
All Trypanosoma cruzi developmental forms present lysosome-related organelles

Molecular and Biochemical Parasitology, 14 (1985) 151-163 151 Elsevier
MBP 00507
T H E E F F E C T O F P R O T E O L Y T I C E N Z Y M E S A N D P R O T E A S E I N H I B I T O R S
O N T H E I N T E R A C T I O N T R Y P A N O S O M A C R U Z ? - F I B R O B L A S T S
MARTA M. PIRAS, DIANA HENRIQUEZ and ROMANO PIRAS. With the technical assistance of STEFANIA NEGRI
Unidad de lnvestigaciones, Centro Medico Docente La Trinidad, A.P. 80474, Caracas 1080-A, Venezuela
(Received 29 May 1984; accepted 14 September 1984)
It has been shown previously that the capability to adhere to and infect fibroblastic cells by Trypanosoma cruzi is expressed only partially in trypomastigotes recently liberated from infected fibroblasts, but these parasites can increase several-fold their adhesion and infectivity by a time-dependent extracellular incuba- tion. It is now shown that polyacrylamide gel electrophoresis patterns of 12SI-labelled surface proteins of the parasites change during the activation process and that protease inhibitors of diverse specificity can block both these changes and the development of adhesion and infectivity. Treatment of fresh trypomastigotes with different proteases increases immediately adhesion and infection. The effect of trypsin has been studied in detail and it was found that this protease stimulates adhesion 4- to 6-fold, even in trypomastigotes obtained and assayed in the absence of serum. Trypomastigotes incubated for various periods and then exposed to trypsin increase their adhesion to values similar to those attained by prolonged incubation of trypomastigotes alone, but infection is stimulated in fresh trypomastigotes only. Trypomastigotes whose development of activation has been inhibited either by (a) protease inhibitors, (b) puromycin, and (c) tunicamycin, and are thereafter trypsinized, show respectively, that: (a) adhesion and infection are restored immediately to the same high values obtained when untreated controls are trypsinized, (b) adhesion is restored, but not infection, and (c) infection is not restored. These results suggest that the adhesion step of T. cruzi trypomastigotes to fibroblastic cells depends on a membrane protein(s) that is (are) already present in an inactive or hidden form in parasites recently liberated from infected fibroblasts. Upon extracellular maturation of these trypomastigotes this proteins(s) is activated or unmasked, probably through an endogenous proteolytic process, whose expression requires protein synthesis. The penetration step requires biosynthesis of a tunicamycin-sensitive glycoprotein(s) of the parasite and its full expression necessitates serum.
Key words: Trypanosoma cruzi; Trypomastigotes; Adhesion; Attachment; Infection; Invasion; Protease; Host-parasite interplay; Trypsin
Abbreviations: MEM, Eagles's minimal essential medium; PBS, Dulbecco's phosphate-buffered saline; FCS, fetal calf serum; SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis; PMSF, phenylmethyl sulphonylfluoride; TLCK, N-a-p-tosyI-L-lysine chloromethyl ketone.
0166-6851/85/$03.30 © 1985 Elsevier Science Publishers B.V. (Biomedical Division)

152
INTRODUCTION
In vitro infection of fibroblastic cells by cell-culture-derived trypomastigotes of Trypanosoma cruzi has been shown to occur only to a limited extent if the parasites are used as soon as they first burst out of infected cells. Extracellular incubation of these recently liberated trypomastigotes brings about, in a time-dependent manner, an increase of the adhesive and infective capabilities of these parasites [ 1]. Puromycin can block this development of adhesion and infectivity, but tunicamycin inhibits the latter only, suggesting that adhesion and penetration are distinct events of the infective process, and that a glycoprotein(s) is probably involved in the penetration step [2].
In order to obtain additional information on the mechanism of the extracellular activation process of T. cruzi trypomastigotes and on the nature of the membrane components involved in adhesion and penetration, several exogenous modifications of the surface membrane of the parasites have been employed [3,4]. The results reported here, using trypsin as a modifying agent, suggest that the adhesion step of cell- culture-derived trypomastigotes to fibroblasts depends on a membrane protein(s) that is already present in an inactive or hidden form in parasites recently liberated from infected fibroblasts. Upon extracellular maturation of the trypomastigotes, this pro- tein(s) is activated or unmasked, probably through an endogenous proteolytic process, whose expression requires protein synthesis. As to the penetration step, this requires biosynthesis of a tunicamycin-sensitive glycoprotein(s) of the parasite and its full expression necessitates serum. Preliminary accounts of these results have been presented [4-6].
MATERIALS AND METHODS
Parasites. Trypomastigotes of T. cruzi (EP strain) recently liberated from infected monolayers of Vero cells were used routinely ('fresh trypomastigotes'). The parasites were obtained as already described [1] and purified by gravity filtration through a Whatman No. 1 filter paper to remove cell debris and loose Vero cells that sometimes are present. Trypomastigotes were incubated in 'complete medium' (MEM/10% FCS/100 units per ml penicillin/100 lag per ml streptomycin) at 37°C to produce, when indicated, their activation [1].
Proteases, protease inhibitors and reagents. Proteases and protease inhibitors were purchased from Sigma Chemical Co. Proteases were dissolved at 1 mg per ml in phosphate-buffered saline (PBS) (Ca 2÷ and Mg 2+ free) and aliquots used to produce a 60 lag per ml final concentration. Stock solutions of protein inhibitors (1 mg per ml) were in water except for pepstatin, N-ct-p-tosyl-L-lysine chloromethyl ketone (TLCK), and phenylmethylsulfonyl fluoride (PMSF), which were in dimethylsulfoxide. Tuni- camycin was kindly supplied by Eli Lilly and iodogen (1,3,4,6-tetrachloro-3ct, 6et-di- phenyl glycoluril) was bought from Pierce.

153
Adhesion and infection assays. They were carried out as already described in detail [ 1,2]. Briefly, parasites were interacted (at 0°C for adhesion and at 37°C for infection) for 2 h with Vero cell monolayers on glass coverslips; at the end of the incubation the coverslips were washed extensively to remove free parasites, the cells and parasites stained, and scoring done under 1 000 X magnification. The adhesion and infection indexes express the average number of extracellular and intracellular parasites per cell, respectively.
Parasite trypsinization. Parasites were washed twice by centrifugation at 300 X g for 10 min in PBS (Ca 2÷ and Mg 2÷ free) and finally resuspended at 50 X 106 per ml. To 2 ml of this suspension were added 50 lal of 2.5 mg per ml trypsin (Difco 1:250) and the mixture incubated at 37°C for 30 min. Soy bean trypsin inhibitor (2 mg) or 2 ml complete medium were added, the parasites washed twice by centrifugation and then resuspended in complete medium. Unless otherwise stated, this was the standard procedure used for trypsinization.
Parasite iodination. Surface membrane proteins of the parasite were radioiodinated by the iodogen procedure [7,8]. Parasites (100 X 106), washed twice by centrifugation (300 × g for 10 min) in PBS, were resuspended in 1 ml of this solution and added to an iodogen-coated (0.2 mg) glass test tube. Carrier-free 1251 (120 ~tCi from New England Nuclear, NEZ-033A) was then added and the reaction allowed to proceed for 5 min at room temperature with constant agitation. After the parasites were transferred to a small plastic conical tube, 1 ml of 5 mM NaI in PBS was added, the parasites centrifuged and used.
Polyacrylamide gel electrophoresis. Aliquots of the parasites were treated with 0.2% (v/v) of Nonidet P-40 in PBS/2 mM TLCK/2 mM PMSF, for 15 min at room temperature. The suspension was then centrifuged (400 × g for 10 min) and the supernatant mixed with an equal volume of a 2 X 'sample buffer' (0.1 M Tris-HC1, pH 6.8/2% (w/v) SDS/0.25 M mercaptoethanol/0.002% bromophenol blue), heated for 2 min at 100°C and then run on a Laemmli [9] discontinuous gel system. Slabs of 1.5 mm thickness of polyacrylamide (4% stacking gel and 8.8 to 16.2% gradient for the separating gel, respectively) were used and run with 0.1% (w/v) SDS/0.39 M glycine- Tris buffer, pH 8.3. The gels were dried (Bio-Rad, model 224) and then autoradiogra- phed at -70°C with a Kodak-X-Omat R-1 film using Kronex Lighting Plus (DuPont) intensifier screens [10].
The results reported in this paper are from one representative experiment, out of 2-4 carried out for each condition, unless otherwise stated. The points for each experiment are always the mean of duplicates, which usually agree within 10%.

154
RESULTS
We have previously shown [6] that extracellular incubat ion of T. cruzi t rypomast i -
gotes for 20 h brings about , in addit ion to a st imulation of parasite adhesion and
infectivity, a change in the P A G E pattern of the T. cruzi surface proteins which can be
labelled with [125I]iodogen. The modificat ions of the pat tern occurred gradually and
concomitant ly with the changes in adhesion and infectivity and consisted mainly in a
decrease of high molecular weight polypeptides, suggesting that this might be the
consequence o f a proteolytic process. Hence, the effect of exogenous protease inhibit-
ors and proteases on this parameter , as well as on the development o f adhesion and
infectivity, was now explored in detail. The results obtained show that, indeed,
protease inhibitors tend to preserve the original polypeptide pattern (Fig. 1), which
shows the same prominent bands reported by Zingales et al. [11]. Protease inhibitors
block also the development o f adhesion and infectivity by approximately 80-95%,
when they are present during the activation process, either individually or as a mixture
1 1 0 "
,~ 9 0 - m C 0
6 0 -
v t . J
r,.
3 0 , , ,
i i o N D D
To T; Fig. 1. Autoradiographs of surface membrane proteins of T. cruzi labelled with [~251]iodogen and separated on SDS-PAGE. Fresh trypomastigotes (To) were labeled and then chased for 7 h in the absence (T7) and presence (T~) of a mixture of protease inhibitors (see Table I). For other experimental details see Materials and Methods.

155
(Table I). The effect is produced by inhibitors of different specificity and is dose dependent: aprotinin, for instance, produces 50% inhibition at 5 lag/ml (not shown). When fresh trypomastigotes were subjected to a mild treatment with exogenous proteases of various specificity, adhesion and infectivity of the parasites were drastic- ally altered (Table II). With the exception of pepsin, all the other proteases tested brought about a stimulation of adhesion and infectivity; trypsin gave usually the highest stimulation (4- to 6-fold for adhesion and 2-fold for infectivity) and therefore the effect of this protease was further investigated. Fig. 2 shows that activation of infectivity can be obtained treating fresh trypomastigotes with concentrations of trypsin differing as much as two orders or magnitude (0.06-6 lag/ml). However, when high concentrations are used the effect is small and, more important, if the parasites have been already activated, trypsin can actually decrease the infectivity, unless very low concentrations are employed (Fig. 2B). Thus, it is not unlikely that a decrease of infectivity could be observed, unless recently released trypomastigotes and the proper trypsin concentration and incubation time (30-60 min) are used. As already shown [6], trypsinized trypomastigotes have a PAGE pattern similar to that of 20 h activated parasites. The stimulation of adhesion by trypsinization of trypomastigotes that arise from cells cultured in serum-free media is observed not only in the presence of FCS (standard assay condition), but also in the absence of serum (Table III). Infectivity of
TABLE I
The effect of different protease inhibitors on the development of adhesion and infectivity of T. cruzi
trypomastigotes.
Additions Parasites per cell
Adhesion index Infection index
None 2.2 3.6
Aprotinin 0.9 1.3
Leupeptin 1.0 1.6
Pepstatin 1.4 2.2
Chymostat in 0.9 1.8
Trypsin inhibitor (soy bean) 0.9 1.9
Antipain 1.0 1.8
PMSF 1.1 1.8
Protease inhibitor mixture 0.7 1.4
Fresh trypomastigotes (having an adhesion and infection index of 0.5 and 1.2, respectively) were activated
for 5 h under standard conditions and with the additions indicated. Protease inhibitors were used at 15 lag/ml, with the exception of PMSF which was 1 raM; the protease inhibitor mixture contained aprotinin,
pepstatin, trypsin inhibitor, PMSF and TLCK. In control experiments it was established that the presence
of inhibitors during the adhesion and infection assays only of fresh (To), partially (Ts) and fully (T24) activated parasites has no effect.

156
TABLE II
The effect on adhesion and infection of treating T. cruzi trypomastigotes with different proteases.
Treatment Percent of control
Adhesion Infection
None 100 100
Trypsin 420 200
Chymotrypsin 221 155
Subtilisin Calsberg 180 116
Nagarse 176 135
Pepsin 67 52
Bromelain 133 161
Papain 273 154
Trypomastigotes were washed twice by centrifugation in Ca 2÷ and Mg2+-free PBS, resuspended in this
solution and treated with 60 lag/ml protease for 30 min at 37°C. Reaction was stopped with 10% FCS and
immediate centrifugation; the parasites were resuspended in 'complete medium' and used for the assays.
RI
X
Z _ o
z 6 q o
I-- U
z 4
A
• - '4 / JJ
~ 0'.0s 015 s'0 s0 200 TRYPSIN (.Uglml)
Fig. 2. The effect of trypsin concentration on the infectivity of fresh trypomastigotes (A) and 20 h activated
parasites (B). Trypsinization was done as indicated in Materials and Methods, except that trypsin (Type XI,
Sigma) was used at the concentrations indicated in the abscisa.

157
TABLE III
The effect of trypsinization on the adhesion and infection of T. cruzi trypomastigotes produced in cells cultured in serum-free medium.
Parasite Parasites per cell treatment
Adhesion Infection
-FCS +FCS -FCS +FCS
None 0.63 0.56 0.41 0.95 Trypsin 3.13 2.61 0.68 2.82
Fresh trypomastigotes, obtained from infected cells which had been switched to MEM alone 24 h prior to bursting, were either trypsinized or not and then used to measure adhesion and infection under standard conditions (+FCS) and in a medium lacking serum (-FCS).
fresh parasites in the absence of serum is only half the value obtained in its presence
and the effect of trypsinization is small when no serum is added during the assay, but it
becomes rather important in its presence. The activation produced by trypsin treat-
ment of trypomastigotes is a property not shared by T. cruz i epimastigotes, since
trypsinization of this form does not modify their inability to adhere and to infect
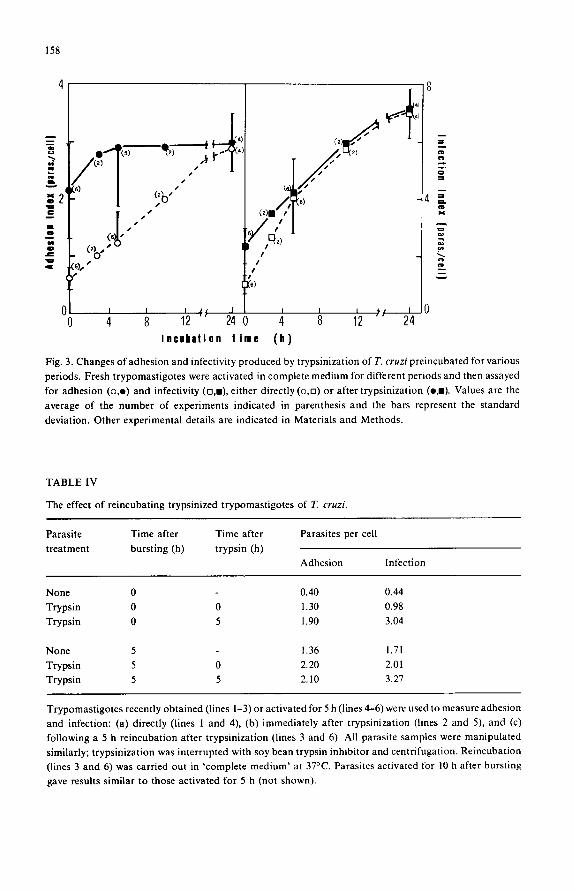
fibroblasts in vitro (not shown). Parasite trypsinization has been used also consecutively to a previous treatment:
Fig. 3 shows the results of treating the parasites with trypsin, after they have been
already activated for different periods. The protease concentration used was chosen as
to produce activation of fresh trypomastigotes, without affecting adversely the activ-
ity of parasite already activated. It can be seen that, regardless of the activation period
(and with the sole exception of fresh trypomastigotes), trypsinization stimulates adhe-
sion to a value which coincides with that obtained by a prolonged incubation of the
parasites. Thus, even trypomastigotes recently liberated from infected fibroblasts can
nearly achieve their maximal adhesive capability upon trypsinization. The kinetics of
infectivity are completely different: trypsinization stimulates infection only if the
trypomastigotes have been activated for less than 5 h; moreover, the infectivity of the fresh trypsinized trypomastigotes is only one-third of that attained by incubating the
parasites for 20-24 h. Table IV shows that reincubation, in the absence of trypsin, of
trypsinized parasites (either fresh or pre-activated for 5 or 10 h) does not decrease their
adhesion or infectivity; on the contrary, the latter continues to increase during this reincubation period. Reincubation of trypsinized parasites, followed by radioiodina-
tion, indicates resynthesis of the 95 000 dalton band (not shown). As already shown in preliminary form [6], trypomastigotes of parasites whose
development of adhesion and infectivity had been blocked by protease inhibitors, puromycin or tunicamycin can restore their full adhesion capability, but not their
158
J
" . ,2 .
if'"," O I i
i, :ll r
f t--a(z)
( 2 ) 1 ~ j(6)
/
/
i I
re)
I e i i ,~/ 24 0 4 8 12 0 4 8 12 24
Incubation lime (h)
= m . o z l
m
4 _ = c o ~,c
Fig. 3. Changes of adhesion and infectivity produced by trypsinization of T. cruzi preincubated for various periods. Fresh trypomastigotes were activated in complete medium for different periods and then assayed for adhesion (o,e) and infectivity (n, i ) , either directly (o,n) or after trypsinization (o j ) . Values are the average of the number of experiments indicated in parenthesis and the bars represent the standard
deviation. Other experimental details are indicated in Materials and Methods.
TABLE IV
The effect of reincubating trypsinized trypomastigotes of T. cruzi.
Parasite Time after Time after Parasites per cell treatment bursting (h) trypsin (h)
Adhesion Infection
None 0 0.40 0.44 Trypsin 0 0 1.30 0.98 Trypsin 0 5 1.90 3.04
None 5 1.36 1.71 Trypsin 5 0 2.20 2.01 Trypsin 5 5 2.10 3.27
Trypomastigotes recently obtained (lines 1-3) or activated for 5 h (lines 4-6) were used to measure adhesion and infection: (a) directly (lines 1 and 4), (b) immediately after trypsinization (lines 2 and 5), and (c) following a 5 h reincubation after trypsinization (lines 3 and 6). All parasite samples were manipulated similarly; trypsinization was interrupted with soy bean trypsin inhibitor and centrifugation. Reincubation (lines 3 and 6) was carried out in 'complete medium' at 37°C. Parasites activated for 10 h after bursting gave results similar to those activated for 5 h (not shown).

DISCUSSION
infectivity, except for the protease inhibitor-treated parasites. Thus, upon trypsiniza-
tion, leupeptin-, antipain-, or trypsin inhibitor-treated t rypomast igotes reacquired
immediately both the adhesion and infective capabilities of trypsinized controls (Fig.
4). Similar experiments were carried out using puromycin and tunicamycin during a 4
and 7 h activation period (Fig. 5). The results indicate that the puromycin-dependent
inhibition of adhesion can be overcome by trypsinization of these parasites, but that
this t reatment does not restore the blocked infectivity; the latter is also observed when
tunicamycin-treated parasites are used.
4
2
..-:.
We have already shown that the invasive capacity of T. cruz i t rypomast igotes
derived f rom cell cultures is not totally expressed in recently released parasites, but
that an extracellular 'ma tu ra t ion ' of these pro tozoa is required to attain this full
capacity [1,6]. The use of RNA, protein, and glycoprotein biosynthesis inhibitors has
suggested that during the 'ma tu ra t ion ' process the following events were taking place:
(a) translation of pre-existing R N A with the information for parasite adhesion, and
A
B
i 3
2
I
O
159
Fig. 4. The effect of trypsinization of T. cruzi activated in the presence of protease inhibitors. Fresh trypomastigotes (open bars) were activated for 5 h either in the absence (control) or in the presence of 15 I~g/ml of the protease inhibitors indicated in the abscisa; the parasites were washed twice by centrifugation, aliquots were treated (cross-hatched bars) or not (single-hatched bars) with trypsin under standard conditions, and then assayed for adhesion (A) and infectivity (B).

160
100
a= 5O
= o .~lOO:
I - - , ;
~ 5o :1
0
I00
'A
t O t 4 t 7 t O t 4
B
D
100
0
t 100
t 7
50 . ~
7, L-J
o7, 100 ~
B
m
Z
Fig. 5. The effect of trypsinization of T. cruzi activated in the presence of puromycin or tunicamycin. Fresh trypomastigotes (To) were activated for 4 and 7 h periods (T4 and T 7 , respectively) either alone (panels A and B) or in the presence of 1.2 lag/ml puromycin (C and D) or 125 p.g/ml tunicamycin (E and F). The parasites were washed by centrifugation and aliquots were untreated (open bars) or trypsinized under standard conditions (hatched bars), and finally assayed for adhesion (A, C, E) and infectivity (B, D, F). The results are expressed as the percentage of the values attained by the 7 h activated and trypsinized control parasites.
(b) t ranscript ion of the RNA and biosynthesis o f the corresponding glycoprotein(s)
necessary for penetrat ion [2,6]. The experiments here reported now suggest that the
protein(s) needed for adhesion was preformed in the parasites, but in an inactive or
hidden form, and that puromycin was probably acting by blocking the biosynthesis of
a protein(s) responsible for the activation or unmasking of these pre-existing mole-
cules, rather than inhibiting directly their synthesis. In fact, trypsin t reatment of
parasites brings about an immediate increase of their adhesive capability to values
similar to those attained by endogenous activation, regardless o f whether: (a) they
have been pre-incubated or not (Fig. 3), (b) the development of adhesion has been blocked or not with protease inhibitors (Fig. 4), or (c) the development has been
blocked with puromycin (Fig. 5). Only a very small p ropor t ion o f the total adhesion capabili ty seems to be synthesized de novo during parasite incubation, according to
the difference between the adhesion index of fully activated parasites and trypsinized
fresh t rypomast igotes (Fig. 3). In contrast , trypsinization o f fresh t rypomast igotes stimulates infectivity 2-fold, but this new value is still one-third of that obtained by
prolonged incubat ion o f the parasites (Fig. 3). Thus, a t ime-dependent process, which is puromycin- and tunicamycin-sensitive and cannot be overcome by trypsinization

161
(Fig. 5), is required to generate most of the infective capability of the parasite. This suggests that a glycoprotein(s) necessary for the penetration step is probably synthesiz- ed de novo during the maturation period. If the limiting event of the infective process of fibroblasts by fresh trypomastigotes is adhesion, then the stimulation of infection by trypsinization of these fresh parasites (Fig. 3) would be due to the elimination of this restriction, rather than to a stimulation of the penetration step per se. After trypsinization, however, the penetration step seems to become limiting, because a de novo synthesis is required to achieve the maximal infective capability. Since activation can be brought about by exogenous proteases and retarded by protease inhibitors, it is tempting to speculate that the endogenous activation occurring during the extracellul- ar incubation is produced by a parasite protease. The changes in surface membrane proteins observed upon radioiodination of the various parasite species are compatible with this suggestion [6], even though it is difficult to assign unequivocally any of these changes to the functional alterations. Also, we have recently detected a protease activity in intact trypomastigotes, which is located in the external surface of the parasites [12]; the activity of this enzyme is inhibited by various protease inhibitors, suggesting that we are dealing either with more than one enzyme or with a non-specific enzyme. The specificity of the parasite activation also is not very stringent, as indicated by the effect of various proteases (Table II) and protease inhibitors (Table I). However, further and independent evidence is desirable to establish more firmly whether the endogenous activation is due to a proteolytic process catalyzed by an inducible surface membrane protease of the parasite.
The effect of proteolytic enzymes, especially trypsin, on different T. cruzi parame- ters has been already explored by several authors, sometimes with apparently conflic- ting results. Thus, Andrews and Colli [13] have reported that exposure of cell-culture-derived trypomastigotes to high levels of FCS inhibited adhesion and infectivity and that these parameters could be restored to their original levels by parasite trypsinization, a treatment which modified only serum-pretreated trypomas- tigotes. In our system, however, no inhibitory effect of serum on adhesion or infectiv- ity could be shown and the presence of serum during the assays, actually stimulated infectivity; moreover, the trypsin-induced activation of adhesion and infection could be obtained even with trypomastigotes liberated from infected cells into a serum-free medium (Table III), suggesting that the activation is not due to the release of an inhibition produced by an interaction between a serum factor and the parasites. Andrews et al. [ 14] have later shown that treatment with 150 ~tg/ml trypsin decreased by more than 90% infection and modified several surface antigens: both parameters recovered upon a 4-5 h incubation and cyciohexamide could block this recovery. These results would be consistent with our finding that trypsinization can decrease infectivity when a high protease concentration is used and no special precautions are taken as to use only fresh trypomastigotes (Fig. 2B). Such a decrease, however, probably reflects a general deterioration, by the protease treatment, of an already activated parasite, rather than a relatively specific change; if this is the case, the

162
polypeptides whose biosynthesis is observed during the recovery period are most likely related to a general parasite recovery, rather than to a specific function. Nogueira et al. [ 15] and Kipnis et al. [ 16] have shown that trypsinization of blood form trypomastigotes substantially stimulates their uptake by macrophages; reincubation of trypsinized parasites restored the low uptake that was characteristic of untreated blood trypomastigotes, and cycloheximide blocked this recovery. This effect of tryp- sin has been interpreted as due to the removal of a surface glycoprotein (M r of 90 000) of the parasite, which acted as an antiphagocytic factor [ 15,17,18]. Using cell-culture- derived trypomastigotes we could not find, upon reincubation of mildly trypsinized parasites, any decrease of their adhesive and infective capability for fibroblasts, but actually we observed a further increase of these parameters (Table IV). Thus, it would seem that the 90000 dalton protein does not play the same role in the interaction between cell-culture-derived trypomastigotes and fibroblasts as it does between blood form parasites and macrophages. In this regard it should be noted that T. cruzi
epimastigotes, which lack the 90 000 dalton protein, are unable to infect fibroblasts, while they can invade macrophages. The eventual role of the 90 000 dalton glycopro- tein in the fibroblast-trypomastigote interaction can be explored now through a different approach, since the presence of neuraminidase activity in the cell surface of fibroblast-derived trypomastigotes has been shown [ 19,20] and its relationship with the 90 000 dalton glycoprotein has been suggested [ 18,21]. Experiments along this line are in progress.
The maturation process would be advantageous for the parasite, since it increasesits ability to invade new fibroblastic cells; however, whether trypomastigotes undergo maturation as they are initially released in vivo is not known. If this phenomenon has an in vivo significance and 'natural ' maturation is brought about by endogenous protease(s), then it is worth considering other in vitro findings on the consequences of trypomastigote treatment with proteases. Thus, it is known that trypomastigotes are not able to activate complement in non-immune sera, but upon trypsinization they become activators of the alternative complement pathway [ 16]. This activation might result eventually in their lysis [16], but we have recently observed that the expression of most of the increased infectivity of mature or trypsinized parasites is complement- dependent ([21] and Henriquez et al., manuscript in preparation). Hence, the effective- ness of the invasive process of fibroblastic cells by T. cruzi trypomastigotes seems to depend on a rather co~mplex interplay of various factors related to the parasites, the medium where the infection occurs and the host cell.
ACKNOWLEDGEMENTS
The authors wish to express their gratitude to Dr. Pablo A. Pulido for his interest and continuous support. This work was financed in part by funds from CONICIT of Venezuela (Project S 1-1303), the UNDP/Wor ld Bank /WHO Special Programme for Research and Training in Tropical Diseases, and CMDLT.

163
REFERENCES
1 Piras, M.M., Piras, R. and Henriquez, D. (1982) Changes in morphology and infectivity of cell culture-
derived trypomastigotes of Trypanosoma cruzi. Mol. Biochem. Parasitol. 6, 67-81. 2 Piras, R., Piras, M.M. and Henriquez, D. (1982) The effect of inhibitors of macromolecular biosynthe-
sis on the in vitro infectivity and morphology of Trypanosoma cruzi trypomastigotes. Mol. Biochem. Parasitol. 6, 83-92.
3 Piras, R., Henriquez, D. and Piras, M.M. (1980) Studies on host-parasite interactions: role of fibroblastic cell surface functions and Trypanosoma cruzi forms in the infective process. In: The Host-Invader Interplay (Van den Bossche, H., ed.), pp. 131-134. Elsevier, Amsterdam.
4 Henriquez•D.•Piras•R.andPiras•M.M.(•98•)Surfacemembraneinteracti•ns•fTrypan•s•macruzi and Vero cells: dissociation of the adhesion and penetration steps. J. Cell Biol. 91, 108a.
5 Henriquez, D., Piras, R., Negri, S. and Piras, M.M. (1982) Efecto de enzimas proteoliticas y de inhibidores de proteasas sobre la interacci6n Trypanosoma cruzi- fibroblasto. Acta Cient. Venez. 33
(Supl. 1), 441. 6 Piras, R., Piras, M.M. and Henrlquez, D. (1983) Trypanosoma cruzi-fibroblast interactions necessary
for cell invasion. In: Cytopathology of Parasitic Disease (Ciba Foundation Symposium No. 99) pp. 31-51, Pitman Books, London.
7 Fraker, F.J. and Speck, J.C. (1978) Protein and cell membrane iodination with a sparingly soluble chloroamide 1,3,4,6-tetrachloro-3a,6a-diphenylglycoluril. Biochem. Biophys. Res. Commun. 80, 849-857.
8 Camargo, E.P., Barbieri, C.L. and Jankevicius, J.V. (1982) Possible artifacts in the radioiodination of surface proteins of trypanosomatids. J. Immunol. Methods 52, 245-253.
9 Laemmli, U.K. (1970) Cleavage of structural proteins during assembly of the head of bacteriophage T4. Nature 227, 680-685.
10 Swanstrom, R. and Shank, P.R. (1978) X-ray intensifying screens greatly enhance the detection by autoradiography of the radioactive isotopes 32p and 125I. Anal. Biochem. 86, 184-192.
11 Zingales, B., Andrews, N.W., Kuwasima, V.Y. and Colli, W. (1982) Cell surface antigens of Trypano- soma cruzi: possible correlation with the interiorization process in mammalian cells. Mol. Biochem. Parasitol. 6, 112-124.
12 Recover, P. and Piras, R. (1983) Actividad prote~.sica frente a ~25I-caseina de tripomastigotes enteros de Trypanosoma cruzi. Acta Cient. Venez. 34 (Supl. 1), 39.
13 Andrews, N.W. and Colli, W. (1982) Adhesion and interiorization of Trypanosoma cruzi in mammal- ian cells. J. Protozool. 29, 264-269.
14 Andre ws• N. W.• Katzin• A.M. and C•••i• w . ( • 984 ) Mapping •f surface g•yc•pr•teins •f Trypan•s•ma cruzi by two-dimensional electrophoresis. Eur. J. Biochem. 140, 599-604.
15 Nogueira, N., Chaplan, S. and Cohn, Z. (1980) Trypanosoma cruzi: factors modifying ingestion and fate of blood-form trypomastigotes. J. Exp. Med. 152, 447-451.
16 Kipnis, T.L., David, J.R., Alper, Ch.A., Sher, A. and Dias Da Silva, W. (1981) Enzymatic treatment transforms trypomastigotes of Trypanosoma cruzi into activators of the alternative complement pathway and potentiates their uptake by macrophages. Proc. Natl. Acad. Sci. U.S.A. 78, 602-605.
17 Nogueira, N., Chaplan, S., Tydings, J.D., Unkless, J. and Cohn, Z. ( 1981) Trypanosoma cruzi: surface antigens of blood culture forms. J. Exp. Med. 153,629-639.
18 Nogueira, N. (1983) Host and parasite factors affecting the-invasion of mononuclear phagocytes by Trypanosoma cruzi. In: Cytopathology of Parasitic Disease (Ciba Foundation Symposium No. 99) pp. 52-73, Pitman Books, London.
19 Pereira, M.E.A. (1983) A developmentally regulated neuraminidase activity in Trypanosoma cruzi. Science 219, 1444-1446.
20 Pereira, M.E.A. (1983) In: Cytopathology of Parasitic Disease (Ciba Foundation Symposium No. 99) p. 68, Pitman Books, London.
21 Piras, M.M., Henfiquez, D., Negri, S. and Piras, R. (1983) Efecto de factores del complemento en la infecci6n de celulas Vero por Trypanosoma cruzi. Acta Cient. Venez. 34 (Supl. 1), 318.
Copyright © 2022 FDOKUMEN




















