
The RGD-dependent and the Hep II binding domains of fibronectin govern the adhesive behaviors of...
10
ELSEVIER Mechanisms of Development 56 (1996) 83-92 The RGD-dependent and the Hep II binding domains of fibronectin govern the adhesive behaviors of amphibian embryonic cells Dominique Alfandaria, Joe Ramosb, Lea Claviliera, Douglas W. DeSimoneb, Thierry Darribh-e%* aUniversite’ P. et M. Curie, U.R.A. CNRS 113.5, Laboratoire de Biologie Moltkulaire et Cellulaire du Dbeloppement, Equipe Adhesion et Migration Cellulaires, 9 Quai Saint-Bernard, 75005 Paris, France bDepartment of Cell Biology, University of Virginia, Box 439, Health Sciences Center, Charlottesville, VA22908, USA Received 2 January 1996; revision received 30 January 1996; accepted 5 February 1996 Abstract In early amphibian development, interactions between fibronectin and both ectoderm and mesoderm cells are critical in the progres- sion of gastrulation movements. In the Pleurodeles Walt1 embryo, it has been established that ectoderm cells of the animal hemisphere organize a fibrillar-extracellular matrix containing fibronectin. Mesoderm cells migrate along the blastocoel roof using these fibronectin fibrils as substratum. Fibronectin is an adhesive glycoprotein which possesses multiple cell-binding domains. From previous studies, it is clear that amphibian ectoderm and mesoderm cells interact with fibronectin in an RGD-dependent manner, whereas the contributions of RGD-independent domains in the adhesive behaviors of gastrula cells has not been defined. To study this question, we have used bacterially expressed Pleurodeles waltl fibronectin-fusion proteins. The approach consisted of in vitro adhesion assays with either iso- lated cells or tissue fragments of embryos dissected at the onset of gastrulation. Tissues were obtained from regions of the embryo which represent presumptive ectoderm cells or from the dorsal-marginal zone which contains cells of the presumptive cephalic, chordal and somitic mesoderm. The results show that both the RGD-dependent and the Hep II domains of fibronectin mediate attachment and spreading of isolated cells. Both regions cooperate to control the proper expansion of a sheet of dorsal mesoderm cells. The Hep II do- main promotes the migration of cells ahead of the mesoderm-cell sheet. Keywords: Fibronectin; Adhesion; Migration; Mesoderm cells; Amphibian embryos 1. Introduction Fibronectin is a dimeric glycoprotein of high molecu- lar mass formed of two subunits joined at the carboxyl terminus by two disulfide bonds. Each subunit consists of three repeated motifs known as type I, II, and III. The repeats are assembled into domains, which have distinct binding activities toward collagen, thrombospondin, fi- brin, heparan sulfate and cell surface receptors. It has been shown that cell adhesion occurs in an Arg-Gly-Asp (RGD)-dependent and -independent manner involving multiple cell recognition sites along the molecule (Yamada, 1991). The RGD sequence, located in the 10th type III module, promotes the adhesion of a number of cell types (Pierschbacher and Ruoslahti, 1984; Yamada and Kennedy, 1984). The RGD sequence is reported to * Corresponding author. Tel.: +33 1 44273690; fax: +33 1 44273445; e-mail: [email protected]. interact with cells via integrin receptors that include a&, and the %& %&3~ %k &$3r %P6? c+Pl, and a8Pl receptors. Two additional sites in the 8th and 9th type III repeats are required for full activity of the RGD sequence and are termed the synergistic sites (Aota et al., 1994; Bowditch et al., 1994). Of these two sites, the synergy site in the 9th type III repeat, along with the RGD sequence, promotes asPI dependent-cell adhesion. Two RGD- independent cell binding domains have been identified in the alternatively spliced IIICS region of some fibronectins (type III connecting segment, also called the V region). These adhesion sites are named CS-1 and CS-5. Their minimal active sequences are, respectively, Leu-Asp-Val (LDV) and Arg-Glu-Asp-Val (REDV, for human fi- bronectin) or Arg-Gly-Asp-Val (RGDV, for bovine and rat fibronectins) (Humphries et al., 1986). Both are rec- ognized by integrins a& or a& which are expressed by a number of cell types including lymphocytes, monocytes and neural crest cells (Wayner et al., 1989, 1992). An- 09254773/96/$15.00 0 1996 Elsevier Science Ireland Ltd. All rights reserved PII: SO925-4773(96)00513-8
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of The RGD-dependent and the Hep II binding domains of fibronectin govern the adhesive behaviors of...

ELSEVIER Mechanisms of Development 56 (1996) 83-92
The RGD-dependent and the Hep II binding domains of fibronectin govern the adhesive behaviors of amphibian embryonic cells
Dominique Alfandaria, Joe Ramosb, Lea Claviliera, Douglas W. DeSimoneb, Thierry Darribh-e%*
aUniversite’ P. et M. Curie, U.R.A. CNRS 113.5, Laboratoire de Biologie Moltkulaire et Cellulaire du Dbeloppement,
Equipe Adhesion et Migration Cellulaires, 9 Quai Saint-Bernard, 75005 Paris, France
bDepartment of Cell Biology, University of Virginia, Box 439, Health Sciences Center, Charlottesville, VA 22908, USA
Received 2 January 1996; revision received 30 January 1996; accepted 5 February 1996
Abstract
In early amphibian development, interactions between fibronectin and both ectoderm and mesoderm cells are critical in the progres- sion of gastrulation movements. In the Pleurodeles Walt1 embryo, it has been established that ectoderm cells of the animal hemisphere organize a fibrillar-extracellular matrix containing fibronectin. Mesoderm cells migrate along the blastocoel roof using these fibronectin fibrils as substratum. Fibronectin is an adhesive glycoprotein which possesses multiple cell-binding domains. From previous studies, it is clear that amphibian ectoderm and mesoderm cells interact with fibronectin in an RGD-dependent manner, whereas the contributions of RGD-independent domains in the adhesive behaviors of gastrula cells has not been defined. To study this question, we have used bacterially expressed Pleurodeles waltl fibronectin-fusion proteins. The approach consisted of in vitro adhesion assays with either iso- lated cells or tissue fragments of embryos dissected at the onset of gastrulation. Tissues were obtained from regions of the embryo which represent presumptive ectoderm cells or from the dorsal-marginal zone which contains cells of the presumptive cephalic, chordal and somitic mesoderm. The results show that both the RGD-dependent and the Hep II domains of fibronectin mediate attachment and spreading of isolated cells. Both regions cooperate to control the proper expansion of a sheet of dorsal mesoderm cells. The Hep II do- main promotes the migration of cells ahead of the mesoderm-cell sheet.
Keywords: Fibronectin; Adhesion; Migration; Mesoderm cells; Amphibian embryos
1. Introduction
Fibronectin is a dimeric glycoprotein of high molecu- lar mass formed of two subunits joined at the carboxyl terminus by two disulfide bonds. Each subunit consists of three repeated motifs known as type I, II, and III. The repeats are assembled into domains, which have distinct binding activities toward collagen, thrombospondin, fi- brin, heparan sulfate and cell surface receptors. It has been shown that cell adhesion occurs in an Arg-Gly-Asp (RGD)-dependent and -independent manner involving multiple cell recognition sites along the molecule (Yamada, 1991). The RGD sequence, located in the 10th type III module, promotes the adhesion of a number of cell types (Pierschbacher and Ruoslahti, 1984; Yamada and Kennedy, 1984). The RGD sequence is reported to
* Corresponding author. Tel.: +33 1 44273690; fax: +33 1 44273445; e-mail: [email protected].
interact with cells via integrin receptors that include a&,
and the %& %&3~ %k &$3r %P6? c+Pl, and a8Pl receptors. Two additional sites in the 8th and 9th type III repeats are required for full activity of the RGD sequence and are termed the synergistic sites (Aota et al., 1994; Bowditch et al., 1994). Of these two sites, the synergy site in the 9th type III repeat, along with the RGD sequence, promotes asPI dependent-cell adhesion. Two RGD- independent cell binding domains have been identified in the alternatively spliced IIICS region of some fibronectins (type III connecting segment, also called the V region). These adhesion sites are named CS-1 and CS-5. Their minimal active sequences are, respectively, Leu-Asp-Val (LDV) and Arg-Glu-Asp-Val (REDV, for human fi- bronectin) or Arg-Gly-Asp-Val (RGDV, for bovine and rat fibronectins) (Humphries et al., 1986). Both are rec- ognized by integrins a& or a& which are expressed by a number of cell types including lymphocytes, monocytes and neural crest cells (Wayner et al., 1989, 1992). An-
09254773/96/$15.00 0 1996 Elsevier Science Ireland Ltd. All rights reserved PII: SO925-4773(96)00513-8

84 D. Alfandari et al. /Mechanisms of Development 56 (1996) 83-92
other RGD-independent binding domain, the Hep II do- main is located in the carboxyl-terminal region of each fibronectin monomer. It possesses a cell-binding activity and an affinity for both heparan sulfate and sulfated glycosaminoglycans (Haugen et al., 1990; Fujita et al., 1995). In its C-terminal end, two sites called FN-C/HI and
FN-C/HI1 promote attachment and spreading of human melanoma and neuroblastema cells (Mould and Hum- phries, 1991). The IDAPS sequence of the type III-14 repeat has also been shown to support cell attachment via the integrin a& (Mould and Humphries, 1991). Finally, recent studies indicate that another RGD- independent domain is able to promote cell attachment. It corresponds to the spliced domain of fibronectin termed
EIIIA (or EDa) (Xia and Culp, 1994, 1995). During amphibian development, the fibronectin-
containing extracellular matrix regulates cell migration and motility (Boucaut et al., 1984; Winklbauer, 1990; Howard et al., 1992; DeSimone, 1994), cell guidance (Shi et al., 1989; Winklbauer and Nagel, 1991) and contains information for left-right axis determination (Yost, 1995). At the onset of gastrulation, the embryo is spherical and contains a cavity in the animal hemisphere, called the blastocoel, which is delimited by cells forming the blasto- coel roof. In Pleurodeles embryos, the blastocoel roof contains two types of tissues interacting with the extra- cellular matrix that contains fibronectin. First, in the up- per part of the animal hemisphere, called the animal cap,
Syn RGD
NH2 COOH
IDAPS
Svn RGD
Homologies
0 Type I
n Type II
Type III
NH2---- COOH
y LDV EIIIB ? EIIIA ’
Kto llil
FPl
FP2
FP3
FP4
FP5
FP6
FP7
Fig. 1. Schematic diagram of one monomer of fibronectin and Pleurodeles waltl fusion proteins. Location of the putative adhesive recognition se- quences are shown. The EIIIA and EIIIB are the type III repeats known to be entirely included or spliced out. The seven fusion proteins are drawn. The
putative cell binding domains that they contain arc indicated. FPl, fusion protein containing the RGD, synergistic sites and the Hep II domains; FP2,
fusion protein containing the RGD and synergy sites; FP3, fusion protein containing the RGD site only; F’F’4, fusion protein containing the synergistic
site; FF’5, fusion protein containing the Hep II binding domain; FP6, fusion protein containing the V-region (IIICS) but lacking the LDV sequence (CSl site); FP7, fusion protein containing the LDV sequence of the V-region.

D. Alfandari et al. /Mechanisms of Development 56 (1996) 83-92
cells are epithelial and elaborate a network of fibronectin fibrils lining their basal surface (Boucaut and Darribere, 1983). These cells undergo epiboly, a morphogenetic process that is critical for ectoderm formation (Shi et al., 1987). Second, in the lower part of the animal hemi- sphere, cells of the marginal zone involute and ingress into the interior of the embryo where they actively mi- grate on the blastocoel roof using fibronectin fibrils as substratum (Boucaut et al., 1984). Together, these mor- phogenetic movements contribute to position the meso- derm. The cells of the marginal zone differ, however, with respect to their position, lineage, developmental fates and also to their way of migration (Delarue et al., 1992). In particular, at least two populations of mesoderm cells can be distinguished in the dorsal region of the marginal zone (called the dorsal marginal zone, DMZ). One, which corresponds to the presumptive head mesoderm (cephalic mesenchyme), contains cells that migrate in a dispersed manner. The second population of cells moves as a cohe- sive tissue behind the head mesoderm and forms the pre- sumptive axial (chordal) and para-axial (somitic) meso- derm. It has been established that fibronectin can support attachment of ectoderm cells and spreading and migration of both populations of dorsal-mesoderm cells, but it is not known whether the RGD-independent adhesive domains of the molecule are relevant in these processes (Boucaut et al., 1984; Darribere et al., 1988). Since ectodermal cells elaborate the network of fibronectin fibrils and dorsal mesoderm cells migrate on it to form the head, axial and para-axial mesoderm, it is important to know how the cell binding domains of fibronectin contribute to the different adhesive behaviors of these cells. Recently, the Pleuro- deles waltl cellular fibronectin has been cloned, providing the possibility of producing fusion proteins to address this question (Clavilier et al., 1993).
In this report, we have examined the capacity of fusion proteins containing one or multiple cell-binding domains to promote attachment and spreading of dispersed cells, and to drive the expansion of a sheet of dorsal-mesoderm cells. We show that ectoderm cells attach but never
spread on any fusion proteins containing the RGD or Hep II domains of fibronectin. We report also that fusion pro- teins containing both the RGD-dependent and the Hep II domains promote full spreading of isolated dorsal- mesoderm cells. Finally, we show that the Hep II domain is needed in the fusion proteins for both cell migration as single cells and expansion of a sheet of dorsal-mesoderm cells.
2. Results
To determine the relative contribution of the RGD- dependent and -independent binding domains of the cel- lular fibronectin in adhesive behaviors of amphibian em- bryonic cells, seven different fusion proteins were pro- duced. They include one or a combination of domains
A
85
I 14 hOUP
Fig. 2. Schematic representation of the in vitro assays. (A) Attachment
and spreading assays of cells from the animal cap (AC) and DMZ of the
embryo. Explants of animal caps or DMZ are dissected as indicated. Cells were dispersed by EDTA treatment and rinsed prior to inoculation
of 3 x lo3 cells into wells of a 96-well plate precoated as indicated in
Experimental procedures. After 90 mm, the plate was inverted for
5 min on another plate to remove unattached cells. Unattached, at-
tached or spreading cells were counted. (B) Expansion assays of DMZ explants. Explants (1 mm in length, 0.8 mm in width) of the DMZ were
dissected from early gastrulae (stage 8a). They were placed face down
on tibronectin or fusion proteins. After 24 h in culture the behavior of
DMZ explants was analyzed. The expansion of the cohesive sheet of
dorsal-mesoderm cells is estimated by scoring the extend of the explant
from the dorsal lip (DL) to the outer edge of the cohesive sheet.
known to possess cell binding activity (Fig. 1). The at- tachment and spreading of isolated ectoderm and dorsal- mesoderm cells (Fig. 2A) and also the expansion of a cohesive sheet of dorsal-mesoderm cells (Fig. 2B) were analyzed.
2.1. The RGD-dependent and Hep II domains promote attachment of dispersed cells
Ectoderm cells plated on fibronectin or fusion proteins FPl (Synergy (syn)-RGD-Hep II), FP2 (Syn-RGD), and FP5 (Hep II) are immobile and spherical. Conversely, on these substrata, dorsal-mesoderm cells initially exhibit a hyaline lobopodium which corresponds to movements of the cytoplasm around the cell body. The cytoplasmic movements decrease and cells change their shape. Lo- bopodia form blebs which progressively extend blunt cytoplasmic projections. After 30 min, the blunt projec- tions are replaced by filiform cytoplasmic protrusions and finally by lamelliform protrusions. In contrast, on BSA and fusion proteins FP3 (RGD), FP4 (Syn), FP6 (LDV-), and FW (LDV) neither ectoderm or dorsal mesoderm cells attach; instead, they form aggregates within 90 min.
To quantify the attachment, cells were seeded on fu- sion proteins and after a 90 min incubation, culures were inverted on a second microwell plate. Cells which do not
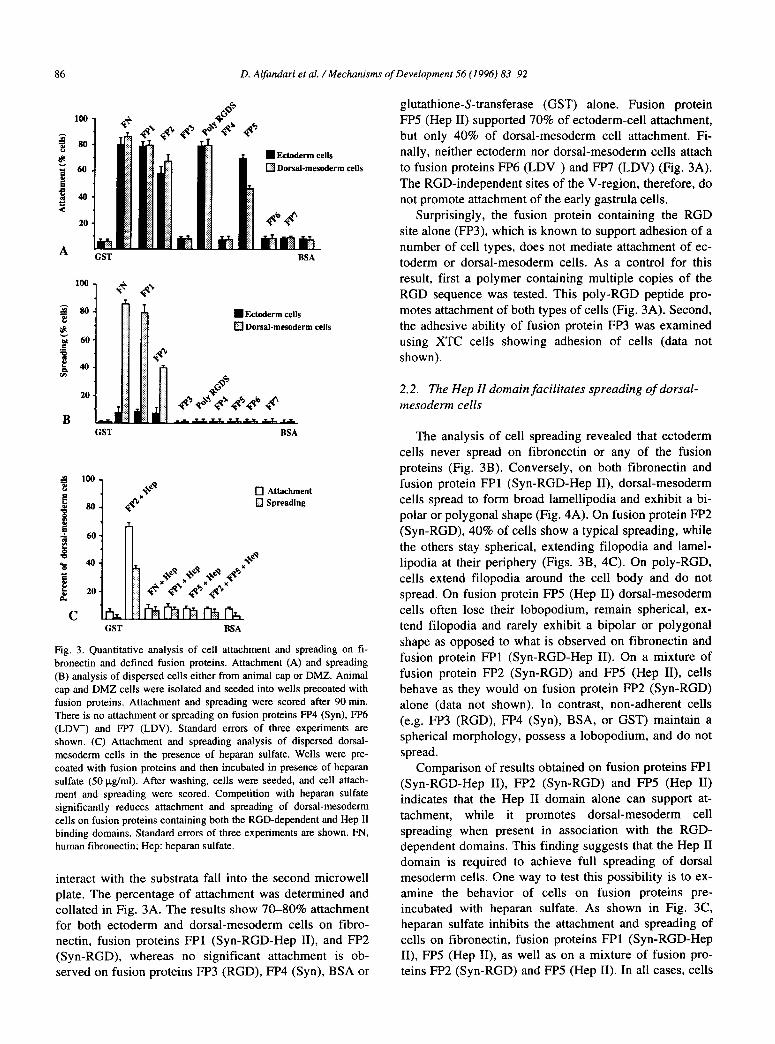
86 D. Alfandari et al. /Mechanisms of Development 56 (I 996) 83-92
P % 80
5 H Eetoderm cells
1 60 q Dorsal-mewderm cells
a
Ij 40
20
A GST BSA
p so- 1
a W Ectoderm cells 0 Dorsal-mesoderm cells
GST BSA
- 60 e I % 40
BSA
q Attachment q Spreading
Fig. 3. Quantitative analysis of cell attachment and spreading on Ii-
bronectin and defined fusion proteins. Attachment (A) and spreading
(B) analysis of dispersed cells either from animal cap or DMZ. Animal
cap and DMZ cells were isolated and seeded into wells precoated with
fusion proteins. Attachment and spreading were scored after 90 min.
There is no attachment or spreading on fusion proteins FP4 (Syn), FP6
(LDV-) and FP7 (LDV). Standard errors of three experiments are shown. (C) Attachment and spreading analysis of dispersed dorsat- mesoderm cells in the presence of heparan sulfate. Wells were pre-
coated with fusion proteins and then incubated in presence of heparan
sulfate (50 kg/ml). After washing, cells were seeded, and cell attach-
ment and spreading were scored. Competition with heparan sulfate
significantly reduces attachment and spreading of dorsal-mesoderm
cells on fusion proteins containing both the RGD-dependent and Hep II binding domains. Standard errors of three experiments are shown. FN, human fibronectin; Hep: heparan sulfate.
interact with the substrata fall into the second microwell plate. The percentage of attachment was determined and collated in Fig. 3A. The results show 70-80% attachment for both ectoderm and dorsal-mesoderm cells on fibro- nectin, fusion proteins FPl (Syn-RGD-Hep II), and FP2 (Syn-RGD), whereas no significant attachment is ob- served on fusion proteins PP3 (RGD), FP4 (Syn), BSA or
glutathione-S-transferase (GST) alone. Fusion protein FP5 (Hep II) supported 70% of ectoderm-cell attachment, but only 40% of dorsal-mesoderm cell attachment. Fi- nally, neither ectoderm nor dorsal-mesoderm cells attach to fusion proteins FP6 (LDV-) and FP7 (LDV) (Fig. 3A). The RGD-independent sites of the V-region, therefore, do not promote attachment of the early gastrula cells.
Surprisingly, the fusion protein containing the RGD site alone (FP3), which is known to support adhesion of a number of cell types, does not mediate attachment of ec- toderm or dorsal-mesoderm cells. As a control for this result, first a polymer containing multiple copies of the RGD sequence was tested. This poly-RGD peptide pro- motes attachment of both types of cells (Fig. 3A). Second, the adhesive ability of fusion protein FP3 was examined using XTC cells showing adhesion of cells (data not shown).
2.2. The Hep II domain facilitates spreading of dorsal-
mesoderm cells
The analysis of cell spreading revealed that ectoderm cells never spread on fibronectin or any of the fusion proteins (Fig. 3B). Conversely, on both fibronectin and fusion protein FPl (Syn-RGD-Hep II), dorsal-mesoderm cells spread to form broad lamellipodia and exhibit a bi- polar or polygonal shape (Fig. 4A). On fusion protein FP2 (Syn-RGD), 40% of cells show a typical spreading, while the others stay spherical, extending filopodia and lamel- lipodia at their periphery (Figs. 3B, 4C). On poly-RGD, cells extend filopodia around the cell body and do not spread. On fusion protein FP5 (Hep II) dorsal-mesoderm cells often lose their lobopodium, remain spherical, ex- tend filopodia and rarely exhibit a bipolar or polygonal shape as opposed to what is observed on fibronectin and fusion protein FPl (Syn-RGD-Hep II). On a mixture of fusion protein FP2 (Syn-RGD) and FP5 (Hep II), cells behave as they would on fusion protein FP2 (Syn-RGD) alone (data not shown). In contrast, non-adherent cells (e.g. FP3 (RGD), FP4 (Syn), BSA, or GST) maintain a spherical morphology, possess a lobopodium, and do not spread.
Comparison of results obtained on fusion proteins FPl (Syn-RGD-Hep II), FP2 (Syn-RGD) and FP5 (Hep II) indicates that the Hep II domain alone can support at- tachment, while it promotes dorsal-mesoderm cell spreading when present in association with the RGD- dependent domains. This finding suggests that the Hep II domain is required to achieve full spreading of dorsal mesoderm cells. One way to test this possibility is to ex- amine the behavior of cells on fusion proteins pre- incubated with heparan sulfate. As shown in Fig. 3C, heparan sulfate inhibits the attachment and spreading of cells on fibronectin, fusion proteins FPl (Syn-RGD-Hep II), FP5 (Hep II), as well as on a mixture of fusion pro- teins FP2 (Syn-RGD) and FP5 (Hep II). In all cases, cells

D. Al&an&n’ et al. /Mechanisms of Development 56 (19%) 83-92 87
Fig. 4. Dorsal-mesoderm cell behaviors on flbronectin and fusion proteins. (A-C) Mesoderm cells were obtained by chemical dissociation of DMZ
explants. Then, they were seeded into wells coated with fusion proteins I71 (Syn-RGD-Hep II) (A), PI’4 (Syn) (B) and PP2 (Syn-RGD) (C). (A) Phase contrast. After 90 min, on fusion protein FPl dorsal-mesoderm cells extend broad lamellipodia and tilopodia (arrowhead). They exhibit a bipolar or
polygonal shape. Some ceils possess blunt projections (arrow). (B) Phase contrast. On fusion protein FP4 which contains only the synergistic site, cells
remain spherical (arrowhead) and are unattached. (C) Phase contrast. On fusion protein PP2 containing the RGD and synergistic sites, dorsal-
mesoderm cells are spherical and extend fine Illopodia around the entire cell body (arrow). Spreading cells are also observed. Bar, 10 pm.
remain spherical and form aggregates within 90 min. Fi- nally, heparan sulfate does no perturb the attachment and spreading of cells on fusion protein FP2 (Syn-RGD; Fig. 3C). This last result suggests that heparan sulfate specifi- cally interferes with attachment and spreading on fibro- nectin and fusion protein FPl (Syn-RGD-Hep II).
2.3. The RGD-dependent and Hep II domains are required for proper expansion of a cohesive sheet of dorsal-mesoderm cells
DMZ was explanted from embryos and cultured on various substrata (Fig. 2B). Three different behaviors were observed. First, on fusion proteins FP3 (RGD), FP4 (Syn), FP6 (LDV-), and FP7 (LDV), explanted mesoderm sheets did not attach nor expand. Within 24 h the DMZ forms a compact ball of cells. Second, on poly RGD and fusion protein FP5 (Hep II), explants attach within 24 h, and the DMZ increases in thickness to form a multilayer of cells that does not expand (Fig. 5A). Finally, on fi- bronectin and fusion protein FPl (Syn-RGD-Hep II), ex- plants adhere rapidly. We observed that within 2 h, the DMZ becomes flattened. Cells, at the marginal edge of the DMZ, extend filopodia and lamellipodia. While flat- tening, the DMZ elongates as a cohesive sheet of cells. Within 6 h, at the periphery of the cell sheet, 30-40 cells break away. They spread, gain a bipolar or polygonal shape and migrate. Within 24 h, a fan-shaped cohesive
cell sheet with dispersed cells at the edges is observed (Fig. 5B). On fusion protein FP2 (Syn-RGD), we observe that the DMZ flattens and expands but no cells detach from the edge of the mesoderm cell sheet (Fig. 5C).
The expansion of the cell sheet was estimated by measuring the length from the dorsal lip of the blastopore to the outer edge of the cell sheet. The cohesive sheet of cells expands as efficiently on fibronectin as on fusion protein FPI (Syn-RGD-Hep II; Fig. 6). In 24 h, there is, on both substrata, an expansion of 1.5 mm. By contrast, DMZ plated on fusion protein FP2 (Syn-RGD) show an expansion of only 0.7 mm. With a substrate composed of both fusion proteins FP2 (Syn-RGD) and FP5 (Hep II), the DMZ explant extends as well as on fusion protein FPl (Syn-RGD-Hep II). Conversely, no expansion of the co- hesive sheet was observed on BSA or bacterially ex- pressed GST. When heparan sulfate was added before seeding DMZ explants, we did not observe significant effect on fusion protein FP2 (Syn-RGD), whereas the expansion of the DMZ explant was reduced on both fi- bronectin and fusion protein FPl (Syn-RGD-Hep II) (Fig. 6). Furthermore, there was a dramatic decrease of the number of dispersed cells ahead of the mesoderm-cell sheet (Fig. 5D,E).
3. Discussion
The major conclusions of this report are: (1) the
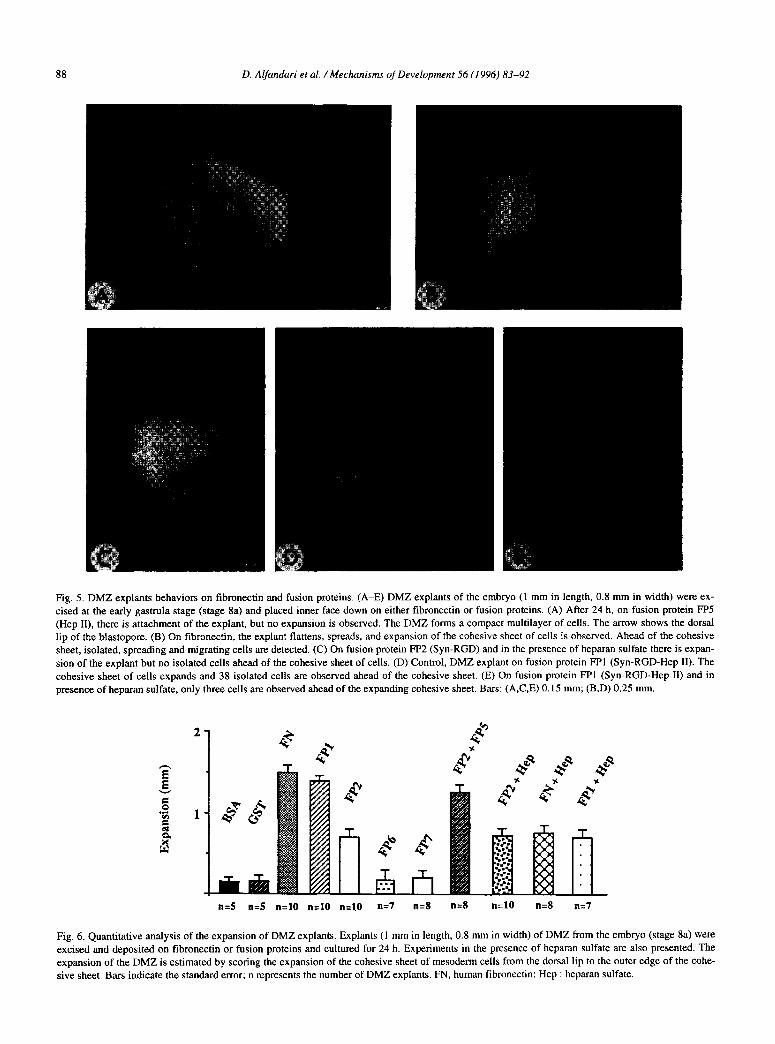
88 D. Alfandari et al. /Mechanisms of Development 56 (1996) 83-92
Fig. 5. DMZ explants behaviors on fibronectin and fusion proteins. (A-E) DMZ explants of the embryo (1 mm in length, 0.8 mm in width) were ex-
cised at the early gastrula stage (stage 8a) and placed inner face down on either tibronectin or fusion proteins. (A) After 24 h, on fusion protein FP5
(Hep II), there is attachment of the explant, but no expansion is observed. The DMZ forms a compact multilayer of cells. The arrow shows the dorsal
lip of the blastopore. (B) On tibronectin, the explant flattens, spreads, and expansion of the cohesive sheet of cells is observed. Ahead of the cohesive sheet, isolated, spreading and migrating cells are detected. (C) On fusion protein FP2 (Syn-RGD) and in the presence of heparan sulfate there is expan-
sion of the explant but no isolated cells ahead of the cohesive sheet of cells. (D) Control, DMZ explant on fusion protein FPl (Syn-RGD-Hep II). The
cohesive sheet of cells expands and 38 isolated cells are observed ahead of the cohesive sheet. (E) On fusion protein FE1 (Syn-RGD-Hep II) and in
presence of heparan sulfate, only three cells are observed ahead of the expanding cohesive sheet. Bars: (A,C,E) 0.15 mm; (B,D) 0.25 mm.
n=S n=5 n=lO n=lO n=lO n=7 n=8 n=8 n=lO n=8 n=7
Fig. 6. Quantitative analysis of the expansion of DMZ explants. Explants (1 mm in length, 0.8 mm in width) of DMZ from the embryo (stage 8a) were
excised and deposited on fibronectin or fusion proteins and cultured for 24 h. Experiments in the presence of heparan sulfate are also presented. The
expansion of the DMZ is estimated by scoring the expansion of the cohesive sheet of mesoderm cells from the dorsal lip to the outer edge of the cohe-
sive sheet. Bars indicate the standard error; n represents the number of DMZ explants. FN, human fibronectin; Hep : heparan sulfate.

D. Alfandari et al. /Mechanism of Development 56 (I 996) 83-92 89
RGD-dependent and the Hep II domains of fibronectin mediate attachment and full spreading of isolated dorsal- mesoderm cells; (2) both domains cooperate to mediate the proper expansion of a sheet of dorsal-mesoderm cells; (3) the Hep II domain facilitates cell migration as single cells ahead of the sheet of mesoderm cells
We find that the fusion protein containing only the Hep II domain promotes attachment of both animal cap and dorsal-mesoderm cells. We report that a fusion pro- tein containing both the Hep II and the RGD-dependent domains supports full expansion of a cohesive sheet of mesoderm and migration of single cells ahead of the co- hesive sheet. We show also that the .presence of heparan sulfate inhibits the attachment of dorsal-mesoderm cells to fusion proteins containing the Hep II domain and re- duces the expansion of the cell sheet as well as the num- ber of dispersed cells. The fate map of the early gastrula indicates that the DMZ contains presumptive cells of the
head, axial and para-axial mesoderm (Delarue et al., 1992). We propose, therefore, that in the culture of DMZ explants, the cohesive sheet of cells corresponds to axial and para-axial mesoderm, while the dispersed cells, ob- served ahead of the cohesive sheet, represent head meso- derm cells. Consequently, the results suggest that at least two domains of fibronectin, the RGD-dependent and the Hep II domains, might be required for complete expan- sion of axial and para-axial mesoderm and dispersion of head mesoderm. They suggest also that proteoglycans containing polysaccharides related to heparan sulfate modulate the adhesive activity of cells. In vivo, intrab- lastocoelic injections of sulfated components or heparan sulfate delay gastrulation which in turn produces anoma- lies in the anterior part of the Xenopus embryo (Slack et al., 1987; Mitani, 1989). The same experiment in Pleuro-
deles delays mesoderm cell migration, resulting in a de- fect in migration of presumptive head mesoderm cells (Alfandari and Darribtre, unpublished results). Injection of heparan sulfatase into Xenopus blastulae produces re- versal of the left-right asymmetry of both heart and gut (Yost, 1992). Although heparan sulfate is not detected in the embryo, proteoglycans containing polysaccharides related to heparan sulfate may be present. It has been shown that in Xenopus and Rana pipiens embryos, heparan sulfate proteoglycans are the major sulfated pro- teoglycans of both the gastrula and neurula (Kosher and Searls, 1973; Johnson, 1978; Yost, 1990; Itoh and Sokol, 1994). In Pleurodeles, there is evidence that cell surface glycoproteins change at the onset of gastrulation, but no data are available concerning proteoglycans (Darribbre et al., 1982; Riou et al., 1986).
The inhibition of attachment of dispersed mesodermal cells, observed with heparan sulfate, suggests that attach- ment could require cell surface proteoglycans or that heparan sulfate perturbs by steric hindrance the interac- tion between Hep II-containing fusion proteins and cell surface receptors. For example, we cannot rule out the
possibility that the fibronectin receptor, integrin tQi, which is reported to interact with the IDAPS sequence in the 14th type III repeat of the Hep II domain, may act in concert with proteoglycans to mediate this adhesive be- havior (Iida et al., 1992). Alternatively, it is possible that cell surface proteoglycans may alter the activity of in- tegrin a& on the mesoderm cell surface. Unfortunately, no information is available concerning integrin olqpi in Pleurodeles. In Xenopus, transcripts encoding Xenopus
integrin a4 subunit are first detected during gastrulation (Whittaker and DeSimone, 1993). Future studies could be directed towards examining if proteoglycans and/or in- tegrin a& are present, and also to define the sequences within the Hep II domain involved in adhesion, spread- ing, and possibly migration of head, axial and para-axial- mesoderm cells.
Fibronectin and fusion proteins containing the RGD- dependent domain promote only attachment of ectoderm cells, but mediate spreading of dorsal-mesoderm cells, suggesting that, according to their position in the embryo, cells behave differently on fibronectin. One explanation of this cell behavior is that a cell may express or activate different fibronectin receptors able to modulate its at- tachment or spreading. It has been shown that the aspi integrin is the major fibronectin receptor, and binds spe- cifically to the RGD sequence (Yamada, 1991). This finding, along with previous data showing that this integrin is expressed at the cell surface of all cells in
Pleurodeles as well as in Xenopus (Darribere et al., 1988; Gawantka et al., 1992; Joos et al., 1995), provides support for the hypothesis that a.& integrin promotes attachment of embryonic cells but is not sufficient to mediate full spreading of dorsal-mesoderm cells. The RGD sequence is also known to be the cell binding site for integrins
a&, a&, c&, avP3, o;Ps, c&, asPi (Edelman et al., 1995). In Pleurodeles, the f3,, a,, a, and a5 integrin subunits have been identified at the onset of gastrulation (Darribtre et al., 1988, 1990; Alfandari et al., 1995). In Xenopus, multiple integrin subunits are co-expressed in early development such as pi, p2, p3, p6 (Ransom et al., 1993), a2, a3, a4, a5, q and am (Whittaker and DeSi- mone, 1993; Lallier et al., personal communication), sug- gesting distinct or redundant adhesive functions espe- cially with fibronectin. We suggest, therefore, the above model to explain the adhesive interactions between cells and fibronectin at the onset of gastrulation. Cells of the animal cap, which in the embryo organize fibronectin fibrils and undergo epiboly, attach but do not spread on fibronectin. We propose that they interact with fibronectin to form fibrils using the RGD and the synergy-sites and at least one integrin, for example integrin a#, . The fact that RGD peptides and antibodies against the integrin a&
perturb in vivo fibronectin fibrillogenesis strengthens this hypothesis (Darribere et al., 1988, 1990, 1992) On the other hand, dorsal-mesoderm cells, which migrate ac- tively during gastrulation, attach and spread on fibronec-

90 D. Alfandari ef al. /Mechanisms of Development 56 (1996) 83-92
tin when RGD-dependent sites are present. Full spreading of dispersed mesoderm cell, complete expansion of a co- hesive sheet of dorsal-mesoderm and dispersion of head mesoderm cells are favored by the presence of the Hep II domain. The Hep II-dependent interaction might involve cell surface proteoglycans and/or an unidentified integrin, whereas RGD-dependent interactions require one or a combination of integrins. In this context, it is interesting
to note that the integrin a, subunit has recently been re- ported to be expressed at the surface of dorsal-mesoderm cells (Alfandari et al., 1995). The integrin a, subunit can associate with several different p subunits including PI, &,, & & and /Is to form receptors for extracellular ma- trix proteins (Edelman et al., 1995). It has been estab- lished that the a, integrin family, including a,P1, a&
a& and a& have specific roles in controlling adhesive cell behaviors (Delannet et al., 1994; Piali et al., 1995). We hypothesize that integrins of the a, family, therefore, might be important in the adhesive behavior of mesoderm cells. Recent work in Xenopus, however, suggests that position-specific changes in integrin-dependent adhesive activity do not require differences in integrin expression, and further, that a single integrin can mediate both the attachment and spreading behavior (Ramos and DeSi- mone, unpublished; Ramos et al., unpublished).
At the onset of gastrulation in Xenopus, fibronectin containing the V-region (V+ fibronectin) has been shown to be the major isoform expressed in the extracellular matrix (DeSimone et al., 1992; Danker et al., 1993). On the other hand, this domain has been described as the sec- ond major cell binding domain containing two sites for adhesion. Their minimal sequences are Leu-Asp-Val (LDV), Arg-Glu-Asp-Val (REDV) or Arg-Gly-Asp-Val (RGDV). These sites support attachment, spreading and migration of fibroblasts, lymphocytes, monocytes and neural crest cells (Humphries et al., 1987; Dufour et al., 1988; Guan and Hynes, 1990; Woods et al., 1993). In Pleurodeles, the LDV sequence of the V-region is con- served and the REDV sequence is replaced by RED1 (Clavilier et al., 1993). This is consistent with an impor- tant role for the V-region and raises the question of its function in the adhesive behaviors of mesoderm cells (Clavilier et al., 1993). Fusion proteins corresponding to the V-region, which either contain or lack the LDV se- quence (FW or FP6, respectively), are not able to pro- mote attachment or spreading of isolated ectoderm and dorsal-mesoderm nor the expansion of a sheet of dorsal- mesoderm. On fusion protein FPl, which contains the RGD-dependent and Hep II binding domains but lacks the LDV sequence, cells of the DMZ attach and spread as efficiently as on fibronectin. We observed also that on a substrate composed of a mixture of fusion proteins FPl and FP7 both containing the LDV sequence, support spreading of dorsal-mesoderm cells similar to that ob- served on fibronectin or FPl alone, suggesting that the fusion protein containing the LDV sequence does not
affect the behavior of dorsal-mesoderm cells. This appar- ent contradiction, between our results and those previ- ously described, may be attributable to conformational changes in fusion protein FP7 (LDV) yielding inaccessi- ble the LDV sequence. Alternatively, we cannot rule out the possibility that the LDV sequence of the V-region has no major functional significance for adhesive interactions of amphibian embryonic cells with fibronectin, at the on- set of gastrulation. In support of this hypothesis, no per- turbation of mesoderm-cell interactions with fibronectin has been observed when FP6 (LDV-) and FP7 (LDV) fusion proteins are injected into the blastocoel before and during gastrulation (unpublished results). Recently, in Xenopus embryos, Ramos and DeSimone (1995) have reported that cells from gastrulae do not attach or spread on the V-region. Our results are consistent with this re- port.
4. Experimental procedures
4. I. Fusion proteins
Fibronectin fusion proteins were constructed by sub- cloning defined regions of cDNA from Pleurodeles cellu- lar fibronectin (Clavilier et al., 1993), into the pGEX-KG plasmid (Guan and Dixon, 1991). Subclones were derived from two cDNA clones. The first, named PFNlO, contains the V-region but lacks the LDV sequence. The other, PFN8, contains the V-region with the LDV sequence.
The cDNA encoding fusion protein Pl (FPl) (Fig. 1) was obtained by inserting the Hint II/Hint II fragment of PFNlO into the Sma I site of pGEX-KG. The cDNA cor- responding to the synergistic sites and the RGD contain- ing fusion protein (FP2) was produced by cutting the PFNlO clone with Hint II and Spe I. The Spe I end was then blunted in order to insert the 1328 bp cDNA into the pGEX-KG Sma I site. Orientation was tested by restric- tion digest, sequence analysis and protein expression. To obtain the cDNA encoding the fusion protein containing the RGD sequence (FP3), the clone PFNlO was first di- gested with Spe I and blunted. The resulting cDNA (784 bp) was then digested with EcoR I and ligated into a
new pBluescript SK- opened with EcoR I and Hint II. This temporary construct was finally digested with BamH I and Xho I, and cloned into pGEX KG. The cDNA for the fusion protein containing the synergistic sites alone (FP4) was produced using a Hint II&OR I fragment (560 bp). A Spe VXmn I fragment of PFNlO was used for the fusion protein FP5. This cDNA was first subcloned into bluescript blunted at the Sac II site and open with Spe I on the other side. This construct was then excised with Sac I and BamH I and cloned into pGEX-KG. In the V region, the insert for the fusion protein without LDV sequence (FP6) was derived from the PFNlO clone by subcloning a Xmn I/Xho I fragment (595 bp) into Sma I and Xho I digested pGEX-KG vector. The cDNA encod-

D. Alfandari et al. /Mechanisms of Development 56 (1996) 83-92 91
ing fusion protein containing the LDV sequence (FP7) was obtained from the digestion of PFN8 with Xba I and
Xho I, and subcloning into pGEX-KG. All fusion proteins were produced by induction with
isopropyl-P-D-thiogalactoside (IPTG, Appligene) and purified as described in Guan and Dixon (1991). Purity of each fusion protein was tested by SDS-PAGE (Laemmli, 1970). One band is observed for each fusion protein ex- cept for FP4 (not shown).
4.2. Preparation of cells and dorsal marginal zones
Embryos of the amphibian urodele Pleurodeles waltl were obtained from natural matings. They were collected and staged as described by Shi and Boucaut (1995). Em- bryos were mechanically dejellied and washed in 10% normal amphibian medium (NAM; Slack, 1984). At the early gastrula stage (stage 8a), the vitelline membrane was removed manually. In order to obtain isolated cells, explants of the animal cap and the dorsal marginal zone were separated by dissection with platinum needles, as shown in Fig. 2A. The tissue fragments were incubated separately in Ca2+, Mg2+-free NAM with 2 mM EDTA. Cells were dispersed by gentle rocking of the explants during 30 min. Finally, isolated cells were washed and incubated in 67% Leibovitz medium (L15, Gibco) at 20°C before seeding on fusion proteins as described be- low.
To obtain in vitro cultures of the DMZ which mimics mesoderm<ell migration observed in vivo, DMZ ex- plants (0.8 mm in width, 1 mm in length) were dissected in 67% L15 from embryos (stage 8a) as described previ- ously (Shi et al., 1989) (Fig. 2B). Explants were placed face-down on the conditioned area of the plastic tissue culture dishes and covered with a coverslip fragment. They were cultured in 67% L15 supplemented with gen- tamycin (25 mg/ml).
4.3. Preparation of substrates
Substrates for cell attachment and spreading assays were prepared in a 96 microwell plate (Nunc). Substrates for DMZ attachment and the expansion assays were pre- pared on plastic tissue culture dishes (Corning, 60 X
15 mm, #25010). Areas of 5-6 mm in diameter were conditioned. Each well or area was coated with human or amphibian plasma fibronectin, or different fusion proteins at concentrations of 10 mg/ml in NAM overnight at 4°C in a humidified chamber. After three washes with NAM, the dishes were incubated with bovine serum albumin (BSA, 0.5 mg/ml) in NAM for at least 60 min to block non-specific binding sites. The surfaces were rinsed ex- tensively with 67% L15. For some experiments, sub- strates were coated with fibronectin or fusion proteins, non-specific binding sites were blocked with BSA and substrates were further incubated with heparan sulfate
(5Opg/ml) for 1 h. The wells were rinsed with 67% L15.
Human fibronectin, fibronectin-like engineered protein polymer (poly RGD) and heparan sulfate were from Sigma.
For some experiments, substrates were coated with fi- bronectin or fusion proteins, non-specific binding sites were blocked with BSA, and heparan sulfate was added for an incubation of 1 h.
4.4. Attachment, spreading and expansion assays
Dispersed ectoderm or dorsal-mesoderm cells (0.4- 0.5 x lo3 cells) were seeded in a microwell plate previ- ously coated with defined fibronectin fusion proteins. Attachment of cells was scored 90 min after seeding as follows. The plate was inverted on a second microwell plate and allowed for 5 min. Then, the two plates were separated. The unbound and the bound cells of each well were scored by manual counting. The percentage of ad- herent cells per well was expressed as: attachment (% cells) = number of bound cells x lOO/number of seeded cells. For each experiment non-specific binding was de- termined by seeding cells on BSA or GST alone.
Cell spreading was measured by direct manual count- ing 90 min after seeding. We scored a cell as being spread when cell assumed a bipolar, triangular or polygonal shape as described in Dar-rib&e et al. (1988).
The expansion of the cohesive sheet of cells, which stems from the DMZ explant, was observed during a 24 h period. The expansion was defined by measuring the length between the dorsal lip and the margin of the cohe- sive sheet of cells. The measurements were done directly with an eyepiece reticule or on photographs.
Acknowledgments
We are very grateful to Pr. J.C. Boucaut for support, encouragement and comments concerning this work. We thank Drs. M. Delarue and D.L. Shi for reading the manu- script and providing insightful recommendations and dis- cussion. We thank P. Grout and C. Arnoult for the excel- lent maintenance in amphibian rearing and XTC cell line. We extend special thanks to M. Skalski for helping in fusion protein production. This work was supported by grants from CNRS (U.R.A. 1135), Universite P. and M. Curie (Paris VI), MRE (92CO653), the Institut Universi- taire de France and by the USPHS (HD 26402, HD 01104).
References
Alfandari, D., Whittaker, C.A., DeSimone, D.W. and Dar&&~, T. (1995) Dev. Biol. 170,249-261.
Aota, S., Nagai, T. and Yamada, K.M. (1991) J. Biol. Chem. 266, 15938-15943.
Aota, S., Nomizu, M. and Yamada, K.M. (1994) J. Biol. Chem. 269, 24756-24761.

92 D. Alfandari et al. /Mechanisms of Development 56 (1996) 83-92
Boucaut, J.C. and Darribere, T. (1983) Cell Tissue Res. 234, 135514.5.
Boucaut, J.C., Darribere, T., Poole, T.J., Aoyama, H., Yamada, K.M.
and Thitry, J.P. (1984) J. Cell Biol. 99,1822-1830.
Bowditch, R.D., Hariharan, M., Tominna, E.F., Smith, J.W., Yamada,
K.M., Getzoff, E.D. and Ginsberg, M.H. (1994) J. Biol Chem. 269,10856-10863.
Clavilier, L., Riou, J.F., Shi, D.L., DeSimone, D.W. and Boucaut, J.C.
(1993) Cell Adhesion Commun. 1, 83-93.
Danker, K., Hacke, H., Ramos, J., DeSimone, D.W. and Wedlich, D.
(1993) Mech. Dev. 44, 155-165.
Darrikre, T., Boucaut, J.C. and Keil-Dlouha, V. (1982) Biochim. Bio-
phys. Acta 686, 145-152.
Darrib&re, T., Yamada, K.M., Johnson, K.E. and Boucaut, J.C. (1988) Dev. Biol. 126, 182-194.
Darribere, T., Guida, K., Larjava, H., Johnson, K.E., Yamada, K.M.,
Thiery, J.P. and Boucaut, J.C. (1990) J. Cell Biol. 110, 1813-1823.
Darribi%e, T., Koteliansky, V.E., Chernousov, M.A., Akiyama, S.K.,
Yamada, K.M., Thiery, J.P. and Boucaut J.C. (1992) Dev. Dyn. 194,63-70.
Delarue, M., Sanchez, S., Johnson, K.E., Darribere, T. and Boucaut,
J.C. (1992) Development 114, 135-146.
Delannet, M., Martin, F., Bossy, B., Cheresh, D.A.. Reichardt, L.F. and
Duband, J.-L. (1994) Development 20.2687-2702.
DeSimone, D.W. (1994) In Takada, Y. (ed.), lntegrins: The Biological
Problems, CRC Press.
DeSimone, D.W., Norton, P.A. and Hynes, R.O. (1992) Dev. Biol. 149,
357-369.
Dufour, S. Duband, J.L., Humphries, M.J., Obara, M., Yamada, K.M.
and Thiery, J.P. (1988) EMBO J. 7,2661-2671.
Edelman, J.M., DiMilla, P.A. and Albelda, S.M. (1995) In Richardson,
P.O. and Steiner, M. (eds.), Principles of Cell Adhesion, CRC Press,
163-186.
Fuji& H., Mohri, H., Kanamori, H., Iwamatsu, A. and Okbo, T. (1995) Exp. Cell Res. 217,484-489.
Gawantka, V., Ellinger-Ziegelbauer, H. and Hausen, P. (1992) Devel-
opment 115.595605.
Guan, J.L. and Hynes, R.O. (1990) Cell, 60,53-63.
Guan, K.L. and Dixon, J.E. (1991) Anal. Biochem. 192,262-267.
Haugen, P.K., McCarthy, J.B., Skubitz, A.P.N., Furcht, L.T. and Le-
toumeau, P.C. (1990) J. Cell Biol. 111.2733-2745.
Howard, J.E., Hirst, E.M. and Smith, J.C. (1992) Mech. Dev. 38, 109-
119.
Humphries, M.J., Akiyama, S.K., Komoriya, A., Olden, K. and Ya-
mada, K.M. (1986) J. Cell Biol. 103,2637-2647.
Humphries, M.J., Komoriya, A., Akiyama, S.K., Olden, K. and Ya-
mada, K.M. (1987) J. Cell Biol. 106, 1289-1297.
lida, J., Skubitz, A.P.N., Furcht, L.T., Wayner. E.A. and McCarthy, J.B.
(1992) J. Cell Biol. 118.431-444.
Itoh, K. and Sokol, S. (1994) Development 120, 2703-2711.
Johnson K.E. (1978) J. Cell Sci. 32, 109-136.
Johnson, K.E., Darribere, T. and Boucaut. J.C. (1993) I. Exp. Zool. 265,4&53.
Joos, T.O., Whittaker, CA., Meng, F., DeSimone, D.W., Gnau, V. and
Hausen, P. (1995) Mech. Dev. 50, 187-199.
Kosher, R.A. and Searles, R.L. (1973) Dev. Biol. 32,[email protected].
Laemmli, U.K. (1970) Nature (London) 227,680-689.
Mitani, S. (1989) Development 107.423435.
Mould, A.P. and Humphries, M.J. (1991) EMBO J. 10,4089-l095.
Piali, L., Hammel, P., Uherek, C., Bachmann, F., Gisler, R.H., Dunon,
D. and Imhof, B.A. (1995) J. Cell Biol. 130.45 1460.
Pietschbacher, M.D. and Ruoslahti, E. (1984) Proc. Natl. Acad. Sci.
USA. 81,5985-5988. Ransom, D.G., Hens, M.D. and DeSimone, D.W. (1993) Dev. Biol.
160,265-275.
Riou, J.F., Darrib&e, T. and Boucaut, J.C. (1986) J. Cell Sci. 82, 23-
40.
Shi, D.L. and Boucaut, J.C. (1995) Int. J. Dev. Biol. 39, 427441.
Shi, D.L., Delarue, M., Dar&&e. T., Riou, J.F. and Boucaut, J.C.
(1987) Development 100, 147-161.
Shi, D.L. Danib&, T., Johnson, K.E. and Boucaut, J.C. (1989) Devel-
opment 105.223-236.
Slack, J.M.W. (1984) J. Embryol. Exp. Morphol. 80, 289-319.
Slack, J.M.W., Darlington, B.G. Heath, J.K. and Godsave, S.F. (1987)
Nature (London) 326,197-200.
Wayner, E.A. and Kovach, N.L. (1992) J. Cell BioI. 116,489-497. Wayner, E.A., Garcia-Pardo, A., Humphries, M.J., McDonald, J.A. and
Carter, W.G. (1989) J. Cell Biol. 109, 1321-1330.
Whittaker, CA. and DeSimone, D.W. (1993) Development 117, 1239-
1249. Winklbauer, R. (1990) Dev. Biol. 142, 155-168.
Winklbauer, R. and Nagel, M. (1991) Dev. Biol. 148.573-582.
Woods, A., McCarthy, J.B., Furcht, L.T. and Couchman, J.R. (1993)
Mol. Biol. Cell 4.605-613.
Xia, P. and Culp, L.A. (1994) Exp. Cell Res. 213, 253-265.
Xia, P. and Gulp, L.A. (1995) Exp. Cell Res., 217.517-527.
Yamada, K.M. (1991) J. Biol. Chem. 266.12809-12812.
Yamada, K.M. and Kennedy, D.W. (1984) J. Cell Biol. 99.29-36.
Yost, H.J. (1990) Development 110, 865-874.
Yost, H.J. (1992) Nature 357, 158-161
Yost, H.J (1995) Cell 82, 689-692.




















