Reproductive biology of the planktonic copepod Temora stylifera
8
Marine Biology 101, 187-194 (1989) Marine Biology ~-veaag ~9e9 Reproductive biology of the planktonic copepod Temora stylifera A. lanora, B. Scotto di Carlo * and P. Mascellaro * Stazione Zoologica, Naples 80121, Italy Abstract Laboratory studies on the reproductive cycle of the plank- tonic copepod Temora stylifera, collected from the Gulf of Naples between October 1987 and March 1988, showed that females oscillated between a light and dark gonadal condi- tion. Histological preparations indicated that the dark con- dition corresponded to a ripe stage of oogenesis as com- pared to the light or unripe maturation state of primary oocytes. The number of eggs released within 24 h was strongly dependent on the reproductive status of the female at the time of incubation. Light unfed females rarely revert- ed to a dark or ripe condition and egg deposition in such cases was almost always zero. Dark females produced eggs within 24 h under all experimental food conditions, revert- ing from dark to light soon after egg deposition. Successive- ly, the length of the interclutch period was strongly depen- dent on the type of food present. A high percentage of infertile eggs was almost always present. We suggest that short-term studies, not taking into consideration the condi- tion of gonads at the time of incubation, will underestimate potential egg production rates. Females maintained for longer than three or four days produced mainly infertile eggs. After remating, females reinitiated the production of fertile eggs but egg viabilities never reached 100% hatching success, indicating poor efficiency in the fertilization of ma- ture oocytes. We hypothesize a possible cause and effect relationship between egg viabilities and seasonal fluctua- tions in population densities. Introduction Studies on reproductive cycles in planktonic copepods are fundamental for the comprehension of mechanisms regulat- ing population densities at sea. Clutch size and rate of clutch production are generally regarded as complex responses to both environmental and physiological conditions. Oscil- lations in the reproductive condition of adult females have * Died tragically at sea during a cruise been well described for freshwater diaptomids. Watras and Haney (1980), Watras (1983) and Williamson and Butler (1987) distinguished between gravid females with enlarged, darkened oviducts as compared to nongravid ones with small, transparent ducts. Dark gonads were filled with ripe gametes. Upon insemination, females reverted to the non- gravid condition by releasing fertilized oocytes that were retained in an egg sac. In the absence of males, females did not form egg sacs and released non-viable eggs directly into the surrounding water. A cycling in the reproductive condition of adult females has also been reported for a number of marine copepod species. Marshall and Orr (1952) identified seven stages of oogenesis for Calanusfmmarchicus, based on differences in size and color of oocytes and dilation of lateral channels of oviducal diverticulae. Runge (1985) used a similar criterion to distinguish four major stages of oogenesis in Calanus pacificus. The proportion of females belonging to the semi- ripe and ripe stages of oogenesis was then used to calculate in situ rates of egg production. Razouls (1974, 1975) de- scribed five maturation stages in the life cycle of adult female Ternora stylifera and four stages for Centropages typicus based on differences in the size of oocytes and oviducal diverticulae. Brief observations on the appearance of the female reproductive tract as an indication of the breeding intensity of adult females are also given by Conover (1967) for Canalus hyperboreus, Katona (1975) for Eurytemora af- finis and Moore and Sander (1976) for T. stylifera and T. turbinata. This study investigates the nature of oscillations in the reproductive condition of one of the more dominant Medi- terranean coastal copepods, Temora stylifera, to assess if differences in the physiological status of wild females deter- mined fluctuations in their reproductive potential. We also examined the implications that reproductive cycles had on the calculation of clutch size and rate of clutch production. Finally, we calculated egg viabilities and performed experi- ments on remating with the objective of obtaining informa- tion on the reproductive biology of this species.
Transcript of Reproductive biology of the planktonic copepod Temora stylifera

Marine Biology 101, 187-194 (1989) Marine Biology �9 ~ - v e a a g ~9e9
Reproductive biology of the planktonic copepod Temora stylifera
A. lanora, B. Scotto di Carlo * and P. Mascellaro *
Stazione Zoologica, Naples 80121, Italy
Abstract
Laboratory studies on the reproductive cycle of the plank- tonic copepod Temora stylifera, collected from the Gulf of Naples between October 1987 and March 1988, showed that females oscillated between a light and dark gonadal condi- tion. Histological preparations indicated that the dark con- dition corresponded to a ripe stage of oogenesis as com- pared to the light or unripe maturation state of primary oocytes. The number of eggs released within 24 h was strongly dependent on the reproductive status of the female at the time of incubation. Light unfed females rarely revert- ed to a dark or ripe condition and egg deposition in such cases was almost always zero. Dark females produced eggs within 24 h under all experimental food conditions, revert- ing from dark to light soon after egg deposition. Successive- ly, the length of the interclutch period was strongly depen- dent on the type of food present. A high percentage of infertile eggs was almost always present. We suggest that short-term studies, not taking into consideration the condi- tion of gonads at the time of incubation, will underestimate potential egg production rates. Females maintained for longer than three or four days produced mainly infertile eggs. After remating, females reinitiated the production of fertile eggs but egg viabilities never reached 100% hatching success, indicating poor efficiency in the fertilization of ma- ture oocytes. We hypothesize a possible cause and effect relationship between egg viabilities and seasonal fluctua- tions in population densities.
Introduction
Studies on reproductive cycles in planktonic copepods are fundamental for the comprehension of mechanisms regulat- ing population densities at sea. Clutch size and rate of clutch production are generally regarded as complex responses to both environmental and physiological conditions. Oscil- lations in the reproductive condition of adult females have
* Died tragically at sea during a cruise
been well described for freshwater diaptomids. Watras and Haney (1980), Watras (1983) and Williamson and Butler (1987) distinguished between gravid females with enlarged, darkened oviducts as compared to nongravid ones with small, transparent ducts. Dark gonads were filled with ripe gametes. Upon insemination, females reverted to the non- gravid condition by releasing fertilized oocytes that were retained in an egg sac. In the absence of males, females did not form egg sacs and released non-viable eggs directly into the surrounding water.
A cycling in the reproductive condition of adult females has also been reported for a number of marine copepod species. Marshall and Orr (1952) identified seven stages of oogenesis for Calanusfmmarchicus, based on differences in size and color of oocytes and dilation of lateral channels of oviducal diverticulae. Runge (1985) used a similar criterion to distinguish four major stages of oogenesis in Calanus pacificus. The proportion of females belonging to the semi- ripe and ripe stages of oogenesis was then used to calculate in situ rates of egg production. Razouls (1974, 1975) de- scribed five maturation stages in the life cycle of adult female Ternora stylifera and four stages for Centropages typicus based on differences in the size of oocytes and oviducal diverticulae. Brief observations on the appearance of the female reproductive tract as an indication of the breeding intensity of adult females are also given by Conover (1967) for Canalus hyperboreus, Katona (1975) for Eurytemora af- finis and Moore and Sander (1976) for T. stylifera and T. turbinata.
This study investigates the nature of oscillations in the reproductive condition of one of the more dominant Medi- terranean coastal copepods, Temora stylifera, to assess if differences in the physiological status of wild females deter- mined fluctuations in their reproductive potential. We also examined the implications that reproductive cycles had on the calculation of clutch size and rate of clutch production. Finally, we calculated egg viabilities and performed experi- ments on remating with the objective of obtaining informa- tion on the reproductive biology of this species.

188 A. Ianora et al.: Reproductive biology of Temora stylifera
Materials and methods
Temora stylifera specimens were collected twice or three times a week during the mornings (09.00 to 11.00 hrs) in coastal waters of the Gulf of Naples from the beginning of October 1987 to March 1988 by towing a 250/trn mesh plankton net obliquely from about 50 m to the surface. The samples were poured into 2-liter glass jars and kept in an insulated box until they reached the laboratory, usually within 1 h of collection. There, adult T. stylifera females were sorted and classified as being either in a dark, ripe reproductive condition or light, unripe condition with the aid of a dissecting microscope.
In experiments measuring egg production rates after 24 h, 10 dark and 12 light females were kept individually in 200 ml crystallizing dishes containing filtered seawater, and 11 dark and 9 light females were incubated in ambient bay water screened through a 40/zm mesh net to remove all eggs and nauplii. An additional 10 dark and 8 light females were separated and kept in containers with ambient bay water that had been enriched with the laboratory-cultured diatom Thalassiosira rotula Meunier (30 to 40/an diam.), given in excess concentrations ranging from 4.7 to 5.3 • 1 0 - 7 cells 1-1. To monitor seasonal changes in egg production rates, 4 to 11 females were incubated in ambient bay water en- riched with T. rotula on 19 sampling dates from October to March. Egg counts were performed after 24 h with an in- verted microscope. Experiments were run at ambient tem- peratures of 18~ from October to December and 14~ from January to March and on a natural window photo- period. Cannibalism under all three experimental conditions was minimal and corrected for by including crumpled mem- branes in egg counts (Dagg 1978).
Egg viability was determined for the first 3 mo by trans- ferring eggs to tissue cluster wells with a wide-mouthed pi- pette for daily observation of egg hatching rates. This meth- od was discontinued after January when we realized that non-viable eggs were much more delicate than viable ones and easily ruptured or disintegrated when transferred. We found it much more convenient to transfer females to new containers with fresh media every day since such handling did not seem to disturb our experimental copepods. Eggs were retained in original crystalliTfing dishes, precounted and checked for hatched nauplii by counting empty egg mem- branes 48 and 72 h after the female had been removed. In cases of disagreement between precount values and empty egg membranes, the precount was taken as the more refiable total production value since infertile eggs tended to disinte- grate rapidly after deposition. Only counts obtained from clutches which were not transferred to tissue cluster wells were used to calculate viability of the population with time.
In order to observe changes in clutch size and interclutch period, we monitored changes in the gross aspect of the gonadal condition of seven females and observed the fre- quency of spawning every 2 h (except for an interval of 8 to 10 h at night) for five consecutive days. Females were classi- fied as dark or light and then incubated in ,ambient bay water enriched with Thalassiosira rotula since the best repro-
ductive performance was obtained for females incubated in this medium.
In remating experiments, we kept ten females incubated individually in ambient bay water with superabundant food for several weeks and when we were certain that females were producing only infe~ile eggs or had stopped producing eggs altogether, males were added to experimental contain- ers. Daily observations were made to record the reinitiation in spawning and the presence of viable eggs. When possible, we recorded the presence of discarded spermatophores that were generally among debris on the container bottom. Such experiments were run until death of the female.
To verify the presence of fertilized eggs, we used a fluorescent dye spe~fic for cellular nuclei. Stock solutions (1 mg ml- 1) of Hoechst 33342 - 2',5'-benzirnidazole, 2'-(4- ethoxyphenyl)-5-(4-methyl-1-piperz 71nyl), trihydrochloride, trihydrate; Sigma- were prepared in distilled water and kept in the dark at 4~ according to the method described by Hinkley et al. (1986). Stock solution (5/tl) was added to 10 ml of filtered seawater containing batches of eggs. Using this method, we colored the nuclei of oocytes dissected from ripe females and eggs collected immediately after deposition. Eggs were kept in this solution for 10 to 20 rain, rinsed several times with filtered seawater and observed with a fluorescent microscope. All experiments were run in a dark- ened room. Eggs with one and two nuclei were separated and kept in tissue cluster wells for several days to observe development.
Histological preparations were made by fixing ten adult females with dark gonads and an equal number of females with transparent gonads for 48 to 60 h at 4 ~ in a mixture of 2.5% gluteraldehyde and 1% paraformaldehyde buffered to pH 7.2 with 0.2 M sodium cacodylate and 20% seawater. Copepods were rinsed for 48 h in the same solution with 0.55 M sucrose (Arnaud et al. 1978). After the dehydration series in alcohol, samples were embedded in epon and sec- tioned with a Reichert ultramicrotome.
Other data such as total copepod counts and all stages of Temora stylifera counts were obtained from biweekly sam- ples collected from 1985 to 1987 at a fixed station in the inner part of the Gulf of Naples (see Scotto di Carlo et al. 1985 for details of sampling site and data collections).
R ~
Temora stylifera was a dominant member of the late summer and autumn copepod assemblage at a fLxed station in coastal waters of the Gulf of Naples where it ranked fourth in terms of numerical abundance. The species was present the year round, constituting about 5% of total copepod numbers but exceeding 25% in periods of maximum abundances. Peak values occurred in September and October, followed by a drastic decline in population numbers in late fall. The spe- cies was very rare in winter and spring. No major year-to- year differences were recorded except for 1985 when adult female and male densities were somewhat higher (Fig. 1).

A. Ianora et al.: Reproductive biology of Temora stylifera 189
From a morphological viewpoint, the appearance of live females differed considerably. Specimens were either trans- lucent or opaque in appearance due to the presence or ab- sence of two dark bands extending from the mid-region o f the cephalosome to the last thoracic segment (Fig. 2). This dark condit ion corresponded to a ripe stage o f oogenesis as compared to the light or unripe maturation state o f primary oocytes. Four different phases o f this oogenic cycle are s h o w n in Fig. 3, T h e First o r previtellogenic phase refers to
light females containing immature oocytes with clearly visi- ble nuclei and nucleoli (Fig. 3 a). Successively, in the semi-
800-
E400 U ~ "o 0"
411
q. ,,~ ~i~ JUV. I I
I I I
I I I [ I I I 1985 "1986 1987
Fig, 1. Temora stylifera. Seasonal fluctuations in population num- ber for females, males and juveniles (CII-CV) sampled from 1985 to 1987 at a fixed coastal station in the Gall" of Naples
Fig. 2. Temora stylifera. Light, tmripe female with transparent go- nads (a) and dark, (b) ripe female with gonads containing mature oocytes ( x 53)
O
& o
O
8
0
,-~
,---2
e~ O
Ca
I
~
u., "x::~
. ~
Ca
. a =
o l
o l
t",l
o l
Cq
r
o
w.
c~ ~d
e-i
190 A. Ianora et al.: Reproductive biology of Temora stylifera
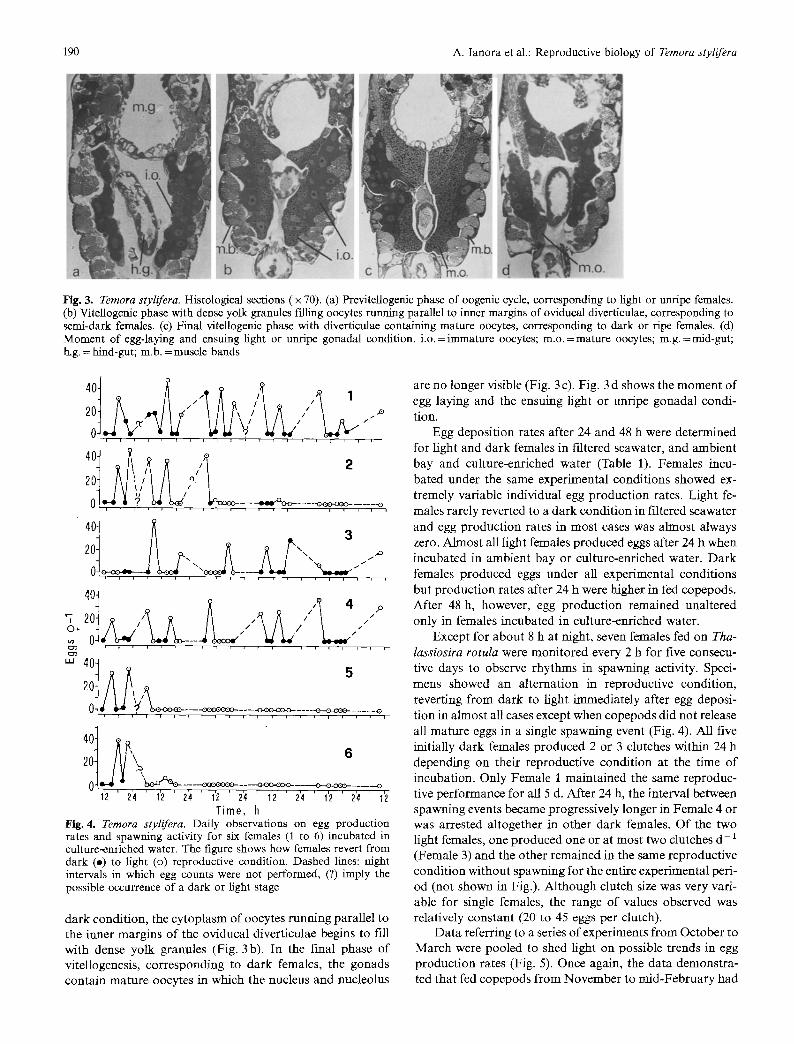
Fig. 3. Temora stylifera. Histological sections ( x 70). (a) Previtellogenic phase of oogenic cycle, corresponding to light or unripe females. (b) Vitellogenic phase with dense yolk granules filling oocytes running parallel to inner margins of oviducal diverticulae, corresponding to semi-dark females. (c) Final vitellogenic phase with diverticulae containing mature oocytes, corresponding to dark or ripe females. (d) Moment of egg-laying and ensuing light or unripe gonadal condition, i.o. =immature oocytes; m.o.=mature oocytes; m.g.=mid-gut; h.g. = hind-gut; m,b. = muscle bands
[ J , [ , r , , , , , , , , , , ( ,
o~.,,4,, ,? .~ ,~ ~ - - - ~ - - ~ - - - ~
20 . \ , ..,o
, ] r ~ , , , ~ , , - , , , , , ~ , , ,
"7 2 O+ /" k~qr
'" 401 o 5
, ,. . . . . ~ - , - , , - ~ y ~ , -:--, , ~ - ? ~ , - : - . - ~
IA; 2 ' ~ 6
0- I ,4 ~ ' ~ - % - - ~ - ~ . ~ o "1'2 ' 2.4 ' 1'2 ' 2'4 ' 1'2 ' 2'4 ' 1'2 ' 2'4 ' 1'2 ' 2'4 ' 1'2
Time, h Fig. 4. Temora sty/if era. Daily observations on egg production rates and spawning activity for six females (t to 6) incubated in culture-enriched water. The figure shows how females revert from dark (e) to light (o) reproductive condition. Dashed lines: night intervals in which egg counts were not performed, (?) imply the possible occurrence of a dark or light stage
dark condition, the cytoplasm of oocytes running parallel to the inner margins of the oviducal diverticulae begins to fill with dense yolk granules (Fig. 3b). In the final phase of vitellogenesis, corresponding to dark females, the gonads contain mature oocytes in which the nucleus and nucleolus
are no longer visible (Fig. 3 c). Fig. 3 d shows the moment of egg laying and the ensuing light or unripe gonadal condi- tion.
Egg deposition rates after 24 and 48 h were determined for light and dark females in fdtered seawater, and ambient bay and culture-enriched water (Table 1). Females incu- bated under the same experimental conditions showed ex- tremely variable individual egg production rates. Light fe- males rarely reverted to a dark condition in filtered seawater and egg production rates in most cases was almost always zero. Almost all fight females produced eggs after 24 h when incubated in ambient bay or culture-enriched water. Dark females produced eggs under all experimental conditions but production rates after 24 h were higher in fed copepods. After 48 h, however, egg production remained unaltered only in females incubated in culture-enriched water.
Except for about 8 h at night, seven females fed on Tha- lassiosira rotula were monitored every 2 h for five consecu- tive days to observe rhythms in spawning activity. Speci- mens showed an alternation in reproductive condition, reverting from dark to light immediately after egg deposi- tion in almost all cases except when copepods did not release all mature eggs in a single spawning event (Fig. 4). All five initially dark females produced 2 or 3 clutches within 24 h depending on their reproductive condition at the time of incubation. Only Female 1 maintained the same reproduc- tive performance for all 5 d. After 24 h, the interval between spawning events became progressively longer in Female 4 or was arrested altogether in other dark females. Of the two light females, one produced one or at most two clutches d - 1 (Female 3) and the other remained in the same reproductive condition without spawning for the entire experimental peri- od (not shown in Fig.). Although clutch size was very vari- able for single females, the range o f values observed was relatively constant (20 to 45 eggs per clutch).
Data referring to a series of experiments from October to March were pooled to shed light on possible trends in egg production rates (Fig. 5). Once again, the data demonstra- ted that fed copepods from November to mid-February had

A. Ianora et al.: Reproductive biology of Temora stylifera 191
140.
120.
E 4 0 .
Z
20.
0
0
0
~ o
~0 0 0 / / O~ 0
o // o ~
Y oO8 ~
0 O
B o o go
I
OCl.
0 0
o 8
0 0
8 o ~ 1 7 6 o o:y !
0 0 0 O 0 0
0
0 0 0
8 ~ 0
p.
i v NOV. DEC. JAN.
o 8 0
0 0
0
0
0 ~ pT.
0
0 0 0
0 7 f f O
o 8
O
O O i i
FEB. MAR.
Fig. 5. Temora stylifera. Relation between different food condititons and individual egg production rates after 24h (o). Mean values (+) given for unfed females (dashed line) and females fed on phyto- plankton (p) and Thalassiosira rotula-enliched food (pT) (continu- ous line) at 18~ from October to December and at 14~ from Janu- ary to March
Table 2. Ternora stylifera. Viability range (%) and mean viability .9=60 of eggs laid by wild females (N= 36) sampled in March 1988 and incubated in caflture-en_riched water for 24 h
Viability range (%) No.
0 5 20-39 3 40-59 8 60-79 4 80-99 10
100 6
Fig. 6. Temora stylifera. (a) Bright-field photogram of an egg im- mediately after deposition, and epifluorescent photogrnmR of (lo) mature oocyte dissected from a ripe female showing female nucleus prior to fertilization, (c) unfertilized egg showing only female nu- cleus, and (d) fertilized egg with both male and female pronuclei ( • 325)
higher mean egg product ion rates than starved ones in Oc- tober and February to March and that the values obtained for individuals incubated in both ambient bay and culture- enriched water were more or less the same. But, more impor- tantly, the da ta indicated that there was no par t icular sea- sonal t rend in egg product ion rates for the sampling period considered, irrespective of the different temperatures at which the copepods were maintained.
Frequency distr ibutions of percentage egg viabil i ty after 24 h are given in Table 2 for 36 ~ l d females sampled in Februa ry and March. Mean egg viabilities for eggs retained in original crystallizing dishes were generally low (60%) even though a high number of females (16) had egg viabili- ties greater than 80%. These da ta were compared to those obtained pr ior to Februa ry when eggs were transferred to tissue cluster wells. This method was later discontinued since it seemed to overestimate viabili ty (82%). Observed differ- ences, however, seemed to reflect not only experimental bias but a d imuni t ion in egg viabili ty due to the presence, in winter, of a number of females that laid non-viable eggs only.
To demonstra te that eggs had been fertilized, we used the fluorescent dye Hoechst 33342 to color the nuclei of oocytes and newly-laid eggs. The results of these experiments dem- onstrate that non-viable eggs had not been fertilized since their aspect was similar to oocytes dissected from ripe fe- males with only one visible female nucleus (Fig. 6b, c). In
192 A. Ianora et al.: Reproductive biology of Temora stylifera
300-
200"
100"
~1B.2.19SB, 4sd i
(~) 200 (~ 22.2.1988,
'~176 l,J l i,
2 0 o -
10.2. 1988, 01 d
I " - -
[ . . . . . . . . . I . . . . . . . . . [ . . . . . . 0 10 20
200"
100 100"
48d 200"
100"
t
t
. . . . . . . . . 1 ... . . . . . . I ........ " I !
v (1) 200-
I ~ 16.2.1988, 48 d
100- 21.3
I I
I . . . . . . . . . I . . . . . . 0 10
T i m e , d
|
22.2.1.988, 28 d
23.3 (I) o"
I . . . . . . . . . I . . . . . . . . . I . . . . . . . . . I 0 10 20 30
22.2.1988, 48d
m .... ,*
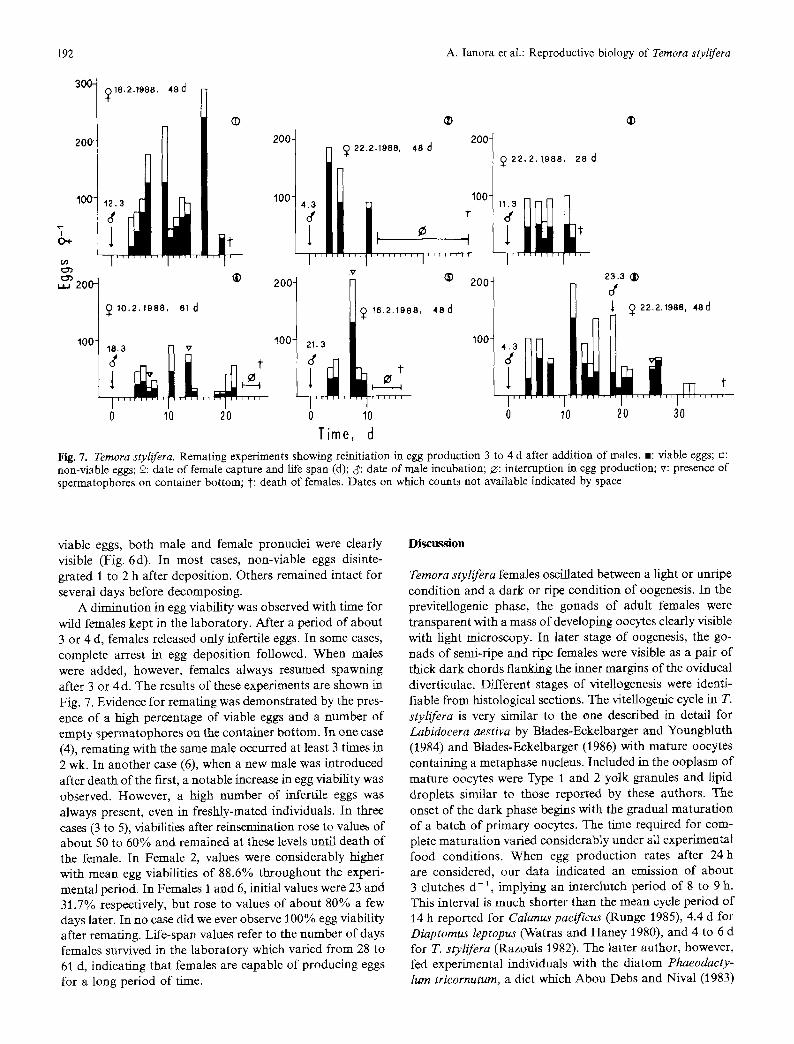
Fig. 7. Ternora stylifera. Remating experiments showing reinitiation in egg production 3 to 4 d after addition of males. -: viable eggs; n: non-viable eggs; 9: date of female capture and life span (d); c~: date of male incubation; ~: interruption in egg production; v: presence of spermatophores on container bottom; t: death of females. Dates on which counts not available indicated by space
viable eggs, both male and female pronuclei were clearly visible (Fig. 6d). In most cases, non-viable eggs disinte- grated 1 to 2 h after deposition. Others remained intact for several days before decomposing.
A diminution in egg viability was observed with time for wild females kept in the laboratory. After a period of about 3 or 4 d, females released only infertile eggs. In some cases, complete arrest in egg deposition followed. When males were added, however, females always resumed spawning after 3 or 4d. The results of these experiments are shown in Fig. 7. Evidence for remating was demonstrated by the pres- ence of a high percentage of viable eggs and a number of empty spermatophores on the container bottom. In one case (4), remating with the same male occurred at least 3 times in 2 wk. In another case (6), when a new male was introduced after death of the first, a notable increase in egg viability was observed. However, a high number of infertile eggs was always present, even in freshly-mated individuals. In three cases (3 to 5), viabilities after reinsemination rose to values of about 50 to 60% and remained at these levels until death of the female. In Female 2, values were considerably higher with mean egg viabilities of 88.6% throughout the experi- mental period. In Females 1 and 6, initial values were 23 and 31.7% respectively, but rose to values of about 80% a few days later. In no case did we ever observe 100% egg viability after remating. Life-span values refer to the number of days females survived in the laboratory which varied from 28 to 61 d, indicating that females are capable of producing eggs for a long period of time.
Olscl ls~on
Temora stylifera females oscillated between a fight or unripe condition and a dark or ripe condition of oogenesis. In the previtellogenlc phase, the gonads of adult females were transparent with a mass of developing oocytes clearly visible with light microscopy. In later stage of oogenesis, the go- nads of semi-ripe and ripe females were visible as a pair of thick dark chords flapking the inner mar~ns of the oviducal diverticulae. Different stages of vitellogenesis were identi- fiable from histological sections. The vitellogenic cycle in T. stylifera is very similar to the one described in detail for Labidocera aestiva by Blades-Eckelbarger and Youngbtuth (1984) and Blades-Eckelbarger (1986) with mature oocytes containing a metaphase nucleus. Included in the ooplasm of mature oocytes were Type 1 and 2 yolk granules and lipid droplets similar to those reported by these authors. The onset of the dark phase begins with the gradual maturation of a batch of primary oocytes. The time required for com- plete maturation varied considerably under all experimental food conditions. When egg production rates after 24 h are considered, our data indicated an emission of about 3 clutches d - t, implying an interclutch period of 8 to 9 h. This interval is much shorter than the mean cycle period of 14 h reported for Calanuspacificus (Runge 1985), 4.4 d for Diaptomus leptopus (Watras and Haney 1980), and 4 to 6 d for T. stylifera (Razouls 1982). The latter author, however, fed experimental individuals with the diatom Phaeodacty- lwn tricornutum, a diet which Abou Debs and Nival (1983)

A. Ianora et al.: Reproductive biology of Temora stylifera 193
report as being poorly suited for this species since egg pro- duction rates in such cases are similar to those obtained in starved conditions.
One of the most common methods to estimate individual rates of egg production for copepod populations is that of incubating females in natural phytoplankton assemblages for periods of about 1 d (Dagg 1978, Dagg et al. 1984, KJm- merer 1984, Runge 1985, Beckmann and Peterson 1986). However, if this method is used without considering the condition of the gonads and status of developing oocytes at the time of incubation, the results obtained will differ con- siderably. In fact, our data indicate that under starved con- ditions, light females will rarely release eggs. Dark females will produce at least one clutch after 24 h even when starved. When fed, we obtained a 3-fold difference in egg production rates after 24 h between light and dark females implying the production of about 3 clutches for dark females and only 1 clutch for light ones, with a clutch size ranging from 20 to 45 eggs. This places in question the conventional practice of counting eggs after 24 h since the method will tend to under- estimate rates of egg production by summing values ob- tained for light females that do not produce eggs with dark ones that will release eggs under most experimental condi- tions. This may be all the more important in cases of species such as Ternora stylifera with a rapid oogenic cycle where this method will result in the summation of 3 clutches. We therefore propose that a more veritable method to assess potential rates of egg production over short time intervals is obtained when egg counts for light and dark experimental females is coupled with information on the incidence of light and dark females sampled at sea in that particular period.
Remating has been demonstrated as necessary to main- tain the production of viable eggs for a number of fresh- water and marine species. Diaptomids must mate before the production of each clutch. In the absence of males, females go through their reproductive cycle releasing infertile eggs directly into the sea rather than retaining viable eggs in a sac (Watras and Haney 1980, Watras 1983, Willlamson and But- ler 1987). Heinle (1970) also reported that female Euryte- mora affinis must mate before the production of each clutch and that, if this does not occur, females will not lay eggs. Katona (1975), however, noted that isolated female E. affi- n/s produced up to five egg sacs long after fertilization even though egg sacs subsequent to the second one did not hatch. Marshall and Orr (1952) and Runge (1984) reported that a single mating was sufficient to fertilize the total eggs pro- duced by females of the species Calanus finmarchicus and C. pacificus, respectively. Wilson and Parrish (1971) and Panfish and Wilson (1978) noted that female Acartia tonsa were capable of producing viable clutches from a single insemination for several days, but remating was necessary for continued reproduction. In a number of remated fe- males, 70% of all eggs laid after 24 h were fertile. Egg viabil- ity thereafter reached values of up to 100% for about 14 d before the rate began to decline. Uye (1981) also reported that a single mating was not sufficient to maintain high egg viabilities for Acartia clausi and that remating resulted in the hatching of 100% of the eggs.
Our data also indicate that remating is necessary for continued production of viable clutches. However, they dif- fer from those reported by other authors in that egg viabill- ties after reinsemination never attained values greater than 80% and remained at these or lower levels thereafter for several days. This seems to denote a poor efficiency in the fertilization of mature oocytes for Temora stylifera. An inef- ficiency in the moment of fertilization resulting in egg viabi- lities of about 70% or lower has also been observed for Centropages typicus females maintained in the laboratory under similar experimental conditions suggesting that this may commonly occur in some copepod species that shed their eggs directly into the surrounding water (unpublished data). That such an inefficiency has not been observed be- fore may in part reflect the inadequacy of the method nor- mally used to obtain egg viability counts. The common prac- tice of transfering eggs from experimental containers tends to overestimate the percentage of viable eggs since infertile ones that degenerate shortly after deposition will not be included in final counts. That this inefficiency of fe~ilization is not a laboratory artefact may be inferred from Ambler's (1985) results on seasonal variability in egg viabilities of Acartia tonsa populations. Ambler found that egg viability varied with season, from nearly 100% in spring to 70% or lower in fall when population abundances for this species were still at an annual maximum, ff remating alone were responsible for oscillations in egg viabilities at sea, it is difficult to explain why such a diminution was observed in periods when the probability of encountering a suitable mate was still quite high. Although incomplete, our results seem to indicate that seasonal oscillations in egg viabilities also occur in T. stylifera.
Whatever the causes for the discarding of unfertilized oocytes, this phenomenon seems to be of common occur- fence at sea and may play an important role in regulating oscillations in population densities. We believe that further studies are warranted to investigate the nature of this type of behavior which, at In-st, would seem an ecological pa- radox since it implies an energetically wasteful mechanism. The fact that it occurs in dominating freshwater diaptomids (Watras and Haney 1980) and marine species such as Cala- nusfmmarchicus (Marshall and Orr 1952, 1972), C. hyper- boreus (Conover 1967), Acartia tonsa (Wilson and Parrish 1971, Ambler 1985), Centropages typicus (Ianora and Scotto di Carlo 1988) and T. stylifera (this study) implies that it may be a biologically worthwhile physiological mechanism which assures a minimum but continuous production even though this entails a less than maximum efficiency.
Acknowledgements. We thank Drs G. D. Grice and C. R. Tomas for their critical evaluation of the manuscript. Thanks are also due to G. Iamunno for the histological sections, G. Gragnaniello for his technical assistance, G. and C. Gargiulo for their photographic work, and M. Di Genova for the drawings.

194 A. Ianora et al.: Reproductive biology of Temora stylifera
Literature cited
Abou Debs, C., Nival, P. (1983). Etude de la ponte et du developp- ment embryonnaire en relation avec la temperature et la norri- ture chez Temora stylifera Dana (Copepoda: Calanoida). J. exp. mar. Biol. Ecol. 72:125-145
Ambler, J. W. (1985). Seasonal factors affecting egg production and viability of eggs ofAcartia tonsa Dana from East Lagoon, Gal- veston, Texas. Estuar. cstl Shelf Sci. 20:743-760
Arnaud, J., Brunet, M., Mazza, J. (1978). Studies on the midgut of Centropages typicus (Copepod, Calanoid). Cell Tissue Res. 187: 333-353
Beckman, B. R., Peterson, W. T. (1986). Egg production by Acartia tonsa in Long Islang Sound. J. Plankton Res. 8:917-925
Blades-Eckelbarger, P. I. (1986). Aspects of internal anatomy and reproduction in the Copepoda. Proc. 2nd int Conf. Copepoda, Ottawa. Syllogeus 58:26-50
Blades-Eckelbarger, P. I., Younbluth, M. J. (1984). The ultra- structure of oogenesis and yolk formation in Labidocera aestiva (copepoda: Calanoida). J. Morphol. 179:33-46
Conover, R. J. (1967). Reproductive cycle, early development, and fecundity in laboratory populations of the copepod Ca/anus hyperboreus. Crnstaccana 13:61-72
Dagg, M. (1978). Estimated,/n sltu, rates of egg production for the copepod Centropages typicus (Kroyer) in the New York Bight. J. exp. mar. Biol. Ecol. 34:183-196
Dagg, M. J., Clarke, M. E., Nishiyama, T., Smith, S. L. (1984). Production and standing stock of copepod nauplii, food items for larvae of the walleye pollock Theregra cha/cogramma in the southern Bering Sea. Mar. Ecol. Progr. Ser. 19:7-16
Heinle, D. R. (1970). Population dynamics of exploited cultures of calanoid copepods. Helgoland. wiss. Meeresunters. 20:360 372
Hinkley, R. E., Wright, B. D., Lynn, J. W. (1986). Rapid visual detection of sperm-egg fusion using the DNA-specific fluoro- chrome Hocchst 33342. Dev. Biol. 118:148-154
Ianora, A., Scotto di Carlo, B. (1988). Observations O n egg produc- tion rates and seasonal changes in the internal morphology of mediterranean populations of Acartia tonsa and Centropages typicus. In: Boxshall G. A., Schminke H. K. (eds.). Biology of Copepods. Hydrobiologia 167/168:247-253
Katona, S. K. (1975). Copulation in the copepod Eurytemora afJ'mis (Poppe, 1880). Crustaceana 28:89-95
gimmerer, W. J. (1984). Spatial and temporal variability in egg production rates of the calanoid copepod Acroca/anus inermis. Mar. Biol. 78:165-169
Marshall, S. M., Orr, A. P. (1952). On the biology of Ca/anus finmarchicus. VII: factors affecting egg production. J. Mar. biol. Ass. U.K. 30:527-547
Marshall, S. M., Orr, A. P (1972). The Biology of a Marine Cope- pod. Springer Verlag, Berlin
Moore, E., Sander, F. (I976). Quantitative and qualitative aspects of the zooplankton and breeding patterns of Copepods at two Caribbean Coral Reef stations. Estuar. cstl Mar. Sci. 4:589-607
Parrish, K. K., Wilson, D. F. (1978). Fecundity studies on Acartia tonsa (Copepoda: Calanoida) in standardize culture. Mar. Biol. 46:65-81
Razouls, S. (1974). Maturit6 sexuelle et frcondit6 chez les femelles de Temora stylifera, coprpode prlagique (Copepoda Cala- noidea). Archs Zool. exp. g~n. 115:387-399
Razouls, S. (1975). Fbconditr, maturit6 sexuelle et diffrrenciation de l'appareil grnital des femelles de deux Cop~odes planctoni- ques: Centropages typicus et Ternora stylifera. Pubbl. Staz. zool. Napoli 39:297-306
Razouls, S. (1982). Etude experimental de la ponte de deux cop& podes prlagiques Temora stylifera et Centropages typicus. II. Dynamique des pontes. Vie Milieu 32:11-20
Runge, J. A. (1984). Egg production of the marine, planktonic copepod, Ca/anus pac/ficus Brodsky: laboratory observations. J. exp. mar. Biol. Ecol. 74:125-145
Runge, J. A. (1985). Relationship of egg production of Ca~anus pac/ficus to seasonal changes in phytoplankton availability in Puget Sound, Washington. Limnol. Oceanogr. 30:382-396
Scotto di Carlo, B., Tomas, C. R., Ianora, A., Marino, D., Mazzoc- chi, M. G., Modigh, M., Montresor, M., Petfillo, L., Ribera d'AlcalA, M., Saggiomo, V., Zingone, A. (1985). Uno studio integrato dell'ecosistema pelagico costiero del Golfo di Napoli. Nova Thalassia 7:99-128
Uye, S.-I. (1981). Fecundity studies of neritic calanoid copepods Acartia clausi Giesbrecht and A. steueri Smirnov: a simple em- pirical model of daily egg production. J. exp. mar. Biol. Ecol. 50: 255-271
Watras, C. J. (1983). Reproductive cycles in diaptomid Copepods: effects of temperature, photocycle, and species on reproductive potential. Can. J. Fish. aquat. Sciences 40:1607-1613
Watras, C. J., Haney, J. F. (1980). Oscillations in the reproductive co~adition of Diaptomus leptopus (Copepoda: Calanoida) and their relation to rates of egg-clutch production. Oecologia 45: 94-103
Williamson, C. E., Butler, N. M. (1987). Temperature, food and mate limitation of copepod reproductive rates: separating the effects of multiple hypotheses. J. Plankton Res. 9:821-836
Wilson, D. F., Parrish, K. K. (1971). Remating in a planktonic marine calanoid copepod. Mar. Biol. 9:202-204
Date of final manuscript acceptance: December 23, 1988. Communicated by M. Sara, Genova