
The Na+/K+/Cl cotransport in C6 glioma cells. Properties and role in volume regulation
9
Eur. J. Biochem. 171,425-433 (1988) (0 FEBS 1988 The Na+/K+/Cl- cotransport in C6 glioma cells Properties and role in volume regulation Olivier CHASSANDE ', Christian FRELTN ', Dariush FARAHIFAR', Thierry JEAN ' and Michel LAZDUNSKI ' Unit6 210 Institut National de la Santt: et de la Recherche Medicale, Facult6 de Mkdecine, Nice (Received August 13/0ctober 12, 1987) - EJB 87 0959 Centre de Biochimie du Centre National de la Recherche Scientifique, Nice The role of the Naf/K+/CI- cotransporter in the regulation of the volume of C6 astrocytoma cells was analyzed using isotopic fluxes and cell cytometry measurements of the cell volume. The system was inhibited by 'loop diuretics' with the following order of potency: benzmetanide > bumetanide > piretanide > furosemide. Under physiological conditions of osmolarity of the incubation media, equal rates of bumetanide-sensitive inward and outward K' fluxes were observed. Blockade of the Na '/Kt/Cl- cotransporter with bumetanide did not lead to a modification in the mean cell volume. When C6 cells were incubated in an hyperosmotic solution, a cell shrinkage was observed. It was accompanied by a twofold increase in the activity of the Na+/K+/Cl- cotransport, which then catalyzed the net influx of K'. In spite of this increased activity, no cell swelling could be measured. Incubation of the cells in an iso-osmotic medium deprived of either Na+, K+ or Cl- also produced cell shrinkage. Large activations (up to tenfold) of the Nat/K+/Cl- cotransport together with a cell swelling back to the normal volume were observed upon returning ion-deprived C6 cells to a physiological solution. This cell swelling was completely prevented in the presence of bumetanide. It is concluded that the Na+/K+/CI- cotransport system is one of the transport systems involved in volume regulation of glial cells. The system can either be physiologically quiescent or active depending on the conditions used. A distinct volume regulating mechanism is the Na+/H exchange system. Vertebrate cells are now thought to regulate their volume by controlling the movements of Na', K+ and C1- across their plasma membrane [l - 31. Abnormal volume regulation is encountered in cytotoxic brain edema. Swelling of ghal cells occurs in various forms of cerebral disorders and is probably an important component of acute and chronic forms of ce- rebral damage [4 - 81. Yet our understanding of the molecular mechanisms that regulate the volume of glial cells is still limited. Bourke et al. 19, 101 first showed that incubation of brain slices in high-K' media in the presence of bicarbonate caused a swelling of astroglia. More recent work by Kempski et al. [8, 111 shows (a) that hypotonically swollen C6 glioma cells do regulate their volume and (b) that energy failure per se was not sufficient to induce cell swelling. Important questions to be answered are (a) What are the transmembrane ion pathways that are involved in volume regulation of glial cells? and (b) How is their activity impaired under pathologi- cal conditions? One volume-sensitive ion pathway in C6 cells has been identified as the Na+/H+ exchange system [12, 131. Another potential transport system for regulating volume is the Na+/K'/Cl- cotransport. Its role in the regulation of the cell volume of a number of non-cerebral cells has been extensively documented (see for reviews 12, 14- 161). There is evidence indicating that this system is present in glial cells and suggestions have been made for its involvement in volume regulation of these cells [6, 17 - 231. The purpose of this paper is to characterize in sufficient detail the basic pharmacological and biochemical properties of the Na+/K ' /Cl- cotransport system of C6 glioma cells and to define some of the conditions in which the system is involved in volume regulation. Correspondence to 0. Chassande, Centre de Biochimie du CNRS, Parc Valrose, F-06034 Nice Cedex, France MATERIALS AND METHODS Materials Dulbecco's modified Eagle's medium and fetal bovine serum were purchased from Gibco. Bumetanide and benzmetanide were kindly provided by Dr L. Feit (Leo Pharmaceuticals, Ballrup, Denmark). Furosemide and ouabain were from Sigma. Piretanide was a kind gift from Dr C. Ellory. Ethylisopropylamiloride was synthesized as pre- viously described [24]. "RbC1 (3 mCi/mg) and 22NaC1 (500 mCi/mg) were from Amersham. 42KC1 (2 mCi/mg), H36Cl (1 mCi/g) and ~-[3,4,5-~H]leucine (60 Ci/mmol) were from the Commissariat A 1'Energie Atomique (Saclay, France). Cell cultures C6 cells were grown in Dulbecco's modified Eagle's me- dium supplemented with 10% fetal bovine serum, 50 units/ml penicillin and 200 pg/ml streptomycin. For ion-flux exper- iments, cells were seeded into 24-well Falcon tissue-culture clusters and used at a near-confluent stage. Cells were labelled with 2 mCi/nil [3H]leucine overnight prior to uptake exper- iments. [3H]Leucineradioactivity was used to correct for cell losses during the washing procedures. Solutions The standard solution used to incubate the cells during biochemical experiments was Earle's salt solution (140 mM NaCI, 5mM KCl, 1.8mM CaC12, 0.8mM MgS04, 5mM glucose buffered at pH 7.4 with 25 mM Hepes/Tris). Hypo- osmotic solutions were obtained by reducing the concen-
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The Na+/K+/Cl cotransport in C6 glioma cells. Properties and role in volume regulation

Eur. J. Biochem. 171,425-433 (1988) (0 FEBS 1988
The Na+/K+/Cl- cotransport in C6 glioma cells Properties and role in volume regulation
Olivier CHASSANDE ', Christian FRELTN ', Dariush FARAHIFAR', Thierry JEAN ' and Michel LAZDUNSKI
' Unit6 210 Institut National de la Santt: et de la Recherche Medicale, Facult6 de Mkdecine, Nice
(Received August 13/0ctober 12, 1987) - EJB 87 0959
Centre de Biochimie du Centre National de la Recherche Scientifique, Nice
The role of the Naf/K+/CI- cotransporter in the regulation of the volume of C6 astrocytoma cells was analyzed using isotopic fluxes and cell cytometry measurements of the cell volume. The system was inhibited by 'loop diuretics' with the following order of potency: benzmetanide > bumetanide > piretanide > furosemide. Under physiological conditions of osmolarity of the incubation media, equal rates of bumetanide-sensitive inward and outward K' fluxes were observed. Blockade of the Na '/Kt/Cl- cotransporter with bumetanide did not lead to a modification in the mean cell volume. When C6 cells were incubated in an hyperosmotic solution, a cell shrinkage was observed. It was accompanied by a twofold increase in the activity of the Na+/K+/Cl- cotransport, which then catalyzed the net influx of K'. In spite of this increased activity, no cell swelling could be measured. Incubation of the cells in an iso-osmotic medium deprived of either Na+, K+ or Cl- also produced cell shrinkage. Large activations (up to tenfold) of the Nat /K+/Cl- cotransport together with a cell swelling back to the normal volume were observed upon returning ion-deprived C6 cells to a physiological solution. This cell swelling was completely prevented in the presence of bumetanide. It is concluded that the Na+/K+/CI- cotransport system is one of the transport systems involved in volume regulation of glial cells. The system can either be physiologically quiescent or active depending on the conditions used. A distinct volume regulating mechanism is the Na+/H +
exchange system.
Vertebrate cells are now thought to regulate their volume by controlling the movements of Na', K + and C1- across their plasma membrane [l - 31. Abnormal volume regulation is encountered in cytotoxic brain edema. Swelling of ghal cells occurs in various forms of cerebral disorders and is probably an important component of acute and chronic forms of ce- rebral damage [4 - 81. Yet our understanding of the molecular mechanisms that regulate the volume of glial cells is still limited. Bourke et al. 19, 101 first showed that incubation of brain slices in high-K' media in the presence of bicarbonate caused a swelling of astroglia. More recent work by Kempski et al. [8, 111 shows (a) that hypotonically swollen C6 glioma cells do regulate their volume and (b) that energy failure per se was not sufficient to induce cell swelling. Important questions to be answered are (a) What are the transmembrane ion pathways that are involved in volume regulation of glial cells? and (b) How is their activity impaired under pathologi- cal conditions? One volume-sensitive ion pathway in C6 cells has been identified as the Na+/H+ exchange system [12, 131. Another potential transport system for regulating volume is the Na+/K'/Cl- cotransport. Its role in the regulation of the cell volume of a number of non-cerebral cells has been extensively documented (see for reviews 12, 14- 161). There is evidence indicating that this system is present in glial cells and suggestions have been made for its involvement in volume regulation of these cells [6, 17 - 231. The purpose of this paper is to characterize in sufficient detail the basic pharmacological and biochemical properties of the Na+/K ' /Cl- cotransport system of C6 glioma cells and to define some of the conditions in which the system is involved in volume regulation.
Correspondence to 0. Chassande, Centre de Biochimie du CNRS, Parc Valrose, F-06034 Nice Cedex, France
MATERIALS AND METHODS
Materials
Dulbecco's modified Eagle's medium and fetal bovine serum were purchased from Gibco. Bumetanide and benzmetanide were kindly provided by Dr L. Feit (Leo Pharmaceuticals, Ballrup, Denmark). Furosemide and ouabain were from Sigma. Piretanide was a kind gift from Dr C. Ellory. Ethylisopropylamiloride was synthesized as pre- viously described [24]. "RbC1 (3 mCi/mg) and 22NaC1 (500 mCi/mg) were from Amersham. 42KC1 (2 mCi/mg), H36Cl (1 mCi/g) and ~-[3,4,5-~H]leucine (60 Ci/mmol) were from the Commissariat A 1'Energie Atomique (Saclay, France).
Cell cultures
C6 cells were grown in Dulbecco's modified Eagle's me- dium supplemented with 10% fetal bovine serum, 50 units/ml penicillin and 200 pg/ml streptomycin. For ion-flux exper- iments, cells were seeded into 24-well Falcon tissue-culture clusters and used at a near-confluent stage. Cells were labelled with 2 mCi/nil [3H]leucine overnight prior to uptake exper- iments. [3H]Leucine radioactivity was used to correct for cell losses during the washing procedures.
Solutions
The standard solution used to incubate the cells during biochemical experiments was Earle's salt solution (140 mM NaCI, 5mM KCl, 1.8mM CaC12, 0.8mM MgS04, 5mM glucose buffered at pH 7.4 with 25 mM Hepes/Tris). Hypo- osmotic solutions were obtained by reducing the concen-

426
tration of NaCl while keeping constant concentrations of KCI, CaCI2, MgS04, glucose and buffer. Hyperosmotic media were obtained by the addition of 200mM mannitol to the incubation solution without changing the ionic concentrations of the solution. Modified Earle's salt solution that contained different concentrations of Na', K+ or C1- were prepared using N-methyl-D-glucamine and methanesulfonate as sub- stituents for cations and CI- respectively. lso-osmotic solutions refer to solutions with an osmolarity of 315 & 10 mOsm/l.
Measurements of cell volume
Cells were harvested from the culture flasks using a CaZ+- free medium, centrifuged, resuspended in Earle's salt solution and kept in suspension at 4°C. Mean cell volumes were deter- mined by flow cytometry using an ATC-3000 cell sorter (Odam). Briefly, the single-cell suspension was forced through an aperture of 120 pm together with a constant electrical current. Passage of a cell through the aperture caused an increased electrical resistance. The height of the resulting volt- age pulse was proportional to the cross-section of the cell. For each measurement, at least 50000 output pulses were analyzed by computer. Acquisition time was usually less than 45 s. Relative volumes were obtained for each experiment.
Influx experiments
Flux experiments were performed in two steps. First, the culture medium was removed by suction and replaced by an equilibration solution of desired composition. After usually 10 min of equilibration, the incubation solution was removed by suction and replaced by the uptake medium that contained the labelled isotope and inhibitors as desired. In all exper- iments using 86Rb', the isotope was used as a tracer for K'. Temperature was 37°C. At the end of the uptake period the incubation medium was removed by suction and the extra- cellular radioactivity was washed using a 0.1 M MgC12 solu- tion [25]. The time of uptake was 1 min unless otherwise indicated. The washing procedure lasted less than 15 s. Cells were then harvested in 0.1 M NaOH and the radioactivity incorporated by the cells was determined. Unless otherwise indicated all incubation media used for 2zNaf , 86Rb+ or 42K+ uptake experiments were supplemented with 1 mM ouabain. Cell proteins were assayed according to Hartree [26] using bovine serum albumin as standard. In routine exper- iments the activity of the Na+/K+/CI- cotransport was mea- sured as the initial rate of bumetanide-sensitive "Rb' uptake. This rate is defined as the difference in the rate of ouabain- insensitive "Rb' uptake measured in the absence and the presence of 0.1 mM bumetanide.
42K+ and "Rb' efflux experiments
For efflux experiments, cells were loaded with 42K' or with "Rb' (8 lCi/ml) and 2 mCi/ml of [3H]leucine overnight in complete culture medium. The culture medium was then removed by suction, cells were rapidly washed with Earle's salt solution and further incubated in the desired medium.
RESULTS
Volume changes in response to anisotonic media
Exposure of C6 cells to a hypotonic medium (100 mOsm/ 1) produced an immediate cell swelling (Fig. 1). Then the cell
volume decreased rapidly back to normal although hypotonicity was maintained (Fig. 1). The inset of Fig. 1 shows that regulatory volume decrease was accompanied by a rapid efflux of 86Rb+ out of the cells. Conversely, exposure of C6 cells to a solution made hyperosmotic by the addition of 200 mM mannitol (540 mOsm/l) produced a rapid cell shrinking with no evidence for recovery for at least 15 rnin (Fig. 1, main panel).
K ' transport pathways under iso-osmotic conditions
Ouabain (2 mM), which blocks the (Na+,K+)ATPase, produced a 39 7% inhibition of the initial rate of x6Rb+ or 42K+ uptake by C6 cells. The value for ouabain inhi- bition of the rate of 86Rb' uptake was observed at 50 pM (data not shown). Bumetanide (0.1 mM), a potent inhibitor of the Nai/K+/C1- cotransport produced a 29 f 5% inhi- bition of the initial rate of 86Rb+ uptake. A mixture of ouabain and bumetanide produced a 70 +_ 5% inhibition of the rate of 86Rb+ uptake. The additivity of the effect indicates that the two drugs act on separate K f uptake pathways.
The right panel of Fig. 2 shows that bumetanide also in- hibited a 4zK+ efflux component in C6 cells. The important point is that the rate of bumetanide-sensitive 4zKf efflux (Fig. 2, right panel) was about the same as the rate of bumetanide-sensitive 4zKf influx (Fig. 2, left panel), in- dicating that under iso-osmotic conditions, the Na + /K'/Cl- cotransport catalyzed no net transport of K' across the mem- brane. In accordance with this finding we observed that C6 cells retained their volume for at least 1 h after blocking the activity of the Na'/Kt /Cl- cotransport with 0.1 mM bumetanide (Fig. 1, main panel).
The injluence of hyperosmolar solutions on K + transport pathways
Fig. 2 compares the rates of 42K+ influx and efflux by C6 cells that have been incubated in a medium made hyperosmo- tic by the addition of 200 mM mannitol. Hyperosmotic con- ditions did not modify the 42K+ efflux component (Fig. 2, right panel). They did not modify the rate of 4zK' influx during the first 5 min of exposure to the hyperosmotic solu- tion, then they increased it and this increase was not observed in the presence of bumetanide (Fig. 2, left panel). The inset A of Fig. 2 presents the dose-response curve for mannitol action on the rate of bumetanide-sensitive 86Rb+ uptake. A maximum activation of 2.2-fold was observed for mannitol concentrations higher than 200 mM. The effect of mannitol on "Rb' uptake was reversed within 5 min following a shift of the cells back to an iso-osmotic medium (Fig. 2, inset B).
The influence of ion deprivation on the volume of C6 cells
Fig. 3 shows that incubation of C6 cells into a medium that contained no Na', no K + or no C1- with osmotic re- placement of cations with N-methylglucamine or of C1- by methanesulfonate produced a cell shrinkage. This shrinkage was the largest in Cl--free and Na+-free media. It was rapidly reversible upon returning cells to a complete Earle's salt solu- tion that contained Na', K f and C1-. Bumetanide (0.1 mM) almost completely prevented the increase in cell volume that followed the shift of the cells back to a physiological salt solution (Fig. 3).
Fig. 4 compares the rates of 42K' influx and efflux by C6 cells during the period of volume increase that followed a

427
15-min exposure to a Na+-free medium. It shows that the rate of K f efflux and the rate of bumetanide-insensitive 42Kf influx were not modified as compared to cells that have been maintained in a physiological salt solution. Only the rate of bumetanide-sensitive 42K+ influx increased and this increase
2.1
Ly
3 2
1.5 F 2 Y
1 .o
0.5
L 1 2 3 4
\ TIME (minutes)
5 10 1 5 ” 3 0 ’ 6 0
TIME (Mlnutes)
Fig. 1. The response of C6 cells to anisotonic media. Main panel: relative cell volumes after an exposure to hypo-osmotic (100 mOsm/l) ( O ) , hyperosmotic (540 mOsm/l) (A) and iso-osmotic solutions (310 mOsm/l) (B) and after exposure to 0.1 mM bumetanide under iso-osmotic conditions (V). Inset: influence of a hypo-osmotic ex- posure on *‘Rb+ content of C6 cells. Experiments were performed as described in Materials and Methods. Hypo-osmotic conditions (100 mOsm/l) (B), iso-osmotic conditions (A)
o:b 0.8
0.8 I I I I I I I
10 20 30 40 0 10 20
TIME (mlnutes)
Fig. 3. Influence of ion deprivation on the volume of C6 cells. Left panel: a cell suspension was centrifuged and resuspended in a Na+- free (upper panel), a Cl--free (middle panel) or a K+-free (lower panel) modified Earle’s salt solution (310 mOsm/l) and the relative cell volumes were determined after different times of incubation. Right panels: cells were equilibrated for 30 min in a Na+-free (upper panel), a CI--free (middle panel) or a K+-free (lower panel) modified Earle’s salt solution, centrifuged and further incubated in iso-osmotic Earle’s salt solution in the absence ( 0 ) or in the presence (B) of 0.1 mM bumetanide. Relative cell volumes were determined after different times of incubation
5 10 TIME (Mlnules)
5 10
TIME (Mlnutos)
Fig. 2. Time courses o f 4 2 K + influx and 42K+ efflux under iso-osmotic and hyperosmotic conditions. Left panel: time courses of 42K+ uptake by ouabain-treated C6 cells under iso-osmotic (B, 0) or hyperosmotic conditions (A, A). Experiments were performed in the absence of (B, A) or the presence (0, A ) of 0.1 mM bumetanide. Right panel: time courses of 42K+ efflux by ouabain-treated C6 cells under iso- osmotic ( , 0) or hyperosmotic conditions (A, A). Experiments were performed in the absence ( , A) or the presence (0, A ) of 0.1 mM bumetanide. Inset A : dose-response curve for mannitol activation of the rate of bumetanide-sensitive 86Rb+ uptake by C6 cells. Cells were equilibrated for 10 min in an Earle’s salt solution supplemented with different concentrations of mannitol. Initial rates of bumetanide-sensitive 86Rbf uptake were then determined using the same medium as the equilibration medium supplemented with 86Rb+ and 1 mM ouabain. Time of uptake was 1 min. Inset B: time course for the deactivation of the Na+/K+/CI- cotransport after returning hyperosmotically shocked cells to an iso-osmotic medium. Cells were exposed for 10 min to a hyperosmotic Earle’s salt solution (200-mM-mannitol-supplemented Earle’s salt solution) and then switched to an iso-osmotic medium for the times indicated. The rates of bumetanide-sensitive “Rb+ uptake were then determined under iso-osmotic conditions using a 1-min uptake period. The rate of bumetanide and ouabain-insensitive s6Rb+ uptake was not modified during the course of the experiments and was subtracted from the experimental data

428
ION DEMIVED
I N. K CI K CI K CI
/ b
1 I 1 1 I 2 4 6 8 10
TIYE (Ylnutos)
Fig. 4. Time courses of 4 2 K + influx and 42K' efflux under iso-osmotic conditions and during volume increase following a IS-min exposure to a Naf-frec, medium. Left panel: time courses of 42K+ uptake by ouabain-treated C6 cells after a 15-min equilibration in complete Earle's salt solution ( W , 0) or in Na+-free modified Earle's salt solution (A, A). All solutions used were iso-osmotic. Experiments were performed in the absence (A, W ) or the presence (0, A ) of 0.1 mM bumetanide. Right panel: time courses of 42K+ efflux by ouabain-treated C6 cells after a 15-min equilibration in complete Earlc's salt solution (U, 0) or in Na+-free modified Earle's salt solution (A, A). All solutions used were iso-osmotic. Experiments were performed in the absence ( W . A) or in presence (0, A ) of 0.1 mM bumetanide. Inset: influencc of different compositions of the equilibration solution on the initial rate of bumetanide-sensitive 86Rbf uptake. Cells were first equilibrated in complete Earle's salt solution, or in modified Earle's salt solutions without Na', K + or CI- under iso-osmotic (310 niOsm/l, open bars) or hyperosmotic (540 mOsm/l, shaded bars) conditions. Cells were then shifted to a complete Earle's salt solution of the same osmolarity as the equilibration solution to measure the activity of the Na+/K+/CI- cotransport. Time of uptake was 1 min. Data are expressed relative to the initial rate of bumetanide-sensitive 86Rb+ uptake, measured in a parallel experiment using a complete Earle's salt solution
correlates with the period of volume increase. The inset of Fig. 4 shows that a 15-min period of either Na', K f or C1- deprivation caused a similar 5 - 6-fold increase in the activity of the Na+/Kf/C1- cotransport upon returning cells to a physiological solution. A larger (8 - 9-fold) increase was ob- served when cells had been deprived of both K + and C1- for 15 min. Finally the inset of Fig. 4 shows that if the period of ion deprivation was achieved using a hyperosmotic medium (540 mOsm/l), still larger (9 - 11 -fold) increases in the activity of the Na+/K+/CI- cotransport were observed. None of these treatments modified the rate of ouabain and bumetanide-insensitive "Rb+ uptake (data not shown).
The left panel of Fig. 5 presents the influence of the time of exposure to a Na', a K+ or a CI--free medium on the activity of the Na+/K+/CI- cotransport. It shows that the system is turned on slowly and that a maximum had not been reached after 15 min although cell shrinking had stabilized (Fig. 3). The right panel of Fig. 5 shows that following a 15- min period of Na' or of C1- deprivation and a shift back to physiological conditions, the Na+/K+/Cl- cotransport turned off within 5 - 10 min. The inset of Fig. 5 indicates that it was not necessary to remove all external Na', K + or C1- from the external medium to turn on the Naf/K+/C1- cotransport. Half-maximum activations of the rate of bumetanide-sensitive 86Rb+ uptake were already observed after a 15-min incubation of the cells in a 10 mM Na' me- dium, a 8 mM C1- medium or a 0.5 mM K + medium.
The pharmacological properties of the Na+/K+/CI- cotransport
Under iso-osmotic conditions the half-maximum inhi- bition of the Na+/Kf/C1- cotransport by bumetanide was
observed at 0.5 pM (Fig. 6). Three other 'loop diuretics' also inhibited the rate of external CI--dependent "Rb+ uptake by C6 cells with half-maximum inhibitions at 0.2 pM, 2 pM and 8 pM for benzmetanide, piretanide and furosemide respectively (Fig. 6) . The insets A and B of Fig. 6 further show that the dose-response curves for bumetanide and piretanide actions on C6 cells were the same under the different con- ditions of activation of the cotransport system. This indicates that very likely the same transporting system was operating under all conditions tested.
Interaction o f t h e N u + , K+/CI- cotransport with Na+lK+ and Cl-
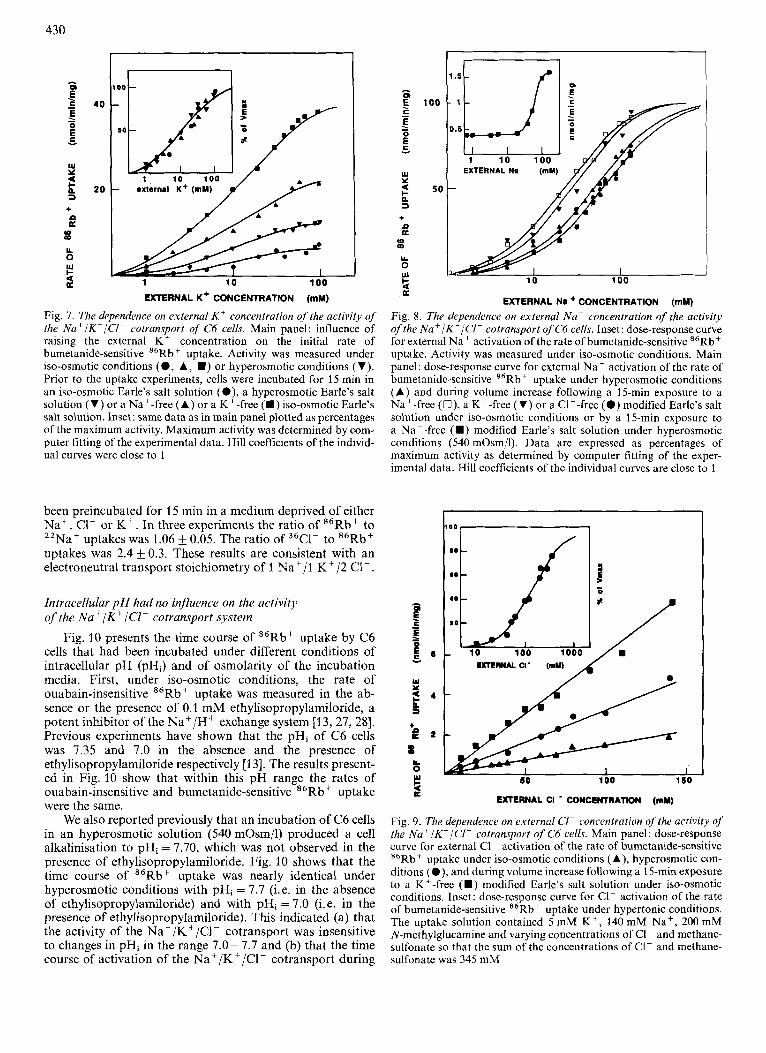
The properties of interaction of the Na+/K+/CI- cotransport with its substrates, Na', K + and C1-, were stud- ied under iso-osmotic conditions and under the different ex- perimental conditions that led to an increase in its activity. Fig. 7 shows the dependence on external K + . Substituting all external K + by N-methylglucamine completely suppressed the rate of buinetanide-sensitive 86Rb+ uptake. The value for external K + activation of the cotransport was the same (10-20 mM) in the different conditions studied (Fig. 7).
Fig. 8 shows the external Na' dependence of the activity of the cotransport system. After a cell shrinkage obtained either by mannitol treatment or by a period of ion deprivation, substituting external Na+ by N-methylglucamine completely suppressed the rate of bumetanide-sensitive 86Rb+ uptake. The value for Na' activation of the cotransporter was observed at 20-50 mM. In contrast we observed that under iso-osmotic conditions, substituting external Na+ by N-methylglucamine reduced the rate of bumetanide-sensitive 86Rb+ uptake by only 75 - 80% (Fig. 8, inset).

429
lo i c""
1 I00
5 0 w 1 10 100
I I I I I I I I I 5 10 15 5 10 15
TIME (Minutes)
Fig. 5. The influence of ion deprivation on the Na+/K+/CI- cotransport. Left panel: cells were equilibrated for various periods of time in a Na+-free ( O ) , a ClFfree ( W ) or a K+-free (A) modified Earle's salt solution and then switched for 1 min to a complete Earle's salt solution to measure the initial rate of bumetanide-sensitive 86Rbi uptake. Data are expressed relative to cells that were maintained in a complete Earle's salt solution. Right panel: cells were equilibrated for 15 rnin in a Na+-free (O) , or a CIF-free (H) modified Earle's salt solution and then switched for various periods of time to a complete Earle's salt solution. The activity of the Na+/K+/Cl- cotransport was then assayed using the same medium. Inset: cells were equilibrated for 15 min in modified Earle's salt solution of different Na+ (O) , K' (A) or CIF ( W ) concentrations and then switched to a complete Earle's salt solution for measurement of the activity of the Na+/K+/CIF cotransport. 100% activity on the ordinate scale represents the activity measured after a 15-min equilibration of the cells in a Na+-free, a K+-free or a CI--free modified Earle's salt solution
I I
c
3 x m
9 7 6 5 4 3 -LOG [ COMPOUND (M)]
Fig. 6. The pharmacological properties of the N a + / K + / C l - cotransport of C6 cells. Main panel: dose-response curves for benzmetanide (O) , bumetanide (A), piretanide ( W ) and furosemide (V) inhibition of the initial rate of ouabain insensitive "Rb' uptake by C6 cells. Experiments were performed under iso-osmotic conditions. Inset A: dose-response curves for piretanide inhibition of the rate of 86Rb+ uptake after exposure of the cells to hyperosmotic conditions (540 mOsm/l) (V) and during volume increase following a 15-min incubation of the cells in a Na+-free ( W ) or a K+-free (A) modified Earle's salt solution. Inset B: dose-response curves for bumetanide inhibition of the rate of 86Rbt uptake after exposure of the cells to hyperosmotic conditions (540 mOsm/l) (V) and during volume increase following a 15-min incubation of the cells in a K+-free modified Earle's salt solution (A)
Substituting external C1F by methanesulfonate completely suppressed the bumetanide-sensitive 86Rb+ uptake com- ponent (Fig. 9). The rate of bumetanide-sensitive 86Rbt up- take was linearly related to the extracellular C1- concentration up to 140 mM under all conditions (Fig. 9, main panel). A saturation of the activity of the system with increasing external C1- concentrations was clearly observed when cells were incu- bated in an hypertonic medium that contained 5 mM K', 140 mM Na' and 200 mM N-methylglucamine (Fig. 9, inset). A computer fitting of the experimental data yielded a
value for C1- activation of the cotransport at 120 mM and a Hill coefficient close to 1.
Transport stoichiometry
Bumetanide was also found to decrease the rate of 22Na+ and of 36Cl- uptake by C6 cells. An estimate of the transport stoichiometry of the Na+/K+/Cl- cotransport system was obtained by comparing the initial rates of bumetanide-sensi- tive 22Nat, 36Cl- and 86Rb+ uptake by C6 cells that had
I 0 z
1 10 100 'W oxtarnal K + (mu) A A A .e,
* * v t I
1 10 100
EXTERNAL K + CONCENTRATION (mM)
Fig. 7. The dependence on external K + concentration of the activity of the Na'/K'/Cl- cotransport of C6 cells. Main panel: influence of raising the external K + concentration on the initial rate of bumetanide-sensitive M6Rb+ uptake. Activity was measured under iso-osmotic conditions (0 , A, W ) or hyperosmotic conditions (V). Prior to the uptake experiments, cells were incubated for 15 min in an iso-osmotic Earle's salt solution (O) , a hyperosmotic Earle's salt solution (V) or a Na' -free (A) or a K +-free (m) iso-osmotic Earle's salt solution. Inset: same data as in main panel plotted as percentages of the maximum activity. Maximum activity was determined by com- puter fitting of the experimental data. Hill coefficients of the individ- ual curves were close to I
been preincubated for 15 min in a medium deprived of either Na', C1- or K'. In three experiments the ratio of *'Rb+ to 22Na' uptakes was 1.06 k 0.05. The ratio of 36Cl- to "Rb' uptakes was 2.4 & 0.3. These results are consistent with an electroneutral transport stoichiometry of 1 Na+/ l K+/2 C1- .
Intracellulur p H had no influence on the activity of the Na+/Kf lC1- cotrunsport system
Fig. 10 presents the time course of "Rb' uptake by C6 cells that had been incubated under different conditions of intracellular pH (pHi) and of osmolarity of the incubation media. First, under iso-osmotic conditions, the rate of ouabain-insensitive "Rb' uptake was measured in the ab- sence or the presence of 0.1 mM ethylisopropylamiloride, a potent inhibitor of the Na+/H+ exchange system [13,27,28]. Previous experiments have shown that the pHi of C6 cells was 1.35 and 7.0 in the absence and the presence of ethylisopropylamiloride respectively [I 31. The results present- ed in Fig. 10 show that within this pH range the rates of ouabain-insensitive and bumetanide-sensitive 86Rb' uptake were the same.
We also reported previously that an incubation of C6 cells in an hyperosmotic solution (540 mOsm/l) produced a cell alkalinisation to pHi = 7.70, which was not observed in the presence of ethylisopropylamiloride. Fig. 10 shows that the time course of "Rb+ uptake was nearly identical under hyperosmotic conditions with pHi = 7.7 (i. e. in the absence of ethylisopropylamiloride) and with pHi = 7.0 (i. e. in the presence of ethylisopropylamiloride). This indicated (a) that the activity of the Na'/K+/Cl- cotransport was insensitive to changes in pHi in the range 7.0 - 7.7 and (b) that the time course of activation of the Na+/K+/Cl- cotransport during
. E 100 - E 2
- W Y 3 50 n 3 + P a In m Y 0 u t 4 a
EXTERNAL No + CONCENTRATION (mM)
Fig. 8. The dependence on external Nu+ Concentration of the activity of the Na+/K-/Cl- cotransport qfC6 cells. Inset: dose-response curve for external Na+ activation of the rate of bumetanide-sensitive M6Rb+ uptake. Activity was measured under iso-osmotic conditions. Main panel: dose-response curve for external Na' activation of the rate of bumetanide-sensitive 86Rb+ uptake under hyperosmotic conditions (A) and during volume increase following a 15-min exposure to a Na+-free (U), a K+-free (V) or a Cl--free ( 0 ) modified Earle's salt solution under iso-osmotic conditions or by a 15-min exposure to a Na+-free (H) modified Earle's salt solution under hyperosmotic conditions (540 mOsm/l). Data are expressed as percentages of maximum activity as determined by computer fitting of the exper- imental data. Hill coefficients of the individual curves are close to 1
0 0 , 1
EXTERNAL CI' (mu)
I 50 100 150
EXTERNAL CI CONCENTRATION (mu)
Fig. 9. The dependence on external CI- concentration of the activity of the Na+/K+/CI- cotransport of C6 cells. Main panel: dose-response curve for external C1- activation of the rate of bumetanide-sensitive "Rb+ uptake under iso-osmotic conditions (A), hyperosmotic con- ditions (O) , and during volume increase following a 15-min exposure to a K+-free ( W ) modified Earle's salt solution under iso-osmotic conditions. Inset: dose-response curve for C1- activation of the rate of bumetanide-sensitive M6Rb+ uptake under hypertonic conditions. The uptake solution contained 5 rnM IS+, 140 mM Na', 200 mM N-methylglucamine and varying concentrations of C1- and methane- sulfonate so that the sum of the concentrations of CI- and methane- sulfonate was 345 mM

43 1
5 10
TIME (Mlnuler)
Fig. 10. The time course of s6Rb+ uptake by ouabain-treated C6 cells; influence of ethylisopropylamiloride. Experiments were performed as described in the legend of Fig. 3 in the absence (0, A, 0, V ) or the presence (0 , A, W , V) of 0.1 mM ethylisopropylamiloride. Iso- osmotic conditions in the absence (0 , 0) or the presence ( A , A) of 0.1 mM bumetanide, hyperosmotic conditions in the absence (0, W) or the presence ( V , V) of 0.1 mM bumetanide
an exposure to a hyperosmotic solution was independent of the prior increase in pHi caused by the activation of the Na+/ H+ exchange system.
DISCUSSION
This paper describes the properties of a loop-diuretics- sensitive Na+/K+/CI- cotransport in C6 glial cells. It further shows that although the system is physiologically quiescent under normal conditions, increasing its activity makes it important for controlling the cell volume.
Properties of the Na+lK+/Cl- cotransporl
A Na+/K+/Cl- cotransport system was identified in C6 cells by showing that an important fraction of the K + uptake was suppressed either when external CI- was substituted by methanesulfonate or when cells were incubated in the presence of loop diuretics. The relative potencies of four loop diuretics in their capacity to inhibit 86Rb+ uptake are: benzmetanide > bumetanide > piretanide > furosemide. This order of potency and the apparent affinities for each of these com- pounds are identical to or very similar to those reported for the Na+/K+/Cl- cotransport of renal [25,29,30] and of non- renal cells (31 - 341.
The biochemical properties of the Naf /K +/Cl- cotrans- port of C6 cells can be defined as follows.
a) The activity of the Na+/K+ /Cl- cotransport increases when the external K + concentration is increased (Fig. 7). The Ko.5 value for K+ activation (15 mM) is similar to values that have been reported in other cell types [25, 33, 351.
b) The rate of bumetanide-sensitive "Rb' uptake is linearly related to the external C1- concentration up to 100 mM (Fig. 9). Similarly no saturation of the rate of 86Rb+
uptake with increasing C1- concentrations has been found for human erythrocytes [35] and fibroblasts [36, 371. Saturation was observed in cultured chick cardiac cells = 56 mM [34]) and in MDCK cells = 58 - 70 mM [36]). Saturation could be observed in C6 cells when experiments were performed at higher ionic strenght in the incubation media (Fig. 9, inset). The value for C1- activation of the Na+/ K+/Cl- cotransport was estimated to 120 mM under these conditions.
c) Substituting external Na+ by N-methylglucamine only suppressed 75 - 80% of the rate of bumetanide-sensitive 86Rb' uptake when C6 cells were incubated under iso-os- motic conditions (Fig. 8). An identical result has previously been reported for chick cardiac cells [34] and for macrophage- like cells [39].
In summary the Na+/K+/CI- cotransport in C6 glioma cells has pharmacological and biochemical properties very similar to those of the system in non-cerebral cells.
The role of the Na+/K+/Cl- cotransport in volume regulation
Under physiological conditions of ionic composition and osmolarity of the incubation media, the Na+/K+/Cl- cotransport of C6 cells catalysed equal rates of K + influx and efflux (Fig. 2). In addition we observed that a prolonged exposure of the cells to bumetanide produced no change in the cell volume (Fig. 1). The system is therefore phy- siologically silent under iso-osmotic conditions and does not contribute to the maintenance of the steady-state volume.
Exposure of C6 cells to a hyperosmolar solution leads to a cell shrinking that does not recover (Fig. 1, main panel) in agreement with previous observations [23, 111. This result could suggest that C6 cells lack a volume-sensitive mechanism for swelling after a cell shrinkage. This paper first shows that after a hyperosmotic shrinking, a twofold activation of the Na+/K+/Cl- cotransport is observed. The system then catalyses the net uptake of K, but its activity is not sufficient to promote regulatory volume increase. We further observed that when C6 cells were deprived of either Na+, K + or CI- for 5 - 15 min, a cell shrinkage ensued (Fig. 4). Cell shrinkage was not due to the blockade of the Na+/K+/Cl- cotransport. This is because (a) long exposures to bumetanide do not lead to a cell shrinkage (Fig. 1) and (b) the dependences on external ion concentrations of the activation of the Na+/K+/Cl- cotransport are different from the dependence of the system for K' transport. Although the exact mechanism of cell shrinkage is unknown at the present time, the important point is that ion-deprived C6 cells rapidly recover their volume when switched back to a physiological salt solution. Volume increase is completely prevented in the presence of bumetanide (Fig. 3) and is associated with a selective and large increase in K' uptake through the Na'/K+/CI- cotransport system (Figs 4 and 5). These experiments therefore indicate that the Na+/K+/CI- cotransport, which seems to be quiescent in C6 cells maintained under iso-osmotic or hyperosmotic con- ditions, can contribute to volume regulation when its activity has been increased to a large extent. Whether or not the system is involved in volume regulation seems to be primarily determined by its absolute activity. When the K+-transporting capacity of the Na+/K+/CI- cotransport is low (for instance after a hyperosmotic shock) the activity of the system does not contribute much to the regulation of the cell volume (Fig. 1). When the system is activated to a larger extent (for instance after a period of ion deprivation), it contributes to an increase in the cell volume (Fig. 3).

432
Mechanisms of activation of the Na'/K+/Cl- cotransport
Activation of the Na'/K+/CI- cotransport was observed after a cell shrinkage induced either by an exposure to an hyperosmotic solution or by a period of ion deprivation. There was, however, no relationship between the extent of the cell shrinking produced by the different experimental con- ditions and the extent of the activation of the Na+/K+/Cl- cotransport. This indicates that shrinkage per se is not the trigger for turning on the Na+/K+/Cl- cotransport. The ob- servation that a hyperosmotic shock potentiates the effect of a period of ion deprivation (Fig. 4, inset) suggests that there may be different ways of activating the Na+/K+/Cl- cotransport of C6 cells. One would be induced by an osmotic cell Shrinkage; another would be induced after cell shrinkage consecutive to a period of ion deprivation. The molecular mechanisms of these activations remain unknown.
An interesting feature of C6 cells is that activation of the NaC/Kf/C1- cotransport by hyperosmotic solutions only occurred after a 5-7-min time lag, which suggests a slow activating process. This contrasts with the situation encountered for the Na+/H+ exchange system, another volume-sensitive ion pathway in C6 cells. Activation of the Na+/H + exchange system immediately followed exposure of C6 cells to a hyperosmotic solution [13]. It led to an increased uptake of Na+, when cells were incubated in the presence of ouabain and to a cell alkalinisation of 0.2 pH unit when cells were incubated in the absence of bicarbonate ions [13]. The delayed activation of the Naf /Kf /CI- cotransport that was observed in this study could be consecutive to that of the Na'/H' exchange system. However, we observed that when C6 cells were submitted to a hyperosmotic shock in the pres- ence of ethylisopropylamiloride to block the N a f / H + ex- change system and the resulting cell alkalinisation [13], a stimulation of the Na'/Kf/C1- cotransport was still ob- served with an identical time course (Fig. 10). These obser- vations indicate that activations of the two membrane cation- transporting systems after hyperosmotic cell shrinkage occur independently of each other and that distinct activation path- ways are triggered by the hyperosmotic cell shrinkage.
After activation by whatever means, the properties of in- teraction of the Na+/K ' / C l ~ cotransport with piretanide, bumetanide, external C1- and K + were not modified. Only the external Na+ dependence of the system was different. The rate of bumetanide-sensitive 86Rb + uptake, which is usually partially dependent on external Na', becomes completely dependent on external Na+ (Fig. 8) and the value for Na' activation of the Na+/K+/Cl- cotransport decreased from a value larger than 140 mM to a value of 20-50 mM depending on the conditions used (Fig. 8). The apparent affn- ities for Na' of the system after activation are similar to those found in chick cardiac cells (40 mM [34]) and in mouse 3T3 cells (69 mM [40]). They are lower than the value found in mouse macrophage like cells (83 mM [39]) and much higher than the value found in human erythrocytes (7 mM [35]), in fibroblasts (35-20 mM [33, 371) and in MDCK cells (15- 24 mM [25]). A change in the properties of interaction of the system with Na' cannot account for the differences in activity found in the different experimental conditions that have been studied here.
Hoffmann [I 51 has recently reviewed the different ionic mechanisms that allow vertebrate cells to increase their vol- ume after a cell shrinking. Two types of mechanism were distinguished. In some cell types volume regulation is mediat- ed by the combined operations of the Na+/H+ exchange
system and a Cl-/HCO; exchanger. This situation is ob- served in Amphiuma red blood cells [3] and lymphocytes [41]. In other cell types the cell volume is regulated by the Na+/ K+/Cl- cotransport. This situation is observed in avian red blood cells [42], Ehrlich ascites cells [14], frog skin [16] and also chick cardiac cells [34] C6 cells seem to be unique in that both the Na+/H+ exchange system [I31 and the Na'/K+/Cl- cotransport are activated after exposure to an hyperosmotic solution. The reasons why glial cell volume control is impaired under ischemic conditions are still unknown. This study does show, however, that the Nat/Kf/C1- cotransport exists as a potential volume-regulating mechanism in a physiologically quiescent state in C6 cells. The activity of the system can be unmasked under some experimental conditions. The study of the molecular mechanisms that lead to such an unmasking will likely prove to be valuable for understanding some aspects of astrocytic swelling in brain edema.
This work was supported by grants from the Centre Nationalde la Recherche Scientlfique (LP 7300) and the Fondation pour In Recherche Medicale. We are grateful to Dr D. Duval for the synthesis of ethylisopropylamiloride, to Dr B. Siesjo for his comments on the manuscript, to Dr A. Baethmann for kindly providing a copy of his manuscripts before publication, to Dr J . P. Breittmayer and M. Fehlmann for advice and to Mrs M. T. Ravier for expert technical assistance.
REFERENCES 1. Hoffmann, E. K., Simonsen, L. 0. & Lambert, I. H. (1984) J .
2. Kregenow, F. M. (1981) Annu. Rev. Physiol. 43,493-505. 3. Cala, P. M. (1983) J . Gen. Physiol. 82, 761 -784. 4. Baethmann, A. (1978) Neurosurg. Rev. I , 85-100. 5. Baethmann, A,, Oettinger, W., Rothenfusser, W., Kempski, 0.
Unterberg, A. & Geiger, R. (1980) Adv. Neurol. 28, 171 -195. 6. Kimelberg, H. K. & Frangakis, M. V. (1986) Adv. Biosci. 61,
7. Kempski, O., Zimmer, M., Neu, A., Rosen, F. v., Jansen, M. &
8. Kempski, O., Staub, F., Rosen, F. v., Zimmer, M., Neu, A. &
9. Bourke, R. S., Kimelberg, H. K. & Nelson, L. R. (1976) Brain
10. Bourke, R. S., Kimelberg, H. K. & Daze, M. A. (1978) Brain Res.
11. Kempski, O., Chaussy, L., Gross, U., Zimmer, M. & Baethmann,
12. Kimelberg. H. K., Biddlecome, S. & Bourke, R. S. (1979) Bruin
13. Jean, T., Frelin, C., Vigne, P. & Lazdunski, M. (1986) Eur. J .
14. Hoffmann, E. K. (1982) Philos. Trans. R. Soc. Lond. 299, 519-
15. Hoffmann, E. K. (1986) Biochim. Biophys. Acta864, 1-31. 16. Ussing, H. H. (1982) Acta Physiol. Scund. 114, 363-369. 17. Johnson, J. H., Dunn, D. P. & Rosenberg, R. N. (1982) Biochem.
Biophys. Res. Commun. 109, 100- 105. 18. Bourke, R. S., Kimelberg, H. K., Nelson, L. R., Barron, K. D.,
Auen, E. L., Popp, A. J. & Waldman, J. B. (1980) in Advances in neurology, vol. 28, Brain edema (Cervos-Navarro, J. & Ferszt, R., eds) pp. 99-109, Raven Press, NY.
Memhr. Biol. 78, 211 -222.
177-186.
Baethmann, A. (1987) Stroke 18, in the press.
Baethmann, A. (1987) Neurochem. Pathol. in the press.
Res. 105, 309 - 323.
154, 196-202.
A. (1983) Brain Res. 279, 217-228.
Res. 173., 11 1 - 124.
Biochem. 160,211 -219.
535.
19. Hertz, L. (1981) J. Cereb. Blood Flow Metab. 1 , 143-153. 20. Walz, W. & Hertz, L. (1982) J . Neurochem. 39, 70-77. 21. Walz, W. & Hinks, E. C. (1985) Brain Res. 343, 44-51. 22. Walz, W., Wuttke, W. & Hertz, L. (1984) Brain Res. 292, 367-
23. Kimelberg, H. K. & Frangakis, M. V. (1985) Bruin Res. 361,
24. Cragoe, E. J. Jr., Woltersdorf, 0. W., Bicking, J. B., Kwong, S.
374.
125- 134.
K. &Jones, J. K. (1967) J. Med. Chem. 10, 66-75.

433
25. Rindler, M. H., McRoberts, J. A. & Saier, M. H. (1982) J . Biol.
26. Hartree, E. F. (1972) Anal. Biochem. 48,422-427. 27. Vigne, P., Frelin, C., Cragoe, E. J. & Lazdunski, M. (1983)
28. Vigne, P., Frelin, C., Cragoe, E. J. & Lazdunski, M. (1984) Mol.
29. Schlatter, E., Greger, R. & Weidtke, C. (1983) Pjliigers Arch. Eur.
30. Forbush, B. & Palfrey, H. C. (1983) J . Biol. Chenz. 258, 11 787-
31. Ellory, J. C. & Stewart, G. W. (1982) Br. J . Pharmacol. 75, 183-
32. Palfrey, H. C., Silva, P. & Epstein, F. H. (1984) Am. J. Physiol.
33. Owen, N. E. & Prastein, M. L. (1985) 1. Biol. Chem. 260, 1445-
Chem. 257,2254- 2259.
Biochem. Biophys. Res. Commun. 116, 86 - 90.
Pharmacol. 25,13 1 - 136.
J . Physiol. 3Y6, 210-217.
11 792.
188.
246, C242 - C246.
1451.
34. Frelin, C., Chdssande, 0. & Lazdunski, M. (1986) Biochem. Bio-
35. Dunham, P. B., Stewart, G. W. & Ellory, J. C. (1980) Proc. Nut1
36. O’Brien, T. G. & Krzeminski, K. (1983) Proc. Nut1 Acad. Sci.
37. Paris, S. & Pouyssegur, J. (1986) J . Biol. Chem. 261, 6177-6183. 38. McRoberts, J. A., Erlinger, S., Rindler, M. J. & Saier, M. H.
39. Bourrit, A,, Atlan, H., Fromer, I., Melmed, R. N. & Lichtstein,
40. Atlan, H., Snyder, D. & Panet, R. (1984) J . Membr. Biol. 81,
41. Grinstein, S., Clarke, C. A. &Rothstein, A. (1983) J. Gen. Physiol.
42. Haas, M., Schmidt, W. F. & Macmanus, T. J. (1982) J. Gen.
phys. Res. Commun. 134, 326-331.
Acad. Sci. USA 77, 171 1 - 1715.
USA 80,4334-4338.
(1982) J . Biol. Chem. 257,2260-2266.
D. (1985) Biochim. Biophys. Acta 817, 85-94.
181 - 188.
82,619-638.
Physiol. 80, 125 - 147.




















