
GM-CSF modulates pulmonary resistance to influenza A infection

The Journal of Immunology
The Inducible Tissue-Specific Expression of the HumanIL-3/GM-CSF Locus Is Controlled by a Complex Array ofDevelopmentally Regulated Enhancers
Euan W. Baxter,* Fabio Mirabella,*,1 Sarion R. Bowers,*,2 Sally R. James,*
Aude-Marine Bonavita,* Elisabeth Bertrand,* Ruslan Strogantsev,†,3 Abbas Hawwari,‡,4
Andrew G. Bert,‡ Andrea Gonzalez de Arce,* Adam G. West,† Constanze Bonifer,*,5 and
Peter N. Cockerill*
The closely linked human IL-3 and GM-CSF genes are tightly regulated and are expressed in activated T cells and mast cells. In this
study, we used transgenic mice to study the developmental regulation of this locus and to identify DNA elements required for its
correct activity in vivo. Because these two genes are separated by a CTCF-dependent insulator, and the GM-CSF gene is regulated
primarily by its own upstream enhancer, the main objective in this study was to identify regions of the locus required for correct
IL-3 gene expression. We initially found that the previously identified proximal upstream IL-3 enhancers were insufficient to
account for the in vivo activity of the IL-3 gene. However, an extended analysis of DNase I-hypersensitive sites (DHSs) spanning
the entire upstream IL-3 intergenic region revealed the existence of a complex cluster of both constitutive and inducible DHSs
spanning the 234- to 240-kb region. The tissue specificity of these DHSs mirrored the activity of the IL-3 gene, and included
a highly inducible cyclosporin A-sensitive enhancer at 237 kb that increased IL-3 promoter activity 40-fold. Significantly,
inclusion of this region enabled correct in vivo regulation of IL-3 gene expression in T cells, mast cells, and myeloid progenitor
cells. The Journal of Immunology, 2012, 189: 000–000.
The closely linked IL-3 and GM-CSF genes are tightlyregulated and are both expressed in a highly inducible andtissue-specific fashion (1). IL-3 and GM-CSF are closely
related cytokines that activate the same signaling pathways andhave similar functions (2, 3). However, they also have important
unique roles in vivo. IL-3 has more wide-ranging functions, as itregulates the proliferation, differentiation, activation, and survivalof hematopoietic progenitor cells, and it promotes the differenti-
ation of mast cells, eosinophils, basophils, neutrophils, monocytes,megakaryocytes, and erythroid cells (2, 3). Although IL-3 is notessential for adult hematopoiesis, it can function as a proin-
flammatory cytokine in vivo, and as a CSF in vitro. During em-bryogenesis, IL-3 is known to play an important role in mobilizingand amplifying the earliest hematopoietic stem cells (HSCs)
emerging from the endothelium of the developing aorta, and it hasbeen shown that IL-3 mutant embryos are deficient in HSCs (4).Within the lumen of the aorta-gonad-mesonephros region of the
embryo, IL-3 is expressed by cells adhering to the endothelialwall, but the identity of these cells remains unknown (4). IL-3 isalso required for expansion of hemangioblasts in the aorta-gonad-
mesonephros region at an even earlier stage of embryogenesis (5).GM-CSF functions primarily as a powerful proinflammatory cy-tokine (3, 6), acting more specifically on granulocyte-macrophagelineage cells and mediating some of the key actions of TNF-a (7).The IL-3, GM-CSF, and IL-5 genes all evolved from a common
ancestor and remain linked within a 1-Mb cluster of cytokine genes(8). The human IL-3 and GM-CSF genes remain just 10.5 kb apart
as a single compact locus, whereas the IL-5 gene lies 465 kbdownstream. Although they are coinduced upon activation in somecell types, such as Th2 cells and mast cells, they are differentially
expressed in other cell types, and are likely to have independentmechanisms of regulation. Furthermore, the IL-5 gene is func-tionally linked to the IL-4/IL-13 locus as part of a region known
as the Th2 cluster, which is regulated by a shared locus controlregion (LCR) (9, 10).Despite their close proximity within the same locus, the human
IL-3 and GM-CSF genes appear to be regulated independently ofeach other as two distinct genes. Although they are coexpressed in
*Leeds Institute of Molecular Medicine, University of Leeds, St. James’s UniversityHospital, Leeds LS9 7TF, United Kingdom; †Institute of Cancer Sciences, WesternInfirmary, University of Glasgow G11 6NT, Scotland, United Kingdom; and‡Centre for Cancer Biology, SA Pathology, Adelaide 5000, South Australia, Australia
1Current address: Institute of Cancer Research, Sutton, U.K.
2Current address: Department of Molecular and Cellular Biology, University of Con-necticut, Storrs, CT.
3Current address: Department of Physiology, University of Cambridge, Cambridge,U.K.
4Current address: Department of Genetics, King Faisal Specialist Hospital and Re-search Centre, Riyadh, Saudi Arabia.
5Current address: School of Cancer Sciences, Institute of Biomedical Research,University of Birmingham, Birmingham, U.K.
Received for publication July 31, 2012. Accepted for publication August 30, 2012.
This work was supported by the Association for International Cancer Research, theBiotechnology and Biological Sciences Research Council, Leukaemia and Lym-phoma Research, the Medical Research Council, the European Community, andthe National Health and Medical Research Council.
Address correspondence and reprint requests to Prof. Peter N. Cockerill at the currentaddress: School of Immunity and Infection, College of Medicine and Dentistry,Institute of Biomedical Research, University of Birmingham, Edgbaston, Birming-ham B15 2TT, U.K. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: BAC, bacterial artificial chromosome; ChIP, chro-matin immunoprecipitation; CML, chronic myeloid leukemia; CNSa, conserved non-coding sequence a; CsA, cyclosporin A; DHS, DNase I-hypersensitive site; HSC,hematopoietic stem cell; LCR, locus control region; MEF, mouse embryonic fibro-blast; MP, myeloid progenitor; PMA/I, PMA and calcium ionophore; TK, thymidinekinase; UTR, untranslated region.
Copyright� 2012 by The American Association of Immunologists, Inc. 0022-1767/12/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1201915
Published September 28, 2012, doi:10.4049/jimmunol.1201915 at U
niversity of Birm
ingham on Septem
ber 28, 2012http://jim
munol.org/
Dow
nloaded from

T cells and mast cells, just GM-CSF is expressed in many types of
nonhematopoietic cells such as endothelial and epithelial cells (2,3). This distinct expression pattern is made possible by a CTCF-dependent insulator located between the two genes (11). The hu-man IL-3 gene is associated with at least two upstream enhancerelements. The most significant of these is a conserved inducibleenhancer at 24.5 kb upstream that functions in both mast cells andT cells, and can therefore help to account for the inducible andtissue-specific regulation of IL-3 expression (1, 12). An additionalnonconserved T cell-specific enhancer of unknown function exists14 kb upstream of the IL-3 gene (1, 13, 14). However, there havenever been any in vivo studies performed to determine just whichregulatory elements in this locus are actually either necessary orsufficient for correct IL-3 gene expression in vivo.The independent expression of the human GM-CSF gene is
controlled by a conserved 23-kb enhancer that supports its effi-cient inducible expression in a transgenic mouse model containingjust a 10-kb segment of the human GM-CSF locus (1, 15–17). TheGM-CSF gene is also associated with additional highly conservedfar downstream sequences, one of which is known to function asa BRG1/NF-kB–dependent enhancer in the mouse, where it wasdefined as conserved noncoding sequence a (CNSa) (18). Thiselement is at +34 kb in the mouse and at +30 kb in the humangenome relative to the start of the GM-CSF gene.The IL-3 24.5- and 214-kb enhancers, and the GM-CSF23-kb
enhancer all form inducible DNase I-hypersensitive sites (DHSs)(1, 12, 14, 16). These enhancers are each activated via cooper-ation between kinase and calcium signaling pathways, which inT cells are linked to the TCR, and they each encompass bindingsites for the Ca2+-inducible transcription factor NFAT, whichsupports chromatin remodeling (1, 19). Induction of these DHSsis suppressed by cyclosporin A (CsA), which blocks the Ca2+-dependent induction of NFAT by inhibiting calcineurin (12, 14,16).IL-3 and GM-CSF can also be aberrantly expressed in myeloid
leukemia, where they function as autocrine growth factors (20). Forexample, IL-3 functions as a mediator of autocrine growth inchronic myeloid leukemia (CML) (21), but the mechanisms re-sponsible for its aberrant expression remain unknown. Additionalstudies are, therefore, needed to identify the significant in vivosources of IL-3, and the DNA elements underlying the normalinduction of activation of IL-3 expression in mature hematopoieticcells and in the developing hematopoietic system, as well as theaberrant induction of IL-3 expression by BCR-ABL signaling inearly myeloid progenitor cells in CML patients (21–23).In this study, we aimed to identify mechanisms controlling the
correct inducible and developmental regulation of the human IL-3gene. We initially found that the proximal 214- and 24.5-kb en-hancer regions were not sufficient to direct efficient IL-3 promoteractivation in transgenic mice. However, a more expansive analysisrevealed that a transgene incorporating a powerful 237-kb induc-ible enhancer was able to direct the correct pattern of inducible anddevelopmentally regulated IL-3 gene expression in vivo.
Materials and MethodsAll experiments involving animals or humans were authorized by researchethics committees.
Transgenic mice
The previously describedB38, C42, andD48 IL-3/GM-CSF transgenicmouselines contain one to six copies of a 130-kb AgeI DNA fragment of bacterialartificial chromosome (BAC) clone CTD2004C12 (24). Line E52 containsone copy of the same BAC fragment, but has a 15.5-kb deletion at the 39 end,terminating 21 kb downstream of the GM-CSF gene, and lacking the +30 kbregion homologous to mouse CNSa +34-kb enhancer element (18). Copy
number was estimated made by Southern blot hybridization of mouse andhuman DNA, with human IL-3 and GM-CSF and mouse CD19 gene probes.
Cell preparation and stimulation
Unless indicated otherwise, all cell stimulation was for 4 h with 20 ng/mlPMA and 2 mM calcium ionophore A23187 (PMA/I). Human cell linesJurkat T cells, HMC-1 mast cells, KG1a myeloblastic cells, and RajiB cells were all cultured, as previously described (25). HMC-1 mast cellswere obtained from J. Butterfield (Mayo Clinic, Rochester, MN) (26).
Mouse cells used in this study include the following: 1) freshly isolatedthymocytes obtained by passing a thymus through a cell strainer; 2) spleen-derived CD19+ve B cells purified on magnetic beads; 3) actively dividingT blast cells prepared from splenic T cells (24); 4) mast cells grown frombone marrow by long-term culture in IL-3 and stem cell factor (25); 5)myeloid progenitor (MP) cells and macrophages derived from fetal liver;and 6) mouse embryonic fibroblasts (MEFs) cultured from embryos. TheMP cells were grown from day 13.5 transgenic mouse fetal liver by cultureat starting density of 1 3 106 cells/ml for 5–8 d in IMDM supplementedwith 10% FCS, 150 mM monothioglycerol, 500 U/ml penicillin and strep-tomycin, 10 ng/ml mouse rIL-3, and 10 ng/ml recombinant mouse stem cellfactor. Nonadherent cells were moved into a new flask each time they wereexpanded so as to separate them from any emerging adherent macrophagesfrom the culture. We confirmed that these cells grew as bunches of roundnonadherent cells, stained for ERMP12 Abs, and were not granular, as isexpected forMP cells. In parallel, the adherent macrophages were maintainedin isolation from the MP cells in the presence of 10% L cell-conditionedmedia containing M-CSF until they reached confluence. The MEFs wereprepared from decapitated, eviscerated day 13.5 transgenic mouse embryosdispersed by digestion for several hours with 0.25% collagenase, followed byculture for 14 d in DMEM plus 5% FCS, splitting the cells each time theyreached confluence. The T lymphoblasts were prepared by culture with CD3/CD28 Ab beads for 2 d, followed by culture in IL-2 for an additional2 d without beads. Liver nuclei were prepared by homogenizing liver innuclei digestion buffer containing 0.3 M sucrose (25), passing it through a70 mM strainer, and centrifugation in an angle rotor at 18,500 3 g througha cushion of digestion buffer containing 1.8 M sucrose for 30 min at 4˚C.
mRNA analyses
mRNA was extracted from cells using TRIzol (Invitrogen), according tothe manufacturer’s instructions. cDNAwas generated from purified mRNAusing Moloney murine leukemia virus reverse transcriptase (Invitrogen),according to the manufacturer’s recommendation. Unless indicated oth-erwise, all the gene expression analyses were performed by quantitativereal-time PCR using SYBR Green I reagents (Applied Biosystems, FosterCity, CA) on an ABI Prism 7500. The data from each set of primers wereanalyzed against a standard curve made with a serial dilution of cDNAfrom PMA/I-stimulated T blast cells or mast cells. Levels of expression ofmRNA encoding both human and mouse GM-CSF and IL-3 were deter-mined relative to mouse GAPDH expression. Semiquantitative analysis ofmRNA was performed using 25 cycles of PCR.
Primers used for mRNA analyses were as follows. Human GM-CSF:forward, 59-CACTGCTGCTGAGATGAATGAAA-39; reverse, 59-GTCT-GTAGGCAGGTCGGCTC-39. Human IL-3: forward, 59-GGACTTCAA-CAACCTCAATGGG-39; reverse, 59-TTGAATGCCTCCAGGTTTGG-39.Mouse GM-CSF: forward, 59-ATGCCTGTCACGTTGAATGAAG-39; re-verse, 59-CAGATCGTTAAGGTGGACCATG-39. Mouse GAPDH: for-ward, 59-TGGTGAAGCAGGCATCTGAG-39; reverse, 59-TGTTGAAG-TCGCAGGAGACAAC-39. Because the sequences of the mouse and hu-man IL-3 and GM-CSF coding regions have diverged considerably, theseprimer sets do not cross-react between the human and mouse mRNAs (andthe human proteins do not recognize the mouse receptors).
DHS analysis
DNase I digestions and analyses were performed on either isolated nuclei orpermeabilized cells, as previously described (24, 25). Samples with optimalextents of DNase I digestion were selected for Southern blot hybridizationanalysis of DHSs using the strategies summarized below.
Probes based on previously published strategies for mapping DHSswere as follows: 1) DHSs upstream of the IL-3 gene210.1-kb BamHI sitewere probed with a 0.7-kb EcoRI/BamHI fragment of DNA (210.1 to210.8kb); 2) DHSs across the IL-3 promoter region were mapped from the26.6-kb SpeI site with a 0.5-kb SpeI/BamHI fragment of DNA; 3) DHSsacross a 9.4-kb region spanning the GM-CSF enhancer and promoter weremapped from an EcoRI site 6.8 kb upstream from the GM-CSF gene usinga 1.5-kb BamHI fragment of DNA as a probe (2); 4) DHSs spanning theinsulator downstream of the IL-3 gene were mapped from a BamHI siteusing a 1-kb BglII/BamHI fragment of DNA (12, 15).
2 DEVELOPMENTAL REGULATION OF THE IL-3/GM-CSF LOCUS
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

Primers used to amplify DNA segments as probes for mapping novelDHSs are described in Supplemental Table I.
Transient transfection assays
Transfection assays and luciferase assays were performed using the lu-ciferase reporter gene plasmid pXPG (27) containing either the 2559- to+50-bp segment of the human IL-3 promoter (pIL3H) or a 229-bp frag-ment of the thymidine kinase (TK) promoter, plus the indicated upstreamDNA fragments, as described previously (25), or a DNA segment spanningthe human IL-3 +4.5-kb DHS. All transfection assays were performed byelectroporation using 4.5 3 106 cells and 5 mg test plasmid. With theexception of Fig. 5D, in each assay cells were cotransfected with 1 mg ofan appropriate Renilla luciferase plasmid to control for transfection effi-ciency. Transfected cells were first cultured for 21 h to equilibrate and thenstimulated for 8 h with 20 ng/ml PMA and 1 or 2 mM A23187.
Construction of plasmids used in transfection assays
DNA fragments spanning the human IL-3 240-, 237-, 234-kb, and +4.5-kb DHSs were amplified from the BAC CTD2004C12 by PCR usingprimers with restriction enzyme sites added to allow cloning, as follows.IL-3 237-kb DHS with XhoI linkers (defined in this study as the X37element because of the XhoI sites): forward, 59-CCCGCTCGAGC-TAAATGCATTTCCCACC-39; reverse, 59-CCCGCTCGAGCTGAGGA-GTGGAGATTACAGG-39. IL-3 234-kb DHS with KpnI linkers: forward,59-GGGGGGTACCGGCCATCATAGAAGCAGAAGG-39; reverse, 59-G-GGGGGTACCTGAATCACAGAGTGCCCACC-39. IL-3 240-kb DHSwith BamHI linkers: forward, 59-CGCGGGATCCAGGGCTCAAAAC-CAACTATGC-39; reverse, 59-CGCGGGATCCGGTGTTTACACCTGC-TCAAGG-39. IL-3 +4.5-kb DHS with BglII and KpnI linkers: forward,59-TTCCCTAGATCTCGCACGTGACAGCTAGTC-39; reverse, 59-CTTGG-TTCTGGTACCAGTCTGGCTTTAAGCTGCAG-39. After amplificationand restriction enzyme digestion, DNA fragments were cloned into eitherpIL3H (12), containing the 2559- to +50-bp segment of the human IL-3promoter in pXPG; pTK229 (12), containing a 229-bp fragment of the TKpromoter in pXPG; or, in the case of the IL-3 +4.5-kb DHS, just the lu-ciferase vector pXPG (27). The combination of BamHI, XhoI, and KpnImade it possible to clone each of the 240-, 237-, and 234-kb DHSs into
pIL3H at the same time and maintain the natural order of these elements inthe genome relative to the IL-3 promoter. The combination of BglII andKpnI made it possible to clone the +4.5-kb DHS in the sense orientationrelative to the human IL-3 and GM-CSF genes in the human genome, andrelative to the luciferase gene in pXPG.
The additional derivatives of the 684-bp X37 segment (the 237-kbelement plus XhoI adapters) as used in Fig. 7B were as follows: 1) The123–684 fragment was generated by BamHI digestion, and the 1–305 and305–684 fragments were generated by PvuII digestion. 2) PCR-generatedsubfragments of the X37 segment were engineered using primers con-taining BglII and KpnI cloning sites, as follows. The 274-bp 84–357segment was created with 59-TTTATAGATCTCTGGGGAAGGGACATC-39 and 59-AATTAGGTACCGGCTAGGTTAGGAAGGAC-39. The 184-bp84–267 segment was created with 59-TTTATAGATCTCTGGGGAAG-GGACATC-39 and 59-AATTAGGTACCATGAGATGACACTGACTC-39.
Primers used for site-directed mutagenesis of X37 were as follows:GATA forward, 59-GGGTGTGGGATCCAGgcAACACAGGATGAGAGG-39 and reverse, 59-CCTCTCATCCTGTGTTgcCTGGATCCCACACCC-39;ETS-1#1 forward, 59-GGGATCCAGATAACACActATGAGAGGTGGG-AGG-39 and reverse, 59-CCTCCCACCTCTCATagTGTGTTATCTGGA-TCCC-39; ETS-1#2 forward, 59-GAAAGAAAGTTCAGGCAAAGct-ACATCTTGCAGGTCTTTCTC-39 and reverse, 59-GAGAAAGACCTG-CAAGATGTagCTTTGCCTGAACTTTCTTTC-39; EGR-1 forward, 59-CA-AACAACCTggtgaACAAAATCATATTG-39 and reverse, 59-CAATATGA-TTTTGTtcaccAGGTTGTTTG-39.
EMSAs and chromatin immunoprecipitation
EMSAs of the IL-3 237-kb enhancer EGR-1 site were performed, aspreviously described (28), and used the following primers: forward strand,59-AGGTCTTTCTCAAACAACCTCCCCCACAAAATCATATTGA-39; re-verse strand, 59-TCAATATGATTTTGTGGGGGAGGTTGTTTGAGAAA-GACCT-39.
EMSAs employed nuclear extracts from Jurkat T cells, cultured withand without stimulation for 4 h with 20 ng/ml PMA and 0.57 mM calciumionophore A23187. The supershift EMSAs included Abs against Sp1(Millipore; 17-601), Sp3 (Santa Cruz Biotechnology; sc644X), or EGR-1(Santa Cruz Biotechnology; sc110X).
FIGURE 1. Developmental regulation
of human IL-3/GM-CSF transgenes. (A)
Map of the 130-kb BAC transgene with
the intact genomic locus showing pre-
viously defined enhancers (E), the insu-
lator (Ins), and a region homologous to
the mouse CNSa enhancer. Lines B38,
C42, and D48 contain the complete 130-
kb transgene, and line E50 has a 15.5-kb
39 deletion, as indicated by the bracket
underneath. Underneath is the map of the
minimal IL-3/Enhancer transgenic construct
containing just the proximal human IL-3
gene elements plus the 214-kb enhancer
linked to a human CD4 reporter gene. (B)
Copy number determination of the 130-kb
transgenes based on segments of the human
IL-3 and GM-CSF genes, using the mouse
CD19 locus as an internal reference, and
human genomic DNA as a two-copy con-
trol. (C) Human IL-3 and GM-CSF mRNA
expression in stimulated transgenic mouse
T blast cells, expressed relative to expression
of the mouse IL-3 and GM-CSF genes,
corrected for copy number. (D) Hema-
topoietic differentiation pathway showing
the predicted sources of IL-3 and GM-CSF.
(E) Inducible expression of human GM-CSF
and IL-3 mRNA in line C42. Columns de-
pict mRNA expression in stimulated cells
relative to mouse GAPDH and corrected
for copy number. Fold induction relative to
nonstimulated cells is shown underneath.
The Journal of Immunology 3
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

EGR-1 chromatin immunoprecipitation (ChIP) assays of the human IL-3promoter in Jurkat cells were performed, as described (28), with enrichmentlevels normalized against a negative control within the human VEZF1gene body, using the following primers: IL-3 promoter forward, 59-GGTTGTGGGCACCTTGCT-39; IL-3 promoter reverse, 59-TCTGTCT-TGTTCTGGTCCTTCGT-39; VEZF forward, 59-GACAGCAGCCGAA-CTTCGTT-39; VEZF reverse, 59-TGGTGCCCGAGGAAGATG-39.
ResultsA 130-kb segment spanning the human IL-3/GM-CSF locussupports correct developmentally regulated and inducible IL-3and GM-CSF gene expression
Our initial aim was to determine whether the previously definedregulatory regions of the human IL-3 locus are sufficient to supportits correct inducible and tissue-specific expression in vivo. Wecreated eight independent mouse lines from a transgene containingthe IL-3 214- and 24.5-kb enhancers and promoter linked toa human CD4 reporter gene (Fig. 1A). However, this combinationof DNA elements was clearly insufficient for in vivo expression
because three lines displayed low-level variegated expression,whereas the remaining five lines showed no expression uponstimulation in T blast cells (Supplemental Fig. 1).To aid identification of additional essential IL-3 gene regula-
tory elements, we performed detailed analyses of a much largertransgene spanning the intact human IL-3/GM-CSF locus. Wecreated several lines of transgenic mice from a 130-kb AgeI ge-nomic DNA fragment (lines B38, C42, D48, and E50; Fig. 1A),which includes the entire upstream 49-kb intergenic region sepa-rating the IL-3 gene from the upstream ACSL6 gene. Lines B38,C42, and D48 also include sequences extending to 36 kb down-stream of the start of the GM-CSF gene, whereas line E50 hasa 15-kb 39 deletion and terminates 21 kb downstream. Transgeneactivity was assessed in actively proliferating T blast cells gen-erated by stimulating spleen T lymphocytes with CD3 and CD28Abs and then culturing them for several days with IL-2 in theabsence of stimulus. These cells were then restimulated withPMA/I to activate TCR signaling pathways and induce cytokine
FIGURE 2. Distribution of tissue-
specific and inducible DHSs in the IL-3/
GM-CSF locus. (A and B) Strategies
employed for mapping DHSs. The in-
direct end-labeling probes used to map
DHSs are marked as horizontal black
arrows. Brackets define the regions be-
tween specific restriction enzyme sites
covered by each probe. (A) Map of
the previously defined inducible DHSs
(vertical black arrows) and constitu-
tive DHSs (vertical gray arrows) located
within proximal regions of the human
IL-3 and GM-CSF genes, as defined in
T cells. The black boxes indicate the
previously defined conserved 24.5-kb
IL-3 and23-kb GM-CSF enhancers. (B)
UCSC genome browser view of con-
served regions upstream of the human
IL-3 gene (http://genome.cse.ucsc.edu),
with the strategies for mapping DHSs
shown underneath. The vertical arrows
indicate the three conserved regions that
were the principal targets for analyses of
DHSs and enhancer function. (C) Sum-
mary of DHS mapping data. Inducible
DHSs are black, stable DHSs are gray,
and weak DHSs are shown as thin arrows.
Levels of inducible IL-3 and GM-CSF
expression relative to GAPDH, taken
from Fig. 1E, are indicated at the left of
each row.
4 DEVELOPMENTAL REGULATION OF THE IL-3/GM-CSF LOCUS
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

gene expression. For all four lines, the IL-3 and GM-CSF geneswere efficiently induced in a copy number-dependent fashion (Fig.1C). The average level of expression was in each case slightlyhigher than the levels determined for the endogenous mouse IL-3and GM-CSF genes (24). Line E50, which lacks the CNSa 39 GM-CSF region, supported approximately the same level of IL-3 ex-pression as the other three lines, and a slightly lower level of GM-CSF expression, but this difference was not considered significant.To define the pattern and mechanisms of developmentally
regulated inducible human IL-3 and GM-CSF gene expressionduring different stages of hematopoietic differentiation, we usedtransgenic mouse line C42 as a model and prepared the followingcell types: 1) thymocytes as a model of immature T cells; 2) ac-tively dividing spleen-derived T blast cells; 3) nonadherent MPcells cultured from fetal liver; 4) mast cells cultured from thebone marrow; 5) adherent macrophages derived from MP cells;and 6) fibroblasts cultured from MEFs. The differentiation path-ways linking these cells are depicted in Fig. 1D. Within thisscheme, HSCs and MP cells potentially give rise to both macro-phages, expected to express just GM-CSF, and mast cells thatefficiently express both IL-3 and GM-CSF.Cells were stimulated with PMA/I, and human IL-3 and GM-
CSF gene mRNA expression was measured relative to mouseGAPDH mRNA (Fig. 1E). The level of induction relative tounstimulated cells is shown underneath this graph depicting levelsmeasured in stimulated cells. As anticipated, the IL-3 and GM-
CSF genes were efficiently induced in activated T blast cells andmast cells. The level of expression was substantially higher inmast cells than in T cells, possibly because of the following: 1)mast cells express GATA-2 in addition to T cell factors such asRunx1, NFAT, and AP-1, and 2) the IL-3/GM-CSF locus includesmany regulatory elements containing GATA motifs (1, 25). This isconsistent with our findings that the GM-CSF enhancer is moreactive, and GM-CSF expression is higher, in cells expressingGATA factors (25).MP cells also produced high levels of GM-CSF mRNA,
equivalent to T blast cells, and moderate levels of IL-3 mRNA. Inagreement with previous studies (3), IL-3 was not expressed inmacrophages or MEFs. However, macrophages and MEFs simi-larly produced barely detectable levels of GM-CSF mRNA, in theorder of 1000-fold lower than T blast cells. This was consistentwith our previous studies of primary CD11b+ myeloid cells de-rived directly from the spleens of GM-CSF transgenic mice, wherethe mouse and human GM-CSF genes were both expressed atsimilar low levels (25). Hence, our findings are inconsistent withthe prevailing dogma that macrophages and fibroblasts are assumedto be significant sources of GM-CSF.
Properties of DHSs located between the IL-3 and GM-CSFgenes
To both identify novel potential regulatory elements, and define thepattern of developmental and inducible regulation of the IL-3/GM-
FIGURE 3. DHS analysis of the human IL-3/
GM-CSF locus in transgenic line C42. (A–F)
Southern blot hybridization analysis of DHSs, as
summarized in Fig. 2C, using the probes defined in
Fig. 2A and 2B. DHSs were mapped before and
after stimulation for 4 h with PMA/I in line C42
mast cells, B cells, thymocytes (Thy), T blast cells
(T BI), MP cells, macrophages (MF), fibroblasts
(MEF), and liver. DHSs are marked by arrows plus
their distances in kb relative to the IL-3 gene
transcription start site.
The Journal of Immunology 5
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

CSF locus, we performed an exhaustive analysis of DHSs in lineC42 in the cell types employed above, both before and afterstimulation (Figs. 2–4), and after stimulation in the presence ofCsA (Supplemental Fig. 2). As additional nonexpressing controls,we included splenic B cells and liver nuclei. For each cell type, theoptimal DNase I-digested samples were selected from a series ofDNase I titrations, and the same set of samples was then used foreach subsequent analysis. This selection was based on the effi-ciency of DNase I digestion and PMA/I induction, as confirmed byexamining the intensities of the DHS bands detected across theubiquitous CTCF sites within the insulator downstream of the IL-3gene (Fig. 3A) and the inducible DHS within the GM-CSF en-hancer (Fig. 3B). DHSs were mapped using the strategies depictedin Fig. 2A. These assays demonstrated strong induction of the23-kb GM-CSF enhancer DHS in T blast cells and mast cells;somewhat weaker induction of this DHS in MP cells, macro-phages, and MEFs; and no induction in thymocytes or B cells thatdo not express GM-CSF.Within the insulator region, we also identified an inducible CsA-
resistant DHS at IL-3 +4.5-kb in GM-CSF–expressing cells inaddition to the ubiquitous DHSs (Supplemental Fig. 2A). Unlikethe other PMA/I-inducible DHSs in the IL-3/GM-CSF locus, thisDHS was not suppressed by CsA, which functions by blocking theCa2+/calcineurin-dependent induction of NFAT. Furthermore, thisDHS functioned as a noncoding RNA promoter that is transcribedtoward the +4.9-kb DHS and the GM-CSF gene (SupplementalFig. 2B). Interestingly, the +4.9-kb DHS, which encompasses alow-affinity CTCF site, was weakest in the cell types that havethe weakest induction of the +4.5-kb DHS and show the lowest
GM-CSF expression. This element may, therefore, function as achromatin opening element for the intergenic region separatingthe GM-CSF gene from the high-affinity CTCF sites within theinsulator.
IL-3 gene activation is associated with a cluster of farupstream DHSs
The above results suggested that the 130-kb BAC transgene isa reliable model to study the regulation of the IL-3/GM-CSF locus.To search for additional essential IL-3 gene regulatory elementswithin this transgene, we used the mapping strategies depicted inFigs. 2B and 4A to identify DHSs within the entire 39-kb regionbetween the 214-kb enhancer and the upstream ACSL6 gene.These assays identified a tightly regulated complex cluster ofnovel far upstream DHSs present only in cells capable of IL-3gene expression (summarized in Fig. 2C) and colocalizing withconserved noncoding sequences (Fig. 2B). This region includeda highly inducible DHS at 237.5 kb, which was strongest in mastcells and T blast cells, weak in MP cells, and absent in all othercell types (Fig. 3C). The237.5-kb–inducible DHS was flanked bystable T blast-specific DHSs located at 240.3 and 233.7 kb, andadditional inducible DHSs present in activated mast cells and Tblast cells at 240.6 and 234.0 kb, which were weak in stimulatedMP cells, and absent in other cell types (Fig. 3D, 3E). Parallelassays of the IL-3 proximal region identified tissue-specific DHSsat 24.1 and 24.5 kb in mast cells, T blast cells, and MP cells;a DHS at21.5 kb specifically in T blast cells; and inducible DHSsat the 24.5-kb enhancer and promoter in T blasts and mast cells(Fig. 3F). The IL-3 234.0-, 237-, and 240.6-kb–inducible DHSs
FIGURE 4. DHSs associated with a LI-
LINE repeat element upstream of the IL-3
gene. (A) UCSC browser view and map
spanning the conserved and repeat regions
upstream of the IL-3 gene (http://genome.
cse.ucsc.edu). Probes used for mapping
DHSs in (B) and (C) are indicated by hori-
zontal arrows, with locations of DHSs in-
dicated as vertical arrows. The 216.4-kb
DHS lies within the 59 UTR of the L1PA13
LINE, and includes motifs for factors such
as NFAT, AP-1, ETS, PU.1, and RUNX1,
similar to the enhancers in this locus. The
223.3-kb DHS lies just outside the LIPA13
LINE, within a coding region segment of
a separate incomplete L1 LINE repeat. The
223.3-kb DHS was also detected using
the 234-kb BamHI probe used in Fig. 3E,
which employed the same BamHI Southern
blot filter as the one depicted here in (C). (B)
Higher resolution mapping of the 223.3-kb
IL-3 DHS in stimulated C42 blast cells
probed from an upstream KpnI site, and
using internal EcoRI and EcoRV sites as
size markers. The sizes of these markers are
indicated on the right-hand side, and the
positions of each band relative to the IL-3
transcription start site are shown on the left.
(C) Mapping DHSs in transgenic tissues
across the repeat-region spanning the 214-
kb enhancer from a BamHI site at 210.3
kb. Shown underneath is an additional DHS
identified at 210.3 kb mapped in the same
DNase I-digested samples, but assayed in a
parallel analysis from a SpeI site at 28.8 kb.
6 DEVELOPMENTAL REGULATION OF THE IL-3/GM-CSF LOCUS
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

were each substantially suppressed by CsA, suggesting a NFAT-dependent mechanism of activation (Supplemental Fig. 2C).We also made the unexpected observation that a full-length LI-
LINE repeat element, embedded within a 25-kb cluster of repeatelements, was associated with inducible DHSs located at223.3 kbin mast cells and T blast cells, and 216.4 kb in mast cells, MPcells, and macrophages (Fig. 4). The 216.4-kb DHS was locatedwithin the 59 untranslated region (UTR), which represents thepromoter of the ancestral retrotransposon. An additional inducibleDHS was detected at 210.3 kb as a prominent DHS in mast cells,and a weaker DHS in MP cells and macrophages (Fig. 4C). The214-kb region, which functions as an enhancer in Jurkat andCEM T cells (12), was not detected as a DHS in any of the celltypes examined in this study.
The IL-3 237-kb DHS functions as a powerful inducibleenhancer
Overall, it was clear that the level of inducible IL-3 expression indifferent cell types correlated very closely with the presence ofa highly complex cluster of tissue-specific DHSs occupying muchof the upstream intergenic region. Because the pattern of inductionof the 237-kb IL-3 DHS closely paralleled the induction of thegene itself, we tested the function of this element as an inducibleenhancer of the IL-3 promoter in luciferase reporter gene assaysin Jurkat T cells stimulated with PMA/I. Parallel assays wereperformed with the adjacent 234- and 240-kb DHSs, and thepreviously defined 214- and 24.5-kb–inducible enhancers. Wefound that the 237-kb element functioned as an exceptionallypowerful enhancer, which increased IL-3 promoter activity by 40-fold (Fig. 5A). This was in the order of 20 times more powerfulthan the 214- and 24.5-kb IL-3 enhancers, and 10 times greaterthan the 23-kb GM-CSF enhancer (11). In contrast, the 234-and 240-kb DHSs contributed no enhancer activity, even whenin combination with the 237-kb enhancer (240 plus 237, and240 plus237 plus 234). The237-kb enhancer was equally activein the reverse orientation, whereas the 240-kb DHS remainedinactive when reversed (data not shown).Because the IL-3 promoter is itself both inducible and tissue
specific, the above assays are unable to assess either of theseparameters for the 237-kb enhancer. To find a model system inwhich we could test these properties, we first screened a panel ofhuman cell lines for the inducible 237-kb DHS. These includedJurkat T cells, HMC1 mast cells, and KG1a myeloid progenitorcells, where in each case the 237-kb DHS was inducible, and RajiB cells and 5637 epithelial cells, where this DHS could not beinduced (Fig. 5B). To assess both tissue specificity and induc-ibility of the 237-kb enhancer, it was assayed in plasmids con-taining the constitutively active TK promoter in parallel with thewell-defined SV40 enhancer, which was used as the referencepoint. The 237-kb enhancer showed the greatest activity in JurkatT cells, where it was twice as active as the SV40 enhancer, andincreased TK promoter activity by 25-fold (Fig. 5C). The 237-kbenhancer was also active, but to a lesser degree, in HMC1 andKG1a cells, where it increased promoter activity by 3- to 6-fold,whereas it was inactive in Raji B cells. The function of the 237-kb enhancer was also strictly inducible because it did not supportany significant increase in TK promoter activity in the absence ofstimulation (Fig. 5D).
FIGURE 5. Analysis of enhancers upstream of the IL-3 gene. (A)
Luciferase reporter gene assays in stimulated Jurkat T cells transiently
transfected with the luciferase plasmid pIL3H containing human IL-3
promoter alone or in conjunction with DNA segments containing the
indicated DHSs. For 237/240 and 234/237/240, the order of ele-
ments relative to the IL-3 promoter is as in the genome. In each case,
values represent the average of two independent constructs. (B) DHS
mapping of the IL-3 237-kb region as in Fig. 3C in human cell lines
before and after stimulation for 4 h with PMA/I. (C) Transient trans-
fection assays of a luciferase plasmid containing the TK promoter alone,
or with either the SV40 or the IL-3 237-kb enhancer in stimulated cell
lines. (D) Transient transfection assays of a luciferase plasmid con-
taining the TK promoter alone, or with the IL-3 237-kb enhancer, in
nonstimulated and stimulated Jurkat cells. In (A) and (C), cotransfected
Firefly luciferase activity is used to correct for transfection efficiency of
firefly luciferase. (D) Depicts just the Firefly luciferase data so as to make it
possible to show the level of induction of the TK promoter. Error bars
indicate SD.
The Journal of Immunology 7
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from
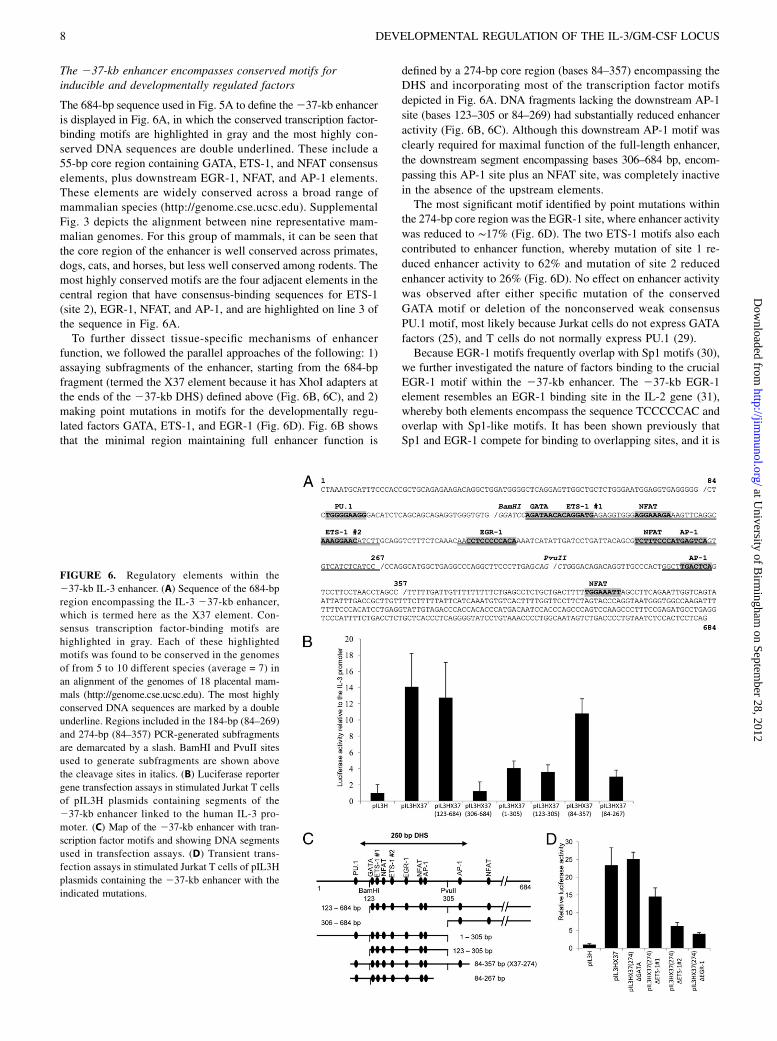
The 237-kb enhancer encompasses conserved motifs forinducible and developmentally regulated factors
The 684-bp sequence used in Fig. 5A to define the237-kb enhanceris displayed in Fig. 6A, in which the conserved transcription factor-binding motifs are highlighted in gray and the most highly con-served DNA sequences are double underlined. These include a55-bp core region containing GATA, ETS-1, and NFAT consensuselements, plus downstream EGR-1, NFAT, and AP-1 elements.These elements are widely conserved across a broad range ofmammalian species (http://genome.cse.ucsc.edu). SupplementalFig. 3 depicts the alignment between nine representative mam-malian genomes. For this group of mammals, it can be seen thatthe core region of the enhancer is well conserved across primates,dogs, cats, and horses, but less well conserved among rodents. Themost highly conserved motifs are the four adjacent elements in thecentral region that have consensus-binding sequences for ETS-1(site 2), EGR-1, NFAT, and AP-1, and are highlighted on line 3 ofthe sequence in Fig. 6A.To further dissect tissue-specific mechanisms of enhancer
function, we followed the parallel approaches of the following: 1)assaying subfragments of the enhancer, starting from the 684-bpfragment (termed the X37 element because it has XhoI adapters atthe ends of the 237-kb DHS) defined above (Fig. 6B, 6C), and 2)making point mutations in motifs for the developmentally regu-lated factors GATA, ETS-1, and EGR-1 (Fig. 6D). Fig. 6B showsthat the minimal region maintaining full enhancer function is
defined by a 274-bp core region (bases 84–357) encompassing theDHS and incorporating most of the transcription factor motifs
depicted in Fig. 6A. DNA fragments lacking the downstream AP-1
site (bases 123–305 or 84–269) had substantially reduced enhancer
activity (Fig. 6B, 6C). Although this downstream AP-1 motif was
clearly required for maximal function of the full-length enhancer,
the downstream segment encompassing bases 306–684 bp, encom-
passing this AP-1 site plus an NFAT site, was completely inactive
in the absence of the upstream elements.The most significant motif identified by point mutations within
the 274-bp core region was the EGR-1 site, where enhancer activity
was reduced to ∼17% (Fig. 6D). The two ETS-1 motifs also each
contributed to enhancer function, whereby mutation of site 1 re-
duced enhancer activity to 62% and mutation of site 2 reduced
enhancer activity to 26% (Fig. 6D). No effect on enhancer activity
was observed after either specific mutation of the conserved
GATA motif or deletion of the nonconserved weak consensus
PU.1 motif, most likely because Jurkat cells do not express GATA
factors (25), and T cells do not normally express PU.1 (29).Because EGR-1 motifs frequently overlap with Sp1 motifs (30),
we further investigated the nature of factors binding to the crucialEGR-1 motif within the 237-kb enhancer. The 237-kb EGR-1element resembles an EGR-1 binding site in the IL-2 gene (31),whereby both elements encompass the sequence TCCCCCAC andoverlap with Sp1-like motifs. It has been shown previously thatSp1 and EGR-1 compete for binding to overlapping sites, and it is
FIGURE 6. Regulatory elements within the
237-kb IL-3 enhancer. (A) Sequence of the 684-bp
region encompassing the IL-3 237-kb enhancer,
which is termed here as the X37 element. Con-
sensus transcription factor-binding motifs are
highlighted in gray. Each of these highlighted
motifs was found to be conserved in the genomes
of from 5 to 10 different species (average = 7) in
an alignment of the genomes of 18 placental mam-
mals (http://genome.cse.ucsc.edu). The most highly
conserved DNA sequences are marked by a double
underline. Regions included in the 184-bp (84–269)
and 274-bp (84–357) PCR-generated subfragments
are demarcated by a slash. BamHI and PvuII sites
used to generate subfragments are shown above
the cleavage sites in italics. (B) Luciferase reporter
gene transfection assays in stimulated Jurkat T cells
of pIL3H plasmids containing segments of the
237-kb enhancer linked to the human IL-3 pro-
moter. (C) Map of the 237-kb enhancer with tran-
scription factor motifs and showing DNA segments
used in transfection assays. (D) Transient trans-
fection assays in stimulated Jurkat T cells of pIL3H
plasmids containing the 237-kb enhancer with the
indicated mutations.
8 DEVELOPMENTAL REGULATION OF THE IL-3/GM-CSF LOCUS
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

proposed that EGR-1 displaces Sp1 to mediate activation of somePMA-inducible genes (32). To determine which of these factorsbind to the enhancer, we performed EMSA and ChIP analyses inPMA/I-stimulated Jurkat cells (Fig. 7A, 7B). In these studies, wealso confirmed induction of both IL-3 and GM-CSF mRNA ex-pression (Fig. 7C). EMSAs revealed that the 237-kb EGR-1 motifbound in vitro to both of the constitutively expressed factors Sp1and Sp3, and to the inducible factor EGR-1 (Fig. 7B). Note,however, that although Sp1 and EGR-1 can both bind in EMSAs,where the probe is in excess, these factors cannot bind simulta-neously to the same site in vivo. We used ChIP assays in Jurkatcells to confirm inducible binding of EGR-1 to the 237-kb en-hancer, but not the Sp1/EGR-1–like element in the IL-3 promoter(Fig. 7A). However, neither Sp1 nor Sp3 was found by ChIP tobind to the 237-kb enhancer in Jurkat cells (data not shown). Wesuggest that the lack of in vivo Sp1 binding prior to activation maybe due to lack of chromatin accessibility within the enhancer innonstimulated cells, whereas EGR-1 may outcompete Sp1 bindingafter activation once the DHS has formed.
DiscussionIndependent regulation of the IL-3/GM-CSF locus
This study establishes that a 130-kb region of the IL-3/GM-CSFlocus includes all the DNA elements that are required for itscorrect developmental and inducible regulation. It also establishesthat the IL-3/GM-CSF locus is controlled independently of thedownstream Th2 cytokine gene cluster encompassing the IL-4, IL-5, and IL-13 genes, which are coregulated as a separate unit by theTh2 LCR (9). This conclusion is further supported by chromatincross-linking (3C) assays that detected direct interactions withinthe nucleus between the Th2 LCR and the IL-4, IL-5, and IL-13genes, but not between the Th2 locus and the IL-3/GM-CSF locus(10). Furthermore, our data provide further evidence suggestingthat the closely linked IL-3 and GM-CSF genes are regulatedindependently of each other. This concept was first proposed onthe basis of the following: 1) the GM-CSF gene is expressed ef-ficiently in vivo as an isolated 10-kb transgene (15); 2) the IL-3
insulator effectively blocks activation of the IL-3 promoter by theGM-CSF enhancer (11); and 3) IL-3 and GM-CSF have over-lapping, but also unique functions in hematopoietic development.There is also the suggestion that coexpressed genes may actuallyrequire independent enhancers, which is based on evidence thatthe b-globin LCR can only activate one gene in this locus at anyone moment in time (33).In this study, we also obtained a fortuitous deletion in line E50
of a conserved sequence 30 kb downstream of the GM-CSF gene,which is homologous to the mouse GM-CSF +34-kb CNSaenhancer (18). However, the activity of this truncated GM-CSFtransgene was not substantially different from the intact trans-genes, suggesting that CNSa is not playing a major role in theinducible activation of the GM-CSF gene in T blast cells.
Identification of elements controlling IL-3 gene expression
Our analyses revealed the existence of a highly complex 40-kbregion extending upstream from the IL-3 gene that encompasses atleast three enhancers and a total of nine additional DHSs that eachhad inducible and/or tissue-specific properties that mirrored theexpression pattern of the IL-3 gene. Whereas it remains likely thatmany of these upstream elements are required for the correct finetuning of IL-3 gene expression, the cluster of DHSs that colo-calized with conserved sequences in the233-kb to 40-kb regionwas particularly prominent as a region closely linked to IL-3 geneexpression. This region is reminiscent of LCRs, which typicallyinclude both enhancer and nonenhancer elements, such as theb-globin LCR, which exists as a complex series of five DHSsspread over at least 22 kb upstream of the b-globin gene cluster(34). However, the IL-3 upstream regulatory region also includesinducible tissue-specific DHSs associated with LI-LINE retro-transposons. These DHSs may also make a significant contributionto the normal regulation of the IL-3 gene as there is already someprecedent for L1 LINE 59UTRs directing transcripts within thehuman genome, and even contributing to tissue specificity (35).This finding is also reminiscent of the observation that a repeatelement has been adopted as a normal component of an enhancerin the Tal1 locus (36), and our previous studies identified aberrantactivation of repeat elements as a factor contributing to aberrantgene expression in Hodgkin’s lymphoma (37).
Regulation of the 237-kb IL-3 enhancer
The 237-kb enhancer stands out as the dominant element con-trolling the IL-3 gene, considering the following: 1) the 237-kbenhancer is 10–to 20 times more powerful than either the 214- or24.5-kb enhancers; 2) the combination of the 214- and 24.5-kbenhancers was insufficient to support IL-3 gene promoter activityin vivo; 3) the 214-kb DHS has never been detected in non-leukemic cells; and 4) the IL-3 240-, 234-, and 24.1-kb DHSslack enhancer activity (12). Like the other previously definedenhancers in the IL-3/GM-CSF locus, the 237-kb enhancer con-tains ideal consensus binding sites for the Ca2+-inducible factorNFAT and the MAPK-inducible factor AP-1. The 237-kb NFATand AP-1 sites are, therefore, likely to represent significant targetsin which the Ca2+ and MAPK TCR signaling pathways convergeto activate the IL-3 locus. However, unlike the GM-CSF enhancerand IL-2 promoter, these NFAT and AP-1 motifs do not exist asclassical composite NFAT/AP-1 binding sites in which thesefactors bind cooperatively (1, 38).The exquisite tissue specificity of the 237-kb enhancer may be
dictated by a highly conserved 55-bp core segment encompassingGATA, ETS-1, and NFAT motifs. In addition, a highly conservedEGR-1 site exists just downstream of this region. The two ETS-1motifs and the EGR-1 binding site all contributed significantly to
FIGURE 7. Transcription factor binding within the 237-kb IL-3 en-
hancer. (A–C) Analyses of Jurkat T cells cultured with and without stim-
ulation for 4 h with 20 ng/ml PMA and 0.57 mM calcium ionophore
A23187. (A) EGR-1 ChIP assay of the237-kb IL-3 enhancer performed as
described (28), with enrichment levels normalized against a negative
control within the VEZF1 gene body. (B) EMSA performed using a probe
encompassing the IL-3 237-kb EGR-1 site and Abs against Sp1, Sp3, and
EGR-1. (C) Semiquantitative real-time PCR analysis of mRNA expression.
The Journal of Immunology 9
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

enhancer function in Jurkat T cells. The PU.1 and GATA motifswere not required for function in Jurkat T cells, which lack thesefactors. However, it is highly significant that essentially all themajor regulatory elements in the IL-3/GM-CSF locus encompassGATA motifs (1, 25). These GATA motifs are more likely to beimportant for promoting inducible expression in progenitor cellsand mast cells than in T cells, as is established for GATA motifs inthe GM-CSF enhancer and promoter (25). These motifs may alsobe critical for IL-3 and GM-CSF expression in the emerging he-mopoietic system in the embryo (4). Furthermore, because GATA-2 expression is typically maintained in leukemic cells arising fromMP cells, these GATA motifs are also likely to contribute to IL-3and GM-CSF expression in myeloid leukemia. Future studies ofthis locus will now be better equipped to identify the elements thataccount for the activation of IL-3 expression in CML in responseto constitutive tyrosine kinase signaling from the BCR-ABL fu-sion protein (21). It is also possible that some DHSs in the locuswill be disease specific, as the IL-3 214-kb enhancer DHS hasonly ever been detected in two cell lines derived from T cellleukemia (12).
The IL-3/GM-CSF locus undergoes a program ofdevelopmental regulation
The specific combination of developmentally regulated transcrip-tion factor motifs found in the 237-kb enhancer, together with themany RUNX motifs found elsewhere in the locus (1), representsan ideal combination adapted to support gene expression in pro-genitor cells, mast cells, and T cells (25, 39), but not in other celltypes. GATA, RUNX, and ETS family factors belong to a tightcluster of factors directing combinatorial control in hematopoieticprogenitor cells (39). However, because the 237-kb DHS isstrictly inducible, it is likely that the binding of the constitutivelyexpressed factors to the enhancer is dependent on the induciblefactors NFAT, AP-1, and EGR-1. This mechanism resemblesthe GM-CSF enhancer in which binding of the tissue-specificfactor RUNX-1 is dependent on accessibility to chromatin createdby NFAT-dependent chromatin remodeling (25).Cells with the capacity to express IL-3 and/or GM-CSF arise
from two different types of progenitor cell, as follows: 1) lymphoidprogenitors that can produce either T cells that express both genesor B cells that express neither gene, and 2) myeloid progenitors thatstart out with some capacity to express both genes, and can producemast cells that upregulate IL-3 and GM-CSF expression, andmonocytes/macrophages that downregulate both genes. Withinthe lymphoid lineage, the 24.1- and 21.5-kb DHSs were constitu-tively present in T blast cells and memory T cells, but absent in boththymocytes, naive T cells, and B cells (24). We also observed somespecific differences between the lymphoid and myeloid path-ways, with the IL-3 240.3- and 21.5-kb DHSs being T cellspecific, and the IL-3 216.4- and 210-kb DHSs being myeloidcell specific. To a very large extent, the inducible expression levelsseen in the different lineages reflect whether these cells had acquiredthese various DHSs.We observed that some specific DHSs in the IL-3 locus, such
as the IL-3 24.5- and 24.1-kb DHSs, begin to be acquired earlyduring the differentiation process in MP cells, and become strongerin IL-3–expressing mast cells, but are extinguished in an alternatedifferentiation pathway leading to macrophages. We previouslyproposed that the IL-3 24.5-, 24.1-, and 21.5-kb DHSs, whichare stably maintained in T blast cells, also play a role in main-taining the locus in a partially active state in memory T cells,priming the locus for activation by inducible enhancers (24). Thesestably maintained DHSs, and the IL-3 promoter (40), each encom-pass motifs for RUNX1, a factor known to be required for IL-3
gene expression and expansion of HSCs in the early embryo (4).Hence, it is significant that the constitutive IL-3 240-, 234-, and210-kb DHSs also each encompass RUNX1 motifs. Once theopen chromatin regions have formed at each of these DHSs, theRUNX elements may be able to maintain stable RUNX1 bindingand thereby maintain the locus in a primed state in both memoryT cells and myeloid cells that have gained the capacity to expressRUNX1. Most of these DHSs also encompass motifs for otherinducible factors that may be responsible for the initial creationof these DHSs.The finding that GM-CSF expression was also strongly down-
regulated during monocyte/macrophage differentiation was some-what unexpected. This does not appear to be a cell culture artifact,as we obtained similar findings in our studies of freshly isolatedspleen CD11b+ monocytic cells stimulated with either PMA/I orLPS (25). Hence, the concept that macrophage-lineage cells pro-duce significant amounts of GM-CSF may be more representativeof macrophage progenitor cells that express GATA-2, and primitivecell lines (25), rather than mature monocytes and macrophagesthat no longer express GATA-2.
AcknowledgmentsWe thank D. Roberts and D. Brooke for making transgenic mice and
H. Tagoh for the CD19 gene probe. We thank Peter Laslo for comments
on the manuscript.
DisclosuresThe authors have no financial conflicts of interest.
References1. Cockerill, P. N. 2004. Mechanisms of transcriptional regulation of the human IL-
3/GM-CSF locus by inducible tissue-specific promoters and enhancers. Crit. Rev.Immunol. 24: 385–408.
2. Alexander, W. S. 1998. Cytokines in hematopoiesis. Int. Rev. Immunol. 16: 651–682.
3. Nicola, N. A. 1989. Hemopoietic cell growth factors and their receptors. Annu.Rev. Biochem. 58: 45–77.
4. Robin, C., K. Ottersbach, C. Durand, M. Peeters, L. Vanes, V. Tybulewicz, andE. Dzierzak. 2006. An unexpected role for IL-3 in the embryonic development ofhematopoietic stem cells. Dev. Cell 11: 171–180.
5. He, W. Y., Y. Lan, H. Y. Yao, Z. Li, X. Y. Wang, X. S. Li, J. Y. Zhang, Y. Zhang,B. Liu, and N. Mao. 2010. Interleukin-3 promotes hemangioblast development inmouse aorta-gonad-mesonephros region. Haematologica 95: 875–883.
6. Gasson, J. C. 1991. Molecular physiology of granulocyte-macrophage colony-stimulating factor. Blood 77: 1131–1145.
7. Basu, S., A. R. Dunn, M. W. Marino, H. Savoia, G. Hodgson, G. J. Lieschke, andJ. Cebon. 1997. Increased tolerance to endotoxin by granulocyte-macrophagecolony-stimulating factor-deficient mice. J. Immunol. 159: 1412–1417.
8. Frazer, K. A., Y. Ueda, Y. Zhu, V. R. Gifford, M. R. Garofalo, N. Mohandas,C. H. Martin, M. J. Palazzolo, J. F. Cheng, and E. M. Rubin. 1997. Computa-tional and biological analysis of 680 kb of DNA sequence from the human 5q31cytokine gene cluster region. Genome Res. 7: 495–512.
9. Lee, G. R., C. G. Spilianakis, and R. A. Flavell. 2005. Hypersensitive site 7 ofthe TH2 locus control region is essential for expressing TH2 cytokine genes andfor long-range intrachromosomal interactions. Nat. Immunol. 6: 42–48.
10. Spilianakis, C. G., M. D. Lalioti, T. Town, G. R. Lee, and R. A. Flavell. 2005.Interchromosomal associations between alternatively expressed loci. Nature 435:637–645.
11. Bowers, S. R., F. Mirabella, F. J. Calero-Nieto, S. Valeaux, S. Hadjur,E. W. Baxter, M. Merkenschlager, and P. N. Cockerill. 2009. A conserved in-sulator that recruits CTCF and cohesin exists between the closely related butdivergently regulated interleukin-3 and granulocyte-macrophage colony-stimulating factor genes. Mol. Cell. Biol. 29: 1682–1693.
12. Hawwari, A., J. Burrows, M. A. Vadas, and P. N. Cockerill. 2002. The human IL-3 locus is regulated cooperatively by two NFAT-dependent enhancers that havedistinct tissue-specific activities. J. Immunol. 169: 1876–1886.
13. Bert, A. G., J. Burrows, A. Hawwari, M. A. Vadas, and P. N. Cockerill. 2000.Reconstitution of T cell-specific transcription directed by composite NFAT/Octelements. J. Immunol. 165: 5646–5655.
14. Duncliffe, K. N., A. G. Bert, M. A. Vadas, and P. N. Cockerill. 1997. A T cell-specific enhancer in the interleukin-3 locus is activated cooperatively by Oct andNFAT elements within a DNase I-hypersensitive site. Immunity 6: 175–185.
15. Cockerill, P. N., A. G. Bert, D. Roberts, and M. A. Vadas. 1999. The human GM-CSF gene is autonomously regulated in vivo by an inducible tissue-specificenhancer. Proc. Natl. Acad. Sci. USA 96: 15097–15102.
10 DEVELOPMENTAL REGULATION OF THE IL-3/GM-CSF LOCUS
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

16. Cockerill, P. N., M. F. Shannon, A. G. Bert, G. R. Ryan, and M. A. Vadas. 1993.The granulocyte-macrophage colony-stimulating factor/interleukin 3 locus isregulated by an inducible cyclosporin A-sensitive enhancer. Proc. Natl. Acad.Sci. USA 90: 2466–2470.
17. Osborne, C. S., M. A. Vadas, and P. N. Cockerill. 1995. Transcriptional regu-lation of mouse granulocyte-macrophage colony-stimulating factor/IL-3 locus. J.Immunol. 155: 226–235.
18. Wurster, A. L., P. Precht, and M. J. Pazin. 2011. NF-kB and BRG1 bind a distalregulatory element in the IL-3/GM-CSF locus. Mol. Immunol. 48: 2178–2188.
19. Johnson, B. V., A. G. Bert, G. R. Ryan, A. Condina, and P. N. Cockerill. 2004.GM-CSF enhancer activation requires cooperation between NFAT and AP-1elements and is associated with extensive nucleosome reorganization. Mol.Cell. Biol. 24: 7914–7930.
20. Russell, N. H. 1992. Autocrine growth factors and leukaemic haemopoiesis.Blood Rev. 6: 149–156.
21. Jiang, X., A. Lopez, T. Holyoake, A. Eaves, and C. Eaves. 1999. Autocrineproduction and action of IL-3 and granulocyte colony-stimulating factor inchronic myeloid leukemia. Proc. Natl. Acad. Sci. USA 96: 12804–12809.
22. Jiang, X., E. Ng, C. Yip, W. Eisterer, Y. Chalandon, M. Stuible, A. Eaves, andC. J. Eaves. 2002. Primitive interleukin 3 null hematopoietic cells transducedwith BCR-ABL show accelerated loss after culture of factor-independencein vitro and leukemogenic activity in vivo. Blood 100: 3731–3740.
23. Holyoake, T. L., X. Jiang, H. G. Jorgensen, S. Graham, M. J. Alcorn, C. Laird,A. C. Eaves, and C. J. Eaves. 2001. Primitive quiescent leukemic cells frompatients with chronic myeloid leukemia spontaneously initiate factor-indepen-dent growth in vitro in association with up-regulation of expression of inter-leukin-3. Blood 97: 720–728.
24. Mirabella, F., E. W. Baxter, M. Boissinot, S. R. James, and P. N. Cockerill. 2010.The human IL-3/granulocyte-macrophage colony-stimulating factor locus isepigenetically silent in immature thymocytes and is progressively activatedduring T cell development. J. Immunol. 184: 3043–3054.
25. Bert, A. G., B. V. Johnson, E. W. Baxter, and P. N. Cockerill. 2007. A modularenhancer is differentially regulated by GATA and NFAT elements that directdifferent tissue-specific patterns of nucleosome positioning and inducible chro-matin remodeling. Mol. Cell. Biol. 27: 2870–2885.
26. Butterfield, J. H., and D. A. Weiler. 1989. In vitro sensitivity of immature human mastcells to chemotherapeutic agents. Int. Arch. Allergy Appl. Immunol. 89: 297–300.
27. Bert, A. G., J. Burrows, C. S. Osborne, and P. N. Cockerill. 2000. Generation ofan improved luciferase reporter gene plasmid that employs a novel mechanismfor high-copy replication. Plasmid 44: 173–182.
28. Dickson, J., H. Gowher, R. Strogantsev, M. Gaszner, A. Hair, G. Felsenfeld, andA. G. West. 2010. VEZF1 elements mediate protection from DNA methylation.PLoS Genet. 6: e1000804.
29. Anderson, M. K., G. Hernandez-Hoyos, R. A. Diamond, and E. V. Rothenberg.1999. Precise developmental regulation of Ets family transcription factors duringspecification and commitment to the T cell lineage. Development 126: 3131–3148.
30. Liu, C., A. Calogero, G. Ragona, E. Adamson, and D. Mercola. 1996. EGR-1,the reluctant suppression factor: EGR-1 is known to function in the regulation ofgrowth, differentiation, and also has significant tumor suppressor activity anda mechanism involving the induction of TGF-beta1 is postulated to account forthis suppressor activity. Crit. Rev. Oncog. 7: 101–125.
31. Skerka, C., E. L. Decker, and P. F. Zipfel. 1995. A regulatory element in thehuman interleukin 2 gene promoter is a binding site for the zinc finger proteinsSp1 and EGR-1. J. Biol. Chem. 270: 22500–22506.
32. Khachigian, L. M., and T. Collins. 1997. Inducible expression of Egr-1-dependent genes: a paradigm of transcriptional activation in vascular endothe-lium. Circ. Res. 81: 457–461.
33. Wijgerde, M., F. Grosveld, and P. Fraser. 1995. Transcription complex stabilityand chromatin dynamics in vivo. Nature 377: 209–213.
34. Dillon, N., and F. Grosveld. 1993. Transcriptional regulation of multigene loci:multilevel control. Trends Genet. 9: 134–137.
35. Matlik, K., K. Redik, and M. Speek. 2006. L1 antisense promoter drivestissue-specific transcription of human genes. J. Biomed. Biotechnol. 2006:71753.
36. Smith, A. M., M. J. Sanchez, G. A. Follows, S. Kinston, I. J. Donaldson,A. R. Green, and B. Gottgens. 2008. A novel mode of enhancer evolution: theTal1 stem cell enhancer recruited a MIR element to specifically boost its activity.Genome Res. 18: 1422–1432.
37. Lamprecht, B., K. Walter, S. Kreher, R. Kumar, M. Hummel, D. Lenze,K. Kochert, M. A. Bouhlel, J. Richter, E. Soler, et al. 2010. Derepression of anendogenous long terminal repeat activates the CSF1R proto-oncogene in humanlymphoma. Nat. Med. 16: 571-579, 1p following 579.
38. Cockerill, P. N., A. G. Bert, F. Jenkins, G. R. Ryan, M. F. Shannon, andM. A. Vadas. 1995. Human GM-CSF enhancer function is associated with co-operative interactions between AP-1 and NFATp/c. Mol. Cell. Biol. 15: 2071–2079.
39. Wilson, N. K., S. D. Foster, X. Wang, K. Knezevic, J. Schutte, P. Kaimakis,P. M. Chilarska, S. Kinston, W. H. Ouwehand, E. Dzierzak, et al. 2010.Combinatorial transcriptional control in blood stem/progenitor cells: genome-wide analysis of ten major transcriptional regulators. Cell Stem Cell 7: 532–544.
40. Taylor, D. S., J. P. Laubach, D. G. Nathan, and B. Mathey-Prevot. 1996. Co-operation between core binding factor and adjacent promoter elements con-tributes to the tissue-specific expression of interleukin-3. J. Biol. Chem. 271:14020–14027.
The Journal of Immunology 11
at University of B
irmingham
on September 28, 2012
http://jimm
unol.org/D
ownloaded from

SUPPLEMENTARY,SECTION,,Supplementary,table,1:,,,Primers,used,to,create,probes,for,DHS,analyses,,
,,
1
SUPPLEMENTAL SECTION
Probes used DNase I-hypersensitive site analysis.
Probes based on previously published strategies for mapping DHSs were as follows:
(i) DHSs upstream of the IL-3 gene -10.1 kb BamHI site were probed with a 0.7 kb EcoRI-
BamHI fragment of DNA (-10.1 to -10.8 kb)1,
(ii) DHSs across the IL-3 promoter region were mapped from the -6.6 kb SpeI site with a 0.5
kb SpeI/BamHI fragment of DNA1,
(iii) DHSs across a 9.4 kb region spanning the GM-CSF enhancer and promoter were mapped
from an EcoRI site 6.8 kb upstream from the GM-CSF gene using a 1.5kb BamHI fragment
of DNA as a probe2,
(iv) DHSs spanning the IL-3 gene insulator downstream of the were mapped from a BamHI
site using a 1-kb BglII/BamHI fragment of DNA2.
Additional strategies developed in this study for probing regions of DNase I hypersensitivity
are detailed in supplementary table 1.
Table S1: Probes for mapping DNase I hypersensitive sites using PCR-generated probes.
Region probed Enzyme and location Enzyme sites and PCR primers used to create probes
Distal 5' IL-3
-40 kb DHS
BamHI -37.7 kb from the IL-3
start, probing upstream
PCR fragment using primers
5' CTGGAATGGAAGGTATGGCTC
and 5' CTGCTGCTGAGATGTCCCTTC
Distal 5' IL-3
-37 kb Enhancer
EcoRI -40.7 kb from the IL-3
start, probing downstream
PCR fragment using primers
5' GAGGTGACTTGGCCACAAACA
and 5' TGGAAATAGGTCCCCTGGTAG
Distal 5' IL-3
-34 kb DHS
BamHI -36.6 kb from the IL-3
start, probing downstream
PCR fragment using primers
5' CTCCCCTTCCATGACTTTTACC
and 5' AGCTAGTGCCCATGTGGTTC
Distal 5' IL-3
-23 kb DHS
KpnI -29.0 kb from the IL-3
start, probing downstream
Restriction enzyme fragment
720 bp KpnI (-29.0 kb) - SfiI (-28.3 kb)
Distal 5' IL-3
-10 kb DHS
SpeI -8.8 kb from the IL-3
start, probing upstream
PCR fragment using primers
5' CCTGGAGGAAGATGTGTGTG
and 5' ACTGCCCTGGAAAAGACAAG

Supplemental figure 1
B
27%
A
Nil PMA/I
IgG control CD4 Ab
CD4
Pro
m.
-1.5
kb
-4.1
kb
-4.5
kb
-14 k
b
E E
IL-3 gene 5’ regulatory elements
E E
IL-3 GM-CSF
E E E Ins
- 14 kb - 4.5 kb - 3 kb
P P
P
Analysis of the proximal human IL-3 enhancers and promoter in transgenic mice
The human IL-3 -14 kb enhancer was linked to a proximal 5.2 kb segment spanning the human
IL-3 -4.5 kb enhancer and the promoter, coupled to a human CD4 reporter gene, and used to
generate 8 independent lines of transgenic mice. The proximal 5.2 kb region of the IL-3 locus
also includes constitutive tissue-specific DHSs located at -4.1 and -1.5 kb. To establish a model
system to measure transgene activities we first generated cultures of T blast cells by stimulating
splenic cells with concanavalin A for two days, followed by at least 2 days culture with IL-2.
These cells were then re-stimulated for 4 h with 20 ng/ml PMA and 2 mM calcium ionophore
(PMA/I), and stained with either IgG control (grey line) or CD4 antibodies (black line). (A)
Representative example of 3 transgenic lines showing low level variegated human CD4
transgene expression. (B) Representative example of 5 transgenic lines showing no CD4
expression. The map underneath depicts the DNA segments used to construct the transgene,
depicting the enhancers (E), the IL-3 promoter (P), and the DHSs (vertical arrows).

Supplemental Figure 2
Cyclosporin A sensitivity of DHSs in the IL-3/GM-CSF locus
A
+2.9
+4.2
+4.9
+4.5
Nil
PM
A/I
PM
A/I
/CsA
BamHI probe EcoRI probe
Nil
PM
A/I
PM
A/I
/CsA
Prom
GME
IL-3 Insulator GM-CSF Enhancer
Fo
ld in
cre
ase in
Lucifera
se a
ctivity
B IL-3 +4.5 kb ncRNA Promoter activity
0
40
80
120
160
pXPG +4.5 kb DHS
IL-3 promoter
IL-3 -37 kb IL-3 -40 kb IL-3 -34 kb
- 37.5
Nil
PM
A/I
PM
A/I
/CsA
6.6
4.4
2.3
marker
DHS
EcoRI -37 kb probe
-40.6
-40.3
DHS
-40.9
6.6
4.4
2.3
marker
9.4
Nil
PM
A/I
PM
A/I
/CsA
BamHI -40 probe
EcoRI
EcoRI
-34.0
-35.0
DHS
-33.7
6.6
4.4
2.3
marker
Nil
PM
A/I
PM
A/I
/CsA
- 23.3
BamHI -34 probe
2.0
9.4
23
C
Cyclosporin A (CsA) sensitivity and function of inducible DHSs in the IL-3/GM-CSF locus.
(A and C) Mapping of DHSs in line C42 transgenic T blast cells, before and after stimulation for 4 h with
PMA/I, and in the presence or absence of 0.1 M CsA, using the probes defined in supplementary figure 2.
In panel C, the positions of HindIII-lambda DNA size markers are shown on the right hand side, and the
positions of DHSs relative to the IL-3 transcription start site are shown on the left.
(B) Luciferase reporter gene transfection assay of a non-coding RNA promoter within the inducible CsA-
resistant IL-3 +4.5 kb DHS. This DHS is the potential source of non-coding transcripts emanating from this
region, which are transcribed in the direction of the GM-CSF gene(4). DNA spanning the DHS was inserted
in pXPG in the same orientation as in the gene and assayed in parallel with the IL-3 promoter plasmid pIL3H
and the empty vector. Transfected cells were assayed after stimulation with PMA/I, and normalized values
were expressed relative to values for pXPG. Assays were performed in triplicate on each of two independent
clones of each construct. Error bars indicate S.D.

Supplemental figure 3
Conservation of transcription factor motifs within the -37 kb IL-3 enhancer
A Vertebrate conversation
B Enhancer Core = PU.1 + GATA + Ets + Egr + central NFAT + AP-1 motifs
PU.1 GATA ETS-1 #1 NFAT
Hu TGGGGAAGGGACATCTCAGCAGCAGAGGT--GGGTGTGGGATCC--AGATAACACAGGATGAGAGGTGGGAGGAAAGA-AAGTTCAGGC
Ch TGGGGAAGGGACATCTCAGCAGCAGAGGT--GGGTGTGGGATCC--AGATAACACAGGATGAGAGGTGGGAGGAAAGA-AAGTTCAGGC
Rh TGGGGAAGGGACGTCTCAGCAGCAGAGGT--GGGTGTGGGATCC--CAATAAGACAGAATGAGAGGTGGGAGGAAAGA-AAGTTCAGGC
Mo CCAGGAAGGGACCT------GGCAGAGGAGTGGGTGTGAGATCC--TGATAA--CAGAAAGACAAGTGGGAGGGAAGAAAAGTTCAGGC
Gu ---------------------GCGGGGGTGCGGGTGTGGGGCCCGGTGATAG--CAGGCAAATGGGTGGG-GGCGGGAAAAGTCCAGGC
Ra ---------------------GAGGGGAA---------AGGACC---AACAGCATAGGATGAGACAGGGGAGGAAGATAAAGTCCAGGC
Do TGGGG-AGGGTTTCATCA---GCAGAAAT--GCATGTAGAATCC--AGATAACACAGAATGAGCAGTGGGAGGAAAGAAAAGTCCAGGC
Ca TGGGGAAGAGCCCCATCA---GCAGATGT--GCATGTAGAATCC--AGATAACACAGGATGAGCAGTGGGAGGAAGGAAAAGTCCAGGC
Ho TGGGGAAGGG-CCCATCA---GCAGAAGT--GGGTGTGGGATCT--AGATAACACAGGATGAGCAGTGGGAGGAAAGAAAAGTCCAGGC
ETS-1 #2 EGR-1 NFAT AP-1
Hu AAAGGAACATCTTGCAGGTCTTTCTCAAACAACCTCCCCCACAAAATCATATTGATCCTGATTACAGCGTCTTTCCCATGAGTCAGTGTC
Ch AAAGGAACATCTTGCAGGTCTTTCTCAAACAACCTCCCCCACAAAATCATATTGATCCTGATTACAGCGTCTTTCCCATGAGTCAGTGTC
Rh AAAGGAACATCTTGCAGGTCTTTCTCAAACAACCTCCCCCGCAAAATCATATTGATCCTGATTACAGCGTCTTTCCCATGAGTCAGTGTC
Mo TAAGGAACA---------TCTTTG------------CCCCATACAGCTGCCCTGATCCCGATCACCGTGTCCTTCCCATGAGTCACTGTC
Gu CAAGGAACAGCTTGCAGGTCTTTGTCTAACATCCTCCCCCGCAGAATCACGCTGATCCTGACTGCAGTGTCTTTCCCCTGAGTCAGGGCC
Ra TGAAGAAGAGCTTCCAGGTCCTCA----GCATCCT-CCCCACAGAACCACACCAGTCCTGATCACCAGGCC--TCCCTCGAGTCAGTGCC
Do AAAGGAACATCTTAAAAGTCTTACTCTACTATCCTCCCCCACAGAATCATACTGGTCCTGATTACAGCTTCTTTCCCATGAGTCAGTGTC
Ca AGAGGAACATATTAAAAGTCTTTCTCTAATATCCTCCCCCACAGAATCATATTGATCCTGATTACAGCATCTTTCCCATGAGTCAATGTC
Ho AAAGGAACATCTTATAGGTCGTTCTCTAATATCCTCCCCCACAAAATCGTGTTGATCCTGATTACAGGGTCTTCCCCATGAGTCAGTATC
C 3’ region of Enhancer = 3’ AP-1 + NFAT motifs
AP-1 NFAT
Hu TGGCTTGACTCAGTCCTTCCTAACCTAGCCTTTTTGATTGTTTTTTTTTCTGAGCCTCTGCTGACTTTTTGGAAATTAGCCTTCAGA
Ch TGGCTTGACTCAGTCCTTCCTAACCTAGCCTTTTTGATTG-TATTTTTTCTGAGCCTCTGCTGACTTTTTGGAAATTAGCCTTCAGA
Rh TGGCTTGACTCAGTCCTTCCTAACCTGGCCTTTTCAGTTG-TTGTTTTTCTGAGCCTCTGCTGACTTTTTGGAAATTGACCTTCAGA
Mo TGGCTCAACTTAGCCCCTTCTAACATGGCCTCTTCTTCTA-CTTTCTCT======================================
Gu TGGCTTGGCTCAGTCCTTCCTCACCTGGCCTTTGCGTGCA-CCCTCTTTCTGAGCCTCAGGTGCCTTCTGGGAAGTTGGGTTTCACA
Ra ================================================================================TTTCAGA
Do TGCCTTGACTCAATCCTCCCT-ATCTGGCTTTTTTTTTTTTTTTTTTTTTTAAACCTCTGCTGCCTTCTTGGAAATTGGGTTTCAGA
Ca =======================================================================================
Ho TTGCTTGACTCAATCCTTCCTAACCTGGCTTTTTTTCTT-------------AACCTCTGCTGCTTCTTTGGAAATTGGGTTTCAGA
(A) UCSC genome browser view of conserved regions within the core of the human IL-3 -37 kb enhancer
(Vertebrate Multiz Alignment & PhastCons Conservation, http://genome.cse.ucsc.edu).
A map of predicted transcription factor binding sites is aligned underneath. (B and C) Alignment of
representative mammalian genome sequences of the central core (B) and 3’ region (C) of the enhancer. Motifs
are highlighted in yellow if the consensus sequence predicted to be required for DNA-binding is conserved.
The DNA sequences are from the human (Hu), chimp (Ch), rhesus (Rh), mouse (Mo), guinea pig (Gu), rabbit
(Ra), dog (Do), cat (Ca) and horse (Ho) reference genomes. Gaps in the alignment are depicted by dashes.
ET
S-1
#1
ET
S-1
#2
AP
-1
PU
.1
AP
-1
NF
AT
EG
R-1
GA
TA
NF
AT
123
305
NF
AT
enhancer core 3’ enhancer region
Copyright © 2022 FDOKUMEN




















