
The karst of the Vaucluse, an exceptional record for the Last Glacial Maximum (LGM) and the...
21
The karst of the Vaucluse, an exceptional record for the Last Glacial Maximum (LGM) and the Late-glacial period palaeoenvironment of southeastern France Evelyne Crégut-Bonnoure a, b, * , Jacqueline Argant c , Salvador Bailon d , Nicolas Boulbes e, f , Claude Bouville g , Jacques Buisson-Catil h , Evelyne Debard i , Emmanuel Desclaux j , Jan Fietzke k , Jean-Baptiste Fourvel b , Nicolas Frèrebeau l , Delphine Kuntz b , Jadwiga Krzepkowska a , Frédéric Laudet b , Thibault Lachenal e , Nicolas Lateur c , Alaric Manzano j , Adrian Marciszak m , Xavier Margarit c , Cécile Mourer-Chauviré n , Julien Oppliger o, p , Thierry Roger j , Amber G.F. Teacher q , Michel Thinon r a Muséum Requien, 67 rue Joseph Vernet 84000 Avignon, France b UMR 5608 TRACE (UTM), 5 allées Antonio Machado, 31058 Toulouse Cedex 1, France c Aix Marseille Université, C.N.R.S, MCC, Laboratoire Méditerranéen de Préhistoire Europe Afrique, U.M.R. 7269, 5 rue du Château de l’Horloge 13094, Aix-en-Provence Cedex 2, France d Muséum national d’Histoire naturelle, UMR 7209, Case 56, 55 rue Buffon, 75005 Paris, France e Université Montpellier 3, UMR 5140 Archéologie des Sociétés Méditerranéennes, 390 avenue de Pérols, 34970 Lattes, France f EPCC-Centre européen de Recherche, 66720 Tautavel, France g Les Calamuchons, chemin de Cabassude, 13530 Trets, France h Centre des monuments nationaux, BP 152, 56343 Carnac, France i Association de Recherche Paléoécologique en Archéologie, Département des Sciences de la Terre, Université Claude Bernard Lyon I, 69622 Villeurbanne Cedex, France j Laboratoire départemental de Préhistoire du Lazaret, 33bis Bd Franck Pilatte, 06000 Nice, France k GEOMAR Helmholtz Center for Ocean Research, Wischhofstr. 1-3, 24148 Kiel, Germany l UMR 5060 IRAMAT-CRP2A, Centre de recherche en Physique appliquée à l’archéologie, Esplanade des Antilles, Université de Bordeaux 3, 33607 Pessac Cedex, France m Division of Palaeozoology, Department of Evolutionary Biology and Ecology, Faculty of Biological Sciences, University of Wroclaw, Wroclaw, Poland n Laboratoire de Géologie de Lyon, Terre, Planètes et Environnement, UMR 5276, Université Claude Bernard Lyon I, Géode, 2 rue Dubois, 69622 Villeurbanne Cedex, France o Muséum d’histoire naturelle, Département d’archéozoologie, Route de Malagnou 1,1211 Genève 6, Suisse, France p Laboratoire d’archéologie préhistorique et anthropologie, Route des Acacias 18,1211 Genève 4, Suisse, France q Environment and Sustainability Institute, University of Exeter, Penryn Campus, Penryn Cornwall TR10 9EZ, Exeter, United Kingdom r Institut Méditerranéen d’Ecologie et de Paléoécologie de la Faculté de Saint-Jérôme Université d’Aix-Marseille Faculté de Saint-Jérôme Case, 461 Avenue Escadrille Normandie-Niemen, 13297 Marseille Cedex 20, France article info Article history: Available online xxx abstract Investigations in four natural traps from southeastern France have provided new and extensive infor- mation on the palaeoenvironment from the Last Glacial Maximum (LGM; 21,000 to 15,075 cal BP), the Late-glacial period (15,075 to 11,490 cal BP) and the beginning of Holocene (11,490 to 2835 cal BP). Radiocarbon and U/Th dates provided precise chronological data. In the Coulet des Roches, an LGM and a near complete Late-glacial period sequence were identified. The infilling preserved skeletons of large mammals, revealing a new faunal assemblage for southeastern France. For the first time, two distinct southern expansions of Dicrostonyx torquatus have been identified, correlated firstly to the LGM and secondly to the Late-glacial period. Dicrostonyx torquatus was found to be associated with Microtus oeconemus during the Oldest Dryas (15,075 to 18,270 cal BP). Morphological adaptations to the cold climate were indicated by some mammals (Vulpes vulpes, Mustela nivalis, Mustela erminea, Equus caballus gallicus). Among the birds, Bubo scandiacus and Pyrrhocorax graculus were found to be abundant. Paly- nological data suggested a very open landscape as well as a cold and rather dry climate. Rangifer tarandus remains were recovered from the Oldest Dryas layers of Aven des Planes. During the Allerød, a wooded * Corresponding author. Muséum Requien, 67 rue Joseph Vernet, 84000 Avignon, France. E-mail address: [email protected] (E. Crégut-Bonnoure). Contents lists available at ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint 1040-6182/$ e see front matter Ó 2013 Elsevier Ltd and INQUA. All rights reserved. http://dx.doi.org/10.1016/j.quaint.2013.10.058 Quaternary International xxx (2013) 1e21 Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst of the Vaucluse, an exceptional record for the Last Glacial Maximum (LGM) and the Late-glacial period palaeoenvironment of southeastern France, Quaternary International (2013), http://dx.doi.org/10.1016/ j.quaint.2013.10.058
-
Upload
culturecommunication-fr -
Category
Documents
-
view
2 -
download
0
Transcript of The karst of the Vaucluse, an exceptional record for the Last Glacial Maximum (LGM) and the...

lable at ScienceDirect
Quaternary International xxx (2013) 1e21
Contents lists avai
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
The karst of the Vaucluse, an exceptional record for the Last GlacialMaximum (LGM) and the Late-glacial period palaeoenvironment ofsoutheastern France
Evelyne Crégut-Bonnoure a,b,*, Jacqueline Argant c, Salvador Bailon d, Nicolas Boulbes e,f,Claude Bouville g, Jacques Buisson-Catil h, Evelyne Debard i, Emmanuel Desclaux j,Jan Fietzke k, Jean-Baptiste Fourvel b, Nicolas Frèrebeau l, Delphine Kuntz b,Jadwiga Krzepkowska a, Frédéric Laudet b, Thibault Lachenal e, Nicolas Lateur c,Alaric Manzano j, Adrian Marciszakm, Xavier Margarit c, Cécile Mourer-Chauviré n,Julien Oppliger o,p, Thierry Roger j, Amber G.F. Teacher q, Michel Thinon r
aMuséum Requien, 67 rue Joseph Vernet 84000 Avignon, FrancebUMR 5608 TRACE (UTM), 5 allées Antonio Machado, 31058 Toulouse Cedex 1, FrancecAix Marseille Université, C.N.R.S, MCC, Laboratoire Méditerranéen de Préhistoire Europe Afrique, U.M.R. 7269, 5 rue du Château de l’Horloge 13094,Aix-en-Provence Cedex 2, FrancedMuséum national d’Histoire naturelle, UMR 7209, Case 56, 55 rue Buffon, 75005 Paris, FranceeUniversité Montpellier 3, UMR 5140 Archéologie des Sociétés Méditerranéennes, 390 avenue de Pérols, 34970 Lattes, Francef EPCC-Centre européen de Recherche, 66720 Tautavel, Franceg Les Calamuchons, chemin de Cabassude, 13530 Trets, FrancehCentre des monuments nationaux, BP 152, 56343 Carnac, FranceiAssociation de Recherche Paléoécologique en Archéologie, Département des Sciences de la Terre, Université Claude Bernard Lyon I,69622 Villeurbanne Cedex, Francej Laboratoire départemental de Préhistoire du Lazaret, 33bis Bd Franck Pilatte, 06000 Nice, FrancekGEOMAR Helmholtz Center for Ocean Research, Wischhofstr. 1-3, 24148 Kiel, GermanylUMR 5060 IRAMAT-CRP2A, Centre de recherche en Physique appliquée à l’archéologie, Esplanade des Antilles, Université de Bordeaux 3,33607 Pessac Cedex, FrancemDivision of Palaeozoology, Department of Evolutionary Biology and Ecology, Faculty of Biological Sciences, University of Wrocław, Wroclaw, Polandn Laboratoire de Géologie de Lyon, Terre, Planètes et Environnement, UMR 5276, Université Claude Bernard Lyon I, Géode, 2 rue Dubois,69622 Villeurbanne Cedex, FranceoMuséum d’histoire naturelle, Département d’archéozoologie, Route de Malagnou 1, 1211 Genève 6, Suisse, Francep Laboratoire d’archéologie préhistorique et anthropologie, Route des Acacias 18, 1211 Genève 4, Suisse, Franceq Environment and Sustainability Institute, University of Exeter, Penryn Campus, Penryn Cornwall TR10 9EZ, Exeter, United Kingdomr Institut Méditerranéen d’Ecologie et de Paléoécologie de la Faculté de Saint-Jérôme Université d’Aix-Marseille Faculté de Saint-Jérôme Case,461 Avenue Escadrille Normandie-Niemen, 13297 Marseille Cedex 20, France
a r t i c l e i n f o
Article history:Available online xxx
* Corresponding author. Muséum Requien, 67 rue JE-mail address: [email protected]
1040-6182/$ e see front matter � 2013 Elsevier Ltd ahttp://dx.doi.org/10.1016/j.quaint.2013.10.058
Please cite this article in press as: Crégut-B(LGM) and the Late-glacial period palaeoej.quaint.2013.10.058
a b s t r a c t
Investigations in four natural traps from southeastern France have provided new and extensive infor-mation on the palaeoenvironment from the Last Glacial Maximum (LGM; 21,000 to 15,075 cal BP), theLate-glacial period (15,075 to 11,490 cal BP) and the beginning of Holocene (11,490 to 2835 cal BP).Radiocarbon and U/Th dates provided precise chronological data. In the Coulet des Roches, an LGM and anear complete Late-glacial period sequence were identified. The infilling preserved skeletons of largemammals, revealing a new faunal assemblage for southeastern France. For the first time, two distinctsouthern expansions of Dicrostonyx torquatus have been identified, correlated firstly to the LGM andsecondly to the Late-glacial period. Dicrostonyx torquatus was found to be associated with Microtusoeconemus during the Oldest Dryas (15,075 to 18,270 cal BP). Morphological adaptations to the coldclimate were indicated by some mammals (Vulpes vulpes, Mustela nivalis, Mustela erminea, Equus caballusgallicus). Among the birds, Bubo scandiacus and Pyrrhocorax graculus were found to be abundant. Paly-nological data suggested a very open landscape as well as a cold and rather dry climate. Rangifer tarandusremains were recovered from the Oldest Dryas layers of Aven des Planes. During the Allerød, a wooded
oseph Vernet, 84000 Avignon, France.m (E. Crégut-Bonnoure).
nd INQUA. All rights reserved.
onnoure, E., et al., The karst of the Vaucluse, an exceptional record for the Last Glacial Maximumnvironment of southeastern France, Quaternary International (2013), http://dx.doi.org/10.1016/

E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e212
Fig. 1. Location of themain archeological localities and thGorges of “La Nesque”: Unang, Abri Edward, Abri Gauthi
Please cite this article in press as: Crégut-B(LGM) and the Late-glacial period palaeoej.quaint.2013.10.058
environment permitted the dispersal of Cervus elaphus, Sus scrofa, Tetrao urogallus as well as reptiles andamphibians. Holocene sequences existed in these two natural traps as in Aven Souche n� 1 and n� 2. Atthis time, a sparsely wooded landscape covered much of the area. Cervus elaphus and Tetrao urogalluswere still present with Canis lupus, Lynx lynx and Felis silvestris. Areas of open landscape were occupiedby Otis tarda. During the Bronze Age (2200 to 800 cal BC) and Iron Age (800 to 50 cal BC), Aven desPlanes, Aven Souche n� 1 and n� 2 were used by humans and had a sepulchral destination.
� 2013 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
Today, climate change is of concern because of its impact on theenvironment. For a long time, scientists have utilized oceanic andcontinental deposits to elucidate long-term climatic patterns inEarth’s history. Among these, the karst infillings have preservedclimatological archives, making climatic and palaeoenvironmentalreconstructions possible.
Recently, numerous studies have been carried out in Europe toclarify the chronology of the end of the Pleistocene. However,complete sequences are scarce, and the climatic changes observed,associated with chronological data, are interpreted differently(Miller, 2012; Van Raden et al., 2013). In southeastern France, the
e natural traps in Vaucluse.1: Chincher. 4: Coulet des Roches, Aven des Pl
onnoure, E., et al., The karst onvironment of southeastern
data collected from archeological caves has remained incompletefor the end of the Pleistocene.
Since 1970, several small pits about ten feet deep, bordering theVaucluse highlands, have been discovered. This area is a mosaic ofplains, plateaus, high reliefs, limestone and siliceous soils. The karstCretaceous limestone area is famous for the great depth of its holesformed during the Messinian Salinity Crisis (MSC) and corre-sponding to the drying up of the Mediterranean Sea (about 6 Ma).However, many of these holes are shallow, and some played therole of natural traps for the local fauna. Four main deposits inproximity retrace the major environmental transformations of thekarst plateau (700 to 800 m-asl): Coulet des Roches, aven desPlanes n� 1 and aven Souche n�1 and n�2.
on 1 on the borderwith Comtadine plain. 2: Luberonmountain: Roquefure, Soubeyras. 3:anes, Aven Souche n� 1 and n� 2. Map modified from Geoatlas and from Olive (2004).
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 3
2. Background
From an archeological point of view, Vaucluse is rich in rockshelters and caves. Some of the infillings weremainly formed duringthe end of the Pleistocene, more precisely during the end of the LateGlacial Maximum (LGM; 21,000 to 15,075 cal BP), the Late-glacial(15,075 to 11,490 cal BP) and the beginning of Holocene (11,490 to2835 cal BP). These infillings preserved fragmentary faunal remainsin association with artefacts (Crégut-Bonnoure, 2008). Thesearcheological sites are located in three distinct geographical units(Fig. 1), and were excavated several times in the 1950s and 1960s(Paccard, 1954, 1956, 1961, 1962, 1963; Gauthier and Paccard, 1962).Consequently, some chronological attributions, such as the radio-carbon dates, are imprecise, and the interpretation of some strati-graphical levels is debatable (Brochier, 1977; Brochier and Livache,2004). The faunal remains are fragmentary and no information onthe vegetation is available. Therefore, the palaeoenvironment of theendof thePleistocene and thebeginningofHolocene ispoorly knownin these southeastern sites, in contrast to southwestern France.
3. New Karst infillings
The discovery of four natural traps has provided the opportunityto study new sedimentary sequences with updated scientific ap-proaches (Fig. 1). The field research and analyses, initiated in 2005,required a multidisciplinary cooperation, especially between
Fig. 2. Coulet des Roches. Cross section. Topographic
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
scientists and spelunkers. This project has provided extensive in-formation on the climate and palaeoenvironments from 21,000 BPto 11,490 cal BP, and into the Iron Age (800 to 50 cal BC). Thevertebrate fauna appears exceptional for this area. The extractedmaterial is preserved in the Museum Requien in Avignon.
3.1. Coulet des Roches
Coulet des Roches was opened by spelunkers from Carpentras in1971 (Jean, 1973, 1977; Daniel, 1979; Parein and Languille, 1981;Parein, 1990; Le Fahler and Perret, 1995). Our investigationsbegan in 2007 (Crégut-Bonnoure et al., 2007a, 2008, 2009a, 2010a,2011a, 2012). The ellipsoidal opening is about 4 � 2 m. The hole is10 m deep andwidens rapidly (Fig. 2). At the bottom there is a largeroom, 10 m long and averaging 3e4 m in width.
The sediments, extracted from 1971 to 1973, have been screenedby our team using 2.25 mm, 1.4 mm and 1.10 mm screen mesh.Thousands of bones were found. A sequence holding three strati-graphic units was identified. Unfortunately, part of this sequencewas destroyed by the spelunkers who did not realize the paleon-tological significance of these levels. At the beginning of our in-vestigations, Unit 1 was located on both on the western wall (5 mlong and 0.3 m deep) and also in the south corner of the room (in alayer 0.4 m deepwith an area of 2.5m2). Unit 2 was an area of about20 m2. Unit 3 was revealed by digging by the spelunkers in thenorthern corner. Stalagmitic floors and flowstones were visible on
al survey: Ch. Boucher, redrawn by J.-B. Fourvel.
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Table 1U/Th dates. For the correction of detrital 230Th, a 230Th/232Th activity ratio of 0.6 � 0.2 was used. Details of the method used can be found in Fietzke et al. (2005). Abbreviation: Stal. floor: Stalagmitic floor.
Sample Comments Ageky
� ky min-Ageky
max-Ageky
U238ppm
� ppm Th232ppb
� ppb Th230ppt
� ppt Th230/Th232dpm/dpm
� dpm/dpm
U238/Th232dpm/dpm
� dpm/dpm
Th230/U238dpm/dpm
� dpm/dpm
Th230excess/U238dpm/dpm
� dpm/dpm
U234/U238dpm/dpm
� dpm/dpm
U234/U238initial
Aven Souche1 Stal. floor 502 31 474.8 537.3 0.621 0.000 7.5725 0.0090 10.2308 0.012 252.250 0.4 253.9 0.3 0.9936 0.0013 0.99123 0.00159 1.00097 0.0009 1.0040Coulet des
RochesJ3 Nr 1
Alveolar layer5.61 0.04 5.6 5.7 0.179 0.000 2.7396 0.0033 0.1579 0.001 10.763 0.1 202.1 0.3 0.0533 0.0003 0.05029 0.00031 1.00429 0.0015 1.0044
Coulet desRoches 2
Stal. floorEast
452 32 424.3 488.3 0.865 0.001 18.1250 0.0693 14.3271 0.043 147.583 0.7 147.8 0.6 0.9985 0.0031 0.99448 0.00327 1.00768 0.0010 1.0275
Coulet desRoches 3
Flowstone 10.7 0.1 10.6 10.7 0.369 0.000 8.7530 0.0141 0.5938 0.002 12.666 0.0 130.5 0.2 0.0971 0.0003 0.09246 0.00034 0.99423 0.0011 0.9941
Coulet desRochesJ4 Nr 4
Stal. floorWest
356 9 346.8 365.5 1.137 0.001 41.4033 0.0867 18.4521 0.028 83.209 0.2 85.0 0.2 0.9786 0.0016 0.97157 0.00199 1.00776 0.0011 1.0212
Comments Age ky � ky min-Age ky max-Age ky Laboratory number
Stalagmitic floor 502 31 474.8 537.3 Axiom 73-12
Stalagmitic floor East 452 32 424.3 488.3 Axiom 75-12Stalagmitic floor West 356 9 346.8 365.5 Axiom 77-12Flowstone 10.7 0.1 10.6 10.7 Axiom 76-12Alveolar layer 5.61 0.04 5.6 5.7 Axiom 74-12
E.Crégut-Bonnoureet
al./Quaternary
Internationalxxx
(2013)1e21
4Pleasecite
thisarticle
inpress
as:Crégut-Bonnoure,E.,et
al.,Thekarst
ofthe
Vaucluse,an
exceptionalrecordfor
theLast
GlacialM
aximum
(LGM)and
theLate-glacial
periodpalaeoenvironm
entof
southeasternFrance,
Quaternary
International(2013),
http://dx.doi.org/10.1016/j.quaint.2013.10.058

E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 5
the walls, and have been dated by Uranium/Thorium methods(Fietzke et al., 2005). The dating of the stalagmitic floors indicatesimportant flows of water at two distinct times in the MiddlePleistocene, during the MIS 13 and the MIS 11 (Schackleton andOpdyke, 1973; Table 1), in relation with the speleogenesis mecha-nisms of the local karst. Unit 1 appeared to be covered by stalag-mitic floors on the western wall, but it continues under thesefloors.
Radiocarbon dating of numerous bones and charcoal date theCoulet infillings to the end of the Pleistocene and the beginning ofthe Holocene (Table 2). Bird remains were very numerous: 5121bird bones were identified. Among them, we found 1858 uniden-tifiable bones of young Corvids. Most of identified bones of adultCorvids belong to the Alpine Chough (Pyrrhocorax graculus). It islikely that the majority of the remains of young Corvids belong tothis species. Nowadays, the Alpine Chough lives on cliff ledges or incaves, sometimes deep down and often in darkness. On that basis,we interpret Coulet des Roches as a potential former nesting site forthe Alpine Chough.
3.1.1. Extracted sedimentsThousands of bones were collected and 78 species were
identified: 18 large mammals, 2 insectivores, 5 bats, 2 lago-morphs, 9 rodents, 24 birds, 11 reptiles, and 7 amphibians(Tables 3 and 4). The faunal assemblage was heterogeneous. Someof the species came from the black loam level, others from thecryoclastic level, and others are more recent. Amphibians andreptiles were abundant: 11,928 bones were identified followingthe general criteria given by Bailon (1991, 1999) and Haller-Probst(1997) for amphibians, and Szyndlar (1984, 1991a,b), Bailon(1991), Barahona and Barbadillo (1997) and Blain (2009) forreptiles. A total of 157 ceramic remains, characteristic of the end
Fig. 3. Coulet des Roches, view of the western wall. Above, cross section (Topographical surloam level” above the “cryoclastic unit”. Scales: 50 cm (vertical) and 1 m (horizontal).
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
of the Neolithic and of the beginning of the Bronze Age (2200 to800 cal BC), were discovered. Burned bones were also found inthis deposit, dating from the Subboreal (5450 to 2835 cal BP):3610 � 40 BP (calibration intCal 09; Beta analytic 288277)(Table 2).
One of the three wolves (Canis lupus) was dated from theAtlantic episode (8835 to 5450 cal BP): 5970 � 40 BP (Beta analytic288945), as was the lynx (Lynx lynx; 6590 � 30 BP and 7260 � 40BP; Beta analytic 330094, 330095). Lynx DNA analyses are inprogress by Ricardo Rodríguez-Valera from Centro Mixto UCM-ISCIII de Evolución y Comportamiento Humanos (Madrid).
3.1.2. Unit 1: Black loam levelThis level was partially destroyed by spelunkers. Its trace was
visible against the walls with two scraps, one of them, on thewestern wall, showing alveolar calcification (¼alveolar layer) inrelationwith the flow of water (Fig. 3). The other onewas located atthe southeastern area of the cavity. A total of 33 species wereidentified: 3 large mammals, 2 insectivores, 3 rodents, 3 bats, 5birds, 12 reptiles, and 5 amphibians (Tables 3 and 4). In the alveolarlayer, no fauna were discovered.
Among the mammals, the Boar (Sus scrofa) was represented byone young animal and the Chamois (Rupicapra rupicapra) by oneadult. The chronological attribution is difficult. This level seemedto cover one of the last climatic oscillations of the Late-glacialperiod: either the end of Allerød, either the beginning ofYounger Dryas (12,710 to 11,490 cal BP). The date of the Red Deer(one adult) was 10,360 � 60 BP (Beta Analytic 267376; Table 2).The presence of the Western Capercaillie (Tetrao urogallus) in-dicates a forest environment under a rather cool climate. On theother hand, the amphibian and reptile association indicates aMediterranean climate with mountain influences (supra-
vey: Ch. Boucher, E. Debard). Down, photography (E. Crégut-Bonnoure). See the “black
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Table 2Coulet des Roches, Les Planes, Aven Souche n� 1. Radiocarbon dates.
Chronozones Not calibratedConventionalage
Calibrated BPIntCal 09
Locality Reference ofthe analysis
COULET DES ROCHES
Datations Dated sample
Conventional age Calibrated BP Calibrated BC Sample Reference
HoloceneSubatlantic Souche 1 BA 288280 2340 � 40 BP 2370e2320 BP 420e370 BC Homo sapiens; She1 506-6
2440e2410 BP 490e460 BC
Subboreal 4700e2700 BP 5450e2835 BP Coulet BA 288277 3610 � 40 BP 4070e4040 BP 2120e2090 BC Burned bone; CdR OBR Extracted sediments3990e3830 BP 2040e1880 BC
Planes Lyon 3141(Poz) 3255 � 35 BP 1616e1640 BC C. elaphus; MR4.01.915
Atlantic 8000e4700 BP 8835e5450 BP Coulet BA 288945 5970 � 40 BP 6900e6720 BP 4950e4770 BC C. lupus; CdR R 575 Extracted sedimentsCoulet Axiom 7412 5610 � 40 BP Flowstone Alveolar layerCoulet BA 330094 6590 � 30 BP 7560e7540 BP 5610e5590 BC Lynx lynx ; CdR R 5548 Extracted sediments
7520e7430 BP 5570e5480 BCCoulet BA 330095 7260 � 40 BP 8170e8000 BP 6220e6050 BC Lynx lynx ; CdR R 5801 Extracted sediments
7990e7980 BP 6040e6030 BC
Boreal 9000e8000 BP 10,085e8835 BP Souche 1 BA 308071 8070 � 40 BP 9070e9060 BP 7120e7110 BC Juniperus communis; charcoal9030e8980 BP 7080e7030 BC She1 2 R12
Preboreal 10,000e9000 BP 11,490e10,085 BP Coulet BA 267382 9270 � 50 BP 10,580e10,270 BP 8630e8320 BC Charcoal Alveolar layerSouche 1 BA 308070 9670 � 40 BP 11,200e11,070 BP 9250e9120 BC C. elaphus adult; She1 4 175 Red deer Unit
10,950e10,870 BC 9000e8920 BPLate-GlacialDryas III 10,800e10,000 BP 12,710e11,490 BP Coulet BA 267376 10,360 � 60 BP 12,560e12,500 BP 10,610e10,550 BC C. elaphus adult; M7 4 Black loam levelAllerød 11,800e10,800 BP 13,600e12,710 BP Coulet Axiom 7612 10,700 � 100 BP Flowstone
Coulet BA 267375 10,750 � 70 BP 12,860e12,690 BP 10,910e10,740 BC C. elaphus young; J3 119 Cryoclastic levelCoulet BA 267383 11,060 � 60 BP 13,090e12,880 BP 11,140e10,940 BC Charcoal; J3 Z ¼ 152; Cryoclastic level
Dryas II 12,000e11,800 BP 13,900e13,600 BP Coulet BA 267377 11,740 � 70 BP 13,750e13,420 BP 11,800e11,470 BC C. elaphus adult; N5 49 Cryoclastic levelBølling 12,700e12,000 BP 15,075e13,900 BP Coulet BA 267381 12,380 � 70 BP 14,720e14,100 BP 12,770e12,150 BC C. ibex ; N3(5) 1 Cryoclastic levelDryas I 15,000e12,700 BP 18,270e15,075 BP Planes Lyon 3138(Poz) 12,660 � 75 BP 13,287e12,550 BC R. tarandus; MR4.01.759 E Level
Coulet BA 267379 13,090 � 70 BP 15,660e15,260 BP 13,720e13,310 BC E. c. gallicus adult ; K4 9 Cryoclastic levelCoulet BA 267380 13,110 � 70 BP 15,690e15,280 BP 13,740e13,340 BC E. c. gallicus adult; N5 76 Cryoclastic levelPlanes Lyon 3139(Poz) 13,360 � 80 BP 14,343e13,513 BC E. c. gallicus E LevelPlanes BA 329247 13,730 � 60 BP 16,910e16,790 BP 14,960e14,840 BC E. c. gallicus ; MR4.01.834 D Level
Last Glacial MaximumCoulet BA 267378 15,070 � 80 BP 18,650e18,070 BP 16,700e16,120 BC E. c. gallicus adult; K3 111 Cryoclastic levelCoulet BA 288279 15,920 � 60 BP 19,210e18,980 BP 17,260e17,030 BC C. ibex female; M3 137-2 Cryoclastic levelCoulet BA 308067 16,380 � 50 BP 19,560e19,430 BP 17,610e17,480 BC C. ibex male; N5 283 Cryoclastic levelCoulet BA 308066 17,950 � 80 BP 21,520e21,320 BP 19,570e19,370 BC E. c. gallicus mare-foetus; N4 289 Cryoclastic levelCoulet BA 329249 18,020 � 120 BP 21,670e21,320 BP 19,720e19,370 BC E. c. gallicus stallion; M4 547 Cryoclastic level
E.Crégut-Bonnoureet
al./Quaternary
Internationalxxx
(2013)1e21
6Pleasecite
thisarticle
inpress
as:Crégut-Bonnoure,E.,et
al.,Thekarst
ofthe
Vaucluse,an
exceptionalrecordfor
theLast
GlacialM
aximum
(LGM)and
theLate-glacial
periodpalaeoenvironm
entof
southeasternFrance,
Quaternary
International(2013),
http://dx.doi.org/10.1016/j.quaint.2013.10.058

Table 3Coulet des Roches, Les Planes, Aven Souche n� 1 and n� 2. Mammal distribution.
Coulet des Roches Aven des Planes Aven Souche n� 1 Aven Souche n� 2
Extractedsediment
Allerød orYoungerDryas
Allerød OldestDryas
LGM Extractedsediment
B Unit C UniOldestDryast
Unit withdomesticfauna
Unit withRed deer
Unit with domesticfauna
CarnivoraCanis lupus �Canis familiaris � � � �Vulpes vulpes � � � � � � � � �Alopex lagopus �Lynx lynx � �Felis silvestris � � �Meles meles � �Martes foina �Mustela (Putorius) putorius � � �Mustela erminea � �Mustela nivalis � � �PerissodactylaEquus gallicus � � � � �ArtiodactylaSus scrofa � � � � � � �Rangifer tarandus � � �Cervus elaphus � � � � � � � �Capreolus capreolus � � � � � �Bos taurus � � � �Ovis aries � � � � � �Capra hircus � � � �Capra ibex � � �Rupicapra rupicapra � �InsectivoraTalpa europaea � � � � �Erinaceus europaeus � �Sorex sp. � � �Sorex minutus � �Sorex araneus � �ChiropteraRhinolophus ferrumequinum � �Rhinolophus euryale �Barbastella barbastellus � �Myotis myotis � �M. myotis/blythii �Plecotus cf. austriacus � �LagomorphaOryctolagus cuniculus � � � � � � � � �Lepus timidus � � � � �Lepus europaeus � � � �RodentiaMarmota marmota � � �Sciurus vulgaris � �Myoxus glis � �Myodes glareolus � � � cf.Eliomys quercinus � � � �Apodemus sylvaticus � � � � �Arvicola amphibius � � � �Arvicola ambibius/sapidus �Microtus arvalis � � � � �Microtus arvalis/agrestis � �Chionomys nivalis � � � � � �Microtus (Terricola)
duodecimcostatus� �
Microtus (Terricola)cf. multiplex
� �
Microtus oeconomus � �Dicrostonyx torquatus � �
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 7
Mediterranean bioclimatic stage), but hot enough during thesummer months to allow for the reproduction of thermophile taxasuch as Montpellier Snake (Malpolon monspessulanus) (i.e. anaverage temperature for summer at or above 22 �C and averageannual temperature exceeding 10 �C; Cheylan et al., 1981;Pleguezuelos, 1997).
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
The palynological data revealed a sparsely wooded landscapewith Pinus, Hazel Tree (Corylus), Elm (Ulmus), Lime (Tilia), Ash(Fraxinus) and Ephedra. Caryophyllaceae and Artemisia were rela-tively abundant (Fig. 4, sample L3). Juniper (Juniperus communis)charcoal was also present, and 14C dating revealed one piece ofcharcoal to be 9240� 50 BP (Beta analytic 26382), corresponding to
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Table 4Coulet des Roches, Les Planes, Aven Souche n� 1 and n� 2. Bird, reptile, and amphibian distribution.
Coulet des Roches Aven des Planes Aven Souche n�1 Aven Souche n� 2
Extractedsediment
Allerød orYounger Dryas
Allerød OldestDryas
LMG Extractedsediment
B Unit C Unit OldestDryas
Unit withdomestic fauna
Unit withRed deer
Unit with domesticfauna
AvesAquila chrysaetos � � � �Buteo buteo �Accipiter nisus �Circus sp. �Circus macrourus �Circus cf. cyneus �Circaetus gallicus �Falco sp. �Falco cf. peregrinus � �Falco cf. columbarius �Lagopus mutus � � �Tetrao urogallus � � � � �Alectoris rufa �Perdix perdix � � �Coturnix coturnix � �Rallus aquaticus � �Otis tarda �Columba livia � �Columba oenas � � �Columba livia-oenas � � � � �Columba palumbus � �Columba sp. �Bubo scandiacus � � �Strix aluco �Apus apus �Calandrella brachydactyla �Coracias garrulus �Hirundo sp. � � � � �Turdus sp. �cf. Turdus merula �Passeriforma indet. �Garrulus glandarius �Pyrrhocorax pyrrhocorax � �Pyrrhocorax graculus � � � � � �Corvus monedula �Corvus corone � �Pyrrhocorax-Corvus �Corvidae indet. � � �ReptiliaChalcides striatus �Lacerta viridis cf.Lacerta bilineata � � �Timon lepidus � �Podarcis cf. muralis � �Anguis fragilis � � �Coronella cf. girondica � �Coronella sp. �Malpolon monspessulanus � � �Rhinechis scalaris � �cf. Zamenis longissimus � �Natrix cf. maura � �Natrix natrix � � �Vipera aspis � � cf. �Vipera sp. cf.
AmphibiaSalamandra salamandra � �Pelodytes punctatus � �Bufo bufo � � � cf.Bufo calamita � �Pelophylax sp. �Rana cf. dalmatina �Rana sp. � � �
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e218
the Holocene: Preboreal (11,490 to 10,085 cal BP). The concretionswere dated by UraniumeThorium and correspond to the Atlanticphase (8835 to 5450 cal BP): 5610 � 40 BP (Table 1). The anthra-cological data indicated the development of Scots Pine (Pinus syl-vestris), White Sorb (Sorbus aria) and Juniper.
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
3.1.3. Unit 2: top of the Cryoclastic levelSixteen species were present (Tables 3 and 4): 2 large mammals,
2 insectivores, 8 rodents, 4 birds. The great variety of rodents whichnowadays occur in temperate and forested habitats, such as denDormouse (Eliomys quercinus), Wood Mouse (Apodemus sylvaticus),
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Fig. 4. Coulet des Roches. Pollen analysis (simplified).
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 9
Bank Vole (Myodes glareolus), Mediterranean Pine Vole (Microtus(Terricola) duodecimcostatus) and Alpine Pine Vole (M. (T.) cf.multiplex), likely to date from the middle or the end of the Late-glacial period: Bølling (15,075 to 13,900 cal BP) or Allerød (13,600to 12,710 cal BP). The absolute dating allows the assignment of thislevel to the Allerød (Table 2). Two Red Deer (Cervus elaphus) werepresent: one was a young animal (8/9 months old) which dated to10,660 � 70 BP (Beta analytic 267375), and the other was a 4e5year old male, dating to 11,740 � 70 BP (Beta analytic 267377)(Table 2).
Charcoal was abundant. The anthracological data indicated thedevelopment of Scots Pine and White Sorb, a thermophile species,which likely actively contributed to the reforestation. One ScotsPine charcoal piece was dated to 11,060 � 60 BP (¼Allerød; Betaanalytic 26783; Table 2). The excavation of the top of the cryoclasticlevel revealed white flowstone on the western wall, dated by theUraniumeThorium method to 10,700 � 100 BP (Axiom n� 75-12;Table 1), and thus in accordance with the Allerød period.
3.1.4. Unit 2: the cryoclastic levelThis level was essentially formed by the accumulation of frag-
ments of limestone of small size grading. This resulted from a thick
Fig. 5. Vulpes vulpes. Ratio-diagram of the length of the limb bones. 0: currentV. vulpes. Abbreviations: juv.: juvenile. Extr.sed.: extracted sediments. Measurementsfor Coulet in Table 3.
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
surface deposit formed during the LGM, which was then graduallywashed away to the bottom of the hole by runoff. In the southcorner of the room, the sediment is soil-like, resulting from waterdraining through a crack. The analysis of clays indicates that thisdeposit is distinct from the previous deposits, and that it consti-tutes a sedimentary unit resulting from waterflows.
At least 29 species are present: 6 large mammals, 1 insectivore,2 lagomorphs, 7 rodents, and 13 birds (Tables 3 and 4). The dis-tribution of the species is variable from the top to the bottom ofthe cryoclastic unit and the radiocarbon dating permits assign-ment to the LGM and the Oldest Dryas (15,075 to 18,270 cal BP)(Table 2). The peripheral position of carcasses, the absence of adraw point and of a central talus fan, combined with the paleo-climatical data, allow us to propose the hypothesis of a drain atthe bottom, channeling bodies out of the opening. The tapho-nomic analysis reveals a significant role of the circulation of waterin the reorganization of skeletons. Dissolution marks (large sur-face S or pits P, bone surface disintegration, bone cracking andcalcite deposits) are numerous. The absence of anatomicalconnection also supports the bone dispersal hypothesis, althoughcomplete individuals are restricted to a small area of 5e6 squaremeters.
Among Carnivores, the Red Fox (Vulpes vulpes) was presentduring the LGM (Minimum Number of Individuals ¼ 2) and theOldest Dryas (MNI¼ 1). The foxes from the Dryas I appeared to havea morphological particularity. They had longer and more slenderlimb bones compared to modern foxes. The lengthening and nar-rowing of the tibiawas more pronounced than for the femur (Fig. 5;Table 5). The DNA analysis does not show evidence of a separatelineage (Teacher et al., 2011). The cohabitation with cold-adaptedspecies supports the conclusion from the DNA analysis that theRed Fox remained in Europe throughout the glaciations, withoutbeing forced into glacial refugia and undergoing subsequent re-emergence and population expansion during warmer periods(Teacher et al., 2011). In horses, the proportions of limb-bones areconsistent with the environmental features (Eisenmann,1984) as inthe current Caprinae (Crégut-Bonnoure, 2002). On that basis, weinterpret the specificity of the Coulet Red Fox as a potential adap-tation to the development of an open landscape under a dryclimate.
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Table 5Vulpes vulpes measurements. Coulet des Roches and Les Planes. Length of the limbbones (in mm).
Length
Humerus Radius Femur Tibia
Vulpes vulpesCurrentN 20 19 24 23Extremes 105.6e130.6 101.3e123.8 114.6e143.2 124e150Mean 121.64 114.42 129.25 137.47
Coulet des RochesLast Glacial Maximum2012 N ¼ 2 139.1 130.1 147 152.42010 N ¼ 2 135.7 127.7 138.8 156.9
Dryas I2008 N ¼ 2 130.3 123.5 134.0 148.0
Extracted sedimentN 1 2 5 4Extremes 126.6 116.0e116.1 126.8e137.5 143.1e151.4Mean e 116.1 132.4 147.1
Les PlanesDryas IN ¼ 1e2 138.9 e 146.8 161.5
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e2110
The Stoat (Mustela erminea) and Weasel (M. nivalis) remains arecharacterized by the presence of small and medium size speci-mens. The Stoat differs from Weasels in having a larger size, a lesspronounced postorbital bar, a larger foramen infra orbitalae, a
Table 6Mustela erminea and Mustela nivalis measurements of the skull (in mm). Coulet des Ro(Brantes, Vaucluse; Holocene). Abbreviations: W zyg: zygomatic width. CBL: condylo ba
Skull. male Coulet des Roches
Mustela nivalis
R 2878 R 8857 R 7057 R 8856 R 435
CBL 40.3 35.2 37.6 34.2 37.3W zyg 21.2 e 19.7 e e
Skull. female Mustela nivalis
Coulet des Roches
R 8858 N3 N4 2
CBL 30.7 33.4W zyg e 17.4
Table 7Mustela erminea andMustela nivalis measurements of the lower M1 (in mm). Coulet des RAbbreviations: L: length; W: width; trig.: trigonid; tal.: talonid.
Female Mustela nivalis
Coulet des Roches
R 3037 R 3039 R 8858 R 8858
LM1 3.88 3.33 3.66 3.63L trig. 2.76 2.52 2.68 2.73L tal. 0.81 0.68 0.93 1.02W trig. 1.55 1.49 1.35 1.33
Male Mustela nivalis e Coulet des Roches
R 3034 R 3035 R 3719 R 3059 R 3063 R 9294 M3 61
LM1 4.01 4.25 4.38 4.53 4.43 4.33 4.19L trig. 2.66 2.94 3.28 3.29 3.43 3.22 2.93L tal. 1.28 1.27 1.09 1.17 1.24 1.29 1.52W trig. 1.32 1.58 1.66 1.90 1.50 1.61 1.43
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
round shape of nasalia (in case of weasel nasalia are oval), a widerposterior part of the crown of lower P4, a longer trigonid, morereduced talonid and strongly marked widening of the crown at thebase of the protoconid of the lower M1 (Heptner and Naumov,1967; Altuna, 1971; Stresemann, 1974; Hugueney, 1975; Clot,1980; Buchalczyk, 1984; Baryshnikov and Golovanova, 1989;Delpech, 1989; Reichstein, 1993a,b; Baryshnikov, 2009). In-dividuals of M. nivalis are slightly smaller than those occurringtoday in France (Heptner and Naumov, 1967) (Fig. 6; Table 6). Thesize of mandibles and long bones were generally comparable withthe dimensions of weasels from other Late Pleistocene Frenchlocalities such as Coudes (Puy-de-Dôme), La Colombière (Poncin,Ain), La Baume de Gigny (Jura), Lourdes Calvaire (Hautes-Pyr-énées) or Grotte du bois de Cantet (Espèche, Hautes-Pyrénées)(Delpech, 1973, 1989; Hugueney, 1975; Clot, 1980; Clot et al., 1984).Stoat material from Coulet des Roches is more comparable withthe medium size subspecies Mustela erminea aestiva which nowoccurs mostly in central Europe. Remains of weasel heavily out-numbered stoat remains (weasel: MNI LGM ¼ 4; MNI OldestDryas ¼ 4 e stoat: MNI LGM ¼ 0; MNI Oldest Dryas ¼ 2) and malesare more numerous than females (remains from extracted sedi-ments included) (Fig. 7; Table 7). Most individuals of both speciesare quite young specimens, in the II (5e6 months old) and III(above 6 months) age classes (Schmidt, 1992). The slightly smallersize of individuals from Coulet des Roches still confirms unstableclimatic conditions and some colder periods during sedimentaccumulation.
ches, Planessard (Saint-Saturnin-d’Apt, Vaucluse; Oldest Dryas), Mont Ventoux 14sal length.
Coulet des Roches Planes sard
Mustela erminea
M4 548 M4 217 N4 164 J3 23 18m
35.8 31.1 46.3 44.2 43.118.8 14.9 26.4 26.0 24.8
Mont Ventoux
M3 217 MV 14
31.0 34.515.2 17.7
oches, Goult (Vaucluse; Holocene), Mont Ventoux 14 (Brantes, Vaucluse; Holocene).
Mont Ventoux 14
R 9295 R 9306 M5 4 MV 14 MV 14
3.81 3.91 3.88 3.82 3.833.26 2.81 2.71 2.88 2.850.80 1.08 1.38 0.99 1.001.69 1.58 1.48 1.42 1.40
-4 N5-N6 R1 M4 217 M4 217 M3-M4 1-2 M4 548 M4 548
4.32 4.10 4.45 4.22 4.23 4.273.10 2.70 2.87 3.00 3.01 3.251.07 1.30 0.71 0.92 1.09 1.101.85 1.40 1.48 1.48 1.67 1.41
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/
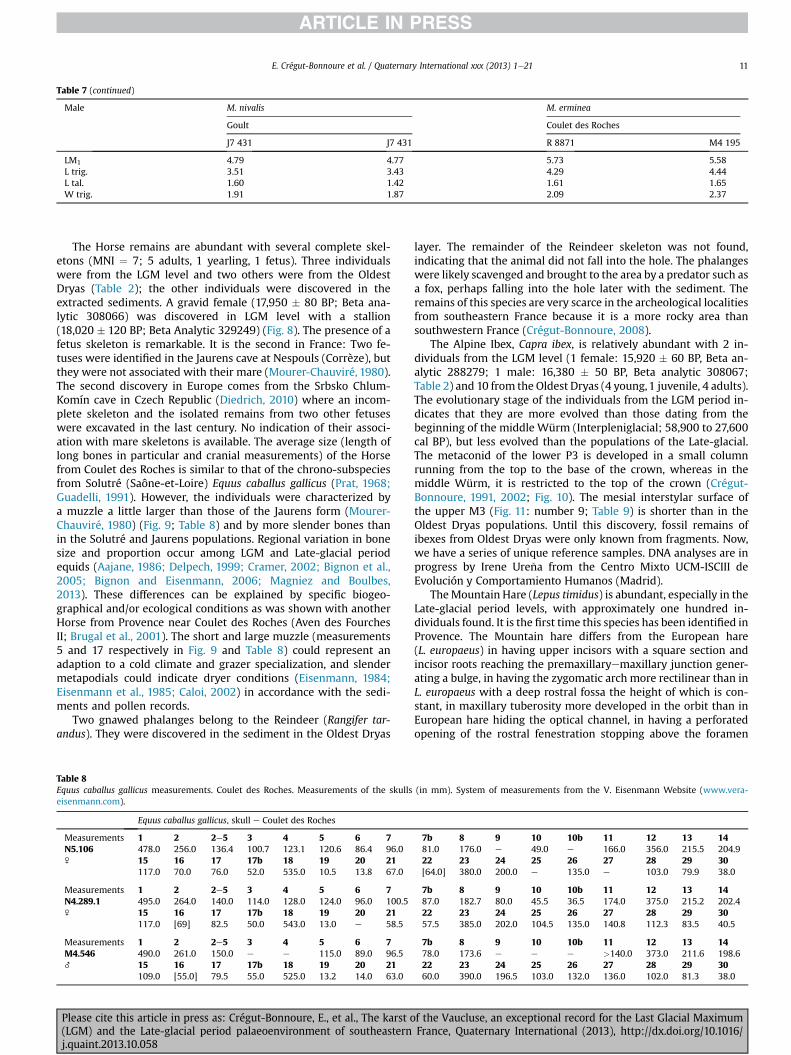
Table 7 (continued)
Male M. nivalis M. erminea
Goult Coulet des Roches
J7 431 J7 431 R 8871 M4 195
LM1 4.79 4.77 5.73 5.58L trig. 3.51 3.43 4.29 4.44L tal. 1.60 1.42 1.61 1.65W trig. 1.91 1.87 2.09 2.37
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 11
The Horse remains are abundant with several complete skel-etons (MNI ¼ 7; 5 adults, 1 yearling, 1 fetus). Three individualswere from the LGM level and two others were from the OldestDryas (Table 2); the other individuals were discovered in theextracted sediments. A gravid female (17,950 � 80 BP; Beta ana-lytic 308066) was discovered in LGM level with a stallion(18,020 � 120 BP; Beta Analytic 329249) (Fig. 8). The presence of afetus skeleton is remarkable. It is the second in France: Two fe-tuses were identified in the Jaurens cave at Nespouls (Corrèze), butthey were not associated with their mare (Mourer-Chauviré, 1980).The second discovery in Europe comes from the Srbsko Chlum-Komín cave in Czech Republic (Diedrich, 2010) where an incom-plete skeleton and the isolated remains from two other fetuseswere excavated in the last century. No indication of their associ-ation with mare skeletons is available. The average size (length oflong bones in particular and cranial measurements) of the Horsefrom Coulet des Roches is similar to that of the chrono-subspeciesfrom Solutré (Saône-et-Loire) Equus caballus gallicus (Prat, 1968;Guadelli, 1991). However, the individuals were characterized bya muzzle a little larger than those of the Jaurens form (Mourer-Chauviré, 1980) (Fig. 9; Table 8) and by more slender bones thanin the Solutré and Jaurens populations. Regional variation in bonesize and proportion occur among LGM and Late-glacial periodequids (Aajane, 1986; Delpech, 1999; Cramer, 2002; Bignon et al.,2005; Bignon and Eisenmann, 2006; Magniez and Boulbes,2013). These differences can be explained by specific biogeo-graphical and/or ecological conditions as was shown with anotherHorse from Provence near Coulet des Roches (Aven des FourchesII; Brugal et al., 2001). The short and large muzzle (measurements5 and 17 respectively in Fig. 9 and Table 8) could represent anadaption to a cold climate and grazer specialization, and slendermetapodials could indicate dryer conditions (Eisenmann, 1984;Eisenmann et al., 1985; Caloi, 2002) in accordance with the sedi-ments and pollen records.
Two gnawed phalanges belong to the Reindeer (Rangifer tar-andus). They were discovered in the sediment in the Oldest Dryas
Table 8Equus caballus gallicus measurements. Coulet des Roches. Measurements of the skullseisenmann.com).
Equus caballus gallicus, skull e Coulet des Roches
Measurements 1 2 2e5 3 4 5 6 7N5.106 478.0 256.0 136.4 100.7 123.1 120.6 86.4 96.0\ 15 16 17 17b 18 19 20 21
117.0 70.0 76.0 52.0 535.0 10.5 13.8 67.0
Measurements 1 2 2e5 3 4 5 6 7N4.289.1 495.0 264.0 140.0 114.0 128.0 124.0 96.0 100.5\ 15 16 17 17b 18 19 20 21
117.0 [69] 82.5 50.0 543.0 13.0 e 58.5
Measurements 1 2 2e5 3 4 5 6 7M4.546 490.0 261.0 150.0 e e 115.0 89.0 96.5_ 15 16 17 17b 18 19 20 21
109.0 [55.0] 79.5 55.0 525.0 13.2 14.0 63.0
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
layer. The remainder of the Reindeer skeleton was not found,indicating that the animal did not fall into the hole. The phalangeswere likely scavenged and brought to the area by a predator such asa fox, perhaps falling into the hole later with the sediment. Theremains of this species are very scarce in the archeological localitiesfrom southeastern France because it is a more rocky area thansouthwestern France (Crégut-Bonnoure, 2008).
The Alpine Ibex, Capra ibex, is relatively abundant with 2 in-dividuals from the LGM level (1 female: 15,920 � 60 BP, Beta an-alytic 288279; 1 male: 16,380 � 50 BP, Beta analytic 308067;Table 2) and 10 from the Oldest Dryas (4 young,1 juvenile, 4 adults).The evolutionary stage of the individuals from the LGM period in-dicates that they are more evolved than those dating from thebeginning of the middle Würm (Interpleniglacial; 58,900 to 27,600cal BP), but less evolved than the populations of the Late-glacial.The metaconid of the lower P3 is developed in a small columnrunning from the top to the base of the crown, whereas in themiddle Würm, it is restricted to the top of the crown (Crégut-Bonnoure, 1991, 2002; Fig. 10). The mesial interstylar surface ofthe upper M3 (Fig. 11: number 9; Table 9) is shorter than in theOldest Dryas populations. Until this discovery, fossil remains ofibexes from Oldest Dryas were only known from fragments. Now,we have a series of unique reference samples. DNA analyses are inprogress by Irene Ure�na from the Centro Mixto UCM-ISCIII deEvolución y Comportamiento Humanos (Madrid).
TheMountain Hare (Lepus timidus) is abundant, especially in theLate-glacial period levels, with approximately one hundred in-dividuals found. It is the first time this species has been identified inProvence. The Mountain hare differs from the European hare(L. europaeus) in having upper incisors with a square section andincisor roots reaching the premaxillaryemaxillary junction gener-ating a bulge, in having the zygomatic arch more rectilinear than inL. europaeus with a deep rostral fossa the height of which is con-stant, in maxillary tuberosity more developed in the orbit than inEuropean hare hiding the optical channel, in having a perforatedopening of the rostral fenestration stopping above the foramen
(in mm). System of measurements from the V. Eisenmann Website (www.vera-
7b 8 9 10 10b 11 12 13 1481.0 176.0 e 49.0 e 166.0 356.0 215.5 204.922 23 24 25 26 27 28 29 30[64.0] 380.0 200.0 e 135.0 e 103.0 79.9 38.0
7b 8 9 10 10b 11 12 13 1487.0 182.7 80.0 45.5 36.5 174.0 375.0 215.2 202.422 23 24 25 26 27 28 29 3057.5 385.0 202.0 104.5 135.0 140.8 112.3 83.5 40.5
7b 8 9 10 10b 11 12 13 1478.0 173.6 e e e >140.0 373.0 211.6 198.622 23 24 25 26 27 28 29 3060.0 390.0 196.5 103.0 132.0 136.0 102.0 81.3 38.0
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/
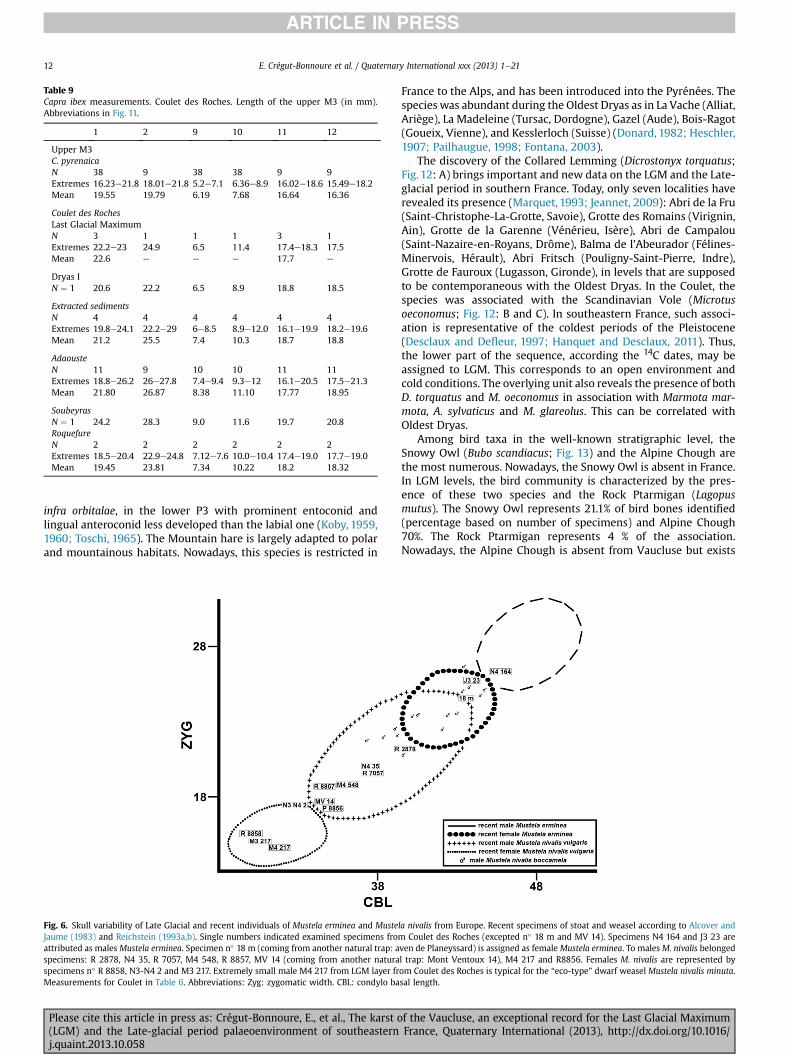
Table 9Capra ibex measurements. Coulet des Roches. Length of the upper M3 (in mm).Abbreviations in Fig. 11.
1 2 9 10 11 12
Upper M3C. pyrenaicaN 38 9 38 38 9 9Extremes 16.23e21.8 18.01e21.8 5.2e7.1 6.36e8.9 16.02e18.6 15.49e18.2Mean 19.55 19.79 6.19 7.68 16.64 16.36
Coulet des RochesLast Glacial MaximumN 3 1 1 1 3 1Extremes 22.2e23 24.9 6.5 11.4 17.4e18.3 17.5Mean 22.6 e e e 17.7 e
Dryas IN ¼ 1 20.6 22.2 6.5 8.9 18.8 18.5
Extracted sedimentsN 4 4 4 4 4 4Extremes 19.8e24.1 22.2e29 6e8.5 8.9e12.0 16.1e19.9 18.2e19.6Mean 21.2 25.5 7.4 10.3 18.7 18.8
AdaousteN 11 9 10 10 11 11Extremes 18.8e26.2 26e27.8 7.4e9.4 9.3e12 16.1e20.5 17.5e21.3Mean 21.80 26.87 8.38 11.10 17.77 18.95
SoubeyrasN ¼ 1 24.2 28.3 9.0 11.6 19.7 20.8RoquefureN 2 2 2 2 2 2Extremes 18.5e20.4 22.9e24.8 7.12e7.6 10.0e10.4 17.4e19.0 17.7e19.0Mean 19.45 23.81 7.34 10.22 18.2 18.32
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e2112
infra orbitalae, in the lower P3 with prominent entoconid andlingual anteroconid less developed than the labial one (Koby, 1959,1960; Toschi, 1965). The Mountain hare is largely adapted to polarand mountainous habitats. Nowadays, this species is restricted in
Fig. 6. Skull variability of Late Glacial and recent individuals of Mustela erminea and MusteJaume (1983) and Reichstein (1993a,b). Single numbers indicated examined specimens froattributed as males Mustela erminea. Specimen n� 18 m (coming from another natural trap: aspecimens: R 2878, N4 35, R 7057, M4 548, R 8857, MV 14 (coming from another naturaspecimens n� R 8858, N3-N4 2 and M3 217. Extremely small male M4 217 from LGM layer frMeasurements for Coulet in Table 6. Abbreviations: Zyg: zygomatic width. CBL: condylo ba
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
France to the Alps, and has been introduced into the Pyrénées. Thespecies was abundant during the Oldest Dryas as in La Vache (Alliat,Ariège), La Madeleine (Tursac, Dordogne), Gazel (Aude), Bois-Ragot(Goueix, Vienne), and Kesslerloch (Suisse) (Donard, 1982; Heschler,1907; Pailhaugue, 1998; Fontana, 2003).
The discovery of the Collared Lemming (Dicrostonyx torquatus;Fig. 12: A) brings important and new data on the LGM and the Late-glacial period in southern France. Today, only seven localities haverevealed its presence (Marquet, 1993; Jeannet, 2009): Abri de la Fru(Saint-Christophe-La-Grotte, Savoie), Grotte des Romains (Virignin,Ain), Grotte de la Garenne (Vénérieu, Isère), Abri de Campalou(Saint-Nazaire-en-Royans, Drôme), Balma de l’Abeurador (Félines-Minervois, Hérault), Abri Fritsch (Pouligny-Saint-Pierre, Indre),Grotte de Fauroux (Lugasson, Gironde), in levels that are supposedto be contemporaneous with the Oldest Dryas. In the Coulet, thespecies was associated with the Scandinavian Vole (Microtusoeconomus; Fig. 12: B and C). In southeastern France, such associ-ation is representative of the coldest periods of the Pleistocene(Desclaux and Defleur, 1997; Hanquet and Desclaux, 2011). Thus,the lower part of the sequence, according the 14C dates, may beassigned to LGM. This corresponds to an open environment andcold conditions. The overlying unit also reveals the presence of bothD. torquatus and M. oeconomus in association with Marmota mar-mota, A. sylvaticus and M. glareolus. This can be correlated withOldest Dryas.
Among bird taxa in the well-known stratigraphic level, theSnowy Owl (Bubo scandiacus; Fig. 13) and the Alpine Chough arethe most numerous. Nowadays, the Snowy Owl is absent in France.In LGM levels, the bird community is characterized by the pres-ence of these two species and the Rock Ptarmigan (Lagopusmutus). The Snowy Owl represents 21.1% of bird bones identified(percentage based on number of specimens) and Alpine Chough70%. The Rock Ptarmigan represents 4 % of the association.Nowadays, the Alpine Chough is absent from Vaucluse but exists
la nivalis from Europe. Recent specimens of stoat and weasel according to Alcover andm Coulet des Roches (excepted n� 18 m and MV 14). Specimens N4 164 and J3 23 areven de Planeyssard) is assigned as female Mustela erminea. To males M. nivalis belongedl trap: Mont Ventoux 14), M4 217 and R8856. Females M. nivalis are represented byom Coulet des Roches is typical for the “eco-type” dwarf weasel Mustela nivalis minuta.sal length.
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Fig. 7. Sexual variability based on the size of lower M1 of the stoat (Mustela erminea) and the weasel (Mustela nivalis). Examined material from Coulet des Roches and Les Planes hasbeen compared with recent central European stoat and weasel and Late Pleistocene specimens of both species from French locality La Baume de Gigny (Delpech, 1989). Singlenumbers on the diagram indicated specimens from Coulet des Roches and Les Planes. Specimens R 8871 and M4 195 are attributed as males M. erminea. Specimens R 3058, R 3037and M3 M4 1 are assigned as females M. erminea. To males M. nivalis belonged specimens: J7 431, R 3059, N5 N6 R1, M4 548, R 9294, R 3719, R 3035, R 3060, M4 217m, M3 M4 1-2, R3034, M3 61-4 and Les Planes. Females M. nivalis are represented by specimens n� R 9295, R 9306, R 3038, M5 41-4, MV 14 and R 3039. Measurements for Coulet in Table 7.Abbreviations: L: length; W: width.
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 13
to the east, in the territories of Drôme, Alpes-de-Haute-Provenceand Hautes-Alpes. This association is characteristic of a very coldclimate, in accordance with the LGM period. In the Oldest Dryaslevel, the Rock Ptarmigan is absent. The Snowy Owl exists in manyLate-glacial period sites of southwestern France (Tyrberg, 1998) asthe Grotte d’Isturiz (Pyrénées-Atlantiques), Abri de Duruthy(Sorde-l’Abbaye, Landes), Espèche (Hautes-Pyrénées), Les Espé-lugues (Lourdes, Hautes-Pyrénées), Gourdan (Haute-Garonne),Grotte des Trois-Frères (Montesquieu-Avantès, Ariège), Massat(Ariège), Le Mas d’Azil (Ariège), La Vache (Alliat, Ariège) (Clot andMourer-Chauviré, 1986), Bois Ragot (Gouaix, Vienne), Bourouilla(Arancou, Pyrénées-Atlantiques), Gabillou (Dordogne), Jaurias(Dordogne), Abri Dufaure (Sorde-l’Abbaye, Landes), La Madeleine(Tursac, Dordogne), La Gare de Couze (Dordogne), Le Morin(Pessac-sur-Gironde, Gironde) and Fostin (Cessac, Gironde)(Laroulandie, 2003). This is the first occurrence for the RockPtarmigan in Vaucluse, and its abundance is striking. This in-dicates an environment under a very cold climate. The Alpine
Fig. 8. Equus caballus gallicus. Gravid mare (above) and stallion in situ. Photography: E.Crégut-Bonnoure. Scale: 30 cm.
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
Chough represents 55.3% of bird bones and the Snowy Owl 28%.This species association is characteristic of a very cold climate, inaccordance with the Oldest Dryas period.
Palynological data from the Oldest Dryas revealed a sparselywooded landscape (Scots Pine, Juniper and Ephedra) with a pre-dominance of herbaceous taxa including taxa commonly encoun-tered in steppe, sometimes reaching a relatively high rate(Caryophyllaceae, Cichorioideae for example) (Fig. 4, sample O3).These characteristics suggest a very open landscape as well as aharsh and rather dry climate.
3.1.5. CommentsA LGM and a quite complete Late-glacial period sequence were
identified for the first time in southeastern France. The Reindeer,the Collared Lemming, the Scandinavian Vole, the Marmot, theMountain Hare, the Grouse and the Snowy Owl, together withnumerous horses, are indicators of a very cold and harsh climate.Before this discovery, such associations were unknown.
The Collared Lemming has been highlighted in at least sevensites dating from the Late Glacial period in southern France,generally dating to the Oldest Dryas. At the Coulet des Roches, twodistinct southern expansions of D. torquatus can be correlatedfirstly to the LGM and secondly to the Oldest Dryas (Fig. 14). Theultimate expansion of the Collared Lemming during the OldestDryas can be explained by the fact that the Alps were a refuge forthis cold-adapted species during the limited and cooler period oftime between the LGM and the Oldest Dryas.
In the Holocene, Wolves, Wildcats (Felis silvestris), EurasianLynxes, Weasels, Red Deers, Roe Deers (Capreolus capreolus) andBoars settled in the forest. Reptiles and amphibians were abundant.They are characteristic of the Mediterranean climate with moun-tain influences.
3.2. Aven des Planes
Aven des Planes is located 3 km from Coulet des Roches. It wasopened by spelunkers from Avignon (S.S.A.) in 1970. The ellipsoidalopening is about 2.70 � 1 m. It is 11 m deep (Fig. 15). Nine visuallevels, which provided a well preserved Pleistocene large mammal
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Fig. 9. Bivariate plot of muzzle breadth and length in some late Pleistocene and Holocene horses. System of measurement, data of Siréjol, Jaurens and Przewalski’s horse from the V.Eisenmann Website (www.vera-eisenmann.com). Other sites for comparison: Fontainhas (Cardoso and Eisenmann, 1989), Mounoï (Boulbes, personal data), Quéroy I (Eisenmann,personal communication). Measurements for Coulet in Table 8.
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e2114
fauna, including complete skeletons, have been recognized byspelunkers (Aujard-Catot, 1973; Aujard-Catot et al., 1979). A surveywas completed in 2005 and 2006, and the extracted sedimentswere screened by our team using 2.25 mm, 1.4 mm and 1.10 mmscreen mesh (Crégut-Bonnoure, 2005, 2006). Additional excavationhas taken place at the bottom with new mapping.
Three main stratigraphical units were identified. At the top, theA Unit is sterile (levels C1 and C2). The B Unit dates from the Ho-locene (levels C3 to C7). At the bottom, the C Unit corresponds tothe Late-glacial period (levels D and E).
The infilling has been partially extracted by spelunkers. Finesediments have been left on the walls. As in Coulet des Roches, apart of the infilling is formed by the accumulation of fragments oflimestone of small size grading.
Fig. 10. Capra ibex. Lower teeth (rigth P3, P4). Note the development of the P3 met-aconid (arrow). Photography O. Ambrosio.
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
3.2.1. Extracted sedimentSeveral hundred bones have been collected and 32 species were
identified: 10 large mammals, 2 insectivores, 2 bats, 3 lagomorphs,5 rodents, 4 birds, 5 reptiles, 1 amphibian (Tables 3 and 4). Thefaunal assemblage is heterogeneous with domestic and wildspecies.
3.2.2. B UnitWild and domestic faunawere present (24 species), with human
remains: 8 large mammals, 2 insectivores, 2 lagomorphs, 5 rodents,5 birds, 1 reptile, 1 amphibian (Tables 3 and 4). The top of the holecorresponds to a human burial (C3 level). Five human individualshave been identified. Based on the archaeological material (ceramicremains, bronze ornament), the accumulation dates from the finalBronze Age (BF 3b; 1,400 to 800 BC) and the beginning of the Iron
Fig. 11. Capra ibex. Ratio-diagram of the upper M3. 0: C. pyrenaica. Abbreviations:Length: 1: occlusal. 2: basal. 9: anterior interstylar surface. 10: posterior interstylarsurface. 11: occlusal, first þ second lobes (lingual side). 12: basal, first þ second lobes(lingual side). References of the measurements in Crégut-Bonnoure (1991, 2002).Measurements in Table 9. See the evolution of the length of the interstylar surfacesfrom the Pleniglacial to the Late-glacial period, the most recent population beingAdaouste and Soubeyras.
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Fig. 12. Rodents from Coulet des roches. A. Dicrostonyx torquatus: lower M1 right (N3R47 z ¼ 322.5); B. Microtus oeconomus: right lower M1 (M4 R324 z ¼ 174.5); C.Microtus oeconomus: left lower M1 (M4 R371 z ¼ 360e366); D. Microtus cf. arvalis: leftlower M1 (M3 R143 z ¼ 3.14); E. Chionomys nivalis: right lower M1 (M3 R153 z ¼ 150);F. Chionomys nivalis: right lower M1 (M4 R324 z ¼ 174.5). Scale in mm.
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 15
Age (Hallstatt C; 800 to 450 BC). The Red Deer (a complete skeleton)discovered by spelunkers in the C7 level was dated by radiocarbon:3255 � 35 BP (IntCal 04; Lyon 3141(Poz); Table 2).
Micromammals were abundant (Number of Remains¼ 608) andwere mainly found in the extracted sediments (NR ¼ 465) andduring the speleological excavation (NR ¼ 94). According to theobservations from the spelunkers, the remains found in theextracted sediment came from the C4 level. The micromammalcommunity, characterized by an abundance of European Snow Vole(Chionomys nivalis), Wood Mouse (A. sylvaticus) and Common Vole(Microtus arvalis), is representative of foot-hill mountains of theperi-mediterranean area. Among bird taxa, the Eurasian Jackdaw(Corvus monedula) was relatively abundant. The presence ofWestern Capercaillie indicates a forest environment.
3.2.3. C UnitFifteen species were identified: 4 largemammals, 2 lagomorphs,
4 rodents, 5 birds (Tables 3 and 4). The Red Fox had elongated limbbones like the Coulet des Roches individuals (Fig. 5). The Reindeerwas represented by the skeleton of a 3e4 year old female (initiallyreferred to as a 2 year oldmale by Aujard-Catot et al., 1979) (Fig. 16).The discovery of a complete skeleton is unique in southeasternFrance. According to the dating (12,660 � 75 BP ; IntCal 04; Lyon3138(Poz); Table 2) its presence in planes could coincide with theOldest Dryas. This species disappeared from southwestern andsoutheastern France around 12,000 BP (14,000 cal BP) (Costamagnoet al., 2008; Bridault and Chaix, 2009). The horses are relatively
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
abundant (MNI ¼ 8) but the skulls were broken. The radiocarbondatewas 13,360� 80 BP (IntCal 04; Lyon 3139(Poz); Table 2) for oneindividual from the bottom (C level), and another individual fromthe D level was dated to 13,730 � 60 BP (IntCal 09; Beta analytic329247; Table 2). The average length of long bones seems to besmaller than those from Coulet des Roches and closer to that of theMagdalenian level horse from Solutré E. caballus arcelini (Guadelli,1991), but several individuals are not mature, and the 14C datingindicate that they are contemporaneous to the Coulet horses.
Sediment samples for palynological analysis were taken fromtwo different sections: at the bottom, Pinus is relatively abundanttogether with Quercus. The herbaceous plants belong to Cichor-ioideae. Charcoal has been collected in these sections. The Pinusbelongs to the P. silvestris group and Quercus to the deciduousgroup. However, pollen and charcoal are scarce and the interpre-tation of the results is difficult.
3.2.4. CommentsAs the infilling was almost totally destroyed, there is a lack of
data about the modalities of the infilling. The main interest of thedeposit lies in the presence of a complete Reindeer skeleton. Thefaunal association of the Oldest Dryas (levels D and E) shows sim-ilarities with Coulet des Roches.
3.3. Aven Souche n�1
Aven Souche n�1 was first opened by spelunkers who rapidlystopped their investigations (Perret, 1975). We do not know whenthis opening occurred and who the spelunkers were. The infilling isquite intact. Today the hole is 6 m deep (Fig. 17). Its upper part,slightly funneled, is about 5 m long and 4 mwide. The excavationsbegan in 2009 (Crégut-Bonnoure et al., 2009b). Three distinct unitswere identified in the homogeneous sediment from the top to thebottom: human burials dating back to the Iron Age, a unit withdomestic and wild species, and a Red Deer accumulation (Crégut-Bonnoure et al., 2010, 2011b). We observed a downward distribu-tion of the remains. The origin of such a distribution can be con-nected to two concomitant factors: karst raking and open scree.
The excavation revealed a significant fissuring of the walls andsmall drains: the sediment was clearly channeled by these open-ings, suggesting that it came from the soil which covered the lapies.The walls showed an intense karstification which could reflect aflooded context with a former dome of truncated dissolution andgrooves. The dating of a stalagmitic floor indicated significant flowsof water during the MIS 15 (Middle Pleistocene; Axiom n� 73-12;Table 1).
3.3.1. Iron Age UnitThe human burials were totally destroyed by spelunkers. Some
bones slid between the spaces of the rocky blocks and werecollected. Three individuals were present (NR ¼ 162): two adultsand a child. The radiocarbon date is 2240 � 40 BP (IntCal04; Betaanalytic 288280; Table 2).
3.3.2. Unit with domestic faunaThis unit is an accumulation of fragmentary bones and rocks
inside black sediment. Twenty five species are identified (Tables 3and 4): 11 large mammals (wilds and domestic), 2 insectivores, 2lagomorphs, 1 rodent, and 9 birds. The importance of the domesticspecies decreases with the depth. The association of Wildcat, RoeDeer, Red Deer, Capercaillie and Eurasian Jay (Garrulus glandarius)reveals a wooded environment while the Great Bustard (Otis tarda)indicates an open landscape. We thus envisage a mosaic landscape.
The weathering marks (Behrensmeyer, 1978) and the action ofcarnivores (with potential den of a small predator) and rodents
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Fig. 13. Bubo scandiacus from Coulet des roches. 1. Right tarsometatarsal, female sized (N4 27-1). 2. Left tarsometatarsal, female sized (K3 78). 3. Right tarsometatarsal, male sized (R4322). 4. Left carpometacarpal, female sized (K4 15). 5. Left carpometacarpal, female sized (M4 40). 6. Right femur (K3 50).7. Left femur (R 4321). Photography Th. Roger. Scale 1 cm.
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e2116
suggest prolonged exposure of the osseous to the outdoors (Fig. 17).Phenomena connected to water (rain, flow) are likely to havestrongly altered the remains and to have partially upset the orga-nization of the assembly. Some bones are burned. Artefacts havebeen discovered (n ¼ 32): three are characteristic of the Neolithic(6000 to 2100 BC). Burned bones and artefacts were pulled into thehole with the sediment. Outside of the hole, some limited areasprovided abundant flint fragments and flakes (including nuclei)
Fig. 14. Dicrostonyx torquatus phases of expa
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
which may be related to a Neolithic flint factory. The artefacts andthe bones of domestic species are in secondary position: the top ofthe hole can be identified as a disposal area.
Pollen grains were not preserved. However charcoal wasnumerous: Quercus sp. (probably Q. pubescens), J. communis, Prunusspinosa, Acer sp., A. opalus, A. campestre, Ulmus minor, Sorbusdomestica, S. aria, P. type sylvestris. The analysis allowed us to definethis as a deciduous forest. Oak and Juniper are dominant. Scots Pine
nsion in the Coulet des Roches infilling.
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Fig. 15. Aven des Planes. Cross section. A: topography. B: archeological levels. topographical survey: Aujard-Catot (1973), redrawn by J.-B. Fourvel.
Fig. 16. Rangifer tarandus. Aven des Planes. MR 4.001.759. A. Skull in norma frontalis. B. Skull in norma lateralis. C. Limb bones; 1. Humerus. 2. Radius. 3. Metacarpal III þ IV. 4. Femur.5. Tibia. 6. Metatarsal III þ IV. Photographies: D. Kuntz.
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 17
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst of the Vaucluse, an exceptional record for the Last Glacial Maximum(LGM) and the Late-glacial period palaeoenvironment of southeastern France, Quaternary International (2013), http://dx.doi.org/10.1016/j.quaint.2013.10.058

Fig. 17. Aven Souche n�1. Cross section (Topographical survey: J.-B. Fourvel and N. Frèrebeau). Global trend of both faunal list and taphonomic observations related to the depth.
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e2118
is rare. The association indicates climatic conditions close to thoseof today, but with a more open landscape.
3.3.3. Red Deer bone assemblage unitThree Red Deer disarticulated skeletons were present at the
bottom (Fig. 17). One, at the top of the accumulation, dates from9670� 40 BP (IntCal09; Beta Analytic 308070; Table 2). At the samedepth, charcoal (Juniper) dates from 8070 � 40 BP (IntCal09; BetaAnalytic 308071; Table 2).
3.3.4. CommentsAt least eight different phases followed one another until the
use of the site as a sepulchral place. The bottom corresponds to anatural trap in which the Red Deer fell. The marks observed onbones indicate burying of the animals over time. The carcasses weregradually dislocated; they split up and dispersed across one meterin thickness. These results illustrate the process of drawing asso-ciated with open masses of fallen rocks, favoring the movement ofbones downwards.
3.4. Aven Souche n� 2
The aven is located 100 m from Aven Souche n�1 and wasopened by spelunkers from S.S.A. in 1972. The spelunkers stoppedthe excavation at the depth of e 2.70 m (Fig. 18). We have no in-formation about the spelunkers’ investigations. A survey wascompleted in 2007 (Crégut-Bonnoure, 2007b). The hole appearsunder a depression, sloping gently towards the Southwest, andbounded to the east by a fault. The infilling was formed by theaccumulation of rocky blocks.
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
3.4.1. The infillingThere was a human burial at the top of the infilling that was
excavated in the 1970s. Our survey collected disarticulated humanbones without any anatomical connection and with a downwarddistribution. Two individuals were identified by Bérangère Perez.Artefacts from the Neolithic (n ¼ 25), identified by StéphaneRenault, were discovered in association with ceramic shards(n ¼ 157) from the Early Bronze Age (2200 to 1600 BC). Domesticand wild species were represented by 16 species (Tables 3 and 4): 8large mammals, 1 insectivore, 2 lagomorphs, 1 rodent, 3 reptiles,and 1 amphibian. The anthropological origin of this accumulationwas not attested. The dimensions of punctures and scoring onRabbit (Oryctolagus cuniculus) and Sheep (Ovis aries) bones indicatethat they result partially from the action of red foxes.
Charcoal is relatively abundant. Only three taxa were identified:Quercus sp. (probably Q. pubescens), S. domestica, A. campestre.These species indicate environmental conditions for the most partidentical to those of today. Oak is dominant and indicates thepresence of a deciduous forest. The presence of Sorb (S. domestica)indicates that the forest was not totally continuous.
3.4.2. CommentsNo intact sedimentary levels were preserved. Remains were
scattered, and dispersal through the rocky scree is significant. Ce-ramics suggest use of the top of the aven by humans, a phenome-non common in the area (Buisson-Catil et al., 1996e1997).
4. Conclusion
These natural traps have provided extensive information on theend of the Glacial period and the beginning of the Holocene,permitting interpretation of the environmental transformations.Among the four localities, the Coulet and the Planes can be
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

Fig. 18. Aven Souche n�2. Plan and cross section. Topographical survey: Ch. Boucher,redrawn by J.-B. Fourvel.
E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 19
considered as references for the knowledge of climate change andthe environment in southeastern France. The chronological datawere well-constrained because of the numerous radiocarbon datesobtained. The LGM and the Oldest Dryas were for the first timeclearly identified in this geographical area.
The data allow us to state that during the LGM the Horse, Ibex,Mountain Hare, Polecat, Weasel, Snowy Owl, Collared Lemmingand Scandinavian Vole were present. This assemblage is unique forsoutheastern France. These species persisted during the OldestDryas. This period is characterized by the appearance of Marmot,Ermine and Reindeer. The Allerød corresponds to the arrival of atemperate fauna.
The excavated material is remarkable particularly regarding itsdiversity and the presence of complete skeletons of large mammaland bird species. Special adaptations to the cold and harsh climateof the LGM were identified for Horse (width of the muzzle andslenderness of the metapodials), Stoat and Weasel (small size).Another adaptation for Red Fox (elongated tibia) seems to occurduring the Oldest Dryas. For the first time, two distinct southernexpansions of D. torquatus, were identified, correlated firstly to theLGM and secondly to the Oldest Dryas. The infillings of these holesare exceptional.
Acknowledgments
We are grateful to International Cave Bear Symposium orga-nizers, Dr. Silviu Constantin and Dr. Marius Robu, who invited us to
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
present this study. We are also grateful to Ph.D. Christopher N. Jassand the anonymous reviewer for their constructive and usefulcomments.
We want to thank all the institutions and their representativeswhich support our research: Xavier Delestre from D.R.A.C-P.A.C.A.,Marie-Josée Roig Ex Minister, Major of Avignon city, the executorsfrom the Calvet Fondation, Alain Gabert, Major of Monieux village,André Faraud, Major of Sault city, Dominique Carru from the Servicedépartemental d’Archéologie de Vaucluse, André Souquières fromthe Association des Amis de Saint-Jean de Sault.
We express our gratitude to Alain Gabert and the Malavard andLhote Families for their authorizations.
The excavations were made in close collaboration with theDepartmental Committee of Speleology from Vaucluse (CDS 84)and the Speleological clubs: Carpentras, Avignon (Société Spéléo-logique d’Avignon and Association Vauclusienne des ExplorationsNouvelles de Jean Nouveau), Bollène, Camaret, Cavaillon, Cour-thezon, Fontaine-de-Vaucluse, Oraison (Departmental Committeeof Speleology from Alpes-de-Haute-Provence).
Special thanks to Christian Bérard, Robert Russ, Maurice Ricci,Michel Baillet and Christian Boucher who insure an effective andcontinuous monitoring of the field operations. We want to thankMargret Baillet who insures the estate management and AlainPlouzen for his logistic support.
We also thank Véra Eisenmann from M.N.H.N. (Paris) and Jean-François Tournepiche from the Angoulême Museum for the com-parison data of the horse from Le Quéroy, Stéphane Renault, fromthe M.M.S.H., who was kind enough to assure the typologicalevaluation of the artefacts from Souche n� 2, as well as PhD stu-dents who stopped their studies: Bérangère Perez who analyzedthe human remains from this site and Alexandre Bournery who hasstudied the birds from Les Planes. And of course, many thanks tothe excavators: without them nothing would be possible.
References
Aajane, A., 1986. Contribution à l’étude des faunes du Würm récent en Haute-Loire.Les niveaux magdaléniens du Rond du Barry. Thèse de 3� cycle, Université deBordeaux I.
Alcover, J.A., Jaume, D., 1983. Sobre el mostel Mustela nivalis Linnaeus 1758 de lesBalears (Carnivora, Mustelidae). Bolletino della Societa Historia Naturale desBalears 27, 145e164.
Altuna, J., 1971. Fauna de Mamiferos de los yacimientos prehistóricos de Guipúzcoa.Munibe 24, 1e464.
Aujard-Catot, R., 1973. Chronique archéologique. II: l’aven des Planes. Bulletin de laSociété spéléologique d’Avignon 10, 27e36.
Aujard-Catot, R., Crégut, E., Gagnière, S., Gruneisen, A., Mourer-Chauviré, C., 1979.Un squelette complet de renne, Rangifer tarandus (Mammalia, Artiodactyla),dans le Pléistocène supérieur du remplissage karstique de l’aven des Planes(Monieux, Vaucluse, France). Nouvelles Archives du Muséum d’histoirenaturelle de Lyon 17, 15e20.
Bailon, S., 1991. Amphibiens et reptiles du Pliocène et du Quaternaire de France etd’Espagne: mise en place et évolution des faunes. Ph.D. Dissertation. Universitéde Paris VII.
Bailon, S., 1999. Différenciation ostéologique des Anoures (Amphibia, Anura) deFrance. In: Desse, J., Desse-Berset, N. (Eds.), Fiches d’Ostéologie Animale pourl’Archéologie, Série C: Varia. Centre de Recherches Archéologiques, CNRS,Valbonne.
Barahona, F., Barbadillo, L.J., 1997. Identification of some Iberian lacertids using skullcharacters. Revista Española de Herpetologia 11, 47e62.
Baryshnikov, G.F., 2009. Pleistocene Mustelidae (Carnivora) from Paleolithic site inKudaro Caves in the Caucasus. Russian Journal of Theriology 8, 75e95.
Baryshnikov, G.F., Golovanova, L.V., 1989. On the Mousterian site Matuzka in theKuban Caucasus. Proceedings of the Zoological Institute 198, 1e55.
Behrensmeyer, A.K., 1978. Taphonomic and ecologic information from boneweathering. Paleobiology 4, 150e162.
Bignon, O., Baylac, M., Vigne, J.D., Eisenmann, V., 2005. Geometric morphometricsand the population diversity of Late Glacial horses in Western Europe (Equuscaballus arcelini): phylogeographic and anthropological implications. Journal ofArchaeological Science 32, 375e391.
Bignon, O., Eisenmann, V., 2006. Western European Late Glacial horses diversity andits ecological implications. In: Mashkour, M. (Ed.), Equids in Time and Space, 9thICAZ Conference, Durham 2002. Oxbow Books, pp. 161e171.
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e2120
Blain, H.-A., 2009. Contribution de la paléoherpétofaune (Amphibia and Squamata)à la connaissance de l’évolution du climat et du paysage du Pliocène supérieurau Pléistocène moyen d’Espagne. Treballs de Museu de Geologia de Barcelona16, 39e170.
Bridault, A., Chaix, L., 2009. Réflexions sur la recomposition des spectres fau-niques dans le massif jurassien et les Alpes françaises du nord durant leTardiglaciaire. In: Pion, G., Mevel, L. (Eds.), La fin du Paléolithique supérieurdans les Alpes du nord françaises et le Jura méridional. Approches cultur-elles et environnementales. Mémoire de la Société Préhistorique Française,L, pp. 59e72.
Brochier, J.E., 1977. Évolution des climats et des paysages vauclusiens au cours duWürmien récent et du Postglaciaire en Vaucluse. Paléoécologie de l’HommeFossile 1, 1e180.
Brochier, J.E., Livache, M., 2004. Les traces des derniers chasseurs-cueilleurs. In:Buisson-Catil, J., Guilcher, A., Hussy, Ch., Olive, M., Pagni, M. (Eds.), Vauclusepréhistorique. Le territoire, les hommes, les cultures et les sites, pp. 111e126.Barthélémy.
Brugal, J.-Ph., Buisson-Catil, J., Helmer, D., 2001. L’aven des Fourches II (Sault,Vaucluse): les derniers chevaux sauvages en Provence. Paleo 13, 73e88.
Buisson-Catil, J., Sauzade, G., Couturaud, A., Hasler, A., Puig, J.-M., Thiebault, S.,1996e1997. Le gisement Bronze final de l’aven des Fourches I à Sault. Contri-bution à l’étude fonctionnelle des cavités karstiques des plateaux de Vaucluse.Documents d’Archéologie Méridionale 19e20, 7e31.
Buchalczyk, T., 1984. Rzad: Drapie _zne e Carnivora. In: Pucek, Z. (Ed.), Klucz dooznaczania ssaków Polski. Pa�nstwowe Wydawnictwo Naukowe, pp. 256e310.
Caloi, L., 2002. The two Equids of the Middle Pleistocene of the site Venosa-Loreto(Southern Italy): functional morphology of the cranial remains. GeologicaRomana 36, 275e287.
Cardoso, J.L., Eisenmann, V., 1989. Equus caballus antunesi, nouvelle sous-espècequaternaire du Portugal. Palaeovertebrata 19 (2), 47e72.
Cheylan, M., Bons, J., Saint-Girons, H., 1981. Evidence of a vernal cycle and pre-nuptial spermatogenetic cycle in a Mediterranean snake Malpolon mon-spessulanus (Reptilia ; Colubridae). Comptes Rendus de l’Académie des Sciences292, 1207e1209.
Clot, A., 1980. La grotte de la Carrière (Gerde, Hautes-Pyrénées). Stratigraphie etpaléontologie des Carnivores. Travaux du Laboratoire de Geologie de l’Uni-versité Paul Sabatier Toulouse 3, 1e502.
Clot, A., Brochet, G., Chaline, J., Desse, G., Evin, J., Granier, J., Mein, P., Mourer-Chauviré, C., Omnes, J., Rage, J.C., 1984. Faune de la grotte préhistorique du boisdu Cantet (Espèche, Hautes-Pyrénées, France). Munibe 36, 33e50.
Clot, A., Mourer-Chauviré, C., 1986. Inventaire systématique des oiseaux quatern-aires des Pyrénées française. Munibe 38, 171e184.
Costamagno, S., Cochard, D., Ferrie, J.-G., Laroulandie, V., Cazals, N., Langlais, M.,Valdeyron, N., Dachary, M., Barbaza, M., Galop, D., Martin, H., Philibert, S., 2008.Nouveaux milieux, nouveaux gibiers, nouveaux chasseurs? Evolution des pra-tiques cynégétiques dans les Pyrénées du Tardiglaciaire au début du Post-glaciaire. Bulletin de la Société Préhistorique Française 105, 17e27.
Cramer, B., 2002. Morphometrische Untersuchungenan Quartären Pferden in Mit-teleuropa. Unpublished PhD thesis. University of Tübingen.
Crégut-Bonnoure, E., 1991. Intérêt biostratigraphique de la morphologie dentaire deCapra (Mammalia, Bovidae). Annales Zoologici Fennici 28, 273e290.
Crégut-Bonnoure, E., 2002. Les Ovibovini, Caprini et Ovini (Mammalia, Artiodactyla,Bovidae, Caprinae) du Plio-Pléistocène d’Europe: systématique, évolution etbiochronologie. Thèse de Doctorat d’Etat ès Sciences. Université de Lyon I.
Crégut-Bonnoure, E., 2005. Monieux: Aven des Planes. Bilan Scientifique de la Ré-gion PACA 2006, 209e210.
Crégut-Bonnoure, E., 2006. Monieux: Aven des Planes. Bilan Scientifique de la Ré-gion PACA 2007, 224e225.
Crégut-Bonnoure, E., 2008. 18 000 ans d’évolution de la faune mammalienne enVaucluse. Bulletin archéologique de Provence 5e6, 45e60.
Crégut-Bonnoure, E., Argant, J., Bailon, S., Boulbes, N., Bournery, A., Laudet, F.,Oppliger, J., Thinon, M., 2007a. Monieux: Coulet des Roches. Bilan Scientifiquede la Région PACA 2008, 245e246.
Crégut-Bonnoure, E., Bailon, S., Lachenal, Th., Perez, B., Thinon, M., 2007b. Monieux:Aven Souche. Bilan Scientifique de la Région PACA 2008, 247e248.
Crégut-Bonnoure, E., Argant, J., Boulbes, N., Debard, E., Magnin, F., 2008. Monieux:Coulet des Roches. Bilan Scientifique de la Région PACA 2009, 223e224.
Crégut-Bonnoure, E., Argant, J., Bailon, S., Boulbes, N., Buisson-Catil, J., Debard, E.,Desclaux, E., Oppliger, J., Roger, Th., Thinon, M., 2009a. Monieux: Coulet desRoches. Bilan Scientifique de la Région PACA, 239e241.
Crégut-Bonnoure, E., Fourvel, J.-B., Argant, J., Bouville, Cl., Buisson-Catil, J., Roger, T.,2009b. Monieux: Aven Souche 1. Bilan Scientifique de la Région PACA, 241e242.
Crégut-Bonnoure, E., Argant, J., Bailon, S., Boucher, Ch., Boulbes, N.,Bournery, A., Buisson-Catil, J., Debard, E., Desclaux, E., Kuntz, D., Lateur, N.,Laudet, F., Manzano, A., Margarit, X., Oppliger, J., Roger, Th., Russ, R.,Thinon, M., 2010a. Monieux: Coulet des Roches. Bilan Scientifique de laRégion PACA, 227e228.
Crégut-Bonnoure, E., Fourvel, J.-B., Roger, Th, 2010b. Monieux: Aven Souche 1. BilanScientifique de la Région PACA, 226e227.
Crégut-Bonnoure, E., Argant, J., Fourvel, J.-B., Roger, T., Boulbes, N., Desclaux, E.,Frèrebeau, N., Manzano, A., 2011a. Monieux: Coulet des Roches. Bilan Scienti-fique de la Région PACA, 223e224.
Crégut-Bonnoure, E., Argant, J., Fourvel, J.-B., Frèrebeau, N., Krzepkowska, J.,Thinon, M., 2011b. Monieux: Aven Souche 1. Bilan Scientifique de la RégionPACA, 223.
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
Crégut-Bonnoure, E., Boulbes, N., Desclaux, E., Frèrebeau,, N., Marciszak, A.,Roger, Th., 2012. Monieux: Coulet des Roches. Bilan scientifique de la régionPACA, 217e218.
Daniel, M., 1979. Et la taupe y creusa son trou. Blousons d’Argile 9, 45e48.Delpech, F., 1973. Présence de « Mustela minuta» Pomel (1853) (¼ « Mustela rixosa »
Bangs, 1896) dans les couches aurignaco-périgordiennes du gisement de Roc deCombe (commune de Payrignac, Lot). Bulletin de l’Association française pourl’étude du Quaternaire 10, 3e6.
Delpech, F., 1989. Les Mustélidés. In: Campy, M., Chaline, J., Vuillemey, M. (Eds.), LaBaume de Gigny (Jura). Gallia Préhistoire 27, 61e68.
Delpech, F., 1999. Biomasse d’Ongulés au Paléolithique et inférences sur la démo-graphie/Ungulate Biomass during the Paleolithic Period and Inferences con-cerning Human Demography. Paleo 11, 19e42.
Desclaux, E., Defleur, A., 1997. Etude préliminaire des micromammifères de laBaume Moula-Guercy à Soyons (Ardèche, France). Systématique, bio-stratigraphie et paléoécologie. Quaternaire 8, 213e223.
Diedrich, C., 2010. Specialized horse killers in Europe: Fetal horse remains in theLate Pleistocene Srbsko Chlum-Komín Cave hyena den in the Bohemian Karst(Czech Republic) and actualistic comparisons to modern African spotted hyenasas zebra hunters. Quaternary International 220, 174e187.
Donard, E., 1982. Recherche sur les léporinés quaternaires (Pléistocène moyen etsupérieur, Holocène). Thèse de 3� cycle. Université de Bordeaux I.
Eisenmann, V., 1984. Sur quelques caractères adaptatifs du squelette d’Equus etleurs implications paléoécologiques. Bulletin du Muséum National d’HistoireNaturelle de Paris 2, 185e195.
Eisenmann, V., Cregut-Bonnoure, E., Moigne, A.-M., 1985. Equus mosbachensis et lesgrands chevaux de la Caune de l’Arago et de Lunel-Viel: craniologie comparée.Bulletin du Muséum National d’Histoire Naturelle de Paris 4, 17e173.
Gauthier, P., Paccard, M., 1962. L’abri Edward (commune de Méthamis e Vaucluse).Cahiers Ligures de Préhistoire et d’Archéologie 11, 1e11.
Guadelli, J.-L., 1991. Les chevaux de Solutré (Saône-et-Loire, France). Cahiers duQuaternaire 16, 261e336.
Fontana, L., 2003. Characterization and exploitation of the Arctic Hare (Lepus tim-idus) during the Magdalenian: surprising data from Gazel Cave (Aude, France)?In: Costamagno, S., Laroulandie, V. (Eds.), Mode de vie au Magdalénien: Apportsde l’archéozoologie. Actes du XIVè Congrès UISPP de Liège (2001), BAR Int. Ser,vol. 1144. Archaeopress, pp. 101e118.
Fietzke, J., Liebetrau, V.A., Eisenhauer, A., Dullo, Ch, 2005. Determination of Uraniumisotope ratios by multi-static MIC-ICP-MS: method and implementation forprecise U- and Th-series isotope measurements. Journal of Analytical AnatomicSpectrometry 20, 395e401.
Haller-Probst, M.S., 1997. Neues Salamandridenmaterial von süddeutschen Fun-dorten. Bayerische Staatssammlung für Palaeontologie und Historische Geo-logie Mitteilungen 37, 53e73.
Hanquet, C., Desclaux, E., 2011. Analyse paléoécologique des communautés demicromammifères de la Caune de l’Arago (Tautavel, France) dans le contextedes migrations de faunes en Europe méridionale au cours du Pléistocènemoyen. Quaternaire 22, 37e47.
Heptner, V.G., Naumov, N.P., 1967. Mlekopitayushchije Sovetskogo Soyuza. In: Partlb. _Zinszczinyje (Kunicy; dopolnitelnychwidow), vol. II. Vysshaya Shkola,Moscow.
Heschler, K., 1907. Die Tierreste im Kesslerloch bei Thayngen. Neue DenkschriftenSchweiz 43, 61e154.
Hugueney, M., 1975. Les Mustelidés (Mammalia, Carnivora) du gisement pléistocènemoyen de la Fage (Corrèze). Nouvelles Archives du Muséum d’Histoire Naturellede Lyon 13, 29e46.
Jeannet, M., 2009. L’environnement tardiglaciaire préalpin: essai de reconstitutionbasée sur le potentiel climatique et écologique des microvertébrés. RevueArchéologique de l’Est 58, 5e56.
Koby, F.E., 1959. Contribution au diagnostic ostéologique différentiel de Lepus tim-idus L. et de Lepus europaeus Pallas. Verhandlungen des NaturforschendenGesellschaft in Basel 70, 19e44.
Koby, F.E., 1960. Contribution à la connaissance des lièvres fossiles, principalementde ceux de la dernière glaciation. Verhandlungen des NaturforschendenGesellschaft in Basel 71, 149e176.
Jean, M., 1973. Le Coulet des Roches n� 1. Blousons d’Argile 5, 6.Jean, M., 1977. Activités sportives. Blousons d’Argile 8, 6e7.Laroulandie, V., 2003. Exploitation des Oiseaux au Magdalénien en France: état des
lieux. In: Costamagno, S., Laroulandie, V. (Eds.), Mode de vie au Magdalénien:Apports de l’archéozoologie/Zooarchaeological. Insights into Magdalenian life-ways, Actes du colloque 6.4 du XIVe Congrès du l’UISPP, Liège, Belgique, 2e8septembre 2001, BAR, vol. 1144, pp. 129e133.
Le Fahler, B., Perret, J.-F., 1995. Aven du Coulet des Roches n� 1 ou du Petit cheval.In: Gaubert, G., Le Fahler, B. (Eds.), Les arcanes de Vaucluse. Hydrogéologie etspéléologie des territoires alimentant en eau la fontaine de Vaucluse. Asso-ciation de Recherches et d’Etudes Hydrogéologiques du Plateau d’Albion,p. 223.
Magniez, P., Boulbes, N., 2013. Environment at the transition Middle and LatePalaeolithic in South of France. The archaeological sequence of the Tournal Cave(Bize-Minervois, France). Quaternary International in press.
Marquet, J.C., 1993. Paléoenvironnement et chronologie des sites du domaineatlantique français d’âge pléistocène moyen et supérieur d’après l’étude desRongeurs. Les cahiers de la Claise 2, 1e345.
Miller, R., 2012. Mapping the expansion of the Northwest Magdalenian. QuaternaryInternational 272e273, 209e230.
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/

E. Crégut-Bonnoure et al. / Quaternary International xxx (2013) 1e21 21
Mourer-Chauviré, C., 1980. Le gisement Pléistocène supérieur de la grotte de Jaurensà Nespouls (Corrèze, France): les Equidés (Mammalia, Perissodactyla). Nou-velles Archives du Muséum d’Histoire Naturelle de Lyon 18, 17e60.
Olive, M., 2004. Géographie du département de Vaucluse. In: Buisson-Catil, J.,Guilcher, A., Hussy, Ch., Olive, M., Pagni, M. (Eds.), Vaucluse préhistorique. Leterritoire, les hommes, les cultures et les sites, p. 14. Barthélémy.
Paccard, M., 1954. La grotte d’Unang (Gorges de La Nesque, Malemort, Vaucluse).Cahiers Ligures de Préhistoire et d’Archéologie 4, 3e27.
Paccard, M., 1956. Du Magdalénien en Vaucluse: l’abri Soubeyras (Ménerbes, Vau-cluse). Cahiers Ligures de Préhistoire et d’Archéologie 5, 1e33.
Paccard, M., 1961. Nouvelle étude statistique des industries de l’abri Soubeyras(Ménerbes, Vaucluse). Cahiers Ligures de Préhistoire et d’Archéologie 10,1e15.
Paccard, M., 1962. L’abri n� 1 de Chinchon (com. de Saumanes de Vaucluse). Bulletinde la Société préhistorique française LVIII, 40e44.
Paccard, M., 1963. Le gisement préhistorique de Roquefure (commune de Bonnieux,Vaucluse). Cahiers Rhodaniens 10, 3e36.
Pailhaugue, N., 1998. Faune et saisons d’occupation de la salle Monique au Mag-dalénien Pyrénéen, grotte de la Vache (Alliat, Ariège, France). Quaternaire 9,385e400.
Parein, R., 1990. Guide spéléo des Monts de Vaucluse. Rimbaud, Cavaillon.Parein, R., Languille, A., 1981. La Haute-Provence souterraine. Echirolles.Perret, J.-F., 1975. Aven de Souche. In: Gaubert, G., Le Fahler, B. (Eds.), Les arcanes de
Vaucluse. Hydrogéologie et spéléologie des territoires alimentant en eau lafontaine de Vaucluse. Association de Recherches et d’Etudes Hydrogéologiquesdu Plateau d’Albion, p. 230.
Pleguezuelos, J.M., 1997. Elaphe scalaris (Schinz, 1822). In: Salvador, A. (Ed.), Reptiles.In: Ramos, M.A., et al. (Eds.), Fauna Ibérica, vol. 10. Museo Nacional de CienciasNaturales. CSIC, Madrid, pp. 390e407.
Prat, F., 1968. Recherche sur les Equidés pléistocènes en France. Thèse de doctorat.Faculté des Sciences de Bordeaux.
Please cite this article in press as: Crégut-Bonnoure, E., et al., The karst o(LGM) and the Late-glacial period palaeoenvironment of southeasternj.quaint.2013.10.058
Reichstein, H., 1993a. Mustela erminea. In: Niethammer, J., Krapp, F. (Eds.), Hand-buch der Säugetiere Europas. Band 5: Raubsäuger e Carnivora (Fissipedia). TeilII: Mustelidae 2, Viverridae, Herpestidae, Felidae. AULA-Verlag, pp. 533e571.
Reichstein, H., 1993b. Mustela nivalis. In: Niethammer, J., Krapp, F. (Eds.), Handbuchder Säugetiere Europas. Band 5: Raubsäuger e Carnivora (Fissipedia). Teil II:Mustelidae 2, Viverridae, Herpestidae, Felidae. AULA-Verlag, pp. 571e627.
Schackleton, N.J., Opdyke, N.D., 1973. Oxygen Isotope and Palaeomagnetic Stratig-raphy of Equatorial Pacific Core V28e238: oxygen isotope temperatures and icevolumes on a 105 year scale. Quaternary Research 3, 39e55.
Schmidt, K., 1992. Skull variability of Mustela nivalis Linnaeus, 1766 in Poland. ActaTheriologica 37, 141e162.
Stresemann, E., 1974. Exkursionsfauna für die Gebiete der DDR und BRD. Teil 3.Wirbeltiere. Volk und Wissen, Berlin.
Szyndlar, Z., 1984. Fossil snakes from Poland. Acta Zoologica Cracoviensia 28, 1e156.Szyndlar, Z., 1991a. A review of Neogene and Quaternary snakes of central and
eastern Europe. Part I: Scolecophidia, Boidae, Colubrinae. Estudia Geologica 47,103e126.
Szyndlar, Z.,1991b. A reviewofNeogene andQuaternary snakes of central and easternEurope. Part II: Natricinae, Elapidae, Viperidae. Estudia Geologica 47, 237e266.
Teacher, A.G.F., Thomas, J.A., Barnes, I., 2011. Modern and ancient red fox (Vulpesvulpes) in Europe show an unusual lack of geographical and temporal struc-turing, and differing responses within the carnivores to historical climaticchange. Evolutionary Biology 11, 214e222.
Toschi, A., 1965. Lagomorpha. Fauna d’Italia 7, 1e47.Tyrberg, T., 1998. Pleistocene Birds of the Palearctic: A Catalogue. Nuttall Ornitho-
logical Club, Cambridge.Van Raden, U.J., Colombaroli, D., Gill, A., Schwander, J., Bernasconi, S.M., Van
Leeuwen, J., Leuenberger, M., Eicher, U., 2013. High-resolution late-glacialchronology for the Gerzensee lake record (Switzerland): d18O correlation be-tween a Gerzensee-stack and NGRIP. Palaeogeography Palaeoclimatology,Palaeoecology 391, 13e24.
f the Vaucluse, an exceptional record for the Last Glacial MaximumFrance, Quaternary International (2013), http://dx.doi.org/10.1016/




















