
The effect of age, pairing, copulation, and 2-Br-?-ergocryptine in the male Mongolian gerbil on the...
10
THE ANATOMICAL RECORD 208:411-420 11984) The Effect of Age, Pairing, Copulation, and 2-Br-~Ergocryptine in the Male Mongolian Gerbil on the Prolactin Cells in the Anterior Pituitary Identified by lmmunocytochemical and Herlant’s Tetrachrome Methods ROBERT ORTMAN AND FRANK PEREZ, JR. Department of Biology, The City College of New York, New York, NY 10031 ABSTRACT Near-adjacent, midsagittal sections of pituitaries of infantile and adult male Mongolian gerbils in several experimental groups (bachelor, paired, tartaric acid-injected, postcopulatory gerbil, and postcopulatory animal injected with 2-bromo-a-ergocryptine [CB-154]), were stained by a modification of Herlant’s tetrachrome and by the peroxidase-labeled antibody method for prolactin. Cell counts of erythrosinophils and cells immunoreactive for prolac- tin were made. Sequential staining procedures and thin adjacent sections were used to correlate the staining results. Erythrosinophils were very rare in the infant pituitaries; they increased (P< .01) in numbers in bachelor pituitaries, remained at the same level (P> .05) in paired animals, increased (P< .01) in the postcopulatory gerbils that were injected with tartaric acid, and increased much less (P < .05) in postcopulatory males that were injected with CB-154. Prolactin cells were present in modest numbers in infant pituitaries; they increased (P< .Ol); in bachelors and reached their highest number in paired animals (P < .001); they remained unchanged (P > .05) in tartaric acid-injected postcopulatory animals but declined (P < .001) in CB-154-injected,postcopula- tory animals. The number of prolactin cells was always significantly greater (P < .001) than the number of erythrosinophils. Correlative studies revealed the erythrosinophils, some of the light blue cells, and some of the chromophobes gave positive immunocytochemical reactions for prolactin. Apparently, CB-154 inhibited the erythrosinophils and the immunoreactive prolactin cells in the postcopulatory male gerbil, as indicated by a reduction in the number, size, and staining intensity of the cells. The role of prolactin in male mammals is not well understood (Hermanns and Hafez, 1981). One approach to this problem is to count prolactin cells at various times in the life history to ascertain periods when these cells are more numerous. Identification of such periods is the starting point for physio- logic experimentation. If the prolactin cells of the subject have not been studied exten- sively (e.g., gerbil) it is probably preferable to employ suitable classical staining methods in parallel with immunocytochemical meth- ods rather than to use either method inde- pendently. In this way, each method can function as a check on the results obtained with the other procedure. The erythrosinophils of the anterior lobe have long been regarded as prolactin cells principally because these cells undergo a dra- matic increase in size and numbers in late pregnancy and lactation and involute after weaning (Pasteels, 1963; Herlant, 1964a,b; Purves, 1966). The function of prolactin (along with adrenal cortical hormones) in ini- Received April 29, 1983; accepted September 13, 1983. Dr. Robert Ortman’s present address is General Delivery, Al- tona, NY 12910. 0 1984 ALAN R. LISS, INC.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The effect of age, pairing, copulation, and 2-Br-?-ergocryptine in the male Mongolian gerbil on the...

THE ANATOMICAL RECORD 208:411-420 11984)
The Effect of Age, Pairing, Copulation, and 2-Br-~Ergocryptine in the Male Mongolian Gerbil on the Prolactin Cells in the Anterior Pituitary Identified by lmmunocytochemical and Herlant’s Tetrachrome Methods
ROBERT ORTMAN AND FRANK PEREZ, JR. Department of Biology, The City College of New York, New York, N Y 10031
ABSTRACT Near-adjacent, midsagittal sections of pituitaries of infantile and adult male Mongolian gerbils in several experimental groups (bachelor, paired, tartaric acid-injected, postcopulatory gerbil, and postcopulatory animal injected with 2-bromo-a-ergocryptine [CB-154]), were stained by a modification of Herlant’s tetrachrome and by the peroxidase-labeled antibody method for prolactin. Cell counts of erythrosinophils and cells immunoreactive for prolac- tin were made. Sequential staining procedures and thin adjacent sections were used to correlate the staining results.
Erythrosinophils were very rare in the infant pituitaries; they increased (P< .01) in numbers in bachelor pituitaries, remained at the same level (P> .05) in paired animals, increased (P< .01) in the postcopulatory gerbils that were injected with tartaric acid, and increased much less (P < .05) in postcopulatory males that were injected with CB-154.
Prolactin cells were present in modest numbers in infant pituitaries; they increased (P< . O l ) ; in bachelors and reached their highest number in paired animals (P < .001); they remained unchanged (P > .05) in tartaric acid-injected postcopulatory animals but declined (P < .001) in CB-154-injected, postcopula- tory animals. The number of prolactin cells was always significantly greater (P < .001) than the number of erythrosinophils.
Correlative studies revealed the erythrosinophils, some of the light blue cells, and some of the chromophobes gave positive immunocytochemical reactions for prolactin.
Apparently, CB-154 inhibited the erythrosinophils and the immunoreactive prolactin cells in the postcopulatory male gerbil, as indicated by a reduction in the number, size, and staining intensity of the cells.
The role of prolactin in male mammals is not well understood (Hermanns and Hafez, 1981). One approach to this problem is to count prolactin cells a t various times in the life history to ascertain periods when these cells are more numerous. Identification of such periods is the starting point for physio- logic experimentation. If the prolactin cells of the subject have not been studied exten- sively (e.g., gerbil) it is probably preferable to employ suitable classical staining methods in parallel with immunocytochemical meth- ods rather than to use either method inde- pendently. In this way, each method can function as a check on the results obtained
with the other procedure. The erythrosinophils of the anterior lobe
have long been regarded as prolactin cells principally because these cells undergo a dra- matic increase in size and numbers in late pregnancy and lactation and involute after weaning (Pasteels, 1963; Herlant, 1964a,b; Purves, 1966). The function of prolactin (along with adrenal cortical hormones) in ini-
Received April 29, 1983; accepted September 13, 1983.
Dr. Robert Ortman’s present address is General Delivery, Al- tona, NY 12910.
0 1984 ALAN R. LISS, INC.

412 R. ORTMAN AND F. PEREZ, JR.
tiation and maintenance of lactation is also well known (Meites and Clemens, 1972). Im- munocytochemical studies have confirmed that the coarsely granulated erythrosino- phils are prolactin cells (Stokes and Boda, 1968; Pasteels et al., 1972; Herbert and Hay- ashida, 1974; Herbert, 1978).
Ortman and Heimann (1980) studied the erythrosinophils of the male gerbil from in- fancy through the postcopulatory period. It was found that erythrosinophils were vir- tually absent in the infant and were more abundant around puberty and in the early postcopulatory period. The injection of the postcopulatory male with 2-Br-a-ergocryp- tine (CB-154), a potent prolactin cell inhibi- tor (Meites and Clemens, 19721, reduced the weight of the seminal vesicles, whereas the weight of this organ was maintained after concurrent injection of CB-154 and ovine pro- lactin. These results implicated endogenous prolactin in the maintenance of this sex ac- cessory organ after copulation. The presump- tion was made that CB-154 suppressed the secretory activity of the prolactin cells, but cytologic confirmation was not attempted.
It was of interest to obtain, after staining with Herlant’s tetrachrome and an immuno- cytochemical method, 1) cytologic confirma- tion of the inhibitory action of CB-154 on the prolactin cells in the postcopulatory gerbil and 2) cell count data on prolactin cells in infantile gerbils and adult male gerbils in different conditions of existence (bachelor, paired, and postcopulatory).
MATERIALS AND METHODS
The colony of Mongolian gerbils (Meriones unguiculatus) was developed from breeding stock obtained from Tumblebrook Farms, Inc. (Westbrookfield, MA). Colony animals were maintained in plastic cages in unisexual groups. Cages were provided with wood shav- ings. Animals were given rat chow and water ad libitum. The procedures employed for pairing adult male and female animals and for obtaining the date of copulations at the postpartum estrus were described previously (Ortman and Heimann, 1980).
The experimental animals were divided into five groups:
Seven infantile males sacrificed when 21 days old.
Six bachelor males (adult animals that had lived in unisexual groups from weaning until sacrifice).
Six uninjected, paired males (male coex- isted with female during pairing test, but no
copulations were observed). A familiariza- tion period of about 1 week preceded the pair- ing test.
Nine postcopulatory males injected daily with CB-154 (Sandoz, Inc.) suspended in 0.5% tartaric acid. The animals received 0.6 mgi 100 gm body weightiday S.C. for 10 days; the first injection was on the day after copula- tion. The range of actual dose was 0.41-0.56 mg. The males were retained with their ma- tes and offspring during the injection period and were killed on the 11th day postcopu- lation.
Six postcopulatory males given 10 daily in- jections of 0.5% tartaric acid.
All gerbils were killed with chloroform. Pi- tuitaries were fixed in Zenker-formol. Five- micron serial, sagittal paraffin sections were cut. Some adjacent sections were cut at 2 and 3 pm for cell identification studies. Cell count studies were carried out with pairs of slides containing near-adjacent sections from the midsagittal region. Sections on one slide were stained by a modification of the Herlant (1960) method, as previously described (Ort- man and Heimann, 19801, to demonstrate tinctorial cell types. Sections of the other slide were stained immunocytochemically by the peroxidase-labeled antibody procedure of Nakane and Pierce (1967) to demonstrate prolactin cells. Sections were exposed to rab- bit antiserum to rat prolactin (NIADDK) di- luted 1:lOO with Tris buffer for 24 hours at 5°C. Following a buffer wash, the sections were treated with goat antirabbit gamma globulin conjugated to horseradish peroxi- dase (Antibodies, Inc.) and diluted 1:50 with Tris buffer for 1.5 hour a t room temperature. Diaminobenzidine tetrahydrochloride (DAB) was used as the substrate in the peroxidase reaction.
Controls for the specificity of the immuno- cytochemical method involved 1) absorption of 1 ml of primary antibody with 30 pg of rat prolactin (NIADDK) for 48 hours a t 5°C or with 35 pg of rat prolactin a t 5°C for 24 hours or a t 37°C for 1 hour. Additional anti- serum was maintained at 37°C for 1 hour without antigen; 2) absorption of 2 ml of pri- mary antibody with 130 pg of rat growth hormone (NIADDK) at 37°C for 1 hour (Midgley, 1966); 3) absorption of 1 ml of pri- mary antibody with 215 pg porcine follicle- stimulating hormone (FSH) (Sigma), 180 pg equine luteinizing hormone (LH) (Cal- Biochem), or 170 pg bovine thyroid-stimulat- ing hormone (TSH) (Sigma) at 5°C for 24 hours; 4) replacement of either the primary

PROLACTIN CELLS OF PITUITARY OF MALE GERBIL 413
or the secondary antibody with Tris buffer; 5 ) replacement of the primary antibody with 1% normal rabbit serum in Tris buffer; 6) replacement of the secondary antibody with goat antimonkey IgG (Cappel) diluted 1 5 0 with Tris buffer at pH 7.6; 7) application of the immunocytochemical method to organs (fixed in Zenker-formol) not known to pro- duce prolactin (thyroid, esophagus, trachea, seminal vesicles and testes); 8) exposure of sections to a DAB solution without prior treatment of sections with the antibody so- lutions; and 9) application of the immunocy- tochemical method to pituitaries (Zenker- formol fixed) of chicken and rat.
Cell counts were performed with an oil im- mersion objective (100 X) on a single repre- sentative section for each specimen (after several stained sections were examined), ac- cording to the procedures described previ- ously (Ortman and Heimann, 1980). Only cells having their nucleus in the plane of the section and touching, or included within, the outer borders of the ocular grid field were counted. The counts obtained from the grid fields involved in the scan of the section were summated to give the relative number of cells for the specimen: Group means were calcu- lated from these values. Levels of statistical significance among the group means were calculated using the Student’s t-test.
Correlation of the prolactin cells with the classical cell types was achieved by 1) placing adjacent sections, cut at 2 or 3 pm, on sepa- rate microslides and staining one with Her- lant’s tetrachrome and the other with the immunocytochemical method and by 2) car- rying out an immunocytochemical staining- destaining and restaining sequence. Destain- ing can be carried out with absolute ethanol if DAB is replaced by 4-chloro-1-naphthol (Pfaltz and Bauer, Inc.) (Nakane, 1968).
RESULTS
The ventral surface of the gerbil pituitary exhibited a morphological feature that was not present on the dorsal side: a subdivision of the anterior lobe into a median region (“lobe”) and two lateral regions (“lobes”). This condition occurred because the median region projected further ventrally than did the lateral regions.
The following parenchymal cell types were present in the anterior lobe after Herlant’s tetrachrome (Ortman and Heimann, 1980): erythrosinophils (coarse, erythrosin-positive secretory granules-appeared red); auranti- phils (fine, orange G-positive granules-ap-
peared yellow); light blue cyanophils; dark blue cyanophils; and chromophobes (“naked” nuclei or with unstaining cytoplasm). The bulk of the erythrosinophils were confined to the median region and appeared to be dis- tributed symmetrically on both sides of the midsagittal plane. Aurantiphils and cyano- phils were abundant in the median and the lateral regions.
Positive staining with the immunocyto- chemical method occurred when the secre- tory granules developed a dark brown or brownish black color in the DAB solution, so that the cell stood out against a generally colorless background. On the basis of several control studies, presented below, it was con- cluded that the immunocytochemical method was probably specific for the immunoreactive prolactin cells (designated prolactin cells). Positive staining did not occur if a) either the primary or secondary antiserum was re- placed with Tris buffer, b) the primary anti- serum was replaced by normal rabbit serum, c) the antibodies to rat prolactin were ab- sorbed with rat prolactin under any of the conditions of temperature and time specified for the absorption studies, d) the secondary antibody was not immunologically suitable to react with the primary antibody, or e) both primary and secondary antisera were omit- ted. Positive staining was not noticeably af- fected when the primary antisera had been absorbed with rat growth hormone, porcine FSH, equine LH, or bovine TSH. Finally, a positive reaction occurred in the anterior lobe of the rat but not in the pituitary of the chicken or in the posterior lobe, thyroid, esophagus, trachea, seminal vesicles or testes of the gerbil.
The study of near-adjacent sections from different parasagittal levels in the lateral and median regions has indicated that the distribution of the immunoreactive prolactin cells paralleled the distribution of the ery- throsinophiIs in the gland. Figure 1 shows the body weights and the mean relative num- bers of erythrosinophils and prolactin cells in the midsagittal plane in the several groups. The mean body weights of the adult gerbil groups did not differ significantly from one another. Erythrosinophils were very rare in the infant pituitary (in this group, the scan- ning method was modified to scan every dor- sal-ventral row on the section because there were so few erythrosinophils); they increased in numbers (P< .01) in bachelor pituitaries remained at the same level (P > .05) in paired animals, increased (P < .01) in postcopulatory
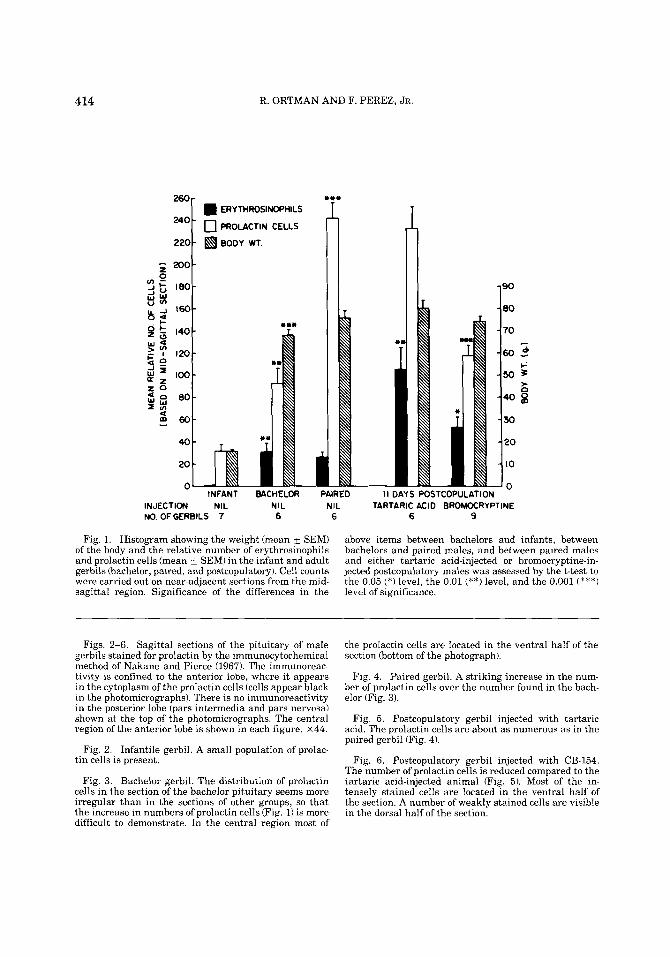
414 R. ORTMAN AND F. PEREZ. JR.
ERYTHROSINOPHILS
PROLACTIN CELLS
140 - 120
loo - 80 -
60-
40-
20 - n-
-
*
1'0 80
70 c
604
2 40 8
w s
30
20
10
n " " INFANT BACHELOR PARED I I DAYS POSTCOPULPTION
INJECTION NIL NIL NIL TARTARIC ACID BROMOCRYPTINE NO. OFGERBILS 7 6 6 6 9
Fig. 1. Histogram showing the weight (mean i SEM) of the body and the relative number of erythrosinophils and prolactin cells (mean k SEM) in the infant and adult gerbils (bachelor, paired, and postcopulatory). Cell counts were carried out on near-adjacent sections from the mid- sagittal region. Significance of the differences in the
above items between bachelors and infants, between bachelors and paired males, and between paired males and either tartaric acid-injected or bromocryptine-in- jected postcopulatory males was assessed by the t-test to the 0.05 (*) level, the 0.01 (**) level, and the 0.001 (***) level of significance.
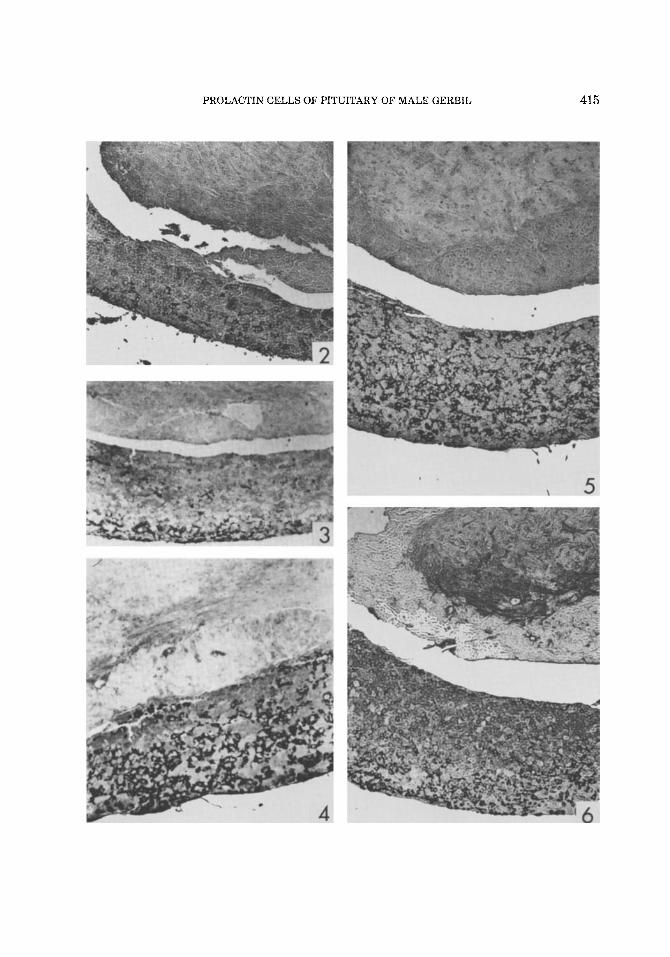
Figs. 2-6. Sagittal sections of the pituitary of male gerbils stained for prolactin by the immunocytochemical method of Nakane and Pierce (1967). The immunoreac- tivity is confined to the anterior lobe, where it appears in the cytoplasm of the prolactin cells (cells appear black in the photomicrographs). There is no immunoreactivity in the posterior lobe (pars intermedia and pars nervosa) shown at the top of the photomicrographs. The central region of the anterior lobe is shown in each figure. ~ 4 4 .
Fig. 2. Infantile gerbil. A small population of prolac- tin cells is present.
Fig. 3. Bachelor gerbil. The distribution of prolactin cells in the section of the bachelor pituitary seems more irregular than in the sections of other groups, so that the increase in numbers of prolactin cells (Fig. 1) is more difficult to demonstrate. In the central region most of
the prolactin cells are located in the ventral half of the section (bottom of the photograph).
Fig. 4. Paired gerbil. A striking increase in the num- ber of prolactin cells over the number found in the bach- elor (Fig. 3).
Fig. 5. Postcopulatory gerbil injected with tartaric acid. The prolactin cells are about as numerous as in the paired gerbil (Fig. 4).
Fig. 6. Postcopulatory gerbil injected with CB-154. The number of prolactin cells is reduced compared to the tartaric acid-injected animal (Fig. 5). Most of the in- tensely stained cells are located in the ventral half of the section. A number of weakly stained cells are visible in the dorsal half of the section.
PROLACTIN CELLS OF PITUITARY OF MALE GERBIL 415
416 R. ORTMAN AND F. PEREZ, JR.

PROLACTIN CELLS OF PITUITARY OF MALE GERBIL 417
gerbils injected with vehicle, and increased much less (P< .05) in postcopulatory males injected with CB-154. There were signifi- cantly more (generally P < .001) prolactin cells than erythrosinophils in infant and adult groups. The prolactin cells were at a low level in the infant (Fig. 2), increased sig- nificantly (P < .01) in the bachelor gerbil (Fig. 31, and further increased in numbers in the paired male (P < .001) (Fig. 4). The number of prolactin cells in tartaric acid-injected, post- copulatory males (Fig. 5) was not signifi- cantly (P > .05) different from the number for the paired males. There was a significant decline (P < .001) in the number of prolactin cells in the postcopulatory male injected with bromocryptine (Fig. 6).
The erythrosinophils in the tartaric acid- injected postcopulatory gerbils stained rela- tively strongly and often were large cells (Fig. 7). After bromocryptine injections, many of these cells were smaller and stained less in- tensely because they contained fewer secre- tory granules (Fig. 8). No obvious effect of the drug on the aurantiphils or the cyano- phils was observed.
To explain the numerical discrepancy be- tween the erythrosinophils and the immuno- reactive prolactin cells, the tinctorial identity of the prolactin cells was investigated by four approaches:
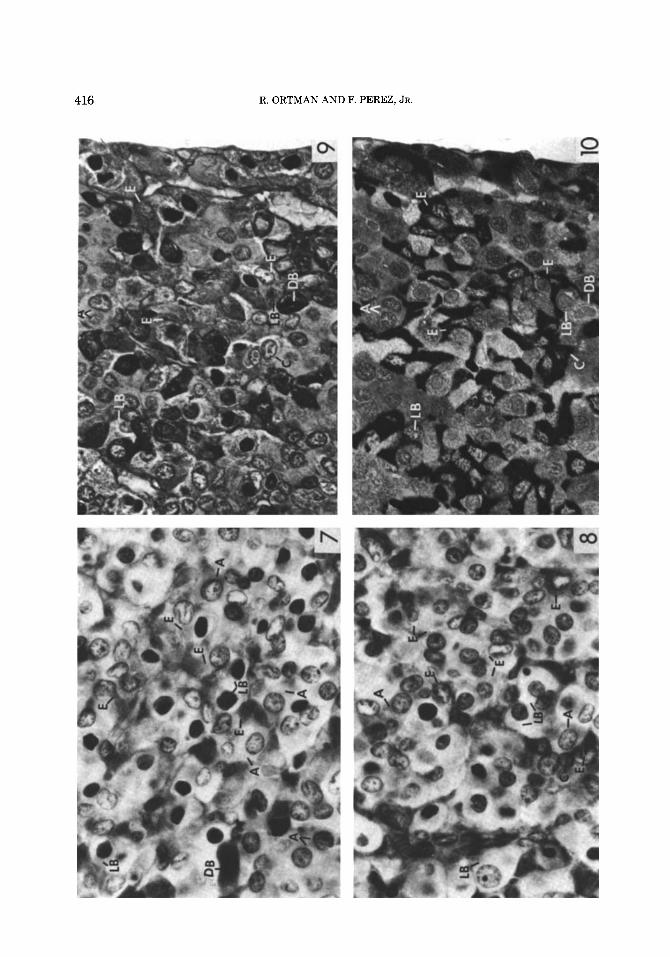
Figs. 7-10. Figures 7-9 were stained by Herlant’s method; Figure 10 was stained immunocytochemically. A, aurantiphil; C, chromophobe; E, erythrosinophil; DB, dark blue cell; and LB, light blue cell.
Fig. 7. Postcopulatory gerbil injected with tartaric acid. Herlant’s method. Photographed with a green fil- ter. The E cells are relatively large and stain strongly. x 175.
Fig. 8. Postcopulatory gerbil injected with CB-154. Herlant’s method. Photographed with a green filter. In general, the E cells are smaller and stain weakly. x 175.
Fig. 9. Paired male. The 2-pm section was stained with Herlant’s method and photographed with a cobalt blue filter. The E cells appear lighter and the LB cells appear darker than cells of the same designation in Figures 7 and 8 that were photographed with a green filter. All the cells labeled are present also in the adja- cent section (Fig. 10). ~ 2 7 6 .
Fig. 10. Paired male. The 2-pm section, adjacent to the section in Figure 9, was stained by the immunocyto- chemical method for prolactin. Observe that the three E cells, the LB cell in the upper left, and the C cell near the lower border are immunoreactive cells. The LB cell near the lower border, the DB cell, and the A cells near the upper border are not stained. ~ 2 7 6 .
1) The sections stained by the Herlant method were destained and carried through the immunocytochemical method. This ap- proach failed because the immunoreactivity was blocked (or lost) by previous exposure to the Herlant method. 2) The sections stained by the immunocytochemical method were counterstained by the Herlant procedure. In sections of adult pituitary, many light and dark blue cyanophils and aurantiphils were apparent. All immunoreactive cells were also stained with erythrosin, giving these cells a muddied appearance. However, with infant pituitary, the flaw in this approach became apparent: There were many more erythrosin- positive and immunoreactive cells than there were erythrosinophils in the infant. 3) By substituting 4-chloro-1-naphthol for DAB, the reaction product was made soluble in ethanol and, after selected cells were recorded, could be removed before staining with tetrach- rome. By this approach it was found that many of the immunoreactive cells were light blue cyanophils. 4) The comparison of adja- cent sections, in which one section was stained by the Herlant method (Fig. 9) and the other by the immunocytochemical method (Fig. 101, has clearly shown that some of the light blue cells, some of the chromo- phobes, and probably all of the erythrosino- phils gave a positive immunocytochemical reaction for prolactin.
Frequently, immunoreactive cells partially surround (cup) nonimmunoreactive cells. A similar observation was made previously on the erythrosinophils (Ortman and Heimann, 1980). Cupping by other tinctorial types of cells has not been observed. Thus, cupping provides additional evidence that erythrosi- nophils are prolactin cells.
DISCUSSION
In the anterior lobe of all mammalian spe- cies that have been studied after staining with Herlant’s tetrachrome, the erythrosino- phils with coarse secretory granules are pro- lactin cells while the finely granulated orange G-positive cells (aurantiphils) are growth hormone cells (Purves, 1966). After Brookes’s (1968) method, which substitutes carmoisine L for erythrosin, the carmoisino- philic cells are the prolactin cells (Goluboff and Ezrin, 1969). Immunocytochemical methods have confirmed the above func- tional identifications that were based on his- tophysiological studies using the Herlant method (Stokes and Boda, 1968; Pasteels et

418 R. ORTMAN AND F. PEREZ, JR.
al., 1972) or employing the Brookes method (Herbert and Hayashida, 1974; Herbert, 1978). Herlant (1964a,b) has also described a second type of erythrosinophil which con- tains a small number of fine erythrosino- philic granules.
The coarsely granulated erythrosinophils and aurantiphils have been found more re- cently in the anterior lobe of adult Indian desert gerbils {Meriones hurrianae) (Kaul and Ramaswami, 1974) and the adult Mongolian gerbil (Meriones unguiculatus) (Ortman and Heimann, 1980). The finely granulated ery- throsinophil has not been observed in ger- bils. The failure of Bagwell and Leavitt (1970) to demonstrate two tinctorial types of acido- phils in the Mongolian gerbil was probably due to their choice of staining methods.
A brief description of the cyanophils of ger- bils is required in order to discuss adequately our immunocytochemical results. Kaul and Ramaswami (1974) and Ortman and Hei- mann (1980) have described light blue cells and dark blue cells. These cells are numer- ous in both regions of the anterior lobe. Us- ing the oxidation alcian blue-periodic acid- Schiff (PAS)-orange G method of Herlant (19601, Heimann (1981) found that the light blue cells of the Mongolian gerbil were sepa- rable into two types: l) alcian blue cells and 2) PAS-purple cells. Finally, he determined, by immunocytochemistry, that the alcian blue cells were thyrotrophs.
Our immunocytochemical method ap- peared to be specific for immunoreactive pro- lactin. Positive reactions were confined to certain cells in the anterior lobe of the gerbil and the rat. No positive reactions were ob- tained in the posterior lobes of these species, in the pituitary of the chicken or in the thy- roid, esophagus, trachea, seminal vesicles, or testes of the gerbil. Nolin (1978), employing the same immunocytochemical method, ob- tained positive reactions in the rat testes after Bouin fixation. The positive reaction in the anterior lobe of the gerbil did not develop when the primary antibody was absorbed by homologous antigen, whereas treatment of the primary antibody with considerably more growth hormone, FSH, LH, or TSH was in- effective in absorbing the antibody.
The cell counts based on the Herlant method consistently underestimated the number of prolactin cells that were present in the pituitary gland. Our study of the iden- tity of the cell types that were immunoreac- tive indicated that two investigative ap-
proaches were useful: a) a sequential stain- ing procedure in which the reaction product, formed in the immunocytochemical method, was removed before the Herlant method was applied. This was necessary because the re- action product, formed when DAB was the substrate, was erythrosinophilic; b) the use of adjacent sections.
We found that erythrosinophils (probably all of them), some chromophobes, and some light blue cells were immunoreactive for pro- lactin. Our finding on erythrosinophils agrees with the results obtained on other mammalian species mentioned above. Im- munoreactivity in some chromophobes is probably due to the greater sensitivity of the immunocytochemical method relative to the Herlant method because of the two steps of amplification of the immunologic response (Nakane and Pierce, 1967; Pearse, 1968). Im- munoreactivity of some light blue cells may reflect a highly developed ergastoplasm (which stains cyanophilically) in actively se- creting prolactin cells that have a low con- tent of secretory granules (Herlant, 1964a; Herlant and Pasteels, 1967).
Ortman and Heimann (1980) reported that erythrosinophils were virtually absent in the anterior lobe of 21-day-old gerbils, although bioassayable prolactin was present. In the present study, we have found a small popu- lation of immunoreactive prolactin cells in these animals. This finding harmonizes with the bioassay data and is explainable on the basis on the increased sensitivity of the method. Pasteels et al. (1972) reported a sim- ilar situation in the infant rhesus monkey. In contrast, Tassel1 and Kennedy (1972) found erythrosinophils as well as aurantiphils in the pituitary of term fetuses of sheep.
The interpretation of the cellular changes observed in the pituitary of paired male ger- bils is hampered by a lack of accompanying prolactin assay data. The dramatic rise in the number of immunoreactive prolactin cells in the paired male was novel and unex- pected, since there was no significant differ- ence in the number of erythrosinophils between bachelor and paired animals.
Pairing is apparently a stressful process inasmuch as an unfamiliar adult male and anestrous female will fight when brought to- gether directly. Since stress is a recognized stimulus of prolactin secretion (Noel et al., 1972; Frantz et al., 1972), it may be that stress increased the number of immunoreac- tive prolactin cells. A second possibility is

PROLACTIN CELLS OF PITUITARY OF MALE GERBIL 419
that stimuli associated with sex caused an increase in the number of prolactin cells in the male. Kamel et al. (1977) have reported that unsuccessful as well as successful mat- ing caused a rise in the blood titers of prolac- tin, luteinizing hormone, and testosterone.
We have confirmed the increase in the number of erythrosinophils in the postcopu- latory male; in addition, we found no change in the number of immunoreactive prolactin cells. Interpretation of these changes is diffi- cult because only behavioral studies on cop- ulation in the gerbil are available as far as we are aware (Kuehn and Zucker, 1968; Dewsbury, 1972; Davis et al., 1974). Hor- mone assays of samples of blood obtained within minutes after copulation have been done on several mammals. Bryant et al. (19701, Convey et al. (1971), and Kamel et al. (1975, 1977) have reported acute elevations in blood prolactin after copulation in goats, dairy bulls, and rats, respectively; but other studies of rats (Spies and Niswender, 1971; Balin and Schwartz, 1976) or humans (Noel et al., 1972) have not found such a change. Apparently, no assay studies have been ex- tended to include the time period used in the present study. The increased number of ery- throsinophils might reflect an increase in prolactin storage. These erythrosinophils may have come from chromophobes or from light blue cells by a process of regranulation.
Most of the erythrosinophils and the im- munoreactive prolactin cells that remained in the pituitary after 10 days of CB-154 treat- ment were small cells with small nuclei; they stained less intensely with both methods. Studying prolactin cells of ectopic rat pitui- taries exposed to CB-154 for 22 days, Graf et al. (1977) reported similar changes in cell numbers and cell size. In the gerbil there was no obvious effect of CB-154 on the auranti- phils or cyanophils. At small or moderate doses, CB-154 is considered a specific inhibi- tor of prolactin cells; the release of other an- terior lobe hormones is not affected (Meites, 1977). Our cytological observations seem to agree with Meites’s position.
It was previously reported (Ortman and Heimann, 1980) that the weight of the sem- inal vesicles was significantly reduced in CB- 154-injected postcopulatory gerbils. This ef- fect was counteracted by concurrent admin- istration of prolactin. It was suggested that prolactin that was secreted during the postcopulatory period was important to the maintenance of the seminal vesicles; proba-
bly the CB-154 blocked the secretion of en- dogenous prolactin. The cytological data obtained in the present study clearly sup- ports this interpretation.
ACKNOWLEDGMENTS
We are pleased to acknowledge the gift of CB-154 by Sandoz, Inc. (East Hanover, NJ), the scientific illustration work of Mrs. G. Fisher, and the photographic and artistic as- sistance of Professor J.J. Lee. Finally, we wish to thank Dr. J.L. Pasteels, Faculte de Medecine, Laboratoire d’Histologie, Univer- site Libre de Bruxelles, Belgium, for helpful suggestions and critical reading of an early draft of the manuscript.
This investigation was supported in part by NIH Biomedical Research Support grants 2 507RRO7132-09 and -10. Frank Perez, Jr., was supported in part by the MBRS Program NIH grant RR-08168.
LITERATURE CITED
Bagwell, J.N., and W.W. Leavitt (1970) The anterior pituitary cel1 types of the Mongolian gerbil. Anat. Rec., 166:274 (Abstr.).
Balin, M.S., and N.B. Schwartz (1976) Effects of mating on serum LH, FSH, and prolactin and accessory tissue weight in male rats. Endocrinology, 98.522-526.
Brookes, L.D. (1968) A stain for differentiating two types of acidophil cells in the rat pituitary. Stain Technol., 43:41-42.
Bryant, G.D., J.L. Linzell, and F.C. Greenwood (1970) Plasma prolactin in goats measured by radioimmu- noassay: The effects of teat stimulation, mating behav- ior, stress, fasting and of oxytocin, insulin and glucose injections. Hormones, 1:26-35.
Convey, E.M., E. Bretschneider, H.D. Hafs, and W.D. Oxender (1971) Serum levels of LH, prolactin and growth hormone after ejaculation in bulls. Biol. Re- prod., 520-24.
Davis, H.N., Jr., D.Q. Estep, and D.A. Dewsbury (1974) Copulatory behavior of Mongolian gerbils (Meriones unguiculatus). Anim. Learn. Behav., 2:69-73.
Dewsbury, D.A. (1972) Patterns of copulatory behavior in male mammals. Quart. Rev. Biol., 47:l-33.
Frantz, A.G., D.L. Kleinberg, and G.L. Noel (1972) Stud- ies on prolactin in man. Recent Prog. Horm. Res., 28:527-590.
Goluboff, L.G., and C. Ezrin (1969) Effect of pregnancy on the somatotroph and the prolactin cell of the human adenohypophysis. J. Clin. Endocrinol. Metab., 29: 1533- 1538.
Graf, K.-J., R. Horowski, and M.F. El Etreby (1977) Ef- fect of prolactin inhibitory agents on the ectopic ante- rior pituitary and the mammary gland in rats. Acta Endocrinol., 85:267-278.
Heimann, A. (1981) A study of the thyrotroph in the male Mongolian gerbil. MD thesis, Yale University School of Medicine.
Herbert, D.C. (1978) Stimulation of prolactin secretion by testosterone in juvenile male Rhesus monkeys. Biol. Reprod., 18:448-453.
Herbert, D.C., and T. Hayashida (1974) Histologic iden- tification and immunochemical studies of prolactin and

420 R. ORTMAN AND F. PEREZ. JR.
growth hormone in the primate pituitary gland. Gen. Comp. Endocrinol..,24:381-397.
Herlant, M. (1960) Etude critique de deux techniques nouvelles destinee a mettre en evidence les differentes categories cellulaires presentes dans la glande pitui- taire. Bull. Micr. Appl., 1Or37-44,
Herlant, M. (1964a) The cells of the adenohypophysis and their functional significance. Int. Rev. Cytol., 17299-382.
Herlant, M. (1964b) Present state of knowledge concern- ing the cytology of the anterior lobe of the hypophysis. In: Proceedings of the Second International Congress of Endocrinology. Excerpta Medica International Con- gress, Series No. 83t468-481.
Herlant, M., and J.L. Pasteels (1967) Histophysiology of human anterior pituitary. Methods Achiev. Exp. Pa- thol, 3:250-305.
Hermanns, U., and E.S.E. Hafez (1981) Prolactin and male reproduction. Arch. Androl., 6:95-125.
Kamel, F., E.J. Mock, W.W. Wright, and A.I. Frankel (1975) Alterations in plasma concentrations of testos- terone, LH, and prolactin associated with mating in the male rat. Horm. Behav., 6:277-288.
Kamel, F., W.W. Wright, E.J. Mock, and A.I. Frankel (1977) The influence of mating and related stimuli on plasma levels of luteinizing hormone, follicle stimulat- ing hormone, prolactin and testosterone in the male rat. Endocrinology, 101:421-429.
Kaul, D.K., and L.S. Rasmaswami (1974) Cytology of the pituitary gland of the Indian desert gerbil Meriones hurrianae Jerdon. Acta Anat., 88.299-319.
Kuehn, R.E., and I. Zucker (1968) Reproductive behavior of the Mongolian gerbil (Meriones unguiculatus). J. Comp. Physiol. Psychol., 66t747-752.
Meites, J. (1977) Evaluation of research on control of prolactin secretion. In: Comparative Endocrinology of Prolactin. H.-D. Dellmann, J.A. Johnson, and D.M. Klachko, eds. Plenum, New York, pp. 135-152.
Meites, J., and J.A. Clemens (1972) Hypothalamic con- trol of prolactin secretion. Vitam. Horm., 30:165-221.
Midgley, A.R., Jr. (1966) Human pituitary luteinizing hormone: An immunohistochemical study. J. Histo- chem. Cytochem., 14:159-166.
Nakane, P.K. (1968) Simultaneous localization of multi- ple tissue antigens using the peroxidase-labeled anti- body method: A study on pituitary glands of the rat. J. Histochem. Cytochem., 16557-560,
Nakane, P.K., and G.B. Pierce, Jr. (1967) Enzyme la- beled antibodies for the light and electron microscope localization of tissue antigens. J. Cell Biol., 33:307- 318.
Noel, G.L., H.K. Suh, J.G. Stone, and A.G. Frantz (1972) Human prolactin and growth hormone release during surgery and other conditions of stress. J. Clin. Endocri- nol. Metab., 35840-851.
Nolin, J.M. (1978) Target cell prolactin. In: Structure and Function of the Gonadotrophins. K.W. McKerns, ed. Plenum, New York, pp. 151-182.
Ortman, R., and A. Heimann (1980) Changes in erythro- sinophils of the anterior lobe of the adenohypophysis during sexual development and after copulation in the male Mongolian gerbil. Biol. Reprod., 22:1245-1253.
Pasteels, J.L. (1963) Recherches morphologiques et ex- perimentales sur la secretion de prolactine. Arch. Biol. (Brussels), 74:439-553.
Pasteels, J.L., P. Gausset, A. Danguy, F. Ectors, C.S. Nicoll, and P. Varavudhi (1972) Morphology of the lac- totropes and somatotropes of man and Rhesus monkey. J. Clin. Endocrinol. Metab., 34:959-967.
Pearse, A.G.E. (1968) Histochemistry, 3rd ed., Vol 1. Wil- liams and Wilkins Company, Baltimore, p. 183.
Purves, H.D. (1966) Cytology of the adenohypophysis. In: The Pituitary Gland G.W. Harris and B.T. Donovan, eds. University of California Press, Berkeley, Vol. 1 pp. 147-232.
Spies, H.G., and G.D. Niswender (1971) Levels of prolac- tin, LH and FSH in the serum of intact and pelvic- neurectomized rats. Endocrinology, 88t937-942.
Stokes, H., and J.M. Boda (1968) Immunofluorescent lo- calization of growth hormone and prolactin in the ad- enohypophysis of fetal sheep. Endocrinology, 83:1362- 1366.
Tassell, R., and J.P. Kennedy (1972) Tinctorial differen- tiation of the cell types in the pars anterior of the sheep. Aust. J. Biol. Sci., 25:1249-1257.




















