase (MPO) activ-ty were determined on supernatants using hydrogeneroxide as substrate for the enzyme. A unit of MPOctivity was defined as that converting 1 �mol of hydro-en peroxide to water in 1 minute at 40°C.37
ProstaglandinsColonic PGE2 and PGJ2 levels were determined
sing enzyme immunoassay kits (Biolink, Barcelona,pain, and Amersham, Buckinghamshire, England, re-pectively). Samples were purified using Amprep minicol-mns (Amersham). After homogenizing colons by soni-ation (Branson Sonifier 250; American Lab Trading,iantic, CT) in the same buffer used for Western blot
nalysis (see following text), the fraction needed for en-yme immunoassay isolation was obtained and PG quan-ification was performed following the manufacturer’snstructions.
Western Blot AnalysisTo determine the levels of NOS-2, cyclooxygenase
COX)-1, COX-2, and PPAR�, tissues were homogenizedt 4°C in 5 vol of buffer containing 320 mmol/L sucrose,
mmol/L DL-dithiothreitol, 10 �g/mL leupeptin, 10g/mL soybean trypsin inhibitor, 2 �g/mL aprotinin,nd 50 mmol/L Tris brought to pH 7.0 at 20°C with HClhomogenization buffer), and supernatants after centrif-gation at 12,000g for 20 minutes were used. For p65/elA fraction of NF-�B, cytosolic and nuclear extractsere obtained (see following text).After centrifugation in a microcentrifuge for 15 min-
tes, the proteins present in the supernatant were loaded20 �g) and size separated in 10% sodium dodecyl sul-ate/polyacrylamide gel electrophoresis (90 mV). The gelsere processed against the antigens and after blottingnto a polyvinylidene difluoride membrane (Millipore,edford, MA) were incubated with specific goat poly-lonal anti-rat COX-1 (1:1000), goat polyclonal anti-ratOX-2 (1:1000), rabbit polyclonal anti-rat NOS-2 (1:00), mouse monoclonal anti-rat PPAR� (1:1000), andabbit polyclonal anti-rat p65/RelA (1:1000) antibodiesall from Santa Cruz Biotechnology, Santa Cruz, CA).roteins recognized by the antibody were visualized on
-ray film by chemiluminescence following the manufac- uurer’s instructions (Amersham lbérica, Madrid, Spain).utoradiographs were quantified by densitometry (Soft-are Total Lab Dynamics Ltd, Phoretix, Newcastle, En-land), and several time expositions were analyzed tonsure the linearity of the band intensities.
Preparation of Cytosolic and NuclearExtractsA modified procedure based on the method of
chreiber et al38 was used. Tissues were homogenizedith 300 �L of buffer (10 mmol/L HEPES, pH 7.9, 1mol/L EDTA, 1 mmol/L ethylene glycol-bis[�-amino-
thyl ether]-N,N,N=,N=-tetraacetic acid, 10 mmol/L KCl, 1mol/L dithiothreitol, 0.5 mmol/L phenylmethylsulfo-
yl fluoride, 0.1 �g/mL aprotinin, 1 �g/mL leupeptin, 1g/mL N�-p-tosyl-L-lysine-chloromethyl ketone, 5 mmol/LaF, 1 mmol/L NaVO4, 0.5 mol/L sucrose, and 10mol/L Na2MoO4). After 15 minutes, Nonidet P40 was
dded to reach a 0.5% concentration. The tubes wereently vortexed for 15 seconds, and nuclei were collectedy centrifugation at 8000g for 5 minutes. Supernatantsere taken as a cytosolic fraction. The pellets were resus-ended in 100 �L of buffer supplemented with 20%lycerol and 0.4 mol/L KCl and gently shaken for 30inutes at 4°C. Nuclear protein extracts were obtained
y centrifugation at 13,000g for 5 minutes, and aliquotsf the supernatant were stored at �80°C. All steps of theractionation were performed at 4°C.
Electron MicroscopyColon segments from 4 rats per group were
xed in 2% glutaraldehyde for 3 hours, cryoprotectedvernight at 4°C in buffer phosphate 0.1 mol/L with0% sucrose, postfixed in 1% osmium tetroxide, stained
n 1% uranyl acetate, and embedded in Epon-Aralditeesin. Ultrathin sections (70 nm) were collected onickel grids. Sections were examined under Zeiss EM-02 (Zeiss, Oberkochen, Germany) transmission elec-ron microscopy. To evaluate changes in tight junc-ions (TJs), the junctional regions of several sections ofongitudinally sectioned villi per animal were exam-ned by a member of the study group blinded to theonditions of the experiment. At least 30 TJs per ani-al were evaluated. Results were expressed as percent-
ge of opened (ie, dilation �20 nm) TJs.
Assessment of Colonic PermeabilityRats were anesthetized with halothane. A catheter
OD, 1 mm) was inserted rectally at 4 cm from the anus.e slowly perfused 1.5 �Ci 51Cr-EDTA (Perkin Elmer
spaña, Madrid, Spain) in 0.5 mL NaCl 0.9% into theolon (0.25 mL/h) to control rats or just after the lastession in stressed animals. After 2 hours, rats were killed
sing sodium pentobarbital and blood was collected by
ctbapba
IsdrhtMamhctclilbt
biLn6ve
aiiwts
a
sagswnhhwaksii
F1kdsdrkSs
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1794 PONFERRADA ET AL GASTROENTEROLOGY Vol. 132, No. 5
ardiac puncture and anticoagulated in the presence ofrisodium citrate (3.15% wt/vol, 1 vol citrate per 9 vollood). The [51Cr]-bound radioactivity was counted usinggamma counter to measure radioactivity of the sam-
les. The permeability was expressed as the ratio betweenlood and total 51Cr instilled and reported as a percent-ge, as previously described.33,34
Bacterial TranslocationRats were anesthetized with sodium pentobarbital
P (320 mg/kg; Vetoquinol), and the abdominal skin washaved and sterilized with an iodine solution. Aftereath, mesenteric lymph nodes, liver, and spleen wereemoved under sterile conditions. After weighing andomogenization, aliquots (2 mL) of serial 10-fold dilu-ions of the suspension were plated onto 5% blood and
acConkey’s agar plates for recovery of aerobic bacteriand Brucella blood agar plates supplemented with vita-in K1 and hemin for anaerobic bacteria. After 24 and 48
ours of incubation at 37°C, for aerobic and anaerobicultures, respectively, colonies were counted. Quantita-ive culture results were expressed as the number ofolony-forming units per gram. Any positive mesentericymph node, liver, or spleen cultures were consideredndicative of bacterial translocation from the intestinalumen. Bacterial strains were identified by Gram stain,iochemical tests, and standard biochemical identifica-ion systems.
Plasma Corticosterone AssayPlasma was obtained within 1 hour of obtaining
lood samples by using a commercially available radio-mmunoassay kit of 131I-labeled rat corticosterone (DPC,os Angeles, CA). The sensitivity of the assay was 10g/mL. The values obtained in control animals (89.3 �.1 ng/mL) matched the kit manufacturer’s expectedalues in adult male Wistar rats at the time of bloodxtraction (�15.00 hours).
Chemicals and Statistical AnalysisChemicals were from Sigma (Barcelona, Spain) or
s indicated. Results are expressed as mean � SE of thendicated number of experiments, and statistical compar-sons were made using the Mann–Whitney test and one-ay analysis of variance followed by the Newman–Keuls
est, as appropriate. P � .05 was considered statisticallyignificant.
ResultsEffect of Immobilization Stress on BodyWeight and Colonic Inflammation and Effectof Pharmacologic PPAR� ActivationRepeated immobilization during 5 days induced
significant weight loss in comparison with non- w
tressed animals (Figure 1A). The macroscopic appear-nce of colonic mucosa was not affected in stressroups compared with control animals (data nothown). Nevertheless, MPO activity in colonic mucosa,hich has been shown to be linearly correlated witheutrophil infiltration,37 resulted in significantlyigher colonic homogenates of rats exposed to acute (6ours) stress and 5 days of immobilization comparedith nonstressed animals (Figure 1B). Intraperitonealdministration of PPAR� agonists rosiglitazone (3 mg/g) and 15d-PGJ2 (120 �g/kg) before each stress ses-ion prevented both weight loss (Figure 1A) and anncrease in MPO (Figure 1B). Administration of ros-glitazone or 15d-PGJ2 to control animals did not alter
igure 1. (A) Effect of immobilization stress during 6 hours, 5 days, and0 days on weight of rats receiving vehicle (saline), rosiglitazone (3 mg/g), or 15d-PGJ2 (120 �g/kg) at the onset of each stress session. Theata represent the mean � SEM of 8 rats. *P � .05 vs control non-tressed animals; #,†P � .05 vs saline. (B) Effect of immobilization stressuring 6 hours, 5 days, and 10 days on colonic MPO activity of ratseceiving vehicle (saline), rosiglitazone (3 mg/kg), or 15d-PGJ2 (120 �g/g) at the onset of each stress session. The data represent the mean �EM of 8 rats. *P � .05 vs control nonstressed animals; #,†P � .05 vsaline.
eight or colonic MPO activity.
vpNysacwsT(da
mssedilPcat
mPaoi4ha
lwsic(cfIA
Fcrr(t*bd(e*
Fe
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 PPAR� AND COLONIC DYSFUNCTION 1795
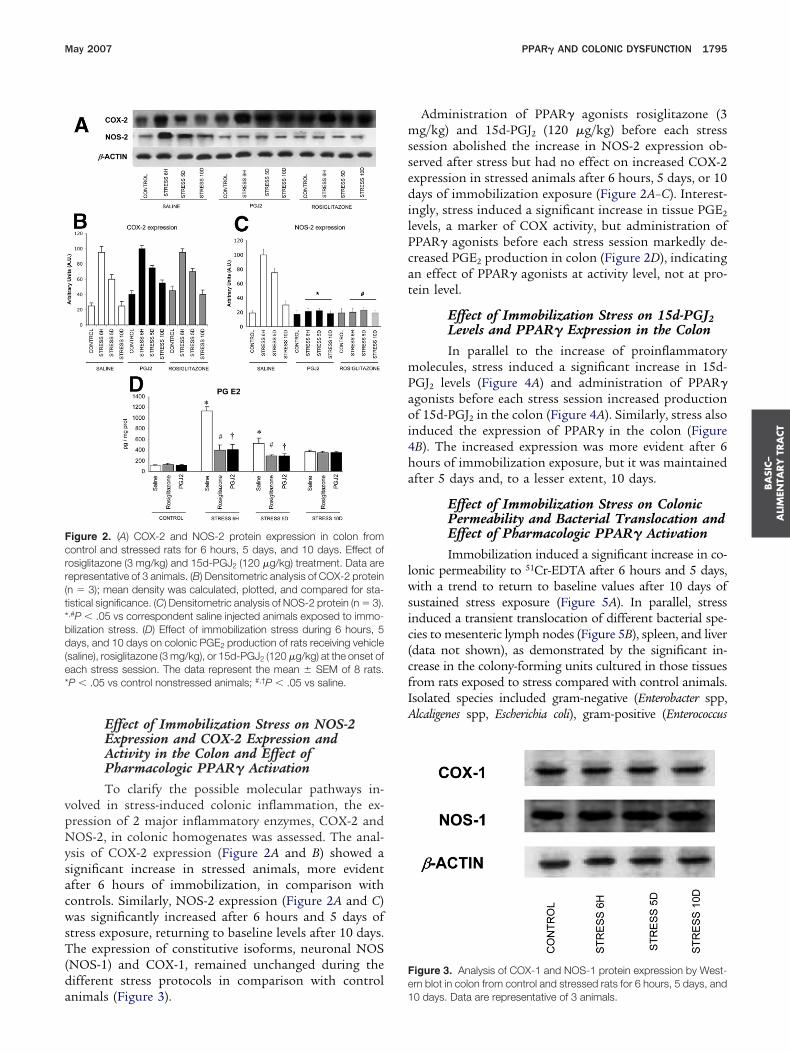
Effect of Immobilization Stress on NOS-2Expression and COX-2 Expression andActivity in the Colon and Effect ofPharmacologic PPAR� ActivationTo clarify the possible molecular pathways in-
olved in stress-induced colonic inflammation, the ex-ression of 2 major inflammatory enzymes, COX-2 andOS-2, in colonic homogenates was assessed. The anal-
sis of COX-2 expression (Figure 2A and B) showed aignificant increase in stressed animals, more evidentfter 6 hours of immobilization, in comparison withontrols. Similarly, NOS-2 expression (Figure 2A and C)as significantly increased after 6 hours and 5 days of
tress exposure, returning to baseline levels after 10 days.he expression of constitutive isoforms, neuronal NOS
NOS-1) and COX-1, remained unchanged during theifferent stress protocols in comparison with control
igure 2. (A) COX-2 and NOS-2 protein expression in colon fromontrol and stressed rats for 6 hours, 5 days, and 10 days. Effect ofosiglitazone (3 mg/kg) and 15d-PGJ2 (120 �g/kg) treatment. Data areepresentative of 3 animals. (B) Densitometric analysis of COX-2 proteinn � 3); mean density was calculated, plotted, and compared for sta-istical significance. (C) Densitometric analysis of NOS-2 protein (n � 3).,#P � .05 vs correspondent saline injected animals exposed to immo-ilization stress. (D) Effect of immobilization stress during 6 hours, 5ays, and 10 days on colonic PGE2 production of rats receiving vehicle
saline), rosiglitazone (3 mg/kg), or 15d-PGJ2 (120 �g/kg) at the onset ofach stress session. The data represent the mean � SEM of 8 rats.P � .05 vs control nonstressed animals; #,†P � .05 vs saline.
nimals (Figure 3). 1
Administration of PPAR� agonists rosiglitazone (3g/kg) and 15d-PGJ2 (120 �g/kg) before each stress
ession abolished the increase in NOS-2 expression ob-erved after stress but had no effect on increased COX-2xpression in stressed animals after 6 hours, 5 days, or 10ays of immobilization exposure (Figure 2A–C). Interest-
ngly, stress induced a significant increase in tissue PGE2
evels, a marker of COX activity, but administration ofPAR� agonists before each stress session markedly de-reased PGE2 production in colon (Figure 2D), indicatingn effect of PPAR� agonists at activity level, not at pro-ein level.
Effect of Immobilization Stress on 15d-PGJ2Levels and PPAR� Expression in the ColonIn parallel to the increase of proinflammatory
olecules, stress induced a significant increase in 15d-GJ2 levels (Figure 4A) and administration of PPAR�gonists before each stress session increased productionf 15d-PGJ2 in the colon (Figure 4A). Similarly, stress also
nduced the expression of PPAR� in the colon (FigureB). The increased expression was more evident after 6ours of immobilization exposure, but it was maintainedfter 5 days and, to a lesser extent, 10 days.
Effect of Immobilization Stress on ColonicPermeability and Bacterial Translocation andEffect of Pharmacologic PPAR� ActivationImmobilization induced a significant increase in co-
onic permeability to 51Cr-EDTA after 6 hours and 5 days,ith a trend to return to baseline values after 10 days of
ustained stress exposure (Figure 5A). In parallel, stressnduced a transient translocation of different bacterial spe-ies to mesenteric lymph nodes (Figure 5B), spleen, and liverdata not shown), as demonstrated by the significant in-rease in the colony-forming units cultured in those tissuesrom rats exposed to stress compared with control animals.solated species included gram-negative (Enterobacter spp,lcaligenes spp, Escherichia coli), gram-positive (Enterococcus
igure 3. Analysis of COX-1 and NOS-1 protein expression by West-rn blot in colon from control and stressed rats for 6 hours, 5 days, and
0 days. Data are representative of 3 animals.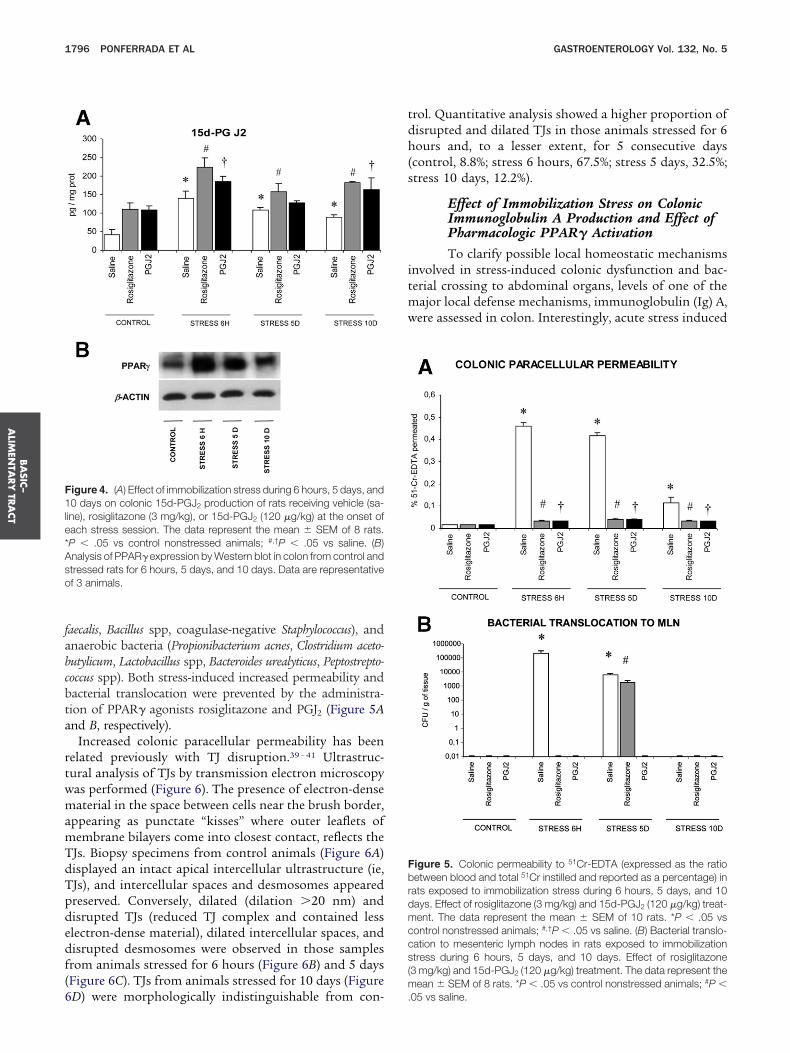
fabcbta
rtwmamTdTpdedf(6
tdh(s
itmw
F1le*Aso
Fbrdmccs(m
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1796 PONFERRADA ET AL GASTROENTEROLOGY Vol. 132, No. 5
aecalis, Bacillus spp, coagulase-negative Staphylococcus), andnaerobic bacteria (Propionibacterium acnes, Clostridium aceto-utylicum, Lactobacillus spp, Bacteroides urealyticus, Peptostrepto-occus spp). Both stress-induced increased permeability andacterial translocation were prevented by the administra-ion of PPAR� agonists rosiglitazone and PGJ2 (Figure 5And B, respectively).
Increased colonic paracellular permeability has beenelated previously with TJ disruption.39 – 41 Ultrastruc-ural analysis of TJs by transmission electron microscopyas performed (Figure 6). The presence of electron-denseaterial in the space between cells near the brush border,
ppearing as punctate “kisses” where outer leaflets ofembrane bilayers come into closest contact, reflects theJs. Biopsy specimens from control animals (Figure 6A)isplayed an intact apical intercellular ultrastructure (ie,Js), and intercellular spaces and desmosomes appearedreserved. Conversely, dilated (dilation �20 nm) andisrupted TJs (reduced TJ complex and contained lesslectron-dense material), dilated intercellular spaces, andisrupted desmosomes were observed in those samplesrom animals stressed for 6 hours (Figure 6B) and 5 daysFigure 6C). TJs from animals stressed for 10 days (Figure
igure 4. (A) Effect of immobilization stress during 6 hours, 5 days, and0 days on colonic 15d-PGJ2 production of rats receiving vehicle (sa-
ine), rosiglitazone (3 mg/kg), or 15d-PGJ2 (120 �g/kg) at the onset ofach stress session. The data represent the mean � SEM of 8 rats.P � .05 vs control nonstressed animals; #,†P � .05 vs saline. (B)nalysis of PPAR� expression by Western blot in colon from control andtressed rats for 6 hours, 5 days, and 10 days. Data are representativef 3 animals.
D) were morphologically indistinguishable from con- .
rol. Quantitative analysis showed a higher proportion ofisrupted and dilated TJs in those animals stressed for 6ours and, to a lesser extent, for 5 consecutive days
control, 8.8%; stress 6 hours, 67.5%; stress 5 days, 32.5%;tress 10 days, 12.2%).
Effect of Immobilization Stress on ColonicImmunoglobulin A Production and Effect ofPharmacologic PPAR� ActivationTo clarify possible local homeostatic mechanisms
nvolved in stress-induced colonic dysfunction and bac-erial crossing to abdominal organs, levels of one of the
ajor local defense mechanisms, immunoglobulin (Ig) A,ere assessed in colon. Interestingly, acute stress induced
igure 5. Colonic permeability to 51Cr-EDTA (expressed as the ratioetween blood and total 51Cr instilled and reported as a percentage) inats exposed to immobilization stress during 6 hours, 5 days, and 10ays. Effect of rosiglitazone (3 mg/kg) and 15d-PGJ2 (120 �g/kg) treat-ent. The data represent the mean � SEM of 10 rats. *P � .05 vs
ontrol nonstressed animals; #,†P � .05 vs saline. (B) Bacterial translo-ation to mesenteric lymph nodes in rats exposed to immobilizationtress during 6 hours, 5 days, and 10 days. Effect of rosiglitazone3 mg/kg) and 15d-PGJ2 (120 �g/kg) treatment. The data represent the
ean � SEM of 8 rats. *P � .05 vs control nonstressed animals; #P �
05 vs saline.
arev
PgcTtbotmmw
tTmomtt
Fdzsc
Ftgfed(falrn
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 PPAR� AND COLONIC DYSFUNCTION 1797
significant decrease in colonic IgA production thateturned to baseline values after 5 days of repeated stressxposure (Figure 7). Both PPAR� agonists evaluated pre-ented the decrease in IgA after acute stress.
igure 7. Effect of immobilization stress during 6 hours, 5 days, and 10ays on colonic IgA synthesis of rats receiving vehicle (saline), rosiglita-one (3 mg/kg), or 15d-PGJ2 (120 �g/kg) at the onset of each stressession. The data represent the mean � SEM of 8 rats. *P � .05 vs
igure 6. Transmission elec-ron micrographs of the TJ re-ion of colonic epithelial cells
rom (A) control rats or animalsxposed to immobilization stressuring (B) 6 hours, (C) 5 days, and
D) 10 days. Immobilization stressor 6 hours and 5 days inducedn increased proportion of TJ di-
atation and disruption (white ar-ows) versus control animals (seeumerical data in the text).
aontrol nonstressed animals; #,†P � .05 vs saline.
Effect of Immobilization Stress onCorticosterone Release and Effect ofPharmacologic PPAR� ActivationTo test the possibility that the effects of selective
PAR� activation were related to interference on theeneral response to stress, we determined plasma corti-osterone levels in all of the experimental groups studied.he effects elicited by the PPAR� ligands after stress in
he colon are independent of the systemic stress response,ecause plasma corticosterone levels after 6 hours, 5 days,r 10 days were not different between the saline, rosigli-azone, or 15d-PGJ2 groups. Similarly, nonstressed ani-
als receiving rosiglitazone or 15d-PGJ2 did not haveodified plasma corticosterone levels when comparedith control (Figure 8).
Effect of Pharmacologic PPAR� Inhibition onAcute Stress-Induced Colonic Inflammationand DysfunctionTo verify the anti-inflammatory role of PPAR� in
he colon in vivo, the specific PPAR� antagonist0070907 was used in different sets of experiments. Ad-inistration of T0070907 (2 mg/kg IP) 1 hour before the
nset of each stress session exacerbated colonic inflam-ation (increased MPO activity [Figure 9A], PGE2 syn-
hesis [Figure 9B], and NOS-2 expression [Figure 9C]) inhose animals treated with PPAR� agonists rosiglitazone
nd 15d-PGJ2. Furthermore, T0070907 blocked the pre-
vcstbdu
asicieadoatPmtsia
tirow
mttcsposwHatosamittPph
sglplpsnpmstpd
dtmaidscis
PrmpsP
Fdtsc
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1798 PONFERRADA ET AL GASTROENTEROLOGY Vol. 132, No. 5
entive effect of PPAR� agonists on the stress-inducedolonic dysfunction as seen by 51Cr-EDTA permeabilitytudies (Figure 9D) and bacterial translocation to mesen-eric lymph nodes (Figure 9E). Moreover, T0070907locked the effects of PPAR� agonists on stress-inducedecreased IgA synthesis in colonic homogenates (Fig-re 9F).
DiscussionThe reported results showed that (1) exposure to
cute (6 hours) or subacute (5 days) immobilizationtress induces weight loss and an increase in MPO activ-ty reflecting neutrophil infiltration of the colonic mu-osa, as well as an increase in the expression of enzymaticnflammatory sources NOS-2 and COX-2; (2) colonicpithelial barrier results morphologically and function-lly altered after stress exposure and IgA productionecrease after stress; (3) stress leads to an early inductionf PPAR� expression and 15d-PGJ2 synthesis in the colon;nd (4) this stress-induced inflammation and dysfunc-ion was prevented with both natural and syntheticPAR� ligands (15d-PGJ2 and rosiglitazone), whereas ad-inistration of an specific PPAR� antagonist reverted
hose effects. On the other hand, subchronic (10 days)tress exposure leads to the normalization of MPO activ-ty, proinflammatory enzyme expression, colonic perme-bility, and colonic IgA production.
Growing evidence from experimental studies supportshat psychological and physical stress can lead to acutentestinal inflammatory response and barrier dysfunctionesulting in bacterial translocation and enhanced uptakef luminal antigens, which, consequently, are associated
igure 8. Effect of immobilization stress during 6 hours, 5 days, and 10ays on corticosterone release of rats receiving vehicle (saline), rosigli-azone (3 mg/kg), or 15d-PGJ2 (120 �g/kg) at the onset of each stressession. The data represent the mean � SEM of 8 rats. *P � .05 vsontrol nonstressed animals; #,†P � NS vs saline.
ith maintained stimulation of lamina propria and sub- P
ucosa immune cells and finally with chronic inflamma-ion.42,43 Interestingly, our results indicate a normaliza-ion of the colonic inflammatory response andonsequent barrier dysfunction after repeated predictabletressful stimuli. The trend to the normalization of MPO,roinflammatory enzymes, and colonic barrier functionbserved in our experiments after 10 days of repeatedtress could be related to an adaptation to the stressorith lack of corticotropin-releasing factor response.owever, taking into account that corticosterone levels
re still significantly higher after 10 days of immobiliza-ion stress compared with control animals, the activationf additional compensatory mechanisms that counteracthort exposure to stress-induced colonic inflammationnd dysfunction is suggested. In this vein, our experi-ents have focused on PPAR� because of the increas-
ngly recognized anti-inflammatory activity of this recep-or44 and because the colon is the organ, together withhe adipose tissue, with the greatest concentration ofPAR� messenger RNA in the organism,20 suggesting aivotal role of this receptor in the regulation of colonicomeostasis.Numerous experimental and clinical studies demon-
trate anti-inflammatory effects of PPAR� agonists in theut; treatment with PPAR� agonists such as thiazo-idinediones, 15d-PGJ2, and other members of the cyclo-entenone family of prostaglandins, or conjugated lino-
eic acid, has been shown to protect against inflammatoryrocesses in the gut.22,23,45– 48 PPAR� heterozygous micehow a significantly enhanced susceptibility to 2,4,6-tri-itrobenzene sulfonic acid–induced colitis,23 PPAR� ex-ression has been found to be impaired in the colonicucosa of patients with ulcerative colitis,49 and a pilot
tudy assessing the efficacy of the PPAR� agonist rosigli-azone in patients with ulcerative colitis has shownromising results that could be the starting point for theesign of future randomized controlled trials.50
PPAR� exerts its anti-inflammatory effects throughifferent mechanisms: inhibition of proinflammatory cy-okines,51 down-regulation of the expression of adhesion
olecules,52 and inhibition of the NOS-2 expression andctivity.53 All these effects seem to be mediated by thenterference with NF-�B transcriptional activity.22,54 In-eed, we have found that the expression of the RelAubunit of NF-�B in nuclear extracts from colonic mu-osa increased after 6 hours and 5 days of stress exposuren comparison with control, but not after 10 days oftress (data not shown).
Among the identified regulatory mechanisms ofPAR� expression, glucocorticoids could play a centralole in the stress-induced up-regulation observed in our
odel. Glucocorticoids induce PPAR� and PPAR� ex-ression in various cell types,55–58 and immobilizationtress has been demonstrated as a potent stimulator ofPAR� expression in the liver.56 On the other hand,
PAR� is considered to be a ligand-activated transcrip-
FW(r1
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 PPAR� AND COLONIC DYSFUNCTION 1799
igure 9. Effect of immobilization stress during 6 hours on (A) colonic MPO activity, (B) PGE2 production, (C) NOS-2 expression (representativeestern blot [inset] and densitometry plot, n � 3), (D) colonic permeability to 51Cr-EDTA, (E) bacterial translocation to mesenteric lymph nodes, and
F) colonic IgA production of rats receiving vehicle (saline), T0070907 (2 mg/kg), rosiglitazone (3 mg/kg), 15d-PGJ2 (120 �g/kg), T0070907 plusosiglitazone, or T0070907 plus 15d-PGJ . The data represent the mean � SEM of 8 rats. *P � .05 vs rosiglitazone-treated animals; #P � .05 vs
25d-PGJ2–treated animals.

tptutes
aacfPptrPbd2eattpbc
sptvrtaflivdarciabndflseitob
mc
vidAdpldIsdfpclgletuctic1t
bsientfm1alma
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1800 PONFERRADA ET AL GASTROENTEROLOGY Vol. 132, No. 5
ion factor, and its regulation, apart from the gene ex-ression level, can occur depending on ligand availability;hus, taking into account the possibility of ligand-stim-lated displacement of the transcriptional corepressorhat is constitutively associated with the PPAR/RXR het-rodimer,59 the stress-induced increase in 15d-PGJ2 ob-erved may explain the increase in PPAR� activation.
Furthermore, our results show that PPAR� activationlso inhibits the stress-induced increased colonic perme-bility, decreased IgA production, and bacterial translo-ation. The results obtained with the PPAR� antagonisturther support the role of this receptor. Activation ofPAR� by butyrate has been shown to reduce colonicermeability, probably through the promotion of intes-inal epithelial cell differentiation and the consequenteinforcement of the TJs.60 Additionally, the effects ofPAR� on the regulation of colonic permeability coulde also related to the ability, shown in this report, ofecreasing NOS-2 expression and PGE2 synthesis. NOS-– derived NO has been shown to increase intestinalpithelial permeability both in vitro61,62 and in vivo63 inn effect that seems to be related to NO-induced cy-oskeleton rearrangement and subsequent TJ dysfunc-ion.61,63– 65 Finally, the release of PGs after lamina pro-ria mast cell degranulation induced by acute stress haseen proposed as a mechanism of stress-induced in-reased colonic permeability.66
The stress-induced increase of 15d-PGJ2 in the coloneems to run parallel with the expression of COX-2, asreviously shown in the brain.36 It has been suggestedhat the enhanced production of 15d-PGJ2 in the acti-ated macrophages during the inflammatory process mayepresent a response that contributes to negative regula-ion of inflammation,67 probably through its ability toctivate PPAR�.28 Apart from the anti-inflammatory ef-ects mediated by 15d-PGJ2 as a high-affinity PPAR�igand, it has shown important PPAR�-independent anti-nflammatory activity by inhibiting transcriptional acti-ation via NF-�B.68,69 In this vein, we have shown aown-regulation of NOS-2 by 15d-PGJ2 in our model, ingreement with previous findings in neurons and mac-ophages.36,70,71 Furthermore, the findings of an in-reased severity of dextran sodium sulfate–induced colitisn rats by pretreatment with selective COX-2 inhibitors72
nd of the induction of a colonic secretory dysfunctiony pre-exposure of normal colonic tissue to PGD2, butot to 15d-PGJ2,73 supports the hypothesis that COX-2–erived 15d-PGJ2 could exert a protective and anti-in-ammatory role in the gut. Interestingly, it has beenhown that PGE2 synthesis predominates during thearly inflammatory step in carrageenin-induced pleurisyn rats, whereas 15d-PGJ2 substitutes PGE2 formation athe end of the process, coincident with the accumulationf macrophages.74 Interestingly, 15d-PGJ2 has recently
een shown to decrease PGE2 synthesis by means oficrosomal PGE synthase down-regulation in chondro-ytes75 and intestinal epithelial cells.76
Although others have found a decrease in salivary andaginal secretion IgA levels after stress exposure,77,78 thiss the first report, to our knowledge, of stress-inducedecrease of IgA production in the colonic mucosa.mong the mechanisms responsible for the stress-in-uced decrease in IgA, endogenous glucocorticoids couldlay a central role. Previous studies have shown that IgA
evels in serum were elevated when increasing doses ofexamethasone were administered to rats; in contrast,gA levels significantly decreased in saliva and vaginalecretions over the same dose range.77 If the stress-in-uced diminished IgA levels in the colonic mucosa resultrom a true decrease of IgA synthesis by lamina proprialasma cells or if there is a lack of transportation to theolonic lumen deserves further investigation. Neverthe-ess, there is evidence supporting the first hypothesis: (1)lucocorticoids significantly reduced estradiol-inducedevels of IgA in uterine secretions with no effect onstradiol-induced levels of the secretory component ofhe polymeric Ig receptor79 and (2) glucocorticoids stim-late secretory component synthesis by hepatocytes inulture.80 In our model, the effect of PPAR� agonists onhe restoration of colonic IgA levels seems to have anndependent component on glucocorticoid secretion, be-ause treatment of stressed rats with rosiglitazone or5d-PGJ2 does not affect plasma corticosterone levels inhose animals.
In summary, the present work shows that acute immo-ilization stress leads to colonic barrier dysfunction thateems to be mediated by the increased expression ofnflammatory enzymes COX-2 and NOS-2. In parallel,arly PPAR� up-regulation and activation by its endoge-ous agonist 15d-PGJ2 as well as direct anti-inflamma-ory effects by 15d-PGJ2 are responsible, at least in part,or the return to baseline values of biochemical inflam-
atory markers and mucosal permeability observed after0 days of stress exposure, representing a physiologic,nti-inflammatory pathway that counteracts initial co-onic inflammation and dysfunction. The pharmacologic
odulation of this pathway represents a promising ther-peutic approach in stress-related intestinal disorders.
References
1. Dickhaus B, Mayer EA, Firooz N, Stains J, Conde F, Olivas TI, FassR, Chang L, Mayer M, Naliboff BD. Irritable bowel syndromepatients show enhanced modulation of visceral perception byauditory stress. Am J Gastroenterol 2003;98:135–143.
2. Wilhelmsen I. The role of psychosocial factors in gastrointestinaldisorders. Gut 2000;47(Suppl 4):iv73–iv75.
3. Mayer EA, Naliboff BD, Chang L, Coutinho SV. V. Stress andirritable bowel syndrome. Am J Physiol Gastrointest Liver Physiol2001;280:G519–G524.
4. Bennett EJ, Tennant CC, Piesse C, Badcock CA, Kellow JE. Levelof chronic life stress predicts clinical outcome in irritable bowel
syndrome. Gut 1998;43:256–261.
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 PPAR� AND COLONIC DYSFUNCTION 1801
5. Duffy LC, Zielezny MA, Marshall JR, Byers TE, Weiser MM, PhillipsJF, Calkins BM, Ogra PL, Graham S. Relevance of major stressevents as an indicator of disease activity prevalence in inflam-matory bowel disease. Behav Med 1991;17:101–110.
6. Levenstein S, Prantera C, Varvo V, Scribano ML, Berto E, AndreoliA, Luzi C. Psychological stress and disease activity in ulcerativecolitis: a multidimensional cross-sectional study. Am J Gastroen-terol 1994;89:1219–1225.
7. Levenstein S, Prantera C, Varvo V, Scribano ML, Andreoli A, LuziC, Arca M, Berto E, Milite G, Marcheggiano A. Stress and exac-erbation in ulcerative colitis: a prospective study of patientsenrolled in remission. Am J Gastroenterol 2000;95:1213–1220.
8. Bitton A, Sewitch MJ, Peppercorn MA, de Edwardess MD, Shah S,Ransil B, Locke SE. Psychosocial determinants of relapse inulcerative colitis: a longitudinal study. Am J Gastroenterol 2003;98:2203–2208.
9. Ilnyckyj A, Shanahan F, Anton P, Cheang M, Bernstein C. Quanti-fication of the placebo response in ulcerative colitis. Gastroen-terology 1997;112:1854–1858.
0. Su C, Lichtenstein G, Krok K, Brensinger C, Lewis J. A metaanaly-sis of the placebo rates of remission and response in clinicaltrials of active Crohn’s disease. Gastroenterology 2004;126:1257–1269.
1. Narducci F, Snape WJ Jr, Battle WM, London RL, Cohen S.Increased colonic motility during exposure to a stressful situa-tion. Dig Dis Sci 1985;30:40–44.
2. Collins SM, McHugh K, Jacobson K, Khan I, Riddell R, Murase K,Weingarten HP. Previous inflammation alters the response of therat colon to stress. Gastroenterology 1996;111:1509–1515.
3. Saunders PR, Hanssen NP, Perdue MH. Cholinergic nerves me-diate stress-induced intestinal transport abnormalities in Wistar-Kyoto rats. Am J Physiol 1997;273:G486–G490.
4. Ferrier L, Mazelin L, Cenac N, Desreumaux P, Janin A, Emilie D,Colombel JF, Garcia-Villar R, Fioramonti J, Bueno L. Stress-in-duced disruption of colonic epithelial barrier: role of interferon-gamma and myosin light chain kinase in mice. Gastroenterology2003;125:795–804.
5. Gue M, Bonbonne C, Fioramonti J, More J, Del Rio-Lacheze C,Comera C, Bueno L. Stress-induced enhancement of colitis inrats: CRF and arginine vasopressin are not involved. Am J Physiol1997;272:G84–G91.
6. Qiu BS, Vallance BA, Blennerhassett PA, Collins SM. The role ofCD4� lymphocytes in the susceptibility of mice to stress-inducedreactivation of experimental colitis. Nat Med 1999;5:1178–1182.
7. Colon AL, Madrigal JL, Menchen LA, Moro MA, Lizasoain I,Lorenzo P, Leza JC. Stress increases susceptibility to oxidative/nitrosative mucosal damage in an experimental model of colitisin rats. Dig Dis Sci 2004;49:1713–1721.
8. McEwen B. Protective and damaging effects of stress mediators.N Engl J Med 1998;338:171–179.
9. Ricote M, Li AC, Willson TM, Kelly CJ, Glass CK. The peroxisomeproliferator-activated receptor-gamma is a negative regulator ofmacrophage activation. Nature 1998;391:79–82.
0. Fajas L, Auboeuf D, Raspe E, Schoonjans K, Lefebvre AM, Sala-din R, Najib J, Laville M, Fruchart JC, Deeb S, Vidal-Puig A, Flier J,Briggs MR, Staels B, Vidal H, Auwerx J. The organization, pro-moter analysis, and expression of the human PPARgamma gene.J Biol Chem 1997;272:18779–18789.
1. Kliewer SA, Xu HE, Lambert MH, Willson TM. Peroxisome prolif-erator-activated receptors: from genes to physiology. Recent ProgHorm Res 2001;56:239–263.
2. Su CG, Wen X, Bailey ST, Jiang W, Rangwala SM, Keilbaugh SA,Flanigan A, Murthy S, Lazar MA, Wu GD. A novel therapy for colitisutilizing PPAR-gamma ligands to inhibit the epithelial inflamma-
tory response. J Clin Invest 1999;104:383–389.3. Desreumaux P, Dubuquoy L, Nutten S, Peuchmaur M, Englaro W,Schoonjans K, Derijard B, Desvergne B, Wahli W, Chambon P,Leibowitz MD, Colombel JF, Auwerx J. Attenuation of colon inflam-mation through activators of the retinoid X receptor (RXR)/perox-isome proliferator-activated receptor gamma (PPARgamma) het-erodimer. A basis for new therapeutic strategies. J Exp Med2001;193:827–838.
4. Pare WP, Glavin GB. Restraint stress in biomedical research: areview. Neurosci Biobehav Rev 1986;10:339–370.
5. Lehmann JM, Moore LB, Smith-Oliver TA, Wilkison WO, WillsonTM, Kliewer SA. An antidiabetic thiazolidinedione is a high affinityligand for peroxisome proliferator-activated receptor gamma(PPAR gamma). J Biol Chem 1995;270:12953–12956.
6. Gupta RA, Brockman JA, Sarraf P, Willson TM, DuBois RN. Targetgenes of peroxisome proliferator-activated receptor gamma incolorectal cancer cells. J Biol Chem 2001;276:29681–29687.
7. Adachi M, Kurotani R, Morimura K, Shah Y, Sanford M, MadisonBB, Gumucio DL, Marin HE, Peters JM, Young HA, Gonzalez FJ.Peroxisome proliferator activated receptor gamma in colonic ep-ithelial cells protects against experimental inflammatory boweldisease. Gut 2006;55:1104–1113.
8. Forman BM, Tontonoz P, Chen J, Brun RP, Spiegelman BM, EvansRM. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for theadipocyte determination factor PPAR gamma. Cell 1995;83:803–812.
9. Kliewer SA, Lenhard JM, Willson TM, Patel I, Morris DC, LehmannJM. A prostaglandin J2 metabolite binds peroxisome proliferator-activated receptor gamma and promotes adipocyte differentia-tion. Cell 1995;83:813–819.
0. Lee G, Elwood F, McNally J, Weiszmann J, Lindstrom M, Amaral K,Nakamura M, Miao S, Cao P, Learned RM, Chen JL, Li Y.T0070907, a selective ligand for peroxisome proliferator-acti-vated receptor gamma, functions as an antagonist of biochemi-cal and cellular activities. J Biol Chem 2002;277:19649–19657.
1. Collin M, Murch O, Thiemermann C. Peroxisome proliferator-activated receptor-gamma antagonists GW9662 and T0070907reduce the protective effects of lipopolysaccharide precondition-ing against organ failure caused by endotoxemia. Crit Care Med2006;34:1131–1138.
2. Leza JC, Salas E, Sawicki G, Russell JC, Radomski MW. Theeffects of stress on homeostasis in JCR-LA-cp rats: the role ofnitric oxide. J Pharmacol Exp Ther 1998;286:1397–1403.
3. Aiko S, Fuseler J, Grisham MB. Effects of nitric oxide synthaseinhibition or sulfasalazine on the spontaneous colitis observed inHLA-B27 transgenic rats. J Pharmacol Exp Ther 1998;284:722–727.
4. Stein J, Ries J, Barrett KE. Disruption of intestinal barrier functionassociated with experimental colitis: possible role of mast cells.Am J Physiol 1998;274:G203–G209.
5. Garcia-Bueno B, Madrigal JL, Lizasoain I, Moro MA, Lorenzo P,Leza JC. Peroxisome proliferator-activated receptor gamma acti-vation decreases neuroinflammation in brain after stress in rats.Biol Psychiatry 2005;57:885–894.
6. Garcia-Bueno B, Madrigal JL, Lizasoain I, Moro MA, Lorenzo P,Leza JC. The anti-inflammatory prostaglandin 15d-PGJ2 de-creases oxidative/nitrosative mediators in brain after acutestress in rats. Psychopharmacology 2005;180:513–522.
7. Bradley PP, Priebat DA, Christensen RD, Rothstein G. Measure-ment of cutaneous inflammation: estimation of neutrophil con-tent with an enzyme marker. J Invest Dermatol 1982;78:206–209.
8. Schreiber E, Matthias P, Muller MM, Schaffner W. Rapid detec-tion of octamer binding proteins with “mini-extracts,” preparedfrom a small number of cells. Nucleic Acids Res 1989;17:6419.
9. Schmitz H, Barmeyer C, Fromm M, Runkel N, Foss HD, Bentzel
CJ, Riecken EO, Schulzke JD. Altered tight junction structure
4
4
4
4
4
4
4
4
4
4
5
5
5
5
5
5
5
5
5
5
6
6
6
6
6
6
6
6
6
6
7
7
7
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1802 PONFERRADA ET AL GASTROENTEROLOGY Vol. 132, No. 5
contributes to the impaired epithelial barrier function in ulcerativecolitis. Gastroenterology 1999;116:301–309.
0. Soderholm JD, Olaison G, Peterson KH, Franzen LE, Lindmark T,Wiren M, Tagesson C, Sjodahl R. Augmented increase in tightjunction permeability by luminal stimuli in the non-inflamed ileumof Crohn’s disease. Gut 2002;50:307–313.
1. Demaude J, Salvador-Cartier C, Fioramonti J, Ferrier L, Bueno L.Phenotypic changes in colonocytes following acute stress oractivation of mast cells in mice: implications for delayed epithe-lial barrier dysfunction. Gut 2006;55:655–661.
2. Collins SM. Stress and the gastrointestinal tract IV. Modulationof intestinal inflammation by stress: basic mechanisms and clin-ical relevance. Am J Physiol Gastrointest Liver Physiol 2001;280:G315–G318.
3. Soderholm JD, Perdue MH. Stress and gastrointestinal tract. II.Stress and intestinal barrier function. Am J Physiol GastrointestLiver Physiol 2001;280:G7–G13.
4. Genolet R, Wahli W, Michalik L. PPARs as drug targets to modu-late inflammatory responses? Curr Drug Targets Inflamm Allergy2004;3:361–375.
5. Saubermann LJ, Nakajima A, Wada K, Zhao S, Terauchi Y, Kad-owaki T, Aburatani H, Matsuhashi N, Nagai R, Blumberg RS.Peroxisome proliferator-activated receptor gamma agonist li-gands stimulate a Th2 cytokine response and prevent acutecolitis. Inflamm Bowel Dis 2002;8:330–339.
6. Takagi T, Naito Y, Tomatsuri N, Handa O, Ichikawa H, Yoshida N,Yoshikawa T. Pioglitazone, a PPAR-gamma ligand, provides pro-tection from dextran sulfate sodium-induced colitis in mice inassociation with inhibition of the NF-kappaB-cytokine cascade.Redox Rep 2002;7:283–289.
7. Katayama K, Wada K, Nakajima A, Mizuguchi H, Hayakawa T,Nakagawa S, Kadowaki T, Nagai R, Kamisaki Y, Blumberg RS,Mayumi T. A novel PPAR gamma gene therapy to control inflam-mation associated with inflammatory bowel disease in a murinemodel. Gastroenterology 2003;124:1315–1324.
8. Bassaganya-Riera J, Reynolds K, Martino-Catt S, Cui Y, Hen-nighausen L, Gonzalez F, Rohrer J, Benninghoff AU, HontecillasR. Activation of PPAR gamma and delta by conjugated linoleicacid mediates protection from experimental inflammatory boweldisease. Gastroenterology 2004;127:777–791.
9. Dubuquoy L, Jansson EA, Deeb S, Rakotobe S, Karoui M, Colom-bel JF, Auwerx J, Pettersson S, Desreumaux P. Impaired expres-sion of peroxisome proliferator-activated receptor gamma in ul-cerative colitis. Gastroenterology 2003;124:1265–1276.
0. Lewis JD, Lichtenstein GR, Stein RB, Deren JJ, Judge TA, Fogt F,Furth EE, Demissie EJ, Hurd LB, Su CG, Keilbaugh SA, Lazar MA,Wu GD. An open-label trial of the PPAR-gamma ligand rosiglita-zone for active ulcerative colitis. Am J Gastroenterol 2001;96:3323–3328.
1. Jiang C, Ting AT, Seed B. PPAR-gamma agonists inhibit produc-tion of monocyte inflammatory cytokines. Nature 1998;391:82–86.
2. Pasceri V, Wu HD, Willerson JT, Yeh ET. Modulation of vascularinflammation in vitro and in vivo by peroxisome proliferator-acti-vated receptor-gamma activators. Circulation 2000;101:235–238.
3. Colville-Nash PR, Qureshi SS, Willis D, Willoughby DA. Inhibitionof inducible nitric oxide synthase by peroxisome proliferator-activated receptor agonists: correlation with induction of hemeoxygenase 1. J Immunol 1998;161:978–984.
4. Kelly D, Campbell JI, King TP, Grant G, Jansson EA, Coutts AG,Pettersson S, Conway S. Commensal anaerobic gut bacteriaattenuate inflammation by regulating nuclear-cytoplasmic shut-tling of PPAR-gamma and RelA. Nat Immunol 2004;5:104–112.
5. Lemberger T, Staels B, Saladin R, Desvergne B, Auwerx J, Wahli
W. Regulation of the peroxisome proliferator-activated receptoralpha gene by glucocorticoids. J Biol Chem 1994;269:24527–24530.
6. Lemberger T, Saladin R, Vazquez M, Assimacopoulos F, Staels B,Desvergne B, Wahli W, Auwerx J. Expression of the peroxisomeproliferator-activated receptor alpha gene is stimulated by stressand follows a diurnal rhythm. J Biol Chem 1996;271:1764–1769.
7. Wu Z, Bucher NL, Farmer SR. Induction of peroxisome prolifera-tor-activated receptor gamma during the conversion of 3T3 fibro-blasts into adipocytes is mediated by C/EBPbeta, C/EBPdelta,and glucocorticoids. Mol Cell Biol 1996;16:4128–4136.
8. Inoue I, Shino K, Noji S, Awata T, Katayama S. Expression ofperoxisome proliferator-activated receptor alpha (PPAR alpha) inprimary cultures of human vascular endothelial cells. BiochemBiophys Res Commun 1998;246:370–374.
9. Daynes RA, Jones DC. Emerging roles of PPARs in inflammationand immunity. Nat Rev Immunol 2002;2:748–759.
0. Kinoshita M, Suzuki Y, Saito Y. Butyrate reduces colonic para-cellular permeability by enhancing PPARgamma activation. Bio-chem Biophys Res Commun 2002;293:827–831.
1. Banan A, Fields JZ, Zhang Y, Keshavarzian A. iNOS upregulationmediates oxidant-induced disruption of F-actin and barrier ofintestinal monolayers. Am J Physiol Gastrointest Liver Physiol2001;280:G1234–G1246.
2. Xu DZ, Lu Q, Deitch EA. Nitric oxide directly impairs intestinalbarrier function. Shock 2002;17:139–145.
3. Han X, Fink MP, Yang R, Delude RL. Increased iNOS activity isessential for intestinal epithelial tight junction dysfunction inendotoxemic mice. Shock 2004;21:261–270.
4. Banan A, Farhadi A, Fields JZ, Zhang LJ, Shaikh M, KeshavarzianA. The delta-isoform of protein kinase C causes inducible nitric-oxide synthase and nitric oxide up-regulation: key mechanism foroxidant-induced carbonylation, nitration, and disassembly of themicrotubule cytoskeleton and hyperpermeability of barrier of in-testinal epithelia. J Pharmacol Exp Ther 2003;305:482–494.
5. Han X, Fink MP, Uchiyama T, Yang R, Delude RL. Increased iNOSactivity is essential for hepatic epithelial tight junction dysfunc-tion in endotoxemic mice. Am J Physiol Gastrointest Liver Physiol2004;286:G126–G136.
6. Santos J, Saunders PR, Hanssen NP, Yang PC, Yates D, Groot JA,Perdue MH. Corticotropin-releasing hormone mimics stress-in-duced colonic epithelial pathophysiology in the rat. Am J Physiol1999;277:G391–G399.
7. Shibata T, Kondo M, Osawa T, Shibata N, Kobayashi M, Uchida K.15-deoxy-delta 12,14-prostaglandin J2. A prostaglandin D2 me-tabolite generated during inflammatory processes. J Biol Chem2002;277:10459–10466.
8. Petrova TV, Akama KT, Van Eldik LJ. Cyclopentenone prostaglan-dins suppress activation of microglia: down-regulation of induc-ible nitric-oxide synthase by 15-deoxy-Delta12,14-prostaglandinJ2. Proc Natl Acad Sci U S A 1999;96:4668–4673.
9. Rossi A, Kapahi P, Natoli G, Takahashi T, Chen Y, Karin M,Santoro MG. Anti-inflammatory cyclopentenone prostaglandinsare direct inhibitors of IkappaB kinase. Nature 2000;403:103–108.
0. Heneka MT, Feinstein DL, Galea E, Gleichmann M, Wullner U,Klockgether T. Peroxisome proliferator-activated receptor gammaagonists protect cerebellar granule cells from cytokine-inducedapoptotic cell death by inhibition of inducible nitric oxide syn-thase. J Neuroimmunol 1999;100:156–168.
1. Li M, Pascual G, Glass CK. Peroxisome proliferator-activatedreceptor gamma-dependent repression of the inducible nitric ox-ide synthase gene. Mol Cell Biol 2000;20:4699–4707.
2. Singh VP, Patil CS, Jain NK, Kulkarni SK. Aggravation of inflam-matory bowel disease by cyclooxygenase-2 inhibitors in rats.
Pharmacology 2004;72:77–84.
7
7
7
7
7
7
7
8
GUM9
eJF
May 2007 PPAR� AND COLONIC DYSFUNCTION 1803
3. Zamuner SR, Warrier N, Buret AG, MacNaughton WK, Wallace JL.Cyclooxygenase 2 mediates post-inflammatory colonic secretoryand barrier dysfunction. Gut 2003;52:1714–1720.
4. Gilroy DW, Colville-Nash PR, Willis D, Chivers J, Paul-Clark MJ,Willoughby DA. Inducible cyclooxygenase may have anti-inflam-matory properties. Nat Med 1999;5:698–701.
5. Bianchi A, Moulin D, Sebillaud S, Koufany M, Galteau MM, NetterP, Terlain B, Jouzeau JY. Contrasting effects of peroxisome-proliferator-activated receptor (PPAR)gamma agonists on mem-brane-associated prostaglandin E2 synthase-1 in IL-1beta-stimu-lated rat chondrocytes: evidence for PPARgamma-independentinhibition by 15-deoxy-Delta12,14prostaglandin J2. Arthritis ResTher 2005;7:R1325–R1337.
6. Schroder O, Yudina Y, Sabirsh A, Zahn N, Haeggstrom JZ, SteinJ. 15-deoxy-Delta12,14-prostaglandin J2 inhibits the expressionof microsomal prostaglandin E synthase type 2 in colon cancercells. J Lipid Res 2006;47:1071–1080.
7. Wira CR, Sandoe CP, Steele MG. Glucocorticoid regulation of thehumoral immune system. I. In vivo effects of dexamethasone onIgA and IgG in serum and at mucosal surfaces. J Immunol 1990;
144:142–146. (8. Guhad FA, Hau J. Salivary IgA as a marker of social stress in rats.Neurosci Lett 1996;216:137–140.
9. Sullivan DA, Underdown BJ, Wira CR. Steroid hormone regulationof free secretory component in the rat uterus. Immunology 1983;49:379–386.
0. Wira CR, Colby EM. Regulation of secretory component by glu-cocorticoids in primary cultures of rat hepatocytes. J Immunol1985;134:1744–1748.
Received May 12, 2006. Accepted January 18, 2007.Address requests for reprints to: Luis Menchén, MD, PhD, Sección de
astroenterología, Servicio de Aparato Digestivo, Hospital Generalniversitario “Gregorio Marañón,” C/ Dr Esquerdo 46, 28007adrid, Spain. e-mail: [email protected]; fax: (34)14265024.Á.P. and J.R.C. contributed equally to this work.Supported by grants from Spanish Ministry of Education and Sci-
nce (DGI SAF 04/27) and Fundación Santander/UCM (PR27/05) (to.C.L.), Asociación Castellana de Aparato Digestivo (ACAD/2004), andundación para la Investigación en Gastroenterología y Hepatología
to L.M.)BA
SIC–
ALI
MEN
TARY
TRA
CT
Copyright © 2022 FDOKUMEN




















