
Burkholderia phytofirmans PsJN Acclimates Grapevine to Cold by Modulating Carbohydrate Metabolism
Upload
independentCategory
view
0download
0
Temporal variability of incidental solar radiation and modulating factorsin a coastal upwelling area (36!S)
Klaudia Hernández a,b,⇑, Beatriz Yannicelli c, Aldo Montecinos d, Marcel Ramos c,Humberto E. González b,e,f, Giovanni Daneri b,e
a Centro de Investigación en Nutrición y Sustentabilidad (CIEN Austral), Casilla correo 1327, Universidad Austral, Campus Pelluco, Puerto Montt, Chileb Centro de Investigaciones Oceanográficas del Pacífico Sudoriental (COPAS), Casilla correo 160-C, Universidad de Concepción, Concepción, Chilec Centro de Estudios Avanzados en Zonas Áridas (CEAZA), Facultad de Ciencias del Mar, Universidad Católica del Norte, Campus Guayacán, Larrondo 1281, Coquimbo, Chiled Departamento de Geofísica (DGEO), Casilla correo 160-C, Universidad de Concepción, Concepción, Chilee Centro de Investigación en Ecosistemas de la Patagonia (CIEP), Francisco Bilbao 449, Coyhaique, Chilef Instituto de Biología Marina, Universidad Austral de Chile, Casilla 567, Valdivia, Chile
a r t i c l e i n f o
Article history:Available online 28 July 2011
a b s t r a c t
In this study, we quantify the annual and synoptic variability of solar radiation (PAR and UVR) forsouthern-central Chile at !36.5!S from 2003 to 2004 and we relate these fluctuations to total ozoneconcentration, cloudiness, and winds. Data considered approximately 2 years of daily visible PAR(400–700 nm), UVA (400–320 nm), and UVB (280–320 nm). Satellite ozone data were also comparedwith a 305/340 ratio. On an annual scale, the visible and UV radiation co-varied in phase, with maximain spring–summer and lower values in autumn–winter. Winds and cloudiness also showed an annualcycle, with predominantly southerly winds and lower cloudiness in summer. On a synoptic scale, solarradiation was coupled with cloudiness and winds, a relationship that is also evidenced over large scaleatmospheric circulation. A 3-month lag was observed between solar radiation and ozone annual cycles;the latter peaked in springtime. The 305/340 ratio was a good ozone index over short time scales, show-ing fluctuations at the same frequencies as ozone and good cross-correlations once the annual cycle wasextracted. The spatial–temporal analysis supports the possibility that the local pattern observed might becommon for other important upwelling sites affected by the same type of atmospheric variability. Finally,this study showed that solar radiation wavelengths fluctuating at similar frequencies as those of upwell-ing favorable winds, are likely to be modulating biological processes in the coastal upwelling area offConcepción over short time scales.
" 2011 Elsevier Ltd. All rights reserved.
1. Introduction
The energy from solar radiation, which acts on several spatialand temporal scales, is the most important factor forcing marineand atmospheric global circulation. Locally (bearing in mind tensof km), solar radiation is related to many coupled bio-physical pro-cesses in the marine ecosystem (McKenzie et al., 2003). On annualand daily scales, coastal circulation and mixing are affected by thedifferential heating/cooling cycles of the sea surface and the adja-cent continent. Besides the direct effect of solar radiation on bio-logical processes, mixing and recirculation also affect productionin coastal marine ecosystems by redistributing nutrients to surface
layers and exporting/retaining organic matter and nutrients. To-gether, these processes shape the mesoscale pelagic and benthicbiological realms.
The irradiance reaching the coastal ocean surface at any giventime (i.e. on an annual or seasonal scale) depends on astronomic,atmospheric, and geographic factors (Seidlitz et al., 2001; McKen-zie et al., 2003), and its penetration in the water column is relatedto stratification and bio-optical conditions (Stramska and Dickey,1998; Neale et al., 2003; Montecino et al., 2004). Four variablesare then strongly related to surface irradiance variability: zenithangle, atmospheric ozone and other gas concentrations, and cloudcover (Cabrera et al., 1995; Lovengreen et al., 2000; Häder et al.,2007). The strong fluctuations in visible (PAR) and UVR spectraobserved from site to site in coastal areas at different latitudes, re-sult from the interaction of broad scale atmospheric circulationand local topography, which regulate cloud cover and, among otherfactors, the surface solar radiation reaching the earth’s surface(Borkowski, 2000; Liao and Frederick, 2005). For instance strato-spheric ozone and cloud cover account for over 70% of surface
0079-6611/$ - see front matter " 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.pocean.2011.07.011
⇑ Corresponding author at: Centro de Investigación en Nutrición y Sustentabil-idad CIEN Austral-Universidad Austral, Campus Pelluco, Puerto Montt Chile, Casilla1327, Chile. Tel.: +56 65 48 7438; fax: +56 65 25 5583.
E-mail addresses: [email protected] (K. Hernández), [email protected] (B. Yannicelli), [email protected] (A. Montecinos), [email protected] (M. Ramos), [email protected] (H.E. González), [email protected](G. Daneri).
Progress in Oceanography 92–95 (2012) 18–32
Contents lists available at SciVerse ScienceDirect
Progress in Oceanography
journal homepage: www.elsevier .com/locate /pocean
UVR variability in Valdivia (39!S, see Lovengreen et al., 2005).Cloud cover is also dependent on ocean–atmospheric interactions(e.g. Garreaud et al., 2008), so incident solar radiation variabilityat coastal sites could feedback on ocean variability, at several fre-
quencies. Understanding how patterns of ground solar radiationare related to global and ‘local’ modulators is not only relevant topredict local effects but also to identify the key factors to considerwhen undertaking regional or interregional comparisons.
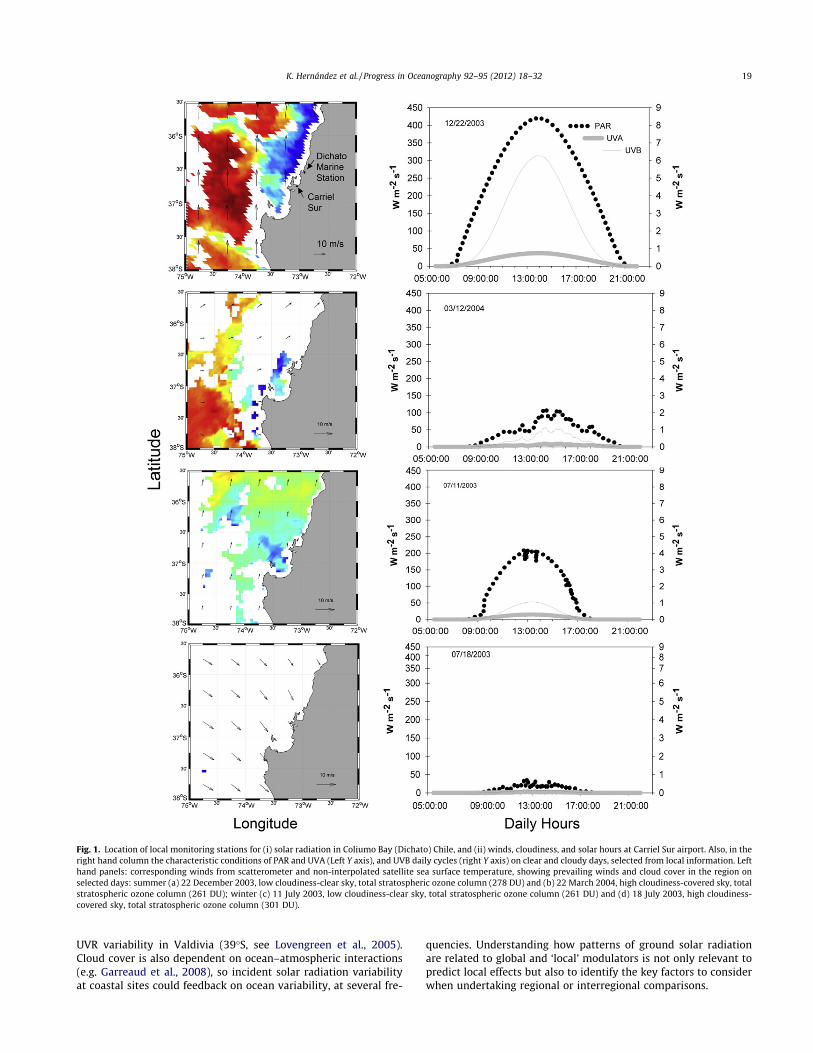
Fig. 1. Location of local monitoring stations for (i) solar radiation in Coliumo Bay (Dichato) Chile, and (ii) winds, cloudiness, and solar hours at Carriel Sur airport. Also, in theright hand column the characteristic conditions of PAR and UVA (Left Y axis), and UVB daily cycles (right Y axis) on clear and cloudy days, selected from local information. Lefthand panels: corresponding winds from scatterometer and non-interpolated satellite sea surface temperature, showing prevailing winds and cloud cover in the region onselected days: summer (a) 22 December 2003, low cloudiness-clear sky, total stratospheric ozone column (278 DU) and (b) 22 March 2004, high cloudiness-covered sky, totalstratospheric ozone column (261 DU); winter (c) 11 July 2003, low cloudiness-clear sky, total stratospheric ozone column (261 DU) and (d) 18 July 2003, high cloudiness-covered sky, total stratospheric ozone column (301 DU).
K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32 19

The patterns of productivity in the coastal ocean are strongly re-lated to solar radiation, which has been differentiated into photo-synthetically active radiation (PAR, 400–750 nm) and ultravioletradiation (UVR, 275–315 nm). Most of the described biological ef-fects of solar radiation at mid and high latitudes have been relatedto PAR limitations on primary production but also to the inhibitionof photosynthesis by UVR (Franklin and Foster, 1997; Villafañeet al., 2001; Häder et al., 2003; Fritz et al., 2008). Solar radiationalso affects community structures, production, and trophic interac-tions at other microbial levels (e.g., virus, prokaryotes; Wilhelmet al., 2003; Church et al., 2004; Hernández et al., 2006). In fact,the relevant processes for autotrophic/heterotrophic/mixotrophicproduction result from many nested non-linear relations.
At an annual scale, experimental results along the western coastof South America from 32!S to 45!S, have suggested annualuni-modal cycles of primary productivity PP in phase with solarradiation (mainly PAR) in natural populations of phytoplankton(Helbling et al., 1993; Montecino and Pizarro, 1995; Dellarosa,1998). Seasonal scale variability in coastal primary producerbiomass has also been described from satellite and in situmeasurements along the eastern south Pacific, although the rela-tive explanatory roles of solar radiation and upwelling intensityhave not been quantitatively explored at such a broad spatial scale.Regarding UVB wavelengths, surface values varying at bothseasonal and synoptic time scales along the Pacific and AtlanticSouth American coasts south of 20! have been suggested to playa role in the UV induced DNA damage and repair processes (Diazet al., 2006; Vernet et al., 2009). Furthermore, incubation experi-ments around 37!S showed deleterious effects of natural levels ofUVB on production, size spectra and DNA structure of bacterio-plankton with different photobiological histories (Hernándezet al., 2006, 2007). North of 42!S organisms deal with several daysof excess radiation while to the South there is a relief from UVRstress (higher frequency of days with UVR levels below climatologyvalues) (Vernet et al., 2009). At lower latitudes, cloud and aerosolmediated modulation prevail, while ozone is the main modulatorof surface UVB at higher latitudes. In fact, UVB oscillations andthe ratios between 305/340 or 305/PAR wavelengths have beenused as indicators of total column ozone (Cabrera et al., 1995;Villafañe et al., 2001; Cede et al., 2002).
The west coast of South America around 37!S remains under theinfluence of the southeast Pacific anticyclone year round, althoughits degree of influence varies seasonally: it is stronger in Spring–Summer when south-southwesterly winds predominate alongthe coast (e.g., Strub et al., 1998; Shaffer et al., 1999; Garreaudet al., 2002). It is well accepted, that this condition drives coastalupwelling and the associated high PP typical of eastern boundarycurrents in the Humboldt Current System (HCS). Synoptic scalewind variability has also been described for the area, with south-erly/northerly wind cycles ranging from 3 to 15 days superim-posed on the seasonal cycle. Biological processes, therefore, aresubject to strong forcing variability due to winds and surface irra-diance cycles (e.g., Dellarosa, 1998; Montero et al., 2007). Addition-ally, the studied area (Concepción, 36!30.80S–73!07.70W) lieswithin the potentially affected, mid-latitude, low ozone area (Kirc-hoff et al., 1996; Cabrera and Fuenzalida, 1999).
So far only monthly averaged data have been used to describeatmospheric connections among dominant wind, atmosphericpressure, cloudiness, and rainfall, as well as the influence thesehave on ground radiation year around in Concepción (Saavedra,1980; Saavedra and Foppiano, 1992; Dellarosa, 1998). Theseauthors pointed out that on a monthly scale, when southerly windsdominated, low cloudiness and rainfall coincided with high groundradiation, i.e., the prevailing summer conditions, and contrastedwith those of winter. Days with high solar radiation selected be-tween 2003 and 2004 had low cloud cover (in both winter and
summer) and coincided with southerly winds. The opposite wastrue for days with low surface solar radiation, evidenced for allwavelengths. The annual cycle of ground radiation at Concepciónhas also been reported using average weekly radiation values(Dellarosa, 1998). Sobarzo et al. (2007) described a short periodof time in autumn (!21 days) when the patterns described formonthly averaged data series also held for daily data.
Physical dynamics within the euphotic layer might be especiallyrelevant in upwelling areas such as Concepción, where weatherconditions have important short-term fluctuations throughoutthe year (Saavedra and Foppiano, 1992). In the Concepción area,PP is highly correlated to micro-phytoplankton biomass (B) (Mon-tero et al., 2007) and both vary seasonally following the seasonalcycles of upwelling favorable conditions and incoming solar radia-tion (McManus and Peterson, 1988; Cuevas et al., 2004; Monteroet al., 2007). Nevertheless, high within season variability character-izes B, PP, the PP/B ratio, as well as bacterial secondary productiv-ity and the export of phytoplankton carbon from the euphotic layer(Montero et al., 2007). In addition, González et al. (2009) showedthat an important part of the observed B variability could be ex-plained by local winds, whose interaction with patterns of incom-ing solar radiation have not been thoroughly explored. Therefore,the potential solar radiation effects (direct and indirect) on bio-physical processes cannot be overlooked over the scales described.A comprehensive characterization of the atmospheric irradiativeforcing, its relationship and feedbacks with other relevant atmo-spheric forcing factors and their potential effect on this costalupwelling area becomes necessary in order to interpret, compareand predict relevant ecosystem properties and functions. In thismanuscript, we quantify the annual and synoptic variability of so-lar radiation (PAR, UVR) for southern-central Chile at 36!S and re-late it to the total ozone concentration, cloudiness, and winds.Furthermore, after establishing those relationships, we evaluatedwhether it was possible to identify an influence of the irradiativepatterns at different time scales on field phytoplankton growthrates. Data considered include almost 2 years of daily visible PAR(400–700 nm), UVA (400–320 nm), and UVB (280–320 nm)measurements.
2. Materials and methods
2.1. Data collection
2.1.1. Solar radiation at ConcepciónGround level irradiances were registered as PAR and UVR wave-
lengths from February 2003 to December 2004 with a ground radi-ometer (GUV-511C Biospherical Instruments) located in ColiumoBay, at the Dichato Marine Biology Station of the University of Con-cepción (36.5!S 72.9!W, Fig. 1). The GUV511 is a temperature-sta-bilized multichannel radiometer that measures downwellingirradiances with moderately narrow bandwidth channels (near10 nm) within the UVR (380, 340, 320, 305 nm). Plus a broad bandchannel with full width at maximum of 300 nm for PAR (400–700 nm). The instrument took one measurement every minute,24 h per day, sent directly to a PC. Raw data were processed byusing software provided by Biospherical Instruments. Daily datawas checked for quality where night values (instrument internalnoise) were subtracted, and calibration constants were applied.No problems were detected with a temperature monitor duringthe study period. The total UVA and UVB irradiances were calcu-lated by following the Orce and Helbling (1997) and Lubin andFrederick (1989) models. The irradiance values were expressed asW m"2 and they were integrated over one day to obtain the dailydose (kJ m"2) of PAR, UVA, and UVB for each individual channel.Calibration was made just once using solar calibration with a
20 K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32

traveling reference GUV-511 (RGUV) following Booth et al. (1994).The reference radiometer was calibrated, under solar light, beforeand after traveling, against the spectroradiometer SUV-100 of theUS National Science Foundation UV Monitoring Network, installedin San Diego, California, USA, followed by a correction of the con-stants with the same model of instrument. The daily 305/340 ratiowas also calculated as a total column ozone proxy (see Frederickand Lubin, 1988; Cabrera et al., 1995; Orce and Helbling, 1997). Fi-nally, the dose and maximum daily irradiance data from almost2 years of measurements were used for the present analysis. Thefew gaps in the time series for all wavelengths in the UV rangeand for PAR channel were filled by linear interpolation to be ableto apply time series analysis on the domain of the frequency andcross-correlations. There were only two gaps of 13 and 10 dayswhich coincided with one of our scales of interest. Filling each ofthese gaps using the parsimonious linear interpolation technique,would only remove the smaller scale variability in less than 0.5%of the time series, and would not comprise more than a couple ofsmall scale cycles out of the total. All the other linear analyses con-ducted (see below) ignored periods where data was lacking.
2.1.2. OzoneData from the EarthProbe satellite (February 2003 to December
2004) is provided for 1 degree latitude and 1.25 degree longitudebins on a grid centered very near Concepción. For details see theNASA satellite ADEUS TOMS data of the Total Ozone Mapping Spec-trometer (TOMS, http://jwocky.gsfc.nasa.gov). Data was expressedin Dobson units (DU).
2.1.3. CloudinessTime series data on cloud cover are usually obtained by two
methods: (i) direct visual observation and (ii) approximationsusing the number of sunshine hours relative to the total amountof expected sunshine hours (García et al., 2000). In our analysis,we used both indices. (i) Direct daily observations were conductedat Carriel Sur airport (36.80S, 73.1!W) by meteorological observersfollowing standard procedures of the International Cloud Atlas(WMO, 1965). (ii) For the cloudiness index (CI), daily hours of sun-shine were obtained from a heliograph (Campbell-Stokes; Meteo-rological Service of Chile, Carriel Sur airport). The expected dailyhours of sun exposure were calculated from sunrise and sunsethours measured by the GUV radiometer and verified with a solarhour calendar (http://www.tutiempo.net/index.php). The daily CI(García et al., 2000) was calculated as: CI = 1 " (number of ‘real’sun hours/number of expected sun hours) (Anabalón et al., 2007).
2.1.4. Coastal windsThe wind magnitude and direction were recorded every 6 h at
Carriel Sur. However, due to the frequency of missing nighttimedata, we used afternoon winds (14:00 and 20:00 local time) in or-der to compute pseudostress components. The higher wind speedsduring the afternoon led to an overestimation of the magnitude ofthe pseudostress.
2.1.4.1. Meridional wind pseudostress. North–South or meridionalwind pseudostress (Zv) was calculated as WV |W| in m2 s"2 (Arcos,1987), where WV is the North–South wind component and W iswind speed (in m s"1). Positive values indicate southerly upwellingfavorable winds, and negative values reflect northerly downwel-ling favorable winds (Arcos, 1987; Pond and Pickard, 1997).
2.1.4.2. Cross-shore wind pseudostress. The East–West or zonal windcomponent is especially strengthened in the afternoon by the dailysea breeze cycle. Therefore, we calculated the zonal wind pseudo-stress (Zu), using the same wind dataset as for the meridional wind
estimation, as Wu |W| in m2 s"2, where Wu is the east–west windcomponent (Pond and Pickard, 1997).
2.2. Data analysis
First, we described the general characteristics of the averageand extreme values of the different variables considered for thestudied seasons. Then the annual cycle of each time series (solarradiation, ozone, CI, pseudostress wind components) was analyzedby fitting harmonics to a 365-day period using the least squaresmethod (Emery and Thomson, 1997). We evaluated the statisticalsignificance of the amplitude of each harmonic by running a MonteCarlo test, randomly reordering the values in the time series, andestimating new amplitudes for the annual harmonic. Since ourinterest was in synoptic scale variability, the residuals of each ser-ies were obtained by subtracting the values predicted with the har-monic function from the observed data. The dominant frequenciesof smaller cycles were identified in the residuals via spectral anal-ysis (by the fast Fourier transform) following Emery and Thomson(1997). Before the spectra were computed, the mean and trendwere removed and a Hamming taper was applied. The relationshipbetween variables was analyzed through linear cross-correlationsin the time domain and coherence/phase analyses in the frequencydomain (Emery and Thomson, 1997). Those analyses were per-formed on the complete series. The relationships were establishedfor: (i) the residuals of the atmospheric variables (ozone, winds, CI)and (ii) the residuals of radiation series and atmospheric variables.Previous efforts described qualitatively the interaction between so-lar radiation and atmospheric variables for Concepción on an an-nual scale using mean monthly values (Saavedra, 1980, 1985;Saavedra and Foppiano, 1992) or across time series covering veryshort periods of the year (Sobarzo et al., 2007).
Winter and summer synoptic conditions differ due to differ-ences in the interaction between the main atmospheric phenom-ena present in the region (southeast Pacific anticyclone,migratory mid-latitude weather systems, and coastal lows).Accordingly, the analysis of entire year data series should be com-plemented with an analysis of split summer/winter data sets withcomposite analysis.
In order to analyze whether the strictly local measurementswould be representative of spatial synoptic scale conditions,large-scale surface circulation and related cloud cover, these wereanalyzed using daily sea level pressure (SLP) at 12 UTC and dailymean total cloud cover from NCEP–NCAR reanalysis (Kalnayet al., 2006) between February 2003 to December 2004. In general,NCEP–NCAR total cloud cover underestimated the total cloudinessat mid- and high latitudes by 10–25% (Bedacht et al., 2006).
We used composite analysis to study large-scale circulation andthe associated cloud cover, along with local observations of differ-ent meteorological parameters when daily moderate to strongmeridional pseudostress was observed at Carriel Sur airport(!37!S). We defined pseudostress as equatorward and polewardwhen the Zv was above and below 10 m2 s"2, respectively. Theanalysis was conducted separately for summer (February, Decem-ber 2003; January–February 2004) and winter (June–August 2003,2004). The statistical significance of the composites was assessedby Monte Carlo experiments at 95% confidence (von Storch andZwiers, 1999).
2.3. Biological implications
In spite of the difficulty of routine field data to estimate primaryproduction related rates (Cullen, 1990), we undertook two differ-ent approaches in order to quantify and evaluate if seasonal andsynoptic PAR variability within the ranges observed contributedto the modulation of in situ phytoplankton growth rates
K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32 21
(Production to biomass ratio). Two models with different degreesof complexity were adjusted to data available from the Station‘‘18’’ time series maintained by the Centre of Oceanographic Re-search of the South Pacific (COPAS) over the Concepción continen-tal shelf (36!300S and 73!070W). We used depth integrated anddepth specific values of PP, total chlorophyll-a, nitrate and ammo-nium measured at standard depths from September 2003–Novem-ber 2005, at quasi monthly periodicity. Primary productivity wasestimated after incubations of in situ light–dark bottles and oxygenevolution measurements (see Montero et al., 2007 for details onprocedures).
Since upwelling (and surface water enrichment) has been con-sidered the main driver of primary productivity in the HCS, a firstand simplest approximation was used to evaluate the role of solarradiation as a co-limiting factor for phytoplankton growth rate, to-gether with available nutrients. We aimed to identify whether thesurface irradiance made a significant contribution to integratedphytoplankton growth rates without smoothing any variability fre-quency (from daily to annual). So, after simple data inspection, avery simple set of linear models was fitted to the integrated PP/Bratio (down to 35 m depth), to surface maximum irradiance andintegrated nitrogen as explanatory variables. Ammonium and ni-trate were added into a single nitrogen (N) pool. We followed amulti-model comparison procedure that allowed us to choose thelinear model that ‘better’ explained observed integrated PP/B ratio:integrated N (single variable); surface irradiance (single variable);integrated N (variable 1) + surface irradiance (two variables). The
choice of the best model was based on the Akaike InformationCriteria (AIC, see Burnham and Anderson, 2002), which evaluatesthe fit of model predictions and data, but penalizes the numberof parameters used to construct the empirical model. The estima-tions were conducted with Statistica 6.0. Generalized linear modelwere identified, and a normal distribution was chosen to build themodels.
A second empirical approach (model 2) was undertaken in orderto evaluate whether the suit of observed PP/B ratios at depth wasbetter explain by: (i) the actual surface irradiance series, (ii) an an-nual mean of surface irradiance, (iii) the corresponding seasonalmean. In this case, we adjusted the parameters of a very simplesemi-mechanistic model to data, and compared the resulting rootsum of squares. The adjusted model included light and nutrientlimitation as well as growth rate temperature dependency. Themodel used was the following:
PP=B ¼ VmT $ a$ IzffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiVmT2 þ a2 $ I2
z
q $ ½N'kN þ ½N'
$ C : Chl-a
where PP is primary production (mg C m3 d"1); B is phytoplanktonstanding stock (mg Chl-a m"3). The terms on the right hand side ofthe equation represent solar and nutrient growth limitation. Alfa isthe initial slope of the production/irradiance curve; Iz (W m"2) isPAR irradiance at depth z (m); VmT is the temperature dependentmaximum growth rate at light saturation, [N] is nitrogen concentra-tion (nitrate plus ammonium, M m"3) and kN is the half saturation
PAR
max
(KJm
-2)
UVB
max
(KJm
-2)
UVA
max
(KJm
-2)
0 90 180 270 360 450 540 630 720
0 90 180 270 360 450 540 630 720
0 90 180 270 360 450 540 630 720
0 90 180 270 360 450 540 630 720
02000400060008000
100001200014000
0200400600800
10001200
JunMar Sept Dec Mar Jun Sept Dec
Day / Month of the yearJan
Clo
udin
ess
inde
x
-0,5
0,0
0,5
1,0
1,5
020406080
100120140160
a
b
c
d
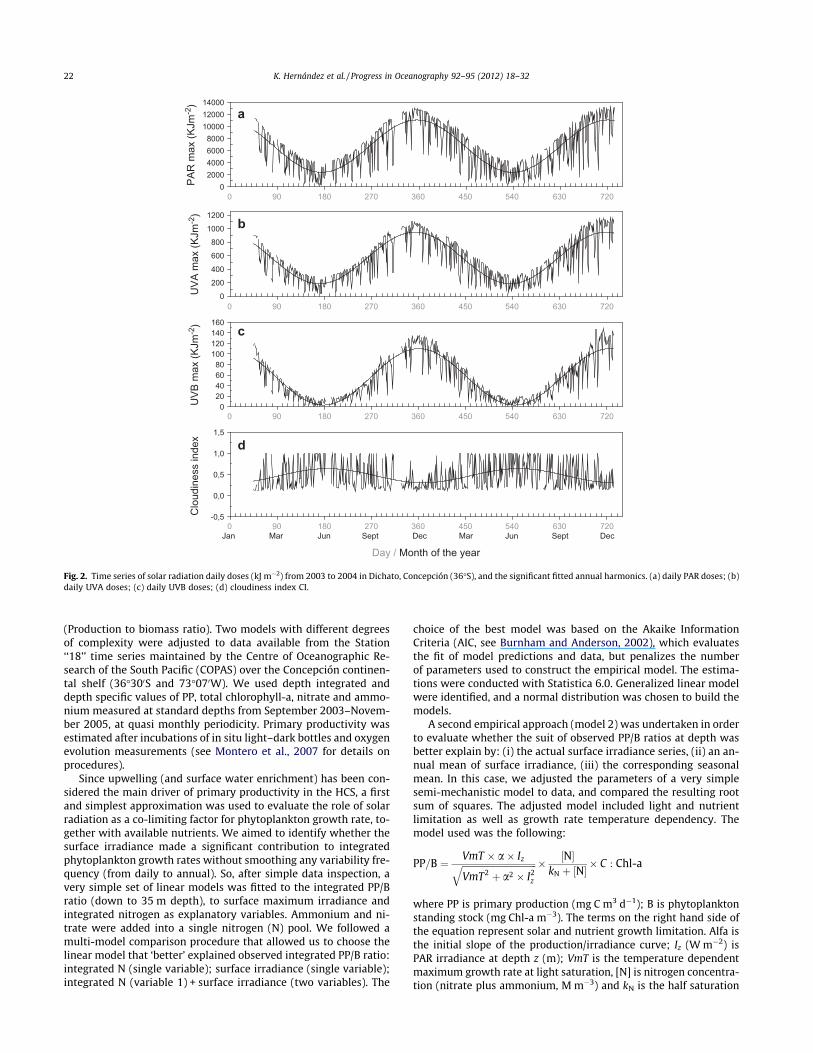
Fig. 2. Time series of solar radiation daily doses (kJ m"2) from 2003 to 2004 in Dichato, Concepción (36!S), and the significant fitted annual harmonics. (a) daily PAR doses; (b)daily UVA doses; (c) daily UVB doses; (d) cloudiness index CI.
22 K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32

constant for nutrient uptake. C:Chl-a was fix in the range of thelower values 20 (Taylor et al., 1997). Although it is known to dependon light and nutrient limitation (Cloern et al., 1995), it was fixed forparsimony regarding our hypothesis and the lack of complimentarydata.
VmT ¼ a ( bT
where T is in situ temperature, a is the maximum growth rate at0 !C (d"1) set to 0.8, while b was set to 1.06 following the literature(Oschilies and Garçon, 1999; Koné et al., 2005). Estimated VmT ran-ged: 1.6–2.6 at observed temperatures, as expected.
Light at depth (Iz) was obtained from the exponential decay ofsurface irradiance. It depended on the water decay constant (kw)of 0.2 and a chlorophyll concentration term (kChl-a ( [Chl-a]) wherekChl-a was set to 0.4. Values were taken from literature (Fashamet al., 1990; Olivieri and Chavez, 2000) since there is no access tolocal estimations. We are aware of k potential variability (Siegelet al., 1995), it was a parsimonious assumption when they have al-ready been successfully used in other cases (Koné et al., 2005) andthere is no other data available. Photo inhibition was not consid-ered. Two parameters were estimated in the model: a and kN.The root sum of squares of estimated and observed PP/B valueswas minimized to obtain parameter values with Excel solver. Theroot sum of squares (RSS) was calculated for each adjusted model.
3. Results
3.1. Annual cycles
As expected, the time series is too short to make general state-ments. Nevertheless some trends are clear for the ca. 2 years ofmeasurements: The annual cycles for PAR, UVA, and UVB(Fig. 2a–c), which covaried in phase. Throughout the study period,the highest irradiance, dose, and variance for the visible and UVranges were in late spring and beginning of summer (2003 and2004 respectively), and lowest in autumn and winter (Table 1).The highest differences among mean seasonal daily doses occurredbetween summer and autumn (2004) for PAR (!5480 kJ m"2), be-tween autumn (2003) and spring (2004) for UVA (543 kJ m"2), and
between summer (2004) and winter (2003, 2004) for UVB(!71 kJ m"2; Fig. 2b, Table 1). The solar radiation data for Concep-ción were highly variable within season. In summer, the PAR/UVA/UVB doses ranged between 4000/500/60 and 13000/1100/130 kJ m"2, whereas in winter, they varied from !61/6/0.4 to5000/400/20 kJ m"2. Most of this day-to-day variability could beexplained by cloudiness (see below). As expected, minimum sum-mer irradiance values on different wavelengths were as low as orlower than wintertime values. The annual harmonic fitted for eachwavelength was significant in all cases and explained a large frac-tion of the total variance (Table 2). The annual cycles of pseudo-stress wind components and cloudiness showed maximum ZundZv (Fig. 3a–b) and minimum CI in summer (Table 1; Fig. 2d). Theannual harmonic was significant for the three variables at 5% con-fidence (Monte Carlo test), but the percentage of the explained var-iance was lower than for solar radiation variables (Table 2).Regarding pseudostress wind components, Zu presented the stron-gest seasonality, with annual amplitudes larger than Zv, but ex-plained three times the variance explained by the annualharmonic of Zv. The lower explained variance of the annual har-monic of Zv and CI denoted the importance of synoptic variabilityin these two variables. Most annual cycles mentioned so far werein phase, positively correlated at time lag 0. The annual CI cyclewas negatively correlated with pseudostress wind componentsand radiative variables (Figs. 2d and 3a–b).
Table 1Solar radiation and atmospheric conditions at Concepción, Chile, from 2003 to 2004.
2003 2004⁄ Summer Autumn Winter Spring Summer Autumn Winter Spring
Ozone Max/Min 302/214 313/214 372/256 353/256 297/239 318/214 368/229 329/239DU X ± Sd 262 ± 13 259 ± 17 301 ± 25 306 ± 20 270 ± 11 260 ± 20 286 ± 30 278 ± 21PAR Max/Min – 7828/296 7560/296 13,104/1606 112,291/2118 8133/375 8554/558 13,177/1541kJ m"2 X ± Sd 3915 ± 2164 4237 ± 1840 9361 ± 3061 9372 ± 2681 3892 ± 2130 3993 ± 2184 9339 ± 3034UVA Max/Min – 599/33 606/33 1105/166 1080/187 675/44 736/66 1162/181kJ m"2 X ± Sd 298 ± 157 342 ± 140 796 ± 242 762 ± 208 324 ± 163 344 ± 177 841 ± 247UVB Max/Min – 68/1 47/2 136/11 136/17 73/2 64/3 147/14kJ m"2 X ± Sd 23 ± 18 22 ± 11 83 ± 31 93 ± 29 25 ± 18 22 ± 16 87 ± 33305/340 Max/Min – 0.11/0.01 0.06/0.02 0.12/0.04 0.15/0.08 0.10/0.02 0.09/0.02 0.14/0.06
X ± Sd 0.05 ± 0.03 0.04 ± 0.01 0.09 ± 0.02 0.12 ± 0.01 0.05 ± 0.02 0.04 ± 0.02 0.10 ± 0.02PAR Max/Min – 372/72 402/24 521/150 515/112 452/43 425/60 603/124W m"2 X ± Sd 227 ± 84 251 ± 80 390 ± 83 382 ± 68 240 ± 99 255 ± 87 422 ± 83UVA Max/Min – 27/3 29/3 44/15 41/9 34/5 36/6 49/14W m"2 X ± Sd 16 ± 6 19 ± 6 33 ± 7 31 ± 5 18 ± 7 20 ± 7 37 ± 6UVB Max/Min – 3.6/0.2 2.9/0.2 7.1/1.3 6.9/1.1 4/0.25 3.6/0.35 7.2/1.5W m"2 X ± Sd 1.5 ± 0.9 1.5 ± 0.6 4.4 ± 1.3 4.9 ± 1.1 1.0 ± 0.1 1.5 ± 0.87 4.9 ± 1.2CC Max/Min 8/0 8/0 8/0 8/0 8/0 8/0 8/0 8/0Oktas X ± Sd 3.2 ± 2.9 5 ± 3 4.69 ± 2.66 4 ± 2 2.61 ± 2.47 4.6 ± 2.7 5 ± 2.54 4 ± 2.6CI Max/Min 1/0.11 1/0.13 1/0.11 1/0.09 1.0/0.09 1.0/0.01 1.0/0.01 0.99/0.12
X ± Sd 0.38 ± 0.3 0.53 ± 0.33 0.55 ± 0.31 0.39 ± 0.30 0.31 ± 0.20 0.56 ± 0.28 0.51 ± 0.30 0.40 ± 0.27Zv Max/Min 59/"168 33/"168 39.5/"132 71/"107 77/"59 31/"115 29/"151 43/"44m2 s"2 X ± Sd 10.0/23 "7.75 ± "30.75 "2.66 ± 26 10 ± 27 9 ± 22.3 "6 ± 23 "11 ± 33 10 ± 19
⁄ Solar radiation values measured from 15 February to 21 March 2003 were not considered as part of the average 2003 summer values.
Table 2Parameters of the annual harmonics fitted for irradiative and meteorologicalvariables. Amplitude is expressed in kJ m"2 for PAR, UVA and UVB; in DU for O3;and in m2 s"2 for Zv and Zu. Phase is expressed in days relative to calendar day. RMS:root mean square. p: significance of the fitted harmonic as evaluated through MonteCarlo analysis over a 1000 randomization cycles. ⁄Significance at the 0.05 level.
Variable PAR UVA UVB O3 Zv Zu CI
Amplitude 4314 379 52.9 22.7 13.5 15.3 0.17Phase "9.8 "13.5 "5 "92.5 "5.96 "9.08 1.93%Variance 69.1 76.1 83.3 36.9 11.7 33.3 13.4RMS 2005 149 16.4 21.3 25.7 15.0 0.29p ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄
DU: dobson units; nd: non dimensional parameter.
K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32 23
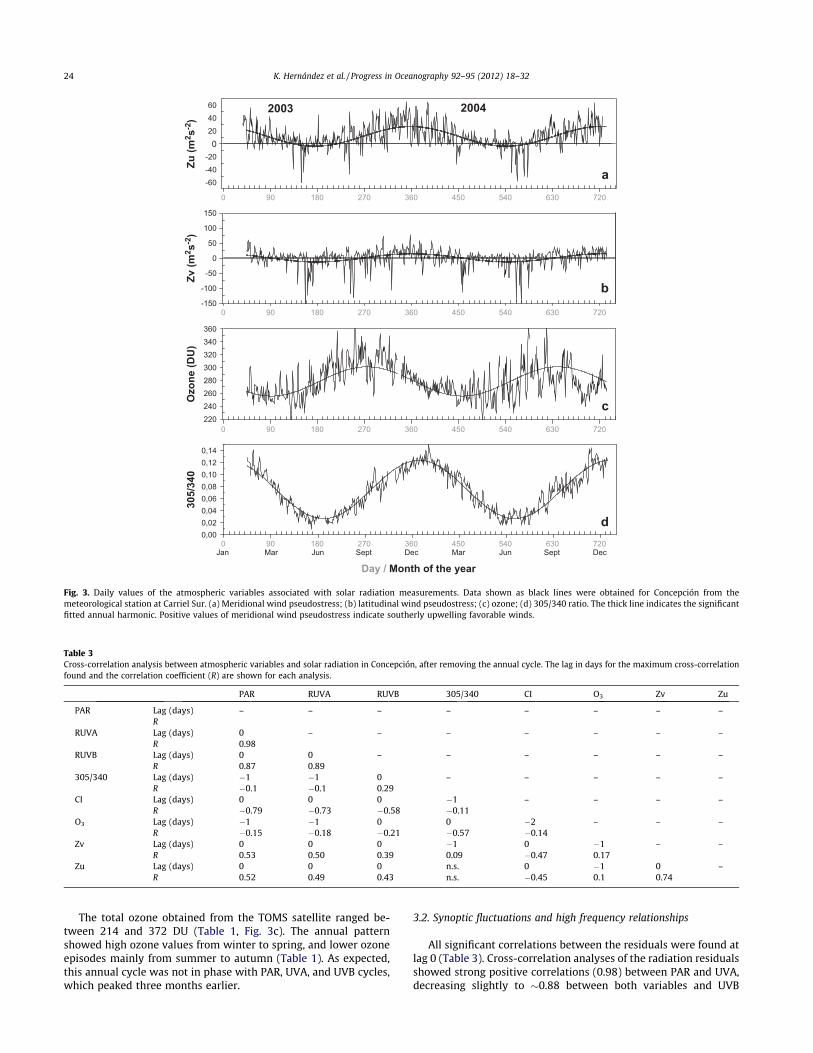
The total ozone obtained from the TOMS satellite ranged be-tween 214 and 372 DU (Table 1, Fig. 3c). The annual patternshowed high ozone values from winter to spring, and lower ozoneepisodes mainly from summer to autumn (Table 1). As expected,this annual cycle was not in phase with PAR, UVA, and UVB cycles,which peaked three months earlier.
3.2. Synoptic fluctuations and high frequency relationships
All significant correlations between the residuals were found atlag 0 (Table 3). Cross-correlation analyses of the radiation residualsshowed strong positive correlations (0.98) between PAR and UVA,decreasing slightly to !0.88 between both variables and UVB
Zv (m
2 s-2
)30
5/34
0
-150
-100
-50
0
50
100
150
Ozo
ne (D
U)
220240260280300320340360
2003 2004
0,000,020,040,060,080,100,120,14
Zu (m
2 s-2
)0 90 180 270 360 450 540 630 720
0 90 180 270 360 450 540 630 720
0 90 180 270 360 450 540 630 720
0 90 180 270 360 450 540 630 720
-60-40-20
0204060
JunMar Sept Dec Mar Jun Sept Dec
Day / Month of the year Jan
a
b
c
d
Fig. 3. Daily values of the atmospheric variables associated with solar radiation measurements. Data shown as black lines were obtained for Concepción from themeteorological station at Carriel Sur. (a) Meridional wind pseudostress; (b) latitudinal wind pseudostress; (c) ozone; (d) 305/340 ratio. The thick line indicates the significantfitted annual harmonic. Positive values of meridional wind pseudostress indicate southerly upwelling favorable winds.
Table 3Cross-correlation analysis between atmospheric variables and solar radiation in Concepción, after removing the annual cycle. The lag in days for the maximum cross-correlationfound and the correlation coefficient (R) are shown for each analysis.
PAR RUVA RUVB 305/340 CI O3 Zv Zu
PAR Lag (days) – – – – – – – –R
RUVA Lag (days) 0 – – – – – – –R 0.98
RUVB Lag (days) 0 0 – – – – – –R 0.87 0.89
305/340 Lag (days) "1 "1 0 – – – – –R "0.1 "0.1 0.29
CI Lag (days) 0 0 0 "1 – – – –R "0.79 "0.73 "0.58 "0.11
O3 Lag (days) "1 "1 0 0 "2 – – –R "0.15 "0.18 "0.21 "0.57 "0.14
Zv Lag (days) 0 0 0 "1 0 "1 – –R 0.53 0.50 0.39 0.09 "0.47 0.17
Zu Lag (days) 0 0 0 n.s. 0 "1 0 –R 0.52 0.49 0.43 n.s. "0.45 0.1 0.74
24 K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32
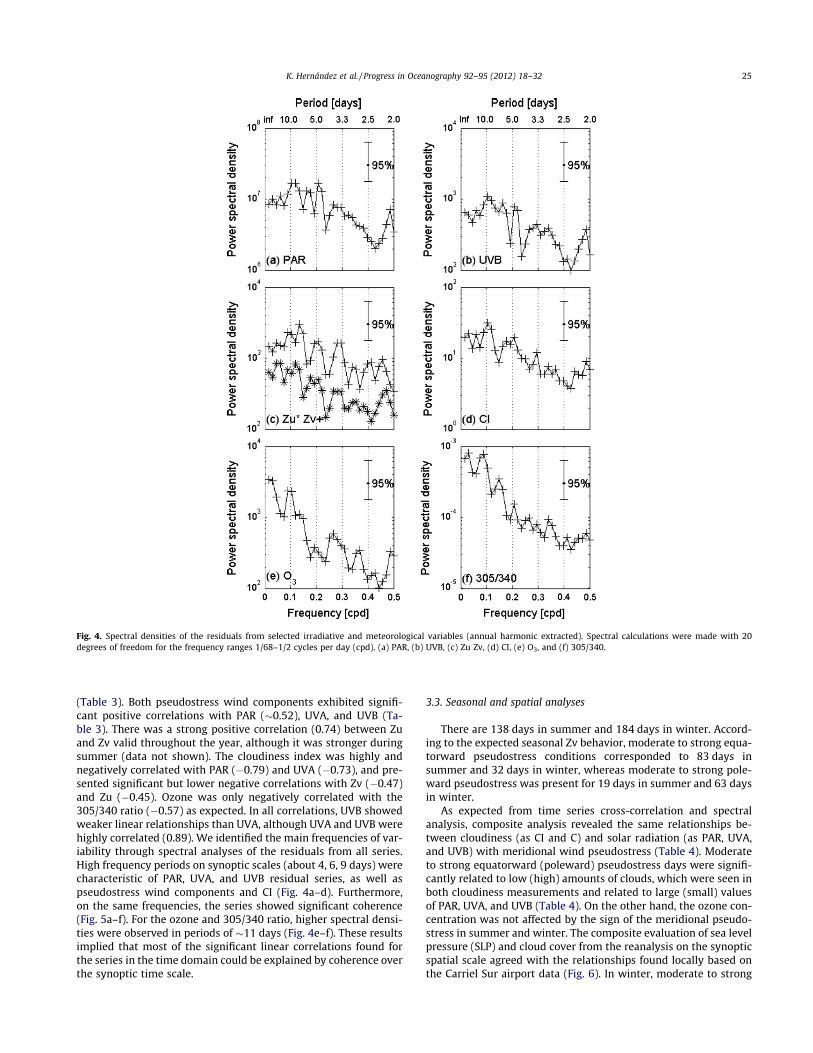
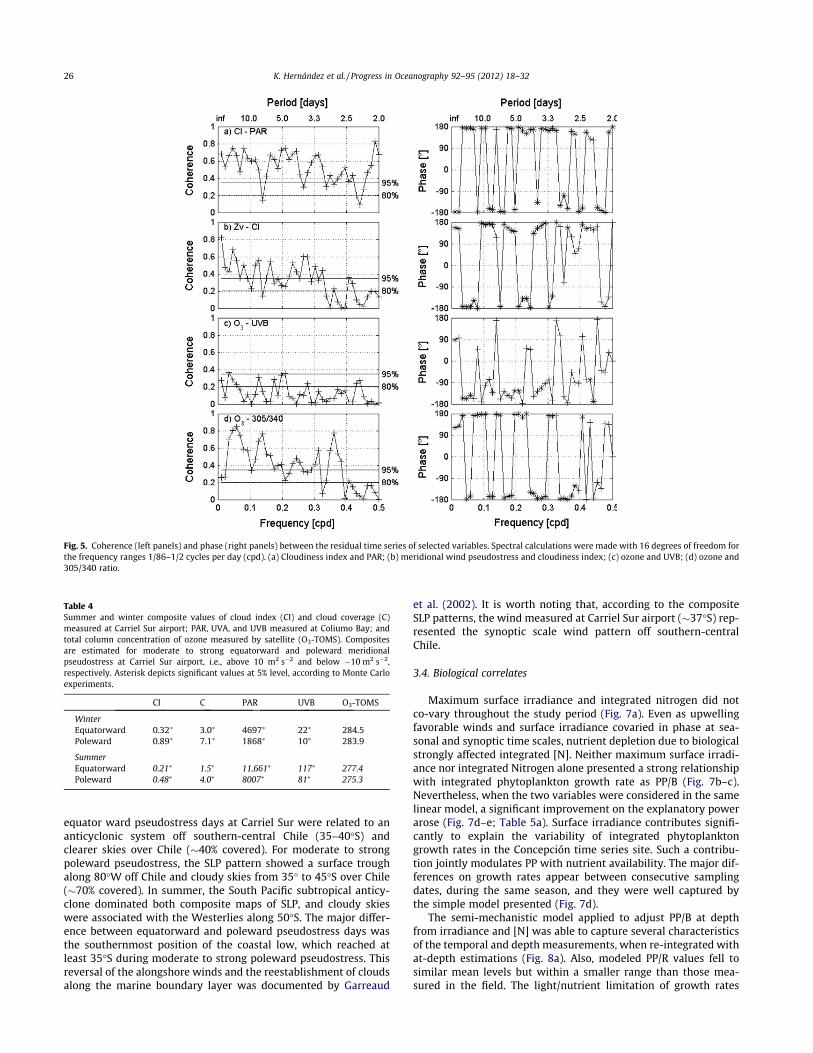
(Table 3). Both pseudostress wind components exhibited signifi-cant positive correlations with PAR (!0.52), UVA, and UVB (Ta-ble 3). There was a strong positive correlation (0.74) between Zuand Zv valid throughout the year, although it was stronger duringsummer (data not shown). The cloudiness index was highly andnegatively correlated with PAR ("0.79) and UVA ("0.73), and pre-sented significant but lower negative correlations with Zv ("0.47)and Zu ("0.45). Ozone was only negatively correlated with the305/340 ratio ("0.57) as expected. In all correlations, UVB showedweaker linear relationships than UVA, although UVA and UVB werehighly correlated (0.89). We identified the main frequencies of var-iability through spectral analyses of the residuals from all series.High frequency periods on synoptic scales (about 4, 6, 9 days) werecharacteristic of PAR, UVA, and UVB residual series, as well aspseudostress wind components and CI (Fig. 4a–d). Furthermore,on the same frequencies, the series showed significant coherence(Fig. 5a–f). For the ozone and 305/340 ratio, higher spectral densi-ties were observed in periods of !11 days (Fig. 4e–f). These resultsimplied that most of the significant linear correlations found forthe series in the time domain could be explained by coherence overthe synoptic time scale.
3.3. Seasonal and spatial analyses
There are 138 days in summer and 184 days in winter. Accord-ing to the expected seasonal Zv behavior, moderate to strong equa-torward pseudostress conditions corresponded to 83 days insummer and 32 days in winter, whereas moderate to strong pole-ward pseudostress was present for 19 days in summer and 63 daysin winter.
As expected from time series cross-correlation and spectralanalysis, composite analysis revealed the same relationships be-tween cloudiness (as CI and C) and solar radiation (as PAR, UVA,and UVB) with meridional wind pseudostress (Table 4). Moderateto strong equatorward (poleward) pseudostress days were signifi-cantly related to low (high) amounts of clouds, which were seen inboth cloudiness measurements and related to large (small) valuesof PAR, UVA, and UVB (Table 4). On the other hand, the ozone con-centration was not affected by the sign of the meridional pseudo-stress in summer and winter. The composite evaluation of sea levelpressure (SLP) and cloud cover from the reanalysis on the synopticspatial scale agreed with the relationships found locally based onthe Carriel Sur airport data (Fig. 6). In winter, moderate to strong
Fig. 4. Spectral densities of the residuals from selected irradiative and meteorological variables (annual harmonic extracted). Spectral calculations were made with 20degrees of freedom for the frequency ranges 1/68–1/2 cycles per day (cpd). (a) PAR, (b) UVB, (c) Zu Zv, (d) CI, (e) O3, and (f) 305/340.
K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32 25
equator ward pseudostress days at Carriel Sur were related to ananticyclonic system off southern-central Chile (35–40!S) andclearer skies over Chile (!40% covered). For moderate to strongpoleward pseudostress, the SLP pattern showed a surface troughalong 80!W off Chile and cloudy skies from 35! to 45!S over Chile(!70% covered). In summer, the South Pacific subtropical anticy-clone dominated both composite maps of SLP, and cloudy skieswere associated with the Westerlies along 50!S. The major differ-ence between equatorward and poleward pseudostress days wasthe southernmost position of the coastal low, which reached atleast 35!S during moderate to strong poleward pseudostress. Thisreversal of the alongshore winds and the reestablishment of cloudsalong the marine boundary layer was documented by Garreaud
et al. (2002). It is worth noting that, according to the compositeSLP patterns, the wind measured at Carriel Sur airport (!37!S) rep-resented the synoptic scale wind pattern off southern-centralChile.
3.4. Biological correlates
Maximum surface irradiance and integrated nitrogen did notco-vary throughout the study period (Fig. 7a). Even as upwellingfavorable winds and surface irradiance covaried in phase at sea-sonal and synoptic time scales, nutrient depletion due to biologicalstrongly affected integrated [N]. Neither maximum surface irradi-ance nor integrated Nitrogen alone presented a strong relationshipwith integrated phytoplankton growth rate as PP/B (Fig. 7b–c).Nevertheless, when the two variables were considered in the samelinear model, a significant improvement on the explanatory powerarose (Fig. 7d–e; Table 5a). Surface irradiance contributes signifi-cantly to explain the variability of integrated phytoplanktongrowth rates in the Concepción time series site. Such a contribu-tion jointly modulates PP with nutrient availability. The major dif-ferences on growth rates appear between consecutive samplingdates, during the same season, and they were well captured bythe simple model presented (Fig. 7d).
The semi-mechanistic model applied to adjust PP/B at depthfrom irradiance and [N] was able to capture several characteristicsof the temporal and depth measurements, when re-integrated withat-depth estimations (Fig. 8a). Also, modeled PP/R values fell tosimilar mean levels but within a smaller range than those mea-sured in the field. The light/nutrient limitation of growth rates
Fig. 5. Coherence (left panels) and phase (right panels) between the residual time series of selected variables. Spectral calculations were made with 16 degrees of freedom forthe frequency ranges 1/86–1/2 cycles per day (cpd). (a) Cloudiness index and PAR; (b) meridional wind pseudostress and cloudiness index; (c) ozone and UVB; (d) ozone and305/340 ratio.
Table 4Summer and winter composite values of cloud index (CI) and cloud coverage (C)measured at Carriel Sur airport; PAR, UVA, and UVB measured at Coliumo Bay; andtotal column concentration of ozone measured by satellite (O3-TOMS). Compositesare estimated for moderate to strong equatorward and poleward meridionalpseudostress at Carriel Sur airport, i.e., above 10 m2 s"2 and below "10 m2 s"2,respectively. Asterisk depicts significant values at 5% level, according to Monte Carloexperiments.
CI C PAR UVB O3-TOMS
WinterEquatorward 0.32⁄ 3.0⁄ 4697⁄ 22⁄ 284.5Poleward 0.89⁄ 7.1⁄ 1868⁄ 10⁄ 283.9
SummerEquatorward 0.21⁄ 1.5⁄ 11,661⁄ 117⁄ 277.4Poleward 0.48⁄ 4.0⁄ 8007⁄ 81⁄ 275.3
26 K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32
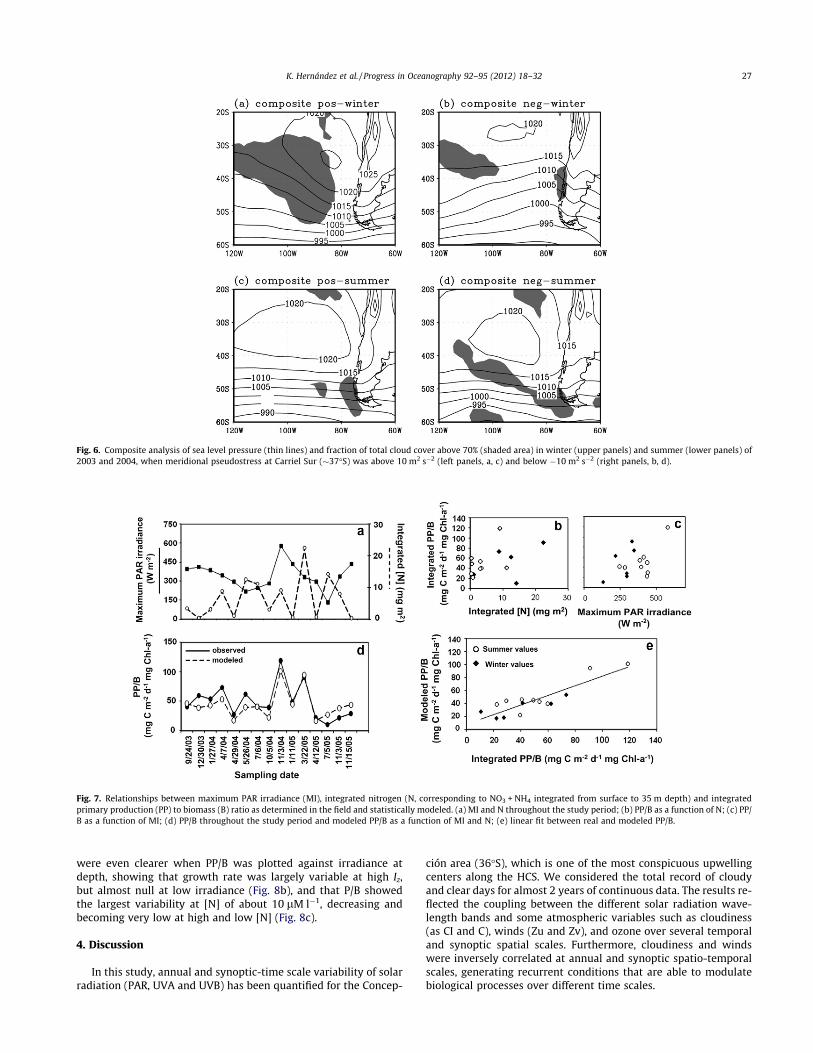
were even clearer when PP/B was plotted against irradiance atdepth, showing that growth rate was largely variable at high Iz,but almost null at low irradiance (Fig. 8b), and that P/B showedthe largest variability at [N] of about 10 lM l"1, decreasing andbecoming very low at high and low [N] (Fig. 8c).
4. Discussion
In this study, annual and synoptic-time scale variability of solarradiation (PAR, UVA and UVB) has been quantified for the Concep-
ción area (36!S), which is one of the most conspicuous upwellingcenters along the HCS. We considered the total record of cloudyand clear days for almost 2 years of continuous data. The results re-flected the coupling between the different solar radiation wave-length bands and some atmospheric variables such as cloudiness(as CI and C), winds (Zu and Zv), and ozone over several temporaland synoptic spatial scales. Furthermore, cloudiness and windswere inversely correlated at annual and synoptic spatio-temporalscales, generating recurrent conditions that are able to modulatebiological processes over different time scales.
Fig. 6. Composite analysis of sea level pressure (thin lines) and fraction of total cloud cover above 70% (shaded area) in winter (upper panels) and summer (lower panels) of2003 and 2004, when meridional pseudostress at Carriel Sur (!37!S) was above 10 m2 s"2 (left panels, a, c) and below "10 m2 s"2 (right panels, b, d).
Fig. 7. Relationships between maximum PAR irradiance (MI), integrated nitrogen (N, corresponding to NO3 + NH4 integrated from surface to 35 m depth) and integratedprimary production (PP) to biomass (B) ratio as determined in the field and statistically modeled. (a) MI and N throughout the study period; (b) PP/B as a function of N; (c) PP/B as a function of MI; (d) PP/B throughout the study period and modeled PP/B as a function of MI and N; (e) linear fit between real and modeled PP/B.
K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32 27

Our results also contributed to depict the latitudinal gradient ofsolar radiation and other meteorological variables along the Pacificcoast of South America (Table 6). Solar radiation at Concepción(!36!S) showed slightly higher maximum values in summer andwinter, as compared to those registered at the Valdivia station(39!S, Lovengreen et al., 2005) following a latitudinal trend. Our re-ported irradiance ranges agreed with previous reports along SouthAmerica (Orce and Helbling, 1997; Cede et al., 2002; Diaz et al.,2006; Vernet et al., 2009) and similar latitudes (e.g. Malaga, Athensand Sierra Nevada, Häder et al., 2007). This latitudinal and seasonalvariability of the different solar radiation wavelengths is usuallyexplained by the solar zenithal angle (SZA), the atmospheric absor-bance related to Rayleigh scattering which enhances it effects withdecreasing wavelengths (UV range), (e.g. see Häder et al., 2007;Vernet et al., 2009). However, among-site variability does notnecessarily follow latitudinal trends. Other sources of among-sitevariability are aerosols, the altitude and the specific climatic–geo-graphic conditions. To illustrate this, Santiago de Chile (33!S; table6) can have maximum PAR levels similar to those of Punta Arenas(53!S) probably due to high levels of pollution. The same type ofsituation has been observed for polluted cities such as Athens(37!N) as compared to places as Erlangen (!49!N) (Häder et al.,
2007). Specific interactions between atmospheric circulation andlocal topography contribute to shape local scale and among-sitevariability as well as departures from latitudinal trends. In spiteof its lower latitude and higher altitude, maximum summer irradi-ance at Jujuy (24, 10!S) can be as low as in Santiago (33.27!S, Diazet al., 2006; Vernet et al., 2009), because of differences in cloudcover and the large oscillations measured in the former.
In our study, the 305/PAR ratio for Concepción showed that ithad the highest value, decreasing towards the north and south(shown in Table 6). This result can be explained by the fact thatthere is a decrease in cloud cover dependence from PAR to UVAto UVB wavelengths, and the 305 wavelength attenuation is moredependent on the distance traveled in the atmosphere due to SolarZenital Angle (Diaz et al., 2006, Häder et al., 2007). The two oppo-site gradients (more PAR attenuation towards the south because ofincreased cloudiness along the western coast of South America,and larger zenith angle effect on the higher latitudes over UVB)would combine to render maximum 305/PAR ratios at mid lati-tudes, such as that of Concepción. PAR–UVA residual correlationwas higher than PAR–UVB and PAR–UVA reflecting the differentialresponse of each wavelength to factors such as cloud and ozone.When our results are compared with those of Vernet et al.(2009), latitudinal patterns become clearer for sites sharing certaingeographical and climate characteristics like distance to the sea,height, relative position in relation to the main wind field and largetopographic features (cordillera), among others. It is also possiblethat the local predominant cloud type could attenuate/enhanceproportionally larger fractions of PAR than at other locations(Lovengreen et al., 2005). These effects could not be evaluated inthe present study, but they should be considered in the future byusing local cloud type data and replicating the present analysisat other sites.
4.1. Winds, cloudiness, and solar radiation
Concepción presented lower variance of PAR, UVA, and UVB inwinter than in summer. The high seasonal and synoptic variabilityof these wavelength bands reflected the interaction of the zenithangle with local meteorological conditions, especially cloud cover,as modulating factors of solar radiation in summer. In Concepción,cloudiness depended heavily on winds (NS), not only on annual butalso on synoptic scales. For instance, southwesterly winds (SW)and clear skies were more frequent in summer and cloudinessresiduals were correlated significantly and positively with winds,also varying within similar periods. In Concepción, the annual wind
Table 5Statistical results for bio-physical models relating in situ phytoplankton growth rateto surface PAR irradiance. (a) Set of models compared to relate integrated (35 m)primary productivity to biomass ratio (mg C m"2 d"1/mg Chl-a"1). Shaded line: bestmodel chosen on the basis of the lowest Akaike Information Criteria (AIC), and itssignificant difference with the model that followed. (b) Fit of model 2 (sea descriptionin the text) to data (PP/B at depth) as forced by measured solar radiation (Real I),Seasonal Irradiance (SI) and Mean Constant Irradiance. Shaded line: best model fit.
df: degrees of freedom; Var: explanatory variables; AIC: Akaike InformationCriteria; LRatio: Likelihood ratio; p: probability level.Alfa: initial slope of the light dependent production/biomass curve, kN: half satu-ration constant for nutrient uptake; N number of data points used for the estima-tion; k: number of estimated parameters; RSS: root sum of squares.
a
b
c
Date
Observed
Modeled
[N]z-1
Iz (W m-2)
Inte
grat
ed P
P/B
(0-1
5m)
(mg
C m
-2d-
1m
g Ch
l-a-1
)
PP/B
z (m
g C
d-1
mg
Chl-a
-1)
Fig. 8. (a) Primary production (PP) to biomass (B) ratio integrated between 0 and 15 m depth, modeled results were integrated after depth specific estimations (model 2); (b)and (c) relationships between production to biomass ratio with irradiance and nitrogen concentration (NO3 + NH4) at depth (z), respectively.
28 K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32

cycle of increasing frequency and strength of SW in Spring–Sum-mer was related to the southward migration of the South Pacificanticyclone (Saavedra and Foppiano, 1992). Superimposed on theannual cycle, perturbations with frequencies similar to those ob-served in the present time series have been associated with thesouthward propagation of coastal lows from 30! to 40!S (Garreaudet al., 2002; Rutllant et al., 2004). The cyclonic lows generatedalong the ocean–atmosphere boundary layer north of Concepción,from where they propagate southward, pass through our studyarea (Garreaud et al., 2002). Besides enhancing/diminishingupwelling favorable winds, the passage of these coastal lows alsogenerated coastal trapped waves with a periodicity of 3–7 days.Synoptic variability was found throughout the year. In winter,storms and alongshore wind reversals were also associated withdisturbances of the polar front that migrated north concomitantlywith the entire South Pacific pressure system (Fuenzalida et al.,2005). Southerly upwelling favorable winds were also associatedwith clear skies and, therefore, stronger radiation. On the contrary,northerly winds were associated with cloudy skies and lower radi-ation. Our results supported previous qualitative descriptions forConcepción (Saavedra, 1980, 1985; Saavedra and Foppiano, 1992)and observations over shorter (one month) periods (Dellarosa,1998).
Since the local patterns and relationships found between theatmospheric variables characterized in this study reflect processesoccurring at regional scales, it is likely that they should hold for sev-eral degrees of latitude along the HCS. A recent study by Garreaudet al. (2011), has described a similar pattern in the variability of,and co-variability between, winds and solar radiation at anotherimportant upwelling site at 30!S in the HCS. At this latitude, the dailyland–ocean warming/cooling cycles have been proposed to modu-late coastal upwelling favorable winds (Rutllant et al., 2004; Bakun,1990). Therefore, an increase in cloud cover could potentially simul-taneously reduce coastal upwelling and surface solar radiation.
The short term pulses of upwelling favorable winds superim-posed on latitudinal dependent seasonality in northern and south-ern Pacific Eastern Boundary Currents have been well described,and both systems share several similar characteristics (Hill et al.,1998). In Monterey Bay in coast of California, upwelling favorablewinds and PAR have also been reported to co-vary at least in shortterm studies. Our results regarding the correlation between atmo-spheric variables, is likely to be a thorough description of a pattern
that could be common to other coasts along the Northern and South-ern Pacific Eastern Boundary Currents (Bedacht et al., 2006).
4.2. UV radiation and ozone
The annual cycle of stratospheric ozone (e.g., 37–38!S) dependson the global circulation of air masses and ozone budgets (Cabreraet al., 1995; Kirchoff et al., 1996; Orce and Helbling, 1997), so asexpected, the annual cycle of ozone and radiation are not in phase(Rowland, 1991). The 305/340 ratio has been used for several yearsas an ozone estimator (Cabrera et al., 1995; Kirchoff et al., 1996;Orce and Helbling, 1997; Liao and Frederick, 2005). However, inour case, the relationship between stratospheric ozone and the305/340 ratio, was significant only after removing the annual cy-cles. The ratio should be used cautiously: if day to day measure-ments are to be used without subtracting the annual harmonic ofthe series, then it is only useful for short periods of time (biweeklyfor example). If it were to be used for longer time series, then theannual harmonic of the 305/340 ratio should be subtracted and theresults should be considered as the residuals of an ozone series.
The relationship between residual winds and cloudiness, andozone was weak and displayed a 1-day lag. Ozone synoptic fluctu-ations (!11 days) were also related to local winds probably be-cause ozone destruction in the lower stratosphere depends onlocal troposphere pressure systems (Stolarsky et al., 1991; Logan,1994; McPeters et al., 1996). This might explain why lower ozonelevels in Concepción were (slightly) associated with SW, whichwere driven by high pressure systems, once again emphasizingthe coupling between high radiation levels (on different wave-lengths) and upwelling favorable conditions in the coastal areaoff Concepción.
4.3. Biological correlates and other potential implications
Since southwesterly winds associated with clear skies also forcecoastal upwelling, the periods with higher surface solar radiationcoincide with the fertilization of the photic layer at both annualand synoptic scales. So far, light limitation of PP has been proposedfor wintertime in this area (Cuevas et al., 2004; Böttjer and Morales2005; Montero et al., 2007); and suggested for northern Chile(Montecino and Pizarro, 1995), but high within-season variabilityhas only been related to upwelling pulses. The relationships
Table 6Daily maximum PAR and UVR values reported for different Chilean cities. The maximum irradiances are expressed in W m"2 during summer and winter.
Location LatitudeSouth
LongitudeWest
Altitude(m)
305 nmWm"2 nm"1
305S/W
320 nmWm"2 nm"1
340 nmWm"2 nm"1
380 nmWm"2 nm"1
PARWm"2 nm"1
305/PARppm
Aricaa 18!200 70!190 20Summer 0.046 1.8 0.372 0.625 0.844 457 100Winter 0.026 0.251 0.52 0.703 370 70Santiagoa 33!330 70!190 5Summer 0.039 7.8 0.346 0.58 0.784 435 89Winter 0.005 0.108 0.202 0.293 174 29Concepciónb 36!500 72!900 5Summer 0.098 2.52 0.40 0.55 1.075 520 188Winter 0.04 0.28 0.42 0.85 425 94Valdiviac 39!480 73!140 30Summer 0.075 3.75 ⁄ 0.66 ⁄ ⁄ ⁄
Winter 0.02 0.03Pto Montta 41!300 72!55 105Summer 0.044 4.0 0.303 0.711 0.988 435 100Winter 0.011 0.144 0.333 0.479 261 42Pta Arenasa 53!08 70!55 33Summer 0.022 7.3 0.292 0.563 0.819 435 50Winter 0.003 0.065 0.174 0.27 174 18
a Cabrera et al. (1995).b Hernández et al. (present work).c Lovengreen et al. (2005).
K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32 29

between winds and surface radiation are non-linear, and furtherefforts are needed and encouraged in order to elucidate their inter-actions with the biological processes of coastal upwelling areas.Nevertheless, the results from our two statistical modeling ap-proaches provide some evidence in support of the idea that bothseasonal and synoptic meteorological variability (on a scale ofdays), through its dual influence on fertilization and solar radiationavailability, could contribute to the large differences observed inthe biological processes in the area. Integrated PP/B ratios in theConcepción time series were better explained when solar radiationwas included as an explanatory variable, improving both winterand summer predictions. In addition, when a more detailed modelconsidering self shading in the water column, in situ temperatureand nutrient profiles to reproduce in situ PP/B data was forced withdifferent surface irradiance series, the best fit was that obtainedwith the real time series.
We are aware that the modeling approach we used to relate PP/B to environmental variables at depth is a gross simplification ofthe system, where biological processes (grazing removal, physio-logical adaptation) and physical advective processes for examplehave not been taken into account. The approach and equationsused in the more complex model though, are similar to those usedin the coupled bio-physical modeling of PP, and other empiricalstudies (Bissett et al., 2001; Koné et al., 2005; Fennel and Neu-mann, 2004), and their shortcomings have been widely discussedand acknowledged in the literature. Our simplification assumed(i) maximum surface irradiance was just an indicator of surfaceradiation day to day forcing variability; (ii) constant parameterswere fitted or fixed with a single value throughout years (a, kM,C:Chl-a). Field studies so far have failed to find significant Sum-mer–Winter variability in bio-optical properties along the coastof Concepción (Montecino et al., 2004); even as the relative contri-bution to total B of diatoms decreases in winter, the same commu-nity was dominant throughout the year in Concepción ‘Station 18’(Anabalón et al., 2007). The choice regarding C:Chl-a ratio mightaccount for the lower variability of modeled vs. measured PP/Band the misfit of some particular situations (e.g. lower than ex-pected PP/Chl-a when Chl-a was extremely high and self-shading.In spite of the roughness of our two approximations, they consti-tute the first quantitative approach of its type for the system,and support the idea that ‘day to day’ irradiance variability mighthave a more important role on phytoplankton growth, and there-fore on carbon fluxes in the system, that previously thought.
Several other biological processes, their stability and budgetsmight be affected by the temporal dynamics of solar radiation ex-plored in this paper. Picoplankton abundance, diversity and func-tion for example, have been related to the depth of the lowerlimit of euphotic zone and the base of the oxycline in the South Pa-cific (Cuevas and Morales, 2006; Galán et al., 2009; Lavin et al.,2010; Molina et al., 2005), where the depth of the oxycline is veryshallow (!30–40 m), and intercepts the euphotic zone in the openocean. Experiments simulating in situ exposure to solar radiationhave shown that PAR could cause an inhibition of the bacterial sec-ondary production in the waters off Concepción. The inhibition wasestimated to be as much as 60% of dark control measurements(Hernández et al., 2006). Considering that at seasonal and synoptictime scales, upwelling and radiations co-vary, the deep communi-ties (with no recent light history) would be rapidly exposed tohigher solar radiation during intense upwelling periods. The ex-pected effects of PAR photoinhibition and reduction in bacterialsecondary production in the surface layer under such specific cir-cumstances could be higher than similar effect caused by grazingCuevas et al. (2004).
The potential effect of short time scale variability in solar radi-ation as a modulator of phytoplankton related parameters has beenlargely relegated in studies from both the North and South Pacific
upwelling systems in favor of that of winds (Carr, 2002; Hendersonand Thomas, 2007). Nevertheless, some reports (Service et al.,1999) indicate that the increase of upwelling favorable windsand light availability occurring synoptically in the Monterey Bay,were followed by an increase in Chl-a after a time lag of 4–7 days.In addition to this complexity, the taxonomic sensibility to solarradiation makes it more difficult to model relationships such asirradiance vs. PP, as shown by Ryther (1956), who used experimen-tal data to describe phytoplankton photosynthesis in function ofirradiance. Following the attempts of many models to calculatePP based on water column chlorophyll content, the incident radia-tion and extinction coefficient can nevertheless not always be ap-plied to all systems for the reasons explained above, e.g. theSouthern California Bight survey (see Fig 5 in Cullen, 1990). Sea-sonally, Vernet et al. (2009) studied biological weighted irradi-ances for (1) DNA damage (key molecules) (2) photosynthesis(ecosystem responses) based on measured radiation along SouthAmerica. Their comparison between summer and winter showedthat at least once a year at any given location, the organisms areexposed to a wide variability in effective damaging UVR. Withinthe latitudinal gradient observed, the highest sensitivity to UVBwas for DNA weighted irradiances, and to UVA for phytoplanktonweighted irradiances, with the specific effective wavelengthchanging between summer and winter. These results suggest thatin South America a predominant condition from high day to day toyearly UVR variability could be described as chronic damaging UVRexposure zone, especially for UVR sensitive biological systems.
5. Conclusion
Our description contributes a relevant environmental factor tobe considered when undertaking oceanographic research in theHCS. Because the surface solar irradiance patterns observed offthe coast of Concepción respond to a regional meteorological sys-tem, they are likely to apply to other relevant upwelling centers.The analysis presented suggests a re-consideration of the role oflight in upwelling systems, because of its potential co-modulatingrole in biological/physical processes over several time scales.Coastal upwelling has been predicted to increase in forthcomingyears, influencing ocean–atmosphere feedbacks. In such a scenario,it is important to elucidate the role of light as bio-physical driver inproductive coastal upwelling areas in order to answer questionsregarding future ecosystem trends.
Acknowledgements
We thank the Chilean meteorological directorate for data oncloudiness, sun hours, and winds. A. Pacheco and L. Cuevas pro-vided technical support for the GUV-511C. We acknowledge Dr.H. Fuenzalida (UCHILE), Dr. Dante Figueroa (DGEO-UDEC), Dr. J.Garcés (UACH), Dr. J. Letelier and R. Montoya (DGEO-UDEC), whocommented on previous stages of the time series analysis. Thiswork was supported by FONDECYT project 1020503 (CONICYT,Chile) and the FONDAP-COPAS Program (CONICYT, Chile). KHacknowledges the support of the Deutschen Akademischen Aust-auschdienst (DAAD), Programa de Mejoramiento de la Calidad yla Equidad de la Educación Superior (MECESUP-UCO, Ministeriode Educación, Chile), and the Graduate School, Universidad de Con-cepción, for the grants obtained.
References
Anabalón, V., Morales, C.E., Escribano, R., Varas, M.V., 2007. The contribution ofnano- and micro-planktonic assemblages in the surface layer (0–30 m) underdifferent hydrographic conditions in the upwelling area off Concepción, centralChile. Progress in Oceanography 75, 396–414.
30 K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32

Arcos, D.F., 1987. Seasonal and Short Time Scale Variability in Copepod Abundanceand Species Composition in An Upwelling Area off Concepción Coast, Chile.Ph.D. Dissertation. State University of New York, Stony Brook. Unpublished.
Bakun, A., 1990. Global climate change and intensification of coastal oceanupwelling. Science 24, 198–201.
Bedacht, E., Gulev, S.K., Macke, A., 2006. Intercomparison of global cloud cover fieldsover oceans from the VOS observations and NCEP/NCAR reanalysis.International Journal of Climatology 27 (13), 1707–1719.
Bissett, W.P., Schofield, O., Glenn, S., Cullen, J.J., Miller, W., Pluddeman, A., Mobley,C., 2001. Resolving the impacts and feedbacks of ocean optics on upper oceanecology. Oceanography 14, 30–53.
Booth, C.R., Lucas, T.B., Morrow, J.H., Weiler, C.S., Penhale, P.A., 1994. The UnitedStates National Science Foundation’s 50 polar network for monitoringultraviolet radiation. American Geophysical Union: Antarctic Research Series62, 17–37.
Borkowski, J., 2000. Homogenization of the Belsk UVB series (1976–1997) and trendanalysis. Journal of Geophysical Research 105, 4873–4878.
Böttjer, D., Morales, C.E., 2005. Microzooplankton grazing in a coastal embaymentoff Concepcion, Chile (!36!S) during non-upwelling conditions. Journal ofPlankton Research 27 (4), 383–391. doi:10.1093/plankt/fbi012.
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Multimodel Inference: APractical Information-Theoretic Approach. Springer Science, USA, 497 pp.
Cabrera, S., Fuenzalida, H., 1999. Interannual variations of global UV radiation inSantiago, Chile (33.5!S). Geophysical Research Letters 26, 2945–2948.
Cabrera, S., Bozzo, S., Fuenzalida, H., 1995. Variations in UV radiation in Chile.Journal of Photochemistry and Photobiology 28, 137–142.
Carr, M.E., 2002. Estimation of potential productivity in Eastern Boundary Currentsusing remote sensing. Deep-Sea Research II 49, 59–80.
Cede, A., Blumthaler, M., Luccini, E., Piacentini, R.D., Nuñez, L., 2002. Effects ofclouds on erythermal and total irradiance as derived from data of the ArgentineNetwork. Geophysical Research Letters 29. doi:10.1029/2002GL015708.
Church, M.J., Ducklow, H.W., Karl, D.M., 2004. Light dependence of [3H] leucineincorporation in the oligotrophic North Pacific Ocean. Applied andEnvironmental Microbiology 70, 4079–4087.
Cloern, J.E., Grenz, C., Vidergar-Lucas, L., 1995. An empirical model for thephytoplankton chlorophyll:carbon ratio to conversion factor betweenproductivity and growth rate. Limnology and Oceanography 40, 1313–1321.
Cuevas, L.A., Morales, C.E., 2006. Nanoheterotroph grazing on bacteria andcyanobacteria in oxic and suboxic waters in coastal upwelling areas offnorthern Chile. Journal of Plankton Research 28 (4), 385–397. doi:10.1093/plankt/fbi124.
Cuevas, L.A., Daneri, G., Jacob, B., Montero, P., 2004. Microbial abundance andactivity in the seasonal upwelling area off Concepción (!36!S), central Chile: acomparison of 20 21 upwelling and non-upwelling conditions. Deep-SeaResearch II 51, 2427–2440.
Cullen, J.J., 1990. On models of growth and photosynthesis in phytoplankton. Deep-Sea Research 37, 667–683.
Dellarosa, V., 1998. Producción primaria anual en sistemas de alta producciónbiológica. Tesis Doctoral en Botánica. Universidad de Concepción, Concepción,Chile, unpublished.
Diaz, S., Camilión, C., Deferrari1, G., Fuenzalida, H., Armstrong, R., Booth, C., Paladini,A., Cabrera, S., Casiccia, C., Lovengreen, C., Pedroni, J., Rosales, A., Zagarese, H.,Vernet, M., 2006. Ozone and UV radiation over Southern South America:climatology and anomalies. Photochemistry and Photobiology 82 (4), 834–843.
Emery, W.J., Thomson, R.E., 1997. Data Analysis Methods in Physical Oceanography.Elsevier Science Ltd., Oxford.
Fasham, M.J.R., Ducklow, H.W., McKelvie, S.M., 1990. A nitrogen based model ofplankton dynamics in the ocean mixed layer. Journal Marine Research 48, 591–639.
Fennel, W., Neumann, T., 2004. In: Halpern, D. (Ed.), Introduction to the Modeling ofMarine Ecosystems. Elsevier Oceanography series 72, berlin, p. 97 pp.
Franklin, L.A., Foster, R.M., 1997. The changing irradiance environment:consequences for marine macrophyte physiology, productivity and ecology.European Journal of Phycology 32, 207–232.
Frederick, J.E., Lubin, D., 1988. The budget of biologically active ultraviolet radiationin the earth–atmosphere system. Journal of Geophysical Research 93 (D4),3825–3832. doi:10.1029/JD093iD04p03825.
Fritz, J.J., Neale, P.J., Davis, R.F., Peloquin, J.A., 2008. Response of Antarcticphytoplankton to solar UVR exposure: inhibition and recovery ofphotosynthesis in coastal and pelagic assemblages. Marine Ecology ProgressSeries 365, 1–16. doi:10.3354/meps07610.
Fuenzalida, H.A., Sánchez, R., Garreaud, R.D., 2005. A climatology of cutoff lows inthe Southern Hemisphere. Journal of Geophysical Research 110, D18101.doi:10.1029/2005JD005934.
Galán, A., Molina, V., Thamdrup, B., Woebken, D., Lavik, G., Kuypers, M.M.M., Ulloa,O., 2009. Anammox bacteria and the anaerobic oxidation of ammonium in theoxygen minimum zone off northern Chile. Deep Sea Research II 56 (16), 1021–1031. doi:10.1016/j.dsr2.2008.09.016.
García, R., Cisneros, J.M., Hernández, E., Manzano, J., Diaz, J., 2000. Time modeling ofdaily UVB values in Madrid, Spain. Journal of Applied Meteorology 39, 117–123.
Garreaud, R.D., Rutllant, J.A., Fuenzalida, H., 2002. Coastal lows along the subtropicalWest Coast of South America: mean structure and evolution. Monthly WeatherReview 130, 75–88.
Garreaud, R., Barichivic, J., Christie, D.A., Maldonado, A., 2008. Interannualvariability of the coastal fog at Fray Jorge relict forests in semiarid Chile.Journal of Geophysical Research Biogeosciences 113, G4G04011.
Garreaud, R.D., Rutllant, J.A., Muñoz, R.C., Rahn, D.A., Ramos, M., Figueroa, D., 2011.VOCALS-CUpEx: The chilean upwelling experiment. Atmospheric Chemistryand Physics 11, 2015–2029. doi:10.5194/acp-11-2015-2011.
González, H.E., Daneri, G., Iriarte, J.L., Yannicelli, B., Menschel, E., Barría, C., Pantoja,S., Lizárraga, L., 2009. Carbon fluxes within the epipelagic zone of the Humboldtcurrent system off Chile: the significance of euphausids and diatoms as keyfunctional groups for the biological pump. Progress in Oceanography 83, 217–227.
Häder, D.P., Kumar, H.D., Smith, R.C., Worrest, R., 2003. Aquatic ecosystems: effectsof solar ultraviolet radiation and interactions with other climatic changefactors. Photochemical and Photobiological Sciences 2, 39–50.
Häder, D.P., Lebert, M., Schuster, M., del Ciampo, L., Helbling, E.W., McKenzie, R.,2007. ELDONET—a decade of monitoring solar radiation on five continents.Photochemistry and Photobiology 83, 1348–1357.
Helbling, E.W., Avaria, S., Letelier, J., Montecino, V., Ramirez, B., Ramos, M., Rojas, W.,Villafañe, V.E., 1993. Respuesta del fitoplancton marino a la radiaciónultravioleta en latitudes medias (33!S). Revista de Biología Marina, Valparaíso28, 219–237.
Henderson, S.A., Thomas, A.C., 2007. Interannual variability in timing of bloominitiation in the California current system. Journal of Geophysical Research 112.doi:10.1029/2006JC003960.
Hernández, K.L., Quiñones, R.A., Daneri, G., Helbling, E.W., 2006. Effects of solarradiation on bacterioplankton assemblages from the highly productive systemoff Central-South Chile. Marine Ecology Progress Series 315, 19–31.
Hernández, K., Quiñones, R., Daneri, G., Farias, M.E., Helbling, E.W., 2007. Solar UV-solar radiation modulates daily production and DNA damage of marinebacterioplankton from a productive upwelling zone (36!S). Journal ofExperimental Marine Biology and Ecology 343 (1), 82–95.
Hill, A.E., Hickey, B.M., Shillington, F.A., Strub, P.T., Brink, K.H., Barton, E.D., Thomas,A.C., 1998. Eastern boundary currents: a pan-regional review. In: Robinson, A.R.,Brink, K.H. (Eds.), The Sea, vol. 11. John Wiley, New York, pp. 29–68.
Kalnay, E., Kanamitsu, M., Kistler, R., Collins, W., Deaven, D., Gandin, L., Iredell, M.,Saha, S., White, G., Woollen, J., Zhu, Y., Leetmaa, A., Reynolds, R., Chelliah, M.,Ebisuzaki, W., Higgins, W., Janowiak, J., Mo, K.C., Ropelewski, C., Wang, J., Jenne,R., Dennis, J., 2006. The NCEP/NCAR reanalysis project. Bulletin of AmericanMeteorology Society 77, 437–471.
Kirchoff, V.W.J.H., Schuch, N.J., Pinheiro, D.K., Harris, J.M., 1996. Evidence for anozone hole perturbation at 30! South. Atmospheric Environment 30, 1481–1488.
Koné, V., Machu, E., Penven, P., Andersen, Garçon, V., Freón, P., Demarcq, H., 2005.Modeling the primary and secondary production of the southern Benguelaupwelling system: a comparative study through two biogeochemical models.Global Biogeochemical Cycles 19, GB4021.
Lavin, P., González, B., Santibáñez, J.F., Scanlan, D.J., Ulloa, O., 2010. Novel lineages ofProchlorococcus thrive within the oxygen minimum zone of the eastern tropicalSouth Pacific. Environmental Microbiology Reports 2 (6), 728–738. doi:10.1111/j.1758-2229.2010.00167.x.
Liao, Y., Frederick, J.E., 2005. The ultraviolet radiation environment of high southernlatitudes: springtime behavior over a decadal timescale. Photochemistry andPhotobiology 81, 320–324.
Logan, J.A., 1994. Trends in the vertical distribution of ozone: an analysis of ozonedata. Journal of Geophysical Research 99, 25553–25585.
Lovengreen, C., Fuenzalida, H., Villanueva, L., 2000. Ultraviolet solar radiation atValdivia, Chile (39.8!S). Atmospheric Environment 34, 4051–4061.
Lovengreen, C., Fuenzalida, H., Videla, L., 2005. On the spectral dependency of UVradiation enhancements due to clouds in Valdivia, Chile (39.8!S). Journal ofGeophysical Research 110, D14207. doi:10.1029/2004JD005372.
Lubin, D., Frederick, J., 1989. Measurements of enhanced springtime ultravioletspringtime radiation at palmer station. Geophysical Research Letters 16, 783–785.
McKenzie, R.L., Björn, L.O., Bais, A., Ilyasd, M., 2003. Changes in biologically activeultraviolet radiation reaching the Earth’s surface. Photochemical andPhotobiological Sciences 2, 5–15.
Mc Manus, G.B., Peterson, W.T., 1988. Bacterioplankton production in the nearshore zone during upwelling off central Chile. Marine Ecology Progress Series43, 11–17.
McPeters, R.D., Hollandsworth, S.M., Flynn, L.E., Herman, J.R., Seftor, C.J., 1996. Long-term ozone trends derived from the 16-year combined Nimbus-7/Meteor-3TOMS version-7 record. Geophysical Research Letters 23, 3699–3702.
Molina, V., Farías, L., Eissler, Y., Cuevas, L.A., Morales, C.E., Escribano, R., 2005.Ammonium cycling under the strong oxygen gradient associated with OxygenMinimum Zone off northern Chile (!23!S). Marine Ecology Progress Series 288,35–43. doi:10.3354/meps288035.
Montecino, V., Pizarro, G., 1995. Phytoplankton acclimation and spectralpenetration of UV irradiance off the central Chilean coast. Marine EcologyProgress Series 121 (1–3), 261–269.
Montecino, V., Astoreca, R., Alarcón, G., Retamal, L., Pizarro, G., 2004. Bio-opticalcharacteristics and primary productivity during upwelling and non-upwellingconditions in a highly productive coastal ecosystem off central Chile (36!S).Deep-Sea Research II 51, 2413–2426.
Montero, P., Daneri, G., Cuevas, L.A., González, H.E., Jacob, B., Lizárraga, L., Menschel,E., 2007. Productivity cycles in the coastal upwelling area off Concepción: theimportance of diatoms and bacterioplankton in the organic carbon flux.Progress in Oceanography 75 (3), 518–530. doi:10.1016/j.pocean.2007.08.013.
Neale, P., Helbling, E.W., Zagarese, H., 2003. Modulation of UVR exposure and effectsby vertical mixing and advection. In: Helbling, E.W., Zagarese, H.E. (Eds.), UV
K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32 31

Effects in Aquatic Organisms and Ecosystems. The Royal Society of Chemistry,Cambridge, pp. 107–134.
Olivieri, R.A., Chavez, F.P., 2000. A model of plankton dynamics for the coastalupwelling system of Monterey Bay, California. Deep Sea Research II 47, 1077–1106.
Orce, V.L., Helbling, E.W., 1997. Latitudinal UVR–PAR measurements in Argentina:extent of the ozone hole. Global Planetary Change 15, 113–121.
Oschilies, A., Garçon, V., 1999. An eddie-permitting coupled physical-biologicalmodel of the North Atlantic: 1. Sensitivity to advection numerics and mixedlayer physics. Global Biogeochemical Cycles 13, 135–160.
Pond, S., Pickard, G.L., 1997. Introductory Dynamical Oceanography, second ed.Oxford, 329 pp.
Rowland, F.S., 1991. Stratospheric ozone depletion – review. Annual Review ofPhysics and Chemistry 42, 731–768.
Rutllant, J., Rosenbluth, B., Hormazabal, S., 2004. Intraseasonal variability in thelocal wind forcing of coastal upwelling off central Chile (30!S). Continental ShelfResearch 24, 789–804. doi:10.1016/j.csr.2004.02.004.
Ryther, J.H., 1956. Photosynthesis in the ocean as a function of light intensity.Limnology and Oceanography 2, 281–286.
Saavedra, N., 1980. La presión y la dirección del viento en Concepción. Tralka (Chile)1, 153–162.
Saavedra, N., 1985. Modelo Climático simple para Concepción. Geoacta 13, 13–26.Saavedra, N., Foppiano, A.J., 1992. Monthly mean pressure model for Chile.
International Journal of Climatology 12, 469–480.Seidlitz, H.K., Thiel, S., Krins, A., Mayer, H., 2001. Solar radiation at the Earth’s
surface. In: Giacomoni, P.U. (Ed.), Comprehensive Series in Photosciences. ‘SunProtection in Man’, vol. 3. Elsevier, Amsterdam, pp. 705–738.
Service, S.K., Rice, J.A., Chavez, F.P., 1999. Relationship between physical andbiological variables during the upwelling period in Monterey Bay, CA. Deep SeaResearch Part II: Topical Studies in Oceanography 45, 1669–1685.
Shaffer, G., Hormazabal, S., Pizarro, O., Salinas, S., 1999. Seasonal and interannualvariability of currents and temperature off central Chile. Journal of GeophysicalResearch 104 (C12), 29951–29961.
Siegel, D.A., Ohlmann, C., Washburn, L., Bidigare, R.R., Nosse, C.T., Fields, E., Zhou, Y.,1995. Solar radiation, phytoplanckton pigments and the radiant heating of theequatorial warm pool. Journal of Geophysical Research 100 (C3), 4885–4891.
Sobarzo, M., Shearman, R.K., Lentz, S., 2007. Near-inertial motions over thecontinental shelf off Concepción, central Chile. Progress in Oceanography 75(3), 348–362.
Stolarsky, R.S., Bloomfield, P., McPeters, R.D., Herman, J.R., 1991. Total ozone trendsdeduced from Nimbus-7 TOMS data. Geophysical Research Letters 18, 1015–1018.
Stramska, M., Dickey, T.D., 1998. Short term variability of the underwater light fieldin the oligotrophic ocean in response to surface waves and clouds. Deep SeaResearch I 45, 1393–1410.
Strub, T., Mesias, J.M., Montecino, V., Rutllant, J., Salinas, S., 1998. Coastal oceancirculation off western South America Coastal segment (6,E). In: Robinson, A.R.,Brink, K.H. (Eds.), The Sea, vol. 11. John Wiley & Sons Inc., BErlin, pp. 273–313.
Taylor, A.H., Geider, R.J., Gilbert, F.J.H., 1997. Seasonal and latitudinal dependenciesof phytoplankton carbon to chlorophyll a ratios: results of a modeling study.Marine Ecology Progress Series 152, 51–66.
Vernet, M., Diaz, S., Fuenzalida, H., Camilión, C., Booth, C., Cabrera, S., Casiccia, C.,Deferrari1, G., Lovengreen, C., Paladini, A., Pedroni, J., Rosales, A., Zagarese, H.,2009. Quality of UVR exposure for different biological systems along alatitudinal gradient. Photochemical and Photobiological Sciences 8, 1329–1345.
Villafañe, V.E., Helbling, E.W., Zagarese, H.E., 2001. Solar ultraviolet radiation and itsimpact on aquatic ecosystems of Patagonia, South America. Ambio 30, 112–117.
von Storch, H., Zwiers, F.W., 1999. Statistical Analysis in Climate Research.Cambridge University Press, New York.
Wilhelm, S.W., Jeffrey, W.H., Dean, A.L., Meador, J., Pakulski, J.D., Mitchell, D., 2003.UV radiation induced DNA damage in marine viruses along a latitudinalgradient in the south eastern Pacific Ocean. Aquatic Microbial Ecology 31, 1–8.
World Meteorological Organization (WMO), 1965. International Cloud Atlas,Abridged.
32 K. Hernández et al. / Progress in Oceanography 92–95 (2012) 18–32
Copyright © 2022 FDOKUMEN




















