
The relationship between water masses and benthic ostracod assemblages in the western Coral Sea,...
22
Palaeogeography, Palaeoclimatology, Palaeoecology, 105 (1993): 245-266 245 Elsevier Science Publishers B.V., Amsterdam The relationship between water masses and benthic ostracod assemblages in the western Coral Sea, Southwest Pacific Thierry Corr6ge t Department of Geology, The Australian National University, PO Box 4, Canberra, Australia (Received January 21, 1993; revised and accepted May 28, 1993) ABSTRACT Correge, T., 1993. The relationship between water masses and benthic ostracod assemblages in the western Coral Sea, Southwest Pacific. Palaeogeogr., Palaeoclimatol., Palaeoecol., 105: 245-266. The bathymetric distribution of 67 ostracod genera identified in 42 sediment samples (water depth range: 370 3776 m) from the western Coral Sea is documented. Statistical analyses of this dataset support preliminary observations that the vertical distribution of the ostracod fauna is largely controlled by the three water masses present in the studied area. Four ostracod assemblages are recognised: assemblage I, consisting mainly of ostracods present in the Deep Water; assemblage II. consisting of ubiquitous ostracods and ostracods mostly confined to the Antarctic Intermediate Water; and assemblages llI and IV, with ostracods characteristic of the Western South Pacific Central Water. Temperature, food supply and dissolved oxygen level are believed to be the most important factors influencing the ostracods bathymetric zonation. The present work es|ablishes the potential of ostracods for use in palaeoceanographic reconstruction in the western Pacific Ocean. Introduction Living organisms, whether they are aquatic or terrestrial, are strongly influenced by and adapted to the type of environment in which they occur. In the deep-sea (bathyal and abyssal zones), the distribution of epi-benthic organisms is a reflection of the conditions that prevail at the sediment/water interface, e.g. temperature, salinity, pressure, dis- solved oxygen and nutrient levels. These are all important factors that affect life on the sea-floor and most are also used to characterise the different water masses present in the world oceans. Thus, it is a logical assumption to expect a relationship between water masses and the structure and distribution of ecological communities. Indeed, extensive studies on benthic foraminifers have demonstrated that their bathymetric zonation is 1Present address: D6partement de G+ologie et Oc6anographie. Universit6 de Bordeaux I, Avenue des Facult6s, Talence Cedex 33405, France. controlled by water masses (e.g. Lohmann, 1978; Schnitker, 1980; Culver and Buzas, 1980, 1981; Culver, 1988; Corliss and Emerson, 1990). Unlike foraminifers, ostracods have received little atten- tion with respect to their relationship with water masses (e.g. Peypouquet and Benson, 1980; Benson et al., 1983; Cronin, 1984; Steineck et al., 1988; Dingle et al., 1989), and it is only recently that a synthesis of all these studies has been attempted for the Atlantic Ocean (Dingle and Lord, 1990). In the latter study, the emphasis was put on the abundance of common ostracod taxa to character- ise the different water masses. With the exception of some preliminary work conducted in the Japan area (Ishizaki and Irizuki, 1990; Zhou, 1993), all the previous work on ostracods and water masses have been from the Atlantic Ocean. Here, I present results from a semiquantitative and statistical analysis of the total ostracod genera collected from 42 bottom samples (water depth range: 370-3776 m) taken in the western Coral Sea (see Fig. 1). The aim of this work is to examine 0031-0182/93/$06.00 ~) 1993 - - Elsevier Science Publishers B.V. All rights reserved.
-
Upload
u-bordeaux -
Category
Documents
-
view
2 -
download
0
Transcript of The relationship between water masses and benthic ostracod assemblages in the western Coral Sea,...

Palaeogeography, Palaeoclimatology, Palaeoecology, 105 (1993): 245-266 245 Elsevier Science Publishers B.V., Amsterdam
The relationship between water masses and benthic ostracod assemblages in the western Coral Sea, Southwest Pacific
T h i e r r y C o r r 6 g e t
Department of Geology, The Australian National University, PO Box 4, Canberra, Australia
(Received January 21, 1993; revised and accepted May 28, 1993)
ABSTRACT
Correge, T., 1993. The relationship between water masses and benthic ostracod assemblages in the western Coral Sea, Southwest Pacific. Palaeogeogr., Palaeoclimatol., Palaeoecol., 105: 245-266.
The bathymetric distribution of 67 ostracod genera identified in 42 sediment samples (water depth range: 370 3776 m) from the western Coral Sea is documented. Statistical analyses of this dataset support preliminary observations that the vertical distribution of the ostracod fauna is largely controlled by the three water masses present in the studied area. Four ostracod assemblages are recognised: assemblage I, consisting mainly of ostracods present in the Deep Water; assemblage II. consisting of ubiquitous ostracods and ostracods mostly confined to the Antarctic Intermediate Water; and assemblages llI and IV, with ostracods characteristic of the Western South Pacific Central Water. Temperature, food supply and dissolved oxygen level are believed to be the most important factors influencing the ostracods bathymetric zonation. The present work es|ablishes the potential of ostracods for use in palaeoceanographic reconstruction in the western Pacific Ocean.
Introduction
Living organisms, whether they are aquatic or terrestrial, are strongly influenced by and adapted to the type o f environment in which they occur. In the deep-sea (bathyal and abyssal zones), the distribution o f epi-benthic organisms is a reflection of the condit ions that prevail at the sediment/water interface, e.g. temperature, salinity, pressure, dis- solved oxygen and nutrient levels. These are all impor tan t factors that affect life on the sea-floor and most are also used to characterise the different water masses present in the world oceans. Thus, it is a logical assumpt ion to expect a relationship between water masses and the structure and distribution o f ecological communities. Indeed, extensive studies on benthic foraminifers have demonst ra ted that their bathymetr ic zonat ion is
1Present address: D6partement de G+ologie et Oc6anographie. Universit6 de Bordeaux I, Avenue des Facult6s, Talence Cedex 33405, France.
controlled by water masses (e.g. Lohmann , 1978; Schnitker, 1980; Culver and Buzas, 1980, 1981; Culver, 1988; Corliss and Emerson, 1990). Unlike foraminifers, ostracods have received little atten- tion with respect to their relationship with water masses (e.g. Peypouque t and Benson, 1980; Benson et al., 1983; Cronin, 1984; Steineck et al., 1988; Dingle et al., 1989), and it is only recently that a synthesis of all these studies has been at tempted for the Atlantic Ocean (Dingle and Lord, 1990). In the latter study, the emphasis was put on the abundance o f c o m m o n ostracod taxa to character- ise the different water masses. With the exception o f some preliminary work conducted in the Japan area (Ishizaki and Irizuki, 1990; Zhou, 1993), all the previous work on ostracods and water masses have been f rom the Atlantic Ocean.
Here, I present results f rom a semiquantitative and statistical analysis o f the total ostracod genera collected f rom 42 bo t t om samples (water depth range: 370-3776 m) taken in the western Coral Sea (see Fig. 1). The aim of this work is to examine
0031-0182/93/$06.00 ~) 1993 - - Elsevier Science Publishers B.V. All rights reserved.

246 T. CORREGE
10 0
110
.~7o
.18 °
,~eo
~4aOE
' GulfofPapu'a ' ~ ' ' ~ . . . . ........... ~ PAPUA I o~ ~,
; ' . .......... -!:-'.. ~ NEW G U I N E A ~ £
: : . . : , , . . . . : - ;~o o "... ---"~, ":~":-:-?:--::-:,:-v:~------~ . . . . " - -
'" i ,' ..... O i" "i ,).; PAPUAN ............ :::'"::;:'-'%?~ ...... ,.: ....... PLATEAU,!..... ....... "".~/ " .... PLATEAU '-' ........ O"-:_':!-:~:-'~--=-~'L-"~-_'-~3.'-.~ .......
,~",:': . . . . . : .......... . - ; . . - . ~ . " . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . :-:--:::-:.:.:.:-:::::-:::.::.-::-:.:;:.~?-'-':.-~,~.--.:~.,...
,~,~;!i.' . . . . . . . . . . . . . . :::-~6~-":~+ .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ::::::::::::::::::::::::::::::::::::::::::::::: .... ",.---:=o:-C.~
> ~i:~ . . . i' "-'°~-" ......... " .............. ~' ......... ',:,,"i,! " ' " ,'-'~ .. ". • . . . . . -"- ~" """
} ~ ":::':'\- ..... : ...... :: * ...... "'::, ", ,~e" CORAL SEA BAS N (
\ "'-::".:'-'-b:: 0 : "; < ~' ' "' ""-:'--::--? .~ ' " G~, ":" • '~-..: ""'-~ ..... :-". -':- "::'-----'"'," .......
L , : ~ , i.", ~, .... . .................... :'~--:::::::::!:-'.::-,:: ....... ,':: .....
~i oO.~.-.. ~ ....... ... ':--:---::... ...... " ~ " ~ ~ <~. ,.~ ',, QUEENSLAND ..... "::::.'-::'.'-: : - - : : : : : : : : : - - - -~
i,',~,, ~^ ~.c. o ; . . . . . . . . . . . . " ........ -:::!--":-:-:-:--:--:::"%T"
~ ...... . ~ . :, :~ i , , . . - - ~ . . . . . . , . : : ~ - ~"~,'~ .~ :" / 1 ', ~ "-- "." ~X "~ ~-",,, ............ " ........ <:: ':: :~ ............... i~.'"
" ~ '%: ...... :' '~ ....... ' : - ' ' ' '" i ' ~ " : : "" ' ...... ':':.'., "., 9 " ..-!-" ,- ...... '- .-.
"~ '~:, " .... ~::" PLATEAU ': "-'--.\ _-/ / "; ",: '.Ca ,:. ,'--,, - ........ -'" .--+~" .-. -"
~, ".:~. o ~_ ,. ,.. ...... ..' ~ , O :grab e ',.,, '~'w'.'" (" ~0 /" "" ..... "" '''-'"
• . . . . . / ":::'": (~ .... "::"-::-.. ::' ~ ............. ...o:;000"-J";~ ...... / ' -"J'
O coretop L%"~ ~ ""~-:'-::'::: ...... "TO-WNSVILLE .... "~ ~ ........ ::: .................. ................. !.: . . . . . .
2~ ~" " ..................... ,, ~ ",.:
Fig. 1. Bathymetric map of the western Coral Sea, with the sample locations.
the possible relationship between ostracod and water mass distribution, and to assess the potential use of ostracods as tracers of past oceanographic conditions. A slightly different approach was taken from that of Dingle and Lord (1990). Here, semi- quantitative methods are used, and equal emphasis is placed on each taxon.
Oceanography and geology of the Coral Sea
Oceanography
Data on the physical oceanography of the west- ern Coral Sea are sparse. The most important sources are summarised in Pickard et al. (1977),
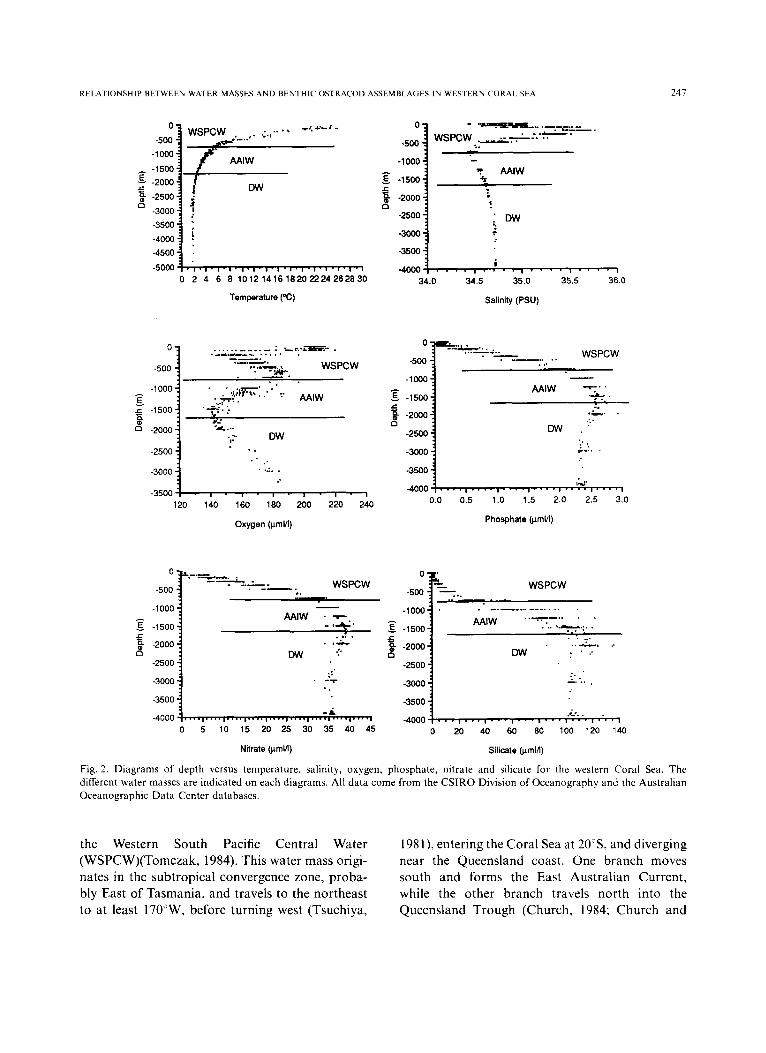
Emery and Meincke (1986), Tomczak (1984), and Tomczak and Hao (1989). Recent cruises con- ducted by the Australian Institute of Marine Sciences and the CSIRO Division of Oceanography have yielded valuable information, but very few results have been published to date. Nevertheless, raw data are available from the CSIRO Division of Oceanography database. Figure 2 shows the depth variation of six parame- ters: temperature, salinity, oxygen, phosphate, nitrate, and silicate. Several water masses can be identified from the surface to the sea floor.
Underneath the mixed layer (in the first 150 m on average), down to approximately 800 m (the total range of the permanent thermocline), lies
R E L A T I O N S H I P B E T W E E N W A T E R M A S S E S A N D B E N T H I C O S T R A C O D A S S E M B L A G E S IN W E S T E R N C O R A L S E A 247
g
W S P C W . . . . . "~" ~ " t .
f " AA IW
DW
0 ¸
-500
-1000
- 1 5 0 0
-2000 1
-2500 ; d
- 3000 I t
-3500" [ -4000"
-4500-
" 50~ " l ' l ' U ' l ' U ' l ' l ' l ' i ' U ' l ' l ' l ' l ' l
0 2 4 6 8 1 0 1 2 1 4 1 6 1 8 20 ~ 24 26 28 30
T e m p e r a t u r e (°C)
0 "
~ o o
-lOOO
, E - 1 5 o o .
2ooo~ 1:3
-2500'
- 3 0 0 0 '
- 3 5 0 0 '
-4000 34.0
" " = = = = P m ~ l l i _ _ , . . ~ - ¢ . . . . . . .
W S P C W . . . . . . . . " " ' ..2"
I". AA IW
DW | ,
i . . . . i . . . . i . . . . i . . . . i
34.5 35.0 35.5 36.0
Salinity (PSU)
0 "
-500
-1000 g
-1500" r~ a -2000.
-25OO,
-3000.
-3500 120
• . L . . , , . ,
. . . . • - -~.:~..,~.,. W S P C W
." .'..:if'}':~',i "" : AA IW " , • ' ~ ; -
"-~. . :.
" . i ¢ , . -
-,:'. DW
g
• : . : . .
i l • | - =
140 160 180 200
O x y g e n (pml/1)
• ! • i
220 240
0 - =~ - .
-500
-1000
E -1500'
2®o! -2500 i
- 3000"
-35O0 -:.
-4000 0.0
• " " ~ - - " W S P C W
AAIW -- ' - ' " " • , ,B - - .
oo
..-•. DW
" : . . .
=:~=. . . . . i . . . . i . . . . i . . . . i . . . . i . . . . i
0.5 1.0 1.5 2.0 2.5 3.0
P h o s p h a t e ~ml/I)
0
-500
-lO00
E.~ -1500
~- -2000"
-2500"
-3000 •
-3500"
W S P C W ;•
AA IW • " , " -
DW "'
• . . p
. i .
A E
-4000 • . . , . . . . , . . . . , . . . . t . . . . , . . . . , . . . . , . . . . , . . . . , 0 5 10 15 20 25 30 35 40 45
a
0 •
-500
-1000
- 1 5 0 0 •
r; ~--- W S P C W
o . . | | - - - - - | |
AAIW " -
DW : ..
- .:.• -3000 . . . . •
- 3 5 0 0 -.:
-4000 . . . , • . . , • - • , • , - - • , - • - , • • ,
0 20 40 60 80 100 120 140
Nitrate Osml/l) Silicate (p.ml/l)
Fig. 2. Diagrams of depth versus temperature, salinity, oxygen, phosphate , nitrate and silicate for the western Coral Sea. The different water masses are indicated on each diagrams. All data come from the C S I R O Division of Oceanography and the Austral ian Oceanographic Data Center databases.
the Western South Pacific Central Water (WSPCW)(Tomczak, 1984)• This water mass origi- nates in the subtropical convergence zone, proba- bly East of Tasmania, and travels to the northeast to at least 170°W, before turning west (Tsuchiya,
1981), entering the Coral Sea at 20°S, and diverging near the Queensland coast. One branch moves south and forms the East Australian Current, while the other branch travels north into the Queensland Trough (Church, 1984; Church and

248 T. CORREGE
Boland, 1983). This water mass is characterised by its high salinity (decreasing with depth), its high oxygen content (increasing with depth), and its low levels of nutrients (e.g. phosphate, nitrate and silicate) (see Fig. 2 and Table 1).
The Antarctic Intermediate Water (AAIW) can be found between 800 and 1700 m (Tomczak, 1984). This water mass originates at the Subpolar Convergence (polar front), situated at 55-60°S. The AAIW travels 3300 km before reaching the Coral Sea, and in the process, its salinity increases by mixing with the WSPCW above and the Deep Water (DW) below (Tomczak, 1984). The flow pattern in the Queensland Trough, as indicated by Scully-Power (1973), Pickard et al. (1977) and Church and Boland (1983) is to the north. At the limit between the AAIW and the DW, at about 1700 m, a level of no motion and oxygen minimum has been postulated by several authors (Rotschi and Lemasson, 1967; Wyrtki, 1962). Figure 2 clearly shows the oxygen minimum level (approxi- mately 140 laml/1) at the base of the AAIW. At this level, the oxygen saturation is only 40%, compared to 50-60% above 1500 m and below 2000 m (Tomczak, 1984). A phosphate maximum layer has also been reported between 1500 m and 2000 m (Rochford 1960), and coincides with a nitrate maximum (see Fig. 2 and Table I).
The Deep Water (DW, below 1700 m) is poorly known, and there are few published data on its origin and behaviour in the Coral Sea Basin. According to Wyrtki (1961), the DW flows north- wards from the East Australian Basin, and over- flows into the Coral Sea Basin at a depth of 2850 m. The DW then continues into the Solomon
TABLE 1
Characteristics of the three water masses present in the studied area
WSPCW AAIW DW
Depth range (m) 150-800 Temperature (°C) 22-6 Salinity (PSU) 35.7-34.5 Oxygen (/~ml/1) 150-190 Phosphate (pml/1) 0.3-2.0 Nitrate (#ml/1) 5-30 Silicate (pml/1) 5-20
800-1700 > 1700 6-2 < 2 34.4-34.7 34.7 190-145 145-170 2.0-2.6 2.3 30-40 40-35 20-100 > 100
Basin through a 3400 m deep channel (Wyrtki, 1961). No data on the movement of the DW are available from the western Coral Sea. The DW is a cold water mass, with a fairly constant salinity and high oxygen and nutrient levels (Fig. 2 and Table 1).
It is important to note that there is no sharp boundary between the AAIW and the DW. The limit is put at 1700 m by Tomczak (1984), but other authors give different figures (e.g. 1500 m in Emery and Meincke, 1986). Therefore, there is probably a smooth and progressive transition between the two water masses in most of the western Coral Sea.
Sedimen tology
Most of the samples used in this study are from the Queensland Trough and the western part of the Queensland Plateau (Fig. 1). The nature of the sedimentation in this region reflects the tropical climate as well as proximity to the Australian continental shelf and the Great Barrier Reef (GBR) to the west (Gardner, 1970; Beiersdorf, 1989). Pelagic sedimentation consists mainly of coccol- iths, foraminifers and pteropods. Pteropods are not preserved in the sediment below approximately 1650 m water depth, which is the level of the Aragonite Compensation Depth in the Western Coral Sea (Corr6ge, unpublished data). Sporadic input from the GBR and the reefs that stud the western margin of the Queensland Plateau also contribute to the sedimentation in the Queensland Trough (Winterer, 1970; Mutter, 1977; Falvey and Taylor, 1974; Harris et al., 1990).
Material and methods
Material
A total of 33 core top samples and 9 grab samples have been analysed for their ostracod content. These samples were collected from the western Coral Sea, and in particular the Queensland Trough area (see Fig. 1). The cores were collected by different institutions between 1967 and 1990, whereas the grab samples were collected by the author during two cruises in July
RELATIONSHIP BETWEEN WATER MASSES AND BENTHIC OSTRACOD ASSEMBLAGES IN WESTERN CORAL SEA 249
1990 and September 1991 on the R/V Franklin (see Table 2). For this study, the ostracods were iden- tified to generic level. Species identification was not attempted for two reasons. Firstly, the number of adult specimens is limited in each sample, and it is often difficult to assign species names to juveniles. Secondly, a database at the species level
TABLE 2
Location and water depth of the 42 samples
Latitude Longitude Water depth (S) (E) (m)
791/84 19°48.08 151°56.87 370 5 1 G C 9 15°41.49 145°49.67 381 791/63 17°58.69 149°34.54 494 791/102 11"24.51 152°28.00 562 590/07 16°20.70 146°06.60 706 690/23 11°29.45 144°07.19 792 690/29 8°44.28 145°34.59 950 ODP 822 16°25.379 146°12.904 955 VM24 160 18°07.00 147°36.00 1007 791/56 18°04.55 147°39.53 1076 590/08 16°18.40 146°09.20 1093 VM24 157 14"57.00 147°55.00 1212 590/09 16°18.20 146°11.20 1242 690/49 14°50.11 148°08.35 1260 VM24 182 17°31.00 147°30.00 1369 690/34 10058.29 145°59.92 1478 590/10A 16°15.50 146°15.00 1489 590/32 16°19.52 146°17.90 1490 5 1 G C 21 15°32.50 146°56.47 1630 590/31 16°15.60 146°24.20 1650 590/23 15°03.50 145°50.00 1670 VM24 172 14°41.00 146°49.00 1690 590/11 16°10.00 146°22.10 1690 590/13 16007.80 146°19.20 1701 VM24 158 15°55.00 146°51.00 1754 VM24 168 16°20.00 146°52.00 1785 590/12 16"05.90 146°30.00 1798 590/30 16°10.80 146°33.00 1798 590/29 15°42.30 145°59.60 1834 590/28 15°43.60 146°01.60 1850 590/15 15°04.10 146017.00 1875 590/22 15°02.30 145°56.60 1907 590/21 15°01.30 145°49.80 1982 690/12 1l°17.07 150°20.94 2023 5 1 G C 18 15°37.27 146°25.98 2060 VM24 183 15°20.00 146°15.00 2215 13 SL 14°45.63 146°17.98 2230 VM24 170 13°31.00 146°53.00 2243 590/17 15°02.70 146°09.30 2369 VM24 171 14°18.00 145°51.00 2714 VM24 184 12°52.00 146°12.00 2992 17SL 14°12.29 148°56.29 3776
would probably involve more than 200 species, rendering it difficult to manage and probably too detailed for my purpose. In addition, it appears that, with the exception of a few genera whose distribution spans a wide bathymetric range (e.g. Argilloecia, Krithe, Cytheropteron, Xestoleberis), each ostracod genus is represented in a relatively narrow depth interval. Results from foraminifer analyses show that the generic level is sufficient to characterise associations related to water masses~ and in some cases, even reduced generic data sets are used (Culver, 1988).
Methods"
Material from the grab samples was sieved onboard the ship through a 100 gm nylon mesh and preserved in ethanol. Rose bengal was added to the material to help reveal living or recently deceased material. In his deep-sea ostracod study, Van Harten (1990) made cautionary comments on the use of rose bengal, mostly because he found very few stained specimens in surface samples. In my material, the staining technique gave quite acceptable results: all the ostracods with soft parts were stained, making them easy to detect in the sediment. Moreover, some empty shells (or some- times with soft parts still remaining) displayed a strong staining. These latter shells were considered to have been recently alive, in contrast to shells that showed an irregular staining, possibly due to bacterial activity on them (Whatley and Wall, 1969). In each core, a sample was taken as close to the surface as possible, usually in the upper 5 cm. The samples were left for several days in diluted hydrogen peroxide (3% solution) to deflo- culate the clay fraction, then washed in a 100 gm sieve and dried in a low temperature oven (40°C). The ostracods were extracted, and their generic abundance recorded in a semiquantitative way as follows (Table 3): Abundance normalized to 5 g of dry raw sediment: -Absent (value 0 in the Table)
Present (value 1 in the Table): 1-5 valves - C o m m o n (value 2 in the Table): 6-25 valves
No genera were represented by more than 25 specimens per 5 g o f dry sediment. The use and advantage of semiquantitative data compared to

2 5 0 T. C O R R E G E
x4~
O~ t 'q
¢',1
¢q
t"-I II o . ,
tr~ t'~
¢q
r..--
r~
¢q
o~ t'-...-
O

RELATIONSHIP BETWEEN WATER MASSES AND BENTHIC OSTRACOD ASSEMBLAGES IN WESTERN CORAL SEA 251
~ E
E
~ E
- E
E
o E
E
.~ ~Z. ~ ~ ~ ~

252 T. CORREGE
,-.3 e ~
r.-..
¢"-I
t ' " -
~ E
t " q
t¢~ j
¢-q
• i ~
R E L A T I O N S H I P B E T W E E N W A T E R M A S S E S A N D B E N T H I C O S T R A C O D A S S E M B L A G E S I N W E S T E R N C O R A L S E A 253
~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ i ~
~ o ~ o ~ o o ~ _
~,-~ o.~
t--q
~ - c-q ¢,q t-q
E , . d , ~
¢~ t-q
~ E
t-'q t"q cq
tr~ ¢'-1
E
~ E
tr~
E

254 T. CORREGE
ud
r--.
~ " o'~ ¢ q t"q
t"..- t"q ¢ q
r - - E
t ,~ ¢-,I
~ E
t",l ¢ q
, . d ~ ,
¢ q ¢ q
t ' q
t"q
r"..-
¢ q
¢ q t"q
t" ' l ~ ¢ q
r'Z.
~ ~1" o"1 t",l

RELATIONSHIP BETWEEN WATER MASSES AND BENTHIC OSTRACOD ASSEMBLAGES IN WESTERN CORAL SEA 255
quantitative one in palaeoceanography is discussed by Sancetta (1979).
The dataset was examined to evaluate the poten- tial relationship between the ostracods and the water masses. The matrix (67 ostracod genera x 42 samples) was then subjected to statistical analyses.
Statistical analyses were performed using the GENSTAT package. Given the nature of the matrix, several multivariate analyses were available, including cluster analysis, principal coordinates analysis, and correspondence analysis (for a description of the different methods and their applications to ecological studies, see Digby and Kempton, 1987). The three methods were run, but only correspondence analysis and cluster analysis were judged to give relevant information, and only the results of these are shown here.
To assess the problem of downslope contamina- tion, the statistical analyses were performed on a reduced database of 31 samples (those with less than 20% of unknown fauna. Unknown taxa are mainly shallow ones, although some could also be present in deeper water). No significant difference was noted between the two sets of results
Results
Ostracod bathymetric distribution
Table 3 records the relative abundance of 67 ostracod genera in the 42 samples. With respect to water masses, 6 samples are situated in the Western South Pacific Central Water (WSPCW), 18 are in the Antarctic Intermediate Water (AAIW), and the remaining 18 are in the Deep Water (DW).
Characteristic ostracod assemblages can be determined for each water mass using data from Table 3. Ubiquitous ostracods (i.e. those present in almost all the samples) are excluded from these assemblages except where they show marked differences in their abundance.
The WSPCW fauna (370-792 m) Although there are only 6 samples in the
WSPCW, some obvious trends emerge from the data. The ostracod fauna is dominated by abun- dant Bairdia s.l., Cytherella, Macrocypris, Polycope
and Pontocypris. Most of these genera are present in the AAIW, but their abundance decreases rapidly with depth, and they are absent or rare in the DW (see Table 3). Less common, but restricted to the WSPCW are Jonesia, Cytheralison, and Hanaiceratina. The rest of the fauna is character- ised by Australoecia, Maddocksella, Monoceratina, Paradoxostoma, and Sclerochilus.
The AAIW fauna (955-1701 m) The Antarctic Intermediate Water exhibits a
different association, with some ostracod genera apparently restricted to this water mass although the poor representation of samples in the WSPCW prevents definite conclusions about upper range limits. The AAIW assemblage comprises Danipusella, "Glyphidocythere" Ayress et al., 1993, Loxocauda, Philoneptunus (all those genera being restricted to AAIW). Other important taxa include Aratrocypris, Henryhowella, Legitimocythere, Microcythere, ?Paracytherois, Pedicythere, Pelecocythere, and Swainocythere. The latter are also present in the the DW, but in smaller numbers.
The DWfauna (1754 3776 m) The Deep Water assemblage is characterised
by Abyssobairdia, Abyssocvthere, Dutoitella, Poseidonamicus, Rugocythereis and Xylocythere.
The indication is therefore that some ostracod genera are confined to a certain depth range, controlled largely by water masses. To investigate further such relationships, statistical analyses were run on the dataset.
Statistical analyses
Correspondence analysis Correspondence analysis on 67 ostracod genera
and the 42 samples was carried out. From an ordination diagram of the species (score 1 versus score 2), four assemblages can be identified (Fig. 3 and Table 4).
Assemblage I: Fauna dominant in the DW and sometimes present in the bottom of the AAIW.
Assemblage II: AAIW and ubiquitous fauna. Assemblage III: Fauna dominant in the top of
the AAIW and the WSPCW. Assemblage IV: WSPCW fauna.

256 T. CORREGE
1 4 III (topAAIW ~ ~ - ~
1.0" I (DW and and WSPCW
0.6. W .... /,,,~ ~'~,~,
- 0.2. " .
4.6- ,~ii~::i:i:;~ ..... ,~ ~ 7 -1.0 ~ ; ~ : . ~i~;:i::':}~" ~
-1 .4 ' II (AAI ubiquitous)
-1.8 • , ' , • , • , - , • , " , " , " ' " ' " -2.2 -1.8 -1.4 -1.0 LO.6 -0.2 0.2 0.6 1.0 1.4 1.8 2.2
Score 1
Fig. 3. Plot of Score I versus Score 2 for the correspondence analysis run on the ostracod fauna. Compact oblique lines: Assemblage I; Dots: Assemblage II; Loose oblique lines: Assemblage III; Crossed lines: Assemblage IV. Note that 2 genera contained in Assemblage IV (Cytheralison and Hanaiceratina) are not shown on the diagram because of their low value for Score 2 (see Table 4).
Although the four assemblages are well defined, their respective fields are not well separated, indi- cating that most of the ostracod genera are not restricted to a single water mass. However, the correspondence analysis corroborates the observa- tion that there is a relationship between the ostra- cod assemblages and the parameters used to define the water masses.
Results from the correspondence analysis on the 42 samples emphasize the limitations of the present work. Plot of score 1 versus score 2 for the depths shows that the WSPCW is easily distinguished from the two other water masses (Fig. 4 and Table 5). Since the scores are a reflection of the ostracod content of each samples, it is possible to identify the WSPCW as a water mass with a specific ostracod fauna. For the AAIW and the DW, although there is a clear trend in the data, some samples are in a transition zone, centered around the boundary between the two water masses (at 1700 m). This boundary coincides with an oxygen minimum and a nutrient maximum (Fig. 2). Therefore, except for temperature, samples collected at equal distances on each side of the boundary will have fairly similar water parameters isee Fig. 2).
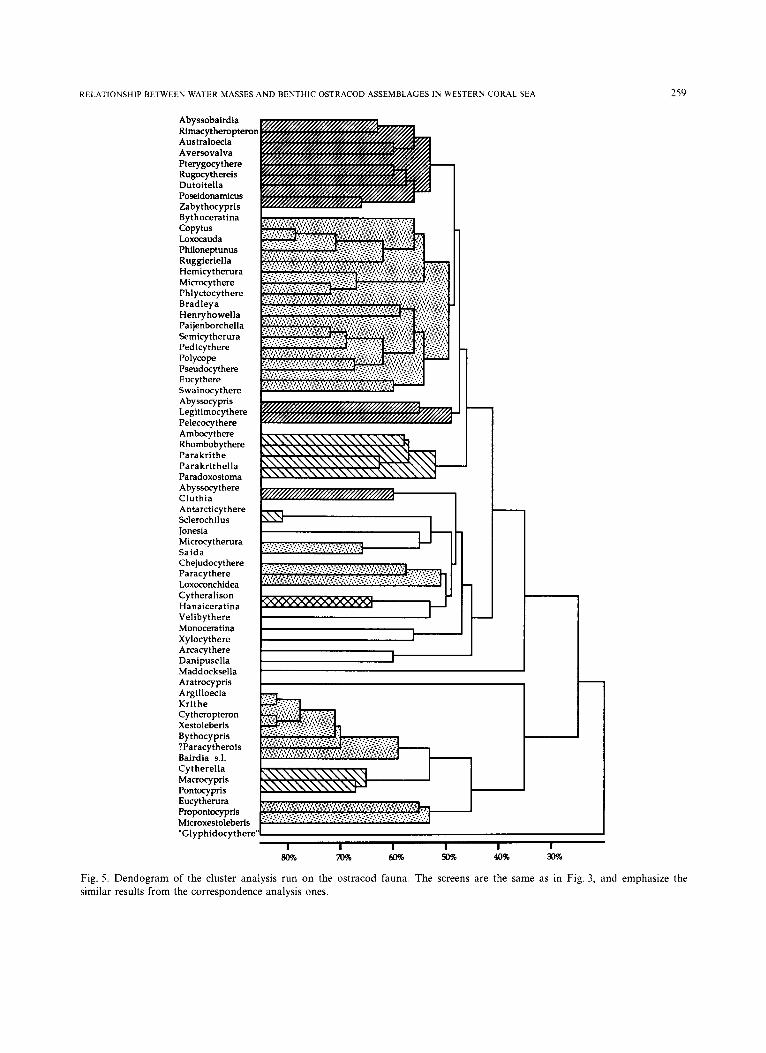
Cluster analysis The 67 ostracod genera were clustered according
to an ecological, quantitative method, without
taking into account zero-zero matches (i.e. absence of 2 ostracods in a sample is not significant com- pared to their presence; see Gower, 1971), and the results are shown in Fig 5. Due to the fact that zero-zero matches were excluded, the major clus- ters appear at fairly low percentages, which is not unexpected, given the high number of zeros (i.e. absence) in the matrix.
Argilloecia, Cytheropteron, Krithe and Xestoleberis have a similarity coefficient of 80%. These 4 genera constitute the background fauna, and are present in almost all the samples. Therefore, they cannot be used at the generic level as tracers of the water masses. The same is true of Bythocypris and ?Paracytherois, which both cluster with the previous group at the 70% level.
The rest of the fauna clusters into groups which are similar to the ones from the correspondence analysis. To help visualise this agreement between the two methods, the same patterns are used in Figs. 3, 4 and 5 to identify the four assemblages.
There are some discrepancies, however, between the two statistical analyses. Some ostracod genera which cluster together are found in different assem- blages (e.g. Monoceratina and Xylocythere). But their respective occurrences in the dataset suggests that it is the cluster analysis which is sometimes giving misleading information. Caution is therefore needed when interpreting results from statistical tests.
Discussion
The problem of downslope contamination
The majority of the ostracod valves encountered during this study were empty, although some ostra- cod carapaces, complete with soft parts were found in the grab samples. Nevertheless, the valves can still be used to assess the present-day bathymetric distribution of the ostracod fauna for several reasons. The sedimentation rate varies between 2.5 cm/kyr and 8 cm/kyr in the Queensland Trough (Gardner, 1970; ODP Leg 133 Initial Reports; Corr6ge, unpublished data), and bioturbation affects the top 7 cm of the sediment on average in the Western Pacific (Peng et al., 1979). As a result, ostracods collected in the top 1 cm of the sediment
RELATIONSHIP BETWEEN WATER MASSES AND BENTHIC OSTRACOD ASSEMBLAGES IN WESTERN CORAL SEA
T A B L E 4
Va lues f o r the correspondence analysis r u n o n the ostracod fauna
257
Correspondence Correspondence Correspondence Correspondence analysis analysis analysis analysis Score 1 Score 2 Score 1 Score 2
Assemblage 1 Abyssobairdia - 0 .7258
Abyssocythere - 1.2187
Aratrocypris - 0 .5867
Arcacythere - 0 .5589
Aversovalva 0.6897 Cluthia - 0 .818
Dutoitella - 0 ,1838
Legitimocythere - 0 .5344
Pelecocythere - 0.8731
Poseidonamicus 0,632
Pterygocythere - 0 . 3 9 4 1
Rimacytheropteron - 0 .364
Rugocythereis - 0 .6463
Xylo~ythere - 1.9743
Zabythocypris - 1.0086
Assemblage I1
Abyssocypris 0.259
Ambocythere 0.1903
Argilloecia - 0 .0376
Australoeda 0.2923
Bairdia s.1. 0 ,1562
Bradleya - 0 .2104
Bythoceratina 0.0905
Bythocypris - 0 .2192
Chejudocythere 0 .5884
Copytus 0.2049
Cytheropteron - 0 .0176
Danipusella 0.2503
Eucythere - 0 ,3577
Eucytherura - 0. 1766
"Glyphidocythere" - 0 . 8 0 1 3
Hemicytherura 0.0757
Henryhowella - 0 ,3348
Krithe - 0 .3004
Loxocauda 0.0915
Loxoconchidea 0.2144 0.4581
0 .1292 Microcythere 0.2803 0 .2886
- O. 1616 Microcytherura - 0 .0774 0 .2038
- O. 1111 Mjcroxestoktberis 0.2031 0 .1957
0 .0096 Paijenborchella 0.0332 0 .2307
- 0 .0565 Paracythere 0.0471 0 .0036
- 0 .2405 ?Paracytherois 0.0278 0 .0733
0 .3123 Parakrithe 0.3225 0 .4136
0 .0585 Pedicythere - 0 .035 0.211
- 0.2199 Philoneptunus 0.4615 0 .5892
0 .0414 Polycope 0.1002 - 0 .547
0 .2143 Pseudocythere 0.1584 - 0 .3683
O. 191 Ruggieriella 0.0863 0 .2177
0 .0918 Saida 0.1571 0 .1322
- 0 .4607 Semicytherura 0.1057 0.1351
- 0.1491 Swainocythere - 0 .359 0 .0723
Xes toleberis 0.1062 - 0 .1166
- 0.0004 Assemblage I I I
0 .0214 Antarcticythere 2.0042 1.2991
0.0871 Cytherella 0.4996 0 .1308
0 .5087 Macrocypris 0 .4656 0 .0702
- 0.1581 Maddocksella 1.2474 1.1343
- 0.1977 Monoceratina - 0 .0968 - 0 .7147
- 0.3967 Paradoxostorna 0.8351 0 .1012
0 .0488 Parakrithella 0.8555 0 ,5773
0 .6855 Phlyctocythere 0.662 0 .2207
0 .4318 Pontocypris 0.4253 - 0 .0478
- 0 .1592 Propontocypris 0.3921 0 .1705
0 .2069 Rhombobythere 0 .5824 0 .0537
- 0 .4702 Sclerochilus 1.8138 1.0023
0.167 Velibythere 0.5312 - 1,5839
- 0.9682 Assemblage IV 0.1303 Cytheralison 2.093 - 7.4 166
0 .0973 Hanaiceratina 1.8784 - 3 .2465
0 .1036 Jonesia 1.7866 - 0 .1759
0 .4563
could be up to 3000 years old. Since sea-level and also the temperature of the deep ocean has been stable for the past 6000-9000 years (Chappell and Shackleton, 1986; Shackleton, 1987; Labeyrie et al., 1989), it is reasonable to assume that the bathymetric distribution of the ostracod fauna studied herein has remained relatively unchanged for that period of time. Indeed, examination of the ostracod fauna throughout several cores (Corr6ge, 1993) shows no major change since the modern-day mode of circulation commenced in
the Western Pacific, 9500 yr B.P. (Labeyrie et al., 1989).
Nevertheless, downslope contamination, mainly through slumping and turbiditic processes, causes a problem that needs to be addressed when investi- gating depth zonation of benthic fauna (Van Harten, 1986; Drapala, 1993). In this study, most ostracod valves are opaque and appear similar in terms of preservation, making it difficult to ascer- tain which have been transported and which are in situ. Shallow ostracods are often sighted, a

258
1.0
0.8 ~
0.6"
0.4"
0.2"
~o 0.0" co -0 .2 " y,
-0 .4 " . ~.~:.:~!:
-0 .8"
-1.0 - . - . . . . . " . . . . . . . . . . -1.0 -0.8 -0.6 -0.4 -0.2 0.0 0.2 0.4 0.6 0.8 1.0
Score 1
[] DW LI AAIW 0 W S P C W
Fig. 4. Plot of Score I versus Score 2 for the correspondence analysis run on the 42 samples (see Table 5 for details). The patterns are the same as in Fig. 3, and reflect the relationship between the assemblages and the water masses. Note that one sample from the WSPCW is not shown on the diagram.
fea ture not found be low 600-800 m (Benson,
1975a). But a p a r t f rom obvious shal low sighted
os t racods , no definit ive conclus ion regard ing pos-
sible downs lope t r anspo r t can be d rawn for the
rest o f the fauna. Some genera which are to da te
only k n o w n f rom shal low env i ronments (e.g.
Danipusella, see Wouters , 1988; Jonesia, see Keij ,
1979, and t axonomic notes in append ix) look very
much in situ in the ba thya l env i ronmen t o f the
Cora l Sea, a l though no s ta ined valve be longing to
those t axa were found in the g rab samples. These
observa t ions suppo r t s imilar ones f rom the eastern
M i d - A t l a n t i c Ridge (Van Har ten , 1990), and show
tha t some taxa have a much wider ba thymet r i c
d i s t r ibu t ion than prev ious ly accepted.
The fact tha t s ta t is t ical analyses give s imilar
results on a reduced da tase t (wi thout the samples
showing obv ious con t amina t i on ) tends to indicate
tha t a l t hough present , downs lope c o n t a m i n a t i o n
is no t a m a j o r p r o b l e m in this s tudy.
Limitation of the present work
The size o f the da t abase (only 42 samples) can
account for some p rob lems identif ied in the present
work. I t is significant tha t 2 samples f rom the same dep th (590/12 and 590/30) have mode ra t e ly
different values for score 1 in the co r re spondence analysis . The downs lope c o n t a m i n a t i o n in one sample is p r o b a b l y respons ib le for this, but some o the r exp lana t ion might also be p roposed , e.g.
T. CORREGE
TABLE 5
Values for the correspondence analysis run on the samples
Correspondence Correspondence analysis analysis Score 1 Score 2
791/84 0.8611 - 2.7114 51GC9 0.8867 0.6772 791/63 0.5767 -0.5712 791/102 0.8805 -0.278 590/07 0.5895 0.1494 690/23 0.3524 -0.1293 690/29 -0.0384 -0.1171 ODP822 0.7624 0.2727 VM24160 0.0229 0.0272 791/56 0.0037 -0.1856 590/08 0.2563 0.2772 VM24157 -0.3296 -0.3539 590/09 0.2864 0.0921 690/49 -0.4228 -0.1377 VM24182 0.0899 0.0371 690/34 -0.197 0.0455 590/10A 0.2435 0.0916 590/32 0.2063 0.0774 51GC21 -0.4437 -0.0204 590/31 0.1492 0.1296 590/23 -0.1514 0.1844 VM24172 -0.3575 -0.1433 590/11 0.1641 0.2395 590/13 -0.2299 0.0035 VM24158 -0.3891 - 0.1147 VM24168 -0.3239 -0.1363 590/12 -0.2218 0.0631 590/30 0.1123 0.0495 590/29 -0.0528 0.1428 590/28 -0.0437 0.2451 590/15 -0.4436 -0.0817 590/22 -0.6137 -0.0034 590/21 0.0956 0.1147 690/12 -0.9588 -0.1299 51GC18 0.035 0.1882 VM24183 -0.2347 0.1228 13SL -0.6656 -0.2069 VM24170 -0.6962 -0.0803 590/17 -0.143 0.1713 VM24171 -0.091 0.0703 VM24184 -0.5019 -0.0561 17SL - 50.7302 0.2835
var iable wate r quali ty. Due to inadequa te knowl-
edge o f the oceanography , it is assumed tha t the b o u n d a r y between the water masses lies at the same dep th t h roughou t the entire Cora l Sea. This a s sumpt ion is p r o b a b l y unreal is t ic (see Wyr tk i ,
RELATIONSHIP BETWEEN WATER MASSES AND BENTHIC OSTRACOD ASSEMBLAGES IN WESTERN CORAL SEA 259
Abyssobairdia Rimacytheropteron Australoecia Aversovalva Pterygocythere Rugocythereis DutoiteUa Poseidonamicus Zabythocypris Bythoceratina Copytus Loxocauda Philoneptunus Ruggieriella Hemicytherura Microcythere Phlyctocythere Bradleya Henryhowella Paijenborchella Semicytherura Pedicythere Polycope Pseudocythere Eucythere Swainocythere Abyssocypris Legitimocythere Pelecocythere Ambocythere Rhombobythere Parakrithe Parakrithella Paradoxostoma Abyssocythere Cluthia Antarcticythere Sclerochilus Jonesia Microcytherura Saida Chejudocythere Paracythere Loxoconchidea Cytheralison Hanaiceratina Velibythere Monoceratina Xylocythere Arcacythere Danipusella Maddocksella Aratrocypris Argilloecia Krithe Cytheropteron Xestoleberis Bythocypris ?Paracytherois Bairdia s.1. Cytherella Macrocypris Pontocypris Eucytherura Propontocypris Microxestoleberis "Glyphidocythere
~,~-~//J////////i/////f/f////////////A ~--~I
I I I I l I 80% 70% ~ 50% 40% Fig. 5. Dendogram of the cluster analysis run on the ostracod fauna. The screens are the same as in Fig. 3, and emphasize the similar results from the correspondence analysis ones.

260 T. CORREGE
1961) and could account for some anomalies in the grouping of the samples.
The relatively restricted size of the dataset cannot provide a high resolution determination of the ostracods distribution, and some genera could therefore be overlooked as tracers of a specific water mass. For example, "Glyphidocythere" Ayress et al. is present in one sample, but examina- tion of numerous Late Pleistocene core samples from the Coral and Tasman Seas indicates that this genus has a very narrow bathymetric range and is restricted to the AAIW (Ayress et al., 1993).
The small size of the samples is most likely responsible for the clustering at low percentages of ostracods which obviously live at the same depth range. Most of the ostracods have a patchy distribution within a depth range, simply because they are scarce in the sediment, and the chance of encountering them in a core sample is small. On the other hand, two genera like Monoceratina and Xylocythere, which have a different bathymetric range (see Fig. 6), are clustered together because they occur together in one sample (out of two for Xylocythere)
Therefore, although the present database is large enough to provide an assessment of the bathymet- ric distribution of the ostracods, it could probably be improved by either increasing the size of the samples, or the number of samples.
Comparison between preliminary observations and statistical analyses
The ostracod assemblages identified for each water mass by examination of Table 3 are very similar to the four assemblages determined from the correspondence analysis. All the genera which were grouped in the DW fauna are included in Assemblage I. However, three genera from the AAIW fauna are also included in Assemblage I (e.g. Aratrocrypris, Legitimoeythere, and Pelecocythere; see Table 4), because of their pres- ence in the lower part of the AAIW. As pointed out earlier, Assemblage I contains ostracod genera mainly from the DW, but also from the lower part of the AAIW. Similarly, three genera which were identified as belonging to the WSPCW fauna (Bairdia, Polycope, and Australoecia) are found
in Assemblage II. These three genera occur in the AAIW, but are always more abundant in the WSPCW, which justifies their presence in the WSPCW fauna. The small number of samples from the WSPCW compared to the number of samples from the AAIW (6 and 18 respectively) is responsible for the appearance of these three genera in Assemblage II, because the correspon- dence analysis cannot take into account the samples number discrepancy between the two water masses.
It appears that, under certain conditions, corre- spondence analysis gives results very similar to those obtained from a visual analysis of the ostra- cod dataset. The main discrepancy between the two methods occurs when two water masses are represented in the dataset by markedly different numbers of samples, as it is the case for the WSPCW and the AAIW. Providing samples are evenly distributed within the water masses, corre- spondence analysis is a very effective method to determined ostracod assemblages related to water masses.
Factors affecting the bathymetric distribution of ostracods
Not all physical and chemical factors of the water masses are equal in their influence on the distribution range of ostracods.
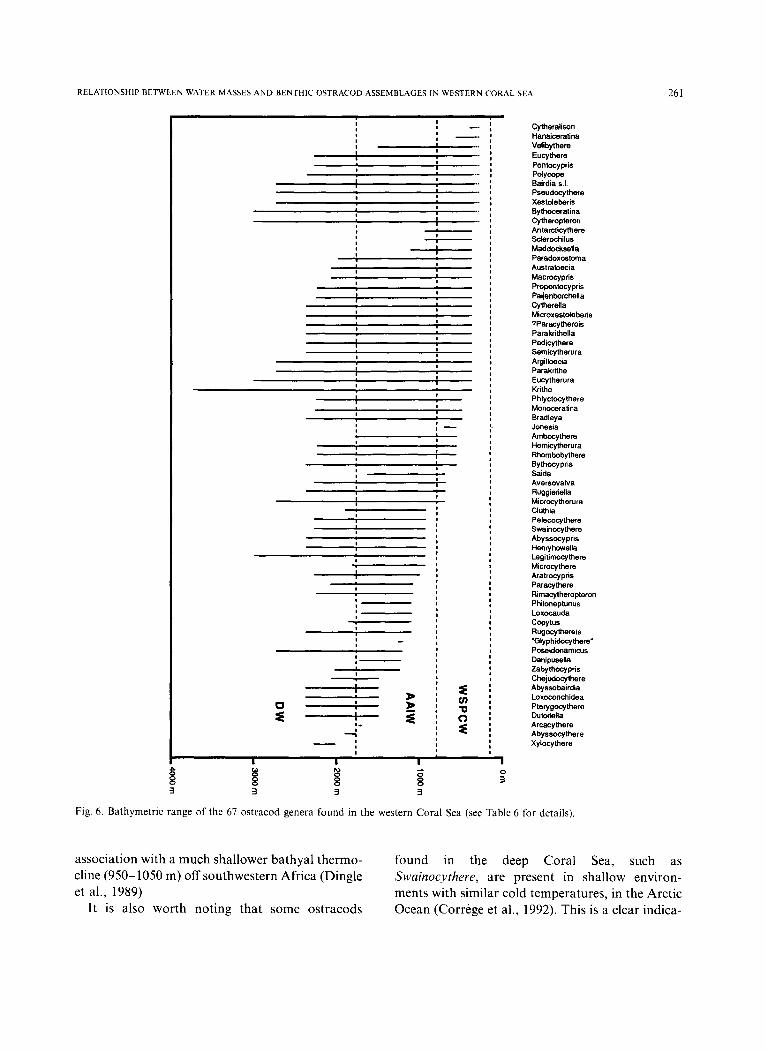
Temperature certainly has a major control on the composition of the ostracod fauna, as noted in previous studies (e.g. Benson, 1975b; Whatley, 1988). The permanent thermocline (150-800 m) is an area where many taxa have their Lower Depth Limit or Upper Depth Limit (see Table 6 and Fig. 6).
The generic abundance ostracod fluctuates between 22 and 34 in the samples shallower than 2215 m, then decreases rapidly below that depth (except for sample 590/17, which contains trans- ported fauna). This sudden drop in abundance below 2215 m can partly be attributed to low temperatures and decreasing food supply. The temperature versus depth curve flattens below 2200 m (bathyal thermocline), and there is only a change of about 0.2°C from 2200 m down to 4784 m (see Fig. 2). Similar faunal changes were recorded in
RELATIONSHIP BETWEEN WATER MASSES AND BENTHIC OSTRACOD ASSEMBLAGES IN WESTERN CORAL SEA 261
> o >
m.
o
I I I
8 8 8 8 3 3 3 3
" U
Cytheralison HanaiceratJna Velibythere Eucythere Pontocypris Polycope Bairdia s.I. Pseudocythere Xestoleberis Bythoceratina Cytheropteron Antarcticythere Sclerochilus Maddocksella Paradoxostoma Australoecia Macrocypris Propontocypris Paijenborchella Cytherella Microxestolebeds ?Paracytherois Parakrithella Pedicythere Sernicytherura Argilloecia Parakrithe Eucytherura Krithe Phlyctocythere Monoceratina Bradleya Jonesia Ambocythere Hernicytherura Rhombobythere Bythocypds Saida Aversovalva Ruggieriella Microcytherura Cluthia Pelecocythere Swainocythere Abyssocypris Henry howella Legitimocythere Microcythere Aratrocypris Paracythere Rimacytheropteron Philoneptunus Loxocauda Copytus Rugocythereis "Glyphidocythere" Poseidonamicus Danipusella Zabythocypris Chejudocythere Abyssobairdia Loxoconchidea Pterygocythere Dutoitella Arcacythere Abyssocythere Xylocy~ere
I
Fig. 6. Bathymetric range of the 67 ostracod genera found in the western Coral Sea (see Table 6 for details).
association w i t h a m u c h shallower bathyal thermo- found in the deep Coral Sea, such as cline (950-1050 m) off southwestern Africa (Dingle Swainocythere, are present in shallow environ- et al., 1989) merits with similar cold temperatures, in the Arctic
It is also worth noting that some ostracods Ocean (Corr6ge et al., 1992). This is a clear indica-
262
TABLE 6
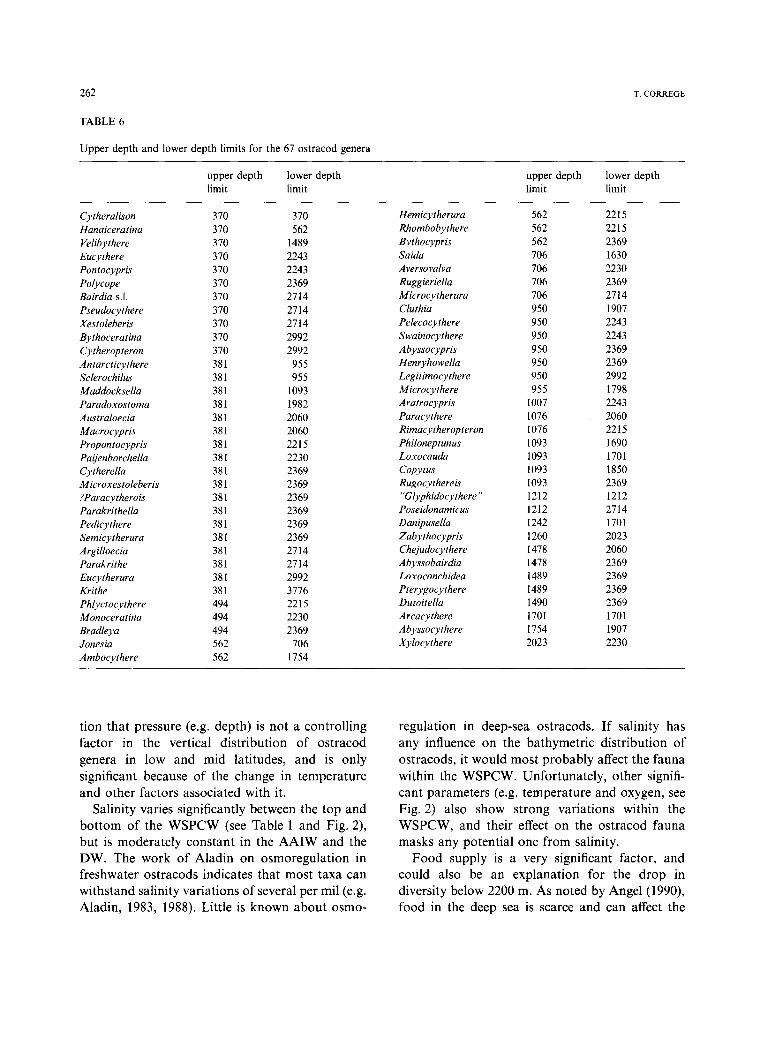
Upper depth and lower depth limits for the
T. CORREGE
67 ostracod genera
upper depth lower depth upper depth lower depth limit limit limit limit
Cytheralison 370 Hanaiceratina 370 Velibythere 370 Eucythere 370 Pon tocypris 370 Polycope 370 Bairdia s.1. 370 Pseudoeythere 370 Xestoleberis 370 Bythoeeratina 370 Cytheropteron 370 Antarcticythere 381 Sclerochilus 381 Maddocksella 381 Paradoxostoma 381 Australoeeia 381 Macrocypris 381 Propontocypris 381 Paijenborchella 381 Cytherella 381 Microxestoleberis 381 ?Paracytherois 381 Parakrithella 381 Pedieythere 381 Semieytherura 381 Argilloecia 381 Parakrithe 381 Eucytherura 381 Krithe 381 Phlyctocythere 494 Monoceratina 494 Bradleya 494 Jonesia 562 Ambocythere 562
370 Hemicytherura 562 2215 562 Rhombobythere 562 2215
1489 Bythocypris 562 2369 2243 Saida 706 1630 2243 Aversovalva 706 2230 2369 Ruggieriella 706 2369 2714 Microcytherura 706 2714 2714 Cluthia 950 1907 2714 Pelecocy there 950 2243 2992 Swainocythere 950 2243 2992 Abyssocypris 950 2369 955 Henryhowella 950 2369 955 Legitimocythere 950 2992
1093 Microcythere 955 1798 1982 Aratrocypris 1007 2243 2060 Paracythere 1076 2060 2060 Rimacytheropteron 1076 2215 2215 Philoneptunus 1093 1690 2230 Loxocauda 1093 1701 2369 Copytus 1093 1850 2369 Rugocythereis 1093 2369 2369 "G lyphidocyther e" 1212 1212 2369 Poseidonamicus 1212 2714 2369 Danipusella 1242 1701 2369 Zabythocypris 1260 2023 2714 Chejudocythere 1478 2060 2714 Abyssobairdia 1478 2369 2992 Loxoconchidea 1489 2369 3776 Pterygocythere 1489 2369 2215 Dutoitella 1490 2369 2230 Arcacythere 1701 1701 2369 Abyssocythere 1754 1907
706 Xylocythere 2023 2230 1754
t ion tha t pressure (e.g. depth) is not a cont ro l l ing fac tor in the vert ical d i s t r ibu t ion o f o s t r acod
genera in low and mid la t i tudes , and is only
significant because o f the change in t empera tu re and o ther factors associa ted with it.
Sal ini ty varies significantly between the top and b o t t o m o f the W S P C W (see Table 1 and Fig. 2), bu t is mode ra t e ly cons tan t in the A A I W and the D W . The work o f A lad in on osmoregu la t ion in
f reshwater os t racods indicates tha t mos t t axa can wi ths tand sal ini ty var ia t ions o f several per mil (e.g.
Alad in , 1983, 1988). Li t t le is k n o w n a b o u t osmo-
regula t ion in deep-sea os t racods . I f sal ini ty has
any influence on the ba thyme t r i c d i s t r ibu t ion o f
os t racods , it wou ld mos t p r o b a b l y affect the fauna
within the W S P C W . Unfor tuna te ly , o ther signifi- cant pa rame te r s (e.g. t empera tu re and oxygen, see
Fig. 2) also show s t rong var ia t ions within the W S P C W , and their effect on the os t racod fauna masks any po ten t ia l one f rom salinity.
F o o d supply is a very significant factor , and could also be an exp lana t ion for the d rop in divers i ty be low 2200 m. As no ted by Angel (1990),
food in the deep sea is scarce and can affect the

RELATIONSHIP BETWEEN WATER MASSES AND BENTHIC OSTRACOD ASSEMBLAGES IN WESTERN CORAL SEA 263
distribution of benthic organisms. Being crusta- ceans, ostracods cannot directly assimilate dis- solved nutrients. Deep-sea ostracods feeding habits remain largely unknown, but it is believed that most are scavengers (Van Morkhoven, 1962). The nutrient profiles (Fig. 2) all show depleted surface waters, and a maximum at about 1800 m (the approximate depth of the A A I W - D W boundary). However, there is almost no primary production in the deep-sea. Photosynthesis is non existant in the aphotic zone (below 500-600 m), and chemo- synthesis is always a local phenomenon, with small global significance (Rowe and Staresinic, 1979). Thus, the nutrient maximum level at 1800 m is probably indicative of a food shortage for ostra- cods. Table 6 and Fig. 6 show that 30% of the genera present in the AAIW disappear before or at 1800 1900 m.
Together with temperature and food supply, dissolved oxygen is probably the most influential factor. The oxygen minimum zone (OMZ) situated at the boundary between the Antarctic Inter- mediate Water and the Deep Water could act as a barrier for vertical migration of some of the fauna. However, this relatively weak OMZ (dissolved oxygen values not less than 140 laml/1) is seemingly responsible for the presence of the transition zone between the AAIW and the DW (Fig. 4). A stronger OMZ would no doubt induce a sharper turnover of the fauna as noted in other studies (e.g. Cronin, 1984; Zhou and Ikeya, 1992).
Conclusions
This study demonstrates that even in an area without marked differences between the water masses (at least between the AAIW and the DW), it is possible to associate ostracod assemblages with water masses. Statistical analyses show that the WSPCW is the water mass with the most distinctive fauna, whereas the AAIW and the DW are more difficult to differentiate on the grounds of their ostracod assemblages. Future studies will compare Late Quaternary associations retrieved from cores to the modern associations, and reconstruct the oceanographic changes which occurred in the western Coral Sea.
Acknowledgements
I would like to thank Patrick De Deckker (ANU) for his constant support on this project and for his review of this article. I also thank Michael Ayress (ANU) for help in the identifica- tion of the ostracod fauna and for his critical reading of an early draft, and Derek Burrage (AIMS) for allowing me to participate on the cruises on the R/V Franklin, and for many helpful discussions on the physical oceanography of the Coral Sea region. The captains and crew of the R/V Franklin made my two cruises a most enjoy- able experience, and I acknowledge their support with gratitude. Statistical analyses were performed with the help of Ross Cunningham and Christine Donnelly (Department of Statistics, ANU). I am indebted to David Jonhson (James Cook University), Peter Davies (University of Sydney) and the Lamont-Doherty Geological Observatory for providing the core samples. Finally, I thank Richard Dingle and Paul Steineck for their excel- lent review of this manuscript. This research was supported by an ANU scholarship.
Appendix
Taxonomic notes
Arcacythere Hornibrook 1952 Rockallia Whatley et al. 1978. has been synony-
raised by Ayress (1991).
"Glyphidocythere" Ayress et al. (1993) Refers to a new paradoxostomatid genus based
on Pseudocythere funafutiensis Chapman 1910 (Plate I, 7, 8).
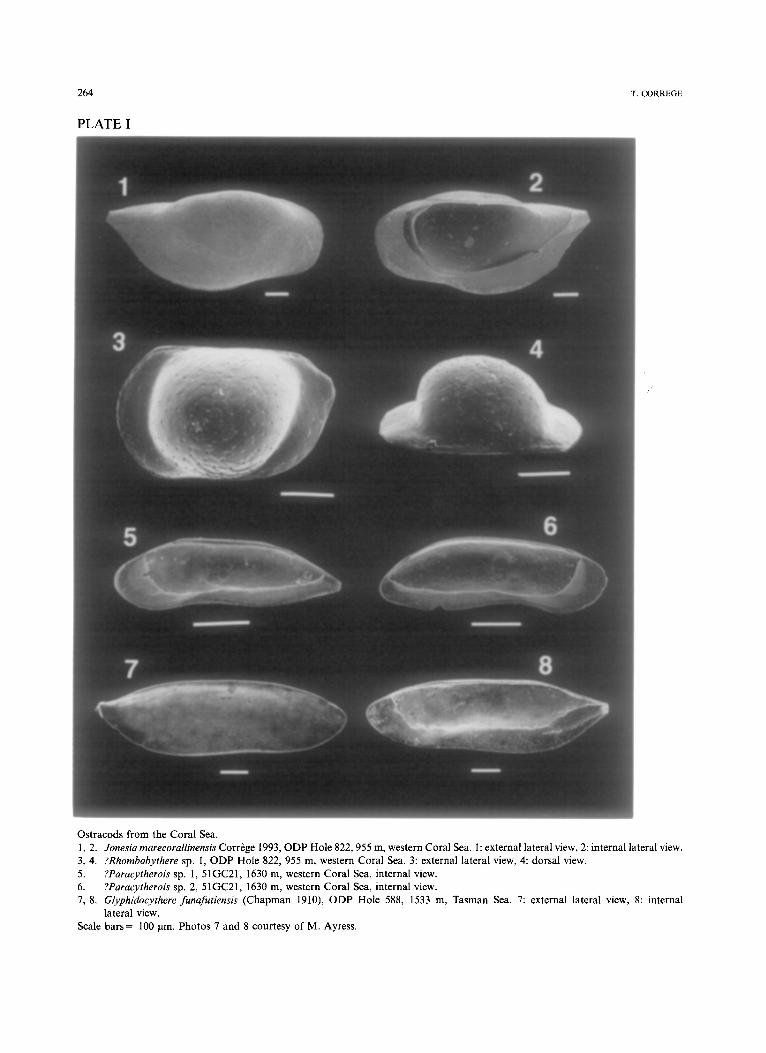
Jonesia Brady 1866 Represented by one species, Jonesia marecoralli-
nensis Corr6ge 1992. This species is closest to species previously refered to Baltraella Pokorny 1968. Following Schornikov, Baltraella is consid- ered to be synonymous with Jonesia (Plate I, 1, 2).
?Paracytherois Muller 1894 The genus recorded here is tentatively assigned
to Paracytherois, although it most probably belongs to a new genus. It has also been found by
264
P L A T E I
T. CORREGE
Ostracods from the Coral Sea. 1, 2. Jonesia marecorallinensis Corr~ge 1993, ODP Hole 822, 955 m, western Coral Sea. 1: external lateral view, 2: internal lateral view. 3, 4. ?Rhombobythere sp. I, ODP Hole 822, 955 m, western Coral Sea. 3: external lateral view, 4: dorsal view. 5. ?Paracytherois sp. 1, 51GC21, 1630 m, western Coral Sea, internal view. 6. ?Paracytherois sp. 2, 51GC21, 1630 m, western Coral Sea, internal view. 7, 8. Glyphidocythere funafutiensis (Chapman 1910), ODP Hole 588, 1533 m, Tasman Sea. 7: external lateral view, 8: internal
lateral view. Scale bars = 100 Ixm. Photos 7 and 8 courtesy of M. Ayress.

RELATIONSHIP BETWEEN WATER MASSES AND BENTHIC OSTRACOD ASSEMBLAGES IN WESTERN CORAL SEA 265
Van H a r t e n (pers . c o m m . ) in the deep A t l a n t i c
O c e a n , w h o also cons ide r s it to b e l o n g to a n e w
genus . T h e s p e c i m e n desc r ibed by N e a l e (1967) as
Xiphichilus gracilis ( C h a p m a n ) 1915 be longs to this
genus (P la te I, 5, 6).
Pterygocythere Hil l 1955
Re fe r s to P. mucronalatum (Brady) 1880, wh ich
has been as s igned to v a r i o u s g e n e r a in the pas t
(see D i n g l e a n d L o r d , 1990, fo r a d iscuss ion) .
Pterygocythere is p r e f e r ed on the basis o f its
iden t ica l in t e rna l fea tures .
?Rhombobythere S c h o r n i k o v 1982
Refe r s to severa l species, t en t a t i ve ly p l aced in
Rhombobythere. S c h o r n i k o v (pers . c o m m . ) sug-
ges ted the e rec t ion o f a s u b - g e n u s to a c c o m o d a t e
these species, bu t this awai t s f u r t h e r s tudy. (Pla te
I, 3, 4)
Rugocythereis D i n g l e et al. 1990
Inc ludes Pennyella N e a l e 1974 a n d
"'Oxycythereis" B e n s o n 1974 (see D i n g l e a n d L o r d ,
1990, for a d iscuss ion) .
Semicytherura W a g n e r 1957
Inc ludes Mayburia Coles a n d W h a t l e y 1989,
wh ich has been s y n o n y m i s e d by Ayres s a n d
C o r r 6 g e (1992).
References
Aladin, N.V., 1983. Salinity adaptations and osmoregulation abilities of the Ostracoda from the Caspian and Aral Seas. Zool. Zh., 62(1): 51-57 (in Russian).
Aladin, N.V., 1988. Reproductive salinity adaptations and salinity dependent features of embryonic development in Ostracoda and Branchiopoda. Zool. Zh., 67(7): 974-982 (in Russian).
Angel, M.V., 1990. Food in the deep ocean. In: R.C. Whatley and C. Maybury (Editors), Ostracoda and Global Events. Chapman and Hall, London, pp. 273-285.
Ayress, M.A., 1991. On Arcacythere Hornibrook, 1952 (Cytheracea, Ostracoda, Crustacea), a senior synonym of Rockallia Whatley, Frame and Whittaker, 1978. J. Micropalaeontol., 10(2): 223-226.
Ayress, M.A. and Corr6ge, T., 1992. On Semicytherura pulchra (Coles and Whatley). In: The Stereo-Atlas of Ostracod Shells. Br. Micropalaeontol. Soc., London, 19(I), pp. 57 60.
Ayress, M.A., Corr~ge, T. and Whatley, R.C., 1993. Glyphidocythere, a new deep marine paradoxostomatid (Ostracoda) genus from the Quaternary and Recent of the Indo-Pacific. J. Micropalaeontol., submitted.
Beiersdorf, H., 1989. Provenance and accumulation rates of
Pliocene and Quaternary sediments from the western Coral Sea. Geol, Rundsch., 78(3): 987 998.
Benson, R.H., 1975a. Morphologic stability in Ostracoda. Bull. Am. Paleontol., 65(282): 13-15.
Benson, R.H., 1975b. The origin of the psychrosphere as recorded in Deep-Sea ostracode assemblages. Lethaia, 8: 69-83.
Benson, R.H., DelGrosso, R.M. and Steineck, P.L., 1983. Ostracode distribution and biofacies, Newfoundland conti- nental slope and rise. Micropaleontology, 29(4): 430-453.
Chappell, J. and Shackleton, N.J., 1986. Oxygen isotopes and sea level. Nature, 324: 137-140.
Church, J.A. and Boland, F.M., 1983. A permanent undercur- rent adjacent to the Great Barrier Reef. J. Phys. Oceanogr., 13:1747 1749.
Church, J.A., 1984. A review of circulation in the western Coral Sea and internal tides on the shelf slope adjacent to the Great Barrier Reef. In: J.T. Baker et al. (Editors), Proceedings of the Great Barrier Reef Conference, Townsville, 1983, pp. 415-420.
Corliss, B.H. and Emerson, S., 1990. Distribution of Rose Bengal stained deep-sea benthic foraminifera from the Nova Scotian continental margin and Gulf of Maine. Deep-Sea Res., 37(3): 381 400.
Corr6ge, T., 1992. On Jonesia marecorallinensis Corr~ge sp. nov. In: The Stereo-Atlas of Ostracod Shells, 19(2), pp. 103-106.
Corr6ge, T., 1993. The evolution of the ostracod fauna in the Queensland Trough (western Coral Sea) since the Last Glacial. In: K.G. McKenzie and P. Jones (Editors), Ostracoda in the Earth and Life Sciences. Balkema, Rotterdam.
Corr+ge, T., Ayress, M.A. and Drapala, V., 1992. On Swainocythere nanseni (Joy and Clark). In: Stereo-Atlas of Ostracod Shells, 19(2), pp. 107-110.
Cronin, T.M., 1984. Bathyal ostracodes from the Florida- Hatteras slope, the Straits of Florida, and the Blake Plateau. Mar. Micropaleontol., 8:89 119.
Culver, S.J., 1988. New foraminiferal depth zonation of the northwestern Gulf of Mexico. Palaios, 3:69 85.
Culver, S.J. and Buzas, M.A., 1980. Foraminifera: distribution of provinces in the western North Atlantic. Science, 209: 687-689.
Culver, S.J. and Buzas, M.A., 1981. Foraminifera: distribution of provinces in the Gulf of Mexico. Nature, 290: 328-329.
Digby, P.G.N. and Kempton, R.A., 1987. Multivariate Analysis of Ecological Communities. Chapman and Hall, London, 206 pp.
Dingle, R.V. and Lord, A.R., 1990. Benthic ostracods and deep water-masses in the Atlantic Ocean. Palaeogeogr., Palaeoclimatol., Palaeoecol., 80: 213-235.
Dingle, R.V., Lord, A.R. and Boomer, I.D., 1989. Ostracod faunas and water masses accross the continental margin off southwestern Africa. Mar. Geol., 88:323 328.
Drapala, V., 1993. The use of ostracods in detecting fine- grained turbidites in deep-sea cores. In: K.G. McKenzie and P. Jones (Editors), Ostracoda in the Earth and Life Sciences. Balkema, Rotterdam.
Emery, W.J. and Meincke, J., 1986. Global water masses: summary and review. Oceanol. Acta. 9(4): 383-391.

266 T. CORREGE
Falvey, D.A. and Taylor, L.W.H., 1974. Queensland Plateau and Coral Sea Basin structural and time stratigraphic pat- terns. Aust. Soc. Explor. Geophys. Bull., 5(4): 123-126.
Gardner, J.V., 1970. Submarine geology of the western Coral Sea. Geol. Soc. Am. Bull., 81: 2599-2614.
Gower, J.C., 1971. A general coefficient of similarity and some of its properties. Biometrics, 27: 857-872.
Harris, P.T., Davies, P.J. and Marshall, J.F., 1990. Late Quaternary sedimentation on the Great Barrier Reef conti- nental shelf and slope east of Townsville, Australia. Mar. Geol., 94: 55-77.
Ishizaki, K. and Irizuki, T., 1990. Distribution of bathyal ostracodes in sediments of Toyama Bay, Central Japan. Cour. Forschungsinst. Senckenberg, 123: 53-67.
Keij, A.J., 1979. On two new Baltraella species (Ostracoda, Bythocytheridae) from the western Indopacific. K. Ned. Akad. Wet. Proc. Ser. B, 82 (1): 33-44.
Labeyrie, L.D., Duplessy, J.C. and Shackleton, N.J., 1989. The major deglacial changes in deep water circulation occur in the Pacific Ocean at the end of the Younger Dryas. Eos, 70 (43): 1146.
Lohmann, G.P., 1978. Abyssal benthonic Foraminifera as hydrographic indicators in the western South Atlantic Ocean. J. Foraminiferal Res., 8 :6-34
Mutter, J.C., 1977. The Queensland Plateau. Bur. Miner. Resour. Aust. Bull., 179.
Neale, J.W., 1967. An ostracod fauna from Halley Bay, Coats Land, British Antarctic Territory. Br. Antarct. Surv. Sci. Rep., 58.
Peng, T.H., Broecker, W.W. and Berger, W.H., 1979. Rates of benthic mixing in deep-sea sediments as determined by radioactive tracers. Quat. Res., 1 I: 141-149.
Peypouquet, J.P. and Benson, R.H., 1980. Les ostracodes actuels des bassins du Cap et d'Angola: Distribution bathy- m&rique en fonction de l'hydrologie. Bull. Inst. G6ol. Bassin Aquitaine, 28: 5-12.
Pickard, G.L., Donguy, J.R., Henin, C. and Rougerie, F., 1977. A review of the physical oceanography of the Great Barrier Reef and the western Coral Sea. Aust. Inst. Mar. Sci. Monogr. Ser., 2
Rochford, D.J., 1960. Some aspects of the deep circulation of the Tasman and Coral Seas. Aust. J. Mar. Freshwater Res., 11: 166-181.
Rotschi, H. and Lemasson, L., 1967. Oceanography of the Coral and Tasman Seas. Oceanogr. Mar. Biol. Annu. Rev., 5: 49-97.
Rowe, G.T. and Staresinic, N., 1979. Sources of organic matter to the deep-sea benthos. Ambio Spec. Rep. R. Swed. Acad. Sci., Stockholm, 6: 19-23.
Sancetta, C., 1979. Use of semiquantitative microfossil data for paleoceanography. Geology, 7: 88-92.
Schnitker, D.,. 1980. Quaternary deep-sea benthic foraminifers and bottom water masses. Annu. Rev. Earth Planet. Sci., 8: 343-370.
Scully-Power, P.D., 1973. Coral Sea budgets in winter. Aust. J. Mar. Freshwater Res., 24: 203-215.
Shackleton, N.J., 1987. Oxygen isotopes, ice volume and sea level. Quat. Sci. Rev., 6: 183-190.
Steineck, P.L., Dehler, D., Hoose, E.M. and McCalla, D., 1988. Oligocene to Quaternary Ostracods of the Central Equatorial Pacific (Leg 85, DSDP-IPOD). In: T. Hanai, N. Ikeya and K. lshizaki (Editors), Evolutionary Biology of Ostracoda. Elsevier, Amsterdam, pp. 597-617.
Tomczak, M., 1984. Coral Sea water masses. In: J.T. Baker et al. (Editors), Proceedings of the Great Barrier Reef Conference, Townsville, 1983, pp. 461-466.
Tomczak, M. and Hao, D., 1989. Water masses in the thermo- cline of the Coral Sea. Deep-Sea Res., 36(10): 1503-1514.
Tsuchiya, M., 1981. The origin of the Pacific Equatorial 13 ° water. J. Phys. Oceanogr., 11: 794-812.
Van Harten, D., 1986. Use of ostracodes to recognize down- slope contamination in paleobathymetry and a preliminary reappraisal of the paleodepth of the Prasas Marls (Pliocene), Crete, Greece. Geology, 14: 856-859.
Van Harten, D., 1990. Modern abyssal ostracod faunas of the area in the North Atlantic and a comparison with the Mediterranean. In: R.C. Whatley and C. Maybury (Editors), Ostracoda and Global Events. Chapman and Hall, London, pp. 321 328.
Van Morkhoven, F.P.C.M., 1962. Post Paleozoic Ostracoda. Elsevier, Amsterdam, 1.
Whatley, R.C. 1988. Ostracoda and palaeogeography. In: P. De Deckker et al. (Editors), Ostracoda in the Earth Sciences. Elsevier, Amsterdam, pp. 103-123.
Whatley, R.C. and Wall, D.R., 1969. A preliminary account of the ecology and distribution of Recent Ostracoda in the southern Irish Sea. In: J. Neale (Editor), The Taxonomy, Morphology and Ecology of Recent Ostracoda. Oliver and Boyd, Edinburgh, pp. 268-298.
Winterer, E.L., 1970. Submarine valley systems around the Coral Sea Basin (Australia). Mar. Geol., 8: 229-244.
Wouters, K., 1988. Two interesting new marine interstitial Ostracoda (Crustacea) from the Comoros, with the descrip- tion of Danipussella gen. nov. Bull. Inst. R. Sci. Nat. Belg. Biol., 58: 85-93.
Wyrtki, K., 1961. The flow of water into the deep sea basins of the western South Pacific Ocean. Aust. J. Mar. Freshwater Res., 12: 1-16.
Wyrtki, K., 1962. The subsurface water masses in the western South Pacific Ocean. Aust. J. Mar. Freshwater Res., 13: 18-47.
Zhou, B., 1993. Distribution of recent ostracode faunas in the Pacific off southwest Japan. (abstract). In: K.G. McKenzie and P. Jones (Editors), Ostracoda in the Earth and Life Sciences. Balkema, Rotterdam.
Zhou, B. and Ikeya, N., 1992. Three species of Krithe (Crustacea: Ostracoda) from Suruga Bay, Central Japan. Trans. Proc. Palaeontol. Soc. Jap., N.S., 166:1097-1115.




















