
Palaeoenvironment of the Acheulian Gesher Benot Ya’aqov Pleistocene lacustrine strata, Northern...
13
Palaeoenvironment of the Acheulian Gesher Benot Ya’aqov Pleistocene lacustrine strata, Northern Israel––lithology, ostracod assemblages and ostracod shell geochemistry A. Rosenfeld a, * , Y. Nathan a , C.S. Feibel b , B. Schilman a , L. Halicz a , N. Goren-Inbar c , R. Siman-Tov a a Geological Survey of Israel, 30 Malkhe Yisrael Street, 95501 Jerusalem, Israel b Department of Anthropology and Geological Sciences, Rutgers University, New Brunswick, NJ 08901, USA c Institute of Archeology, Hebrew University, 91240 Jerusalem, Israel Received 23 July 2003; accepted 31 October 2003 Abstract The Acheulian archaeological site Gesher Benot Ya’aqov (GBY) is located along the shoreline of a Pleistocene Lake, the GBY Lake, within the Jordan Rift valley. The uniqueness of the site is expressed in its complex sedimentary sequence, rich fossil assemblages, and in the first appearance of lithic technologies not previously seen in Eurasia. The site is reliably dated at the Matuyama–Brunhes geomagnetic polarity chron boundary (780 kA), which occurs within the lake sediments. The sedimentary sequence at GBY exhibits a pronounced cyclicity. A single first order cycle begins with a lower conglomerate followed by lacustrine beds, overlain by an upper conglomerate that closes the cycle. Second-order cyclicity is apparent in shifts from littoral to deeper water facies, with five such cycles, identified within the first order cycle. The diatom and ostracod distri- butions indicate a freshwater–oligohaline environment of deposition for the lower part of the section and oligohaline–meio-meso- haline waters in the upper part of the section. In addition, the C and O isotopes and the Sr/Ca and Mg/Ca ratios of the ostracod (Candona neglecta) shells exhibit three geochemical zones supporting the paleoecological biozonation. They reflect a cooler period for the lower part of the sequence (interval I), changing into warmer climate at the Matuyama–Brunhes boundary and upward (interval II). Further gradual temperature increase in the upper interval III caused the retraction and desiccation of the GBY Lake. Ó 2003 Elsevier Ltd. All rights reserved. 1. Introduction The Acheulian archaeological site of Gesher Benot Ya’aqov (GBY) is located along the Jordan River (Fig. 1) in the northern part of the African-Syrian Rift Sys- tem. It is situated within a narrow north–south oriented valley at the southeastern end of the Hula Basin (Fig. 1), in northern Galilee, Israel (33°00 0 30 00 N, 35°37 0 30 00 E). GBY has been known for its artefacts and fossils since 1934 (Stekelis et al., 1937; Stekelis, 1960) and documents hominid activities along the margins of this palaeo-Hula Lake. The GBY palaeo-lake is one of a series of lakes, which existed in the Hula Basin, the northern extension of the Dead Sea Rift Valley. Excavations undertaken in 1989–1997 south of the Benot Ya’aqov bridge (southern margins of the GBY Lake) recovered a rich assemblage of lithic artefacts and vertebrate fossils (Goren-Inbar and Belitzky, 1989; Goren-Inbar et al., 1992). In addition, the site contains unusually well pre- served organic remains such as fossil wood, seeds, fruits and pollen. The uniqueness of the site is reflected in the complex sedimentary sequence, rich fossil record, a lithic artefact assemblage and a boundary, between magnetic polarity zones, which was correlated with the Matuyama–Bruhnes chron boundary (Goren-Inbar et al., 2000). This marks the movement of early hominid technologies out of Africa around 780 kA (date of the Matuyama–Bruhnes chron boundary). During the recent excavations, geological work on the stratigraphy and sedimentology of the site was accompanied by systematic sampling of the sequence for microfossil remains. * Corresponding author. E-mail address: [email protected] (A. Rosenfeld). 0899-5362/$ - see front matter Ó 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.jafrearsci.2003.10.008 Journal of African Earth Sciences 38 (2004) 169–181 www.elsevier.com/locate/jafrearsci
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Palaeoenvironment of the Acheulian Gesher Benot Ya’aqov Pleistocene lacustrine strata, Northern...

Journal of African Earth Sciences 38 (2004) 169–181
www.elsevier.com/locate/jafrearsci
Palaeoenvironment of the Acheulian Gesher Benot Ya’aqovPleistocene lacustrine strata, Northern Israel––lithology,ostracod assemblages and ostracod shell geochemistry
A. Rosenfeld a,*, Y. Nathan a, C.S. Feibel b, B. Schilman a, L. Halicz a,N. Goren-Inbar c, R. Siman-Tov a
a Geological Survey of Israel, 30 Malkhe Yisrael Street, 95501 Jerusalem, Israelb Department of Anthropology and Geological Sciences, Rutgers University, New Brunswick, NJ 08901, USA
c Institute of Archeology, Hebrew University, 91240 Jerusalem, Israel
Received 23 July 2003; accepted 31 October 2003
Abstract
The Acheulian archaeological site Gesher Benot Ya’aqov (GBY) is located along the shoreline of a Pleistocene Lake, the GBY
Lake, within the Jordan Rift valley. The uniqueness of the site is expressed in its complex sedimentary sequence, rich fossil
assemblages, and in the first appearance of lithic technologies not previously seen in Eurasia. The site is reliably dated at the
Matuyama–Brunhes geomagnetic polarity chron boundary (�780 kA), which occurs within the lake sediments.
The sedimentary sequence at GBY exhibits a pronounced cyclicity. A single first order cycle begins with a lower conglomerate
followed by lacustrine beds, overlain by an upper conglomerate that closes the cycle. Second-order cyclicity is apparent in shifts
from littoral to deeper water facies, with five such cycles, identified within the first order cycle. The diatom and ostracod distri-
butions indicate a freshwater–oligohaline environment of deposition for the lower part of the section and oligohaline–meio-meso-
haline waters in the upper part of the section. In addition, the C and O isotopes and the Sr/Ca and Mg/Ca ratios of the ostracod
(Candona neglecta) shells exhibit three geochemical zones supporting the paleoecological biozonation. They reflect a cooler period
for the lower part of the sequence (interval I), changing into warmer climate at the Matuyama–Brunhes boundary and upward
(interval II). Further gradual temperature increase in the upper interval III caused the retraction and desiccation of the GBY Lake.
� 2003 Elsevier Ltd. All rights reserved.
1. Introduction
The Acheulian archaeological site of Gesher Benot
Ya’aqov (GBY) is located along the Jordan River (Fig.
1) in the northern part of the African-Syrian Rift Sys-
tem. It is situated within a narrow north–south oriented
valley at the southeastern end of the Hula Basin (Fig. 1),
in northern Galilee, Israel (33�0003000N, 35�3703000
E).GBY has been known for its artefacts and fossils since
1934 (Stekelis et al., 1937; Stekelis, 1960) and documents
hominid activities along the margins of this palaeo-Hula
Lake. The GBY palaeo-lake is one of a series of lakes,
which existed in the Hula Basin, the northern extension
of the Dead Sea Rift Valley. Excavations undertaken
* Corresponding author.
E-mail address: [email protected] (A. Rosenfeld).
0899-5362/$ - see front matter � 2003 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jafrearsci.2003.10.008
in 1989–1997 south of the Benot Ya’aqov bridge
(southern margins of the GBY Lake) recovered a rich
assemblage of lithic artefacts and vertebrate fossils
(Goren-Inbar and Belitzky, 1989; Goren-Inbar et al.,
1992). In addition, the site contains unusually well pre-
served organic remains such as fossil wood, seeds, fruits
and pollen. The uniqueness of the site is reflected in
the complex sedimentary sequence, rich fossil record, alithic artefact assemblage and a boundary, between
magnetic polarity zones, which was correlated with the
Matuyama–Bruhnes chron boundary (Goren-Inbar
et al., 2000). This marks the movement of early hominid
technologies out of Africa around 780 kA (date of
the Matuyama–Bruhnes chron boundary). During the
recent excavations, geological work on the stratigraphy
and sedimentology of the site was accompanied bysystematic sampling of the sequence for microfossil
remains.

Fig. 1. Location map.
170 A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181
Non-marine ostracods have been used extensively as
proxy indicators of palaeosalinities, palaeotemperatures
and lacustrine palaeoenvironments (e.g., Chivas et al.,
1985, 1986; De Deckker and Forester, 1988; Holmes and
Horne, 1999). The aim of this study is the reconstruction
of the palaeoenvironments within the palaeo-GBY Lake
during the time of the early hominid occupation. We
report here on a detailed analysis of the fossil ostracodfaunas from GBY and their geochemical signatures as
well as supporting data from sedimentological studies
and diatom analysis.
One of our aims is to contribute data on the associ-
ation between the hominids and lake-margin environ-
ments. A detailed reconstruction of the lake properties
could add much to the understanding of a well-known
phenomenon of repetitive Palaeolithic occupationsalong lake shores that are known from Africa and
Eurasia. The site of GBY, one of the few documented in
the Levant, attests to the fact that the ‘‘Levantine cor-
ridor’’ was a unique ecological niche, which provided
areas rich in resources and was supportive of the
Acheulian hominid mode of living.
2. Geological setting
2.1. General
The Hula Basin and the Jordan Valley are the
northern extension of the Dead Sea–Red Sea–East
African Rift System (Fig. 1). The Rift in the Levant
represents a north–south trending depressed morpho-structure that constitutes a natural passageway between
Africa and Eurasia. Subsidence in the Hula Basin began
in the late Neogene, and continues to the present
(Horowitz and Horowitz, 1985). Freshwater lakes and
marshes have existed more or less continuously within
the basin, and their sediments are locally interstratified
with basalt flows. Late Quaternary to Recent sediments
of the Hula Lake have been the subject of numerousinvestigations (e.g., Cowgill, 1969, 1973, 1994; Racek,
1974; Kafri and Lang, 1987). The chemical and isotopic
composition of gastropod shells has been used to
reconstruct the environments of deposition of Quater-
nary freshwater lakes in the area (Rosenthal et al., 1989;
Moshkovitz and Magaritz, 1987).

A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181 171
At GBY, Quaternary strata are exposed along bothbanks of the Jordan River. Faulting and folding asso-
ciated with the left-lateral Dead Sea Transform have
deformed those sediments (Goren-Inbar and Belitzky,
1989) tilting strata up to 45�. Exposures in the vicinity of
the site include four different Pleistocene formations
(Horowitz, 1973), along with at least two basalt flows.
2.2. Sedimentology
The Pleistocene littoral sediments exposed in exca-
vations at the GBY site (southern margins of the GBY
Lake), are part of the Benot Ya’aqov Formation
(Horowitz, 1973). They comprise some 35 m of inter-
bedded calcareous muds, coquinas and gravels (Figs. 2
and 3). The sediments reflect deposition along the lit-
toral zone of the GBY Lake (one of the Hula palaeo-
lakes); both subaerial and subaqueous deposits arepresent, reflecting lake-level fluctuations along the
palaeo-shoreline.
Subaerial components include both channel facies
and palaeosols. At low lake levels, gravel-bed streams
trangressed the site. They deposited basalt boulder to
cobble-conglomerates in tabular sheets. The matrix of
the clast-supported conglomerates is a hash of frag-
mented molluscs and sand, indicating close proximity tothe lake margin at the time of deposition. These con-
glomerates are prominent at the base and top of the
section exposed at the site. Scattered boulders are also
associated with most of the lake margin sands and
coquinas (shell beds), possibly introduced to the site by
flash floods. The fine-grained lacustrine facies are
Fig. 2. Stratigraphic sections of GBY (trenches). The figure shows the corr
obtain the composite section (Fig. 3).
punctuated by several pedogenic profiles, documentingsoil development during brief desiccation events. All
other facies reflect deposition in beach and shallow
water environments.
The lacustrine facies document deposition in two
broad settings: the higher energy strand line and the low
energy offshore environment. Molluscan coquinas and
pebbly sands are characteristic of the lake margin set-
ting, and it is this facies association which preserves thebulk of the stone tool, and fossil vertebrate remains. The
high-energy deposits appear to be dominated by accre-
tion during storm events (Feibel, 2001). In these
deposits, tabular beds of 10–20 cm thickness occur,
often with few or no internal primary structures.
Archaeological accumulations are concentrated at the
interfaces between such depositional units, while few
stone tools or bones occur within them. Thus the unitsthemselves are interpreted to reflect short-term (storm)
event stratification, while the interfaces between them
reflect depositional hiatuses over which accumulation of
stone-tool discard and fossil bone occurred. Deposits of
the low-energy offshore settings are predominantly
marls (54–68% CaCO3) with a well-preserved organic
component (3–21% OM) with only a minor detrital
contribution. These marls occur in thick packages (1–5m), but internally show a high degree of variability in
particle size, carbonate content and organic matter,
reflecting local and short-term variability in the char-
acter of aquatic vegetation, currents, oxygenation and
other parameters.
The lacustrine sediments contain a rich and varied
assemblage of aquatic invertebrates (molluscs, ostracods,
elations between the beds in the various trenches which were used to

Fig. 3. Composite section at GBY, showing the lithology; the Matuyama–Brunhes geomagnetic boundary; the sedimentological cycles; the
occurrence of the Acheulian artefacts; the trends of the d18O, d13C, Sr/Ca and Mg/Ca ratios for Candona neglecta valves as well as the ostracod
population density, biozones and palaeosalinity.
172 A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181
charophytes, and diatoms). Two types of carbonate mud
are present: organic matter-rich (12–21% OM) and or-
ganic matter-poor (3–8% OM). The organic matter-rich
carbonate muds reflect deposits in low-energy poorlyoxygenated settings, and are generally grey to greyish-
brown. Other than ostracods, they are relatively impov-
erished in calcareous fossils. All the carbonate of this type
of mud is calcite; the endogenic calcite in this type of mud
originates from ostracod valves. Nearly all the sediments
with abundant ostracod faunas are of this type of car-
bonate mud (organic-rich). The organic-poor carbonate
muds, deposited in high-energy open circulating waterssettings, are brown to dark grayish brown. They are rich
in gastropod shells and poor in ostracods. The carbonate
of this type of mud is mostly calcite but contains signifi-
cant amounts of aragonite (gastropod shells). The dif-
ferences between the two types of muds are probably due
to the different environment of deposition, most of the
OM is oxidized in the high-energy settings. The organic
matter-poor, carbonate muds grade into molluscan sandsand coquinas (shell-beds) in which carbonate content
may exceed 90%. These however tend to be poor in os-
tracods, probably because they are largely wave sorted
and wave deposited sediments.
2.3. Age
The sedimentary sequence at GBY lies above the
Yarda Basalt, which has been dated at �900 kA (Hei-
man et al., 1987; Goren-Inbar et al., 1992). A magnetic
polarity transition within the sequence (Fig. 3) is cor-
related with the Matuyama–Brunhes geomagnetic
polarity chron boundary at �780 kA (Bassinot et al.,
1994; Tauxe et al., 1996; Verosub et al., 1998; Goren-
Inbar et al., 2000). The sequence exhibits a pronounced
cyclicity in sedimentary character. A single first-ordercycle begins with the lower conglomerate followed by
lacustrine beds and closes with the upper conglomerate.
This cycle is shown in the composite stratigraphic
sequence (Fig. 3). Second-order cyclicity is apparent in
shifts from littoral to offshore facies, with five such
cycles found within the larger pattern. The magnitude,
character and relationships of these signals have been
interpreted to represent climatic forcing of lake levelfluctuations at the Milankovitch periodicities of roughly
20 and 100 kA (Feibel et al., 1998). This is supported
by the age of the Yarda Basalt (Heiman et al., 1987) and
the detailed analysis of the magnetic characteristics of
the sediments (Verosub et al., 1998, 1999; Goren-Inbar
et al., 2000), which suggest a span of slightly less than
100–150 kA for a 24 m segment of the composite sec-
tion. The combination of cyclostratigraphy and thechron boundary allow direct correlations of the GBY
sequence to oscillations in and around marine Oxygen
Isotope Stage 19.
3. Samples and methods
The samples were obtained from six trenches, whose
locations are given in Goren-Inbar et al. (2000). Fig. 2illustrates the stratigraphic sections of the six trenches,
and their correlation is the basis of the composite
stratigraphic section (Fig. 3). The correlation of the

A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181 173
various trenches was made on the basis of a magneto-stratigraphic study of 155 oriented samples from tren-
ches II–VI (Goren-Inbar et al., 2000). Fig. 3 shows the
position of the second-order cycles, the position of
the Matuyama–Brunhes boundary and the lithology.
The dominance of coarser lithologies at certain levels,
made up mainly of gastropods (Melanopsis sp., Vivipa-
rus sp.) and their debris, resulted in minor gaps in rep-
resentation. More than forty samples (from TrenchesVI, II and III, Table 1) were examined for ostracods.
Only twenty-seven contained enough ostracods to carry
out geochemical analyses. Relatively large samples of
sediment (up to 500 g) had to be washed to obtain en-
ough material (ostracods) for the palaeontological and
geochemical analyses.
All the geochemical analyses were performed at the
Geological Survey of Israel. Only adult specimens of theostracod Candona neglecta were employed in the geo-
chemical study. This species was chosen because of its
abundance in the samples and because of the detailed
experimental studies of Xia et al. (1997a,b) on a species
of the same genus (Candona rawsoni). The shell chem-
istry of Candona species has already been used for a
similar palaeoenvironmental reconstruction (Anadon
et al., 1994). Ostracod valves for the geochemical studywere hand-picked from the sieved separates, and special
care was taken to avoid contamination. A non-metallic
brush was used for picking the ostracods into the ana-
lyzed tubes to avoid metallic contamination. The os-
tracods were washed twice with double de-ionized water,
and then dissolved in ultra-pure dilute hydrochloric
acid. Sr, Ca and Mg concentrations were determined by
ICP-MS (Perkin Elmer, Sciex Elan 6000). Blanks resultswere lower than the detection limit, and the results of
one sample analyzed in triplicate were within a 10%
range. All results were obtained as ratios to avoid errors
due to weighing very small samples.
Sixteen oxygen and carbon isotopic measurements of
the ostracod species (Candona neglecta) were done on a
VG ISOCARB system coupled to a SIRA-II mass-
spectrometer, according to the method of Shackleton(1974). In the Isocarb, the samples (five to eight adult
valves each) were reacted successively under vacuum in
a reservoir of �100% orthophosphoric acid (H3PO4) at
90 �C. Calibration of the mass spectrometer to the
international PDB Vienna (Pee Dee Belemnite) car-
bonate standard was done through the National Bureau
of Standards (NBS) 9, following Coplen (1988).
Instrument precision was better than ±0.1& for bothd13C and d18O values, and the external reproducibility
based on duplicate measurements of reference standard
was 6 0.05&.
Thirty samples (from Trenches II–IV) were examined
for diatoms. Only two samples yielded a few diatoms.
For diatom analyses the samples were treated with HCl
and H2O2 through the standard procedure, with re-
peated washing in distilled water. Slides were preparedby using Naphrax high resolution mounting medium.
Fifteen bulk samples were examined by XRD, with a
Philips PW-1820/00 diffractometer and Cu Ka radiation
using a curved graphic monochromator. This was done
mainly to identify the CaCO3 polymorph present in the
samples (calcite or aragonite).
4. Results
4.1. Ostracods
The photographs of 10 ostracod species together with
the algae Chara are shown in Plate 1. To describe water
salinity we follow Wagner (1957, p. 108) who used the
Valikangas (1933) classification: Freshwater up to 0.5&;oligohaline water 0.5–3&; meio-mesohaline water 3–
10&; pleio-mesohaline 10–16.5&. Euryhaline describes
a species which has a tolerance for a rather wide range of
salinity.
4.1.1. Brief taxonomy and ecology
Cytheromorpha fuscata (Brady, 1868), (Plate 1, Figs.
12–14) is known to dwell only in an oligohaline–
mesohaline environment (Elofson, 1969), probably 0.6–
17&, in water depth of 0–30 m.
Heterocypris salina (Brady, 1868) (Plate 1, Fig. 15).
This species habitat is uniquely in an oligo–mesohalinewater environment, from 0.5& to 13& (Neale, 1964;
Henderson, 1990).
Ilyocypris gibba (Ramdohr, 1808) (Plate 1, Fig. 7)
lives in a freshwater to oligohaline environment of
deposition from 0.1& to 0.8& (Neale, 1964), in shallow
lakes and streams.
Ilyocypris bradyi (Sars, 1890) (Plate 1, Fig. 8) prefers
to live in cooler waters and is found in permanentstreams. It lives in freshwater to slightly mesohaline
water (Henderson, 1990).
Cyprideis torosa (Jones, 1850) is euryhaline and can
tolerate all the salinity range from fresh-water to
hypersaline, from 0.2& to 80&, in water depth of 0–30
m (Rosenfeld and Vesper, 1977).
Candona neglecta (Sars, 1887) (Plate 1, Figs. 1–3)
lives in freshwater to oligohaline environments of softand muddy sediments. The species is able to survive even
in places where water dries out (Wagner, 1957). This
species dominates (high population density) the whole
section. This is the main reason why this species was
chosen for geochemical (trace elements and stable iso-
topes) analyses.
Candona compressa (Koch, 1838) (Plate 1, Fig. 4)
generally inhabits freshwater to oligohaline environ-ments (up to 1&, Neale, 1964).
Candona angulata (Mueller, 1900) (Plate 1, Fig. 5)
dwells in freshwater to mesohaline water. According to

Table 1
Ostracod distribution and population density
Year/
sample
no.
Trench-
layer
Depth
(m)
Cypri-
deis
torosa
Candona
neglecta
Ilyocy-
pris
gibba
Ilocypris
bradyi
Candona
angulata
Darwinula
stevensoni
Dolero-
cypris
sp.
Cythero-
morpha
fuscata
Candona
com-
pressa
Hetero-
cypris
salina
Ilyocy-
pris sp.
Total
speci-
mens
Ostracod
biozones
Palaeo-
environment
96/201 VI-2 6.4 72� 9 3 7 2 93 2B Oligo-
haline–
mesohaline
203 VI-4 7.0 95 6 11 1 13 126
204 VI-4 8.0 10 2 4 16
205 VI-4 8.7 92 2 11 10 115
206 VI-5 9.3 11 1 1 13
207 VI-6 9.5 28 2 1 9 2 42
208 VI-7 9.8 65 3 5 14 8 2 97
209 VI-8 10.4 1 1
210 VI-9 11.0 2 3 10 15
211 VI-10 11.7 10 2 18 30
96/213 VI-12 13.0 3 3 5 10 9 30
95/54 II-3 13.2 18 5 1 3 24 4 37
56 II-5 14.0 22 11 2 35 2A
82 II-7 16.6 112 1 1 114
58 II-7 16.7 48 9 57
59 II-8 17.0 3 1 4
60 II-8 17.3 40 5 5 11 61
61 II-9 18.2 95 8 103
63 II-11 19.1 51 5 2 4 62 1 Freshwater–
oligohaline65 II-13 19.9 2 2
67 II-15 20.6 51 51
69 II-17 21.3 360 6 2 6 374
70 III-1 23.2 14 3 2 19
72 III-3 24.6 5 5
75 III-6 27.4 6 1 7
77 III-8 30.8 1 4 1 6
78 III-9 31.0 15 1 16
95/79 III-10 31.3 2 2
*Number of specimens per 350 g of sediment sample.
174
A.Rosen
feldet
al./JournalofAfrica
nEarth
Scien
ces38(2004)169–181
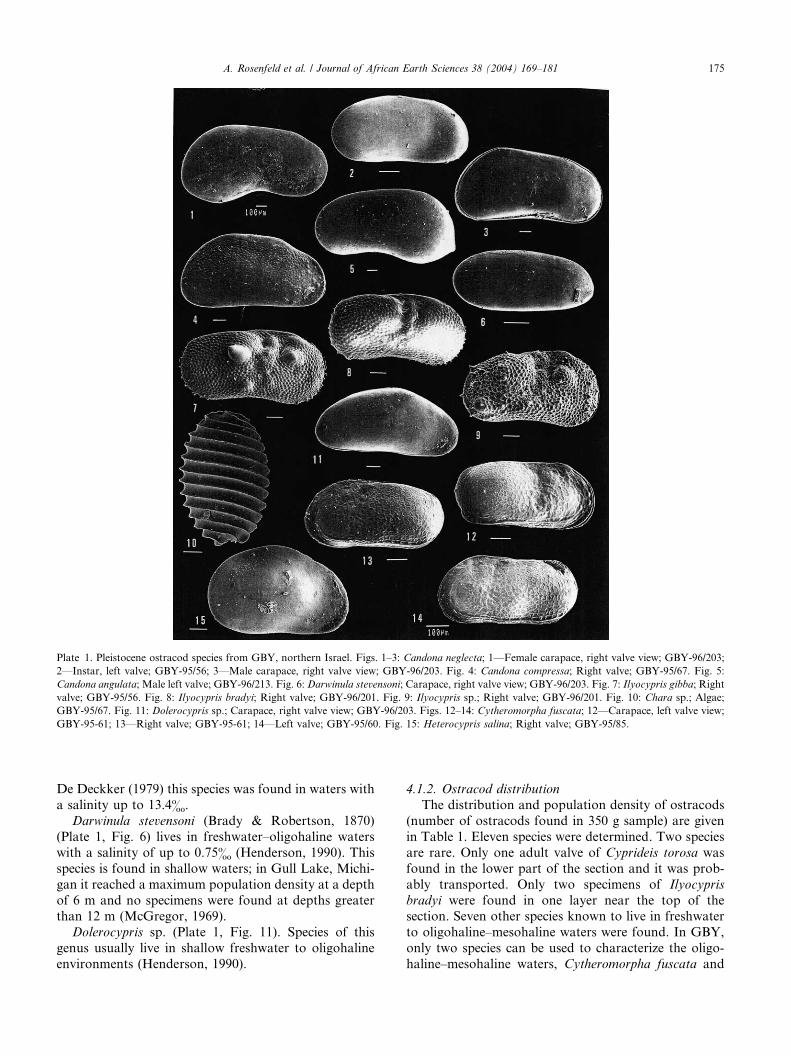
Plate 1. Pleistocene ostracod species from GBY, northern Israel. Figs. 1–3: Candona neglecta; 1––Female carapace, right valve view; GBY-96/203;
2––Instar, left valve; GBY-95/56; 3––Male carapace, right valve view; GBY-96/203. Fig. 4: Candona compressa; Right valve; GBY-95/67. Fig. 5:
Candona angulata; Male left valve; GBY-96/213. Fig. 6: Darwinula stevensoni; Carapace, right valve view; GBY-96/203. Fig. 7: Ilyocypris gibba; Right
valve; GBY-95/56. Fig. 8: Ilyocypris bradyi; Right valve; GBY-96/201. Fig. 9: Ilyocypris sp.; Right valve; GBY-96/201. Fig. 10: Chara sp.; Algae;
GBY-95/67. Fig. 11: Dolerocypris sp.; Carapace, right valve view; GBY-96/203. Figs. 12–14: Cytheromorpha fuscata; 12––Carapace, left valve view;
GBY-95-61; 13––Right valve; GBY-95-61; 14––Left valve; GBY-95/60. Fig. 15: Heterocypris salina; Right valve; GBY-95/85.
A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181 175
De Deckker (1979) this species was found in waters with
a salinity up to 13.4&.
Darwinula stevensoni (Brady & Robertson, 1870)
(Plate 1, Fig. 6) lives in freshwater–oligohaline waters
with a salinity of up to 0.75& (Henderson, 1990). This
species is found in shallow waters; in Gull Lake, Michi-
gan it reached a maximum population density at a depth
of 6 m and no specimens were found at depths greaterthan 12 m (McGregor, 1969).
Dolerocypris sp. (Plate 1, Fig. 11). Species of this
genus usually live in shallow freshwater to oligohaline
environments (Henderson, 1990).
4.1.2. Ostracod distribution
The distribution and population density of ostracods
(number of ostracods found in 350 g sample) are given
in Table 1. Eleven species were determined. Two species
are rare. Only one adult valve of Cyprideis torosa was
found in the lower part of the section and it was prob-
ably transported. Only two specimens of Ilyocypris
bradyi were found in one layer near the top of thesection. Seven other species known to live in freshwater
to oligohaline–mesohaline waters were found. In GBY,
only two species can be used to characterize the oligo-
haline–mesohaline waters, Cytheromorpha fuscata and

176 A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181
Heterocypris salina, both occur only in oligo–mesohaline waters; neither occur in freshwater. Both
their presence and absence are significant.
4.2. Diatoms
Two samples only (95/70 and 95/71), both from the
lower part of the section yielded a few diatoms with
relatively very low quantity. Most of the diatom valves
are very badly preserved, as a result of dissolution and/
or of mechanical breakage. Almost all the species were
identified at least to the genus stage mainly based onEhrlich (1995) and Krammer and Lange-Bertalot (1986,
1988, 1991a,b). Table 2 lists almost all the diatom spe-
cies that were found in the two samples and Plate 2
shows photographs of the diatoms.
These two samples yielded apart from diatoms, fish
teeth, gastropods and sponge spicules. Most of the
diatoms belong to the Fragilaria group. The diatom
assemblages of these two samples may be correlated tothe assemblages of the Hula Basin (Ehrlich, 1973):
sample GBY/95/70 to the assemblage of H1 zone and
sample GBY/95/71 to the H4 zone. Both assemblages
Table 2
Diatoms in samples 95/70 and 95/71
Species/sample no. 95/70
Achnanthes sp. Xa
? Achnanthes minutissima X
Amphora cf. libyca –b
? Anomoeoneis sphaerophora –
Aulacoseira cf. ambigua A single va
Aulacoseira cf. granulata X
Cocconeis placentula var. euglypta –
Cocconeis placentula var. lineata X
Cyclotella ? ocellata –
Cymbella cf. aspera –
Ellerbeckia arenaria X
Ephithemia sp. –
Ephithemia adnata –
Ephithemia frickei X
Epithemia turgida –
Eunotia cf. pectinalis –
Fragilaria spp. X
Fragilaria cf. brevistriata X
Fragilaria cf. pinnata X
Fragilaria construens var. construens X
Fragilaria construens var. venter X
? Fragilaria virescens X
Gomphonema sp. –
Gomphonema cf. insigne –
Navicula sp. X
Nitzschia amphibia –
Opephora martyi X
Pinnularia sp. X
Synedra sp. X
Synedra ? ulna –
aX¼present.b –¼Not detected.
indicate a very shallow freshwater, eutrophic, alkalineenvironment.
These two very badly preserved assemblages were
probably the result of breakage by currents and waves
and/or from being biologically ingested and digested.
Molluscs feed mainly on epilithic and epiphytic algae,
hence most of these diatom species, which are also epi-
lithic and epiphytic were probably ingested by molluscs.
4.3. Geochemical analyses
The results of the geochemical analyses are given inTable 3. Fig. 3 shows the trends of the d18O, d13C, andSr/Ca and Mg/Ca ratios along most of the section.
According to these results, the section may be divided
into three parts. I––a lower part in which the Sr/Ca and
Mg/Ca curves exhibit similar trends, while the d13C and
d18O have opposite trends. II––a middle part where the
Sr/Ca and Mg/Ca curves show opposite trends, while the
d13C and d18O have similar trends. III––an upper partwhere the relationships between the curves are not clear.
All the samples analysed by XRD contained appre-
ciable amounts of calcite, with ostracods constituting
95/71
X
–
X
X
lve –
X
X
X
X
X
X
X
X
–
X
X
–
X
–
X
X
–
X
X
X
X
X
X
X
X

Plate 2. Pleistocene diatom species from GBY, northern Israel. Fig. 1: Fragilaria construens var. venter (Ehrnberg) Grunow, 1881. Sample no. GBY-
95-71. Fig. 2: Fragilaria construens var. construens (Ehrnberg) Grunow, 1862 (broken valve). Sample no. GBY-95-70. Fig. 3: Fragilaria construens
var. construens (Ehrnberg) Grunow, 1862. Sample no. GBY-95-70. Fig. 4: Opephora martyi Heribaud, 1902. Sample no. GBY-95-70. Fig. 5:
Fragilaria cf. pinnata Ehrenberg, 1843. Sample no. GBY-95-70. Fig. 6: Cocconeis placentula var. lineata (Ehrenberg) Van Heurck, 1880–1885. Sample
no. GBY-95-70. Fig. 7: Ellerbeckia arenaria (Moore) Crawford, 1988. [The magnification in LM · 1000]. Sample no. GBY-95-70. Fig. 8: Aulacoseira
cf. granulata (Ehrenberg) Simonsen, 1979. Sample no. GBY-95-70. Fig. 9: Chrysophyte cyst. Sample no. GBY-95-71.
A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181 177
most of the endogenic, biogenic component. Aragonite
was identified only in samples, which contained gastro-
pod debris.
5. Discussion and palaeoecology
5.1. Ostracod biozones and population density
According to the ostracod distribution (Table 1), the
two ostracod species Heterocypris salina and Cythero-
morpha fuscata, which do not occur in freshwater, occur
only in the upper part of the section. The ostracod
Cytheromorpha fuscata var. significans that occurs infreshwater (Alm, 1916; quoted by Elofson, 1969, p. 209)
was not found in the GBY samples. All the ostracods
occurring in the lower part of the section can live in
freshwater to oligohaline environments of deposition.
Therefore a division into two parts according to ostra-
cod distribution may be discerned within the GBY sec-
tion: (1) a lower freshwater to oligohaline part up to 3&,
from �19 to 31 m, and (2) an oligohaline to oligohaline–
meio–mesosaline, up to 10&, upper part from �6 to 18
m. This division is a reflection of the first-order cycle, thefreshwater–oligohaline phase marks the beginning of the
lake and the slight increase in salinity is an indication of
a drier climate leading towards the lake desiccation. The
boundary between the freshwater–oligohaline ostracod
zone 1 and the oligohaline–meio–mesohaline zone 2
coincides more or less with the Matuyama–Brunhes
boundary (Fig. 3).
A secondary twofold division may be noticeable in theupper oligohaline–meio–mesohaline biozone 2: a lower
2A zone, with oligohaline waters, from 18 to 13.5 m

Table 3
Geochemistry results trace elements and C and O isotopes of Candona neglecta valves
Sample Layer Depth (m) Ca (ppm)a Mg (ppb)a Sr (ppb)a Sr/
Cab · 10�3
Sr/Mgb Mg/
Cab · 10�2
d13C &
PDB
d18O &
PDB
96/201 VI-2 6.4 2.406 9.82 2.13 0.406 0.060 0.673 )3.03 )5.2896/203 VI-4 7.0 2.752 8.57 2.39 0.397 0.077 0.514 +3.62 )5.0896/204 VI-4 8.0 4.521 13.10 3.93 0.398 0.833 0.478 )5.57 )4.5796/205 VI-4 8.7 1.216 4.83 1.25 0.471 0.072 0.656 )5.20 )4.7196/208 VI-7 9.8 11.142 28.44 12.02 0.494 0.117 0.421 )4.05 )4.7396/211 VI-10 11.7 2.697 11.73 2.27 0.385 0.054 0.718 )8.58 )4.9896/213 VI-12 13.0 3.038 86.98 2.88 0.433 0.009 4.724 )8.22 )4.6895/56 II-5 14.0 1.416 5.10 1.45 0.468 0.079 0.595 – –
95/82 IV-7 16.6 17.467 30.01 30.77 0.806 0.284 0.284 )3.26 )4.3095/58 II-7 16.7 6.304 11.14 10.90 0.791 0.271 0.292 )3.77 )4.4795/60 II/8 17.3 6.035 14.75 6.90 0.462 0.114 0.403 )7.02 )3.8195/61 II/9 18.2 7.144 19.72 5.47 0.351 0.077 0.455 )7.05 )4.1595/63 II/11 19.1 4.385 12.58 3.70 0.386 0.082 0.473 )5.94 )3.8395/67 II-15 20.6 2.602 10.53 3.31 0.582 0.064 0.716 )1.54 )4.3695/69 II-17 21.3 7.776 33.42 7.71 0.454 0.086 0.284 +0.97 )5.2895/70 III-1 23.2 0.721 3.13 0.97 0.616 0.086 0.716 )3.70 )5.0995/77 III-8 30.8 – – – – – – )2.82 )4.97aConcentration in solution (10 ml, see methods).bMolar ratio.
178 A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181
(occurrence of both Cytheromorpha fuscata and Hetero-
cypris salina) and an upper 2B zone, from 13.5 to 6.5 m
(absence of Cytheromorpha fuscata and presence of only
Heterocypris salina). The absence of Cytheromorpha
fuscata in the 2B zone may indicate a higher salinity in
this zone. It should be emphasized that we are evaluat-
ing the highest salinity to be only about 8–10&. Thissubdivision reflects the second order cyclicity. These two
parts may be correlated to the third and fourth second-
order sedimentary cycles.
Other findings strengthen the interpretation that the
lower part of the section had a freshwater–oligohaline
environment of deposition. The two diatom assemblages
(H1 and H4 of Ehrlich, 1973) found in the lower part of
the GBY sequence indicate a very shallow freshwater,eutrophic, alkaline environment. A poor epiphytic
assemblage characterizes zone H1. Zone H4 is domi-
nated by epiphytic species and by benthic–planktic
forms together with abundant sponge spicules and
chrysophyte cysts. Gasse et al. (1987, pp. 9–10) observed
a reasonably good correlation between chrysophyte
contents and the frequencies of freshwater organisms.
Thus we think that the specimens of the algae Chara sp.(Plate 1, Fig. 10), which occur only in the lower part of
the section (Sample 95/67) should indicate low salinity.
Furthermore, it should be noted that Candonid spe-
cies, which originate in cooler climates (Martens and
Ortal, 1999), represent about 60% of the ostracods
present in the GBY section, while they represent only
some 10% of the recent non-marine ostracods in Israel.
This is in accord with the conclusion of Martens andOrtal (1999) which states that most of the Mediterra-
nean served as refugium for Western European faunas
during glacial phases.
5.2. Geochemical zones
There is a strong relationship between the Sr/Ca and
Mg/Ca ratios of the ostracods and of the waters in
which they moulted (De Deckker et al., 1999, see also
references therein). Most of the relationships between
the two curves (Sr/Ca and Mg/Ca) in Fig. 3 may beexplained by the peaks in the deposition of biogenic
aragonite due to the relative abundance of gastropods in
the sediment, because aragonite is much richer in Sr
than calcite. The precipitation of aragonite lowers the
Sr/Ca ratio in the lake waters while precipitation of
calcite increases it. The precipitation of both calcite and
aragonite increases the Mg/Ca ratios. This is true even in
an open lake where the concentrations are diluted butthe cation ratios do not change significantly.
It is possible to distinguish three sections along the
Sr/Ca and Mg/Ca curves. I––A lower interval from �21
to �18 m where the two curves have similar trends. II––
A large middle interval from �17 to �9 m where the
relations are not clear-cut but in the main the Sr/Ca
ratios have opposite trends to those of the Mg/Ca ratios.
III––A small upper interval from �9 to �6 m where thetwo curves have again similar trends. In the lower
interval of the curve the endogenic, biogenic calcite is
low magnesium calcite (ostracod shells); therefore pre-
cipitation of this calcite increases both ratios. In the
middle–upper interval the endogenic, biogenic carbon-
ate is mainly aragonite (gastropod shells). Formation of
aragonite gastropod shells (rich in Sr) lowers the Sr/Ca
ratio while it increases the Mg/Ca ratio in the lake wa-ters. In the uppermost part it is again low magnesium
calcite. This explains the opposite trends of the Sr/Ca
and Mg/Ca ratios in parts of interval II and the well-

A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181 179
defined covariant trends in intervals I and III. Thealternation of relatively high and low Sr/Ca and Mg/Ca
ratios suggests alternating conditions which are consis-
tent with a small lake subject to rapid fluctuations. The
lower interval (I) may be correlated to the second sedi-
mentary cycle while the third interval (III) may be cor-
related to the fourth sedimentary cycle.
The variability of the d13C values of Candona neglecta
valves is greater than that of the oxygen isotopic values(Fig. 3). The values range between )9& and 3& with a
general cyclical pattern. The d13C values of ostracod
carbonate are mainly controlled by the d13C of dissolved
inorganic carbon (DIC) in the ambient lake water. At
the same time d13C values seem to be systematically
biased by microhabitat and vital effects (Von Graffen-
stein et al., 1999). In the bottom waters, both respiration
and oxidation of the organic matter that formed at thesurface waters at times of high productivity decrease the
oxygen concentration, raise the DIC and lower the d13Cof the DIC (Deuser and Hunt, 1969; Shackleton and
Vincent, 1978). There are three clear peaks of negative
value for d13C: )7.1& at 18.2 m, )8.6& at 11.7 m and
)5.6& at 8 m (Fig. 3). These values indicate high pro-
ductivity and it can be seen that they are found near the
beginning of the secondary sedimentary cycles 2, 3 and4. The lower and upper peaks of the negative d13Ccoincide with the ostracod population density peaks
(Table 1). The highest negative values (middle peak)
coincide with the exceptionally high Mg/Ca value (Table
3, Fig. 3) due to very high productivity (formation of
biogenic carbonate). We are aware of the exceptionally
high positive value of sample 96/203 (Table 3 and Fig.
3). Nevertheless, this is a single measurement on whichwe could not make a duplicate because of lack of os-
tracods, therefore apart from the fact that it agrees with
the lowering of productivity towards the top of the
section, we are unable to draw further conclusions.
The d18O values vary from )5.3& to )3.8&. They
show a trend to heavier isotopes in the interval between
21.3 and 17.3 m ()5.3& to )3.8&) and an opposite
trend in the interval 17.3–6.4 m ()3.8& to )5.3&). Weinterpret the d18O variations as mainly due to tempera-
ture changes and not due to concentration (evaporation)
or dilution (rain). We base this on the small salinity
changes inferred by the ostracod assemblages and the
fact that the GBY Lake was for most of its history an
open lake (Horowitz, 1979; Moshkovitz and Magaritz,
1987). Moshkovitz and Magaritz (1987), from their
study of C and O isotope records of middle and latePleistocene molluscs, reached the conclusion of high
productivity and found no evidence for high evapora-
tion in the Hula basin. Our results indicate a decrease in
temperature followed by an increase. The most promi-
nent climatic event seems to have occurred at the tran-
sition between intervals I and II, where the trends invert.
The decrease in temperature indicates the beginning of
the lake and the increase in temperature is consistentwith the interpreted retraction leading to its desiccation
toward the upper part of the sequence.
We are aware that there might have been short peri-
ods when the water level in the GBY Lake was so low
that it was lower than the exit, leading to the formation
of a closed lake during these short periods. We have
no direct evidence from the ostracod distribution or
their chemistry since they were formed at high stands,nevertheless the occurrence of pedogenic profiles indi-
cating brief desiccation events (see sedimentology) sup-
ports this possibility.
6. Conclusions
The cyclicity observed in the GBY sediments is
also recorded by the ostracod distribution, population
density and chemistry of their valves. The ostracod
distribution indicates at least two biozones, a lower
freshwater–oligohaline and an upper oligohaline–meso-
haline zone.
Climatic changes during the time interval sampledreflect (d18O values) a lowering of the temperature fol-
lowed by a gradual warming. This agrees with Horo-
witz’s (1979, 2001) synthesis, which concludes that
during pluvial periods the climate was cooler and these
periods are characterized by an expansion of the lakes in
the Jordan and Hula basins. During interpluvial peri-
ods, the climate was hotter and these periods are char-
acterized by the retraction of these lakes and thedesiccation of some of them.
The sedimentology, ostracod population density and
the ostracod valve chemistry (Sr/Ca and Mg/Ca ratios,
d13C and d18O) all exhibit a cyclicity, which reflects the
expansion and retraction of the lake. Five such cycles
were observed (Fig. 3). The detailed reconstruction of
the lake properties adds much to the understanding of a
well-known phenomenon of repetitive palaeolithicoccupations along lake shores, which are known from
Africa and Eurasia. The results of this study are viewed
as an additional evidence for the notion that the dis-
persal ‘‘Out of Africa’’ was a mechanism of repetitive
waves.
Acknowledgements
We are grateful to Mr. M. Dvorachek for the SEM
micrographs, Mrs. T. Beer for preparing and washing the
samples and Mr. Y. Levy for printing the photos. We
thank Mr. S. Hoyland for preparing the location map,
and Mrs. N. Shragay and B. Cohen for their technicalassistance. Last but not least we thank Dr. I. Boomer for
his careful review and remarks, which considerably im-
proved an earlier version of the manuscript. The Irene

180 A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181
Levi Sala Care Foundation and the L.S.B. LeakeyFoundation supported fieldwork at GBY.
References
Anadon, P., Utrilla, R., Julia, R., 1994. Paleoenvironmental recon-
struction of a Pleistocene lacustrine sequence from faunal assem-
blages and ostracode shell geochemistry, Baza Basin, SE Spain.
Palaeogeography, Palaeoclimatology, Palaeoecology 111, 191–205.
Bassinot, F.C., Labeyrie, L.D., Vincent, E., Quideleur, X., Shackelton,
N.J., Lancelot, Y., 1994. The astronomical theory of climate and
the age of the Brunhes–Matuyama reversal. Earth Planetary
Science Letters 126, 91–108.
Chivas, A.R., De Decker, P., Shelley, J.M.G., 1985. Strontium content
of ostracods indicates lacustrine paleosalinity. Nature 316, 251–
253.
Chivas, A.R., De Decker, P., Shelley, J.M.G., 1986. Magnesium
content of non-marine ostracod shells: a new paleosalinometer and
paleothermometer. Palaeogeography, Palaeoclimatology, Palaeo-
ecology 54, 43–61.
Coplen, T.B., 1988. Normalization of oxygen and hydrogen isotope
data. Chemical Geology (Isotope Geoscience Section) 72, 293–297.
Cowgill, U.M., 1969. The waters of Merom: a study of Lake Huleh. I.
Introduction and general stratigraphy of a 54 m core. Archive fur
Hydrobiology 66, 249–272.
Cowgill, U.M., 1973. The waters of Merom: a study of Lake Huleh. II.
The mineralogy of a 54 m core. Archive fur Hydrobiology 71, 421–
474.
Cowgill, U.M., 1994. The waters of Merom: a study of Lake Huleh.
IX. The minor chemical constituents of a 54 m core. Archive fur
Hydrobiology, Supplement-Band 99, 97–153.
De Deckker, P., 1979. The middle Pleistocene fauna of the West
Runton freshwater bed, Norfolk. Palaeontology 22, 293–316.
De Deckker, P., Forester, R.M., 1988. The use of ostracods to
reconstruct continental paleoenvironmental records. In: De Deck-
ker, P. et al. (Eds.), Ostracoda in the Earth Sciences. Elsevier
Science Publishers, Amsterdam, New York, pp. 175–199.
De Deckker, P., Chivas, R.A., Shelley, J.M.G., 1999. Uptake of Mg
and Sr in the euryhaline ostracod Cyprideis determined from in
vitro experiments. Palaeogeography, Palaeoclimatology, Palaeo-
ecology 48, 105–116.
Deuser, W.G., Hunt, J.M., 1969. Stable isotope ratios of dissolved
inorganic carbon in the Atlantic. Deep-Sea Research 16, 221–
225.
Ehrlich, A., 1973. Quaternary diatoms of the Hula Basin (northern
Israel). Israel Geology Survey Bulletin 58, 1–39.
Ehrlich, A., 1995. Atlas of the Inland-water Diatom Flora of Israel.
The Israel Academy of Sciences and Humanities, 166.
Elofson, O., 1969. Marine Ostracoda of Sweden, with special consid-
eration of Skagerrak. Translated from German (Zur kenntris der
marinen ostracoden Schwedens, mit besonder Berucksichtung
Skageraks, Zoologiska Bidray fran Uppsala, 1941) by Israel
Program for scientific translation, Jerusalem, p. 286.
Feibel, C.S., 2001. Archaeological sediments in lake margin environ-
ments. In: Stein, J.K., Farrand, W.R. (Eds.), Sediments in
Archaeological Context. University of Utah Press, Salt Lake City,
pp. 127–148.
Feibel, C.S., Goren-Inbar, N., Verosub, K.L., Saragusti, I., 1998.
Gesher Benot Ya’akov, Israel: New evidence for its stratigraphic
and sedimentologic context. Paleoanthropology Society, Annual
Meeting, Seatle. Journal of Human Evolution, 34, A7.
Gasse, F., Fontes, J.Ch., Plaziat, J.C., Carbonel, P., Kacsmarska, I.,
De Deckker, P., Soulie-Marsche, I., Callot, Y., Dupeuble, P.A.,
1987. Biological remains, geochemistry and table isotopes for the
reconstruction of environmental and hydrological changes in the
Holocene from north Sahara. Palaeogeography, Palaeoclimato-
logy, Palaeoecology 60, 1–46.
Goren-Inbar, N., Belitzky, S., 1989. Structural position of the
Pleistocene Gesher Benot Ya’aqov site in the Dead Sea rift zone.
Quaternary Research 31, 371–376.
Goren-Inbar, N., Belitzky, S., Verosub, K., Werker, E., Kislev, M.,
Heimann, A., Carmi, I., Rosenfeld, A., 1992. New discoveries at
the Middle Pleistocene Acheulian site of Gesher Benot Ya’aqov.
Quaternary Research 38, 117–128.
Goren-Inbar, N., Feibel, C.S., Verosub, K.I., Melamed, Y., Kislev,
M.E., Tchernov, E., Saragusti, I., 2000. Pleistocene milestones on
the out-of Africa corridor at Gesher Benot Ya’aqov, Israel. Science
289, 944–947.
Heiman, A., Mor, D., Steinitz, G., 1987. K–Ar dating of the Ramat
Korazim basalts, Dead Sea Rift. Israel Geological Society, Annual
Meeting, 1987, Mitzpe Ramon, Abstracts, pp. 54–56.
Henderson, P.A., 1990. Fresh water Ostracods. The Linenean Society
of London and The Estuarine and Coastal Science Association
Universal Book Services/Dr. W. Backhuys, Oegstgeest, The Nether-
lands. 228 pp.
Holmes, J.A., Horne, D.J. (Eds.), 1999. Non-marine ostracoda:
evolution and environment. In: Palaeogeography, Palaeoclimato-
logy, Palaeoecology, 148, p. 185. Special issue.
Horowitz, A., 1973. Development of the Hula Basin, Israel. Israel
Journal of Earth Sciences 22, 107–139.
Horowitz, A., 1979. The Quaternary of Israel. Academic Press,
New York. 394 pp.
Horowitz, A., 2001. The Jordan Rift Valley. A.A. Balkema Publishers,
Lisse, Tokyo. 730 pp.
Horowitz, A., Horowitz, M., 1985. Subsurface late Cenozoic palynos-
tratigraphy of the Hula Basin, Israel. Pollen et Spores 27, 365–
390.
Kafri, U., Lang, B., 1987. New data on the Late Quartenary fill of
the Hula Basin, Israel. Israel Journal of Earth Sciences 36, 73–
81.
Krammer, K., Lange-Bertalot, H., 1986. Bacillariophyceae, part:
Naviculaceae. In: Ettl, H., Gerloff, J., Heynig, H., Mollenhauer,
D. (Eds.), Susswasserflora vom Mitteleuropa. Gustav Fischer
Verlag, Jena. 876 pp.
Krammer, K., Lange-Bertalot, H., 1988. Bacillariophyceae, part 2:
Bacillariaceae, Epithemiaceae, Surrirellaceae. In: Ettl, H., Gerloff,
J., Heynig, H., Mollenhauer, D. (Eds.), Susswasserflora vom
Mitteleuropa. Gustav Fischer Verlag, Jena. 596 pp.
Krammer, K., Lange-Bertalot, H., 1991a. Bacillariophyceae, part 3:
Centrales, Fragilariaceae, Eunotiaceae. In: Ettl, H., Gerloff, J.,
Heynig, H., Mollenhauer, D. (Eds.), Susswasserflora vom Mitte-
leuropa. Gustav Fischer Verlag, Jena. 576 pp.
Krammer, K., Lange-Bertalot, H., 1991b. Bacillariophyceae, part 4:
achnanthaceae, kristische ergo-nzungen zu navicula (lineolatae)
und gomphonema gesamtliteraturverzeichnis. Teil 1-4. In: Ettl, H.,
Gerloff, J., Heynig, H., Mollenhauer, D. (Eds.), Susswasserflora
vom Mitteleuropa. Gustav Fischer Verlag, Jena. 437 pp.
Martens, K., Ortal, R., 1999. Diversity and zoogeography of inland
water ostracoda (Crustacea) in Israel (Levant). Israel Journal of
Zoology 45, 159–173.
McGregor, D.L., 1969. The reproductive potential, life history
and parasitism of the freshwater ostracod Darwinula Stevenstoni
(Brady and Robertson). In: Neale, N.W. (Ed.), The Taxonomy,
Morphology and Ecology of Recent Ostracoda. Oliver and Boyd,
Edinburgh.
Moshkovitz, S., Magaritz, M., 1987. Stratigraphy and isotope records
of Middle and Late Pleistocene mollusks from a continuous
corehole in the Hula Basin, northern Jordan Valley, Israel.
Quaternary Reseach 28, 226–237.
Neale, J.W., 1964. Some factors influencing the distribution of recent
British Ostracoda. Plubblicationi della Stazione Zoologica di
Napoli: 33 Supplemento, pp. 247–307.

A. Rosenfeld et al. / Journal of African Earth Sciences 38 (2004) 169–181 181
Racek, A.A., 1974. The study of Merom: A study of Lake Huleh. IV.
Spicular remains of freshwater sponges (Porifera). Archive fur
Hydrobiology 74 (2), 137–158.
Rosenfeld, A., Vesper, B., 1977. The variability of the sieve pores in
recent and fossil species of Cyprideis torosa (Jones, 1850), as an
indicator for salinity and paleosalinity. In: Loeffler, H., Danielopol,
D. (Eds.), Aspects of Ecology and Zoogeography of Recent and
Fossil Ostracoda. Dr. W Junk Publishers, The Hague, pp. 55–68.
Rosenthal, Y., Katz, A., Tchernov, E., 1989. The reconstruction of
Quarternary freshwater lakes from the chemical and isotopic
composition of Gastropod shells: the Dead Sea Rift, Israel.
Palaeogeography, Palaeoclimatology, Palaeoecology 74, 241–253.
Shackleton, N.J., 1974. Attainment of isotopic equilibrium between
ocean water and the benthonic foraminifera genus Uvigerina:
isotopic changes in the ocean during the last glacial. CNRS,
Colloque International 219, 203–209.
Shackleton, N.J., Vincent, E., 1978. Oxygen and carbon isotopes
studies in recent foraminifera from the SW Indian Ocean. Marine
Micropaleontology 3, 1–13.
Stekelis, M., 1960. The palaeolithic deposits of Jisr Banat Yaqub.
Bulletin of the Research Council of Israel 69, 61–87.
Stekelis, M., Picard, L., Bate, D.M.A., 1937. Jisr Banat Ya’qub.
Quarterly of the Department of Antiquities in Palestine 6, 214–215.
Tauxe, L., Herbert, T., Shackelton, N.J., Kok, Y.S., 1996. Astronom-
ical calibration of the Matuyama–Brunhes boundary: Conse-
quences for magnetic remanence acquisition in marine carbonates
and the Asian loess sequences. Earth Science Planetary Letters 140,
133–146.
Verosub, K.L., Goren-Inbar, N., Feibel, C.S., Saragusti, I., 1998.
Location of the Matayuma/Brunhes boundary in the Gesher Benot
Ya’aqov archaeological site, Israel. Paleoanthropology Society
Annual Meeting, Seatle. Journal of Human Evolution, 34, A22.
Verosub, K.L., Goren-Inbar, N., Feibel, C.S., Saragusti, I., 1999.
Magnetostratigraphic dating of the Gesher Benot Ya’aqov archae-
ological site, Jordan River Valley, Israel. XV INQUA Congress,
Durban, South Africa.
Von Graffenstein, U., Erlenkeuser, H., Trimborn, P., 1999. Oxygen
and carbon isotopes in modern fresh-water ostracod valves:
assessing vital offsets and autoecological effects of interest for
paleoclimate studies. Palaeogeography, Palaeoclimatology, Palaeo-
ecology 148, 133–152.
Wagner, C.W., 1957. Sur les Ostracodes du Quaternaire Recent des
Pays-Bas et leur Utilisation dans l’Etude Geologique des depots
Holocenes. Mouton &Co., �S-Gravenhage, 259 p.
Xia, J., Ito, E., Engstrom, D.R., 1997a. Geochemistry of ostracode
calcite: Part 1. An experimental determination of oxygen iso-
tope fractionation. Geochimica et Cosmochimica Acta 61, 377–
382.
Xia, J., Engstrom, D.R., Ito, E., 1997b. Geochemistry of ostracode
calcite: Part 2. The effects of water chemistry and seasonal
temperature variation on Candona rawsoni. Geochimica et Cosmo-
chimica Acta 61, 383–391.




















