
Environmental management and open-air experiments in Brazilian Amazonia
Upload
eastangliaCategory
view
0download
0
Surface wildfires in central Amazonia: short-term impacton forest structure and carbon loss
Torbjørn Haugaasen*, Jos Barlow, Carlos A. PeresCentre for Ecology, Evolution and Conservation, School of Environmental Sciences,
University of East Anglia, Norwich NR4 7TJ, UK
Received 8 November 2001; received in revised form 15 July 2002; accepted 7 October 2002
Abstract
Changes in forest structure were examined 10–15 months after an unprecedented understorey wildfire burnt previously
undisturbed primary forest in central Brazilian Amazonia, following the severe 1997–1998 El Nino dry season. On the basis of
20 0.25 ha plots (10 m � 250 m) in both burnt and unburnt forest, we found marked differences in the overall live biomass,
canopy openness and understorey vegetation. On average, 36% of all trees equal to or greater than 10 cm DBH were found to be
dead in the burnt forest, and there was also a near-complete mortality in all pre-burn saplings. Using an allometric equation to
predict biomass mortality we estimate that the tree mortality rates found would commit an additional 25.5 t C/ha to be released
from these BFs. The dramatic increase of aboveground dead biomass in BF is of major global concern because of the increased
flux of CO2 to the atmosphere, which has a role in enhancing the greenhouse effect and promoting climate change.
# 2002 Published by Elsevier Science B.V.
Keywords: Aboveground biomass; Carbon emissions; El Nino; Fire disturbance; Global warming; Surface fires
1. Introduction
Tropical forests are widely known to harbour the
highest species diversities in the world, but are cur-
rently under enormous pressure from rising human
populations (Pahari and Murai, 1999; Cincotta et al.,
2000), growing agricultural practices (Dale and Pear-
son, 1997), logging (Nepstad et al., 1999a,b; Putz et al.,
2001) and other disturbances such as hunting (Robin-
son and Bennett, 2000; Peres, 2000). In the Brazilian
Amazonia alone, a forest area the size of France has
been cleared in the last three decades (Fearnside,
1999), affecting an area nearly twice as large through
edge effects and forest fragmentation (Skole and
Tucker, 1993). In these areas, synergistic interactions
between structural and non-structural forms of dis-
turbance continue to erode biodiversity (Peres, 2001).
Since the mid 1980s, however, tropical forests have
been haunted by yet another threat which has steadily
increased in importance as greater areas of forest
become affected each year—surface wildfires.
Tropical rain forests are normally thought to be fire-
resistant environments because of high atmospheric
and soil moisture levels, and major wildfires have been
historically rare events (Turcq et al., 1998). Charcoal
dating evidence suggests that catastrophic fires have
occurred in the Amazon only four times in the past two
millennia, at 1500, 1000, 700 and 400 BP (Sanford
Forest Ecology and Management 6154 (2002) 1–11
* Corresponding author. Tel.: þ44-1603-591099;
fax: þ44-1603-507719.
E-mail address: [email protected] (T. Haugaasen).
0378-1127/02/$ – see front matter # 2002 Published by Elsevier Science B.V.
PII: S 0 3 7 8 - 1 1 2 7 ( 0 2 ) 0 0 5 4 8 - 0

et al., 1985) whilst the pollen record suggests that
there was a period of fires lasting from 7000 to
4000 years BP (Turcq et al., 1998). However, as a
result of recent climatic oscillations and large-scale
disturbances such as logging and fragmentation, fire is
becoming an increasingly common pantropical phe-
nomenon (Goldhammer, 1999). In Amazonia, surface
wildfires threaten a much greater area of forest than
does deforestation per se (Nepstad et al., 1999a;
Cochrane, 2001), even though current deforestation
rates in the Amazon are the fastest anywhere (Whit-
more, 1997).
In contrast to the far more detectable canopy fires,
surface fires rarely exceed 10–30 cm in height under
normal fuel and humidity conditions, burning the fine
and course litter on the forest floor (Holdsworth and
Uhl, 1997; Cochrane and Schulze, 1999; Nepstad et al.,
1999a). However, these apparently innocuous fires
have been shown to have serious detrimental effects
on both the forest structure (Peres, 1999; Barbosa and
Fearnside, 1999) and the vertebrate fauna (Kinnaird
and O’Brien, 1998; Haugaasen, 2000; Barlow et al.,
2002; Peres et al., in press), because of their extremely
rare occurrence in evolutionary time (Uhl and Kauff-
man, 1990).
By killing a substantial amount of the forest bio-
mass, the fires increase the carbon flux to the atmo-
sphere (Barbosa and Fearnside, 1999), enhancing the
greenhouse effect and promoting climate change. In
particular, this source of carbon release has been
severely underestimated or disregarded in estimates
of the carbon contribution to the atmosphere asso-
ciated with land cover changes in Amazonia (Fearn-
side, 1997a; Laurance et al., 1998).
In this paper we present tree mortality data and
subsequent changes in forest structure following a
large surface fire in central Brazilian Amazonia. We
provide estimates of the aboveground dead biomass
(AGDB) resulting from this unprecedented wildfire,
and estimate the amount of carbon that will eventually
be released from these fires (committed carbon emis-
sions). We also discuss the degradation and impover-
ishment of tropical forests in light of potentially
irreversible ecosystem transitions given a scenario
of more severe or more frequent El Nino events.
Finally, we consider whether accidental fires are likely
to become a permanent feature of many previously
undisturbed neotropical forest ecosystems.
2. Methods
2.1. Study area
The study was conducted on both banks of the Rio
Maro, a tributary of the Rio Arapiuns which flows into
the mouth of the Rio Tapajos, westernmost Para,
Brazil (028440S; 558410W; see Barlow et al. (2002)
or Peres et al. (in press) for a map of the study area and
sample plots). The work took place from October 1998
to March 1999, around 1 year after an accidental
wildfire swept through large parts of the forest on
both sides of the Rio Maro.
Average annual rainfall at the nearest meteorologi-
cal station (Santarem) is 2041 mm epr year (range ¼1287�2538 mm per year 1992–1997; INFRAERO,
1998), with a pronounced dry season typically lasting
3–5 months, normally from July to November. The
recent fires occurred in November–December of 1997,
at the end of the longest dry season in living memory.
A rainless period of 110 days, linked to the 1997–1998
El Nino Southern Oscillation (ENSO) event led to leaf
abscission and permitted sunlight to penetrate the
understorey, allowing normally fire-resistant forests
to become flammable. The soils in this region are
typically sandy (podzolic) and highly permeable,
which further increases the risk of breaching the forest
flammability threshold (Hammond and ter Steege,
1998).
2.2. Data collection
Data were obtained within 20 0.25 ha plots (10 m�250 m) located in burnt and unburnt forest across the
Maro river basin. Sixteen plots were placed >500 m
from the clearly distinguishable fireline, and were
equally split between burnt and unburnt forest on both
banks of the river. A further four plots were placed
perpendicular to, and cutting across the fireline so that
each half of these plots (0.125 ha) lay in either burnt or
unburnt forest. The four forest treatments are hereafter
referred to as BF (burnt forest), UF (unburnt forest),
BFI (burnt forest interface) and UFI (unburnt forest
interface). In order to compare the BF and UF plots
away from the fireline with those cutting across the
fireline interface, tree and basal area densities are
expressed on a per hectare basis. Biomass and carbon
values were converted likewise.
2 T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11
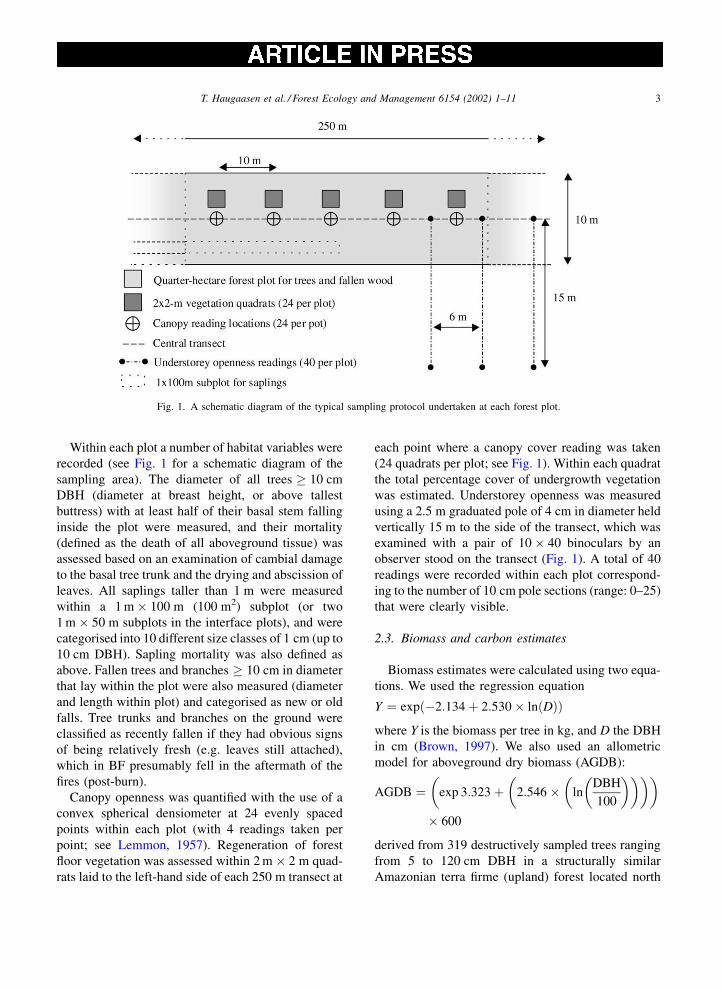
Within each plot a number of habitat variables were
recorded (see Fig. 1 for a schematic diagram of the
sampling area). The diameter of all trees � 10 cm
DBH (diameter at breast height, or above tallest
buttress) with at least half of their basal stem falling
inside the plot were measured, and their mortality
(defined as the death of all aboveground tissue) was
assessed based on an examination of cambial damage
to the basal tree trunk and the drying and abscission of
leaves. All saplings taller than 1 m were measured
within a 1 m � 100 m (100 m2) subplot (or two
1 m � 50 m subplots in the interface plots), and were
categorised into 10 different size classes of 1 cm (up to
10 cm DBH). Sapling mortality was also defined as
above. Fallen trees and branches � 10 cm in diameter
that lay within the plot were also measured (diameter
and length within plot) and categorised as new or old
falls. Tree trunks and branches on the ground were
classified as recently fallen if they had obvious signs
of being relatively fresh (e.g. leaves still attached),
which in BF presumably fell in the aftermath of the
fires (post-burn).
Canopy openness was quantified with the use of a
convex spherical densiometer at 24 evenly spaced
points within each plot (with 4 readings taken per
point; see Lemmon, 1957). Regeneration of forest
floor vegetation was assessed within 2 m � 2 m quad-
rats laid to the left-hand side of each 250 m transect at
each point where a canopy cover reading was taken
(24 quadrats per plot; see Fig. 1). Within each quadrat
the total percentage cover of undergrowth vegetation
was estimated. Understorey openness was measured
using a 2.5 m graduated pole of 4 cm in diameter held
vertically 15 m to the side of the transect, which was
examined with a pair of 10 � 40 binoculars by an
observer stood on the transect (Fig. 1). A total of 40
readings were recorded within each plot correspond-
ing to the number of 10 cm pole sections (range: 0–25)
that were clearly visible.
2.3. Biomass and carbon estimates
Biomass estimates were calculated using two equa-
tions. We used the regression equation
Y ¼ expð�2:134 þ 2:530 � lnðDÞÞ
where Y is the biomass per tree in kg, and D the DBH
in cm (Brown, 1997). We also used an allometric
model for aboveground dry biomass (AGDB):
AGDB ¼ exp 3:323 þ 2:546 � lnDBH
100
� �� �� �� �
� 600
derived from 319 destructively sampled trees ranging
from 5 to 120 cm DBH in a structurally similar
Amazonian terra firme (upland) forest located north
Fig. 1. A schematic diagram of the typical sampling protocol undertaken at each forest plot.
T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11 3

of Manaus, Brazil (Santos, 1996). While there are
many potential problems with using DBH to estimate
forest biomass and carbon (e.g. Fearnside, 1997b), the
Santos equation formulated through the destructive
harvesting of 319 trees near Manaus is considered to
be the most representative of the Brazilian Amazon,
being further supported by results from another site in
eastern Amazonia (Araujo et al., 1997). We therefore
focus our discussion on the results from this equation,
although include the Brown equation for its compara-
tive value. Following Fearnside (1997a) and Brown
(1997), the carbon content of biomass was assumed to
be 50%.
2.4. Data analysis
One-way ANOVAs (with Tukey’s HSD post-hoc
test) were used to test for differences in the forest
structure between the four treatments (Table 1). Data
were checked for normality, and subsequently left
untransformed. In order to conduct the examination
of size-dependent mortality (Fig. 2), data from all
plots was pooled, and split into two categories accord-
ing to whether the forest had burnt or not. This pooling
of all forest plots minimised the stochastic variation in
tree densities inherent within relatively small 0.25 ha
plots, and was necessary to increase the number of
trees sampled in the larger (and hence rarer) DBH
classes. Data for saplings was treated similarly.
Paired-t-tests were used to compare the results of
the two biomass equations.
3. Results
3.1. Tree mortality
A total of 2643 standing trees were measured in the
20 forest plots, with 1341 being found in the UF
(2.5 ha) and 1302 in BF (2.5 ha). The abundance of
trees was strongly size-dependent, and both forest types
showed a reverse J distribution curve for the tree
assemblage as a whole (Fig. 2a). Trees smaller than
20 cm DBH accounted for 74% of all trees in all BF and
Table 1
Habitat data obtained from four groups of forest plots (*, y: subsets from Tukey’s HSD)a
Mean (S.E.) per plot in d.f. F P
UF ðn ¼ 8Þ UFI ðn ¼ 4Þ BFI ðn ¼ 4Þ BF ðn ¼ 8Þ
No. trees per hectare 533.5 19.2 548.0 21.8 528.0 64.7 519.0 20.8 3, 20 0.16 0.92
No. trees dead per hectare 24.0* 3.1 28.0* 4.0 152.0y 41.2 184.0y 9.2 3, 20 34.69 <0.001
Basal area per hectare 121.2 7.3 110.9 5.6 123.1 28.1 105.9 9.2 3, 20 0.49 0.69
Basal area dead per hectare 6.2* 1.2 6.5* 1.3 17.8y 2.1 25.7y 2.5 3, 20 24.53 <0.001
Brown equation
Total biomass per hectare 349.9 33.2 285.4 23.2 342.4 99.1 282.6 32.9 3, 20 0.67 0.58
Dead biomass per hectare 16.9* 4.5 15.1* 3.5 38.6*,y 6.6 593*,y 7.9 3, 20 11.18 <0.001
Potential Carbon loss per hectare 8.5* 2.3 7.6* 1.8 19.3*,y 3.3 29.7y 4 3, 20 11.18 <0.001
Santos equation
Total biomass per hectare 424.3 40.7 344.6 28.4 414.5 120.7 341.7 40.1 3, 20 0.67 0.58
Dead biomass per hectare 20.5* 5.6 18.2* 4.3 46.4*,y 8 71.4y 9.6 3, 20 10.93 <0.001
Potential Carbon loss per hectare 10.2* 2.8 9.1* 2.1 23.2*,y 4 35.7y 4.8 3, 20 10.93 <0.001
Other habitat data
All fallen wood per hectare (m3) 45.7 9.1 50.1 8.5 30.4 3.2 45.2 5.9 3, 20 0.81 0.51
Newly fallen wood per hectare (m3) 0.8* 0.4 2.3* 1.7 12.9y 3.3 10*,y 2.8 3, 20 6.58 0.003
% Saplings dead 4.4* 1.0 3.1* 0.7 63.1y 13.2 76y 5.7 3, 20 47.03 <0.001
% Canopy gap 4.7* 0.6 5.5* 0.5 19.5y 6.2 19.2y 2.1 3, 20 11.34 <0.001
% Ground vegetation cover 20.6 3.5 25.7 3.6 50.9 15.7 48.5 6.9 3, 20 4.59 0.01
Understorey vegetation openness 4.5 0.8 6.6 1.3 9.6 3 9.1 1.2 3, 20 2.98 0.06
a UF: unburnt forest >500 m from the fireline; UFI: unburnt forest plots adjoining and perpendicular to the fireline; BF: burnt forest
>500 m from the fireline; BFI: burnt forest plots adjoining and perpendicular to the fireline.
4 T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11

UF plots. There was no significant difference between
the mean overall number of trees per hectare or the
mean basal area per hectare in any forest type (Table 1).
As expected, however, tree mortality, was clearly
greater in BF plots; 36% of all trees in BF plots were
classified as dead, compared to only 4.5% of those in
UF plots. The number of dead trees per hectare and
the mean dead basal area per hectare were thus
significantly greater in the BF plots than in the
UF plots. Distance to the fireline did not appear
Fig. 2. (a) Frequency distribution of the size of standing trees expressed in terms of DBH in all BF and UF plots. Relationship between tree
DBH for UF and BF plots and tree survival (b), and (c) the estimated AGDB, using both the Brown (1997) and Santos (1996) equations.
T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11 5

to have a significant effect on tree mortality, and the
interface plots were not significantly different from
those >500 m from the fireline in either BF or UF
(Table 1).
In order to examine size-dependent mortality, all
trees were classed according to their DBH. Within the
BF, there was a strong positive relationship between
the DBH class of the tree and the percentage of live
trees (r2 ¼ 0:77, F1;10 ¼ 33:7, P ¼< 0:001), with
smaller stems succumbing to higher mortality
(Fig. 2b). No such relationship was apparent in UF
plots, where trees appeared to have a mortality peak
around 30–35 cm DBH (Fig. 2b).
3.2. Sapling mortality
Mortality of saplings averaged 76% in the eight BF
plots and 63.1% in the four BFI plots. Both of these
means were significantly different from the 3 to 4%
background mortality rate found in the UF treatments
(Table 1). The proportion of dead saplings in each
DBH class, however, was not significantly related to
stem diameter in either BF or UF plots (P < 0:5 in
both the cases).
3.3. Fallen dead wood
There was no significant difference between the
total volume of fallen wood per hectare in the four plot
categories (Table 1), though the mean values were
variable, ranging from 30.4 to 50.1 m3 per hectare.
However, the BFIs had a significantly greater number
of recently fallen trees than either unburnt forest
treatment (UF and UFI).
3.4. Canopy openness and understorey
regeneration
Canopy openness in all BF plots was significantly
greater than that in UF (Table 1), this being largely
explained by the high density of the leafless crowns of
dead canopy trees left standing in the BF. Although
there was significant differences in the percentage
ground vegetation cover in the understorey, the Tukey’s
post-hoc test showed that no treatment was signifi-
cantly different from another. However, understorey
vegetation cover was found to be significantly related
to percentage canopy openness (Fig. 3). The regen-
eration of the BF understorey was highly variable in
Fig. 3. Relationship between canopy openness and understorey vegetation cover for forest plots >500 m from the fireline. Error bars represent
standard errors. Regression line was fitted to mean values.
6 T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11

terms of structure and species composition, though
Cecropia spp., Palicourea guianensis (Rubiaceae),
and Aparisthmium cordatum (Euphorbiaceae) were
the most common pioneer tree species.
3.5. Forest biomass and potential carbon loss
Using either the Brown or Santos equations to
calculate tree biomass, there were no significant dif-
ferences between the mean total biomass density
across any of the forest treatment types (Table 1).
However, there were significant differences between
the mean dead biomass per hectare, with both equa-
tions displaying similar trends. The eight BF plots
contained a significantly greater dead biomass per
hectare than the UF and UFI, while the BFI was
intermediate and did not differ significantly from
any of the other forest treatments. Fig. 2c shows
how the total AGDB is distributed among each
DBH class comparing the two equations.
The Santos equation added considerable tonnage to
the biomass estimates, and the total estimates for both
UF and BF from each equation were significantly
different (mean Santos UF S:E: ¼ 424:3 40:7ðn ¼ 8Þ; mean Brown UF S:E: ¼ 349:9 33:2ðn ¼ 8Þ; paired-t ¼ 9:6, d:f: ¼ 7, P ¼< 0:001; mean
Santos BF S:E: ¼ 341:7 40:1 ðn ¼ 8Þ; mean
Brown BF S:E: ¼ 282:6 32:9 ðn ¼ 8Þ; paired-
t ¼ 8:2, d:f: ¼ 7, P ¼< 0:001).
The total potential carbon loss (due to the increase
in dead AGDB) in BF plots was 3.5 times than that
found in the equivalent UF plots, an increase of
25.5 t C/ha using the Santos equation. The difference
across the interface was less dramatic, though the total
potential loss in the BFI was still 2.5 times than that of
the UFI, an increase of 14.1 t C/ha using the Santos
equation.
4. Discussion
4.1. Tree mortality and biomass
The mortality of 36% of all trees � 10 cm DBH in
the BF plots was 4.5 times greater than that reported
for seven post-burn plots of 750 m2 at three forest sites
in Roraima (Barbosa and Fearnside, 1999) but closely
matches that of previously logged eastern Amazonian
forests near Paragominas (Holdsworth and Uhl, 1997;
Cochrane and Schulze, 1999; Table 2). This study thus
shows that seasonal central Amazonian forests such as
around the Maro can be severely affected by surface
wildfires during strong El Nino events, even if their
history of logging disturbance has been negligible.
Mortality and AGDB estimates in this study may also
have been underestimated. The study by Peres (1999)
carried out in the same region immediately after the
fires estimated a standing dead biomass at 43.3 t/ha, a
figure almost 40% lower than our estimates, suggest-
ing that there had been an increase in mortality in the
following year. We may expect this increase in mor-
tality to continue following exposure to sublethal
thermal stress; other studies in both Amazonia (Holds-
worth and Uhl, 1997) and Sumatra (Sunarto, 2000)
have shown mortality to increase for up to 2 years after
the initial fire disturbance as injured trees gradually
succumb to infections of fungi or other pathogens, and
eventually die whether standing or fallen. Further-
more, the occurrence of both visually and acoustically
detected tree falls plots recorded during the time of the
study was five times higher in the BF than UF (J.
Barlow, unpublished data), further opening up the
forest canopy and knocking over trees that had sur-
vived the initial fire. Another cause of underestimation
comes from the exclusion of woody lianas and vines in
Table 2
Mortality of individuals �10 DBH (n/ha) and aboveground
biomass (t/ha) from five different studies in Amazonian Brazil
Mortality
(n/ha)
Aboveground
biomass (t/ha)
Study
Dead 147 71 Cochrane and
Schulze (1999)% Dead 38 35.5
Totals 384 200
Dead 161 Holdsworth and
Uhl (1997)% Dead 44
Totals 367
Dead 46 17.4 Barbosa and
Fearnside (1999)% Dead 7.9 7.9
Totals 585 219.7
Dead 68 16.1 Santos et al.
(1998)% Dead 16
Totals 425
Dead 184 71.4a This study
% Dead 35.5 20.9a
Totals 519 341.7a
a Numbers derived with the Santos equation.
T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11 7

this study. Despite being extremely susceptible to fire
damage (Cochrane and Schulze, 1999), large (�10 cm
DBH) lianas were fairly rare in this region and were
not considered here.
Tree mortality in the BF plots appeared to be
strongly size-dependent, with small stems (10–
20 cm DBH) in BF suffering higher mortality. Uhl
and Kauffman (1990) suggest that this size-dependent
mortality may be a consequence of differences in bark
thickness, as thicker bark is better able to protect the
cambium layer from heat stress. This is consistent with
a recent study in the Arapiuns basin where bark
thickness appeared to be an important morphological
correlate of tree survival (Barlow et al., in press). In
contrast, tree mortality in UF plots peaked at 30–
40 cm DBH, which was presumably due to other
causes.
4.2. Edge and interface effects
Initially we predicted that burn intensity (and hence
tree mortality) may have been lower close to the
fireline, as interviews with local people indicated that
the fires had been reduced in intensity by the rains that
eventually extinguished them. Furthermore, as edges
are associated with an increase in tree mortality in
tropical forest fragments (Mesquita et al., 1999) we
also expected a higher mortality and treefall rate in the
UFI close to the fireline. While we consider it note-
worthy that these predictions were supported numeri-
cally (Table 1), this must be qualified by the small
sample sizes at the interfaces, and the lack of statistical
support.
4.3. Sapling mortality and understorey regeneration
As expected from the size-dependent mortality of
trees, mortality was very high amongst the pre-burn
saplings in all BF plots matching other reports on post-
fire sapling mortality (Holdsworth and Uhl, 1997).
Following the fires, the greatly increased light envir-
onment in the understorey seemed to favour the rapid
growth of early successional tree species, although in
other areas that had apparently been severely burned
the growth of aggressive bamboo and sedges appeared
to inhibit seedling regeneration, which is confirmed by
findings elsewhere in the Amazon (Nepstad et al.,
1991).
4.4. Positive feedback mechanisms
One of the most alarming features of these fires is
their potential to trigger a positive feedback system,
potentially leading to the progressive impoverishment
and degradation of vast expanses of tropical forest
(Cochrane et al., 1999; Nepstad et al., 1999b). The
high post-burn tree mortality significantly reduced
canopy cover, and will therefore create a hotter and
drier microclimate as more solar radiation passes
through to the understorey, increasing the rate of fuel
drying (Uhl and Buschbacher, 1985; Uhl and Kauff-
man, 1990; Holdsworth and Uhl, 1997; Nepstad et al.,
1999b). Furthermore, the dead and dying trees that fall
to the forest floor add to the combustible fuel layer. We
calculate that nearly 14 m3 /ha of coarse fuel has been
added to the forest floor in BF just 1 year after the fire,
and the greatly augmented treefall rate suggests that
much more will be added in the near future. Further-
more, given the standing and fallen dead trees, sap-
lings and lianas, fuel continuity will be greatly
enhanced from the forest floor up to the canopy.
Combined with another prolonged dry season, these
factors could increase the risk of a further fire in these
forests. The resulting second burn is likely to be much
more severe than the first, and Cochrane et al. (1999)
expect up to 98% of all remaining trees to be suscep-
tible to recurrent fires. Moreover, pioneer saplings
may be no more likely to survive a second burn than
non-pioneers (Cochrane and Schulze, 1999) meaning
that if fire return rates are frequent enough, new
regeneration will effectively be prevented from reach-
ing maturity and these forest ecosystems will shift
towards scrub-savannahs where arborescent plants
will be far less prevalent. Indeed, our experience in
forest areas that local people report to have burnt twice
support these predictions, as they are heavily domi-
nated by bamboo (Guadua spp.), with very few stand-
ing trees remaining (J. Barlow, unpublished data).
Without drastic fire prevention measures aimed at
excluding fire from local agricultural systems, it seems
unlikely that these forests will ever recover in time to
prevent a subsequent fire. Many years of regrowth are
necessary for BFs to recover the fire resistance of
primary forests (Cochrane and Schulze, 1998) because
tall trees are needed to establish the full shade and
moist microclimate typical of the primary forest inter-
ior. Selectively logged forests have been found to take
8 T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11

5–6 years (Mason, 1996) and 7–12 years (Johns, 1989)
to return to approximately pre-disturbance conditions
and we could expect a similar or greater time-span
after fire disturbance. Therefore, the evidence that El
Nino events are increasing in frequency and severity in
recent years (Trenberth and Hoar, 1996; Timmermann
et al., 1999) does not bode well for the future of these
forests.
4.5. Carbon loss
Substantial carbon loss is derived from surface
wildfires through the long-term decomposition of
unburnt dead biomass and the initial combustion of
dead leaves, twigs and branches on the forest floor,
with fine branches and the leaf litter making up as
much as 70% of all fuel on the forest floor (Chambers
et al., 2000). However, the small litter and rootmat on
the forest floor comprise only around 3.7–8.0% of the
aboveground biomass (Kauffman et al., 1995) and
saplings and small trees (<10 cm DBH) account for
only 6.2% of the total AGDB resulting from a surface
fire in our study region (Peres, 1999). We therefore
emphasise the potential (or committed) carbon loss
from the large tree mortality (�10 cm DBH), which is
likely to be far more important to the overall residual
fuel load. In the absence of a subsequent fire, carbon
will be released from the standing and fallen dead
wood through bacterial decomposition and termite
activity, which occurs largely over the first decade
following death (Melillo et al., 1996; Fearnside,
1997a). Although there was an almost 8-fold increase
in tree mortality in the BF plots, the size-dependent
mortality (which selected for smaller trees; see Fig. 2b)
meant that this only resulted in a 3.5-fold increase in
the amount of dead biomass and potential carbon loss.
Our AGDB estimates closely resemble those found by
Cochrane and Schulze (1999) in eastern Amazonia,
but far exceeds those from other studies such as
Barbosa and Fearnside (1999) and Santos et al.
(1998; Table 2).
Using the Santos equation the potential loss of an
additional 25.5 t C/ha in BF (above the background
level of 10.2 t C/ha in unburnt controls) dramatically
increases the amount of carbon released from Ama-
zonian forests. Deforestation activities in the Brazilian
Amazon emit approximately 250�350 � 106 t C
annually (Fearnside, 1999), or 4–5.5% of the annual
global flux of carbon to the atmosphere caused by
human activities, which in 1998 was an estimated
6318 � 106 t C (Worldwatch News Brief, 1999).
However, if our results (based on short-term estimates
of tree mortality) are extrapolated to the rest of the
Brazilian Amazon, the burning of all the primary
forests that were prone to surface wildfires after the
1998 dry season (270,000 km2; cf. Nepstad et al.,
1999b) would commit 6885 � 106 t C to be released,
supporting the assertion of Nepstad et al. (1999b) that
committed carbon estimates from the Brazilian Ama-
zon could be doubled during strong El Nino years such
as 1998. Furthermore, fire degrades both the forest
structure and species composition and will presum-
ably disrupt the important role Amazonian forest play
as a sink for global carbon emissions (Chambers et al.,
2001).
However, caution should be applied in making wide
extrapolations across Amazonia from a restricted net-
work of sampling sites. Soils in the study area were
characterised by sand fractions, and therefore may
support a lower standing biomass than other Amazo-
nian forests on clay soils (Laurance et al., 1999).
However, the lower standing biomass may be offset
by the increased root biomass for the acquisition of
soil nutrients (Wilson and Tilman, 1991), an important
point as total root and soil carbon represent almost half
the total carbon content in South American tropical
forests (Dixon et al., 1994), and little is known about
belowground carbon loss. Furthermore, much of the
forests at risk of burning will also be on sandy soils
because of their poor water retention capacity (Ham-
mond and ter Steege, 1998).
5. Conclusion
In brief, perhaps the most important ecological
effect of surface fires is that they increase the like-
lihood that fire will become a permanent feature of the
forest ecosystem, because of decreasing intervals
between consecutive El Nino events and the long
process of regrowth which is necessary for secondary
forest to recover the fire resistance of primary forests.
Consequently, the fires disrupt what is a well-balanced
carbon cycle between earth and atmosphere (Pielke
et al., 1998) as closed-canopy forests hold more
carbon per unit area in vegetation and soil than any
T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11 9

of the ecosystems replacing them (Melillo et al.,
1996).
Fearnside (1999) acknowledges that the avoidance
of ‘natural’ disasters represents a major factor in the
carbon balance of tropical forests, and one that should
be addressed in global warming mitigation strategies,
because the vast areas involved ensures that carbon
emissions are substantial. However, surface fires in
neotropical forests are often portrayed as fairly innoc-
uous, with low-impact fires resulting in a relatively
small percentage of dead trees (e.g. Fearnside, 1999).
In this paper we have shown that 36% of all midstorey
and canopy trees may die as a result of these fires,
representing a loss of an extra 21.6 t C/ha. Surface
fires are of huge global concern both in terms of
biodiversity conservation and ecosystem services,
and should be taken more seriously than ever before
because of the vast areas that will become prone to
fires at the end of future El Nino dry seasons.
Acknowledgements
This study was funded by the Centre for Applied
Biodiversity Science (CABS) of Conservation Inter-
national and the Josephine Bay and Michael Paul
Foundation. Jos Barlow’s fieldwork was funded by
a NERC Ph.D. studentship at the University of East
Anglia. We are especially thankful to the political
leadership of the Reserva Extrativista do Tapajos-
Arapiuns and the villagers of Cachoeira do Maro,
Sao Jose and Porto Rico for allowing us to conduct
this study, and to the local assistance of Nan, Arnei and
Torozinho. Both Rionaldo and Edith de Santos pro-
vided enormous logistical assistance during all stages
of the project.
References
Araujo, T.M., Higuchi, N., Junior, J.A.D., 1997. Comparacao de
Metodos para Determinar Biomassa na Regiao Amazonica.
Anais da Academia Brasileira de Ciencias 68 (1).
Barbosa, R.I., Fearnside, P.M., 1999. Incendios na Amazonia
Brasileira: estimativa da emissao de gases do efeito estufa pela
queima de diferentes ecossistemas de Roraima na passagem do
evento ‘‘El Nino’’ (1997/1998). Acta Amazonica 29, 513–534.
Barlow, J., Haugaasen, T., Peres, C.A., 2002. Effects of ground fires
on understorey bird assemblages in Amazonian forests. Biol.
Conserv. 105, 157–169.
Barlow, J., Lagan, B.O., Peres, C.A., in press. Morphological
correlates of fire-induced tree mortality in a central Amazonian
forest. J. Trop. Ecol.
Brown, S., 1997. Estimating biomass and biomass change of
tropical forests. FAO Forestry Paper 134.
Chambers, J.Q., Higuchi, N., Schimel, J.P., Ferreira, L.V., Melack,
J.M., 2000. Decomposition and carbon cycling of dead trees in
tropical forests of the central Amazon. Oecologia 122, 380–
388.
Chambers, J.Q., Higuchi, N., Tribuzi, E.S., Trumbore, S.E., 2001.
Carbon sink for a century. Nature 410, 429.
Cincotta, R.P., Wisnewski, J., Engelman, R., 2000. Human
population in the biodiversity hotspots. Nature 404, 990–992.
Cochrane, M.A., 2001. In the line of fire—understanding the
impacts of tropical forest fires. Environment 43, 28–38.
Cochrane, M.A., Schulze, M.D., 1998. Forest fires in the Brazilian
Amazon. Conserv. Biol. 12, 948–950.
Cochrane, M.A., Schulze, M.D., 1999. Fire as a recurrent event in
tropical forests of the eastern Amazon: effects on forest
structure, biomass, and species composition. Biotropica 31, 2–
16.
Cochrane, M.A., Alencar, A., Schulze, M.D., Souza, C.M.,
Nepstad, D.C., Lefebvre, P., Davidson, E.A., 1999. Positive
feedbacks in the fire dynamic of closed canopy tropical forests.
Science 284, 1832–1835.
Dale, V.H., Pearson, S.M., 1997. Quantifying habitat fragmentation
due to land-use change in Amazonia. In: Laurance, W.F.,
Bierregaard, R.O. (Eds.), Tropical Forest Remnants: Ecology,
Management, and Conservation of Fragmented Communities.
University of Chicago Press, Chicago, pp. 400–409.
Dixon, R.K., Brown, S., Houghton, R.A., Solomon, A.M., Trexler,
M.C., Wisniewski, J., 1994. Carbon pools and flux of global
forest ecosystems. Science 263, 185–190.
Fearnside, P.M., 1997a. Greenhouse gases from deforestation in
Brazilian Amazonia: net committed emissions. Clim. Change
35, 321–360.
Fearnside, P.M., 1997b. Wood density for estimating forest biomass
in Brazilian Amazonia. For. Ecol. Manage. 90, 59–87.
Fearnside, P.M., 1999. Forests and global warming mitigation in
Brazil: opportunities in the Brazilian forest sector for responses
to global warming under the clean development mechanism.
Biomass Bioenergy 16, 171–189.
Goldhammer, J.G., 1999. Forests on fire. Science 284, 1782–
1783.
Hammond, D.S., ter Steege, H., 1998. Propensity of fire in Guianan
rainforests. Conserv. Biol. 12, 944–947.
Haugaasen, T., 2000. Effects of ground fires on understorey birds in
central Amazonia, Brazil. M.Sc. Thesis. University of East
Anglia, Norwich.
Holdsworth, A.R., Uhl, C., 1997. Fire in Amazonian selectively
logged rain forest and the potential for fire reduction. Ecol.
Appl. 7, 713–725.
INFRAERO, 1998. Infraero rainfall data. Superintendencia da
Infraero, Sala AIS/SBSN, Santarem Airport, Santarem.
Johns, A.D., 1989. Recovery of a peninsular Malaysian rainforest
avifauna following selective timber logging: the first twelve
years. Forktail 4, 89–105.
10 T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11

Kauffman, J.B., Cummings, D.L., Ward, D.E., Babbitt, R., 1995.
Fire in the Brazilian Amazon. 1. Biomass, nutrient pools, and
losses in slashed primary forests. Oecologia 104, 397–408.
Kinnaird, M.F., O’Brien, T.G., 1998. Ecological effects of wildfire
on lowland rainforest in Sumatra. Conserv. Biol. 12, 954–956.
Laurance, W.F., Laurance, S.G., Delamonica, P., 1998. Tropical
forest fragmentation and greenhouse gas emissions. For. Ecol.
Manage. 110, 173–180.
Laurance, W.F., Fearnside, P.M., Laurance, S.G., Delamonica, P.,
Lovejoy, T.E., Rankin-de Merona, J., Chambers, J.Q., Gascon,
C., 1999. Relationship between soils and Amazon forest biomass:
a landscape-scale study. For. Ecol. Manage. 118, 127–138.
Lemmon, P.E., 1957. A new instrument for measuring forest
overstory density. J. For. 55, 667–668.
Mason, D., 1996. Responses of Venezuelan understorey birds to
selective logging, enrichment strips and vine cutting. Biotro-
pica 28, 296–309.
Melillo, J.M., Houghton, R.A., Kicklighter, D.W., McGuire, A.D.,
1996. Tropical deforestation and the global carbon budget.
Ann. Rev. Energy Environ. 21, 293–310.
Mesquita, R.C.G., Delamonica, P., Laurance, W.F., 1999. Effect of
surrounding vegetation on edge-related tree mortality in
Amazonian forest fragments. Biol. Conserv. 91, 129–134.
Nepstad, D.C., Uhl, C., Serrao, E.A.S., 1991. Recuperation of a
degraded Amazonian landscape—forest recovery and agricul-
tural restoration. Ambio 20, 248–255.
Nepstad, D.C., Moreira, A.G., Alencar, A.A., 1999a. Flames in the
Rain Forest: Origins, Impacts and Alternatives to Amazonian
Fires. The Pilot Program to Conserve the Brazilian Rain Forest,
Brasilia, Brazil.
Nepstad, D.C., Verıssimo, A., Alencar, A., Nobre, C., Lima, E.,
Lefebvre, P., Schlesinger, P., Potter, C., Moutinho, P., Mendoza,
E., Cochrane, M., Brooks, V., 1999b. Large-scale impoverish-
ment of Amazonian forests by logging and fire. Nature 398,
505–508.
Pahari, K., Murai, S., 1999. Modelling for prediction of global
deforestation based on the growth of human population. ISPRS
J. Photogramm. Remote Sens. 54, 317–324.
Peres, C.A., 1999. Ground fires as agents of mortality in a central
Amazonian forest. J. Trop. Ecol. 15, 535–541.
Peres, C.A., 2000. Effects of subsistence hunting on vertebrate
community structure in Amazonian forests. Conserv. Biol. 14,
240–253.
Peres, C.A., 2001. Synergistic effects of subsistence hunting and
habitat fragmentation on Amazonian forest vertebrates. Con-
serv. Biol. 15, 1490–1505.
Peres, C.A., Barlow, J., Haugaasen, T., in press. Vertebrate
assemblage responses to wildfire disturbance in a central
Amazonian forest. Oryx.
Pielke, R.A., Avissar, R., Raupach, M., Dolman, A.J., Zeng, X.,
Denning, A.S., 1998. Interactions between the atmosphere and
terrestrial ecosystems: influence on weather and climate.
Global Change Biol. 4, 461–475.
Putz, F.E., Blate, G.M., Redford, K.H., Fimbel, R., Robinson, J.,
2001. Tropical forest management and conservation of
biodiversity: an overview. Conserv. Biol. 15, 7–20.
Robinson, J.G., Bennett, E.L. (Eds.), 2000. Hunting for Sustain-
ability in Tropical Forests. Columbia University Press, New
York.
Sanford, R.L., Saldarriaga, J., Clark, K.E., Uhl, C., Herrera, R.,
1985. Amazon rain-forest fires. Science 227, 53–55.
Santos, J., 1996. Analise de modelos de regressao para estimar a
fitomassa de floresta tropical umida de terra-firme da Amazonia
Central. Ph.D. Thesis. Universidade Federal de Vicosa, Minas
Gerais, Brazil.
Santos, J.R., Pardi Lacruz, M.S., Araujo, L.S., Xaud, H.A.M.,
1998. El proceso de queima de biomassa de bosque tropical y
de sabanas en la Amazonia Brasileira: experiencias de
monitioreo com dados opticos y de microondas. Revista Serie
Geografica 7, 97–108.
Skole, D., Tucker, C., 1993. Tropical deforestation and habitat
fragmentation in the Amazon: satellite data from 1978–1988.
Science 260, 1905–1910.
Sunarto, S., 2000. Survival of plant communities and dynamics of
forest ecosystems in response to fire: a long-term study from
Sumatra. M.Sc. Thesis. University of East Anglia, Norwich.
Timmermann, A., Oberhuber, J., Bacher, A., Esch, M., Latif, M.,
Roeckner, E., 1999. Increased El Nino frequency in a climate
model forced by future greenhouse warming. Nature 398, 694–
697.
Trenberth, K.E., Hoar, T.J., 1996. The 1990–1995 El Nino-southern
oscillation event: longest on record. Geophys. Res. Lett. 23,
57–60.
Turcq, B., Sifeddine, A., Martin, L., Absy, M.L., Soubies, F.,
Suguio, K., Volkmer-Ribeiro, C., 1998. Amazonia rainforest
fires: a lacustrine record of 7000 years. Ambio 27, 139–142.
Uhl, C., Buschbacher, R., 1985. A disturbing synergism between
cattle ranch burning practices and selective tree harvesting in
the eastern Amazon. Biotropica 14, 265–268.
Uhl, C., Kauffman, J.B., 1990. Deforestation, fire susceptibility,
and potential tree responses to fire in the eastern Amazon.
Ecology 71, 37–449.
Whitmore, T.C., 1997. Tropical forest disturbance, disappearance,
and species loss. In: Laurance, W.F., Bierregaard, R.O. (Eds.),
Tropical Forest Remnants: Ecology, Management, and Con-
servation of Fragmented Communities. University of Chicago
Press, Chicago, pp. 3–12.
Wilson, S.D., Tilman, D., 1991. Components of plant competitions
along an experimental gradient of nitrogen availability. Ecology
72, 1050–1065.
Worldwatch News Brief, 1999. World carbon emissions fall. http://
www.worldwatch.org/alerts/990727.html.
T. Haugaasen et al. / Forest Ecology and Management 6154 (2002) 1–11 11
Copyright © 2022 FDOKUMEN




















