Surface protein patterns govern morphology, proliferation, and expression of cellular markers but...
13
Surface Protein Patterns Govern Morphology, Proliferation, and Expression of Cellular Markers but Have No Effect on Physiological Properties of Cortical Precursor Cells Anna K. Magnusson, 1 Pontus Linderholm, 1,2 Christian Vieider, 2 Mats Ulfendahl, 1 and Anna Erlandsson 1 * 1 Center for Hearing and Communication Research, Department of Clinical Neuroscience, Karolinska Institutet, Stockholm, Sweden 2 Acreo AB, Electrum 236, Kista, Sweden The ability to differentiate and give rise to neurons, astrocytes, and oligodendrocytes is an inherent feature of neural stem cells, which raises hopes for cell-based therapies of neurodegenerative diseases. However, there are many hurdles to cross before such regimens can be applied clinically. A considerable challenge is to elucidate the factors that contribute to neural differen- tiation. In this study, we evaluated the possibility of steering neuronal maturation by growing cortical pre- cursor cells on microscale surface patterns of extracel- lular matrix (ECM) proteins. When the cells were encouraged to extend processes along lines of ECM proteins, they displayed a much more mature morphol- ogy, less proliferation capacity, and greater expression of a neuronal marker in comparison with cells grown in clusters on ECM dots. This implied that the growth pat- tern alone could play a crucial role for neural differen- tiation. However, in spite of the strikingly different mor- phology, when performing whole-cell patch-clamp experiments, we never observed any differences in the functional properties between cells grown on the two patterns. These results clearly demonstrate that mor- phological appearances are not representative meas- ures of the functional phenotype or grade of neuronal maturation, stressing the importance of complementary electrophysiological evidence. To develop successful transplantation therapies, increased cell survival is criti- cal. Because process-bearing neurons are sensitive and break easily, it would be of clinical interest to explore further the differentiating capacity of the cells cultured on the ECM dot pattern, described in this arti- cle, which are devoid of processes but display the same functional properties as neurons with mature morphology. V V C 2008 Wiley-Liss, Inc. Key words: stem cells; neural differentiation; extra- cellular matrix molecules; electrophysiology Neural stem/precursor cells have many properties that make them excellent candidates for cell-based treat- ments of injuries in the central nervous system. How- ever, a number of important questions regarding the control of neural differentiation remain to be answered before clinical trials can be initiated. To date, no method has been presented that satisfactorily controls differentia- tion of neural precursor cells into neurons, astrocytes, and oligodendrocytes. Because this is crucial for safe and effective transplantations, improved understanding of all parameters that regulate neural differentiation is of high importance. The presence of various extrinsic factors, such as growth factors or genetic manipulations, have been shown to affect the differentiation potential to some extent (McKay, 1997; Gage, 2000; Glaser et al., 2005; Erlandsson et al., 2006). For example, ciliary neu- rotrophic factor (CNTF) and leukemia inhibitory factor (LIF) shift the differentiation to an astrocytic fate (Johe et al., 1996; Bonni et al., 1997) whereas addition of trii- dothyronine (T3) or insulin-like growth factor 1 (IGF-1) Contract grant sponsor: European Commission (FP6 Integrated Project EUROHEAR); Contract grant number: LSHG-CT-20054-512063; Contract grant sponsor: Swedish Foundation for Strategic Research (BioX program); Contract grant sponsor: Swedish Research Council; Contract grant sponsor: Tysta Skolan; Contract grant sponsor: Petrus and Augusta Hedlund Foundation; Contract grant sponsor: Swedish Society for Medical Research; Contract grant sponsor: A ˚ ke Wiberg Foundation; Contract grant sponsor: Magnus Bergvall Foundation. *Correspondence to: Dr. Anna Erlandsson, Center for Hearing and Communication Research, M1:00, Karolinska University Hospital, Solna, SE-171 76 Stockholm, Sweden. E-mail: [email protected] Received 24 October 2007; Revised 13 December 2007; Accepted 21 January 2008 Published online 1 April 2008 in Wiley InterScience (www. interscience.wiley.com). DOI: 10.1002/jnr.21688 Journal of Neuroscience Research 86:2363–2375 (2008) ' 2008 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Surface protein patterns govern morphology, proliferation, and expression of cellular markers but...

Surface Protein Patterns GovernMorphology, Proliferation, and Expressionof Cellular Markers but Have No Effect onPhysiological Properties of CorticalPrecursor Cells
Anna K. Magnusson,1 Pontus Linderholm,1,2 Christian Vieider,2 Mats Ulfendahl,1
and Anna Erlandsson1*1Center for Hearing and Communication Research, Department of Clinical Neuroscience, KarolinskaInstitutet, Stockholm, Sweden2Acreo AB, Electrum 236, Kista, Sweden
The ability to differentiate and give rise to neurons,astrocytes, and oligodendrocytes is an inherent featureof neural stem cells, which raises hopes for cell-basedtherapies of neurodegenerative diseases. However,there are many hurdles to cross before such regimenscan be applied clinically. A considerable challenge is toelucidate the factors that contribute to neural differen-tiation. In this study, we evaluated the possibility ofsteering neuronal maturation by growing cortical pre-cursor cells on microscale surface patterns of extracel-lular matrix (ECM) proteins. When the cells wereencouraged to extend processes along lines of ECMproteins, they displayed a much more mature morphol-ogy, less proliferation capacity, and greater expressionof a neuronal marker in comparison with cells grown inclusters on ECM dots. This implied that the growth pat-tern alone could play a crucial role for neural differen-tiation. However, in spite of the strikingly different mor-phology, when performing whole-cell patch-clampexperiments, we never observed any differences in thefunctional properties between cells grown on the twopatterns. These results clearly demonstrate that mor-phological appearances are not representative meas-ures of the functional phenotype or grade of neuronalmaturation, stressing the importance of complementaryelectrophysiological evidence. To develop successfultransplantation therapies, increased cell survival is criti-cal. Because process-bearing neurons are sensitiveand break easily, it would be of clinical interest toexplore further the differentiating capacity of the cellscultured on the ECM dot pattern, described in this arti-cle, which are devoid of processes but display thesame functional properties as neurons with maturemorphology. VVC 2008 Wiley-Liss, Inc.
Key words: stem cells; neural differentiation; extra-cellular matrix molecules; electrophysiology
Neural stem/precursor cells have many propertiesthat make them excellent candidates for cell-based treat-ments of injuries in the central nervous system. How-ever, a number of important questions regarding thecontrol of neural differentiation remain to be answeredbefore clinical trials can be initiated. To date, no methodhas been presented that satisfactorily controls differentia-tion of neural precursor cells into neurons, astrocytes,and oligodendrocytes. Because this is crucial for safe andeffective transplantations, improved understanding of allparameters that regulate neural differentiation is of highimportance. The presence of various extrinsic factors,such as growth factors or genetic manipulations, havebeen shown to affect the differentiation potential tosome extent (McKay, 1997; Gage, 2000; Glaser et al.,2005; Erlandsson et al., 2006). For example, ciliary neu-rotrophic factor (CNTF) and leukemia inhibitory factor(LIF) shift the differentiation to an astrocytic fate (Joheet al., 1996; Bonni et al., 1997) whereas addition of trii-dothyronine (T3) or insulin-like growth factor 1 (IGF-1)
Contract grant sponsor: European Commission (FP6 Integrated Project
EUROHEAR); Contract grant number: LSHG-CT-20054-512063;
Contract grant sponsor: Swedish Foundation for Strategic Research
(BioX program); Contract grant sponsor: Swedish Research Council;
Contract grant sponsor: Tysta Skolan; Contract grant sponsor: Petrus and
Augusta Hedlund Foundation; Contract grant sponsor: Swedish Society
for Medical Research; Contract grant sponsor: Ake Wiberg Foundation;
Contract grant sponsor: Magnus Bergvall Foundation.
*Correspondence to: Dr. Anna Erlandsson, Center for Hearing and
Communication Research, M1:00, Karolinska University Hospital, Solna,
SE-171 76 Stockholm, Sweden. E-mail: [email protected]
Received 24 October 2007; Revised 13 December 2007; Accepted 21
January 2008
Published online 1 April 2008 in Wiley InterScience (www.
interscience.wiley.com). DOI: 10.1002/jnr.21688
Journal of Neuroscience Research 86:2363–2375 (2008)
' 2008 Wiley-Liss, Inc.

increases the number of oligodendocytes in cultures(Johe et al., 1996; Glaser et al., 2005). Neuron-specificinduction is more difficult to achieve, and, at present,no single efficient soluble factor has been identified. Forexample, platelet-derived growth factor (PDGF) was ear-lier suggested to be involved in neuronal differentiationbut has more recently been shown to maintain the pro-liferation of precursor cells with an immature morphol-ogy (Erlandsson et al., 2001, 2006). It has been demon-strated that other extrinsic factors, such as the stiffness ofthe surface or the presence of different extracellular ma-trix (ECM) proteins, are also involved in the regulationof neural differentiation (Engler et al., 2004; Discheret al., 2005; Georges and Janmey, 2005; Yeung et al.,2005). It is, for instance, clear from many studies thatsignal transduction by matrix molecules through integ-rins plays an important role in neural differentiation(Campos, 2005; Leone et al., 2005).
Studies of three-dimensional scaffolds and matricesindicate that a 3D network of ECM proteins has a posi-tive effect on neuronal differentiation (Silva et al., 2004;Ma et al., 2004). Various molecules have been success-fully used for the patterning of cell culture surfaces,including synthetic molecules (e.g., poly-lysine) andphysiological adhesion molecules such as laminin, as wellas ECM gels, which are a mixture of many differentECM proteins. Most studies conducted to date havefocused on cell attachment and axonal outgrowth. Forexample, adhesion and outgrowth of hippocampal neu-rons has been shown to be sensitive to poly-lysine pat-terns, and micropatterns of laminin create a highly or-dered Schwann cell matrix (Wyart et al., 2002; Schma-lenberg and Uhrich, 2005).
The aim of the present study was to investigate therole of cell morphology in the regulation of neural dif-ferentiation, a parameter that to our knowledge hasnever been studied in this context. Cortical precursorcells were cultured on ECM protein patterns, by micro-contact printing applied to polystyrene substrates, a tech-nique that has previously been successfully used for stud-ies of neuronal networks (Vogt et al., 2003, 2005a). Thepatterns were designed so that the cells either were re-stricted to 20 lm dots or had the possibility of formingprocesses along lines connecting the dots (see Fig. 1).The morphology of a cell is traditionally thought to be asecondary result of other cellular processes, such as cellsignaling and differentiation. We asked whether it canalso be the other way around, so that neural precursorcells that are grown on a surface with a microscale pat-tern that allows cell spreading develop more mature fea-tures than cells grown on a pattern that prevents out-growth of neural processes.
MATERIALS AND METHODS
Microcontact Printing
Microcontact printing stamps were produced from poly-dimethylsiloxane (PDMS) by photolithography and molding.
The mask design was carried out in a CAD program, and thepattern was transferred to a chromium mask. The master wasmade by irradiating a thin layer of the photoresist, SU8,through the mask (making the irradiated areas chemicallyharder then the unexposed areas), and the unexposed areaswere dissolved in a developer. After hardening and antisticktreatment, the PDMS prepolymer was poured onto the masterand cured. Stamps with two different types of pattern werefabricated; 20-lm-wide squares with a distance of 80 lm(dots) or the same squares connected with 3-lm-wide lines(dots 1 lines). The stamps were thoroughly washed toremove uncured PDMS and were made hydrophilic by a briefexposure to oxygen plasma. Until use, the stamps were storedin ethanol to prevent hydrophobic recovery. Unoxidized pol-ystyrene dishes (60 3 15 mm; Falcon) or glass-bottom micro-well dishes (for the electrophysiology experiments; MatTekInc.) were patterned with extracellular matrix gel (ECM gel;Sigma, St. Louis, MO), diluted 1:10 in DMEM and supple-mented with 10 lg/ml poly-D-lysine (Sigma; Fig. 1A). Drystamps were placed in the ECM ink for 30 sec and rinsedtwice in PBS and twice in H2O. The stamps were left to dryat room temperature with the inked side facing up for 20–30min. To receive optimal moisture, the stamps were held at48C for 10 min and then immediately pressed to the substratefor 1 min. After removal of the stamps, the dishes were rinsedonce in DMEM before the cells were plated on the surface.
Cell Culture
The experiments were performed in conformity withEC Council Directive 86/89/ECC and were authorized bythe local Animal Care and Use Committee in agreement withSwedish law. Embryonic cortex was dissected in Hanks’ bal-anced salt solution (HBSS; Invitrogen, Carlsbad, CA) fromtimed-pregnant C57/BL6 mice (B&K) on embryonic day 14(E14; E1 was defined as the day of copulatory plug). The tis-sue was gently triturated using a 1-ml pipette, and larger cellclumps were allowed to sediment for 10 min, after which thecell suspension was centrifuged. The pellet was resuspended inDMEM-glutamax (Invitrogen) supplemented with B27 (1:50;Invitrogen), Hepes (Invitrogen), penicillin/streptomycin (Invi-trogen), 10 ng/ml fibroblast growth factor-2 (FGF2; Pepro-Tech), and 20 ng/ml epidermal growth factor (EGF; BD, SanJose, CA). The cortical cells were plated at a cell density of10–20 cells/ll and were expanded in suspension as neuro-spheres for 11–12 days prior to the experiments. At day 4 andday 8 after dissection, the neurospheres were passaged. Thespheres were then centrifuged and carefully resuspended inHBSS. After a 15-min incubation at 378C the neurosphereswere centrifuged again and thoroughly resuspended in culturemedia using a Pasteur pipette, approximately 50 times, until asingle-cell suspension was received. The cells were used forexperiments 3–4 days after the second passage. Neurosphereswere treated, as described above, to become single cells, andthe cells were plated on patterned surfaces at a concentrationof 0.125 million cells/ml (4 ml/60-mm dish) in cell culturemedia without the mitogenes FGF2 and EGF. On the day af-ter plating, the medium was changed to remove cells that hadnot attached to the surface.
2364 Magnusson et al.
Journal of Neuroscience Research
Immunocytochemistry
Differentiating cortical precursor cells cultured on dot ordot 1 lines patterns were fixed in 4% paraformaldehyde(PFA) at day 1, 4, or 6 after plating. Cells were permeabilizedwith 0.2% Triton X-100 in PBS (this step was excluded forthe O4 staining) and incubated in blocking solution with 5%normal goat serum (Dako, Carpinteria, CA) in PBS for 1 hr
at room temperature. Immunostainings were performed using
the primary antibodies: mouse nestin (1:50; Chemicon, Teme-
cula, CA), mouse bIII-tubulin (Covance, Berkeley, CA;
1:200), rabbit glial fibrillary acidic protein (GFAP; Dako;
1:400), mouse IgM O4 (1:50; Chemicon), rabbit laminin
(Sigma; 1:100), mouse laminin (Sigma; 1:100), and the sec-
ondary antibodies Alexa Flour 488 and Alexa Fluor 546
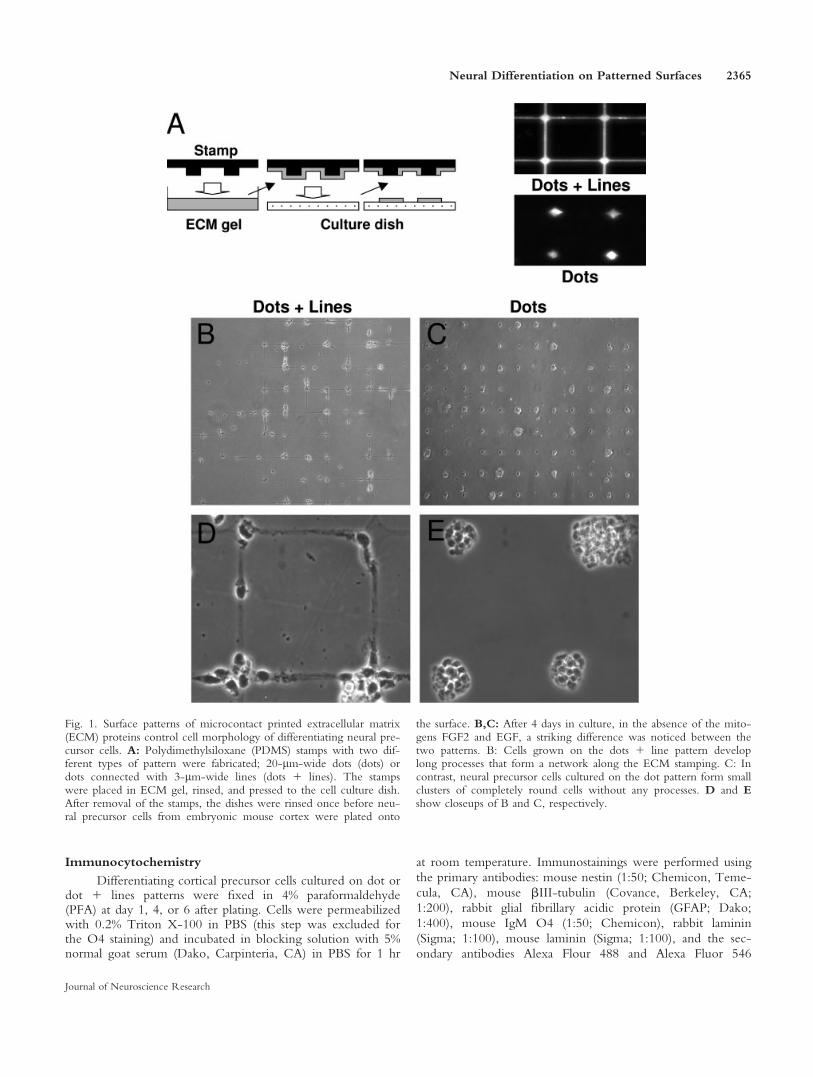
Fig. 1. Surface patterns of microcontact printed extracellular matrix(ECM) proteins control cell morphology of differentiating neural pre-cursor cells. A: Polydimethylsiloxane (PDMS) stamps with two dif-ferent types of pattern were fabricated; 20-lm-wide dots (dots) ordots connected with 3-lm-wide lines (dots 1 lines). The stampswere placed in ECM gel, rinsed, and pressed to the cell culture dish.After removal of the stamps, the dishes were rinsed once before neu-ral precursor cells from embryonic mouse cortex were plated onto
the surface. B,C: After 4 days in culture, in the absence of the mito-gens FGF2 and EGF, a striking difference was noticed between thetwo patterns. B: Cells grown on the dots 1 line pattern developlong processes that form a network along the ECM stamping. C: Incontrast, neural precursor cells cultured on the dot pattern form smallclusters of completely round cells without any processes. D and Eshow closeups of B and C, respectively.
Neural Differentiation on Patterned Surfaces 2365
Journal of Neuroscience Research

against mouse IgG, mouse IgM or rabbit IgG (Invitrogen;1:200). Incubation was performed at 48C overnight with pri-mary antibodies and at room temperature for 1 hr with sec-ondary antibodies. The cultures were rinsed three times inphosphate-buffered saline (PBS) after the antibody incuba-tions. Cell nuclei were labeled with 40-6-diamidino-2-phenyl-indole (DAPI) for 10 min (Molecular Probes, Eugene, OR).
Proliferation Assay
For the bromodeoxyuridine (BrdU) labeling assay, cellswere pulsed with 10 mM BrdU (Amersham, ArlingtonHeights, IL) for 20 hr before fixation in ice-cold acid ethanol(90% ethanol, 5% acetic acid). Incorporation of BrdU wasdetected by using monoclonal anti-BrdU antibodies (Amer-sham), followed by Alexa Fluor 546 anti-mouse IgG second-ary antibodies (Invitrogen; 1:200), as described above.
Cell Counting
After the immunostainings, the number of cells express-ing BrdU, nestin, bIII-tubulin, GFAP, or O4 and the totalnumber of DAPI-labeled cells were counted in eight different200-lm 3 200-lm fields/culture dishes. The cells werecounted in two dishes per experiment, and each experimentwas performed in triplicate. The number of positive cells ispresented as the percentage of the total number of cells, andthe values are expressed as means 6 SD in the text and asmeans 6 95% confidence intervals (c.i.) in the figures. Stu-dent’s unpaired t-test was used for evaluation of statistical sig-nificance (*P < 0.05, **P < 0.01, ***P < 0.001).
Electrophysiology
For the electrophysiological experiments, the cells wereplated on glass-bottom microwell dishes (MatTek Inc.)according to procedures described above. Prior to the experi-ments, the cell culture media was exchanged for artificial cere-brospinal fluid (aCSF) containing (in mM): 125 NaCl, 2.5KCl, 1.25 NaH2PO4, 26 NaHCO3, 25 glucose, 2 CaCl2, and1 MgCl2. The aCSF was bubbled with carbogen gas (95%O225% CO2) to generate a pH of 7.4.
The cortical precursor cells were viewed with aninverted microscope (Zeiss Axiovert, Jena, Germany) using a340 oil-immersion objective (Neofluar; Zeiss), a 32 Optovar(Zeiss), and infrared-differential interference optics equippedwith an infrared sensitive digital camera (C3077; Hamamatsu,Hamamatsu, Japan). Whole-cell current and voltage-clamprecordings were made at room temperature with a Multiclamp700B amplifier (Molecular Devices, Sunnyvale, CA). Borosili-cate glass microelectrodes (GC150F-10; Harvard Apparatus,Cambridge, United Kingdom) were pulled on a vertical elec-trode puller (PP-830; Narashige, Tokyo, Japan) yielding afinal tip resistance of 5–9 MX. The internal pipette solutioncontained (in mM): 130 K-gluconate, 5 KCl, 10 HEPES, 1EGTA, 2 Na2-ATP, 2 Mg-ATP, 0.3 Na3-GTP, 10 Na2-phosphocreatinine, adjusted to pH 7.3 with KOH.
The bridge balance was applied for current-clamprecordings. For voltage-clamp recordings, the series resistance(dots 1 lines: 28.8 6 10.0 MX, n 5 19; dots: 29.5 6 12.2MX, n 5 22; P 5 0.84) was compensated by 70–80% and
monitored throughout the experiment. The voltages have notbeen corrected for the junction potential.
The signals were filtered with a low-pass four-pole Bes-sel filter at 10 kHz, sampled at 20 kHz, and digitized with aDigidata 1440A interface (Molecular Devices). Stimulus gen-eration, data acquisition, and off-line analysis of data wereperformed in pClamp software (version 10.0; MolecularDevices).
The input resistance (Rin) and the membrane time con-stant (smembrane) were estimated from the voltage responses tosmall current injections (610 pA) around rest by, respectively,calculating the slope of current-voltage plot or fitting a singleexponential function. The membrane capactitance (Cm) wasderived from the formula: s 5 R 3 C.
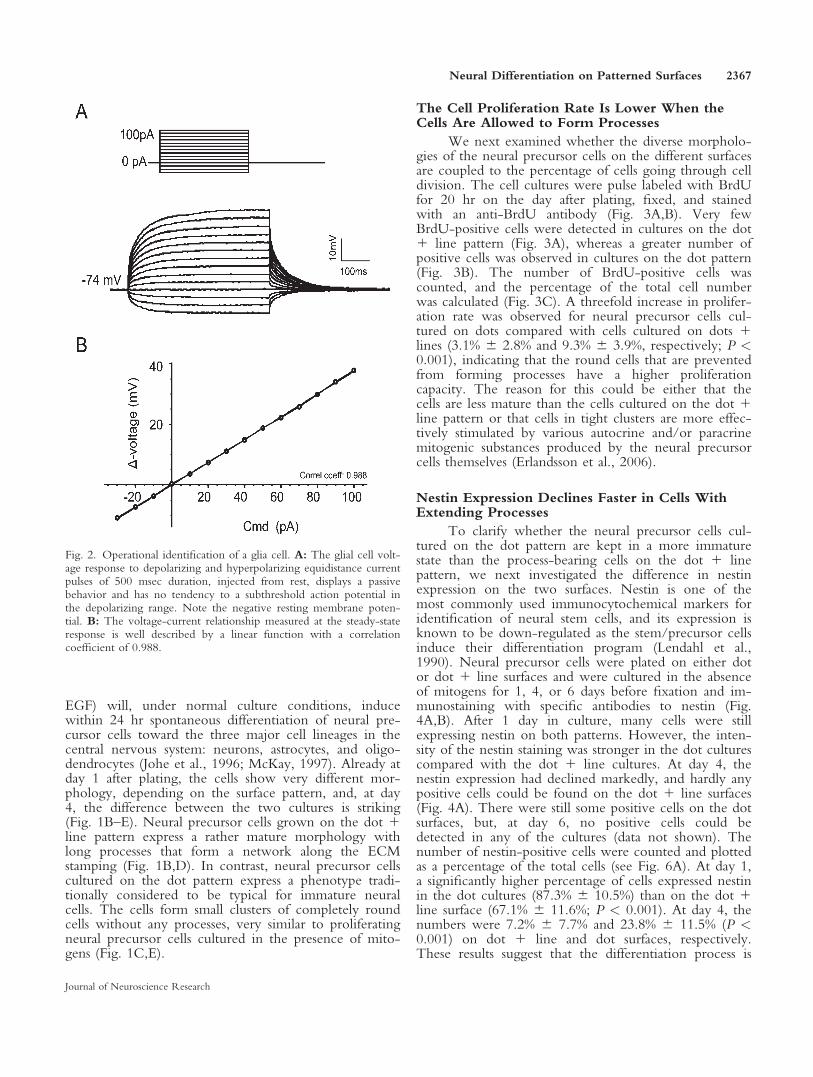
These precursor cells can give rise to both neurons andglia cells, so the latter cell type was naturally encountered dur-ing the experiments. These cells were, however, easy to dis-tinguish from ‘‘neuron-like’’ cells, in that they had a muchmore negative membrane potential and were electrophysiolog-ically passive (Fig. 2). This type of linear voltage-current rela-tionship is typical of astroglial cells (Berninger et al., 2007). Incontrast, the putative neuronal precursors had a more positivemembrane potential and displayed outward rectification upondepolarization (see Results). With these electrophysiologicalcriteria, we could easily target ‘‘neuron-like’’ cells withoutusing any immunohistochemical cellular markers. Controlexperiments showed that cortical cells from embryonic tissuehave the capacity to form mature neurons when differentiatedon a confluent ECM layer in the presence of serum. After 4days of differentiation, the membrane potential was still rela-tively depolarized and only subthreshold potentials wereobserved upon depolarization. In contrast, after 6 days of dif-ferentiation, action potentials were evoked by the depolariza-tion in three of five ‘‘neuron-like’’ cells (data not shown).These results indicate that the cells develop neural electricalproperties after approximately 1 week, which provided uswith an appropriate time window for studying the effects ofECM patterns on neural differentiation. The electrophysiolog-ical results are expressed as means 6 SD in the text and asmeans 6 95% c.i. in the figures. The levels of significancewere determined by Student’s unpaired t-test (P < 0.05 wasconsidered statistically significant).
RESULTS
Cell Morphology of Differentiating NeuralPrecursor Cells Can Be Directed by an ECMSurface Pattern
To investigate the role of cell morphology in theregulation of neural differentiation, we created two dif-ferent surface patterns using microcontact printing ofECM and poly-D-lysine (Vogt et al., 2003, 2005b). Thepatterns were designed so that the cells either were re-stricted to 20-lm squares (dots) or had the possibility toform processes along 3-lm-wide lines connecting thesquares (dots 1 lines; Fig. 1A). Neural precursor cellsfrom embryonic mouse cortex were cultured on the twodifferent surfaces in serum-free media, without any addi-tion of mitogens. Removal of the mitogens (FGF and
2366 Magnusson et al.
Journal of Neuroscience Research
EGF) will, under normal culture conditions, inducewithin 24 hr spontaneous differentiation of neural pre-cursor cells toward the three major cell lineages in thecentral nervous system: neurons, astrocytes, and oligo-dendrocytes (Johe et al., 1996; McKay, 1997). Already atday 1 after plating, the cells show very different mor-phology, depending on the surface pattern, and, at day4, the difference between the two cultures is striking(Fig. 1B–E). Neural precursor cells grown on the dot 1line pattern express a rather mature morphology withlong processes that form a network along the ECMstamping (Fig. 1B,D). In contrast, neural precursor cellscultured on the dot pattern express a phenotype tradi-tionally considered to be typical for immature neuralcells. The cells form small clusters of completely roundcells without any processes, very similar to proliferatingneural precursor cells cultured in the presence of mito-gens (Fig. 1C,E).
The Cell Proliferation Rate Is Lower When theCells Are Allowed to Form Processes
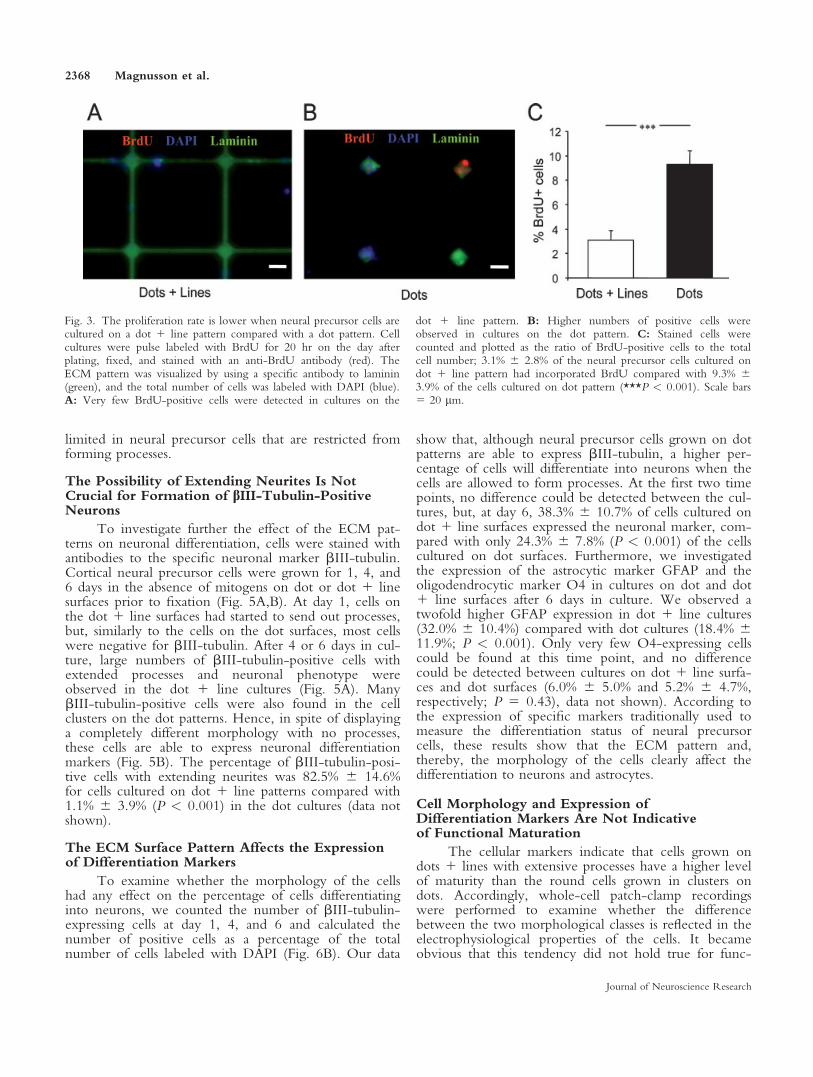
We next examined whether the diverse morpholo-gies of the neural precursor cells on the different surfacesare coupled to the percentage of cells going through celldivision. The cell cultures were pulse labeled with BrdUfor 20 hr on the day after plating, fixed, and stainedwith an anti-BrdU antibody (Fig. 3A,B). Very fewBrdU-positive cells were detected in cultures on the dot1 line pattern (Fig. 3A), whereas a greater number ofpositive cells was observed in cultures on the dot pattern(Fig. 3B). The number of BrdU-positive cells wascounted, and the percentage of the total cell numberwas calculated (Fig. 3C). A threefold increase in prolifer-ation rate was observed for neural precursor cells cul-tured on dots compared with cells cultured on dots 1lines (3.1% 6 2.8% and 9.3% 6 3.9%, respectively; P <0.001), indicating that the round cells that are preventedfrom forming processes have a higher proliferationcapacity. The reason for this could be either that thecells are less mature than the cells cultured on the dot 1line pattern or that cells in tight clusters are more effec-tively stimulated by various autocrine and/or paracrinemitogenic substances produced by the neural precursorcells themselves (Erlandsson et al., 2006).
Nestin Expression Declines Faster in Cells WithExtending Processes
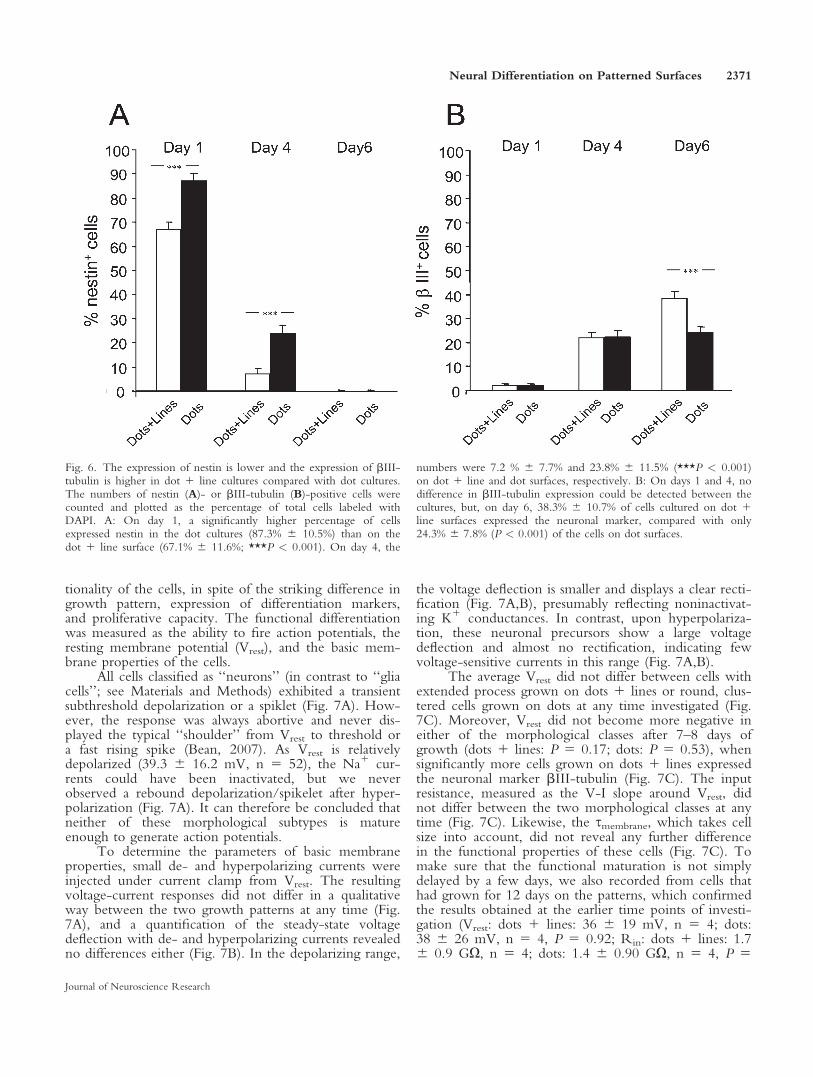
To clarify whether the neural precursor cells cul-tured on the dot pattern are kept in a more immaturestate than the process-bearing cells on the dot 1 linepattern, we next investigated the difference in nestinexpression on the two surfaces. Nestin is one of themost commonly used immunocytochemical markers foridentification of neural stem cells, and its expression isknown to be down-regulated as the stem/precursor cellsinduce their differentiation program (Lendahl et al.,1990). Neural precursor cells were plated on either dotor dot 1 line surfaces and were cultured in the absenceof mitogens for 1, 4, or 6 days before fixation and im-munostaining with specific antibodies to nestin (Fig.4A,B). After 1 day in culture, many cells were stillexpressing nestin on both patterns. However, the inten-sity of the nestin staining was stronger in the dot culturescompared with the dot 1 line cultures. At day 4, thenestin expression had declined markedly, and hardly anypositive cells could be found on the dot 1 line surfaces(Fig. 4A). There were still some positive cells on the dotsurfaces, but, at day 6, no positive cells could bedetected in any of the cultures (data not shown). Thenumber of nestin-positive cells were counted and plottedas a percentage of the total cells (see Fig. 6A). At day 1,a significantly higher percentage of cells expressed nestinin the dot cultures (87.3% 6 10.5%) than on the dot 1line surface (67.1% 6 11.6%; P < 0.001). At day 4, thenumbers were 7.2% 6 7.7% and 23.8% 6 11.5% (P <0.001) on dot 1 line and dot surfaces, respectively.These results suggest that the differentiation process is
Fig. 2. Operational identification of a glia cell. A: The glial cell volt-age response to depolarizing and hyperpolarizing equidistance currentpulses of 500 msec duration, injected from rest, displays a passivebehavior and has no tendency to a subthreshold action potential inthe depolarizing range. Note the negative resting membrane poten-tial. B: The voltage-current relationship measured at the steady-stateresponse is well described by a linear function with a correlationcoefficient of 0.988.
Neural Differentiation on Patterned Surfaces 2367
Journal of Neuroscience Research
limited in neural precursor cells that are restricted fromforming processes.
The Possibility of Extending Neurites Is NotCrucial for Formation of bIII-Tubulin-PositiveNeurons
To investigate further the effect of the ECM pat-terns on neuronal differentiation, cells were stained withantibodies to the specific neuronal marker bIII-tubulin.Cortical neural precursor cells were grown for 1, 4, and6 days in the absence of mitogens on dot or dot 1 linesurfaces prior to fixation (Fig. 5A,B). At day 1, cells onthe dot 1 line surfaces had started to send out processes,but, similarly to the cells on the dot surfaces, most cellswere negative for bIII-tubulin. After 4 or 6 days in cul-ture, large numbers of bIII-tubulin-positive cells withextended processes and neuronal phenotype wereobserved in the dot 1 line cultures (Fig. 5A). ManybIII-tubulin-positive cells were also found in the cellclusters on the dot patterns. Hence, in spite of displayinga completely different morphology with no processes,these cells are able to express neuronal differentiationmarkers (Fig. 5B). The percentage of bIII-tubulin-posi-tive cells with extending neurites was 82.5% 6 14.6%for cells cultured on dot 1 line patterns compared with1.1% 6 3.9% (P < 0.001) in the dot cultures (data notshown).
The ECM Surface Pattern Affects the Expressionof Differentiation Markers
To examine whether the morphology of the cellshad any effect on the percentage of cells differentiatinginto neurons, we counted the number of bIII-tubulin-expressing cells at day 1, 4, and 6 and calculated thenumber of positive cells as a percentage of the totalnumber of cells labeled with DAPI (Fig. 6B). Our data
show that, although neural precursor cells grown on dotpatterns are able to express bIII-tubulin, a higher per-centage of cells will differentiate into neurons when thecells are allowed to form processes. At the first two timepoints, no difference could be detected between the cul-tures, but, at day 6, 38.3% 6 10.7% of cells cultured ondot 1 line surfaces expressed the neuronal marker, com-pared with only 24.3% 6 7.8% (P < 0.001) of the cellscultured on dot surfaces. Furthermore, we investigatedthe expression of the astrocytic marker GFAP and theoligodendrocytic marker O4 in cultures on dot and dot1 line surfaces after 6 days in culture. We observed atwofold higher GFAP expression in dot 1 line cultures(32.0% 6 10.4%) compared with dot cultures (18.4% 611.9%; P < 0.001). Only very few O4-expressing cellscould be found at this time point, and no differencecould be detected between cultures on dot 1 line surfa-ces and dot surfaces (6.0% 6 5.0% and 5.2% 6 4.7%,respectively; P 5 0.43), data not shown). According tothe expression of specific markers traditionally used tomeasure the differentiation status of neural precursorcells, these results show that the ECM pattern and,thereby, the morphology of the cells clearly affect thedifferentiation to neurons and astrocytes.
Cell Morphology and Expression ofDifferentiation Markers Are Not Indicativeof Functional Maturation
The cellular markers indicate that cells grown ondots 1 lines with extensive processes have a higher levelof maturity than the round cells grown in clusters ondots. Accordingly, whole-cell patch-clamp recordingswere performed to examine whether the differencebetween the two morphological classes is reflected in theelectrophysiological properties of the cells. It becameobvious that this tendency did not hold true for func-
Fig. 3. The proliferation rate is lower when neural precursor cells arecultured on a dot 1 line pattern compared with a dot pattern. Cellcultures were pulse labeled with BrdU for 20 hr on the day afterplating, fixed, and stained with an anti-BrdU antibody (red). TheECM pattern was visualized by using a specific antibody to laminin(green), and the total number of cells was labeled with DAPI (blue).A: Very few BrdU-positive cells were detected in cultures on the
dot 1 line pattern. B: Higher numbers of positive cells wereobserved in cultures on the dot pattern. C: Stained cells werecounted and plotted as the ratio of BrdU-positive cells to the totalcell number; 3.1% 6 2.8% of the neural precursor cells cultured ondot 1 line pattern had incorporated BrdU compared with 9.3% 63.9% of the cells cultured on dot pattern (***P < 0.001). Scale bars5 20 lm.
2368 Magnusson et al.
Journal of Neuroscience Research
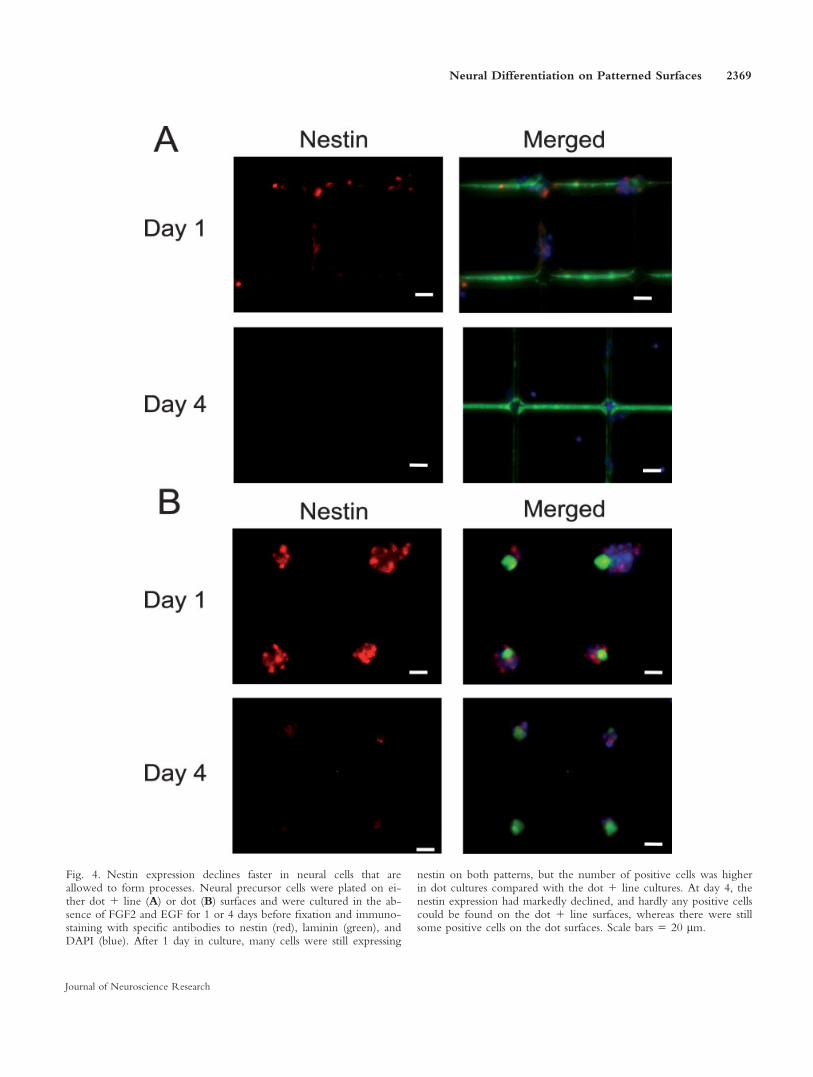
Fig. 4. Nestin expression declines faster in neural cells that areallowed to form processes. Neural precursor cells were plated on ei-ther dot 1 line (A) or dot (B) surfaces and were cultured in the ab-sence of FGF2 and EGF for 1 or 4 days before fixation and immuno-staining with specific antibodies to nestin (red), laminin (green), andDAPI (blue). After 1 day in culture, many cells were still expressing
nestin on both patterns, but the number of positive cells was higherin dot cultures compared with the dot 1 line cultures. At day 4, thenestin expression had markedly declined, and hardly any positive cellscould be found on the dot 1 line surfaces, whereas there were stillsome positive cells on the dot surfaces. Scale bars 5 20 lm.
Neural Differentiation on Patterned Surfaces 2369
Journal of Neuroscience Research
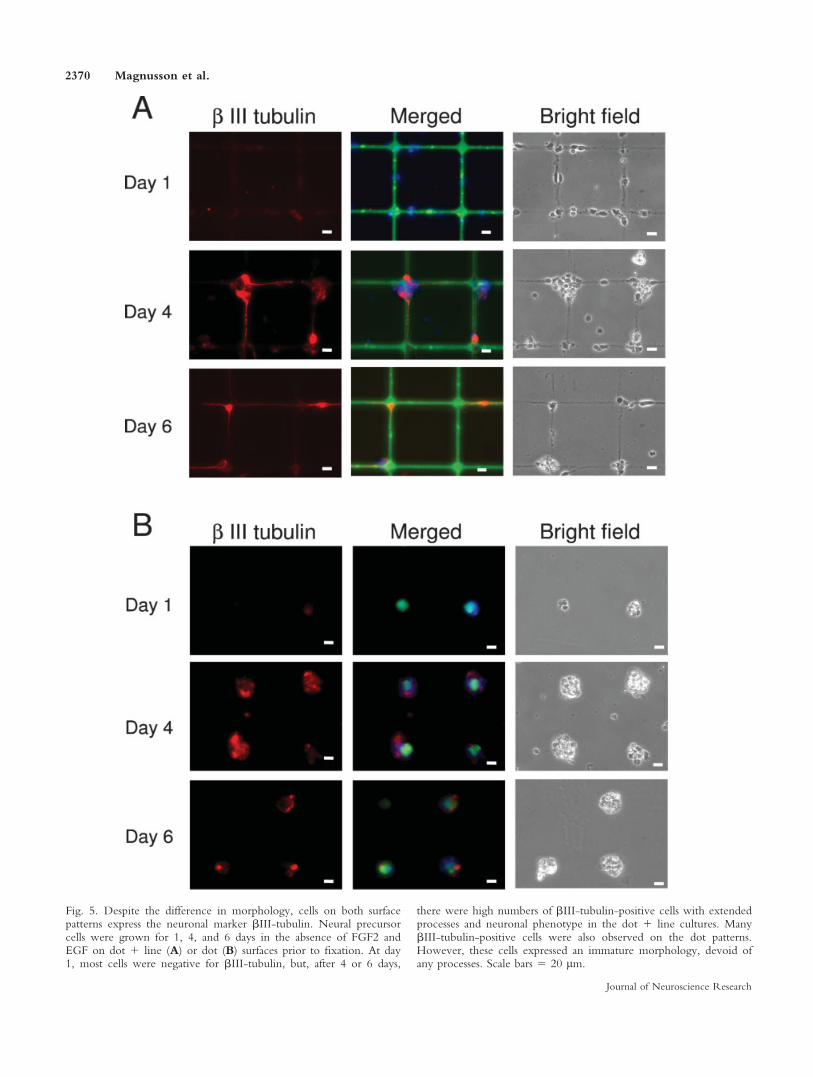
Fig. 5. Despite the difference in morphology, cells on both surfacepatterns express the neuronal marker bIII-tubulin. Neural precursorcells were grown for 1, 4, and 6 days in the absence of FGF2 andEGF on dot 1 line (A) or dot (B) surfaces prior to fixation. At day1, most cells were negative for bIII-tubulin, but, after 4 or 6 days,
there were high numbers of bIII-tubulin-positive cells with extendedprocesses and neuronal phenotype in the dot 1 line cultures. ManybIII-tubulin-positive cells were also observed on the dot patterns.However, these cells expressed an immature morphology, devoid ofany processes. Scale bars 5 20 lm.
2370 Magnusson et al.
Journal of Neuroscience Research
tionality of the cells, in spite of the striking difference ingrowth pattern, expression of differentiation markers,and proliferative capacity. The functional differentiationwas measured as the ability to fire action potentials, theresting membrane potential (Vrest), and the basic mem-brane properties of the cells.
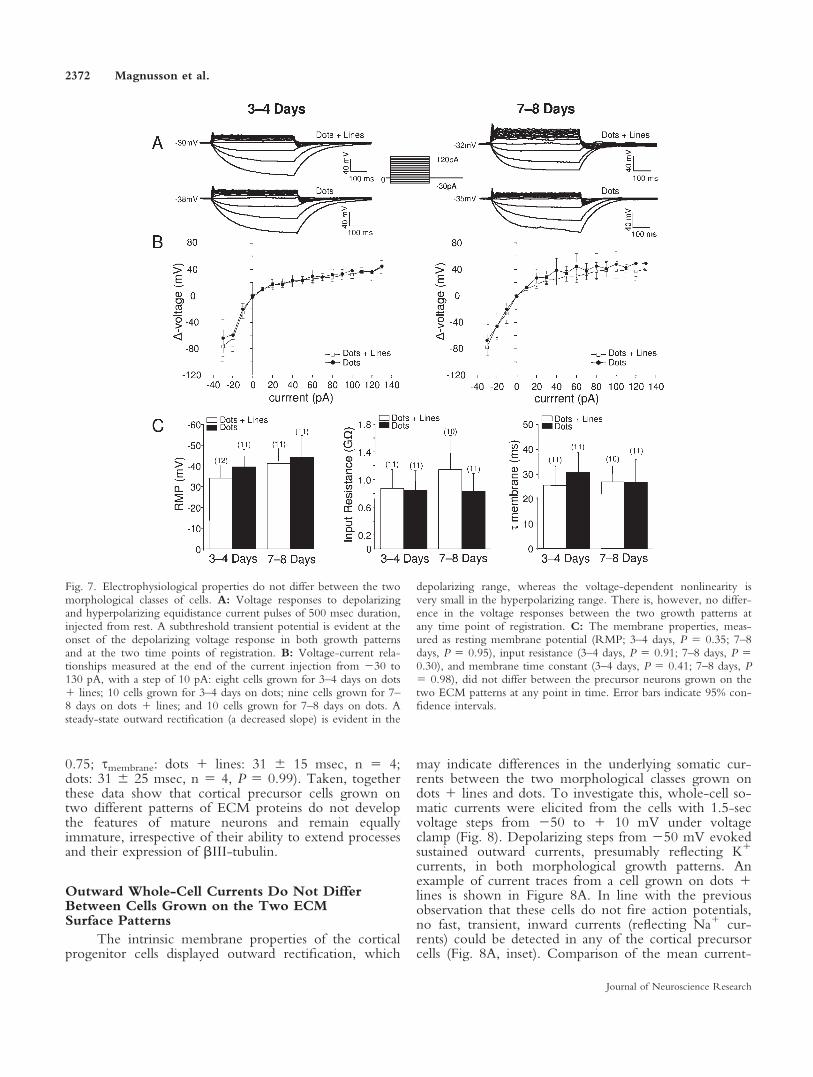
All cells classified as ‘‘neurons’’ (in contrast to ‘‘gliacells’’; see Materials and Methods) exhibited a transientsubthreshold depolarization or a spiklet (Fig. 7A). How-ever, the response was always abortive and never dis-played the typical ‘‘shoulder’’ from Vrest to threshold ora fast rising spike (Bean, 2007). As Vrest is relativelydepolarized (39.3 6 16.2 mV, n 5 52), the Na1 cur-rents could have been inactivated, but we neverobserved a rebound depolarization/spikelet after hyper-polarization (Fig. 7A). It can therefore be concluded thatneither of these morphological subtypes is matureenough to generate action potentials.
To determine the parameters of basic membraneproperties, small de- and hyperpolarizing currents wereinjected under current clamp from Vrest. The resultingvoltage-current responses did not differ in a qualitativeway between the two growth patterns at any time (Fig.7A), and a quantification of the steady-state voltagedeflection with de- and hyperpolarizing currents revealedno differences either (Fig. 7B). In the depolarizing range,
the voltage deflection is smaller and displays a clear recti-fication (Fig. 7A,B), presumably reflecting noninactivat-ing K1 conductances. In contrast, upon hyperpolariza-tion, these neuronal precursors show a large voltagedeflection and almost no rectification, indicating fewvoltage-sensitive currents in this range (Fig. 7A,B).
The average Vrest did not differ between cells withextended process grown on dots 1 lines or round, clus-tered cells grown on dots at any time investigated (Fig.7C). Moreover, Vrest did not become more negative ineither of the morphological classes after 7–8 days ofgrowth (dots 1 lines: P 5 0.17; dots: P 5 0.53), whensignificantly more cells grown on dots 1 lines expressedthe neuronal marker bIII-tubulin (Fig. 7C). The inputresistance, measured as the V-I slope around Vrest, didnot differ between the two morphological classes at anytime (Fig. 7C). Likewise, the smembrane, which takes cellsize into account, did not reveal any further differencein the functional properties of these cells (Fig. 7C). Tomake sure that the functional maturation is not simplydelayed by a few days, we also recorded from cells thathad grown for 12 days on the patterns, which confirmedthe results obtained at the earlier time points of investi-gation (Vrest: dots 1 lines: 36 6 19 mV, n 5 4; dots:38 6 26 mV, n 5 4, P 5 0.92; Rin: dots 1 lines: 1.76 0.9 GX, n 5 4; dots: 1.4 6 0.90 GX, n 5 4, P 5
Fig. 6. The expression of nestin is lower and the expression of bIII-tubulin is higher in dot 1 line cultures compared with dot cultures.The numbers of nestin (A)- or bIII-tubulin (B)-positive cells werecounted and plotted as the percentage of total cells labeled withDAPI. A: On day 1, a significantly higher percentage of cellsexpressed nestin in the dot cultures (87.3% 6 10.5%) than on thedot 1 line surface (67.1% 6 11.6%; ***P < 0.001). On day 4, the
numbers were 7.2 % 6 7.7% and 23.8% 6 11.5% (***P < 0.001)on dot 1 line and dot surfaces, respectively. B: On days 1 and 4, nodifference in bIII-tubulin expression could be detected between thecultures, but, on day 6, 38.3% 6 10.7% of cells cultured on dot 1line surfaces expressed the neuronal marker, compared with only24.3% 6 7.8% (P < 0.001) of the cells on dot surfaces.
Neural Differentiation on Patterned Surfaces 2371
Journal of Neuroscience Research
0.75; smembrane: dots 1 lines: 31 6 15 msec, n 5 4;dots: 31 6 25 msec, n 5 4, P 5 0.99). Taken, togetherthese data show that cortical precursor cells grown ontwo different patterns of ECM proteins do not developthe features of mature neurons and remain equallyimmature, irrespective of their ability to extend processesand their expression of bIII-tubulin.
Outward Whole-Cell Currents Do Not DifferBetween Cells Grown on the Two ECMSurface Patterns
The intrinsic membrane properties of the corticalprogenitor cells displayed outward rectification, which
may indicate differences in the underlying somatic cur-rents between the two morphological classes grown ondots 1 lines and dots. To investigate this, whole-cell so-matic currents were elicited from the cells with 1.5-secvoltage steps from 250 to 1 10 mV under voltageclamp (Fig. 8). Depolarizing steps from 250 mV evokedsustained outward currents, presumably reflecting K1
currents, in both morphological growth patterns. Anexample of current traces from a cell grown on dots 1lines is shown in Figure 8A. In line with the previousobservation that these cells do not fire action potentials,no fast, transient, inward currents (reflecting Na1 cur-rents) could be detected in any of the cortical precursorcells (Fig. 8A, inset). Comparison of the mean current-
Fig. 7. Electrophysiological properties do not differ between the twomorphological classes of cells. A: Voltage responses to depolarizingand hyperpolarizing equidistance current pulses of 500 msec duration,injected from rest. A subthreshold transient potential is evident at theonset of the depolarizing voltage response in both growth patternsand at the two time points of registration. B: Voltage-current rela-tionships measured at the end of the current injection from 230 to130 pA, with a step of 10 pA: eight cells grown for 3–4 days on dots1 lines; 10 cells grown for 3–4 days on dots; nine cells grown for 7–8 days on dots 1 lines; and 10 cells grown for 7–8 days on dots. Asteady-state outward rectification (a decreased slope) is evident in the
depolarizing range, whereas the voltage-dependent nonlinearity isvery small in the hyperpolarizing range. There is, however, no differ-ence in the voltage responses between the two growth patterns atany time point of registration. C: The membrane properties, meas-ured as resting membrane potential (RMP; 3–4 days, P 5 0.35; 7–8days, P 5 0.95), input resistance (3–4 days, P 5 0.91; 7–8 days, P 50.30), and membrane time constant (3–4 days, P 5 0.41; 7–8 days, P5 0.98), did not differ between the precursor neurons grown on thetwo ECM patterns at any point in time. Error bars indicate 95% con-fidence intervals.
2372 Magnusson et al.
Journal of Neuroscience Research
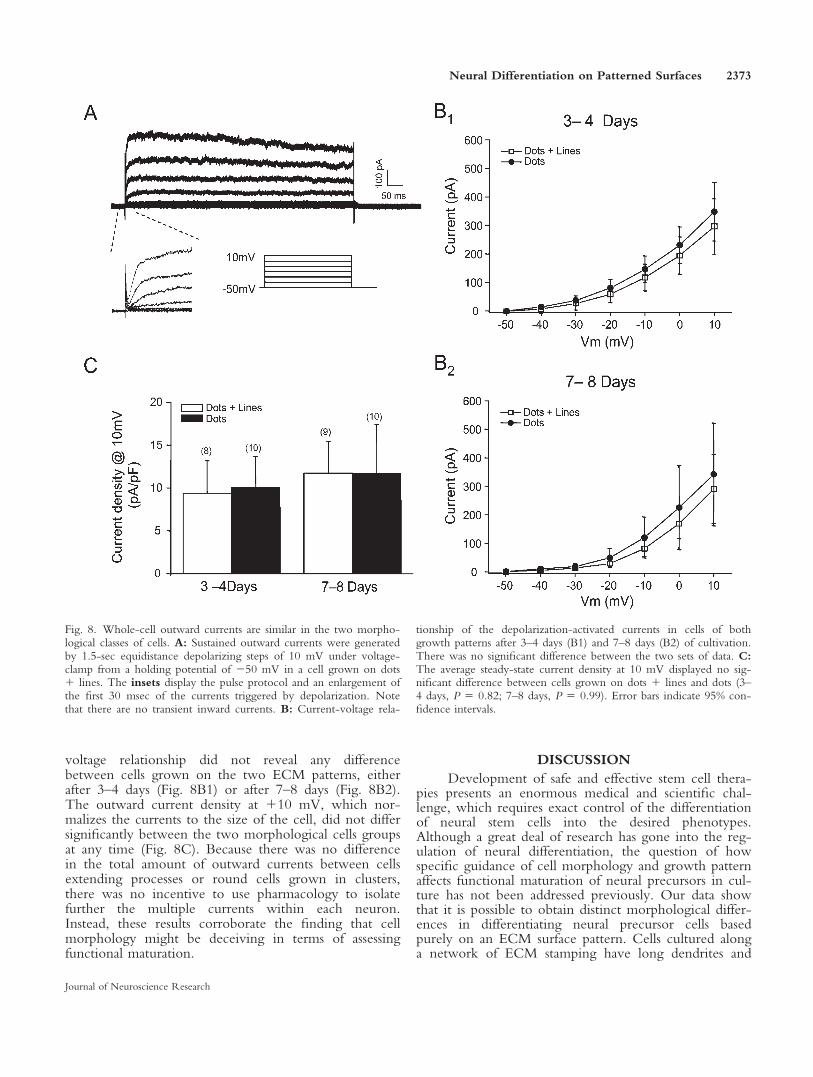
voltage relationship did not reveal any differencebetween cells grown on the two ECM patterns, eitherafter 3–4 days (Fig. 8B1) or after 7–8 days (Fig. 8B2).The outward current density at 110 mV, which nor-malizes the currents to the size of the cell, did not differsignificantly between the two morphological cells groupsat any time (Fig. 8C). Because there was no differencein the total amount of outward currents between cellsextending processes or round cells grown in clusters,there was no incentive to use pharmacology to isolatefurther the multiple currents within each neuron.Instead, these results corroborate the finding that cellmorphology might be deceiving in terms of assessingfunctional maturation.
DISCUSSION
Development of safe and effective stem cell thera-pies presents an enormous medical and scientific chal-lenge, which requires exact control of the differentiationof neural stem cells into the desired phenotypes.Although a great deal of research has gone into the reg-ulation of neural differentiation, the question of howspecific guidance of cell morphology and growth patternaffects functional maturation of neural precursors in cul-ture has not been addressed previously. Our data showthat it is possible to obtain distinct morphological differ-ences in differentiating neural precursor cells basedpurely on an ECM surface pattern. Cells cultured alonga network of ECM stamping have long dendrites and
Fig. 8. Whole-cell outward currents are similar in the two morpho-logical classes of cells. A: Sustained outward currents were generatedby 1.5-sec equidistance depolarizing steps of 10 mV under voltage-clamp from a holding potential of 250 mV in a cell grown on dots1 lines. The insets display the pulse protocol and an enlargement ofthe first 30 msec of the currents triggered by depolarization. Notethat there are no transient inward currents. B: Current-voltage rela-
tionship of the depolarization-activated currents in cells of bothgrowth patterns after 3–4 days (B1) and 7–8 days (B2) of cultivation.There was no significant difference between the two sets of data. C:The average steady-state current density at 10 mV displayed no sig-nificant difference between cells grown on dots 1 lines and dots (3–4 days, P 5 0.82; 7–8 days, P 5 0.99). Error bars indicate 95% con-fidence intervals.
Neural Differentiation on Patterned Surfaces 2373
Journal of Neuroscience Research

show other neuron-like features, such as low prolifera-tion rate and expression of the cellular marker bIII-tubulin. In contrast, neural precursor cells cultured onsimple dots of ECM proteins retain an immature mor-phology completely devoid of extending processes.These cells have a higher proliferation capacity and alower expression of bIII-tubulin compared with the pro-cess-bearing cells cultured on the ECM network pattern.However, electrophysiological investigations of the dif-ferentiating neural precursors on the two patternsrevealed that they are unable to fire action potentials andhave a depolarized Vrest, a high Rin, and a slowsmembrane, all of which indicate immaturity (Picken Bah-rey and Moody, 2003; Zhang, 2004). The developmentof electrical properties in the cells thus ceases at animmature stage irrespective of the neural-like growthpattern or time in culture. Taken together, these dataclearly demonstrate that, to be able to determine the dif-ferentiation status of neural cells, it is not enough to basethe investigations on expression of the immunospecificmarkers and the morphological phenotype, but electro-physiological examinations are also needed.
Improved understanding of all parameters that reg-ulate differentiation into the various neural lineages,including contacts between the cell and its surroundings,is of high importance. Much interest has been focusedon how the extrinsic environment influences neural fatechoice. In vitro studies of neural stem/precursor cellsderived from the central nervous system show that thecells are responsive to the addition of single extrinsicdeterminants, such as CNTF, PDGF, T3, FGF2, EGF(Johe et al., 1996; Bonni et al., 1997; McKay, 1997;Gage, 2000; Erlandsson et al., 2001, 2006; Glaser et al.,2005). Other studies have elucidated the intrinsic devel-opmental program that promotes neural lineage in stem/precursor cells (e.g., Nieto et al., 2001; Hanashimaet al., 2002; Berninger et al., 2007). Recently, variousbiomaterials and three-dimensional matrices have beenused to investigate how the ECM controls neural fatechoice. In a seminal study, Silva et al. (2004) demon-strated that an artificial scaffold containing the lamininepitope IKVAV could direct differentiation of neuralprogenitor cells into neurons while suppressing astrocytedifferentiation. In another study, the number of ECMproteins, in combination with different growth factors,was screened (Nakajima et al., 2007). In line with a pre-vious study, it was observed that effects of the growthfactors were frequently altered depending on the type ofcoimmobilized matrices, suggesting that a cross-talkbetween integrins and growth factor receptors may becritical to neural differentiation (ffrench-Constant andColognato, 2004; Nakajima et al., 2007). Our data dem-onstrate that manipulation of the growth pattern, basedon the nature of the cell culture surface alone, is able toinfluence differentiation of the morphological and im-munohistochemical characteristics of mouse cortical pre-cursor cells but not their electrophysiological maturation.How stem/precursor cells develop the electrical pheno-type of a neuron is therefore an important issue. In a
broader sense, the intrinsic membrane excitability, i.e.,how neurons produce action potentials and integratetheir synaptic input, is dependent on an intrinsic devel-opmental program but also on extrinsic factors such asspecific growth factors (Baines and Pym, 2006). On amechanistic level, though, relatively little is knownabout how this development occurs. Some parallels maybe drawn from studies of how mature neurons adapttheir intrinsic electrical properties to altered externalconditions, so-called homeostatic plasticity (Desai, 2003).It appears that an increase in membrane excitability isproduced by coordinated changes in the voltage-gatedNa1 currents (increases) and persistent K1 currents(decreases; Desai et al., 1999). The notion that the de-velopmental expression of Na1 channels and their den-sity is critical for functional neurogenesis is also sup-ported by the in vivo development of electrical proper-ties of Drosophila embryonic central neurons, in whichthe first action potentials coincide with the appearanceof an inward Na1 current (Baines and Bate, 1998). Elec-trophysiological studies of neuron-specific ion currentsin embryo- and adult-derived neural stem/precursor cellsin vitro have shown that it is possible to acquire physio-logical properties similar to those of mature neuronsunder differentiating conditions (Sah et al., 1997; Piperet al., 2000; Cho et al., 2002; Song et al., 2002). How-ever, in line with the present study, several studies high-light the fact that morphological and immunohistochem-ical appearances may not be predictive of functional ma-turity (Balasubramaniyan et al., 2004; Pagani et al., 2006;Donato et al., 2007).
As stem/precursor cells are considered to hold greatpotential for treatment of a wide range of central nerv-ous conditions, it is important to investigate the func-tional properties of such cells. Most disorders within theCNS are caused by the loss or damage of a specific cellpopulation, and predifferentiation of neural precursorcells to a specific cell type and function prior to in vivotransplantation is therefore preferable. However, even ifit was fully understood how to achieve a highly purifiedpopulation of neurons suitable for transplantations, thesecells would still have to survive, migrate to the damagedarea, and replace the missing cells. A general problemwith stem cell-based therapies is that most grafted cellsundergo apoptosis within a few days following transplan-tation (Zawada et al., 1998; Sortwell et al., 2000;Emgard et al., 2003). Transplantation of mature neuronswould be extremely difficult, because these cells expresslong processes that break very easily, strongly limitingthe cell survival chances and potential success of suchtherapy. The fact that the round, less brittle cells cul-tured on the dot pattern are able to express neuronalmarkers (but to a lower extent) and have exactly thesame functional properties as cells with a mature neuro-nal morphology suggests that these cells might be able todifferentiate further to become mature, nonprocess-bear-ing neurons in the presence of the correct extrinsic fac-tors. Hence, further studies of the differentiation capacityof cells cultured on the dot pattern in combination with
2374 Magnusson et al.
Journal of Neuroscience Research

the addition of various growth factors to the cell culturemedia would be of clinical interest.
REFERENCES
Baines RA, Bate M. 1998. Electrophysiological development of central
neurons in the Drosophila embryo. J Neurosci 18:4673–4683.
Baines RA, Pym EC. 2006. Determinants of electrical properties in
developing neurons. Semin Cell Dev Biol 17:12–19.
Balasubramaniyan V, de Haas AH, Bakels R, Koper A, Boddeke HW,
Copray JC. 2004. Functionally deficient neuronal differentiation of
mouse embryonic neural stem cells in vitro. Neurosci Res 49:261–265.
Bean BP. 2007. The action potential in mammalian central neurons. Nat
Rev Neurosci 8:451–465.
Berninger B, Costa MR, Koch U, Schroeder T, Sutor B, Grothe B,
Gotz M. 2007. Functional properties of neurons derived from in vitro
reprogrammed postnatal astroglia. J Neurosci 27:8654–8664.
Bonni A, Sun Y, Nadal-Vicens M, Bhatt A, Frank DA, Rozovsky I,
Stahl N, Yancopoulos GD, Greenberg ME. 1997. Regulation of glio-
genesis in the central nervous system by the JAK-STAT signaling path-
way. Science 278:477–483.
Campos LS. 2005. Beta1 integrins and neural stem cells: making sense of
the extracellular environment. Bioessays 27:698–707.
Cho T, Bae JH, Choi HB, Kim SS, McLarnon JG, Suh-Kim H, Kim
SU, Min CK. 2002. Human neural stem cells: electrophysiological
properties of voltage-gated ion channels. Neuroreport 13:1447–1452.
Desai NS. 2003. Homeostatic plasticity in the CNS: synaptic and intrinsic
forms. J Physiol (Paris) 97:391–402.
Desai NS, Rutherford LC, Turrigiano GG. 1999. Plasticity in the intrin-
sic excitability of cortical pyramidal neurons. Nat Neurosci 2:515–520.
Discher DE, Janmey P, Wang YL. 2005. Tissue cells feel and respond to
the stiffness of their substrate. Science 310:1139–1143.
Donato R, Miljan EA, Hines SJ, Aouabdi S, Pollock K, Patel S, Edwards
FA, Sinden JD. 2007. Differential development of neuronal physiologi-
cal responsiveness in two human neural stem cell lines. BMC Neurosci
8:36.
Emgard M, Hallin U, Karlsson J, Bahr BA, Brundin P, Blomgren K.
2003. Both apoptosis and necrosis occur early after intracerebral grafting
of ventral mesencephalic tissue: a role for protease activation. J Neuro-
chem 86:1223–1232.
Engler AJ, Griffin MA, Sen S, Bonnemann CG, Sweeney HL, Discher
DE. 2004. Myotubes differentiate optimally on substrates with tissue-
like stiffness: pathological implications for soft or stiff microenviron-
ments. J Cell Biol 166:877–887.
Erlandsson A, Enarsson M, Forsberg-Nilsson K. 2001. Immature neurons
from CNS stem cells proliferate in response to platelet-derived growth
factor. J Neurosci 21:3483–3491.
Erlandsson A, Brannvall K, Gustafsdottir S, Westermark B, Forsberg-
Nilsson K. 2006. Autocrine/paracrine platelet-derived growth factor
regulates proliferation of neural progenitor cells. Cancer Res 66:8042–
8048.
Ffrench-Constant C, Colognato H. 2004. Integrins: versatile integrators
of extracellular signals. Trends Cell Biol 14:678–686.
Gage FH. 2000. Mammalian neural stem cells. Science 287:1433–1438.
Georges PC, Janmey PA. 2005. Cell type-specific response to growth on
soft materials. J Appl Physiol 98:1547–1553.
Glaser T, Perez-Bouza A, Klein K, Brustle O. 2005. Generation of puri-
fied oligodendrocyte progenitors from embryonic stem cells. FASEB J
19:112–114.
Hanashima C, Shen L, Li SC, Lai E. 2002. Brain factor-1 controls the
proliferation and differentiation of neocortical progenitor cells through
independent mechanisms. J Neurosci 22:6526–6536.
Johe KK, Hazel TG, Muller T, Dugich-Djordjevic MM, McKay RD.
1996. Single factors direct the differentiation of stem cells from the fetal
and adult central nervous system. Genes Dev 10:3129–3140.
Lendahl U, Zimmerman LB, McKay RD. 1990. CNS stem cells express
a new class of intermediate filament protein. Cell 60:585–595.
Leone DP, Relvas JB, Campos LS, Hemmi S, Brakebusch C, Fassler R,
Ffrench-Constant C, Suter U. 2005. Regulation of neural progenitor
proliferation and survival by beta1 integrins. J Cell Sci 118:2589–2599.
Ma W, Fitzgerald W, Liu QY, O’Shaughnessy TJ, Maric D, Lin HJ,
Alkon DL, Barker JL. 2004. CNS stem and progenitor cell differentia-
tion into functional neuronal circuits in three-dimensional collagen gels.
Exp Neurol 190:276–288.
McKay R. 1997. Stem cells in the central nervous system. Science
276:66–71.
Nakajima M, Ishimuro T, Kato K, Ko IK, Hirata I, Arima Y, Iwata H.
2007. Combinatorial protein display for the cell-based screening of bio-
materials that direct neural stem cell differentiation. Biomaterials 28:
1048–1060.
Nieto M, Schuurmans C, Britz O, Guillemot F. 2001. Neural bHLH
genes control the neuronal vs. glial fate decision in cortical progenitors.
Neuron 29:401–413.
Pagani F, Lauro C, Fucile S, Catalano M, Limatola C, Eusebi F, Grassi
F. 2006. Functional properties of neurons derived from fetal mouse
neurospheres are compatible with those of neuronal precursors in vivo.
J Neurosci Res 83:1494–1501.
Picken Bahrey HL, Moody WJ. 2003. Early development of voltage-
gated ion currents and firing properties in neurons of the mouse cere-
bral cortex. J Neurophysiol 89:1761–1773.
Piper DR, Mujtaba T, Rao MS, Lucero MT. 2000. Immunocytochemi-
cal and physiological characterization of a population of cultured human
neural precursors. J Neurophysiol 84:534–548.
Sah DW, Ray J, Gage FH. 1997. Regulation of voltage- and ligand-gated
currents in rat hippocampal progenitor cells in vitro. J Neurobiol 32:95–110.
Schmalenberg KE, Uhrich KE. 2005. Micropatterned polymer substrates
control alignment of proliferating Schwann cells to direct neuronal
regeneration. Biomaterials 26:1423–1430.
Silva GA, Czeisler C, Niece KL, Beniash E, Harrington DA, Kessler JA,
Stupp SI. 2004. Selective differentiation of neural progenitor cells by
high-epitope density nanofibers. Science 303:1352–1355.
Song HJ, Stevens CF, Gage FH. 2002. Neural stem cells from adult hip-
pocampus develop essential properties of functional CNS neurons. Nat
Neurosci 5:438–445.
Sortwell CE, Pitzer MR, Collier TJ. 2000. Time course of apoptotic cell
death within mesencephalic cell suspension grafts: implications for
improving grafted dopamine neuron survival. Exp Neurol 165:268–277.
Vogt AK, Lauer L, Knoll W, Offenhausser A. 2003. Micropatterned sub-
strates for the growth of functional neuronal networks of defined ge-
ometry. Biotechnol Prog 19:1562–1568.
Vogt AK, Brewer GJ, Offenhausser A. 2005a. Connectivity patterns in
neuronal networks of experimentally defined geometry. Tissue Eng
11:1757–1767.
Vogt AK, Wrobel G, Meyer W, Knoll W, Offenhausser A. 2005b. Syn-
aptic plasticity in micropatterned neuronal networks. Biomaterials
26:2549–2557.
Wyart C, Ybert C, Bourdieu L, Herr C, Prinz C, Chatenay D. 2002.
Constrained synaptic connectivity in functional mammalian neuronal net-
works grown on patterned surfaces. J Neurosci Methods 117:123–131.
Yeung T, Georges PC, Flanagan LA, Marg B, Ortiz M, Funaki M, Zahir
N, Ming W, Weaver V, Janmey PA. 2005. Effects of substrate stiffness
on cell morphology, cytoskeletal structure, and adhesion. Cell Motil
Cytoskeleton 60:24–34.
Zawada WM, Zastrow DJ, Clarkson ED, Adams FS, Bell KP, Freed CR.
1998. Growth factors improve immediate survival of embryonic dopa-
mine neurons after transplantation into rats. Brain Res 786:96–103.
Zhang ZW. 2004. Maturation of layer V pyramidal neurons in the rat
prefrontal cortex: intrinsic properties and synaptic function. J Neuro-
physiol 91:1171–1182.
Neural Differentiation on Patterned Surfaces 2375
Journal of Neuroscience Research