
The CD28Related Molecule ICOS Is Required for Effective T Cell–Dependent Immune Responses
11
Immunity, Vol. 13, 95–105, July, 2000, Copyright 2000 by Cell Press The CD28-Related Molecule ICOS Is Required for Effective T Cell–Dependent Immune Responses CTLA-4, delivers a negative signal to the activated T cell, opposing CD28-mediated costimulation (Walunas et al., 1994). The importance of this pathway is further illustrated in mice deficient in CTLA-4 that exhibit pro- found lymphoproliferative defects characterized by Anthony J. Coyle,* ‡ Sophie Lehar,* Clare Lloyd,* Jane Tian,* Tracy Delaney,* Stephen Manning,* Trang Nguyen,* Tim Burwell,* Helga Schneider, ² Jose Angel Gonzalo,* Michael Gosselin,* Laura Rudolph Owen,* Christopher E. Rudd, ² and Jose Carlos Gutierrez-Ramos* ‡ polyclonal T cell activation and a high frequency of cells expressing activation/memory T cell antigens (Tivol et * Department of Biology Inflammation Division al., 1995). More recently, CTLA-4 engagement has been demonstrated to augment antitumour immunity (Leach Millennium Pharmaceuticals, Inc. Cambridge, Massachusetts 02139 et al., 1996), to regulate autoimmune diabetes (Luhder et al., 1998), and to provide an important signal leading ² Department of Cancer Immunology and AIDS Dana-Farber Cancer Institute and to T cell tolerance (Perez et al., 1997). Recent in vitro experiments have suggested that the Department of Pathology Harvard Medical School dependency on CD28/B7-mediated costimulation is greatly influenced by the antigenic experience of the T Boston, Massachusetts 02115 cell. Although antigen-inexperienced CD4 1 T cells re- quire CD28-mediated signaling for IL-2 production and clonal expansion (McKnight et al., 1994; Seder et al., Summary 1994), optimal activation of recently activated T helper subsets occurs independently of CD28 ligation While CD28 is critical for expansion of naive T cells, (Schweitzer and Sharpe, 1998). In addition, although recent evidence suggests that the activation of ef- CD28 costimulation has been reported to promote dif- fector T cells is largely independent of CD28/B7. We ferentiation to the Th2 phenotype in vitro (Rulifson et suggest that ICOS, the third member of the CD28/ al., 1997), studies performed in CD28 gene–targeted CTLA-4 family, plays an important role in production mice have demonstrated that at least under some cir- of IL-2, IL-4, IL-5, and IFNg from recently activated T cumstances, normal Th2 effector immune responses cells and contributes to T cell–dependent B help in can be generated, suggesting that other costimulatory vivo. Inhibition of ICOS attenuates lung mucosal in- signals are important in T helper effector function flammation induced by Th2 but not Th1 effector popu- (Brown et al., 1996; Gause et al.; 1997, Wu et al., 1998). lations. Our data indicate a critical function for the In this context, members of the tumor necrosis factor third member of the CD28 family in T cell–dependent (TNF):TNF receptor family that include OX40, OX40L, immune responses. RANK:RANKL, and 4–1BB:4–1BBL have been impli- cated as regulators of T cell function (Bachmann et al., Introduction 1999; Chen et al., 1999; DeBenedette et al., 1999; Kopf et al., 1999). The majority of these studies indicate that In vitro and in vivo studies of T cell activation have led these molecules provide a costimulatory signal for acti- to the widely accepted hypothesis that in addition to vation of naive T cells. signals delivered to the T cell receptor (TCR) by MHC2/ In an attempt to identify novel genes that may function peptide complexes, costimulatory signals by antigen- as costimulators of effector populations and more im- presenting cells (APCs) are critical for complete immune portantly may regulate T helper subpopulations, we gen- activation, the absence of which results in an abortive erated subtractive libraries from activated murine Th1 immune response (Schwartz, 1990; Bretscher, 1999). versus Th2 clones. One of the Th2-specific cDNA clones The most important costimulatory signal delivered to we identified exhibited 36.5% homology to mCD28 and resting T cells occurs upon CD28 engagement by B7 38.5% homology to mCTLA-4 and was identical to the molecules (Jenkins et al., 1991; Harding et al., 1992). A recently described third member of the CD28/CTLA-4 large number of studies have attempted to address the family, ICOS (Hutloff et al., 1999; Yoshinaga et al., 1999). importance of CD28-mediated costimulation using a va- In the present study, we report that although ICOS is riety of in vivo model systems. These data have sug- induced upon activation, repetitive antigenic stimulation gested that CD28 engagement is critical in determining in the presence of IL-12 leads to downregulation of ICOS susceptibility to disease progression following infection mRNA in Th1 cells, resulting in overexpression in Th2 with Leishmania major (Corry et al., 1994) in the main- effector cells. Functionally, ICOS predominates over tenance of long-term allograft survival (Sayegh et al., CD28 in regulating cytokine production from recently 1995), for mediating experimental autoimmune enceph- activated T cells but not from naive T cells where CD28- alomyelitis (Cross et al., 1995), for T cell–dependent anti- dependent signaling predominates. In vivo, ICOS also body production (Linsley et al., 1992), and in the devel- delivers an important signal for T cell–dependent B cell opment of allergic lung disease (Keane-Myers et al., activation in a secondary immune response. However, 1997; Tsuyuki et al., 1997; Padrid et al., 1998). In contrast when highly differentiated T helper cells are generated, to CD28, the second member of this family of molecules, ICOS contributes in part to Th2 and not Th1 effector function in vitro. Moreover, in a model of allergic airway ‡ To whom correspondence should be addressed (e-mail: coyle@ mpi.com and [email protected]). inflammation mediated exclusively by adoptive transfer
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of The CD28Related Molecule ICOS Is Required for Effective T Cell–Dependent Immune Responses

Immunity, Vol. 13, 95–105, July, 2000, Copyright 2000 by Cell Press
The CD28-Related Molecule ICOSIs Required for EffectiveT Cell–Dependent Immune Responses
CTLA-4, delivers a negative signal to the activated Tcell, opposing CD28-mediated costimulation (Walunaset al., 1994). The importance of this pathway is furtherillustrated in mice deficient in CTLA-4 that exhibit pro-found lymphoproliferative defects characterized by
Anthony J. Coyle,*‡ Sophie Lehar,* Clare Lloyd,*Jane Tian,* Tracy Delaney,* Stephen Manning,*Trang Nguyen,* Tim Burwell,* Helga Schneider,†Jose Angel Gonzalo,* Michael Gosselin,*Laura Rudolph Owen,* Christopher E. Rudd,†and Jose Carlos Gutierrez-Ramos*‡ polyclonal T cell activation and a high frequency of cells
expressing activation/memory T cell antigens (Tivol et*Department of BiologyInflammation Division al., 1995). More recently, CTLA-4 engagement has been
demonstrated to augment antitumour immunity (LeachMillennium Pharmaceuticals, Inc.Cambridge, Massachusetts 02139 et al., 1996), to regulate autoimmune diabetes (Luhder
et al., 1998), and to provide an important signal leading†Department of Cancer Immunology and AIDSDana-Farber Cancer Institute and to T cell tolerance (Perez et al., 1997).
Recent in vitro experiments have suggested that theDepartment of PathologyHarvard Medical School dependency on CD28/B7-mediated costimulation is
greatly influenced by the antigenic experience of the TBoston, Massachusetts 02115cell. Although antigen-inexperienced CD41 T cells re-quire CD28-mediated signaling for IL-2 production andclonal expansion (McKnight et al., 1994; Seder et al.,Summary1994), optimal activation of recently activated T helpersubsets occurs independently of CD28 ligationWhile CD28 is critical for expansion of naive T cells,(Schweitzer and Sharpe, 1998). In addition, althoughrecent evidence suggests that the activation of ef-CD28 costimulation has been reported to promote dif-fector T cells is largely independent of CD28/B7. Weferentiation to the Th2 phenotype in vitro (Rulifson etsuggest that ICOS, the third member of the CD28/al., 1997), studies performed in CD28 gene–targetedCTLA-4 family, plays an important role in productionmice have demonstrated that at least under some cir-of IL-2, IL-4, IL-5, and IFNg from recently activated Tcumstances, normal Th2 effector immune responsescells and contributes to T cell–dependent B help incan be generated, suggesting that other costimulatoryvivo. Inhibition of ICOS attenuates lung mucosal in-signals are important in T helper effector functionflammation induced by Th2 but not Th1 effector popu-(Brown et al., 1996; Gause et al.; 1997, Wu et al., 1998).lations. Our data indicate a critical function for theIn this context, members of the tumor necrosis factorthird member of the CD28 family in T cell–dependent(TNF):TNF receptor family that include OX40, OX40L,immune responses.RANK:RANKL, and 4–1BB:4–1BBL have been impli-cated as regulators of T cell function (Bachmann et al.,Introduction1999; Chen et al., 1999; DeBenedette et al., 1999; Kopfet al., 1999). The majority of these studies indicate thatIn vitro and in vivo studies of T cell activation have ledthese molecules provide a costimulatory signal for acti-to the widely accepted hypothesis that in addition tovation of naive T cells.signals delivered to the T cell receptor (TCR) by MHC2/
In an attempt to identify novel genes that may functionpeptide complexes, costimulatory signals by antigen-as costimulators of effector populations and more im-presenting cells (APCs) are critical for complete immuneportantly may regulate T helper subpopulations, we gen-activation, the absence of which results in an abortiveerated subtractive libraries from activated murine Th1immune response (Schwartz, 1990; Bretscher, 1999).versus Th2 clones. One of the Th2-specific cDNA clonesThe most important costimulatory signal delivered towe identified exhibited 36.5% homology to mCD28 andresting T cells occurs upon CD28 engagement by B738.5% homology to mCTLA-4 and was identical to themolecules (Jenkins et al., 1991; Harding et al., 1992). Arecently described third member of the CD28/CTLA-4large number of studies have attempted to address thefamily, ICOS (Hutloff et al., 1999; Yoshinaga et al., 1999).importance of CD28-mediated costimulation using a va-In the present study, we report that although ICOS isriety of in vivo model systems. These data have sug-induced upon activation, repetitive antigenic stimulationgested that CD28 engagement is critical in determiningin the presence of IL-12 leads to downregulation of ICOSsusceptibility to disease progression following infectionmRNA in Th1 cells, resulting in overexpression in Th2with Leishmania major (Corry et al., 1994) in the main-effector cells. Functionally, ICOS predominates overtenance of long-term allograft survival (Sayegh et al.,CD28 in regulating cytokine production from recently1995), for mediating experimental autoimmune enceph-activated T cells but not from naive T cells where CD28-alomyelitis (Cross et al., 1995), for T cell–dependent anti-dependent signaling predominates. In vivo, ICOS alsobody production (Linsley et al., 1992), and in the devel-delivers an important signal for T cell–dependent B cellopment of allergic lung disease (Keane-Myers et al.,activation in a secondary immune response. However,1997; Tsuyuki et al., 1997; Padrid et al., 1998). In contrastwhen highly differentiated T helper cells are generated,to CD28, the second member of this family of molecules,ICOS contributes in part to Th2 and not Th1 effectorfunction in vitro. Moreover, in a model of allergic airway‡ To whom correspondence should be addressed (e-mail: coyle@
mpi.com and [email protected]). inflammation mediated exclusively by adoptive transfer
Immunity96
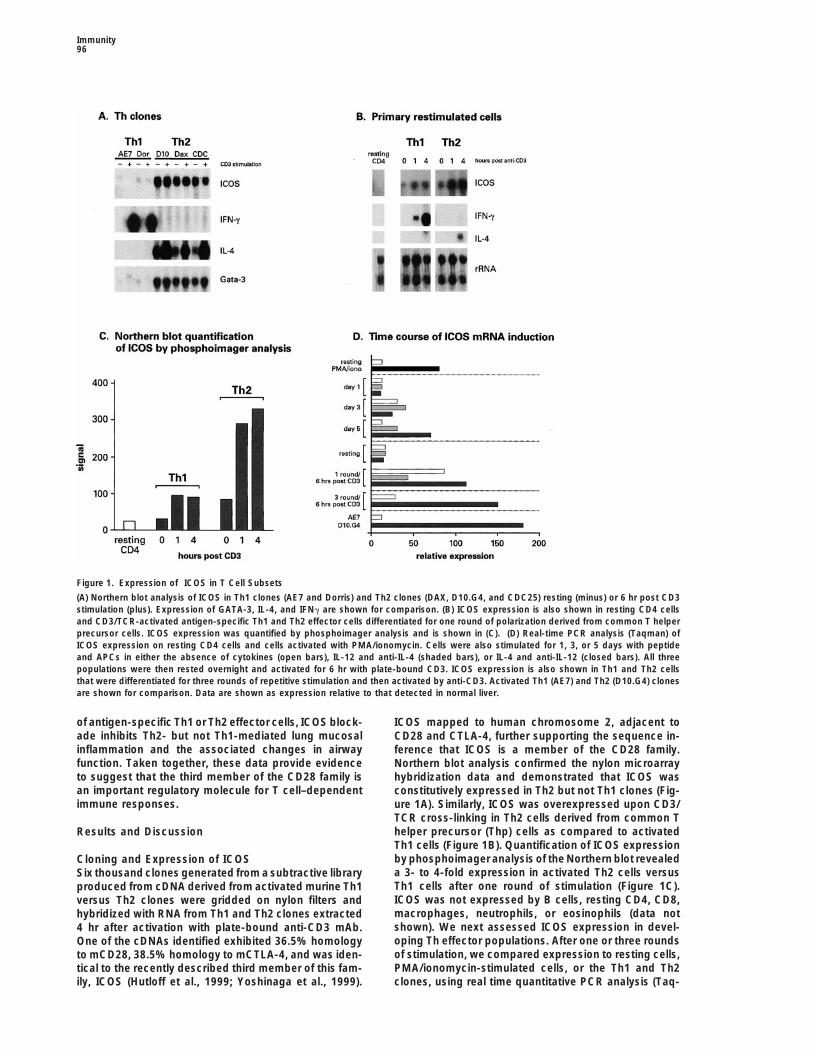
Figure 1. Expression of ICOS in T Cell Subsets
(A) Northern blot analysis of ICOS in Th1 clones (AE7 and Dorris) and Th2 clones (DAX, D10.G4, and CDC25) resting (minus) or 6 hr post CD3stimulation (plus). Expression of GATA-3, IL-4, and IFNg are shown for comparison. (B) ICOS expression is also shown in resting CD4 cellsand CD3/TCR-activated antigen-specific Th1 and Th2 effector cells differentiated for one round of polarization derived from common T helperprecursor cells. ICOS expression was quantified by phosphoimager analysis and is shown in (C). (D) Real-time PCR analysis (Taqman) ofICOS expression on resting CD4 cells and cells activated with PMA/ionomycin. Cells were also stimulated for 1, 3, or 5 days with peptideand APCs in either the absence of cytokines (open bars), IL-12 and anti-IL-4 (shaded bars), or IL-4 and anti-IL-12 (closed bars). All threepopulations were then rested overnight and activated for 6 hr with plate-bound CD3. ICOS expression is also shown in Th1 and Th2 cellsthat were differentiated for three rounds of repetitive stimulation and then activated by anti-CD3. Activated Th1 (AE7) and Th2 (D10.G4) clonesare shown for comparison. Data are shown as expression relative to that detected in normal liver.
of antigen-specific Th1 or Th2 effector cells, ICOS block- ICOS mapped to human chromosome 2, adjacent toade inhibits Th2- but not Th1-mediated lung mucosal CD28 and CTLA-4, further supporting the sequence in-inflammation and the associated changes in airway ference that ICOS is a member of the CD28 family.function. Taken together, these data provide evidence Northern blot analysis confirmed the nylon microarrayto suggest that the third member of the CD28 family is hybridization data and demonstrated that ICOS wasan important regulatory molecule for T cell–dependent constitutively expressed in Th2 but not Th1 clones (Fig-immune responses. ure 1A). Similarly, ICOS was overexpressed upon CD3/
TCR cross-linking in Th2 cells derived from common Thelper precursor (Thp) cells as compared to activatedResults and DiscussionTh1 cells (Figure 1B). Quantification of ICOS expressionby phosphoimager analysis of the Northern blot revealedCloning and Expression of ICOSa 3- to 4-fold expression in activated Th2 cells versusSix thousand clones generated from a subtractive libraryTh1 cells after one round of stimulation (Figure 1C).produced from cDNA derived from activated murine Th1ICOS was not expressed by B cells, resting CD4, CD8,versus Th2 clones were gridded on nylon filters andmacrophages, neutrophils, or eosinophils (data nothybridized with RNA from Th1 and Th2 clones extractedshown). We next assessed ICOS expression in devel-4 hr after activation with plate-bound anti-CD3 mAb.oping Th effector populations. After one or three roundsOne of the cDNAs identified exhibited 36.5% homologyof stimulation, we compared expression to resting cells,to mCD28, 38.5% homology to mCTLA-4, and was iden-PMA/ionomycin-stimulated cells, or the Th1 and Th2tical to the recently described third member of this fam-
ily, ICOS (Hutloff et al., 1999; Yoshinaga et al., 1999). clones, using real time quantitative PCR analysis (Taq-

ICOS and T Cell–Dependent Responses97
Figure 2. ICOS Associates with PI3K and Ex-hibits Lipid Hydrolysis Activity
Left panel: Jurkat cells, stably transfectedwith empty PCDNA3 vector (lanes 1–3),ICOS (lanes 4–6), or the deletion mutantICOSd180C (lanes 7–9) were either left un-stimulated (lanes 1, 4) or stimulated with anti-CD28 (lanes 2, 3) or anti-Flag mAbs (lanes 5,6, 8, 9) and rabbit anti-mouse antibody for 2min (lanes 2, 5, 8) and 7.5 min (lanes 3, 6, 9).Following activation, cells were lysed, im-munoprecipitated with CD28 mAb (lanes 1–3)or Flag mAb (lanes 4–9), and subjected toimmunoblotting with p85 antiserum. The cor-responding band recognized in cell lysateserved as a positive control (lane 10). Rightpanel: cells were stimulated with anti-CD28
(lane 12) or anti-Flag mAbs (lanes 13, 14) and rabbit anti-mouse antibody for 2 min, lysed, immunoprecipitated with CD28 mAb (lane 12) orFlag mAb (lanes 13, 14), and subjected to a lipid kinase assay. Lane 11 represents a negative control; lane 15 represents a positive controlfor PI 3K.
man, Perkin Elmer). RNA was prepared from CD41 T cross-linking (lanes 12 and 13). Deletion of the cyto-plasmic tail (ICOS d180C) (lanes 7–9 and lane 14) incells on days 1, 3, and 5 after peptide/APC stimulation
in the presence of either IL-12 and anti-IL-4 (Th1 polariz- ICOS transfectants ablated both PI3K association andlipid kinase activity. These data demonstrate that CD28ing), IL-4 and anti-IL-12 (Th2 polarizing), or in the ab-
sence of exogenous cytokines (neutral condition). Cells and ICOS share some common downstream signalingelements, although the precise significance of the con-were then rested overnight and activated with plate-
bound CD3. As shown in Figure 1D, ICOS was induced stitutive association of PI3K to ICOS in the absence ofreceptor ligation remains to be elucidated. However,24 hr after activation of CD4 cells with ionomycin/PMA
stimulation. Likewise, ICOS mRNA was increased by while the CD28 pYMNM site also binds the SH2 domainof Grb-2, albeit at 10-fold lower avidity than PI3K, ICOSday 3 following stimulation with specific antigen inde-
pendent of the addition of exogenous cytokines. In failed to recruit Grb-2 (data not shown), supporting pre-vious data that arginine (N-193) is required for Grb-2agreement with the Northern analysis, our Taqman data
confirmed the 3- to 4- fold overexpression in Th1 versus association (Schneider et al., 1995). It remains to bedetermined whether ICOS can either associate with orTh2 cells. After three repetitive stimulations, ICOS was
overexpressed 10- to 12-fold on Th2 versus Th1 and activate specific intracellular cascades.was comparable to that observed in the clones. Thus, inagreement with data recently described for both humanand murine expression (Hutloff et al., 1999; Yoshinaga Inhibition of ICOS Attenuates Cytokine Production
In Vitroet al., 1999), ICOS is induced upon activation. However,as cells undergo differentiation to subpopulations of Th In the first series of in vitro experiments, we compared
the requirement of CD28 versus ICOS for activation ofeffector cells with repetitive antigenic stimulation, ICOSmRNA expression is downregulated on Th1 cells, re- resting antigen-specific CD41 T cells using either the
extracellular domains of CTLA-4 or ICOS fused to humansulting in overexpression in Th2 effector populationsconsistent with our original identification from the SH- IgG1. Inhibition of CD28 but not ICOS dramatically atten-
uated the proliferative response and production of IL-2PCR library generated from Th1 and Th2 clones.(Figure 3A). These observations are consistent with theobservations that unlike CD28, ICOS ligation fails toICOS and CD28 Share Some Common
Signaling Mechanisms provide a costimulatory signal for IL-2 production (Hut-loff et al., 1999). However, while ICOS in combinationWe next investigated whether ICOS and CD28 interact
with common intracellular signaling proteins. CD28 has with CD3/TCR activation has been reported to stimulateT cell proliferative responses in T cells from CD28-defi-previously been shown to bind the lipid kinase phospha-
tidylinositol 3-kinase (PI3K) by means of a phosphotyro- cient mice (Yoshinaga et al., 1999), we were unable toshow any role of ICOS in mediating the proliferativesine-based motif, pYMNM, which serves as a binding
domain for the Src homology domain (SH2 domain) of response to specific antigen presented by APCs. In con-trast to the critical role of CD28 for activation of restingPI3K (Prasad et al., 1994; Truitt et al., 1994). Upon bind-
ing to CD28, PI3K is sequestered to the inner face of T cells, signals delivered by CD28 are not required forsecretion of either IFNg or IL-4 secretion from recentlythe plasma membrane, where it can act on its target
substrates. ICOS possesses a similar YMFM motif (resi- activated T cells, as suggested by recent studies usingAPCs deficient in both B7-1 and B7-2 (Schweitzer anddues 181–184) and is therefore also potentially suitable
for binding to the p85 subunit of PI 3K. As seen in Figure Sharpe, 1998). As ICOS does not bind to either B7-1 orB7-2 (Yoshinaga et al., 1999), we hypothesized that ICOS2, immunoprecipitation of ICOS, like CD28 (lanes 1–3),
coprecipitated a 85 kDa band that corresponds to the and its interaction with its ligand may contribute to cyto-kine secretion from effector T cells. To investigate thisp85 subunit of PI3K as detected by anti-p85 immu-
noblotting (lanes 4–6) and lipid kinase activity upon issue further, we activated Thp cells with peptide and
Immunity98
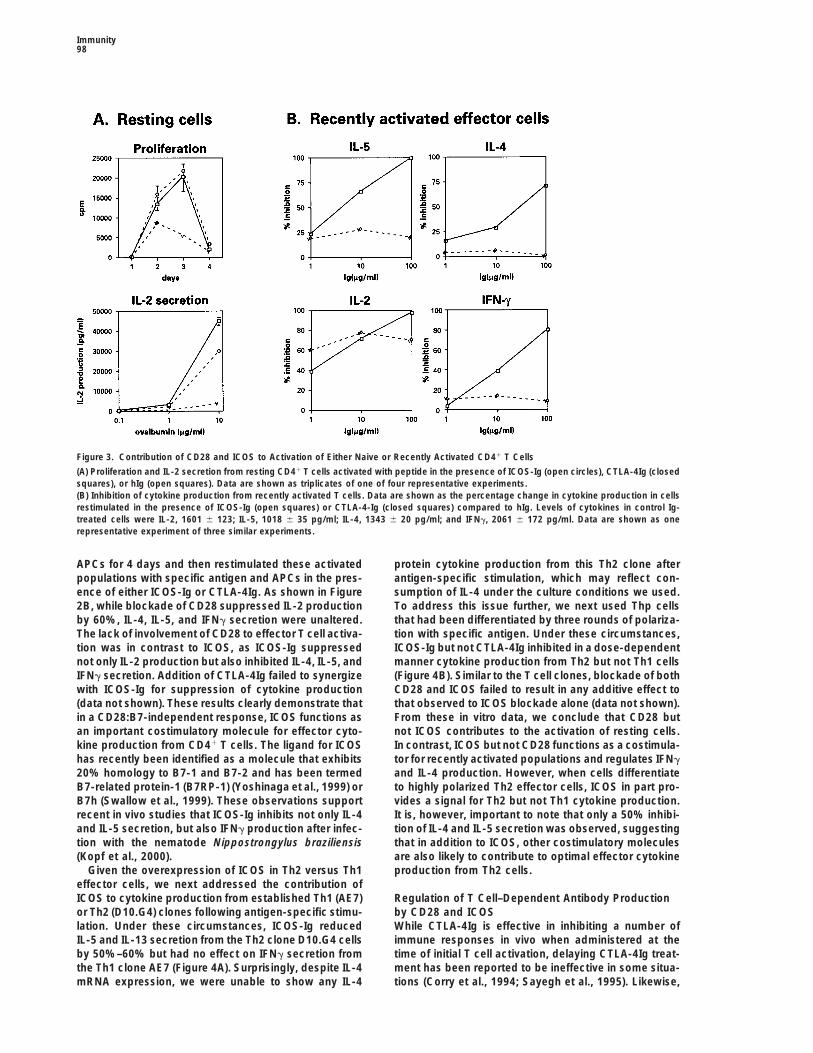
Figure 3. Contribution of CD28 and ICOS to Activation of Either Naive or Recently Activated CD41 T Cells
(A) Proliferation and IL-2 secretion from resting CD41 T cells activated with peptide in the presence of ICOS-Ig (open circles), CTLA-4Ig (closedsquares), or hIg (open squares). Data are shown as triplicates of one of four representative experiments.(B) Inhibition of cytokine production from recently activated T cells. Data are shown as the percentage change in cytokine production in cellsrestimulated in the presence of ICOS-Ig (open squares) or CTLA-4-Ig (closed squares) compared to hIg. Levels of cytokines in control Ig-treated cells were IL-2, 1601 6 123; IL-5, 1018 6 35 pg/ml; IL-4, 1343 6 20 pg/ml; and IFNg, 2061 6 172 pg/ml. Data are shown as onerepresentative experiment of three similar experiments.
APCs for 4 days and then restimulated these activated protein cytokine production from this Th2 clone afterantigen-specific stimulation, which may reflect con-populations with specific antigen and APCs in the pres-
ence of either ICOS-Ig or CTLA-4Ig. As shown in Figure sumption of IL-4 under the culture conditions we used.To address this issue further, we next used Thp cells2B, while blockade of CD28 suppressed IL-2 production
by 60%, IL-4, IL-5, and IFNg secretion were unaltered. that had been differentiated by three rounds of polariza-tion with specific antigen. Under these circumstances,The lack of involvement of CD28 to effector T cell activa-
tion was in contrast to ICOS, as ICOS-Ig suppressed ICOS-Ig but not CTLA-4Ig inhibited in a dose-dependentmanner cytokine production from Th2 but not Th1 cellsnot only IL-2 production but also inhibited IL-4, IL-5, and
IFNg secretion. Addition of CTLA-4Ig failed to synergize (Figure 4B). Similar to the T cell clones, blockade of bothCD28 and ICOS failed to result in any additive effect towith ICOS-Ig for suppression of cytokine production
(data not shown). These results clearly demonstrate that that observed to ICOS blockade alone (data not shown).From these in vitro data, we conclude that CD28 butin a CD28:B7-independent response, ICOS functions as
an important costimulatory molecule for effector cyto- not ICOS contributes to the activation of resting cells.In contrast, ICOS but not CD28 functions as a costimula-kine production from CD41 T cells. The ligand for ICOS
has recently been identified as a molecule that exhibits tor for recently activated populations and regulates IFNgand IL-4 production. However, when cells differentiate20% homology to B7-1 and B7-2 and has been termed
B7-related protein-1 (B7RP-1) (Yoshinaga et al., 1999) or to highly polarized Th2 effector cells, ICOS in part pro-vides a signal for Th2 but not Th1 cytokine production.B7h (Swallow et al., 1999). These observations support
recent in vivo studies that ICOS-Ig inhibits not only IL-4 It is, however, important to note that only a 50% inhibi-tion of IL-4 and IL-5 secretion was observed, suggestingand IL-5 secretion, but also IFNg production after infec-
tion with the nematode Nippostrongylus braziliensis that in addition to ICOS, other costimulatory moleculesare also likely to contribute to optimal effector cytokine(Kopf et al., 2000).
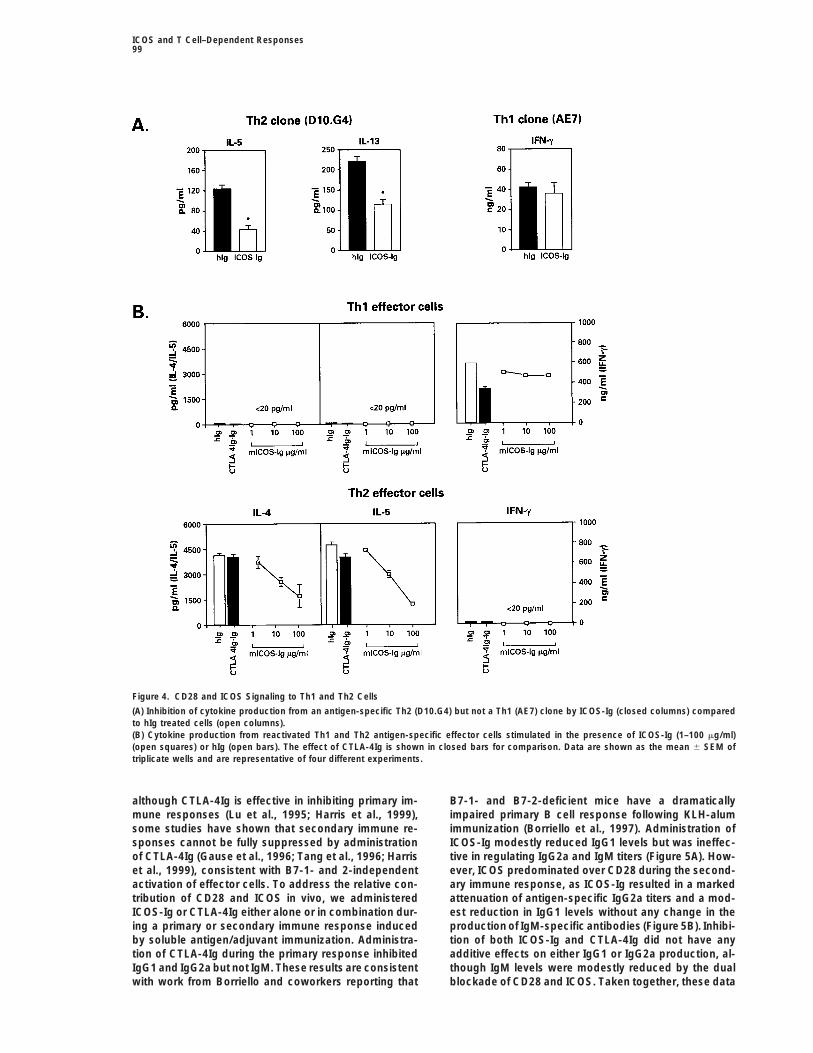
Given the overexpression of ICOS in Th2 versus Th1 production from Th2 cells.effector cells, we next addressed the contribution ofICOS to cytokine production from established Th1 (AE7) Regulation of T Cell–Dependent Antibody Production
by CD28 and ICOSor Th2 (D10.G4) clones following antigen-specific stimu-lation. Under these circumstances, ICOS-Ig reduced While CTLA-4Ig is effective in inhibiting a number of
immune responses in vivo when administered at theIL-5 and IL-13 secretion from the Th2 clone D10.G4 cellsby 50%–60% but had no effect on IFNg secretion from time of initial T cell activation, delaying CTLA-4Ig treat-
ment has been reported to be ineffective in some situa-the Th1 clone AE7 (Figure 4A). Surprisingly, despite IL-4mRNA expression, we were unable to show any IL-4 tions (Corry et al., 1994; Sayegh et al., 1995). Likewise,
ICOS and T Cell–Dependent Responses99
Figure 4. CD28 and ICOS Signaling to Th1 and Th2 Cells
(A) Inhibition of cytokine production from an antigen-specific Th2 (D10.G4) but not a Th1 (AE7) clone by ICOS-Ig (closed columns) comparedto hIg treated cells (open columns).(B) Cytokine production from reactivated Th1 and Th2 antigen-specific effector cells stimulated in the presence of ICOS-Ig (1–100 mg/ml)(open squares) or hIg (open bars). The effect of CTLA-4Ig is shown in closed bars for comparison. Data are shown as the mean 6 SEM oftriplicate wells and are representative of four different experiments.
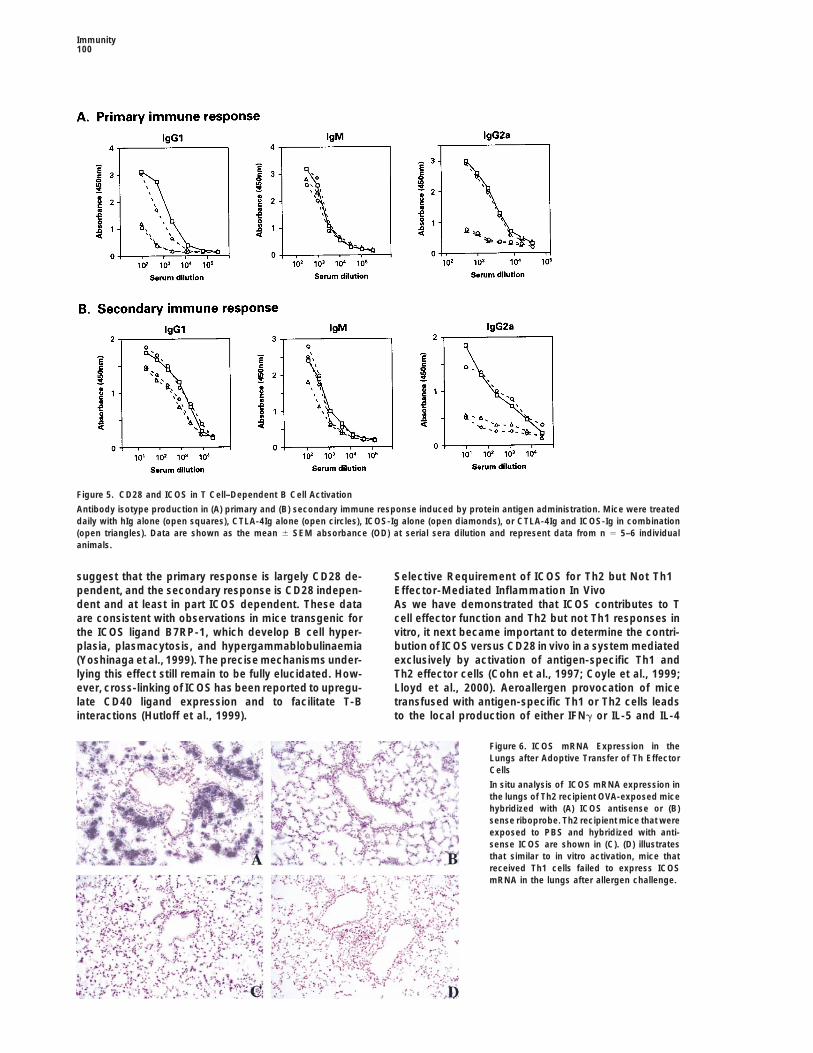
although CTLA-4Ig is effective in inhibiting primary im- B7-1- and B7-2-deficient mice have a dramaticallyimpaired primary B cell response following KLH-alummune responses (Lu et al., 1995; Harris et al., 1999),
some studies have shown that secondary immune re- immunization (Borriello et al., 1997). Administration ofICOS-Ig modestly reduced IgG1 levels but was ineffec-sponses cannot be fully suppressed by administration
of CTLA-4Ig (Gause et al., 1996; Tang et al., 1996; Harris tive in regulating IgG2a and IgM titers (Figure 5A). How-ever, ICOS predominated over CD28 during the second-et al., 1999), consistent with B7-1- and 2-independent
activation of effector cells. To address the relative con- ary immune response, as ICOS-Ig resulted in a markedattenuation of antigen-specific IgG2a titers and a mod-tribution of CD28 and ICOS in vivo, we administered
ICOS-Ig or CTLA-4Ig either alone or in combination dur- est reduction in IgG1 levels without any change in theproduction of IgM-specific antibodies (Figure 5B). Inhibi-ing a primary or secondary immune response induced
by soluble antigen/adjuvant immunization. Administra- tion of both ICOS-Ig and CTLA-4Ig did not have anyadditive effects on either IgG1 or IgG2a production, al-tion of CTLA-4Ig during the primary response inhibited
IgG1 and IgG2a but not IgM. These results are consistent though IgM levels were modestly reduced by the dualblockade of CD28 and ICOS. Taken together, these datawith work from Borriello and coworkers reporting that
Immunity100
Figure 5. CD28 and ICOS in T Cell–Dependent B Cell Activation
Antibody isotype production in (A) primary and (B) secondary immune response induced by protein antigen administration. Mice were treateddaily with hIg alone (open squares), CTLA-4Ig alone (open circles), ICOS-Ig alone (open diamonds), or CTLA-4Ig and ICOS-Ig in combination(open triangles). Data are shown as the mean 6 SEM absorbance (OD) at serial sera dilution and represent data from n 5 5–6 individualanimals.
suggest that the primary response is largely CD28 de- Selective Requirement of ICOS for Th2 but Not Th1Effector-Mediated Inflammation In Vivopendent, and the secondary response is CD28 indepen-
dent and at least in part ICOS dependent. These data As we have demonstrated that ICOS contributes to Tcell effector function and Th2 but not Th1 responses inare consistent with observations in mice transgenic for
the ICOS ligand B7RP-1, which develop B cell hyper- vitro, it next became important to determine the contri-bution of ICOS versus CD28 in vivo in a system mediatedplasia, plasmacytosis, and hypergammablobulinaemia
(Yoshinaga et al., 1999). The precise mechanisms under- exclusively by activation of antigen-specific Th1 andTh2 effector cells (Cohn et al., 1997; Coyle et al., 1999;lying this effect still remain to be fully elucidated. How-
ever, cross-linking of ICOS has been reported to upregu- Lloyd et al., 2000). Aeroallergen provocation of micetransfused with antigen-specific Th1 or Th2 cells leadslate CD40 ligand expression and to facilitate T-B
interactions (Hutloff et al., 1999). to the local production of either IFNg or IL-5 and IL-4
Figure 6. ICOS mRNA Expression in theLungs after Adoptive Transfer of Th EffectorCells
In situ analysis of ICOS mRNA expression inthe lungs of Th2 recipient OVA-exposed micehybridized with (A) ICOS antisense or (B)sense riboprobe. Th2 recipient mice that wereexposed to PBS and hybridized with anti-sense ICOS are shown in (C). (D) illustratesthat similar to in vitro activation, mice thatreceived Th1 cells failed to express ICOSmRNA in the lungs after allergen challenge.
ICOS and T Cell–Dependent Responses101
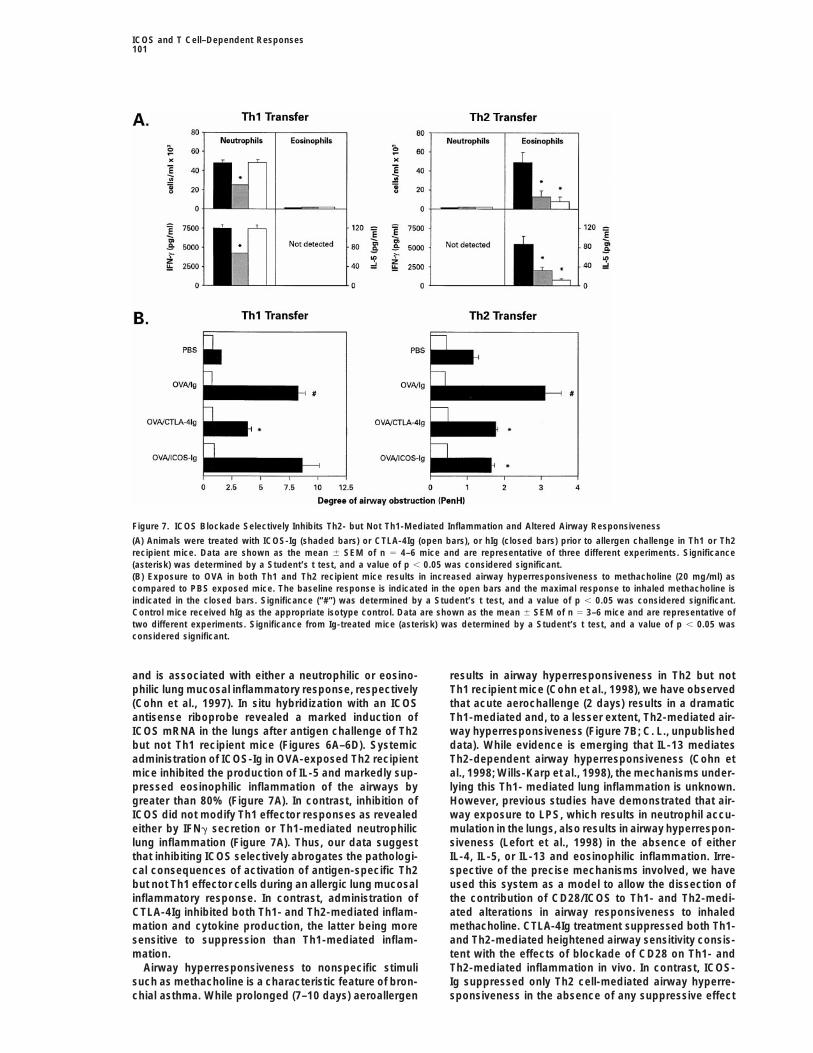
Figure 7. ICOS Blockade Selectively Inhibits Th2- but Not Th1-Mediated Inflammation and Altered Airway Responsiveness
(A) Animals were treated with ICOS-Ig (shaded bars) or CTLA-4Ig (open bars), or hIg (closed bars) prior to allergen challenge in Th1 or Th2recipient mice. Data are shown as the mean 6 SEM of n 5 4–6 mice and are representative of three different experiments. Significance(asterisk) was determined by a Student’s t test, and a value of p , 0.05 was considered significant.(B) Exposure to OVA in both Th1 and Th2 recipient mice results in increased airway hyperresponsiveness to methacholine (20 mg/ml) ascompared to PBS exposed mice. The baseline response is indicated in the open bars and the maximal response to inhaled methacholine isindicated in the closed bars. Significance (“#”) was determined by a Student’s t test, and a value of p , 0.05 was considered significant.Control mice received hIg as the appropriate isotype control. Data are shown as the mean 6 SEM of n 5 3–6 mice and are representative oftwo different experiments. Significance from Ig-treated mice (asterisk) was determined by a Student’s t test, and a value of p , 0.05 wasconsidered significant.
and is associated with either a neutrophilic or eosino- results in airway hyperresponsiveness in Th2 but notTh1 recipient mice (Cohn et al., 1998), we have observedphilic lung mucosal inflammatory response, respectively
(Cohn et al., 1997). In situ hybridization with an ICOS that acute aerochallenge (2 days) results in a dramaticTh1-mediated and, to a lesser extent, Th2-mediated air-antisense riboprobe revealed a marked induction of
ICOS mRNA in the lungs after antigen challenge of Th2 way hyperresponsiveness (Figure 7B; C. L., unpublisheddata). While evidence is emerging that IL-13 mediatesbut not Th1 recipient mice (Figures 6A–6D). Systemic
administration of ICOS-Ig in OVA-exposed Th2 recipient Th2-dependent airway hyperresponsiveness (Cohn etal., 1998; Wills-Karp et al., 1998), the mechanisms under-mice inhibited the production of IL-5 and markedly sup-
pressed eosinophilic inflammation of the airways by lying this Th1- mediated lung inflammation is unknown.However, previous studies have demonstrated that air-greater than 80% (Figure 7A). In contrast, inhibition of
ICOS did not modify Th1 effector responses as revealed way exposure to LPS, which results in neutrophil accu-mulation in the lungs, also results in airway hyperrespon-either by IFNg secretion or Th1-mediated neutrophilic
lung inflammation (Figure 7A). Thus, our data suggest siveness (Lefort et al., 1998) in the absence of eitherIL-4, IL-5, or IL-13 and eosinophilic inflammation. Irre-that inhibiting ICOS selectively abrogates the pathologi-
cal consequences of activation of antigen-specific Th2 spective of the precise mechanisms involved, we haveused this system as a model to allow the dissection ofbut not Th1 effector cells during an allergic lung mucosal
inflammatory response. In contrast, administration of the contribution of CD28/ICOS to Th1- and Th2-medi-ated alterations in airway responsiveness to inhaledCTLA-4Ig inhibited both Th1- and Th2-mediated inflam-
mation and cytokine production, the latter being more methacholine. CTLA-4Ig treatment suppressed both Th1-and Th2-mediated heightened airway sensitivity consis-sensitive to suppression than Th1-mediated inflam-
mation. tent with the effects of blockade of CD28 on Th1- andTh2-mediated inflammation in vivo. In contrast, ICOS-Airway hyperresponsiveness to nonspecific stimuli
such as methacholine is a characteristic feature of bron- Ig suppressed only Th2 cell-mediated airway hyperre-sponsiveness in the absence of any suppressive effectchial asthma. While prolonged (7–10 days) aeroallergen

Immunity102
of Th1-mediated alterations in airway function (Figure asthma, transplantation, and autoimmune responses,where activated T cells are believed to contribute to7B). It is important to note that these in vivo observations
with CTLA-4Ig are not necessarily in conflict with CD28- disease progression.independent activation of effector cells in vitro. In vivo,
Experimental Procedureseffective immune response requires not only the appro-priate costimulatory signals necessary for optimal cyto-
Generation of Th1 and Th2 Clones and Effector Populationskine production, but also the coordinated migration of Murine antigen-specific Th1 (AE7 and Dorris) and Th2 (D10.G4, DAX,T cells into the lymph nodes and target tissues, a pro- and CDC25) clones were stimulated every 10–14 days with specificcess that is critically regulated by chemokines and their peptide- and splenic mitomycin C–treated APCs and IL-2 (100
U/ml). Cells were activated with anti-CD3 mAb (2C11, PharMingen)receptors (Randolph et al., 1999). In this context, it isand poly(A)1 RNA isolated. To generate effector populations frominteresting to note that the upregulation of the chemo-T helper precusors (Thp) cells, mice expressing the transgene forkine receptors CCR4 and CCR8 on Th2 cells and CXCR3the DO11.10 ab-TCR, which recognizes residues 323–339 of chicken
on Th1 requires not only TCR ligation but also CD28 ovalbumin (OVA) in association with I-Ad, were used (Dr. D. Loh,engagement (D’Ambrosio et al., 1998). Therefore, atten- Washington University, St. Louis, MO). Resting TCR-transgenicuated migration of effector cells may at least in part CD41 T cells were cultured in complete RPMI 1640 with OVA 323-339
(10 mg/ml) and mitomycin C–treated splenocytes in a ratio of 1:2account for the effectiveness of CTLA-4Ig treatment ineither in the absence of exogenous cytokines (neutral conditions)vivo versus the lack of effect in vitro. In this context,or in the presence of IL-12 (10 ng/ml) and anti-IL-4 mAb (11B11, 40we have recently reported that inhibition of signalingmg/ml), or IL-4 (10 ng/ml) and anti-IL-12 mAb (TOSH 2, 3 mg/ml) to
through CCR4 inhibits Th2 cytokine production and eo- polarize to Th1 or Th2 phenotypes, respectively. After 5 days, cellssinophilic inflammation of the airways as a consequence were washed and incubated overnight in medium alone. RNA wasof attenuated migration/accumulation of Th2 cells (Lloyd either extracted from these resting cells or after activation by plate-
bound CD3 (2C11, 10 mg/ml) for either 1 or 4 hr.et al., 2000). Whether CTLA-4Ig inhibits the upregulationof CCR4 and/or CCR8 on T cells leading to reduced
Identification and Cloning of ICOSaccumulation is, however, speculative and remains toA Th2-specific library was generated using the Clontech PCR-Selectbe further investigated.cDNA Subtraction Kit (Clontech, Palo Alto, CA). Poly(A)1 RNA (2.0mg) from activated Th2 clones was used as “tester” and 2.0 mgof poly(A)1 RNA from activated Th1 clones used as “driver.” PCRConclusionsproducts from the library were cloned into the PCRII cloning vectorThis report provides evidence that the third member of(Invitrogen, San Diego, CA). Six thousand plasmid DNA from individ-the CD28 family, ICOS, is an important costimulatoryual clones were spotted onto nylon filters, denatured, and hybridiza-receptor for both recently activated cells and for Th2tion was performed with single stranded probes generated from Th1
but not Th1 effector cells. The contribution of ICOS to and Th2 RNA by reverse transcription using 33P- labeled dCTP.effective T cell–mediated immune responses and the Clones showing a differential hybridization to Th2 versus Th1 cDNA
were selected and sequenced. One of the novel cDNAs exhibitedfunction consequences of ICOS inhibition would be criti-a significant homology to mCD28. Differential expression of ICOScally influenced by both the nature of the immune re-cDNA was subsequently confirmed by Northern blot analysis withsponse and the timing of intervention with anti-ICOS1.0 mg of poly(A)1 RNA of Th1 and Th2 clones or activated Th1 andblockade strategies. In this context, we have recentlyTh2 cells derived from common Thp cells. This novel clone consisted
reported that ICOS contributes to effective T cell– of a 965 bp cDNA and was used to obtain the full-length clone usingmediated responses after infection with vesticular sto- a cDNA library prepared from murine Th2 cells and cDNAs cloned
into a lambda phage expression vector, ZIPLOX (GIBCO–BRL,matitis virus (VSV) but not lymphocytic choriomeningitusGaithersburg, MD). The cDNAs corresponding to the 2.2 kb tran-virus (LCMV) unless CD28 signaling is abrogated (Kopfscript isolated encode a predicted 22.7 kDa protein that shareset al., 2000). Inhibition of ICOS would be effective in36.5% identity with human CD28 and exhibits 69% to the recentlysuppressing the function of recently activated T helperidentified human gene ICOS.
cells, inhibiting the secretion of both IL-4 and IFNg. How- The restricted pattern of ICOS expression was extended usingever, under circumstances where strong immune devia- real-time quantitative PCR analysis (Taqman). In brief, an oligonucle-
otide probe was designed to anneal to the ICOS gene between twotion occurs, the contribution of ICOS to T cell activationPCR primers. The probe was then fluorescently labeled with FAMwould be restricted to Th2 helper cells. Our data suggest(reporter dye) on the 59 end and with TAMRA (quencher dye) on thethat CD28:B7-1 and B7-2 interactions are critical for39 end. A similar probe and PCR primers were designed forthe initiation of an effective immune response, whereasmGAPDH. The probe for this gene incorporated VIC as the reporter
ICOS:B7RP-1 is required at later stages and predomi- dye. PCR reactions were run that included the primers and probesnates over CD28 for secondary immune responses. This for these two genes as well as cDNA made from various cells and
tissues. As the polymerase moves across the gene during the reac-hypothesis is supported by the recent demonstrationtion, it cleaves the dye from one end of each probe, which causesthat administration of B7RP-1Ig enhances responses ina fluorescent emission that is measured by the Sequence Detectora model of cutaneous hypersensitivity when adminis-7700. The emissions recorded for each cDNA can then be convertedtered at the time of challenge but not during the initialinto the level of expression for ICOS normalized to the expression
sensitization phase. In addition, ICOS has also been of mGAPDH. Splenic CD4, CD8, and CD3 cells were purified byreported to be expressed on CD41CD44hiCD69low cells, negative selection (R&D Systems). B cells were isolated by positive
selection by labeling cells with rat anti-CD19 and separating the cellwhich are best defined as memory T cells (Yoshinagausing rat Ig- coupled beads (MACS Microbeads, Miltenyi Biotec,et al., 1999) that undergo rapid expansion on antigenCA). Neutrophils were obtained from the peritoneum after thioglycol-reexposure, independent of B7 and CD40 ligation (Lon-late treatment and macrophages were obtained as the adherentdon et al., 2000). Whether ICOS can modify memoryfraction of peritoneal cells. Finally, eosinophils (92% purity) were
T cell activation is currently under investigation in our isolated as the nonadherent population of peritoneal cells obtainedlaboratory. Finally, our data highlight the potential thera- from IL-5 transgenic mice. Expression of ICOS was determined on
resting CD4 cells or 24 hr after activation with PMA and ionomycin,peutic benefit of inhibiting ICOS in diseases such as

ICOS and T Cell–Dependent Responses103
in developing Th populations on days 1, 3, and 5 and after one mM Tris-HCl (pH 7.5) with 0.5 M LiCl, and twice with TNE (10 mMTris-HCl [pH 7.5], 150 mM NaCl, and 1 mM EGTA). The lipid kinaseround or three rounds of antigenic stimulation under the influence
of the appropriate cytokines. reaction was carried out on the beads using soybean PI liposomesand g [32P]ATP (20 mCi) as described (Prasad et al., 1994).
Mapping ICOS to Human Chromosome 2ICOS-specific primers (forward-TGCTTGACAGCCCAACAGCC, re- Regulation of T Cell Function by CD28 and ICOSverse-ATATTCTTGTAGGGAGAATATAGGC) were used to amplify a Resting TCR-transgenic CD41 T cells were cultured with OVA323–339159 bp PCR product from human control cell line DNA and multiple (10 mg/ml) and mitomycin C–treated splenocytes in a ratio of 1:2faint larger bands from control hamster cell line DNA by PCR. These under neutral conditions in the presence of CTLA-4Ig or ICOS-Igprimers were used to amplify the 90 DNAs in duplicate from the (100 mg/ml) or human Ig and pulsed with [3H]thymidine on days 1–4.Stanford TNG Radiation Hybrid Panel. Products were run on 2% Supernatants were collected on day 3 for measurement of IL-2 byagarose gels, poststained with SYBR Gold (1:10,000 dilution in 13 ELISA (R&D Systems). To study the contribution of CD28 and ICOSTBE), and scanned on a Molecular Dynamics 595 Fluorimager. ICOS on recently activated populations, CD4 cells were activated for 4was found to map to human chromosome 2, 8 cR50000 from the Stan- days with APC/peptide and then reactivated in 96-well plates (5 3ford Human Genome Center framework marker SHGC-13145, with 104/well) together with mitomycin C–treated 1 3 105 splenocytes ina LOD score for linkage of 8.99. This region corresponds to the the presence of ICOS-Ig or CTLA-4 Ig alone or in combination (1–100cytogenetic location 2q33. Further, markers designed from se- mg/ml), or human Ig as the control. Cytokine production was thenquence from human CTLA4 and CD28 were found to map 37.2 cR50000 assessed. To determine the effect of ICOS on effector cells, CD4and 103.8 cR50000 from ICOS, respectively, with a most probable gene cells were differentiated to Th1 and Th2 cells for three rounds oforder of ICOS-CTLA4-CD28. repetitive stimulation with 10 mg/ml OVA peptide in the presence of
either hIg, CTLA-4Ig, or ICOS-Ig (1–100 mg/ml). IL-4, IL-5, and IFNg
were measured by ELISA.Generation of ICOS-Ig Fusion ProteinsA DNA sequence containing the extracellular domain of ICOS wasPCR-amplified and cloned into a vector containing the CD5 signal
T Cell–Dependent B Cell Immune Responsessequence and the human IgG1 constant region (ICOS-Ig). COS cellsMale BALB/c mice (20 g) were immunized in the footpad on day 0were transiently transfected using lipofectamine (GIBCO), and thewith 10 mg OVA in 4 mg/alum. On day 8, mice were boosted with 100recombinant protein was purified over a protein A column. The puritymg OVA/alum administered subcutaneously. CTLA-4Ig and ICOS-Igof ICOS-Ig was subsequently assessed by coomassie-stained SDS–alone or in combination were administered daily i.p. either duringPAGE and was determined to be greater than 90%. The identifythe primary response or during the secondary response. Human Igof the ICOS-Ig was further confirmed by mass spectrometry bywas used as a control. On day 18, mice were bled and antigen-comparing the trypsin peptides generated from the extracted gelspecific IgG1, IgG2a, and IgM were measured by specific ELISAband to a theoretical trypsin digest (peptide mass fingerprinting by(PharMingen).MALDI-TOF analysis).
Adoptive Transfer of Antigen-Specific T Cells In VivoGeneration of Stable Jurkat Cells Expressing ICOSRecipient normal BALB/c mice were injected i.v. with 2 3 106 Th1Jurkat cells were transfected by electroporation with 2 mg ICOS-or Th2 effector cells. Twenty-four hours later, mice were exposedFlag DNA subcloned in pcDNA3 (Invitrogen). After a 2 week periodto an aerosol of ovalbumin (50 mg/ml) for 20 min on 2 consecutiveof drug selection, high Flag-positive expressing cells were sorteddays. One hour prior to allergen exposure, recipient mice were in-by flow cytometry (FACStar Vantage, Becton Dickinson), subcloned,jected i.v. with either 100 mg ICOS-Ig, CTLA-4Ig, or hIg (Sigma, St.and stable lines generated. ICOS and empty vector cells were thenLouis). Twenty-four hours later, a BAL was performed and cytokineselected on the basis of comparable surface expression of hCD3levels in the lavage fluid measured by ELISA. Lungs were removedand hCD28.24 hr after antigen challenge, inflated with 10% neutral bufferedformalin, and paraffin embedded. Four-micrometer sections were
Immunoprecipitation and Immunoblottingprehybridized for 2–4 hr, after which 35S-labeled ICOS sense and
For immunoprecipitations, Jurkat ICOS-Flag and empty vector-antisense riboprobes (1.2 3 106 cpm/slide) were added and hybrid-
transfected cells were either left unstimulated or stimulated withized overnight. Slides were dipped in emulsion, exposed for 2 weeks,
anti-CD28 (10 mg/ml, 4B10) or anti-Flag mAb (10 mg/ml) and rabbitdeveloped, and counterstained with hematoxylin.
anti-mouse antibody (5 mg/ml) for 2 and 7.5 min. Cells were thenlysed in ice-cold lysis buffer containing 1% Triton X-100 (v/v) in 20mM Tris-HCl (pH 8.3) and 150 mM NaCl. The lysis buffer contained Measurement of Airway Hyperresponsiveness1 mM PMSF, 1 mM Na4VO3, 10 mM NaF, and 1mM Na4P2O7. Lysates Airway responsiveness was measured in both Th1 and Th2 recipientwere incubated for 20 min on ice before centrifugation at 15,000 3 mice 24 hr after the last aerosol challenge by recording respiratoryg for 15 min at 48C. Postnuclear lysates were incubated for 1 hr pressure curves by whole body plethysmography (Buxco, EMKAwith agitation at 48C with the indicated monoclonal antibody. Protein Technologies, France) in response to 20 mg/ml of methacholineA-Sepharose beads (30 ml, Pharmacia) swollen and washed in lysis inhaled for 1 min. Airway responsiveness was expressed in en-buffer were added and incubated for 1 hr at 48C. The beads were hanced pause (Penh), a calculated value which correlates with mea-washed three times in cold lysis buffer and proteins were eluted by surement of airway resistance, impedance, and intrapleural pressureboiling for 5 min in SDS sample buffer, separated by SDS–PAGE, in the same mouse. Penh 5 (Te/TR1) 3 Pef/Pif (Te 5 expirationand transferred to nitrocellulose for immunoblotting. The mem- time, Tr 5 relaxation time, Pef 5 peak expiratory flow, Pif 5 peakbranes were blocked with 5% milk in TBS (10 mM Tris-HCl (pH 7.6), inspiratory flow) as described in detail elsewhere (Hamelman et al.,150 mM NaCl) and incubated with p85 antiserum. Bound antibody was 1997; Tsuyuki et al., 1997).revealed with horseradish peroxidase (HRP)-conjugated anti-rabbitantibody using enhanced chemiluminescence (ECL, Amersham).
Acknowledgments
Lipid Kinase Assay We would like to thank Mr. Chris Groves for advice on the FACSJurkat ICOS-Flag and empty vector-transfected cells were either and Steve Lin, Michael Pickard, and Louis Poisson for excellentleft unstimulated or stimulated with anti-CD28 (10 mg/ml, 4B10) or technical assistance. We would also like to acknowledge Mr. Kevinanti-Flag mAb (10 mg/ml) and rabbit anti-mouse antibody (5 mg/ml) Godout for production and purification of the ICOS-Ig fusion pro-for 2 and 7.5 min. Cells were solubilized in 1% Triton X-100 (v/v) teins. We are indebted to Dr. M. Kopf for helpful discussions.based lysis buffer with protease and phosphatase inhibitors andsubjected to precipitation as described above. Immune complexeswere washed three times with the lysis buffer, three times with 100 Received September 23, 1999; revised May 23, 2000.

Immunity104
References Harris, N.L., Peach, R.J., and Ronchese, F. (1999). CTLA4-Ig inhibitsoptimal T helper 2 cell development but not protective immunity ormemory response to Nippostrongylus brasiliensis. Eur. J. Immunol.Bachmann, M.F., Wong, B.R., Josien, R., Steinman, R.M., Oxenius,29, 311–316.A., and Choi, Y. (1999). TRANCE, a tumor necrosis factor family
member critical for CD40 ligand-independent T helper cell activa- Hutloff, A., Dittrich, A.M., Beier, H.C., Eljaschewitsch, B., Kraft, R.,tion. J. Exp. Med. 189, 1025–1031. Anagnostopoulos, I., and Kroczek, R.A. (1999). ICOS is an inducibleBorriello, F., Sethna, M.P., Boyd, S.D., Schweitzer, A.N., Tivol, E.A., T-cell co-stimulator structurally and functionally related to CD28.Jacoby, D., Strom, T.B., Simpson, E.M., Freeman, G.J., and Sharpe Nature 397, 263–266.A.H. (1997). B7-1 and B7-2 have overlapping, critical roles in immu- Jenkins, M.K., Taylor, P.S., Norton, S.D., and Urdahl, K.B. (1991).noglobulin class switching and germinal center formation. Immunity CD28 delivers a costimulatory signal involved in antigen-specific6, 303–313. IL-2 production by human T cells. J. Immunol. 8, 2461–2466.Bretscher, P.A. (1999). A two-step, two-signal model for the primary
Keane-Myers, A., Gause, W.C., Linsley, P.S., Chen, S.J., and Wills-activation of precursor helper T cells. Proc. Natl. Acad. Sci. USA
Karp, M. (1997). B7–CD28/CTLA-4 costimulatory pathways are re-96, 185–190.
quired for the development of T helper cell 2-mediated allergic air-Brown, D.R., Green, J.M., Moskowitz, N.H., Davis, M., Thompson, way responses to inhaled antigens. J. Immunol. 158, 2042–2049.C.B., and Reiner, S.L. (1996). Limited role of CD28-mediated signals
Kopf, M., Ruedl, C., Schmitz, N., Gallimore, A., Lefrang, K., Ecabert,in T helper subset differentiation. J. Exp. Med. 3, 803–810.B., Odermatt, B., and Bachmann, M.F. (1999). OX-40 deficient mice
Chen, A.I., McAdam, A.J., Buhlmann, J.E., Scott, S., Lupher, M.L., are defective in Th cell proliferation but are competent in generatingGreenfield, E.A., Baum, P.R., Fanslow, W.C., Calderhead, D.M., B cell and CTL responses after virus infection. Immunity 11, 699–708.Freeman, G.J., and Sharpe, A.H. (1999). OX40 ligand has a critical
Kopf, M., Coyle, A.J., Schmitz, N., Barner, M., Oxenius, A., Gallimore,costimulatory role in dendritic cell:T cell interactions. Immunity 11,A., Gutierrez-Ramos, J.C., and Bachmann, M.F. (2000). Inducible689–698.costimulator protein (ICOS) controls T helper cell subset polarization
Cohn, L., Homer, R.J., Marinov, A., Rankin, J., and Bottomly, K. after virus and parasite infection. J. Exp. Med. 192, 1–10.(1997). Induction of airway mucus production by T helper 2 (Th2)
Leach, D.R., Krummel, M.F., and Allison, J.P. (1996). Enhancement ofcells: a critical role for interleukin 4 in cell recruitment but not mucusantitumor immunity by CTLA-4 blockade. Science 271, 1734–1736.production. J. Exp. Med. 10, 1737–1747.Lefort, J., Singer, M., Leduc, D., Renesto, P., Nahori, M.A., Huerre,Cohn, L., Tepper, J.S., and Bottomly, K. (1998). IL-4-independentM., Creminon, C., Chignard, M., and Vargaftig, B.B. (1998). Systemicinduction of airway hyperresponsiveness by Th2, but not Th1, cells.administration of endotoxin induces bronchopulmonary hyperreac-J. Immunol. 161, 3813–3816.tivity dissociated from TNF-alpha formation and neutrophil seques-Corry, D.B., Reiner, S.L., Linsley, P.S., and Locksley, R.M. (1994).tration into the murine lungs. J. Immunol. 161, 474–480.Differential effects of blockade of CD28–B7 on the development ofLinsley, P.S., Wallace, P.M., Johnson, J., Gibson, M.G., Greene, J.L.,Th1 or Th2 effector cells in experimental leishmaniasis. J. Immunol.Ledbetter, J.A., Singh, C., and Tepper, M.A. (1992). Immunosuppres-153, 4142–4148.sion in vivo by a soluble form of the CTLA-4 T cell activation mole-Coyle, A.J., Lloyd, C., Eriksson, C., Tian, J., Nguyen, T., Wang, L.cule. Science 257, 792–795.Ottoson, P., Persson, P., Lunblad, L., Delaney, T., et al. (1999). T1/
ST2 plays a critical role in Th2 cell function and regulates allergen Lloyd, C.M., Delaney, T., Nguyen, T.,Tian, T., Martinez-A, C., Coyle,induced lung pathology and airway hyperresponsiveness. J. Exp. A.J., and Gutierrez-Ramos, J.C. (2000). CC chemokine receptorMed. 190, 895–902. (CCR)3/eotaxin is followed by CCR4/monocyte-derived chemokine
in mediating pulmonary T helper lymphocyte type 2 recruitment afterCross, A.H., Girard, T.J., Giacoletto, K.S., Evans, R.J., Keeling, R.M.,serial antigen challenge in vivo. J. Exp. Med. 191, 265–274.Lin, R.F., Trotter, J.L., and Karr, R.W. (1995). Long-term inhibition
of murine experimental autoimmune encephalomyelitis using CTLA- London, C.A., Lodge, M.P., and Abbas, A.K. (2000). Functional re-4-Fc supports a key role for CD28 costimulation. J. Clin. Invest. 6, sponses and costimulator dependence of memory CD41 T cells. J.2783–2789. Immunol. 164, 265–272.D’Ambrosio, D., Iellem, A., Bonecchi, R., Mazzeo, D., Sozzani, S., Lu, P., Zhou, X.D., Chen, S.J., Moorman, M., Schoneveld, A., Morris,Mantovani, A., and Sinigaglia F. (1998). Selective up-regulation of S., Finkelman, F.D., Linsley, P., Claassen, E., and Gause, W.C. (1995).chemokine receptors CCR4 and CCR8 upon activation of polarized Requirement of CTLA-4 counter receptors for IL-4 but not IL-10human type 2 Th cells. J. Immunol. 161, 5111–5115. elevations during a primary systemic in vivo immune response. J.
Immunol. 154, 1078–1087.DeBenedette, M.A., Wen, T., Bachmann, M.F., Ohashi, P.S., Barber,B.H., Stocking, K.L., Peschon, J.J., and Watts, T.H. (1999). Analysis Luhder, F., Hoglund, P., Allison, J.P., Benoist, C., and Mathis, D.of 4–1BB ligand (4–1BBL)-deficient mice and of mice lacking both (1998). Cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) regu-4–1BBL and CD28 reveals a role for 4–1BBL in skin allograft rejection lates the unfolding of autoimmune diabetes. J. Exp. Med. 187,and in the cytotoxic T cell response to influenza virus. J. Immunol. 427–432.163, 4833–4841.
McKnight, A.J., Perez, V.L., Shea, C.M., Gray, G.S., and Abbas, A.K.Gause, W.C., Lu, P., Zhou, X.D., Chen, S.J,. Madden, K.B., Morris, (1994). Costimulator dependence of lymphokine secretion by naiveS.C., Linsley, P.S., Finkelman, F.D., and Urban, J.F. (1996). H. poly- and activated CD41 T lymphocytes from TCR transgenic mice. J.gyrus: B7-independence of the secondary type 2 response. Exp. Immunol. 11, 5220–5225.Parasitol. 84, 264–273.
Padrid, P.A., Mathur, M., Li, X., Herrmann, K., Qin, Y., Cattamanchi,Gause, W.C., Chen, S.J., Greenwald, R.J., Halvorson, M.J., Lu, P.,
A., Weinstock, J., Elliott, D., Sperling, A.I., and Bluestone, J.A. (1998).Zhou, X.D., Morris, S.C., Lee, K.P., June, C.H., Finkelman, F.D., et al.
CTLA4Ig inhibits airway eosinophila and hyperresponsiveness by(1997). CD28 dependence of T cell differentiation to IL-4 production
regulating the development of Th1/Th2 subsets in a murine modelvaries with the particular type 2 immune response. J. Immunol. 158,
of asthma. Am. J. Respir. Cell. Mol. Biol. 4, 453–462.4082–4087.
Perez, V.L., Van Parijs, L., Biuckians, A., Zheng, X.X., Strom, T.B.,Hamelmann, E., Schwarze, J., Takeda, K., Oshiba, A., Larsen, G.L.,and Abbas, A.K. (1997). Induction of peripheral T cell tolerance inIrvin, C.G., and Gelfand, E.W. (1997). Noninvasive measurement ofvivo requires CTLA-4 engagement. Immunity 6, 411–417.airway responsiveness in allergic mice using barometric plethys-Prasad, K.V., Cai, Y.C., Raab, M., Duckworth, B., Cantley, L., Shoel-mography. Am. J. Respir. Crit. Care Med. 156, 766–775.son, S.E., and Rudd, C.E. (1994). T-cell antigen CD28 interacts withHarding, F.A., McArthur, J.G., Gross, J.A., Raulet, D.H., and Allison,the lipid kinase phosphatidylinositol 3-kinase by a cytoplasmicJ.P. (1992). CD28-mediated signalling co-stimulates murine T cellsTyr(P)-Met-Xaa-Met motif. Proc. Natl. Acad. Sci. USA 91, 2834–2838.and prevents induction of anergy in T-cell clones. Nature 356,
607–609. Randolph, D.A., Huang, G., Carruthers, C.J., Bromley, L.E., and

ICOS and T Cell–Dependent Responses105
Chaplin, D.D. (1999). The role of CCR7 in TH1 and TH2 cell localiza-tion and delivery of B cell help in vivo. Science 286, 2159–2162.
Rulifson, I.C., Sperling, A.I., Fields, P.E., Fitch, F.W., and Bluestone,J.A. (1997). CD28 costimulation promotes the production of Th2cytokines. J. Immunol. 15, 658–665.
Sayegh, M.H., Akalin, E., Hancock, W.W., Russell, M.E., Carpenter,C.B., Linsley, P.S., and Turka, L.A. (1995). CD28–B7 blockade afteralloantigenic challenge in vivo inhibits Th1 cytokines but sparesTh2. J. Exp. Med. 5, 1869–1874.
Schneider, H., Cai, Y.C., Prasad, K.V., Shoelson, S.E., and Rudd,C.E. (1995). T cell antigen CD28 binds to the GRB-2/SOS complex,regulators of p21ras. Eur. J. Immunol. 4, 1044–1050.
Schwartz, R.H. (1990). A cell culture model for T lymphocyte clonalanergy. Science 248, 1349–1356.
Schweitzer, A.N., and Sharpe, A.H. (1998). Studies using antigen-presenting cells lacking expression of both B7-1 (CD80) and B7-2(CD86) show distinct requirements for B7 molecules during primingversus restimulation of Th2 but not Th1 cytokine production. J.Immunol. 6, 2762–2771.
Seder, R.A., Germain, R.N., Linsley, P.S., and Paul, W.E. (1994).CD28-mediated costimulation of interleukin 2 (IL-2) production playsa critical role in T cell priming for IL-4 and interferon gamma produc-tion. J. Exp. Med. 1, 299–304.
Swallow, M.M., Wallin, J.J., and Sha, W.C. (1999). B7h, a novelcostimulatory homolog of B7.1 and B7.2, is induced by TNFalpha.Immunity 11, 423–432.
Tang, A., Judge, T.A., Nickoloff, B.J., and Turka, L.A. (1996). Sup-pression of murine allergic contact dermatitis by CTLA4Ig. Toleranceinduction of Th2 responses requires additional blockade of CD40-ligand. J. Immunol. 1, 117–125.
Tivol, E.A., Borriello, F., Schweitzer, A.N., Lynch, W.P., Bluestone,J.A., and Sharpe, A.H. (1995). Loss of CTLA-4 leads to massivelymphoproliferation and fatal multiorgan tissue destruction, reveal-ing a critical negative regulatory role of CTLA-4. Immunity 3,541–547.
Truitt, K.E., Hicks, C.M., and Imboden, J.B. (1994). Stimulation ofCD28 triggers an association between CD28 and phosphatidylinosi-tol 3-kinase in Jurkat T cells. J. Exp. Med. 3, 1071–1076.
Tsuyuki, S., Tsuyuki, J., Einsle, K., Kopf, M., and Coyle, A.J. (1997).Costimulation through B7-2 (CD86) is required for the induction ofa lung mucosal T helper cell 2 (TH2) immune response and alteredairway responsiveness. J. Exp. Med. 9, 1671–1679.
Walunas, T.L., Lenschow, D.J., Bakker, C.Y., Linsley, P.S., Freeman,G.J., Green, J.M., Thompson, C.B., and Bluestone, J.A. (1994).CTLA-4 can function as a negative regulator of T cell activation.Immunity 1, 405–413.
Wills-Karp, M., Luyimbazi, J., Xu, X., Schofield, B., Neben, T.Y., Karp,C.L., and Donaldson, D.D. (1998). Interleukin-13: central mediatorof allergic asthma. Science 282, 2258–2261.
Wu, Y., Zhou, Q., Zheng, P., and Liu, Y. (1998). CD28-independentinduction of T helper cells and immunoglobulin class switches re-quires costimulation by the heat-stable antigen. J. Exp. Med. 187,1151–1156.
Yoshinaga, S.K., Whoriskey, J.S., Khare, S.D., Sarmiento, U., Guo,J., Horan, T., Shih, G., Zhang, M., Coccia, M.A., Kohno, T., et al.(1999). T-cell co-stimulation through B7RP-1 and ICOS. Nature 402,827–832.




















