
BAGE: a new gene encoding an antigen recognized on human melanomas by cytolytic T lymphocytes
Upload
independentCategory
view
6download
0
EJCa European Journal of Cell Biology 76, 25-32 (1998, May) . © Gustav Fischer Verlag· Jena 25
The leucine-based motif DDQxxLI is recognized both for internalization and basolateral sorting of invariant chain in MDCK cells
Anne Simonsen2), Bjprn Bremnes2
), Tommy W. Nordeng, Oddmund Bakke 1)
Department of Biology, University of Oslo, Oslo/Norway
Received April 7, 1997 Accepted December 9, 1997
Endocytosis - epithelial cell polarity - invariant chain -sorting signals
The major histocompatibility complex (MHC) class II-associated invariant chain (Ii) contains signals for transport to endocytic compartments where the class II molecules bind antigenic peptides for presentation to CD4+ T cells. Two leucine-based signals in the Ii cytoplasmic tail can be independently recognized for endosomal sorting of Ii, and we have recently shown that each signal is sufficient for basolateral sorting and internalization of Ii in polarized Madine Darby Canine Kidney (MDCK) II cells. The recognition motif for endosomal sorting is complex and consists of two critical leucine-like residues as well as surrounding amino acids. Here, we have analyzed the importance of residues surrounding the membrane-distal leucine-based signal in basolateral sorting and internalization of Ii in MDCK II cells. We find that the DDQxxLI motif is involved in both sorting events indicating the presence of similar signal recognition components both at the TGN and at the plasma membrane. The identical motif is required for endosomal localization and internalization of Ii also in simian COS cells and the human He La and Ml cells.
Abbreviations: DMEM Dulbecco's modified Eagle's medium. - FcRIl Immunoglobulin Fc receptor. - FCS Fetal calf serum. - GaM-FITC Goat anti-mouse IgG conjugated to fluorescein isothiocyanate. -Ii Invariant chain. - immuno-FM Immuno-fluorescence microscopy.LDL-R Low density lipoprotein receptor. - MDCK Madin Darby Canine Kidney. - MHC Major histocompatibility complex. - PBS phosphate-buffered saline. - PBS-CM PBS-CaCb-MgCb - TGN trans-Goigi network.
1) Dr. Oddmund Bakke, Division of Molecular Biology, Department of Biology, University of Oslo, P. O. Box 1950 Blindern, N-0316 Oslol Norway. 2) Present address: Department of Biochemistry, The Norwegian Radiumhospital, Montebello, N-03lO OslolNorway.
Introduction
The plasma membrane of polarized cells is divided into apical and basolateral domains with distinct protein and lipid composition [52, 53]. In MDCK cells, proteins destined for the plasma membrane are generally sorted in the trans-Golgi network (TGN) for direct delivery to the respective domains [60]. Sorting signals involved in the transport of newly synthesized membrane proteins from the TGN to the basolateral surface have been identified in cytoplasmic domains of several proteins and these signals are generally divided into two major classes. The first is apparently related to signals recognized for internalization via clathrin coated pits and contains a critical tyrosine [7, 11, 15, 16, 19,25,27,45,46,64] or leucine residue [14,27,54]. The second class of basolateral sorting signals is not related to internalization signals, but can still be tyrosinedependent [12, 25, 27, 31, 56, 57] or -independent [2, 8, 9,17, 39, 54]. Basolateral sorting signals also appear to mediate recycling of internalized proteins from endosomes back to the basolateral plasma membrane [3, 26, 30], indicating that the sorting machinery at different sorting stations within the cell recognizes elements of the same signal.
The precise three-dimensional structure adopted by basolateral sorting signals is not known, but recent data indicate that some basolateral signals adopt similar conformations as several known internalization signals [2, 20, 33, 62]. However, the precise requirements for basolateral sorting and internalization are not necessarily identical [16, 20, 25, 27, 45], indicating that amino acids in the vicinity of the tyrosine or leucine residues determine whether the motif functions as a signal for endocytosis, basolateral sorting, or both. Acidic residues downstream of the tyrosine are required for basolateral sorting, but not for internalization, of the low density lipoprotein receptor (LDL-R) [27], whereas basolateral sorting and internalization of the immunoglobulin Fc receptor II (FcRII) by its leucine-based motif appears to be independent on acidic residues [14]. However, acidic amino acids are important for endosomal sorting of several proteins containing tyrosine- or leucine-based motifs such as invariant chain (Ii) [33,44], furin [59] and the cation-dependent mannose 6-phosphate receptor [28].

26 A. Simonsen, B. Bremnes, T. W. Nordeng, O. Bakke
The cytoplasmic tail of Ii contains two independent endosomal sorting signals, a leucine7-isoleucine8 (LI)-based motif and a methionine16-leucine17 (ML)-based motif [6, 36], both also shown to mediate rapid internalization of Ii from the cell surface [6, 49]. The LI motif lies within an a-helix, whereas the ML motif is part of a ~-turn [33]. We have recently shown that both motifs can independently mediate internalization and basolateral sorting of Ii in MDCK cells [54]. The function of these motifs in Ii internalization depends on their amino acid context: the context of the membrane-distal LI motif was identified as (D)DQxxLI in COS cells [33] or DDxxxLI in He La cells [44], where x could be mutated to alanine with no effect on internalization.
In this study we have analyzed the context of the LI motif required for internalization and basolateral sorting of Ii in MDCK cells. We found that the DDQxxLI motif is involved in both sorting events. As this motif is somewhat different from what was previously found in COS and He La cells, we wanted to determine whether these differences were due to cell typeor species-specific differences. We have compared the context of the LI motif in MDCK, COS, He La and Ml cells and found that the same DDQxxLI motif was involved in endosomal sorting and internalization of Ii in these cell lines. Together our data indicate that similar sorting motifs can be recognized both in the TGN and at the plasma membrane and suggest that the machinery that recognizes endosomal sorting signals is conserved among cell types and species.
Materials and methods
Cell culture Madin Darby Canine Kidney (MDCK) (strain II) epithelial cells, simian COS fibroblasts, human HeLa cells, and human M1 fibroblasts were grown in Dulbecco's modified Eagle's medium (DMEM) (Bio Whittaker, Walkersville, MD, USA) supplemented with 10 % fetal calf serum (FCS) (Gibco, Paisley, England). To form tight epithelial monolayers, MDCK cells (3.3 x lOs/cm2) were grown for 4-5 days on Transwell polycarbonate filter units (Costar, Cambridge, MA, USA) (pore size of 0.4 [lm) prior to the experiments. The relative surface area of the apical and the basolateral plasma membrane in MDCK cells grown under the present conditions is 1: 1 (apicallbasolateral) [58].
Plasmid constructions cDNA encoding human p33Ii and the deletion mutants 812-29Ii and 821-29Ii [41] were subcloned into the expression vector RSV.5 (neo) [23]. Point mutations in the cytoplasmic tail of Ii were introduced by site directed mutagenesis in a single-stranded M13 mp19 vector by the method of Kunkel [18]. The cytoplasmic tail region used for mutagenesis was controlled by DNA sequencing and then exchanged with the same non-mutated region in the Ii constructs. The mutated cDNA (812-29IiD2A , 812-29IiD3A, 812-29IiQ4A, 812_29IiR5A.D6A, 812-29IiL7A, 812-29IiI8A, 812-29Iis9A, 821_29Iis9A.L17A, Iis9A.Ll7A) were then subcloned into the RSY.5 (neo) vector.
Transfection and clone selection MDCK cells were stably transfected by the DNA-calcium phosphate procedure essentially as described [61]. Cells expressing the neomycin resistance marker were selected in the presence of 0418 (geneticin) (0.5 mg/ml active weight) (Sigma, St. Louis, MO, USA) in the culture medium. Stably transfected cell lines were established by isolating resistant colonies using cloning cylinders. Colonies expressing the protein of interest were identified by immuno-fluorescence microscopy and radio immunoprecipitation assays, as described.
For transient expression in COS, HeLa and Ml cells, cells were transfected by a DEAE-dextran procedure, as described [6].
Immunofluorescence microscopy (immuno-FM) Stably transfected MDCK cells were grown on filters and processed for immuno-FM essentially as described by Berod et al. [5]. The mouse monoclonal antibody (mAb) BU45 [63] (The Binding Site, Birmingham, UK), recognizing the luminal domain of human Ii was used. The polarized plasma membrane distribution or internalization of the transfected proteins were visualized adding BU45, diluted in DMEM-1 %FCS-lOmM Hepes, to the apical or the basolateral side of the monolayer for 1 hour at 4°C or 37°C, respectively. The cells were then washed extensively with phosphate-buffered saline (PBS) containing 0.1 mM CaCl2 and 1 mM MgCIz (PBS-CM) and 2 % FCS, fixed by the pH shift paraformaldehyde fixation procedure [5] and permeabilized by 0.05 % Triton X-lOO. The total cellular protein distribution was visualized by labeling of fixed and permeabilized cell mono layers with BU45 and FITC-conjugated goat anti-mouse IgG antibodies (GaMFITC) (Dianova, Hamburg, FRG) for 1 hour at 3rC in a humidified chamber.
The intracellular protein localization in unpolarized MDCK cells and in transiently transfected COS, HeLa and Ml cells was visualized by labeling of cells grown on coverslips with BU45 and GaM-FITC after fixation in methanol for 5 min at -20°C. Internalization of the transfected proteins was analyzed by incubation of cells grown on coverslips with BU45 for 30 min at 37°C before fixation and labeling with GaM-FITC. As a control, non-transfected cells were used in all experiments, and no specific labeling could be detected. Fluorescence was detected using a Nikon fluorescence microscope (Nikon, Tokyo, Japan).
Radiolabeling and immunoprecipitation Biosynthesis of the transfected proteins was examined by metabolic labeling of stably transfected MDCK cells grown in 35-mm wells, followed by immunoprecipitation analysis. The cells were starved for 30 min in MEM lacking methionine and cysteine and then labeled at 37°C for 20 min in the same medium supplemented with 200 [lCi [3SS]methionine/cysteine per ml. The cells were then placed on ice, washed three times in ice-cold PBS and lysed in ice-cold lysis buffer (50 mM TrisHCI, pH 7.5, 150 mM NaCl, 1 % NP-40) containing a cocktail of protease inhibitors (4 [lg/ml PMSF, 2 [lg/ml antipain, 2 [lg/ml leupeptin, and 1 [lg/ml pepstatin A) for 30 min. Samples were centrifuged (10000g) for 10 min at 4°C to remove cell nuclei and cellular debris. The lysate was incubated with BU45 for at least 2 hours at 4°C. Dynabeads® rat anti-mouse IgGI (M-450) (Dynal, Oslo, Norway) (0.5 mg Dynabeads® per lysate from 1 x 106 cells) were added and incubation at 4°C continued for 1-2 hours. The beads were then washed three times in 1.5 ml ice-cold PBS/I % NP-40 by using a Dynal Magnetic Particle Concentrator (MPC®-M) to collect the beads after each washing step. The beads were then resuspended in 25 [ll sample buffer and boiled for 5 min. Proteins bound to the beads were analyzed by SDSpolyacrylamide gel electrophoresis on a 12 % polyacrylamide gel. Fluorography of the gel was performed using Amplify, as directed by the manufacturer (Amersham, Buckinghamshire, England).
No detectable Ii was precipitated from non-transfected cells. The amount of precipitated protein, compared to the amount of total labeled cellular protein of each mutant cell line was quantitated by counting 1110 of the precipitated material and VHXl of the total lysate in a ~-counter. The level of expression between the different cell lines varied by less than a factor 4 and at least two independent clones with different expression levels were analyzed for each construct. The results of the various assays did not depend on the expression level.
Internalization of 1251-labeled antibodies Mouse ascites BU45 was purified and 12sI-labeled as described [54]. Stably transfected MDCK cells grown in 35-mm wells were incubated with 125I_BU45 in DMEM-1 % FCS-lOmM Hepes at 4°C for 2h. The cells were then washed six times in ice-cold PBS-2 % FCS and chased in DMEM-lO % FCS at 37°C for different periods of time (0, 1,5, 15, or 30 min), each performed in duplicate. The cells were then cooled on ice and treated twice with 0.5 M acetic acid 0.15 M NaCl (pH 2.5) for 7 min. This step removed 95-98 % of the antibody bound to the surface

EJCI
after internalization. The cells were then lysed in 1 M NaOH. The acid wash and the lysate were counted in a y-counter. Internalized antibody was calculated from the amount of 125I_BU45 (cpm) present in the lysate relative to the total amount of 125I_BU45 bound before the chase period (cpm in lysate + cpm in acid wash). Non-specific binding of 125r_ BU45 was typically less than 1 % of the specific activity, as determined by binding to cells transfected with an irrelevant cDNA or by quenching binding with a lOO-fold excess of non-labeled antibody. We used intact BU45 antibody, as internalization of 125I-labeled BU45 and of 125I_labeled BU45 Fab-fragments is equivalent [49].
Surface binding of 1251_labeled antibodies The polarized cell surface distribution of the transfected proteins was analyzed by binding of 125I_BU45 to cells grown on Transwell units for 4-5 days prior to the experiments. Binding assays were all performed on ice in the cold-room. Cells were cooled on ice and 125I-BU45, diluted in DMEM-l % FCS-lOmM Hepes, was added to either the apical or the basolateral side of the monolayer for 2 hours. Cell monolayer tightness was assessed by measuring the radioactivity in the medium of both chambers, and generally less than 0.1 % of the added radioactivity had diffused through the monolayer. Unbound antibody was removed by extensive washing (5 x) in PBS-CM-2 % FCS. The filters were then excised, and bound radioactivity was measured in a ycounter. Non-specific binding was determined by quenching with a 100-fold excess of non-labeled antibody. These values, which generally represented less than 10 % of the total binding, were subtracted from the total bound radioactivity to give the specific binding. Total binding ranged from 2 x 104_2 X 105 cpm for proteins that were expressed stably at the plasma membrane and from 2 x 103-2 X 104 cpm for proteins having efficient internalization signals. The given values represent the mean ± standard error (SE), obtained from at least three independent experiments, each performed in duplicate.
Results
Expression and intracellular localization of A 12-291i mutants in non-polarized MDCK cells We have recently shown that a leucine-based motif in the membrane distal part of the Ii cytoplasmic tail can mediate internalization and basolateral sorting of Ii in polarized MDCK cells [54]. Here we wanted to study whether both of these functions depend on similar residues surrounding Leu7. We performed an alanine-scan mutagenesis on an Ii deletion mutant, containing 11 N-terminal residues of the cytoplasmic tail (A12-29Ii), as outlined in Fig. 1. This construct and the mutants A12_29IiD2A A12-29IiD3A A12-29IiQ4A A 12_29IiR5A-D6A, A12-29IiL7A , A12-29IiI8A and 'A 12-29Iis9A (Fig. 1), were transfected stably in MDCK cells. Biosynthesis of the mutant proteins was examined by metabolic labeling of the cells for 20min with a mixture of [35S] methionine/cysteine. Ii was immunoprecipitated with the BU45 antibody. All A12-29Ii constructs had the expected molecular mass of 30 kDa (data not shown). The amount of precipitated protein was compared to total labeled cellular protein by counting in a B-counter. When Ii is overexpressed in MDCK cells it is distributed both basolaterally and apically (Nordeng and Bakke, manuscript in preparation). We therefore used mutant clones with similar or lower expression levels than A12-29Ii (Table I) to avoid a potential saturation of the sorting machinery.
Intracellular localization of the stably expressed proteins in non-polarized MDCK cells was analyzed by immuno-FM. A12-29Ii and A12_29IiR5A-D6A were localized in intracellular vesicles, with no detectable plasma membrane staining (Fig.
Signals for polarized sorting of Ii 27
Metl~Asp2~Asp3~Gln4_Arg5_Asp6_Leu7_I1euS_Ser~_Asnlll_Asnll_Glull_Gln13_Leu14_Prol!_Metl~_Leu17_
Gly18_ArgI9_Arglll.ProZI_Gly2l.Ala23.Pro24_Glu25.SerU_LYs27.CysU.SerZ9_Arg30 ....
Ii- human MDDQRDLISNNEQLPMLGRRPGAPESKCSR ...
t112·29Ii MDDQRDLISNN RRR.
'" 12.29IiDlA MADQRDLISNN RRR ..
'" 12·29IiD3A MDAQRDLISNN RRR.
'" 12_29IiQ4A MDDARDLISNN RRR ...
~ 12_29IiRSA.D6A MDDQAALISNN RRR ...
'" 12.29IiL 7 A MDDQRDAISNN RRR ...
'" 12-291i'" MDDQRDLASNN RRR ...
'" 12_29IiS9A MDDQRDLIANN RRR ..
~ 21_29Iis9A-L17A MDDQRDLIANNEQLPMAGRR ___ RRR .. .
nS9A-L17A MDDQRDLIANNEQLPMAGRRPGAPESKCSR .. .
Fig. 1. Invariant chain amino acid sequence. Amino acid sequence of the N-terminal cytoplasmic tail of wild-type human p33 Ii (in threeletter amino acid code) and various Ii deletion and point mutants (in one-letter amino acid code). In ~12-29Ii and ~2l-29Ii, the sequence encoding residues 12-29 and 21-29 of the Ii cytoplasmic tail was deleted, as indicated by a line. In the constructs ~12-29IiD2A, ~12-29IiD3A , ~12-29IiQ4A, ~12_29IiR5A-D6A, ~12-29IiL7A, ~12-29IiI8A, ~12-29Iis9A, ~21_29Iis9A-Ll7A, and Iis9A-LI7Awere the indicated residues mutated to alanine, as shown in bold letters.
2). In contrast, a predominant plasma membrane localization of A12-29IiD2A, A12-29IiD3A, A 12-29IiQ4A, A 12-29IiL7A , A12-29IiI8A and A12_29Iis9A was detected (Fig. 2), whereas A12-29Iis9A showed some vesicular staining. These data indicate that the amino acids ASp2, ASp3, Gln4, Leu7, Ileu8, and Ser9
are important for endosomal sorting of A 12-29Ii, whereas Arg5-Asp6 can be mutated to alanine with no detectable effect on endosomal sorting of A12-29Ii.
Internalization of A 12-291i mutants in nonpolarized MDCK cells Endosomal sorting of Ii can occur directly from the TGN or via internalization from the plasma membrane. In order to analyze the efficiency of sorting of different mutant A12-29Ii
Tab. I. Biosynthesis of ~12-29Ii mutants III stably transfected MDCKcells.
Cell line
~12-29Ii ~ 12_291iD2A
~ 12_291iD3A
~12-29IiQ4A ~ 12_29IiR5A-D6A ~ 12_291iL7A
~ 12-291i18A
~ 12-291iS9A
Precipitated protein (x 10-3 of total ± SE)
1.40±0.23 1.35 ± 0.15 1.50±0.20 0.74±0.07 0.90±0.22 1.14±0.12 0.35±0.03 1.19±0.24
Cells expressing Ii, ~ 12-29Ii, ~ 12-29IiD2A, ~ 12-29IiD3A, ~ 12-29IiQ4A, ~ 12-29IiR5A-D6A, ~ 12-29IiL7A, ~ 12-291i18A, or ~ 12-291iS9A were labeled with [35S]methionine/cysteine for 20 min, before lysis and immunoprecipitation of labeled protein with BU45 and RAM IgG1 Dynabeads, followed by SDS-PAGE and fluorography. The amount of precipitated protein compared to total labeled cellular protein was quantitated by counting a fraction of the precipitated material and a fraction of the total lysate in a ~-counter. For each mutant, the clone having the highest level of expression is shown.

28 A. Simonsen, B. Bremnes, T. W. Nordeng, O. Bakke
L'.12-29Ii
L'.12-29Ii
DZA
L'.12-29Ii
D3A
L'.12-29Ii
Q4A
Fig. 2. Intracellular localization of L'.12-29Ii mutants. Immunofluorescence micrographs showing the intracellular distribution of L'.12-29Ii, ~12-29IiD2A, M2-29IiD3A, M2-29IiQ4A, ~12_29IiR5A.D6A, ~12-
proteins via the plasma membrane, the internalization of the proteins was quantified by the uptake of cell surface bound 125I-Iabeled BU45 in non-polarized cells. ~12-29Ii internalized with the same kinetics as the wild-type Ii (Fig. 3): approximately 40 % of the surface labeled protein was internalized after 5 min and approximately 80 % was internalized after 30 min. Interestingly, 125I_BU45 was internalized more efficiently in ~ 12_29IiR5A.D6A transfected cells as 70 % of the surfacebound antibody was internalized after 5 min (Fig. 3). Mutation of ASp2 to Ala decreased the rate of internalization, as only 13 % of surface-labeled ~12_29IiD2A was internalized in 5 min, whereas 60 % was internalized after 30 min chase (Fig. 3). The rest of the mutants (M2-29IiD3A, ~12-29IiQ4A, ~12-29IiL7A, ~12_29IiI8A, ~12_29IiS9A) internalized slowly, as less than 25 % of the surface-bound 125I_BU45 was internalized after 30 min (Fig. 3). From this we conclude that ASp2, ASp3, Gln4, Leu 7, Ileu8, and Ser9 are important for efficient internalization of ~12-29Ii in MDCK cells, whereas mutation of Arg5-Asp6 does not reduce the internalization efficiency.
Polarized surface distribution of A 12-291i mutants To analyze the steady state polarized surface distribution of the transfected Ii mutants, MDCK cells were grown on perme-
L'.12-29Ii
RSA,D6A
L'.12-29Jj
L7A
L'.I2-29Jj
1SA
L'.12-29Ii
S9A
29IiL7A , ~12-29IiI8A, ~12_29Iis9A in stably transfected MDCK cells. The cells were fixed in methanol and labeled with BU45 and GaMFITC. Bar 20 f.lm.
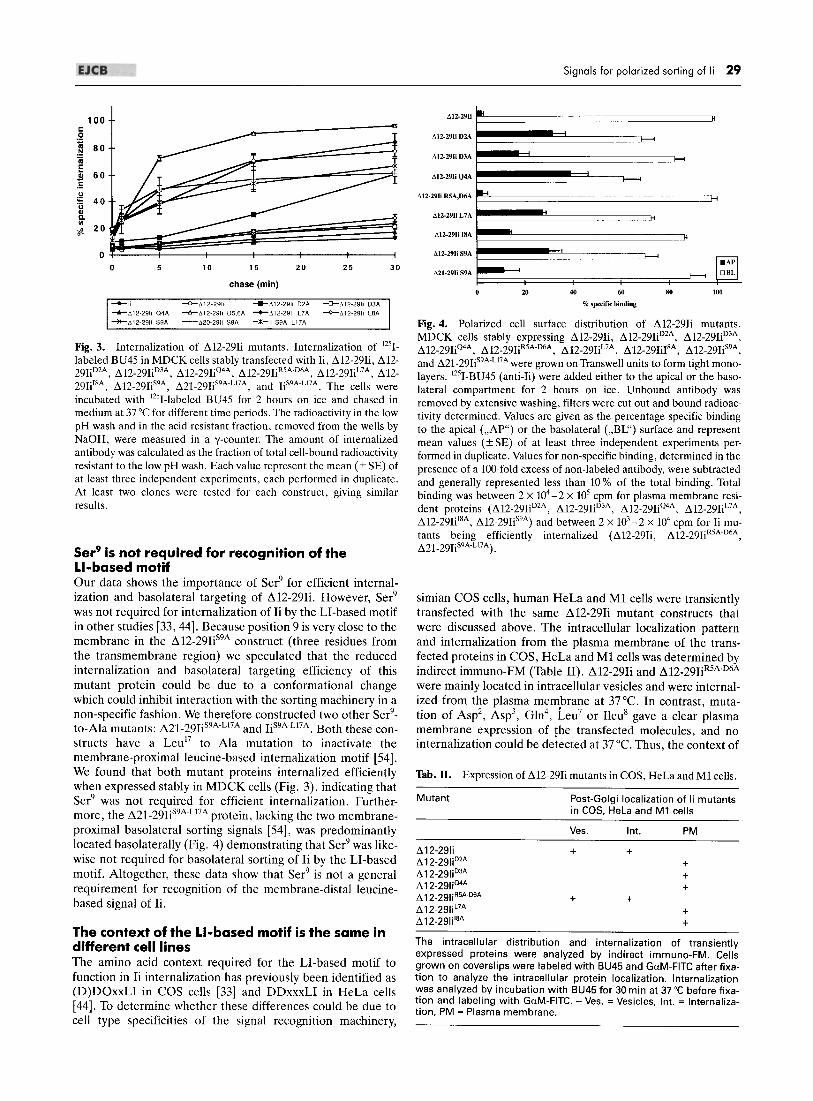
able filter supports to form tight epithelial monolayers, and 125I-Iabeled BU45 was added to the apical or the basolateral side of the cell monolayer at 4°C. After removal of unbound antibody, the filters were excised and bound radioactivity was determined in a y-counter. The percentage of specific binding to the apical or the basolateral surface is shown in Fig. 4. 98 % of the ~12-29Ii and ~12_29IiR5A.D6A molecules expressed on the surface were located basolaterally. The mutant proteins ~12_29IiD2A, ~12-29IiQ4A, ~12_29IiL7A, and ~12_29Iis9A were detected at both apical and basolateral surfaces (30 % apical), whereas mutation of ASp3 or Ileu8 to alanine had an intermediate effect on the basolateral sorting, as 18 % and 13 % of the surface-expressed proteins were detected apically, respectively. The polarized surface distribution of the mutant proteins obtained by this method was corroborated by confocal immuno-FM of polarized cells surface-labeled with BU45 (micrographs not shown). Furthermore, at 37°C BU45 was internalized from the basolateral surface of cells expressing ~12-29Ii and ~12_29IiR5A.D6A, but no internalization could be detected from the apical side (micrographs not shown). Together, these data show that ASp2, ASp3, Gln4, Leu7, Ileu8
and Ser9, but not Arg5 and ASp6, are important for efficient basolateral sorting of the ~12-29Ii construct.
EJCB
1 00 c 0
~ 80 .!:! iii E
60 ~ .S
" ;;::: 40 '0 ., a. Ul
"# 20
0 0 10 15
chase (min)
20 25
~Ii -<>-612-2911 _612-291i D2A ~D..12-29Ii 03A -'-612-29Ii Q4A -6-612-29Ii D5,6A ~l!.12-29Ij L7A -<>-612-29Ii LBA
~L\12-29Ii S9A -+-620-29Ii S9A -X-Ii S9A, L17A
30
Fig. 3. Internalization of ~12-29Ii mutants. Internalization of 1251_ labeled BU45 in MDCK cells stably transfected with Ii, M2-29Ii, ~12-29IiD2A, ~12_29IiD3A, ~12-29IiQ4A, ~12_29IiR5A.D6A, ~12_29IiL7A, ~12-29IiI8A, ~12-29Iis9A, ~21_29Iis9A.L17A, and IiS9A.L17A. The cells were incubated with 12sI-labeled BU45 for 2 hours on ice and chased in medium at 37°C for different time periods. The radioactivity in the low pH wash and in the acid resistant fraction, removed from the wells by NaOH, were measured in a y-counter. The amount of internalized antibody was calculated as the fraction of total cell-bound radioactivity resistant to the low pH wash. Each value represent the mean (± SE) of at least three independent experiments, each performed in duplicate. At least two clones were tested for each construct, giving similar results.
Ser9 is not required for recognition of the LI-based motif Our data shows the importance of Ser9 for efficient internalization and basolateral targeting of A12-29Ii. However, Ser9
was not required for internalization of Ii by the LI-based motif in other studies [33, 44]. Because position 9 is very close to the membrane in the A12-29Iis9A construct (three residues from the transmembrane region) we speculated that the reduced internalization and basolateral targeting efficiency of this mutant protein could be due to a conformational change which could inhibit interaction with the sorting machinery in a non-specific fashion. We therefore constructed two other Ser9
-
to-Ala mutants: A21_29Iis9A.L17A and Iis9A.L17A. Both these con-structs have a Leu!7 to Ala mutation to inactivate the membrane-proximal leucine-based internalization motif [54]. We found that both mutant proteins internalized efficiently when expressed stably in MDCK cells (Fig. 3), indicating that Ser9 was not required for efficient internalization. Furthermore, the A21_29Iis9A.L17A protein, lacking the two membraneproximal basolateral sorting signals [54], was predominantly located basolaterally (Fig. 4) demonstrating that Ser9 was likewise not required for basolateral sorting of Ii by the LI-based motif. Altogether, these data show that Ser9 is not a general requirement for recognition of the membrane-distal leucinebased signal of Ii.
The context of the LI-based motif is the same in different cell lines The amino acid context required for the LI-based motif to function in Ii internalization has previously been identified as (D)DQxxLI in COS cells [33] and DDxxxLI in He La cells [44]. To determine whether these differences could be due to cell type specificities of the signal recognition machinery,
Signals for polarized sorting of Ii 29
t>12·29Ii II.
M2·29IiD2A
t>12·29IiD3A J--<
M2-29I;Q4A
il12·29Ii R5A,D6A
t>12·29Ii L7A
~ :::J< D..12-29li 18A
612-29Ij S9A
~1-29IiS9A ~ DBL
20 40 80 100
% specific binding
Fig. 4. Polarized cell surface distribution of ~12-29Ii mutants. MDCK cells stably expressing ~12-29Ii, ~12-29IiD2A, ~12_29IiD3A, ~12-29IiQ4A, ~12_29IiR5A.D6A, ~12-29IiL7A, ~12-29IiI8A, ~12_29Iis9A,
and ~21_29Iis9A.L17A were grown on Transwell units to form tight monolayers. 125I_BU45 (anti-Ii) were added either to the apical or the basolateral compartment for 2 hours on ice. Unbound antibody was removed by extensive washing, filters were cut out and bound radioactivity determined. Values are given as the percentage specific binding to the apical ("AP") or the basolateral ("BL") surface and represent mean values (± SE) of at least three independent experiments performed in duplicate. Values for non-specific binding, determined in the presence of a lOO-fold excess of non-labeled antibody, were subtracted and generally represented less than 10 % of the total binding. Total binding was between 2 x 104 - 2 X 105 cpm for plasma membrane resident proteins (~12-29IiD2A, ~12-29IiD3A, ~12-29IiQ4A, ~12_29IiL7A,
~12-29IiI8A, ~12-29Iis9A) and between 2 x 103_2 X 104 cpm for Ii mutants being efficiently internalized (~12-29Ii, ~12_29IiR5A.D6A, ~21_29Iis9A'L17A) .
simian COS cells, human HeLa and Ml cells were transiently transfected with the same A12-29Ii mutant constructs that were discussed above. The intracellular localization pattern and internalization from the plasma membrane of the transfected proteins in COS, HeLa and Ml cells was determined by indirect immuno-FM (Table II). A12-29Ii and A12_29IiR5A.D6A were mainly located in intracellular vesicles and were internalized from the plasma membrane at 37°C. In contrast, mutation of Asp2, Asp3, Gln4
, Leu7 or IleuS gave a clear plasma membrane expression of the transfected molecules, and no internalization could be detected at 37°C. Thus, the context of
Tab. II. Expression of ~12-29Ii mutants in COS, HeLa and Ml cells.
Mutant
~12-29Ii
812-291iD2A
~ 12-291iD3A
~ 12-291iQ4A 812_29IiR5A.D6A 812-291iL7A
~ 12-291i18A
Post-Golgi localization of Ii mutants in COS, HeLa and Ml cells
Ves.
+
+
Int.
+
+
PM
+ + +
+ +
The intracellular distribution and internalization of transiently expressed proteins were analyzed by indirect immuno-FM. Cells grown on coverslips were labeled with BU45 and GaM-FITC after fixation to analyze the intracellular protein localization. Internalization was analyzed by incubation with BU45 for 30 min at 37°C before fixation and labeling with GaM-FITC. - Ves. = Vesicles, Int. = Internalization, PM = Plasma membrane.

30 A. Simonsen, B. Bremnes, T. W. Nordeng, O. Bakke
the leucine-based motif required for endosomal localization and internalization of ~12-29Ii in COS , HeLa, M1 and MDCK cells appears to be identical.
Discussion
In this paper we show that the same residues surrounding the LI motif are involved in the efficient internalization and basolateral sorting of Ii in MDCK cells. ASp2, Gln4 and Leu 7 were required both for efficient internalization and basolateral sorting, whereas ASp3 and I1eu8 were required for efficient internalization, but had only moderate effect on the steady-state basolateral localization of Ii. On the other hand , mutating Args-Asp6 to alanine did not result in reduced efficiency of internalization and basolateral sorting compared to wild-type Ii. Interestingly, the Args-Asp6 mutant internalized even more efficiently than wild-type Ii , suggesting that tentative sorting signals are not necessarily optimal for maximal endocytosis.
We find the Ser9 residue to be required for efficient internalization of ~12-29Ii, but not of ~21-29Ii or full-length Ii. Thus, we conclude that Ser9 is not required for internalization of Ii by the LI motif. The replacement of Ser9 with Ala may result in conformational changes that are critical only in ~12-29Ii due to the proximity of the motif to the plasma membrane. It is noteworthy though that the serine residue is conserved in mouse, rat, human [4], bovine [35], and chicken Ii (Bremnes, B., Rode, M., Gedde-Dahl, M., Ness, S. and Bakke, 0., submitted) . Ser9 has also been found to be phosphorylated in HeLa cells (Anderson, H . A. and Roche, P.A., personal communication), suggesting that Ser9 may playa role in the intracellular routing of Ii.
The functional endosomal sorting motif DDQxxLI found in MDCK cells is not identical to the functional contexts (D)DQxxLI and DDxxxLI previously identified in COS cells [33] and HeLa cells [44], respectively. By transient transfection of simian COS cells and human HeLa and M1 cells with the same Ii mutants used for stable transfection of MDCK cells, we have shown that the same leucine-based motif was in fact recognized for endosomal sorting and internalization in all these cell lines, suggesting that the sorting machinery is conserved between the cell types studied. The different context requirements found in previous studies using the same cell lines could be due to the use of different Ii constructs. For instance, in a recent study using a chimeric molecule containing the Ii cytoplasmic tail fused to the transmembrane and luminal domains of the transferrin receptor (TfR) , the LIbased motif was reported to mediate rapid internalization, but not to be involved in basolateral sorting in MDCK cells [37]. These results should be interpreted with caution as Arneson and Miller [1] have shown that Ii is not sorted efficiently to endosomes unless it is in the trimeric form in which at least two of the tails contained the sorting signals whereas the IiTfR fusion protein is likely to form a monomer or dimer. We also have some evidence that the third basolateral sorting motif in the membrane-proximal part of the Ii cytoplasmic tail is only effective in the trimeric conformation, as a fusion protein with this Ii region and the transmembrane and luminal domain of neuraminidase, which forms a tetramer, is sorted to the apical surface [54]. Altogether, this indicates that the overall three-dimensional structure of the Ii molecule is important for recognition of the leucine-based motifs by the sorting machinery.
l:iij~;t!:iii~;~":~i ,E':1I
Several studies agree that in addition to the leucineisoleucine residues , acidic N-terminal amino acids are essential for active internalization. Acidic residues are also involved in endosomal sorting of other proteins [44, 51, 55, 59] and could serve to increase the affinity for the Golgi adaptor complex AP-1 [28]. Interestingly, basolateral sorting, but not internalization, of LDL-R is influenced by clusters of acidic residues C-terminal to the two tyrosine-based basolateral sorting motifs [27]. However, both basolateral sorting and internalization of FcRII by a leucine-based motif appear not to depend on the nearby acidic residues [14]. Clusters of two or more acidic residues C-terminal of tyrosine, phenylalanine or leucine residues are found in the cytoplasmic domains of many proteins that are targeted to the basolateral surface of MDCK cells [22, 29, 32, 34, 40, 42, 43, 48] , but the significance of the acidic residues in basolateral sorting of these proteins has not been studied. Thus, the precise requirements for a general acidic-xxx-Ieucine/tyrosine motif seem to differ between various proteins.
In MDCK cells, newly synthesized molecules are generally sorted in the TGN for direct delivery to the apical or the basolateral plasma membrane [60]. From the results presented in this paper we may conclude that the same DDQxxLI motif is recognized both at the TGN and at the plasma membrane. Other proteins also rely on a common tyrosine or leucine residue for internalization and basolateral sorting, although the amino acid context of the signal often differs [16, 21, 25, 27, 45]. A medium chain of the adaptor protein (AP)-complexes has recently been demonstrated to interact with the tyrosinebased signals [38, 47]. No such interaction was demonstrated for the leucine-based signals in their system, but other results indicate that also leucine-based motifs can bind to the APcomplexes [10, 13] and in particular to the medium chains (T. Bremnes, V. Lauvrak, B. Lindqvist and O. Bakke, submitted; D . G. Rodionov and O. Bakke, submitted). In a recent study Salamero et al. [50] found that a~-li expression recruited AP-1 adaptor complexes to Golgi membranes. This recruitment was dependent on the Ii cytoplasmic tail, again suggesting that AP-1 interacts with leucine signals. In a recent study it was concluded that common saturable components recognize tyrosine-based sorting signals for internalization and sorting to different post-Golgi compartments [24]. Thus, similar sorting motifs may be recognized by the sorting machinery at different cellular locations and direct a protein into different intracellular pathways.
However, we cannot rule out the possibility that the newly synthesized Ii molecules are targeted from the TGN to endosomes where they may be sorted for delivery to the apical or the basolateral plasma membrane. Proteins can be sorted in early endosomes for recycling to the same plasma membrane domain, transcytosed to the opposite plasma membrane domain or transported to later endocytic compartments or to other intracellular compartments. All these transport steps may depend on specific sorting signals. The amino acid context of the leucine-based motif recognized at the plasma membrane might differ from the context required for transport to late endocytic compartments from sorting endosomes. As we are not able to distinguish between Ii molecules which have internalized once or several times during the chase period in our internalization assay, the observed inhibition of Ii internalization by point mutations could also be due to inhibition of transport from early endosomes to late endosomes, leading to increased recycling of Ii. However, as we could not detect any

EJtB
internalization of anti-Ii antibodies at 3rC by immuno-FM, we suggest that the DDQxxLI motif is important for Ii internalization at the plasma membrane.
In this report we have shown that the same sorting motif may be recognized at two different locations in the cell and that the sorting machinery recognizing this motif seems to be conserved between cell types and species. These data may give insights into the nature of both basolateral signals and clathrin-mediated endocytosis signals, which eventually could lead to the identification of components of the sorting machinery and to an understanding of the complex mechanism of intracellular sorting in polarized cells.
Acknowledgements. We thank Hege Hardersen and Maifrid Roe for technical assistance and Tor Gjoen and Dmitrii Rodionov for critical reading of the manuscript. This work was supported by grants from the Research Council of Norway and the Norwegian Cancer Society.
References
[1] Arneson, L. S., J. Miller: Efficient endosomal localization of major histocompatibility complex class II-invariant chain complexes requires multimerization of the invariant chain targeting sequence. J. Cell BioI. 129, 1217-1228 (1995).
[2] Aroeti, B., P. A. Kosen, 1. D. Kuntz, F. E. Cohen, K. E. Mostov: Mutational and secondary structural analysis of the basolateral sorting signal of the polymeric immunoglobulin receptor. J. Cell BioI. 123, 1149-1160 (1993).
[3] Aroeti, B., K. E. Mostov: Polarized sorting of the polymeric immunoglobulin receptor in the exocytotic and endocytotic pathways is controlled by the same amino acids. EMBO J. 13, 2297-2304 (1994).
[4] Bakke, 0., B. Dobberstein: MHC class II-associated invariant chain contains a sorting signal for endosomal compartments. Cell 63, 707-716 (1990).
[5] Berod, A., B. K. Hartman, J. F. Pujol: Importance of fixation in immunohistochemistry: use of formaldehyde solutions at variable pH for the localization of tyrosine hydroxylase. J. Histochern. Cytochem. 29, 844-850 (1981).
[6] Bremnes, B., T. Madsen, M. Gedde-Dahl, O. Bakke: An LI and ML motif in the cytoplasmic tail of the MHC-associated invariant chain mediate rapid internalization. J. Cell Sci. 107, 2021-2032 (1994).
[7] Brewer, C. B., M. G. Roth: A single amino acid change in the cytoplasmic domain alters the polarized delivery of influenza virus hemagglutinin. J. Cell BioI. 114, 413-421 (1991).
[8] Casanova, J. E., G. Apodaca, K. E. Mostov: An autonomous signal for basolateral sorting in the cytoplasmic domain of the polymeric immunoglobulin receptor. Cell 66, 65-75 (1991).
[9] Dargemont, C., A. LeBivic, S. Rothenberger, B.Iacopetta, L. C. Kuhn: The internalization signal and the phosphorylation site of transferrin receptor are distinct from the main basolateral sorting information. EMBO J. 12, 1713-1721 (1993).
[10] Dietrich, J., J. Kastrup, B. L. Nielsen, N.Odum, C. Geisler: Regulation and function of the CD3gamma DxxxLL motif: A binding site for adaptor protein-l and adaptor protein-2 in vitro. J. Cell BioI. 138,271-281 (1997).
[11] Geffen, 1., C. Fuhrer, B. Leitinger, M. Weiss, K. Huggel, G. Griffiths, M. Spiess: Related signals for endocytosis and basolateral sorting of the asialoglycoprotein receptor. J. BioI. Chern. 268,20772-20777 (1993).
[12] Haass, C., E. H. Koo, A. Capell, D. B. Teplow, D. J. Selkoe: Polarized sorting of ~-amyloid precursor protein and its proteolytic products in MDCK cells is regulated by two independent signals. J. Cell BioI. 128,537-547 (1995).
[13] Heilker, R., U. Manning-Krieg, J. F. Zuber, M. Spiess: In vitro binding of clathrin adaptors to sorting signals correlates with
[14]
[15]
[16]
[17]
[18]
[19]
[20]
[21]
[22]
[23]
[24]
[25]
[26]
[27]
[28]
[29]
[30]
[31]
Signals for polarized sorting of Ii 31
endocytosis and basolateral sorting. EMBO J. 15, 2893-2899 (1996). Hunziker, w., C. Furney: A di-leucine motif mediates endocytosis and basolateral sorting of macrophage IgG Fc receptors in MDCK cells. EMBO J. 13,2963-2969 (1994). Hunziker, W., C. Harter, K. Matter, 1. Mellman: Basolateral sorting in MDCK cells requires a distinct cytoplasmic domain determinant. Cell 66, 907-920 (1991). Honing, S., W. Hunziker: Cytoplasmic determinants involved in direct lysosomal sorting, endocytosis, and basolateral targeting of rat Igp120 (lamp-I) in MDCK cells. J. Cell BioI. 128, 321-332 (1995). Kundu, A., D. P. Nayak: Analysis of the Signals for polarized transport of influenza virus (A/WSN/33) neuraminidase and human transferrin receptor, type II transmembrane proteins. J. Virol. 68, 1812-1818 (1994). Kunkel, T. A., J. D. Roberts, R. A. Zakour: Rapid and efficient site-specific mutagenesis without phenotypic selection. Methods Enzymol. 154,367-382 (1987). Le Bivic, A., Y. Sambuy, A. Patzak, N. Patil, M. Chao, E. Rodriguez-Boulan: An internal deletion in the cytoplasmic tail reverses the apical localization of human NGF receptor in transfected MDCK cells. J. Cell BioI. 115, 607-618 (1991). Lehmann, L. E., W. Eberle, S. Krull, V. Prill, B. Schmidt, C. Sander, K. von Figura, C. Peters: The internalization signal in the cytoplasmic tail of lysosomal acid phosphatase consists of the hexapeptide PGYRHV. EMBO J. 11, 4391-4399 (1992). Lin, S., H. Y. Nairn, M. G. Roth: Tyrosine-dependent basolateral sorting signals are distinct from tyrosine-dependent internalization signals. J. BioI. Chern. 272, 26300-26305 (1997). Lobel, P., N. M. Dahms, S. Kornfeld: Cloning and sequence analysis of the cation-independent mannose 6-phosphate receptor. J. BioI. Chern. 263, 2563-2570 (1988). Long, E.O., S. Rosen-Bronson, D. R. Karp, M. Malnati, R. P. Sekaly, D. Jaraquemada: Efficient cDNA expression vectors for stable and transient expression of HLA-DR in transfected fibroblast and lymphoid cells. Hum. Immunol. 31, 229-235 (1991). Marks, M. S., L. Woodruff, H. Ohno, J. S. Bonifacino: Protein targeting by tyrosine- and di-Ieucine-based signals: Evidence for distinct saturable components. J. Cell BioI. 135, 341-354 (1996). Matter, K., W. Hunziker, 1. Mellman: Basolateral sorting of LDL receptor in MDCK cells: the cytoplasmic domain contains two tyrosine-dependent targeting determinants. Cell 71, 741-753 (1992). Matter, K., J. A. Whitney, E. M. Yamamoto, 1. Mellman: Common signals control low density lipoprotein receptor sorting in endosomes and the Golgi complex of MDCK cells. Cell 74, 1053-1064 (1993). Matter, K., E. M. Yamamoto, 1. Mellman: Structural requirements and sequence motifs for polarized sorting and endocytosis of LDL and Fc receptors in MDCK cells. J. Cell BioI. 126, 991-1004 (1994). Mauxion, F., R. LeBorgne, H. Munier-Lehmann, B. Hoflack: A casein kinase II phosphorylation site in the cytoplasmic domain of the cation-dependent mannose 6-phosphate receptor determines the high affinity interaction of the AP-1 Golgi assembly proteins with membranes. J. BioI. Chern. 271, 2171-2178 (1996). McClelland, A., L. C. Kuhn, F. H. Ruddle: The human transferrin receptor gene: genomic organization, and the complete primary structure of the receptor deduced from a cDNA sequence. Cell 39, 267-274 (1984). Mellman, 1., E. Yamamoto, J. A. Whitney, M. Kim, W. Hunziker, K. Matter: Molecular sorting in polarized and non-polarized cells: Common problems, common solutions. J. Cell Sci. 106, Supp!. 17, 1-7 (1993). Miettinen, H. M., S. N. Edwards, M. Jalkanen: Analysis of transport and targeting of syndecan-1: effect of cytoplasmic tail deletions. Mol. BioI. CeliS, 1325-1339 (1994).

32 A. Simonsen, B. Bremnes, T. W. Nordeng, O. Bakke
[32]
[33]
[34]
[35]
[36]
[37]
[38]
[39]
[40]
(41]
[42]
[43]
[44]
[45]
[46]
[47]
[48]
Mostov, K. E., M. Friedlander, G. Blobel: The receptor for transepithelial transport of IgA and IgM contains multiple immunoglobulin-like domains. Nature 308, 37-43 (1984). Motta, A., B. Bremnes, M. A. C. Morelli, R. W. Frank, G. Saviano, O. Bakke: Structure-activity relationship of the leucinebased sorting motifs in the cytosolic tail of the major histocompatibility complex-associated invariant chain. J. BioI. Chern. 270,27165-27171 (1995) . Nagafuchi, A. , Y. Shirayoshi, K. Okazaki, K. Yasuda, M. Takeichi: Transformation of cell adhesion properties by exogenously introduced E-cadherin cDNA. Nature 329, 341-343 (1987). Niimi, M., Y. Nakai, Y. Aida: Identification of bovine invariant chain (Ii) gene by nucleotide sequencing. Biochem. Biophys. Res. Commun. 222,7-12 (1996). Odorizzi, C. G., I. S. Trowbridge, L. Xue, C. R. Hopkins, C. D. Davis, J. F. Collawn: Sorting signals in the MHC class II invariant chain cytoplasmic tail and transmembrane region determine trafficking to an endocytic processing compartment. J. Cell BioI. U6, 317-330 (1994). Odorizzi, G., I. S. Trowbridge: Structural requirements for major histocompatibility complex class II invariant chain trafficking in polarized Madin-Darby canine kidney cells. J. BioI. Chern. 272, 11757-11762 (1997). Ohno, H. , J. Stewart, M. C. Fournier, H. Bosshart, I. Rhee, S. Miyatake, T. Saito, A. Gallusser, T. Kirchhausen, J. S. Bonifacino: Interaction of tyrosine-based sorting signals with clathrinassociated proteins. Science 269, 1872-1875 (1995). Okamoto, C. T., S. P. Shia, C. Bird, K. E. Mostov, M. G. Roth: The cytoplasmic domain of the polymeric immunoglobulin receptor contains two internalization signals that are distinct from its basolateral sorting signal. J. BioI. Chern. 267, 9925-9932 (1992). Oshima, A., C. M. Nolan, J . W. Kyle, J. H. Grubb, W. S. Sly: The human cation-independent mannose 6-phosphate receptor. Cloning and sequence of the full-length cDNA and expression of functional receptor in COS cells. J . BioI. Chern . 263, 2553-2562 (1988). Pieters, J . , O. Bakke, B. Dobberstein: The MHC class 11-associated invariant chain contains two endosomal targeting signals within its cytoplasmic tail. J. Cell Sci. 106,831-846 (1993) . Pohlmann, R, C. Krentler, B. Schmidt, W. Schroder, G. Lorkowski, J. Culley, G. Mersmann, C. Geier, A. Waheed, S. Gottschalk: Human lysosomal acid phosphatase: cloning, expression and chromosomal assignment. EMBO J. 7,2343-2350 (1988). Pohlmann, R., G. Nagel, B. Schmidt, M. Stein, G. Lorkowski, C. Krentler, J. Cully, H. E. Meyer, K. H. Grzeschik, G. Mersmann: Cloning of a cDNA encoding the human cationdependent mannose 6-phosphate-specific receptor. Proc. Nat. Acad. Sci. USA 84,5575-5579 (1987) . Pond, L., L. A. Kuhn, L. Teyton, M. P. Schutze, J. A. Tainer, M. R. Jackson, P. A. Peterson: A role for acidic residues in dileucine motif-based targeting to the endocytic pathway. J. BioI. Chern. 270, 19989-19997 (1995). Prill, V., L. Lehmann, K. von Figura, C. Peters: The cytoplasmic tail of lysosomal acid phosphatase contains overlapping but distinct signals for basolateral sorting and rapid internalization in polarized MDCK cells. EMBO J. U, 2181-2193 (1993). Rajasekaran, A. K., J . S. Humphrey, M. Wagner, G. Miesenbock, A. LeBivic, 1. S. Bonifacino, E. Rodriguez-Boulan: TGN38 recycles basolaterally in polarized Madin-Darby canine kidney cells. Mol. BioI. Cell 5, 1093-1103 (1994) . Rapoport, I., M. Miyazaki, W. Boll, B. Duckworth, L. C. Cantley, S. Shoelson, T. Kircbhausen : Regulatory interactions in the recognition of endocytic sorting signals by AP-2 complexes . EMBO 1.16,2240-2250 (1997). Ringwald, M., R Schuh, D. Vestweber, H. Eistetter, F. Lottspeich, 1. Engel, R. Dolz, F. Jahnig, 1. Epplen, S. Mayer: The
(49)
[50]
[51]
[52]
[53]
[54]
[55]
[56]
[57]
[58]
[59]
[60]
[61]
[62]
[63]
[64]
structure of cell adhesion molecule uvomorulin. Insights into the molecular mechanism of Ca2+ -dependent cell adhesion. EMBO 1. 6,3647-3653 (1987). Roche, P. A., C. L. Teletski , E. Stang, O. Bakke, E. O. Long: Cell surface HLA-DR-invariant chain complexes are targeted to endosomes by rapid internalization. Proc. Natl. Acad. Sci. USA 90,8581-8585 (1993). Salamero, 1., R. Le Borgne, C. Saudrais, B. Goud, B. Hoflack: Expression of major histocompatibility complex class II molecules in HeLa cells promotes the recruitment of AP-l Golgispecific assembly proteins on Golgi membranes. J. BioI. Chern . m, 30318-30321 (1996) . Schafer, w., A. Stroh, S. BerghOfer, 1. Seiler, M. Vey, M.L. Kruse, H. F. Kern, H.-D . Klenk, W. Garten: Two independent targeting signals in the cytoplasmic domain determine transGolgi network localization and endosomal trafficking of the proprotein convertase furin . EMBO 1.14,2424-2435 (1995). Simons, K., S. D. Fuller: Cell surface polarity in epithelia. (Review). Annu. Rev. Cell BioI. 1,243-288 (1985). Simons, K., G. van Meer: Lipid sorting in epithelial cells . (Review). Biochemistry 27, 6197-{i202 (1988). Simonsen, A., E . Stang, B. Bremnes, M. Roe , K. Prydz, O. Bakke: Sorting of MHC class II molecules and the associated invariant chain (Ii) in polarized MOCK cells. 1. Cell Sci . 110, 597-{iQ9 (1997). Takahashi , S., T. Nakagawa, T. Banno, T. Watanabe, K. Murakami, K. Nakayama: Localization offurin to the trans-Golgi network and recycling from the cell surface involves Ser and Tyr residues within the cytoplasmic domain. 1. BioI. Chern. 270, 28397-28401 (1995). Thomas, D. C., C.B.Brewer, M.G.Roth: Vesicular stomatitis virus glycoprotein contains a dominant cytoplasmic basolateral sorting signal critically dependent upon a tyrosine. 1. BioI. Chern. 268, 3313-3320 (1993) . Thomas , D. c. , M. G. Roth: The basolateral targeting signal in the cytoplasmic domain of glycoprotein G from vesicular stomatitis virus resembles a variety of intracellular targeting motifs related by primary sequence but having diverse targeting activities. 1. BioI. Chern. 269, 15732-15739 (1994). Van Genderen, I. L" G. van Meer, 1. W. Slot, H. 1. Geuze, W. F. Voorhout: Subcellular localization of Forssman glycolipid in epithelial MDCK cells by immuno-electronmicroscopy after freeze-substitution. 1. Cell BioI. 115, 1009-1019 (1991). Voorhees, P., E.Deignan, E. Van Donselaar, 1. Humphrey, M. S. Marks, P. 1. Peters, J . S. Bonifacino: An acidic sequence within the cytoplasmic domain of furin functions as a determinant of trans-Golgi network localization and internalization from the cell surface. EMBO 1.14,4961-4975 (1995). Wandinger-Ness, A., M. K. Bennett, C. Antony, K. Simons: Distinct transport vesicles mediate the delivery of plasma membrane proteins to the apical and basolateral domains of MDCK cells. 1. Cell BioI. 111, 987-1000 (1990). Wigler, M., A. Pellicer, S. Silverstein, R. Axel, G. Urlaub, L. Chasin: DNA-mediated transfer of the adenine phosphoribosyltransferase locus into mammalian cells. Proc. Natl. Acad . Sci. USA 76, 1373-1376 (1979) . Wilde, A., C. Dempsey, G. Banting: The tyrosine-containing internalization motif in the cytoplasmic domain ofTGN38/41 lies within a nascent helix. J . BioI. Chern. 269,7131-7136 (1994). Wraight , C.l. , P.vanEndert, P.Moller, J .Lipp, N.R.Ling, I. C. MacLennan, N. Koch, G. Moldenhauer: Human major histocompatibility complex class II invariant chain is expressed on the cell surface. 1. BioI. Chern. 265, 5787-5792 (1990). Yokode, M., R. K. Pathak, R. E. Hammer, M. S. Brown, 1. L. Goldstein, R G. Anderson: Cytoplasmic sequence required for basolateral targeting of LDL receptor in livers of transgenic mice. 1. Cell BioI. 117, 39-46 (1992).
Copyright © 2022 FDOKUMEN




















