
Mucosal Lymphoid Infiltrate Dominates Colonic Pathological Changes in Murine Experimental...
13
136 • JID 2005:192 (1 July) • Martino et al. MAJOR ARTICLE Mucosal Lymphoid Infiltrate Dominates Colonic Pathological Changes in Murine Experimental Shigellosis Maria Celeste Martino, 1 Giacomo Rossi, 2 Irene Martini, 1 Ivan Tattoli, 1 Damiana Chiavolini, 3 Armelle Phalipon, 4 Philippe J. Sansonetti, 4 and Maria Lina Bernardini 1 1 Dipartimento di Biologia Cellulare e dello Sviluppo, Universita ` La Sapienza, Rome, 2 Facolta ` di Medicina Veterinaria, Universita ` di Camerino, Matelica, and 3 Dipartimento di Microbiologia, Universita ` di Siena, Siena, Italy; 4 Pathoge ´nie Microbienne Mole ´culaire, Institut Pasteur, Paris, France Background. Shigella species are invasive human pathogens that cause acute rectocolitis by triggering a dysregulated inflammatory reaction in the colonic and rectal mucosa. Because mice are naturally resistant to shigellosis, there is no mouse model that mimics human disease. We explore the susceptibility of intestinal flora– depleted mice to shigellosis after intragastric infection with Shigella strains. Methods. Mice given 5 g/L streptomycin as a beverage were infected intragastrically with cfu of either 8 1 10 invasive or noninvasive Shigella strains. Results. We found that invasive Shigella strains persist up to 30 days in feces, whereas the persistence of noninvasive Shigella strains was reduced. Colonization primarily involves the colon and the cecum and, to a lesser extent, the ileum. The hallmark of inflammation in the intestinal tissue is a dramatic expansion of the lymphoid follicles, in which a high apoptotic index is recorded. Conclusions. We provide a murine model in which shigellae are able to reach their natural tissue target: the colon. Moreover, the absence of polymorphonuclear leukocyte recruitment and of epithelial cell lesions reveal some aspects of shigellosis that are usually hidden by the prevalence of this cell population. This novel model may contribute to the identification of new targets for vaccines and therapies. Shigellosis is an acute rectocolitis [1] provoked by the ingestion of as few as 10–100 Shigella colony-forming units [2], and 1150 million cases occur every year [3]. Shigellae invade the colonic mucosa by translocating through M cells of the follicle-associated epithelium that covers lymphoid nodules dispersed on the colo- rectal surface [4]. From there, shigellae penetrate in- testinal epithelial cells (IECs), in which they inject ef- fector proteins via a type III secretion apparatus [5]. These events initiate inflammation through the apop- totic killing of infected macrophages [6], due to the activation of caspase-1, resulting in the parallel release Received 5 October 2004; accepted 11 February 2005; electronically published 31 May 2005. Financial support: European Commission (grant QLK2-1999-00938); Italian Ministero dell’Istruzione, Universita ` e Ricerca (grant PRIN2002). Reprints or correspondence: Dr. Maria Lina Bernardini, Dipartimento di Biologia Cellulare e dello Sviluppo, Universita ` La Sapienza, Via dei Sardi 70, 00185 Rome, Italy ([email protected]). The Journal of Infectious Diseases 2005; 192:136–48 2005 by the Infectious Diseases Society of America. All rights reserved. 0022-1899/2005/19201-0019$15.00 of mature interleukin (IL)–1b [7] and, possibly, a de- creased expression of IL-1 receptor antagonist [8]. In- vaded IECs produce proinflammatory cytokines and chemokines, particularly IL-8, a potent activator of poly- morphonuclear leukocytes (PMNLs) [9, 10], after in- tracellular sensing of a peptidoglycan motif [11] via nucleotide-binding oligomerization domain protein 1 [12]; inflammation thus increases in intensity and ex- tends according to the progression of shigellae in the epithelium. This is an essential feature, because mucosal inflammation disrupts the epithelial barrier and facil- itates further invasion of the epithelium by shigellae [13–15]. PMNLs eventually eliminate invasive shigellae [16] by blocking their escape from the phagocytic vac- uole. In summary, eradication of shigellae at the innate stage of the immune response occurs at the expense of massive inflammatory destruction of the epithelium. Although mice are the ideal animals to use in the ex- perimental study of human infectious diseases, because of their natural resistance to intestinal infection by Shi- gella species, to date, alternative animal systems have by guest on May 23, 2016 http://jid.oxfordjournals.org/ Downloaded from
Transcript of Mucosal Lymphoid Infiltrate Dominates Colonic Pathological Changes in Murine Experimental...

136 • JID 2005:192 (1 July) • Martino et al.
M A J O R A R T I C L E
Mucosal Lymphoid Infiltrate DominatesColonic Pathological Changes in MurineExperimental Shigellosis
Maria Celeste Martino,1 Giacomo Rossi,2 Irene Martini,1 Ivan Tattoli,1 Damiana Chiavolini,3
Armelle Phalipon,4 Philippe J. Sansonetti,4 and Maria Lina Bernardini1
1Dipartimento di Biologia Cellulare e dello Sviluppo, Universita La Sapienza, Rome, 2Facolta di Medicina Veterinaria,Universita di Camerino, Matelica, and 3Dipartimento di Microbiologia, Universita di Siena, Siena, Italy;4Pathogenie Microbienne Moleculaire, Institut Pasteur, Paris, France
Background. Shigella species are invasive human pathogens that cause acute rectocolitis by triggering adysregulated inflammatory reaction in the colonic and rectal mucosa. Because mice are naturally resistant toshigellosis, there is no mouse model that mimics human disease. We explore the susceptibility of intestinal flora–depleted mice to shigellosis after intragastric infection with Shigella strains.
Methods. Mice given 5 g/L streptomycin as a beverage were infected intragastrically with cfu of either81 � 10invasive or noninvasive Shigella strains.
Results. We found that invasive Shigella strains persist up to 30 days in feces, whereas the persistence ofnoninvasive Shigella strains was reduced. Colonization primarily involves the colon and the cecum and, to a lesserextent, the ileum. The hallmark of inflammation in the intestinal tissue is a dramatic expansion of the lymphoidfollicles, in which a high apoptotic index is recorded.
Conclusions. We provide a murine model in which shigellae are able to reach their natural tissue target: thecolon. Moreover, the absence of polymorphonuclear leukocyte recruitment and of epithelial cell lesions revealsome aspects of shigellosis that are usually hidden by the prevalence of this cell population. This novel model maycontribute to the identification of new targets for vaccines and therapies.
Shigellosis is an acute rectocolitis [1] provoked by the
ingestion of as few as 10–100 Shigella colony-forming
units [2], and 1150 million cases occur every year [3].
Shigellae invade the colonic mucosa by translocating
through M cells of the follicle-associated epithelium
that covers lymphoid nodules dispersed on the colo-
rectal surface [4]. From there, shigellae penetrate in-
testinal epithelial cells (IECs), in which they inject ef-
fector proteins via a type III secretion apparatus [5].
These events initiate inflammation through the apop-
totic killing of infected macrophages [6], due to the
activation of caspase-1, resulting in the parallel release
Received 5 October 2004; accepted 11 February 2005; electronically published31 May 2005.
Financial support: European Commission (grant QLK2-1999-00938); Italian Ministerodell’Istruzione, Universita e Ricerca (grant PRIN2002).
Reprints or correspondence: Dr. Maria Lina Bernardini, Dipartimento di BiologiaCellulare e dello Sviluppo, Universita La Sapienza, Via dei Sardi 70, 00185 Rome,Italy ([email protected]).
The Journal of Infectious Diseases 2005; 192:136–48� 2005 by the Infectious Diseases Society of America. All rights reserved.0022-1899/2005/19201-0019$15.00
of mature interleukin (IL)–1b [7] and, possibly, a de-
creased expression of IL-1 receptor antagonist [8]. In-
vaded IECs produce proinflammatory cytokines and
chemokines, particularly IL-8, a potent activator of poly-
morphonuclear leukocytes (PMNLs) [9, 10], after in-
tracellular sensing of a peptidoglycan motif [11] via
nucleotide-binding oligomerization domain protein 1
[12]; inflammation thus increases in intensity and ex-
tends according to the progression of shigellae in the
epithelium. This is an essential feature, because mucosal
inflammation disrupts the epithelial barrier and facil-
itates further invasion of the epithelium by shigellae
[13–15]. PMNLs eventually eliminate invasive shigellae
[16] by blocking their escape from the phagocytic vac-
uole. In summary, eradication of shigellae at the innate
stage of the immune response occurs at the expense of
massive inflammatory destruction of the epithelium.
Although mice are the ideal animals to use in the ex-
perimental study of human infectious diseases, because
of their natural resistance to intestinal infection by Shi-
gella species, to date, alternative animal systems have
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from
A Murine Model to Study Human Shigellosis • JID 2005:192 (1 July) • 137
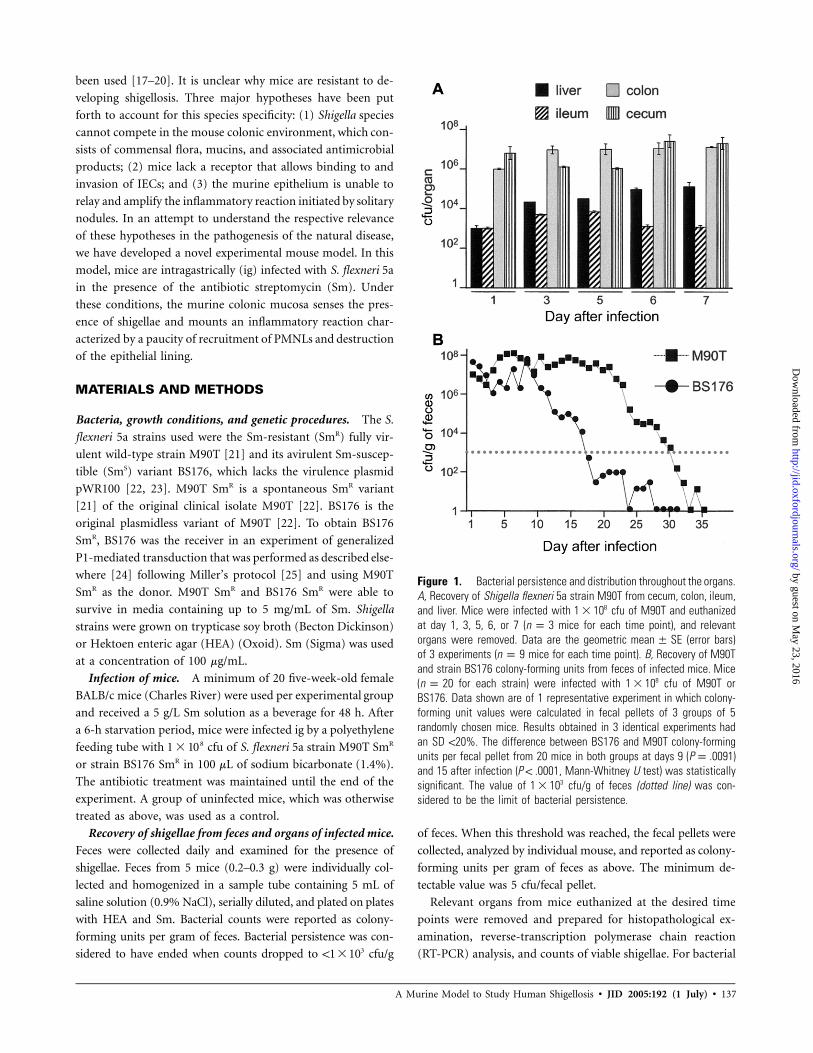
Figure 1. Bacterial persistence and distribution throughout the organs.A, Recovery of Shigella flexneri 5a strain M90T from cecum, colon, ileum,and liver. Mice were infected with cfu of M90T and euthanized81 � 10at day 1, 3, 5, 6, or 7 ( mice for each time point), and relevantn p 3organs were removed. Data are the geometric (error bars)mean � SEof 3 experiments ( mice for each time point). B, Recovery of M90Tn p 9and strain BS176 colony-forming units from feces of infected mice. Mice( for each strain) were infected with cfu of M90T or8n p 20 1 � 10BS176. Data shown are of 1 representative experiment in which colony-forming unit values were calculated in fecal pellets of 3 groups of 5randomly chosen mice. Results obtained in 3 identical experiments hadan SD !20%. The difference between BS176 and M90T colony-formingunits per fecal pellet from 20 mice in both groups at days 9 (P p .0091)and 15 after infection ( , Mann-Whitney U test) was statisticallyP ! .0001significant. The value of cfu/g of feces (dotted line) was con-31 � 10sidered to be the limit of bacterial persistence.
been used [17–20]. It is unclear why mice are resistant to de-
veloping shigellosis. Three major hypotheses have been put
forth to account for this species specificity: (1) Shigella species
cannot compete in the mouse colonic environment, which con-
sists of commensal flora, mucins, and associated antimicrobial
products; (2) mice lack a receptor that allows binding to and
invasion of IECs; and (3) the murine epithelium is unable to
relay and amplify the inflammatory reaction initiated by solitary
nodules. In an attempt to understand the respective relevance
of these hypotheses in the pathogenesis of the natural disease,
we have developed a novel experimental mouse model. In this
model, mice are intragastrically (ig) infected with S. flexneri 5a
in the presence of the antibiotic streptomycin (Sm). Under
these conditions, the murine colonic mucosa senses the pres-
ence of shigellae and mounts an inflammatory reaction char-
acterized by a paucity of recruitment of PMNLs and destruction
of the epithelial lining.
MATERIALS AND METHODS
Bacteria, growth conditions, and genetic procedures. The S.
flexneri 5a strains used were the Sm-resistant (SmR) fully vir-
ulent wild-type strain M90T [21] and its avirulent Sm-suscep-
tible (SmS) variant BS176, which lacks the virulence plasmid
pWR100 [22, 23]. M90T SmR is a spontaneous SmR variant
[21] of the original clinical isolate M90T [22]. BS176 is the
original plasmidless variant of M90T [22]. To obtain BS176
SmR, BS176 was the receiver in an experiment of generalized
P1-mediated transduction that was performed as described else-
where [24] following Miller’s protocol [25] and using M90T
SmR as the donor. M90T SmR and BS176 SmR were able to
survive in media containing up to 5 mg/mL of Sm. Shigella
strains were grown on trypticase soy broth (Becton Dickinson)
or Hektoen enteric agar (HEA) (Oxoid). Sm (Sigma) was used
at a concentration of 100 mg/mL.
Infection of mice. A minimum of 20 five-week-old female
BALB/c mice (Charles River) were used per experimental group
and received a 5 g/L Sm solution as a beverage for 48 h. After
a 6-h starvation period, mice were infected ig by a polyethylene
feeding tube with cfu of S. flexneri 5a strain M90T SmR81 � 10
or strain BS176 SmR in 100 mL of sodium bicarbonate (1.4%).
The antibiotic treatment was maintained until the end of the
experiment. A group of uninfected mice, which was otherwise
treated as above, was used as a control.
Recovery of shigellae from feces and organs of infected mice.
Feces were collected daily and examined for the presence of
shigellae. Feces from 5 mice (0.2–0.3 g) were individually col-
lected and homogenized in a sample tube containing 5 mL of
saline solution (0.9% NaCl), serially diluted, and plated on plates
with HEA and Sm. Bacterial counts were reported as colony-
forming units per gram of feces. Bacterial persistence was con-
sidered to have ended when counts dropped to !1�103 cfu/g
of feces. When this threshold was reached, the fecal pellets were
collected, analyzed by individual mouse, and reported as colony-
forming units per gram of feces as above. The minimum de-
tectable value was 5 cfu/fecal pellet.
Relevant organs from mice euthanized at the desired time
points were removed and prepared for histopathological ex-
amination, reverse-transcription polymerase chain reaction
(RT-PCR) analysis, and counts of viable shigellae. For bacterial
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from

138 • JID 2005:192 (1 July) • Martino et al.
Table 1. Histopathological scores for lesions in intestinal tis-sues from mice infected with Shigella flexneri 5a strain M90T orBS176 and from uninfected control mice.
Group,hours afterinfection,organ
Score for
Transcytosisa/epithelialdamageb
Interstitial areas
NeutrophilscMononuclear
cellsdLymphoidfolliclese
M90T24
Colon 1 2 1 1Cecum 0 0 0 1
72Colon 4 1 3 3Cecum 2 0 1 2
120Colon 3 1 4 3Cecum 2 0 2 2
BS17624
Colon 0 0 0 0Cecum 0 0 0 0
72Colon 1 0 1 1Cecum 0 0 0 0
120Colon 0 0 2 1Cecum 0 0 1 0
Control24
Colon 0 0 0 0Cecum 0 0 0 0
72Colon 0 0 1 0Cecum 0 0 0 0
120Colon 0 0 0 1Cecum 0 0 0 0
NOTE. A total of 6 mice were used in each group. for epithelialP p .0022damage in colon from M90T-infected mice vs. BS176-infected mice at 72 and120 h after infection and for lymphoid follicle activation of colon of M90T-infected mice vs. BS176-infected mice at 72 and 120 h after infection (Mann-Whitney U test).
a Scoring for transcytosis across the epithelium: 0, !10 cells; 1, 10–19 cells;2, 20–49 cells; 3, 50–100 cells; 4, 1100 cells.
b Scoring for epithelial damage: 0, absence of cellular damage; 1, presenceof cytoplasmic vacuolization in some epithelial cells; 2, diffuse cellular vacu-olization; 3, diffuse vacuolization and necrosis/detachment of singular/sporadicepithelial cells; 4, diffuse and severe cellular vacuolization with areas of cellulardetachment (groups of cells with microerosive status).
c Scoring for no. of neutrophils per high-power field: 0, �4 cells; 1, 5–19cells; 2, 20–49 cells; 3, 50–100 cells; 4, 1100 cells.
d Scoring for no. of mononuclear cells per high-power field: 0, �49 cells;1, 50–100 cells; 2, 101–200 cells; 3, 201–600 cells; 4, 1600 cells.
e The mean areas of 10 different randomly selected follicles in gut-associ-ated lymphoid tissues for each sample was measured. Scoring for mean areaof activated lymphoid follicles: 0, none; 1, up to 0.17 mm2; 2, up to 0.32 mm2;3, up to 2.07 mm2.
counts, tissue sections were placed in ice-cold saline solution
(5 mL for liver, 4 mL for colon and ileum, and 2 mL for spleen
and cecum) and homogenized, and serial dilutions were plated
on plates with HEA and Sm. Bacterial counts were reported as
colony-forming units per organ. The minimal detectable values
were 50 cfu/liver, 40 cfu/ileum, 40 cfu/colon, 20 cfu/spleen, and
20 cfu/cecum.
Histopathological analysis. Samples for histopathological
and immunohistochemical (IH) analysis were taken from the
same areas of the organ of interest from 6 randomly chosen
mice in each experimental group. For histopathological ex-
amination, 3-mm sections were deparaffinized, rehydrated, and
stained with hematoxylin-eosin. The following monoclonal an-
tibodies (MAbs) were used: mouse anti–S. flexneri 5a lipopoly-
saccharide (LPS; dimeric IgA; 6 mg/mL) [18], rat anti–murine
CD3+ T lymphocytes, rat anti–murine CD8+ T lymphocytes
(KT15), rat anti–murine macrophages (F4/80 antigen), rat anti–
dendritic/interdigitating cells (Midc-8) (all from Serotec), rat
anti–murine CD5+ B lymphocytes, mouse anti–murine CD21+
B lymphocytes, rat anti–murine CD32+CD16+ NK cells, and
rabbit polyclonal IgG anti–mouse IL-2Ra (CD25) (all from Santa
Cruz Biotechnology). For IH analysis, tissue sections were
treated as described elsewhere [26]. Binding of antibodies was
shown by use of avidin-biotin immunoperoxidase or avidin-
biotin alkaline phosphatase (both from Vector Laboratories)
with biotin-conjugated rabbit anti–rat IgG (Vector Laborato-
ries) and biotinylated goat anti–mouse IgG (AO433; DAKO)
(both at 1:200), respectively. The reactions were visualized with
3-1-diaminobenzidine (DAB; Sigma), Very Intense Purple, or
Vector Blue and Vector Red (all from Vector Laboratories). For
multiple stainings, the first chromogen used was DAB, with or
without nickel addition, the second was VIP, the third was
Vector Blue, and the last was Vector Red, with Meyer’s he-
matoxylin used as a nuclear counterstain.
The number of inflammatory cells and of lymphoid aggre-
gates was assessed at �400 and �100 magnification, respec-
tively, was scored as described by Dixon et al. [27], and was
customized for murine samples as described by Happonen et
al. [28]. The proapoptotic effect induced by S. flexneri was
highlighted through TUNEL colorimetric staining (DeadEnd;
Promega), in accordance with the manufacturer’s instructions.
RNA extraction and RT-PCR. Total RNA from homoge-
nized ileum or colon of 3 mice per experimental group was
extracted using Trizol solution (Invitrogen), in accordance with
the manufacturer’s instructions. RT of total RNA (1 mg) and
cDNA PCR was performed using the Super-Script One-Step
RT-PCR with Platinum Taq (Invitrogen), in accordance with
the manufacturer’s instructions. PCR conditions, primers, and
densitometry are described elsewhere [24].
ELISA. Peripheral blood was obtained from the caudal vein
at 24, 48, 72, and 96 h after infection. IL-6 levels in serum were
determined by solid-phase ELISA (R&D Systems). The absor-
bance was measured at 450 nm, and concentrations were de-
termined by interpolation of a standard calibration curve.
Analysis of data. The exact Mann-Whitney U test was used
for statistical analysis of bacterial counts in feces and the cal-
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from
A Murine Model to Study Human Shigellosis • JID 2005:192 (1 July) • 139
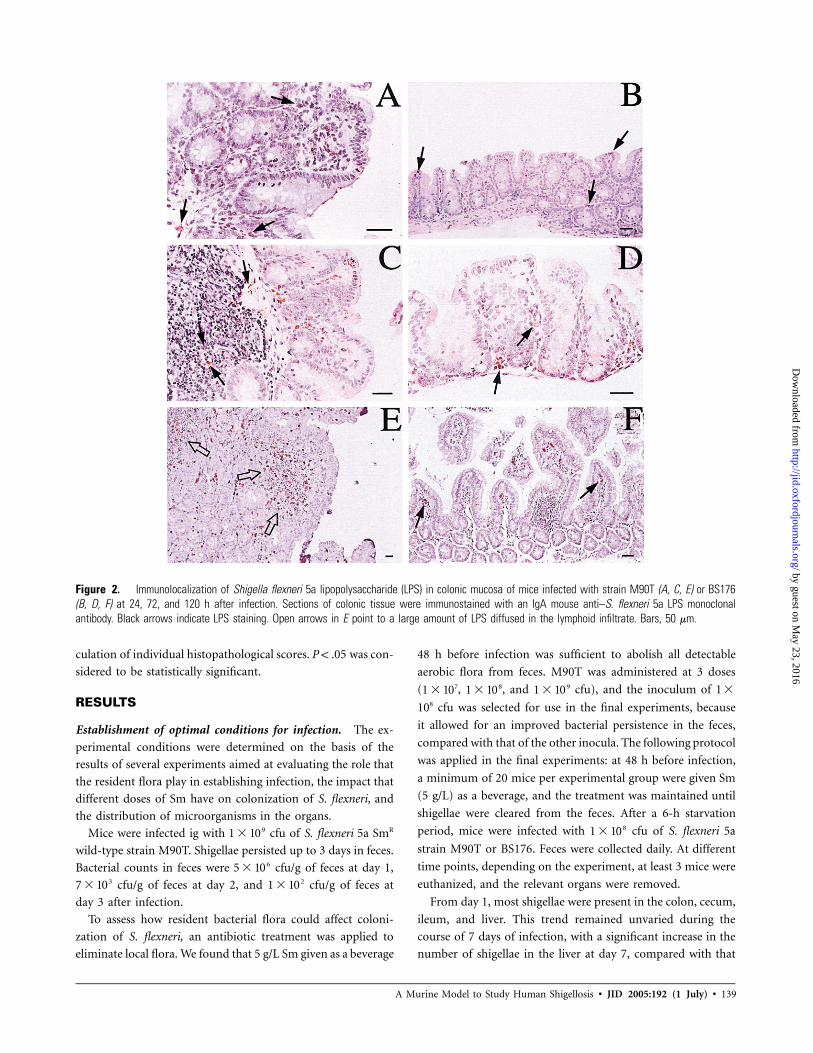
Figure 2. Immunolocalization of Shigella flexneri 5a lipopolysaccharide (LPS) in colonic mucosa of mice infected with strain M90T (A, C, E) or BS176(B, D, F) at 24, 72, and 120 h after infection. Sections of colonic tissue were immunostained with an IgA mouse anti–S. flexneri 5a LPS monoclonalantibody. Black arrows indicate LPS staining. Open arrows in E point to a large amount of LPS diffused in the lymphoid infiltrate. Bars, 50 mm.
culation of individual histopathological scores. was con-P ! .05
sidered to be statistically significant.
RESULTS
Establishment of optimal conditions for infection. The ex-
perimental conditions were determined on the basis of the
results of several experiments aimed at evaluating the role that
the resident flora play in establishing infection, the impact that
different doses of Sm have on colonization of S. flexneri, and
the distribution of microorganisms in the organs.
Mice were infected ig with cfu of S. flexneri 5a SmR91 � 10
wild-type strain M90T. Shigellae persisted up to 3 days in feces.
Bacterial counts in feces were cfu/g of feces at day 1,65 � 10
cfu/g of feces at day 2, and cfu/g of feces at3 27 � 10 1 � 10
day 3 after infection.
To assess how resident bacterial flora could affect coloni-
zation of S. flexneri, an antibiotic treatment was applied to
eliminate local flora. We found that 5 g/L Sm given as a beverage
48 h before infection was sufficient to abolish all detectable
aerobic flora from feces. M90T was administered at 3 doses
( , , and cfu), and the inoculum of 1 �7 8 91 � 10 1 � 10 1 � 10
108 cfu was selected for use in the final experiments, because
it allowed for an improved bacterial persistence in the feces,
compared with that of the other inocula. The following protocol
was applied in the final experiments: at 48 h before infection,
a minimum of 20 mice per experimental group were given Sm
(5 g/L) as a beverage, and the treatment was maintained until
shigellae were cleared from the feces. After a 6-h starvation
period, mice were infected with cfu of S. flexneri 5a81 � 10
strain M90T or BS176. Feces were collected daily. At different
time points, depending on the experiment, at least 3 mice were
euthanized, and the relevant organs were removed.
From day 1, most shigellae were present in the colon, cecum,
ileum, and liver. This trend remained unvaried during the
course of 7 days of infection, with a significant increase in the
number of shigellae in the liver at day 7, compared with that
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from

140
Figu
re3.
Hist
opat
holo
gica
lcha
ract
eriza
tion
ofco
loni
cep
ithel
ium
and
lam
ina
prop
ria(L
P).A
–C,H
emat
oxyl
in-e
osin
(HE)
stai
ning
ofco
loni
cse
ctio
nsfro
mm
ice
infe
cted
with
Shig
ella
flexn
eri5
ast
rain
M90
T(A
)or
BS17
6(B
)and
from
unin
fect
edco
ntro
lmic
e(C
)at
120
haf
ter
infe
ctio
n.In
A,ar
row
head
spo
int
totra
nscy
tosi
sof
poly
mor
phon
ucle
arle
ukoc
ytes
(PM
NLs
)thr
ough
out
the
epith
eliu
m.I
nB,
the
arro
whe
adin
dica
tes
aPM
NL
unde
rth
eLP
.D–F
,HE
stai
ning
ofch
orio
nse
ctio
nsfro
mm
ice
infe
cted
with
M90
T(D
)at
72h
afte
rin
fect
ion
orBS
176
(E)a
t12
0h
afte
rinf
ectio
nan
dfro
mun
infe
cted
cont
rolm
ice
(F).
G–I,
Imm
unoh
isto
chem
ical
char
acte
rizat
ion
ofth
ein
ters
titia
lmon
ocyt
ein
filtra
tein
sect
ions
ofth
eLP
from
mic
ein
fect
edw
ithM
90T
(G)o
rBS
176
(H)a
ndfro
mun
infe
cted
cont
rolm
ice
(I)at
120
haf
teri
nfec
tion.
Avid
in-b
iotin
imm
unop
erox
idas
ela
belin
gof
F4/8
0-po
sitiv
em
acro
phag
esim
mun
osta
ined
with
anIg
Am
ouse
anti–
S.fle
xner
i5a
LPS
mon
oclo
nala
ntib
ody
(bro
wn
and
arro
whe
ads)
and
CD3+
Tly
mph
ocyt
es(b
lue)
.A–C
,Bar
s,25
mm
.D–I
,Bar
s,50
mm
.
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from
141
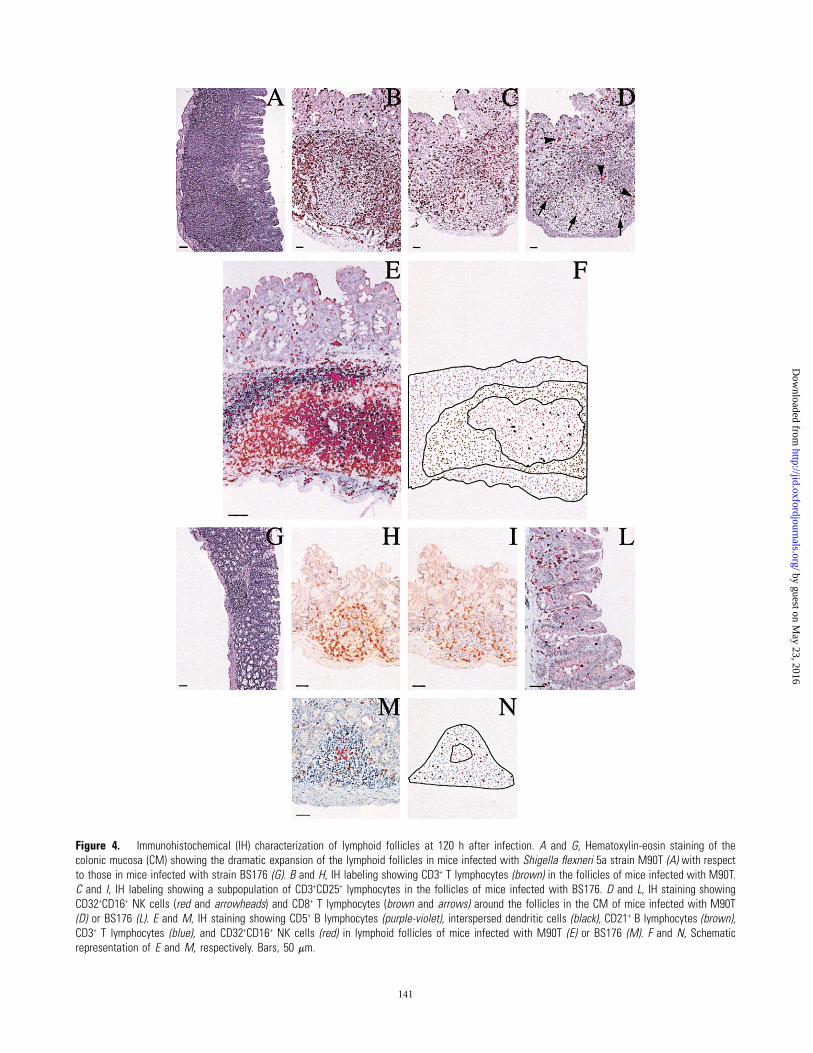
Figure 4. Immunohistochemical (IH) characterization of lymphoid follicles at 120 h after infection. A and G, Hematoxylin-eosin staining of thecolonic mucosa (CM) showing the dramatic expansion of the lymphoid follicles in mice infected with Shigella flexneri 5a strain M90T (A) with respectto those in mice infected with strain BS176 (G). B and H, IH labeling showing CD3+ T lymphocytes (brown) in the follicles of mice infected with M90T.C and I, IH labeling showing a subpopulation of CD3+CD25+ lymphocytes in the follicles of mice infected with BS176. D and L, IH staining showingCD32+CD16+ NK cells (red and arrowheads) and CD8+ T lymphocytes (brown and arrows) around the follicles in the CM of mice infected with M90T(D) or BS176 (L). E and M, IH staining showing CD5+ B lymphocytes (purple-violet), interspersed dendritic cells (black), CD21+ B lymphocytes (brown),CD3+ T lymphocytes (blue), and CD32+CD16+ NK cells (red) in lymphoid follicles of mice infected with M90T (E) or BS176 (M). F and N, Schematicrepresentation of E and M, respectively. Bars, 50 mm.
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from

142 • JID 2005:192 (1 July) • Martino et al.
Figure 5. Apoptosis assessment in the colonic tissue. TUNEL analysis of sections of the colonic mucosa from mice infected with Shigella flexneri5a strain M90T (A) or BS176 (B) and from uninfected control mice (C). Open arrows point to the stained cells. Bars, 50 mm.
Table 2. Apoptotic index in gut-associated lymphoid tissue (GALT) follicles and the lamina propria (LP).
Group
Apoptotic index, mean � SE, %
In GALT folliclesa In the LPb
24 h 72 h 120 h 24 h 72 h 120 h
M90T 0.48 � 0.12 2.20 � 0.65 5.44 � 0.37c 6.60 � 0.32 10.96 � 0.23 9.64 � 0.17BS176 0.53 � 0.21 0.72 � 0.19 0.89 � 0.23 8.87 � 0.15 7.42 � 0.18 8.58 � 0.31Control 0.56 � 0.11 0.41 � 10.09 0.53 � 0.17 6.28 � 0.19 6.44 � 0.21 8.56 � 0.22
a The apoptotic index was calculated as the percentage of TUNEL-positive cells (i.e., the number of TUNEL-positive mono-nuclear cells divided by the total number of lymphoid cells in GALT follicles, � 100) at 24, 72, and 120 hours after infection. A
of cells was counted for each sample.mean � SE 2203� 212b The apoptotic index was calculated as the percentage of TUNEL-positive cells (i.e., the number of TUNEL-positive mono-
nuclear cells divided by the total number of lymphoid cells in diffuse infiltrate in the LP, � 100) at 24, 72, and 120 hours afterinfection. A of cells was counted for each sample.mean � SE 656� 97
c for values at 120 h vs. values at 24 h after infection.P ! .0001
at day 1 (figure 1A). Histopathological analysis confirmed rel-
evant morphological changes at early time points in the liver,
colon, and, to a lesser extent, cecum (data not shown).
Bacterial persistence. To assess whether invasiveness plays
a role in bacterial persistence and distribution, we compared
ig infection with M90T and infection with its noninvasive var-
iant SmS strain BS176. M90T persisted in feces for a mean�
SD of days, whereas BS176 persisted for a mean30.2 � 2.5
�SD of days (figure 1B). After 7 days, the number19.0 � 2.2
of BS176 colony-forming units in feces rapidly decreased,
whereas the number of M90T colony-forming units remained
high. Similarly, during the first 7 days of infection, bacterial
counts in the colon and cecum did not vary for either strain,
with the number of colony-forming units of BS176 being ∼1
log lower than the number of colony-forming units of M90T
in these organs (data not shown).
Histopathological features. Histopathological analysis was
performed on tissue sections removed from the colon and ce-
cum of mice euthanized at 24, 72, and 120 h after infection.
The severity of tissue inflammation was evaluated by recording
qualitative and quantitative alterations of the intestinal mucosa
as reported elsewhere [27, 28]. These alterations were quantified
and recorded as the mean of scores calculated for each lesion,
and these scores are reported in table 1.
In the intestinal tissue of M90T-infected mice, we observed a
relative paucity of PMNLs, compared with the massive numbers
of mononuclear cells and the dramatic expansion of the lym-
phoid follicles of gut-associated lymphoid tissues (GALT). In the
intestinal tissue of BS176-infected mice, only a moderate number
of mononuclear cells were found at 120 h after infection.
Localization of the shigellae in the tissues. The distribu-
tion of shigellae in the gastrointestinal tract was highlighted by
immunostaining for S. flexneri LPS. The cecal and colonic con-
tents of M90T-infected mice were strongly stained for S. flexneri
LPS. However, although a scant positive result for S. flexneri
LPS was found in the cecal mucosa, a stronger presence was
detected in the colonic mucosa (CM). In the CM, at 24 h after
infection, LPS was mainly found in the cytoplasm of macro-
phages located in the central portion of villi beyond the epi-
thelial layer (figure 2A). At 72 h after infection, LPS was de-
tected in the deep chorion (figure 2C), whereas, at 120 h after
infection, a large amount of LPS was found diffused within the
CM and in scattered lymphoid cells of the activated follicles
(figure 2E). In BS176-infected mice, small quantities of LPS
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from
A Murine Model to Study Human Shigellosis • JID 2005:192 (1 July) • 143
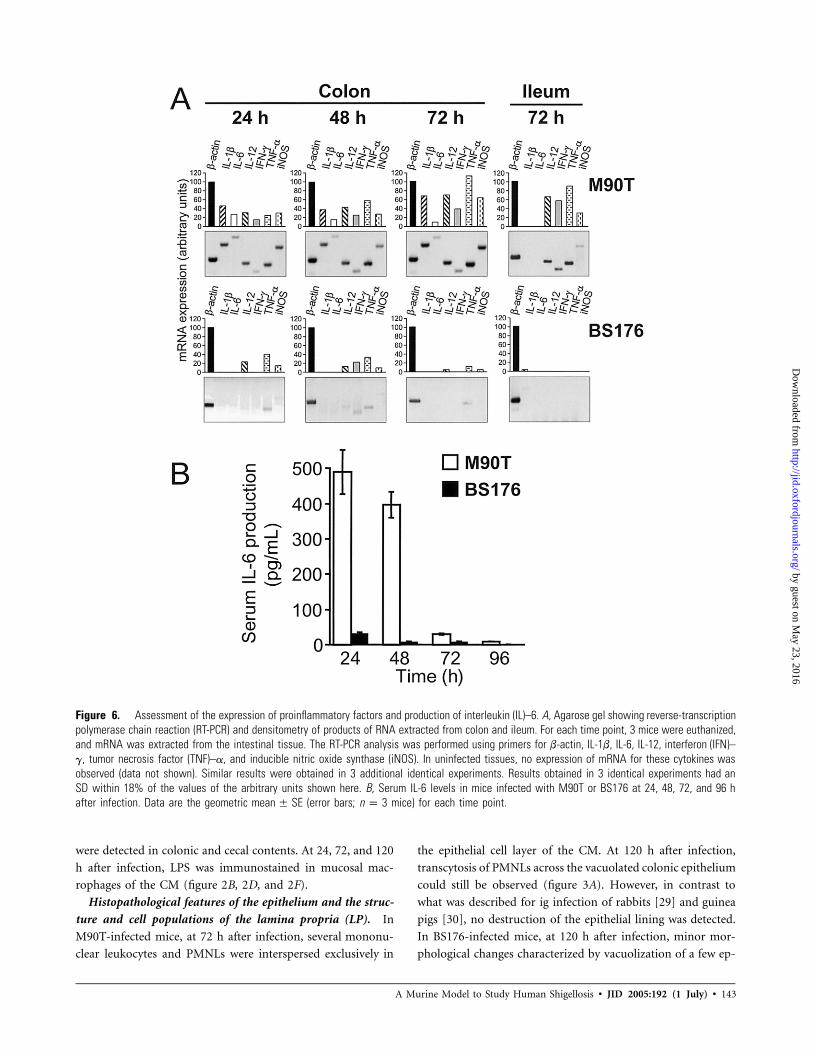
Figure 6. Assessment of the expression of proinflammatory factors and production of interleukin (IL)–6. A, Agarose gel showing reverse-transcriptionpolymerase chain reaction (RT-PCR) and densitometry of products of RNA extracted from colon and ileum. For each time point, 3 mice were euthanized,and mRNA was extracted from the intestinal tissue. The RT-PCR analysis was performed using primers for b-actin, IL-1b, IL-6, IL-12, interferon (IFN)–g, tumor necrosis factor (TNF)–a, and inducible nitric oxide synthase (iNOS). In uninfected tissues, no expression of mRNA for these cytokines wasobserved (data not shown). Similar results were obtained in 3 additional identical experiments. Results obtained in 3 identical experiments had anSD within 18% of the values of the arbitrary units shown here. B, Serum IL-6 levels in mice infected with M90T or BS176 at 24, 48, 72, and 96 hafter infection. Data are the geometric (error bars; mice) for each time point.mean � SE n p 3
were detected in colonic and cecal contents. At 24, 72, and 120
h after infection, LPS was immunostained in mucosal mac-
rophages of the CM (figure 2B, 2D, and 2F).
Histopathological features of the epithelium and the struc-
ture and cell populations of the lamina propria (LP). In
M90T-infected mice, at 72 h after infection, several mononu-
clear leukocytes and PMNLs were interspersed exclusively in
the epithelial cell layer of the CM. At 120 h after infection,
transcytosis of PMNLs across the vacuolated colonic epithelium
could still be observed (figure 3A). However, in contrast to
what was described for ig infection of rabbits [29] and guinea
pigs [30], no destruction of the epithelial lining was detected.
In BS176-infected mice, at 120 h after infection, minor mor-
phological changes characterized by vacuolization of a few ep-
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from

144 • JID 2005:192 (1 July) • Martino et al.
Figure 7. Immunohistochemical (IH) analysis of tissue sections of liver. Hematoxylin-eosin staining of tissue sections of liver from mice infectedwith Shigella flexneri 5a strain M90T (A) or BS176 (B) and from uninfected control mice (C) at 72 h after infection are shown. IH staining shows S.flexneri lipopolysaccharide in the liver from mice infected with M90T (D) or BS176 (E) and from uninfected control mice (F) at 120 h after infection.In A, open arrows indicate a microgranuloma resulting in mononuclear cell aggregates with few neutrophils. In B, open arrows point to small aggregatesof lymphocytes. Bars, 50 mm.
Table 3. Histopathological scores for liver frommice infected with M90T.
Hours afterinfection
Score
Microgranulomasa Degenerationb
24 0 072 1 0120 2 2
NOTE. Liver from both BS176-infected mice and unin-fected control mice had scores of 0 in both parameters ana-lyzed at all time points.
a Microgranulomas were defined as well-circumscribed cellaggregates composed of �5 mononuclear phagocytes. Scor-ing for mean area of granuloma extension: 0, none; 1, up to0.008 mm2; 2, up to 0.042 mm2; 3, up to 0.4 mm2.
b Scoring for no. of hepatocytes showing degeneration (i.e.,cloudy swelling and/or vacuolar degeneration) per high-powerfield: 0, none; 1–19 cells; 2, 20–49 cells; 3, 50–200 cells.
ithelial cells in the absence of transcytosis of inflammatory cells
were observed (figure 3B).
At 24 h after infection, the LP of M90T-infected mice showed
an inflammatory infiltrate characterized by a few PMNLs and a
significant number of mononuclear cells, consisting of CD3+ T
lymphocytes, a few plasma cells, and macrophages (data not
shown). At 72 h after infection, the mononuclear cell–infiltrating
population increased (figure 3D), and a small number of PMNLs
persisted. At 120 h after infection, the colonic villi were enlarged,
and the LP appeared infiltrated by a large number of CD3+ T
lymphocytes (histopathological score, 4) and scattered macro-
phages that were positively immunostained by mouse anti–S.
flexneri 5a LPS (figure 3G). In BS176-infected mice, at 120 h
after infection, the CM showed no significant infiltrate within
the LP (figure 3E). Only a few CD3+ T lymphocyte infiltrates
(histopathological score, 1) in association with a moderate pres-
ence (histopathological score, 2) of interspersed LPS-positive
macrophages (figure 3H) were observed. No alteration in the
epithelium (figure 3C), no cellular infiltrate (figure 3F), and a
few or no CD3+ T lymphocytes were observed in the corre-
sponding tissue from the uninfected control mice (figure 3I).
Characterization of the expanded follicles in the CM. The
main sign of the presence of shigellae in the colonic tissue was
the dramatic expansion of the lymphoid follicles. At 24 h after
infection, we observed the presence of lymphoid aggregates in
the CM of M90T-infected mice. The size of these well-structured
follicles reached a peak at 120 h after infection (figure 4A). Their
mantle area was covered by CD3+ T lymphocytes, primarily rep-
resented by CD3+CD25+ lymphocytes (figure 4B and 4C) and by
many CD3+CD8+ lymphocytes (figure 4D) interspersed with a
large number of CD32+CD16+ NK cells. The central area con-
tained large arachnid-shaped dendritic cells (DCs) interspersed
in a large population of CD5+ B lymphocytes and a few CD3+
T lymphocytes. An area of CD21+ B lymphocytes was immu-
nostained around this central portion. These well-structured fol-
licles, localized in the deep chorion, were consistently accom-
panied by CD32+CD16+ NK cells (figure 4E and 4F) and by a
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from

A Murine Model to Study Human Shigellosis • JID 2005:192 (1 July) • 145
Figure 8. Schematic representation of different steps of Shigella flexneri colonization of the gut mucosa and the liver. IFN, interferon; IL, interleukin;LPS, lipopolysaccharide; TNF, tumor necrosis factor.
mixed population of CD3+ T lymphocytes that formed a diffuse,
superficial, and subepithelial infiltrate, as described above. In the
CM of BS176-infected mice, there were small to moderate lym-
phoid aggregates (figure 4G) not organized as a structured follicle.
As was observed in M90T-infected mice, at 120 h after infection,
a certain number of CD3+CD25+ lymphocytes covered the mantle
area of the lymphoid aggregates (figure 4H and 4I). At this time
point, CD3+CD8+ lymphocytes constituted the prevalent lym-
phocyte population, whereas few or no CD32+CD16+ NK cells
were detected (figure 4L). The lymphoid aggregates mainly re-
sulted in CD3+ T, CD21+ B, and CD5+ B lymphocytes (figure
4M and 4N).
Apoptosis of infected cells in the lymphoid follicles of the LM.
The amount and the localization of apoptotic cells in the CM
was assessed by a TUNEL analysis (figure 5 and table 2). In the
M90T-infected mice, from 24 to 120 h after infection, an in-
creasing number of TUNEL-positive cells was present in the
activated GALT (figure 5A). In contrast, in the BS176-infected
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from

146 • JID 2005:192 (1 July) • Martino et al.
mice, a few TUNEL-positive lymphoid cells were detected (figure
5B). In uninfected control mice, a few TUNEL-positive cells were
observed in the rare solitary lymphoid follicles of the CM (figure
5C), whereas a few apoptotic cells were counted in unstimulated
lymphoid cells of the GALT (table 2).
Expression of proinflammatory cytokines in colonic tissue
at early time points of infection and serum IL-6 production.
We assessed the expression of IL-1b, IL-6, IL-12, interferon
(IFN)–g, tumor necrosis factor (TNF)–a, and inducible nitric
oxide synthase (iNOS) mRNAs by RT-PCR in the colon and
ileum of mice infected with M90T or BS176 and of uninfected
control mice at 24, 48, and 72 h after infection (figure 6A). In
the ileum, a considerable expression of mRNA for IFN-g, TNF-
a, and IL-12 was seen exclusively in M90T-infected mice only
at 72 h after infection.
At 24 h after infection, in M90T-infected mice, a measurable
expression of all these cytokines was observed in colonic tissues,
whereas, in BS176-infected mice, only low levels of IL-12, TNF-
a, and iNOS were detected. In M90T-infected mice, this trend
remained unchanged, with a progressive increase in the expres-
sion of mRNA for these cytokines—with the exception of mRNA
for IL-6, whose low expression slightly decreased from that ob-
served at 48 h after infection—until 72 h after infection. In
BS176-infected mice, at 48 h after infection, weak expression of
mRNA for IL-12, TNF-a, IFN-g, and iNOS was present, whereas,
at 72 h after infection, no or minimal expression of mRNA for
these cytokines was observed (figure 6A). In M90T-infected mice,
serum IL-6 production was high at 24 h after infection and then
disappeared at 72 h after infection (figure 6B).
Damage by shigellae and localization in the liver. In M90T-
infected mice, at 120 h after infection, the liver showed hepatic
cell death and well-defined hepatic microgranulomas with mono-
nuclear cell aggregates consisting of epithelioid and lymphoid
cells (figure 7A). LPS was detected within the cytoplasm of many
macrophages located in some central areas of these microgran-
ulomatous lesions and in Kupffer cells (figure 7D). In BS176-
infected mice, the liver did not show significant morphological
changes, and little or no infiltration of mononuclear cells was
observed (figure 7B). A positive response to LPS was detected
only in Kupffer cells, in the absence of significant lesions (figure
7E). The lesions are scored in table 3.
DISCUSSION
In this study, we addressed 3 key hypotheses about the inability
of mice to develop shigellosis. First, we examined whether the
endogenous flora could protect the intestinal tissue from Shi-
gella infection. We found that treatment of mice with Sm is a
prerequisite for colonization of the intestinal tract by shigellae,
which is in accordance with the role played by the intestinal
flora in preventing oral infection with various enteropathogens,
such as Salmonella species [31–33]. Then, we investigated the
ability of Shigella species to invade the intestinal tissue. As in
humans, the colon and eventually the cecum of the Sm-treated
mice were the main targets for the invasion of a wild-type
Shigella strain. However, in contrast to human shigellosis, no
lesions or only a few abscesses were observed in the mice, de-
spite the presence of a high number of shigellae. Finally, we
considered the hypothesis that the murine epithelium may be
unable to respond to intracellular shigellae by recruiting an
inflammatory infiltrate. Interestingly, the presence of a mono-
nuclear infiltrate and a dramatic expansion of lymphoid folli-
cles dispersed along the CM and the cecal mucosa were the
main features of the infected intestine, thus demonstrating that
the intestinal epithelium is able to sense shigellae and to mount
an inflammatory reaction. However, in contrast to the situation
in human shigellosis, and in accordance with the paucity of
lesions observed, a moderate number of PMNLs was observed
in the mice.
In the rabbit ligated ileal loop model of Shigella infection,
inhibition of IL-8 prevents the transmigration of PMNLs, thereby
reducing the number of tissue lesions at the expense of bacterial
dissemination in deeper layers of the CM [9]. In mice, the
delivery of recombinant human IL-8 together with invasive
Shigella strains through intraluminal instillation induces mu-
cosal lesions similar to those seen in human disease [20]. In
the present study, at 24 h after infection, shigellae were isolated
from the liver, in which they induced microgranulomas, as is
schematically shown in figure 8. These findings suggest that the
absence of a murine equivalent of human IL-8 [34] might con-
tribute to the prevention of the deleterious consequences of
inflammation and favor the septic dissemination of shigellae
into the liver. This confirms the results of our study showing
that, in mice infected intravenously, shigellae reach the liver,
where they induce microgranulomatous lesions [26]: again, the
absence of PMNLs is a feature of these lesions.
In the CM of Sm-treated mice, several cell populations par-
ticipate in the mononuclear infiltrate. CD32+CD16+ NK cells are
localized superficially and subepithelially in the CM of M90T-
infected mice. NK cells producing IFN-g play an essential role
in the resistance in mice after primary Shigella intranasal infection
[35], whereas, in humans, high levels of mRNA for IFN-g are
found in the LP only during late-phase shigellosis [36].
In M90T-infected mice, only mature DCs migrating to sec-
ondary lymphoid organs, such as the expanded follicles, were
observed. In M90T-infected mice, apoptosis of macrophages
and the subsequent release of shigellae or their material in the
expanded follicles might contribute to the maturation of DCs.
Mature DCs can produce IL-12 and stimulate NK cells again
[37], thus amplifying their activation and proliferation, the
release of cytokines such as TNF-a and IFN-g, and their bac-
teriolytic activity [38]. In accordance with this hypothesis, mRNA
for both IL-12 and IFN-g was detected only in colonic tissues
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from

A Murine Model to Study Human Shigellosis • JID 2005:192 (1 July) • 147
of M90T-infected mice. In the rectal mucosa of humans with
shigellosis [39], a large expansion of CD3+ T lymphocytes is
observed. In mice, the large CD3+ T lymphocyte population
primarily consists of a subset of CD3+CD25+ lymphocytes.
CD4+CD25+ lymphocytes are a subset of naturally activated
lymphocytes, CD45RBlow, which control inflammatory re-
sponses to commensal bacteria [40] by acting as a negative
regulator of the immune response [41]. This cell population
might play an important role in controlling the magnitude and
the deleterious consequences of the murine intestinal tissue
response to shigellae. Whether CD3+CD25+ lymphocytes play
a role during natural shigellosis is still unknown. In the present
study, a large number of CD3+CD8+ lymphocytes was present
in the LP, particularly in the cortical areas of the expanded
follicles of M90T-infected mice. This cell population is the T
cell subset predominantly involved in the inflammation [39]
of human rectal mucosa. Finally, in the present study, CD5+ B
lymphocytes were diffusely distributed in the expanded central
areas of follicles. In mice, CD5+ B lymphocytes predominate
in the gut mucosa [42] and play a crucial role in the eradication
of pathogens by stimulating the innate immune system via the
classic pathway of complement activation [43].
In conclusion, it must be stressed that ig infection of Sm-
treated mice is, to our knowledge, the first model of shigellosis
in which the pathogens spontaneously interacted with the co-
lonic tissue that represents the natural target of their infection.
Furthermore, in accordance with the results of studies of in-
tragastric infection of the macaque Macaca mulatta, the dis-
tribution of bacteria in this tissue is essentially concentrated
around lymph nodes [4].
The study of infected murine colonic tissue in which PMNLs
are poorly recruited helps us to understand the other cell pop-
ulations involved in inflammation, their interactions, and their
interplay with bacteria. This may facilitate the search for new
vaccines and therapeutic approaches to shigellosis.
Acknowledgments
We thank Giorgina Levi and Stanley Stewart, for help in manuscriptpreparation, and Giuseppe Bertini, Piero Piccoli, and Giancarlo Cortese,for care of the mice.
References
1. Mathan VI, Mathan MM. Intestinal manifestations of invasive diar-rheas and their diagnosis. Rev Infect Dis 1991; 13(Suppl 4):S311–3.
2. DuPont HL, Levine MM, Hornick RB, Formal SB. Inoculum size inshigellosis and implications for expected mode of transmission. J InfectDis 1989; 159:1126–8.
3. Kotloff KL, Winickoff JP, Ivanoff B, et al. Global burden of Shigellainfections: implications for vaccine development and implementationof control strategies. Bull World Health Organ 1999; 77:651–6.
4. Sansonetti PJ, Arondel J, Fontaine A, d’Hauteville H, Bernardini ML.ompB (osmo-regulation) and icsA (cell-to-cell spread) mutants of Shi-
gella flexneri: vaccine candidates and probes to study the pathogenesisof shigellosis. Vaccine 1991; 9:416–22.
5. Buttner D, Bonas U. Port of entry: the type III secretion translocon.Trends Microbiol 2002; 10:186–92.
6. Zychlinsky A, Prevost MC, Sansonetti PJ. Shigella flexneri induces ap-optosis in infected macrophages. Nature 1992; 358:167–9.
7. Zychlinsky A, Fitting C, Cavaillon JM, Sansonetti PJ. Interleukin 1 isreleased by murine macrophages during apoptosis induced by Shigellaflexneri. J Clin Invest 1994; 94:1328–32.
8. Arondel J, Singer M, Matsukawa A, Zychlinsky A, Sansonetti PJ. In-creased interleukin 1 (IL-1) and imbalance between IL-1 and IL-1receptor antagonist during acute inflammation in experimental shi-gellosis. Infect Immun 1999; 67:6056–66.
9. Sansonetti PJ, Arondel J, Huerre M, Harada A, Matsushima K. Inter-leukin-8 controls bacterial transepithelial translocation at the cost ofepithelial destruction in experimental shigellosis. Infect Immun 1999;67:1471–80.
10. Philpott DJ, Yamaoka S, Israel A, Sansonetti PJ. Invasive Shigella flexneriactivates NF-kappaB through a lipopolysaccharide-dependent innateintracellular response and leads to IL-8 expression in epithelial cells.J Immunol 2000; 165:903–14.
11. Girardin SE, Boneca IG, Carneiro LA, et al. Nod1 detects a uniquemuropeptide from gram negative bacterial peptidoglycan. Science 2003;300:1584–7.
12. Girardin SE, Tournebize R, Mavris M, et al. CARD4/Nod1 mediatesNF-kappaB and JNK activation by invasive Shigella flexneri. EMBORep 2001; 2:736–42.
13. Perdomo OJ, Cavaillon JM, Huerre M, Ohayon H, Gounon P, San-sonetti PJ. Acute inflammation causes epithelial invasion and mucosaldestruction in experimental shigellosis. J Exp Med 1994; 180:1307–19.
14. Sansonetti PJ, Arondel J, Cavaillon JM, Huerre M. Role of IL-1 in thepathogenesis of experimental shigellosis. J Clin Invest 1995; 96:884–92.
15. Sansonetti PJ, Phalipon A, Arondel J, et al. Caspase-1 activation of IL-1beta and IL-18 are essential for Shigella flexneri-induced inflammation.Immunity 2000; 12:581–90.
16. Weinrauch Y, Drujan D, Shapiro SD, Weiss J, Zychlinsky A. Neutrophilelastase targets virulence factors of enterobacteria. Nature 2002; 417:91–4.
17. Voino-Yasenetsky MV, Voino-Yasenetskaya MK. Experimental pneu-monia caused by bacteria of Shigella group. Acta Morphol Acad SciHung 1962; 11:439–54.
18. Phalipon A, Kaufmann M, Michetti P, et al. Monoclonal immuno-globulin A antibody directed against serotype-specific epitope of Shi-gella flexneri lipopolysaccharide protects against murine experimentalshigellosis. J Exp Med 1995; 182:769–78.
19. Fernandez MI, Thuizat A, Pedron T, Neutra M, Phalipon A, SansonettiPJ. A newborn mouse model for the study of intestinal pathogenesisof shigellosis. Cell Microbiol 2003; 5:481–91.
20. Singer M, Sansonetti PJ. IL-8 is a key chemokine regulating neutrophilrecruitment in a new mouse model of Shigella-induced colitis. J Im-munol 2004; 173:4197–206.
21. Allaoui A, Mounier J, Prevost C, Sansonetti PJ, Parsot C. icsB: a Shigellaflexneri virulence gene necessary for the lysis of protrusions duringintercellular spread. Mol Microbiol 1992; 6:1605–16.
22. Sansonetti PJ, Kopecko DJ, Formal SB. Involvement of a plasmid inthe invasive ability of Shigella flexneri. Infect Immun 1982; 35:852–60.
23. Buchrieser C, Glaser P, Rusniok C, et al. The virulence plasmid pWR100and the repertoire of proteins secreted by the type III secretion ap-paratus of Shigella flexneri. Mol Microbiol 2000; 38:760–71.
24. Cersini A, Martino MC, Martini I, Rossi G, Bernardini ML. Analysisof virulence and inflammatory potential of Shigella flexneri purinebiosynthesis mutants. Infect Immun 2003; 71:7002–13.
25. Miller JH. A short course in bacterial genetics. Cold Spring Harbor,NY: Cold Spring Harbor Laboratory Press, 1992.
26. Martino MC, Rossi G, Tattoli I, et al. Intravenous infection of virulentshigellae causes fulminant hepatitis in mice. Cell Microbiol 2005; 7:115–27.
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from

148 • JID 2005:192 (1 July) • Martino et al.
27. Dixon MF, Genta RM, Yardley JF, Correa P. Classification and gradingof gastritis: the updated Sydney System. Am J Surg Pathol 1996; 20:1161–81.
28. Happonen I, Linden J, Saari S, et al. Detection and effects of helico-bacters in healthy dogs and dogs with signs of gastritis. J Am Vet MedAssoc 1998; 213:1767–74.
29. Etheridge ME, Hoque AT, Sack DA. Pathologic study of a rabbit modelfor shigellosis. Lab Anim Sci 1996; 46:61–6.
30. Bernardini ML, Arondel J, Martini I, Aidara A, Sansonetti PJ. Param-eters underlying successful protection with live attenuated mutants inexperimental shigellosis. Infect Immun 2001; 69:1072–83.
31. Brown KJ, Tannock GW, Eyres RA, Elliott RB, Lines R. Colonizationof Salmonella typhimurium and Shigella flexneri III of the gastrointes-tinal tract of mice treated with beta-2-thienylalanine and streptomycin.Antonie Van Leeuwenhoek 1979; 45:531–46.
32. Que JU, Casey SW, Hentges DJ. Factors responsible for increased sus-ceptibility of mice to intestinal colonization after treatment with strep-tomycin. Infect Immun 1986; 53:116–23.
33. Barthel M, Hapfelmeier S, Quintanilla-Martinez L, et al. Pretreatmentof mice with streptomycin provides a Salmonella enterica serovar Ty-phimurium colitis model that allows analysis of both pathogen andhost. Infect Immun 2003; 71:2839–58.
34. Zhang Z, Jin L, Champion G, Seydel KB, Stanley SL Jr. Shigella infection
in SCID-HU-INT mice: role for neutrophils in containing bacterialdissemination in human intestine. Infect Immun 2001; 69:3240–7.
35. Way SS, Borczuk AC, Goldberg MB. An essential role for gammainterferon in innate resistance to Shigella flexneri infection. Infect Im-mun 1998; 66:1342–8.
36. Raqib R, Ljungdahl A, Lindberg AA, Andersson U, Andersson J. Localentrapment of interferon gamma in the recovery from Shigella dysen-teriae type 1 infection. Gut 1996; 38:328–36.
37. Moretta A. Natural killer cells and dendritic cells: rendezvous in abusedtissues. Nat Rev Immunol 2002; 2:957–60.
38. Gerosa F, Baldani-Guerra B, Nisii C, Marchesini V, Carra G, TrinchieriG. Reciprocal activating interaction between natural killer cells anddendritic cells. J Exp Med 2002; 195:327–33.
39. Islam D, Veress B, Bardhan PK, Lindberg AA, Christensson B. In situcharacterization of inflammatory responses in the rectal mucosae ofpatients with shigellosis. Infect Immun 1997; 65:739–49.
40. Singh B, Read S, Asseman C, et al. Control of intestinal inflammationby regulatory T cells. Immunol Rev 2001; 182:190–200.
41. Shevach EM. CD4+CD25+ suppressor T cells: more questions thananswers. Nat Rev Immunol 2002; 2:389–400.
42. Kantor AB, Herzemberg LA. Origin of murine B-cell lineages. AnnuRev Immunol 1993; 11:501–38.
43. Carroll MC, Prodeus AP. Linkages of innate and adaptive immunity.Curr Opin Immunol 1998; 10:36–40.
by guest on May 23, 2016
http://jid.oxfordjournals.org/D
ownloaded from














