
Changes in intestinal tight junction permeability associated with
Upload
independentCategory
view
1download
0
t(
m
d
Virology 279, 201–209 (2001)doi:10.1006/viro.2000.0708, available online at http://www.idealibrary.com on
Human Immunodeficiency Virus Type 1 VPU Protein Affects Sindbis Virus GlycoproteinProcessing and Enhances Membrane Permeabilization
Marıa Eugenia Gonzalez1 and Luis Carrasco
Centro de Biologıa Molecular CSIC-UAM, Universidad Autonoma de Madrid, Cantoblanco, 28049 Madrid, Spain
Received July 3, 2000; returned to author for revision August 8, 2000; accepted October 11, 2000
The human immunodeficiency virus type 1 (HIV-1) Vpu is an integral membrane protein that forms oligomeric structuresin membranes. Expression of vpu using Sindbis virus (SV) as a vector leads to permeabilization of plasma membrane tohydrophilic molecules and impaired maturation of wild type SV glycoproteins in BHK cells. The 6K protein is a membraneprotein encoded in the SV genome that facilitates budding of virus particles and regulates transport of viral glycoproteinsthrough the secretory pathway. Some of these functions were assayed with a SV mutant containing a partially deleted 6Kgene. Transfection of BHK cells with pSVD6K vector rendered defective SVD6K virus, which had lower membrane perme-abilization, impaired glycoprotein processing, and deficient virion budding. Replacement of 6K function by HIV-1 Vpu in SVD6Kwas tested by cloning the vpu gene under a duplicated late promoter (pSVD6KVpu). The presence of the vpu gene in the6K-deleted virus enhances membrane permeability, modifies glycoprotein precursor processing, and facilitates infectiousvirus particle production. Restoration of infectivity of 6K-deleted SV by Vpu was evidenced by increased PFU production andcytopathic effect on infected cells. The modification of SVD6K glycoprotein maturation by Vpu was reflected in augmentedprocessing of B precursor and impairment of PE2 cleavage. Taken together, our data support the notion that HIV-1 Vpu andSV 6K proteins share some analogous functions. © 2001 Academic Press
Key Words: HIV-1; Vpu; accessory protein; membrane permeability; glycoprotein processing; membrane proteins; viralpathogenesis; Sindbis virus; 6K; alphavirus vector; gene expression.
c1i(isiECMipwtotapteipimc
INTRODUCTION
Vpu is a regulatory protein encoded by human immuno-deficiency virus type 1 (HIV-1) (Cohen et al., 1988; Strebel etal., 1988). Vpu is an integral type I membrane phosphopro-tein that forms homo-oligomers in membranes (Maldarelliet al., 1993). Vpu has not been found in virus particles.Immunofluorescence microscopic studies in mammaliancells showed major perinuclear localization of Vpu (Klimkaitet al., 1990; Kimura et al., 1994) as well as a minor localiza-ion at the cell membrane in a later stage of infectionSchubert et al., 1992; Friborg et al., 1995). Vpu synthesized
by E. coli cells appears in a crude membrane fraction,integrally associated with the outer membrane (Gonzalezand Carrasco, 1998). The dual location of Vpu is intriguing,with many observations pointing to the presence of Vpu inthe endoplasmic reticulum (ER)–Golgi network and also inplasma membrane.
Recent reports implicated Vpu with the modification ofmembrane functionality. Recombinant Vpu forms cation-specific channels when inserted into lipid bilayers andalso when expressed in E. coli or in Xenopus oocytes(Ewart et al., 1996; Schubert et al., 1996). In addition, Vpu
odifies membrane permeability upon expression in E.
mp
1 To whom correspondence and reprint requests should be ad-ressed. Fax: 34 91 3974799. E-mail: [email protected].
201
oli and mammalian cells (Gonzalez and Carrasco,998). Replication of Vpu-deficient HIV-1 gives rise to
ntracytoplasmic particles with aberrant morphologyKlimkait et al., 1990), associated with the cell surface ofnfected cells and connected to each other by shorttalks (Iwatani et al., 1997). On the other hand, Vpu
nteracts with the cytoplasmic domain of CD4 within theR and this specific interaction induces degradation ofD4 (Chen et al., 1993; Lenburg and Landau, 1993).oreover, the presence of Vpu in the ER–Golgi network
s required for efficient processing of envelope gp160rotein (Willey et al., 1992b). In addition, Vpu interferesith transport of soluble gp120 and secretion of G pro-
ein of VSV, and diminishes expression of MHC-I andther membrane proteins. Changes mediated by Vpu in
he biosynthesis (Coady et al., 1998), trafficking (Vincentnd Jabbar, 1995; Kerkau et al., 1997), or stability ofroteins (Willey et al., 1992a) were previously proposed
o explain inhibition of these membrane proteins. Thisxtensive array of inhibitory effects suggests a close
nteraction between Vpu and the secretory pathway. Ex-eriments using CD4/Vpu hybrids demonstrated that Vpu
n the context of CD4 is capable of reaching the plasmaembrane (Raja and Jabbar, 1996). The lack of an extra-
ellular domain and any maturation markers in Vpu
akes it technically difficult to follow the progress of therotein through the secretory pathway.
0042-6822/01 $35.00Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

202 GONZALEZ AND CARRASCO
Alphavirus vectors are convenient tools to study thefunctioning of cellular compartments involved in the se-cretory pathway. There is evidence that Semliki Forestvirus vector produces high levels of HIV-1 envelope gly-coproteins, which are proteolytically processed, folded,and transported to the cell membrane (Paul et al., 1993).Indeed, glycoproteins of HIV-1 and alphavirus envelopesshare the secretory pathway for transport from the ER tothe cell surface (reviewed in Einfeld, 1996; Strauss andStrauss, 1994). Both viruses synthesize a precursor of theenvelope protein in the endoplasmic reticulum, andthese proteins are glycosylated, modified by chaperone,and oligomerized before reaching the Golgi compart-ment. Both envelope precursors acquired Endo-H resis-tance in the Golgi and proteolytic cleavage by furin yieldsmature glycoproteins. Moreover, as previously noted,Vpu and alphavirus 6K proteins share structural andfunctional similarities (Carrasco, 1995) and also causeplasma membrane alterations (Sanz et al., 1994; Gonza-lez and Carrasco, 1998).
To explore potential modification of normal proteinprocessing through the secretory pathway, Vpu was ex-pressed using Sindbis virus (SV) vector. Interestingly, wefind that proteolytic processing of SV 6K-truncatedpolyprotein can be complemented in trans by HIV-1 Vpuprotein. We also find that Vpu complements membranepermeabilization capacity of 6K-defective Sindbis virus.These results are consistent with the idea that the dis-ruption of membrane integrity and the modification ofprotein-processing compartments by Vpu are connected.
RESULTS
Expression of vpu by recombinant Sindbis virus
To analyze the functionality of a recombinant viralprotein, it is desirable that the levels of its expression becomparable to a genuine viral infection. Some prokary-otic and eukaryotic transient expression systems thatproduce a high level of a single protein are available(Sanz et al., 1994; Gonzalez and Carrasco, 1998). For thatpurpose, we used a Sindbis virus vector that expressedthe vpu gene under the control of an additional SV sub-genomic promoter rendering pSVVpu plasmid (Fig. 1A).This construct allowed the possibility of introducing intocells the in vitro RNA transcripts from pSVWT andpSVVpu. Both pSVWT and pSVVpu plasmids were lin-earized, in vitro-transcribed using T7 RNA polymerase,and capped. BHK-21 cells were transfected with in vitro-made RNA transcripts. Cells electroporated with RNAfrom pSVVpu will produce two separate late mRNAsencoding SV glycoprotein precursor and Vpu protein.Thus, synthesis of SV structural proteins is not stoichio-metric with synthesis of Vpu protein. Immunofluores-cence analysis of transfected cells showed that over 90%
of cells were positive for Vpu protein (Fig. 1B). Levels ofVpu synthesis were high enough to be detected by au-toradiography of the SDS–PAGE of radioactive-labeledcells (Fig. 1C). This was confirmed by Western blot anal-ysis (Fig. 1D). Levels of Vpu were comparable to thoseobtained for other recombinant genes using this expres-sion system (Hahn et al., 1992).
Enhancement of membrane permeability byrecombinant SVVpu: effects on glycoproteinmaturation
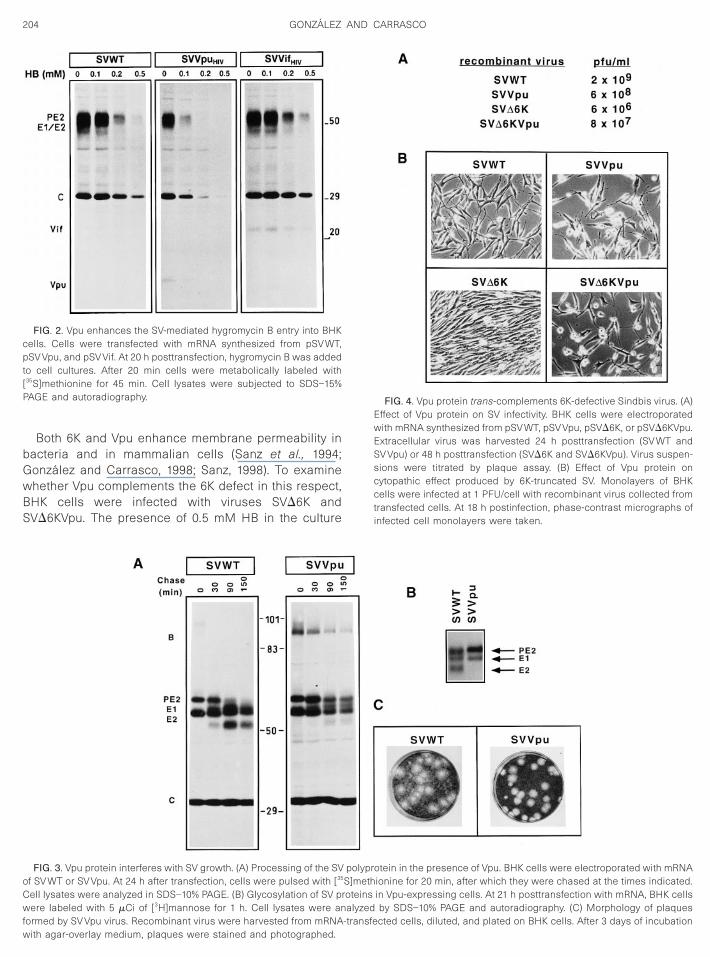
The modification of membrane permeability by Vpuwas analyzed by using the SV expression vector in BHKcells. The plasma cell membrane is nonpermeable tohydrophilic molecules like hygromycin B (HB), a potentinhibitor of translation (Carrasco, 1995). Membrane bar-rier integrity of cells transfected with SVWT and SVVpuRNAs was tested by analyzing protein synthesis of cellstreated with different concentrations of HB (Fig. 2). SDS–PAGE analysis of cell lysates demonstrated that treat-ment with 0.5 mM HB blocks protein synthesis in SV-transfected cells, consistent with previous data usingalphavirus-infected cells (Munoz et al., 1985). Notably,SVVpu-transfected cells were more sensitive to the inhi-bition of translation by HB. Enhancement of membranepermeability was not found, however, when other heter-ologous proteins, such as HIV-1 Vif protein, were synthe-sized (Fig. 2).
Interference of Vpu protein with maturation of SV gly-coproteins was studied by pulse-chase experiments (Fig.3A). Cells transfected with RNA transcripts that ex-pressed wild type SV genes showed that proteolyticprocessing of PE2 started before 90 min of chase andwas completed 1 h later, yielding mature E2 glycoprotein.By contrast, SV glycoprotein processing was hamperedin SVVpu-transfected cells. Thus, the B precursor ofabout 98 kDa containing all glycoproteins was observedafter the pulse and was processed soon after. In addi-tion, the PE2 precursor remained uncleaved, even after150 min of chase, such that mature E2 was not detected.To further analyze PE2 processing, cells were labeledwith radioactive mannose (Fig. 3B). Mannose-labeledPE2 precursor and E1 glycoproteins were found whetheror not Vpu was present in transfected cells. Notably,mature E2 was not detected in SVVpu-transfected cells.To test whether the defects of glycoprotein processingaffected virus production, virus plaque analysis was car-ried out (Fig. 3C). The amount of virus produced by eitherSVWT- or SVVpu-transfected cells was similar (see be-low). The plaque size was also equivalent in both in-stances, although the presence of Vpu rendered a moredefined contour of plaques. In addition, we observed thata more pronounced cytopathic effect was induced uponSVVpu infection of BHK cells. Our conclusion from these
experiments is that HIV-1 Vpu enhances membrane per-meability over that observed by SVWT infection and Vpu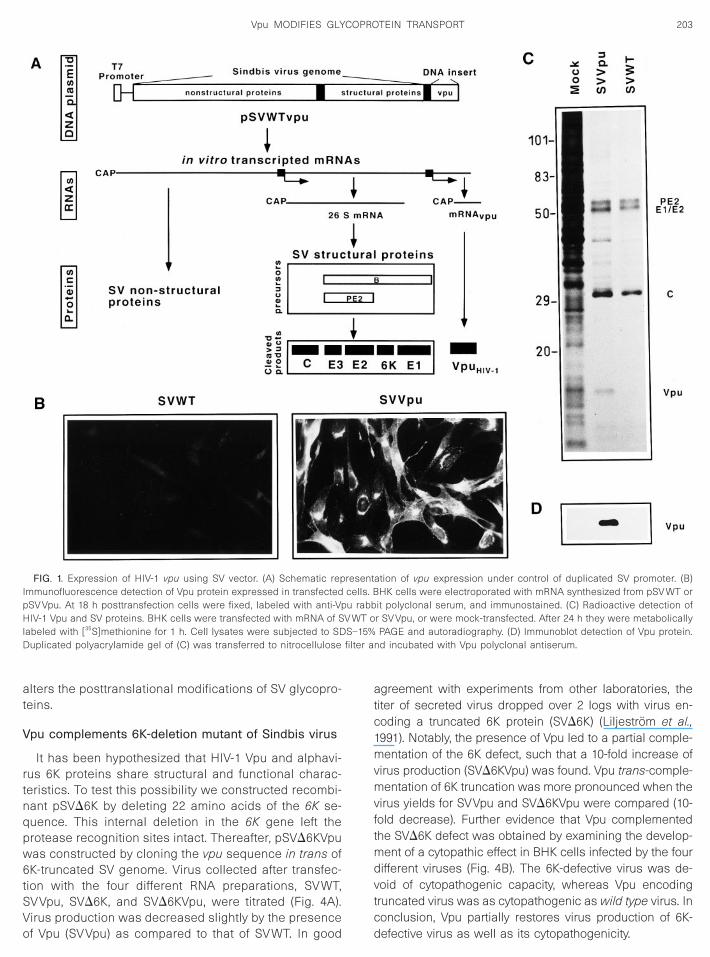
IpHl S–15%D filter a
203Vpu MODIFIES GLYCOPROTEIN TRANSPORT
alters the posttranslational modifications of SV glycopro-teins.
Vpu complements 6K-deletion mutant of Sindbis virus
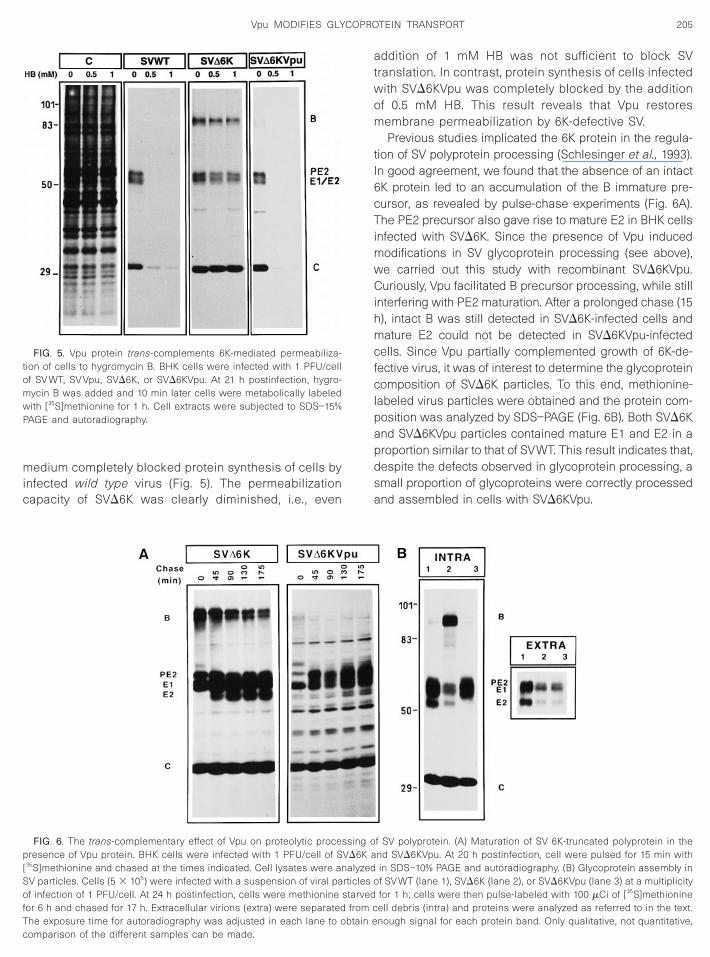
It has been hypothesized that HIV-1 Vpu and alphavi-rus 6K proteins share structural and functional charac-teristics. To test this possibility we constructed recombi-nant pSVD6K by deleting 22 amino acids of the 6K se-quence. This internal deletion in the 6K gene left theprotease recognition sites intact. Thereafter, pSVD6KVpuwas constructed by cloning the vpu sequence in trans of6K-truncated SV genome. Virus collected after transfec-tion with the four different RNA preparations, SVWT,SVVpu, SVD6K, and SVD6KVpu, were titrated (Fig. 4A).
FIG. 1. Expression of HIV-1 vpu using SV vector. (A) Schematic repmmunofluorescence detection of Vpu protein expressed in transfectedSV Vpu. At 18 h posttransfection cells were fixed, labeled with anti-VpIV-1 Vpu and SV proteins. BHK cells were transfected with mRNA of S
abeled with [35S]methionine for 1 h. Cell lysates were subjected to SDuplicated polyacrylamide gel of (C) was transferred to nitrocellulose
Virus production was decreased slightly by the presenceof Vpu (SVVpu) as compared to that of SVWT. In good
agreement with experiments from other laboratories, thetiter of secreted virus dropped over 2 logs with virus en-coding a truncated 6K protein (SVD6K) (Liljestrom et al.,1991). Notably, the presence of Vpu led to a partial comple-mentation of the 6K defect, such that a 10-fold increase ofvirus production (SVD6KVpu) was found. Vpu trans-comple-mentation of 6K truncation was more pronounced when thevirus yields for SVVpu and SVD6KVpu were compared (10-fold decrease). Further evidence that Vpu complementedthe SVD6K defect was obtained by examining the develop-ment of a cytopathic effect in BHK cells infected by the fourdifferent viruses (Fig. 4B). The 6K-defective virus was de-void of cytopathogenic capacity, whereas Vpu encodingtruncated virus was as cytopathogenic as wild type virus. In
ation of vpu expression under control of duplicated SV promoter. (B)HK cells were electroporated with mRNA synthesized from pSV WT or
it polyclonal serum, and immunostained. (C) Radioactive detection ofr SV Vpu, or were mock-transfected. After 24 h they were metabolicallyPAGE and autoradiography. (D) Immunoblot detection of Vpu protein.
nd incubated with Vpu polyclonal antiserum.
resentcells. Bu rabb
V WT o
conclusion, Vpu partially restores virus production of 6K-defective virus as well as its cytopathogenicity.
Cwfw
transfected cells. At 18 h postinfection, phase-contrast micrographs ofinfected cell monolayers were taken.
204 GONZALEZ AND CARRASCO
Both 6K and Vpu enhance membrane permeability inbacteria and in mammalian cells (Sanz et al., 1994;Gonzalez and Carrasco, 1998; Sanz, 1998). To examinewhether Vpu complements the 6K defect in this respect,BHK cells were infected with viruses SVD6K andSVD6KVpu. The presence of 0.5 mM HB in the culture
FIG. 2. Vpu enhances the SV-mediated hygromycin B entry into BHKcells. Cells were transfected with mRNA synthesized from pSV WT,pSV Vpu, and pSV Vif. At 20 h posttransfection, hygromycin B was addedto cell cultures. After 20 min cells were metabolically labeled with[35S]methionine for 45 min. Cell lysates were subjected to SDS–15%PAGE and autoradiography.
FIG. 3. Vpu protein interferes with SV growth. (A) Processing of the SVof SV WT or SV Vpu. At 24 h after transfection, cells were pulsed with [35
ell lysates were analyzed in SDS–10% PAGE. (B) Glycosylation of SV prere labeled with 5 mCi of [3H]mannose for 1 h. Cell lysates were an
polyprotein in the presence of Vpu. BHK cells were electroporated with mRNAS]methionine for 20 min, after which they were chased at the times indicated.oteins in Vpu-expressing cells. At 21 h posttransfection with mRNA, BHK cellsalyzed by SDS–10% PAGE and autoradiography. (C) Morphology of plaques
ormed by SV Vpu virus. Recombinant virus were harvested from mRNA-transfeith agar-overlay medium, plaques were stained and photographed.
FIG. 4. Vpu protein trans-complements 6K-defective Sindbis virus. (A)Effect of Vpu protein on SV infectivity. BHK cells were electroporatedwith mRNA synthesized from pSV WT, pSV Vpu, pSVD6K, or pSVD6KVpu.Extracellular virus was harvested 24 h posttransfection (SV WT andSV Vpu) or 48 h posttransfection (SVD6K and SVD6KVpu). Virus suspen-sions were titrated by plaque assay. (B) Effect of Vpu protein oncytopathic effect produced by 6K-truncated SV. Monolayers of BHKcells were infected at 1 PFU/cell with recombinant virus collected from
cted cells, diluted, and plated on BHK cells. After 3 days of incubation
c
mw
205Vpu MODIFIES GLYCOPROTEIN TRANSPORT
medium completely blocked protein synthesis of cells byinfected wild type virus (Fig. 5). The permeabilization
apacity of SVD6K was clearly diminished, i.e., even
FIG. 5. Vpu protein trans-complements 6K-mediated permeabiliza-tion of cells to hygromycin B. BHK cells were infected with 1 PFU/cellof SV WT, SV Vpu, SVD6K, or SVD6KVpu. At 21 h postinfection, hygro-
ycin B was added and 10 min later cells were metabolically labeledith [35S]methionine for 1 h. Cell extracts were subjected to SDS–15%
PAGE and autoradiography.
FIG. 6. The trans-complementary effect of Vpu on proteolytic procepresence of Vpu protein. BHK cells were infected with 1 PFU/cell of S[35S]methionine and chased at the times indicated. Cell lysates were anSV particles. Cells (5 3 105) were infected with a suspension of viral paof infection of 1 PFU/cell. At 24 h postinfection, cells were methioninefor 6 h and chased for 17 h. Extracellular virions (extra) were separated
The exposure time for autoradiography was adjusted in each lane to obtain ecomparison of the different samples can be made.addition of 1 mM HB was not sufficient to block SVtranslation. In contrast, protein synthesis of cells infectedwith SVD6KVpu was completely blocked by the additionof 0.5 mM HB. This result reveals that Vpu restoresmembrane permeabilization by 6K-defective SV.
Previous studies implicated the 6K protein in the regula-tion of SV polyprotein processing (Schlesinger et al., 1993).In good agreement, we found that the absence of an intact6K protein led to an accumulation of the B immature pre-cursor, as revealed by pulse-chase experiments (Fig. 6A).The PE2 precursor also gave rise to mature E2 in BHK cellsinfected with SVD6K. Since the presence of Vpu inducedmodifications in SV glycoprotein processing (see above),we carried out this study with recombinant SVD6KVpu.Curiously, Vpu facilitated B precursor processing, while stillinterfering with PE2 maturation. After a prolonged chase (15h), intact B was still detected in SVD6K-infected cells andmature E2 could not be detected in SVD6KVpu-infectedcells. Since Vpu partially complemented growth of 6K-de-fective virus, it was of interest to determine the glycoproteincomposition of SVD6K particles. To this end, methionine-labeled virus particles were obtained and the protein com-position was analyzed by SDS–PAGE (Fig. 6B). Both SVD6Kand SVD6KVpu particles contained mature E1 and E2 in aproportion similar to that of SVWT. This result indicates that,despite the defects observed in glycoprotein processing, asmall proportion of glycoproteins were correctly processedand assembled in cells with SVD6KVpu.
f SV polyprotein. (A) Maturation of SV 6K-truncated polyprotein in thend SVD6KVpu. At 20 h postinfection, cell were pulsed for 15 min within SDS–10% PAGE and autoradiography. (B) Glycoprotein assembly inf SV WT (lane 1), SVD6K (lane 2), or SVD6KVpu (lane 3) at a multiplicityfor 1 h; cells were then pulse-labeled with 100 mCi of [35S]methionineell debris (intra) and proteins were analyzed as referred to in the text.
ssing oVD6K a
alyzedrticles ostarved
from c
nough signal for each protein band. Only qualitative, not quantitative,
atvNcTpSwvcoa
M
eco1a1bsbbsem
bp
mcdVbp
G
ttccm
206 GONZALEZ AND CARRASCO
DISCUSSION
The analogies of 6K and Vpu were previously noticed(Carrasco, 1995). Both proteins are small and hydropho-bic, and both interact with membranes. The functioningof Vpu and 6K was related with virion production, mem-brane permeabilization, and glycoprotein processing. Ev-idence of the extent of these similarities between Vpuand 6K is further provided by our present results, show-ing that replacement of 6K by Vpu in the SVD6K en-hances virion production, membrane permeability, andthe cytopathic effect. In addition, Vpu restores proteolyticprocessing of B, the glycoprotein precursor that wasaccumulated by cells infected with 6K-deleted virus,while it inhibits the furin-mediated cleavage of PE2.
Virus production
The mechanism by which Vpu enhances the release ofvirus particles is intriguing. Vpu increases the pinching-off from the plasma membrane of infectious particles andvirus-like particles of HIV. Moreover, Vpu augments virusparticle production from other retroviruses as divergentas Visna virus and Mo-MLV (Klimkait et al., 1990; Paul et
l., 1998; Gottlinger et al., 1993). The 6K protein affectswo different steps during the SV replication cycle: (i)irion release from the plasma membrane (Gaedigk-itschko et al., 1990) and (ii) the transport of vesicles
ontaining SV glycoproteins (Schlesinger et al., 1993).ransport of 6K protein through secretory cellular com-artments is well characterized (reviewed in Strauss andtrauss, 1994), whereas the precise mechanism byhich 6K contributes to virus assembly and infectivity of
irions is obscure. The number of infectious virus parti-les (6K-deleted SV) was 10-fold higher in the presencef Vpu. This finding adds further evidence to the potentialnalogy of Vpu and 6K proteins.
embrane permeabilization
Vpu disrupts membrane integrity in prokaryotic andukaryotic cells (Gonzalez and Carrasco, 1998). Ionhannel activity of Vpu on model membranes was previ-usly documented (Ewart et al., 1996; Marassi et al.,999). Besides, 6K is also a membrane-active proteinble to enhance membrane permeability (Sanz et al.,994). SVD6K mutants show defects in promoting mem-rane permeability (Sanz and Carrasco, unpublished re-ults). The capacity to permeabilize the plasma mem-rane and to induce the cytopathic effect were recoveredy 6K-deleted virus carrying the vpu gene. This repre-ents a good model system to test other viral proteinsndowed with the capacity to enhance membrane per-eability.A model explaining the role of Vpu channels in the
udding process suggests that insertion of Vpu at thelasma membrane alters the resting potential and pro-
otes fusion and release of virions. Accordingly, Vpuhannels can be involved in the enhancement of 6K-eleted particle release. The complementation of 6K withpu suggests that Vpu induces a modification on mem-ranes similar to 6K that may influence budding of virusarticles from the cell membrane.
lycoprotein processing
Vpu can affect the sorting of proteins from the secre-ory pathway by direct protein–protein interactionshrough specific Vpu-responding-elements or by modifi-ation of secretory compartments by means of the ionhannel activity. In accord with defects in the secretion ofembrane proteins (Kerkau et al., 1997; Willey et al.,
1992b; Vincent and Jabbar, 1995), we found that Vpuimpaired the processing of PE2 glycoprotein. These ob-servations suggest that Vpu modifies the functioning ofcellular compartments involved in the secretory pathway.A previous report showed that the expression of a viralprotein endowed with ion channel activity, such as influ-enza M2, also results in the modification of viral glyco-protein transport (Sakaguchi et al., 1996). Self-assemblyof Vpu monomers into noncovalent oligomers, resem-bling potassium channels, can alternate during vesiculartransport with monomeric structures (Willbold et al.,1997; Moore et al., 1998; Marassi et al., 1999). Thus, theproportion of oligomeric forms of Vpu during its transportto the plasma membrane can determine the availabilityof ion channels that modify each cellular compartment.This then helps to explain why Vpu affects the process-ing of SV glycoproteins at multiple sites.
It is noteworthy that Vpu trans-complements a 6K-deficient SV because, in the wild type virus, the 6K geneis translated as part of a precursor that gives rise to thestructural glycoproteins. As soon as 6K is covalentlyseparated from E1 and PE2, it becomes associated withthe PE2/E1 complex. The sequence-specific interactionsbetween 6K and the PE2/E1 complex allows the trans-port of 6K protein to the cell membrane, where it be-comes assembled into virions, although with a lowerstoichiometric proportion than that of mature glycopro-teins (Gaedigk-Nitschko and Schlesinger, 1990). Thus,the sequence divergence between 6K and Vpu shouldprevent the specific interaction of Vpu with PE2/E1 com-plexes during assembly of 6K-deleted virus. The stabilityof PE2/E1 complex is critical for furin-mediated cleavage(Strauss and Strauss, 1994). Conceivably, the absence of6K/heterodimer-specific interactions may cause the re-duction of virus production that we observed withSVD6KVpu, as compared to that of SVWT. In SVD6K, Vpucomplemented some 6K functions but not sequence-specific interactions. Actually, a SV chimeric virus inwhich the 6K gene had been replaced with that from
Ross River virus gave rise to a 10-fold reduction of as-sembled virus (Yao et al., 1996). The approximately two-
r
cc
eaebtdu
207Vpu MODIFIES GLYCOPROTEIN TRANSPORT
fold decrease in the yield of SVWT titer by Vpu could bethe arithmetical result of the increase of virus productionby the enhancement of membrane permeabilization andthe decrease of virus assembly resulting from the ab-sence of specific interactions of Vpu with glycoproteincomplexes. Therefore, the partial complementation of6K-deleted mutant by Vpu provides a model to study thefunctionality of other viral proteins related to 6K.
MATERIALS AND METHODS
Plasmids
pSVWT, a plasmid containing a full-length copy of theSindbis virus genome downstream from the T7 promoter,was constructed from pTE392J (Hahn et al., 1992) byeplacing the SacI–SpeI fragment containing the SP6
promoter with the corresponding region from pT7TOTOplasmid (generously provided by Dr. C. M. Rice, St. Louis,MO). pSVD6K, containing Sindbis virus genome with atruncated 6K protein, was obtained by an intermediatesubcloning of an NdeI–BamHI fragment. This fragmentwas obtained in two pieces by PCR amplification from thepSVWT template. Oligonucleotides used for the 59 fragmentwere: 59Sub6K (CCCGGGCATATGGGATGGCCACACGAAA-ATAGTACAG) and 39Del6K (GGCGCGCATGCCTGGACCCA-GAAGAACGGCTGACT). Oligonucleotides used for the 39fragment were: 59Del6K (CCCGGGGCATGCGCCGGCGC-CTACCTGGCGAAGGTA) and 39Sub6K (GGCGCCGGATC-CTTATTAGACGTACGCCTCACTCATCTGGCT). Each ampli-fied fragment was doubled digested with NdeI and SphI orSphI and BamHI, respectively (restriction sites of oligonu-
leotides are underlined) and then ligated. The fragmentontaining a new internal SphI site and compatible cohe-
sive ends was subcloned into pET11 using NdeI andBamHI restriction sites. Digestion of the pET11 constructwith BssHII and SplI rendered a fragment containing amutated 6K (deletion of 24–45 amino acids and conversionof Leu22 to Ala) that was used to replace the wild type 6Ksequence in pSVWT. pSVVpu and pSVD6Vpu were ob-tained by subcloning the NcoI–BamHI fragment containingvpu sequence from pTM1vpu (Gonzalez and Carrasco,1998) into pSVWT or pSVD6K, using the shuttle phagemidvector pH392J1 (Hahn et al., 1992). A procedure similar tothat used for pSVVpu construction was followed for pSVVifconstruction; the vif gene was PCR-amplified from the BH10clone of HIV-1 (Hahn et al., 1984) using oligonucleotidescontaining 59NcoI and 39BamHI restriction sites.
In vitro transcription and transfections
Plasmid containing T7 promoter was linearized withXhoI before use as a template for in vitro transcription,which was performed in 25 ml at 37°C for 2 h. Reactionmixtures contained: 2.5 ml DNA template; 40 mM Tris–
HCl (pH 7.9); 6 mM MgCl2; 2 mM spermidine–HCl; 5 mMdithiothreitol; 1 mM each ATP, CTP, and UTP; 500 mMWc
GTP; 10 U of RNAsin; and 0.5 ml of T7 RNA polymerase.In addition, 1 mM of 7mG(59)ppp(59)G (New EnglandBioLabs, Beverly, MA) was added to the reaction mixtureto obtain capped RNAs. BHK-21 cells were transfectedwith capped RNAs by electroporation, following a previ-ously reported protocol (Liljestrom et al., 1991). Briefly,2 3 106 cells were resuspended in 0.4 ml of phosphate-buffered saline (PBS) and mixed with 25 ml of transcrip-tion product at room temperature. The mixture was elec-troporated in a 0.2-cm cuvette by two consecutive pulsesat 1500 V and 25 mF with a Gene Pulser (Bio-Rad, Her-cules, CA). The transfected cells were diluted 1:20 withDMEM with 5% FCS and plated.
Cell culture and viruses
Baby hamster kidney (BHK-21) cells were cultured inDulbecco’s modified Eagle’s medium (DMEM) with 5%heat-inactivated fetal calf serum (FCS). Sindbis virus waspropagated in BHK cells. Plaque assay was performedon BHK cells; confluent monolayers were infected withserial virus dilutions, covered with 0.7% agar-overlaidmedium, and incubated for 3 days. Following standardprocedures, cell monolayers were fixed with 10% form-aldehyde and plaques stained with 2% crystal violet in10% formaldehyde.
Immunoblot analysis
Proteins were separated by SDS–PAGE (10% poly-acrylamide) and transferred onto a nitrocellulose filter.The filter was blocked by incubation in 3% nonfat dry milkin washing solution (0.05% Tween in PBS), at room tem-perature for 1 h. Blocked membrane was washed threetimes with washing solution and then incubated with theprimary antibody at room temperature for 2 h. After re-moving antibody, the filter was washed three times withwashing solution and incubated with biotin-conjugatedsecondary antibody at room temperature for 1 h. Afterwashing three times, streptavidin-conjugated peroxidasewas coupled to biotin-labeled immune complex. Thefilter was then incubated with luminol-luciferin solutionand the light emission from the chemiluminescence re-action was captured on an Agfa X-ray film.
Metabolic labeling of proteins
Cell monolayers were labeled with L-[35S]methionine(1000 Ci/mmol) or D-[2,6-3H]mannose (38 Ci/mmol) (Am-
rsham, Arlington Heights, IL) in methionine-free DMEMnd glucose-free DMEM, respectively. In pulse/chasexperiments, cells were methionine starved for 30 minefore radioactive labeling, the metabolic labeling was
erminated by washing cell monolayers with fresh me-ium, and cells were maintained in DMEM containingnlabeled methionine for the appropriate chase period.
hen noted, proteins incorporated in mature viral parti-les were separated from total viral proteins by sequen-

tibPf
I
ewfwPPaPc1mA
C
C
C
E
E
F
G
G
G
G
H
H
I
K
K
L
L
M
M
M
M
P
208 GONZALEZ AND CARRASCO
tial low- and high-speed centrifugations. Briefly, cellmonolayers together with culture medium were centri-fuged at 2000 g for 10 min. Pelleted cells were sus-pended in sample buffer (160 mM Tris–HCl, pH 6.8, 100mM dithiothreitol, 2% SDS, 0.03% bromphenol blue, and11.6% glycerol), heated to 100°C for 4 min, and analyzedby SDS–10% PAGE and autoradiography. The superna-tant was clarified by centrifugation at 10,000 g for 10 min.Precipitated debris was removed and the supernatantcontaining viral particles was layered on top of 1/10volume of PBS with 20% sucrose (w/v), and centrifuged at100,000 g for 2 h at 4°C, in a Beckman TL100 ultracen-rifuge (Beckman Coulter, Fullerton, CA). Pellets contain-ng partially purified virus were suspended in sampleuffer and viral proteins were resolved by SDS–10%AGE and visualized by autoradiography on Agfa X-ray
ilm.
mmunofluorescence assay
Very dilute cell monolayers were grown on glass cov-rslips. Prior to immunostaining, cell monolayers wereashed with PBS and fixed with 4% paraformaldehyde
or 10 min at room temperature. Cells were washed twiceith PBS and permeabilized with 0.2% Triton X-100 inBS for 2 min at room temperature. After washing withBS, cells were incubated in PBS with primary polyclonalntiserum for 1 h at 37°C. Cells were then washed withBS three times and incubated in PBS with rhodamine-onjugated goat anti-rabbit IgG (Pierce, Rockford, IL) forh at 37°C. The coverslips were placed on glass slides,ounted with Mowiol solution, and viewed with a Zeissxiovert 35 microscope (Zeiss, Thornwood, NY).
ACKNOWLEDGMENTS
We thank Dr. Miguel Angel Sanz for construction of pSV WT plasmidand mutant form pSVD6K. This work was supported by Fondo deInvestigacion Sanitaria (Grant 98/0644) and Comunidad de Madrid(Grant 07B/0002/1997). We also acknowledge the institutional grant tothe CBM of Fundacion Areces.
REFERENCES
Carrasco, L. (1995). Modification of membrane permeability by animalviruses. Adv. Virus Res. 45, 61–112.
hen, M.-Y., Maldarelli, F., Karczewski, M. K., Willey, R. L., and Strebel,K. (1993). Human immunodeficiency virus type 1 Vpu protein inducesdegradation of CD4 in vitro: The cytoplasmic domain of CD4 contrib-utes to Vpu sensitivity. J. Virol. 67, 3877–3884.
oady, M. J., Daniel, N. G., Tiganos, E., Allain, B., Friborg, J., Lapointe,J. Y., and Cohen, E. A. (1998). Effects of Vpu expression on Xenopusoocyte membrane conductance. Virology 244, 39–49.
ohen, E. A., Terwilliger, E. F., Sodroski, J. G., and Haseltine, W. A.(1988). Identification of a protein encoded by the vpu gene of HIV-1.Nature 334, 532–534.
infeld, D. (1996). Maturation and assembly of retroviral glycoproteins.Curr. Top. Microbiol. Immunol. 214, 133–176.
wart, G. D., Sutherland, T., Gage, P. W., and Cox, G. B. (1996). The Vpu
protein of human immunodeficiency virus type 1 forms cation-selec-tive ion channels. J. Virol. 70, 7108–7115.riborg, J., Ladha, A., Gottlinger, H., Haseltine, W. A., and Cohen, E. A.(1995). Functional analysis of the phosphorylation sites on the hu-man immunodeficiency virus type 1 Vpu protein. J. Acquir. ImmuneDefic. Syndr. Hum. Retrovirol. 8, 10–22.
aedigk-Nitschko, K., Ding, M., Aachlevy, M., and Schlesinger, M. J.(1990). Site-directed mutations in the Sindbis virus 6K protein revealsites for fatty acylation and the underacylated protein affects virusrelease and virion structure. Virology 175, 282–291.
aedigk-Nitschko, K., and Schlesinger, M. J. (1990). The Sindbis virus6K protein can be detected in virions and is acylated with fatty acids.Virology 175, 274–281.
onzalez, M. E., and Carrasco, L. (1998). The human immunodeficiencyvirus type 1 Vpu protein enhances membrane permeability. Biochem-istry 37, 13710–13719.
ottlinger, H. G., Dorfman, T., Cohen, E. A., and Haseltine, W. A. (1993).Vpu protein of human immunodeficiency virus type 1 enhances therelease of capsids produced by gag gene constructs of widelydivergent retroviruses. Proc. Natl. Acad. Sci. USA 90, 7381–7385.
ahn, B. H., Shaw, G. M., Arya, S. K., Popovic, M., Gallo, R. C., andWong-Staal, F. (1984). Molecular cloning and characterization of theHTLV-III virus associated with AIDS. Nature 312, 166–169.
ahn, C. S., Hahn, Y. S., Braciale, T. J., and Rice, C. M. (1992). InfectiousSindbis virus transient expression vectors for studying antigen pro-cessing and presentation. Proc. Natl. Acad. Sci. USA 89, 2679–2683.
watani, Y., Song, S. K., Wang, L. Q., Planas, J., Sakai, H., Ishimoto, A.,and Cloyd, M. W. (1997). Human immunodeficiency virus type 1 Vpumodifies viral cytopathic effect through augmented virus release.J. Gen. Virol. 78, 841–846.
erkau, T., Bacik, I., Bennink, J. R., Yewdell, J. W., Hunig, T., Schimpl, A.,and Schubert, U. (1997). The human immunodeficiency virus type 1(HIV-1) vpu protein interferes with an early step in the biosynthesis ofmajor histocompatibility complex (MHC) class I molecules. J. Exp.Med. 185, 1295–1305.
imura, T., Nishikawa, M., and Ohyama, A. (1994). Intracellular mem-brane traffic of human immunodeficiency virus type 1 envelopeglycoproteins: Vpu liberates Golgi-targeted gp160 from CD4-depen-dent retention in the endoplasmic reticulum. J. Biochem. 115, 1010–1020.
Klimkait, T., Strebel, K., Hoggan, M. D., Martin, M. A., and Orenstein,J. M. (1990). The human immunodeficiency virus type 1-specificprotein vpu is required for efficient virus maturation and release.J. Virol. 64, 621–629.
enburg, M. E., and Landau, N. R. (1993). Vpu-induced degradation ofCD4: Requirement for specific amino acid residues in the cytoplas-mic domain of CD4. J. Virol. 67, 7238–7245.
iljestrom, P., Lusa, S., Huylebroeck, D., and Garoff, H. (1991). In vitromutagenesis of a full-length cDNA clone of Semliki Forest virus: Thesmall 6.000-molecular-weight membrane protein modulates virusrelease. J. Virol. 65, 4107–4113.aldarelli, F., Chen, M.-Y., Willey, R. L., and Strebel, K. (1993). Humanimmunodeficiency virus type 1 Vpu protein is an oligomeric type Iintegral membrane protein. J. Virol. 67, 5056–5061.arassi, F. M., Ma, C., Gratkowski, H., Straus, S. K., Strebel, K., Oblatt-Montal, M., Montal, M., and Opella, S. J. (1999). Correlation of thestructural and functional domains in the membrane protein Vpu fromHIV-1. Proc. Natl. Acad. Sci. USA 96, 14336–14341.oore, P. B., Zhong, Q. F., Husslein, T., and Klein, M. L. (1998). Simu-lation of the HIV-1 Vpu transmembrane domain as a pentamericbundle. FEBS Lett. 431, 143–148.unoz, A., Castrillo, J. L., and Carrasco, L. (1985). Modification ofmembrane permeability during Semliki Forest virus infection. Virol-ogy 146, 203–212.
aul, M., Mazumder, S., Raja, N., and Jabbar, M. A. (1998). Mutationalanalysis of the human immunodeficiency virus type 1 Vpu transmem-brane domain that promotes the enhanced release of virus-like
particles from the plasma membrane of mammalian cells. J. Virol. 72,1270–1279.
S
W
Y
209Vpu MODIFIES GLYCOPROTEIN TRANSPORT
Paul, N. L., Marsh, M., McKeating, J. A., Schulz, T. F., Liljestrom, P.,Garoff, H., and Weiss, R. A. (1993). Expression of HIV-1 envelopeglycoproteins by Semliki Forest virus vectors. AIDS Res. Hum. Ret-roviruses 9, 963–970.
Raja, N. U., and Jabbar, M. A. (1996). The human immunodeficiencyvirus type 1 Vpu protein tethered to the CD4 extracellular domain islocalized to the plasma membrane and is biologically active in thesecretory pathway of mammalian cells: Implications for the mecha-nisms of Vpu function. Virology 220, 141–151.
akaguchi, T., Leser, G. P., and Lamb, R. A. (1996). The ion channelactivity of the influenza virus M2 protein affects transport though theGolgi apparatus. J. Cell Biol. 133, 733–747.
Sanz, M. A. (1998). Caracterizacion funcional de la proteına 6K deAlfavirus. El virus Sindbis como vector de expresion. Ph.D. Thesis,Universidad Autonoma de Madrid.
Sanz, M. A., Perez, L., and Carrasco, L. (1994). Semliki Forest virus 6Kprotein modifies membrane permeability after inducible expressionin Escherichia coli cells. J. Biol. Chem. 269, 12106–12110.
Schlesinger, M. J., London, S. D., and Ryan, C. (1993). An in-frameinsertion into the Sindbis virus 6K gene leads to defective proteolyticprocessing of the virus glycoproteins, a trans-dominant negativeinhibition of normal virus formation, and interference in virus shut offof host-cell protein synthesis. Virology 193, 424–432.
Schubert, U., Ferrer-Montiel, A. V., Oblatt-Montal, M., Henklein, P.,
Strebel, K., and Montal, M. (1996). Identification of an ion channelactivity of the Vpu transmembrane domain and its involvement in theregulation of virus release from HIV-1-infected cells. FEBS Lett. 398,12–18.
Schubert, U., Schneider, T., Henklein, P., Hoffmann, K., Berthold, E.,Hauser, H., Pauli, G., and Porstmann, T. (1992). Human-immunodefi-ciency-virus-type-1-encoded Vpu protein is phosphorylated by ca-sein kinase II. Eur. J. Biochem. 204, 875–883.
Strauss, J. H., and Strauss, E. G. (1994). The alphaviruses: Gene ex-pression, replication, and evolution. Microbiol. Rev. 58, 491–562.
Strebel, K., Klimkait, T., and Martin, M. A. (1988). A novel gene of HIV-1,vpu, and its 16-kilodalton product. Science 241, 1221–1223.
Vincent, M. J., and Jabbar, M. A. (1995). The human immunodeficiencyvirus type 1 Vpu protein: A potential regulator of proteolysis andprotein transport in the mammalian secretory pathway. Virology 213,639–649.
Willbold, D., Hoffmann, S., and Rosch, P. (1997). Secondary structureand tertiary fold of the human immunodeficiency virus protein U (Vpu)cytoplasmic domain in solution. Eur. J. Biochem. 245, 581–588.
Willey, R. L., Maldarelli, F., Martin, M. A., and Strebel, K. (1992a). Humanimmunodeficiency virus type 1 Vpu protein regulates the formation ofintracellular gp160-CD4 complexes. J. Virol. 66, 226–234.illey, R. L., Maldarelli, F., Martin, M. A., and Strebel, K. (1992b). Humanimmunodeficiency virus type 1 Vpu protein induces rapid degrada-tion of CD4. J. Virol. 66, 7193–7200.
ao, J. S., Strauss, E. G., and Strauss, J. H. (1996). Interactions between
PE2, E1, and 6K required for assembly of alphaviruses studied withchimeric viruses. J. Virol. 70, 7910–7920.Copyright © 2022 FDOKUMEN




















