The carboxyl-terminal one-third of bovine rhodopsin: Its structure and function
10
V~.\ir,u Rc\. Vol. 22. pp. 1429 to 1438. 1982 Printed in Great Bntain 0043-6989,82~121429-I0$03.00~0 Pergamon Pres\ Ltd THE CARBOXYL-TERMINAL ONE-THIRD OF BOVINE RHODOPSIN: ITS STRUCTURE AND FUNCTION PAUL A. HARGRAVE’, J. HUGH MCDOWELL’, ELIZABETHC. SIEMIATKOWSK~-JUSZ~ZAK’. SHAO-LING FONG’, HERMANN K~;~HN~; JANET K. WANG’. DONNAR. CURTIS’, J.K.MOHANA RAO~, PATRICK ARGOSY and RICHARDJ.FELDMANN" ‘Department of Medical Biochemistry (School of Medicine) and Department of Chemistry and Bio- chemistry (College of Science), Southern Illinois University at Carbondale, Carbondale, IL 62901. U.S.A. “Baylor College of Medicine, Department of Ophthalmology, Texas Medical Center. Houston, TX 77025. U.S.A. -‘Institut fur Neurobiologie der Kernforschungsanlage, Postfach 1913, D-5170 Jtilich, West Germany ‘Department of Biological Sciences. Purdue University, West Lafayette. IN 47907. U.S.A. 5Computer Center Branch. Division of Computer Research and Technology, National Institutes of Health, Bethesda, MD 20205. U.S.A. Abstract-The covalent sequence of the carboxyl-terminal one-third of bovine rhodopsin has been determined. It is 108 amino acids in length and consists of three hydrophilic regions exposed to the aqueous environment at membrane surfaces and two hydrophobic regions which penetrate the disc membrane lipid bilayer. This carboxyl-terminal third of rhodopsin contains the retinal binding site, sites of phosphorylation, the fast-reacting sulfhydryl group and the thermolytic fragment F2. Hydrophobic helices in the sequence have been predicted by a new method developed for application to membrane proteins and a hypothetical “helical hairpin” space-filling working model has been constructed. INTRODUCTION In the first event in vision, rhodopsin’s chromophore, 1 I-cis-retinal, absorbs a photon and is isomerized to the all-trans form. This leads to permeability changes in the plasma membrane of the rod cell and to visuat excitation. To elucidate how the photoreceptor pro- tein rhodopsin functions in affecting rod cell bio- chemistry through response to light, we have con- ducted studies of rhodopsin’s structure and several of its chemical properties. In this article we summarize the results of our current studies on the structure and functional properties of the carboxyl-terminal region of rhodopsin. THE SEQUENCE OF THE CARBOXYL-TERMINAL ONE-THIRD OF RHODOPSIN HAS BEEN DETERMINED The carboxyl-terminal amino acid of bovine rho- dopsin has been previously reported to be resistant to identification (Albrecht, 1957; HelIer, 1968). However, application of the method of hydrazinolysis showed alanine to be the carboxyl-terminal amino acid (Har- grave and Fong. 1977). A nine amino acid tryptic pep- tide which contained alanine at its carboxyl-terminus was isolated and demonstrated to be the carboxyl- terminal peptide of rhodopsin (Hargrave and Fong, Stereo pictures in this paper can be viewed using an inex- pensive stereoviewer such as that sold by The Hubbard Scientific Co., Northbrook, Illinois (stereoscope). Abhreciutions: ROS-rod outer segments: des(l’ - la’)rho- dopsin--the rhodopsin molecule minus its 12 carboxyl- terminal amino acids; NAPT-nitroazidophenyl taur- ine; AP--azidopyrene; IASiodoacetamidosalicylate. 1977). Knowledge of the primary structure of this region was extended by the isolation and sequence determination of a 31”amino acid carboxyl-terminal peptide from the cyanogen bromide cleavage of rho- dopsin (Fong, 1978). Additional peptides were charac- terized which served to extend the carboxyl-terminal sequence to 40 amino acids (Hargrave or ~1.. 1980). Recently the laboratory of Virmaux at Strasbourg reported the isolation and partial characterization of a large peptide formed by cleavage of rhodopsin at tryptophanyl residues (Pellicone et ul., 1980). This peptide overlapped with our sequence and extended knowledge of this region of the molecule to 83 amino acids. To elucidate further the sequence of rhodopsin we have prepared two large fragments from rhodop- sin by limited proteolysis with thermolysin (Hargrave rt at., 1980). Fragment F2, which originates from the carboxyl-terminal region of rhodopsin, has been sequenced starting from its amino terminus for more than 50 residues in our laboratory (Siemiatkowski- Juszczak. 1981). These results allowed us to extend the known sequence of rhodopsin’s carboxyl-terminal region to 108 amino acids, approximately one-third of the entire protein (Fig. 1). RHODOPSIN IS PHOSPHORYLATED NEAR ITS CARBOXYL-TERMINUS It is well-known that rhodopsin is phosphorylated by a rod-cell kinase in a light-dependent reaction (Kuhn and Dreyer, 1972; Bownds et nt., 1972; Frank et al., 1973). Serine and threonine residues were found to be sites of phosphorylation (Kuhn and Dreyer, 1972) and the composition of a small phosphopeptide was reported (Hargrave er al., 1972). It became clear 1429
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The carboxyl-terminal one-third of bovine rhodopsin: Its structure and function

V~.\ir,u Rc\. Vol. 22. pp. 1429 to 1438. 1982 Printed in Great Bntain
0043-6989,82~121429-I0$03.00~0 Pergamon Pres\ Ltd
THE CARBOXYL-TERMINAL ONE-THIRD OF BOVINE RHODOPSIN: ITS STRUCTURE AND FUNCTION
PAUL A. HARGRAVE’, J. HUGH MCDOWELL’, ELIZABETH C. SIEMIATKOWSK~-JUSZ~ZAK’. SHAO-LING FONG’, HERMANN K~;~HN~; JANET K. WANG’. DONNA R. CURTIS’,
J.K.MOHANA RAO~, PATRICK ARGOSY and RICHARDJ.FELDMANN"
‘Department of Medical Biochemistry (School of Medicine) and Department of Chemistry and Bio- chemistry (College of Science), Southern Illinois University at Carbondale, Carbondale, IL 62901. U.S.A. “Baylor College of Medicine, Department of Ophthalmology, Texas Medical Center. Houston, TX
77025. U.S.A. -‘Institut fur Neurobiologie der Kernforschungsanlage, Postfach 1913, D-5170 Jtilich, West Germany
‘Department of Biological Sciences. Purdue University, West Lafayette. IN 47907. U.S.A. 5Computer Center Branch. Division of Computer Research and Technology, National Institutes of
Health, Bethesda, MD 20205. U.S.A.
Abstract-The covalent sequence of the carboxyl-terminal one-third of bovine rhodopsin has been determined. It is 108 amino acids in length and consists of three hydrophilic regions exposed to the aqueous environment at membrane surfaces and two hydrophobic regions which penetrate the disc membrane lipid bilayer. This carboxyl-terminal third of rhodopsin contains the retinal binding site, sites of phosphorylation, the fast-reacting sulfhydryl group and the thermolytic fragment F2. Hydrophobic helices in the sequence have been predicted by a new method developed for application to membrane proteins and a hypothetical “helical hairpin” space-filling working model has been constructed.
INTRODUCTION
In the first event in vision, rhodopsin’s chromophore, 1 I-cis-retinal, absorbs a photon and is isomerized to the all-trans form. This leads to permeability changes in the plasma membrane of the rod cell and to visuat excitation. To elucidate how the photoreceptor pro- tein rhodopsin functions in affecting rod cell bio- chemistry through response to light, we have con-
ducted studies of rhodopsin’s structure and several of its chemical properties. In this article we summarize the results of our current studies on the structure and functional properties of the carboxyl-terminal region of rhodopsin.
THE SEQUENCE OF THE CARBOXYL-TERMINAL
ONE-THIRD OF RHODOPSIN HAS BEEN
DETERMINED
The carboxyl-terminal amino acid of bovine rho- dopsin has been previously reported to be resistant to identification (Albrecht, 1957; HelIer, 1968). However, application of the method of hydrazinolysis showed alanine to be the carboxyl-terminal amino acid (Har-
grave and Fong. 1977). A nine amino acid tryptic pep- tide which contained alanine at its carboxyl-terminus was isolated and demonstrated to be the carboxyl- terminal peptide of rhodopsin (Hargrave and Fong,
Stereo pictures in this paper can be viewed using an inex- pensive stereoviewer such as that sold by The Hubbard Scientific Co., Northbrook, Illinois (stereoscope).
Abhreciutions: ROS-rod outer segments: des(l’ - la’)rho- dopsin--the rhodopsin molecule minus its 12 carboxyl- terminal amino acids; NAPT-nitroazidophenyl taur- ine; AP--azidopyrene; IASiodoacetamidosalicylate.
1977). Knowledge of the primary structure of this region was extended by the isolation and sequence determination of a 31”amino acid carboxyl-terminal
peptide from the cyanogen bromide cleavage of rho- dopsin (Fong, 1978). Additional peptides were charac- terized which served to extend the carboxyl-terminal sequence to 40 amino acids (Hargrave or ~1.. 1980).
Recently the laboratory of Virmaux at Strasbourg reported the isolation and partial characterization of a large peptide formed by cleavage of rhodopsin at tryptophanyl residues (Pellicone et ul., 1980). This peptide overlapped with our sequence and extended knowledge of this region of the molecule to 83 amino acids. To elucidate further the sequence of rhodopsin we have prepared two large fragments from rhodop- sin by limited proteolysis with thermolysin (Hargrave
rt at., 1980). Fragment F2, which originates from the carboxyl-terminal region of rhodopsin, has been sequenced starting from its amino terminus for more than 50 residues in our laboratory (Siemiatkowski- Juszczak. 1981). These results allowed us to extend the known sequence of rhodopsin’s carboxyl-terminal region to 108 amino acids, approximately one-third of the entire protein (Fig. 1).
RHODOPSIN IS PHOSPHORYLATED NEAR
ITS CARBOXYL-TERMINUS
It is well-known that rhodopsin is phosphorylated by a rod-cell kinase in a light-dependent reaction (Kuhn and Dreyer, 1972; Bownds et nt., 1972; Frank
et al., 1973). Serine and threonine residues were found to be sites of phosphorylation (Kuhn and Dreyer, 1972) and the composition of a small phosphopeptide was reported (Hargrave er al., 1972). It became clear
1429

108’ 96’
,.. Alo Thr . Thr Gin . Lys - Ala - Glu - Lys . Glu . Vol Thr Arg - Met . Vol 118 Ile . Met Vol . Ile Alo - Phe - Leu - ‘\ J
73’ 71’ ’ 66’
ile Xxx . Trp - Leu - Pro - Tyr Ala . Gly . Vol . Ala Phe - Tyr Ile Phe Thr His Gin Gly Ser izp . Phe - Gly .
s3l
Pro - tie . Phe - Met . Thr - lie Pro Alo Phe . Phe Ala Lyt - Thr . Ser - Alo . Vol Tyr Asn . Pro - Vol . Ila . Tyr - \
40’ 39’ 33’ 31’ 11’
Ile - Met - Met - Asn - Lys Gln . Phe - Arg _ Asn - Cys - Met . Vol . Thr - Thr . Leu Cys - Cys . Gly Lys - Asn - Pro ‘I /
1s’ 14’ 13’ II’ II’ 9’ 7’ 61 4’ 1’
Leu . Gly Asp - Asp - Glu Ala . Ser. Thr . Thr - Vol Ser Lys Thr . Glu Thr . Ser. Gln - Vol - Ala Pro. Alo
Fig. I. Sequence of the carhoxyl-terminal one-third of bovine rhodopsin. Until the complete covalent sequence of the protein has been determined, we are numbering the polypeptide chain starting from the carbo~yl-terminal residue as 1’. Amino acids whtch are specifically referred to in the text have their
position in the sequence shown by number.
that phosphoryiation occurred in the carboxyl- terminal region of the molecule when the 32P of ther- molysin-digested 32P-ROS was found to be associated
with fragment F2 (Hargrave and Fong, 1977). Over 854; of the 3zP was found to be located in a 31-amino acid carboxyl-terminal peptide (Fong. 1978; Hargrave
et d., 1980). It is likely that each of the seven amino acids Ser”‘, Thrf4’, Thr13’. Ser”‘. Thr”. Thr” and Se?“ can be phosphory~ated in highly phosphorylated rhodopsin.
RETINAL IS BOLWU TO Lys””
The retinal chromophore of rhodopsin may be reductively linked to the protein by exposing the native protein to fight in the presence of sodium boro- hydride (Bownds and Wald. 1965). This yields a reti- nyl lysine which is located in a region of the amino acid sequence adjacent to amino acids threonine, pro-
line, alanine, isoieucine and phenylalanine (Bownds, 1967). To further elucidate the binding site, rhodopsin was bleached and then regenerated with C3H]1 I-cis retinal. digested in the memb~dne with thermoIysit1 to
form the Fl-,F2 complex, which was then reduced with sodium borohydride (Wang et cd., 1980). This and an earlier report (Pober and Stryer. 1975) demon- strate that the site of retinal attachment is located in F2. When chromatographically purified [3H]retinal rhodopsin in detergent solution was reduced, and the [3~]retinyl-opsin digested with pronase. a dipeptide Aia-[3H]retinyl-Lys was recovered (Wang it al., 1980). Only a single Ala-Lys is present in F2. the lysine 53 amino acids from the carboxyl terminus. This Lyss3’ is surrounded by the amino acids in the composition of retinyl-peptides reported by Bownds (1967) (see Fig. 1) and thus seems securely identified.
CYSTEINE”’ IS HIGHLY REACTIVE
A variety of reagents may be used to selectively
modify cysteine sulfhydryl groups of rhodopsin in the membrane (deGrip ef ul., 1975). Alkylating agents such as iodoacetic acid and iodoacetamidosalicylate
(IAS) appear to show some specificity. incorporating only a single mole of reagent per mole of rhodopsin (McDowell ~1 trl.. 1979). IAS was found to modify principally Cy?“’ (McDowell and Griffith, 1978). Although two cysteines may be modified by reaction of the reagent l~“ethy~maleimide with rhodopsin in the dark. and one more mole of cysteine reacts follow- ing illumination (McDowell ct ~1.. 1979), these two additional cysteines arc not located in the 1’.-108’ sequence (McDowell and Hargrave. unpublished ex- periments). Only under denaturing conditions have we found that additional cysteines in this region of the molecule (presumably Cys’“’ and/or Cys2”) can be modified by alkylating agents.
Limited proteolysis has proven useful in the study of rhodopsin’s structure and topography and is becoming useful in probing its functions. Proteolysis with thermolysin. first reported by Saari (1974). was shown by Pober and Stryer (1975) to yield a large glycopeptide fragment (Fl) and a smaller fragment. F2. When an amino-terminal peptide of rhodopsin was shown to contain oligosaccharide attachment sites (Hargrave. 1977). this placed Fl at the amino terminus and F2 at the carboxyl-terminus. Hargrave and Fong (1977) found that the earliest proteolytic event, prior to fornlatjon of FI and F2. was digestion at rhodopsin’s carboxyl-terminus releasing the two soluble peptides Val-Ala-Pro-Ala and Val-Ser-Lys- Thr-Glu-Thr-Ser-Cln. This resulted from proteolysis at the bonds Thr’3’iVal’2 and Gln”‘LVai4’, in accord with the known preference of thermolysin for cleavage on the amino-terminal side of residues with

The carboxyl-terminal one-third of bovine rhodopsin
hydrophobic side chains. Further sites of proteolysis
became apparent when fragment F2, prepared by more extensive proteolysis of ROS membranes, was
isolated and characterized (Siemiatkowski-Juszczak, 1981). The amino-terminus of F2 was shown to be Ala”” and the carboxyl-terminus Pro”’ (Fig. I), thus demonstrating cleavage of the bonds Xxx109’~Ala’08‘
and Pro2”LLeu2”.
PROTEOLYSIS SHOWS A RHODOPSIN SURFACE
CONFORMATIONAL CHANGE FOLLOWING
METARHODOPSIN I
Several lines of evidence suggest that rhodopsin undergoes a change in conformation following ex-
posure to light. For example, several ROS ~ripheral membrane proteins undergo binding to ROS mem- branes following exposure of the membranes to light (Kiihn, 1978, 1980). When rhodopsin in ROS is digested with thermolysin producing the spectrally intact Fl--F2 complex, the light-dependence of
GTPase binding to the membranes is destroyed (Kiihn and Hargrave, 1981). These observations sug- gest that rhodopsin is changing in response to light to allow protein binding to occur, and that the sites for the binding are located on rhodopsin’s cytoplasmic surface. Rhodopsin’s light-dependent phosphorylation
may also be explained by conformational change. Rhodopsin can only be phosphorylated following illumination, and the kinase is not itself light depen- dent (McDowell and Kiihn, 1977). Therefore it seems reasonable that rhodopsin must undergo a change in conformation which would then make it a substrate for the kinase. Since one of the sites of cleavage of thermolysin is at Val”’ (see previous section) which is located within the phosphorylatable carboxyl- terminal region, we investigated the use of thermoly- sin as a probe for a light-dependent change in the conformation of that region. It was found that con- version of membrane bound rhodopsin to des(1’ - 1~)rhodopsin occurs - 1.7 times faster when rhodopsin has been illuminated than when it has not (Kiihn et al., 1982). This enhanced susceptibility to proteolysis following illumination presumably results from a change in conformation which affects the car- boxyl-terminal region. Using special conditions in which Metarhodopsin I or Metarhodopsin II, respect- ively, were the predominant photoproducts, we found no difference in the proteolytic susceptibility of Metarhodopsin I compared to unbleached rhodopsin, but an increase of - 1.5 times for Metarhodopsin II compared to unbleached rhodopsin. Apparently the change in conformation leading to enhanced suscepti-
bility to proteolysis and interaction with the kinase, must occur after the decay of Metarhodopsin I.
RHODOPSIN’S CARBOXYL-TERMINAL THIRD
CONTAINS BOTH HYDROPHII~IC AND
HYDROPHOBIC REGiONS
Rhodopsin has been chemically modified in disk
Y.H. 22!12 -”

PALL A. HARGRAVI ct trl
Fig. 3. A model for the arrangement of rhodopsin thermolytje fragment F.2 {residues 12’ l(W) in the membrane lipid bilayer. Use of circles and diamonds to designate amino acid polarity is the same as rn
Fig. 2.
membranes with the highly reactive membrane-imper- meant nitrene formed from nitroazidophenyltaurine (,NAPT) (Mas et al., 1980). Thermolytic fragment F2 becomes labeled by NAPT and several peptides. in- cluding the carboxyl-terminal peptides 1’ 3 1’ and 32’.-39’. have been shown to incorporate the reagent. This suggests that the peptide sequence I’-39’ as well as other portions of the carboxyl-terminal region are aqueous accessible and located at the disc cytoplas- mic surface. F2 is also labeled by a hydrophobic re-
agent, azidopyrene (AP) (Smith et cd.. 1981). This re- agent is restricted to the membrane lipid bilayer and reacts with molecular species within it (Smith et ul..
1981). The results of this study with AP, and that of a study with a similar reagent (Klip et al.. 1976) provide chemical evidence that a significant portion of the sequence of F2 is embedded in the lipid bilayer. The hydrophobic reagent AP appears to react with regions of rhodopsin not labeled by the hydrophilic reagent NAPT and vice versa. It thus appears that both hydrophilic and hydrophobic domains exist within the sequence of the carboxyl-terminal one- third of rhodopsin.
When the rhodopsin sequence l’-108’ is coded to emphasize which residues are hydrophilic and which
are hydrophobic, it becomes clear that the sequence contains two predominantly hydrophobic stretches (Fig. 2). A 24-amino acid region (73’ -96’) and a 27-amino acid region (40’.-66’) are flanked by blocks rich in hydrophilic amino acids. This information, in conjunction with the chemical and enzymatic data presented above. suggests a model for the arrange- ment of this sequence in the membrane as shown in
Fig. 3. Ala I’*’ and Pro”’ are placed at the membrane
cytoplasmic surface due to their accessibility to the macromolecular probe thermolysin. In addition. the sequence 1’.-39’ is reactive toward the water-soluble reagent NAPT. thereby also placing it external to the lipid bilayer. The hydrophobic sequences 40’~66’ and 73’ 96’ are shown traversing the lipid bilayer. con- nected by the hydrophilic segment 67-72’ which is shown in contact with the intradisc membrane aqueous surface. The lengths of the proposed mem- brane-spanning segments are appropriate based upon information available from studies on bacteriorho- dopsin (Engelman et al., 1980) and M 13 coat protein (Wickner. 1976). This model predicts that the site of retinal attachment, Lys’“‘. is located near the middle of the lipid bilayer. at the midpoint of transmembrane segment segment 40’66’.
The carboxyl-terminal one-third of bovine rhodopsin 1433
5.0
4.0
3.0
2.0
r r
E Ix) L :: z 0
.I! z = Q -1.0
z
E 3 -2.0 v)
-3.0
-4.0
- 5.0
I-
T
0 20 40 60 SO /
100 120 140 I60 180 200 220 240 260 Amino acid number
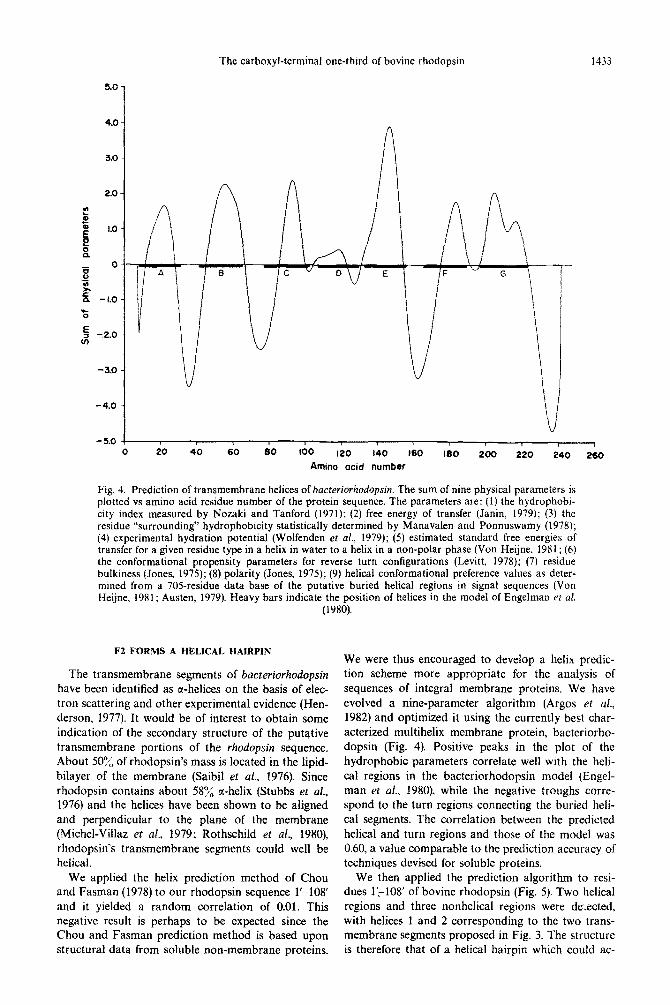
Fig. 4. Prediction of transmembrane helices of bacreriorhodopsin. The sum of nine physical parameters is plotted vs amino acid residue number of the protein sequence. The parameters are: (I) the hydrophobi- city index measured by Nozaki and Tanford (1971); (2) free energy of transfer (Janin, 1979); (3) the residue “surrounding” hydrophobicity statistically determined by Manavalen and Ponnuswamy (1978); (4) experimental hydration potential (Wolfenden et al., 1979); (5) estimated standard free energies of transfer for a given residue type in a helix in water to a helix in a non-polar phase (Von Heijne, 1981; (6) the conformational propensity parameters for reverse turn configurations (Levitt, 1978); (7) residue bulkiness (Jones, 1975); (8) polarity (Jones, 1975); (9) helical conformational preference values as deter- mined from a 70Sresidue data base of the putative buried helical regions in signal sequences (Von Heijne, 1981; Austen, 1979). Heavy bars indicate the position of helices in the model of Engelman et al.
(1980).
F2 FORMS A HELICAL HAIRPiN
The transmembrane segments of bacteriorhodopsin have been identified as a-helices on the basis of elec- tron scattering and other experimental evidence (Hen- derson, 1977). It would be of interest to obtain some indication of the secondary structure of the putative transmembrane portions of the rhodopsin sequence. About 50% of rhodopsin’s mass is located in the lipid- bilayer of the membrane (Saibi! ef al., 1976). Since rhodopsin contains about 58% a-helix (Stubbs et al., 1976) and the helices have been shown to be aligned and ~rpendicular to the plane of the membrane (~ichel-Villaz et at., 1979; Rothschild et al., 1980), rhodopsin’s transmembrane segments could well be helical.
We applied the helix prediction method of Chou and Fasman (1978) to our rhodopsin sequence l’-108’ and it yielded a random correlation of 0.01. This negative result is perhaps to be expected since the Chou and Fasman prediction method is based upon structural data from soluble non-membrane proteins.
We were thus encouraged to develop a helix predic- tion scheme more appropriate for the analysis of sequences of integral membrane proteins. We have evolved a nine-parameter algorithm (Argos et al., 1982) and optimized it using the currently best char- acterized multihelix membrane protein, bacteriorho- dopsin (Fig. 4). Positive peaks in the plot of the hydrophobic parameters correlate well with the heli- cal regions in the bacteriorhodopsin model (Engel- man er al., 1980), while the negative troughs corre- spond to the turn regions connecting the buried heli- cal segments. The correlation between the predicted helical and turn regions and those of the model was 0.60, a value comparable to the prediction accuracy of techniques devised for soluble proteins.
We then applied the prediction algorithm to resi- dues 1:-108’ of bovine rhodopsin (Fig. 5). Two helical regions and three nonhelical regions were de.ected, with helices 1 and 2 corresponding to the two trans- membrane segments proposed in Fig. 3. The structure is therefore that of a helical hairpin which could ac-

AMINO ACID NUMBER
Fig. 5. Predictiofl of tmnsmembrane helices for the bovine rhodopsin sequence 1’~ 104’. The sum of physical par-
ameters plotted is described in the legend to Fig. 3.
Fig. 6. Stereo drawings of a skeletal working model for the structure of bovine rhodopsin thermolytic fragment F2. The sequence 22’ 108’ was arranged so that residues 73’.-96’ (helix I) and 4Whh’ (helix 2) were given cp angles (amide N C 2) and $ angles (C-Y L-arhonyl C) for an cx helix. d, and ti angles of residues 97’ 10X’ and 22’-39’ are arbitrary. The peptide chain was severed at 66’.- 67’ and the two helices were ahgned. Q and r(, angles for the regment 67’--72’ were adjusted in an attempt to reform the peptide
chain. A firont view is shown as in Fig. XA.
A
Fig. 7. Helices of bovine rhodopsin F2: (A) the skeletal model of Fig. 6 is viewed by looking from above directly down the axes of the helices; (B) helical wheel representations of F2 heliccs I and 2 (Schiffer and Edmundson, 1967). The wheel is the projection of all amino acid side chain positians aiong the helix axis onto a plane. The aromatic residues of helix 1 are circled and the arrow ghows the helix face where most are located. The hydrophobic residues of helix 2 are circled and the arrow shows the 180 arc of their
predominant location. 1434

1435


The carboxyl-terminal one-third of bovine rhodopsin 1437
comphsh spontaneous ~netration of the hydrophobic portion of a membrane bilayer as proposed by Engel- man and Steitz (1981). It is of interest to note that both helix G of bacteriorhodopsin and helix 2 of bovine rhodopsin F2 exhibit the same discontinuity in the predictive parameters (see Fig. 5). Both helices contain the site of retinal attachment; Lys2i6 in bac- teriorhodopsin (Bayley et al., 1981) and Lyss3’ in bovine rhodopsin (Wang et al., 1980). It is interesting to speculate that a special environment may exist in the lysine vicinity which may be important in func- tions of the protein following bleaching.
A THREE-DIMENSIONAL WORKING MODEL FOR F2
To visualize better how the amino acids in F2 helices 1 and 2 might be arranged with respect to one another in space, a stereo skeletal model of the sequence was generated (Fig. 6). The structure is hy- pothetical; currently unavailable X-ray crystallo- graphic data would be required in order to fix the positions of atoms in space within the helices and the relative relationships of the helices. The “front” view of fragment F2 (Fig. 6) shows it as a helical hairpin which would penetrate the membrane, reverse direc- tion and traverse the membrane again. The view from the top down the axis of the helices (Fig. 7A), shows that the easily identifiable aromatic side chains are not uniformly distributed about the surface of the helices. When the sequences of both helices are exam- ined by presentation as helical wheels (Fig. 7B), it is observed that helix 1 has 5 of its 6 aromatic residues in one arc and that helix 2 has 12 of 15 of its most hydrophobic residues on one 180” surface. It can only be surmised that such differences in polarity in helix surfaces must reflect requirements for helix-helix and helix-lipid interactions.
The skeletal protein models of F2 were converted to a space-filling form in which each of the atoms was represented by a sphere of appropriate radius (Feld- mann et al., 1981). Various views of F2 are presented in Fig. 8. It is hoped that such concrete presentations of proposed secondary structure for rhodopsin will assist in future studies on the protein to understand the environment of retinal as well as suggest experi- ments designed to test specific models.
Acknnwirdyements-This work has been supported by grants EY 1275 and EY27S5 from the National Institutes of Health and NATO grant RG076.80 (to P.A.H.). P.A. and J.K.M. acknowledge-support from the National Institutes of Health (GM 27682) and the American Cancer Society (FRA 173).
REFERENCES
Albrecht G. (1957) Terminal amino acids of rhodopsin. 3. hiol. Chem. 229, 477-487.
Argos P., Mohana Rao J. K. and Hargrave P. A. (1982) Structural prediction of memnrane-bound proteins. Eur. J. Biochem. To be published.
Austen B. M. (1979) Predicted secondary structures of
amino-terminal extension sequences of secreted proteins. FEBS L&t. 103, 308-313.
Bayley H., Huang K.-S.. Radhakrishnan R., Ross A. H., Takagaki Y. and Khorana H. G. (1981) Site of attach- ment of retinal in bacteriorhodopsin. froc. natn. Acad. Sci. U.S.A. 78, 2225-2229.
Bownds D. and Wald G. (1965) Reaction of the rhodopsin chromophore with sodium borohydride. Nature 205, 254257.
Bownds D. (1967) Site of attachment of retinal in rhodop- sin. Nature 216, 1178-1181.
Bownds D., Dawes J.. Miller J. and Stahlman M. (1972) Phosphorylation of frog photoreceptor membranes induced by light. Nature he; Eiol. 237, 125-127.
Chou P. Y. and Fasman C. D. (1978) Prediction of the secondary structure of proteins from their amino acid sequence. In Adwtcrs in Enzymology (Edited by Meister A.), Vol. 47, pp. 45-148. Wiley, New York.
deGrip W. J.. Bonting S. L. and Daemen F. J. M. (1975) Biochemical aspects of the visual process. XXVIII. Classification of sulfhydryl groups in rhodopsin and other photoreceptor membrane proteins. Biochim. bio- phys. Acta 396, 104-l 15.
Engelman D. M. and Steitz T. A. (1981) The spontaneous insertion of proteins into and across membranes: the helical hairpin hypothesis. Cell 23, 411-422.
Engelman D. M., Henderson R.. McLachlan A. D. and Wallace B. A. (19801 Path of the polypeptide in bacterio- rhodopsin. Proc. tram. Acad. Sci. U.S.A. 77, 2023-2027.
Feldmann R. J., Potter M. and Glaudemans C. P. J. (1981) A hypothetical space-~lling model of the V-regions of the galactan-binding myeloma immunoglobuiin 5539. M&c. ~?~~zr~}~. 18, 683-69X.
Fong S.-L. (1978) Partial structural characterization of bovine rhodopsin-a photoreceptor protein. Ph.D. dis- sertation. Southern Illinois University.
Frank R. N., Cavanagh H. D. and Kenyon K. R. (1973) Light-stimulated phosphorylation of bovine visual pig- ments by adenosine triphosphate. J. hiol. Chem. 248, 596609.
Hargrave P. A. (1977) The amino-terminal tryptic peptide of bovine rhodopsin. A glycopeptide containing two sites of oligosaccharide attachment. Biochim. hiophys. Acta 492, 83-94.
Hargrave P. A. and Fong S.-L. (1977) The amino- and carboxyl-terminal sequence of bovine rhodopsin. J. .supramolec Struct. 6. 559-570.
Hargrave P. A., Kuhn H. and Dreyer W. J. (1972) Light- dependent phosphorylation of rhodopsin by ATP. Fedn Proc. Fedn Am. Sots exp. Biol. 31, 462.
Hargrave P. A.. Fong S.-L., McDowell J. H.. Mas M. T., Curtis D. R.. Wang J. K.. Juszczak E. and Smith D. P. (1980) The partial primary structure of bovine rhodopsin and its topography in the retinal rod cell disc membrane. Nwrochem. Int. 1, 231-244.
Heller J. (1968) Structure of visual pigments. 1. Purification, molecular weight and composition of bovine visual pigment,,,. Biochemistry 7, 2906-2913.
Henderson R. (1977) The purple membrane from Halobac- trrium ~~~~obi~rn. Ann. Rw. kophys. Bioengng 6, 87- 109.
Janin J. (1979). Surface and inside volumes in alobular oro- teins. Matar<> 277, 491-492.
Jones D. D. (1975) Amino acid properties and side-chain orientation in proteins: A cross correlation approach. J. &or. Biol. 50, 167-f 83.
Klip A.. Darszon A. and Montal M. (1976) Labelling of rhodopsin moieties confined to the membrane lipid bilayer. Biochem. hiophys. Res. Commun. 72. 1350-1358.
Kuhn H. (1978) Light-regulated binding of rhodopsin kinase and other proteins to cattle photoreceptor mem- branes. Biochemistry 17, 4389-4395.
Kuhn H. (1980) Light- and GTP-regulated interaction of GTPase and other proteins with bovine photoreceptor membranes. Nrrturr 283, 587-589.

1438 PAUL A. HARGRAVE et al.
Kuhn H. and Dreyer W. J. (1972) Light dependent phos- phorylation of rhodopsin by ATP. FEES Lett. 20, 1-6.
Kuhn H. and Hargrave P. (1981) Light-induced binding of guanosinetriphosphatase to bovine photoreceptor mem- branes: effect of limited proteolysis of the membranes. Biochemistry 20, 241@2417.
Kuhn H.. Mommertz 0. and Harnrave P. A. (1982) Light- dependent ~onformational change at rhodopsin’s cite- plasmic surface detected by increased susceptibility to proteolysis. Biochim. hiophys. Acta 679, 95-100.
Levitt M. (1978). Conformational preference of amino acids in globular proteins. Biochemistry 17, 4277.--4285.
Manavalan P. and Ponnuswamy P. K. (1978) Hydrophobic character of amino acid residues in globular proteins. Narurr 275, 673.-674.
Mas M. T., Wang J. K. and Hargrave P. A. (1980) Topo- graphy of rhodopsin in rod outer segment disk mem- branes. Photochemical labeling with N-(4-Azido-2-nitro- phenyl)-2-aminoethanesulfonate. Biochemistry 19, 684-692.
McDowell .I. H. and Kuhn H. (1977) Light-induced phos- phorylation of rhodopsin in cattle photoreceptor mem- branes: substrate activation and inactivation, Biochemis-
try 16, 40544060. McDowell J. H. and Griffith K. (1978). Localization of a
dark reactive cysteine in rhodopsin. Incest. Ophthal. vis- ml sci. 17, Suppl. 1, 125.
McDowell J. H.. Mas M. T., Griffith K. D. and Hargrave P. A. (1979) The reactivity of the suhhydryl groups of rhodopsin in rod outer segment membranes. Vision Res. 19, 1143-l 145.
Michel-Villaz M., Saibil H. R. and Chabre M. (1979) Orientation of rhodopsin r-helices in retinal rod outer segment membranes studied by infrared linear dich- roism. Proc. n&n. Acad. Sci. U.S.A. 76, 4405-4408.
Nozaki Y. and Tanford C. (1971) The solubility of amino acids and two glycine peptides in aqueous ethanol and dioxane soluttons. f. hinl. Chem. 246, 221 I-221 7.
Pelticone C., Bouillon P. and Virmaux N. (1980) Purifica- tion et sequence partielle dun polypeptide hydrophobe de la rhodopsine bovine fragmentee par le BNPS-sca-
tole. C.r. hehd. SZanc. Acad. Sci.. Paris Ser. D 290, 567-569.
Pober J. S. and Stryer t. (1975) Light dissociates enzymati- catly-cleaved rhodopsin into two different fragments. J. molec. Biol. 95, 477-481.
Rothschild K. J., Sanches R., Hsiao T. L. and Clark N. A. (1980) A spectroscopic study of rhodopsin alpha-helix orientation. ~ioph~s. J. 31, 53-64.
Saari J. C. (1974) The accessibility of bovine rhodopsin m photoreceptor membranes. J. Cell Biol. 63, 480-491.
Saibil H., Chabre M. and Worcester D. (1976). Neutron diffraction studies of retinal rod outer segment mem- branes. Nature 262, 26& 270.
Schiffer M. and Edmundson A. B. (1967) Use of helical wheels to represent the structures of proteins and to identify segments with helical potential. Bioplty.s. J. 7, 121~135.
Siemiatkowski-Juszczak E. C. (1981) Limited proteolysis of bovine rhodopsin and sequence determination of the F2 fragment. Ph.D. dissertation. Southern Illinois Univer- sity.
Smith D. P., Kilbourn M. R., McDowell J. H. and Har- grave P. A. (1981) Topography of rhodopsin in retinal rod outer segment disk membranes. Photochemical labeling with 1-azidopyrene. Biochemistry 20, 2417 -2424.
Stubbs G. W., Smith H. G. and Litman B. J. (1976). Alkyd glucosides as effective soiubilizing agents for bovine rho- dopsin. A comparison with several commonly used detergents. Eiochim. hiophqs. Acta 426, 4656.
Von Heijne G. (1981) On the hydrophobic nature of signal sequences. Eur. J. &o&m. 116, 419-422.
Wang J. K.. McDowell J. H. and Hargrave P. A. (1980) Site of attachment of 1 I-cis-retinal in bovjne rhodopsin. Bio- chemistr_r 19, 511 I-5I 17.
Wickner W. (1976) Asymmetric orientation of phage Ml3 coat protein in Escherichia w/i cytoplasmic membranes and in synthetic lipid vesicles, Proc. nutn A<,crd. Sci. U.S.A. 73, 1159-1163.
Wolfenden R. V., Cullis P. M. and Southgate C. C. F. (1979) Water, protein Folding and the genetic code. Science 206, 575-577.