
Identification and Characterization of Proteolytic Activities from ...
114
Identification and Characterization of Proteolytic Activities from S. mutans that Hydrolyzes Dentinal Collagen Matrix Bo Huang, DMD, PhD A thesis submitted in conformity with the requirements for the degree of MSc Faculty of Dentistry University of Toronto © Copyright by Bo Huang (2021)
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Identification and Characterization of Proteolytic Activities from ...

Identification and Characterization of Proteolytic Activities from S.
mutans that Hydrolyzes Dentinal Collagen Matrix
Bo Huang, DMD, PhD
A thesis submitted in conformity with the requirements
for the degree of MSc
Faculty of Dentistry
University of Toronto
© Copyright by Bo Huang (2021)

ii
Identification and Characterization of Proteolytic Activities from S. mutans that
Hydrolyzes Dentinal Collagen Matrix
A thesis submitted in conformity with the requirements
for the degree of MSc (2021)
Bo Huang, DMD, PhD
Faculty of Dentistry
University of Toronto
Abstract
Objective: To measure the proteolytic activity of S. mutans, its discrete fractions, and proteases
towards demineralized human dentin.
Methods: Demineralized human dentin slabs were incubated with either medium, cultures
(overnight or newly inoculated) of S. mutans UA159, or different bacterial fractions (intracellular,
supernatant or bacterial membrane). Media from each condition was analyzed for a collagen
degradation marker, hydroxyproline. Three potential proteolytic enzymes (SMU_759, SMU_761
and SMU_1438c) from S. mutans UA159 were expressed and their activity toward dentinal
collagen was measured based on hydroxyproline analysis.
Results: Media only and bacterial membrane had no activity towards dentinal collagen. Overnight
culture of S. mutans had the highest degradative activity (p<0.05), followed by supernatant and
intracellular component, and newly inoculated culture (p<0.05). SMU_759 had the highest
degradative activity towards dentinal collagen, followed by SMU_761 (p<0.05). SMU_1438c
showed no collagen degradative activity (p<0.05).
Conclusion: S. mutans dentinal collagen degradation could potentially contribute to caries
formation.

iii
Acknowledgments
I would like to thank my supervisors, Dr. Yoav Finer and Dr. Dennis Cvitkovitch, for giving me
this opportunity to work with them. They have been excellent mentors. This thesis would not have
been possible without their support. I would also like to thank my committee members, Dr.
Christopher McCulloch and Dr. Paul Santerre, whose insights and suggestions helped me to
improve the quality of this project.
I have been lucky to work with great people in the Dr. Finer’s laboratory, who have created a great
work environment: Dr. Cameron Stewart, Russel Gitalis, Dr. Ousama Damlaj. I would also like to
thank members in Dr. McCulloch’s laboratory.
I am also very grateful to my family for their support throughout this process, in particular my
husband Liang Ren, who has always been there for me, encourages me, guides me and understands
me. And, thank you, my lovely children, Claire and Ajax, for your hugs, kisses, smiles and kind
letters. At the last, but not the least, I would love to express my sense of gratitude to my sister,
Youning, who has been so caring and supportive during the 3 years.

iv
Table of Contents
Abstract ......................................................................................................................................... II
List of tables................................................................................................................................. VI
List of figures ............................................................................................................................. VII
List of abbreviations ................................................................................................................ VIII
Preface .......................................................................................................................................... IX
Chapter 1 introduction ................................................................................................................. 1
1.1 INTRODUCTION ..................................................................................................................... 1
1.2 HYPOTHESES ....................................................................................................................... 3
1.3 OBJECTIVES ........................................................................................................................ 3
Chapter 2 literature review .......................................................................................................... 5
The potential role of bacterial proteases in caries and periodontitis pathogenesis ................ 5
2.1 ABSTRACT.............................................................................................................................. 5
2.2 INTRODUCTION ..................................................................................................................... 6
2.3 TOOTH AND SUPPORTING STRUCTURES ................................................................................ 8
2.4 BACTERIAL ASSOCIATED ORAL DISEASES .......................................................................... 15
2.5 BACTERIAL PROTEASES ...................................................................................................... 21
2.6 CONCLUSIVE REMARKS ....................................................................................................... 31
Chapter 3 manuscript ............................................................................................................... 33
Streptococcus mutans proteolytic activity degrade dentinal collagen..................................... 33
3.1 ABSTRACT............................................................................................................................ 33
3.2 INTRODUCTION ................................................................................................................... 35
3.3 MATERIALS AND METHODS ................................................................................................. 36
3.3.1 generic and specific mmp-like activity of s. Mutans ua159 ......................................... 36
3.3.2 soluble type i collagen degradation by s. Mutans ua159............................................. 37
3.3.3 dentinal collagen degradation by s. Mutans ua159 and its discrete fractions ............ 38
3.4 RESULTS .............................................................................................................................. 39
3.4.1 the generic and specific mmp-like activity of s. Mutans ua159 ................................... 39
3.4.2 soluble type i collagen degradation by s. Mutans ua159............................................. 40
3.4.3 dentinal collagen degradation by s. Mutans ua159 and its discrete fractions ............ 41
3.4 DISCUSSION ......................................................................................................................... 42
3.5 CONCLUSION ....................................................................................................................... 47
Chapter 4 manuscript ............................................................................................................... 48
Characterization of proteolytic activity and identification of responsible proteolytic
enzymes of streptococcus mutans towards dentinal collagen................................................... 48
4.1 ABSTRACT............................................................................................................................ 48
4.2 INTRODUCTION ................................................................................................................... 50
4.3 MATERIALS AND METHODS ................................................................................................. 51
4.3.1 characterization of proteolytic activity of intracellular proteins of s. Mutans ............ 51
4.3.2 verification of dentinal collagen degradation by intracellular proteins of s. Mutans
using sds-page and mass spectrometry ................................................................................. 52

v
4.3.3 bioinformative analysis of putative genes of collagen-degrading proteases in s.
Mutans ua159........................................................................................................................ 53
4.3.4 protein identification of putative collagen-degrading proteases in s. Mutans ua 159 54
4.3.5 cloning, expression and purification of bacterial collagen-degrading proteases ....... 54
4.3.6 degradation of dentinal collagen by smu_759, smu_761 and smu_1438c .................. 56
4.4 RESULTS .............................................................................................................................. 57
4.4.1 characterization of proteolytic activity of intracellular proteins of s. Mutans ............ 57
4.4.2 verification of dentinal collagen degradation by intracellular proteins of s. Mutans
using sds-page and mass spectrometry ................................................................................. 58
4.4.3 bioinformative analysis of putative genes of collagenolytic/gelatinolytic proteases in s.
Mutans ua159........................................................................................................................ 61
4.4.4 protein identification of putative collagen-degrading proteases in s. Mutans ua 159 61
4.4.5 cloning, expression and purification of bacterial collagenolytic/gelatinolytic proteases
............................................................................................................................................... 61
4.4.6 degradation of dentinal collagen by smu_759, smu_761 and smu_1438c .................. 62
4.5 DISCUSSION ......................................................................................................................... 63
4.5.1 the characteristics of collagenolytic/gelatinolytic activity of s. Mutans intracellular
proteins ................................................................................................................................. 64
4.5.2 the specific collagenolytic/gelatinolytic proteases ...................................................... 67
4.6 CONCLUSION ....................................................................................................................... 69
Chapter 5 general discussion and summary ........................................................................... 71
5.1 THE POTENTIAL CONTRIBUTION OF PROTEOLYTIC ACTIVITY OF S. MUTANS TO COLLAGEN
DEGRADATION IN CARIES FORMATION..................................................................................... 71
5.2 THE CHARACTERISTICS OF COLLAGENOLYTIC/GELATINOLYTIC ACTIVITY OF S. MUTANS
INTRACELLULAR ENZYMES ...................................................................................................... 73
5.3 THE SPECIFIC COLLAGENOLYTIC/GELATINOLYTIC ENZYMES .......................................... 75
Chapter 6 conclusions and future studies ............................................................................... 76
Chapter 7 reference .................................................................................................................. 81
8. Supplemental information.................................................................................................... 100
8.1 PREPARATION OF DISCRETE FRACTIONS OF S. MUTANS .................................................. 100
8.1.1 intracellular components ........................................................................................... 100
8.1.2 membrane pellets ....................................................................................................... 100
8.2 HOMOLOGY DETECTION AND 3D MODEL STRUCTURAL ANALYSIS OF SMU_759, SMU_761
AND SMU_1438C...................................................................................................................... 101
8.3 PUTATIVE COLLAGENASE GENE SEQUENCES.................................................................... 102

vi
List of Tables
Table 2.1: Major collagen types ...................................................................................................12
Table 2.2: General properties of biofilms and microbial communities ...................................... 16
Table 2.3: Selected examples of bacterial proteases, their preferred cleavage sites and example
inhibitors .......................................................................................................................................22
Table 2.4: proteases involved in generation of energy source......................................................24
Table 2.5: Enzymatic Activities of P. gingivalis, P. asaccharolyticus, and P. endodontalis.......28

vii
List of Figures
Fig. 2.1: Diagram of dental plaque..................................................................................................6
Fig. 2.2: Diagram of tooth structure ...............................................................................................9
Fig. 2.3: The supporting tissues of tooth and the cellular components.........................................14
Fig. 3.1: MMP-like activity of intact (left) and lysed (right) S. mutans UA 159 .........................40
Fig. 3.2: Hydroxyproline production after incubation of soluble type I collagen with S. mutans
UA159 ...........................................................................................................................................41
Fig. 3.3: Isolated hydroxyproline from media after incubation of dentinal collagen slabs with
O/N and NEW S. mutans UA159 and its discrete fractions .........................................................42
Fig. 4.1: The primers and pCOLDII vector information for gene expression ..............................56
Fig. 4.2: Hydroxyproline production from dentinal collagen slabs treated with various methods
and incubated by intracellular proteins of S. mutans UA159 or media ........................................58
Fig. 4.3: Identification of the dentin collagen degradation products following digestion with the
extracted intracellular proteins of S. mutans UA159.....................................................................59
Fig. 4.4: Identification of peptide sequence from dentinal collagen degradation by S. mutans
UA159 intracellular proteins..........................................................................................................60
Fig. 4.5: SDS-PAGE analysis of purified enzymes.......................................................................62
Fig. 4.6: Hydroxyproline production after incubation of dentinal collagen with SMU_759,
SMU_761 and SMU_1438c...........................................................................................................63
Fig. 8.1: The identification and analysis of proteinases from S. mutans UA159........................101

viii
List of Abbreviations
ANOVA Analysis of Variance
CDM Chemically Defined Media
MS Mass Spectrometry
OD Optical Density
O/N Overnight
PBS Phosphate Buffered Saline
PCR Polymerase Chain Reaction
SEM Scanning Electron Microscopy
THYE Todd-Hewitt-Yeast Extract
TYEG Tryptone Yeast Extract Supplement with 0.1% Glucose Broth
UV Ultraviolet
UPLC Ultra Performance Liquid Chromatography
WT Wild-type

ix
Preface
This dissertation is submitted in the form of manuscript-based thesis for the degree of Master of
Science at the University of Toronto. The research described herein was conducted in the faculty
of dentistry at U of T between September 2017 and August 2020. This work is original, except
where acknowledgments and references to previous work are made.
Dissertation formatChapter 1: A general introduction, hypotheses, and objectives of the current
project. Chapter 2: Detailed literature review of the topics pertaining to the research problem.
Chapters 3& 4: Compilations of the experimental data that will be submitted. The manuscripts are
presented in form with possible minor changes to include additional experimental details.Chapter
5: A general discussion of all the experimental data obtained in the study. Chapter 6: Conclusions
and future directionsChapter 7: ReferencesChapter 8: Supplementary data in the study that was
not included in the publications.

1
Chapter 1 Introduction
1.1 Introduction
Dental caries, also called tooth decay, is one of the most prevalent chronic diseases and with
significant impact throughout the lifetime [1, 2]. Every year, more than 160 million dental
procedures are required to restore recurrent caries at the margins of restorations at a cost of over
34 billion dollars in the North America [3, 4].
Tooth dentin is comprised of two major components, inorganic minerals and organic collagen
which is mainly dentinal type I collagen [5]. The dental caries process is defined as
demineralization of inorganic minerals, mainly hydroxyapatite, by acid by-products from
cariogenic bacteria, such as Streptococcus mutans (S. mutans), which results in the exposure of
organic dentinal collagen. It was suggested that dentinal collagen degradation due to proteolytic
activity follows demineralization and complements the initial degradative effect of bacterial acids
on dentinal mineral structure, contributing to initiation and progression of primary and recurrent
(secondary) caries [6-9].
The main potential sources of proteolytic enzymes that could contribute to dentinal collagen
degradation are endogenous proteases present in dentin [10-12], the oral microflora [6, 7, 13, 14],
and neutrophils [15]. Previous studies mainly focused on the role of endogenous proteases, matrix
metalloproteinases (MMPs) in the degradation process of dentinal collagen [8, 16, 17]. However,
the contribution of endogenous MMPs to dentin degradation is controversial due to their limited
amount and activity in dentin compared to bacteria and neutrophils [18-20]. In addition, the
activation status of dentinal MMP is unclear [21].
Bacterial collagenases were identified and reported as virulence factors contributing to human

2
disease [22]. Extensive research has been carried out to investigate the key roles of bacterial
collagenase in host colonization [22, 23]. The most well-known microbial collagenases are from
Clostridium [24], followed by Bacillus and Vibrio [22, 25]. Oral bacterial collagenolytic proteases
were identified, characterized and reported as virulence factors contributing to inflammatory
periodontal disease [26]. Human isolates of S. mutans have been shown to cause extensive loss of
bone and the breakdown of the periodontal ligament in gnotobiotic rats [23]. The collagenolytic
activity of this organism was later confirmed using rat tail tendons as the substrate [27]. Two
extracellular S. mutans proteases were isolated that are capable of hydrolyzing synthetic collagen
substrate PZ-Pro-Leu-Gly-Prop-Arg (PZ-PLGPA) and furylacryloyl-Leu-Gly-Pro-Ala (FALGPA)
[28-30], suggesting that these enzymes may contribute to the breakdown of the collagen
component of both dentin and cementum in the formation of caries or secondary caries [28]. In
addition, it’s been reported that two putative collagenases are expressed by S. mutans UA 159
isolated from root caries [31]. These studies suggest a potential role of the bacterial proteolytic
activity in caries formation. However, none of these studies have directly linked specific
collagenolytic/gelatinolytic activity of S. mutans to human dentinal collagen degradation, and only
limited data exist regarding the verification, characterization and the level of specific
collagenolytic/gelatinolytic activity from cariogenic bacteria, nor its mechanism of caries
pathogenesis.
Considering the reported high activity, high efficiency and continuous production of bacterial
collagenolytic enzymes [13, 23, 30], further exploration of the effect of proteolytic activity of the
cariogenic species S. mutans on dentinal degradation and its potential impact on the pathogenesis
of caries and secondary caries is warranted. With pilot studies that putative collagenase genes and
degradative activity were reported in S. mutans [26, 30, 32], the aim of the current study was to

3
investigate proteolytic activity of S. mutans towards type I collagen and demineralized human
dentin, to assess bacterial expressed proteases with collagen-degrading activities in discrete
bacterial fractions, to characterize the proteolytic activity of S. mutans, and to elaborate the
degradation mechanisms.
1.2 Hypotheses
1.2.1 Central Hypotheses
• S. mutans UA 159 is capable of degrading type I collagen and dentinal collagen; the expressed
bacterial proteins have specific enzymatic activity that is different from endogenous
collagenases and contribute to tooth structural destruction
1.2.2 Specific Hypotheses
• S. mutans UA 159 has proteolytic activity that degrades soluble type I collagen
• S. muatns UA 159 produces both intracellular and extracellular proteolytic enzymes that
degrade dentinal collagen
• The whole-cell proteolytic enzymes or specific enzymes identified from S. muatns UA 159
present collagenolytic activity or gelatinolytic activity towards dentinal collagen by
exhibiting characteristic substrate specificity
1.3 Objectives
• To investigate the collagenolytic/gelatinolytic activity of S. mutans UA 159 towards type I
collagen
• To investigate the collagenolytic/gelatinolytic activity of S. mutans UA 159 towards
demineralized human dentinal collagen
• To investigate the collagenolytic/gelatinolytic activity of different fractions of S. mutans UA
159 cells

4
• To characterize proteolytic activity of intracellular proteins of S. muatns UA 159
• To identify specific proteolytic enzymes from S. muatns UA 159 which may contribute to
dentinal collagen degradation
• To elaborate the pathogenic role of S. mutans UA 159 in the degradation of dentinal collagen
by identification and characterization of e specific proteolytic activity of S. mutans.

5
Chapter 2 Literature Review
The potential role of bacterial proteases in caries and periodontitis pathogenesis
2.1 Abstract
Caries and periodontitis are the most common oral disease that have been managed by dental
clinicians on a daily basis. Although specific pathogenic bacteria are identified to be associated
with these diseases, the manifestation of microbial pathogenesis is dependent on complex events
and processes in the host. The current understanding of dental caries defines this disease as the
demineralization of the tooth tissues due to the acid produced by sugar-fermenting
microorganisms. Thus, caries is considered a diet- and pH-dependent process. However, more and
more studies suggest the involvement of proteolytic activity of host cells and bacteria in caries
formation and progression. On the other hand, although host derived proteases have been identified
as the primary etiology for tissue destruction in periodontitis, bacterial proteases could still play
an importance role in understanding disease processes. Unlike diseases attributed to bacterial
toxins which are rather specific to each toxin in the disease manifestation, caries and periodontitis
have been attributed to microbial proteases that are non-specific and very complex. In this review,
we describe the oral structures (tooth and supporting tissue) that is affected by caries and
periodontitis and the current understanding of caries and periodontitis and their associated
pathogenic bacteria. We will elaborate on the contribution of bacterial proteases as virulence
factors in disease initiation and progression in terms of colonization, acquisition of growth
nutrients, evasion of host defenses and tissue destruction. This will allow us to deepen our
understanding of the complex roles of bacteria in disease pathogenesis, to clarify the concept of
multifactorial etiology and to justify the interest of recent investigations in bacterial proteases as
virulence factors.

6
2.2 Introduction
More than 700 bacterial species exist in the oral cavity and the dominant bacteria are streptococcal
species, with other common inhabitants such as Veillonella, Gamella, Rothia,
Fusobacterium, Neisseria, Corynebacterium and Porphyromonas [33-35]. These oral bacteria
survive in the form of a biofilm, also known as dental plaque, which is a complex microbial
community adherent to human soft and hard tissues and responsible for multiple human diseases
[36]. Based on the locations, dental plaque can be classified as supragingival or subgingival with
different proportions of bacterial species (Fig.2.1).
Fig. 2.1: Diagram of dental plaque.
In the oral cavity, dental caries and periodontitis are the most common dental plaque (biofilm)-
related diseases and are associated with supragingival or subgingival biofilm, respectively. Dental
clinicians manage these two diseases on a daily basis due to their significant impact on oral health
status in our community [37]. Over the past 40 years, oral microbiologists have identified specific
bacteria or bacterial groups in the biofilm as etiological agents responsible for dental caries and

7
periodontitis based on their virulence in disease pathogenesis [38-40]. The common bacterial
virulence factors include bacterial invasion, colonization, biofilm formation, evasion of host
immune defense and destruction of tissue structure, which are accomplished by numerous classes
of bacterial end products and proteases [38, 41-43]. Therefore, microbial proteases have received
increased attention to better understand the manifestation of the microbial virulence in disease
pathogenesis. Although the current understanding of dental caries considers this disease as the
result of demineralization of the tooth tissues due to the acid produced by cariogenic bacteria, and
the bacterial virulence factors have been identified and include their ability to produce and tolerate
acids, multiple studies suggest the involvement of proteolytic activity of host cells and bacteria on
the tooth’s demineralized organic tissue destruction in caries formation and progression [44, 45].
On the other hand, the contribution of proteolytic enzymes in periodontitis has been well studied
and host derived proteases have been identified as a major etiological factor in the pathogenesis of
tissue destruction. However, bacterial proteases still play crucial roles due to their direct and
indirect impact on disease initiation and progression [46].
Unlike diseases attributed to bacterial toxins which are rather specific to each toxin in the disease
manifestation, caries and periodontitis, which represent most of disease states attributed to the
microbial proteases are non-specific and very complex. Without accurate understanding of the
involvement of bacterial proteases in disease pathogenesis, it would be difficult to acknowledge
the contribution of specific bacteria to various stages and aspects of pathogenic processes. As a
result, at the clinical level, it is difficult to formulate efficient and effective prevention and
management protocols for disease control. Although this project focus on the proteolytic activity
of cariogenic bacteria on the caries formation, most fundamental and extensive information
regarding collagenolytic/gelatinolytic activity of oral bacteria are from periodontal pathogens.

8
Therefore, this review aims to describe the oral structures (tooth and supporting periodontal tissue)
that is affected in caries and periodontitis, the current understanding of caries and periodontitis and
their associated pathogenic bacteria, and elaborate on the contribution of bacterial proteases as
virulence factors in disease initiation and progression in terms of colonization, acquisition of
growth nutrients, evasion of host defenses and tissue destruction. This would allow to deepen our
understanding of the complex roles of bacteria in disease pathogenesis, to clarify the concept of
multifactorial etiology and to justify the interest of recent investigations of bacterial proteases as
virulence factors.
2.3 Tooth and supporting structures
Clinically, the tooth has two parts, clinical crown and root (Fig.2.2). Each part has distinct
components: the crown is composed of enamel and dentine that shield pulp tissue, and the anatomic
root is covered with cementum as outer layer and dentin as middle layer which shield pulp tissue.
Dentin is the major component that covers pulp tissue from crown to root [47].
The tooth is suspended in the alveolar socket by collagen fibers known as the periodontal ligament,
which are embedded in both alveolar bone and the cementum [48]. The periodontal ligament, the
tooth root, and the alveolar bone socket are defined as the periodontium [49]. These structures are
also known as the supporting structures. Overlying these supporting structures are the gingiva and
the alveolar mucosa (Fig.2.2).

9
Fig. 2.2: Diagram of tooth structure (DT: dentin tubules): SEMs of enamel and cementum show
mineral phase structures; SEMs of dentin show organic matrix, collagen fibers.
2.3.1 Mineral phase of tooth structure
Teeth are composed of enamel, pulp–dentine complex, and cementum (Fig.2.2). The enamel,
dentin and cementum are calcified hard tissue that are mineralized with hydroxyapatite (HA),
which is a crystalline calcium phosphate [50, 51]. The structure of enamel is unique, with 96% of
HA, and the remainder are composed of organic phase and water Since enamel has no residual
cellular components, damage to the enamel structure cannot be actively repaired [52]. Dentin
contains a lower percentage of HA (70%), 20% organic component and 10% water, while
cementum has 50% HA and 50% organic phase and water. Both cementum and dentin have higher
content of organic phase and cellular components that assist in maintenance and repair of their
structures [52]. Since the solubility of HA is pH dependent, and each unit decrease in pH increases
results with a 10-fold of increased solubility of HA [53], pH fluctuations in the oral cavity
significantly affect oral hard tissue. Previous studies have confirmed the critical pH for enamel is
5.4, at which the HA starts dissolving due to the unsaturated calcium and phosphate in saliva or

10
plaque fluids [53]. For dentin and cementum, the critical pH was determined to be 6.7 due to the
different calcium and phosphate saturation conditions [54]. Thus, the root surface is much more
susceptible to acid challenge than enamel. The acidic attacks occur through two primary means:
dietary acid consumed through food or drink and microbial acid attack from bacteria present in the
mouth.
Regardless of the source of acids, the demineralization process initiates when oral pH drops
below the critical pH (pH 5.5). However, demineralization is a reversible process and the
demineralized HA crystal can re-grow under the favorable oral environment for remineralization,
above the critical pH for the respective tissue [55]. Therefore, the demineralization and
remineralization of tooth structures are continuous processes that are significantly affected by
various biological factors in saliva and oral bacteria [52].
2.3.2 Organic phase of tooth structure
Collagen is a rod-like molecule, roughly 300 nm long, comprised of two α1(I) left-handed helix
polypeptide chains and one α2(I) left-handed helix polypeptide chain twisted around a common
axis to form a major right-handed helix [56, 57]. Within triple helical domain, there is a common
triplet sequence Gly- X-Y, where Gly is glycine and X and Y are often proline and hydroxyproline.
The integrity of collagen is maintained by hydrogen bonding between helical chains and inter- and
intramolecular cross-links [58].
There are several genetically distinct collagen types which are categorized by length, triple-helical
domains, the ratios of hydroxylated to non-hydroxylated residues, and the degree of hydroxylysine
glycosylation [59]. Their relative amounts differ among tissues (Table 2.1) [58, 60]. Type I
collagen fiber is the most abundant in human tissues.
In human tooth dentin and cementum, the organic matrices contain collagen, mainly type I, and
non-collagenous proteins (NCPs) including phosphoproteins, proteoglycans, and acidic

11
glycoproteins [61-63]. Enamel contains less than 4 % organic matter which is made up of 90%
amelogenin, the non-collagenous protein [64]. Collagen in enamel matrix is considered virtually
completely removed during the enamel’s maturation process. Traces of collagen in enamel are
most likely types I and V collagen [65, 66]. However, in dentin and cementum, collagen content
is much higher. Type I collagen is the primary component of the organic portion, accounting for
85% in dentin, with the types III and V collagen as the remainder. Among other non-collagenous
proteins, phosphoprotein is the major content accounting for 50% of the non-collagenous part [67].
Despite comprising a minor portion of tooth structures, it is commonly believed that the organic
matrices play important roles in tooth formation, mineralization and maintenance. The NCPs not
only act as inhibitors, initiators, promotors, and/or stabilizers of mineral deposition [68], they also
play roles in maintaining collagen integrity. The function of phosphoproteins in dentin
remineralization was proposed due to the reported electrostatically binding to collagen and calcium
irons [61, 69]. It has been confirmed that proteoglycans formulate and maintain collagen structures
serving as nuclei for organization of collagen fibrils [70, 71]. Collagen molecules are chemically
cross-linked to each other and act as scaffold and active protective sheath coating the HA crystallite
in the tooth structure [55].The collagen cross-links in dentin are unique due to the molecular
distribution and characteristics of NCPs resultant with reducible and non-reducible intermolecular
cross-links [72, 73]. The non-reducible cross-link is critical to maintain collagen integrity, since it
ties collagen chains into triple helical structure by pyridinoline induced tri-functional cross-link
between peptides [61, 74]. In addition, it has been reported that the reducible cross-link constituted
by dihydroxylysinonorleucine or hydroxylysinonorleucine disappeared in carious dentin, which
indicates irreversible destruction of collagen fibers that cannot be repaired by remineralization [73,
75, 76].

12
Previous studies have pointed out that dentinal collagen is more stable in acidic or degradative
environments compared to other tissue collagens [74]. This high mechanical strength and chemical
resistance to enzymatic digestion and acid challenge is primarily due to their covalent cross-
linkages, mineral coating and special interaction of collagen with NCPs [45]. Consequently, any
process that results in degradation, structural disruption or loss of integrity of collagen or NCPs is
likely to have a significant impact on tooth structural integrity.
Table 2.1: Major collagen types
Type Molecule Composition Structural Features Representative Tissues
Fibrillar Collagens
I [α1(I)]2[α2(I)] 300-nm-long fibrils Skin, tendon, bone,
ligaments, dentin, interstitial
tissues
II [α1(II)]3 300-nm-long fibrils Cartilage, vitreous humor
III [α1(III)]3 300-nm-long fibrils; often with type I Skin, muscle, blood vessels
V [α1(V)]3 390-nm-long fibrils with globular N-
terminal domain; often with type I
Similar to type I; also cell
cultures, fetal tissues
Fibril-Associated Collagens
VI [α1(VI)] [α2(VI)] Lateral association with type I;
periodic globular domains
Most interstitial tissues
IX [α1(IX)][α2(IX)][α3(IX)] Lateral association with type II; N-
terminal globular domain; bound
glycosaminoglycan
Cartilage, vitreous humor;
Sheet-Forming Collagens

13
Type Molecule Composition Structural Features Representative Tissues
IV [α1(IV)]2[α2(IV)] Two-dimensional network All basal laminaes
SOURCE: K. Kuhn, 1987, in R. Mayne and R. Burgeson, eds., Structure and Function of
Collagen Types, Academic Press, p. 2; M. van der Rest and R. Garrone, 1991, FASEB J. 5:2814.
Used with permission from the publishers.
2.3.3 Supporting structure (periodontium)
There are four principle components: cementum, alveolar bone, gingiva and periodontal ligament
(Fig. 2.3), as periodontium providing support and maintaining tooth function. Each of the
components is distinct in locations, tissue architecture, chemical and biochemical composition [77].
Bone and cementum are both calcified tissues that consist of mineral matter, mainly HA, and
collagen fibrils (Fig.2.2) [78]. HA content of cementum is 45% to 50%, which is less than that of
bone (65%), dentin (70%) or enamel (97%). The major source of collagen fibers in cementum are
Sharpey fibers as embedded portion of principle fibers of periodontal ligaments, which is
composed of type I collagen (90%) coated with type III collagen (5%) [79, 80]. In addition, there
are fibers that belong to the cementum matrix and non-collagenous components of the interfibrillar
ground substance, such as proteoglycans, glycoproteins, and phosphoproteins. For alveolar bone,
the inorganic matter is composed principally of the HA, along with hydroxyl, carbonate, citrate,
and trace amounts of other ions [81, 82]. The organic matrix consists mainly of collagen type I
(90%), with small amounts of non-collagenous proteins such as osteocalcin, osteonectin, bone
morphogenetic protein, phosphoproteins, and proteoglycans. Unlike cementum which has
acellular (primary) and cellular (secondary) matter [83], bone is enriched with vascular and
cellular matter responsible for constant remodeling to force, oral environment and host conditions.
On the other hand, the gingiva and periodontal ligaments contain no mineral phase and are
predominately comprised of organic matter. The gingiva is composed of the overlying stratified
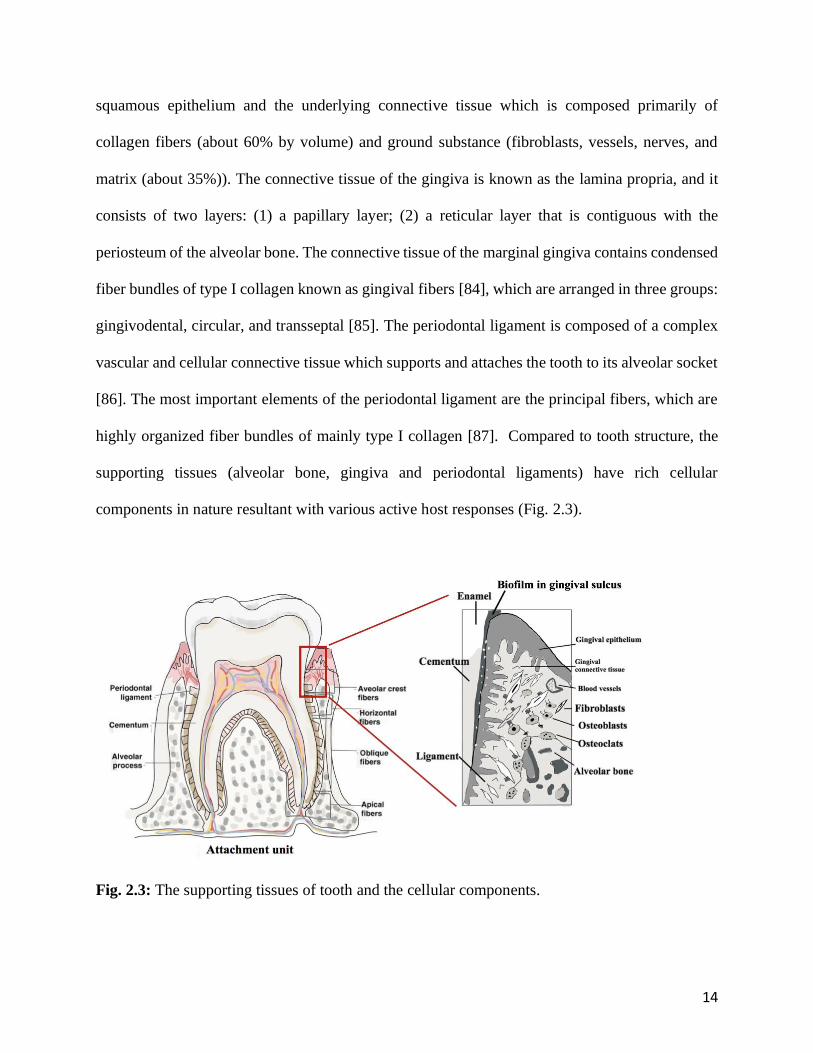
14
squamous epithelium and the underlying connective tissue which is composed primarily of
collagen fibers (about 60% by volume) and ground substance (fibroblasts, vessels, nerves, and
matrix (about 35%)). The connective tissue of the gingiva is known as the lamina propria, and it
consists of two layers: (1) a papillary layer; (2) a reticular layer that is contiguous with the
periosteum of the alveolar bone. The connective tissue of the marginal gingiva contains condensed
fiber bundles of type I collagen known as gingival fibers [84], which are arranged in three groups:
gingivodental, circular, and transseptal [85]. The periodontal ligament is composed of a complex
vascular and cellular connective tissue which supports and attaches the tooth to its alveolar socket
[86]. The most important elements of the periodontal ligament are the principal fibers, which are
highly organized fiber bundles of mainly type I collagen [87]. Compared to tooth structure, the
supporting tissues (alveolar bone, gingiva and periodontal ligaments) have rich cellular
components in nature resultant with various active host responses (Fig. 2.3).
Fig. 2.3: The supporting tissues of tooth and the cellular components.

15
2.4 Bacterial associated oral diseases
For a normal, healthy human being, the total body bacterial population is 10-fold of the human
cells [77]. Bacterial colonization starts at birth and within 2 weeks, a nearly mature microbiota is
established. The entire human microbiota is a very complex collection of hundreds of different
species of bacteria [88]. The microorganisms found in the human oral cavity have been referred to
as the oral microbiota, surviving in the form of biofilm which is an ecological community of
commensal, symbiotic, and pathogenic bacteria as determinants of oral health and disease [89].
2.4.1 Dental plaque
Dental plaque, also known as oral or dental biofilm, is defined as the diverse community of micro-
organisms adherent to the tooth surface and embedded in an extracellular matrix of polymers of
host and microbial origin [90]. The biofilms exhibit enhanced pathogenic properties, which are
more than the sum of the same organisms growing in planktonic form (Table 2.2) [91]. For
example, bacteria in biofilm exhibit surviving advantages such as up to 1,000-fold more antibiotics
resistances, altered metabolic behavior and different stress responses [92-94]. This is due to the
fact that species in biofilms are not randomly distributed, but highly spatially and functional
structured with circulatory systems [91]. As a result, in this highly organized bacterial community
are able to maximize their adaptation mechanisms through biofilm regulation of gene expression,
cell-cell communication and gene transfer [90]. The general properties of biofilm are summarized
in table 2.2 [90].
In the past decades, there have been more than 700 different bacterial species found in the oral
cavity [36]. In general, the dominant bacteria of the oral cavity are streptococcal species, with
other common inhabitants such as Veillonella, Gamella, Rothia, Fusobacterium, Neisseria,
Corynebacterium and Porphyromonas [33-35]. Based on the locations of biofilm, either

16
supragingival or subgingival, the proportion of bacterial species varies. For example, similar to
supragingival plaque, a dominant species subgingivally are Actinomyces, however significantly
higher proportions and counts of anaerobic species were found in the subgingival plaque [95]. The
majority of the microflora benefits health, while only the minority of bacterial species are harmful
and are referred to as pathogenic species [96]. In the oral environment, dental caries (“cavities”)
and periodontitis (“gums disease”) are the most common bacteria-associated diseases. Dental
caries and periodontitis, are considered to be caused at least in part by bacteria. Over the past 40
years, oral microbiologists have identified mutans group streptococci as etiological agents of
dental caries due to their ability to form biofilm, produce acid and leading to destructive tooth
demineralization [97]. It has been also acknowledged that the anaerobic bacteria Porphyromonas
gingivalis (P. gingivalis) and Tannerella forsythia (T. forsythia) are prime agents in the
development of chronic periodontitis [98].
Table 2.2: General properties of biofilms and microbial communities
SOURCE: P. Marsh, 2004, Dental plaque as a microbial biofilm, Caries research 38(3) 204-211.
Used with permission from the publishers.

17
2.4.2 Caries and secondary (recurrent) caries
Dental caries is a multifactorial disease caused by bacteria and influenced by diet, hygiene, tooth
structural integrity and host immune responses [1]. This disease results with a destructive condition
of the dental hard tissues due to the demineralization of the mineral matters in enamel, dentin or
cementum due to desaturation in low pH, which is modulated by acids [99]. Several acid-
generating/producing bacteria have been isolated from biofilm and have been linked to caries
pathogenesis [100]. Bacterial acids are the initial step demineralizing the mineral portion in enamel
and dentin. In dentin, the demineralization process also exposes dentinal collagen to endogenous
and exogenous proteases, which leads to further structural destruction [10, 16, 101].
Secondary caries is defined as the recurrent caries developed along the restoration-tooth interface.
The prevalence of secondary caries has been a major concern, since it is the primary cause (31-
70%) of restoration replacements [102-104]. Over all, dental caries is one of the most prevalent
disease in the world, with more than 160 million dental procedures to restore caries or secondary
caries at a cost of over 34 billion dollars in the North American [3, 4]. The etiology of primary and
secondary caries, are both bacteria associated disease and characterized as tooth demineralization
due to acid formation [105]. However, the existing restorative material is the additional
determinant for the initiation and progression of secondary caries due to the interaction between
cariogenic bacteria and restorative materials and the restoration-tooth interface [106, 107]. Resin
composites as the most popular restorative materials in dentistry [102, 104, 108], but has a higher
secondary caries rate compared to other materials [4, 102, 106, 109-112], which could be related
to the aforementioned interactions between the material and the bacteria.

18
2.4.2.1 Cariogenic bacteria: Streptococcus mutans (S. mutans)
Cariogenic bacteria are considered as a group of microorganisms directly associated with the
pathogenesis of dental caries. Out of the 700 bacterial species that colonize and persist in the oral
cavity, S. mutans is one of the few species that have been consistently linked to caries formation
[97, 113]. The main virulence factors for S. mutans are its ability to form biofilm (dental plaque)
to survive and persist in continuously changed oral environment [114], producing acid
(acidogenicity) and tolerant acidic environments (aciduricity) [115].
The virulence factors associated with adhesion of S. mutans within biofilm have been extensively
investigated: sucrose-independent and sucrose-dependent adhesion of S. mutans are modulated by
self-produced proteins or protease [116-122]. In addition to the proteins and enzymes that
contribute to bacterial adhesion, several proteins have been involved in the metabolism of various
carbohydrates providing an energy resource [123-126]. It has been reported that quorum-sensing
systems encoded by comCDE, have an effect on the capacity of biofilm formation [127-130].
Acidogenicity has been identified as another virulence factor, since S. mutans consumes dietary
carbohydrates and produces various acidic products including lactate, formate or acetate that
decreases biofilm pH, leading to tooth demineralization and caries development [131]. Although
other oral streptococci have ability to produce acid, S. mutans is the one of a few that has the ability
to maintain its function at low pH levels (pH 4.4) which inhibits growth of other oral species [132].
This property is defined as aciduricity or acid-tolerance. S. mutans’ survival capacities in
challenged environments are considered as virulence factors associated with bacterial
pathogenicity. The two-component signal transduction systems (TCSTSs or TCSs) are widely
adopted to regulate its virulent performance by sensing environmental stimuli and responding
accordingly [133]. In S. mutans, 13 TCSTSs and one orphan regulator have been reported. A

19
typical two-component regulatory system contains a membrane-associated, histidine kinase sensor
protein, which senses the environmental conditions, and a cytoplasmic response regulator, which
allows the bacteria to regulate diverse physiological responses through the adjustment of regulator-
target genes expression. [133]. All these virulence factors are executed by different classes of
bacterial proteases which will be discussed in detail in next section (section 2.5.1).
2.4.3 Periodontal disease
Periodontal disease is one of the major causes of tooth loss in adults [37, 134, 135]. Based on the
most current report, 46% of US adults, representing 64.7 million people, had periodontitis [136].
It is a complex infectious disease resulting from interplay of bacterial infection and the host
response to the bacterial challenge that lead to destruction of periodontal ligament and alveolar
bone with clinical presentations of deep probing pockets and tooth mobility [77]. The periodontal
health is normally protected and maintained by intact gingival, sulcular and junctional epithelia
(Fig.2.3) that act as an effective defensive barrier; the underlying connective tissue consisting of
highly organized collagen fibers, proteoglycans and serum-derived components; and host immune
cells such as macrophages and leukocytes as innate defense to bacterial invasion [137]. It is now
generally accepted that a few specific bacteria present virulence factors that cause periodontitis
which include bacterial product-induced tissue toxicity [138, 139], bacterial enzymes that cause
direct tissue destruction [140] and bacteria stimulated host inflammatory responses as the result of
host innate immune defense [137]. The inflammatory process is considered as the major
contributor to the pathogenesis of periodontitis [141, 142]. Previous studies have reported that
some bacterial soluble components are able to diffuse through the epithelium and stimulate the
production of cytokines such as interleukin (IL)-1, IL-6, IL-8 and tumour necrosis factor (TNF),
which are believed to be major mediators of inflammatory disease such as periodontitis [143-145].

20
The immune cells, such as neutrophils are recruited and activated by these mediators such as IL-
8, and release granule enzymes and other intra- and extracellular enzymes contributing to damage
of periodontal supporting tissue [144-146]. Although more recent studies have placed greater
emphasis on the host cells as major contributors to periodontitis rather than bacteria, the dental
plaque formed in the gingival sulcus on enamel or cementum is still considered as a prerequisite
factor for the initiation and accelerating factor for the progression of chronic periodontitis (Fig.2.3).
In addition, the wide spectrum of hydrolytic enzymes furnished by oral bacteria still have direct
effect on tissue pathological change [26, 147].
2.4.3.1 Bacterial species that are associated with periodontitis
Although the nature of periodontitis is very complex and it is not a simple infection caused by one
or two specific pathogenic bacteria that could provide the basis for the diagnosis, several gram
negative bacteria have been linked to the initiation and progression of the periodontal disease
process [148]. The bacteria associated with periodontal diseases reside within the subgingival
biofilms, which consists of more than 500 different species [95, 149]. Among all the bacteria, P.
gingivalis, Actinobacillus actinomycetemcomitans (A actinomycetemcomitans), T. denticola and
Bacteroides forsythus (B. forsythus) are detected in high level using immunocytochemistry and
DNA probing in patient with periodontitis [33].
P. gingivalis has been implicated in chronic and severe adult periodontitis [150], T. denticola in
acute necrotizing ulcerative gingivitis[151] and A. actinomycetemcomitans in localized juvenile
periodontitis [152, 153]. B. forsythus plays an important role in the progression of advanced and
recurrent periodontitis, [39, 154]. Among several periodontal pathogenic bacteria, P. gingivalis
and A. actinomycetemcomitans are the most well-studied ones. Their virulence factors are
associated with the ability to produce tissue toxic fatty acids, lipopolysaccharide which stimulates

21
host immune responses [145, 155], and extracellular proteases which facilitate bacterial invasion,
nutrition acquisition and evasion of host immune defense [156-159]. The mechanisms and
contributions of the proteases will be discussed in detail in next section (section 2.5.2).
2.5 Bacterial proteases
According to the Nomenclature Committee of the International Union of Biochemistry and
Molecular Biology, proteases are classified as a subgroup of hydrolases. However, it is difficult to
assign nomenclature to proteases based on general rules due to their huge diversity of action and
structure. Proteases can be further classified into various categories based on different criteria such
as their site of action on protein substrates, their amino acid sequences and pH optima [160, 161].
The most common classification is based on their catalytic site, such as (1) serine proteases (e.g.
trypsin-like enzymes), (2) cysteine proteases (e.g. gingipains), (3) aspartic proteases (e.g. Candida
albicans Saps), and (4) metallo-proteases (e.g. microbial keratinases). Table 2.3 shows a series of
example target proteases of pathogenic bacteria, including certain oral organisms. Each protease
has its own preferred cleavage site: some of these have broad specificity such trypsin-like proteases
cleaving peptide bonds following Lys or Arg [162], while others are very specific such as IgA
protease, which cleaves the hinge region of the immunoglobulin molecule [163]. All these four
types of proteases were found in pathogenic bacteria and their individual or collective actions are
regarded as virulence factors that play critical roles in disease pathogenesis. The primary function
of bacterial proteases is nutrition acquisition for bacterial growth and proliferation by digesting
host tissue [43, 164]. These proteolytic enzymes also facilitate bacterial invasion and act as defense
system for bacteria against host immune responses that inactivate host protease inhibitors,
degradation of host macromolecules and disruption of host cellular signaling network [165].

22
Table 2.3: Selected examples of bacterial proteases, their preferred cleavage sites and example
inhibitors
Source [166]: Allaker, Robert P., and CW Ian Douglas. "Novel anti-microbial therapies for
dental plaque-related diseases." International journal of antimicrobial agents 33.1 (2009): 8-13.
Used with permission from the publishers.
2.5.1 S. mutans proteases and enzymes involvement in caries
Bacterial proteases are not only closely associated with three well-established virulence factors in
cariogenesis: biofilm formation, aciduricity, and acidogenicity [38], but also involved in other
perspectives contributing to caries and secondary caries formation and progression such as tissue
and material degradation [13, 28, 99, 167].
2.5.1.1 Bacteria adhesion and biofilm formation
The ability of S. mutans are to form biofilms with solid and stable structure enable the bacteria to
resist biological, mechanical and immune attack, improve their survival and increase their
pathogenicity. Virulence factors that are associated with the adhesion of S. mutans within biofilm
have been extensively investigated. There are two pathways associated with adhesion of S. mutans:

23
sucrose-independent and sucrose-dependent adhesion. The sucrose-independent adhesion is
mostly influenced by antigen I/II, a surface protein [116], while proteases of S. mutans,
glucosyltransferases (GTFs) encoded by gtfB, gtfC, and gtfD, govern the sucrose-dependent
adhesion by the synthesis of water-soluble and water-insoluble glucans as extracellular polymer
facilitating bacterial adhesion [117-119]. Although there are other non-enzymatic proteins, such
as glucan-binding proteins A (Gbp A) and glucan-binding proteins D (Gbp D) that are involved in
bacterial adhesion, the GTF-mediated adhesion is considered as the major mechanism for bacteria
binding to tooth surface and to each other [120-122, 168]. GtfC produces a mixture of soluble and
insoluble glucans adsorbed to enamel within saliva pellicle [118, 169] which facilitates binding.
GtfB binds to bacteria such as Actinomyces viscosus, Lactobacillus casei and S. mutans, promoting
cell clustering, enhancing cohesion of plaque and establishing 3D architecture of multi-species
microcolonies [170-172], and is responsible for the formation of biofilm with highly differentiated
structures [173]. GtfD forms a soluble, readily metabolizable polysaccharide and acts as a primer
for GtfB [174]. .
2.5.1.2 Bacterial survival – energy acquisition
In addition to the proteases contributing to bacterial adhesion, other proteases are involved in the
metabolism of various carbohydrates, thus providing energy resource for cariogenic bacteria and
therefore, are also considered as virulence factors. Fructosyltransterase (Ftf) and extracellular
dextranase (DexA) are able to synthesize energy for bacteria; while fructanase (FruA) is able to
digest exogenous carbohydrate into utilizable energy source (table 2.4) [123-126]. Other proteases,
such as sucrose phosphorylase (GtfA) and an intracellular dextranase (DexB), play roles in the
further energy transportation into cells [175].

24
Table 2.4: proteases involved in generation of energy source
Source[126]: J.A. Banas, Virulence properties of Streptococcus mutans, Front Biosci 9(10)
(2004) 1267-77. Used with permission from the publishers.
2.5.1.3 Acidogenicity (acid production) and aciduricity (acid tolerance)
S. mutans consumes dietary carbohydrates and produces various fermentation products including
lactate, formate, acetate, and ethanol. The precise distribution of fermentation products depends
on the growth conditions. When glucose is abundant, lactate (pKa 3.8) is the major end-product
that causes a decrease in the biofilm pH, leading to tooth demineralization and caries development
[131]. The enzyme produced by S. mutans, lactate dehydrogenase (Ldh), is responsible for lactic
acid production and is recognized as a virulence factor of S. mutans, since reduced cariogenicity
was observed on animal models inoculated with Ldh mutant strain [176-178].
To adapt to pH challenged environments, the gene or protein expression patterns are modified by
S. mutans, in an acid tolerance response (ATR) [179, 180]. There are two critical mechanisms that
S. mutans uses to survive at low pHs. The first one is maintaining intracellular pH to avoid external
proton penetration and disruption to cytoplasmic enzymes functions. Studies have shown that the
membrane-bound proton-translocating, F1F0-ATPase proton pump could be up-expressed to
maintain a pH gradient across the cytoplasmic membrane [132, 181, 182]. The second mechanism
is DNA repair, and its importance for bacterial survival of acid shock is well-established [183]. In
S. mutans, one DNA repairing enzyme encoded by uvrA, was not only confirmed to be responsible
for the recovery of pH-induced DNA damage, but also for the bacterial growth at moderately acidic
pH. In addition to these enzymes, other proteases actively contribute to the aciduricity of S. mutans
by contributing to the physical barriers which play critical roles in blocking acidic molecules and

25
maintaining cellular proton gradient. The first barrier is extracellular polysaccharide matrix (EPS)
of biofilm which is regulated by GFTs [184, 185]. The second barrier is the integrity of cellular
membrane. It has been proposed that diacylglycerol kinase, is involved in phospholipid turnover
and therefore affecting the membrane’s structure [186].
Other proteases have been involved in ATR to advance bacterial survival in acidic environment.
In S. mutans, the clpP gene is up-regulated at low pH [187]. The encoded caseinolytic protease,
ClpP associates with other members of the Clp ATPase family and acts as a serine protease can
remove abnormal proteins that accumulate during stress conditions and allow the recycling of
amino acids from non‐essential proteins during starvation [188]. Another proteinase (ClpL) from
the ClpP family was shown to be up‐regulated at pH 5.0 to facilitate acid‐tolerant growth as part
of the stress response of S. mutans [189].
2.5.1.4 Tooth and dental biomaterial degradation by bacterial enzymes
Protease induced dentin degradation has been proposed as an important element in pathological
process of caries [11, 16, 190]. Host derived proteases, such as dentinal MMPs have been mostly
investigated and attracted most attention due to the their true collagenolytic nature [11, 191-193],
Because multiple studies indicate that cariogenic bacterial proteases play multiple important roles
in caries formation, especially dentin caries [28, 99], the proteolytic degradative activity of
cariogenic bacteria is likely a significant contributor to caries. First, host MMPs are secreted in
latent form and buried in dentin matrix which need to be activated to function. Bacterial proteases
are reported to activate these dormant MMPs [9, 18, 194]. As a result, bacterial proteases indirectly
contribute to tissue destruction in caries progression. Second, multiple studies suggested that
several degradative enzymes of S. mutans have direct roles in dentin collagen degradation. Human
isolates of S. mutans have been shown to cause extensive loss of bone and the breakdown of the

26
periodontal ligament in gnotobiotic rats [23]. The collagenolytic activity of this organism was later
confirmed using rat tail tendons as the substrate [27]. Two extracellular S. mutans proteases were
isolated that are capable of hydrolyzing synthetic collagen substrate PZ-Pro-Leu-Gly-Prop-Arg
(PZ-PLGPA) and furylacryloyl-Leu-Gly-Pro-Ala (FALGPA) [28-30], suggesting that these
enzymes may contribute to the breakdown of the collagen component of both dentin and cementum
in the formation of caries or secondary caries [28]. In addition, in a recent study, two putative
collagenases are found to be expressed by S. mutans UA 159 isolated from root caries suggesting
the involvement of bacterial proteases in caries progression [31]. Although the direct contribution
of bacterial protease in tooth structural destruction has been proposed, more elaborate studies are
required and necessary to clarify the mechanisms and to establish the virulence role of bacterial
proteases in caries pathogenesis.
On the other hand, the degradative effect of S. mutans toward dental restorative materials have
been well established, [195, 196]. The esterase SMU_118c is expressed by S. mutans under acidic
conditions, and can hydrolyze resin monomers and polymerized resin composite dental restorative
materials, while retaining its activity in an acidic pH for an extended period. As such, SMU_118c
could contribute to the biodegradation of the restoration-tooth interface, allowing for further
bacterial invasion and potentially promoting formation of secondary caries and restoration failure
[196].
2.5.2 Bacterial proteases as virulence factors in periodontal disease
In order for P. gingivalis and other periodontal pathogenic bacteria to survive and proliferate
within the periodontal pocket, they have been evolved to produce various intra- or extracellular
proteases to facilitate nutrients accumulation, to aid host invasion and to avoid host immune attack.
These proteases are substantial part of the infective armamentarium of the bacteria [197, 198] such

27
as gingipains K (alias Kgp) and R (RgpA and RgpB)[199], which have been well studied as major
virulence factors of the pathogen [46].
2.5.2.1 Nutrient acquisition for growth and proliferation
In order to survive and proliferate in the oral environment, pathogenic bacteria require sufficient
and continuous nutrition supply. The common ways to obtain nutrition are either through
degradation of host connective tissue or proteolysis of plasma exudate.
P. gingivalis and A. actinomycetemcomitans are capable of direct digestion of host tissue such as
collagen by collagenases, that result in the release of amino acid which can be utilized as nutrients
by the resident bacteria [140, 200, 201]. Although not all periodontal pathogens possess true
collagenases, studies show T. denticola, Fusobacterium, Veillonella and even a Bacillus cereus
spp. isolated from the subgingival biofilm elaborate a large number of proteolytic enzymes (Table
2.5) that include hyaluronidase, chondroitin-4-sulfatase, heparinase and a variety of proteases,
peptidases, and aminopeptidases. All of the enzymes are capable of degrading host
macromolecules into their subunit structure for use as carbon and energy sources [202].
However, it has been reported that the most effective way of nutrient acquisition is to transport
nutrients released as plasma proteins [164]. The arginine-specific gingipains (RGPs), from P.
gingivalis can directly act on kininogens leading to overproduction of bradykinin (BK) which is
peptide hormone [46, 203, 204]. The binding of BK to receptors on vascular endothelial cells leads
to increased capillary permeability by contraction of endothelial cells and capillary leakage. As a
result, this process facilitates the accumulation of nutrients from host plasma and aid the
intravascular dissemination of pathogens [205, 206]. At the same time, other bacterial proteases
are produced to aggregate and utilize the nutrients derived from the host. For example, at least two
proteases are proposed to be involved in iron/heme acquisition process which is required for P.

28
gingivalis growth, oxygen tolerance and virulent factors expression [207-210]: the lysine-specific
gingipain (KGP) from P. gingivalis agglutinates red blood cells, then, hemolysin from P. gingivalis,
lyses red blood cells and release the hemoglobin molecule; the freed hemoglobin molecules are
bond to cellular surface of P. gingivalis by KGP for its eventual transport and utilization into the
cell [211, 212].
Table 2.5: Enzymatic Activities of P. gingivalis, P. asaccharolyticus, and P. endodontalis
SOURCE: S.C. Holt, T.E. Bramanti, Factors in virulence expression and their role in periodontal
disease pathogenesis, Critical Reviews in Oral Biology & Medicine 2(2) (1991) 177-281. Used
with permission from the publishers.

29
2.5.2.2 Stimulation and inactivation of host immune response
Another very critical role of bacterial degradative proteases is to defend host immune attack which
has been identified as one of the virulence factors of periodontal pathogenic bacteria [213].
Although there is no protease from S. mutans has been linked to evasion of host immune responses,
a protease from another oral streptococcus, S. sanguis has been isolated that can degrade IgA1
which leads to functional loss of the immunoglobulin [214, 215].
Bacterial proteases are not only involved in the initiation of periodontal disease characterized by
the influx of significant amounts of polymorphonuclear leukocytes into the affected periodontal
region, the release of lysosomal contents, and an accompanying breakdown of associated tissue
[216], but also contribute to the progression of the disease due to their degradative effect on
proteinase inhibitors resulting in rapid and uncontrolled periodontal tissue destruction [147].
The gingipain proteases have been linked to the initiation and progression of periodontal disease
due to their ability to stimulate host immune defense causing tissue destruction and to inactivate
host defense facilitating survival and propagating the destructive processes. Although RGPs
activate the complement pathway which is a primary innate host defense against invasive
pathogens by recruitment of neutrophils [217], other P. gingivalis-derived proteinases are able to
silence the phagocytic effect of the recruited neutrophils by impairing the receptors on neutrophil
surfaces and degrading some components in the complement pathway such as complement
proteins C3, C4, and C5 [218-221]. As a result, instead of killing the pathogens, the recruited
neutrophils die and degranulate at the infected sites, releasing host hydrolases, such as
metalloproteinases and cathepsins G which degrade host connective tissue. This host proteases-
derived tissue degradation process has been widely accepted as the initiative step of pathogenesis
of periodontitis [202]. In addition, the degradative effect of proteases (RGPs, KGP and other

30
proteolytic enzymes) from P. gingivalis do not only target the complement components, they are
also able to rapidly degrade other cytokines such as TNF-, IFN-, IL-6 (regulates differentiation
of B cells) and IL-1 (host response to pathogens) which are critical signal molecules for immune
cells recruitment and regulation [222-224]. The degraded cytokines lost their function to initiate
cell communication, leading to delayed or decreased host defense responses [164, 223]. Even at
the later stage of host defense stage, trypsin-like protease of P. gingivalis are able to digest most
classes of immunoglobulins, including IgA, IgG and IgM [213]. This is likely to be a major
detriment in the maintenance of antibody function by the host. In addition to immunoglobulins,
tissue proteases inhibitors are another class of molecules produced by host to protect tissue
integrity by modulating enzymatic activity. However, multiple proteases isolated from P.
gingivalis can completely digest the host protease inhibitors (a-1-antitrypsin, antichymotrypsin,
2-macroglobulin, antithrombin III, antiplasmin and cystatin C), thus reducing their protective
effect. As a result, without the protease inhibitors, the uncontrolled destructive proteases
continuously degrade host connective tissue, leading to progression stage of periodontitis.
Although extensive studies focused on P. gingivalis, it is clear by now that other pathogens possess
similar proteolytic activity and contribute to the pathogenesis of periodontitis. For example,
chymotrypsin-like protease of T. denticola has been reported as having degradative effect on
proteases inhibitors a2- macroglobulin and cystatin C [225].
2.5.2.3 Contribution of host tissue degradation to periodontal disease
It is important to recognize that the degradation of the elastin and collagen components of
periodontal soft tissue by host-derived proteases is the primary etiology of periodontitis. However,
the question still remains as to the direct and indirect roles of the bacterial proteases in the
bone and tissue destruction.

31
Firstly, it has been proposed that thiol enzymes of P. gingivalis are not only able to up-regulate
the synthesis of MMPs by fibroblast and epithelial cells, but also able to activate the latent forms
of host MMPs, which could be considered as a significant contributor to host proteases-induced
tissue destruction in periodontitis [226].
Secondly, the direct degradative effect from periodontal pathogenic bacteria has been widely
studied and well documented. The collagenases isolated from P. gingivalis and A.
actinomycetemcomitans have been linked to type I collagen degradation in dentin and gingival
tissue leading to periodontal pocket formation with attachment loss [26, 147, 200, 227]. Then, the
degraded gelatin and collagen fragments can be further hydrolyzed by gelatinase and trypsin-like
enzyme from P. gingivalis and T. denticola [228, 229]. In addition to type I collagen,
chymotrypsin-like enzyme from T. denticola membrane could degrade type IV collagen, laminin,
and fibronectin [230]. P. gingivalis, A. actinomycetemcomitans and T. denticola also possess
fibrinolytic activity, which destroy fibrin, breech the host fibrin barrier and to evade into deeper
tissue [231, 232]. Although the correlation of proteases from P. gingivalis with alveolar bone
resorption has not been clearly defined, it has been proposed that alkaline phosphatases of P.
gingivalis can function as phosphoprotein phosphatase that hydrolyzes phosphoserine that could
lead to alveolar bone resorption [233, 234]. In addition, P. gingivalis and the other oral Bacteroides
spp. also produce significant amounts of phospholipid degrading enzymes (phospholipase A)
which may lead to bone resorption [235, 236].
2.6 Conclusive remarks
Dental caries and periodontitis, as the most common oral diseases, have significant impact on
human oral health status. They present as destructive conditions of the mineral and organic matrix
of tooth structure and its supporting tissues which are directly or indirectly caused by pathogenic

32
bacteria. The above literature review covers the current understanding of the mechanism of caries
and periodontitis, their associated specific pathogenic bacteria and emphasized on the contribution
of bacterial proteases in disease pathogenesis. This review points out to bacterial proteases as
executors of virulence factors that play various roles at different stages and aspects in disease
development. For example, bacterial collagenolytic and gelatinolytic enzymes have significant
effect at caries progression. As well, multiple periodontal pathogenic bacteria share common
enzymatic activities stimulating host responses to initiate periodontitis, while other specific
bacteria present with distinct degradative effect to host immune components and host tissue which
contribute to disease progression. In addition, this review summarized those specific proteases
based on previously identified bacterial virulence factors in disease pathogenesis which elaborates
the mechanism of disease development at protein level.

33
Chapter 3 Manuscript
Streptococcus mutans proteolytic activity degrade dentinal collagen
Bo Huang1, Christopher McCulloch1, J. Paul Santerre1,2, Dennis G. Cvitkovitch1,2, Yoav
Finer1,2
1Faculty of Dentistry, University of Toronto, Ontario, Canada
2Institute of Biomaterials and Biomedical Engineering, University of Toronto
3.1 Abstract
Objectives: to explore the role of S. mutans whole cell and discrete fractions in the degradation
of dentinal collagen and to locate potential responsible proteases
Materials & Methods:
MMP-like activities from intact or lysed S. mutans UA159 were measured using fluorimetric
assays. Soluble type I collagen was incubated in chemically defined medium (CDM) alone or with
overnight (O/N) culture of S. mutans UA159, or 1:100 newly inoculated culture of S. mutans
UA159 (NEW). Human dentin slabs (DS) were demineralized in 10% phosphoric acid, then
incubated in of ¼ Todd-Hewitt-Yeast extract (THYE) medium alone or with one of the following:
O/N S. mutans culture; NEW S. mutans culture; intracellular proteins of O/N culture; supernatant
(cell-free fraction) from O/N culture; or bacterial membrane. Media from all above incubated
groups were analyzed for the collagen degradation marker hydroxyproline.
Results: Intact and lysed S. mutans UA 159 showed similar trend of MMP-like activity with
highest generic and MMP9-like activity, followed by MMP1-, MMP2-, and MMP8-like activity.
Generic and MMP1-like activity of lysed bacteria was significantly higher than intact bacteria
(p<0.05). O/N degraded soluble type I collagen at a higher rate than NEW (p<0.05). O/N culture
had the highest degradative activity towards dentinal collagen, followed by supernatant (cell-free

34
fraction), intracellular components, and NEW culture (p<0.05). Media only and bacterial
membrane did not degrade dentinal collagen.
Conclusion: Several sources of proteolytic activity from S. mutans enable the cariogenic
bacterium to degrade type I and dentinal collagen and may play a role in the pathogenesis of dental
caries.

35
3.2 Introduction
Dental caries, or tooth decay, is one of the most prevalent chronic diseases affecting millions with
significant impact throughout the lifetime [1, 2]. Every year, more than 160 million dental
procedures are required to restore primary and recurrent (secondary) caries at the margins of
restorations at a cost of over 34 billion dollars in the North America [3, 4].
Tooth dentin is comprised of two major components, inorganic minerals and organic collagen
which is mainly dentinal type I collagen [5]. Dental caries is defined as demineralization of the
inorganic minerals, mainly hydroxyapatite, by acid end-products from cariogenic bacteria, such as
Streptococcus mutans (S. mutans), which results in the exposure of the organic dentinal collagen.
It was suggested that dentinal collagen degradation due to proteolytic activity follows
demineralization and complements the initial degradative effect of bacterial acids on dentinal
mineral structure, contributing to initiation and progression of primary and recurrent caries [6-9].
The main potential sources of proteolytic activities that could contribute to dentinal collagen
degradation are endogenous dentinal proteases [10-12], the oral microflora [6, 7, 13, 14], and
neutrophils [15]. Previous studies mainly focused on the role of degradative activities from
endogenous matrix metalloproteinases (MMPs) in the hydrolysis of dentinal collagen [8, 16, 17].
However, the contribution of endogenous MMPs to dentin degradation is controversial due to their
limited amount and activity in dentin compared to bacteria and neutrophils [18-20]. In addition,
the activation status of dentinal MMP is unclear [21].
Bacterial proteolytic activities have been well investigated due to their roles in nutrient acquisition,
bacterial invasion and tissue destruction in human diseases [26, 27]. Human isolates of S. mutans
have been shown to cause extensive bone loss and the breakdown of the periodontal ligament in
gnotobiotic rats [23], suggested to be related to the bacteria’s proteolytic activity, suggested by its
ability to degrade rat tail tendons [27]. However, none of these studies have directly linked specific

36
proteolytic activity of S. mutans to human dentinal collagen degradation, and there are no data
about the possible locations of the proteases responsible for the collagenolytic/gelatinolytic
activity from the cariogenic bacteria, S. mutans.
Based on the above, further exploration of the effect of proteolytic activity of the cariogenic
bacteria S. mutans on dentinal degradation and its potential impact on the pathogenesis of caries
and secondary caries is warranted. The aim of the current study was to explore the role of S. mutans
whole cell and bacterial fractions in the degradation of type I collagen and dentinal collagen. The
hypothesis is that S. mutans has specific proteolytic activities, located in different cell extract
fractions of the bacterium that can degrade soluble type I and dentinal collagen.
3.3 Materials and Methods
3.3.1 Generic and specific MMP-like activity of S. mutans UA159
MMP-like activity from S. mutans UA159 was measured using generic and specific MMP1, 2, 8,
and 9 Assay Kits (SensoLyte ® 520 Generic MMP Activity Kit *Fluorimetric*, SensoLyte ® 520
MMP1, 2, 8, and 9 Assay Kit *Fluorimetric*, AnaSpec, San Jose, CA, USA) following the
manufacturer’s instructions [14, 15]. Overnight (O/N) cultures of S. mutans UA159 (were cultured
in chemically defined media (CDM) at 37oC for 12 hours. Whole bacteria cells from O/N culture
were separated and collected. Some of the cells were disrupted using an ultrasonic homogenizer
(20kHz, Branson Ultrasonics™ Sonifier™ SFX150 Cell Disruptor, Fisher Scientific). The whole
bacteria or lysed cells were incubated, respectively with the generic or specific MMP1, 2, 8, and
9 substrates at 37oC for 30 min. MMP9 (20 ng) was prepared and incubated with generic substrate
as a positive control to validate the assay kit. The assays were performed in a 96-well plate, and
the activities were quantified using fluorimetric plate reader (Cytation Multi - Mode Reader,
BioTek, Vermont, USA). Fluorescence values were normalized to assay buffer with the substrate
(background).

37
Statistical analysis: All experiments were performed in triplicate and results are presented as
relative fluorescence units (RFU). One-way analyses of variance (ANOVA) and Scheffe’s
multiple comparison tests (p < 0.05) were performed to validate differences in fluorescence
productions of intact and lysed bacteria against various MMP substrates. Homogeneity of variance
and normality were verified with Leven’s and Shapiro-Wilk tests, respectively.
3.3.2 Soluble type I collagen degradation by S. mutans UA159
Type I soluble rat tail collagen (3 mg/mL) (Life Technologies, Burlington, ON) was mixed with
10X phosphate buffered saline (PBS) to final concentration of 300 g/mL. The pH was adjusted
to 7.0 by 1N NaOH. Then, 50 L of solution was dispensed into 96-well plate and incubated at
37°C in humidified incubator for 30–40 min. until a firm gel is formed. Overnight (O/N) cultures
of S. mutans UA159 were prepared using chemically defined medium (CDM) as described above
(2.1). Then, collagen gels (n=3/group) were exposed to 200 μL of the following at 37oC for 24
hours:
1) 125 CDU/mL Clostridium histolyticum (C. histolyticum) collagenase (0.2 mg) (positive
control to validate the assay)
2) CDM medium (negative control to exclude degradative effect from medium)
3) Overnight (O/N) culture of S. mutans UA159 (OD600 = 0.8)
4) 1:100 fresh inoculated culture (NEW) of S. mutans UA159 (OD600 = 0.8)
Media from each well were collected for hydroxyproline assay by Ultra Performance Liquid
Chromatography and mass spectrometry (UPLC-MS, ThermoFisher LTQ, SPARC Biocentre at
the Hospital for Sick Children) assay as described previously [237] and for calculation of the dry
weight of cells for standardization..
Statistical analysis: Background measurements from the negative control values (media only)

38
were subtracted from the values of the experimental groups. A Student’s t-test was used to
determine differences of isolated hydroxyproline between experimental groups incubated with
O/N cultures and fresh-inoculated cultures of S. mutans (p < 0.05).
3.3.3 Dentinal collagen degradation by S. mutans UA159 and its discrete fractions
O/N cultures of S. mutans UA159 were prepared using ¼ Todd-Hewitt-Yeast extract (THYE) at
37oC for 12 hours. 1:100 fresh inoculated cultures were prepared using ¼ THYE at 37oC for 4
hours. Whole bacteria cells and supernatant (cell-free fraction) from O/N culture were separated
and collected. Part of the cells were disrupted using an ultrasonic homogenizer (20kHz, Branson
Ultrasonics™ Sonifier™ SFX150 Cell Disruptor, Fisher Scientific), then intracellular and
bacterial membrane fractions of S. mutans were separated, concentrated and collected (protocol in
Supplemental information 8.1).
Dentin slabs (width x length x thick: 3 mm x 3 mm x1 mm) were prepared from human molars
(University of Toronto Human Ethics Protocol #25793), demineralized in 10% phosphoric acid
for 18 hours, and then were incubated (n=3/group) with 200 L of one of the following at 37oC
for 2 weeks:
1) O/N S. mutans UA159 (OD600 = 0.8)
2) 1:100 fresh inoculated (NEW) S. mutans UA159 (OD600 = 0.8)
3) Intracellular protein fraction of lysed O/N S. mutans UA159
4) Supernatant (cell-free fraction) from O/N culture
5) Bacterial membrane fraction of lysed O/N S. mutans UA159
6) ¼ THYE medium (negative control)

39
Media from each group were collected and analyzed for hydroxyproline as described above in
2.2 [237].
Statistical Analysis: One-way analyses of variance (ANOVA) and Scheffe’s multiple comparison
tests (p < 0.05) were performed to validate differences in isolated hydroxyproline from dentin
samples among experimental groups incubated with different cultures and bacterial fractions.
Homogeneity of variance and normality were verified with Leven’s and Shapiro-Wilk tests,
respectively.
3.4 Results
3.4.1 The generic and specific MMP-like activity of S. mutans UA159
Intact and lysed S. mutans UA 159 showed similar trend of MMP-like activity (Fig. 3.1).
Background RFU were subtracted from data. Highest readings were for MMP9 substrate (7353.3
± 1968.7 RFU) and generic substrate (7378.3 ± 1122.6 RFU) for the lysed cells group, followed
by MMP9 substrate (5636.3 ± 924.0 RFU) for intact cells (p < 0.05). Lower values were measured
for MMP1 (3158.3 ± 622.7 RFU and 4375.3 ± 251.6 RFU, p < 0.05), MMP2 (2911.4 ± 969.0 RFU
and 2713.2 ± 930.1 RFU, p > 0.05), and MMP8 (1989.0 ± 1058.4 RFU and 2691.2 ±1160.0 RFU,
p > 0.05) substrates for both intact and lysed cells, respectively. Generic and MMP1-like of lysed
bacteria was significantly higher than intact bacteria (p < 0.05). The assay was validated by using
MMP9 with the generic substrate (data not shown).
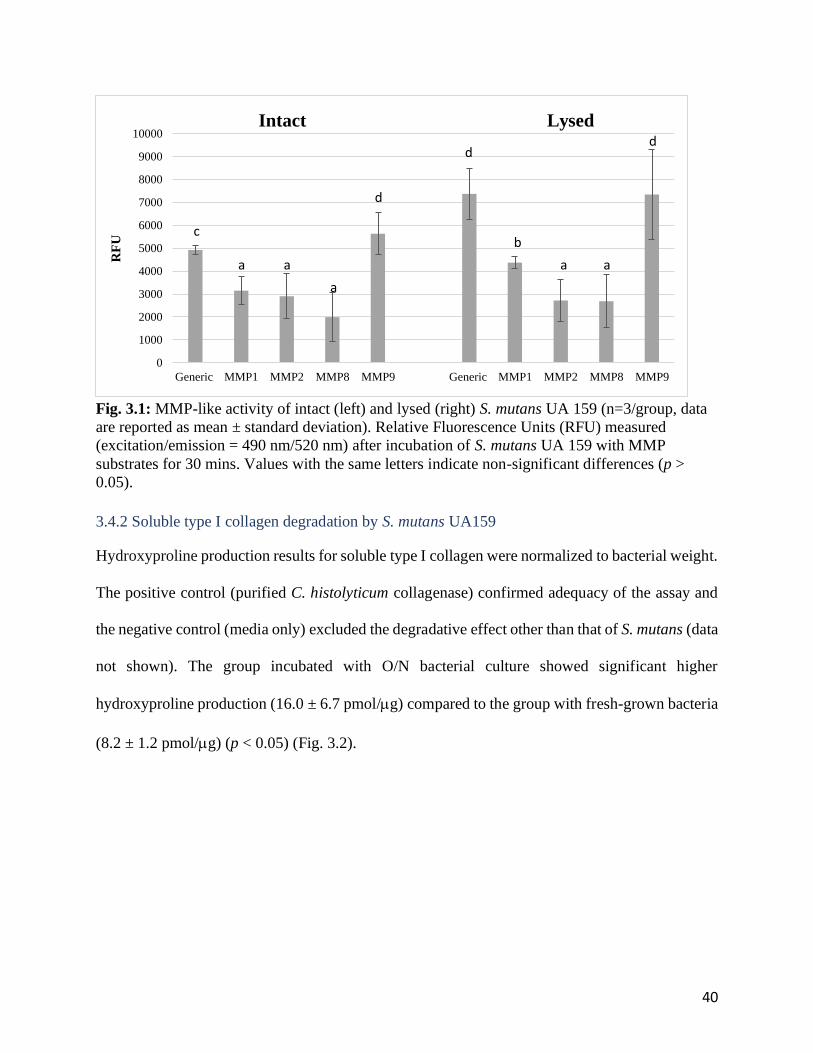
40
Fig. 3.1: MMP-like activity of intact (left) and lysed (right) S. mutans UA 159 (n=3/group, data
are reported as mean ± standard deviation). Relative Fluorescence Units (RFU) measured
(excitation/emission = 490 nm/520 nm) after incubation of S. mutans UA 159 with MMP
substrates for 30 mins. Values with the same letters indicate non-significant differences (p >
0.05).
3.4.2 Soluble type I collagen degradation by S. mutans UA159
Hydroxyproline production results for soluble type I collagen were normalized to bacterial weight.
The positive control (purified C. histolyticum collagenase) confirmed adequacy of the assay and
the negative control (media only) excluded the degradative effect other than that of S. mutans (data
not shown). The group incubated with O/N bacterial culture showed significant higher
hydroxyproline production (16.0 ± 6.7 pmol/g) compared to the group with fresh-grown bacteria
(8.2 ± 1.2 pmol/g) (p < 0.05) (Fig. 3.2).
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
Generic MMP1 MMP2 MMP8 MMP9 Generic MMP1 MMP2 MMP8 MMP9
RF
UIntact Lysed
c
a a
a
d
d
b
a a
d

41
Fig. 3.2: Hydroxyproline production after incubation of soluble type I collagen with S. mutans
UA159 (n=3/group; data are reported as mean ± standard deviation). * indicate non-significant
differences (p > 0.05)
3.4.3 Dentinal collagen degradation by S. mutans UA159 and its discrete fractions
The proteolytic activity of S. mutans UA159 and its discrete fractions towards dentinal collagen
are depicted in Fig. 3.3. The results were normalized to mass of collected components. The
medium and bacterial membrane fraction had no activity towards dentinal collagen. The O/N
culture of S. mutans has the highest degradative activity towards dentinal collagen producing 178.5
± 9.0 pmol/g hydroxyproline (p < 0.05), followed by that of supernatant (129.8 ± 1.2 pmol/g)
and intracellular component (82.8 ± 11.2 pmol/g) (p < 0.05). The least hydroxyproline (29.1 ±
5.3 pmol/g) was released from dentin slabs incubated with new inoculated S. mutans UA159 (p
< 0.05).
0
10
20
30
40
50
60
Positive control O/N NEW
Hydro
xypro
line
(pm
ol/
µg)
*
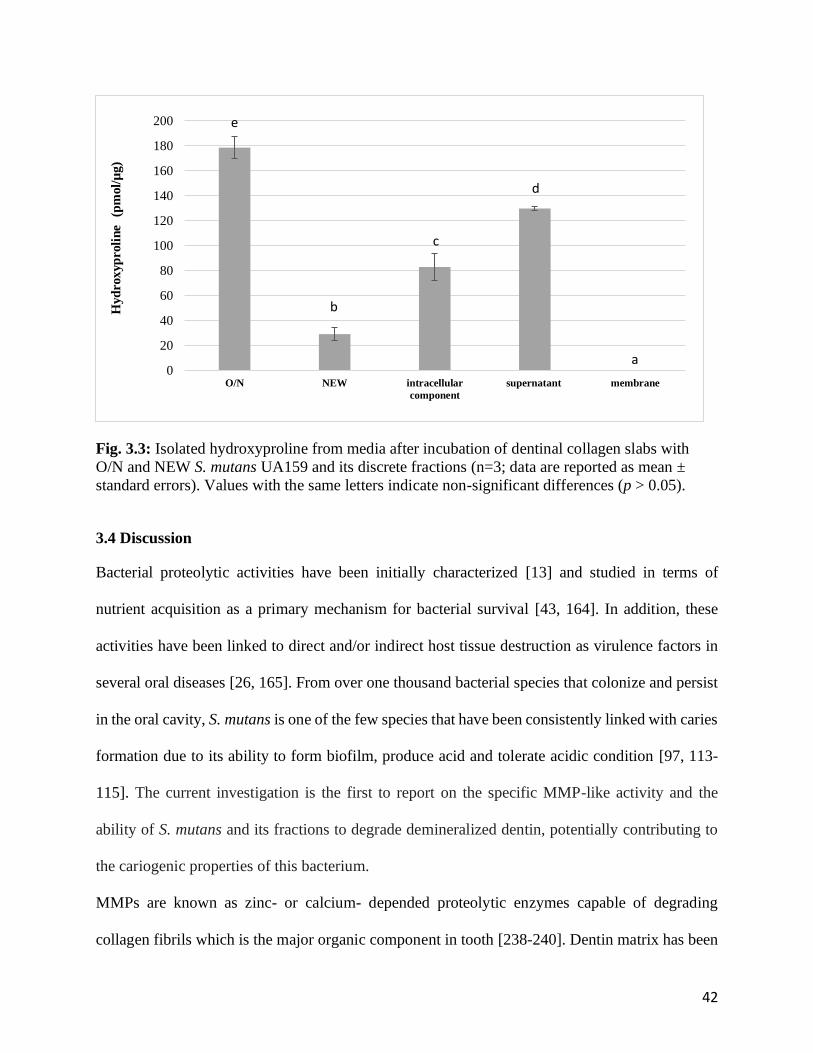
42
Fig. 3.3: Isolated hydroxyproline from media after incubation of dentinal collagen slabs with
O/N and NEW S. mutans UA159 and its discrete fractions (n=3; data are reported as mean ±
standard errors). Values with the same letters indicate non-significant differences (p > 0.05).
3.4 Discussion
Bacterial proteolytic activities have been initially characterized [13] and studied in terms of
nutrient acquisition as a primary mechanism for bacterial survival [43, 164]. In addition, these
activities have been linked to direct and/or indirect host tissue destruction as virulence factors in
several oral diseases [26, 165]. From over one thousand bacterial species that colonize and persist
in the oral cavity, S. mutans is one of the few species that have been consistently linked with caries
formation due to its ability to form biofilm, produce acid and tolerate acidic condition [97, 113-
115]. The current investigation is the first to report on the specific MMP-like activity and the
ability of S. mutans and its fractions to degrade demineralized dentin, potentially contributing to
the cariogenic properties of this bacterium.
MMPs are known as zinc- or calcium- depended proteolytic enzymes capable of degrading
collagen fibrils which is the major organic component in tooth [238-240]. Dentin matrix has been
0
20
40
60
80
100
120
140
160
180
200
O/N NEW intracellular
component
supernatant membrane
Hyd
roxyp
roli
ne
(pm
ol/
µg)
a
b
c
d
e

43
shown to contain at least five MMPs: stromelysin-1 (MMP3) [241], true collagenases (MMP1 and
MMP8) [12, 242] and gelatinases A and B (MMP2 and MMP9 respectively) [10]. Once activated,
these peptidases are responsible for the intrinsic auto-degenerative process of dentinal degradation
[18, 239, 243-246]. The extrinsic sources of MMP involved in dentinal degradation include
neutrophils and oral bacteria [15, 247]. It is assumed the contribution of extrinsic MMPs to dentin
destruction is more profound due to their greater amount and higher degradative efficiency [15,
247], which could play an important role in dentin organic matrix degradation in caries process
[16, 238]. In the current investigation fluorometric MMP assay kits were used as a first diagnostic
tool to analyze possible MMP-like activity of S. mutans. This kit uses a 5-FAM (fluorophore) and
QXL520™ (quencher) labeled FRET peptide substrates which mimic collagen backbone structure
for continuous measurement of enzymatic activities. In an intact FRET substrate, the fluorescence
of 5-FAM is quenched. Upon the cleavage of the FRET peptide by proteins with MMP-like
activities, the fluoresce of 5-FAM is recovered, and can be continuously monitored, measured and
provided as RFU. In caries lesions, as reported, the degradation of collagen is caused by the
combined efforts of multiple MMPs including MMP1, MMP8, MMP2 and MMP9 [12, 238, 248].
The collagenases (MMP1 and 8) cleave native type I, II and III collagens with intact triple-helical
structures, then the gelatinases (MMP2 and 9) further digest degraded collagen fragments or
denatured collagen [249, 250]. Accordingly, generic and specific MMP1, 2, 8, 9 activity of S.
mutans were tested in the current study.
In the current investigation, both intact and lysed cells show activity towards all MMP substrates,
suggesting possible collagenolytic (MMP1-and MMP8-like) and gelatinolytic (MMP2- and
MMP9-like) activity of S. mutans toward dentinal collagen, similarly to that of an oral pathogenic
bacterium, Enterococcus faecalis [247]. The MMP-like activity of E. faecalis has been verified as

44
a virulence factor contributing to formation of periapical lesion [247]. However, unlike E. faecalis
which has highest MMP8-like activity [247], the highest activity of S. mutans was found toward
MMP9 substrate, indicating dominant gelatinolytic activity. While both intact and lysed bacterial
cells show significant MMP-like activity, suggesting proteolytic activity for from both intracellular
and extracellular origins, the higher activity of lysed bacteria towards the generic and MMP1
substrates indicates the involvement of intracellular proteolytic enzymes within the bacterial cells.
It should be emphasized that, unlike dentinal MMPs, neither intact nor lysed bacteria required
activation procedure in order to digest the MMP substrates in the current assay, indicating that the
relevant proteases from S. mutans are in their active forms. Although the MMP-like activity test
utilized in the current study provides useful information about potential proteolytic activity of the
bacterium, the specificity and efficiency of S. mutans proteolytic activity cannot be entirely
concluded based on the synthetic MMPs substrates in this assay, since they lack structural features
and integrity of real native collagen.
The specificity and activity/efficiency of bacterial proteases varies towards different substrates
[251, 252]. Soluble type I collage is considered a relevant and practical substrate to further
investigate the degradative activity of S. mutans towards dentinal collagen, since around 90% of
the organic matrix in dentin is type I collagen [253]. The amino acid sequence of type I collagen
contains glycines, and rich in hydroxyproline and proline. Although bacterial collagenolytic
enzymes cleave collagen at different sites and generating multiple degradation fragments, the
release of hydroxyproline has been used as a suitable and reliable parameter to indicate and
quantify collagen degradation [15, 254-256]. The results of the current investigation showed a
significant increase of hydroxyproline release from type I collagen in the presence of both
overnight and newly inoculated S. mutans cultures, and no hydroxyproline release from media

45
alone, suggesting that the bacterium is the source of this protease activity and is capable of
degrading soluble type I collagen.
In the current study, the higher degradation of type I collagen by O/N S. mutans cultures compared
with NEW culture suggests a growth-phase dependency of degradative capacity of the bacterium.
This can be explained by the autolysis of S. mutans in its later growth stage to facilitate cell wall
turnover, cell division, assembly of secretion systems, resuscitation of dormant cells and micro
fratricide [257-260]. As a result, more intracellular enzymes are released into incubation medium,
contributing to enhanced collagen degradation. This explanation is also supported by the results of
the MMP-like activity in the current investigation, where increased intracellular proteases release
from the lysed cells resulted with increased measured MMP-like activity from this group compared
to that of intact cells. The increased collagen degradative activity of overnight cultures could also
be explained by the increased selective proteases production in the late growth stage of S. mutans,
which is part of bacterial adaptation strategies, where some oral pathogenic bacteria could digest
host tissue such as collagen to allow the release of amino acids as their nutrients [140, 200, 201,
257, 261].
Compared to soluble rat tail type I collagen, human dentin collagen has more complex structure at
different hierarchical levels [262, 263]. These cross-linked structures represent a state of collagen
molecules that are more resistant to enzymatic degradation than collagen in cartilage and tendon
[74, 263]. Therefore, further experiments were carried out to verify the proteolytic activity of S.
mutans towards demineralized human dentinal collagen. Although the contribution of dentinal
MMPs on dentinal collagen degradation has been reported [11], the clinical significance of their
activity is questionable due to their lower activity levels [264, 265]. In the current study, there was
no hydroxyproline release from control groups, media only group in which the dentinal MMPs are

46
the only possible source proteases. This finding supports previous statement that MMPs has
insignificant long-term effect on dentinal collagen degradation [266] due to the limited amount of
MMPs and its inactive form in dentin [10, 265, 267]. On the other hand, the significant
hydroxyproline release from overnight and fresh-inoculated S. mutans confirmed their ability to
degrade demineralized human dentin, corroborating the results for rat tail type I collagen. The
amount of hydroxyproline released from demineralized dentin was several folds more than that
from soluble type I collagen, and could be due to the prolonged incubation time (1 day vs. 14 days).
This suggests the degradative proteases are stable and can maintain their activity through extended
incubation time.
In order to further locate the responsible proteases for dentinal collagen degradation, activity from
discrete bacterial fractions and the media were investigated. Supernatants from S. mutans O/N
cultures showed higher degradative activity than intracellular components, suggesting that most of
the proteases may be secreted or released extracellularly. This finding is supported by previous
reports that most of bacterial collagenase are extracellular proteins involving bacterial invasion
[268-271]. However, it cannot be assumed that the proteases were secreted, since intracellular
proteases could be released into extracellular environment by S. mutans autolytic activity
mentioned above [259, 272]. The autolyzed bacteria count for 30% to 40% of all population in S.
mutans biofilm which significantly contributes to extracellular/supernatant proteolytic activity
[273]. In addition, it has been verified that there is significant proportion of proteases located
intracellularly based on the high hydroxyproline release from dentin samples incubated with
intracellular proteins of S. mutans.

47
3.5 Conclusion
The current investigation verified the proteolytic activity of a clinical isolate of S. mutans, a major
pathogen involved in the pathogenesis of dental caries, and the ability of this bacterium in to
degrade dentinal collagen. The initial analysis suggest that the bacterial proteases originate from
both intra- and extracellular origins. Further characterization of the bacterium degradative activity,
degradative mechanisms, and identification of specific proteases that are involved in this process
is needed.

48
Chapter 4 Manuscript
Characterization of proteolytic activity and identification of responsible proteolytic
enzymes of Streptococcus mutans towards dentinal collagen
Bo Huang1, Lida Sadeghinejad1, Walter Siqueira, Christopher McCulloch1, J. Paul
Santerre1,2, Dennis G. Cvitkovitch1,2, Yoav Finer1,2
1Faculty of Dentistry, University of Toronto, Ontario, Canada
2Institute of Biomaterials and Biomedical Engineering, University of Toronto
4.1 Abstract
Objectives: to explore the possible mechanisms of Streptococcus mutans (S. mutans) in the
degradation of dentinal collagen, to characterize proteolytic activity of S. mutans and to identify
responsible specific proteolytic enzymes.
Materials & Methods: Human dentin slabs (DS) from molar teeth were non-demineralized (ND)
or demineralized with either lactic acid (LADS) or phosphoric acid (PADS). Non-demineralized,
LADS or PADS±heat-treatment (HT) were incubated with intracellular proteins (IP) of S. mutans
UA159. The sequence of degraded human dentin collagen fragment was analyzed by SDS-PAGE
and Mass spectrometry. SMU_759, SMU_761 and SMU_1438c, putative enzymes with possible
collagenolytic/gelatinolytic activities were selected based on a search in the genome of S. mutans
UA159 and their expression profile that was analyzed by proteomics. These S. mutans enzymes
were cloned and expressed in E. coli BL21. The enzyme degradation effect of PADS was analyzed
by quantification of degradation by-products, hydroxyproline.
Results: Highest release of hydroxyproline by intracellular proteins from DS was measured from
lactic acid treated group, IP+LADS+HT (414.4±69.9pmol/µg) (p < 0.05), the least amount was
from demineralized DS in buffer, PBS + PADS (0.2±0.01pmol/µg) (p < 0.05). SDS-PAGE/MS
confirmed the presence of collagen fragments from 1 chain of type I collagen. SMU_759 had the

49
highest degradative activity towards dentinal collagen (219.0 ± 11.2 pmol/g), followed by
SMU_761 (76.8 ± 15.3 pmol/g), while SMU_1438c had no activity towards dentin (p<0.05).
Conclusion: The demineralization process of tooth structure is a critical step for further bacterial
protease induced dentinal collagen degradation and several specific proteases have been isolated
with degradative activities that could contribute dentinal structure destruction and caries formation.

50
4.2 Introduction
Collagens are the most abundant proteins in human body structures as primary extracellular matrix
in the ultra-structure of organs and tissues. The type I collagen fiber is the most abundant in human
tissues. It is a rod-like molecule, roughly 300 nm long, comprised of two α1(I) and one α2(I) left-
handed helix polypeptide chains twisted around a common axis to form a major right-handed helix
[56, 57]. Within the triple helical domain, there is a common triplet sequence glycine-X-Y, where
X and Y are often proline and hydroxyproline [58]. In the human tooth, the organic matrices
contain collagen, mainly type I, and non-collagenous proteins (NCPs) including phosphoproteins,
proteoglycans, and acidic glycoproteins [61-63].
Collagen degradation is involved in both physiological and pathological processes [23, 194].
Collagenases are a group of proteases which are capable of cleaving native collagen under
physiological conditions. Subsequently, the gelatin in the degraded collagen fragments can be
further hydrolyzed by gelatinase. Bacterial collagenases were identified and reported as virulence
factors contributing to human disease [22]. Extensive research has been carried out to investigate
the key roles of bacterial collagenase in host colonization [22, 23]. The most well-known microbial
collagenases are from Clostridium [24], followed by Bacillus and Vibrio [22, 25]. Oral bacterial
collagenolytic proteases were identified, characterized and reported as virulence factors
contributing to periodontal disease [26]. Two extracellular S. mutans proteases that are capable of
hydrolyzing synthetic collagen substrate, PZ-Pro-Leu-Gly-Prop-Arg (PZ-PLGPA) and
furylacryloyl-Leu-Gly-Pro-Ala (FALGPA) were isolated [28]. However, their definitive role in
the breakdown of the collagen component of tooth structure has been controversial due to the non-
representative synthetic substrates used to characterize these enzymes [28]. Recently, our team
have reported on the degradative activity of S. mutans toward human dentinal collagen degradation,
suggesting the potential pathogenic role of the bacterium in tooth structure distraction and caries

51
progression (paper 1). And, it has been postulated that the responsible proteolytic enzymes are
located both intra- and extracellularly (paper 1). A recent study reported the expression of putative
collagenases by S. mutans UA 159 isolated from root caries [31], however these proteases have
yet to be characterized.
Considering the reported high activity, high efficiency and continuous production of bacterial
collagenolytic enzymes [13, 23, 30], further exploration of the characterization of proteolytic
activity of S. mutans and identification of responsible proteolytic enzymes on dentinal degradation
and their potential impact on the pathogenesis of caries is warranted. Building on studies that
reported on putative collagenase genes and degradative activity from S. mutans [26, 30, 32], the
aim of the current study was to elaborate the pathogenic role of S. mutans in the degradation of
dentinal collagen by identification and characterization of e specific proteolytic activity of S.
mutans. The hypothesis is S. mutans has collagenolytic and/or gelatinolytic proteolytic enzymes
that degrade dentinal collagen.
4.3 Materials and Methods
4.3.1 Characterization of proteolytic activity of intracellular proteins of S. mutans
S. mutans UA 159 was cultured in ¼ THYE at 37oC for 12 hours. This overnight (O/N) S. mutans
cells were disrupted using an ultrasonic homogenizer (20kHz, Branson Ultrasonics™ Sonifier™
SFX150 Cell Disruptor, Fisher Scientific) and intracellular proteins were separated, concentrated
and collected (protocol in Supplemental information 8.1). Total protein concentration was assessed
by Micro Bicinchoninic Acid (Micro BCA) assay [274] and equal protein amount (75g) was
collected for further incubation experiments.
Dentin slabs (DS) (width x length x thick: 3 mm x 3 mm x1 mm) were prepared from human
molars (University of Toronto Human Ethics Protocol #25793) and pre-treated with one of the
following protocols (n=3/group):

52
1) demineralized with 10% phosphoric acid (PA)(Ricca Chemical Company, Arlington, TX,
USA) for 18 hours (PADS) [275]
2) demineralized with 10% phosphoric acid for 18 hours, then denatured collagen by boiling
for 5 mins (heat treatment) (PADS+HT)
3) demineralized with 0.1 M lactic acid for 36 hours (LADS) [276]
4) demineralized with 0.1 M lactic acid for 36 hours, then boiled for 5 mins (LADS + HT)
5) Non-demineralized (ND)
6) Non-demineralized, denatured collagen by boiling for 5 mins (ND+HT)
Pretreated dentin samples mentioned above were incubated either with PBS (control) or extracted
intracellular proteins (IP) (experimental groups) at 37oC for 2 weeks. Media from each well were
collected for hydroxyproline assay by Ultra Performance Liquid Chromatography and mass
spectrometry (UPLC-MS, Waters Aquity, Waters Corporation, Milford, MA) as described
previously [237] .
Statistical Analysis: Homogeneity of variance and normality were verified with Leven’s and
Shapiro-Wilk tests, respectively, then one-way analyses of variance (ANOVA) and Scheffe’s
multiple comparison tests (p < 0.05) were performed to validate differences in hydroxyproline
productions among control and experimental groups.
4.3.2 Verification of dentinal collagen degradation by intracellular proteins of S. mutans using
SDS-PAGE and Mass Spectrometry
Equal amount of intracellular proteins (75 g) were collected from O/N culture of S. mutans UA
159 as described above (2.1) [274]. Dentin slabs were prepared as described above (2.1) and
demineralized in 10% phosphoric acid for 18 hours, then incubated at 37oC for 2 weeks, with one
of the following (n=3/group):
1) C. histolyticum collagenase (positive control)

53
2) PBS (negative control)
3) 75 g of protein (experimental groups)
In addition, pure C. histolyticum collagenase and extracted intracellular proteins of S. mutans were
incubated for 2 weeks without dentin samples as benchmark.
The media containing dentinal collagen degradation products from all groups were separated.
Verification of collagen degradation products within the incubation media was by 15% sodium
dodecyl sulfate -polyacrylamide gel electrophoresis (SDS-PAGE) as described previously [251].
Briefly, the suspected collagen degradation fragments were presented as peptide bands on SDS-
PAGE gel. Two bands of interest from experimental groups were collected, digested and analyzed
by mass spectrometry as described previously [277] by using LC-MS/MS (ThermoFisher LTQ,
SPARC Biocentre at the Hospital for Sick Children). All MS/MS samples were analyzed using
MS-Amanda Proteome Discoverer (Research Institute of Molecular Pathology, Vienna, Austria;
version AmandaPeptideIdentifier in Proteome Discoverer 2.2.0.388).
4.3.3 Bioinformative analysis of putative genes of collagen-degrading proteases in S. mutans
UA159
Putative genes of collagen-degrading proteases were searched in the S. mutans UA159 genome
database at The National Center for Biotechnology Information (NCBI)
(http://www.ncbi.nlm.nih.gov). Nucleotide and deduced amino-acid sequences were analyzed
using MacVector software. The presence of signal peptide was searched using the default settings
of Gram-negative bacteria on the SignalP Server 4.1 to predict if the protein of interest was
secreted [278]. Furthermore, template search was performed for the deduced amino-acid sequences
using Phyre2 (http://www.sbg.bio.ic.ac.uk/~phyre2/html/page.cgi?id =index) [279]. The
templates with the highest scoring crystal structure were then selected for analysis.

54
4.3.4 Protein Identification of putative collagen-degrading proteases in S. mutans UA 159
18-hour biofilm of S. mutans UA 159 were cultured in TYEG medium buffered at pH 5.5 with
MES (Sigma-Aldrich, St. Louis, MO, USA) at 37oC. Then, the biofilm cells were collected and
disrupted using a homogenizer (Thermo Savant, FastPrep FP 101) and the proteins were collected.
Equal protein amount (20 µg) were dried, denatured, and reduced for 2 hours by the addition of
200 µL of 4 M urea, 10 mM dithiothreitol (DTT), and 50 mM NH4HCO3, pH 7.8. After four-fold
dilution with 50 mM NH4HCO3, pH 7.8, tryptic digestion was carried out for 18 h at 37oC,
following the addition of 2% (w/w) sequencing-grade trypsin (Promega, Madison, WI, USA).
Peptide separation and mass spectrometric analyses were carried out as previously described [274,
280]. The obtained MS/MS spectra were searched against the streptococci protein database to
verify expression of putative collagen-degrading proteases (Swiss Prot and TrEMBL, Swiss
Institute of Bioinformatics, Geneva, Switzerland, http://ca.expasy.org/sprot/) using SEQUEST
algorithm in Proteome Discoverer 1.3 software (Thermo Scientific, San Jose, CA, USA).
4.3.5 Cloning, expression and purification of bacterial collagen-degrading proteases
Basic Local Alignment Search Tool (BLAST) identified several putative esterase genes in the S.
mutans UA159 genome database. The gene candidates (SMU_759, SMU_761, SMU_1438c), were
selected based on the results of section 2.4 above, were PCR amplified from S. mutans UA159
genomic DNA by using designed primers (Fig. 4.1), then cloned into the pCOLDII vector as
previously described (empty vector was used as a control) [281], providing an N-terminal hexaHis
tag (His6) and the factor Xa cleavage site (IEGR). The enzymes were expressed in E. coli BL21
(DE3) then harvested for further protein isolation and purification (Bio Basic Inc, Wuhan, China
PRC). Cells were re-suspended in binding buffer [50 mM Hepes (pH 7.5), 100/300 mM NaCl, 10
mM imidazole and 2% glycerol (v/v)], lysed using a sonicator, and cell debris was removed via
centrifugation at 30,000G (Eppendorf, Hamburg, Germany). Cleared lysate was loaded onto a 5
55
mL Ni-NTA column (QIAGEN, Dusseldorf, Germany) pre-equilibrated with binding buffer, and
washed. Proteins were eluted using the above buffer with 20 mM or 250 mM imidazole. Fractions
containing the protein of interest were identified by 12.5% SDS-polyacrylamide gel
electrophoresis and further purified via gel filtration on a HiLoad 16/60 Superdex75 prep-grade
column [10 mM Hepes (pH 7.5) and 50 mM KCl].
SMU_759
SMU_761

56
SMU_1438c
Fig. 4.1: The primers and pCOLDII vector information for gene expression.
4.3.6 Degradation of dentinal collagen by SMU_759, SMU_761 and SMU_1438c
Dentin slabs (DS) were prepared as described above (4.3.1) and were demineralized in 10%
phosphoric acid for 18 hours. Then, 200 L of collagen gels were mixed with PBS, polymerized
(pH 7.4) and coated in 48-well plate at 37 for 12 hours. The collagen samples (3 mg/mL, n=3/group)
or DS were exposed to 200 L of 1 mg/mL of SMU_759, SMU_761 or SMU_1438c, respectively,
or 200 μL of 125 1 mg/mL C. histolyticum collagenase (0.2 mg of C. histolyticum collagenase)
(positive control), or 200 μL of PBS (negative control) at 37oC for 24 hours (collagen gels) or 2
weeks (dentin slabs). Medium from each well was collected and filtered for hydroxyproline assay
as described above (4.3.1) [237].
Statistical Analysis: Homogeneity of variance and normality were verified with Leven’s and
Shapiro-Wilk tests, respectively, then one-way analyses of variance (ANOVA) and Scheffe’s
multiple comparison tests (p < 0.05) were performed to validate differences in hydroxyproline
productions among controls incubated with PBS and the experimental groups incubated with
SMU_759 SMU_761 or SMU_1438c.

57
4.4 Results
4.4.1 Characterization of proteolytic activity of intracellular proteins of S. mutans
Hydroxyproline release by S. mutans intracellular proteins vary depending on different
pretreatment of dentin collagen specimens (Fig. 4.2). The lowest amounts of hydoxyproline were
released from the phosphoric acid demineralized group incubated without presence of intracellular
proteins, PBS + PADS (0.2±0.01pmol/µg), followed by lactic acid demineralized group, PBS +
LADS (0.3±0.1pmol/µg) (p < 0.05). Low amounts of hydroxyproline were measured from the
non-demineralized dentin incubated with intracellular proteins of S. mutans (IP + ND ± HT) (p <
0.05); heat treatment had no significant effect on the release of degradation product (IP + ND +
HT (19.5±12.5pmol/µg) vs. (IP + ND - HT) (13.12±12.62pmol/µg) (p > 0.05). Higher amounts of
hydroxyproline were detected for specimens demineralized with phosphoric acid with and without
heat treatment (IP + PADS ± HT) (p < 0.05), and there is no significant difference between heated
and non-heated groups, IP + PADS + HT (111.2±9.9pmol/µg) vs. IP + PADS – HT
(106.6±11.7pmol/µg (p > 0.05). The highest release was measured for dentin specimens that were
demineralized by lactic acid with and without heat-treatment (IP + LADS ± HT) (p < 0.05), and
there is no significant difference between heated and non-heated groups, IP + LADS + HT
(414.4±69.9pmol/µg) vs. IP + LADS – HT (398.9±53.8pmol/µg) (p > 0.05). There was no
significant effect of heat-treatment on hydroxyproline release in both phosphoric acid and lactic
acid demineralized groups (p > 0.05).

58
Fig. 4.2: Hydroxyproline production from dentinal collagen slabs treated with various methods
and incubated by intracellular proteins of S. mutans UA159 or media (n=3; data are reported as
mean ± standard deviation). Values with the same letters indicate non-significant differences (p >
0.05). DS (demineralized Slab); ND (Non-demineralized Dentin); PA (Phosphoric Acid
demineralized); LA (Lactic acid demineralized); HT (Heat treatment)
4.4.2 Verification of dentinal collagen degradation by intracellular proteins of S. mutans using
SDS-PAGE and Mass Spectrometry
The degradation fragments of dentinal collagen by intracellular proteins are presented as peptide
bands on the SDS-PAGE gel image (Fig. 4.3). The bands in lane 1 represent degraded collagen
fragments by C. histolyticum collagenase; in lane 2 (negative control), there are only two distinct
bands close to each other around 120 KDa presenting typical 1- and 2- chains of type 1 collagen,
and there is a smear spreading from 120 KDa to 8 KDa. Lane 3 and 4, the bands represent S.
mutans UA159 intracellular proteins and degraded dentinal collagen fragments. In lane 5 the bands
represent the pure C. histolyticum collagenase. In lane 6, the bands represent the intracellular
proteins extracted from S. mutans UA159. By comparing to dentin specimens incubated with PBS
(controls, lane 2) and extracted bacterial protein of S. mutans UA159 (lane 6), there are multiple
extra bands distinctly presented in lane 3 and 4 (Fig. 4.3), which could be considered as degraded
0
100
200
300
400
500
600
PBS+ PADS IP+ PADS IP+ PADS +
HT
PBS+ LADS IP + LADS IP + LADS +
HT
IP + ND IP + ND +
HT
Hyd
roxyp
roli
ne
(pm
ol/
µg )
a b
d d
ee
c c
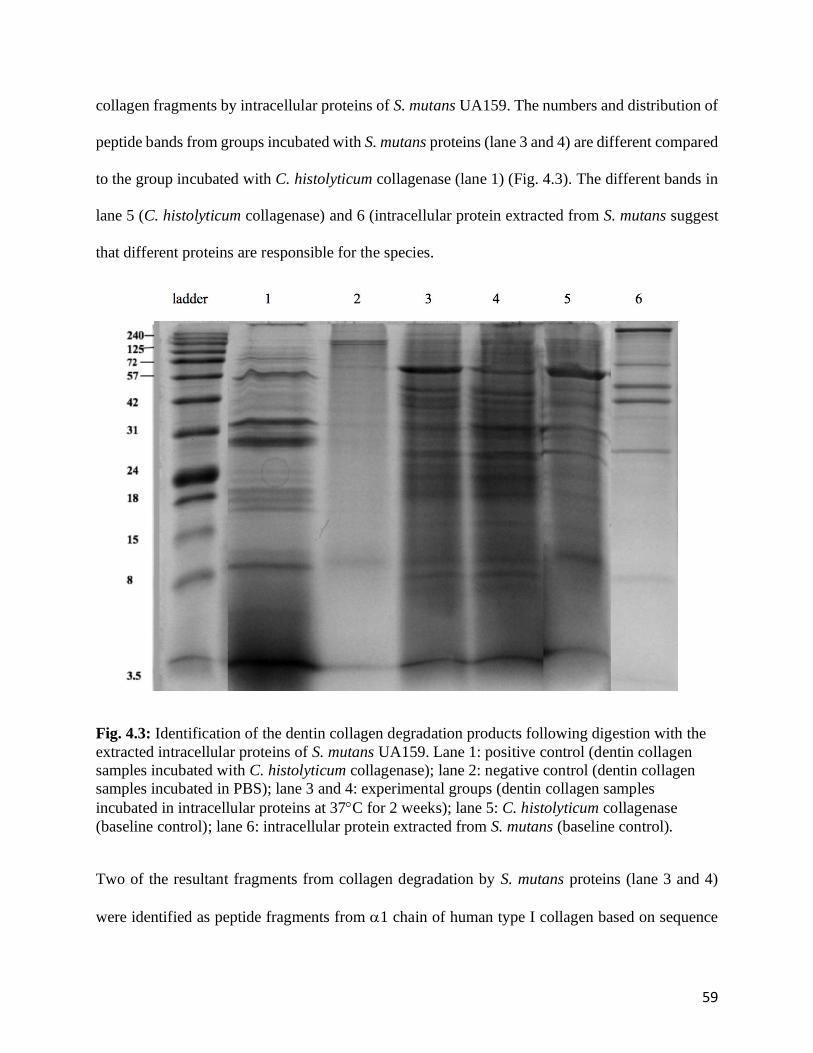
59
collagen fragments by intracellular proteins of S. mutans UA159. The numbers and distribution of
peptide bands from groups incubated with S. mutans proteins (lane 3 and 4) are different compared
to the group incubated with C. histolyticum collagenase (lane 1) (Fig. 4.3). The different bands in
lane 5 (C. histolyticum collagenase) and 6 (intracellular protein extracted from S. mutans suggest
that different proteins are responsible for the species.
Fig. 4.3: Identification of the dentin collagen degradation products following digestion with the
extracted intracellular proteins of S. mutans UA159. Lane 1: positive control (dentin collagen
samples incubated with C. histolyticum collagenase); lane 2: negative control (dentin collagen
samples incubated in PBS); lane 3 and 4: experimental groups (dentin collagen samples
incubated in intracellular proteins at 37C for 2 weeks); lane 5: C. histolyticum collagenase
(baseline control); lane 6: intracellular protein extracted from S. mutans (baseline control).
Two of the resultant fragments from collagen degradation by S. mutans proteins (lane 3 and 4)
were identified as peptide fragments from 1 chain of human type I collagen based on sequence

60
homology and specificity analysis (Fig. 4.4a.). These resultant peptide sequences were highlighted
in the sequence of human type I collagen 1 chain (Fig. 4.4b).
a
b
Fig. 4.4: Identification of peptide sequence from dentinal collagen degradation by S. mutans
UA159 intracellular proteins. The origin of degraded fragments was identified based on
sequence homology against human type I collagen (Fig. 4.4a); peptide sequences were
highlighted in sequences of human type I collagen 1 chain (Fig. 4.4b).

61
4.4.3 Bioinformative analysis of putative genes of collagenolytic/gelatinolytic proteases in S.
mutans UA159
Five putative collagenase or gelatinase gene were identified from S. mutans UA159 genome
database based; SMU_1438c and SMU_1784c were identified as genes of Zn-dependent protease;
prpO peptidase was identified as a gene of zinc metalloproteinase; SMU_759 and SMU_761 were
identified as genes of protease related to collagenases. Signal peptides were reported for SMU_759
and SMU_761, indicating that both of them are secreted proteins.
4.4.4 Protein Identification of putative collagen-degrading proteases in S. mutans UA 159
Bacterial proteases SMU_759, SMU_761 and SMU_1438c, coded by putative gene SMU_759,
SMU_761 and SMU_1438c showed a consistent elution from S. mutans UA159, confirming their
expression at the protein level, and therefore were chosen for further investigations. Further details
regarding each protein structure and mass are provided in the supplemental document.
4.4.5 Cloning, expression and purification of bacterial collagenolytic/gelatinolytic proteases
Two intracellular (SMU_759, SMU_761) and one trans-membrane (SMU_1438c) enzymes were
expressed in E. coli BL21 as soluble enzymes. Homogeneity of the purified protein was confirmed
by SDS-PAGE as a single molecular subunit mass (Fig.4. 5). The molecular mass of SMU_759 is
33 KDa, which is similar to the molecular mass calculated from the deduced amino-acid sequence.
The molecular mass of SMU_761 is 58 KDa, which is higher than predicted value. The molecular
mass of SMU_1438c is 28 KDa as predicted.
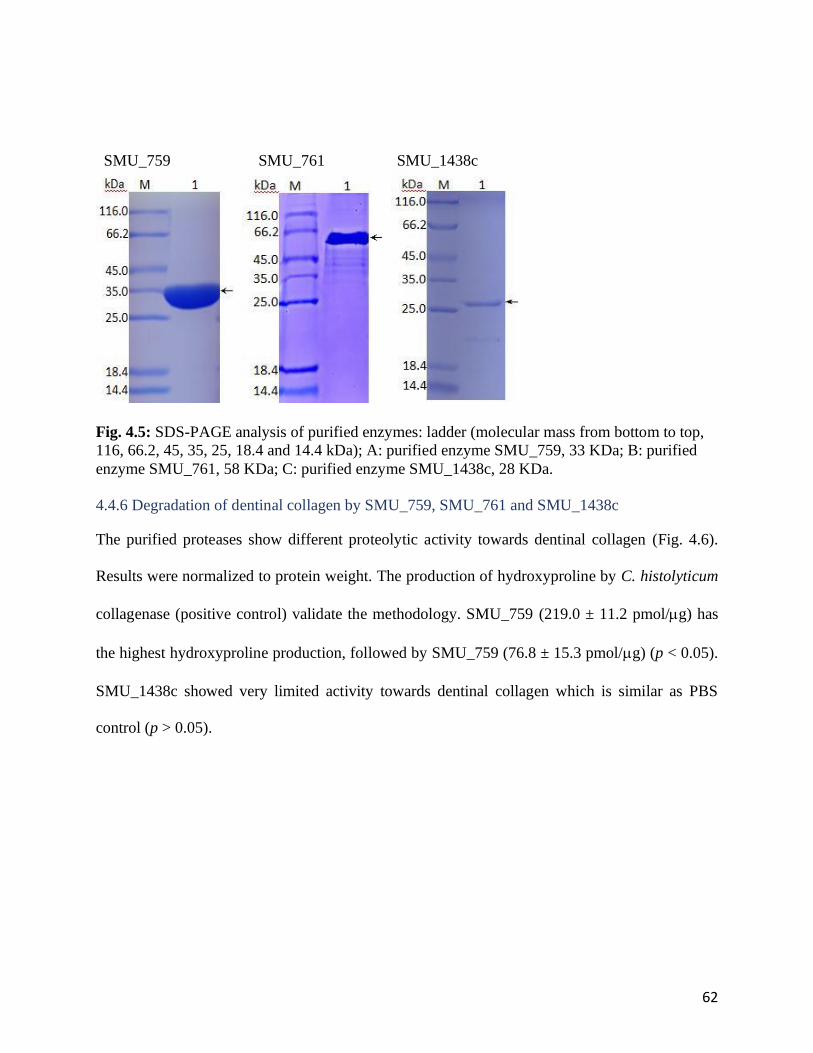
62
SMU_759 SMU_761 SMU_1438c
Fig. 4.5: SDS-PAGE analysis of purified enzymes: ladder (molecular mass from bottom to top,
116, 66.2, 45, 35, 25, 18.4 and 14.4 kDa); A: purified enzyme SMU_759, 33 KDa; B: purified
enzyme SMU_761, 58 KDa; C: purified enzyme SMU_1438c, 28 KDa.
4.4.6 Degradation of dentinal collagen by SMU_759, SMU_761 and SMU_1438c
The purified proteases show different proteolytic activity towards dentinal collagen (Fig. 4.6).
Results were normalized to protein weight. The production of hydroxyproline by C. histolyticum
collagenase (positive control) validate the methodology. SMU_759 (219.0 ± 11.2 pmol/g) has
the highest hydroxyproline production, followed by SMU_759 (76.8 ± 15.3 pmol/g) (p < 0.05).
SMU_1438c showed very limited activity towards dentinal collagen which is similar as PBS
control (p > 0.05).

63
Fig. 4.6: Hydroxyproline production after incubation of dentinal collagen with SMU_759,
SMU_761 and SMU_1438c (n=3; data are reported as mean ± standard errors). Values with the
same letters indicate non-significant differences (p > 0.05).
4.5 Discussion
Collagen degradation takes place during various physiological and pathological conditions, such
as bone and embryonal development, malignant tumor invasion, wound repair, pathogenic
microorganism invasion and chronic periodontal inflammation [23]. In the oral environment,
dentinal (endogenous) collagenases, bacterial collagenolytic proteases and neutrophils have been
linked to destructive collagen degradation leading to various oral diseases [11, 251, 282]. Although
the collagenolytic activity of S. mutans toward collagen [13, 26] and dentinal collagen (paper 1)
have been investigated, the current investigation is the first to report the identification and initial
characterization of specific proteolytic bacterial enzymes and their activity toward dentinal
collagen.
0
50
100
150
200
250
PBS SMU_759 SMU_761 SMU_1438c
Hyd
roxyp
roli
ne (
pm
ol/
µg
)
a
c
b
a

64
4.5.1 The characteristics of collagenolytic/gelatinolytic activity of S. mutans intracellular
proteins
The dentin demineralization procedure by phosphoric acid was adopted by this study due to its
clinical relevance to etch-and-rinse bonding protocol of dental restorative procedures. In this
procedure, 32 to 37% phosphoric acid was used to expose collagen fibril meshwork for
micromechanical retention of adhesive resins [283]. Dentin demineralization in the current study
was also performed with lactic acid, produced by the cariogenic bacteria, S. mutans and responsible
for pathological demineralization in the caries process [38, 131]. Previous studies reported that
latent dentinal MMPs can be activated by mild etching acids and then could initiate dentinal
collagen degradation [284, 285]. However, the etching procedure is transient and superficial,
therefore the amount of activated MMPs by etching procedure is limited. The lack of
hydroxyproline found for dentin slab incubated ion PBS after demineralization by phosphoric acid
in the current investigation support this notion.
It has also been claimed that bacterial acids can activate MMPs by cleaving their pro-domains,
which could link MMPs to collagen destruction in primary and secondary caries formation [238].
However, MMPs are neutral proteases, and are not able to stay active at low pH [20, 244, 286].
The previous studies put the merits on the recovery of MMPs activity by pH neutralization due to
dentinal or salivary fluid fluctuation [8, 238, 287]. Considering the depth of caries and the limited
access to the deep area beneath restoration of secondary caries, the buffering effect of dentinal
fluid or saliva to local pH in the carious areas is questionable. In other words, MMPs are
continuously exposed to an acidic environment in deep carious areas created by constant lactic
acid production by cariogenic bacteria without sufficient buffering [288-291]. Since it was
previously reported that low pH denatures MMPs, a reasonable doubt can be raised regarding
dentinal MMPs’ contribution to caries or secondary caries formation [20, 286]. This is also

65
supported by current finding that there is rarely any hydroxyproline production from lactic acid
treated samples when there is no intercellular protein present in the incubation media, suggesting
virtually no degradative impact on dentin by endogenous MMPs alone.
Although most bacterial proteases cannot be accounted for true collagenase which directly
hydrolyze native collagen molecules with unique specificity [192, 194, 292], it is important to
stress that a large number of bacterial proteases have the capacity to hydrolyze single-stranded and
denatured collagen polypeptides [22]. The current investigation suggests a mechanism for S.
mutans proteases utilization of its lactic acid production as a “pre-treatment” of dentin to allow the
bacterium to degrade dentinal collagen, potentially contributing to the caries process; the results
of the current investigation, demonstrate that S. mutans proteases could not degrade non-
demineralized/non-acid-pretreated dentin slabs as demonstrated by the very low level of
hydroxyproline release. Even after heat-treatment which denatured the collagen fibers, the non-
demineralized dentin was still resistant to degradation, while both phosphoric acid and lactic acid
demineralized dentin released significantly more hydroxyproline. These findings suggested there
are two stages of tissue destruction in caries [293, 294]: acid-initiated demineralization of dentine
minerals provides access for bacterial proteases, followed by collagen breakdown in which acid
may also play a critical role to aid bacterial proteases induced degradation.
It has been reported that acids could release non-collagenous proteins (NCPs) which are part of
organic matrix of tooth dentin other than collagen [69, 295]. NCPs play critical roles in maintain
collagen integrity by serving as nuclei for organization of collagen fibrils [70, 71] and inducing
collagen intermolecular cross-links [72, 73]. As a result, the acid-induced NCPs release alters the
collagen macrostructural and conformation [44, 276] and thereby increase the susceptibility of the
collagen molecules to enzymatic degradation [296-299]. Other studies also suggested that acids

66
could change inter-chain bonds affecting inter- and intramolecular cross-links of collagen [300,
301]. The current investigation corroborates that and suggest that bacterial acid had a destructive
effect on dentinal collagen structural integrity, while the heat-denaturing treatment after acid-
demineralization of dentin samples did not significantly increase hydroxyproline production (Fig.
4.2). In addition, the current study also showed that the type of acid used for demineralization
affected the degradation of dentinal collagen by bacterial proteases; demineralizing the dentin by
lactic acid released significantly more hydroxyproline compared to that of phosphoric acid. It is
assumed that lactic acid, as a by-product from cariogenic bacteria, has more destructive effects on
collagen structures by either releasing larger amount of NCPs [302] or reducing cross-linking of
collagen [300, 301]. The above findings are supported by previous study, suggesting that lactic
acid is a prerequisite for non-specific proteases, such as bacterial proteases, to degrade
ethylenediaminetetraacetic acid (EDTA) demineralized dentin samples [276].
Bacterial collagenolytic proteases have a broader range of specificity, but their products are
hydrolyzed at various specific peptide bonds [25, 303, 304]. The main source of knowledge on
bacterial collagenases is based on multiple studies on the enzymes produced by C. histolyticum
[24, 270, 304]. In the current study, the purified collagenase from C. histolyticum was used as a
positive control to analyze the degradative effect and pattern of proteases from S. mutans. As a
baseline, the type I collagen released from demineralized dentin incubated with PBS buffer were
collected and presented on the SDS-PAGE gel as two distinct bands indicating typical 1- and
2-chains of Type I collagen. The absence of A chain which is characteristic 3/4-cleavage products
by true collagenase cleavage of intact type I collagen suggests the limited effect of dentinal
endogenous MMPs to dentinal collagen degradation [12]. The smear presented on the SDS-PAGE
gel from the demineralized dentin incubated with PBS buffer indicates possible NCPs release after

67
demineralization procedure or denatured collagen fragments. The degraded collagen fragments by
S. mutans proteases presented as multiple bands on SDS-PAGE gel. Comparing to C. histolyticum
collagenase treated dentin collagen, the numbers and distribution of bands from samples treated
with S. mutans intracellular proteins are different. This indicates that the cleavage sites on collagen
targeted by S. mutans proteases are different from that of C. histolyticum, and that different
enzymes are responsible for the cleavage. The primary-structural analysis of two resultant collagen
fragments derived from dentinal collagen upon digestion with S. mutans intracellular proteins
confirmed the degraded peptides were from 1 chain of type I collagen, suggesting that the
enzymes might preferably act on certain peptide sequences. Although not all of the possible
cleavage sites in collagen were determined, several preferred amino acids were suggested as
cleavage sites, including Lys, Gly, Ser and Arg, which have been reported also for other bacterial
collagenolytic proteases [25, 268, 269]. However, the proteases’ specificity cannot be identified
due to mixed effect of multiple proteases in the intracellular components. As a result, specific
collagenolytic/gelatinolytic proteases from S. mutans were synthesized for more detailed
investigation.
4.5.2 The specific collagenolytic/gelatinolytic proteases
Out of five putative genes identified by bioinformative search and based on the proteomics
analyses that verified that their coded proteases have been produced by S. mutans UA159, three
genes, SMU_759, SMU_761, and SMU_1438c, were selected to be expressed. SMU_1438c, was
identified as an interstitial collagenase with structural similarity to human pro-MMP-1 containing
the HEXXH peptide consensus sequence usually found in metalloproteinases [22, 305]. SMU_759
and SMU_761, were listed belonging to U32 family of collagenases, which relates to virulence
factors of various human-pathogenic bacteria [306, 307] and were previously reported to exist in

68
root caries lesions (31). The U32 family of collagenases is one of a few that the catalytic domain
and structure have not been fully described [308]. The most well-studied U32 family member from
oral pathogen is PrtC isolated from Porphyromonas gingivalis (P. gingivalis), which plays a
critical role in periodontal tissue destruction and bacterial invasion [251]. Signal sequence
prediction suggested both SMU_759 and SMU_761 are secreted proteins containing
transmembrane domains which was similar to PrtC from P. gingivalis [147], and, that their
predicted structures have the characteristic compact distorted open barrel made up of -strands
and may function in protein binding [147, 309]. However, the secretion mechanism is not clear
and still under investigation [310, 311].
The SDS-PAGE result indicated that the synthesized proteases were in monomeric form, and that
their molecular masses ranged from 28 kDa to 58 kDa, similarly to all reported microbial
collagenolytic proteases [22]. Both SMU_759 and SMU_761 are capable of degrading
demineralized dentinal collagen. The highest activity was found for SMU_759, which is
comparable to the activity of overnight whole cells (paper 1), and SMU_761 showed similar
activity as supernatant (cell-free fraction from O/N culture) (Paper 1). This finding indicates that
both enzymes are major contributors to proteolytic activity of S. mutans towards dentinal collagen.
As discussed above, the acid demineralized dentin could have lost its original interstitial structures,
so SMU_659 and SMU_761 may not be considered as true collagenases. This finding is similar to
a previous report that one recombinant U32 peptidase of a non-pathologic bacteria was only
capable of degrading heat-denatured collagen [308]. Although the U32 family has been recognized
as a collagenase group, multiple studies have reported conflicting results regarding their activity
against collagen substrates [308, 312, 313]. Based on protein sequence and structure analysis,
previous studies have identified several U32 family members which showed significant

69
heterogeneity of substrate specificity. The PrtC from P. gingivalis degraded soluble collagen but
not gelatin [147]. In contrast, the other U32 family member from Pseudoalteromonas agarivorans
(P. agarivorans) is capable of cleaving native collagen and gelatin [312]. In addition, other factors
such as ions also play role in U32 proteases activity, which could also explain the different
degradative susceptibility of dentin slabs following lactic acid versus phosphoric acid
pretreatments. The recombinant Filifactor alocis U32 protease (PrtFAC) interacted with and
degraded type I collagen in a Ca2+ dependent manner similar to the P. agarivorans U32 collagenase
[313], while Zn2+ showed inhibitory effect [147]. Considering the complexity of oral conditions,
further characterization of SMU_759 and SMU_761, previously found in caries lesions should be
investigated in simulated oral conditions.
On the other hand, SMU_1438c showed no activity to dentinal collagen. Although it was identified
as a collagenase with structural similarity to human pro-MMP-1 by bio-informative analysis, the
relatively low molecular mass of SMU_1438c (28 KDa) does not indicate any collagen-binding
domain [22]. In addition, it may need activation if it is in a pro-MMP-like form [21]. Due to the
lack of structural information and limited knowledge of the catalytic domain of these proteases,
the reasons for the difference in degradation efficiency between the three enzymes tested in the
current investigation are presently unknown. Additional investigation will be required to determine
the cleavage site specificity and degradation efficiency of the specific proteases for further
characterization.
4.6 Conclusion
The current study not only confirmed the pathogenic role of S. mutans in dentinal collagen
degradation, that could affect affecting dentin structure and potentially involving in caries process
by verifying its proteolytic activity and identifying specific proteases, but also suggest the

70
contribution of acid as a factor in the degradation process of dentinal collagen. In addition, the
preliminary characterization of SMU_759 and SMU_761 proteases, previously isolated from root
caries lesions provides more information on the current investigations of U32 collagenase family
which is responsible for multiple bacterial infectious disease, including caries.

71
Chapter 5 General Discussion and Summary
Collagen degradation takes place during various physiological and pathological conditions, such
as bone and embryonal development, malignant tumor invasion, wound repair, pathogenic
microorganism invasion and chronic periodontal inflammation [23]. In oral environment, both
endogenous collagenases and bacterial collagenolytic proteases have been linked to destructive
collagen degradation leading to various oral diseases [11, 251, 282]. From over one thousand
bacterial species that colonize and persist in the oral cavity, S. mutans is one of the few species
that has been consistently linked with caries formation [97, 113-115]. Although collagenolytic
activity of S. mutans has been investigated [13, 26], its ability to degrade demineralized dentin has
not been previously demonstrated and the specific proteolytic activity has never been characterized.
The current investigation is the first to report on the ability of S. mutans and its cellular fractions
to degrade demineralized dentin, the identification and initial characterization of specific
proteolytic bacterial enzymes and their activity toward dentinal collagen, potentially contributing
to the cariogenic properties of this bacterium.
5.1 The potential contribution of proteolytic activity of S. mutans to collagen degradation in
caries formation
MMPs are known as zinc- or calcium- depended proteolytic enzymes capable of degrading
collagen fibrils which is the major organic component in tooth [238-240]. Dentin matrix has been
shown to contain endogenous collagenases and gelatinases. [12, 241, 242]. The extrinsic sources
of MMP involved in dentinal degradation include neutrophils and oral bacteria [15, 247]. In the
current investigation fluorometric MMP assay kits were used as a first diagnostic tool to analyze
possible MMP-like activity of S. mutans. Both intact and lysed cells show activity towards all
MMP substrates, suggesting possible collagenolytic and gelatinolytic activity of S. mutans toward
dentinal collagen, similarly to that of an oral pathogenic bacterium, Enterococcus faecalis [247].

72
While both intact and lysed bacterial cells show significant MMP-like activity, suggesting
proteolytic activity for from both intracellular and extracellular origins.
Since synthetic MMPs substrates in this assay, since they lack structural features and integrity of
real native collagen, the specificity and activity/efficiency of bacterial proteases were tested
against type I collagen, which composes of around 90% of the organic matrix in dentin [253]. The
results of the current investigation showed a significant increase of hydroxyproline release from
soluble type I collagen in the presence of both overnight and newly inoculated S. mutans cultures,
and no hydroxyproline release from media alone, suggesting that the bacterium is the source of
this protease activity and is capable of degrading soluble type I collagen.
The higher degradation of type I collagen by O/N S. mutans cultures compared with NEW culture
suggests a growth-phase dependency of degradative capacity of the bacterium which can be can
be explained by the autolysis of S. mutans and bacterial adaptation strategies in its later growth
stage [140, 200, 201, 257-261]. In addition, overnight and fresh-inoculated S. mutans showed their
ability to degrade demineralized human dentin which has more complex structure at different
hierarchical levels [262, 263]. In the current study, there was no hydroxyproline release from
control groups, media only group in which the dentinal MMPs are the only possible source of
proteases. This finding supports previous statement that MMPs have insignificant long-term effect
on dentinal collagen degradation [266] due to the limited amount of MMPs and their inactive form
in dentin [10, 265, 267].
In order to further locate the responsible proteases for dentinal collagen degradation, activity from
discrete bacterial fractions and the media were investigated. Supernatants (cell-free fraction) from
S. mutans O/N cultures showed higher degradative activity than intracellular components. It can
be assumed that the proteases were secreted or released into extracellular environment by S.

73
mutans autolytic activity mentioned above [259, 272].
5.2 The characteristics of collagenolytic/gelatinolytic activity of S. mutans intracellular
enzymes
The dentin demineralization procedures by phosphoric acid and lactic acid were adopted by this
study due to its clinical relevance to dental restorative procedures and biological relevance to
cariogenic bacteria acid production in oral environment, respectively [38, 131, 283]. Previous
studies reported that latent dentinal MMPs can be activated by mild etching acids and then could
initiate dentinal collagen degradation [284, 285]. However, current finding that there is rarely any
hydroxyproline production from acid treated samples when there is no intercellular protein present
in the incubation media, suggesting virtually no degradative impact on dentin by endogenous
MMPs alone. This might be explained by the etching procedure being transient and superficial,
therefore the amount of activated MMPs by etching procedure is limited, and the MMPs were not
activated by bacterial acid as reported before [20, 238, 244, 286, 288-291].
Although most bacterial proteases cannot be considered true collagenases which directly hydrolyze
native collagen molecules with unique specificity [192, 194, 292], it is important to stress that a
large number of bacterial proteases have the capacity to hydrolyze single-stranded and denatured
collagen polypeptides [22]. The results of the current investigation demonstrate that S. mutans
proteases could not degrade heat-treated non-demineralized/non-acid-pretreated dentin slabs,
while both phosphoric acid and lactic acid demineralized dentin released significantly more
hydroxyproline. These findings suggested there are two stages of tissue destruction in caries [293,
294]: acid-initiated demineralization of dentine minerals provides access for bacterial proteases,
followed by collagen breakdown in which acid may also play a critical role to aid bacterial
proteases induced degradation. It has been reported acids can case release non-collagenous
proteins (NCPs) which alters the collagen macrostructural and conformation [44, 276], and change
inter-chain bonds affecting inter- and intramolecular cross-links of collagen [300, 301], thereby

74
increases the susceptibility of the collagen molecules to enzymatic degradation [296-299]. The
current study also showed that the type of acid affected the degradation of dentinal collagen by
bacterial proteases; demineralizing the dentin by lactic acid released significantly more
hydroxyproline compared to that of phosphoric acid. It is assumed that lactic acid, as a by-product
from cariogenic bacteria, has more destructive effects on collagen structures by either releasing
larger amount of NCPs [302] or reducing cross-linking of collagen [300, 301]. The above findings
are supported by a previous study, suggesting that lactic acid is a prerequisite for non-specific
proteases, such as bacterial proteases, to degrade ethylenediaminetetraacetic acid (EDTA)
demineralized dentin samples [276]. However, without definitive structural analysis of dentinal
collagen after demineralization, it is very hard to draw any conclusion.
Bacterial collagenolytic proteases have a broader range of specificity than MMPs, but their
products are hydrolyzed at various specific peptide bonds [25, 303, 304]. In the current study, the
well-studied collagenases from C. histolyticum was used as a positive control to analyze the
degradative effect and pattern of proteases from S. mutans. Comparing to C. histolyticum
collagenase treated dentin collagen, S. mutans intracellular proteins showed different cleavage
sites on collagen. The primary-structural analysis of two resultant collagen fragments derived from
dentinal collagen confirms the degraded peptides are from 1 chain of type I collagen, and, several
preferred amino acids were suggested as cleavage sites, including Lys, Gly, Ser and Arg, which
have been reported also for other bacterial collagenolytic proteases [25, 268, 269]. However, the
proteases’ specificity cannot be identified due to mixed effect of multiple proteases in the
intracellular components. As a result, specific collagenolytic/gelatinolytic proteases from S.
mutans were cloned and expressed for more detailed investigation.

75
5.3 The specific collagenolytic/gelatinolytic enzymes
Three putative genes, SMU_759, SMU_761, and SMU_1438c, were selected to be expressed, since
their coded proteases have been produced by S. mutans UA159. SMU_1438c, was identified as an
interstitial collagenase with structural similarity to human pro-MMP-1 [22, 305]. SMU_759 and
SMU_761, were listed belonging to U32 family of collagenases, which relates to virulence factors
of various human-pathogenic bacteria [306, 307] and were previously isolated from root caries
lesions (31). The most well-studied U32 family member from oral pathogen is PrtC isolated from
Porphyromonas gingivalis (P. gingivalis), which plays a critical role in periodontal tissue
destruction and bacterial invasion [251]. Signal sequence prediction suggested both SMU_759 and
SMU_761 are secreted proteins containing transmembrane domain which was similar to the results
obtained for the prtC from P. gingivalis [147], and, that their predicted structures have the
characteristic compact distorted open barrel made up of -strands and may function in protein
binding [147, 309]. However, the secretion mechanism is not clear and still under investigation
[310, 311].
Both SMU_759 and SMU_761 are capable of degrading demineralized dentinal collagen. The
highest activity was found for SMU_759, which is comparable to the activity of overnight whole
cells. As discussed above, the acid demineralized dentin could have lost its original interstitial
structures, so SMU_659 and SMU_761 may not be considered as true collagenases. This finding
is similar to a previous report that one recombinant U32 peptidase of a non-pathologic bacteria
was only capable of degrading heat-denatured collagen [308]. Although the U32 family has been
recognized as a collagenase group, multiple studies have reported conflicting results regarding
their activity against collagen substrates [308, 312, 313]. In addition, other factors such as ions
also play role in U32 proteases activity, which could also explain the different degradative

76
susceptibility of dentin slabs following lactic acid versus phosphoric acid pretreatments. The
recombinant Filifactor alocis U32 protease (PrtFAC) interacted with and degraded type I collagen
in a Ca2+ dependent manner similar to the P. agarivorans U32 collagenase [313], while Zn2+
showed an inhibitory effect [147]. Considering the complexity of oral conditions, further
characterization of SMU_759 and SMU_761 should be investigated in simulated oral conditions.
On the other hand, SMU_1438c showed no activity to dentinal collagen. Although it has identified
as collagenase with structural similarity to human pro-MMP-1 by bio-informative analysis, the
relatively low molecular mass of SMU_1438c (28 KDa) does not indicate any collagen-binding
domain [22]. In addition, it may need activation if it is in a pro-MMP-like form [21]. Due to the
lack of structural information and limited knowledge of the catalytic domain of these proteases,
the reasons for the difference in degradation efficiency between the three enzymes tested in the
current investigation are presently unknown. Additional investigation will be required to determine
the cleavage site specificity and degradation efficiency of the specific proteases for further
characterization.
Chapter 6 Conclusions and Future Studies
Conclusions

77
• S. mutans UA 159 has proteolytic activity capable of degrading soluble Type I collagen and
dentinal collage, confirming the pathogenic role of S. mutans in dentinal collagen
degradation that may contribute to caries and secondary caries formation.
• The proteolytic activity of S. mutans UA 159 towards collagen is growth-phase dependent,
which may due to the autolysis of S. mutans in its later growth stage to facilitate cell wall
turnover, cell division, assembly of secretion systems, resuscitation of dormant cells and
micro fratricide [257-260] or by the increased selective proteases production in the late
growth stage of S. mutans, which is part of bacterial adaptation strategies, where some oral
pathogenic bacteria could digest host tissue such as collagen to allow the release of amino
acids as their nutrients [140, 200, 201, 257, 261].
• S. mutans collagenolytic/gelatinolytic proteases originate from both intra- and extracellular
origins, which have been reported in other bacterial pathogens [268-271]. The secreted
proteases could explain the histochemical changes of non-demineralized bacteria-free
deep zone of caries lesions in animal model [314], which is considered as a contributing
factor to caries development.
• The demineralization process of tooth structure is a critical step for further bacterial protease
induced dentinal collagen degradation, since non-demineralized dentinal slabs were more
resistant to intracellular proteases induced dentinal collagen degradation. Different acids
have distinct effects on dentinal collagen degradation induced by intracellular proteases
of S. mutans UA 159, which is assumed due to their various effects on collagen structures
by either releasing larger amount of NCPs [302] or reducing cross-linking of collagen [300,
301].

78
• The current investigation is the first to report about the ability of SMU_759 and SMU_761 to
degrade dentinal collagen, supporting previous report about the involvement of these
proteases in [31] caries and secondary caries formation.
Future directions
1. In order to characterize the enzymatic activity and compare to other bacterial collagenase,
additional investigation will be required to determine the cleavage site specificity.
Proposed methods:
• Degraded collagen fragments will be analyzed by comparing SDS-PAGE gel
patterns with well-known bacterial collagenases [315]
• Prime and non-prime cleavage site specificity will be profiled using Proteomic
Identification of protease Cleavage Sites (PICS), a mass spectrometry-based method
utilizing database searchable proteome-derived peptide libraries [316]
2. In order to predict the clinical significance of degradative activity of the specific proteases
to dentinal collagen destruction, additional investigation will be required to determine the
kinetics, stability and inhibition of the specific proteases.
Proposed methods:
• Proteases kinetics will be investigated as described before [317] using soluble type
I collagen under different conditions such as temperature, pH and metal ions
• Protease stability will be investigated by analyzing hydroxyproline production
from type I collagen at different time points

79
• The inhibition of proteases will be further explored by development of specific
antibody inhibitors targeting on SMU_759 or SMU_761 or using generic proteases
inhibitors [318, 319]
3. To further categorize enzymatic protease into collagenases or gelatinases, the collagen
structural integrity after demineralization needs to be determined. As a result, further
structural analysis of demineralized dentin will be required to confirm the effects of various
acid on the collagen structural integrity.
Proposed methods:
• SEM and TEM analysis investigate the macrostructure changes of dentinal
collagen after demineralization [320, 321]
• Fourier transform infrared spectroscopy (FTIR) with attenuated total reflectance
(ATR) and environmental scanning electron microscopy (ESEM) with energy
dispersive X-ray spectrometry (EDX) are useful for analyzing the changes in the
degree of dentine mineralization and the collagen modifications after chemical
treatments [322, 323]
4. Further investigation (gene KO and complementary experiments) will be required to
confirm the role of SMU_759 and SMU_761 in bacterial degradative activity towards
dentinal collagen.
Proposed methods:
• The gene knock-out and complementary strains of S. mutans will be constructed
as described before [128, 324]. Then, the degradation experiments of type I

80
collagen and dentinal collagen will be repeated with the KO and complementary
stains of S. mutans UA 159

81
Chapter 7 Reference
[1] O. Fejerskov, E. Kidd, Dental caries: the disease and its clinical management, John Wiley &
Sons2009.
[2] R.H. Selwitz, A.I. Ismail, N.B. Pitts, Dental caries, The Lancet 369(9555) (2007) 51-59.
[3] R. Frankenberger, F. Garcia-Godoy, P.E. Murray, A.J. Feilzer, N. Krämer, Risk aspects of
dental restoratives: from amalgam to toothcolored materials, World J Stomatol 2(1) (2013) 1-11.
[4] S.E. Kopperud, A.B. Tveit, T. Gaarden, L. Sandvik, I. Espelid, Longevity of posterior dental
restorations and reasons for failure, Eur J Oral Sci 120(6) (2012) 539-48.
[5] F. Brudevold, L.T. Steadman, F.A. Smith, Inorganic and organic components of tooth
structure, Annals of the New York Academy of Sciences 85(1) (1960) 110-132.
[6] R.F. Sognnaes, Introduction to the problem of caries, Annals of the New York Academy of
Sciences 131(2) (1965) 687-689.
[7] W. Armstrong, Further studies on the action of collagenase on sound and carious human
dentin, Journal of dental research 37(6) (1958) 1001-1015.
[8] C. Chaussain, T. Boukpessi, M. Khaddam, L. Tjaderhane, A. George, S. Menashi, Dentin
matrix degradation by host matrix metalloproteinases: inhibition and clinical perspectives toward
regeneration, Frontiers in physiology 4 (2013) 308.
[9] C. Chaussain-Miller, F. Fioretti, M. Goldberg, S. Menashi, The role of matrix
metalloproteinases (MMPs) in human caries, Journal of Dental Research 85(1) (2006) 22-32.
[10] L.N. Niu, L. Zhang, K. Jiao, F. Li, Y.X. Ding, D.Y. Wang, M.Q. Wang, F.R. Tay, J.H.
Chen, Localization of MMP-2, MMP-9, TIMP-1, and TIMP-2 in human coronal dentine, J Dent
39(8) (2011) 536-42.
[11] A. Mazzoni, L. Tjäderhane, V. Checchi, R. Di Lenarda, T. Salo, F.R. Tay, D.H. Pashley, L.
Breschi, Role of Dentin MMPs in Caries Progression and Bond Stability, Journal of Dental
Research 94(2) (2015) 241-251.
[12] M. Sulkala, T. Tervahartiala, T. Sorsa, M. Larmas, T. Salo, L. Tjäderhane, Matrix
metalloproteinase-8 (MMP-8) is the major collagenase in human dentin, Archives of oral biology
52(2) (2007) 121-127.
[13] R.J. Jackson, D.V. Lim, M.L. Dao, Identification and analysis of a collagenolytic activity in
Streptococcus mutans, Curr Microbiol 34(1) (1997) 49-54.
[14] M. Marashdeh, R. Gitalis, C. Lévesque, Y. Finer, Endodontic pathogens possess
collagenolytic properties that degrade human dentine collagen matrix, International endodontic
journal 52(4) (2019) 416-423.
[15] R. Gitalis, L. Zhou, M.Q. Marashdeh, C. Sun, M. Glogauer, Y. Finer, Human neutrophils
degrade methacrylate resin composites and tooth dentin, Acta biomaterialia 88 (2019) 325-331.
[16] A. Hedenbjork-Lager, L. Bjorndal, A. Gustafsson, T. Sorsa, L. Tjaderhane, S. Akerman, D.
Ericson, Caries correlates strongly to salivary levels of MMP-8, Caries Res 49(1) (2015) 1-8.
[17] A. Mazzoni, L. Tjäderhane, V. Checchi, R. Di Lenarda, T. Salo, F. Tay, D. Pashley, L.
Breschi, Role of Dentin MMPs in Caries Progression and Bond Stability, Journal of dental
research 94(2) (2015) 241-251.
[18] A. Van Strijp, D. Jansen, J. DeGroot, J. Ten Cate, V. Everts, Host-derived proteinases and
degradation of dentine collagen in situ, Caries Res 37(1) (2000) 58-65.
[19] R. Osorio, M. Yamauti, E. Osorio, M.E. Ruiz-Requena, D. Pashley, F. Tay, M. Toledano,
Effect of dentin etching and chlorhexidine application on metalloproteinase-mediated collagen
degradation, Eur J Oral Sci 119(1) (2011) 79-85.

82
[20] D.H. Pashley, F. Tay, C. Yiu, M. Hashimoto, L. Breschi, R.M.d. Carvalho, S. Ito, Collagen
degradation by host-derived enzymes during aging, Journal of dental research 83(3) (2004) 216-
221.
[21] T. Sorsa, L. Tjäderhane, T. Salo, Matrix metalloproteinases (MMPs) in oral diseases, Oral
Dis 10(6) (2004) 311-8.
[22] A.S. Duarte, A. Correia, A.C. Esteves, Bacterial collagenases–a review, Critical Reviews in
Microbiology 42(1) (2016) 106-126.
[23] D.J. Harrington, Bacterial collagenases and collagen-degrading enzymes and their potential
role in human disease, Infection and immunity 64(6) (1996) 1885.
[24] E. Yoshida, H. Noda, Isolation and characterization of collagenases I and II from
Clostridium histolyticum, Biochimica et Biophysica Acta (BBA)-Enzymology and Biological
Oxidation 105(3) (1965) 562-574.
[25] M. Okamoto, Y. Yonejima, Y. Tsujimoto, Y. Suzuki, K. Watanabe, A thermostable
collagenolytic protease with a very large molecular mass produced by thermophilic Bacillus sp.
strain MO-1, Applied microbiology and biotechnology 57(1-2) (2001) 103-108.
[26] S.Z. Dung, Effects of mutans streptococci, Actinomyces species and Porphyromonas
gingivalis on collagen degradation, Zhonghua Yi Xue Za Zhi (Taipei) 62(11) (1999) 764-74.
[27] L. Rosengren, B. Winblad, Proteolytic activity of Streptococcus mutans (GS-5), Oral
Surgery, Oral Medicine, Oral Pathology 42(6) (1976) 801-809.
[28] D.J. Harrington, R.R. Russell, Identification and characterisation of two extracellular
proteases of Streptococcus mutans, FEMS Microbiol Lett 121(2) (1994) 237-41.
[29] R.J. Jackson, M.L. Dao, D.V. Lim, Modified FALGPA assay for cell-associated
collagenolytic activity, Journal of microbiological methods 21(2) (1995) 209-215.
[30] S. Despres, H. Métivier, R. Weill, [Degradation of collagen by the cariogenic bacteria,
Streptococcus mutans], C R Seances Acad Sci D 290(1) (1980) 41-4.
[31] N. Damé-Teixeira, C.C.F. Parolo, M. Maltz, A.G. Rup, D.A. Devine, T. Do, Gene
expression of bacterial collagenolytic proteases in root caries, Journal of Oral Microbiology
10(1) (2018) 1424475.
[32] S. Argimón, P.W. Caufield, Distribution of putative virulence genes in Streptococcus
mutans strains does not correlate with caries experience, J Clin Microbiol 49(3) (2011) 984-92.
[33] S. Socransky, A. Haffajee, M. Cugini, C. Smith, R. Kent Jr, Microbial complexes in
subgingival plaque, Journal of clinical periodontology 25(2) (1998) 134-144.
[34] D.J. Bradshaw, P.D. Marsh, G.K. Watson, C. Allison, Role of Fusobacterium nucleatum and
coaggregation in anaerobe survival in planktonic and biofilm oral microbial communities during
aeration, Infection and immunity 66(10) (1998) 4729-4732.
[35] S.S. Socransky, A.D. Haffajee, Dental biofilms: difficult therapeutic targets, Periodontology
2000 28(1) (2002) 12-55.
[36] P.D. Marsh, Dental plaque: biological significance of a biofilm and community life‐style,
Journal of clinical periodontology 32 (2005) 7-15.
[37] K.R. Phipps, V.J. Stevens, Relative contribution of caries and periodontal disease in adult
tooth loss for an HMO dental population, Journal of public health dentistry 55(4) (1995) 250-
252.
[38] S. Hamada, T. Koga, T. Ooshima, Virulence factors of Streptococcus mutans and dental
caries prevention, Journal of dental research 63(3) (1984) 407-411.
[39] C.H. Lai, M. Listgarten, M. Shirakawa, J. Slots, Bacteroides forsythus in adult gingivitis
and periodontitis, Oral microbiology and immunology 2(4) (1987) 152-157.

83
[40] E. Könönen, H.P. Müller, Microbiology of aggressive periodontitis, Periodontology 2000
65(1) (2014) 46-78.
[41] P. Fives‐Taylor, D. Meyer, K. Mintz, Virulence factors of the periodontopathogen
Actinobacillus actinomycetemcomitans, Journal of periodontology 67 (1996) 291-297.
[42] J.C. Fenno, B.C. McBride, Virulence factors of oral treponemes, Anaerobe 4(1) (1998) 1-
17.
[43] H. Maeda, Role of microbial proteases in pathogenesis, Microbiology and immunology
40(10) (1996) 685-699.
[44] S.-Z. Dung, R. Gregory, Y. Li, G. Stookey, Effect of lactic acid and proteolytic enzymes on
the release of organic matrix components from human root dentin, Caries research 29(6) (1995)
483-489.
[45] T.Z. Dung, A.H. Liu, Molecular pathogenesis of root dentin caries, Oral diseases 5(2)
(1999) 92-99.
[46] J. Potempa, J. Travis, Porphyromonas gingivalis proteinases in periodontitis, a review, Acta
Biochim. Pol 43 (1996) 455-465.
[47] D.H. Pashley, Clinical correlations of dentin structure and function, Journal of Prosthetic
Dentistry 66(6) (1991) 777-781.
[48] R.C. Williams, Periodontal disease, New England Journal of Medicine 322(6) (1990) 373-
382.
[49] M.I. CHO, P.R. Garant, Development and general structure of the periodontium,
Periodontology 2000 24(1) (2000) 9-27.
[50] M. Staines, W. Robinson, J. Hood, Spherical indentation of tooth enamel, Journal of
materials science 16(9) (1981) 2551-2556.
[51] B.K. Berkovitz, G.R. Holland, B.J. Moxham, Oral Anatomy, Histology and Embryology E-
Book, Elsevier Health Sciences2017.
[52] J. Hicks, F. Garcia-Godoy, C. Flaitz, Biological factors in dental caries: role of saliva and
dental plaque in the dynamic process of demineralization and remineralization (part 1), Journal
of Clinical Pediatric Dentistry 28(1) (2004) 47-52.
[53] C. Dawes, What is the critical pH and why does a tooth dissolve in acid?, Journal-Canadian
Dental Association 69(11) (2003) 722-725.
[54] P. Hoppenbrouwers, F. Driessens, J. Borggreven, The mineral solubility of human tooth
roots, Archives of Oral Biology 32(5) (1987) 319-322.
[55] E.A.A. Neel, A. Aljabo, A. Strange, S. Ibrahim, M. Coathup, A.M. Young, L. Bozec, V.
Mudera, Demineralization–remineralization dynamics in teeth and bone, International journal of
nanomedicine 11 (2016) 4743.
[56] G. Ramachandran, Stereochemistry of collagen, International journal of peptide and protein
research 31(1) (1988) 1-16.
[57] W. Landis, M. Song, A. Leith, L. McEwen, B. McEwen, Mineral and organic matrix
interaction in normally calcifying tendon visualized in three dimensions by high-voltage electron
microscopic tomography and graphic image reconstruction, Journal of structural biology 110(1)
(1993) 39-54.
[58] R. Maynes, Structure and function of collagen types, Elsevier2012.
[59] S. Ayad, R. Boot-Handford, M. Humphries, K. Kadler, A. Shuttleworth, The extracellular
matrix factsbook, Elsevier1998.
[60] H. Lodish, A. Berk, S.L. Zipursky, P. Matsudaira, D. Baltimore, J. Darnell, Molecular cell
biology 4th edition, National Center for Biotechnology Information, Bookshelf (2000).

84
[61] C. Walters, D.R. Eyre, Collagen crosslinks in human dentin: increasing content of
hydroxypyridinium residues with age, Calcified tissue international 35(1) (1983) 401-405.
[62] M. Macdougall, M. Zeichner-david, H.C. Slavkin, Characterization of extracellular and
nascent dentin phosphoproteins, Connective tissue research 22(1-4) (1989) 697-703.
[63] Y. Takagi, H. Nagai, S. Sasaki, Difference in noncollagenous matrix composition between
crown and root dentin of bovine incisor, Calcified tissue international 42(2) (1988) 97-103.
[64] A. Nanci, Ten Cate's Oral Histology-E-Book: Development, Structure, and Function,
Elsevier Health Sciences2017.
[65] J. Eastoe, Enamel protein chemistry-past, present and future, Journal of dental research
58(2_suppl) (1979) 753-764.
[66] S. Weidmann, D. Eyre, Amino acid composition of enamel protein in the fully developed
human tooth, Caries research 1(4) (1967) 349-355.
[67] P.S. Hart, T.C. Hart, Disorders of human dentin, Cells Tissues Organs 186(1) (2007) 70-77.
[68] G. Orsini, A. Ruggeri Jr, A. Mazzoni, F. Nato, L. Manzoli, A. Putignano, R. Di Lenarda, L.
Tjäderhane, L. Breschi, A review of the nature, role, and function of dentin non‐collagenous
proteins. Part 1: proteoglycans and glycoproteins, Endodontic Topics 21(1) (2009) 1-18.
[69] S. McCurdy, B. Clarkson, R. Speirs, F. Feagin, Phosphoprotein extraction from the
dentine/cementum complex of human tooth roots, Archives of oral biology 35(5) (1990) 347-
357.
[70] M.B. Mathews, The interaction of collagen and acid mucopolysaccarides. A model for
connective tissue, Biochemical Journal 96(3) (1965) 710.
[71] S. Chandrasekhar, H.K. Kleinman, J.R. Hassell, G.R. Martin, J.D. Termine, R.L. Trelstad,
Regulation of type I collagen fibril assembly by link protein and proteoglycans, Collagen and
related research 4(5) (1984) 323-337.
[72] Y. Kuboki, G.L. Mechanic, Comparative molecular distribution of cross-links in bone and
dentin collagen. Structure-function relationships, Calcified tissue international 34(1) (1982) 306-
308.
[73] Y. Kuboki, M.L. Tanzer, G.L. Mechanic, Isolation of polypeptides containing the
intermolecular cross-link δ, δ′-dihydroxylysinonorleucine from dentin collagen, Archives of
biochemistry and biophysics 158(1) (1973) 106-115.
[74] A. Veis, R.J. Schlueter, The macromolecular organization of dentine matrix collagen. I.
Characterization of dentine collagen, Biochemistry 3(11) (1964) 1650-1657.
[75] Y. Kuboki, K. Ohgushi, T. Fusayama, Collagen biochemistry of the two layers of carious
dentin, Journal of dental research 56(10) (1977) 1233-1237.
[76] A. LINDE, Differences between non‐collagenous protein content of rat incisor and
permanent bovine dentin, European Journal of Oral Sciences 96(3) (1988) 188-198.
[77] M.G. Newman, H. Takei, P.R. Klokkevold, F.A. Carranza, Carranza's clinical
periodontology, Elsevier health sciences2011.
[78] D.D. Bosshardt, K.A. Selvig, Dental cementum: the dynamic tissue covering of the root,
Periodontology 2000 13(1) (1997) 41-75.
[79] P. Bartold, Turnover in periodontal connective tissues: dynamic homeostasis of cells,
collagen and ground substances, Oral diseases 1(4) (1995) 238-253.
[80] L. Rao, H. Wang, R. Kalliecharan, J. Heersche, J. Sodek, Specific immunohistochemical
localization of type I collagen in porcine periodontal tissues using the peroxidase-labelled
antibody technique, The Histochemical Journal 11(1) (1979) 73-82.

85
[81] M.J. Glimcher, U.A. Friberg, P.T. Levine, The identification and characterization of a
calcified layer of coronal cementum in erupted bovine teeth, Journal of ultrastructure research
10(1-2) (1964) 76-88.
[82] M.J. Glimcher, The nature of the mineral component of bone and the mechanism of
calcification, Instructional course lectures 36 (1987) 49-69.
[83] B. Gottlieb, Biology of the cementum, The Journal of Periodontology 13(1) (1942) 13-17.
[84] G. Romanos, J. Bernimoulin, Collagen as a basic element of the periodontium:
immunohistochemical aspects in the human and animal. 1. Gingiva and alveolar bone,
Parodontologie (Berlin, Germany) 1(4) (1990) 363-375.
[85] R. Kronfeld, Histopathology of the Teeth: And Their Surrounding Structures, Lea &
Febiger1955.
[86] M. McKee, S. Zalzal, A. Nanci, Extracellular matrix in tooth cementum and mantle dentin:
localization of osteopontin and other noncollagenous proteins, plasma proteins, and
glycoconjugates by electron microscopy, The Anatomical Record: An Official Publication of the
American Association of Anatomists 245(2) (1996) 293-312.
[87] K. Romaniuk, SOME OBSERVATIONS OF FINE STRUCTURE OF HUMAN
CEMENTUM, JOURNAL OF DENTAL RESEARCH, AMER ASSOC DENTAL RESEARCH
1619 DUKE ST, ALEXANDRIA, VA 22314, 1967, pp. 152-&.
[88] W. Moore, L.V. Holdeman, Special problems associated with the isolation and
identification of intestinal bacteria in fecal flora studies, The American journal of clinical
nutrition 27(12) (1974) 1450-1455.
[89] E.A. Grice, J.A. Segre, The human microbiome: our second genome, Annu Rev Genomics
Hum Genet 13 (2012) 151-70.
[90] P. Marsh, Dental plaque as a microbial biofilm, Caries research 38(3) (2004) 204-211.
[91] J.W. Costerton, Z. Lewandowski, D.E. Caldwell, D.R. Korber, H.M. Lappin-Scott,
Microbial biofilms, Annual review of microbiology 49(1) (1995) 711-745.
[92] R.A. Burne, Y.-Y.M. Chen, J.E. Penders, Analysis of gene expression in Streptococcus
mutans in biofilms in vitro, Advances in dental research 11(1) (1997) 100-109.
[93] B.D. Hoyle, J.W. Costerton, Bacterial resistance to antibiotics: the role of biofilms, Progress
in Drug Research/Fortschritte der Arzneimittelforschung/Progrès des recherches
pharmaceutiques, Springer1991, pp. 91-105.
[94] D.G. Cvitkovitch, Quorum sensing and biofilm formation in Streptococcal infections,
Journal of Clinical Investigation 112(11) (2003) 1626-1632.
[95] B.J. Paster, S.K. Boches, J.L. Galvin, R.E. Ericson, C.N. Lau, V.A. Levanos, A.
Sahasrabudhe, F.E. Dewhirst, Bacterial diversity in human subgingival plaque, J Bacteriol
183(12) (2001) 3770-3783.
[96] R. Huang, M. Li, R.L. Gregory, Bacterial interactions in dental biofilm, Virulence 2(5)
(2011) 435-444.
[97] W.J. Loesche, Role of Streptococcus mutans in human dental decay, Microbiology and
molecular biology reviews 50(4) (1986) 353.
[98] J. Slots, M.A. Listgarten, Bacteroides gingivalis, Bacteroides intermedius and
Actinobacillus actinomycetemcomitans in human periodontal diseases, Journal of clinical
periodontology 15(2) (1988) 85-93.
[99] A. Simón-Soro, P. Belda-Ferre, R. Cabrera-Rubio, L. Alcaraz, A. Mira, A tissue-dependent
hypothesis of dental caries, Caries research 47(6) (2013) 591-600.

86
[100] S.H. Kaufmann, M.W. Steward, Topley and Wilson's microbiology and microbial
infections, Arnold2005.
[101] V. Ballal, S. Rao, A. Bagheri, V. Bhat, T. Attin, M. Zehnder, MMP-9 in dentinal fluid
correlates with caries lesion depth, Caries research 51(5) (2017) 460-465.
[102] P.E. Murray, L.J. Windsor, T.W. Smyth, A.A. Hafez, C.F. Cox, Analysis of pulpal
reactions to restorative procedures, materials, pulp capping, and future therapies, Critical
Reviews in Oral Biology & Medicine 13(6) (2002) 509.
[103] T. Beazoglou, S. Eklund, D. Heffley, J. Meiers, L.J. Brown, H. Bailit, Economic impact of
regulating the use of amalgam restorations, Public Health Rep 122(5) (2007) 657-63.
[104] J.W. Simecek, K.E. Diefenderfer, M.E. Cohen, An evaluation of replacement rates for
posterior resin-based composite and amalgam restorations in US Navy and Marine Corps
recruits, The Journal of the American Dental Association 140(2) (2009) 200.
[105] W.J. Loesche, Dental caries: a treatable infection, University of Michigan School of
Dentistry1982.
[106] K. Sunnegårdh-Grönberg, J.W. van Dijken, U. Funegård, A. Lindberg, M. Nilsson,
Selection of dental materials and longevity of replaced restorations in Public Dental Health
clinics in northern Sweden, Journal of dentistry 37(9) (2009) 673-678.
[107] I. Mjör, J. Moorhead, Selection of restorative materials, reasons for replacement, and
longevity of restorations in Florida, The Journal of the American College of Dentists 65(3)
(1998) 27.
[108] C.W. Wakefield, K.R. Kofford, Advances in restorative materials, Dental Clinics of North
America 45(1) (2001) 7.
[109] I.A. MJÖR, Clinical diagnosis of recurrent caries, The Journal of the American Dental
Association 136(10) (2005) 1426-1433.
[110] J.A. Soncini, N.N. Maserejian, F. Trachtenberg, M. Tavares, C. Hayes, The longevity of
amalgam versus compomer/composite restorations in posterior primary and permanent teeth:
findings From the New England Children's Amalgam Trial, J Am Dent Assoc 138(6) (2007)
763-72.
[111] J.-P. Van Nieuwenhuysen, W. D'hoore, J. Carvalho, V. Qvist, Long-term evaluation of
extensive restorations in permanent teeth, Journal of dentistry 31(6) (2003) 395-405.
[112] N.A. Chrysanthakopoulos, Reasons for placement and replacement of resin-based
composite restorations in Greece, Journal of dental research, dental clinics, dental prospects 5(3)
(2011) 87.
[113] R. Seemann, M. Bizhang, I. Klück, J. Loth, J.-F. Roulet, A novel in vitro microbial-based
model for studying caries formation–development and initial testing, Caries research 39(3)
(2005) 185-190.
[114] Y.H. Li, P.C. Lau, J.H. Lee, R.P. Ellen, D.G. Cvitkovitch, Natural genetic transformation
of Streptococcus mutans growing in biofilms, J Bacteriol 183(3) (2001) 897-908.
[115] Y.H. Li, M.N. Hanna, G. Svensater, R.P. Ellen, D.G. Cvitkovitch, Cell density modulates
acid adaptation in Streptococcus mutans: implications for survival in biofilms, J Bacteriol
183(23) (2001) 6875-84.
[116] B. House, J.V. Kus, N. Prayitno, R. Mair, L. Que, F. Chingcuanco, V. Gannon, D.G.
Cvitkovitch, D. Barnett Foster, Acid-stress-induced changes in enterohaemorrhagic Escherichia
coli O157 : H7 virulence, Microbiology 155(Pt 9) (2009) 2907-18.
[117] P. Khalichi, D.G. Cvitkovitch, J.P. Santerre, Effect of composite resin biodegradation
products on oral streptococcal growth, Biomaterials 25(24) (2004) 5467-72.

87
[118] N. Hanada, H.K. Kuramitsu, Isolation and characterization of the Streptococcus mutans
gtfC gene, coding for synthesis of both soluble and insoluble glucans, Infect Immun 56(8) (1988)
1999-2005.
[119] N. Hanada, H.K. Kuramitsu, Isolation and characterization of the Streptococcus mutans
gtfD gene, coding for primer-dependent soluble glucan synthesis, Infection and immunity 57(7)
(1989) 2079-2085.
[120] K. Krastel, D.B. Senadheera, R. Mair, J.S. Downey, S.D. Goodman, D.G. Cvitkovitch,
Characterization of a glutamate transporter operon, glnQHMP, in Streptococcus mutans and its
role in acid tolerance, J Bacteriol 192(4) (2009) 984-93.
[121] J.A. Banas, R. Russell, J. Ferretti, Sequence analysis of the gene for the glucan-binding
protein of Streptococcus mutans Ingbritt, Infection and immunity 58(3) (1990) 667-673.
[122] D. Shah, R. Russell, A novel glucan-binding protein from Streptococcus mutans,
Streptococcal genetics. 6th ed. Asheville, NC: American Society for Microbiology (2002) 75.
[123] S. Colby, R. Russell, Sugar metabolism by mutans streptococci, Journal of Applied
Microbiology 83(S1) (1997).
[124] R. Burne, Y.-Y. Chen, D. Wexler, H. Kuramitsu, W. Bowen, Cariogenicity of
Streptococcus mutans strains with defects in fructan metabolism assessed in a program-fed
specific-pathogen-free rat model, Journal of dental research 75(8) (1996) 1572-1577.
[125] S. Colby, G. Whiting, L. Tao, R. Russell, Insertional inactivation of the Streptococcus
mutans dexA (dextranase) gene results in altered adherence and dextran catabolism,
Microbiology 141(11) (1995) 2929-2936.
[126] J.A. Banas, Virulence properties of Streptococcus mutans, Front Biosci 9(10) (2004) 1267-
77.
[127] A. Yoshida, H.K. Kuramitsu, Multiple Streptococcus mutans genes are involved in biofilm
formation, Applied and environmental microbiology 68(12) (2002) 6283-6291.
[128] Y.-H. Li, N. Tang, M.B. Aspiras, P.C. Lau, J.H. Lee, R.P. Ellen, D.G. Cvitkovitch, A
quorum-sensing signaling system essential for genetic competence in Streptococcus mutans is
involved in biofilm formation, Journal of bacteriology 184(10) (2002) 2699-2708.
[129] Y.-H. Li, P.C. Lau, N. Tang, G. Svensäter, R.P. Ellen, D.G. Cvitkovitch, Novel two-
component regulatory system involved in biofilm formation and acid resistance in Streptococcus
mutans, Journal of bacteriology 184(22) (2002) 6333-6342.
[130] Z.T. Wen, R.A. Burne, Functional Genomics Approach to Identifying Genes Required for
Biofilm Development by Streptococcus mutans, Applied and environmental microbiology 69(1)
(2003) 722.
[131] S.G. Dashper, E.C. Reynolds, Lactic acid excretion by Streptococcus mutans,
Microbiology 142(1) (1996) 33-39.
[132] G. Bender, E. Thibodeau, R. Marquis, Reduction of acidurance of streptococcal growth
and glycolysis by fluoride and gramicidin, Journal of Dental Research 64(2) (1985) 90-95.
[133] A.M. Stock, V.L. Robinson, P.N. Goudreau, Two-component signal transduction, Annual
review of biochemistry 69(1) (2000) 183-215.
[134] E. Reich, K.A. Hiller, Reasons for tooth extraction in the western states of Germany,
Community dentistry and oral epidemiology 21(6) (1993) 379-383.
[135] S. Nuvvula, V.K. Chava, S. Nuvvula, Primary culprit for tooth loss!!, Journal of Indian
Society of Periodontology 20(2) (2016) 222.

88
[136] P.I. Eke, B.A. Dye, L. Wei, G.D. Slade, G.O. Thornton‐Evans, W.S. Borgnakke, G.W.
Taylor, R.C. Page, J.D. Beck, R.J. Genco, Update on prevalence of periodontitis in adults in the
United States: NHANES 2009 to 2012, Journal of periodontology 86(5) (2015) 611-622.
[137] K.S. Kornman, R.C. Page, M.S. Tonetti, The host response to the microbial challenge in
periodontitis: assembling the players, Periodontology 2000 14(1) (1997) 33-53.
[138] C. Tsai, W. McArthur, P. Baehni, B. Hammond, N. Taichman, Extraction and partial
characterization of a leukotoxin from a plaque-derived Gram-negative microorganism, Infection
and Immunity 25(1) (1979) 427-439.
[139] R.E. Singer, B.A. Buckner, Butyrate and propionate: important components of toxic dental
plaque extracts, Infection and Immunity 32(2) (1981) 458-463.
[140] R.J. Gibbons, J.B. MacDonald, Degradation of collagenous substrates by Bacteroides
melaninogenicus, Journal of bacteriology 81(4) (1961) 614.
[141] T. Ogawa, H. Uchida, S. Hamada, Porphyromonas gingivalis fimbriae and their synthetic
peptides induce proinflammatory cytokines in human peripheral blood monocyte cultures, FEMS
microbiology letters 116(2) (1994) 237-242.
[142] E. Gemmell, G. Seymour, Interleukin 1, interleukin 6 and transforming growth factor‐β
production by human gingival mononuclear cells following stimulation with Porphyromonas
gingivalis and Fusobacterium nucleatum, Journal of periodontal research 28(2) (1993) 122-129.
[143] H.A. Schenkein, Host responses in maintaining periodontal health and determining
periodontal disease, Periodontology 2000 40(1) (2006) 77-93.
[144] M. Bickel, The role of interleukin-8 in inflammation and mechanisms of regulation,
Journal of periodontology 64(5 Suppl) (1993) 456-460.
[145] M. Wilson, K. Reddi, B. Henderson, Cytokine‐inducing components of
periodontopathogenic bacteria, Journal of periodontal research 31(6) (1996) 393-407.
[146] P. Kruger, M. Saffarzadeh, A.N. Weber, N. Rieber, M. Radsak, H. von Bernuth, C.
Benarafa, D. Roos, J. Skokowa, D. Hartl, Neutrophils: between host defence, immune
modulation, and tissue injury, PLoS pathogens 11(3) (2015).
[147] T. Kato, N. Takahashi, H.K. Kuramitsu, Sequence analysis and characterization of the
Porphyromonas gingivalis prtC gene, which expresses a novel collagenase activity, Journal of
bacteriology 174(12) (1992) 3889-3895.
[148] W. Moore, L. Holdeman, R. Smibert, D. Hash, J. Burmeister, R. Ranney, Bacteriology of
severe periodontitis in young adult humans, Infection and Immunity 38(3) (1982) 1137-1148.
[149] A.A. Salyers, D.D. Whitt, Bacterial pathogenesis: a molecular approach, ASM press
Washington, DC1994.
[150] J. Slots, Importance of black-pigmented Bacteroides in human periodontal disease,
Hostparasite interactions in periodontal disease (1982) 27-45.
[151] W.J. Loesche, S.A. Syed, B.E. Laughon, J. Stoll, The bacteriology of acute necrotizing
ulcerative gingivitis, Journal of periodontology 53(4) (1982) 223-230.
[152] J. Slots, B.G. Rosling, Suppression of the periodontopathic microflora in localized juvenile
periodontitis by systemic tetracycline, Journal of clinical periodontology 10(5) (1983) 465-486.
[153] J.J. Zambon, Actinobacillus actinomycetemcomitans in human periodontal disease, Journal
of clinical periodontology 12(1) (1985) 1-20.
[154] M. Yoneda, T. Hirofuji, N. Motooka, K. Nozoe, K. Shigenaga, H. Anan, M. Miura, H.
Kabashima, A. Matsumoto, K. Maeda, Humoral immune responses to S-layer-like proteins of
Bacteroides forsythus, Clin. Diagn. Lab. Immunol. 10(3) (2003) 383-387.

89
[155] T. Kurita-Ochiai, K. Fukushima, K. Ochiai, Butyric acid-induced apoptosis of murine
thymocytes, splenic T cells, and human Jurkat T cells, Infection and immunity 65(1) (1997) 35-
41.
[156] G.A. Barkocy-Gallagher, N. Han, J.M. Patti, J. Whitlock, A. Progulske-Fox, M.S. Lantz,
Analysis of the prtP gene encoding porphypain, a cysteine proteinase of Porphyromonas
gingivalis, Journal of Bacteriology 178(10) (1996) 2734-2741.
[157] G.S. Bedi, T. Williams, Purification and characterization of a collagen-degrading protease
from Porphyromonas gingivalis, Journal of Biological Chemistry 269(1) (1994) 599-606.
[158] M.S. Lantz, R.D. Allen, P. Ciborowski, S.C. Holt, Purification and immunolocalization of
a cysteine protease from Porphyromonas gingivalis, Journal of periodontal research 28(7) (1993)
467-469.
[159] R.P. Darveau, A. Tanner, R.C. Page, The microbial challenge in periodontitis,
Periodontology 2000 14(1) (1997) 12-32.
[160] M.B. Rao, A.M. Tanksale, M.S. Ghatge, V.V. Deshpande, Molecular and biotechnological
aspects of microbial proteases, Microbiol. Mol. Biol. Rev. 62(3) (1998) 597-635.
[161] N.J. Veloorvalappil, B.S. Robinson, P. Selvanesan, S. Sasidharan, N.U.
Kizhakkepawothail, S. Sreedharan, P. Prakasan, S.J. Moolakkariyil, B. Sailas, Versatility of
microbial proteases, Advances in enzyme research 2013 (2013).
[162] D. Grenier, D. Mayrand, Selected characteristics of pathogenic and nonpathogenic strains
of Bacteroides gingivalis, Journal of clinical microbiology 25(4) (1987) 738-740.
[163] M.R. Batten, B.W. Senior, M. Kilian, J.M. Woof, Amino acid sequence requirements in
the hinge of human immunoglobulin A1 (IgA1) for cleavage by streptococcal IgA1 proteases,
Infection and immunity 71(3) (2003) 1462-1469.
[164] J. Travis, J. Potempa, Bacterial proteinases as targets for the development of second-
generation antibiotics, Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular
Enzymology 1477(1-2) (2000) 35-50.
[165] R. Grayson, C. Douglas, J. Heath, A. Rawlinson, G. Evans, Activation of human matrix
metalloproteinase 2 by gingival crevicular fluid and Porphyromonas gingivalis, Journal of
clinical periodontology 30(6) (2003) 542-550.
[166] R.P. Allaker, C.I. Douglas, Novel anti-microbial therapies for dental plaque-related
diseases, International journal of antimicrobial agents 33(1) (2009) 8-13.
[167] B. Huang, W. Siqueira, D.G. Cvitkovitch, Y. Finer, Esterase from a Cariogenic Bacterium
Hydrolyzes Dental Resins, Acta biomaterialia (2018).
[168] W.H. Bowen, H. Koo, Biology of Streptococcus mutans-derived glucosyltransferases: role
in extracellular matrix formation of cariogenic biofilms, Caries Res 45(1) (2011) 69-86.
[169] H. Aoki, T. Shiroza, M. Hayakawa, S. Sato, H.K. Kuramitsu, Cloning of a Streptococcus
mutans glucosyltransferase gene coding for insoluble glucan synthesis, Infect Immun 53(3)
(1986) 587-94.
[170] H. Koo, J. Xiao, M. Klein, J. Jeon, Exopolysaccharides produced by Streptococcus mutans
glucosyltransferases modulate the establishment of microcolonies within multispecies biofilms,
Journal of bacteriology 192(12) (2010) 3024-3032.
[171] M.I. Klein, J. Xiao, B. Lu, C.M. Delahunty, J.R. Yates, H. Koo, Streptococcus mutans
protein synthesis during mixed-species biofilm development by high-throughput quantitative
proteomics, PLoS One 7(9) (2012) e45795.

90
[172] J. Xiao, M.I. Klein, M.L. Falsetta, B. Lu, C.M. Delahunty, J.R. Yates, A. Heydorn, H.
Koo, The exopolysaccharide matrix modulates the interaction between 3D architecture and
virulence of a mixed-species oral biofilm, PLoS Pathog 8(4) (2012) e1002623.
[173] K. Fujita, M. Matsumoto‐Nakano, S. Inagaki, T. Ooshima, Biological functions of glucan‐binding protein B of Streptococcus mutans, Oral microbiology and immunology 22(5) (2007)
289-292.
[174] W. Krzyściak, A. Jurczak, D. Kościelniak, B. Bystrowska, A. Skalniak, The virulence of
Streptococcus mutans and the ability to form biofilms, European Journal of Clinical
Microbiology & Infectious Diseases 33(4) (2014) 499-515.
[175] L. Tao, I.C. Sutcliffe, R. Russell, J. Ferretti, Cloning and expression of the multiple sugar
metabolism (msm) operon of Streptococcus mutans in heterologous streptococcal hosts,
Infection and immunity 61(3) (1993) 1121-1125.
[176] R. Fitzgerald, B. Adams, H. Sandham, S. Abhyankar, Cariogenicity of a lactate
dehydrogenase-deficient mutant of Streptococcus mutans serotype c in gnotobiotic rats, Infection
and immunity 57(3) (1989) 823-826.
[177] A.T. Brown, C.L. Wittenberger, Fructose-1, 6-diphosphate-dependent lactate
dehydrogenase from a cariogenic streptococcus: purification and regulatory properties, Journal
of bacteriology 110(2) (1972) 604-615.
[178] J.D. Hillman, Lactate dehydrogenase mutants of Streptococcus mutans: isolation and
preliminary characterization, Infection and immunity 21(1) (1978) 206-212.
[179] G. Svensäter, U.B. Larsson, E. Greif, D. Cvitkovitch, I. Hamilton, Acid tolerance response
and survival by oral bacteria, Molecular Oral Microbiology 12(5) (1997) 266-273.
[180] Y. Ma, T.M. Curran, R.E. Marquis, Rapid procedure for acid adaptation of oral lactic-acid
bacteria and further characterization of the response, Canadian journal of microbiology 43(2)
(1997) 143-148.
[181] I. Hamilton, N. Buckley, Adaptation by Streptococcus mutans to acid tolerance, Molecular
Oral Microbiology 6(2) (1991) 65-71.
[182] S. Dashper, E. Reynolds, pH regulation by Streptococcus mutans, Journal of dental
research 71(5) (1992) 1159-1165.
[183] R.G. Quivey, R.C. Faustoferri, K.A. Clancy, R.E. Marquis, Acid adaptation in
Streptococcus mutans UA159 alleviates sensitization to environmental stress due to RecA
deficiency, FEMS microbiology letters 126(3) (1995) 257-261.
[184] K. McNeill, I. Hamilton, Acid tolerance response of biofilm cells of Streptococcus mutans,
FEMS microbiology letters 221(1) (2003) 25-30.
[185] S. Hata, H. Mayanagi, Acid diffusion through extracellular polysaccharides produced by
various mutants of Streptococcus mutans, Archives of oral biology 48(6) (2003) 431-438.
[186] Y. Yamashita, T. Takehara, H. Kuramitsu, Molecular characterization of a STreptococcus
mutans mutant altered in environmental stress responses, Journal of bacteriology 175(19) (1993)
6220-6228.
[187] J.A. Lemos, R.A. Burne, Regulation and physiological significance of ClpC and ClpP in
Streptococcus mutans, Journal of bacteriology 184(22) (2002) 6357-6366.
[188] G.T. Robertson, W.-L. Ng, J. Foley, R. Gilmour, M.E. Winkler, Global transcriptional
analysis of clpP mutations of type 2 Streptococcus pneumoniae and their effects on physiology
and virulence, Journal of bacteriology 184(13) (2002) 3508-3520.
[189] A.C. Len, D.W. Harty, N.A. Jacques, Stress-responsive proteins are upregulated in
Streptococcus mutans during acid tolerance, Microbiology 150(5) (2004) 1339-1351.

91
[190] M. Sulkala, J. Wahlgren, M. Larmas, T. Sorsa, O. Teronen, T. Salo, L. Tjäderhane, The
effects of MMP inhibitors on human salivary MMP activity and caries progression in rats,
Journal of dental research 80(6) (2001) 1545-1549.
[191] A.R. Hannas, J.C. Pereira, J.M. Granjeiro, L. Tjäderhane, The role of matrix
metalloproteinases in the oral environment, Acta Odontologica 65(1) (2007) 1-13.
[192] H. Birkedal-Hansen, W. Moore, M. Bodden, L. Windsor, B. Birkedal-Hansen, A. DeCarlo,
J. Engler, Matrix metalloproteinases: a review, Critical Reviews in Oral Biology & Medicine
4(2) (1993) 197-250.
[193] H. Nagase, J.F. Woessner, Matrix metalloproteinases, Journal of Biological chemistry
274(31) (1999) 21491-21494.
[194] H. Nagase, Matrix metalloproteinases, Zinc metalloproteases in health and disease, CRC
Press1996, pp. 173-224.
[195] M. Bourbia, D. Ma, D.G. Cvitkovitch, J.P. Santerre, Y. Finer, Cariogenic bacteria degrade
dental resin composites and adhesives, J Dent Res 92(11) (2013) 989-94.
[196] B. Huang, W.L. Siqueira, D.G. Cvitkovitch, Y. Finer, Esterase from a cariogenic
bacterium hydrolyzes dental resins, Acta biomaterialia 71 (2018) 330-338.
[197] J. Potempa, R.N. Pike, Bacterial peptidases, Contributions to microbiology 12 (2005) 132-
180.
[198] G. Dubin, J. Koziel, K. Pyrc, B. Wladyka, J. Potempa, Bacterial proteases in disease–role
in intracellular survival, evasion of coagulation/fibrinolysis innate defenses, toxicoses and viral
infections, Current pharmaceutical design 19(6) (2013) 1090-1113.
[199] R.J. Lamont, H.F. Jenkinson, Life below the gum line: pathogenic mechanisms
ofPorphyromonas gingivalis, Microbiol. Mol. Biol. Rev. 62(4) (1998) 1244-1263.
[200] P. Robertson, M. Lantz, P. Marucha, K. Kornman, C. Trummel, S. Holt, Collagenolytic
activity associated with Bacteroides species and Actinobacillus actinomycetemcomitans, Journal
of periodontal research 17(3) (1982) 275-283.
[201] H. Suido, M. Nakamura, P. Mashimo, J. Zambon, R. Genco, Arylaminopeptidase activities
of oral bacteria, Journal of dental research 65(11) (1986) 1335-1340.
[202] S.C. Holt, T.E. Bramanti, Factors in virulence expression and their role in periodontal
disease pathogenesis, Critical Reviews in Oral Biology & Medicine 2(2) (1991) 177-281.
[203] J. Potempa, N. Pavloff, J. Travis, Porphyromonas gingivalis: a proteinase/gene accounting
audit, Trends in microbiology 3(11) (1995) 430-434.
[204] H. Maeda, T. Yamamoto, Pathogenic mechanisms induced by microbial proteases in
microbial infections, Biological chemistry Hoppe-Seyler 377(4) (1996) 217-226.
[205] Y. Sakata, T. Akaike, M. Suga, S. Ijiri, M. Ando, H. Maeda, Bradykinin generation
triggered by Pseudomonas proteases facilitates invasion of the systemic circulation by
Pseudomonas aeruginosa, Microbiology and immunology 40(6) (1996) 415-423.
[206] K. Maruo, T. Akaike, T. Ono, H. Maeda, Involvement of bradykinin generation in
intravascular dissemination of Vibrio vulnificus and prevention of invasion by a bradykinin
antagonist, Infection and immunity 66(2) (1998) 866-869.
[207] Y. Shi, D.B. Ratnayake, K. Okamoto, N. Abe, K. Yamamoto, K. Nakayama, Genetic
analyses of proteolysis, hemoglobin binding, and hemagglutination of Porphyromonas gingivalis
construction of mutants with a combination of rgpA, rgpB, kgp, and hagA, Journal of Biological
Chemistry 274(25) (1999) 17955-17960.
[208] K. Okamoto, K. Nakayama, T. Kadowaki, N. Abe, D.B. Ratnayake, K. Yamamoto,
Involvement of a lysine-specific cysteine proteinase in hemoglobin adsorption and heme

92
accumulation by Porphyromonas gingivalis, Journal of Biological Chemistry 273(33) (1998)
21225-21231.
[209] C. Genco, W. Simpson, R. Forng, M. Egal, B. Odusanya, Characterization of a Tn4351-
generated hemin uptake mutant of Porphyromonas gingivalis: evidence for the coordinate
regulation of virulence factors by hemin, Infection and immunity 63(7) (1995) 2459-2466.
[210] W. Simpson, C.-Y. Wang, J. Mikolajczyk-Pawlinska, J. Potempa, J. Travis, V.C. Bond,
C.A. Genco, Transposition of the Endogenous Insertion Sequence Element IS1126 Modulates
Gingipain Expression inPorphyromonas gingivalis, Infection and immunity 67(10) (1999) 5012-
5020.
[211] J. Carlsson, J. Höfling, G. Sundqvist, Degradation of albumin, haemopexin, haptoglobin
and transferrin, by black-pigmented Bacteroides species, Journal of medical microbiology 18(1)
(1984) 39-46.
[212] T.E. Bramanti, S.C. Holt, Iron-regulated outer membrane proteins in the periodontopathic
bacterium, Bacteroides gingivalis, Biochemical and biophysical research communications 166(3)
(1990) 1146-1154.
[213] C.S. Fishburn, J.M. Slaney, R. Carman, M. Curtis, Degradation of plasma proteins by the
trypsin‐like enzyme of Porphyromonas gingivalis and inhibition of protease activity by a serine
protease inhibitor of human plasma, Oral microbiology and immunology 6(4) (1991) 209-215.
[214] M. Kilian, J. Mestecky, M. Russell, Defense mechanisms involving Fc-dependent
functions of immunoglobulin A and their subversion by bacterial immunoglobulin A proteases,
Microbiological reviews 52(2) (1988) 296.
[215] A.G. Plaut, The IgA1 proteases of pathogenic bacteria, Annual review of microbiology
37(1) (1983) 603-622.
[216] N.S. Taichman, Potential mechanisms of tissue destruction in periodontal disease, Journal
of dental research 47(6) (1968) 928-928.
[217] J.A. Wingrove, R. DiScipio, Z. Chen, J. Potempa, J. Travis, T. Hugli, Activation of
complement components C3 and C5 by a cysteine proteinase (gingipain-1) from Porphyromonas
(Bacteroides) gingivalis, Journal of Biological Chemistry 267(26) (1992) 18902-18907.
[218] H.A. Schenkein, The effect of periodontol proteolytic Bacteroides species on proteins of
the human complement system, Journal of periodontal research 23(3) (1988) 187-192.
[219] G.K. SUNDQVIST, J. CARLSSON, B.F. HERRMANN, J.F. HÖFLING, A.
VÄÄTÄINEN, Degradation in vivo of the C3 protein of guinea‐pig complement by a pathogenic
strain of Bacteroides gingivalis, European Journal of Oral Sciences 92(1) (1984) 14-24.
[220] M.A. Jagels, J. Travis, J. Potempa, R. Pike, T.E. Hugli, Proteolytic inactivation of the
leukocyte C5a receptor by proteinases derived from Porphyromonas gingivalis, Infection and
immunity 64(6) (1996) 1984-1991.
[221] A. Lala, A. Amano, H.T. Sojar, S.J. Radel, E. Denardin, Porphyromonas gingivalis
trypsin-like protease: a possible natural ligand for the neutrophil formyl peptide receptor,
Biochemical and biophysical research communications 199(3) (1994) 1489-1496.
[222] C.C. Calkins, K. Platt, J. Potempa, J. Travis, Inactivation of tumor necrosis factor-α by
proteinases (gingipains) from the periodontal pathogen, Porphyromonas gingivalis implications
of immune evasion, Journal of biological chemistry 273(12) (1998) 6611-6614.
[223] J. Fletcher, S. Nair, S. Poole, B. Henderson, M. Wilson, Cytokine degradation by biofilms
of Porphyromonas gingivalis, Current microbiology 36(4) (1998) 216-219.

93
[224] J. Fletcher, K. Reddi, S. Poole, S. Nair, B. Henderson, P. Tabona, M. Wilson, Interactions
between periodontopathogenic bacteria and cytokines, Journal of periodontal research 32(1)
(1997) 200-205.
[225] D. Grenier, Degradation of host protease inhibitors and activation of plasminogen by
proteolytic enzymes from Porphyromonas gingivalis and Treponema denticola, Microbiology
142(4) (1996) 955-961.
[226] A.A. DeCarlo, H.E. Grenett, G.J. Harber, L.J. Windsor, M.K. Bodden, B. Birkedal‐Hansen, H. Birkedal‐Hansen, Induction of matrix metalloproteinases and a collagen‐degrading
phenotype in fibroblasts and epithelial cells by secreted Porphyromonas gingivalis proteinase,
Journal of periodontal research 33(7) (1998) 408-420.
[227] D. Mayrand, D. Grenier, Detection of collagenase activity in oral bacteria, Canadian
journal of microbiology 31(2) (1985) 134-138.
[228] B. Pellat, T. Planchenault, C. Pellerin, V. Keil-Dlouha, A comparison of fibronectinolytic
activities from several oral bacteria, Journal de biologie buccale 17(4) (1989) 255-261.
[229] D. Mayrand, S.C. Holt, Biology of asaccharolytic black-pigmented Bacteroides species,
Microbiological reviews 52(1) (1988) 134.
[230] V.-J. Uitto, D. Grenier, E. Chan, B. McBride, Isolation of a chymotrypsinlike enzyme from
Treponema denticola, Infection and immunity 56(10) (1988) 2717-2722.
[231] M. Wikström, G. Dahlén, A. Linde, Fibrinogenolytic and fibrinolytic activity in oral
microorganisms, Journal of clinical microbiology 17(5) (1983) 759-767.
[232] F. Saglie, F. Carranza Jr, M. Newman, L. Cheng, K. Lewin, Identification of tissue‐invading bacteria in human periodontal disease, Journal of periodontal research 17(5) (1982)
452-455.
[233] T. Binder, J. Goodson, S. Socransky, Gingival fluid levels of acid and alkaline
phosphatase, Journal of Periodontal Research 22(1) (1987) 14-19.
[234] R. Frank, J. Voegel, Bacterial bone resorption in advanced cases of human periodontitis,
Journal of periodontal research 13(3) (1978) 251-261.
[235] J. Bulkacz, J.F. Erbland, J. MacGregor, Phospholipase A activity in supernatants from
cultures of Bacteroides melaninogenicus, Biochimica et Biophysica Acta (BBA)-Lipids and
Lipid Metabolism 664(1) (1981) 148-155.
[236] J. Bulkacz, N. MG, S. SS, Phospholipase A activity of micro-organisms from dental
plaque, (1979).
[237] B.A. Bidlingmeyer, S.A. Cohen, T.L. Tarvin, Rapid analysis of amino acids using pre-
column derivatization, J Chromatogr 336(1) (1984) 93-104.
[238] L. Tjaderhane, H. Larjava, T. Sorsa, V.J. Uitto, M. Larmas, T. Salo, The Activation and
Function of Host Matrix Metalloproteinases in Dentin Matrix Breakdown in Caries Lesions,
Journal of Dental Research 77(8) (1998) 1622-1629.
[239] D. Pashley, F. Tay, C. Yiu, M. Hashimoto, L. Breschi, R. Carvalho, S. Ito, Collagen
degradation by host-derived enzymes during aging, Journal of Dental Research 83(3) (2004)
216-221.
[240] T. Ingman, T. Sorsa, O. Lindy, H. Koski, Y.T. Konttinen, Multiple forms of
gelatinases/type IV collagenases in saliva and gingival crevicular fluid of periodontitis patients,
Journal of Clinical Periodontology 21(1) (1994) 26-31.
[241] T. Boukpessi, S. Menashi, L. Camoin, J. Tencate, M. Goldberg, C. Chaussain-Miller, The
effect of stromelysin-1 (MMP-3) on non-collagenous extracellular matrix proteins of

94
demineralized dentin and the adhesive properties of restorative resins, Biomaterials 29(33)
(2008) 4367-4373.
[242] L. Randall, R. Hall, Temperospatial expression of matrix metalloproteinases 1, 2, 3, and 9
during early tooth development, Connective tissue research 43(2-3) (2002) 205-211.
[243] D. Arola, R. Reprogel, Effects of aging on the mechanical behavior of human dentin,
Biomaterials 26(18) (2005) 4051-4061.
[244] Y. Nishitani, M. Yoshiyama, B. Wadgaonkar, L. Breschi, F. Mannello, A. Mazzoni, R.M.
Carvalho, L. Tjäderhane, F.R. Tay, D.H. Pashley, Activation of gelatinolytic/collagenolytic
activity in dentin by self etching adhesives, European journal of oral sciences 114(2) (2006) 160-
166.
[245] L.W. Boushell, M. Kaku, Y. Mochida, R. Bagnell, M. Yamauchi, Immunohistochemical
localization of matrixmetalloproteinase-2 in human coronal dentin, Archives of oral biology
53(2) (2008) 109-116.
[246] A. Mazzoni, D.H. Pashley, F.R. Tay, P. Gobbi, G. Orsini, A. Ruggeri Jr, M. Carrilho, L.
Tjäderhane, R. Di Lenarda, L. Breschi, Immunohistochemical identification of MMP 2 and
MMP 9 in human dentin: Correlative FEI SEM/TEM analysis, Journal of Biomedical Materials
Research Part A 88(3) (2009) 697-703.
[247] Marashdeh, R. Gitalis, C. Lévesque, Y. Finer, 1, Enterococcus faecalis possesses
Collagenolytic Activity that Degrades Human Dentin Collagen Matrix, 2018.
[248] D. Wang, L. Zhang, F. Li, Y. Chen, J. Chen, [Quantitative detection of matrix
metalloproteinase-2 in normal coronal dentine of young people], Zhonghua kou qiang yi xue za
zhi = Zhonghua kouqiang yixue zazhi = Chinese journal of stomatology 49(4) (2014) 244-6.
[249] L. Tjäderhane, F.D. Nascimento, L. Breschi, A. Mazzoni, I.L. Tersariol, S. Geraldeli, A.
Tezvergil-Mutluay, M.R. Carrilho, R.M. Carvalho, F.R. Tay, Optimizing dentin bond durability:
control of collagen degradation by matrix metalloproteinases and cysteine cathepsins, Dental
Materials 29(1) (2013) 116-135.
[250] S. Perumal, O. Antipova, J.P. Orgel, Collagen fibril architecture, domain organization, and
triple-helical conformation govern its proteolysis, Proceedings of the National Academy of
Sciences 105(8) (2008) 2824-2829.
[251] N. Takahashi, T. Kato, H.K. Kuramitsu, Isolation and preliminary characterization of the
Porphyromonas gingivalis prtC gene expressing collagenase activity, FEMS microbiology letters
84(2) (1991) 135-138.
[252] P. Mäkinen, D. Clewell, F. An, K. Mäkinen, Purification and substrate specificity of a
strongly hydrophobic extracellular metalloendopeptidase (" gelatinase") from Streptococcus
faecalis (strain 0G1-10), Journal of Biological Chemistry 264(6) (1989) 3325-3334.
[253] L. Tjäderhane, M.A.R. Buzalaf, M. Carrilho, C. Chaussain, Matrix metalloproteinases and
other matrix proteinases in relation to cariology: the era of ‘dentin degradomics', Caries research
49(3) (2015) 193-208.
[254] A. Fawzy, L. Nitisusanta, K. Iqbal, U. Daood, L. Beng, J. Neo, Characterization of
riboflavin-modified dentin collagen matrix, Journal of dental research 91(11) (2012) 1049-1054.
[255] M.Q. Marashdeh, R. Gitalis, C. Levesque, Y. Finer, Enterococcus faecalis Hydrolyzes
Dental Resin Composites and Adhesives, Journal of endodontics (2018).
[256] D.J. Epasinghe, C.K.Y. Yiu, M.F. Burrow, N. Hiraishi, F.R. Tay, The inhibitory effect of
proanthocyanidin on soluble and collagen-bound proteases, Journal of dentistry 41(9) (2013)
832-839.

95
[257] E.G. Smith, G.A. Spatafora, Gene Regulation in S. mutans: Complex Control in a
Complex Environment, Journal of Dental Research 91(2) (2012) 133-141.
[258] J.A. Lemos, R.A. Burne, A model of efficiency: stress tolerance by Streptococcus mutans,
Microbiology 154(Pt 11) (2008) 3247-55.
[259] D. Dufour, C.M. Lévesque, Cell death of Streptococcus mutans induced by a quorum-
sensing peptide occurs via a conserved streptococcal autolysin, Journal of bacteriology 195(1)
(2013) 105-114.
[260] W. Vollmer, B. Joris, P. Charlier, S. Foster, Bacterial peptidoglycan (murein) hydrolases,
FEMS microbiology reviews 32(2) (2008) 259-286.
[261] G.H. Bowden, Y.H. Li, Nutritional influences on biofilm development, Adv Dent Res
11(1) (1997) 81-99.
[262] I. Kramer, The distribution of collagen fibrils in the dentine matrix, British dental journal
91(1) (1951) 1.
[263] L.E. Bertassoni, Dentin on the nanoscale: Hierarchical organization, mechanical behavior
and bioinspired engineering, Dental Materials 33(6) (2017) 637-649.
[264] A. Almahdy, G. Koller, S. Sauro, J. Bartsch, M. Sherriff, T. Watson, A. Banerjee, Effects
of MMP inhibitors incorporated within dental adhesives, Journal of dental research 91(6) (2012)
605-611.
[265] Y. Shimada, S. Ichinose, A. Sadr, M. Burrow, J. Tagami, Localization of matrix
metalloproteinases (MMPs‐2, 8, 9 and 20) in normal and carious dentine, Australian dental
journal 54(4) (2009) 347-354.
[266] A. Almahdy, G. Koller, S. Sauro, J.W. Bartsch, M. Sherriff, T.F. Watson, A. Banerjee,
Effects of MMP inhibitors incorporated within dental adhesives, J Dent Res 91(6) (2012) 605-
11.
[267] H. Birkedal-Hansen, Proteolytic remodeling of extracellular matrix, Current opinion in cell
biology 7(5) (1995) 728-735.
[268] K. Watanabe, Collagenolytic proteases from bacteria, Applied microbiology and
biotechnology 63(5) (2004) 520-526.
[269] N. Tsuruoka, T. Nakayama, M. Ashida, H. Hemmi, M. Nakao, H. Minakata, H. Oyama, K.
Oda, T. Nishino, Collagenolytic serine-carboxyl proteinase from Alicyclobacillus sendaiensis
strain NTAP-1: purification, characterization, gene cloning, and heterologous expression, Appl.
Environ. Microbiol. 69(1) (2003) 162-169.
[270] K. Yoshihara, O. Matsushita, J. Minami, A. Okabe, Cloning and nucleotide sequence
analysis of the colH gene from Clostridium histolyticum encoding a collagenase and a gelatinase,
Journal of bacteriology 176(21) (1994) 6489-6496.
[271] Z.E. Juarez, M.W. Stinson, An extracellular protease of Streptococcus gordonii hydrolyzes
type IV collagen and collagen analogues, Infection and immunity 67(1) (1999) 271-278.
[272] Y. Liu, R.A. Burne, The major autolysin of Streptococcus gordonii is subject to complex
regulation and modulates stress tolerance, biofilm formation, and extracellular-DNA release, J
Bacteriol 193(11) (2011) 2826-37.
[273] J.A. Perry, D.G. Cvitkovitch, C.M. Lévesque, Cell death in Streptococcus mutans biofilms:
a link between CSP and extracellular DNA, FEMS Microbiol Lett 299(2) (2009) 261-6.
[274] W.L. Siqueira, M. Bakkal, Y. Xiao, J.N. Sutton, F.M. Mendes, Quantitative proteomic
analysis of the effect of fluoride on the acquired enamel pellicle, PLoS One 7(8) (2012) e42204.

96
[275] K.A. Agee, A. Prakki, T. Abu-Haimed, G.H. Naguib, M.A. Nawareg, A. Tezvergil-
Mutluay, D.L. Scheffel, C. Chen, S.S. Jang, H. Hwang, Water distribution in dentin matrices:
bound vs. unbound water, Dental Materials 31(3) (2015) 205-216.
[276] S.Z. Dung, Y. Li, A.J. Dunipace, G.K. Stookey, Degradation of insoluble bovine collagen
and human dentine collagen pretreated in vitro with lactic acid, pH 4.0 and 5.5, Arch Oral Biol
39(10) (1994) 901-5.
[277] J.R. Krieger, P. Taylor, M.F. Moran, C.J. McGlade, Comprehensive identification of
phosphorylation sites on the Numb endocytic adaptor protein, Proteomics 15(2-3) (2015) 434-46.
[278] T.N. Petersen, S. Brunak, G. Von Heijne, H. Nielsen, SignalP 4.0: discriminating signal
peptides from transmembrane regions, Nature methods 8(10) (2011) 785-786.
[279] L.A. Kelley, S. Mezulis, C.M. Yates, M.N. Wass, M.J. Sternberg, The Phyre2 web portal
for protein modeling, prediction and analysis, Nat Protoc 10(6) (2015) 845-58.
[280] L. Sadeghinejad, D.G. Cvitkovitch, W.L. Siqueira, J. Merritt, J.P. Santerre, Y. Finer,
Mechanistic, genomic and proteomic study on the effects of BisGMA-derived biodegradation
product on cariogenic bacteria, Dent Mater 33(2) (2017) 175-190.
[281] W.H. Eschenfeldt, S. Lucy, C.S. Millard, A. Joachimiak, I.D. Mark, A family of LIC
vectors for high-throughput cloning and purification of proteins, Methods Mol Biol 498 (2009)
105-15.
[282] R. Häyrinen-Immonen, T. Sorsa, D. Nordström, M. Malmström, Y. Konttinen,
Collagenase and stromelysin in recurrent aphthous ulcers (RAU), International journal of oral
and maxillofacial surgery 22(1) (1993) 46-49.
[283] N. Nakabayashi, Hybridization of dental hard tissues, The quality of hybridized dentin
(1998).
[284] A. Mazzoni, P. Scaffa, M. Carrilho, L. Tjäderhane, R. Di Lenarda, A. Polimeni, A.
Tezvergil-Mutluay, F. Tay, D.H. Pashley, L. Breschi, Effects of etch-and-rinse and self-etch
adhesives on dentin MMP-2 and MMP-9, Journal of dental research 92(1) (2013) 82-86.
[285] F.R. Tay, D.H. Pashley, R.J. Loushine, R.N. Weller, F. Monticelli, R. Osorio, Self-etching
adhesives increase collagenolytic activity in radicular dentin, Journal of Endodontics 32(9)
(2006) 862-868.
[286] A. Mazzoni, D.H. Pashley, Y. Nishitani, L. Breschi, F. Mannello, L. Tjäderhane, M.
Toledano, E.L. Pashley, F.R. Tay, Reactivation of inactivated endogenous proteolytic activities
in phosphoric acid-etched dentine by etch-and-rinse adhesives, Biomaterials 27(25) (2006) 4470-
4476.
[287] A. Hedenbjörk-Lager, K. Hamberg, V. Pääkkönen, L. Tjäderhane, D. Ericson, Collagen
degradation and preservation of MMP-8 activity in human dentine matrix after demineralization,
Archives of oral biology 68 (2016) 66-72.
[288] J. Featherstone, J. Duncan, T. Cutress, A mechanism for dental caries based on chemical
processes and diffusion phenomena during in-vitro caries simulation on human tooth enamel,
Archives of oral biology 24(2) (1979) 101-112.
[289] J. De Soet, F. Toors, J. De Graaff, Acidogenesis by oral streptococci at different pH
values, Caries research 23(1) (1989) 14-17.
[290] C. Sansone, J. Van Houte, K. Joshipura, R. Kent, H. Margolis, The association of mutans
streptococci and non-mutans streptococci capable of acidogenesis at a low pH with dental caries
on enamel and root surfaces, Journal of dental research 72(2) (1993) 508-516.
[291] M.L. Snyder, J.J. Teachout, Acid production of oral bacteria associated with dental caries,
Journal of Dental Research 21(5) (1942) 461-466.

97
[292] L. Ravanti, V.-M. Kähäri, Matrix metalloproteinases in wound repair, International journal
of molecular medicine 6(4) (2000) 391-798.
[293] E. Johansen, H.F. Parks, Electron-microscopic observations on soft carious human dentin,
Journal of Dental Research 40(2) (1961) 235-248.
[294] B. Nyvad, O. Fejerskov, An ultrastructural study of bacterial invasion and tissue
breakdown in human experimental root-surface caries, Journal of dental research 69(5) (1990)
1118-1125.
[295] B. Klont, J. Ten Cate, Release of organic matrix components from bovine incisor roots
during in vitro lesion formation, Journal of dental research 69(3) (1990) 896-900.
[296] R. Frank, Structural events in the caries process in enamel, cementum, and dentin, Journal
of dental research 69(2_suppl) (1990) 559-566.
[297] R. Frank, P. Steuer, J. Hemmerle, Ultrastructural study on human root caries, Caries
research 23(4) (1989) 209-217.
[298] K. Selvig, Ultrastructural changes in human dentine exposed to a weak acid, Archives of
oral biology 13(7) (1968) 719-IN9.
[299] B. Klont, J. Damen, J. Ten Cate, Degradation of bovine incisor root collagen in an in vitro
caries model, Archives of oral biology 36(4) (1991) 299-304.
[300] P.F. Davison, D.J. Cannon, L.P. Andersson, The effects of acetic acid on collagen cross-
links, Connective Tissue Research 1(3) (1972) 205-216.
[301] J. Rosenblatt, B. Devereux, D.G. Wallace, Injectable collagen as a pH-sensitive hydrogel,
Biomaterials 15(12) (1994) 985-95.
[302] A.H. van der Linden, M. Booij, J.J. ten Bosch, J. Arends, Protein and mineral changes in
bovine enamel during in-vitro demineralization, Arch Oral Biol 32(2) (1987) 75-80.
[303] U. Eckhard, E. Schönauer, P. Ducka, P. Briza, D. Nüss, H. Brandstetter, Biochemical
characterization of the catalytic domains of three different Clostridial collagenases, Biological
chemistry 390(1) (2009) 11-18.
[304] H.E. Van Wart, Clostridium collagenases, Handbook of proteolytic enzymes,
Elsevier2004, pp. 416-419.
[305] G.I. Goldberg, S.M. Wilhelm, A. Kronberger, E.A. Bauer, G.A. Grant, A.Z. Eisen, Human
fibroblast collagenase. Complete primary structure and homology to an oncogene
transformation-induced rat protein, Journal of Biological Chemistry 261(14) (1986) 6600-6605.
[306] S.A. Carlson, Z.P. McCuddin, M.T. Wu, SlyA regulates the collagenase-mediated
cytopathic phenotype in multiresistant Salmonella, Microbial pathogenesis 38(4) (2005) 181-
187.
[307] Q. Wu, C. Li, C. Li, H. Chen, L. Shuliang, Purification and characterization of a novel
collagenase from Bacillus pumilus Col-J, Applied biochemistry and biotechnology 160(1) (2010)
129.
[308] A. Jasilionis, A. Kaupinis, M. Ger, M. Valius, D. Chitavichius, N. Kuisiene, Gene
expression and activity analysis of the first thermophilic U32 peptidase, Central European
Journal of Biology 7(4) (2012) 587-595.
[309] S. Trillo-Muyo, A. Jasilionis, M.J. Domagalski, M. Chruszcz, W. Minor, N. Kuisiene, J.L.
Arolas, M. Solà, F.X. Gomis-Rüth, Ultratight crystal packing of a 10 kDa protein, Acta
Crystallogr D Biol Crystallogr 69(Pt 3) (2013) 464-70.
[310] H. Zhao, X. Li, D.E. Johnson, H.L. Mobley, Identification of protease and rpoN-associated
genes of uropathogenic Proteus mirabilis by negative selection in a mouse model of ascending
urinary tract infection, Microbiology 145(1) (1999) 185-195.

98
[311] C.-K. Yang, H.E. Ewis, X. Zhang, C.-D. Lu, H.-J. Hu, Y. Pan, A.T. Abdelal, P.C. Tai,
Nonclassical protein secretion by Bacillus subtilis in the stationary phase is not due to cell lysis,
Journal of bacteriology 193(20) (2011) 5607-5615.
[312] S. Bhattacharya, S. Bhattacharya, R. Gachhui, S. Hazra, J. Mukherjee, U32 collagenase
from Pseudoalteromonas agarivorans NW4327: Activity, structure, substrate interactions and
molecular dynamics simulations, International journal of biological macromolecules 124 (2019)
635-650.
[313] O. Chioma, A.W. Aruni, T.A. Milford, H.M. Fletcher, Filifactor alocis collagenase can
modulate apoptosis of normal oral keratinocytes, Molecular oral microbiology 32(2) (2017) 166-
177.
[314] Y. Kobayashi, M. Ozeki, A. Ogawa, S. Matsumoto, M. Sanjo, T. Moriyama, Invasion of
Streptococcus mutans, Streptococcus intermedius and Propionibacterium acnes into the teeth of
gnotobiotic rats, Caries Research 26(2) (1992) 132-138.
[315] E.N. Baramova, J.D. Shannon, J.B. Bjarnason, J.W. Fox, Identification of the cleavage
sites by a hemorrhagic metalloproteinase in type IV collagen, Matrix 10(2) (1990) 91-97.
[316] U. Eckhard, P.F. Huesgen, H. Brandstetter, C.M. Overall, Proteomic protease specificity
profiling of clostridial collagenases reveals their intrinsic nature as dedicated degraders of
collagen, Journal of proteomics 100 (2014) 102-114.
[317] S.K. Mallya, K.A. Mookhtiar, H.E. Van Wart, Kinetics of hydrolysis of type I, II, and III
collagens by the class I and IIClostridium histolyticum collagenases, Journal of protein
chemistry 11(1) (1992) 99-107.
[318] K. Matsushita, T. Imamura, S. Tancharoen, S. Tatsuyama, M. Tomikawa, J. Travis, J.
Potempa, M. Torii, I. Maruyama, Selective inhibition of Porphyromonas gingivalis growth by a
factor Xa inhibitor, DX‐9065a, Journal of periodontal research 41(3) (2006) 171-176.
[319] T. Imamura, K. Matsushita, J. Travis, J. Potempa, Inhibition of Trypsin-Like Cysteine
Proteinases (Gingipains) fromPorphyromonas gingivalis by Tetracycline and Its Analogues,
Antimicrobial agents and chemotherapy 45(10) (2001) 2871-2876.
[320] T. Starborg, N.S. Kalson, Y. Lu, A. Mironov, T.F. Cootes, D.F. Holmes, K.E. Kadler,
Using transmission electron microscopy and 3View to determine collagen fibril size and three-
dimensional organization, Nature protocols 8(7) (2013) 1433.
[321] M. Hashimoto, F.R. Tay, H. Ohno, H. Sano, M. Kaga, C. Yiu, H. Kumagai, Y. Kudou, M.
Kubota, H. Oguchi, SEM and TEM analysis of water degradation of human dentinal collagen,
Journal of Biomedical Materials Research Part B: Applied Biomaterials: An Official Journal of
The Society for Biomaterials, The Japanese Society for Biomaterials, and The Australian Society
for Biomaterials and the Korean Society for Biomaterials 66(1) (2003) 287-298.
[322] A. Terzi, E. Storelli, S. Bettini, T. Sibillano, D. Altamura, L. Salvatore, M. Madaghiele, A.
Romano, D. Siliqi, M. Ladisa, Effects of processing on structural, mechanical and biological
properties of collagen-based substrates for regenerative medicine, Scientific reports 8(1) (2018)
1-13.
[323] B. de Campos Vidal, M.L.S. Mello, Collagen type I amide I band infrared spectroscopy,
Micron 42(3) (2011) 283-289.
[324] P.C. Lau, C.K. Sung, J.H. Lee, D.A. Morrison, D.G. Cvitkovitch, PCR ligation
mutagenesis in transformable streptococci: application and efficiency, J Microbiol Methods
49(2) (2002) 193-205.

99
[325] G. Svensater, J. Welin, J.C. Wilkins, D. Beighton, I.R. Hamilton, Protein expression by
planktonic and biofilm cells of Streptococcus mutans, FEMS Microbiol Lett 205(1) (2001) 139-
46.
[326] M.J. Hughes, J.C. Moore, J.D. Lane, R. Wilson, P.K. Pribul, Z.N. Younes, R.J. Dobson, P.
Everest, A.J. Reason, J.M. Redfern, F.M. Greer, T. Paxton, M. Panico, H.R. Morris, R.G.
Feldman, J.D. Santangelo, Identification of major outer surface proteins of Streptococcus
agalactiae, Infect Immun 70(3) (2002) 1254-9.
[327] D.E. Kling, L.C. Madoff, J.L. Michel, Subcellular fractionation of group B Streptococcus,
Biotechniques 27(1) (1999) 24-6, 28.
[328] L.A. Kelley, M.J. Sternberg, Protein structure prediction on the Web: a case study using
the Phyre server, Nature protocols 4(3) (2009) 363.

100
8. Supplemental information
8.1 Preparation of discrete fractions of S. mutans
8.1.1 Intracellular components
Intracellular component fractions were prepared as described previously [259, 325] with
modifications. S. mutans UA 159 were grown in THYE (pH 7.0) at 37°C (OD600 = 0.8). The cells
were harvested by centrifugation at 5,000 × g for 10 min and washed with PBS. Then, the bacterial
cells were re-suspended in PBS and subjected to ultrasonic homogenizer (20kHz, Branson
Ultrasonics™ Sonifier™ SFX150 Cell Disruptor, Fisher Scientific) for 3 min with cooling in
between. Intact cells were sedimented by centrifugation at 16000 x g for 5 min at 4°C. The resultant
intracellular components were released into the supernatant which was separated from the
sedimented fraction, collected and stored at -80°C until required.
8.1.2 Membrane pellets
For membrane preparation, S. mutans UA 159 were grown in THYE (pH 7.0) at 37°C (OD600 =
0.8). Membrane-enriched protein fractions were prepared as described previously [326, 327], with
some modifications. The bacterial cells were harvested by centrifugation at 5,000 × g for 10 min
and washed with PBS. The cells were re-suspended in buffer A (20% sucrose, 20 mM Tris [pH
7.0], 10 mm MgCl2), and then in order to reduce cell wall protein contamination, mutanolysin (1
mg; Sigma, St. Louis, MO) and lysozyme (18 mg; Sigma, St. Louis, MO) were added and the
mixture was incubated at 37°C with gentle agitation. Protoplast formation was monitored using
Gram staining. Once cell wall digestion was complete, mixtures were centrifuged as described
above. The protoplasts were washed once with buffer A, and each cell pellet was suspended in
buffer B (10 mM Tris [pH 8.1], 50 mM MgCl2, 10 mM glucose). The protoplasts were lysed.
Debris and unbroken protoplasts were removed by centrifugation at 6,000 × g for 10 min at 4°C,
and the supernatant was ultracentrifuged at 41,000 × g for 45 min at 4°C. The pellet was suspended

101
in buffer C (10 mM Tris [pH 8.1], 50 mM NaCl, 20 mM MgCl2) at 4°C and ultracentrifuged at
105,000 × g for 45 min at 4°C. The supernatant was decanted, and the membrane-enriched pellet
was washed twice with buffer D (20 mM Tris [pH 7.2], 10 mM MgCl2) and stored at - 80°C until
it was used.
8.2 Homology detection and 3D model structural analysis of SMU_759, SMU_761 and
SMU_1438c
Based on previous proteomic analysis (data not published), three proteases coded by SMU_759,
SMU_761 and SMU_1438c were expressed in S. mutans UA159.
Based on homology detection and 3D model structural analysis by Phyre2
(http://www.sbg.bio.ic.ac.uk/~phyre2/html/page.cgi?id =index) [328], SMU_759 and SMU_761
belong to bacterial collagenase U32 family and there are no signal peptide (Fig. 8.1). The predicted
molecular masses for SMU_759 and SMU_761 are 35 KDa and 48 KDa respectively. The
SMU_1438c is predicted as an interstitial collagenase with trans-membrane helices (Fig. 8.1). The
estimated molecular mass is 28 KDa.
SMU_759
SMU_761
SMU_1438c
Fig. 8.1: The identification and analysis of proteinases from S. mutans UA159. SMU_759 was
identified as 35KDa peptide from intracellular component of S. mutans UA159; SMU_761 was

102
identified as 58KDa peptide from intracellular component of S. mutans UA159, ans SMU_1438c
was identified as 28KDa collagenase with transmembrane domain of S. mutans UA159.
8.3 Putative Collagenase Gene sequences
SMU_1438c Zn-dependent protease [ Streptococcus mutans UA159 ]
Sequence: NC_004350.2 (1370234.1370932, complement)
NCBI Reference Sequence: NC_004350.2
GenBank Graphics
>gi|347750429:c1370932-1370234 Streptococcus mutans UA159 chromosome, complete
genome
ATGAAGACTTTCATTAAAATTTTGTTTTTTATTCCTAGACTAATATGGAATATTATTT
GGAGCATCATTAAAACTCTTATTATTCTTGCTGCTATTATTTTTGCTTTTCTTTATTTT
ACAAATAATCATAAAACAGACTTGGAATCAACGATCTCCGAACAATGGAATAAGAT
AACGACTTTTTTTAGTAATGACTTTAGTTTGCCGGATACGATGTCTAAATTATCAACA
GATAATTATAAACATGAAGCAGGGAGTCGTTGGAGTCAAAATAGTGCTAGTGTTTAT
ATAGCTTCTACTGATAAAACTATTGTTAAGGCTTACCAAACTGCTCTGGCTAATTGG
AATGCAACAGGAAGTTTCACTTTTAATATCATTTCGGATAAAGCATCAGCAGATATT
ACAGCAAAGGATTATTCTGATGTAAATTCGCAAGCCGCTGGTTTAGCTGAAACAGA
AACGAATGCAGTGACCAATCGCATGAGTCATGTTGATGTTAAGCTTAATCGCTACTA
TCTTTTAGATGCAAGTTACGGTTATAGTTTTGATAGGATAGTCCACACAGCAGAGCA
TGAGTTAGGGCATGCTATAGGACTTGATCATGATGATAAAGAAACTTCGGTAATGGC
CTCATCAGGTTCCTACAATGGTATTCAAACAGTTGATATAACTGCTGTTAAGAAGCT
TTACGCTAATTAA
SMU_1784c possible membrane-associated Zn-dependent proteases [ Streptococcus mutans
UA159 ]
Gene ID: 1028984, updated on 26-Jun-2015
NCBI Reference Sequence: NC_004350.2
GenBank Graphics
>gi|347750429:c1692541-1691282 Streptococcus mutans UA159 chromosome, complete
genome
ATGTCAGGACTAATAGCTTTTATTATTATCTTTGGAATTATAGTTCTTGTCCATGAAT
TTGGTCATTTCTATTTTGCTAGAAAATCAGGAATTTTGGTTCGGGAATTTGCCATTGG
TATGGGACCGAAAATTTTTGCACATCAAGGAAAAGACGGTACAGCTTATACCATTCG
AATTTTGCCTTTAGGTGGCTATGTCCGTATGGCTGGCTGGGGAGAAGATACTAGCGA
AATTAAAACAGGGATACCTGCCGCTTTGACGCTTAATAAAGCAGGTGTGGTCACTCG
TATTGACCTTTCTGACAGGCAAGTGGACAAGACGGCCTTGCCTATAAATGTGACAGC
TTATGATTTAGAAGACAAATTAGAGATTACAGGACGCGTTCTTGAAGAAACTAAGA
CTTATCCAGTGGATCATGATGCAACAATTGTTGAAGAGGATGGAACAGAAATTCGC
ATTGCACCGCTAGATGTGCAATATCAAAAGGCTAGTATTTGGGGACGTTTAATCACT
AATTTTGCAGGTCCCATGAATAACTTTATTTTAGGTATTTTTGTTTTTGCCCTCTTGAT
TTTTGTGCAAGGCGGTGTTCAGGATTCTTCAAGCAATCATGTGCGTGTGACTCCTAA
CAGTGCTGTAGCTAAGCTCGGACTTAAGAATAATGATCAAATTTTACAAATTGGGAA
AAACAAAGTTCATAATTGGAATGATCTCACTAATGCGGTTGCTAAGTCAACTAGTAA
TTTGAAAAAGAAAGAAGCTATTCCAGTTAAGGCTAAGACTCAAGGAAGCGTAAAAA

103
CTTTAAAAGTCATCCCTAAAAAAGTTAATGGGAATTACGTTATTGGTGTCATGCCAA
GTATGAAAACAGGATTTGGGGATAAAATTGTTGGTGCCTTTAAGATGTCTTGGGACG
GCGCTTTTGTTATCTTGAATGGTCTTAAAGGGCTAATCCTACAGCCAAGTCTCAATA
AATTAGGTGGTCCTGTTGCGATTTATCAACTGAGTAATACAGCTGCTAGAGAAGGTT
TTGCAAGAGTCCTTGAATTAATGGCTATGCTTTCTATTAATCTGGGTATTTTTAATTT
GTTGCCTATTCCTGCTCTTGATGGTGGTAAAATTTTAATCAATTTTATAGAAGTTATT
CGAAAAAAACCGCTCAAACAAGAGACAGAAACCTATATTACCCTCGCTGGTGTTCTT
ATTATGGTTGCGCTTATGATTGCAGTAACTTGGAATGATATCATGCGAGCATTTTTCT
AA
SMU_759 protease [ Streptococcus mutans UA159 ]
Gene ID: 1028149, updated on 26-Jun-2015
NCBI Reference Sequence: NC_004350.2
GenBank Graphics
>gi|347750429:710298-711224 Streptococcus mutans UA159 chromosome, complete genome
ATGGAAAAAATTGTTATCACTGCGACTGCAGAATCTATTGAACAAGTTAAAGAATTA
CTGACAAGTGGTGTTGACCGTATTTATGTTGGTGAGAAAGATTATGCGCTTCGTTTA
CCGCATGCGTTTAGCTATGATGACTTAAGAAAAATTGCTAGCTTGGTTCATGAAGCT
GGTAAAGAATTAACGGTTGCTGCTAATGCACTAATGCATCAAGAAATGATGGACAA
TATTAAACCATTTTTAGAATTAATGAAGGAAATTCAGGTAGATTACTTAGTGGTTGG
TGATGCAGGTGTTTTTTATGTCAATAAGCGTGATGGTTATCATTTTAAACTCATTTAT
GATACCTCTGTTTTTGTCACCTCTAGTCGTCAAGTTAATTTTTGGGGCCAACACGGTG
CGGTAGAAGCTGTTTTGGCACGTGAAATTCCTTCGGAAGAACTGTTTGAAATGTCCA
AAAATCTGGAAATTCCTGCAGAAGTCTTAGTTTACGGTGCTTCTGTCATTCATCATTC
CAAGCGACCTTTAATACAGAATTATTATAATTTTACTCACATTGATGATGAGAAGAC
AAGAGAACGCGGTCTGTTCTTATCAGAACCAAATGATCCTAAATCGCACTATTCTAT
ATATGAAGATAAACACGGCACTCATATTTTTATCAATAATGATATTGATTTGATGAC
CAAATTGCCTGAATTGATTAATCATCATTACAATCATTGGAAATTAGATGGTATCTA
TTGTCCAGGACATAATTTTGTTGAGATTGTTCAACTTTTTGTTAAAGCAAGAGATATG
ATCGAAGCTGGGACTTTTACGCAAGATCAGGCTTTTCTTTTCGATGAACAAATTAGA
AAGCTTCATCCAGCTGGTCGTGGTTTAGATACAGGATTTTATGAGCTTGATCCGCAA
ACAGTTAAGTAA
SMU_761 protease [ Streptococcus mutans UA159 ]
Gene ID: 1028148, updated on 27-Jun-2015
NCBI Reference Sequence: NC_004350.2
GenBank Graphics
>gi|347750429:711496-712782 Streptococcus mutans UA159 chromosome, complete genome
ATGACAAAACAATTAAAACGCCCAGAAGTGCTATCGCCTGCTGGGACTTTAGAAAA
ATTAAAAGTTGCTGTTAACTATGGAGCAGATGCTGTTTTTGTTGGCGGACAAGCTTA
TGGTTTGCGCAGTCGTGCAGGTAACTTTTCGATGGAAGAAATGGCTGAAGGAATTAA
TTATGCTCATGATCATGGGGTCAAGGTTTATGTGGCTGCTAACATGGTAACTCATGA
GGGCAATGAAATAGGAGCCGGTGCATGGTTTCGTGAATTACGCGACTTAGGTCTAG
ATGCAGTTATTGTATCGGATCCAGCCCTTATTGCGATTTGTGCGACAGATGCACCTG
GTTTGGAAATTCATTTGTCAACTCAAGCTTCATCCACTAACTATGAAACCTTTGAATT

104
TTGGAAAGAACTGGGCTTGACACGTGTTGTTTTAGCGCGTGAAGTCACAATGGCAGA
ACTAGCTGAGATTCGTAAGCGTACGAGTGTTGAAATTGAAGCCTTTGTTCATGGGGC
AATGTGTATTTCTTATTCAGGACGCTGTGTACTTTCCAATCATATGAGTCATCGCGAT
GCTAATCGTGGTGGTTGTTCACAATCTTGTCGTTGGAAATACAATCTTTATGATATGC
CTTTCGGTCAAGAAAGACGGTCATTGAAAGGTGAAGTACCAGAGGAATTTTCAATG
TCAGCTGTTGATATGTGCATGATTGAAAATATTCCAGACATGATTGAAAATGGTGTT
GATAGCCTTAAAATTGAAGGACGTATGAAGTCTATTCACTATGTTTCGACGGTCACA
AATTGTTACAAGGCGGCTGTCAATGCCTATCTGGAAAGCCCTCAAGCATTTGAAGCT
ATCAAACAAGATTTGATTGACGAATTGTGGAAAGTCGCTCAGCGTGAATTGGCTACA
GGTTTCTATTACCAAACACCTACTGAAAATGAACAGCTTTTTGGAGCTCGTCGTAAA
ATTCCCCAATATAAATTTGTCGGTGAAGTGGTTGATTTTGATGAGCCAAGTATGACA
GCAACTATTCGTCAGCGTAATGTCATTAATGAGGGGGATCGGGTTGAATTCTACGGA
CCTGGTTTCCGTCATTTTGAAACCTTTATTACAGATTTACATGATGCGGATGGTCAAA
AAATTGAACGTGCGCCAAAACCGATGGAGTTATTGACAATTACGGTACCACAGGAA
GTCAAAGCAGGTGATATGATTCGTGCCTGCAAGGAAGGCTTGGTCAATCTTTACAAA
GAAGATGGCAGCAGCCTTACTGTTAGAACTTAA
pepO peptidase (Zinc metalloproteinase) [ Streptococcus mutans UA159 ]
Gene ID: 1029222, updated on 26-Jun-2015
NCBI Reference Sequence: NC_004350.2
GenBank Graphics
>gi|347750429:c1910118-1908223 Streptococcus mutans UA159 chromosome, complete
genome
ATGGTACGTTTACAAGATGATTTTTATAACGCAGTCAATGGCCAGTGGGAAGAGGC
AGCGGTCATTCCTGATGATAAACCACGGACGGGTGGCTTTTCTGACTTGGCTGATGA
TATTGAAGATTTAATGTTAGAAACTACTGACAAGTGGCTAGATGGGAAAGATGTTCC
TGATGATAGTATTTTACAAAATTTTGTGAAGTTCCATCGTCAGGTGGCGGACTATGA
TGCGCGTGAAGAGACGGGTGTTAAGCCAGTGCTGCCTCTCATTGAAGAATATAAGA
GTCTAACTTCTTTTGCTGATTTTGCTTCCAACATAGCCACTTATGAAATGGCTGGCAA
GCCTAATGAGCTTCCTTTTGGTGTGGCACCGGATTTTATGAATGCACAAATGAATGT
GCTTTGGGCAGAGGCTCCAAATCTTATTTTACCAGATACCACTTATTATGCTGAAGG
TAATGACAAAGGTAAGGAACTGCTTGCTAAGTGGCGTACGATGCAAGAGGAACTTT
TGCCTAAGTTTGGTTTTGAAGAAGCAGAAATTAAAGATCTTCTAGATAAGGTGCTTA
CTTTAGATGCCAAATTGGCTCAATATGTTCTTTCCAGTGAGGAATCATCAGAATATG
TGAAGCTTTATCATCCTTATGATTGGGCTGATTTTACCAAATTAACACCAGAACTGC
CTTTAGATGCGATTTTTACACAGATTTTAGGTCAAAAACCAGATAAAGTTATCGTTC
CTGAAGAGCGTTTTTGGACAAATTTTGCAGCTGAATTTTATTCAGAAAAAAATTGGC
CTTTCTTAAAAGCTACCTTAGTTTTAGCTGCAGCAAGTTCTTACAATTCTTACCTGAC
AGATGATATTCGTATCCTTTCAGGAAGCTATAATCGTGCTCTTTCAGGGACACCTCA
AGCTATGGGTAAGAAAAAAGCCGCTTTTTATCTGGCTCAGGGCCCTTATAATCAAGC
GCTCGGTCTTTGGTACGCTGGCGAGAAATTTTCTCCTGAGGCAAAGAAAGACGTGGA
AGCTAAAGTGGCAACTATGATTGAGGTTTATAAAGAACGTTTGCATAAGACGGACT
GGTTGGCTCAAGAAACGCGTAATAAGGCTATTACCAAACTCAATGTCATAACGCCTC
ATATTGGTTATCCAGAACAATTACCCAAGACTTATGCTCAAAAGATTATTGACGACA
ATCTCAGTCTAGTGGAAAATGCTCAAAATTTGGCTAAAATCTCAATTGCCTATAATT

105
GGAGCAAGTGGAATCAACCAGTTGATCGCAGTGAATGGCATATGCCAGCTCACATG
GTTAATGCTTACTATGATCCGCAGCAAAATCAAATTGTCTTTCCAGCGGCTATTTTGC
AGGCACCATTTTATTCATTGGAGCAATCTTCATCTGCTAATTACGGTGGCATTGGTGC
TGTCATTGCCCATGAAATCTCTCACGCTTTTGATACGAATGGCGCTTCCTTTGATGAA
AATGGCAGTCTTAACAACTGGTGGACTGATGAAGATTATGCGGCTTTTAAAAAGCGT
ACAGACAGAGTTGTTGAACAGTTTGAAGGACTTGATTCTTATGGTGCTAAGGTCAAC
GGTCAGCTAACTGTTTCGGAAAATGTGGCTGATCTTGGTGGCCTTGCCTGTGCTCTTG
AAGCTGCCAAACGTGAAGCAGATTTTTCTGTCCGTGATTTCTTTATTAATTTTGCAAC
GATCTGGCGCATGAAAGCACGCGACGAATATATGCAAATGCTAGCAAGTATTGACG
TTCATGCTCCAGCTAAATGGCGGACCAATGTTACAATTACCAACTTTGACGAATTCC
ACCAAGAATTTGCGGTTAAAGAAGGTGATGGCATGTGGCGTGATGAAGATAAACGT
GTTATTATTTGGTAG




















