
The deep-sea copepod fauna of the Southern Ocean: patterns and processes
18
Hydrobiologia 453/454: 37–54, 2001. R.M. Lopes, J.W. Reid & C.E.F.Rocha (eds), Copepoda: Developments in Ecology, Biology and Systematics. © 2001 Kluwer Academic Publishers. Printed in the Netherlands. 37 The deep-sea copepod fauna of the Southern Ocean: patterns and processes Peter Ward & Rachael S. Shreeve British Antarctic Survey, NERC, High Cross, Madingley Rd., Cambridge CB3 0ET, U.K. E-mail: [email protected] Key words: deep-sea copepods, diversity, distribution, production, metabolism, Southern Ocean Abstract In recent years, much attention has been paid to the Antarctic epipelagic fauna, as a result of the desire to increase our knowledge of ecosystem function and resource management. Unfortunately, our understanding of the polar pelagic deep-sea has not progressed as fast, and in common with many other parts of the world’s deep ocean, knowledge is still fragmentary. As yet, we have an incomplete but evolving knowledge of species presence and distribution, but very little idea of how the extreme seasonality seen in the Southern Ocean might influence the deep-water fauna. An examination is made of species distribution and diversity, in relation to the latitudinal cline seen in many benthic groups, and the historical perspective offered by changing circulation patterns and sea temperature through geological time. Although a number of important frontal systems are found within the circumpolar Southern Ocean, the boundary is marked by the Sub-Tropical Convergence, which appears to be the major biogeographic boundary between it and surrounding provinces. Evidence for seasonality in various families is reviewed in light of what we know and can infer about their biology and particularly in respect of their bathymetric distribution, which in some groups appears to change with latitude. Introduction In general, little is known about the pelagic deep-sea, despite the fact that depths below 1 km comprise 62% of the surface of the earth and 79% of the volume occupied by life (Childress, 1995). The deep-sea has traditionally been of more interest to taxonomists than ecologists, primarily because many of the species have been rarely sampled, and because of the diffi- culties and costs of obtaining sufficient material for experimental work. There are some notable exceptions where deep-water sites have received detailed study, e.g. the time series undertaken in the Rockall Trough (Mauchline, 1992, 1994), the SOND cruise’s study on the Abyssal Madeira Plain (Roe, 1972 a,b,c,d), and the physiological studies of Crustacea (includ- ing copepods) undertaken in the California Current by Childress and co-workers (Childress, 1975,1977; Thuesen et al., 1998). Several other benchmark studies and syntheses have specifically investigated or have in- cluded data on patterns of distribution and diversity in deep-sea copepods (Grice & Hulsemann, 1965; Brod- sky, 1967; Owre & Foyo, 1967; Scotto Di Carlo et al., 1984; Hirota, 1995). Patterns of biomass distribution have also been studied (Angel & Baker, 1982; Roe, 1988; Kopplemann & Weikert, 1992, 1999). In the Southern Ocean, work on deep-water cope- pods has been almost exclusively of a taxonomic nature (Vervoort, 1951, 1957; Park, 1978, 1980, 1982; Bradford & Jillet, 1980; Bradford et al., 1983; Fontaine, 1988; Bradford-Grieve, 1994), although a limited number of studies have addressed questions of diversity, diet and feeding ( Hopkins, 1985, 1987; Hopkins & Torres, 1988, 1989; Øresland & Ward, 1993). Seasonal work, which is difficult under the best of circumstances, has hardly taken place at all. It is not the aim of this paper to reveal new data and change this situation at a stroke, but rather to review what we do know of taxonomic diversity and phys- ical and production processes in the Southern Ocean, and to use this as a basis to compare with other parts of the world ocean. Diversity in any community is a
-
Upload
bournemouth -
Category
Documents
-
view
2 -
download
0
Transcript of The deep-sea copepod fauna of the Southern Ocean: patterns and processes

Hydrobiologia 453/454: 37–54, 2001.R.M. Lopes, J.W. Reid & C.E.F. Rocha (eds), Copepoda: Developments in Ecology, Biology and Systematics.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
37
The deep-sea copepod fauna of the Southern Ocean: patterns andprocesses
Peter Ward & Rachael S. ShreeveBritish Antarctic Survey, NERC, High Cross, Madingley Rd., Cambridge CB3 0ET, U.K.E-mail: [email protected]
Key words: deep-sea copepods, diversity, distribution, production, metabolism, Southern Ocean
Abstract
In recent years, much attention has been paid to the Antarctic epipelagic fauna, as a result of the desire to increaseour knowledge of ecosystem function and resource management. Unfortunately, our understanding of the polarpelagic deep-sea has not progressed as fast, and in common with many other parts of the world’s deep ocean,knowledge is still fragmentary. As yet, we have an incomplete but evolving knowledge of species presence anddistribution, but very little idea of how the extreme seasonality seen in the Southern Ocean might influence thedeep-water fauna. An examination is made of species distribution and diversity, in relation to the latitudinalcline seen in many benthic groups, and the historical perspective offered by changing circulation patterns andsea temperature through geological time. Although a number of important frontal systems are found within thecircumpolar Southern Ocean, the boundary is marked by the Sub-Tropical Convergence, which appears to bethe major biogeographic boundary between it and surrounding provinces. Evidence for seasonality in variousfamilies is reviewed in light of what we know and can infer about their biology and particularly in respect of theirbathymetric distribution, which in some groups appears to change with latitude.
Introduction
In general, little is known about the pelagic deep-sea,despite the fact that depths below 1 km comprise 62%of the surface of the earth and 79% of the volumeoccupied by life (Childress, 1995). The deep-sea hastraditionally been of more interest to taxonomists thanecologists, primarily because many of the specieshave been rarely sampled, and because of the diffi-culties and costs of obtaining sufficient material forexperimental work. There are some notable exceptionswhere deep-water sites have received detailed study,e.g. the time series undertaken in the Rockall Trough(Mauchline, 1992, 1994), the SOND cruise’s studyon the Abyssal Madeira Plain (Roe, 1972 a,b,c,d),and the physiological studies of Crustacea (includ-ing copepods) undertaken in the California Currentby Childress and co-workers (Childress, 1975,1977;Thuesen et al., 1998). Several other benchmark studiesand syntheses have specifically investigated or have in-cluded data on patterns of distribution and diversity in
deep-sea copepods (Grice & Hulsemann, 1965; Brod-sky, 1967; Owre & Foyo, 1967; Scotto Di Carlo et al.,1984; Hirota, 1995). Patterns of biomass distributionhave also been studied (Angel & Baker, 1982; Roe,1988; Kopplemann & Weikert, 1992, 1999).
In the Southern Ocean, work on deep-water cope-pods has been almost exclusively of a taxonomicnature (Vervoort, 1951, 1957; Park, 1978, 1980,1982; Bradford & Jillet, 1980; Bradford et al., 1983;Fontaine, 1988; Bradford-Grieve, 1994), although alimited number of studies have addressed questionsof diversity, diet and feeding ( Hopkins, 1985, 1987;Hopkins & Torres, 1988, 1989; Øresland & Ward,1993). Seasonal work, which is difficult under the bestof circumstances, has hardly taken place at all.
It is not the aim of this paper to reveal new data andchange this situation at a stroke, but rather to reviewwhat we do know of taxonomic diversity and phys-ical and production processes in the Southern Ocean,and to use this as a basis to compare with other partsof the world ocean. Diversity in any community is a

38
function of historical, physical and biological inter-specific interactions (Hayward, 1986). In this context,we wish to examine (1) the past historical/geologicalperspective to see how long Antarctica has been a coldocean, and what effects this may have had on speci-ation/colonisation; (2) present day patterns of diversitywith a view to seeing how Antarctica fits into the ex-tensive literature on latitudinal clines; (3) to see howpresent day circulation patterns may influence distri-bution; and (4) examine production regimes and to seewhether seasonality is prevalent or not.
In a very real sense, many of the commonly oc-curring copepods in polar waters are essentially deep-water species, which although spending a part of theirlife in surface waters during the summer, pass up to 7–8 months at depth. However, in respect of this paper,we will largely ignore these and restrict our interest tothose taxa that have exclusively deep-sea distributionsand rarely, if at all, appear in the top 200 m of thewater column.
Review
Before examining the factors which may be influen-tial in determining deep-sea diversity in the SouthernOcean, we briefly review some of the general featuresof the deep-sea environment and trends of zooplanktondistribution with depth.
Physical and biological characteristics of deep water
Continuity and lack of variation in the water columncharacterise the deep ocean (Tyler, 1988). It is tem-porally stable, largely constant in many of its physico-chemical characteristics, and, with the exception ofhydrothermal vents and some isolated areas such asthe abyssal Red Sea where temperatures are high andhorizontally homogeneous (Tyler, 1988; Childress,1995). Surface light does not penetrate below 1000 m,and oxygen concentrations, although low, particu-larly between 1000 and 1200 m, are rarely limiting.Temperature in the deep sea below the depth of thepermanent thermocline is mostly ≤ 4 ◦C, and belowapproximately 1000 m seasonal change is negligible.The only physical property that changes dramaticallywith depth is pressure. The deep ocean has been coldfor almost 40 million years and oxic for 90 millionyears (Childress, 1995). This relative stability throughrecent geological time has been generally implicatedas a major reason for high deep-sea diversity, although
it is now appreciated that there are marked compon-ents of seasonality in the deep sea in terms of physicalevents and vertical fluxes, which may also influencecommunity diversity and structure and function ( Hay-ward, 1986; Tyler, 1988). Equally, there are knownto be spatial variations in the biomass of organismsand species presence/absence, which probably reflectsurface production (Hayward, 1986).
Deep sea distribution of biomass
Over much of the world ocean, patterns of zooplank-ton distribution with depth are largely similar. At asite on the Abyssal Madeira Plain, an exponentialdecline in biomass with increasing depth was recor-ded, with more than 80% of plankton biomass and>95% of micronektonic biomass occurring in the top1000 m of a 5000 m water column (Roe, 1988). Sim-ilar patterns have repeatedly been observed at stationsin the Atlantic, Pacific and Indian Oceans and theMediterranean Sea (Wishner, 1980; Angel & Baker,1982; Scotto Di Carlo et al., 1984, 1991). Kopple-mann & Weikert (1992), re-analysed such patternsfrom 7 studies, and concluded that a power model fitsall data sets better than an exponential one and thatthere were no significant differences in the slopes ofbiomass decline with depth between studies, despitethe use of different net mesh sizes. They concludedthat this was a clear indication that material fluxeswere similar in most bathypelagic systems in theworld ocean. More recently, Kopplemann & Weikert(1999), demonstrated significant differences in slopesor y-intercepts between spring and summer profiles atdepths >1000 m at the BIOTRANS site, with summerbiomass and abundance being higher. The explanationgiven for this increase was a reproductive responserelated to food falls of phyto-detritus occurring dur-ing the spring bloom. Phyto-detritus falls do not occurin the oligotrophic waters over the Abyssal MadeiraPlain, and the summer profiles of Roe (1988) wereaccordingly found to be similar to the spring dataof Kopplemann & Weikert (1999). In the SouthernOcean, a decline of biomass with depth is clearlyseen in spring and summer Longhurst Hardy Plank-ton Recorder (LHPR) profiles taken in the top 1000 maround South Georgia Island (Fig. 1), although inwinter, biomass is more evenly distributed down thewater column (Foxton, 1956). In the top 1000 m,biomass appears to show little overall change dur-ing the course of a year (Foxton, 1956; Hopkins,1971; Atkinson, 1990), probably because of a com-
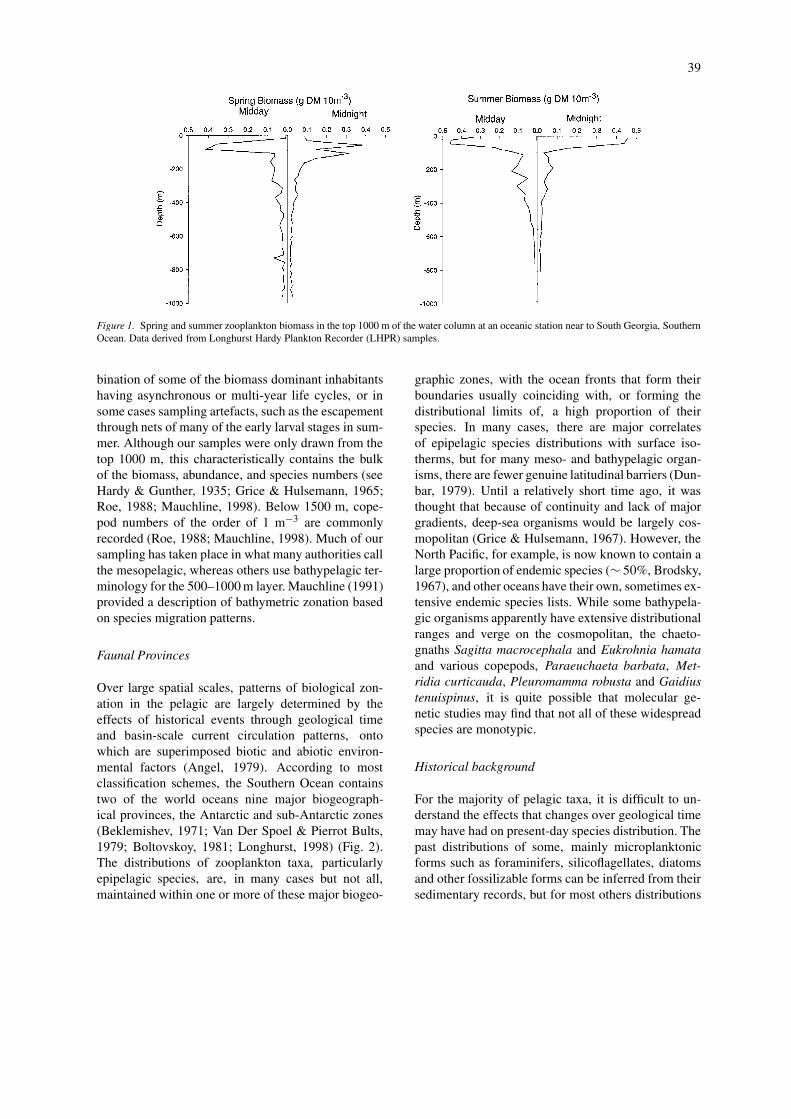
39
Figure 1. Spring and summer zooplankton biomass in the top 1000 m of the water column at an oceanic station near to South Georgia, SouthernOcean. Data derived from Longhurst Hardy Plankton Recorder (LHPR) samples.
bination of some of the biomass dominant inhabitantshaving asynchronous or multi-year life cycles, or insome cases sampling artefacts, such as the escapementthrough nets of many of the early larval stages in sum-mer. Although our samples were only drawn from thetop 1000 m, this characteristically contains the bulkof the biomass, abundance, and species numbers (seeHardy & Gunther, 1935; Grice & Hulsemann, 1965;Roe, 1988; Mauchline, 1998). Below 1500 m, cope-pod numbers of the order of 1 m−3 are commonlyrecorded (Roe, 1988; Mauchline, 1998). Much of oursampling has taken place in what many authorities callthe mesopelagic, whereas others use bathypelagic ter-minology for the 500–1000 m layer. Mauchline (1991)provided a description of bathymetric zonation basedon species migration patterns.
Faunal Provinces
Over large spatial scales, patterns of biological zon-ation in the pelagic are largely determined by theeffects of historical events through geological timeand basin-scale current circulation patterns, ontowhich are superimposed biotic and abiotic environ-mental factors (Angel, 1979). According to mostclassification schemes, the Southern Ocean containstwo of the world oceans nine major biogeograph-ical provinces, the Antarctic and sub-Antarctic zones(Beklemishev, 1971; Van Der Spoel & Pierrot Bults,1979; Boltovskoy, 1981; Longhurst, 1998) (Fig. 2).The distributions of zooplankton taxa, particularlyepipelagic species, are, in many cases but not all,maintained within one or more of these major biogeo-
graphic zones, with the ocean fronts that form theirboundaries usually coinciding with, or forming thedistributional limits of, a high proportion of theirspecies. In many cases, there are major correlatesof epipelagic species distributions with surface iso-therms, but for many meso- and bathypelagic organ-isms, there are fewer genuine latitudinal barriers (Dun-bar, 1979). Until a relatively short time ago, it wasthought that because of continuity and lack of majorgradients, deep-sea organisms would be largely cos-mopolitan (Grice & Hulsemann, 1967). However, theNorth Pacific, for example, is now known to contain alarge proportion of endemic species (∼ 50%, Brodsky,1967), and other oceans have their own, sometimes ex-tensive endemic species lists. While some bathypela-gic organisms apparently have extensive distributionalranges and verge on the cosmopolitan, the chaeto-gnaths Sagitta macrocephala and Eukrohnia hamataand various copepods, Paraeuchaeta barbata, Met-ridia curticauda, Pleuromamma robusta and Gaidiustenuispinus, it is quite possible that molecular ge-netic studies may find that not all of these widespreadspecies are monotypic.
Historical background
For the majority of pelagic taxa, it is difficult to un-derstand the effects that changes over geological timemay have had on present-day species distribution. Thepast distributions of some, mainly microplanktonicforms such as foraminifers, silicoflagellates, diatomsand other fossilizable forms can be inferred from theirsedimentary records, but for most others distributions

40
Figure 2. Marine biogeographic zones of the World Ocean. After Herman (1979).1= Arctic, 2= Subarctic, 3= N. Transitional, 4= N. Subtropical,5= Tropical, 6= S. Subtropical, 7=S. Transitional, 8=Subantarctic, 9=Antarctic.
can only be studied by looking at living assemblages(Boltovskoy, 1994). We are helped to an extent by thefact that living assemblages by and large show varyingdegrees of fidelity to present day water mass distribu-tions, and that changes in marine biogeographic his-tory are tied to paleoceanographic changes. Thus, byanalogy, surface water-mass cladograms based on thepaleoceanographic literature may be compared withfossil record and present day distributions, and infer-ences drawn about gene-flow (White, 1994). The Ant-arctic province has been in existence for many millionsof years. Characteristic assemblages are associatedwith Antarctic, sub-Antarctic and southern subtropical(temperate) regions, separated by the Antarctic PolarFront (APF) and the sub-Tropical Convergence (STC),respectively. The APF marks the divide between as-semblages dominated by siliceous forms (principallydiatoms) and calcareous assemblages (foraminifers),and biodiversity is low compared to tropical and tem-perate areas (Kennett, 1978). The glacial, climatic andpaleoceanographic development of the present South-ern Ocean took place over a long period of time,
commencing in the Tertiary. The movement of con-tinents, the opening of the Drake Passage, and theformation and later development of the Antarctic Cir-cumpolar Current (ACC) had the effect of thermallyisolating Antarctica by decoupling the warmer sub-tropical gyres from the colder Antarctic and sub-Antarctic gyres. This led to increased glaciation andthe development of ice sheets, which brought aboutfurther cooling of waters, the development of sea-ice,and meteorologically forced upwelling of nutrient richwaters. The development of the circumpolar currentalso largely removed within-Antarctic provinciality.Major events in the development of the SouthernOcean through geological time are described belowand summarised in Table 1.
Australia and Antarctica were joined 65–55 MaBP, and the continent was largely unglaciated. Aus-tralia then began to drift north, although circumpolarflow was still blocked. In the Eocene (55–38 Ma BP),the Southern Ocean was relatively warm, and Ant-arctica was still largely unglaciated. By 38 Ma BP,substantial sea-ice had begun to form, resulting in a

41
Table 1. Summary of stages in the development of the Southern Ocean through geological time.Largely based on data in Kennett (1978). Other valuable and detailed sources include Pirrie et al.(1997) and Crame (1999)
Age: Million years before Epoch Significant features during the
present (Ma BP) development of the Southern Ocean
∼ 65 Paleocene Australia and Antarctica joined as
a single continent. Tasman Sea open.
55–38 Eocene Australia moves north. Antarctic
continent unglaciated.
Sub-Antarctic sea temperature ∼19 ◦C
at beginning of epoch falls to ∼11 ◦C by
end.
∼ 38 Eocene/Oligocene Global cooling. Sea-ice forms.
Boundary Production of Antarctic bottom water
starts. Circum-antarctic flow still
restricted.
38–22 Oligocene Antarctic glaciated. Antarctic
Circumpolar Current (ACC) develops
(25–22 my BP), leading to decoupling
and thermal isolation from
surrounding seas.
22–5 Miocene Initial development of the Antarctic
Polar Front (APF). Clear differences
seen in Antarctic and sub-Antarctic
microfossil assemblages. Sub-Antarctic
surface temperature ∼10 ◦C.
6–4 Late Miocene Global cooling resulting in 300 km
northwards movement of APF.
Increase in biogenic productivity.
5–2 Pliocene Continued global cooling.
1.8 – present day Quaternary Increases in ice cover and high
biogenic productivity.
drop in water temperature of ∼5 ◦C to about present-day values. A thermohaline circulation, much liketoday’s, was initiated during this period. At the begin-ning of the Oligocene (38–22 Ma BP), the large-scaleproduction of Antarctic bottom water started and sub-Antarctic surface temperatures fell to ∼7 ◦C , similarto today, and by inference Antarctic surface water tem-peratures fell as well. Circum-antarctic flow developedwith the opening of the Drake Passage during the lateOligocene 25 Ma BP, resulting in decoupling from
other systems and increasing thermal isolation. ThePaleogene-Neogene transition (Neogene 22 Ma BP topresent) was marked by the development of distinctand permanent latitudinal belts from the tropics to thepoles. It is important to note that in preceding periods,although at times the fossil record indicates latitudinalclines, they were often less conspicuous, with specieshaving wider distributions. So it is really only at thebeginning of the Neogene that present patterns becameestablished. There were also increases in siliceous pro-

42
duction at this time, which have been linked to the de-velopment of the APF. By the middle Miocene (14–11Ma BP), the Antarctic ice cap formed, and sedimenta-tion patterns and rates began to reflect the developmentof the APF and the continuing formation of sea ice.The Antarctic has exhibited circumpolar planktonicbiogeographic communication since the middle Ceno-zoic (∼ 25–30 Ma BP). In the late Miocene (5 MaBP), ice volume increased related to global cooling,and there was an associated 300-km northwards move-ment of the APF. The Quaternary represented a periodof high biogenic productivity between the APF andthe Antarctic divergence, and on the continent therewas widespread glaciation, although no ice-cap waspresent.
So there has been relative stability over the lastfew tens of millions of years, at least in terms oflarge-scale circulation patterns, leading to the assump-tion that an equilibrium of sorts has been reached,where species presence/absence reflects major tec-tonic events which have subsequently been influencedby productive patterns.
The question thus arises of how diverse the South-ern Ocean copepod fauna is and the degree of endem-icity it exhibits.
Diversity in the Southern Ocean
Biodiversity studies are currently the focus of muchattention in the Southern Ocean (Arntz et al., 1997;Clarke & Crame, 1997). As elsewhere, they are gener-ally concerned with benthic biodiversity, with pelagicstudies not being as well addressed. Biodiversity atthe deep-sea floor has had two principal themes; firstto investigate variation in diversity with depth andlatitude, and second to examine the mechanisms main-taining diversity (Thistle, 1998). The polewards clinein species richness of terrestrial plants and animalsmay not be mirrored throughout the benthic marineenvironment (Clarke & Crame, 1997). The picturethat is emerging is that whilst some groups such asnatant decapods, and bivalve and gastropod molluscsmay exhibit this trend, other taxa, particularly inhab-itants of shallow-water soft-bottom communities, maynot, and further there are numerous and differing reas-ons for high or low species numbers across taxa (seeArntz et al., 1997 for an up-to-date review). Studiesare often hindered by a lack of sampling comparabil-ity across methods and scales, taxonomic uncertainty,and the information with which to make meaningfulcomparisons.
Are we then better able to divine general trendsin pelagic taxa and copepods in particular? Here toothere are taxonomic uncertainties and methodologicaldifficulties (sampling protocol and efforts). However,biodiversity studies provide fairly convincing evid-ence that for many pelagic taxa there is a decreasein the number of species towards the poles in bothhemispheres. Of the better-studied taxa, the latitud-inal distribution of euphausiids in the Atlantic showsa clear polewards decline in species numbers in bothhemispheres, with major changes in the south occur-ring at the STC and APF (Mackintosh, 1934; John,1936; Mauchline & Fisher, 1969). This pattern is seenrepeatedly across a whole variety of taxa in both hemi-spheres, Chaetognatha (Pierrot-Bults, 1997), Ptero-poda (Van Der Spoel & Boltovskoy, 1981), Salpa andAppendicularia (Esnal & Castro, 1977), all showinga greater number of species at lower latitudes. Thispattern is not restricted to epipelagic dwellers, butcharacterises bathypelagic species as well (ostracods,fish and decapods: see Angel, 1991). Whether thesepatterns represent smooth or steplike transitions is notalways clear. Angel (1991) showed such declines for anumber of taxa from 10–65 ◦N, which are apparentlysmooth. However, for many species frontal regionsmark the limits of their distribution and a clear steplikedistribution pattern is seen. Which pattern is observedis probably dependent on the extent to which spe-cies represent surface or deep-sea communities. In theSouthern Ocean, the STC and the APF are frontalregions that appear to act as faunal barriers for a vari-ety of taxa (John, 1936; Mackintosh, 1937; Kennett,1978), although by no means all. Atkinson & Sinclair(2000) suggested that the APF may not represent a sig-nificant barrier for all copepod species, and a few suchas Oithona similis and Ctenocalanus spp., actuallyreach peak abundances there. The same is also truefor a number of diatom species (Kennett, 1978). TheSTC appears to be the major biogeographic boundaryin the Southern Ocean, and clear transitions in spe-cies compositions can be seen for a variety of copepodtaxa (Bradford & Jillett, 1980; Bradford et al., 1983;Bradford -Grieve, 1994).
Notwithstanding the various species distributionsdescribed above, the question of whether copepoddiversity decreases with increasing latitude in thesouthern hemisphere is not easily answered.
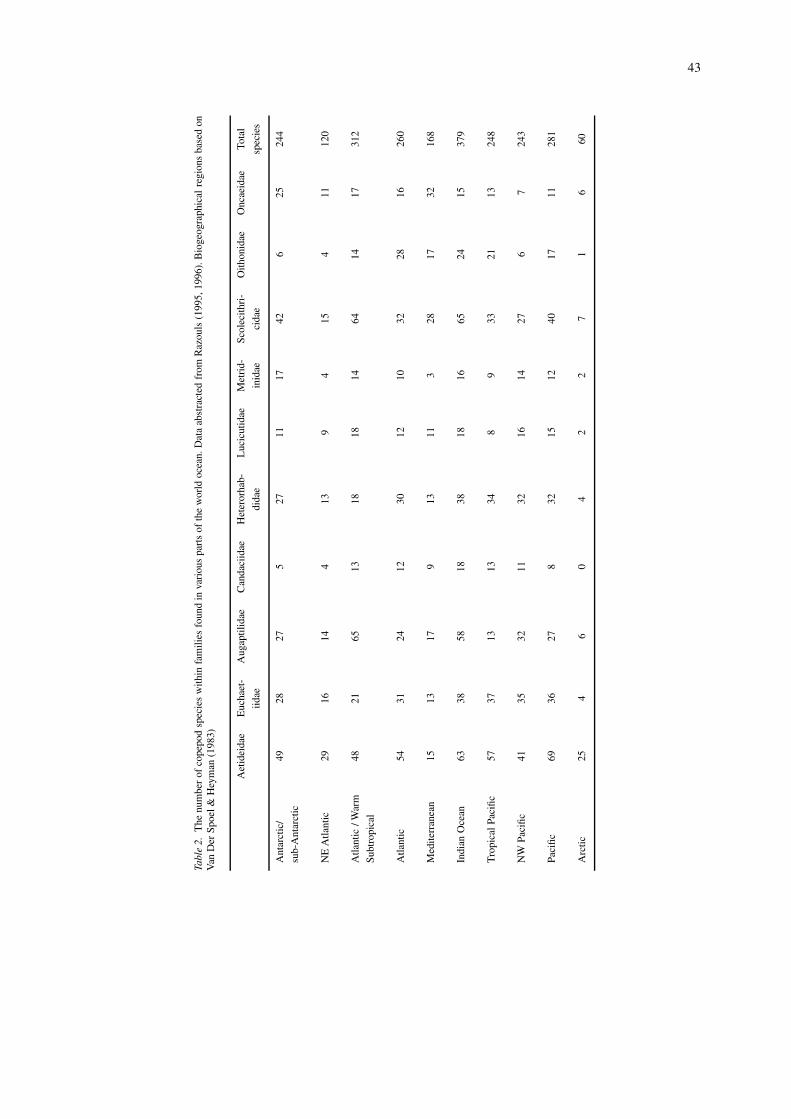
A comparison of the number of species within vari-ous copepod families occurring in different parts ofthe world ocean is made in Table 2, which has beencompiled from Razouls (1995, 1996). While some
43
Tabl
e2.
The
num
ber
ofco
pepo
dsp
ecie
sw
ithin
fam
ilies
foun
din
vari
ous
part
sof
the
wor
ldoc
ean.
Dat
aab
stra
cted
from
Raz
ouls
(199
5,19
96).
Bio
geog
raph
ical
regi
ons
base
don
Van
Der
Spoe
l&H
eym
an(1
983)
Aet
idei
dae
Euc
haet
-A
ugap
tilid
aeC
anda
ciid
aeH
eter
orha
b-L
ucic
utid
aeM
etri
d-Sc
olec
ithri
-O
ithon
idae
Onc
aeid
aeTo
tal
iidae
dida
ein
idae
cida
esp
ecie
s
Ant
arct
ic/
4928
275
2711
1742
625
244
sub-
Ant
arct
ic
NE
Atla
ntic
2916
144
139
415
411
120
Atla
ntic
/War
m48
2165
1318
1814
6414
1731
2
Subt
ropi
cal
Atla
ntic
5431
2412
3012
1032
2816
260
Med
iterr
anea
n15
1317
913
113
2817
3216
8
Indi
anO
cean
6338
5818
3818
1665
2415
379
Tro
pica
lPac
ific
5737
1313
348
933
2113
248
NW
Paci
fic41
3532
1132
1614
276
724
3
Paci
fic69
3627
832
1512
4017
1128
1
Arc
tic25
46
04
22
71
660

44
families such as the Acartiidae and the Paracalanidae,for example, are either absent or poorly representedin the Southern Ocean, they have been ignored on thebasis of their depth distribution, and the comparisonhas been restricted to those families whose membersare largely inhabitants of the deep-sea. One problemthat is immediately apparent is that, in some cases, weare comparing the entire species lists for regions ofdifferent areal extents and water mass influences. Fur-ther, it is not always possible to ascertain the degreeof taxonomic rigour that has been applied to collec-tions. Nonetheless, taking Razouls’ data at face value,diversity in the Southern Ocean broadly seems to beon a par with many of the other areas cited in Table 2.It is also difficult to discern any major differences withrespect to the degree that families are represented byspecies numbers. Some families that have receivedfuller taxonomic study appear less diverse than trop-ical and boreal counterparts (Euchaetidae) but otherssuch as the Oncaeidae and Scolecithricidae are sur-prisingly diverse. In the Southern Ocean, despite thecircumpolar distribution of the majority of species(Baker, 1954), there are some regional differencesin species composition. Some euchaetiid species areendemic to the Ross Sea (Fontaine, 1988), whereasothers such as Paralabidocera antarctica are largelyassociated with sea-ice (Tanimura et al., 1996).
The extent to which regional differences betweenthe Antarctic and sub-Antarctic in particular have beenobscured in Razouls’ treatment of the region as awhole is unclear. It would be surprising if there wereno faunal differences comparable to those seen in theepipelagic, given that in places the region extends inexcess of 30 degrees of latitude. For instance, at amore localised level in the Antarctic, Hopkins (1985)recorded a total of 64 species in 5 replicate tows from0–1000 m. He attributed this moderate diversity to amixing of cold polar, sub-Antarctic and deep waterfaunas. Around South Georgia, again in the Antarc-tic proper, 115 species have been identified in RMT1samples taken down to 2000 m (Atkinson, 1990) and100 species recorded by Hardy & Gunther (1935) fromthe same region. How representative of the Antarcticalone these modest totals are remains to be seen.
A few predominantly deep-sea families have re-ceived more detailed treatment such as the euchaetiids(Park, 1978, 1994a,b) and the related Aetideidae(Park, 1978; Markhaseva, 1998). Both families havea worldwide distribution although other regions onlyremotely share the faunistic composition of the Ant-arctic. The euchaetiids have an endemic Antarctic
species group, and of a total of 27 species known tooccur in Antarctic and sub-Antarctic waters, 12 (44%)are endemic. The Antarctic is only remotely related toother provinces, the number of species shared betweenit and any other region being uniformly <13% (Park,1994a). The Antarctic aetideiids are moderately di-verse, but reduced in number relative to the tropicaland boreal faunas of the Pacific and Atlantic. A faunaabove 500 m is virtually absent from Polar basins,and overall they too show a low level of relatednessto the other provinces, although the deepwater fauna(>500 m) shows a closer relationship to the borealAtlantic (Markhaseva, 1998). For many planktonicgroups there are stronger E-W than N-S affinities, pre-sumed to be a result of past connectivity of the world’swarm oceans (see Dunbar, 1979; Shih, 1979).
The degree of endemicity shown by other copepodtaxa is less clear, although for some it might be high.High endemism in dinoflagellates (80%), tintinnids(100%) and also in siliceous microfossil assemblagesshows that past isolation has been effective enoughto prevent any major migrations (Boltovskoy, 1994).Part of the problem of the biogeography of deep waterspecies is comparative rarity in samples, and the diffi-culty of being able to assign them with any confidenceinto restricted parts of ranges based on very few oc-currences. Recent reviews of Antarctic scolecithriciidsand scaphocalaniids by Park (1980, 1982) illustratethis problem. Based on the Eltanin collections, 5 newspecies of Scolecithricella and 5 scaphocalanids haverecently been described, which may or may not turnout to be endemic. Similarly among the Oncaeidae,samples from 5 hauls have provided descriptions of18 new species of Oncaea, 2 of Conaea, and 2 newspecies placed in a different genus (Heron, 1977). Ant-arctic benthos is also known to have a large number ofendemics (White, 1984).
The affinities shown by other taxa are interesting.Brodsky (1967) indicated that about 20% of previ-ously known forms from the North Pacific were alsoAntarctic forms, in addition to a large proportionof endemic species (50%) which show greatest af-finity with Atlantic taxa. A total of >280 speciesplus a degree of relatedness shown by many sug-gests, according to Brodsky (1967), an ancient originand active speciation. How many of these North Pa-cific/Antarctic forms have subsequently proved to bemore widespread is unclear, but several listed byBrodsky, Metridia brevicauda, Gaussia princeps andScaphocalanus subbrevicornis have also been recor-ded form the Atlantic (Roe, 1972c). This is not sur-

45
Figure 3. The Southern Ocean. After Nowlin (1991).
prising, given that the circumpolarity of the SouthernOcean potentially connects the deepwater fauna of theworld ocean. Some widespread species distributionsare achieved through deep-sea cold water connections,the chaetognaths Eukrohnia hamata and E. bathypela-gica (Van Soest, 1979), which are present in bothhemispheres through the phenomenon of tropical sub-mergence. The same could be true for the widespreadMetridia curticauda, which has been mainly found atdepths >1000 m in the Sargasso Sea, between 660 and960 m in the Atlantic (Roe, 1972c), but at lesser depths(400 and 600 m) in the Southern Ocean (Ward et al.,1995; Ward & Shreeve, 1999).
Given the above picture, we will briefly examinethe present-day physical characteristics of the South-ern Ocean to see how plankton may maintain their
distribution within it, and what effects the contrastingregimes may have on diversity.
The Southern Ocean
The Southern Ocean, south of the subtropical front,encompasses over 20% of the world ocean (Nowlin,1991). It is a region where wind stress drives the east-wards flowing ACC, which has the largest volumetransport of any current in the world ocean (mean135×106 m3 s−1) (Nowlin, 1991). Within this broadcircumpolar band are the full ocean depth frontal jetsof the APF and sub-Antarctic Front (SAF) (Cunning-ham et al., 1997), across which sinking and subductionproduces intermediate water, which spreads through-out the world ocean. Exchange of water occurs at all

46
Figure 4. Schematic section across the Southern Ocean, illustratingvertical and horizontal flow. Adapted from Dunbar (1979). APF =Antarctic Polar Front, AD = Antarctic Divergence.
depths across these zonal fronts, although deep andabyssal flow from the Southern Ocean may be locallyrestricted by bathymetry and density gradients (Now-lin, 1991). The northern edge of the ACC connectswith the Atlantic, Indian, and Pacific Oceans, and itssouthern boundary with the East Wind Drift (EWD)and the subpolar gyres of the Weddell and Ross Seas(Figs. 3 and 4). Typically surface temperatures rangefrom –1 ◦C to +3 ◦C in summer.
Given this physical situation, how are distributionsmaintained in the Southern Ocean?
Horizontal distributions
The way in which some plankton assemblages aremaintained within gyre systems can be easily envis-aged. Continuity of prevailing circulation patterns andbehavioural characteristics serve to keep organismslargely within geographical limits. In the SouthernOcean, the flow of the ACC is in a predominantly east-erly direction, although there is a northerly componentforcing water up to the APF. Seasonal migration intothe warm deep water (WDW), which has a southerlyset, serves to maintain the distribution of many spe-cies that are epipelagic during the Antarctic summer(Mackintosh, 1937). It is less clear how deep-waterplankton maintain their distribution. For some suchas Pleuromamma robusta and one or two euphausiids(Euphausia frigida, E. triacantha), energetic dailymigrations may suffice to maintain limits, but suchspecies are relatively few. For deep water planktonresident in the WDW and which have no effectivediurnal or seasonal migratory behaviour, their meansof maintaining distributional limits is not at all clear.To avoid continually being offset to the south, such
Figure 5. Distribution of the Euchaetidae with respect to depth andlatitude in the south-west Pacific. Solid lines are records from theSW Pacific, dotted lines represent likely distribution from existingrecords. STC = Sub Tropical Convergence, SAF = Sub AntarcticFront, PF = Polar Front. Adapted from Bradford et al. (1983).
animals must either move into the surface waters ordown into the Antarctic bottom water. In most cases,net sampling regimes have only sampled within theWDW down to 1000–2000 m, and so how many of thespecies present in the top 1000 m have distributionsthat extend to the bottom water is unknown. South-wards movements may account for the presence ofspecies that are supplied from a core region outsidethe Southern Ocean, but clearly not for endemics.
For many deep water copepods, the APF does notappear to form a faunal barrier (Atkinson & Sinclair,2000). Many species, including surface dwellers, havedistributions that range across both the Antarctic andsub-Antarctic. If we define the Southern Ocean asall areas lying within the West Wind Drift (WWD),then the STC becomes the most important faunisticboundary for many species. This is exemplified inBradford & Jillett (1980), Bradford et al. (1983)and Bradford-Grieve (1994), where the distributionof many members of the Scolecithricidae, Aetideidae,

47
Figure 6. Distribution of the Scolecithricidae with respect to depthand latitude in the south-west Pacific. Symbols as for Figure 5.Adapted from Bradford et al. (1983).
and Euchaetidae are coincident with the approximateposition of the STC (Figs. 5, 6 and 7). There aresome Southern Ocean species that are regionally re-stricted within gyre systems and shelf seas (Fontaine,1988), but how far these regional restrictions may ap-ply to other species is unknown, although Schulz (thisconference) has discovered the existence of new supra-benthic species in the Weddell Sea. It seems better toadopt the null hypothesis that the majority of deep wa-ter species have a circumpolar distribution until provedotherwise.
Vertical distributions
Despite the low overall abundance of the pelagic deepsea fauna, there is generally greater species diversitythan in the epipelagic, particularly at mesopelagicdepths. Despite the apparent lack of variation in the
Figure 7. Distribution of the Aetideidae with respect to depth andlatitude in the south-west Pacific. Symbols as for Fig. 5. Adaptedfrom Bradford & Jillet (1980).
environment, it appears that there is a wide degree ofniche separation between species. Unfortunately, thisis rarely described, as most deep-water surveys haveused a rather coarse sampling resolution. Araskevich(1978) studied feeding in relation to mouthpart struc-ture for a number of copepod families, and on thisbasis described vertical zonation in the western NorthPacific in terms of trophic preference. Filter-feedersdominated biomass in the top 500 m, below which,down to 1000 m, mixed omnivores and carnivoreswere present in approximately equal numbers. Below1000 m to around 4000 m, carnivores were dominantwhereas at depths >4000 m, omnivores were dom-inant. Studies with the LHPR around South Georgiain the top 1000 m during austral spring and summeroffer a partial view of niche separation and overlap.However, its small mouth area means that abund-ances of rare species are often insufficient to draw

48
any conclusions. Where they are present in sufficientnumbers, the indications are that most species havehighly structured distributions, with a wide verticalrange seen at any time of day, and broad overlapbetween family members. In common with a numberof other studies, whilst peak abundance and biomassoccurs in the top 100 m, secondary peaks can beobserved within the top 1000 m, usually between200 and 500 m (see also Grice & Hulsemann, 1965;Roe, 1972a). Such maxima can represent the day-time residence depths of diurnally migrating species(Pleuromamma robusta), or be present regardless oftime of day (Metridia curticauda, Gaidius tenuispinusand Heterorhabdus spp: see Ward et al., 1995). Mem-bers of the family Metridinidae dominated biomass inthe top 1000 m throughout the year. The dominance ofthese largely migratory forms in the mesopelagic andupper bathypelagic in many parts of the world oceanhas been noted previously by Roe (1972d), Kopple-mann & Weikert (1999) and others. Beyond theseobvious features, the pattern is one of a number ofspecies within each taxa having distinctly differentdepth distributions, although in some horizons thereis a great deal of overlap, with the suggestion thatfor many species their depth range is extensive (seealso Grice & Hulsemann, 1965). For some families,where sufficient data are available, it is clear that thereare changes in the depth horizons occupied by speciesmembers over latitudinal gradients. This is exempli-fied in Figure 5, redrawn from Bradford et al. (1983)(their Fig. 101), which clearly shows the absence of anepipelagic euchaetiid fauna south of the STC, in ad-dition to a fundamentally different bathypelagic faunasouth of the SAF, which may be related to the presenceof a seasonally migrating prey resource.
Recent work has recognised the importance of ver-tical fluxes in the deep-sea and the appearance of aseasonal signal in the deep ocean (Rowe, 1981; Honjo,1996).
Seasonality in the deep sea
Super-imposed on the physical and chemical uniform-ity of the deep-sea is the sedimentation of materialfrom the surface waters. The idea that deep-sea in-habitants are dependent on surface sedimentation forfood is an old one. Rowe (1981) defined six categoriesof food fall (see Tyler, 1988), which includes moults,faecal material, phyto-detritus and also the vertical mi-gration of zooplankton, which could be important ona seasonal basis. Fluxes of small particles that derive
from surface production are probably the most import-ant source of organic matter. Seasonal changes in ratesand types of flux have been established for a number oftime-series, indicating that there are detectable signalsin the deep-sea environment (Honjo, 1996).
Characteristics of productivity in the Southern Ocean
In the Southern Ocean, there are relatively few stud-ies on sedimentation rates. Shallow sites have bettercoverage, with sediment trapping showing higher re-coveries in summer during open water phytoplanktonblooms, and low rates under winter fast ice typical ofpolar environments (Wassmann et al., 1991; Cripps &Clarke, 1998). Ungrazed diatoms are important duringthe early part of the phytoplankton bloom, althoughzooplankton pigment signatures are present. Large mi-crophages such as krill and salps (sensu Fortier etal., 1994), may repackage efficiently and cause rapidflux (see Cadée, 1992; Cadée et al., 1992), althoughat other times the Antarctic pelagial can behave as ahighly efficient retention system, where losses are low(Smetacek et al., 1990). Wefer et al. (1988) indicatedthat major periods of flux in the Bransfield Strait werein December and January with fluxes in other monthsbeing of the order of 10–1000 times less. Fluxes inthose 2 months were close to the annual world aver-age (1.5 g m−1 d−1); although see Karl et al. (1991),who indicated a wider range in measured flux rates.There was little relationship between the presence ofice-algae and flux seen in Wefer’s investigation, andmost studies showed a clear seasonal signal of shortduration, albeit with regional variation.
We must conclude that although much of the sur-face production may be exported, via sedimentation(large diatoms are characteristic), or repackaged bylarge grazers, its brief duration and the fact that thesystem can at times be retentive, must mean that fora large part of the year food derived from this sourcemust be scarce in the deep-sea of the Southern Ocean.Major questions arising from this are whether patternsof surface production exert a significant influence onspecies composition in the deep-sea, and to what ex-tent do they give rise to seasonality in physiologicalprocesses amongst the inhabitants.
Angel (1997) and others have noted that for manypelagic taxa, marked changes in species compositionand diversity occur at around the 40th parallel in bothhemispheres (the Ortmann line, 15 ◦C surface iso-therm). The significance of this seems to be that inboth hemispheres these latitudes mark the point at

49
which there is a change from a permanently stratifiedsurface layer on the equatorwards side, to seasonalbreakdown of the thermocline on the pole-wards side(Longhurst, 1998). The production cycle in the formersystem is retentive, only weakly seasonal, and gener-ally shows a 2–3 fold increase in winter. In the latterthere is a tendency for a summer and an autumn bloomin the lower latitudes and a single bloom in the higherones. All of the main families with deep-sea represent-atives are present in the Southern Ocean, although it isdifficult to determine whether they are poorly repres-ented in relation to other parts of the world ocean (seeTable 2). There is as yet little evidence for an overallreduction in the numbers of many omnivorous spe-cies (sensu Arashkevich, 1978), such as the aetideiidsand scolecithriciids, south of the STC which might belinked with a different production regime, and it wouldappear that there is no shortage of niches for this typeof organism. Carnivorous genera such as the raptorialEuchaetidae and the piercing and sucking Heterorhab-didae and Candaciidae are reasonably well representedin south polar waters. Given the number of seasonalmigrants and the higher occupancy of mesopelagicdepths during winter (Atkinson, 1990), this can per-haps be viewed in the context of a year-round supplyfood. Dietary data for four dominant species of theEuchaetiidae indicates year-round feeding, with littledifference in amounts taken with time of year (Øres-land & Ward, 1993). Mixed feeding groups such asthe Scolecithricidae and the Aetideidae are also reas-onably well represented, although currently relativelyfew Augaptilidae have been recorded. It would appearthat the spectrum of niches available to deep-sea cope-pods in the Southern Ocean is broadly the same aselsewhere.
Growth and reproduction
Seasonality of growth and reproduction is clearly seenwithin the Antarctic epipelagic copepod fauna (seeAtkinson, 1998), and also the shallow water benthos(Clarke, 1988). The limited data available for deep-seaorganisms from other regions indicate that seasonalitymay be a characteristic of some but not all groups.Kopplemann & Weikert (1999) observed temporalchanges in zooplankton abundance below 1000 m inthe NE Atlantic, and linked this reproductive responseto flux events of the spring bloom. In the upper bathy-pelagic zone, higher summer:spring ratios of manytaxa were found, including copepods. This feature wasmost pronounced in the 1050–1650 m layers, where
the Metridinidae comprised 14% of copepod standingstock in spring but 59% in summer.
A comparison of spring and summer stage fre-quency distributions of some of the most commonlyoccurring deep water copepod species taken in LHPRsamples around South Georgia is shown in Table 3.Few differences which are readily interpretable as aseasonal response are shown by many, although thereis a decline in the proportion of males in the popula-tion from spring to summer, and a marked increase inthe proportion of females for Pleuromamma robustaand Metridia curticauda, and a clear period of springrecruitment in both Paraeuchaeta antarctica and P.biloba.
Mauchline (1991) demonstrated different patternsof growth in epi-, meso- and bathypelagic micronek-tonic crustaceans and fish occurring in the RockallTrough. Mesopelagic species had less pronouncedwinter checks to growth than epi-pelagic species, andtended to breed seasonally. Bathypelagic species, bycontrast, displayed linear growth and aseasonal breed-ing. Using data on sex ratios, spermatophore place-ments, and the incidence of egg masses among speciesof the carnivorous Euchaetiidae, Mauchline (1994) hasshown variable patterns, with a mixture of annual, bi-annual and continuous breeders being present. A sim-ilar approach for species of Paraeuchaeta taken duringsummer and winter surveys around South Georgia isshown in Table 4. Sex ratios favour females at bothtimes of year, and with the exception of Paraeuchaetaantarctica, the percentage of females with spermato-phores attached is generally <10–12%. Summer andwinter peaks in reproduction have been demonstratedfor E. antarctica in fjord locations (Ward & Robins,1987), and the high proportion of females with sper-matophores attached in both seasons is noteworthyin this respect. However, a clear resolution of thequestion of seasonality will only come about throughincreased sampling coverage.
Metabolism
The majority of the information that we have on meta-bolic processes in the deep-sea comes from groupsother than copepods. There has been general agree-ment that oxygen consumption among a diverse groupof micronektonic Southern Ocean crustaceans includ-ing ostracods, amphipods and decapods is consider-ably reduced at depth relative to the surface (Ikeda,1988; Torres et al., 1994a). This decline, exceedingthat which might be expected following appropriate

50
Table 3. Comparison of the percentage contribution of copepodite stages with respect to spring and summer for 7 species ofdeep-water copepods abundant in LHPR samples (0–1000 m) taken around South Georgia
Species Spring Summer
Copepodite stage
♂ ♀ V IV III II I ♂ ♀ V IV III II I
Paraeuchaeta antarctica 1 2 2 2 6 17 69 0 0 36 33 21 7 3
Paraeuchaeta biloba 1 46 44 90 0 0 0 0 21 11 43 11 14 0
Heterorhabdus spp. 32 24 36 4 0 5 0 22 17 31 25 6 0 0
Gaidius tenuispinus 0 7 42 52 0 0 0 0 31 35 35 0 0 0
Gaidius intermedius 0 46 36 18 0 0 0 0 64 33 3 0 0 0
Pleuromamma robusta 46 18 31 5 0 0 0 11 86 2 1 0 0 0
Metridia curticauda 31 24 34 6 4 1 0 8 72 9 9 2 0 0
Table 4. Sex ratio and reproductive characteristics of 4 commonly occurring species ofthe genus Paraeuchaeta taken from around South Georgia, during winter and summercruises
Species Female:Male % Females with % Females with No. females
spermatophores evidence of egg examined
attached sac attachment
Summer
P. antarctica 2.7 42.0 5.1 603
P. biloba 3.7 10.0 13.0 337
P. rasa 8.1 3.4 0.5 894
P. farrani 4.6 4.4 20.0 293
Winter
P. antarctica 2.5 39.0 34.6 3342
P. biloba 3.9 12.0 6.0 627
P. rasa 12.6 7.2 1.0 666
P. farrani 9.2 22.6 31.0 119
Q10 and mass adjustments, is not seen as a generaladaptation to life in the deep-sea, as many other phylafail to demonstrate it (Childress, 1995). A decline inmuscle mass and locomotory ability has also beenobserved, but only in sighted predators or prey. Alink between them appears to be a reduction in theneed to use visual information on predators or prey atsubstantial distances from an organism, resulting in alocomotory decline and a corresponding reduction inmetabolic rate (Childress, 1995). Thuesen et al. (1998)have recently demonstrated that such a decline is notseen for a range of deep-sea copepods, although therewere differences within groups which may relate tophylogeny and basic activity patterns. In terms of en-zyme activity, lactate dehydrogenase (LDH) levels (ameasure of anaerobic scope) indicated that the major-
ity of species were anaerobically poised, suggestingthat glycolytic energy was important. Categorisingspecies according to basic morphological patterns andbehaviours suggested that ‘muscular sinkers’, whichhave to swim constantly to avoid sinking, had lowerLDH levels than ‘thin-muscled floaters’, which adopta sit and wait strategy and have better developed burstescape responses.
Seasonality of metabolic processes in many spe-cies of copepods is little understood. A number ofpotential overwintering mechanisms have been elabor-ated by Torres et al. (1994b) for a variety of Antarcticmacroplankton. First, diapause and the use of storedlipid reserves, similar to the approach adopted byherbivorous copepods. Second, a drop in metabolicrate and combustion of body reserves coupled with

51
opportunistic feeding but no true diapause. Third, abusiness as usual strategy in which metabolic activityis not modulated appreciably. The second strategy wasfound to be typical of a number of species of amphi-pods and euphausiids, with combustion of body mass,opportunistic feeding and a lowered metabolic ratetogether serving as a viable overwintering strategy.Comparing crustacean metabolism from a variety ofdeepwater ecosystems, Torres et al. (1994a) concludedthat there was an absence of cold adaptation, with sim-ilar species showing comparable rates irrespective ofgeography. This is perhaps not too surprising giventhe world-wide similarity in deep ocean temperatures.The extent to which Southern Ocean deep-water cope-pods exhibit these characteristics, as with so much elseconcerning the south polar deep water fauna, awaitsfurther research.
Concluding remarks
As our knowledge of the physics of the deep oceanincreases and its importance to world climatic functionis beginning to be recognised and understood, it is vi-tally important that we give greater consideration to itsbiology. In Polar regions particularly, despite decreas-ing zooplankton concentrations with depth, a signific-ant proportion of biomass is located in the meso- andbathypelagial (Ward & Shreeve, 1999), and as suchthese communities will play an important role in en-ergy transfer and recycling. In the Greenland Sea, thefact that Paraeuchaeta populations may control pop-ulation growth of Calanus (Auel, 1999), serves wellto underline the fact that our scientific interest shouldextend considerably beyond the euphotic zone. At atime when there is an unparalleled interest in biod-iversity, efforts should be made to make maximum useof existing institute and museum collections to resolvetaxonomic problems, and to increase our knowledge ofspecies occurrence and diversity. Support should alsobe given to initiatives such as the benthic boundarythematic programme (BENBO) of the U.K.’s NaturalEnvironment Research Council, which aims to studythe role of zooplankton in deep-ocean processes.
References
Angel, M. V., 1979. Zoogeography of the Atlantic Ocean. In VanDer Spoel S. & A. C. Pierrot-Bults (eds), Zoogeography andDiversity in Plankton. Edward Arnold-London, Bunge ScientificPublishers, Utrecht: 168–190.
Angel, M. V., 1991. Variations in time and space: is biogeographyrelevant to studies of long-time scale change? J. mar. biol. Ass.U.K. 71: 191–206.
Angel, M. V., 1997. Pelagic biodiversity. In Ormond, R. F. G.J. D.Gage & M. V. Angel (eds), Marine Biodiversity, Patterns andProcesses. Cambridge Univ. Press, Cambridge: 35–68.
Angel, M. V. & A. De C. Baker, 1982. Vertical standing cropof plankton and micronekton at three stations in the north-eastAtlantic. Biol. Oceanogr. 2: 1–30.
Arashkevich, Ye. G., 1978. Some characteristics of feeding ofcopepods. Tr. Inst. Okeanol. 112: 118–125 (in Russian).
Arntz, W. E., J. Gutt & M. Klages, 1997. Antarctic marine biod-iversity: an overview. In Battaglia, B. J. Valencia & D. W. H.Walton (eds) Antarctic Communities, Species, Structure andSurvival. Cambridge Univ. Press, Cambridge: 3–14.
Atkinson, A., 1990. The ecology and distribution of zooplanktonaround the island of South Georgia, Antarctica. Ph. D. Thesis,British Antarctic Survey, Cambridge, 260 pp.
Atkinson, A., 1998. Life cycle strategies of epipelagic copepods inthe Southern Ocean. J. mar. Syst. 15: 289–311.
Atkinson, A. & J. D. Sinclair, 2000. Zonal distribution and seasonalvertical migration of copepod assemblages in the Scotia Sea. Pol.Biol. 23: 46–58.
Auel, H., 1999. The ecology of arctic deep-sea copepods(Euchaetidae and Aetideidae). Aspects of their distribution,trophodynamics and effect on the carbon flux. Berichte zurPolarforscung 319: 1–97.
Baker, A. De C., 1954. The circumpolar continuity of Antarcticplankton species. Discovery Rep. 27: 201–218
Beklemishev, C. W., 1971. Distribution of plankton as related to mi-cropaleontology. In Funnel B. M. & W. R. Reidel (eds), The Mi-cropaleontology of Oceans. Cambridge Univ. Press, Cambridge:75–87.
Boltovskoy, D., 1981. Caracteristicas biologicas del AtlanticoSudoccidental. In Boltovskoy D. (ed.), Atlas Del Zooplanc-ton del Atlantico Sudoccidental y Metodos de Trabajo conel Zooplancton Marino. Instituto Nacional de Investigación yDesarrollo Pesquero, Mar del Plata: 239–251.
Boltovskoy, D., 1994. The sedimentary record of pelagic biogeo-graphy. Prog. Oceanogr. 34: 135–160.
Bradford-Grieve, J. M., 1994. The marine fauna of New Zea-land: Pelagic calanoid Copepoda: Megacalanidae, Calanidae,Paracalanidae, Mecynoceridae, Eucalanidae, Spinocalanidae,Clausocalanidae. N. Z. Oceanogr. Inst. Mem. 102: 1–160.
Bradford, J. M., L. Haakonssen & J. B. Jillett, 1983. The mar-ine fauna of New Zealand: Pelagic calanoid copepods: Fam-ilies Euchaetidae, Phaennidae, Scolecithricidae, Diaixidae andTharybidae. N. Z. Oceanogr. Inst. Mem. 90: 1–150.
Bradford, J. M. & J. B. Jillett, 1980. The marine fauna of NewZealand: Pelagic calanoid copepods: Family Aetideidae. N. Z.Oceanogr. Inst. Mem. 86: 1–102.
Brodsky, K. A., 1950. Calanoida of the far eastern seas and polarbasin of the USSR. (English Translation 1967, Israel Programfor Scientific Translations). Jerusalem, 440 pp.
Cadée, G. C., 1992. Organic carbon in the upper layer and its sed-imentation during the ice-retreat period in the Scotia-WeddellSea, 1988. Polar Biol. 12: 253–259.
Cadée, G. C., H. González & S. B. Schnack-Schiel, 1992. Krill dietaffects faecal string settling. Polar Biol. 12: 75–80.
Childress, J. J., 1975. The respiratory rates of midwater crustaceansas a function of depth occurrence and relation to the oxygen min-imum layer off Southern California. Comp. Biochem. Physiol.50A: 787–799.

52
Childress, J. J., 1977. Effects of pressure, temperature and oxy-gen on the oxygen consumption rate of the midwater copepodGaussia princeps. Mar. Biol. 39: 19–24.
Childress, J. J., 1995. Are there physiological and biochemical ad-aptations of metabolism in deep-sea animals. Trends Ecol. Evol.10: 30–36.
Clarke, A., 1988. Seasonality in the Antarctic marine environment.Comp. Biochem. Physiol. 90B: 461–473.
Clarke, A. & J. A. Crame, 1997. Diversity, latitude and time: Pat-terns in the shallow sea. In Ormond, R. F. G. J. D. Gage, M.V. Angel (eds), Marine Biodiversity, patterns and processes.Cambridge Univ. Press, Cambridge: 122–147.
Crame, J. A., 1999. An evolutionary perspective on marine faunalconnections between southernmost South America and Antarc-tica. Scientia Marina 63, supplement 1: 1–14.
Cripps, G. C. & A. Clarke, 1998. Seasonal variation in the biochem-ical composition of particulate material collected by sedimenttraps at Signy Island, Antarctica. Polar Biol. 20: 414–423.
Cunningham, S. A., M. J. Griffiths, B. A. King & M. Brandon, 1997.Comparison of bottom tracking and profiling LADCP data in asection across the ACC at Drake Passage. International WorldOcean Circulation Experiment newsletter 26: 39–40.
Dunbar, M. J., 1979. The relation between oceans. In Van Der SpoelS. & A. C. Pierrot-Bults (eds), Zoogeography and diversity inplankton. Edward Arnold-London, Bunge Scientific Publishers,Utrecht: 112–125.
Esnal, G. B. & R. J. Castro, 1977. Distributional and biometricalstudy of appendicularians from the West South Atlantic Ocean.Hydrobiologia 56: 241–246.
Fontaine, M., 1988. Taxonomy and distribution of the antarcticaspecies group of the genus Euchaeta (Copepoda: Calanoida).In Kornicker L. S. (ed.), Biology of the Antarctic Seas XIX.American Geophysical Union, Washington. Antarct. Res. Ser.47: 27–57.
Fortier, L., J. Le Fevre & L. Legendre, 1994. Export of biogeniccarbon to fish and to the deep ocean: the rôle of large planktonicmicrophages. J. Plankton. Res. 16: 809–839.
Foxton, P., 1956. The distribution of the standing stock of zooplank-ton in the Southern Ocean. Discovery Rep. 28: 191–236.
Grice, G. D. & K. Hulsemann, 1965. Abundance, vertical distribu-tion and taxonomy of calanoid copepods at selected stations inthe northeast Atlantic. J. Zool. 146: 213–262.
Grice, G. D. & K. Hulsemann, 1967. Bathypelagic calanoid cope-pods of the western Indian Ocean. Proc. U.S. nat. Mus. 122:1–67.
Hardy, A. C. & E. R. Gunther, 1935. The plankton of the SouthGeorgia whaling grounds and adjacent waters. Discovery Rep.11: 1–435.
Hayward, T. L., 1986. Variability in production and the role of dis-turbance in two pelagic ecosystems. UNESCO Tech. Ser. mar.Sci. 49: 133–140.
Herman, Y., 1979 Plankton distributions in the past. In Van DerSpoel S. & A. C. Pierrot-Bults (eds), Zoogeography and Di-versity in Plankton. Edward Arnold-London, Bunge Scientificpublishers, Utrecht: 29–49.
Heron, G. A., 1977. Twenty-six species of Oncaeidae (Cope-poda: Cyclopoida) from the southwest Pacific-Antarctic area. InPawson D. L. (ed.), Biology of the Antarctic Seas VI, AmericanGeophysical Union, Washington. Antarct. Res. Ser. 26: 37–96.
Hirota, Y., 1995. The Kuroshio. Part III. Zooplankton. In Ansell, A.D. R. N. Gibson & M. Barnes (eds), Oceanogr. mar. biol. Ann.Rev. 33: 151–220.
Honjo, S., 1996. Flux of particles to the interior of the open oceans.In Ittekkot, V. P. Schäfer, S. Honjo & P. J. Depetris (eds), ParticleFlux in the Ocean, John Wiley, Chichester: 91–154.
Hopkins, T. L., 1971. Zooplankton standing crop in the Pacificsector of the Antarctic. Antarct. Res. Ser. 17: 347–362.
Hopkins, T. L., 1985. The zooplankton community of CrokerPassage, Antarctic Peninsula. Polar Biol. 4: 161–170.
Hopkins, T. L., 1987. Midwater food web in McMurdo Sound, RossSea, Antarctica. Mar. Biol. 96: 93–106.
Hopkins, T. L. & J. J. Torres, 1988. The zooplankton community inthe vicinity of the ice edge, western Weddell Sea. Polar Biol. 9:79–87.
Hopkins, T. L. & J. J. Torres, 1989. Midwater food web in the vicin-ity of a marginal ice zone in the western Weddell Sea. Deep-SeaRes. 36: 543–560.
Ikeda, T., 1988. Metabolism and chemical composition of crusta-ceans from the Antarctic mesopelagic zone. Deep-Sea Res. 35:1991–2002.
John, D. D., 1936. The southern species of the genus Euphausia.Discovery Rep. 14: 193–324.
Karl, D. M., B. D. Tilbrook & G. Tien, 1991. Seasonal couplingof organic matter production and particle flux in the westernBransfield Strait, Antarctica. Deep-Sea Res. 38: 1097–1126.
Kennett, J. P., 1978. The development of planktonic biogeographyin the Southern Ocean during the Cenozoic. Mar. Micropal. 3:301–345.
Kopplemann, R. & H. Weikert, 1992. Full-depth zooplankton pro-files over the deep bathyal of the NE Atlantic. Mar. Ecol. Prog.Ser. 86: 263–272.
Kopplemann, R. & H. Weikert, 1999. Temporal changes of deep-sea mesozooplankton abundance in the temperate NE Atlanticand estimates of the carbon budget. Mar. Ecol. Prog. Ser. 179:27–40.
Longhurst, A. R., 1998. Ecological geography of the sea. AcademicPress, San Diego: 398 pp.
Mackintosh, N. A., 1934. Distribution of the macroplankton in theAtlantic sector of the Antarctic. Discovery Rep. 9: 65–160.
Mackintosh, N. A., 1937. The seasonal circulation of the Antarcticmacroplankton. Discovery Rep. 16: 365–412.
Markhaseva, E. L., 1998. Geographical distribution of aetideidcopepods (Copepoda: Calanoida) in the world ocean. In Pierrot-Bults, A. C. & S. Van Der Spoel (eds), Pelagic Biogeography In-ternational Conference on Pelagic Biology II. IntergovernmentalOceanographic Commision Workshop Report 142: 250–256.
Mauchline, J., 1991. Some modern concepts in deep-sea pelagicstudies: Patterns of growth in the different horizons. In Mauch-line J. & T. Nemoto (eds), Marine Biology, its Accomplishmentand Future Prospect. Hokusen-sha, Tokyo: 107–130.
Mauchline, J., 1992. Restriction of body size spectra within speciesof deep-sea plankton. Mar. Ecol. Prog. Ser. 90: 1–8.
Mauchline, J., 1994. Seasonal variation in some population para-meters of Euchaeta species (Copepoda: Calanoida). Mar. Biol.120: 561–570.
Mauchline, J., 1998. The biology of calanoid copepods. AcademicPress, Adv. mar. Biol. 33: 1–710.
Mauchline, J. & L. R. Fisher, 1969. The biology of euphausiids.Academic Press, Adv. Mar. Biol. 7: 1–454.
Nowlin, D. W. Jr., 1991. On water mass exchange between theSouthern Ocean and the World Ocean, emphasis on the Atlanticsector. Mar. Chem. 35: 1–7.
Øresland, V. & P. Ward, 1993. Summer and winter diet of fourcarnivorous copepod species around South Georgia. Mar. Ecol.Prog. Ser. 98: 73–78.

53
Owre, H. B. & M. Foyo, 1967. Copepods of the Florida Current.Fauna Caribaea 1: 1–137.
Park, T., 1978. Calanoid copepods (Aetideidae and Euchaetidae)from antarctic and subantarctic waters. In Pawson D. L. (ed.),Biology of the Antarctic Seas VII. American Geophysical Union,Washington. Antarct. Res. Ser. 27: 91–290.
Park, T., 1980. Calanoid copepods of the genus Scolecithricellafrom Antarctic and Subantarctic waters. In Kornicker L. S. (ed.),Biology of the Antarctic Seas IX. American Geophysical Union,Washington. Antarct. Res. Ser. 31: 25–79.
Park, T., 1982. Calanoid copepods of the genus Scaphocalanusfrom Antarctic and Subantarctic waters. In Kornicker L. S. (ed.),Biology of the Antarctic Seas XI. American Geophysical Union,Washington. Antarct. Res. Ser. 34: 75–127.
Park, T., 1994a. Geographic distribution of the bathypelagic genusParaeuchaeta (Copepoda, Calanoida). In Ferrari, F. D. & B. P.Bradley (eds), Ecology and Morphology of Copepods. KluwerAcademic Publishers, Dordrecht. Dev. Hydrobiol. 102/Hydrobi-ologia 292/293: 317–332.
Park, T., 1994b. Taxonomy and distribution of the marine calanoidcopepod family Euchaetidae. Bull. Scripps Inst. Oceanogr. 29:1–203.
Pierrot-Bults, A. C., 1997. Biological diversity in oceanic macrozo-oplankton: More than counting species. In Ormond, R. F. G. J.D. Gage & M. V. Angel (eds), Marine Biodiversity, Patterns andProcesses. Cambridge University Press, Cambridge: 69–93.
Pirrie, D., J. A. Crame, J. B. Riding, A. R. Butcher & P. D. Taylor,1997. Miocene glaciomarine sedimentation in the northern Ant-arctic Peninsula region: the stratigraphy and sedimentology ofthe Hobbs Glacier Formation, James Ross Island. Geol. Mag.136: 745–762.
Razouls, C., 1995. Diversité et répartition géographique chez lescopépodes pélagiques, 1. Calanoida. Ann. Institut Oceanogr.,Paris 71: 81–404.
Razouls, C., 1996. Diversité et répartition géographique chez lescopépodes pélagiques, 2. Platycopioida, Misophrioida, Mor-monilloida, Cyclopoida, Poecilostomatoida, Siphonostomatoida,Harpacticoida, Monstrilloida. Ann. Institut Oceanogr., Paris 72:5–149.
Roe, H. S. J., 1972a. The vertical distributions and diurnal migra-tions of calanoid copepods collected on the SOND cruise, 1965.I. The total population and general discussion. J. mar. biol. Ass.U.K. 52: 277–314.
Roe, H. S. J., 1972b. The vertical distributions and diurnal migra-tions of calanoid copepods collected on the SOND cruise, 1965.II. Systematic account: Families Calanidae up to and includingthe Aetideidae. J. mar. biol. Ass. U.K. 52: 315–343.
Roe, H. S. J., 1972c. The vertical distributions and diurnal mi-grations of calanoid copepods collected on the SOND cruise,1965.III. Systematic account: Families Euchaetidae up to andincluding Metridiidae J. mar. biol. Ass. U.K. 52: 525–552.
Roe, H. S. J., 1972d. The vertical distributions and diurnal migra-tions of calanoid copepods collected on the SOND cruise, 1965.IV. Systematic account of families Lucicutiidae to Candaciidae.The relative abundance of the numerically most important gen-era. J. mar. biol. Ass. U.K. 52: 1021–1044.
Roe, H., 1988. Midwater biomass profiles over the Madeira AbyssalPlain and the contribution of copepods. In Boxshall, G. A.& H. K. Schminke (eds), Biology of Copepods. Kluwer Aca-demic Publishers, Dordrecht. Dev. Hydrobiol. 47/Hydrobiologia167/168: 169–181.
Rowe, G. T., 1981. The deep-sea ecosystem. In Longhurst A. R.(ed.), Analysis of Marine Ecosystems. Academic Press, London:235–267.
Scotto Di Carlo, B., A. Ianora, E. Fresi & J. Hure, 1984. Verticalzonation patterns for Mediterranean copepods from the surfaceto 3000 m at a fixed station in the Tyrrhenian Sea. J. PlanktonRes. 6: 1031–1056.
Scotto Di Carlo, B., A. Ianora, M. G. Mazzocchi & M. Scardi, 1991.Atlantis II Cruise: uniformity of deep copepod assemblages inthe Mediterranean Sea.. J. Plankton. Res. 13: 263–277.
Shih, C. T., 1979. East-west diversity. In Van Der Spoel S. & A.C. Pierrot-Bults (eds), Zoogeography and Diversity in Plankton.Edward Arnold-London, Bunge Scientific Publishers, Utrecht:87–102.
Smetacek, V., R. Scharek & E.-M.. Nöthig, 1990. Seasonal andregional variation in the pelagial and its relationship to thelife history cycle of krill. In Kerry K. R. & G. Hempel (eds),Antarctic Ecosystems. Ecological Change and Conservation,Springer-Verlag, Berlin & Heidelberg: 105–114.
Tanimura, A., T. Hoshiai & M. Fukuchi, 1996. The life cyclestrategy of the ice-associated copepod, Paralabidocera ant-arctica (Calanoida, Copepoda) at Syowa Station, Antarctica.Antarc. Sci. 8: 257–266.
Thistle, D., 1998. Harpacticoid copepod diversity at two physicallyreworked sites in the deep sea. Deep-Sea Res. Part II 45: 13–24.
Thuesen, E. V., C. B. Miller & J. J. Childress, 1998. Ecophysiolo-gical interpretation of oxygen consumption rates and enzymaticactivities of deep-sea copepods. Mar. Ecol. Prog. Ser. 168:95–107.
Torres, J. J., A. V. Aarset, J. Donnelly, T. L. Hopkins, T. M. Lancroft& D. G. Ainley, 1994a. Metabolism of Antarctic micronektonicCrustacea as a function of depth of occurrence and season. Mar.Ecol. Prog. Ser. 113: 207–219.
Torres, J. J., J. Donnelly, T. L. Hopkins, T. M. Lancraft, A.V. Aarset& D. G. Ainley, 1994b. Proximate composition and overwinter-ing strategies of Antarctic micronektonic Crustacea.. Mar. Ecol.Prog. Ser. 113: 221–232.
Tyler, P. A., 1988. Seasonality in the deep-sea. Oceanogr. mar. biol.Ann. Rev. 26: 227–258.
Van Der Spoel, S. & D. Boltovskoy, 1981. Pteropoda. In BoltovskoyD. (ed.), Atlas del Zooplancton del Atlantico Sudoccidentaly Metodos de Trabajo con el Zooplancton Marino. InstitutoNacional de Investigación y Desarrollo Pesquero, Mar del Plata:493–531.
Van Der Spoel, S. & P. P. Heyman, 1983. A comparative atlas ofzooplankton. biological patterns in the oceans. Springer-Verlag,Berlin: 186 pp.
Van Der Spoel, S. & A. C. Pierrot-Bults, 1979. Zoogeography ofthe Pacific Ocean. In Van Der Spoel S. & A. C. Pierrot-Bults(eds), Zoogeography and Diversity in Plankton. Edward Arnold-London, Bunge Scientific Publishers, Utrecht: 293–327.
Van Soest, R .W. M., 1979. North-South diversity. In Van Der SpoelS. & A. C. Pierrot-Bults (eds), Zoogeography and Diversity inPlankton. Edward Arnold-London, Bunge Scientific Publishers,Utrecht: 103–111.
Vervoort, W., 1951. Plankton copepods from the Atlantic sectorof the Antarctic. Verhandelingen der Koninklijke NederlandseAkademie van Wetenschappen Afdeeling Natuurkunde, Section2, 47: 1–156.
Vervoort, W., 1957. Copepods from Antarctic and sub-Antarcticplankton samples. British New Zealand Ant. Res. Exped., Ser.B 3: 1–160.
Ward, P., A. Atkinson, A. W. A. Murray, A. G. Wood, R. Willi-ams & S. A. Poulet, 1995. The summer zooplankton communityat South Georgia: biomass, vertical migration and grazing. Pol.Biol. 15: 195–208.

54
Ward, P. & D. B. Robins, 1987. The reproductive biology ofEuchaeta antarctica Giesbrecht (Copepoda: Calanoida) at SouthGeorgia. J. exp. mar. Biol. Ecol. 108: 127–145.
Ward, P. & R. S. Shreeve, 1999. The spring mesozooplankton com-munity at South Georgia: a comparison of shelf and oceanic sites.Pol. Biol. 22: 289–301.
Wassmann, P., R. Peinert & V. Smetacek, 1991. Patterns of produc-tion and sedimentation in the boreal and polar Northeast Atlantic.Polar Res. 10: 209–228.
Wefer, G., G. Fischer, D. Füetterer & R. Gersonde, 1988. Seasonalparticle flux in the Bransfield Strait, Antarctica. Deep-Sea Res.35: 891–898.
White, B. N., 1994. Vicariance biogeography of the open-oceanPacific. Prog. Oceanogr. 34: 257–282.
White, M. G., 1984. Marine benthos. In Laws R. M. (ed.), AntarcticEcology, Vol. 2. Academic Press, London: 421–461.
Wishner, K. F., 1980. The biomass of deep-sea benthopelagicplankton. Deep-Sea Res. 27: 203–216.




















