
Thalamocortical oscillations in a genetic model of absence seizures
11
Thalamocortical oscillations in a genetic model of absence seizures Giovanna D’Arcangelo, 1,2 Margherita D’Antuono, 3 Giuseppe Biagini, 4 Richard Warren, 5 Virginia Tancredi 2 and Massimo Avoli 1,3 1 Montreal Neurological Institute and Department of Neurology & Neurosurgery, McGill University, 3801, University St., Montre ´al, QC, H3A 2B4, Canada 2 Dipartimento di Neuroscienze, Universita ` degli Studi di Roma ‘Tor Vergata’, 00173 Roma, Italy 3 IRCCS Neuromed, 86077, Pozzilli (Isernia), Italy 4 Dipartimento di Scienze Biomediche, Universita ` degli Studi di Modena e Reggio Emilia, 41100 Modena, Italy 5 De ´ partement de Psychiatrie, Universite ´ de Montre ´ al and Centre de Recherche Fernand-Seguin, Montre ´al, QC, H1N 3V2, Canada Keywords: 4-aminopyridine, epileptiform activity, extracellular potassium, GABA Abstract We used field potential recordings in an in vitro thalamocortical slice preparation to compare the rhythmic oscillations generated by reciprocally connected networks of the thalamus and cerebral cortex obtained from epileptic (> 160 days old) WAG/Rij and age-matched, nonepileptic control (NEC) rats. To increase neuronal excitability, and thus to elicit spontaneous field potential activity in vitro, we applied medium containing: (i) zero [Mg 2+ ]; (ii) high [K + ] (8.25 mM); or (iii) low concentrations of the K + channel blocker 4-aminopyridine (4AP, 0.5–1 mM). Of these procedures, only the last was effective in triggering oscillatory activity that depended on the type of tissue. Thus, during 4AP application: (i) sequences of fast (intraburst frequency 9.5–16.1 Hz) and slower (5–8.9 Hz) field potential oscillations (FPOs) were recorded in WAG/Rij slices (n = 23), but (ii) only fast FPOs were seen in NEC slices (n = 7). Slower FPOs in WAG/Rij slices reflected a larger degree of thalamocortical synchronization than fast FPOs, and disappeared after surgical separation of cortex and thalamus (n = 5); under these conditions fast FPOs continued to occur in thalamus only. In addition, fast and slower FPOs disappeared in all areas of the WAG/Rij slice during thalamic application of the excitatory amino acid receptor antagonist kynurenic acid (n = 3), while fast FPOs continued to occur in thalamus when kynurenic acid was applied to the cortex (n = 4). Bath application of the N-methyl-D-aspartic acid (NMDA) receptor antagonist 3,3-(2-carboxypiperazine-4-yl)-propyl-1-phosphonate (CPP) abolished slower FPOs in WAG/Rij cortex and thalamus (n = 6) without infuencing fast FPOs recorded in WAG/Rij (n = 6) or NEC slices (n = 4). Moreover, cortical application of CPP (n = 6) abated slower FPOs although they persisted following CPP application to the thalamus (n = 7). Our data demonstrate that highly synchronized, slower FPOs can occur during 4AP application in WAG/Rij but not in NEC slices. This activity, which may represent an in vitro hallmark of thalamocortical epileptogenicity, requires the function of reciprocally connected thalamic and cortical networks and depends on cortical NMDA receptor-mediated mechanisms. Introduction Generalized spike and wave (SW) discharges occurring at 2.5–4 Hz in patients presenting with primary generalized epilepsies are commonly associated with brief periods of impaired consciousness (the absence seizures). Over the last few decades, several investiga- tors have analysed the mechanisms of SW discharges, most often by using models in which normal animals were made ‘epileptic’ by convulsant treatment or by brain electrical stimuli (see for review Avoli et al., 1990, 2001; Snead, 1995). These studies have suggested that SW discharges originate from oscillatory thalamocortical inter- actions similar to those involved in sleep spindles. However, the modalities by which thalamocortical interactions lead to the slow rhythm of SW discharge remain elusive. SW discharges in humans occur spontaneously and have a genetic origin (Lennox & Lennox, 1960; Metrakos & Metrakos, 1961). Hence, knowledge of the pathophysiogenesis of generalized SW activity should gain from naturally occurring genetic animal models of absence seizures. These models, which include the Genetic Absence Epilepsy in Rats from Strasbourg (GAERS) (Vergnes et al., 1990) and the Wistar Albino Glaxo/Rijswijk rat (WAG/Rij) (van Luijtelaar & Coenen, 1986; Peeters et al., 1990), are characterized by generalized SW discharges at 6–10 Hz that respond to antiepileptic drugs in a way similar to what has been reported in patients with primary generalized epilepsy. These genetically determined absence seizures have been analysed with microphysiological (including intracellular) recordings that were obtained in vivo from cerebral cortex and from different thalamic nuclei (Pinault et al., 1998, 2001; Seidenbecher et al., 1998; Charpier et al., 1999; Seidenbecher & Pape, 2001; Staak & Pape, 2001; Meeren et al., 2002; Slaght et al., 2002). These studies, recently reviewed by Crunelli & Leresche 2002), have (i) shown the occurrence of specific patterns of cortical Correspondence: Dr M. Avoli, 1 Montreal Neurological Institute, as above. E-mail: [email protected] Received 7 October 2002, accepted 14 October 2002 doi:10.1046/j.1460-9568.2002.02411.x European Journal of Neuroscience, Vol. 16, pp. 2383–2393, 2002 ª Federation of European Neuroscience Societies
Transcript of Thalamocortical oscillations in a genetic model of absence seizures

Thalamocortical oscillations in a genetic model of absenceseizures
Giovanna D'Arcangelo,1,2 Margherita D'Antuono,3 Giuseppe Biagini,4 Richard Warren,5 Virginia Tancredi2 andMassimo Avoli1,3
1Montreal Neurological Institute and Department of Neurology & Neurosurgery, McGill University, 3801, University St.,
MontreÂal, QC, H3A 2B4, Canada2Dipartimento di Neuroscienze, UniversitaÁ degli Studi di Roma `Tor Vergata', 00173 Roma, Italy3IRCCS Neuromed, 86077, Pozzilli (Isernia), Italy4Dipartimento di Scienze Biomediche, UniversitaÁ degli Studi di Modena e Reggio Emilia, 41100 Modena, Italy5DeÂpartement de Psychiatrie, Universite de MontreÂal and Centre de Recherche Fernand-Seguin, MontreÂal, QC, H1N 3V2,
Canada
Keywords: 4-aminopyridine, epileptiform activity, extracellular potassium, GABA
Abstract
We used ®eld potential recordings in an in vitro thalamocortical slice preparation to compare the rhythmic oscillations generatedby reciprocally connected networks of the thalamus and cerebral cortex obtained from epileptic (> 160 days old) WAG/Rij and
age-matched, nonepileptic control (NEC) rats. To increase neuronal excitability, and thus to elicit spontaneous ®eld potential
activity in vitro, we applied medium containing: (i) zero [Mg2+]; (ii) high [K+] (8.25 mM); or (iii) low concentrations of the K+
channel blocker 4-aminopyridine (4AP, 0.5±1 mM). Of these procedures, only the last was effective in triggering oscillatory activitythat depended on the type of tissue. Thus, during 4AP application: (i) sequences of fast (intraburst frequency 9.5±16.1 Hz) and
slower (5±8.9 Hz) ®eld potential oscillations (FPOs) were recorded in WAG/Rij slices (n = 23), but (ii) only fast FPOs were seen
in NEC slices (n = 7). Slower FPOs in WAG/Rij slices re¯ected a larger degree of thalamocortical synchronization than fastFPOs, and disappeared after surgical separation of cortex and thalamus (n = 5); under these conditions fast FPOs continued to
occur in thalamus only. In addition, fast and slower FPOs disappeared in all areas of the WAG/Rij slice during thalamic
application of the excitatory amino acid receptor antagonist kynurenic acid (n = 3), while fast FPOs continued to occur inthalamus when kynurenic acid was applied to the cortex (n = 4). Bath application of the N-methyl-D-aspartic acid (NMDA)
receptor antagonist 3,3-(2-carboxypiperazine-4-yl)-propyl-1-phosphonate (CPP) abolished slower FPOs in WAG/Rij cortex and
thalamus (n = 6) without infuencing fast FPOs recorded in WAG/Rij (n = 6) or NEC slices (n = 4). Moreover, cortical application
of CPP (n = 6) abated slower FPOs although they persisted following CPP application to the thalamus (n = 7). Our datademonstrate that highly synchronized, slower FPOs can occur during 4AP application in WAG/Rij but not in NEC slices. This
activity, which may represent an in vitro hallmark of thalamocortical epileptogenicity, requires the function of reciprocally
connected thalamic and cortical networks and depends on cortical NMDA receptor-mediated mechanisms.
IntroductionGeneralized spike and wave (SW) discharges occurring at 2.5±4 Hz
in patients presenting with primary generalized epilepsies are
commonly associated with brief periods of impaired consciousness
(the absence seizures). Over the last few decades, several investiga-
tors have analysed the mechanisms of SW discharges, most often by
using models in which normal animals were made `epileptic' by
convulsant treatment or by brain electrical stimuli (see for review
Avoli et al., 1990, 2001; Snead, 1995). These studies have suggested
that SW discharges originate from oscillatory thalamocortical inter-
actions similar to those involved in sleep spindles. However, the
modalities by which thalamocortical interactions lead to the slow
rhythm of SW discharge remain elusive.
SW discharges in humans occur spontaneously and have a genetic
origin (Lennox & Lennox, 1960; Metrakos & Metrakos, 1961).
Hence, knowledge of the pathophysiogenesis of generalized SW
activity should gain from naturally occurring genetic animal models
of absence seizures. These models, which include the Genetic
Absence Epilepsy in Rats from Strasbourg (GAERS) (Vergnes et al.,
1990) and the Wistar Albino Glaxo/Rijswijk rat (WAG/Rij) (van
Luijtelaar & Coenen, 1986; Peeters et al., 1990), are characterized by
generalized SW discharges at 6±10 Hz that respond to antiepileptic
drugs in a way similar to what has been reported in patients with
primary generalized epilepsy. These genetically determined absence
seizures have been analysed with microphysiological (including
intracellular) recordings that were obtained in vivo from cerebral
cortex and from different thalamic nuclei (Pinault et al., 1998, 2001;
Seidenbecher et al., 1998; Charpier et al., 1999; Seidenbecher &
Pape, 2001; Staak & Pape, 2001; Meeren et al., 2002; Slaght et al.,
2002). These studies, recently reviewed by Crunelli & Leresche
2002), have (i) shown the occurrence of speci®c patterns of cortical
Correspondence: Dr M. Avoli, 1Montreal Neurological Institute, as above.E-mail: [email protected]
Received 7 October 2002, accepted 14 October 2002
doi:10.1046/j.1460-9568.2002.02411.x
European Journal of Neuroscience, Vol. 16, pp. 2383±2393, 2002 ã Federation of European Neuroscience Societies
and thalamic neuron activity during SW discharges, (ii) characterized
the GABA receptor subtypes involved in the periods of neuronal
silence that are recorded in thalamic cells, and (iii) identi®ed a
primary role of the cortex in the initiation of SW discharges.
However, the relative contributions provided by thalamic and cortical
networks to the occurrence of SW discharges in these genetic models
of absence seizures remain to date unclear.
We have recently adapted the thalamocortical slice preparation
originally described in mice by Agmon & Connors (1991) to obtain
rat slices in which reciprocal thalamocortical connectivity is suf®-
ciently preserved to express rhythmic oscillations resembling sleep
spindles (Tancredi et al., 2000; Biagini et al., 2001). Here, we have
used this in vitro approach to study the thalamocortical interactions
occurring in epileptic (> 160 days old) WAG/Rij and in age-matched,
nonepileptic control (NEC) rats. To this end, we used simultaneous
®eld potential recordings in thalamocortical slices that were bathed in
medium containing zero [Mg2+], high [K+] (8.25 mM), or low
concentrations of 4-aminopyridine (4AP; 0.5±1 mM). Only 4AP
application proved to be capable of triggering oscillatory patterns
that were different in WAG/Rij and NEC animals. Therefore, we
focused on this in vitro model of thalamocortical synchronization to
identify the contributions of cerebral cortex and thalamus to these
rhythms and to characterize their sensitivity to excitatory amino acid
receptor antagonists.
Materials and methods
Epileptic WAG/Rij (> 160 day-old) and age-matched NEC rats (Crl/
Wi/br/Wistar obtained from Charles River) were used according to
the procedures established by the European Union and Canadian
Councils of Animal Care. By analysing the behaviour of the WAG/
Rij rats housed before the day of the in vitro experiment, we could
document that these animals displayed frequent episodes (duration 4±
10 s) of behavioural arrest accompanied by myoclonic twitches.
Previous studies performed in this genetic strain have demonstrated
that these clinical events correspond to bilaterally generalized SW
discharges at 6±10 Hz (van Luijtelaar & Coenen, 1986; Meeren et al.,
2002).
Both epileptic WAG/Rij and age-matched NEC rats were decapi-
tated under halothane anaesthesia, and their brains were quickly
removed and cooled in cold, oxygenated arti®cial cerebral spinal ¯uid
(ACSF; see below) in which NaCl was isoosmotically replaced with
sucrose to increase slice viability (Aghajanian & Rasmussen, 1989;
Zhang et al., 1996). Thalamocortical slices (550±650 mm) were
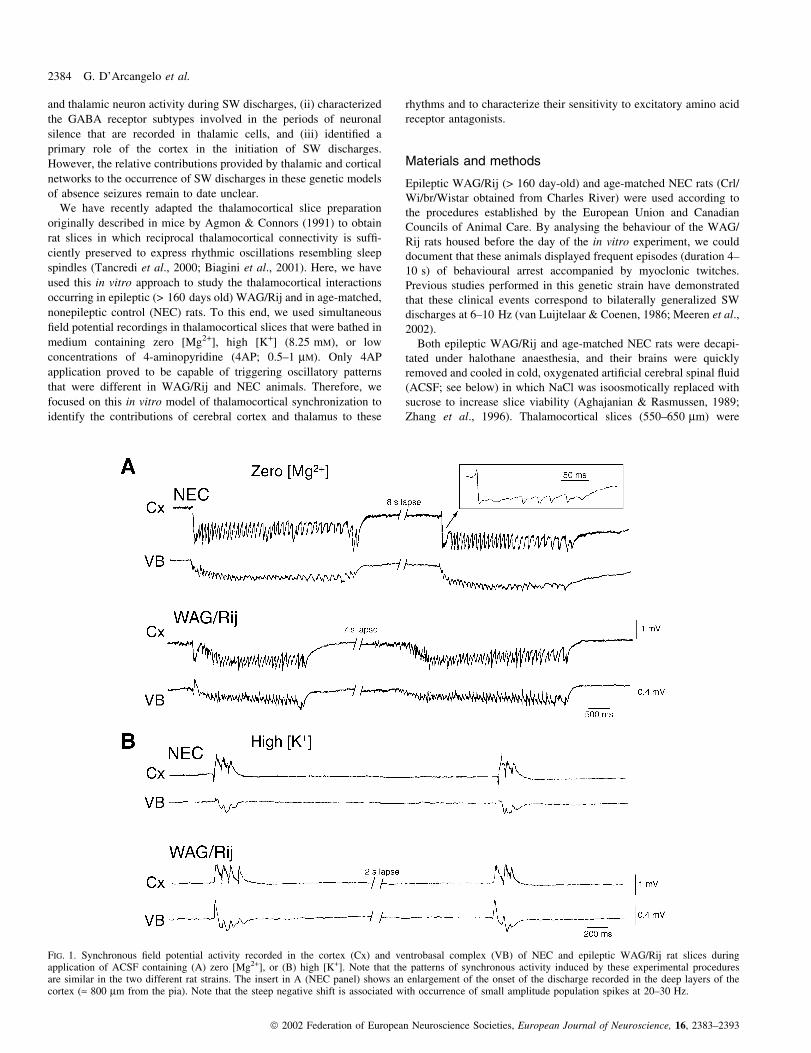
FIG. 1. Synchronous ®eld potential activity recorded in the cortex (Cx) and ventrobasal complex (VB) of NEC and epileptic WAG/Rij rat slices duringapplication of ACSF containing (A) zero [Mg2+], or (B) high [K+]. Note that the patterns of synchronous activity induced by these experimental proceduresare similar in the two different rat strains. The insert in A (NEC panel) shows an enlargement of the onset of the discharge recorded in the deep layers of thecortex (» 800 mm from the pia). Note that the steep negative shift is associated with occurrence of small amplitude population spikes at 20±30 Hz.
2384 G. D'Arcangelo et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

obtained using procedures previously described by us (Tancredi et al.,
2000; Biagini et al., 2001). Slices were transferred to a tissue
chamber where they lay in an interface between oxygenated ACSF
and humidi®ed gas (95%O2 : 5% CO2) at 32±35 °C (pH 7.4). ACSF
composition, unless otherwise speci®ed, was (in mM) NaCl, 124;
KCl, 2; KH2PO4, 1.25; MgSO4, 1; CaCl2, 2; NaHCO3, 26; and
glucose, 10. In some experiments MgSO4 was omitted to obtain a
nominally Mg2+- free medium (hereafter termed zero [Mg2+]
medium) while in other experiments KCl was elevated to 7 mM to
reach a total K+ concentration of 8.25 mM (high [K+] medium).
Chemicals were obtained from Sigma (St Louis, USA) with the
exception of the NMDA receptor antagonist 3,3-(2-carboxypiper-
azine-4-yl)-propyl-1-phosphonate (CPP), which was purchased from
Tocris Cookson (Langford, UK).
Field potentials were recorded with ACSF-®lled electrodes
(resistance 2±8 MW) which were positioned under visual control in
the cerebral cortex, the ventrobasal complex (VB) and/or the nucleus
reticularis of the thalamus (nRT). The cortical recording electrode
was positioned in most of the experiments at a depth presumably
corresponding to layer IV. Signals were fed to high-impedance
ampli®ers and were processed through second-stage ampli®ers with
®ltering capability. The high-pass and low-pass ®lters were often set
at 0.2 and 100 Hz, respectively. Field potential recordings were
displayed on a Gould recorder and at times digitized to obtain average
power spectra of selected epochs (overall length > 1 s). In some
experiments a bipolar stainless steel electrode was used to deliver
extracellular stimuli (50±150 ms; < 200 mA) to the cerebral cortex,
the thalamus or the internal capsule.
Pathway transections were made under direct visual guidance with
micromanipulator-mounted blades. In some experiments kynurenic
acid (5 mM, Sigma; diluted in ACSF) or CPP (0.2 mM, diluted in
ACSF) were pressure-applied to restricted areas of the slice by
employing a superfusion system similar to that used by Biagini et al.
(2001). In brief, a broken-tip theta glass pipette (diameter 1.5 mm)
was connected through two Te¯on tubes to a peristaltic pump with
¯ow rate set at » 0.5 mL/min and used as a push±pull cannula. This
pipette was mounted on a micromanipulator and positioned over the
cortex or the thalamus. The area of the slice covered by the drug
applied through the push±pull cannula was < 1 mm2. To avoid
diffusion of the drug from the superfused area to distant regions,
slices were placed in such a way that the treated area was downstream
with respect to the ACSF ¯ow. The spread of the applied solution was
visualized by including phenol red in the superfused medium.
Our work is based on data from 42 and 19 thalamocortical slices
that were obtained from epileptic WAG/Rij and NEC rat brains,
respectively. Measurements are expressed as mean 6 SD and n
indicates the number of experiments. Statistical analysis of the data
was performed with Student's t-test. Data were considered signi®-
cantly different if P < 0.05.
Results
Synchronous activity in NEC and WAG/Rij rat thalamocorticalslices
Thalamocortical slices obtained from either NEC or epileptic WAG/
Rij rats did not generate spontaneous ®eld potential activity when
bathed in normal ACSF (cf. Zhang et al., 1996; Tancredi et al., 2000;
Biagini et al., 2001). Therefore, we used experimental procedures
known to increase neuronal excitability in vitro to obtain synchronous
events similar to those found to occur spontaneously in vivo. In a ®rst
series of experiments we analysed the effects induced by super-
perfusing slices with zero [Mg2+] medium. As reported by Zhang
et al. (1996), Mg2+ wash-out induced sequences of ®eld potential
oscillations (FPOs) that had intraoscillatory frequencies ranging from
6.5 to 15 Hz, lasted 1.3±4 s and were recorded in synchronicity in
thalamus and cortex (Fig. 1A). The onset of these FPOs was often
characterized in cortical DC recordings by a steep negative shift
associated with a brief (< 200 ms) discharge of small-amplitude
population spikes at 20±30 Hz (Fig. 1A, expanded trace in the NEC
panel). Moreover, there was a tendency for the FPOs to decrease their
intraoscillatory frequency through the sequence. As summarized in
Table 1, FPOs induced by zero [Mg2+] medium had similar
characteristics (i.e. duration, rate of occurrence, and intraoscillatory
frequency) in thalamocortical slices obtained from epileptic WAG/
Rij (n = 5 slices) and NEC (n = 6) rats. During prolonged application
of zero [Mg2+] medium (particularly after 4±5 h), slices obtained
from either rat strain could produce synchronous epileptiform
discharges which closely resembled electrographic seizures (not
illustrated, but see Zhang et al., 1996).
Similarly to what was observed in slices superfused with zero
[Mg2+] medium, application of ACSF containing high [K+] induced
synchronous activity that was similar in the two types of tissue. In
both NEC (n = 3) and epileptic WAG/Rij (n = 4) thalamocortical
slices it consisted of brief (< 1 s) FPOs at 12±17 Hz which occurred
in both cortex and VB (Fig. 1B). However, the rate of occurrence of
these synchronous discharges was consistently higher in NEC slices,
even though this difference was not signi®cant (P < 0.08). The
quantitative characteristics of the spontaneous events induced by
superfusing NEC and epileptic WAG/Rij slices with high [K+]
medium are also summarized in Table 1.
Unlike the experiments performed with zero [Mg2+] or high [K+]
medium, bath application of low concentrations of 4AP (0.5±1 mM)
induced spontaneous FPOs which had strain-dependent features. In
NEC slices (n = 7), 4AP caused only one pattern of FPO, which
occurred in thalamus and/or cortex at 9.2±16.3 Hz (12.8 6 1.9 Hz,
n = 5) (Fig. 2A, NEC panel) (cf. Tancredi et al., 2000; Biagini et al.,
2001). In contrast, in epileptic WAG/Rij thalamocortical slices
(n = 23), 4AP induced two coexisting patterns of FPOs: one type was
characterized by frequencies of 9.5±16.1 Hz (13.1 6 2.1 Hz, n = 19)
while the other had frequencies ranging from 5.0 to 8.9 Hz
(7.7 6 1.1 Hz, n = 19) (Fig. 2A, WAG/Rij panel). These two types
of rhythms will be hereafter termed fast and slower FPOs, respect-
ively. It should be emphasized that in both NEC and epileptic WAG/
Rij thalamocortical slices the ranges of FPO frequencies were
narrower in any given experiment.
Power spectra obtained from segments of fast and slower FPOs
recorded in the cortex and thalamus of an epileptic WAG/Rij slice are
shown in Fig. 2B. By plotting the peak values of the power spectra
TABLE 1. Characteristics of the synchronous activity recorded in NEC and
epileptic WAG/Rij rat thalamocortical slices
Slices(n)
Intraoscillatoryfrequency (Hz)
Duration(s)
Occurrence(events/min)
Zero [Mg2+]NEC 6 7.8 6 2.5 1.9 6 0.7 4.7 6 0.6WAG/Rij 5 8.2 6 2.4 1.7 6 0.5 4.8 6 0.7
High [K+]NEC 3 14.3 6 0.6 0.23 6 0.15 27.6 6 2.9WAG/Rij 4 13.8 6 2.2 0.20 6 0.09 25.1 6 12.2
Slices were bathed in medium containing zero [Mg2+] or high [K+].Measurements were obtained from the cortical recording.
Thalamocortical oscillations in vitro 2385
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

obtained during segments of fast and slower FPOs recorded in the
cortex and VB of each of 19 WAG/Rij slices, we found that the
frequency of the two rhythms was highly correlated (r2 = 0.84) and
displayed a slower : fast frequency ratio of » 0.61 (Fig. 2C). In
addition, slower FPOs were characterized by amplitudes that were
larger than those seen in the same experiment during the fast FPOs
(Figs 2A and 3). As illustrated in Fig. 2A (WAG/Rij panel), the
slower FPOs were often identi®ed ®rst in the cortical recording,
becoming apparent in the thalamic area after 1±3 cycles. However,
this ®nding depended on the cortical and thalamic sites from which
recordings were obtained. Further quantitative details of the FPOs
induced by 4AP in the two types of thalamocortical slices are
provided in Table 2.
Fast FPOs generated by epileptic WAG/Rij or NEC slices could
often occur independently in cortex and thalamus, and appeared
simultaneously in these two structures only when the recording
electrodes were placed within speci®c areas of the cortical or
thalamic sectors (Figs 2 and 3A). In contrast, the slower FPOs
recorded in epileptic WAG/Rij slices were often synchronous in both
structures. This characteristic is further shown in Fig. 3A, which
illustrates fast and slower FPOs successively recorded from different
cortical and thalamic areas in the same slice. As illustrated in Fig. 3B,
FIG. 2. An application of 4AP discloses different patterns of ®eld potential oscillations in the somatosensory cortex (Cx) and ventrobasal complex (VB) ofNEC and epileptic WAG/Rij thalamocortical slices. (A) Only fast FPOs (9.3±10 Hz) are recorded from the NEC slice; in addition, these oscillations occur(a) independently in Cx and VB unless (b) recording sites with intact functional connectivity are identi®ed. In contrast, both fast (10±14 Hz) and slower (6.9±7.8 Hz) FPOs are recorded in an epileptic WAG/Rij thalamocortical slice; note that in this experiment as well, fast FPOs occur independently in Cx and VB,while slower FPOs occur synchronously in cortex and thalamus at these recording sites. Note also that the sequence of slower FPOs in this speci®c sampleappears to initiate in the cortical recording. (B) Power spectra of the rhythmic oscillatory activity recorded in the epileptic WAG/Rij rat slice shown in A andlabelled 1±4. The peak values of the power spectra are shown in each panel. (C) Plot of the power spectrum peaks of fast and slower FPOs recorded in thecerebral cortex (Cx) and the ventrobasal complex (VB) in each of 19 experiments. Note that there is a close relationship (r2 = 0.84) between the peaks of thefast and slower FPOs, with a ratio of » 0.6.
2386 G. D'Arcangelo et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

both fast and slower FPOs generated by epileptic WAG/Rij
thalamocortical slices were also recorded in nRT (n = 5), where
they appeared to be synchronized with those seen in VB. In these
experiments as well, the slower FPOs recorded in nRT displayed
amplitudes that were larger than those seen during fast FPOs.
Moreover, fast FPOs occurred synchronously in nRT and VB even
when they were not re¯ected in the cortical recordings (Fig. 3B,
asterisk). In these experiments we also found that single-shock
electrical stimuli delivered into the white matter could readily elicit
slower FPOs that were similar to those occurring spontaneously
(Fig. 3B, triangle).
Contribution of thalamic and cortical networks to 4AP-inducedoscillations in epileptic WAG/Rij slices
Having established that 4AP application can disclose different
patterns of thalamocortical activity in NEC and epileptic WAG/Rij
slices, we analysed the role played by cortical and thalamic networks
in elaborating the FPOs recorded in epileptic WAG/Rij rat slices.
First, we studied the changes caused by surgical isolation of the
cerebral cortex and thalamus (n = 5) on both spontaneous and
stimulus-induced slower FPOs. As shown in Fig. 4, isolated cortical
networks did not generate either fast or slower FPOs. In contrast, after
cutting the connections between the cerebral cortex and thalamus we
FIG. 3. (A) Different degrees of synchronization characterize fast (column a) and slower (column b) FPOs induced by 4AP application in an epileptic WAG/Rij thalamocortical slice. Position of the recording electrodes in the frontoparietal cortex (Cx) and in VB are numbered in the insert. Note that fast SLOs aresynchronized in cortex and thalamus only in panel a2, while synchronization is always present for the slower FPOs. (B) Slow speed chart recording of thespontaneous ®eld potential activity recorded in the cerebral cortex (Cx), ventrobasal complex (VB) and nucleus reticularis thalami (nRT) of an epilepticWAG/Rij rat thalamocortical slice during 4AP application. Note that synchronous, fast FPOs are recorded in both thalamic locations, but not in the cortex;also, slower FPOs occur in all recordings both spontaneously and in response to single-shock electrical stimulation of the white matter (m).
TABLE 2. Characteristics of the synchronous activity induced by 4APin NEC and epileptic WAG/Rij rat thalamocortical slices
Slices(n)
Intraoscillatoryfrequency (Hz)
Duration(s)
Occurrence(events/min)
Fast Slower Fast Slower Fast Slower
NEC 7 12.8 6 1.9 ± 2.1 6 0.54 ± 5.7 6 1.3 ±WAG/Rij 19 13.1 6 2.1 7.7 6 1.1 1.9 6 0.6 3.2 6 0.9 2.6 6 1.0 3.4 6 0.8
Measurements were obtained from the cortical recording.
Thalamocortical oscillations in vitro 2387
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

could still record sequences of fast FPOs in VB and nRT, both
spontaneously and following single-shock electrical stimuli delivered
in the thalamus (Fig. 4).
Next, we used local application of the glutamatergic antagonist
kynurenic acid to cortical or thalamic areas of the slice to selectively
decrease the excitability of either neuronal network, and thus to
establish their contribution to the FPOs recorded in epileptic WAG/
Rij slices. As illustrated in Fig. 5A, applying kynurenic acid to the
cortex (n = 4) abolished both fast and slower FPOs in the cerebral
cortex while only fast FPOs continued to occur in the thalamic areas.
FIG. 4. Effects induced by cutting the connections between the cerebral cortex and thalamus on the slow ®eld potentials oscillations induced by 4AP in anepileptic WAG/Rij rat thalamocortical slice. (A) Drawings depict the thalamocortical slice under control conditions (left panel) and after complete separationof the thalamus from the cortex with two cuts performed with a micromanipulator-mounted blade (right panel). Recordings were obtained simultaneouslyfrom the cortex (Cx), the ventrobasal complex (VB) and the nucleus reticularis (nRT). FPOs shown in B occurred spontaneously before and after the cut,while those in C were induced by single electrical stimuli delivered into the internal capsule (m) or into the white matter underlying the cortex (d). Notethat, after surgical separation, both spontaneous and stimulus-induced FPOs are absent from the cortex, while they are still observed in VB and nRT, thoughwith higher frequencies than under control conditions.
2388 G. D'Arcangelo et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

Hence, depressing cortical excitability transformed thalamic slower
FPOs into fast FPOs. In contrast, application of this glutamatergic
antagonist to the thalamus made both fast and slower FPOs disappear
in thalamic and cortical areas (n = 3) (Fig. 5B). Under these
conditions focal electrical stimuli in the internal capsule induced
®eld potential responses in the cerebral cortex and VB that lasted
< 180 ms and were characterized by an isolated negative or positive±
negative transient (Fig. 5B, triangle in `Kynurenic acid in VB + nRT'
panel). The effects induced by either cortical or thalamic application
of kynurenic acid were reversible following removal of the pipette
and 20±30 min of washout of the glutamatergic receptor antagonist
from the site of application (not shown). The graphs in Fig. 5C
summarize the results obtained in these experiments.
NMDA receptor-dependent characteristics of the 4AP-inducedoscillations
Sequences of slower FPOs induced by 4AP in WAG/Rij thalamo-
cortical slices (n = 6) became less frequent and eventually disap-
peared when increasing concentrations of the NMDA receptor
antagonist CPP were added to the bathing medium. The blockade
of these slower FPOs was seen concomitantly in cortex and VB, and
it was paralleled by an increase in the recurrence of fast FPOs; such
an effect was more striking with CPP concentrations that completely
abolished the slower oscillatory activity (Fig. 6). In contrast, no
measurable changes were observed when similar concentrations of
CPP were tested in NEC slices on the 4AP-induced fast FPOs (n = 4)
(Fig. 6).
In order to identify the site of action of CPP within the WAG/Rij
thalamocortical slice, we administered this NMDA receptor antag-
onist locally in the cerebral cortex or VB + nRT. In six experiments,
application of CPP to the cerebral cortex markedly decreased and
eventually abolished slower FPOs, while the occurrence of fast FPOs
displayed a tendency to increase (Fig. 7A and C). In contrast, local
application of CPP to the thalamic areas (n = 7) did not modify the
rate of occurrence of either fast or slower FPOs (Fig. 7B and C).
Discussion
We have found that rat thalamocortical slices maintained in vitro in
the presence of minimal concentrations of 4AP generated patterns of
thalamocortical oscillations with electrographic features which
depend on the animal strain. Speci®cally, fast FPOs could occur in
both types of tissue, while slower FPOs were only recorded in
epileptic WAG/Rij rat slices. In addition, slower FPOs were of larger
amplitude than the fast FPOs in both cortex and thalamus, suggesting
that larger neuronal populations were presumably involved in the
slower thalamocortical rhythm. In keeping with this view, cortical
and thalamic slower FPOs occurred in a synchronous fashion over
much of the cortical and thalamic areas investigated with ®eld
FIG. 5. Effects induced by local application of the glutamatergic antagonist kynurenic acid to the cortex or thalamus on FPOs recorded in two slices obtainedfrom epileptic WAG/Rij rats. (A) Kynurenic acid application to the cortex abolishes slower FPOs while fast FPOs continue to occur in VB. (B) In contrast,kynurenic acid application to the thalamus (VB + nRT) makes both fast and slower FPOs disappear in thalamic and cortical areas; note that under theseconditions focal electrical stimuli in the internal capsule (m) induces an isolated ®eld potential response lasting > 200 ms. (C) Summary of the effects inducedby kynurenic acid application to the cortex (n = 4 slices) and thalamus (n = 3) on the fast and slower FPOs recorded in cortex (upper panel) and VB (lowerpanel).
Thalamocortical oscillations in vitro 2389
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

potential recordings, but fast FPOs were synchronized only when
recording from speci®c locations (cf. Tancredi et al., 2000; Biagini
et al., 2001).
Epileptic WAG/Rij rats present with ictal behavioural and EEG
manifestations that are similar to those seen in patients with absence
seizures (van Luijtelar & Coenen, 1986; Peeters et al., 1990), even
though the generalized SW activity in this and in another well-
characterized genetic rat model (i.e. the GAERS) is characterized by
intraburst frequencies that are higher than in humans (i.e. 6±10 Hz vs.
2.5±4 Hz, respectively). Indeed, Destexhe et al. (1999) have proposed
by using mathematical modelling of SW discharges that this may be
caused by differences in low-threshold Ca2+ current and/or GABAB
receptor function. In vivo studies have shown that generalized SW
discharges in WAG/Rij rats re¯ect highly synchronized oscillations in
thalamocortical networks (Staak & Pape, 2001; Meeren et al., 2002).
Similar ®ndings have been obtained in GAERS rats (Seidenbecher
et al., 1998) and in the feline generalized penicillin (FGPE) model
(Avoli et al., 1983), in which transformation of sleep spindles into
generalized SW discharges is accompanied by a progressive increase
in ®ring correlation between cortical and thalamic cells (McLachlan
et al., 1984). Therefore, even though generalized SW discharges
could not (for obvious reasons) be reproduced in the slice preparation,
we are inclined to propose that the strain-dependent ability to
generate slower SLOs, along with their high degree of thalamocor-
tical synchronization, may represents an in vitro hallmark of the
epileptogenicity that characterizes WAG/Rij rats.
The oscillatory patterns recorded in epileptic WAG/Rij rat
thalamocortical slices, regardless of their frequency and degree of
synchronicity, require the function of thalamic networks, whose
output activity must reach cortical targets. Accordingly, no rhythmic
oscillatory activity was generated by cortical neurons when the cortex
was surgically separated from the thalamus or when the broad
spectrum glutamatergic antagonist kynurenic acid was locally applied
to VB-nRT. Similar ®ndings have been reported by us while studying
fast thalamocortical oscillations induced by 4AP in slices obtained
from young control rats (Tancredi et al., 2000). Moreover, early work
in the FGPE model has demonstrated that both EEG spindles and
generalized SW discharges disappear during transient depression of
thalamic function by local microinjection of KCl (Avoli & Gloor,
1981). It is, indeed, well established that spindle activity originates in
the thalamus and relies on the function of nRT networks (Steriade
et al., 1985). Interestingly, nRT cells are also intimately involved in
the generalized SW discharges recorded in GAERS rats (Slaght et al.,
2002). It should, however, be emphasized that, after intracortical
injection of bicuculline, SW activity at 2±3 Hz is recorded in the
cortex of cats which had undergone total ipsilateral thalamectomy
with abolition of sleep spindles (Steriade & Contreras, 1998) Hence,
these data demonstrate that in this experimental model cortical
networks may sustain SW discharges even in the absence of
thalamocortical inputs.
An important issue in the pathophysiogenesis of generalized SW
discharges in patients with absence epilepsy is whether the
synchronous thalamocortical oscillations that accompany the electro-
graphic pattern originate in cortex or thalamus. We have reported
here that only fast FPOs were recorded from VB or nRT after surgical
separation from the cortex and that slower FPOs disappeared in all
areas during cortical application of a glutamatergic receptor antag-
onist. Therefore, our data suggest that the ability of epileptic WAG/
Rij thalamocortical slices to generate slower FPOs depends on the
involvement of cortical networks in this type of rhythm and
speci®cally on their ability to drive both relay (i.e. VB) and nRT
cells via corticothalamic inputs (Deschenes & Hu, 1990; Deschenes
et al., 1998). This view is in keeping with the ability of cortical
spreading depression to change thalamic SW discharges into spindles
in cats injected with large doses of penicillin (Avoli & Gloor, 1982a).
Moreover, penicillin-induced SW discharges in cats are absent in the
thalamus following removal of the cortex (Avoli & Gloor, 1982b).
FIG. 6. Summary of the effects induced by bath application of twoconcentrations of the NMDA receptor antagonist CPP on the rate ofoccurrence of fast and slower FPOs in epileptic WAG/Rij thalamocorticalslices (n = 4) and on the fast FPOs recorded in NEC tissue (n = 3). Notethat CPP reduced and eventually abolished the slower FPOs in epilepticWAG/Rij thalamocortical slices, whereas fast FPOs increased their rate ofoccurrence. In contrast, CPP did not modify the pattern of fast FPOsrecorded in NEC slices. *P < 0.05.
2390 G. D'Arcangelo et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

Recent evidence obtained in slices of the ferret visual thalamus has
demonstrated that the frequency of thalamic network oscillations is
changed by varying the strength of corticothalamic tract stimuli that
were triggerd by the spontaneous activity of thalamic relay cells in
the geniculate nucleus (Bal et al., 2000; Blumenfeld & McCormick,
2000). In particular, it has been shown that weak feedback activation
leads to oscillations at 6±10 Hz, while strong corticothalamic
feedback stimuli cause a more synchronized, slower (2±4 Hz)
oscillatory process. Intracellular recordings obtained in vivo from
GAERS have shown that during SW discharges nRT cells generate
robust discharges of action potentials (presumably in response to
corticothalamic inputs) that cause a tonic hyperpolarization in the
majority of thalamocortical neurons (Pinault et al., 1998; Slaght et al.,
2002). These data are in keeping with an earlier intracellular study of
the activity of thalamocortical neurons in anaesthetized cats that
spontaneously developed SW activity at 2±4 Hz (Steriade &
Contreras, 1995). It was proposed in this study that the hyperpolar-
ization of thalamocortical cells and the concomitant ®ring silence
may represent the cellular correlate of the unresponsiveness seen
during absence seizures.
A leading role of cortical networks in generalized SW discharges
of epileptic WAG/Rij rats has also been proposed by Meeren et al.
(2002), who have employed non-linear association analysis of the
EEG signals recorded in vivo from multiple thalamic and cortical
sites during SW discharges. Interestingly, these experiments have
identi®ed a `consistent cortical focus' of SW activity within the peri-
oral region of the somatosensory cortex. Indeed, this cortical area was
presumably included in most of the thalamocortical slices used by us
in the present study. It must be emphasized that both the cortical
origin of SW discharges and the ability of cortical networks to lead
the thalamus during this rhythm have been identi®ed in several
studies performed in cats by Steriade and collaborators (Steriade &
Amzica, 1994; Steriade & Contreras, 1995; Neckelmann et al.,
1998).We have also established here that the generation of slower
FPOs in epileptic WAG/Rij thalamocortical slices depends on the
function of NMDA receptor-mediated mechanisms. In line with this
view, slower FPOs were no longer recorded during bath application
of the competitive NMDA receptor antagonist CPP, while the
occurrence of fast FPOs increased. Interestingly, CPP application to
NEC thalamocortical slices treated with 4AP did not cause any
change in the rate of occurrence of the fast FPOs, thus suggesting that
the increase in fast FPO occurrence in epileptic WAG/Rij rats during
CPP application may re¯ect the block of slower FPOs. NMDA
receptors have been proposed as contributing to SW discharge
generation in epileptic WAG/Rij (Peeters et al., 1989, 1990) and
GAERS (Pumain et al., 1992; Koerner et al., 1996) rats. However, it
was unclear from these in vivo studies whether NMDA dysfunction
resided in the cortex (Pumain et al., 1992), the thalamus (Koerner
et al., 1996) or, perhaps, in both structures. By using local application
of CPP to cortex or thalamus we have demonstrated that slower FPOs
induced by 4AP in epileptic WAG/Rij thalamocortical slice disappear
only when CPP is cortically applied. Therefore, these data demon-
strate that NMDA receptor-mediated mechanisms responsible for the
occurrence of slower FPOs should reside in the cortex. In addition,
this pharmacological evidence further supports the role of cortical
networks in generating slower FPOs, and suggests that NMDA
receptors do not play any major role in the genesis of fast FPOs.
It should be emphasized that the speci®c role played by NMDA
receptor-mediated mechanisms in the genesis of slow FPOs in WAG/
Rij rat slices is supported by the inability of the other two procedures
tested in this study to elicit different, strain-dependent patterns of
thalamocortical oscillations. It is well established that both zero
[Mg2+] and, to a lesser extent, medium containing high [K+], lead to
activation of NMDA receptor-mediated conductances. Hence, both
FIG. 7. (A and B) Effects induced by local application of CPP to the cerebral cortex and thalamus (VB + nRT) of an epileptic WAG/Rij thalamocortical slicethat was continuously superfused with 4AP-containing medium. Note that during application of CPP to the cortex, slower FPOs disappeared in both cortexand thalamus while the fast FPOs continued to occur in both areas. (C) Summary histograms of the rate of occurrence of 4AP-induced fast and slower FPOsrecorded in cerebral cortex (upper panel) and VB (lower panel) of epileptic WAG/Rij thalamocortical slices under control conditions and during localapplication of CPP to the cortex (n = 4) or to VB + nRT (n = 5).
Thalamocortical oscillations in vitro 2391
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

manipulations could mask the subtle differences in NMDA receptor
function that are present in the cortex of WAG/Rij rat as compared
with NECs.
In conclusion, we have demonstrated here that epileptic WAG/Rij,
but not NEC, rat thalamocortical slices can produce sequences of
slower oscillatory rhythms during bath application of 4AP. The
occurrence of these events requires reciprocally connected and
functional thalamic and cortical networks. In particular, our data
indicate that thalamic networks are necessary to provide rhythmo-
genesis that, however, is not expressed at a slower intraoscillatory
frequency unless corticothalamic inputs are operant. Therefore, an
important determinant for the production of generalized SW
discharges in the WAG/Rij rat resides in the cortex which is
overexcitable (Luhmann et al., 1995) and endowed with the ability to
promote a fast intracortical spread of the epileptiform activity
(Meeren et al., 2002). Overall, as originally proposed by Steriade &
Contreras (1998), our data point at the cortex as a target for
antiepileptic drug action in this genetic rodent model of generalized
SW discharges and presumably in human absence epilepsy.
Acknowledgements
This study was supported by the Canadian Institutes of Health Research (GrantMOP-43964), the National Research Council of Italy and the SavoyFoundation. M.D. and R.A.W. were recipients of a Fellowship from theFragile X Research Foundation of Canada and of a Scholarship from the Fondsde la Recherche en Sante du QueÂbec, respectively. We thank Dr Florin Amzicafor reading an early draft of this paper and Toula Papadopoulos for secretarialassistance.
Abbreviations
4AP, 4-aminopyridine; ACSF, arti®cial cerebrospinal ¯uid; CPP, 3,3-(2-carboxypiperazine-4-yl)-propyl-1-phosphonate; FPOs, ®eld potential oscilla-tions; GAERS, Genetic Absence Epilepsy in Rats from Strasbourg; NEC,nonepileptic control; nRT, nucleus reticularis of the thalamus; SW, spike andwave; VB, ventrobasal complex; WAG/Rij, Wistar Albino Glaxo/Rijswijk.
References
Aghajanian, G.K. & Rasmussen, K. (1989) Intracellular studies in the facialnucleus illustrating a simple new method for obtaining viable motoneuronsin adult rat brain slices. Synapse, 3, 331±338.
Agmon, A. & Connors, B.W. (1991) Thalamocortical responses of mousesomatosensory (barrel) cortex in vitro. Neuroscience, 41, 365±379.
Avoli, M. & Gloor, P. (1981) The effects of transient functional depression ofthe thalamus on spindles and on bilateral synchronous epileptic dischargesof feline generalized penicillin epilepsy. Epilepsia, 22, 443±452.
Avoli, M. & Gloor, P. (1982a) Interaction of cortex and thalamus in spike andwave discharges of feline generalized penicillin epilepsy. Exp. Neurol., 76,96±217.
Avoli, M. & Gloor, P. (1982b) Role of the thalamus in generalized penicillinepilepsy: observations on decorticated cats. Exp. Neurol., 77, 386±402.
Avoli, M., Gloor, P., Kostopoulos, G. & Gotman, J. (1983) An analysis ofpenicillin-induced generalized spike and wave discharges usingsimultaneous recordings of cortical and thalamic single neurons. J.Neurophysiol., 50, 819±837.
Avoli, M., Gloor, P., Kostopoulos, G. & Naquet, R. (1990) GeneralizedEpilepsy: Neurobiological Approaches. Birkhauser, Boston.
Avoli, M., Rogawski, M.A. & Avanzini, G. (2001) Generalized epilepticdisorders: an update. Epilepsia, 42, 445±457.
Bal, T., Debay, D. & Destexhe, A. (2000) Cortical feedback controls thefrequency and synchrony of oscillations in the visual thalamus. J. Neurosci.,20, 7478±7488.
Biagini, G., D'Antuono, M., Tancredi, V., Motalli, R., Louvel, J.,D'Arcangelo, G., Pumain, R., Warren, R. & Avoli, M. (2001)Thalamocortical connectivity in a rat brain slice preparation: Participation
of the ventrobasal complex to synchronous activities. Thalamus RelatedSystems, 1, 169±179.
Blumenfeld, H. & McCormick, D.A. (2000) Corticothalamic inputs control thepattern of activity generated in thalamocortical networks. J. Neurosci., 20,5153±5162.
Charpier, S., Leresche, N., Deniau, J.M., Mahon, S., Hughes, S.W. & Crunelli,V. (1999) On the putative contribution of GABAB receptors to the electricalevents occurring during spontaneous spike and wave discharges.Neuropharmacology, 38, 1699±1706.
Crunelli, V. & Leresche, N. (2002) Childhood absence epilepsy: genes,channels, neurons and networks. Nature Rev. Neurosci., 3, 371±382.
Deschenes, M. & Hu, B. (1990) Electrophysiology and pharmacology of thecorticothalamic input to lateral thalamic nuclei: an intracellular study in thecat. Eur. J. Neurosci., 2, 140±152.
Deschenes, M., Veinante, P. & Zhang, Z.W. (1998) The organization ofcorticothalamic projections: reciprocity versus parity. Brain. Res. Rev., 28,286±308.
Destexhe, A., McCormick, D.A. & Sejnowski, T.J. (1999) Thalamic andthalamocortical mechanisms underlying 3 Hz spike-wave discharges. Prog.Brain Res., 121, 289±307.
Koerner, C., Danober, L., Boehrer, A., Marescaux, C. & Vergnes, M. (1996)Thalamic NMDA transmission in a genetic model of absence epilepsy inrats. Epilepsy Res., 25, 11±19.
Lennox, W.G. & Lennox, M.A. (1960) Epilepsy and Related Disorders. LittleBrown, Boston.
Luhmann, H.J., Mittmann, T., van Luijtelaar, G. & Heinemann, U. (1995)Impairment of intracortical GABAergic inhibition in a rat model of absenceepilepsy. Epilepsy Res., 22, 43±51.
van Luijtelaar, E.L.J.M. & Coenen, A.M.L. (1986) Two types ofelectrocortical paroxysms in an inbred strain of rats. Neurosci. Lett., 70,393±397.
McLachlan, R.S., Avoli, M. & Gloor, P. (1984) Transition from spindles togeneralized spike and wave discharges in the cat: simultaneous single cellrecordings in cortex and thalamus. Exp. Neurol., 85, 413±425.
Meeren, H.K., Pijn, J.P., Van Luijtelaar, E.L., Coenen, A.M. & Lopes da Silva,F.H. (2002) Cortical focus drives widespread corticothalamic networksduring spontaneous absence seizures in rats. J. Neurosci., 22, 1480±1495.
Metrakos, K. & Metrakos, J.D. (1961) Genetics of convulsive disorders, II:genetic and electroencephalographic studies in centrencephalic epilepsy.Neurology, 11, 464±483.
Necklmann, D., Amizida, F. & Steriade, M. (1998) Spike-wave complexes andfast components of cortically generated seizures. III. J. Neurophysiol., 80,1480±1494.
Peeters, B.W.M.M., van Rijn, C.M., van Luijtelaar, E.L.J.M. & Coenen,A.M.L. (1989) Antiepileptic and behavioural actions of MK-801in an animal model of spontaneous absence epilepsy. Epilepsy Res., 3,178±181.
Peeters, B.W.M.M., van Rijn, C.M., Vossen, J.M.H. & Coenen, A.M.L. (1990)Involvement of NMDA receptors in non-convulsive epilepsy in WAG/Rijrats. Life. Sci., 47, 523±529.
Pinault, D., Leresche, N., Charpier, S., Deniau, J.-M., Marescaux, C., Vergnes,M. & Crunelli, V. (1998) Intracellular recordings in thalamic neuronesduring spontaneous spike and wave discharges in rats with absenceepilepsy. J. Physiol. (Lond.), 509, 449±456.
Pinault, D., Vergnes, M. & Marescaux, C. (2001) Medium-voltage 5±9-Hzoscillations give rise to spike-and-wave discharges in a genetic model ofabsence epilepsy: in vivo dual extracellular recording of thalamic relay andreticular neurons. Neuroscience, 105, 181±201.
Pumain, R., Louvel, J., Gastard, M., Kurcewicz, I. & Vergnes, M. (1992)Responses to N-methyl-D-aspartate are enhanced in rats with Petit-mal-likeseizures. J. Neural. Transm. Suppl., 35, 97±108.
Seidenbecher, T. & Pape, H.C. (2001) Contribution of intralaminar thalamicnuclei to spike-and-wave-discharges during spontaneous seizures in agenetic rat model of absence epilepsy. Eur. J. Neurosci., 13, 1537±1546.
Seidenbecher, T., Staak, R. & Pape, H.C. (1998) Relations between corticaland thalamic cellular activities during absence seizures in rats. Eur. J.Neurosci., 10, 1103±1112.
Slaght, S.J., Leresche, N., Deniau, J., Crunelli, V. & Charpier, S. (2002)Activity of thalamic reticular neurons during spontaneous geneticallydetermined spike and wave discharges. J. Neurosci., 22, 2323±2334.
Snead, O.C. (1995) Basic mechanisms of generalized absence seizures. Ann.Neurol., 37, 146±157.
Staak, R. & Pape, H.C. (2001) Contribution of GABA (A) and GABA (B)receptors to thalamic neuronal activity during spontaneous absence seizuresin rats. J. Neurosci., 21, 1378±1384.
2392 G. D'Arcangelo et al.
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393

Steriade, M. & Amzica, F. (1994) Dynamic coupling among neocorticalneurons during evoked and spontaneous spike-wave seizure activity. J.Neurophysiol., 72, 2051±2069.
Steriade, M. & Contreras, D. (1995) Relations between cortical and thalamiccellular events during transition from sleep patterns to paroxysmal activity.J. Neurosci., 15, 623±642.
Steriade, M. & Contreras, D. (1998) Spike-wave complexes and fastcomponents of cortically generated seizures. I. Role of neocortex andthalamus. J. Neurophysiol., 80, 1439±1455.
Steriade, M., Deschenes, M., Domich, L. & Muelle, C. (1985) Abolition ofspindle oscillations in thalamic neurons disconnected from nucleusreticularis thalami. J. Neurophysiol., 54, 1473±1497.
Tancredi, V., Biagini, G., D'Antuono, M., Louvel, J., Pumain, R. & Avoli, M.(2000) Spindle-like thalamocortical synchronization in a rat brain slicepreparation. J. Neurophysiol., 84, 1093±1097.
Vergnes, M., Marescaux, C., Depaulis, A., Micheletti, G. & Warter, J.M.(1990) Spontaneous spike-wave discharges in wistar rats: a model of geneticgeneralized convulsive epilepsy. In Avoli, M., Gloor, P., Kostopoulos, G. &Naquet, R. (eds.), Generalized Epilepsy: Neurobiological Approaches.BirkhaÈuser, Boston, pp. 238±253.
Zhang, Y.-F., Gibbs, J.W. III & Coulter, D.A. (1996) Anticonvulsant effectson spontaneous thalamocortical rhythms in vitro: ethosuximide,trimethadione and dimethadione. Epilepsy Res., 23, 15±36.
Thalamocortical oscillations in vitro 2393
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 2383±2393




















