
The 5HT 2A serotonin receptor enhances cell viability, affects cell cycle progression and activates...
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of The 5HT 2A serotonin receptor enhances cell viability, affects cell cycle progression and activates...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
The 5-HT2A serotonin receptor enhances cell viability, affects cell cycleprogression and activates MEKeERK1/2 and JAK2eSTAT3 signallingpathways in human choriocarcinoma cell lines
T. Oufkir, M. Arseneault, J.T. Sanderson, C. Vaillancourt*
INRS-Institut Armand-Frappier, Université du Québec, Laval, QC H7V 1B7, Canada
a r t i c l e i n f o
Article history:Accepted 25 February 2010
Keywords:Serotonin5-HT- receptorJAKeSTATMAPKJEG-3BeWoCell proliferationCell cyclePlacenta
a b s t r a c t
Previous results from our group have demonstrated the expression of the 5-HT2A receptor and amitogeniceffect of serotonin in human trophoblast. The objectives of the present studywere to investigate the role ofthe 5-HT2A receptor in trophoblast cells and to determine the signalling pathways activated by thisreceptor. We investigated the effect of (�)-2,5-dimethoxy-4-iodoamphetamine hydrochloride (DOI),a selective 5-HT2A agonist, on cell cycle progression and cell viability in BeWo and JEG-3 cells. We alsoinvestigated, by co-immunoprecipitation and western blot analysis, the involvement of the MEKeERK1/2and JAK2eSTAT3 signalling pathways following activation of the placental 5-HT2A receptor. Our resultsshowed a concentration-dependent increase of cell viability by DOI, which was reversed by ketanserin,a selective 5-HT2A receptor antagonist. Furthermore, activation of the 5-HT2A receptor by DOI increasedcell entry into the G2/M and S phase (DNA synthesis) in BeWo and JEG-3 cells, respectively. In addition,stimulation of BeWo and JEG-3 cells by DOI activated both the MEKeERK1/2 and the JAK2eSTAT3 sig-nalling pathways. This study demonstrated that the 5-HT2A receptor increases cell viability and affects cellcycle progression in human trophoblast cell lines as well as activates the MEKeERK1/2 and JAK2eSTAT3intracellular signalling pathways, which are related to survival, differentiation, migration and invasion.These findings indicate that serotonin through the activation of the 5-HT2A receptor is a key regulator ofplacentation and may play a role in the pathophysiology of certain pregnancy disorders associated withalterations in placental development, such as preeclampsia, gestational diabetes and preterm birth.
Crown Copyright � 2010 Published by Elsevier Ltd. All rights reserved.
1. Introduction
Serotonin (5-hydroxytryptamine, 5-HT) is produced and storedin several tissues, including the central nervous system (CNS),blood platelets and gastrointestinal enterochromaffin cells [1]. Thismonoamine acts as a neurotransmitter, hormone and growth factorto regulate a wide range of cellular processes, such as cell differ-entiation, cell growth, cell proliferation, gene expression, apoptosisand cell survival [2,3]. Serotonin regulates many behavioural andphysiological processes and acts as a developmental signal in earlyembryogenesis and regulation of fetal development and pregnancywell-being [3e6]. Studies have suggested that serotonin is involvedin placental development (placentation) and function [7e10].
Serotonin exerts its biological activities by binding to multipleserotonin receptor classes. Among these, the 5-HT2 class comprises
three subtypes: 5-HT2A, 5-HT2B and 5-HT2C [11]. The 5-HT2Areceptor subtype is widely distributed in peripheral tissues,including the human trophoblast [8]. This serotonin receptorsubtype has been shown to regulate cell differentiation, prolifera-tion and migration in different cell types including glomerularmesangial, skeletal muscle and vascular smooth muscle cells[12e15]. Stimulation of the 5-HT2A receptor may exert its effect viathe janus kinases/signal transducer and activator of transcription(JAKeSTAT) and extracellular signal-regulated kinase 1/2 (ERK1/2)pathways [13,14,16]. Direct activation of JAK2eSTAT3 throughstimulation of the 5-HT2A receptor has been demonstrated toenhance cell differentiation and proliferation in rat skeletal musclecells [13]. To date, no study has characterized the signal trans-duction pathways initiated by serotonin via its 5-HT2A receptor introphoblast cells.
Both the mitogen-activated protein kinases (MAPKs) and theJAK/STAT pathways are implicated in cell proliferation, differentia-tion, survival, and apoptosis [17,18]. The JAK2-STAT3 and ERK1/2(p42/44-MAPK)pathwayshavebeen shown totargetgenes involvedin the regulation of cell proliferation and viability in different cell
* Corresponding author. INRS-Institut Armand-Frappier, 531 blvd des Prairies,Laval, QC H7V 1B7, Canada. Tel.: þ1 450 687 5010 x8812; fax: þ1 450 686 5309.
E-mail address: [email protected] (C. Vaillancourt).
Contents lists available at ScienceDirect
Placenta
journal homepage: www.elsevier .com/locate/placenta
0143-4004/$ e see front matter Crown Copyright � 2010 Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.placenta.2010.02.019
Placenta 31 (2010) 439e447

Author's personal copy
types including the trophoblast [19,20]. Moreover, activation ofERK1/2 promotes villous cytotrophoblast differentiation (syncyti-alization) [21,22], while STAT3 is involved in extra-villous cyto-trophoblasts invasion [23,24]. Activationof ERK1/2pathwayhas alsobeen shown to increase extra-villous cytotrophoblast invasion andmigration [25,26]. Taken together, these data suggest that the 5-HT2A receptor regulates placental trophoblast cellular processesthrough the activation of JAK-STAT and/or MAPK pathways.
Previously we demonstrated the expression of the 5-HT2Areceptor in human primary villous trophoblast and in BeWo andJEG-3 trophoblastic-like cell lines [7,8]. In this study, in order toexplore the signalling mechanism and the role of the 5-HT2Areceptor in trophoblast cells, we evaluated the effect of (�)-2,5-dimethoxy-4-iodoamphetamine hydrochloride (DOI) on cellviability, cell cycle progression and activation of the MEKeERK1/2and the JAK2eSTAT3 pathways in BeWo and JEG-3 cell lines.
2. Materials and methods
2.1. Materials
Minimum essential medium Eagle's (MEM Eagle), nutrient mixture F12 HamKaighn's modification (Ham's F-12K), MEM/F-12K, antibiotics (penicillin andstreptomycin), sodium pyruvate, trypsin, ethylenediaminetetraacetic acid (EDTA),trypsin I, protease and phosphatase inhibitor cocktails, acrylamide/bis-acrylamidesolution, (�)-2,5-dimethoxy-4-iodoamphetamine hydrochloride (DOI), ketanserin,and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reagent,were all purchased from SigmaeAldrich (Oakville, ON, Canada). Fetal bovine serum(FBS) was obtained fromHyclone Laboratories (Logan, UT, USA). RNasewas obtainedfrom Roche Diagnostics (Laval, QC, Canada). MACS� separation columns and mMACSProtein G microbeads were obtained from Miltenyi Biotec (Auburn, CA, USA). Cell-Bind T75- and T25- flasks and 6- and 96-wells microplates were purchased fromCorning Life Science (Corning, NY, USA). Bicinchoninic acid (BCA) protein assayreagent was obtained from Pierce Biotechnology (Rockford, IL, USA). Polyvinylidenedifluoride (PVDF) membranes, horseradish peroxidase (HRP)-conjugated secondaryantibodies, Immobilon Western Chemiluminescent HRP Substrate, and Re-Blot PlusMild Antibody Stripping Solution were obtained from Millipore (Mississauga, ON,Canada). Polyclonal antibodies against 5-HT2A (H-75), JAK2 (sc-34479) and phos-pho-MEK1/2 (sc-7995) for western blot analysis were obtained from Santa CruzBiotechnology (CA, USA), while those against MEK1/2 (05-747), phospho-ERK1/2(05-481), ERK1/2 (06-182), STAT3 (06-596), and phospho-STAT3 (Tyr-705) (05-485)were purchased from Millipore. Monoclonal antibody against glyderaldehyde-3-phosphate dehydrogenase (GAPDH, MAB374) was purchased from Millipore. Allother reagents and chemicals were purchased from SigmaeAldrich or FisherScientific (Ottawa, ON, Canada).
2.2. Cell culture
BeWo and JEG-3 human placental choriocarcinoma cell lines were obtainedfrom the American Type Culture Collection (ATCC) (Rockville, MD, USA). BeWo cellswere maintained in MEM/Ham's F-12K containing 10% FBS, 2 mM L-glutamine,100 U/mL penicillin and 100 mg/mL streptomycin. JEG-3 cells were maintained inMEM Eagle containing 10% FBS, 2 mM L-glutamine, 1 mM sodium pyruvate,100 U/mL penicillin and 100 mg/mL streptomycin. Cells were maintained in 75 cm2
culture flasks in a humidified atmosphere of 5% CO2 at 37 �C. Cells were split whenthey reached about 90% confluence, using 0.5% trypsin.
2.3. MTT cell viability assay
The conversion of MTT to formazan, a measure of the mitochondrial reductiveactivity of the cell was used to determine the effect of serotonin, DOI and ketanserinon human placental cell viability. BeWo and JEG-3 cells were seeded in 96-wellplates at a density of 5�103 cells/well (final volume of 100 mL/well). After 24 h, thecells were treated with increasing concentrations of DOI (0, 5, 10, 15, 20, 25, and40 mM) or with 0.1 mM ketanserin (selective 5-HT2A antagonist) before addition of20 mM (BeWo) and 25 mM (JEG-3) DOI or with 10 mM ketanserin before addition of20 mM (BeWo) and 40 mM (JEG-3) serotonin. The experiment was realized in pres-ence of 0.5% (DOI) or 10% FBS (serotonin) and DOI and serotonin were diluted infresh medium containing 0.5% or 10% FBS respectively. After 48 h of incubation at37 �C in 5% CO2, the culture mediumwas removed and 8 mL MTT reagent was addedto 50 mL of fresh culture medium at a final concentration of 0.64 mg/mL. Cells wereincubated for 4 h at 37 �C in 5% CO2. After incubation, medium containing MTT wasremoved and the resulting formazan crystals were dissolved in 120 mL of MTTdetergent solution (95% isopropanol, 1% HCl 12 N, and 4% Triton X-100) by pipettingup and down 30 times. The absorbance was measured spectrophotometrically at
a wavelength of 570 nm (with a reference wavelength of 690 nm) using a Spec-traMax M5 with SoftMax Pro v5 software (Molecular Devices, Sunnyvale, CA, USA).Each assay included blank wells (culture medium without cells) and control wells(culture medium with cells). Cell viability (% of control) was expressed using thefollowing formula:
�Að570 nm-690 nmÞ of each treated well=Að570 nm�690 nmÞ average of control wells
�
� 100
2.4. Cell cycle analysis by flow cytometry
BeWo and JEG-3 (2.5�105 cells) were transferred to T25 flasks. Forsynchronization, cells were kept for 24 h in culture medium supplemented with0.5% FBS and then were incubated with 20 mM (BeWo) and 25 mM (JEG-3) DOI for48 h. Cells were fixed using ice-cold 70% ethanol/30% PBS for at least 1 h on ice. Fixedcells were harvested by centrifugation and resuspended in 800 mL of PBS. Aftera second wash in PBS, 20 mL of RNase (10 mg/mL) and 100 mL of propidium iodide(400 mg/mL) were added. DNA content was then assessed using a Becton DickinsonFACSCalibur Flow Cytometer (Becton Dickinson Inc, Oakville, ON, Canada). Propi-dium iodide was excited at 488 nm and fluorescence was read at 640 nm. Cells weregated bymeasuring forward and orthogonal light scatter and a DNA histogram of cellnumber against fluorescence was obtained. DNA histograms were analyzed usingWinMDI (multiple document interface for windows), Version 2.9 for flowcytometry.
2.5. Protein extraction
After FBS deprivation (0.5% vs. 10% FBS) for 12 h, BeWo and JEG-3 cells werestimulated for 0, 5, 15, 30, and 60 min with 20 mM and 25 mM of DOI (diluted inmedium containing 0.5% FBS) respectively. For total protein extraction, cells werewashed twice with cold PBS, harvested by scraping in 500 mL of cold lysis buffer(50 mM TriseHCl pH 7.4, 175 mM NaCl, 1 mM EDTA) containing 5 mL ProteaseInhibitor Cocktail and 5 mL Phosphatase Halt� Inhibitor Cocktail and transferred toa microcentrifuge tube. The lysates were agitated for 30 min at 4 �C and centrifugedfor 15 min at 21000� g at 4 �C. The supernatants (total proteins) were carefullyremoved and stored at �80 �C until analysis. Protein concentrations were measuredusing theBCAprotein assay reagent according to the instructionsof themanufacturer.
2.6. Co-immunoprecipitation
For the co-immunoprecipitation analyses, 100 mg of total protein lysates weremixed with 3 mL of anti-JAK2 or anti-5-HT2A antibodies and 50 mL of proteinG-conjugated microbeads (Miltenyi Biotech). The lysates were then incubated for30 min on ice and applied to the MACS� separation columns in a magnetic field(ThermoMACS� Separation Unit Miltenyi Biotech). The columns were washed withhigh salt solution (500 mM NaCl in lysis buffer) followed by four washes with lowsalt buffer (TriseHCl 50 mM pH 7.4, 137 mM NaCl, 0.1% Triton X-100). Boundproteins were eluted using SDS buffer (50 mM TriseHCl pH 6.8, 50 mM DTT, 1% SDS,0.005% bromophenol blue, 10% glycerol) pre-heated to 95 �C and stored at �80 �C.
2.7. Immunoblotting
Total proteins diluted in SDS buffer were heated for 10 min at 95 �C for dena-turation. Then, total and immunoprecipitated proteins were fractionated by SDS-polyacrylamide gel electrophoresis on 10% separating and 4% stacking gels, followedby transfer to PVDF membranes. The membranes were blocked in TBS-T (TBS:20 mM TriseHCl pH 7.4, 150 mM NaCl, 0.05% (v/v) Tween 20) containing 5% skimmilk for 1 h at room temperature. Membranes were then incubated with theprimary antibody diluted in 5% skim milk in TBS-T as follows: anti-5-HT2A receptor(1:250 for 3 h at room temperature), anti-MEK1/2 (1:1000 overnight at 4 �C), anti-p-MEK1/2 (1:2000 for 2.5 h at room temperature), anti-ERK1/2 (1:1000 overnight at4 �C), anti-p-ERK1/2 (1:1000 overnight at 4 �C), anti-JAK2 (1:1000 for 3 h at roomtemperature), anti-STAT3 (1:200 for 2 h at room temperature), anti-p-STAT3(1:1000 overnight at 4 �C), or anti-GAPDH (1:20 000 for 30 min at room tempera-ture). Membranes were washed three times with TBS containing 0.1% Tween 20,then incubated with HRP-conjugated anti-rabbit IgG (1:5000) or anti-mouse IgG(1:5000) as secondary antibodies for 1 h at room temperature and then washed3 times with TBS-T 0.05%. Bands were visualized with Immobilon Western Chemi-luminescent HRP Substrate and their intensities were quantified by densitometricanalysis using FluorChem HD2 software version 6.0 (Alpha Innotech, San Leandro,CA). In certain instances, the PVDF membranes were stripped with Re-Blot plus Mildsolution at room temperature for 15 min, washed twice for 5 min with TBS-T, andquickly rinsed 5 times in water before reprobing with another antibody.
2.8. Statistical analysis
For the cell viability assays, datawere presented asmeans� SEM of at least threeseparate experiments with 6 replicates. For cell cycle experiments, data were
T. Oufkir et al. / Placenta 31 (2010) 439e447440

Author's personal copy
presented as means� SEM of at least three separate experiments. For both cellviability and cell cycle analysis, statistically significant differences were determinedby Student's t test using Prism version 5.0 (GraphPad, San Diego, CA). For densito-metric analysis, data were presented as means� SD and statistical significance wasdetermined by one-way ANOVA followed by NewmaneKeuls post-hoc test. Aprobability value of P< 0.05 was considered statistically significant.
3. Results
3.1. Activation of the 5-HT2A receptor promotes viability of BeWoand JEG-3 cells
Cell viability in BeWo and JEG-3 was increased by DOI ina concentration-dependent manner after 48 h of treatment (Fig. 1Aand B). A maximal increase of 25� 3% (P< 0.001) with 20 mM of
DOI in BeWo and of 11�4% (P< 0.05) with 25 mM in JEG-3 cells wasobserved, compared to untreated cells under low (0.5% FBS) serumconditions. Higher concentrations of DOI decreased viability ofBeWo and JEG-3 cells, probably due to receptors down-regulation.Under conditions with 10% FBS, DOI did not significantly stimulatecell viability (data not shown). Therefore, we used the conditions of20 mM of DOI for BeWo, 25 mM of DOI for JEG-3 as well as 0.5% lowserum to characterize the signalling pathways activated by theplacental 5-HT2A receptor.
Pre-treatment with ketanserin (0.1 mM), a selective 5-HT2Areceptor antagonist, completely blocked the effect of DOI on cellviability in both trophoblast-like cell lines (Fig. 1C and D). Increasedcell proliferation by serotonin, which we described previously [7],was also abolished by 10 mM ketanserin in both cell lines (Fig. 1E
100
110
120
130
MT
T a
ctiv
ity
(%
o
f c
on
tro
l)
DOI (µM)
0
***
******
*
*****
**
100
110
120
130
5 10 15 20 25 40 5 10 15 20 25 40
MT
T a
ctiv
ity
(%
o
f c
on
tro
l)
DOI (µM)
0
** ***
***
A B
100
110
120
130
MT
T a
ctiv
ity
(%
o
f c
on
tro
l)
DOI DOI
+
Ketanserin
Ketanserin
****
C
0
100
110
120
130
MT
T a
ctiv
ity
(%
o
f c
on
tro
l)
DOI DOI
+
Ketanserin
Ketanserin
***
D
0
100
110
120
130
MT
T activity (%
o
f co
ntro
l)
Serotonin Serotonin
+
Ketanserin
Ketanserin
100
110
120
130
MT
T activity (%
o
f co
ntro
l)
Serotonin Serotonin
+
Ketanserin
Ketanserin
E F
***
***
0 0
Fig. 1. Effect of serotonin and 5-HT2A agonist and antagonist on the proliferation of BeWo and JEG-3 cells. (A) BeWo and (B) JEG-3 cells were treated with increasing concentrationsof DOI (selective 5-HT2A agonist) and cell proliferation was determined by MTT assay as described in Materials and Methods. Choriocarcinoma cells were treated with 0.1 mMketanserin (selective 5-HT2A antagonist) before addition of (C) 20 mM (BeWo) and (D) 25 mM (JEG-3) DOI or with 10 mM ketanserin before addition of (E) 20 mM (BeWo) and (F)40 mM (JEG-3) serotonin. The experiment was repeated three times with at least six replicates per experiment in presence of 0.5% (DOI; AeD) or 10% FBS (serotonin; E, F). Data areexpressed as mean� SEM. Statistically significant differences (*P< 0.05; **P< 0.01; ***P< 0.001: ****P< 0.0001) were determined by Student's t test.
T. Oufkir et al. / Placenta 31 (2010) 439e447 441

Author's personal copy
and F). Ketanserin alone had no effect on viability of either celllines. Cell size and conformation did not change during DOI, sero-tonin or ketanserin treatment.
3.2. Activation of the 5-HT2A receptor affects the cell cycle of BeWoand JEG-3 cells
To characterize the effect of 5-HT2A receptor activation on cellcycle distribution of BeWo and JEG-3 cells, changes in cell DNAcontent were measured by flow cytometry. Forty-eight hourstreatment of BeWo with 20 mM DOI induces a substantial increasein number of cells in G2/M phase (21.19%� 1.79 P< 0.05 vs.15.96%� 2.56; P< 0.01), while JEG-3 treated with 25 mM DOI wereat a higher proliferative state as indicated by a greater number ofcells in S phase (31.82%� 1.09 vs. 25.28%� 2.71; P< 0.01) (Fig. 2).Both cell lines treated for 24 h present a similar rate of DNA content(data not shown). The analysis of cell cycle showed neitherapoptotic nor necrotic cells in the absence (control) and in thepresence of DOI. In addition, the flow cytometry analyses showedthat the viability of BeWo and JEG-3 cells was not affected by DOItreatment as indicated by the absence of propidium iodide FL2-A'sfluorescence in the sub-G1 phase of cell cycle (Fig. 2).
3.3. Activation of the 5-HT2A receptor by DOI induces MEK1/2 andERK1/2 phosphorylation in BeWo and JEG-3 cells
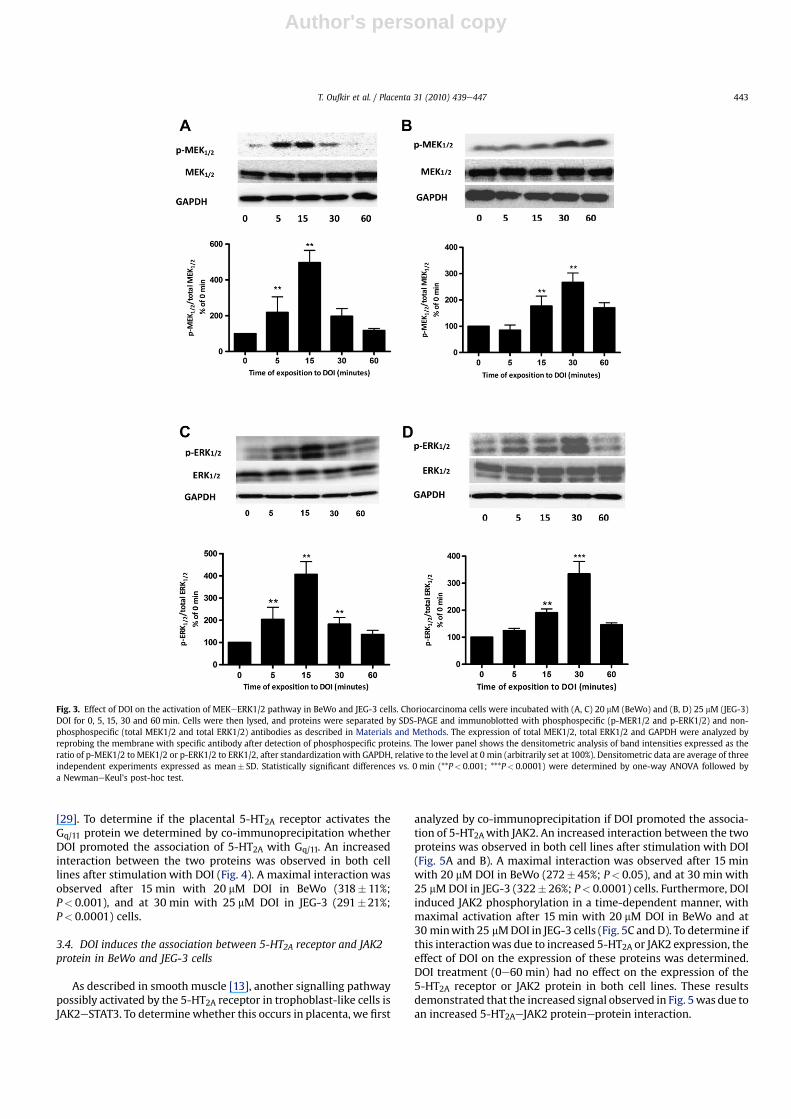
BeWo and JEG-3 cells treated with DOI significantly increasedthe phosphorylation of MEK1/2 in a time-dependent manner(Fig. 3A and B). Maximal phosphorylationwas reached after 15 minwith 20 mM DOI for BeWo (450� 60% of 0 min; P< 0.05), and at30 min with 25 mM DOI for JEG-3 (270� 35% of 0 min; P< 0.001).DOI had no effect on the expression of the non-phosphorylatedform of MEK1/2.
We further determined the effect of DOI on the phosphorylationof ERK1/2 (p42/44-MAPK), the target of p-MEK1/2. As expected,DOI increased the phosphorylation of ERK1/2 in both cell lines ina time-dependent manner. Similar to what we observed forp-MEK1/2, the activation of ERK1/2 reached its maximum after15 min of incubation with 20 mM DOI (410� 47%; P< 0.001) and at30 min incubation with 25 mM DOI (363� 33%; P< 0.0001) inBeWo and JEG-3 cells, respectively (Fig. 3C and D). DOI had no effecton expression of the non-phosphorylated form of ERK1/2.
Involvement of protein kinase C (PKC) in ERK1/2 activation hasbeen described by others [27,28]. Classically, the 5-HT2A receptoractivates the PLC-b/PKC signalling pathway through a Gq/11 protein
Fig. 2. Effect of DOI on cell cycle distribution of BeWo and JEG-3 cells. Choriocarcinoma cell lines were incubated with (A) 20 mM (BeWo) and (B) 25 mM (JEG-3) DOI for 48 h. Cellswere stained for DNA content with propidium iodide. The relative percentage (%) of the cells in each cycle's phase is presented in (C). Each value represents the mean� SEM fromthree independent experiments with different cell passages. Statistically significant differences for each cell cycle phase from control (*P< 0.05, **P< 0.01) were determined byStudent's t test. Y-axis: events or number of cells and X-axis: FL-2A, fluorescence intensity of propidium iodide.
T. Oufkir et al. / Placenta 31 (2010) 439e447442
Author's personal copy
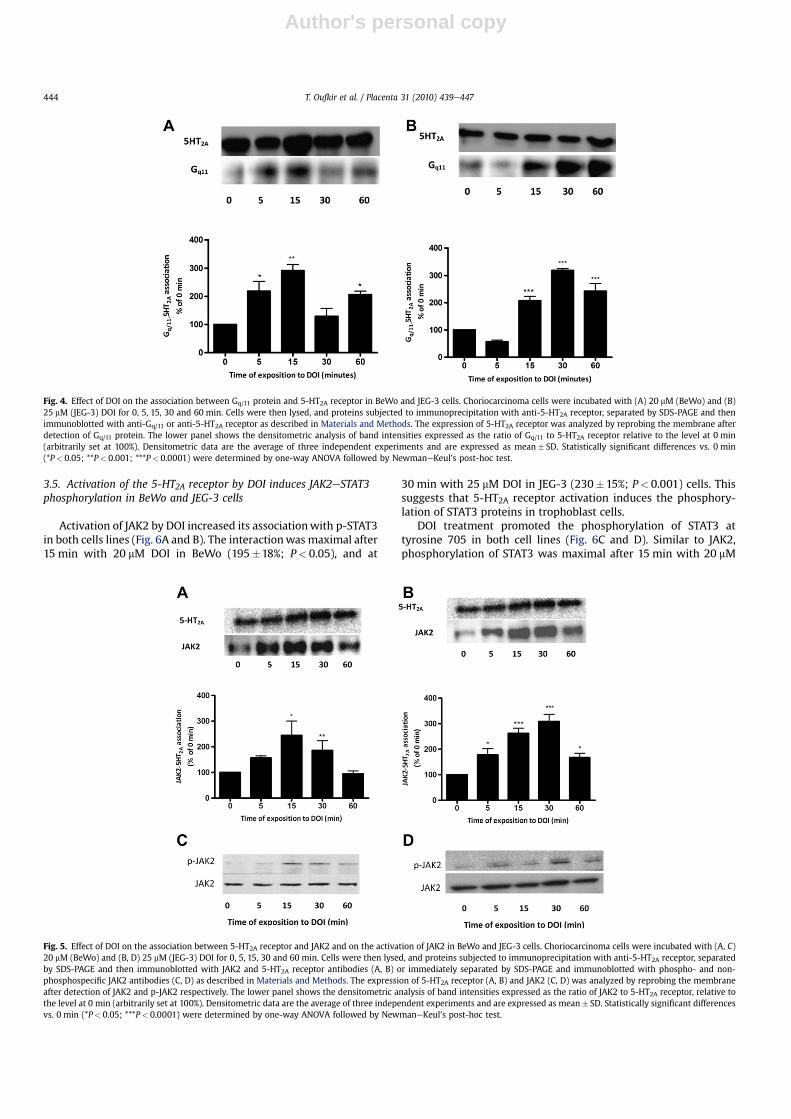
[29]. To determine if the placental 5-HT2A receptor activates theGq/11 protein we determined by co-immunoprecipitation whetherDOI promoted the association of 5-HT2A with Gq/11. An increasedinteraction between the two proteins was observed in both celllines after stimulation with DOI (Fig. 4). A maximal interaction wasobserved after 15 min with 20 mM DOI in BeWo (318� 11%;P< 0.001), and at 30 min with 25 mM DOI in JEG-3 (291�21%;P< 0.0001) cells.
3.4. DOI induces the association between 5-HT2A receptor and JAK2protein in BeWo and JEG-3 cells
As described in smooth muscle [13], another signalling pathwaypossibly activated by the 5-HT2A receptor in trophoblast-like cells isJAK2eSTAT3. To determine whether this occurs in placenta, we first
analyzed by co-immunoprecipitation if DOI promoted the associa-tion of 5-HT2Awith JAK2. An increased interaction between the twoproteins was observed in both cell lines after stimulation with DOI(Fig. 5A and B). A maximal interaction was observed after 15 minwith 20 mM DOI in BeWo (272� 45%; P< 0.05), and at 30 min with25 mMDOI in JEG-3 (322� 26%; P< 0.0001) cells. Furthermore, DOIinduced JAK2 phosphorylation in a time-dependent manner, withmaximal activation after 15 min with 20 mM DOI in BeWo and at30 minwith 25 mMDOI in JEG-3 cells (Fig. 5C and D). To determine ifthis interactionwas due to increased 5-HT2A or JAK2 expression, theeffect of DOI on the expression of these proteins was determined.DOI treatment (0e60 min) had no effect on the expression of the5-HT2A receptor or JAK2 protein in both cell lines. These resultsdemonstrated that the increased signal observed in Fig. 5was due toan increased 5-HT2AeJAK2 proteineprotein interaction.
Fig. 3. Effect of DOI on the activation of MEKeERK1/2 pathway in BeWo and JEG-3 cells. Choriocarcinoma cells were incubated with (A, C) 20 mM (BeWo) and (B, D) 25 mM (JEG-3)DOI for 0, 5, 15, 30 and 60 min. Cells were then lysed, and proteins were separated by SDS-PAGE and immunoblotted with phosphospecific (p-MER1/2 and p-ERK1/2) and non-phosphospecific (total MEK1/2 and total ERK1/2) antibodies as described in Materials and Methods. The expression of total MEK1/2, total ERK1/2 and GAPDH were analyzed byreprobing the membrane with specific antibody after detection of phosphospecific proteins. The lower panel shows the densitometric analysis of band intensities expressed as theratio of p-MEK1/2 to MEK1/2 or p-ERK1/2 to ERK1/2, after standardization with GAPDH, relative to the level at 0 min (arbitrarily set at 100%). Densitometric data are average of threeindependent experiments expressed as mean� SD. Statistically significant differences vs. 0 min (**P< 0.001; ***P< 0.0001) were determined by one-way ANOVA followed bya NewmaneKeul's post-hoc test.
T. Oufkir et al. / Placenta 31 (2010) 439e447 443
Author's personal copy
3.5. Activation of the 5-HT2A receptor by DOI induces JAK2eSTAT3phosphorylation in BeWo and JEG-3 cells
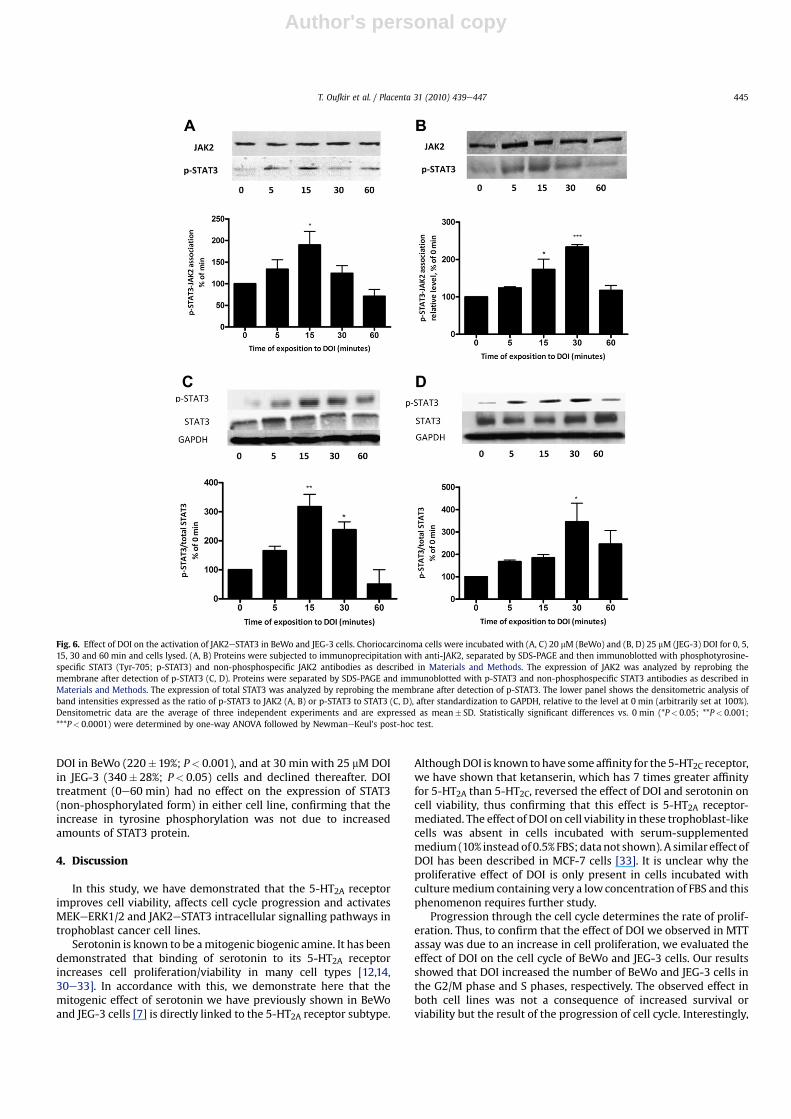
Activation of JAK2 by DOI increased its associationwith p-STAT3in both cells lines (Fig. 6A and B). The interactionwas maximal after15 min with 20 mM DOI in BeWo (195�18%; P< 0.05), and at
30 min with 25 mM DOI in JEG-3 (230�15%; P< 0.001) cells. Thissuggests that 5-HT2A receptor activation induces the phosphory-lation of STAT3 proteins in trophoblast cells.
DOI treatment promoted the phosphorylation of STAT3 attyrosine 705 in both cell lines (Fig. 6C and D). Similar to JAK2,phosphorylation of STAT3 was maximal after 15 min with 20 mM
Fig. 4. Effect of DOI on the association between Gq/11 protein and 5-HT2A receptor in BeWo and JEG-3 cells. Choriocarcinoma cells were incubated with (A) 20 mM (BeWo) and (B)25 mM (JEG-3) DOI for 0, 5, 15, 30 and 60 min. Cells were then lysed, and proteins subjected to immunoprecipitation with anti-5-HT2A receptor, separated by SDS-PAGE and thenimmunoblotted with anti-Gq/11 or anti-5-HT2A receptor as described in Materials and Methods. The expression of 5-HT2A receptor was analyzed by reprobing the membrane afterdetection of Gq/11 protein. The lower panel shows the densitometric analysis of band intensities expressed as the ratio of Gq/11 to 5-HT2A receptor relative to the level at 0 min(arbitrarily set at 100%). Densitometric data are the average of three independent experiments and are expressed as mean� SD. Statistically significant differences vs. 0 min(*P< 0.05; **P< 0.001; ***P< 0.0001) were determined by one-way ANOVA followed by NewmaneKeul's post-hoc test.
Fig. 5. Effect of DOI on the association between 5-HT2A receptor and JAK2 and on the activation of JAK2 in BeWo and JEG-3 cells. Choriocarcinoma cells were incubated with (A, C)20 mM (BeWo) and (B, D) 25 mM (JEG-3) DOI for 0, 5, 15, 30 and 60 min. Cells were then lysed, and proteins subjected to immunoprecipitation with anti-5-HT2A receptor, separatedby SDS-PAGE and then immunoblotted with JAK2 and 5-HT2A receptor antibodies (A, B) or immediately separated by SDS-PAGE and immunoblotted with phospho- and non-phosphospecific JAK2 antibodies (C, D) as described in Materials and Methods. The expression of 5-HT2A receptor (A, B) and JAK2 (C, D) was analyzed by reprobing the membraneafter detection of JAK2 and p-JAK2 respectively. The lower panel shows the densitometric analysis of band intensities expressed as the ratio of JAK2 to 5-HT2A receptor, relative tothe level at 0 min (arbitrarily set at 100%). Densitometric data are the average of three independent experiments and are expressed as mean� SD. Statistically significant differencesvs. 0 min (*P< 0.05; ***P< 0.0001) were determined by one-way ANOVA followed by NewmaneKeul's post-hoc test.
T. Oufkir et al. / Placenta 31 (2010) 439e447444
Author's personal copy
DOI in BeWo (220�19%; P< 0.001), and at 30 min with 25 mM DOIin JEG-3 (340� 28%; P< 0.05) cells and declined thereafter. DOItreatment (0e60 min) had no effect on the expression of STAT3(non-phosphorylated form) in either cell line, confirming that theincrease in tyrosine phosphorylation was not due to increasedamounts of STAT3 protein.
4. Discussion
In this study, we have demonstrated that the 5-HT2A receptorimproves cell viability, affects cell cycle progression and activatesMEKeERK1/2 and JAK2eSTAT3 intracellular signalling pathways introphoblast cancer cell lines.
Serotonin is known to be amitogenic biogenic amine. It has beendemonstrated that binding of serotonin to its 5-HT2A receptorincreases cell proliferation/viability in many cell types [12,14,30e33]. In accordance with this, we demonstrate here that themitogenic effect of serotonin we have previously shown in BeWoand JEG-3 cells [7] is directly linked to the 5-HT2A receptor subtype.
AlthoughDOI is known tohave someaffinity for the 5-HT2C receptor,we have shown that ketanserin, which has 7 times greater affinityfor 5-HT2A than 5-HT2C, reversed the effect of DOI and serotonin oncell viability, thus confirming that this effect is 5-HT2A receptor-mediated. The effect of DOI on cell viability in these trophoblast-likecells was absent in cells incubated with serum-supplementedmedium(10% instead of 0.5% FBS; datanot shown). A similar effect ofDOI has been described in MCF-7 cells [33]. It is unclear why theproliferative effect of DOI is only present in cells incubated withculturemedium containing very a lowconcentration of FBS and thisphenomenon requires further study.
Progression through the cell cycle determines the rate of prolif-eration. Thus, to confirm that the effect of DOI we observed in MTTassay was due to an increase in cell proliferation, we evaluated theeffect of DOI on the cell cycle of BeWo and JEG-3 cells. Our resultsshowed that DOI increased the number of BeWo and JEG-3 cells inthe G2/M phase and S phases, respectively. The observed effect inboth cell lines was not a consequence of increased survival orviability but the result of the progression of cell cycle. Interestingly,
Fig. 6. Effect of DOI on the activation of JAK2eSTAT3 in BeWo and JEG-3 cells. Choriocarcinoma cells were incubated with (A, C) 20 mM (BeWo) and (B, D) 25 mM (JEG-3) DOI for 0, 5,15, 30 and 60 min and cells lysed. (A, B) Proteins were subjected to immunoprecipitation with anti-JAK2, separated by SDS-PAGE and then immunoblotted with phosphotyrosine-specific STAT3 (Tyr-705; p-STAT3) and non-phosphospecific JAK2 antibodies as described in Materials and Methods. The expression of JAK2 was analyzed by reprobing themembrane after detection of p-STAT3 (C, D). Proteins were separated by SDS-PAGE and immunoblotted with p-STAT3 and non-phosphospecific STAT3 antibodies as described inMaterials and Methods. The expression of total STAT3 was analyzed by reprobing the membrane after detection of p-STAT3. The lower panel shows the densitometric analysis ofband intensities expressed as the ratio of p-STAT3 to JAK2 (A, B) or p-STAT3 to STAT3 (C, D), after standardization to GAPDH, relative to the level at 0 min (arbitrarily set at 100%).Densitometric data are the average of three independent experiments and are expressed as mean� SD. Statistically significant differences vs. 0 min (*P< 0.05; **P< 0.001;***P< 0.0001) were determined by one-way ANOVA followed by NewmaneKeul's post-hoc test.
T. Oufkir et al. / Placenta 31 (2010) 439e447 445

Author's personal copy
cell cycle analysis combined with MTT viability assay showed thatDOI induced BeWo cell viability and at the same time decreased thenumber of cell that underwent DNA synthesis (S phase). Thedifferential effects of DOI on BeWo and JEG-3 cells may reflectcertain genetic differences between the cell lines, even though theyare both derived from choriocarcinomas. For example, BeWo areable to fuse spontaneously [34] causing a blockade of cell cycleprogression, thus counteracting the proliferative effect of DOI. JEG-3cells, on the other hand, are highly proliferative and invasive andfuse poorly. Cell proliferation is internally regulated by a balance ofcell cycle activator and inhibitor proteins.We are currently pursuingdetailed studies of the cell cycle to determine which signallingpathways (e.g. cyclins, cyclin kinases.) are involved in the action ofDOI on cell cycle progression of BeWo and JEG-3 cells. Cyclins aretargeted following ERK1/2 nuclear translocation and appear to bedifferentially regulated between BeWo and JEG-3.
BeWo and JEG-3 cells possess different characteristics and donot always respond similarly to various physiological or pharma-cological stimuli. One study showed that insulin treatmentincreased the expression of endothelin-B receptor in BeWo cells,but decreased it in JEG-3 [35]. Another study showed that in BeWocells, increased invasion was associated with a decrease in celldifferentiation, whereas in JEG-3 cells the opposite occurred [36].Those findings are in agreement with our results showing differ-ential concentration- and time-courses for DOI-induced cellproliferation and activation of signalling cascades in the two celllines. The greater mitogenic effect of DOI in BeWo than in JEG-3cells may be explained by a higher density of 5-HT2A receptors inBeWo cells. Binding studies could verify this hypothesis.
Our study has demonstrated that 5-HT2A receptor activation byDOI stimulates MEK1/2 and ERK1/2 phosphorylation in bothtrophoblast-like cell lines. Previous studies have demonstrated theinvolvement of PKC in 5-HT2A receptor activation of ERK1/2 [30,31].As with the activation of MEK1/2 and ERK1/2, DOI promotes theinteraction between 5-HT2A and Gq/11 in both cell lines, suggestingthat the placental 5-HT2A receptor activates MEK1/2eERK1/2through the Gq/11-PLC-b-PKC-Ras signalling pathway. The activa-tion of the MEK1/2eERK1/2 pathway by the 5-HT2A receptor couldalso be the result of a PLC-PKC-independent mechanism, such asthe activation of calmodulin, Src, phosphoinositide-3 kinase (PI3K),or the epidermal growth factor (EGF) receptor tyrosine kinase[32,37,38]. This remains open to further investigation.
The ERK1/2 signalling pathway has been shown to inducevillous trophoblast cell differentiation [21,22], and extra-villoustrophoblast migration [25,26]. Our present study suggests that the5-HT2A receptor is involved in trophoblast development (invasion/migration, proliferation, and differentiation) through activation ofthe ERK1/2 signalling pathway.
The JAK2eSTAT3 signalling cascade has been reported to becoupled to the 5-HT2A receptor in skeletal muscles [13]. We havepresently demonstrated in BeWo and JEG-3 cells that 5-HT2Areceptor activation increases the association between JAK2 andSTAT3 leading to the phosphorylation of STAT3 on tyrosine 705.5-HT2A receptor activation by DOI also increased the phosphoryla-tion of JAK2. For the immunoprecipitation with JAK2, we used anantibody that recognized total JAK2 (non- andphosphorylated) [39].Thus, these results reflect the association of p-JAK2 with STAT3,since only p-JAK2would be able to interact with and activate STAT3.The phosphorylation of this conserved C-terminal tyrosine isrequired for STAT3 dimerization and translocation to the nucleuswhere it binds to target DNA sequences. Genes regulated by STAT3include anti-apoptotic and proliferation-associated genes such asBcl-xL, Bcl-2, Fas, cyclin D1, survivin, and c-Myc [40]. In addition,STAT3 has been found to induce angiogenesis, and increase cellmigration and invasion [41]. Our findings suggest that serotonin
may regulate trophoblast development (invasion/migration, prolif-eration, and apoptosis) through activation of STAT3. In support ofthis, it has been shown that STAT3 activation plays a crucial role inthe invasive phenotype of trophoblast cells [23] and correlateswith choriocarcinoma cell invasiveness [24]. The activation ofJAK2eSTAT3 by the 5-HT2A receptor also appears to be involved inmyogenic differentiation in rat fetal myoblast cells [13].
The MAPK signalling pathway has also been shown to interactwith STAT [42]. JAK2 phosphorylates STAT3 on tyrosine leading todimerization and nuclear localization, whereas MAPK phosphory-lates STAT3 on serine 727, leading to maximal transcriptionalactivation of STAT3 [42]. Given that serotonin through its 5-HT2Areceptor activates the MAPK pathway, STATs and MAPK may coop-erate inmediating theproliferative responseofBeWoand JEG-3cellsto 5-HT2A receptor agonists, although this remains to be studied.
The results of this study are important for a better under-standing of placental health and pregnancy well-being. A numberof studies suggest that serotonin might be implicated in placentaldevelopment and function. We recently showed a decrease in both5-HT2A receptor and serotonin transporter expression in humanplacental tissues from gestational diabetes vs. normal pregnancy[8]. Moreover, it has been suggest that serotonin may play a crucialrole in preeclampsia and ketanserin has been described as a drugthat appears to be tailored for treating pregnancy-induced hyper-tensive disorders [43,44], suggesting a role for serotonin and its5-HT2A receptor in the pathophysiology of these diseases. Forexample, several studies have emphasized the importance oftrophoblast homeostasis in pregnancy health. Specifically, alter-ations in trophoblast development (differentiation, invasion,proliferation, apoptosis) are associated with pregnancy disorders,such as preeclampsia, gestational diabetes, intrauterine growthrestriction and preterm birth [45]. It is important to better under-stand the mechanisms of 5-HT2A receptor function in the tropho-blast cell processes, in order to identify its role in placental function,fetal development and pregnancy well-being.
In summary, this study demonstrated that 5-HT2A receptorimproves cell viability and affect cell cycle progression in humanchoriocarcinoma cell lines and represents the first report that theactivation of the 5-HT2A receptor in trophoblast cells activatesintracellular signal, MEKeERK1/2 and JAK2eSTAT3 pathways, thatcan be related to survival, differentiation, migration and invasion.These findings suggest that serotonin through the activation of the5-HT2A receptor may be an important regulator of placentation andplay a role in the pathophysiology of pregnancy disorders associ-ated with alterations in placental development, such aspreeclampsia, gestational diabetes and preterm birth. Furtherstudies are necessary to better understand and characterize thesignal transduction pathways initiated by serotonin via its placental5-HT2A receptor and their role in regulating human placentation.
Acknowledgements
This study was financed by a Natural Sciences and EngineeringResearch Council of Canada (NSERC) grant to C Vaillancourt(262011) and a Canadian Institutes of Health Research (CIHR) grantto JT Sanderson and C. Vaillancourt (IGO 93972) as well as an NSERCstudentship to M. Arseneault.
References
[1] Jonnakuty C, Gragnoli C. What do we know about serotonin? J Cell Physiol2008;217:301e6.
[2] Azmitia EC. Serotonin neurons, neuroplasticity, and homeostasis of neuraltissue. Neuropsychopharmacology 1999;21:33Se45S.
[3] Buznikov GA, Lambert HW, Lauder JM. Serotonin and serotonin-likesubstances as regulators of early embryogenesis and morphogenesis. CellTissue Res 2001;305:177e86.
T. Oufkir et al. / Placenta 31 (2010) 439e447446

Author's personal copy
[4] Cote F, Fligny C, Bayard E, Launay JM, Gershon MD, Mallet J, et al. Maternalserotonin is crucial for murine embryonic development. Proc Natl Acad SciUSA 2007;104:329e34.
[5] Bolte AC, van Geijn HP, Dekker GA. Pathophysiology of preeclampsia and therole of serotonin. Eur J Obstet Gynecol Reprod Biol 2001;95:12e21.
[6] Radenkovic M, Radunovic N, Momcilov P, Grbovic L. Altered response ofhuman umbilical artery to 5-HT in gestational diabetic pregnancy. PharmacolRep 2009;61:520e8.
[7] Sonier B, Lavigne C, Arseneault M, Ouellette R, Vaillancourt C. Expression ofthe 5-HT2A serotoninergic receptor in human placenta and choriocarcinomacells: mitogenic implications of serotonin. Placenta 2005;26:484e90.
[8] Viau M, Lafond J, Vaillancourt C. Expression of placental serotonin transporterand 5-HT2A receptor in normal and gestational diabetes mellitus pregnancies.Reprod Biomed Online 2009;19:207e15.
[9] Huang WQ, Zhang CL, Di XY, Zhang RQ. Studies on the localization of 5-hydroxytryptamine and its receptors in human placenta. Placenta 1998;19:655e61.
[10] Bottalico B, Larsson I, Brodszki J, Hernandez-Andrade E, Casslen B, Marsal K,et al. Norepinephrine Transporter (NET), Serotonin Transporter (SERT),Vesicular Monoamine Transporter (VMAT2) and Organic Cation Transporters(OCT1, 2 and EMT) in human placenta from pre-eclamptic and normotensivepregnancies. Placenta 2004;25:518e29.
[11] Hoyer D, Hannon JP, Martin GR. Molecular, pharmacological and functionaldiversity of 5-HT receptors. Pharmacol Biochem Behav 2002;71:533e54.
[12] Nebigil CG, Garnovskaya MN, Spurney RF, Raymond JR. Identification of a ratglomerular mesangial cell mitogenic 5-HT2A receptor. Am J Physiol 1995;268(1 Pt 2):F122e7.
[13] Guillet-Deniau I, Burnol AF, Girard J. Identification and localization of a skel-etal muscle secrotonin 5-HT2A receptor coupled to the Jak/STAT pathway.J Biol Chem 1997;272:14825e9.
[14] Banes AKL, Shaw SM, Tawfik A, Patel BP, Ogbi S, Fulton D, et al. Activation ofthe JAK/STAT pathway in vascular smooth muscle by serotonin. Am J Physiol2005;288:C805e12.
[15] Azmitia EC. Modern views on an ancient chemical: serotonin effects on cellproliferation, maturation, and apoptosis. Brain Res Bull 2001;56:413e24.
[16] Watts SW, Yang P, Banes AK, Baez M. Activation of Erk mitogen-activatedprotein kinase proteins by vascular serotonin receptors. J Cardiovasc Phar-macol 2001;38:539e51.
[17] Liebmann C. Regulation of MAP kinase activity by peptide receptor signallingpathway: paradigms of multiplicity. Cell Signal 2001;13:777e85.
[18] Imada K, Leonard WJ. The JakeSTAT pathway. Mol Immunol 2000;37:1e11.[19] Forbes K, Westwood M, Baker PN, Aplin JD. Insulin-like growth factor I and II
regulate the life cycle of trophoblast in the developing human placenta. Am JPhysiol 2008;294:C1313e22.
[20] Marino VJ, Roguin LP. The granulocyte colony stimulating factor (G-CSF)activates Jak/STAT and MAPK pathways in a trophoblastic cell line. J CellBiochem 2008;103:1512e23.
[21] Daoud G, Amyot M, Rassart E, Masse A, Simoneau L, Lafond J. ERK1/2 and p38regulate trophoblasts differentiation in human term placenta. J Physiol 2005;566:409e23.
[22] Vaillancourt C, Lanoix D, Le Bellego F, Daoud G, Lafond J. Involvement of MAPKsignalling in human villous trophoblast differentiation. Mini Rev Med Chem2009;9:962e73.
[23] Poehlmann TG, Fitzgerald JS, Meissner A, Wengenmayer T, Schleussner E,Friedrich K, et al. Trophoblast invasion: tuning through LIF, signalling viaStat3. Placenta 2005;26(Suppl. A):S37e41.
[24] Corvinus FM, Fitzgerald JS, Friedrich K, Markert UR. Evidence for a correlationbetween trophoblast invasiveness and STAT3 activity. Am J Reprod Immunol2003;50:316e21.
[25] Chakraborty C, Gleeson LM, McKinnon T, Lala PK. Regulation of humantrophoblast migration and invasiveness. Can J Physiol Pharmacol 2002;80:116e24.
[26] Prast J, Saleh L, Husslein H, Sonderegger S, Helmer H, Knofler M. Humanchorionic gonadotropin stimulates trophoblast invasion through extracellu-larly regulated kinase and AKT signaling. Endocrinology 2008;149:979e87.
[27] Rozengurt E. Mitogenic signaling pathways induced by G protein-coupledreceptors. J Cell Physiol 2007;213:589e602.
[28] Naor Z, Benard O, Seger R. Activation of MAPK cascades by G-protein-coupledreceptors: the case of gonadotropin-releasing hormone receptor. TrendsEndocrinol Metab 2000;11:91e9.
[29] Raymond JR, Mukhin YV, Gelasco A, Turner J, Collinsworth G, Gettys TW, et al.Multiplicity of mechanisms of serotonin receptor signal transduction. Phar-macol Ther 2001;92:179e212.
[30] Greene EL, Houghton O, Collinsworth G, Garnovskaya MN, Nagai T, Sajjad T,et al. 5-HT(2A) receptors stimulate mitogen-activated protein kinase via H(2)O(2) generation in rat renal mesangial cells. Am J Physiol Renal Physiol2000;278:F650e8.
[31] Grewal JS, Mukhin YV, Garnovskaya MN, Raymond JR, Greene EL. Serotonin 5-HT2A receptor induces TGF-beta1 expression in mesangial cells via ERK:proliferative and fibrotic signals. Am J Physiol 1999;276:F922e30.
[32] Watts SW. Activation of the mitogen-activated protein kinase pathway via the5-HT2A receptor. Ann N Y Acad Sci 1998;861:162e8.
[33] Sonier B, Arseneault M, Lavigne C, Ouellette RJ, Vaillancourt C. The 5-HT2Aserotoninergic receptor is expressed in the MCF-7 human breast cancer cellline and reveals a mitogenic effect of serotonin. Biochem Biophys Res Com-mun 2006;343:1053e9.
[34] Mounier C, Barbeau B, Vaillancourt C, Lafond J. Endocrinology and cellsignaling in human villous trophoblast. Methods Mol Biol 2009;550:89e102.
[35] BilbanM, Barth S, CervarM,Mauschitz R, Schaur RJ, Zivkovic F, et al. Differentialregulation of endothelin secretion and endothelin receptormRNA levels in JAR,JEG-3, and BeWo choriocarcinoma cell lines and in human trophoblasts, theirnonmalignant counterpart. Arch Biochem Biophys 2000;382:245e52.
[36] Hohn HP, Linke M, Ugele B, Denker HW. Differentiation markers and inva-siveness: discordant regulation in normal trophoblast and choriocarcinomacells. Exp Cell Res 1998;244:249e58.
[37] Quinn JC, Johnson-Farley NN, Yoon J, Cowen DS. Activation of extracellular-regulated kinase by 5-hydroxytryptamine(2A) receptors in PC12 cells isprotein kinase C-independent and requires calmodulin and tyrosine kinases.J Pharmacol Exp Ther 2002;303:746e52.
[38] Banes A, Florian JA, Watts SW. Mechanisms of 5-hydroxytryptamine(2A)receptor activation of the mitogen-activated protein kinase pathway invascular smooth muscle. J Pharmacol Exp Ther 1999;291:1179e87.
[39] Thompson BR, Mazurkiewicz-Munoz AM, Suttles J, Carter-Su C, Bernlohr DA.Interaction of adipocyte fatty acid-binding protein (AFABP) and JAK2: AFABP/aP2 as a regulator of JAK2 signaling. J Biol Chem 2009;284:13473e80.
[40] Bromberg J. Stat proteins and oncogenesis. J Clin Invest 2002;109:1139e42.[41] Yu H, Kortylewski M, Pardoll D. Crosstalk between cancer and immune cells:
role of STAT3 in the tumour microenvironment. Nat Rev Immunol 2007;7:41e51.
[42] Bowman T, Garcia R, Turkson J, Jove R. STATs in oncogenesis. Oncogene2000;19:2474e88.
[43] Bolte AC, van Eyck J, Gaffar SF, van Geijn HP, Dekker GA. Ketanserin for thetreatment of preeclampsia. J Perinat Med 2001;29:14e22.
[44] Weiner CP, Socol ML, Vaisrub N. Control of preeclamptic hypertension byketanserin, a new serotonin receptor antagonist. Am J Obstet Gynecol 1984;149:496e500.
[45] Regnault TRH, Galan HL, Parker TA, Anthony RV. Placental development innormal and compromised pregnancies e a review. Placenta 2002;23:S119e29.
T. Oufkir et al. / Placenta 31 (2010) 439e447 447




















