
Congenital brain abnormalities: Pictorial essay - JournalAgent
Upload
independentCategory
view
0download
0
Journal of General Virology (1994), 75, 97%987. Printed in Great Britain 979
Neurological abnormalities associated with feline immunodeficiency virus infection
T. R. Phil l ips, 1 0 . Prospero-Garc ia , ~ D. L. Puao i , 1 D . L. Lerner, ~ H. S. F o x , 1 R. A. Olmsted , 3 F. E. B l o o m , 1 S. J. Henriksen 1 and J. H. Elder 2
Departments of 1Neuropharmacology and 2 Molecular Biology, The Scripps Research Institute, 10666 North Torrey Pines Road, La Jolla, California 92037 and 3 Cancer and Infectious Disease Research, Up john Laboratories, Kalamazoo, Michigan, U.S.A.
Specific pathogen-free cats were infected with the Maryland strain of FIV (FIV-MD) for the purpose of assessing the effects of FIV infection on the central nervous system (CNS). Two separate studies were performed, involving a total of 13 infected cats and six age-matched, sham-inoculated controls. All animals infected with FIV-MD seroconverted by 8 weeks post- infection and virus was recovered from peripheral blood mononuclear cells of all infected cats. All of the infected animals had lower absolute CD4 + cell counts and decreased CD4+/CD8 ÷ ratios. Virus was recovered from the cerebrospinal fluid (CSF) of certain infected in- dividuals, and antiviral antibody and pleocytosis were evident in the CSF of the majority of infected cats.
Additionally, virus was recovered from tissue explants from the cerebellum, midbrain and brainstem of one sacrificed FIV + eat. Specific neurological changes in- eluded anisocoria, delayed righting reflex and delayed pupillary reflex, as well as delayed visual and auditory evoked potentials, and marked alterations in sleep patterns similar to those reported for human immuno- deficiency virus (HIV)-positive individuals. Histological evaluation revealed the presence of perivascular cuffing and glial nodules in FIV-infected cats. These results indicate that FIV causes an acute neurological disease that closely resembles the early neurological effects of HIV infection in humans and should serve well as an animal model for lentivirus-induced CNS disease.
Introduct ion
The incidence of AIDS is continually increasing through- out the world. Up to 80 % of human immunodeficiency virus (HIV)-infected people have brain lesions typical of this viral infection and up to 40% develop HIV-1- associated cognitive/motor complex (Elder & Sever, 1988). However, the underlying pathophysiology of the central nervous system (CNS) effects are not fully understood. To gain a better appreciation of the neurological aspects of the disease and to devise therapeutic approaches directed towards this form of the disease, animal models will be required.
Feline immunodeficiency virus (FIV), first described in 1987 (Pedersen et al., 1987), is known to cause a disease in cats that is very similar to human AIDS. As in HIV infection, neurological involvement is also noted with FIV infection (Pedersen et al., 1987; Dow et al., 1990; Egberink et al., 1988; Hurtrel et al., 1992; Podell et al., 1993). Dow and co-workers were the first to report that cats experimentally infected with FIV developed a productive infection of the brain when inoculated by either the intracerebral or peripheral route (Dow et al., 1990). Virus was isolated from tissue explants from the
cerebral cortex, caudate nucleus, midbrain, cerebellum, rostral and caudal brainstem but not fi'om the choroid plexus (Dow et aI., 1990). Pathological examination of two cats, at 7 months post-inoculation, revealed peri- vascular infiltrates of mononuclear cells, diffuse gliosis, glial nodules and white matter pallor. These lesions were primarily located in the caudate, midbrain and rostral brainstem (Dow et al., 1990). Primary cultures of the microglia and astrocytes were productively infected with FIV in vitro. However, similar cultures of neurons, oligodendrocytes and choroid plexus cells showed no evidence of infection. Another study examined the time course of the FIV-induced CNS lesions (Hurtrel et al., 1992) and demonstrated that CNS lesions were seen as early as 1 month post-infection when the virus was given intracerebrally, and as early as 2 months post-infection when the virus was inoculated intravenously. CNS entry for FIV, like HIV (Davis et al., 1992), is an early event, even when inoculation is peripheral (Podell et al., 1993; Wheeler et al., 1992). Thus it can be concluded from these studies that FIV infects the CNS and causes pathological lesions that are similar to HIV-induced pathology.
In addition to the neuropathological changes associ-
0001-2112 © 1994 SGM

980 T. R. Phillips and others
ated with FIV, we show that experimental infection with the Maryland strain ofFIV (FIV-MD; also known as the Mount Airy strain) produced readily observable neuro- logical function abnormalities early in the course of the infection. In this communication, we report alterations in pupil diameter, righting reflex, pupillary reflex, auditory evoked responses, visual evoked responses and sleep architecture in FIV-infected cats.
M e t h o d s
Animals. Specific pathogen-free (SPF) cats were obtained from Liberty Laboratories. Two separate studies were performed. In Study 1, six SPF 5-month-old animals were placed into either a control group (two animals) or a treatment group (four animals). The two groups of cats were communally housed in different rooms. This method of housing fosters animal interactions and helps to create a mentally stimulating environment. Study II was conducted in a manner similar to Study I, with the exception that the cats were 10 weeks old at the time of infection and that the control group consisted of four animals, whereas the FIV-inoculated group contained nine animals. Eight o f the latter group were inoculated with virus-containing tissue culture supernatant as detailed below; one cat (no. 11) received 1 ml whole blood from cat no. 1 of Study I. Seroconversion rate and disease course in cat no. 11 were similar to those observed in the other infected animals.
Virus inoculations. FIV-MD was employed in these studies. This viral strain was obtained from a young (2 years old) feline leukaemia virus (FeLV)-negative cat. This cat was severely ill with signs that are consistent with the FIV disease syndrome. The virus was isolated and amplified by co-cultivating this cat's peripheral blood mononuclear cells (PBMCs) with SPF cat PBMCs. The TCIDs0 was determined by serial dilution of the virus in SPF cat PBMCs and monitoring the dilutions for reverse transcriptase (RT) activity. The TCIDs0 was then calculated by determining the dilution at which 50 % of the cultures had RT activity (Reed & Muench, 1938). On the day before and the day of infection, both groups were injected with methylprednisolone at 4.0 mg/kg. This pretreatment of the animals with a glucocorticosteroid, prior to infection, has been used with FeLV to enhance the extent of the viral infection (Rojko et al., 1979). The infected group was intra- venously inoculated with 1000 TC1Ds0 units of the FIV-MD strain. The control group received an injection of tissue culture medium from uninfected cells.
Physical and neurological examinations. All cats received routine physical and neurological examinations approximately once a month for the duration of the studies. The neurological examination consisted of an overall assessment of their mental state, behaviour, gait and posture. Additionally, palpebral, menace, pupillary, patellar, hopping, wheel-barrowing and righting reflexes were assessed as were eye positions, lip tone, tone of the masticatory muscles and conscious proprioception (Ettinger, 1983).
Virus isolation. Ten ml of whole blood was collected by jugular puncture. The mononuclear cells were isolated over a Ficoll-Hypaque gradient as previously described (Pedersen et al., 1987). Cells were cultured in RPMI containing 10% fetal bovine serum with 1000 U of recombinant human interleukin-2 (IL-2), kindly provided by Hoffmann-La Roche. At weekly intervals, 5 ml of culture supernatant was assayed for the presence of particle-associated RT activity, as previously described (Pedersen et al., 1987). Cultures were presumed to be negative if RT activity was not detected by the sixth week of incubation. Virus isolation from the brain was performed by first placing the animal into a deep plane of anaesthesia (Ketalar, ketamine
hydrochloride, 15 mg/kg), perfusing the brain with cold saline, rapidly removing the brain and making explant cultures that were co-cultivated with SPF cat PBMCs. Cerebrospinal fluid (CSF) was obtained by cisterna magnum puncture. Virus isolation from the CSF was performed by pelleting of cells from the CSF and separately co- cultivating the CSF cells or cell-free CSF with indicator PBMCs and monitoring for RT activity.
Antibody determination. FIV antibody titre was determined by ELISA. Briefly, 50 ng of gradient-purified FIV was added to each well (96-well half-area ELISA plate, Costar). The antigen, in PBS, was allowed to dry onto the plate and was then methanol-fixed. The plate was then blocked with BLOTTO (Johnson et al., 1983) for 1 h at 37 °C. The sera were added to the plate in twofold dilutions and allowed to incubate for 1 h at 37 °C. The plate was then washed with PBS-0.05 % Tween 20. Rabbit anti-feline IgG-peroxidase conjugate (Fisher Biotech) was added at a dilution of 1 : 1000 and incubated for 1 h at 37 °C. The plate was washed, developed using 2,2'-azino-bis(3- ethylbenzthiazoline-6-sulphonic acid) chromogenic substrate (ABTS) and read at 405 nm. Specificities of the antibodies for FIV proteins were established by Western blot analysis using gradient-purified virus as target antigen, as previously described (Johnson et al., 1983).
CD4 + and CD8 + cell determinations. Lymphocyte subtyping was performed as follows. Whole blood was fractionated over a Ficoll- Paque (Pharmacia) gradient. The PBMCs were collected and washed twice with RPMI and counted. Two-million cells were taken from each specimen. One-million cells were labelled and the other million cells were used as non-labelled controls. The cells were incubated for 30 min on ice with the anti-feline CD8-biotin-conjugated antibody (Southern Biotechnology Associates). The cells were washed twice with cold Earle's balanced salt solution (EBSS). Following this wash, the cells were incubated in the dark for 30 min at 4 °C with anti-feline CD4 FITC-labelled antibody (Southern Biotechnology Associates), as well as avidin-conjugated phycoerythrin (Southern Biotechnology Associ- ates). The cells were then washed twice in EBSS and resuspended in 1 ml of EBSS with 1% formaldehyde. The percentages of CD4 + and CD8 ÷ cells were determined by fluorescence-activated cell sorting (FACS). Absolute CD4 + and CD8 + cell counts were calculated by multiplying the total lymphocyte number by the percentages of each subtype.
Auditory and visual evoked potentials. Ketamine hydrochloride-(10 mg/kg intramuscular) anaesthetized felines were aseptically fitted with subcutaneous monopolar needle electrodes (TECA Intropak DMG-50) to record brainstem auditory evoked potentials (BAEPs) and visual evoked potentials (VEPs) differentially. The two active electrodes were placed centrally near the vertex and on the midline over the frontal sinus, respectively. The ground (earth) electrode was placed in the nuchal muscles. For BAEPs, felines were then fitted with bilateral polyethylene ear tubes placed into the external auditory canal. A ' Y ' - connector attached the two tubes to the central sound source (Grass Instruments Audio Amplifier). A computer program written using National Instruments software ( 'LabView') for the Macintosh II microcomputer was used to generate the stimuli and to collect the data. The sound stimulus consisted of a 70 dB click generated by a 100 gs input to the audio amplifier. Raw signals were amplified by a Grass P- 511 instrumentation amplifier filtered between 300 and 3000 Hz and averaged using custom software operating on a Macintosh II series computer. Stimuli were delivered binaurally at a frequency of 10 per second for a total of 1024 stimuli. On-line averaging of the signals allowed us to repeat trials within a recording session to determine the stability of the evoked responses.
For VEPs, stimuli were elicited by a Grass clinical photostimulator set on the highest intensity (setting 12). Two methods were employed. First, cats were binocularly stimulated at a rate of one stimulus flash per second with the light source 30 cm from the nose. Responses to 100
FIV and neurological abnormalities 981
stimuli were averaged for each recording session. One replication was done for each experiment to evaluate stability of the evoked events. Ambient room lighting remained on. As with auditory stimulation the visual stimuli and data acquisition were controlled by a Macintosh II computer. For the second method, each eye was individually stimulated (monocular stimulation) while the other eye was manually covered by an opaque cup.
Evoked events were analysed on-line using signal averaging capa- bilities of a Macintosh II microcomputer using custom-made LabView software. Averaged peak latencies and amplitudes were calculated and compared individually between animals and as group means using ANOVA and Sheff~ post hoc statistical tests.
Sleep studies. Two felines from the first experimental group (one infected and one control) were anaesthetized with ketamine hy- drochloride, intubated and surgically prepared for sleep recording by implanting cortical stainless steel electrodes in the cranial vault. In addition, bipolar stainless screw electrodes were placed in the frontal sinus to record eye movements. Bipolar nichrome leads (250 gm in diameter) insulated except for the wire tips were placed in the nuchal musculature to monitor electromyographic activity. All leads were cemented to the cranial vault by dental cement and brought together in a connector assembly (amphenol) for later monitoring of electrical activity. Felines were given 2 weeks of recovery and subsequently acclimatized for a minimum of 3 days to an environmental chamber (BRS/LVE Large animal cubicle 89 cm height x 86 cm width x 74 cm depth) containing a 60 cmx 60 cm rug and a litter pan, prior to sleep recording. Felines were sequentially recorded for 3 consecutive days from 8 a.m. to 5 p.m. Two channels of electroencephalographic (EEG) activity were recorded on a Grass model 78 polygraph in addition to one channel each of electromyographic and eye movement activity. EEG activity was simultaneously recorded on an instrumentation tape recorder (Vetter) for later off-line analysis of EEG frequency spectra.
EEG data analysis included the initial visual inspection of the recorded channels of the EEG to determine on-going quality and appearance of spontaneous waveforms. Further analysis included the determination of latency, duration, and percentage of total sleep time of discrete sleep states. These measures were calculated and displayed off-line using computer software (Somnibus).
Histopathology. Animals were deeply anaesthetized with ketamine hydrochloride/Rompine, followed by perfusion through the left ventricle with either 4 % paraformaldehyde (one control and one FIV- infected cat), or a sterile physiological saline solution (one FIV-infected cat) drained through the right atrium. For the animals perfused with paraformaldehyde, sections of brain and spinal cord, as well as vital organs, were post-fixed in 4 % paraformaldehyde for 6 h, transferred to 70% ethanol, and embedded in paraffin. Representative samples of brain were taken aseptically for culture from the saline-perfused FIV- infected cat (see above). Other sections of brain, spinal cord and vital organs were immersion-fixed in 10% zinc-formalin for 24 h, trans- ferred to 70 % ethanol and embedded in paraffin wax. Five gm sections from each block were stained with haematoxylin/eosin, and luxol fast blue/periodic acid Schiff stains for microscopical evaluation.
Results
Two independent studies were initiated to assess the infection of the CNS by FIV. In Study I, four animals were infected with 1000 TCIDs0 units of FIV-MD and two non-contact controls were inoculated with tissue culture supernatant from uninfected cells. In study II, nine animals were infected with FIV-MD and four sham- inoculated controls were employed. All of the infected
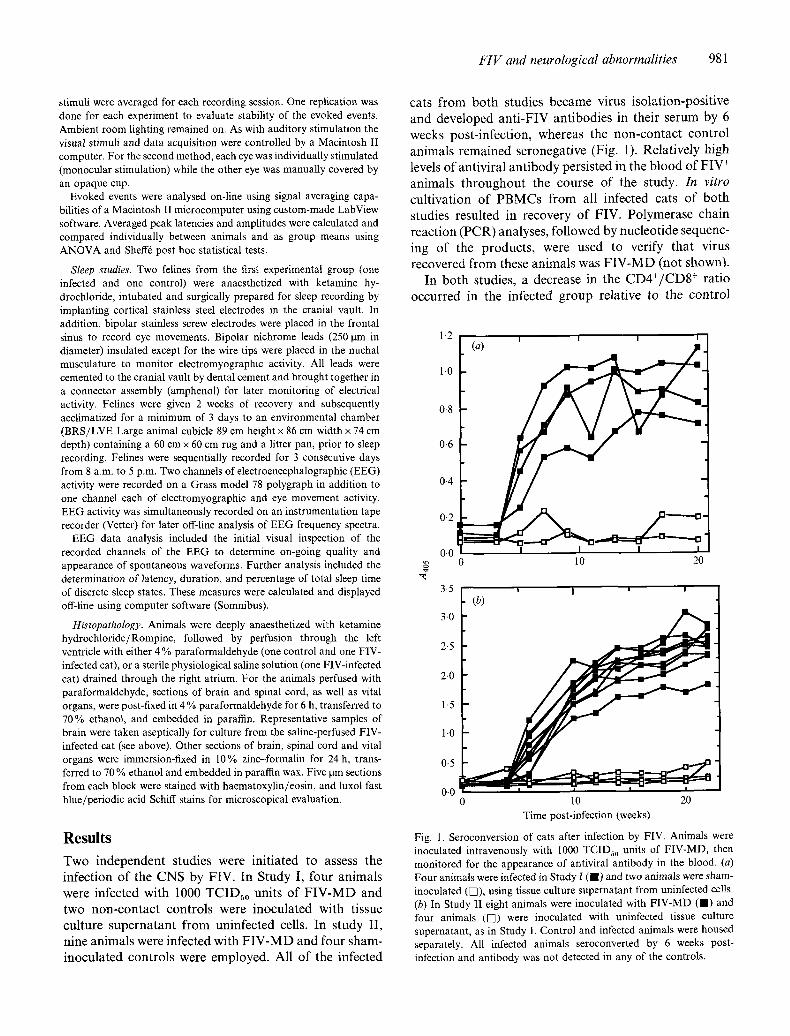
cats from both studies became virus isolation-positive and developed anti-FIV antibodies in their serum by 6 weeks post-infection, whereas the non-contact control animals remained seronegative (Fig. 1). Relatively high levels of antiviral antibody persisted in the blood of FIV + animals throughout the course of the study. In vitro cultivation of PBMCs from all infected cats of both studies resulted in recovery of FIV. Polymerase chain reaction (PCR) analyses, followed by nucleotide sequenc- ing of the products, were used to verify that virus recovered from these animals was FIV-MD (not shown).
In both studies, a decrease in the CD4+/CD8 + ratio occurred in the infected group relative to the control
(a) i
1.0
0-8
0.6
0.4
0-2 -"-'---O
0.0
0-0 i
0 10 20 Time post-infection (weeks)
Fig. 1. Seroconversion of cats after infection by FIV. Animals were inoculated intravenously with 1000 TCIDso units of FIV-MD, then monitored for the appearance of antiviral antibody in the blood. (a) Four animals were infected in Study I ( I ) and two animals were sham- inoculated (V]), using tissue culture supernatant from uninfected cells. (b) In Study II eight animals were inoculated with FIV-MD ( I ) and four animals (VI) were inoculated with uninfected tissue culture supernatant, as in Study I. Control and infected animals were housed separately. All infected animals seroconverted by 6 weeks post- infection and antibody was not detected in any of the controls.
982 T. R. Phillips and others
x
v v
0 1 3 10 13 17 25 29 Time post-infection (months)
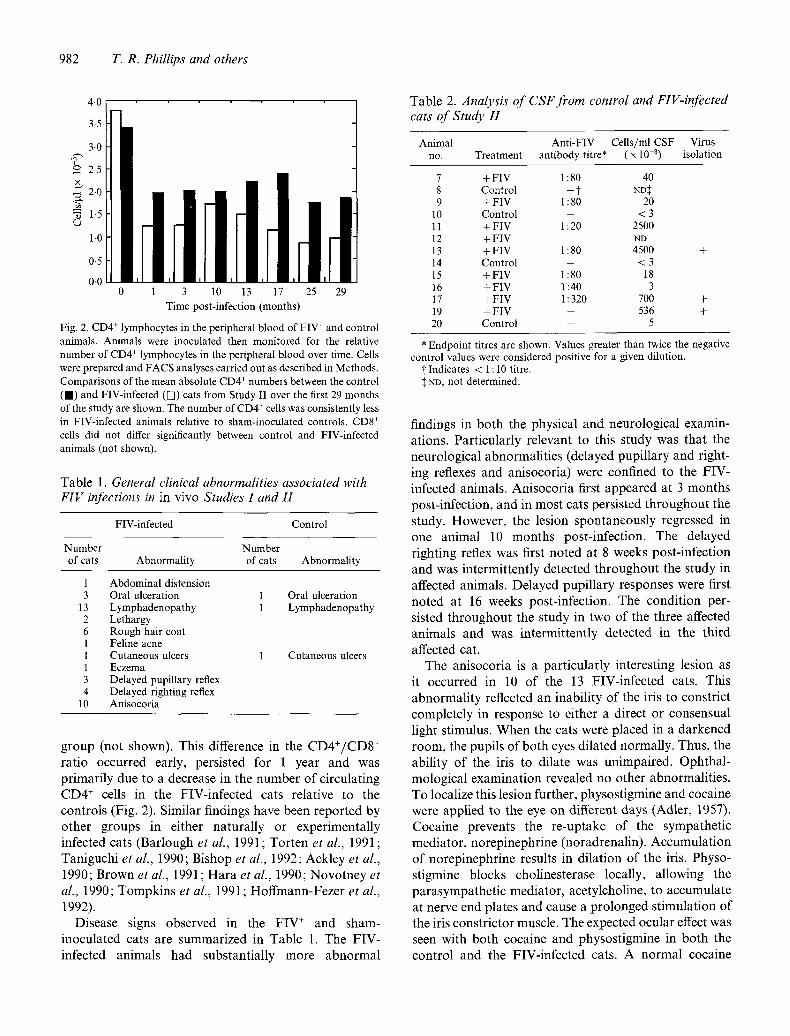
Fig. 2. CD4 + lymphocytes in the peripheral blood of FIV + and control animals. Animals were inoculated then monitored for the relative number of CD4 + lymphocytes in the peripheral blood over time. Cells were prepared and FACS analyses carried out as described in Methods. Comparisons of the mean absolute CD4 + numbers between the control ( I ) and FIV-infected ([]) cats from Study I1 over the first 29 months of the study are shown. The number of CD4 + cells was consistently less in FIV-infected animals relative to sham-inoculated controls. CD8 + cells did not differ significantly between control and FIV-infected animals (not shown).
Table 1. General clinical abnormalities associated with FIV infections in in vivo Studies I and H
FIV-infected Control
Number Number of cats Abnormality of cats Abnormality
1 Abdominal distension 3 Oral ulceration
13 Lymphadenopathy 2 Lethargy 6 Rough hair coat 1 Feline acne 1 Cutaneous ulcers 1 Eczema 3 Delayed pupillary reflex 4 Delayed righting reflex
10 Anisocoria
1 Oral ulceration 1 Lymphadenopathy
1 Cutaneous ulcers
g roup (not shown). This difference in the C D 4 + / C D 8 + ra t io occur red early, pers is ted for 1 year and was p r imar i ly due to a decrease in the number o f c i rcula t ing C D 4 + cells in the FIV- in fec ted cats relat ive to the con t ro l s (Fig. 2). S imi lar f indings have been r epor t ed by o ther g roups in ei ther na tura l ly or exper imenta l ly infected cats (Bar lough et al., 1991 ; Tor t en et al., 1991 ; Tan iguch i et al., 1990; Bishop et al., 1992; Ack ley et al., 1990; Brown et al., 1991 ; H a r a et al., 1990; N o v o t n e y et al., 1990; T o m p k i n s et al., 1991; H o f f m a n n - F e z e r et al., 1992).
Disease signs observed in the F I V + and sham- inocu la ted cats are summar i zed in Table 1. The F IV- infected animals had subs tan t ia l ly more a b n o r m a l
Table 2. Analysis of CSF from control and FIV-infected cats of Study H
Animal Anti-FIV Cells/ml CSF Virus no. Treatment antibody titre* ( x 10 a) isolation
7 + FIV 1 : 80 40 8 Control - ? ND~ 9 + FIV I : 80 20
10 Control - < 3 11 + FIV 1 : 20 2500 1 2 + F I V - ND 13 + FIV 1 : 80 4500 14 Control - < 3 15 +FIV 1:80 18 16 +FIV 1:40 3 17 + FIV 1 : 320 700 19 +FIV - 536 20 Control - 5
+
+ +
* Endpoint titres are shown. Values greater than twice the negative control values were considered positive for a given dilution.
i Indicates < 1 : 10 titre. ND, not determined.
f indings in bo th the physical and neuro log ica l examin- at ions. Par t i cu la r ly re levant to this s tudy was tha t the neuro log ica l abnormal i t i e s (de layed pup i l l a ry a n d r ight- ing reflexes and an isocor ia ) were conf ined to the F IV- infected animals . An i soco r i a first a p p e a r e d at 3 mon ths post - infect ion, and in mos t cats pers is ted t h r o u g h o u t the s tudy. However , the lesion spon taneous ly regressed in one an imal 10 mon ths pos t - infec t ion . The de layed r ight ing reflex was first no ted at 8 weeks pos t - in fec t ion and was in te rmi t ten t ly detected t h r o u g h o u t the s tudy in affected animals . De layed pup i l l a ry responses were first no ted at 16 weeks post - infect ion. The cond i t ion per- sisted t h r o u g h o u t the s tudy in two o f the three affected an imals and was in te rmi t ten t ly de tec ted in the th i rd
affected cat. The an i socor ia is a pa r t i cu la r ly interes t ing lesion as
it occur red in 10 o f the 13 FIV- in fec ted cats. This a b n o r m a l i t y reflected an inabi l i ty o f the iris to const r ic t comple te ly in response to ei ther a direct or consensua l l ight st imulus. W h e n the cats were p laced in a d a r k e n e d room, the pupi ls o f bo th eyes d i la ted normal ly . Thus, the abi l i ty o f the iris to di la te was un impai red . Oph tha l - molog ica l examina t ion revealed no o ther abnormal i t i es . To local ize this lesion fur ther , physos t igmine and cocaine were app l i ed to the eye on different days (Adler , 1957). Coca ine prevents the r e -up take o f the sympa the t i c med ia to r , no rep inephr ine (noradrenal in) . A c c u m u l a t i o n o f no rep inephr ine results in d i la t ion o f the iris. Physo- s t igmine b locks chol ines terase local ly, a l lowing the p a r a s y m p a t h e t i c media to r , acetylchol ine, to accumula te at nerve end pla tes and cause a p r o l o n g e d s t imula t ion o f the iris cons t r ic to r muscle. The expected ocula r effect was seen with bo th cocaine and physos t igmine in bo th the con t ro l and the FIV- in fec ted cats. A n o r m a l cocaine

FIV and neurological abnormalities 983
Fig. 3. Haematoxylin- and eosin-stained, paraffin-embedded sections from cat no. 12. (a) Glial nodules in parahippocampal gyrus. (b) Perivascular inflammatory infiltrate in brainstem. (c) Perivascular inflammatory infiltrate in parahippocampal gyrus.
984 T. R. Phillips and others
25 20 15 10 5 0 5
-10 15
~ - 2 0 .~ 25
25 e'~ E 20 < 15
10 5 0 5
-10 -15 -20
25
i i i I I i 1 1 i i , , i , , ,
(a) P4 .ill P3 .i~a:. P5
Stimulus :k "] I'. " 2 ' ? * ' ~ " • I P1 v ] ' t , t 4 ,
V.'" "" - ', 1 / '
" " " / ' x -. ~' \ , (C\ t . ~ !
2 4 6
(b) P3 P4.:~. P5 t,i /rn~llus
V ',./-" '1
0 2 4 6 Latency (ms)
(c) P1 P2 P3 P4 P5
Latency Araplit~de L~sl )ey Amplitude0 aV) LatenCYlms) Ampliludeip.v) L(.,s )aenc ¢unplitucle(/sV) LaleneY(ms) Amplitude(gV) Groups Y (ms) (gV)
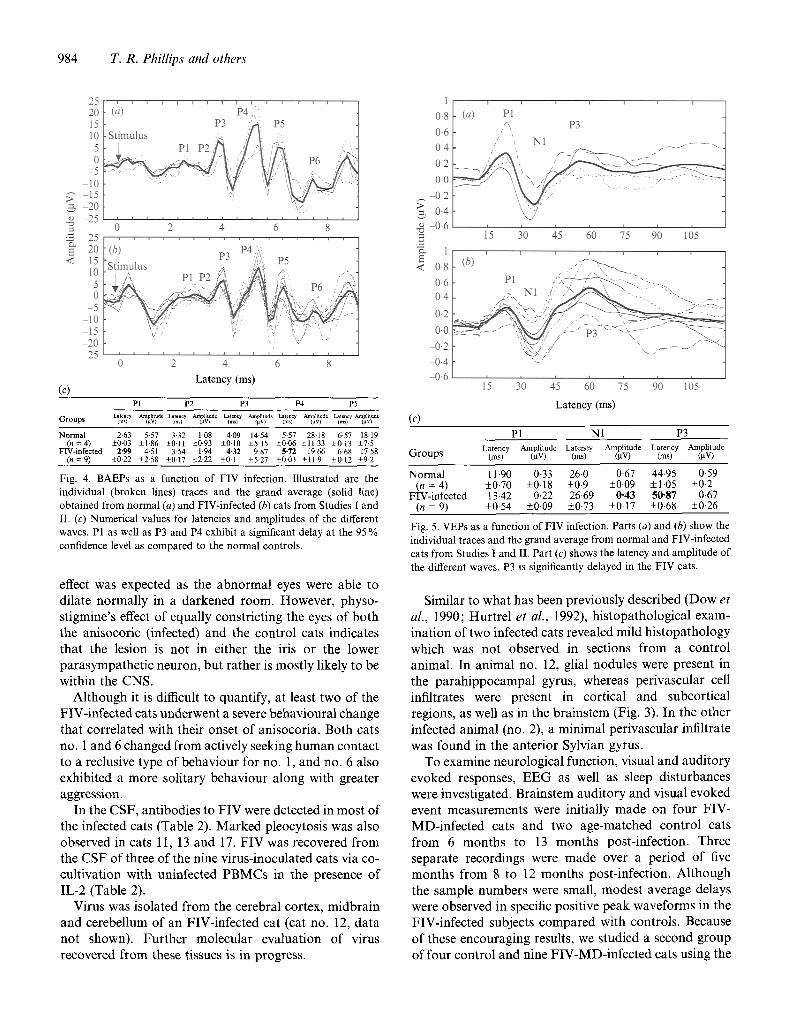
Normal 2-63 5-57 3.32 1.08 4-09 14.54 5.57 28-18 6.57 18-19 ( n = 4 ) _+0.03 _+1.86 -+0.11 _+0.93 _+0.10 -+5-15 _+0-06 _+11.33 -+0.13 _+7.5
FIV-infected 2.99 4.51 3-54 1.94 4.32 9.87 5-72 19.66 6.68 1758 ( n = 9 ) _-4-0.22 _+2.58 -+0.17 _+2.22 -+0.1 -+5.27 _+0-03 _+11-9 -+0.12 _+9.2
Fig. 4. BAEPs as a function of FIV infection. Illustrated are the individual (broken lines) traces and the grand average (solid line) obtained from normal (a) and FIV-infected (b) cats from Studies 1 and II. (c) Numerical values for latencies and amplitudes of the different waves. P1 as well as P3 and P4 exhibit a significant delay at the 95 % confidence level as compared to the normal controls.
effect was expected as the abnormal eyes were able to dilate normal ly in a darkened room. However , physo- stigmine's effect o f equally constricting the eyes o f bo th the anisocoric (infected) and the control cats indicates that the lesion is not in either the iris or the lower parasympathet ic neuron, but rather is most ly likely to be within the CNS.
Al though it is difficult to quantify, at least two of the FIV-infected cats underwent a severe behavioural change that correlated with their onset o f anisocoria. Both cats no. 1 and 6 changed f rom actively seeking h u m a n contact to a reclusive type o f behaviour for no. 1, and no. 6 also exhibited a more solitary behaviour along with greater aggression.
In the CSF, antibodies to F I V were detected in mos t o f the infected cats (Table 2). Marked pleocytosis was also observed in cats 11, 13 and 17. F IV was recovered f rom the CSF of three o f the nine virus-inoculated cats via co- cult ivation with uninfected P B M C s in the presence o f IL-2 (Table 2).
Virus was isolated f rom the cerebral cortex, midbrain and cerebellum o f an FIV-infected cat (cat no. 12, da ta not shown). Fur ther molecular evaluat ion o f virus recovered f rom these tissues is in progress.
1
0.8 0.6 0.4 0-2 0-0
,--, 4)-2
.~ 4).6 ,= c~ 1
< 0-8 0.6 0.4
0.2 0.0
4).2 M).4~
4)-6
t j i i J
(a) P1 /G P3
L / i
;5 30 45 ;0 45 910 165 i i t i t i i
(b)
P1 : , ~ ~ - ' - , "- ...
' 3'0 ' ' ' 9'0 6 5 15 45 60 75 Latency (ms)
(c) P1 N1 P3
Latency Amplitude Latency Amplitude Latency Amplitude Groups (ms) (gv) (ms) (~v) (ms) (gv)
Normal 11.90 0-33 26-0 0.67 44.95 049 (n = 4) +_0.70 +0.18 +0.9 +_0.09 +1.05 +_0.2
FIV-infected 13-42 0.22 26.69 0-43 50-87 0-67 (n = 9) _+0.54 _+0-09 _+0-73 _+0.17 _+0.68 _+0-26
Fig. 5. VEPs as a function of FIV infection. Parts (a) and (b) show the individual traces and the grand average from normal and FIV-infected cats from Studies I and II. Part (c) shows the latency and amplitude of the different waves. P3 is significantly delayed in the FIV cats.
Similar to what has been previously described (Dow et al., 1990; Hurtrel et al., 1992), his topathological exam- inat ion o f two infected cats revealed mild h is topathology which was no t observed in sections f rom a control animal. In animal no. 12, glial nodules were present in the pa rah ippocampal gyrus, whereas perivascular cell infiltrates were present in cortical and subcortical regions, as well as in the brainstem (Fig. 3). In the o ther infected animal (no. 2), a minimal perivascular infiltrate was found in the anterior Sylvian gyrus.
To examine neurological function, visual and audi tory evoked responses, E E G as well as sleep disturbances were investigated. Brainstem audi tory and visual evoked event measurements were initially made on four FIV- MD-infected cats and two age-matched control cats f rom 6 months to 13 months post-infection. Three separate recordings were made over a period o f five months f rom 8 to 12 months post-infection. Al though the sample numbers were small, modest average delays were observed in specific positive peak waveforms in the FIV-infected subjects compared with controls. Because o f these encouraging results, we studied a second group of four control and nine F I ¥ - M D - i n f e c t e d cats using the

F I V and neurological abnormali t ies 985
Wake 1
2 REM l
9 10 11 12 13 14 15 16
Wake 1
2 REM
i
t _ 7 , ~-AI
17 8 9 1'0 1'1 1'2 13 14 15 16
Time (11)
Fig. 6. Illustration of the sleep architecture in one normal (no. 3) and one FIV-infected (no. 1) cat. Latency for sleep onset is substantially increased. Slow-wave sleep 2 is augmented, whereas rapid eye movement (REM) sleep frequency is reduced.
methods described above. These tests were conducted initially 6 months post-inoculation. Fig. 4 and 5 illustrate the electrophysiological findings in this new cohort of subjects. Similar to the pilot investigation, significant delays were observed in specific components of the FIV- MD-infected cats' auditory and visual evoked responses compared to those of controls. Particularly striking is the delay in the auditory 'P3 ' and 'P4 ' waveforms of the FIV-infected cats compared to controls (Fig. 4) and the visually evoked 'P3 ' wave in the FIV-infected group (Fig. 5). Similar findings have recently been reported by Podell et al. (1993).
In addition to the evoked potential studies, one control cat and one FIV-infected cat were surgically implanted with permanent nichrome screw electrodes to assess further the potential EEG abnormalities and sleep-wake changes previously observed in human HIV-infected subjects (St. Kubicki et al., 1988; Norman et al., 1988, 1990, 1992). Nine electrodes were placed in the skull and nuchal musculature and were mounted in an acrylic headmount to monitor behavioural state. Preliminary analysis of the raw EEG did not suggest major differences in the two cats. However, sleep-related slowing of the dominant EEG frequencies was greater in the FIV- infected animal. Analysis of 8 h sleep recordings from the two subjects revealed dramatic changes in sleep para- meters in the FIV-infected cat compared to the control. Fig. 6 illustrates the changes in the sleep hypnogram in the control and FIV-infected cats for one representative 8 h sleep recording. The architectural changes in sleep in the FIV-infected cat mimic those reported for HIV- infected humans and include an overall disruption in
sleep architecture and displacement of slow-wave sleep (SWS) toward the later portion of the recording period.
Discussion The findings of this study indicate that FIV infection of the feline CNS closely parallels the effects that HIV exerts on the human nervous system. In this study, we show that FIV produces a neurological disease early in the course of infection. Examination of the infected cats revealed numerous functional abnormalities including delayed righting and pupillary reflexes, anisocoria, altered visual and auditory evoked potentials and abnormal sleep patterns. These findings are consistent with those described for HIV infection in humans which can cause alterations in both visual (Malessa et al., 1989) and auditory (Goodin et al., 1990; Boccellari et al., 1993; Smith et al., 1988; Pagano et al., 1992; Rosenhall et al., 1989; Koralnik et al., 1990; Jabbari et al., 1993) evoked potentials. In addition, abnormal sleep patterns, similar to the findings of this study, have also been reported in HIV-infected individuals (Aldrich et al., 1988; St. Kubicki et al., 1988; Rothenberg et at., 1990; Norman et al., 1988, 1990, 1992). Finally, decreased coordination and slowed reflexes are common in HIV-infected indi- viduals (Karlsen et al., 1992; Orefice et al., 1991). Thus, it would appear that FIV infection of the feline CNS causes neurological dysfunction that is similar to the effects HIV exerts on the human nervous system (also see Podell et al., 1993).
Although anisocoria has been associated with HIV (Poutiainen et al., 1991) and FIV infection (Hopper et al., 1989), it has not been a predominantly reported clinical finding with either viral infection. The high incidence of anisocoria observed in this study may reflect a viral strain predilection to cause anisocoria or possibly a genetic susceptibility of the cats used in this study to develop this lesion. An alternative possibility is that anisocoria may have been overlooked in a number of studies and thus the incidence with HIV infection may be higher. Also, since we observed reversion of the lesion in some individuals in the late acute phase, it may be that this abnormality reverts in a number of HIV-infected individuals prior to close examination by a physician.
FIV produced detectable neurological abnormalities early during the infection. At least some of these altered parameters were reversible. However, no reversion of the delayed auditory and visual evoked potentials has been observed in our studies to date. Thus, these indices remain consistent for the monitoring of FIV infection of the CNS. However, the demonstration that some of the neurological functions studied returned to normal and that there is minimal histopathology early in the course of the disease indicates that if effective therapy were

986 T. R. Phillips and others
initiated early in the disease process, prior to permanent impairment, that neurological dysfunction may be prevented or possibly returned to normal. Additional evidence to support this assertion is that 3'-azido-3'- deoxythymidine (AZT) has been shown to have an ameliorating effect on the HIV-induced CNS pathology (Pizzo et aL, 1988; DeCarli et al., 1991). Thus, the results of these studies indicate that FIV is a good model for examining the early neurological effects of lentiviruses and that FIV may serve as an effective system to test therapeutic approaches that are directed toward the neurological form of the disease.
The authors wish to thank Drs M. B. A. Oldstone and M. Buchmeier for advice and encouragement. The excellent veterinary assistance of Dr Elizabeth Ford is also acknowledged. We thank Hoffman-LaRoche for the generous donation of IL-2 used in this study. This work was supported by grant 1P50 MH47680 from the National Institute of Mental Health.
References ACKLEY, C.D., YAMAMOTO, J.K., LEVY, N., PEDERSEN, M.C. &
COOPER, M. D. (1990). Immunologic abnormalities in pathogen-free cats experimentally infected with feline immunodeficiency virus. Journal of Virology 64, 5652-5655.
ADLER, F.H. (1957). The eyelids. In Gifford's Textbook of Oph- thalmology, pp. 173-175. Philadelphia: W. B. Saunders.
ALDRICH, M.S., ROGERS, A.E. & ANGELL, K. (1988). Excessive daytime sleepiness as a presenting manifestation of HIV infection. Sleep Research 17, 137.
BARLOUGH, J. E., ACKLEY, C. D., GEORGE, J. W., LEVY, N., ACEVEDO, R., MOOP, E, P.F., RIDEOUT, B.A., COOPER, M.D. & PEDERSEN, N.C. (1991). Acquired immune dysfunction in cats with exper- imentally induced feline immunodeficiency virus infection: com- parison of short-term and long-term infections. Journal of Acquired Immune Deficiency Syndromes 4, 219-227.
BISHOP, S.A., WILLIAMS, N.A., GRUTFYDD-JONES, T.J., HARBOUR, D.A. & STOKES, C.R. (1992). Impaired T-cell priming and proliferation in cats infected with feline immunodeficiency virus. AIDS 6, 287 293.
BOCCELLARI, A.A., DILLEY, M.O., YINGLING, C.D., CHAMBERS, D.B., TAUBER, M.A., MOSS, A.R. & OSMOND, D.H. (1993). Relationships of CD4 counts to neurophysiological function in HIV- 1-infected homosexual men. Archives of Neurology 50, 517-521.
BROWN, W. C., BISSEY, L., LOGAN, K. S., PEDERSEN, N.C., ELDER, J.H. & COLLISSON, E.W. (1991). Feline immunodeficiency virus infects both CD4 + and CD8 + T lymphocytes. Journal of Virology 65, 3359-3364.
DAVIS, L. E., HJELLE, B. L., MILLER, V. E., PALMER, D. L., LLEWELLYN, A. L., MERLIN, T. L., YOUNG, S. A., MILLS, R. G., WACHSMAN, W. & WILEY, C.A. (1992). Early viral brain invasion in iatrogenic human immunodeficiency virus infection. Neurology 42, 173(~1739.
DECARLI, C., FUGATE, L., FALLOON, J., EDDY, J., KATZ, D.A., FRIEDLAND, R. P., RAPOPORT, S. I., BROUWERS, P. & P1ZZO, P. A. (1991). Brain growth and cognitive improvement in children with human immunodeficiency virus-induced encephalopathy after 6 months of continuous infusion zidovudine therapy. Journal of Acquired Immune Deficiency Syndromes 4, 585-592.
Dow, S.W., Poss, M.L. & HOOVER, E.A. (1990). Feline immuno- deficiency virus: a neurotropic lentivirus. Journal ofA cquired Immune Deficiency Syndromes 3, 658-668.
EGBERINK, H., EDERVEEN, J., KOOLEN, M., LUTZ, H. & HORZINEK, M.C. (1988). Infections with feline T-lymphotropic lentivirus. Tijdsehrift voor Diergeneeskunde 113, 93%943.
ELDER, G. A. & SEVER, J. L. (1988). AIDS and neurological disorders: an overview. Annals of Neurology 23, Supplement $4-6.
ETTINGER, S.J. (1983). Textbook of Veterinary Internal Medicine: Diseases of the Dog and Cat, 2nd edn. Philadelphia: W. B. Saunders.
GOODIN, D. S., AMINOFF, M. J., CHERNOFF, D. N. & HOLLANDER, H. (1990). Long latency event-related potentials in patients infected with human immunodeficiency virus. Annals of Neurology 27, 414-419.
HARA, Y., ISHIDA, T , EJIMA, H., TAGAWA, M., MOTOYOSHI, S., TOMODA, I., SHIM1ZU, M. & SHICHINOHE, K. (1990). Decrease in mitogen-induced lymphocyte proliferative responses in cats infected with feline immunodeficiency virus. Nippon Juigaku Zasshi 52, 573 579.
HOEFMANN-FEZER, G., THUM, J., ACKLEY, C., HERBOLD, M., MYSLI- WIETZ, J., THEFELD, S, HARTMANN, K. & K.RAFT, W. (1992). Decline in CD4 ÷ cell numbers in cats with naturally acquired feline immunodeficiency virus infection. Journal of Virology 66, 1484-1488.
HOPPER, C.D., SPARKES, A.H., GRUFEYDD-JONES, T.J., CRISPIN, S. M., MUIR, P., HARBOtJR, D. A. & STOKES, C. R. (1989). Clinical and laboratory findings in cats infected with feline immunodeficiency virus. Veterinary Record 125, 341 346.
HURTREL, M., GANIERE, J. P., GUELFI, J. E., CHAKRABARTI, L., MAIRE, M.A., GRAY, F., MONTAGNIER, L. & HLrRTREL, B. (1992). Com- parison of early and late feline immunodeficiency virus encephalo- pathies. AIDS 6, 399-406.
JABBARI, l . , COATS, M., SALAZAR, A., MARTIN, A., SCHEROKMAN, I . & LAWS, W. (1993). Longitudinal study of LEG and evoked potentials in neurologically asymptomatic HIV infected subjects. Electro- encephalogr aph y and Clinical Neur ophysiology 86, 145-151.
JOHNSON, D.A., GAUTSCH, J.W., SPORTSMAN, R. & ELDER, J.H. (1983). Improved technique utilizing non-fat dry milk for analysis of protein and nucleic acids transferred to nitrocellulose. Gene Analysis Techniques 1, 3-7.
KARLSEN, N.R., REINVANG, I. & FROLAND, S.S. (1992). Slowed reaction time in asymptomatic HIV-positive patients. Acta Neuro- logica Scandinavica 86, 242-246.
KORALNIK, I.J., BEAUMANOIR, R., HAUSLER, A., KOHLER, SAFRAN, A.B., DELACOUX, R., VIBERT, D., MAYER, E., BURKHARD, P., NAHORY, A., MAGISTRIS, M., SANCHES, J., MYERS, P., PACCOLAT, F., QUOEX, F., GABRIEL, V., PERRrN, L., MERMILLOD, B., GAUTHIER, G., WALDVOGEL, F. & HIRSHEL, B. (1990). A controlled study of early neurologic abnormalities in men with asymptomatic human immuno- deficiency virus infection. New England Journal of Medicine 323, 864-870.
MALESSA, R., HEUSER, L. & BROCKENM~YER, N. (1989). Evoked potentials in neurologically asymptomatic persons during the early stages of HIV infection. EEG-EMG 20, 257-266.
NORMAN, S. E., RESNICK, L., COHN, M. A., DUARA, R., HERBST, J. & BERGER, J. R. (1988). Sleep disturbances in HIV-seropositive patients. Journal of the American Medical Association 260, 922.
NORMAN, S. E., CHEDIAK, A. D., FREEMAN, C., KtEL, M., MENDEZ, A., DUNCAN, R., SIMONEAU, J. & NOLAN, B. (1992). Sleep disturbances in men with asymptomatic human immunodeficiency (HIV) in- fection. Sleep 15, 150-155.
NORMAN, S., DEMIROZC, M. & CrIEDIAK, A. (1990). Disturbed sleep architecture (NREMIREM sleep cycles) in HIV-infected healthy men. Sleep Research 19, 339.
NOVOTNEY, C., ENGLISH, R.V., HOUSMAN, J., DAVIDSON, M.G., NASISSE, M.P., JENG, C.R., DAVIS, W.C. & TOMPKINS, M.B. (1990). Lymphocyte population changes in cats naturally infected with feline immunodeficiency virus. AIDS 4, 1213 1218.
OREEICE, G., TROISI, E., CARRIERI, P. B., CHIRIANNI, A., DEMARINIS, T., MARASCO, G., RUB1NO, S. & CAMPANELLA, G. (1991). Archaic reflexes in neurologically asymptomatic HIV positive subjects. International Conference on AIDS 7, 184 (abstract no. M.B.2010).
PAGANO, M. A., CAHN, P. E., GARAU, M. L., MANGONE, C. A., FIGINI, H.A., YORIO, A. A., DELLEPIANE, M. C., AMORES, M. G., PEREZ, H. M. & CASIRO, A. D. (1992). Brain-stem auditory evoked poten- tials in human immunodeficiency virus sero-positive patients with and without acquired immunodeficiency syndrome. Archives of Neurology 49, 16(~169.
PEDERSEN, N. C., HO, E. W., BROWN, M. L. & YAMAMOTO, J. K. (1987). Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science 235, 790-793.
PIzzo, P. A., EDDY, J., FALLOON, J., BALIS, F. M., MURPHY, R. F.,

FIV and neurological abnormalities 987
Moss, H., WOLTERS, P., BROUWERS, P., JAROSINSKI, P., RUBIN, M., BRODER, S., YARCHOAN, R., BRUNETTI, A., MAHA, M., NUSINOFF- LERHMAN, S. & POPLACK, D. (1988). Effect of continuous intravenous infusion of zidovudine (AZT) in children with symptomatic HIV infection. New England Journal of Medicine 319, 889-896.
PODELL, M., OGLESBEE, M., MATI-IES, L., KRAKOWKA, S., OLMSTED, R. & LAFRADO, L. (1993). AIDS-associated encephalopathy with experimental feline immunodeficiency virus infection. Journal of Acquired Immune Deficiency Syndromes 6, 758-771.
POUTIAINEN, E., HALTIA, M., ELOVAARA, I., LAHDEVIRTA, J. & IIVAnAINEN, M. (1991). Dementia associated with human immuno- deficiency virus: subcortical or cortical? Acta Psychiatrica Scandin- avica 83, 297-301.
REED, L. J. & Mtn~NCH, H. (1938). A simple method of estimating fifty percent endpoints. American Journal of Hygiene 27, 493~,97.
ROJKO, J. L., HOOVER, E. A., MATHES, L. E., KRAKOWKA, S. t~; OLSEN, R.G. (1979). Influence of adrenal corticosteroids on the sus- ceptibility of cats to feline leukemia virus infection. Cancer Research 39, 3789 3791.
ROSENHALL, V. C., HAKANSSON, G. B., LOWHAGEN, P., HANNER, P. & JONSSON, F. K. (1989). Otoneurological abnormalities in asympto- matic HIV seropositive patients. Acta Neurologica Scandinavica 79, 140-145.
ROTHENBERG, S., ZOZULA, R., FUNESTI, J. & McAULIFFE, V. (1990). Sleep habits in asymptomatic HIV-seropositive men. Sleep Research 19, 342.
SMITH, T. J., JAKOBSEN, J., GAUB, S., HELWIG-LARSEN, S. tf¢~ TROJABORG, W. (1988). Clinical and electrophysiological studies of human immunodeficiency virus-seropositive men without AIDS. Annals of Neurology 23, 295-297.
ST. KUBICKI, H., HENKES, H., TERSTEGGE, K., TERSTEGGE, K & RUF, B. (1988). AIDS related sleep disturbances a preliminary report. HIV and the Nervous System, pp. 97-105. Edited by H. St Kubicki, H. Henkes, U. Bienzle & H. D. Pohle. Stuttgart: Gustav Fischer.
TANIGUCHI, A., ISHIDA, T., WASmZtJ, T. & TOMODA, I. (1990). Humoral immune response to T cell dependent and independent antigens in cats infected with feline immunodeficiency virus. Journal of Veterinary Medical Science 53, 333-335.
TOMPKINS, M. B., NELSON, P.D., ENGLISH, R.V. & NOVOTNEY, C. (1991). Early events in the immunopathogenesis of feline retrovirus infections. Journal of the American Veterinary Medical Association 199, 1311-1315.
TORTEN, M., FRANCHINI, M., BARLOUGH, J. E., GEORGE, J. W., MOZES, E., LUTZ, H. & PZDERSEN, N.C. (1991). Progressive immune dysfunction in cats experimentally infected with feline immuno- deficiency virus. Journal of Virology 65, 2225-2230.
WHEELER, D.W., MITCHELL, T.W., GASPER, P. W., BARR, M.C. & WHALEN, L. R. (1992). Feline Immunodeficiency Virus. Proceedings of an International Conference. Davis, California, 4 to 7 September, 1991. Veterinary Immunology and lmmunopathotogy 35, t-224.
(Received 24 September 1993 ; Accepted 13 December 1993)
Copyright © 2022 FDOKUMEN




















