
Danthron Triggers ROS and Mitochondria-Mediated Apoptotic Death in C6 Rat Glioma Cells Through...
11
ORIGINAL PAPER Danthron Triggers ROS and Mitochondria-Mediated Apoptotic Death in C6 Rat Glioma Cells Through Caspase Cascades, Apoptosis-Inducing Factor and Endonuclease G Multiple Signaling Shang-Ming Chiou • Chiz-Hao Chiu • Su-Tso Yang • Jai-Sing Yang • Hui-Ying Huang • Chao-Lin Kuo • Po-Yuan Chen • Jing-Gung Chung Received: 26 January 2012 / Revised: 2 April 2012 / Accepted: 27 April 2012 / Published online: 17 May 2012 Ó Springer Science+Business Media, LLC 2012 Abstract This research focused on the induction of cytotoxic effects by danthron, a natural anthraquinone derivative on C6 rat glioma cells through exploring the means of cell death and the effects on mitochondrial function. We found that danthron decreased the percentage of viable C6 cells and induced cell morphological changes in a dose-and time-dependent manner. The morphological and nuclei changes (DAPI staining) in C6 cells were observed using a contrast-microscope and fluorescence microscopy, respectively. The results suggest that cell death of C6 cells which are induced by danthron is closely related to apoptotic death. Danthron decreased the level of mitochondrial membrane potential (DW m ), stimulated the release of cytochrome c from mitochondria to cytosol and promoted the levels of caspase-9 and caspase-3, or induced the release of AIF and Endo G from mitochondria. Based on both observations, we suggest that the danthron-pro- voked apoptotic death of C6 cells is mediated through the mitochondria-dependent pathway. Furthermore, our results also indicated that danthron triggered apoptosis through reactive oxygen species (ROS) production which were increased after 1h exposure of danthron, which was reversed by the ROS scavenger N-acetyl-L-cysteine (NAC). As a consequence, danthron-mediated cell death of C6 cells via ROS production, mitochondrial transmembrane poten- tial collapse and releases of cytochrome c, AIF and Endo G. Taken together, danthron was demonstrated to be Po-Yuan Chen and Jing-Gung Chung made equal contribution to this work. S.-M. Chiou Department of Functional Neurosurgery, Gamma Knife Center, China Medical University Hospital, Taichung 404, Taiwan C.-H. Chiu P.-Y. Chen J.-G. Chung Department of Biological Science and Technology, China Medical University, Taichung 404, Taiwan S.-T. Yang Department of Radiology, China Medical University Hospital, Taichung 404, Taiwan S.-T. Yang School of Chinese Medicine, China Medical University, Taichung 404, Taiwan J.-S. Yang Department of Pharmacology, China Medical University, Taichung 404, Taiwan H.-Y. Huang Department of Nutrition, China Medical University, Taichung 404, Taiwan C.-L. Kuo School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, China Medical University, Taichung 404, Taiwan P.-Y. Chen (&) J.-G. Chung (&) Department of Biological Science and Technology, China Medical University, No 91, Hsueh-Shih Road, Taichung 40402, Taiwan e-mail: [email protected] J.-G. Chung e-mail: [email protected] J.-G. Chung Department of Biotechnology, Asia University, Taichung 413, Taiwan 123 Neurochem Res (2012) 37:1790–1800 DOI 10.1007/s11064-012-0792-3
Transcript of Danthron Triggers ROS and Mitochondria-Mediated Apoptotic Death in C6 Rat Glioma Cells Through...

ORIGINAL PAPER
Danthron Triggers ROS and Mitochondria-Mediated ApoptoticDeath in C6 Rat Glioma Cells Through Caspase Cascades,Apoptosis-Inducing Factor and Endonuclease G MultipleSignaling
Shang-Ming Chiou • Chiz-Hao Chiu •
Su-Tso Yang • Jai-Sing Yang • Hui-Ying Huang •
Chao-Lin Kuo • Po-Yuan Chen • Jing-Gung Chung
Received: 26 January 2012 / Revised: 2 April 2012 / Accepted: 27 April 2012 / Published online: 17 May 2012
� Springer Science+Business Media, LLC 2012
Abstract This research focused on the induction of
cytotoxic effects by danthron, a natural anthraquinone
derivative on C6 rat glioma cells through exploring the
means of cell death and the effects on mitochondrial
function. We found that danthron decreased the percentage
of viable C6 cells and induced cell morphological changes
in a dose-and time-dependent manner. The morphological
and nuclei changes (DAPI staining) in C6 cells were
observed using a contrast-microscope and fluorescence
microscopy, respectively. The results suggest that cell
death of C6 cells which are induced by danthron is closely
related to apoptotic death. Danthron decreased the level of
mitochondrial membrane potential (DWm), stimulated the
release of cytochrome c from mitochondria to cytosol and
promoted the levels of caspase-9 and caspase-3, or induced
the release of AIF and Endo G from mitochondria. Based
on both observations, we suggest that the danthron-pro-
voked apoptotic death of C6 cells is mediated through the
mitochondria-dependent pathway. Furthermore, our results
also indicated that danthron triggered apoptosis through
reactive oxygen species (ROS) production which were
increased after 1 h exposure of danthron, which was
reversed by the ROS scavenger N-acetyl-L-cysteine (NAC).
As a consequence, danthron-mediated cell death of C6 cells
via ROS production, mitochondrial transmembrane poten-
tial collapse and releases of cytochrome c, AIF and Endo
G. Taken together, danthron was demonstrated to bePo-Yuan Chen and Jing-Gung Chung made equal contribution to this
work.
S.-M. Chiou
Department of Functional Neurosurgery, Gamma Knife Center,
China Medical University Hospital, Taichung 404, Taiwan
C.-H. Chiu � P.-Y. Chen � J.-G. Chung
Department of Biological Science and Technology,
China Medical University, Taichung 404, Taiwan
S.-T. Yang
Department of Radiology, China Medical University Hospital,
Taichung 404, Taiwan
S.-T. Yang
School of Chinese Medicine, China Medical University,
Taichung 404, Taiwan
J.-S. Yang
Department of Pharmacology,
China Medical University, Taichung 404, Taiwan
H.-Y. Huang
Department of Nutrition, China Medical University,
Taichung 404, Taiwan
C.-L. Kuo
School of Chinese Pharmaceutical Sciences and Chinese
Medicine Resources, China Medical University, Taichung 404,
Taiwan
P.-Y. Chen (&) � J.-G. Chung (&)
Department of Biological Science and Technology,
China Medical University, No 91, Hsueh-Shih Road,
Taichung 40402, Taiwan
e-mail: [email protected]
J.-G. Chung
e-mail: [email protected]
J.-G. Chung
Department of Biotechnology, Asia University,
Taichung 413, Taiwan
123
Neurochem Res (2012) 37:1790–1800
DOI 10.1007/s11064-012-0792-3

effective in killing C6 rat glioma cells via the ROS-pro-
moted and mitochondria-dependent apoptotic pathways.
Keywords Danthron � C6 rat glioma cells � Apoptosis �ROS � Caspase cascades � Mitochondria
Abbreviations
DWm Mitochondrial membrane potential
AIF Apoptosis-inducing factor
DAPI 40,6-Diamidino-2-phenylindole
DMSO Dimethyl sulfoxide
Endo G Endonuclease G
FBS Fetal bovine serum
GRP78 Glucose-regulated proteins 78
PARP Poly ADP ribose polymerase
PBS Phosphate buffer saline
PI Propidium iodide
ROS Reactive oxygen species
SDS-PAGE Sodium dodecylsulfate polyacrylamide gel
electrophoresis
Introduction
Brain tumor is one of the most threatening malignant
cancers in humans today and notoriously hard to treat.
Glioma is a tumor of dismal prognosis and ultimately
progresses to form high-grade malignancies with local
infiltration of the brain [1, 2]. Unfortunately, high-grade
glioma is difficult and their prognosis is low. The treatment
of high-grade glioma is limited for resection followed by
radio/chemotherapy, resulting in a median survival of
14 months [3]. Currently, the treatments for glioma include
surgery, radiotherapy, chemotherapy or a combination of
multiple modalities but the results are still not satisfactory.
The new for development of new treatment strategies for
glioma patients is urgent. The current clinical anticancer
drugs have been demonstrated to cause cell death through
interfering the processes of the cell cycle and induction of
apoptosis [4]. Therefore, one of the best strategies for
chemoprevention or chemotherapy of agents is to induce
apoptosis in cancer cells [5–9].
Danthron (1,8-dihydroxyanthraquinone), an anthraqui-
none derivative, is a possible human carcinogen (IARC
2B), and has been withdrawn from the market as a laxative
by the US Food and Drug Administration (FDA) [10–13].
Recently, it was reported that the oxidative stress may be a
major contributing pathway in the genotoxicity of danthron
[14]. It was reported that danthron functions as a retinoic X
receptor antagonist by stabilizing tetramers of the receptor
[15]. In our laboratory, we have reported that danthron not
only induced DNA damage and inhibited DNA repair
genes expression [16] but triggered apoptosis [17] in
human brain glioblastoma multiforms GBM 8401 cancer
cells. Our report also found that danthron-induced DNA
damage and apoptotic cell death was involved in mito-
chondrial depolarization and caused the activation of cas-
pase cascades in SNU-1 human gastric cancer cells [5].
Until now, there is no information to address danthron-
induced cell death (apoptosis) in C6 rat glioma cells. In the
present study, we try to examine whether danthron inhibits
the growth and induces apoptosis in C6 cells. Our finding
provides insight into the mechanism of danthron-induced
mitochondria-mediated apoptosis via caspase cascades,
apoptosis-inducing factor (AIF) and endonuclease G (Endo
G)-dependent pathways in C6 cells in vitro.
Materials and Methods
Chemicals and Reagents
Danthron, dimethyl sulfoxide (DMSO), propidium iodide
(PI) and agarose were purchased from Sigma-Aldrich
Corp. (St. Louis, MO, USA). The primary antibodies were
obtained as follows: antibodies for BAX, BCL-2, cyto-
chrome c, APAF-1, AIF, Endo G, caspase-3, -8 and -9,
PARP, p53, GRP78, GADD153, caspase-12 and b-Actin
were purchased from Cell Signaling Technology (Beverly,
MA, USA) and the secondary antibodies, which were
horseradish peroxidase (HRP)-linked goat anti-mouse IgG
and goat anti-rabbit IgG were purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA). The protein assay
kit was purchased from Bio-Rad (Hercules, CA, USA).
Cell Culture
C6 rat glioma cell line was obtained from the Food
Industry Research and Development Institute (Hsinchu,
Taiwan). The culture medium consisted of Kaighn’s
modification of Ham’s F-12 (KMHF) medium, 10 %
fetal bovine serum (FBS), 1 % penicillin–streptomycin
(100 Units/ml penicillin and 100 lg/ml streptomycin) and
2 mM L-glutamine which were obtained from Invitrogen
Life Technologies (Carlsbad, CA, USA). C6 cells were
cultured in KMHF medium as monolayers and maintained
in an incubator with a humidified atmosphere of 95 % air
and 5 % CO2 at 37 �C [8].
Danthron Treatment
In all treatments of experiments, danthron was initially
dissolved in DMSO and made up to the maximum final
concentration of 0.2 % (v/v) in the complete cell culture
medium (KMHF). The sub-cultured cells from experiments
were treated with either varying concentrations of danthron
Neurochem Res (2012) 37:1790–1800 1791
123

or 0.2 % DMSO in media alone that served as control
[5, 17].
Assessment of Cell Morphology and Viability
Approximately 2 9 105 cells/well of C6 cells were cul-
tured onto 12-well plates in KMHF culture medium and
each well was then treated individually with 0, 10, 25, 50,
75 and 100 lM of danthron for 24, 48 and 72 h. For cell
morphological changes examination, cells onto the plate
were directly examined and were photographed under a
phase-contrast microscope [18]. For determination of the
percentage of viable cells, cells were collected from each
well and added PBS containing PI (4 lg/ml). Thereafter,
nonviable cells were stained by a PI dye exclusion
(indicative of an intact membrane) and displayed brighter
fluorescence than the unstained (viable cells). Cells were
immediately determined and counted by flow cytometric
analysis (BD Biosciences, FACSCalibur, Franklin Lakes,
NJ, USA) as previously described [19–21].
DAPI Staining for Apoptosis
Approximately 2 9 105 cells/well of C6 cells were placed
in 12-well plates with KMHF medium as described above
for 24 h. Cells were treated individually with 0, 10, 25, 50,
75 and 100 lM of danthron for a 48-h incubation. Cells
were stained by using 6-diamidino-2-phenylindole dihy-
drochloride (DAPI, Invitrogen Life Technologies) as
described previously [22, 23]. The cells were examined and
were photographed under a fluorescence microscope.
DNA Fragmentation by Agarose Gel Electrophoresis
A total 5 9 105 C6 cells/well on 6-well plates were cul-
tured with or without 50 lM of danthron for 48 h. DNA
was isolated using DNA Purification kit (Genemark tech-
nology Co., Ltd, Tainan, Taiwan) and ladder formation
assays were performed by DNA agarose gel electrophoresis
as described previously [24, 25].
Determinations for Reactive Oxygen Species (ROS),
Mitochondrial Membrane Potential (DWm) and Ca2?
Release
A density of 2 9 105 cells/well of C6 cells were placed
onto 12-well plates and treated with 50 lM danthron at
37 �C, 5 % CO2 and 95 % air for 0, 1, 3, 6, 12 and 24 h to
detect the changes of ROS, DWm and Ca2? release. Cells
were harvested from each treatment by centrifugation
(1,500 rpm) and then re-suspended in 500 ll of DCFH-DA
(10 lM) for ROS determination, in 500 ll of DiOC6
(100 nmol/l) for the levels of DWm and in 500 ll of Fluo-3
(2.5 lg/ml) for Ca2? release at 37 �C for 30 min and
immediately analyzed by flow cytometry [26–28]. Or cells
were pretreated with 20 mM NAC (a ROS scavenger,
Sigma-Aldrich Corp.) and then were treated with or with-
out danthron for 24 h before cells were harvested for
measuring the percentage of viable cells using a PI
exclusion method and flow cytometry as described above.
Apoptosis-Associated Proteins Levels Utilizing
Western Blotting Analysis
C6 cells at the density of 5 9 105 cells/well were placed
onto 6-well plates and treated individually with 50 lM
danthron for 0, 6, 12, 24 and 48 h. Cells were harvested
from each treatment for the total protein determination and
then for Western blotting analysis. The protein levels of
BAX, BCL-2, cytochrome c, APAF-1, AIF, Endo G, cas-
pase-3, -8 and -9, PARP, p53, GRP78, GADD153, caspase-
12 and b-Actin expressions were examined using sodium
dodecylsulfate polyacrylamide gel electrophoresis (SDS-
PAGE) and Western blotting as described previously
[5, 19, 20].
Caspase-3, -8 and -9 mRNA Gene Expressions
by Real-Time PCR
C6 cells (5 9 105 cells/well) were plated in 6-well plates
and placed with KMHF medium containing with or without
50 lM danthron for a 24-h treatment. Cells were then
collected and total RNA was isolated using the Qiagen
RNeasy Mini Kit (Qiagen, Valencia, CA, USA). RNA
samples were reverse-transcribed with High Capacity
cDNA Reverse Transcription Kit according to the standard
protocol of the supplier (Applied Biosystems, Foster City,
CA, USA). Quantitative PCR was done, and 29 SYBR
Green PCR Master Mix (Applied Biosystems) and 200 nM
of forward and reverse primers (rat caspase-3: F-AG
GCCGACTTCCTGTATGCTT, R-AGCATGGCGCAAA
GTGACT; rat caspase-8: F-TCCTTAGTTCTCTCAGTT
GCCTTTC, R-CCGGGCTCACTTCCTGTTC; rat cas-
pase-9: F-CTGTCCCGTGAAGCAAGGAT, R-CCCACG
TCTCCTCCAACCT; rat GAPDH: F-GGTGGACCTCAT
GGCCTACA, R-CAGCAACTGAGGGCCTCTCT) were
applied as previously described [29, 30]. Each assay was
run on an Applied Biosystems 7300 Real-Time PCR sys-
tem in triplicate and expression fold-changes were derived
using the comparative CT method.
Confocal Laser Scanning Microscopy for Protein
Translocation
C6 cells at a density of 1 9 105 cells/well were cultured on
four-well chamber slides for treatment without or with
1792 Neurochem Res (2012) 37:1790–1800
123
50 lM danthron for 24 h. Then cells on the slides were
fixed in 4 % formaldehyde in PBS for 15 min, permeabi-
lized with 0.3 % Triton-X 100 in PBS for 1 h with
blocking of nonspecific binding sites using 2 % BSA as
described previously [31]. Primary antibodies against AIF,
Endo G and cytochrome c (1:100 dilution) (green fluores-
cence) were used to stain the fixed cells overnight, washed
twice with PBS before being stained with secondary anti-
body (FITC-conjugated goat anti-mouse IgG at 1:100
dilution), and followed by DNA and mitochondria staining
with PI (red fluorescence) and MitoTracker (Invitrogen
Life Technologies) (red fluorescence). All samples were
microphotographed using a Leica TCS SP2 Confocal
Spectral Microscope as described previously [18, 31].
Statistical Analysis
The statistical analyses of the data were represented as
means ± SD in triplicate (n = 3) and one-way ANOVA
followed by the Dunnett’s test was used to analyze dif-
ferences between danthron-treated and control groups. The
p value of less than 0.05 was considered a significant
difference.
Results
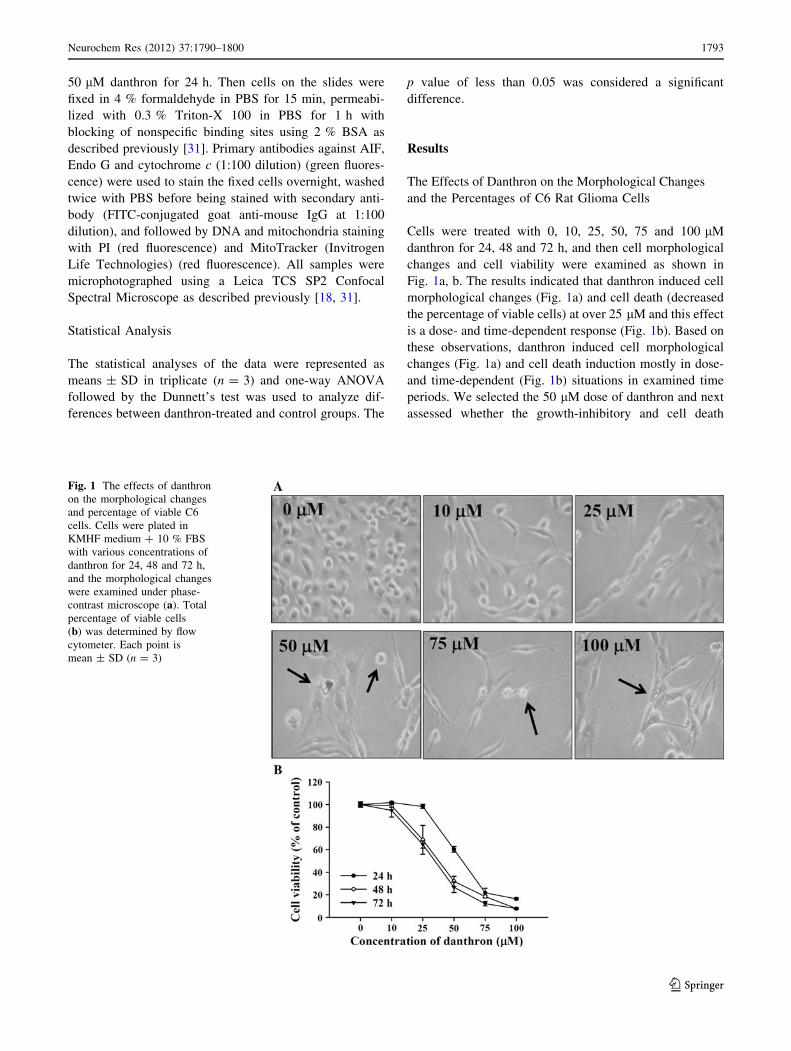
The Effects of Danthron on the Morphological Changes
and the Percentages of C6 Rat Glioma Cells
Cells were treated with 0, 10, 25, 50, 75 and 100 lM
danthron for 24, 48 and 72 h, and then cell morphological
changes and cell viability were examined as shown in
Fig. 1a, b. The results indicated that danthron induced cell
morphological changes (Fig. 1a) and cell death (decreased
the percentage of viable cells) at over 25 lM and this effect
is a dose- and time-dependent response (Fig. 1b). Based on
these observations, danthron induced cell morphological
changes (Fig. 1a) and cell death induction mostly in dose-
and time-dependent (Fig. 1b) situations in examined time
periods. We selected the 50 lM dose of danthron and next
assessed whether the growth-inhibitory and cell death
Fig. 1 The effects of danthron
on the morphological changes
and percentage of viable C6
cells. Cells were plated in
KMHF medium ? 10 % FBS
with various concentrations of
danthron for 24, 48 and 72 h,
and the morphological changes
were examined under phase-
contrast microscope (a). Total
percentage of viable cells
(b) was determined by flow
cytometer. Each point is
mean ± SD (n = 3)
Neurochem Res (2012) 37:1790–1800 1793
123
effects of danthron are accompanied by its effect on
apoptotic cell death.
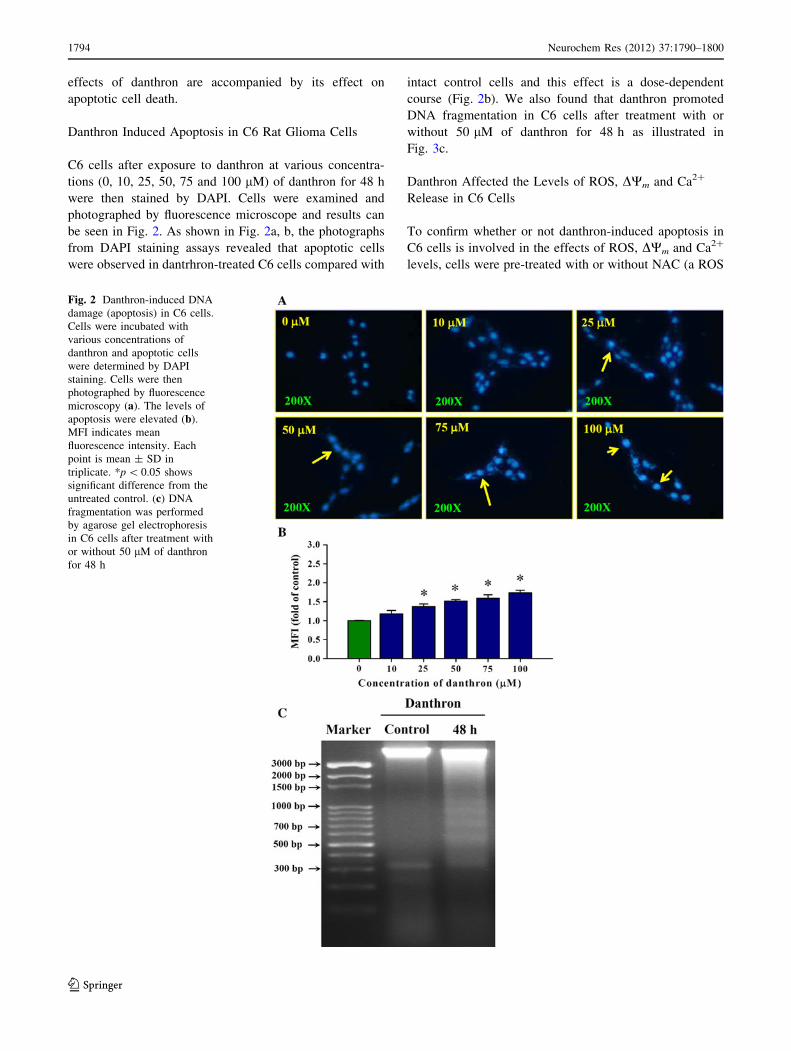
Danthron Induced Apoptosis in C6 Rat Glioma Cells
C6 cells after exposure to danthron at various concentra-
tions (0, 10, 25, 50, 75 and 100 lM) of danthron for 48 h
were then stained by DAPI. Cells were examined and
photographed by fluorescence microscope and results can
be seen in Fig. 2. As shown in Fig. 2a, b, the photographs
from DAPI staining assays revealed that apoptotic cells
were observed in dantrhron-treated C6 cells compared with
intact control cells and this effect is a dose-dependent
course (Fig. 2b). We also found that danthron promoted
DNA fragmentation in C6 cells after treatment with or
without 50 lM of danthron for 48 h as illustrated in
Fig. 3c.
Danthron Affected the Levels of ROS, DWm and Ca2?
Release in C6 Cells
To confirm whether or not danthron-induced apoptosis in
C6 cells is involved in the effects of ROS, DWm and Ca2?
levels, cells were pre-treated with or without NAC (a ROS
Fig. 2 Danthron-induced DNA
damage (apoptosis) in C6 cells.
Cells were incubated with
various concentrations of
danthron and apoptotic cells
were determined by DAPI
staining. Cells were then
photographed by fluorescence
microscopy (a). The levels of
apoptosis were elevated (b).
MFI indicates mean
fluorescence intensity. Each
point is mean ± SD in
triplicate. *p \ 0.05 shows
significant difference from the
untreated control. (c) DNA
fragmentation was performed
by agarose gel electrophoresis
in C6 cells after treatment with
or without 50 lM of danthron
for 48 h
1794 Neurochem Res (2012) 37:1790–1800
123

scavenger) and exposed to 50 lM danthrone for various
time periods. The levels of ROS production were analyzed
and quantified by flow cytometry. The results from Fig. 3a
demonstrated that danthron induced ROS production quite
early and time-dependently and pretreatment with NAC led
to decrease the dead cells of C6 cells after treatment with
danthron (Fig. 3d). Figure 3b shows that danthron pro-
moted the loss of DWm in C6 cells and this effect also is a
time-dependent manner (Fig. 3b). The results from Fig. 3c
indicated that danthron induced Ca2? production was sig-
nificantly increased from 1 h treatment and up to 24 h to
produce high levels of Ca2? (Fig. 3c).
Danthron Altered the Apoptosis-Associated Proteins
and Gene Levels in C6 Rat Glioma Cells
Our data in Figs. 1 and 2 already indicated that danthron
induced cell death of C6 cells (apoptotic death). For
Fig. 3 Danthron affected the
levels of ROS, DWm and Ca2?
in C6 cells. Cells were
incubated with 50 lM danthron
for 0, 1, 3, 6, 12 and 24 h and/or
pre-treated with NAC before
stained by DCFH-DA for ROS
levels determination (a) after
treatment, stained with DiOC6
for the DWm level determination
(b) and stained by Fluo-3/AM
and the Ca2? level
(c) determined as described in
‘‘Materials and Methods’’. Cells
were harvested for the
determination of percentage of
viable cells (d). Each point is
mean ± SD of three
experiments. *Values are
significantly different from the
untreated control (p \ 0.05)
Neurochem Res (2012) 37:1790–1800 1795
123

determining the signaling pathways of danthron-induced
apoptosis of C6 cells via a mitochondria-dependent
pathway, cells were treated with 50 lM danthron for
various time periods, harvested and measured for the total
protein levels from each treatment using Western blotting
analysis. The results are shown in Fig. 4, which indicated
that danthron promoted the expressions of BAX (Fig. 4a),
cytochrome c, APAF-1, AIF and Endo G (Fig. 4b), cas-
pase-3, -8 and -9 and p53 (Fig. 4c), GRP78, GADD153
and caspase-12 (Fig. 4d), but it decreased the level of
BCL-2 (Fig. 4a). As shown in Fig. 4c, treatment of cells
with danthron resulted in a time-dependent increase in
caspase-3, caspase-8, and caspase-9 with strongest effects
at 12, 24 and 48 h, suggesting a possible involvement of
caspases activation in the apoptotic effect of danthron in
C6 cells in vitro. Alternatively, it can be seen in Fig. 4e
that danthron treatment for 24 h led to a significant
increase for caspase-3, -8 and -9 gene expressions in C6
cells.
Danthron Trafficked the Proteins Translocation in C6
Rat Glioma Cells
Cells were cultured on 4-well chamber slides and were then
treated with or without 50 lM danthron for 24 h and then
were individually stained by anti-AIF, Endo G and cyto-
chrome c. The results shown in Fig. 5 indicated that dan-
thron promoted the expression of AIF (Fig. 5a), Endo G
(Fig. 5b) and cytochrome c (Fig. 5c) and all samples also
translocated from mitochondria to nuclei or cytoplasma in
C6 cells.
Discussion
Although it has been shown that danthron induced geno-
toxicity and has been reported by the FDA [12, 13], our
previous studies do show that danthron can induce apop-
tosis in cancer cell lines [5, 16, 17, 32, 33]. Our earlier
Fig. 4 Danthron altered the
apoptosis-associated proteins
levels in C6 cells. A total of
5 9 105 C6 cells/ml cells were
treated with 50 lM danthron for
0, 6, 12, 24 and 48 h. Cells were
harvested from each sample and
associated proteins were
determined by Western blotting.
The levels of BAX and BCL-2
(a), cytochrome c, APAF-1, AIF
and Endo G (b), caspase-3, -8
and -9, PARP and p53 (c),
GRP78, GADD153 and
caspase-12 (d) expressions were
examined using sodium
dodecylsulfate polyacrylamide
gel electrophoresis (SDS-
PAGE) and Western blotting.
(e) Cells were individually
treated with 50 lM danthron for
24 h, and then cells were
harvested for real-time PCR to
determine the gene expression
of caspase-3, -8 and -9 as
described in ‘‘Materials and
Methods’’. *p \ 0.05 indicates
significant difference between
control and treated cells
1796 Neurochem Res (2012) 37:1790–1800
123
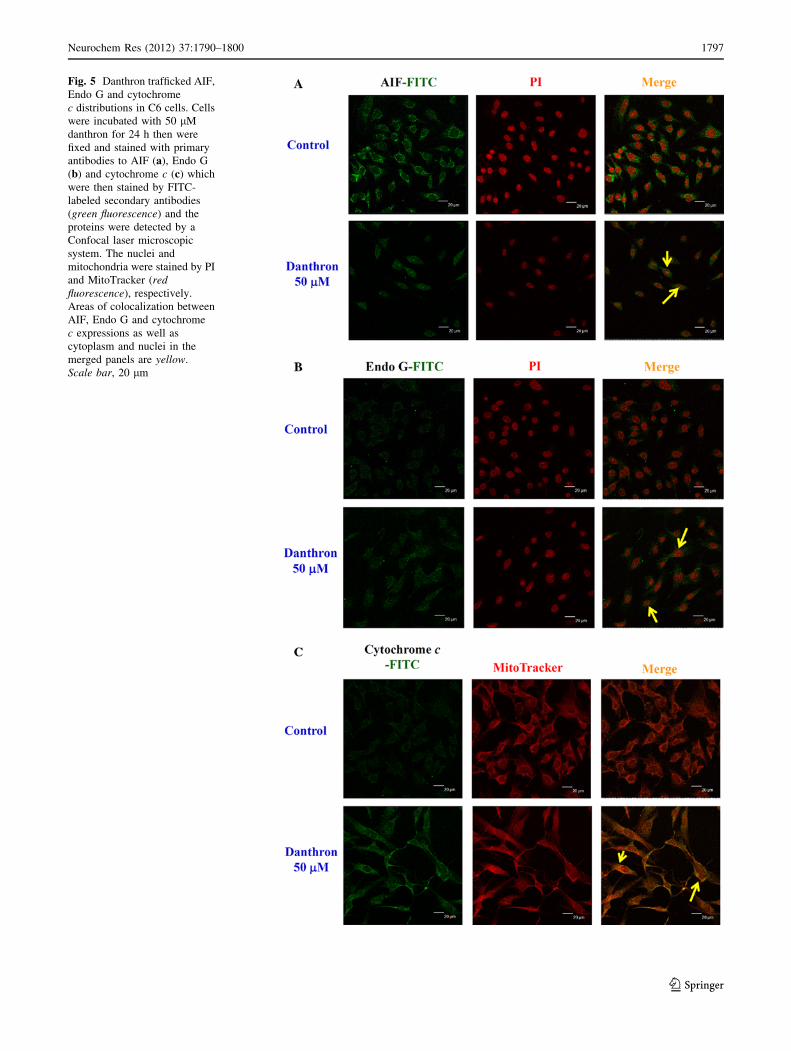
Fig. 5 Danthron trafficked AIF,
Endo G and cytochrome
c distributions in C6 cells. Cells
were incubated with 50 lM
danthron for 24 h then were
fixed and stained with primary
antibodies to AIF (a), Endo G
(b) and cytochrome c (c) which
were then stained by FITC-
labeled secondary antibodies
(green fluorescence) and the
proteins were detected by a
Confocal laser microscopic
system. The nuclei and
mitochondria were stained by PI
and MitoTracker (redfluorescence), respectively.
Areas of colocalization between
AIF, Endo G and cytochrome
c expressions as well as
cytoplasm and nuclei in the
merged panels are yellow.
Scale bar, 20 lm
Neurochem Res (2012) 37:1790–1800 1797
123

study has also demonstrated that danthron at 10–75 lM
had no significant effect on cell viability in rat embryo
aortic smooth muscle cell line (A10), normal fetal osteo-
blast cell line (hFOB) and normal hepatocyte cell line
(Chang liver). We found that no significantly adverse effect
levels of renal, hepatic and immune response in danthron-
treated nude mice at the lower dose [5].
The present study herein provides evidence that dan-
thron-induced apoptosis in C6 rat glioma cells is involved
in the mitochondria-dependent pathways. Furthermore, the
data suggest that induction of cytotoxicity is via apoptotic
death in C6 cells (Fig. 2). The results from flow cytometric
assay already showed that danthron promoted the ROS
production in C6 cells (Fig. 3a). The ROS generation is a
common hall mark for cancer cells due to cancer cells
response to oxidative stress which led to ROS production
[34, 35]. Furthermore, over production of ROS could affect
the levels of DWm accompanying with mitochondrial-
associated event including the apoptosis [36, 37]. Our
results as shown in Fig. 4b also demonstrated that danthron
decreased the level of DWm and these effects are time-
dependent. It was reported that mitochondria plays a crit-
ical role in apoptotic cell death [38]. Thus, mitochondrial
target is becoming the approach to screen therapeutic
agents against cancer [39–41].
It was reported that the accumulation of ROS could lead
to the collapse of mitochondrial membrane potential in
cancer cells, leading to the release of cytochrome c [42]
even AIF and Endo G releases from mitochondria and
finally cause cell apoptosis [39, 43]. To confirm if ROS is
one of the reasons for induction of cell death in C6 cells
after exposure to danthron, NAC was used to pretreat the
C6 cells and then followed by danthron treatment. Cells
were then harvested for the measurements of viable cells
in comparison to danthron treatment alone sample. The
results showed that NAC, a ROS scavenger, significantly
reduced danthron-induced cell death (Fig. 3d), and pro-
tected in vitro mitochondria from depolarization. Our data
showed that ROS was involved and may play a critical role
in danthron-induced apoptosis of C6 cells.
We also examined the apoptosis-associated proteins
levels in C6 cells after exposure to danthron. Results in
Fig. 4a indicated that danthron up-regulated the levels of
pro-apoptotic protein (BAX) but down-regulated the level
of anti-apoptotic protein (BCL-2). BAX and BCL-2 pro-
teins belong to the BCL-2 family and both proteins play an
important role in mitochondrial death cascade [5, 44] and
the ratio of BAX/BCL-2 proteins have been reported to
affect the levels of DWm in several cancer cell lines [6, 45].
BAX translocates to the mitochondria and integrates into
Fig. 6 Schematic diagram
showed danthron-induced
signaling on the activation of
apoptotic machinery in C6 rat
glioma cells
1798 Neurochem Res (2012) 37:1790–1800
123

the outer mitochondrial membrane for causing the disrup-
tion of DWm before leading to the release of cytochrome
c into the cytosol followed by the activation of caspase-9
and -3 [46, 47], but the release of AIF and Endo G from
mitochondria into the nuclei caused apoptosis [5, 48].
However, BCL-2 prevents the process of BAX function via
preserving mitochondrial integrity. Therefore, the ratio of
BAX/BCL-2 has been demonstrated to play a crucial role
for the sustenance of drug-induced apoptosis in the mito-
chondria-mediated apoptotic pathway [6, 45].
Furthermore, the results from Fig. 4b demonstrated that
danthron promoted the levels of cytochrome c, AIF and
Endo G in C6 cells. We also used confocal laser micro-
scope to examine the translocations of cytochrome c, AIF
and Endo G and results. Our data showed that danthron
promoted the releases of cytochrome c, AIF and Endo G in
C6 cells (Fig. 5). Based on these actions, danthron-induced
apoptosis of C6 cells is mediated through mitochondria-
dependent pathway. The possible signal pathways of dan-
thron-induced apoptosis in C6 rat glioma cells was sum-
marized in Fig. 6, which indicated that danthron induced
ROS production, dysfunction of mitochondria, and then led
to the releases of cytochrome c, AIF and Endo G during
cell apoptosis.
Acknowledgments This study was supported by the grant
CMU100-TC-05 from China Medical University and by a research
grant from the National Science Council of Republic of China (Tai-
wan) (NSC 97-2815-C-039-034-B).
References
1. Giese A, Westphal M (2001) Treatment of malignant glioma: a
problem beyond the margins of resection. J Cancer Res Clin
Oncol 127:217–225
2. Wen PY, Kesari S (2008) Malignant gliomas in adults. N Engl J
Med 359:492–507
3. Lefranc F, Rynkowski M, DeWitte O, Kiss R (2009) Present and
potential future adjuvant issues in high-grade astrocytic glioma
treatment. Adv Tech Stand Neurosurg 34:3–35
4. Kao ST, Yeh CC, Hsieh CC, Yang MD, Lee MR, Liu HS et al
(2001) The Chinese medicine Bu-Zhong-Yi-Qi-Tang inhibited
proliferation of hepatoma cell lines by inducing apoptosis via G0/
G1 arrest. Life Sci 69:1485–1496
5. Chiang JH, Yang JS, Ma CY, Yang MD, Huang HY, Hsia TC
et al (2011) Danthron, an anthraquinone derivative, induces DNA
damage and caspase cascades-mediated apoptosis in SNU-1
human gastric cancer cells through mitochondrial permeability
transition pores and Bax-triggered pathways. Chem Res Toxicol
24:20–29
6. Sawada M, Nakashima S, Banno Y, Yamakawa H, Hayashi K,
Takenaka K et al (2000) Ordering of ceramide formation, caspase
activation, and Bax/Bcl-2 expression during etoposide-induced
apoptosis in C6 glioma cells. Cell Death Differ 7:761–772
7. Kuo TC, Yang JS, Lin MW, Hsu SC, Lin JJ, Lin HJ et al (2009)
Emodin has cytotoxic and protective effects in rat C6 glioma
cells: roles of Mdr1a and nuclear factor kappaB in cell survival.
J Pharmacol Exp Ther 330:736–744
8. Chen TC, Lai KC, Yang JS, Liao CL, Hsia TC, Chen GW et al
(2009) Involvement of reactive oxygen species and caspase-
dependent pathway in berberine-induced cell cycle arrest and
apoptosis in C6 rat glioma cells. Int J Oncol 34:1681–1690
9. Lung FD, Tsai JY (2003) Grb2 SH2 domain-binding peptide
analogs as potential anticancer agents. Biopolymers 71:132–140
10. (2011) Danthron Rep Carcinog 128–129
11. (2002) Danthron (1,8-dihydroxyanthraquinone). Rep Carcinog
10:76–77
12. Gilbertson WE, Lessing M (1988) Danthron alarm, FDA
response: crucial OTC drug control. Mil Med 153:487–488
13. (1990) IARC Monographs programme on the evaluation of the
carcinogenic risk of chemicals to humans. Pharmaceutical Drugs
IARC Monogr Eval Carcinog Risk Chem Hum 50:265–275
14. Zhang Z, Fu J, Yao B, Zhang X, Zhao P, Zhou Z (2011) In vitro
genotoxicity of danthron and its potential mechanism. Mutat Res
722:39–43
15. Zhang H, Zhou R, Li L, Chen J, Chen L, Li C et al (2011)
Danthron functions as a retinoic X receptor antagonist by stabi-
lizing tetramers of the receptor. J Biol Chem 286:1868–1875
16. Lu HF, Lai TY, Hsia TC, Tang YJ, Yang JS, Chiang JH et al
(2010) Danthron induces DNA damage and inhibits DNA repair
gene expressions in GBM 8401 human brain glioblastoma mul-
tiforms cells. Neurochem Res 35:1105–1110
17. Lu HF, Wang HL, Chuang YY, Tang YJ, Yang JS, Ma YS et al
(2010) Danthron induced apoptosis through mitochondria- and
caspase-3-dependent pathways in human brain glioblastoma
multiforms GBM 8401 cells. Neurochem Res 35:390–398
18. Yu FS, Yang JS, Yu CS, Lu CC, Chiang JH, Lin CW et al (2011)
Safrole induces apoptosis in human oral cancer HSC-3 cells.
J Dent Res 90:168–174
19. Wu SH, Hang LW, Yang JS, Chen HY, Lin HY, Chiang JH et al
(2010) Curcumin induces apoptosis in human non-small cell lung
cancer NCI-H460 cells through ER stress and caspase cascade-
and mitochondria-dependent pathways. Anticancer Res 30:2125–
2133
20. Lu CC, Yang JS, Huang AC, Hsia TC, Chou ST, Kuo CL et al
(2010) Chrysophanol induces necrosis through the production of
ROS and alteration of ATP levels in J5 human liver cancer cells.
Mol Nutr Food Res 54:967–976
21. Coder DM (2001) Assessment of cell viability. Curr Protoc
Cytom. Editorial board: Paul Robinson J, managing editor…[et al.] Chapter 9:Unit 9.2
22. Lee JH, Li YC, Ip SW, Hsu SC, Chang NW, Tang NY et al
(2008) The role of Ca2? in baicalein-induced apoptosis in human
breast MDA-MB-231 cancer cells through mitochondria- and
caspase-3-dependent pathway. Anticancer Res 28:1701–1711
23. Lin SS, Huang HP, Yang JS, Wu JY, Hsia TC, Lin CC et al
(2008) DNA damage and endoplasmic reticulum stress mediated
curcumin-induced cell cycle arrest and apoptosis in human lung
carcinoma A-549 cells through the activation caspases cascade-
and mitochondrial-dependent pathway. Cancer Lett 272:77–90
24. Ji BC, Yu CC, Yang ST, Hsia TC, Yang JS, Lai KC et al (2012)
Induction of DNA damage by deguelin is mediated through
reducing DNA repair genes in human non-small cell lung cancer
NCI-H460 cells. Oncol Rep 27:959–964
25. Kuo CL, Wu SY, Ip SW, Wu PP, Yu CS, Yang JS et al (2011)
Apoptotic death in curcumin-treated NPC-TW 076 human
nasopharyngeal carcinoma cells is mediated through the ROS,
mitochondrial depolarization and caspase-3-dependent signaling
responses. Int J Oncol 39:319–328
26. Yeh RD, Chen JC, Lai TY, Yang JS, Yu CS, Chiang JH et al
(2011) Gallic acid induces G0/G1 phase arrest and apoptosis in
human leukemia HL-60 cells through inhibiting cyclin D and E,
and activating mitochondria-dependent pathway. Anticancer Res
31:2821–2832
Neurochem Res (2012) 37:1790–1800 1799
123

27. Chou ST, Peng HY, Chang CT, Yang JS, Chung HK, Yang ST
et al (2011) Zanthoxylum ailanthoides Sieb and Zucc. extract
inhibits growth and induces cell death through G2/M-phase arrest
and activation of apoptotic signals in colo 205 human colon
adenocarcinoma cells. Anticancer Res 31:1667–1676
28. Wu PP, Chung HW, Liu KC, Wu RS, Yang JS, Tang NY et al
(2011) Diallyl sulfide induces cell cycle arrest and apoptosis in
HeLa human cervical cancer cells through the p53, caspase- and
mitochondria-dependent pathways. Int J Oncol 38:1605–1613
29. Ho YT, Yang JS, Li TC, Lin JJ, Lin JG, Lai KC et al (2009)
Berberine suppresses in vitro migration and invasion of human
SCC-4 tongue squamous cancer cells through the inhibitions of
FAK, IKK, NF-kappaB, u-PA and MMP-2 and -9. Cancer Lett
279:155–162
30. Ji BC, Hsu WH, Yang JS, Hsia TC, Lu CC, Chiang JH et al
(2009) Gallic acid induces apoptosis via caspase-3 and mito-
chondrion-dependent pathways in vitro and suppresses lung
xenograft tumor growth in vivo. J Agric Food Chem 57:
7596–7604
31. Chen JC, Lu KW, Tsai ML, Hsu SC, Kuo CL, Yang JS et al
(2009) Gypenosides induced G0/G1 arrest via CHk2 and apop-
tosis through endoplasmic reticulum stress and mitochondria-
dependent pathways in human tongue cancer SCC-4 cells. Oral
Oncol 45:273–283
32. Chen YL, Lu HF, Hung FM, Huang AC, Hsueh SC, Liu CM et al
(2011) Danthron inhibits murine WEHI-3 cells in vivo, and
enhances macrophage phagocytosis and natural killer cell cyto-
toxic activity in leukemic mice. In Vivo 25:393–398
33. Lin CC, Chen JT, Yang JS, Lu HF, Hsu SC, Tan TW et al (2009)
Danthron inhibits the migration and invasion of human brain
glioblastoma multiforme cells through the inhibition of mRNA
expression of focal adhesion kinase, Rho kinases-1 and metallo
proteinase-9. Oncol Rep 22:1033–1037
34. Sun Y, Chen J, Rigas B (2009) Chemopreventive agents induce
oxidative stress in cancer cells leading to COX-2 overexpression
and COX-2-independent cell death. Carcinogenesis 30:93–100
35. Chen Y, McMillan-Ward E, Kong J, Israels SJ, Gibson SB (2008)
Oxidative stress induces autophagic cell death independent of
apoptosis in transformed and cancer cells. Cell Death Differ
15:171–182
36. Pathak N, Khandelwal S (2007) Role of oxidative stress and
apoptosis in cadmium induced thymic atrophy and splenomegaly
in mice. Toxicol Lett 169:95–108
37. Chatterjee S, Kundu S, Bhattacharyya A (2008) Mechanism of
cadmium induced apoptosis in the immunocyte. Toxicol Lett
177:83–89
38. Kumar B, Kumar A, Pandey BN, Mishra KP, Hazra B (2009)
Role of mitochondrial oxidative stress in the apoptosis induced by
diospyrin diethylether in human breast carcinoma (MCF-7) cells.
Mol Cell Biochem 320:185–195
39. Galluzzi L, Larochette N, Zamzami N, Kroemer G (2006)
Mitochondria as therapeutic targets for cancer chemotherapy.
Oncogene 25:4812–4830
40. Armstrong JS (2007) Mitochondrial medicine: pharmacological
targeting of mitochondria in disease. Br J Pharmacol 151:
1154–1165
41. Pilkington GJ, Parker K, Murray SA (2008) Approaches to mi-
tochondrially mediated cancer therapy. Semin Cancer Biol
18:226–235
42. Lee I, Bender E, Kadenbach B (2002) Control of mitochondrial
membrane potential and ROS formation by reversible phos-
phorylation of cytochrome c oxidase. Mol Cell Biochem
234–235:63–70
43. Galluzzi L, Joza N, Tasdemir E, Maiuri MC, Hengartner M,
Abrams JM et al (2008) No death without life: vital functions of
apoptotic effectors. Cell Death Differ 15:1113–1123
44. Qi F, Li A, Zhao L, Xu H, Inagaki Y, Wang D et al (2010)
Cinobufacini, an aqueous extract from Bufo bufo gargarizans
Cantor, induces apoptosis through a mitochondria-mediated
pathway in human hepatocellular carcinoma cells. J Ethnophar-
macol 128:654–661
45. Cheng AC, Tsai ML, Liu CM, Lee MF, Nagabhushanam K, Ho
CT et al (2010) Garcinol inhibits cell growth in hepatocellular
carcinoma Hep3B cells through induction of ROS-dependent
apoptosis. Food Funct 1:301–307
46. Muscolini M, Cianfrocca R, Sajeva A, Mozzetti S, Ferrandina G,
Costanzo A et al (2008) Trichostatin A up-regulates p73 and
induces Bax-dependent apoptosis in cisplatin-resistant ovarian
cancer cells. Mol Cancer Ther 7:1410–1419
47. Lee DH, Kim C, Zhang L, Lee YJ (2008) Role of p53, PUMA,
and Bax in wogonin-induced apoptosis in human cancer cells.
Biochem Pharmacol 75:2020–2033
48. Yang SH, Chien CM, Lu MC, Lin YH, Hu XW, Lin SR (2006)
Up-regulation of Bax and endonuclease G, and down-modulation
of Bcl-XL involved in cardiotoxin III-induced apoptosis in K562
cells. Exp Mol Med 38:435–444
1800 Neurochem Res (2012) 37:1790–1800
123




















