
Effects of carotenoid availability during laying on reproduction in the blue tit

Dietary effects on carotenoidcomposition in the marineharpacticoid copepod Nitokra lacustris
ADELAIDE C. E. RHODES*
NORTHWEST FISHERIES SCIENCE CENTER, NATIONAL OCEANIC AND ATMOSPHERIC ADMINISTRATION, 2725 MONTLAKE BLVD. E, SEATTLE,WASHINGTON 98112, USA
*CORRESPONDING AUTHOR: [email protected]
Received April 13, 2006; accepted in principle September 12, 2006; accepted for publication October 27, 2006; published online December 5,2006
Communicating editor: K.J. Flynn
Nitokra lacustris, a euryhaline harpacticoid copepod, was reared on two diets, which varied in
composition and relative quantities of carotenoid pigments, and analyzed for differences in tissue
carotenoid composition. Carotenoid compositions were analyzed by means of reverse-phase high
performance liquid chromatography (RP-HPLC). Total amounts of unesterified astaxanthin were
quantified in the two populations by means of an isocratic HPLC procedure. Carotenoid
composition and total astaxanthin content for adult copepods differed between the two diets.
Copepod populations fed the live microalga Tetraselmis suesica averaged 696 mg carotenoids
(g dry weight)21 and 185 mg astaxanthin (g dry weight)21 while copepod populations fed a
formulated diet with high levels of lycopene, beta-carotene and alpha-carotene averaged 24.3 mg
carotenoids (g dry weight)21 and 650 mg astaxanthin (g dry weight)21. Neither diet contained
canthaxanthin or astaxanthin, suggesting that the copepods were synthesizing the astaxanthin from
precursor pigments. The absence of canthaxanthin and echinenone coupled with the presence of
other intermediary carotenoid pigments suggests that the carotenoid conversion pathway of N. lacus-
tris differs from that described for calanoid copepods and other marine crustaceans. N. lacustris
may utilize the b-carotene! zeaxanthin! b-doradexanthin (adonixanthin)! astaxanthin
pathway of conversion rather than the b-carotene! echninenone! canthaxanthin! astax-
anthin pathway proposed for other copepods. Thus, diets containing higher amounts of b-carotene
and zeaxanthin would be more likely to produce high levels of astaxanthin in this species.
I N T RO D U C T I O N
Animals which are not able to biosynthesize the keto-carotenoids (e.g. canthaxanthin and astaxanthin) fromother carotenoids must acquire them directly from theirdiets. Flamingoes are an example of this phenomenon,because they will lose their pink coloration if theybecome deficient in the ketocarotenoids obtained fromthe small crustaceans they consume (Goodwin, 1984).Marine crustaceans, like other animals, must convertdietary sources of carotenoids, usually b-carotene, intothe ketocarotenoids astaxanthin and canthaxanthin(Paanakker and Hallegraef, 1978). The ability of marinecrustaceans to perform this bioconversion is essential to
the marine food web, as many fish species cannotconvert dietary carotenoids to ketocarotenoids and mustthus accumulate astaxanthin directly from their diets(Katayama et al., 1972; Torrissen and Christiansen, 1995).
Nitokra lacustris has a striking reddish-orange color-ation which can be observed easily through a micro-scope, suggesting that it contains the ketocarotenoidsastaxanthin and/or canthaxanthin that are found inother crustaceans and copepods. This study consideredthe differences in the overall pigment composition andquantity of astaxanthin present in the harpacticoidcopepod N. lacustris fed a natural algal diet versus aformulated diet composed of flax seed oil, yeast,vitamin C, vitamin B complex and vegetable juice.
doi:10.1093/plankt/fbl068, available online at www.plankt.oxfordjournals.org
# The Author 2006. Published by Oxford University Press. All rights reserved. For permissions, please email: [email protected]
JOURNAL OF PLANKTON RESEARCH j VOLUME 29 j SUPPLEMENT 1 j PAGES i73–i83 j 2007 by guest on January 11, 2014
http://plankt.oxfordjournals.org/D
ownloaded from

Neither diet contained astaxanthin nor canthaxanthin,but both contained a variety of precursor carotenoidpigments that could be converted to astaxanthin bycrustaceans. This study utilized reverse-phase highperformance liquid chromatography (RP-HPLC) toconduct the qualitative analyses and an isocratic HPLCprocedure to conduct the quantitative analyses of thepigments contained in N. lacustris copepods fed twodifferent diets.
Role of carotenoid pigments inharpacticoid copepods
Carotenoids are isoprenoid tetraterpenes containing 40carbon atoms that are synthesized by plants, algae andcertain type of bacteria and fungi (Britton, 1995).Copepods and other marine crustaceans may utilizeastaxanthin and other carotenoid pigments as protec-tants against photooxidation and as vitamin A precur-sors. Astaxanthin, the most commonly detectedcarotenoid in crustacean organs, has been associatedwith reproductive and visual physiology as well as thestabilization of membranes and proteins (Goodwin,1984; Linan-Cabello et al., 2002). For marine planktoniccopepods that can escape the photic zone, astaxanthinmay act as an antioxidant for the protection of unsatu-rated storage lipids (Juhl et al., 1996; Lotocka et al.,2004). It has been hypothesized that the rapid oxidationof carotenoids decreases the availability of free radicalsto react with unsaturated fatty acid molecules and con-sequently prevent damage to membranes (Woodallet al., 1997; Lotocka et al., 2004).Astaxanthin cannot be synthesized de novo in cope-
pods (Matsuno, 1989; Andersson et al., 2003). Severalpathways of astaxanthin synthesis from precursorcarotenoids have been suggested for crustaceans(Bandaranayake and Gentien, 1982; Goodwin, 1984;Berticat et al., 2000; Linan-Cabello, 2002) and fish(Katayama et al., 1970; Hata and Hata, 1972; Hsu et al.,1972). Katayama et al. (1973) classified aquatic animalsinto three categories based upon astaxanthin biosyn-thetic capability: (i) fish that cannot biosynthesize astax-anthin from b-carotene, lutein or zeaxanthin, but whichcan transfer dietary astaxanthin to body astaxanthin,(ii) fish, such as red carp and goldfish, that can biocon-vert astaxanthin from lutein or zeaxanthin, but notfrom b-carotene, and (iii) crustaceans (e.g. prawns) thatcan bioconvert b-carotene into astaxanthin.The metabolic pathway for the conversion of
b-carotene to astaxanthin that has been proposed formost crustaceans relies on the intermediates echinenoneand canthaxanthin (Goodwin, 1984). Canthaxanthinhas been identified in the tropical reef copepod Euchaeta
russelli (Bandaranayake and Gentien, 1982) and thecalanoid copepods Acartia bifilosa (Lotocka andStyczynska-Jurewicz, 2001) and Pseudocalanus acuspes
Giesbrecht (Lotocka et al., 2004). Canthaxanthin andthe intermediate 40-hydroxyechinenone have beenreported in the cyclopoid copepod Cyclops kolensis
Lilljeborg (Czeczuga et al., 2000). However, the inter-mediates canthaxanthin and echinenone have not beenobserved in the harpacticoid copepods Tigriopus brevicor-nis (Davenport et al., 2004) or Canuella perplexa
(Buffan-Dubau et al., 1996). The analysis of pigmentsundertaken in this study allows for further examinationof the pigment composition and potential pathways uti-lized by N. lacustris for biosynthesis of astaxanthin fromdietary precursors.
M E T H O D
Copepod production
N. lacustris (originally collected from a dockside planktontow at the Gulf Coast Marine Lab in Panacea, FL,USA) has been reared in the laboratory for severalyears on two diets: (i) Tetraselmis suecica or (ii) a formu-lated feed containing the carotenoid pigments lycopene,a-carotene, b-carotene, lutein, phytoene and phyto-fluene. The copepod cultures are non-axenic, but areperiodically treated with a sodium hypochlorite solution(6.7 mM NaOCl) to remove protozoans and bacteria.The formulated diet has been successfully used to rearN. lacustris in captivity for extended periods, and no sig-nificant differences in population growth rates, size ofanimals, or fatty acid composition have been documen-ted between the populations fed the two different diets(Rhodes, 2003; Rhodes and Boyd, 2005).
The prasinophyte Tetraselmis suecica contains thepigments trans-neoxanthin, violaxanthin, lutein, chloro-phyll b, chlorophyll a and a-carotene as identified byEgeland et al. (1995) and Bustillos-Guzman et al. (2002).The carotenoid content of the formulated feed wasassumed to be composed mainly of the pigments foundin a tomato-based vegetable juice (V-8 juice, Campbell’sSoup, Ohio). The juice blend of tomatoes, beets, celery,carrots, lettuce, parsley, watercress and spinach results ina formulated feed which contains lycopene (87.8%w/w), a-carotene (2.7%), b-carotene (7.6%), lutein(1.5%) and zeaxanthin (0.4%) (Tonucci et al., 1995;Arab et al., 2002).
Copepods were reared in six 10 L trays under identi-cal environmental conditions (salinity 27, 208C, 12-hlight:12-h dark (General Electric Cool White No.F15T8-CW). Three trays were fed the algae and three
JOURNAL OF PLANKTON RESEARCH j VOLUME 29 j SUPPLEMENT 1 j PAGES i73–i83 j 2007
i74
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from

were fed the formulated feed ad libitum. The algaecontinued to grow throughout the experiment, and theformulated feed remained suspended with the assistanceof aeration. The cultures were reared for 10 days andterminated by harvesting all adult copepods through a105 mm mesh and starving for 24 h in fresh seawater toallow them to void enteric contents. Duplicate samplesfor each replicate were rinsed with sterile deionizedwater to remove excess salt before filtration onto aglass fiber filter (Fisherbrand GF/C, Fisher Scientific,Pittsburgh, PA). As the copepods were killed by the fil-tration process, all procedures from this point forwardwere conducted under yellow fluorescent light (PhilipsF40GO) to prevent pigment destruction.
Astaxanthin quantification by isocraticHPLC method
The difference in the quantity of astaxanthin betweencopepod populations fed the two diets was measuredusing an isocratic HPLC procedure on each of thesix populations. Duplicate samples of �5000 copepodsfrom each of the six populations were pooled, filteredand lyophilized at 2708C for 36 h (Model No. 6201–3218; The Virtis Company, Inc., Gardiner, NY).Lyophilized samples were extracted in 10 mL of 100%HPLC-grade acetone in a glass tissue homogenizer tubekept on ice, flushed with nitrogen and set aside for0.5 h in the dark at 2208C. Samples were centrifugedfor 5 min at 200 g (Marathon Micro A; Fisherbrand).All supernatant was collected and the pellet wasextracted two more times. Collected supernatant wasevaporated under a nitrogen flush in the dark, weighedand reconstituted in the injection solvent [60% tetra-hydrofuran (THF), 40% methanol].Samples were injected with a Waters U6K loop injec-
tor (Waters, Massachusetts) into a 3.5 mm WatersSymmetry C18 (4.6 � 150 mm) column with a flowrate of 1.4 mL/min controlled by a Waters Model 510pump. The isocratic mobile phase was made up ofacetonitrile-methanol-chloroform (100:20:5, v/v/v) andthe ultraviolet visible detector (Spectroflow 757; KratosInstruments, New York) was set at 474 nm. Astaxanthinpeaks were identified by comparison to an astaxanthinstandard (Sigma A9335). The standard was preparedunder yellow fluorescent lighting and injected on thesame day as the samples. All samples were run on thesame day.Astaxanthin was quantified using an external stan-
dard curve plotting astaxanthin peak area versusmg mL21 of astaxanthin injected into the HPLC. Peakareas for each the cis- and trans-astaxanthin isomerswere compared to a standard curve developed using an
astaxanthin standard prepared from crystallizedastaxanthin (Sigma A 9335). The standard curve wasprepared by serial dilution of the stock solution [500 mgastaxanthin (mL solvent)21] to final concentrations of100, 50, 25, 10 and 5 mg astaxanthin (mL solvent)21.All sample concentrations fell within this standard curve(R2 ¼ 99%). Areas under the peaks that correlated tocis-astaxanthin and trans-astaxanthin were compared tothe standard curve to determine the concentration ofastaxanthin in mg (mL solvent)21. Results were con-verted into mg astaxanthin (g dry weight)21 in order tofacilitate comparison. Astaxanthin values from theduplicate samples were averaged for each of the threereplicates in each treatment. Concentrations ofcis-astaxanthin, trans-astaxanthin and total astaxanthinwere compared within and between the three popu-lations assigned to each treatment by analysis ofvariance (ANOVA).
Carotenoid separation by RP-HPLC
To analyze the complete carotenoid composition of thecopepods with RP-HPLC, a representative sample ofapproximately five thousand N. lacustris adults eachfrom a culture fed live Tetraselmis and a culture fed theformulated feed were collected and filtered in the samemanner as described previously. Pigments were col-lected from freshly harvested samples by placing thefilters containing the animals in 5 mL THF and sonicat-ing for 5 min. After centrifugation at 200 g for 5 min at48C, the supernatant was drawn off. One mL of super-natant was evaporated under nitrogen flush in thedark and reconstituted in a 200 mL mixture of 1:4 ethylacetate:ethanol for RP-HPLC injection.
Peaks identities from the RP-HPLC analysis wereanalyzed with the assistance of Craft Technologies(Wilson, NC, USA). In brief, this technique involvedRP-HPLC on a 5 mm C30 Column (YMC carotenoidcolumn 25 cm � 4.6 mm, 5 mm diameter; YMC,Wilmington, NC, USA) using HPLC-grade solventsand a linear tertiary gradient system (100% isopropylalcohol:1 N Methanol with 0.15 mM ammoniumacetate:100 % THF) at a flow rate of 1 mL/min.Pigments were quantified at 450 nm and spectrallycharacterized on a Waters 991 M photodiode arraydetector system (Waters; Milford, MA) which recordedspectra at 5 nm intervals (Millenium Software) at a rateof 300 spectra per minute. Data were downloaded andanalyzed using Microsoft Excel 97 software (Microsoft;Redmond, WA).
Peaks were identified at Craft Technologies by com-parison to the spectra and retention times under thesame conditions of a standard mix containing lutein,
A. C. E. RHODES j DIETARY EFFECTS ON COPEPOD CAROTENOIDS
i75
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from

lycopene, b-cryptoxanthin, a-carotene and b-carotene,b-carotene isomers (Hoffman-LaRoche Co. and SigmaChemical Co.), as well as a control mixture preparedfrom homogenized sea cucumber. The sea cucumbercontrol had identifiable peaks for b-carotene, cantha-xanthin, astaxanthin, zeaxanthin, phoenicoxanthin(adonirubin) and echinenone. Peak identifications wereconfirmed by comparison to published absorptionspectra from various sources (Nelis and De Leenheer,1988; Canjura, 1990; Britton, 1995; Matsuno andTsushima, 1995; Yuan and Chen, 1997; Hyvarinenand Hynninen, 1999; Berticat et al., 2000; Ston andKosakowska, 2002). Relative amounts of carotenoids werequantified by dividing peak area at 450 nm by the relativeresponse factors for each type of carotenoid (Britton,1995). The response factors were provided by CraftTechnology for their system and controls at 450 nm.
R E S U LT S
Qualitative carotenoid analysisby RP-HPLC
The separation peaks at 470 nm for copepods fed thelive alga Tetraselmis (Fig. 1) differed from the separationpeaks at 470 nm for copepods fed the formulated feed(Fig. 2). Both copepod populations contained astax-anthin as well as several carotenoid and chlorophyllpigments (Table I, Figs. 1 and 2). Copepod populationsfed the two diets had pigment compositions that
reflected their diets (Table I). Both populations ofcopepods contained unesterified astaxanthin, antherax-anthin, b-carotene, a ketocarotenoid peak characteristicof b-doradexanthin and chlorophylls a and b.Copepods fed the live alga contained the unique caro-tenoid pigments violaxanthin, lutein, zeaxanthin, anda-carotene (Fig. 1). The copepods fed the live microal-gae also exhibited significant amounts of threeunknown carotenoids (peaks 1, 4 and 7) as well asastaxanthin esters (four peaks labeled 13). Only cope-pods fed the formulated diet contained the pigments1,1-carotene, lycopene and cis-lycopene as well as twounique unidentified carotenoids (peaks 5 and 6) (Fig. 2).
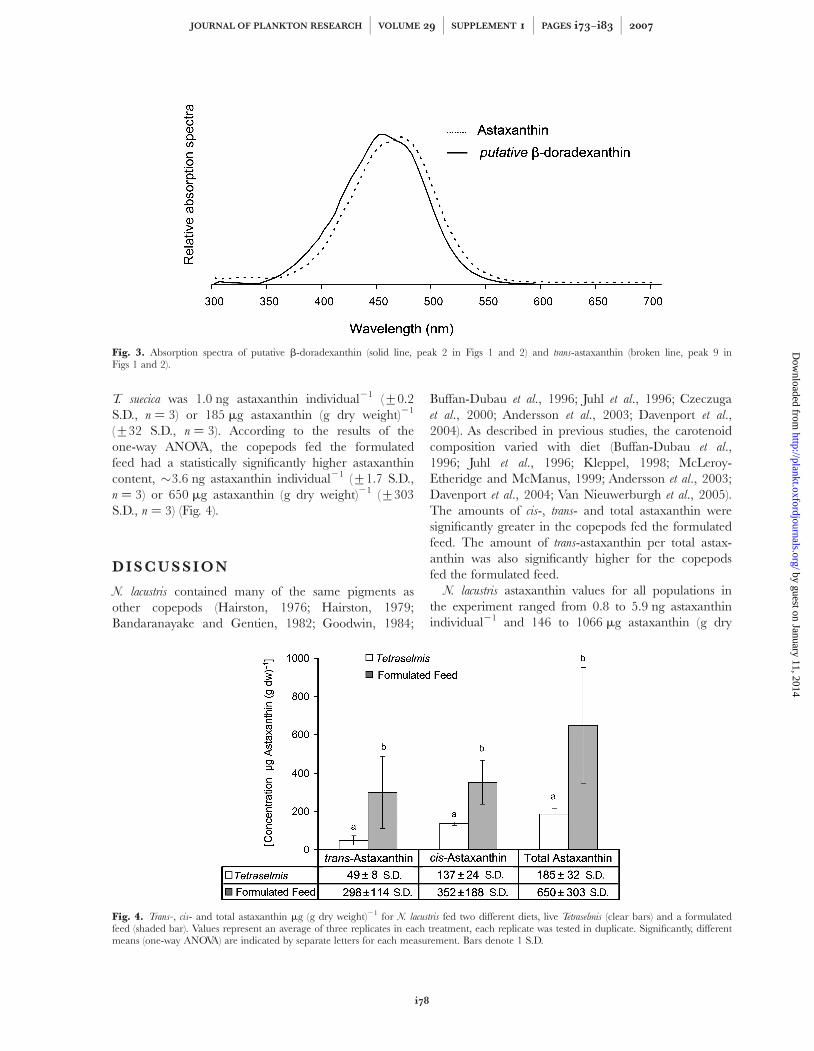
A single peak was identified as unesterified astax-anthin in both sets of animals based on its retentiontime and absorption spectra (Figs 1 and 2). Two distinctspectra corresponding to cis- and trans-astaxanthincoeluted in this single peak during the RP-HPLCmethod (peaks 8 and 9). The isocratic HPLC methodwas able to separate these two peaks and it was possibleto determine their relative concentrations. Other peakswere found with similar absorption spectra, whichcorrespond to astaxanthin esters and isomers (Yuanand Chen, 1998). Canthaxanthin or echinenone werenot detected, based on retention time, absorptionspectra, and comparison to the controls. Based onretention time and the absorption spectra, peak 2 wasputatively identified as the intermediate compoundb-doradexanthin (adonixanthin) found when zeaxanthinis converted to astaxanthin in the crayfish Astacus lepto-
dactylus (Berticat et al., 2000) (Fig. 3).
Fig. 1. Retention time and absorbance at 470 nm for copepods fed the live alga T. suecica. All peak identifications are provided in Table I. Peaksidentified as carotenoids are numbered. Peaks that represent the same compounds are given the same number on all figures. Unlabeled peaksare chlorophyll-related peaks.
JOURNAL OF PLANKTON RESEARCH j VOLUME 29 j SUPPLEMENT 1 j PAGES i73–i83 j 2007
i76
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from

During preparation of the formulated feed, 236 mLof V-8 juice, which contains 17 mg lycopene per serving(Arab et al., 2002; Campbell’s Soup, Ohio) were dilutedinto 1500 mL total volume. The copepods were fed onemL (L culture media)21 of this dilute solution once,corresponding to 11 mg lycopene (L culture media)21
available to the copepods. About 2000 adult copepodswere harvested from each liter of media; hence, about5.5 ng lycopene was available to each copepod over theentire 10 days. Based on the relative amount of lycopeneto astaxanthin determined from the RP-HPLC analysis(Table I), the copepods contain about 2.7 ng lycopeneindividual21 after 10 days in cultures. Lycopene was notfound in the copepods fed the live algal diet.
Quantification of astaxanthin in twocopepod populations
ANOVA analyses of the isocratic HPLC resultsindicated that the tanks fed the formulated feedhad significantly higher amounts of trans- andcis-astaxanthin, as well as a higher proportion of trans-astaxanthin when compared to total astaxanthincontent; 47% (+4% S.D., n ¼ 3) versus 26% (+2%S.D., n ¼ 3), p , 0.05 (Fig. 4). The dry weights andamount of lipid per adult copepod did not differ signifi-cantly between treatments and had an average value of5.5 mg dry weight individual21 (+0.7 S.D., n ¼ 6) and0.56 mg lipid individual21 (+0.2 S.D., n ¼ 6). The totalamount of astaxanthin in copepods fed live microalgae
Fig. 2. Retention time and absorbance at 470 nm for copepods fed the formulated feed. All peak identifications are provided in Table I. Peaksidentified as carotenoids are numbered. Peaks that represent the same compounds are given the same number on all figures. Unlabeled peaksare chlorophyll-related peaks. Inset provided to scale peak 2.
Table I: Carotenoid composition of two populations of copepods fed different diets
Peak number Retention time(min)
Identification absorption maximaabsorption maxima Copepods fed TetraselmisTetraselmis(% of identified carotenoids)
Copepods fed formula(% of identified carotenoids)
1 4.49 Unknown ketocarotenoid 450 (470) 3.02 —2 5.19 b-doradexanthin (adonixanthin) 450 475 14.42 67.443 5.5 Violaxanthin 430 455 485 1.11 —4 5.91, 10.88 Lutein-like 425 450 475 5.45 —5 5.91 cis-ketocarotenoid 330 440 465 — 10.306 7.09 Unknown ketocarotenoid 445 465 — 4.327 8.43 Unknown carotenoid 450 475 1.82 —8 and 9 8.8, 8.92 cis- and trans-Astaxanthin (coeluted) 475 26.58 2.6810 9.62 Antheraxanthin 415 445 475 1.26 6.3511 10.11 Lutein 420 445 475 8.36 —12 12.62 Zeaxanthin 420 450 470 0.70 —13 17.51, 21.92, 31.47, 36.95 Astaxanthin ester 475 15.68 —14 23.63 1,1-carotene 415 440 470 — 0.2715 26.39 b,1-carotene (a carotene) 445 475 5.12 —16 29.12 b,b-carotene (b carotene) 430 455 480 2.42 4.1717 29.88 9-cis b-carotene 425 445 475 2.25 1.1218 38.82 cis-Lycopene (365) 445 470 500 — 1.0919 40.15 Astacene 485 11.81 0.2420 41.88 Lycopene 450 475 505 — 2.02
A. C. E. RHODES j DIETARY EFFECTS ON COPEPOD CAROTENOIDS
i77
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from
T. suecica was 1.0 ng astaxanthin individual21 (+0.2S.D., n ¼ 3) or 185 mg astaxanthin (g dry weight)21
(+32 S.D., n ¼ 3). According to the results of theone-way ANOVA, the copepods fed the formulatedfeed had a statistically significantly higher astaxanthincontent, �3.6 ng astaxanthin individual21 (+1.7 S.D.,n ¼ 3) or 650 mg astaxanthin (g dry weight)21 (+303S.D., n ¼ 3) (Fig. 4).
D I S C U S S I O N
N. lacustris contained many of the same pigments asother copepods (Hairston, 1976; Hairston, 1979;Bandaranayake and Gentien, 1982; Goodwin, 1984;
Buffan-Dubau et al., 1996; Juhl et al., 1996; Czeczugaet al., 2000; Andersson et al., 2003; Davenport et al.,2004). As described in previous studies, the carotenoidcomposition varied with diet (Buffan-Dubau et al.,1996; Juhl et al., 1996; Kleppel, 1998; McLeroy-Etheridge and McManus, 1999; Andersson et al., 2003;Davenport et al., 2004; Van Nieuwerburgh et al., 2005).The amounts of cis-, trans- and total astaxanthin weresignificantly greater in the copepods fed the formulatedfeed. The amount of trans-astaxanthin per total astax-anthin was also significantly higher for the copepodsfed the formulated feed.
N. lacustris astaxanthin values for all populations inthe experiment ranged from 0.8 to 5.9 ng astaxanthinindividual21 and 146 to 1066 mg astaxanthin (g dry
Fig. 3. Absorption spectra of putative b-doradexanthin (solid line, peak 2 in Figs 1 and 2) and trans-astaxanthin (broken line, peak 9 inFigs 1 and 2).
Fig. 4. Trans-, cis- and total astaxanthin mg (g dry weight)21 for N. lacustris fed two different diets, live Tetraselmis (clear bars) and a formulatedfeed (shaded bar). Values represent an average of three replicates in each treatment, each replicate was tested in duplicate. Significantly, differentmeans (one-way ANOVA) are indicated by separate letters for each measurement. Bars denote 1 S.D.
JOURNAL OF PLANKTON RESEARCH j VOLUME 29 j SUPPLEMENT 1 j PAGES i73–i83 j 2007
i78
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from

weight)21. This falls within the middle of the range ofvalues reported for other copepods. In a survey of theontogenetic stages of Acartia bifilosa and Pseudocalanus
acuspes, Lotocka et al. (2004) found that nauplii had thehighest concentration of unesterified astaxanthin, withmean respective concentrations of 427 mg (g dryweight)21 and 321 mg (g dry weight)21. The amount ofastaxanthin reported for Calanus pacificus by Juhl et al.(1996) (11 to 92 ng astaxanthin female21) equals � 65to 541 mg astaxanthin (g dry weight)21 since C. pacificus
adult females dry weight averages 170 mg (Frost, 1972),which is 30 times heavier than N. lacustris. Hairston(1980) found 5 mg pigment (g dry weight)21 forDiaptomus nevadensis, or about 250 ng individual21,which is an order of magnitude larger than what isfound in N. lacustris. However, D. nevadensis inhabitshigh alpine lakes that are exposed to more ultravioletradiation resulting in higher astaxanthin content(Hairston, 1976).Tigriopus brevicornis, a bright orange harpacticoid
copepod found in rocky tide pools, contained10.35 mg astaxanthin (g dry weight)21 when raised inthe laboratory with a substratum of Enteremorpha sp.and a diet of Tetraselmis sp, exceeding the naturallevels found in wild-harvested T. brevicornis [4.86 mg(g dry weight)21] (Davenport et al., 2004). Interestingly,no astaxanthin was detected in the laboratory rearedT. brevicornis when raised in the dark on a diet ofbaker’s yeast (Davenport et al., 2004). Conversely,Canuella perplexa, a harpacticoid copepod which lives inthe sediments of salt marshes, was reported to containvery low amounts of astaxanthin, about 0.0044 to0.012 ng astaxanthin individual21 when harvestedfrom the wild (Buffan-Dubau et al., 1996), which istwo orders of magnitudes less than what is found inlaboratory-reared N. lacustris. N. lacustris populations fedthe two different diets fall in the middle range ofastaxanthin content for marine calanoid and harpacti-coid copepods.
Transfer of pigments from foodto copepods
All Tetraselmis sp. pigments identified byBustillos-Guzman et al. (2002) and Egeland et al. (1995)were found in the copepods fed Tetraselmis in thisexperiment except trans-neoxanthin. Many of the samechlorophyll degradation products found in the copepodtissues and fecal pellets of the Pseudodiaptomous euryhalinus
in Bustillo-Guzman et al.’s (2002) study were also foundin the copepod tissues of N. lacustris: chlorophyllide a,phaeophytin b, phaeophytin a and pyrophaeophytin a.Juhl et al. (1996) found phaeophorbides and
phaeophytins in Calanus pacificus that were reported tobe derived from the digestive breakdown of chlorophyllfrom algae in the diet. McLeroy-Etheridge andMcManus (1999) also reported the rapid breakdown ofchlorophyll into phaeopigments by copepods when foodwas limiting. This phenomenon has been found inharpacticoid copepods as well, Canuella perplexa whichsubsists on diatoms, cyanobacteria and/or green micro-algae converts ingested chlorophyll a into phaeophytinand phaeophorbide-like compounds (Buffan-Dubauet al., 1996).
The formulated feed had an effect on the copepodcarotenoid composition, most notably in the amount oflycopene. The formulated feed contains mostly lycopene(87% w/w), which equates to an availability of �5.5 nglycopene per individual copepod for the duration of theexperiment. Copepods fed the live algae did notcontain lycopene; whereas the copepods fed the formu-lated feed contain about 2.7 ng lycopene individual21,suggesting that the copepods were most likely bioaccu-mulating the lycopene. It is not possible to determinefrom this experiment alone how much the carotenoidcontent of the copepods depends on bioconversionversus bioaccumulation. However, due the absenceof any astaxanthin in the diets, it is possible to state thatN. lacustris bioconverts astaxanthin from precursors inthe diet.
Role of ketocarotenoids incopepod metabolism
Some evidence for a conversion pathway involving theputatively identified b-doradexanthin (adonixanthin) isfound in the extremely large peak (peak 2) found pre-ceding free astaxanthin (Figs 1 and 2). These peaks haveabsorption spectra very close to spectra observed in thefreshwater crayfish for b-doradexanthin (Katayama et al.,1970; Berticat et al., 2000) (Fig. 3). b-doradexanthin hasalso been found to be more polar than astaxanthin inother HPLC analyses (Linan-Cabello et al., 2002), whichmatches the observation from this study that the putativeb-doradexanthin elutes before astaxanthin. Furthermore,no absorption spectra found in the survey correspondedwith the intermediates canthaxanthin and echinenone,suggesting that the copepods may be using an alterna-tive bioconversion pathway.
It has been suggested that it is the structure of astax-anthin and canthaxanthin which makes them moreeffective as antioxidants (Terao, 1989); hence, othercarotenoids with similar structures should share some ofthe same properties. It is usually a minor carotenoid inrelation to astaxanthin and canthaxanthin(Linan-Cabello et al., 2002). If peak 2 is indeed
A. C. E. RHODES j DIETARY EFFECTS ON COPEPOD CAROTENOIDS
i79
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from

b-doradexanthin (adonixanthin), it may be serving asan intermediate between zeaxanthin and astaxanthin.b-doradexanthin has a substituent group at the C-4,which b-carotene and zeaxanthin do not (Katayamaet al., 1970). The position of this group may have aneffect on the antioxidant capacity of carotenoids(Woodall et al., 1997). For example, astaxanthin andcanthaxanthin have been found to have greater freeradical quenching ability than zeaxanthin or b-carotenein vitro (Terao, 1989). The antioxidant properties ofb-doradexanthin (adonixanthin) have not been tested inrelation to the other ketocarotenoids.The large proportion of the peak 2 ketocarotenoid
putatively identified as b-doradexanthin in the cope-pods fed the formulated feed suggests that astaxanthinmay not be the only ketocarotenoid that serves as anantioxidant for N. lacustris. The harpacticoid copepodN. lacustris is an epibenthic detritovore found in saltmarshes, and will migrate up into the water columnperiodically. The role of the pigmentation in N. lacustris
may be more than solely photoprotective, as has beendescribed in high radiation environments for tide pooldwelling harpacticoid copepods (Davenport et al., 2004).For example, Lotocka et al. (2004) hypothesized thatcarotenoid molecules such as astaxanthin serve as aphysiological oxygen replacement mechanism forupwardly migrating calanoid copepods which arerapidly combusting lipid materials as they swim.
Astaxanthin bioconversion pathways
The exact pathway of bioconversion has not been deter-mined for most aquatic animals. Thommen andWackernagel (1964) first suggested the pathwayb-carotene! echninenone! canthaxanthin!astaxanthin (Fig. 5). This pathway has been found to bethe most probable one for conversion of dietary caro-tenoids to canthaxanthin in the brine shrimp Artemia
salina (Czygan, 1968; Hsu et al., 1970). However, Artemiaare not able to continue the conversion process to astax-anthin (Hsu et al., 1970).Two alternative pathways which do not rely on
canthaxanthin and echinenone for the conversion ofthe carotenoids b-carotene and lutein to astaxanthinuse b-doradexanthin (adonixanthin) as an intermediate.The complete conversion of lutein to astaxanthin hasnot been definitively proven for any animal, becausethe bioconversion of a-doradexanthin into its isomerb-doradexanthin has not been demonstrated (Ohkuboet al., 1999). However, it has been demonstrated thatzeaxanthin is metabolised into astaxanthin viab-doradexanthin (adonixanthin) in goldfish (Hata andHata, 1972; Ohkubo et al., 1999), the deep sea red crab
Geryon quinquedens (Kuo et al., 1976) and the crayfishAstacus leptodactylus Eschscholtz (Berticat et al., 2000).One occurrence of b-doradexanthin (adonixanthin) hasbeen reported in a species of copepod surveyed fromthe Great Barrier Reef (Bandaranayake and Genien,1982). However, this copepod also contained the inter-mediate canthaxanthin, which makes it difficult todetermine which, if any, pathway is being used toproduce astaxanthin.
The bioconversion of astaxanthin from b-caroteneand zeaxanthin cannot be specifically elucidated fromthe information presented in this paper. However, someevidence for a conversion pathway involving the keto-carotenoid b-doradexanthin (adonixanthin) is foundwhen the absorption spectra for the ketocarotenoidpeak (peak 2) preceding the free astaxanthin peaks(peaks 8 and 9 in Figs 1 and 2) are analyzed. This peakhas absorption spectra different from the astaxanthinspectra observed in the RP-HPLC phase of the exper-iment (Fig. 3), and which is almost identical to spectraobserved in the freshwater crayfish for b-doradexanthin(Katayama et al., 1970; Berticat et al., 2000).b-doradexanthin has also been found to be more polarthan astaxanthin in other HPLC analyses, whichmatches the observation from this study that the puta-tive b-doradexanthin elutes before astaxanthin.
Furthermore, no absorption spectra found in thesurvey corresponded with the intermediates canthax-anthin and echinenone utilized by other crustaceans(e.g. shrimp, lobster and crabs) to convert b-carotene to
Fig. 5. Suggested pathways for b-carotene conversion to astaxanthin(Thommen and Wackernagel, 1964; Goodwin, 1984; Katayama et al.,1970; Bandaranayake and Gentien, 1982; Berticat et al., 2000,Linan-Cabello, 2002). Black arrows indicate the pathway proposed formost crustaceans. White arrows indicate variations from this mainpathway in other crustaceans. The arrow with a dotted line is ahypothesized pathway for lutein to astaxanthin. The arrows that areshaded gray indicate an alternative conversion pathway proposed forcrustaceans that may not rely on echinenone and canthaxanthin asintermediates.
JOURNAL OF PLANKTON RESEARCH j VOLUME 29 j SUPPLEMENT 1 j PAGES i73–i83 j 2007
i80
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from

astaxanthin (Goodwin, 1984; Linan-Cabello, 2002). Theastaxanthin conversion from dietary carotenoids may befollowing an alternative pathway to the one proposedfor other copepods (Lotocka and Styczynska-Jurewicz,2001; Lotocka et al., 2004). All experiments were con-ducted on adult copepods, so it is not possible to deter-mine whether all life stages are deficient in thesecarotenoids. It is possible that the carotenoid compo-sition of different life stages might contain alternativepigments, as is found in some calanoid species whichcontain canthaxanthin primarily in the early life stagesand not in the adult stages (Lotocka et al., 2004). Due tothe sensitivity of the RP-HPLC procedure, it is notlikely that these carotenoids were present and notdetected, unless they have an almost instantaneous life-span during the conversion process.The amount of astaxanthin conversion by the cope-
pods depended on the diet. Even though the copepodsfed the formulated feed had a lower compositional ratioof astaxanthin, copepods fed the formulated feed hadhigher amounts of free astaxanthin than copepods fedthe live alga Tetraselmis due to the high overall quantitiesof carotenoid. A spectral absorbance peak characteristicof b-doradexanthin (adonixanthin) found in both treat-ments suggests that N. lacustris converts b-carotene toastaxanthin using an alternative pathway that has beenfound in crayfish and deep sea crabs; however, furtherclarification on the identify of this ketocarotenoidpeak needs to be done using a method such as massspectrometry. Copepods fed the live alga Tetraselmis hadproportionately twice as much unesterified astaxanthin(27%) in comparison to the putatively identifiedb-doradexanthin (14%), as well as various astaxanthinesters. In contrast, the copepods fed the formulated feedcontained a very large proportion of the putativelyidentified b-doradexanthin (67%) in comparison tounesterified astaxanthin (3%). This may indicate thatthe enzymatic conversion from this intermediate toastaxanthin may be limited, or it may not be necessaryto convert the ketocarotenoid potentially identified asb-doradexanthin to astaxanthin if it fulfills the sameantioxidant role as astaxanthin for the copepods.Because the amount and proportion of astaxanthin
depended on the diet, diets containing higher amountsof b-carotene and zeaxanthin would be more likelyto produce high levels of astaxanthin in this species.The copepod populations fed the formulated diet stillcontained more astaxanthin, despite the lowerrelative proportions due to the extremely high contentof the ketocarotenoid potentially identified asb-doradexanthin. The fact that N. lacustris diets affectfinal carotenoid composition has implications for theupper trophic levels. In mammals, cis-isomers of
astaxanthin are preferentially absorbed (Østerlie et al.,2000). In contrast, lower digestibility of cis-astaxanthinrelative to trans-astaxanthin has been observed inAtlantic halibut (Hippoglossus hippoglossus) and Atlanticsalmon (Bjerkeng and Berge, 2000). This difference indigestibility suggests that fish species may have differentmechanisms than mammals for astaxanthin transportinto the enterocytes and incorporation into lipoproteins(Bjerkeng and Berge, 2000). Alterations in the basecarotenoid composition of the lower trophic food levelscould therefore affect the digestibility and nutritionalvalue of copepods to natural predators.
Nitokra lacustris is a good model organism for furtherresearch on carotenoid bioconversion in marine harpac-ticoid copepods, as it will readily consume inert and livefeeds. Future work will hopefully allow for manipulationof dietary carotenoid concentrations to determine whichprecursors are directly correlated to astaxanthin pro-duction. The relative importance of environmental con-ditions such as light cycles versus nutritional conditionscan also be tested using harpacticoid copepods, assuggested by the work of Davenport et al. (Davenportet al., 2004), which showed that diet as well as rearingconditions directly affected astaxanthin content inTigriopus brevicornis. Understanding how copepod caro-tenoid compositions are affected by their diets will helpelucidate the vital role that copepods play in the transferof astaxanthin and other ketocarotenoids to uppertrophic levels in the marine environment.
AC K N OW L E D G E M E N T S
I thank Dr Leon Boyd and Ruth Watkins at NC StateUniversity for their guidance and assistance in develop-ing this project. I would also like to thank the personnelat Craft Technologies, Inc., especially John Estes, fortechnical assistance in the preparation of the samplesand use of their HPLC system. I would also like toacknowledge Dr Peter Ferket, Dr Sam Mozley andDr Donna Wolcott of North Carolina State University,Raleigh, NC, USA and Ron Johnson of the NationalOceanic and Atmospheric Administration NorthwestFisheries Science Center, Seattle, WA, USA for theirhelpful comments on the manuscript.
R E F E R E N C E S
Andersson, M., Van Nieuwerburgh, L. and Snoeijs, P. (2003) Pigmenttransfer from phytoplankton to zooplankton with emphasis onastaxanthin production in the Baltic Sea food web. Mar. Ecol. Prog.
Ser. 254, 213–224.
A. C. E. RHODES j DIETARY EFFECTS ON COPEPOD CAROTENOIDS
i81
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from

Arab, L., Steck-Scott, S. and Fleishauer, A. T. (2002) Lycopene andthe lung. Exp. Biol. Med. 227, 894–899.
Bandaranayake, W. M. and Gentien, P. (1982) Carotenoids of Temoraturbinata, Centropages furcatus, Undinula vulgaris and Euchaeta russelli.Comp. Biochem. Phys. B, 72, 409–414.
Berticat, O., Negre-Sadargues, G. and Castillo, R. (2000) The metab-olism of astaxanthin during the embryonic development of the cray-fish Astacus leptodactylus Eschscholtz (Crustacea, Astacidea). Comp.
Biochem. Phys. B, 127, 309–318.
Bjerkeng, B. and Berge, G. M. (2000) Apparent digestibility coeffi-cients and accumulation of astaxanthin E/Z isomers in Atlanticsalmon (Salmo salar L.) and Atlantic halibut (Hippoglossus hippoglossusL.). Comp. Biochem. Physiol. B, 127, 423–432.
Britton, G. (1995) UV/V is spectroscopy in carotenoids. In Britton,G., Liaaen-Jensen, S. and Pfander, H. (eds.), Carotenoids. Vol. 1B.Chapter 2. Birkhauser Verlag, Basel, 13–62.
Buffan-Dubau, E., de Wit, R. and Castel, J. (1996) Feeding selectivityof the harpacticoid Canuella perplexa in benthic muddy environmentsdemonstrated by HPLC analysis of chlorin and carotenoid pig-ments. Mar. Ecol. Prog. Ser., 137, 71–82.
Bustillos-Guzman, J., Lopez-Cortes, D., Mathus, M. E. et al. (2002)Dynamics of pigment degradation by the copepodite stage ofPseudodiaptomus euryhalinus feeding on Tetraselmis suecica. Mar. Biol.140, 143–149.
Canjura, F. L. (1990) Separation, identification and degradation kinetics of
chlorophylls and chlorophyllides in spinach. MS Thesis. North CarolinaState University, Raleigh, NC, USA.
Czeczuga, B., Kozlowska, M. and Czeczuga-Semeniuk, E. (2000)Adaptive role of carotenoids and carotenoproteins in Cyclops kolensis
Lilljeborg (Crustacea: Copepoda) specimens to extremely eutrophi-cal conditions. Folia Biol. Krakow, 48, 77–84.
Czygan, F. C. (1968) On metabolism of carotenoids in crustaceanArtemia salina. Z Naturforsch Pt B, 23, 1367–1368.
Davenport, J., Healy, A., Casey, N. et al. (2004) Diet-dependent UVARand UVBR resistance in the high shore harpacticoid copepodTigriopus brevicornis. Mar. Ecol. Prog. Ser., 276, 299–303.
Egeland, E. S., Eikrem, W., Throndsen, J. et al. (1995) Carotenoidsfrom further prasinophytes. Biochem. Syst. Ecol., 23, 747–755.
Frost, B. W. (1972) Effects of size and concentration of food particleson the feeding behaviour of the marine planktonic copepod Calanus
pacificus. Limnol. Oceanogr., 17, 805–815.
Goodwin, T. W. (1984) The Biochemistry of the Carotenoids, 2nd edn.Vol. 2. Chapman and Hall, London, 64–96.
Hairston, N. G. (1976) Photoprotection by carotenoid pigments inthe copepod Diaptomus nevadensis. Proc. Natl. Acad. Sci. USA, 73,971–974.
Hairston, N. G. (1979) The effect of temperature on carotenoid photo-protection in the copepod Diaptomus nevadensis. Comp. Biochem. Phys. A,62, 445–448.
Hairston, N. G. (1980) On the diel variation of copepod pigmentation.Limnol. Oceanogr., 25, 742–747.
Hata, M. and Hata, M. (1972) Carotenoid pigment in gold fish. IV.Carotenoid metabolism. B. Jpn. Soc. Sci. Fish., 38, 331–338.
Hsu, W. J., Chichester, C. O. and Davies, B. H. (1970) The metab-olism of b -carotene and other carotenoids in the brine shrimp,Artemia salina L. (Crustacea: Branchiopoda). Comp. Biochem. Phys.,32, 69–79.
Hsu, W. J., Rodriguez, D. B. and Chichester, C. O. (1972) Biosynthesisof astaxanthin—VI. Conversion of [c-14] lutein and [c-14]b-carotene in goldfish. Int. J. Biochem., 3, 333–338.
Hyvarinen, K. and Hynninen, P. H. (1999) Liquid chromatographicseparation and mass spectrometric identification of chlorophyll ballomers. J. Chromatogr. A, 837, 107–116.
Juhl, A. R., Ohman, M. D. and Goericke, R. (1996) Astaxanthin inCalanus pacificus: assessment of pigment-based measures of omnivory.Limnol. Oceanogr., 41, 1198–1207.
Katayama, T., Yokoyama, H. and Chichester, C. O. (1970) The bio-synthesis of astaxanthin. I. The structure of a-doradexanthin andb-doradexanthin. Int. J. Biochem., 1, 438–444.
Katayama, T., Shintani, K., Shimaya, M. et al. (1972)Biosynthesis of astaxanthin-IX. Transformation of labeledastaxanthin from diet of sea bream, Chrysophrys major
Temminck and Schlegel, to their body astaxanthin. B. Jpn. Soc.Sci. Fish., 38, 1399–1403.
Katayama, T., Kunisaki, Y., Shimaya, M. et al. (1973) The biosynthesisof astaxanthin. XIV. The conversion of labelled b-carotene-15.150-3H2 into astaxanthin in the crab, Portunus trituberculatus. Comp.
Biochem. Phys. B, 46, 269–272.
Kleppel, G. S. (1998) Plant and animal pigments as trophodynamicindicators. In Soule, D. F. and Kleppel, G. S. (eds), Marine Organisms
as Indicators. Springer-Verlag, New York, 73–90.
Kuo, H. C., Lee, T. C., Lee, C. O. et al. (1976) The carotenoids in thedeep sea red crab. Comp. Biochem. Phys. B, 54, 387–390.
Linan-Cabello, M. A., Paniagua-Michel, J. and Hopkins, P. M. (2002)Bioactive roles of carotenoids and retinoids in crustaceans. Aquacult.Nutr., 8, 299–309.
Lotocka, M. and Styczynska-Jurewicz, E. (2001) Astaxanthin, canthax-anthin and astaxanthin esters in the copepod Acartia bifilosa
(Copepoda, Calanoida) during ontogenetic development.Oceanologia, 43, 487–497.
Lotocka, M., Styczynska-Jurewicz, E. and Bledzki, L. A. (2004)Changes in carotenoid composition in different developmentalstages of copepods: Pseudocalanus acuspes Giesbrecht and Acartia spp.J. Plankton Res., 26, 159–166.
Matsuno, T. (1989) Animal carotenoid. In Krinsky, N. I.,Mathews-Roth, M. M. and Taylor, F. (eds), Carotenoids Chemistry andBiology. Plenum Press, New York, 59–74.
Matsuno, T. and Tsushima, M. (1995) Comparative biochemicalstudies of carotenoids in sea cucumbers. Comp. Biochem. Phys. B,111, 597–605.
McLeroy-Etheridge, S. L. and McManus, G. B. (1999) Food type andconcentration affect chlorophyll and carotenoid destruction duringcopepod feeding. Limnol. Oceanogr., 44, 2005–2011.
Miki, W. (1991) Biological functions and activities of animal caroten-oids. Pure Appl. Chem., 63, 141–146.
Nelis, H. J. C. F. and De Leenheer, A. P. (1988) Reversed-phase liquidchromatography of astacene. J. Chromatogr., 452, 535–542.
Ohkubo, M., Tsushima, M., Maoka, T. et al. (1999) Carotenoids andtheir metabolism in the goldfish Carassius auratus (Hibuna). Comp.
Biochem. Phys. B, 124, 333–340.
Østerlie, M., Bjerkeng, B. and Liaaen-Jensen, S. (2000) Plasmaappearance and distribution of astaxanthin E/Z and R/S isomers inplasma lipoproteins of men after single dose administration of astax-anthin. Nutr. Biochem., 11, 482–490.
JOURNAL OF PLANKTON RESEARCH j VOLUME 29 j SUPPLEMENT 1 j PAGES i73–i83 j 2007
i82
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from

Paanakker, J. F. and Hallegraeff, G. M. (1978) A comparative study onthe carotenoid pigmentation of the zooplankton of Lake Maarsseveen(Netherlands) and of Lac Pavin (Auvergne, France). I. Chromatographiccharacterization of carotenoid. Comp. Biochem. Phys. B, 60, 51–58.
Rhodes, A. (2003) Methods for mass culture of Nitokra lacustris, a marineharpacticoid copepod. In Browman, H. I. and Skiftesvik, A. B. (eds),The Big Fish Bang. Proceedings of the 26th Annual Larval FishConference. Institute of Marine Research, Bergen, Norway, 449–465.
Rhodes, A. and Boyd, L. (2005) Formulated feeds for harpacticoidcopepods: implications for population growth and fatty acid compo-sition. In Lee, C. S., O’Bryen, P. J. and Marcus, N. H. (eds),Copepods in Aquaculture. Blackwell Publishing, Oxford, UK, 61–73.
Shimidzu, N., Goto, M. and Miki, W. (1996) Carotenoids as singletoxygen quenchers in marine organisms. Fish. Sci., 62, 134–137.
Ston, J. and Kosakowska, A. (2002) Phytoplankton pigments desig-nation—an application of RP-HPLC in qualitative and quantitativeanalysis. J. Appl. Phycol., 14, 205–210.
Terao, J. (1989) Antioxidant activity of b-carotene-related carotenoidsin solution. Lipids, 24, 659–661.
Thommen, H. and Wackernagel, H. (1964) Zum vorkommen vonketo-carotinoiden in crustacean. Naturwissenschaften, 51, 87–88.
Tonucci, L. H., Holden, J. M., Beechert, G. R. et al. (1995) Carotenoidcontent of thermally processed tomato-based food products. J. Agr.Food Chem., 43, 579–586.
Torrissen, O. J. and Christiansen, R. (1995) Requirements for caroten-oids in fish diets. J. Appl. Ichthyol., 11, 225–230.
Van Nieuwerburgh, L., Wanstrand, I., Liu, J. G. et al. (2005)Astaxanthin production in marine pelagic copepods grazing on twodifferent phytoplankton diets. J. Sea Res., 5, 147–160.
Woodall, A. A., Lee, S. W., Weesie, R. J. et al. (1997) Oxidation ofcarotenoids by free radicals: relationship between structure andreactivity. Biochim. Biophys. Acta, 1336, 33–42.
Yuan, J. P. and Chen, F. (1997) Identification of astaxanthin isomersin Haematococcus lacustris by HPLC-photodiode array detection.Biotechnol. Tech., 11, 455–459.
Yuan, J. P. and Chen, F. (1998) Chromatographic separation and puri-fication of trans-astaxanthin from the extracts of Haematococcus
pluvialis. J. Agr. Food Chem., 46, 3371–3375.
A. C. E. RHODES j DIETARY EFFECTS ON COPEPOD CAROTENOIDS
i83
by guest on January 11, 2014http://plankt.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















