
Successful Treatment of Human Acute T-Cell Leukemia in SCID Mice Using the Anti-CD7-deglycosylated...
9
[CANCER RESEARCH 52, 1314-1321, March I. 1992] Successful Treatment of Human Acute T-Cell Leukemia in SCID Mice Using the Anti-CD7-deglycosylated Ricin A-Chain Immunotoxin DA71 Burkhard Jansen, Daniel A. Vallera, Waclaw B. Jaszcz, Dat Nguyen, and John H. Kersey2 Bone Marrow Transplantation Program [D. A. V.,J. H. K.¡and Departments of Laboratory Medicine/Pathology [B. J., W. B. ]., D. N., J. H. K.], Pediatrics [J. H. K.J , and Therapeutic Radiology; Section on Experimental Cancer Immunology [D. A. V.], University of Minnesota, Minneapolis, Minnesota 55455 ABSTRACT The study of new therapeutic approaches for refractory human leuke mia has been hampered by the lack of relevant in vivo models with disseminated disease, particularly T acute lymphoblastic leukemia (T- ALL). In the present study we evaluated methods for establishing and therapy of a human T-ALL cell line (MT-ALL) in 73 SCID mice. MT- ALL is a T<ell receptor a/ß+, CD3+, and CD7* leukemia cell line, derived from a patient with refractory disease and early death. Injection of 5 x IO7 MT-ALL cells i.v. caused disseminated human leukemia in hematopoietic and nonhematopoietic organs in 100% of SCID mice (n = 9) leading to death or terminal disease at 65 to 70 days after a uniform clinical course. To study possible therapeutic approaches for dissemi nated leukemia we utilized an immunotoxin, DA7, constructed by chem ically linking the mouse IgG2b anti-CD7(3AlE) monoclonal antibody which recognizes a pan-T-cell marker expressed on almost all T-cell leukemias to deglycosylated ricin A-chain, a catalytic plant toxin and inhibitor of protein synthesis. Administration of DA7 led to >5 log kill of clonogenic MT-ALL cells in vitro and selectively inhibited protein synthesis. DA7 was administered to mice at a dose of 10 Mg/mouse/day for 5 consecutive days starting 8 days after i.v. inoculation of leukemia. The immunotoxin therapy resulted in significant long term survival over 348 days compared to untreated or control mice treated with anti-CD7 antibody and deglycosylated ricin A-chain which were all dead by day 70 (P < 0.001). Even after more than 11 months there was no evidence of disease in 82% of the DA7 treated animals. SCID mice given i.p. injections (n = 9) developed an i.p. tumor mass but demonstrated metas tasis outside the peritoneum with disseminated leukemia in hematopoietic and nonhematopoietic organs, a finding different from most conventional nude mouse models. The leukemia was fatal in 100% and killed the animals at 68-95 days. SCID mice given i.p. injections of MT-ALL completely responded to therapy with DA7, resulting in survival of 100% of the animals (n = 10) at 216 days (/' < 0.001 compared to untreated animals). Anti-CD7 antibody, deglycosylated ricin A-chain, and a control anti-melanoma immunotoxin (INDI-RIA) showed no therapeutic effect. We conclude that DA7 is an effective in vivo therapeutic agent against human MT-ALL ¡n the SCID mouse system, suggesting potential use fulness for therapy of humans with poor prognosis T-cell leukemia. INTRODUCTION Mice homozygous for the SCID3 mutation on chromosome 16 have a severe combined immunodeficiency caused by the inability to correctly rearrange their immunoglobulin and T- cell receptor genes (1-5). The SCID mutation leads to an Received 10/7/91 ; accepted 12/17/91. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1This work was supported by Grams CA-49721, CA-21737, CA-31618, and CA-36725 from the National Cancer Institute/NIH and the Childrens Cancer Research Fund. J. H. K. is the recipient of an Outstanding Investigator Award (CA-49721) from the National Cancer Institute (CGCTR no. 69). 2 To whom requests for reprints should be addressed. 3The abbreviations used are: SCID, severe combined immunodeficiency; DA7, immunotoxin constructed with deglycosylated (D) ricin A-chain toxin (A) and anti-CD7 monoclonal antibody (7); IT, immunotoxin; dgRTA, deglycosylated ricin A-chain; FACS, fluorescence-activated cell sorter; MoAb, monoclonal anti body; FCS, fetal calf serum; FITC, fluorescein isothiocyanate; ALL, acute lym phoblastic leukemia; TCR, T-cell receptor; IMDM, Iscove's modifiai Dulbecco's medium; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; 1C«,, the concentration of DA7 inhibiting the protein synthesis to 50% of control. inability to repair DNA damage induced by ionizing radiation as well as interfering with rearrangement of the immunoglob ulin and T-cell receptor genes (6). Despite the impairment in lymphocyte differentiation, the SCID mouse hematopoietic microenvironment is intact (7), allowing the analysis of differ entiation of the murine lymphoid system and also studies of the human lymphocyte development and biology after reconsti tution with human lymphoid cells (8-12). SCID mice can also be used as a model for Epstein-Barr virus associated lympho- proliferative disorders of human B-cell origin (13-15). Acquired immunodeficiency syndrome (16-17), human lupus erythema- tosus (18), and human cutaneous T-cell lymphoma (19) have been studied in the SCID mouse. Human leukemias of non-T- cell origin have been established in SCID mice (20, 21). Re cently Cesano et al. (22) reported the engraft nient of human T- ALL following i.p. injection in SCID mice. We have previously noted that the SCID mouse provides an excellent microenvi ronment for the growth of human t(4;l 1) acute leukemia (23). In the present studies, we report a model system for human T-ALL, established by injecting the human TCR a/ß+, CD3+, CD7+ T-ALL cell line MT-ALL (24) i.p. and i.v. into SCID mice. In addition, we report the first successful treatment of disseminated human T-ALL in vivo using the anti-CD7(3AlE)- dgRTA IT, DA7, in our model system. The MoAb 3A1E recognizes the CD7 antigen, expressed on essentially all human T-cell leukemias and leukemia progenitors (25). RTA is a toxic portion (M, 30,000) of the plant toxin ricin derived from the castor seed plant Ricinis communis. Currently under clinical evaluation, RTA immunotoxins have demonstrated efficiency when used for therapy of steroid resistant graft versus host disease (26) and B-cell malignancies (27). The toxin was chem ically deglycosylated to reduce its reactivity with the reticulo- endothelial system (28). In this study, therapy using DA7 resulted in long term survival of SCID mice bearing dissemi nated human T-cell leukemia. MATERIALS AND METHODS Experimental Animals. Pathogen free C.B-17 (n = 70) and C3H (n = 3) seid/seid (SCID) mice, 4-7 weeks old, were obtained from the NIH-Fredrick Cancer Research Center (Fredrick, MD), from the Jack son Laboratory (Bar Harbor, ME) or from Taconic Farms, Inc. (Ger- mantown, NY). The animals were housed in laminar flow racks or microisolator cages under specific pathogen free conditions and re ceived autoclaved food, water and an antibiotic prophylaxis with sul- famethoxazole and trimethoprim. Tumor Cells. The previously described human TCR a//8*, CD3% CD7+ T-ALL cell line (MT-ALL) (24) with potential for multilineage differentiation, derived from a patient with refractory disease, was maintained in IMDM (Gibco Laboratories, Grand Island, NY) supple mented with 20% FCS (HyClone Laboratories, Inc., Logan, UT), 2 IHML-glutamine (Gibco), 100 units/ml penicillin, 100 ng/ml strepto mycin (Gibco), 25 mM 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid buffer (Sigma Chemical Co., St. Louis, MO), and 2 g/liter Na- HCOa in a humidified 5% CO2-95% ambient air atmosphere at 37°C. Generation of Human T-Cell Leukemia in SCID Mice. Five x 10' to 5x 10' human MT-ALL cells were washed with phosphate buffered 1314 Research. on January 4, 2015. © 1992 American Association for Cancer cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Successful Treatment of Human Acute T-Cell Leukemia in SCID Mice Using the Anti-CD7-deglycosylated...

[CANCER RESEARCH 52, 1314-1321, March I. 1992]
Successful Treatment of Human Acute T-Cell Leukemia in SCID Mice Using theAnti-CD7-deglycosylated Ricin A-Chain Immunotoxin DA71
Burkhard Jansen, Daniel A. Vallera, Waclaw B. Jaszcz, Dat Nguyen, and John H. Kersey2
Bone Marrow Transplantation Program [D. A. V., J. H. K.¡and Departments of Laboratory Medicine/Pathology [B. J., W. B. ]., D. N., J. H. K.], Pediatrics [J. H. K.J, and Therapeutic Radiology; Section on Experimental Cancer Immunology [D. A. V.], University of Minnesota, Minneapolis, Minnesota 55455
ABSTRACT
The study of new therapeutic approaches for refractory human leukemia has been hampered by the lack of relevant in vivo models withdisseminated disease, particularly T acute lymphoblastic leukemia (T-ALL). In the present study we evaluated methods for establishing andtherapy of a human T-ALL cell line (MT-ALL) in 73 SCID mice. MT-ALL is a T<ell receptor a/ß+,CD3+, and CD7* leukemia cell line,
derived from a patient with refractory disease and early death. Injectionof 5 x IO7 MT-ALL cells i.v. caused disseminated human leukemia in
hematopoietic and nonhematopoietic organs in 100% of SCID mice (n =9) leading to death or terminal disease at 65 to 70 days after a uniformclinical course. To study possible therapeutic approaches for disseminated leukemia we utilized an immunotoxin, DA7, constructed by chemically linking the mouse IgG2b anti-CD7(3AlE) monoclonal antibodywhich recognizes a pan-T-cell marker expressed on almost all T-cellleukemias to deglycosylated ricin A-chain, a catalytic plant toxin andinhibitor of protein synthesis. Administration of DA7 led to >5 log killof clonogenic MT-ALL cells in vitro and selectively inhibited proteinsynthesis. DA7 was administered to mice at a dose of 10 Mg/mouse/dayfor 5 consecutive days starting 8 days after i.v. inoculation of leukemia.The immunotoxin therapy resulted in significant long term survival over348 days compared to untreated or control mice treated with anti-CD7antibody and deglycosylated ricin A-chain which were all dead by day 70(P < 0.001). Even after more than 11 months there was no evidence ofdisease in 82% of the DA7 treated animals. SCID mice given i.p.injections (n = 9) developed an i.p. tumor mass but demonstrated metastasis outside the peritoneum with disseminated leukemia in hematopoieticand nonhematopoietic organs, a finding different from most conventionalnude mouse models. The leukemia was fatal in 100% and killed theanimals at 68-95 days. SCID mice given i.p. injections of MT-ALLcompletely responded to therapy with DA7, resulting in survival of 100%of the animals (n = 10) at 216 days (/' < 0.001 compared to untreatedanimals). Anti-CD7 antibody, deglycosylated ricin A-chain, and a controlanti-melanoma immunotoxin (INDI-RIA) showed no therapeutic effect.We conclude that DA7 is an effective in vivo therapeutic agent againsthuman MT-ALL ¡nthe SCID mouse system, suggesting potential usefulness for therapy of humans with poor prognosis T-cell leukemia.
INTRODUCTION
Mice homozygous for the SCID3 mutation on chromosome
16 have a severe combined immunodeficiency caused by theinability to correctly rearrange their immunoglobulin and T-cell receptor genes (1-5). The SCID mutation leads to an
Received 10/7/91 ; accepted 12/17/91.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by Grams CA-49721, CA-21737, CA-31618, andCA-36725 from the National Cancer Institute/NIH and the Childrens CancerResearch Fund. J. H. K. is the recipient of an Outstanding Investigator Award(CA-49721) from the National Cancer Institute (CGCTR no. 69).
2To whom requests for reprints should be addressed.3The abbreviations used are: SCID, severe combined immunodeficiency; DA7,
immunotoxin constructed with deglycosylated (D) ricin A-chain toxin (A) andanti-CD7 monoclonal antibody (7); IT, immunotoxin; dgRTA, deglycosylatedricin A-chain; FACS, fluorescence-activated cell sorter; MoAb, monoclonal antibody; FCS, fetal calf serum; FITC, fluorescein isothiocyanate; ALL, acute lymphoblastic leukemia; TCR, T-cell receptor; IMDM, Iscove's modifiai Dulbecco'smedium; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis;1C«,,the concentration of DA7 inhibiting the protein synthesis to 50% of control.
inability to repair DNA damage induced by ionizing radiationas well as interfering with rearrangement of the immunoglobulin and T-cell receptor genes (6). Despite the impairment inlymphocyte differentiation, the SCID mouse hematopoieticmicroenvironment is intact (7), allowing the analysis of differentiation of the murine lymphoid system and also studies ofthe human lymphocyte development and biology after reconstitution with human lymphoid cells (8-12). SCID mice can alsobe used as a model for Epstein-Barr virus associated lympho-proliferative disorders of human B-cell origin (13-15). Acquiredimmunodeficiency syndrome (16-17), human lupus erythema-tosus (18), and human cutaneous T-cell lymphoma (19) havebeen studied in the SCID mouse. Human leukemias of non-T-cell origin have been established in SCID mice (20, 21). Recently Cesano et al. (22) reported the engraft nient of human T-ALL following i.p. injection in SCID mice. We have previouslynoted that the SCID mouse provides an excellent microenvironment for the growth of human t(4;l 1) acute leukemia (23).
In the present studies, we report a model system for humanT-ALL, established by injecting the human TCR a/ß+,CD3+,CD7+ T-ALL cell line MT-ALL (24) i.p. and i.v. into SCID
mice. In addition, we report the first successful treatment ofdisseminated human T-ALL in vivo using the anti-CD7(3AlE)-dgRTA IT, DA7, in our model system. The MoAb 3A1Erecognizes the CD7 antigen, expressed on essentially all humanT-cell leukemias and leukemia progenitors (25). RTA is a toxicportion (M, 30,000) of the plant toxin ricin derived from thecastor seed plant Ricinis communis. Currently under clinicalevaluation, RTA immunotoxins have demonstrated efficiencywhen used for therapy of steroid resistant graft versus hostdisease (26) and B-cell malignancies (27). The toxin was chemically deglycosylated to reduce its reactivity with the reticulo-endothelial system (28). In this study, therapy using DA7resulted in long term survival of SCID mice bearing disseminated human T-cell leukemia.
MATERIALS AND METHODS
Experimental Animals. Pathogen free C.B-17 (n = 70) and C3H (n= 3) seid/seid (SCID) mice, 4-7 weeks old, were obtained from theNIH-Fredrick Cancer Research Center (Fredrick, MD), from the Jackson Laboratory (Bar Harbor, ME) or from Taconic Farms, Inc. (Ger-mantown, NY). The animals were housed in laminar flow racks ormicroisolator cages under specific pathogen free conditions and received autoclaved food, water and an antibiotic prophylaxis with sul-famethoxazole and trimethoprim.
Tumor Cells. The previously described human TCR a//8*, CD3%CD7+ T-ALL cell line (MT-ALL) (24) with potential for multilineage
differentiation, derived from a patient with refractory disease, wasmaintained in IMDM (Gibco Laboratories, Grand Island, NY) supplemented with 20% FCS (HyClone Laboratories, Inc., Logan, UT), 2IHML-glutamine (Gibco), 100 units/ml penicillin, 100 ng/ml streptomycin (Gibco), 25 mM 4-(2-hydroxyethyl)-l-piperazineethanesulfonicacid buffer (Sigma Chemical Co., St. Louis, MO), and 2 g/liter Na-HCOa in a humidified 5% CO2-95% ambient air atmosphere at 37°C.
Generation of Human T-Cell Leukemia in SCID Mice. Five x 10' to5 x 10' human MT-ALL cells were washed with phosphate buffered
1314
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

TREATMENT OF A HUMAN ACUTE T-CELL LEUKEMIA IN SCID MICE
saline and injected i.v. in the lateral tail vein or i.p. into SCID mice.Leukemia bearing animals were sacrificed in their terminal stages ofdisease (2-4 days prior to expected death).
Flow Cytometric Analysis and Histology. For the preparation of viablecell suspensions, organs were cut into small pieces and minced througha metal screen. Leukemia and hematopoietic mouse cells of freshlyexcised organs and tissues were separated on a Ficoll-Hypaque densitygradient and stained with FITC labeled anti-CD45 antibody (BectonDickinson, Mountainview, CA). The fluorescence patterns were analyzed by using a FACStar Plus (Becton Dickinson) flow cytometer. Thepercentage of positive cells was determined by subtracting the bindingmeasured following staining with an isotype control from the bindingmeasured following staining with anti-CD45 antibody for each experimental sample. Excised tissues were fixed in 5% buffered formalin,routinely processed and embedded in paraffin, sectioned, and stainedwith hematoxylin and eosin for histopathological examination.
Preparation of the Anti-T-Cell Immunotoxin DA7. The anti-CD7(3A 1E) MoAb was secreted by the hybridoma cell line 3A l E 12H7,generously provided by Dr. Bart Haynes, Duke University, Durham,NC. Anti-CD7(3AlE) MoAb, a mouse IgG2b, was purified fromBALB/c ascites fluid. It was concentrated by ammonium sulfate (50%)precipitation and dialyzed against 0.01 mol/liter sodium phosphate(pH 7.5) and 0.15 mol/liter sodium chloride. The dialysate was appliedto a high performance liquid chromatography protein A column (Bio-Rad, Richmond, CA) and eluted with citrate, pH 3.5. The dgRTA waspurchased from Inland Laboratories (Austin, TX). DA7 was producedby linking the purified MoAb to dgRTA. The purification and conjugation technique has been described previously in detail (29, 30). Inbrief, the MoAb was activated with Ar-succinimidyl-3-(2-pyridyldi-thio)propionate and then reacted with dithiothreitol-treated dgRTA.The IT was purified by high performance liquid chromatography on aTSR G3000SW size exclusion column. The modification in the carbohydrate of the IT was performed as described previously (31). Thefinal product was suspended in 0.15 mol/liter sodium phosphate-0.1mol/liter sodium chloride (pH 7.5) at a concentration of approximately1.0 mg/ml. The purity of DA7 was assessed by SDS-PAGE. As controlsthe anti-melanoma IT (IND1-RTA) (Xoma Co., Berkeley, CA) andanti-Thy-1.2-RTA IT (anti-mouse pan T-cell) (32) linked by the sameprocedure were used.
Gel Analysis. Analysis was performed on a 4-20% SDS-PAGE plate(Daiichi Pure Chemicals Co., Tokyo, Japan). Five Mg of IT weresuspended in sample buffer (40 mmol/liter Tris buffer, pH 6.8-1.5%SDS-7.5% glycero-0.005% bromophenol blue), boiled for 5 min, andadded to the plate. Electrophoresis was performed using a constantcurrent (28 mA) for a running time of 100 min in electrode buffer(0.025 mol/liter Tris buffer-0.192 mol/liter glycine-0.1% SDS). Theplates were stained with Coomassie blue, dried, and scanned using aDu-62 spectrophotometer with gel scanning accessory (Beckman Instruments, Fullerton, CA). The analysis was performed at 595 nm.Quantitation of the bands was performed by densitometric analysisusing the Gel Scan Soft Pack Module Software Package (BeckmanInstruments).
Binding Studies. The binding of DA7, anti-CD7(3AlE) MoAb, orthe control anti-Thy-1.2-RTA IT to MT-ALL cells was determined bystandard indirect immunofluorescence techniques. For in vitro analysis,IO6 MT-ALL cells were suspended in phosphate buffered saline con
taining 5% fetal calf serum and 0.01% sodium azide. The cells wereincubated with varying concentrations (0.0001-10 Mg/ml) of MoAb orIT for 30 min at 4°Cwith 50 ¿ilof a 1:20 dilution of FITC conjugated
goat anti-mouse IgG. Cells were washed three times, diluted in bufferas described above, and analyzed using a FACScan (Becton Dickinson)flow cytometer. The percentage of positive fluorescence was determinedfrom histograms by subtracting background binding determined byreacting FITC labeled goat anti-mouse IgG with control ascites fluidtreated MT-ALL cells from experimental samples.
Protein Synthesis Inhibition Assay. Our protein synthesis assay hasbeen reported previously (33). MT-ALL cells were incubated with 10-30,000 ng/ml of DA7 or anti-Thy-1.2-RTA control IT at 37°Cfor 8 h
with or without the addition of a blocking concentration (150 Mg/ml)of anti-CD5 (T101) or anti-CD7 (3A1E) MoAb for 30 min at 4°Cin
RPMI 1640 in the presence of 20 mmol NH4CI. After three washingswith leucine free RPMI 1640, cells were incubated overnight in fiatbottomed 96-well plates (Costar, Cambridge, MA) in leucine free RPMI1640 containing 20% dialyzed FCS and 1% penicillin/streptomycin.The cells were then pulsed for 6 h at 37T with 1 MCi [3H]leucine
(Amersham, Arlington Heights, IL) per well. The pulsed cells wereharvested onto glass filter paper using an automated harvester (Brandel,Gaithersburg, MD). The filter paper was dried and the [3H]leucine
incorporated into the cells was determined by using standard scintillation counting techniques. The results were expressed as percentages ofthe untreated control.
Clonogenic Assay. Neoplastic target cells were assayed for clonogenicgrowth as described previously (34). In brief, MT-ALL cells wereincubated for 8 h in IMDM at 37°C.The incubation media contained
DA7 or IND1-RTA IT at concentrations of 0.01-100 Mg/ml. Cellswere washed and plated in limiting dilution (dilution factor a = 5) inIMDM supplemented with 20% FCS, 2 mM L-glutamine , 100 units/ml penicillin, and 100 Mg/ml streptomycin in a humidified 5% CO2-95% ambient air atmosphere at 37°C.Two hundred M' from each
dilution were placed into 6 replicate wells. After 14 days in cultureunder standard conditions, the number of wells showing growth (colonies >20 cells ) was counted using an inverted phase microscope. Thefrequency of wells with growth was used to estimate the most probablenumber of remaining clonogenic cells in the original suspension. Leukemia cell elimination by DA7 or control IT was expressed as log kill.
Tumor Treatment with Immunotoxins. In the first series of experiments animals given i.p. injections of 5 x IO7 MT-ALL cells were
treated with 5 consecutive daily i.p. injections of 10 MgDA7 (total of 3mg/kg). The treatment was started 1 day after i.p. leukemia challenge.Based on encouraging results of the i.p. studies combined with ourprevious experience with in vivo administration of RTA IT in murinemodels (32, 35), SCID mice given i.v. injections of 5 x IO7 MT-ALL
cells were treated i.v. with the same regimen beginning at a later timepoint (day 8 after leukemia inoculation). Equivalent amounts of uncon-jugated anti-CD7(3AlE) MoAb mixed with dgRTA or anti-humanmelanoma IT were used as controls. The survival data were analyzedusing log rank statistical methods (36).
RESULTS
Characteristics of Growth of the Human MT-ALL after i.v.Injection into SCID Mice
Injection of 5 x IO7 cells of the human TCR a/ß+,CD3+,CD7+ T-ALL leukemia cell line MT (MT-ALL) i.v. caused
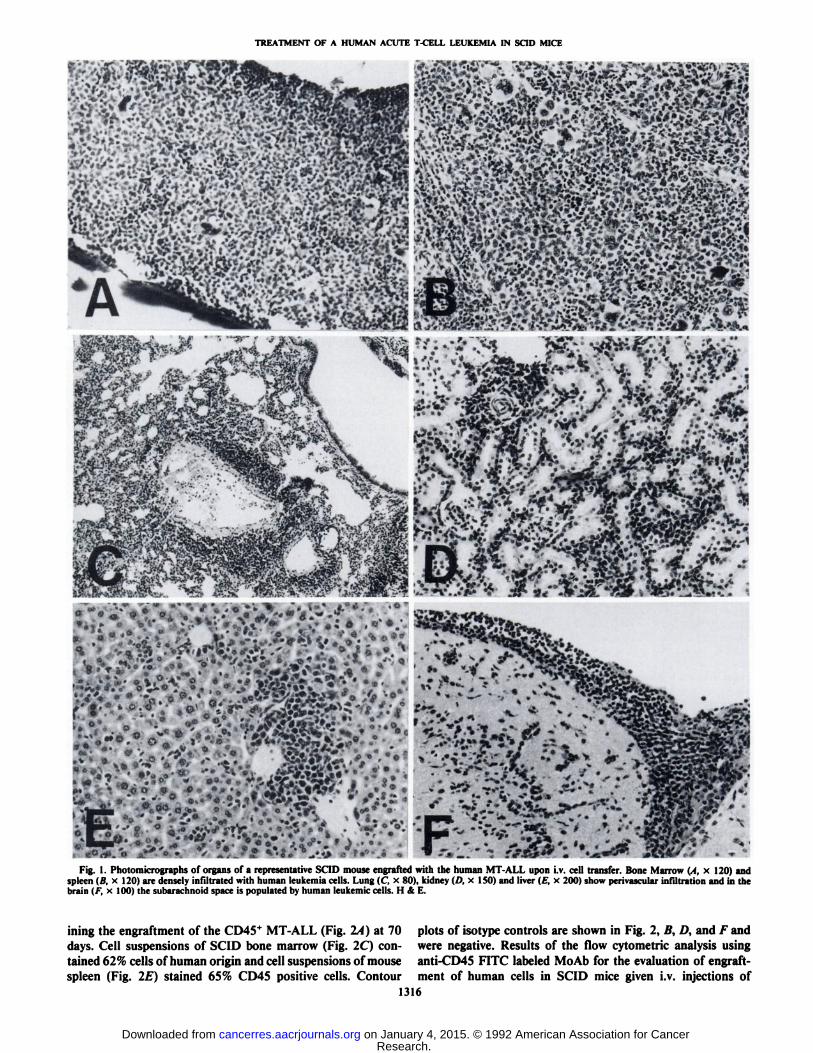
disseminated human leukemia in 9 of 9 (100%) SCID mice andled to the death or terminal disease of the animals after 65-70days. Clinically the animals showed weight loss, rough fur, ahumped back, and paralysis of the hind legs. The leukemia wasdetected by flow cytometric analysis of cell suspensions ofmouse organs using FITC labeled anti-CD45 antibody and byhistopathology as early as 51 days after injection of leukemiacells. Fig. 1 shows involved mouse organs of a representativepreterminal animal sacrificed at 70 days. The bone marrow(Fig. IA) showed extensive infiltration of leukemia cells andthe splenic structure (Fig. \B) was obscured by the diffuseaccumulation of MT-ALL cells. The lungs showed massiveMT-ALL infiltrates around the pulmonary vessels (Fig. 1C).Similar infiltrates were present in subpleural areas and also inthe alveolar walls. The kidney (Fig. \D) had perivascular aswell as dispersed interstitial leukemic cell infiltrates. The liver(Fig. If) showed human leukemia cells in the portal spaces, inthe sinuses, and around the central veins. The brain (Fig. IF)showed massive subarachnoid infiltrates of leukemic cells; smallfoci of perivascular infiltrates in the superficial parts of thecortex could also be detected. Also lymph nodes and blood wereextensively involved. Fig. 2 shows FACS contour plots exam-
1315
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from
TREATMENT OF A HUMAN ACUTE T-CELL LEUKEMIA IN SC1D MICE
S^^SSft ' l^u So? •¿�«-'
'.i.&a»&:i^ >¿»*v.J¡f* .-¿F«/
Fig. 1. Photomicrographs of organs of a representative SCID mouse engrafted with the human MT-ALL upon i.v. cell transfer. Bone Marrow (A, x 120) andspleen (B, x 120) are densely infiltrated with human leukemia cells. Lung (C, x 80), kidney (/'. x ISO) and liver (/•:.x 200) show perivascular infiltration and in thebrain I/, x 100) the subarachnoid space is populated by human leukemic cells. H & E.
ining the engraftment of the CD45+ MT-ALL (Fig. 14) at 70 plots of isotype controls are shown in Fig. 2, A, D, and F and
days. Cell suspensions of SCID bone marrow (Fig. 2C) con- were negative. Results of the flow cytometric analysis usingtained 62% cells of human origin and cell suspensions of mouse anti-CD45 FITC labeled Mo Ab for the evaluation of engraft-
spleen (Fig. 2E) stained 65% CD45 positive cells. Contour ment of human cells in SCID mice given i.v. injections of1316
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

TREATMENT OF A HUMAN ACUTE T-CELL LEUKEMIA IN SCID MICE
"MT Call Line100% B
"MT-Cell Line0%
Antl-CD45 FITC
Bone Marrow62%
Control FITC
Bone Marrow0%
Antl-CD45 FITC
Spleen65%
Control FITC
ìè&c?Spleen
0%Control
FITCAntl-CD45 FITC
Fig. 2. Contour plots from flow cytometric analysis examining the engraftmentof the human MT-ALL in a representative SCID mouse. In vitro grown MT-ALL cells and cell suspensions of bone marrow and spleen from a SCID mousegiven i.v. injections of human leukemia and sacrificed in its terminal stage ofdisease were analyzed by immunofluorescence using the MoAb anti-CD45 FITC(A, C, E). Isotype controls were used (B, D, F).
human MT-ALL revealed 28-75% human cells in the mousebone marrow, 13-65% human cells in the spleen, and 29-64%human cells in the liver of animals sacrificed in the terminalstage at 65-70 days.
In additional experiments we evaluated the number of cellsnecessary to cause disseminated human leukemia in SCID mice.Groups of three animals given i.v. injections of 5 x IO6, 5 xIO5, and 5 x IO4 MT-ALL cells developed terminal human
leukemia after 60 ±5 days, 78.7 ±3.3 days, and 79.7 ±3.9days (median survival time ±SE). All animals in these groupsdeveloped terminal human leukemia within 85 days. One ofthree mice given injections of 5 x IO3MT-ALL cells developed
terminal leukemia after 91 days.
Characteristics of Growth of the Human MT-ALL after i.p.Injection into SCID Mice
Of the animals given i.p. injections of 5 x IO7 human MT-
ALL cells, 9 of 9 (100%) developed progressive tumors whichled to terminal disease after 68-95 days. The leukemia grew asa bulky tumor in the peritoneal cavity and displaced and invadedabdominal organs, kidney, and mediastinum. The leukemia wasalso detectable in later stages of the disease in bone marrow,spleen, lymph nodes, lung, brain, and peripheral blood with adistribution pattern similar to the one observed in animals giveni.v. injections. Clinically, the i.p. injected leukemia did notcause the paralysis noted in the animals that received leukemiaupon i.v. transfer. Cell suspensions from organs of animalssacrificed in the terminal stages of the disease showed evidenceof human cells in the bone marrow (4-50%), in the spleen (10-
82%), and in the liver (15-93%) based on flow cytometricanalysis using the FITC labeled human MoAb anti-CD45.
Studies of Immunotoxin Effects on MT-ALL in Vitro
Gel Analysis of DA7. The SDS-PAGE gel profile of DA7revealed five major IT bands exceeding M, 150,000 (MoAb).Scanning showed that the largest bands (on a percentage basis)for the IT were likely 2:1 (toxin:MoAb) species at M, 210,000(28%) and 3:1 species at MT 240,000 (33%). Smaller amountsof Mr 180,000, A/r 270,000, and A/r 300,000 likely representing1:1, 4:1, and 5:1 species, respectively, were present (9-18%).Gel scanning revealed <2% free MoAb and <0.5% free dgRTA(data not shown).
Binding of anti-CD7 antibody or DA7 to Target Cells in Vitro.The binding of unconjugated anti-CD7 (3A IE) MoAb or DA7to MT-ALL cells in vitro was determined by indirect immunofluorescence at various protein concentrations (Table 1). TheIT and the MoAb bound to human MT-ALL cells in a similardose dependent fashion reaching 100% binding (saturation) ata concentration of l ßg/mlof the respective reagent. The meanfluorescence intensity of MT-ALL cells bound to the MoAb orthe IT was also dose dependent and comparable (160 and 121,respectively, at concentrations of 1 ng/ml). The control anti-Thy-1.2-RTA IT showed only 1-4% binding.
Effect of DA7 on the Clonogenic Growth of Human MT-ALLCells. To determine the potency of DA7, MT-ALL cells wereincubated with various concentrations of IT for 8 h at 37°Cand
then plated in replicates in the absence of NH4C1 potentiation.Comparison of the colony counts for DA7 treated versus non-treated controls gave a >5.6 log (SE < 0.3) kill of clonogenicMT-ALL cells on day 14 for concentrations of 10, 30, and 100Mg/ml of IT (Table 2). The control anti-melanoma IT led to 0.7log killing of clonogenic MT-ALL cells at concentrations of 30and 100 Mg/ml. In separate experiments DA7 was found to beeffective against the human TCR y/o+ cell line Jurkat. A 3.7
log (SE = 0.2) killing of clonogenic Jurkat cells was observed
Table 1 Binding of DA7 to MT-ALL cells in vitro according to flow cytometricanalysis
The binding of DA7, anti-CD7(3AlE) MoAb, or the control anti-Thy-1.2-RTA IT to MT-ALL cells was determined by standard indirect immunofluorescence techniques as described in "Materials and Methods." The percentage of
positive fluorescence was determined from histograms by subtracting backgroundbinding determined by reacting FITC labeled goat anti-mouse IgG with controlascites fluid treated MT-ALL cells from experimental samples.
BindingofPBS-controlDA73A1EAnti-ThyI.2-RTAConcentration(fg/mi)-0.00010.0010.010.11100.00010.0010.010.11100.00010.0010.010.1110%of cells
positive2231994100100322195100100142111Mean
channelfluorescence
intensity89111640121ISO1822134516016610126888
1317
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

TREATMENT OF A HUMAN ACUTE T-CELL LEUKEMIA IN SCID MICE
Table 2 In vitro anti-Ieukemic activity of DA7 agonist clonogenic MT-ALL cells
Neoplastic target cells were assayed for clonogenic growth as described in"Materials and Methods." After 14 days in culture under standard conditions,
the number of wells showing growth (colonies >20 cells) was counted using aninverted phase microscope. The frequency of wells with growth was used toestimate the most probable number of remaining clonogenic cells in the originalsuspension. Leukemia cell elimination by DA7 or control IT was expressed aslog kill.
ImmunotoxintreatmentNoneDA7IND1-RTAConcentration(Mg/ml)-0.01
0.1110
3010030
100Log-kill
(SE <0.3)-0.7
0.72.2
>5.6>5.6>5.60.7
0.7
Table 3 Effect of DA7 on protein synthesis of MT-ALL cells as measured bytritimeli leucine incorporation
MT-ALL cells were incubated with various amounts of DA7 or anti-Thy-1.2-RTA control IT at for 8 h with or without the addition of a blocking concentration(150 (ig/ml) of anti-CD5(T101) or anti-CD7(3AlE) MoAb for 30 min in thepresence of 20 mmol NH4CI. After three washings with leucine-free RPMI 1640,cells were incubated overnight, pulsed for 6 h with [3H]leucine and then harvestedand counted as described in "Materials and Methods." The cpm/well were
determined from triplicate samples and results were expressed as percentages ofthe protein synthesis of the untreated control.
Protein synthesis (% ofcontrol)Concentration
ofIT(ng/ml)10100
1,00010,00030,000DA73
2111DA7
blockedwith anti-CD786
736027
1DA7
blockedwith anti-CD55
1111Thy-1.2-RTA68
64706366
at a concentration of 10 /ug/ml of IT in the absence of NH4C1potentiation.
In Vitro Protein Synthesis Inhibiting Activity of DA7. Toevaluate the cytotoxic potential of DA7 we incubated MT-ALLcells with various concentrations of IT for 8 h at 37°C(Table
3). At concentrations of 10, 100, 1,000,10,000, and 30,000 ng/ml DA7 reduced the protein synthesis activity in the presenceof 20 mmol NH4C1 to 3, 2, 1, 1, and 1% of the untreatedcontrol, respectively. The IC50 was <10 ng/ml. MT-ALL cellsincubated for 30 min at 37'C with 150 ¿ig/mlanti-CD7(3AlE)
MoAb before adding DA7 showed an IC50 of 4000 ng/ml,suggesting a more than 400-fold decreased protein synthesis inunblocked versus specifically blocked MT-ALL cells treatedwith DA7. Blocking with anti-CD5 (T101) MoAb had no effecton the protein synthesis inhibition (ICso < 10 ng/ml). Whentreated with the same levels of a control IT, anti-Thy-1.2-RTA,the protein synthesis of MT-ALL cells as compared to untreated controls was 63-70%. Thus, the IC50 for anti-Thyl.2-RTA was at least 3,000 times higher than the IC50 for DA7.
Effects of DA7 in SCID Mice with Human MT-ALL Givenby i.v. Injection. We next studied the effect of DA7 on thecourse of MT-ALL in SCID mice. The animals were given i.v.injections of human MT-ALL cells and treated i.v. (n = 11) for5 consecutive days with a daily dose of 10 ng of DA7 startingon day 8 after injection of cells. Two of these treated animalswere sacrificed with terminal human leukemia 132 days afterinjection of cells. The remaining 9 animals showed no evidenceof disease 348 days after injection of MT-ALL (Fig. 3). In acontrol group of animals receiving no treatment, all died before
70 days of leukemia determined by histology and flow cytome-tric analysis using anti-CD45 antibody. In another controlgroup 5 leukemia bearing animals were treated with a mixtureof unconjugated anti-CD7(3AlE) MoAb and dgRTA toxin atcomparable concentrations; all animals in this control groupdied within 70 days of human leukemia with an organ involvement and a distribution pattern of the tumor identical to untreated animals. The product limit estimate and standard errorof the probability of survival at 348 days post-i.v. MT-ALLchallenge was 82 ±12% for DA7 treated animals. This wassignificantly better than the O ±0% of untreated control mice(P < 0.001) or the O ±0% of control mice treated with amixture of anti-CD7(3AlE) MoAb and dgRTA (P < 0.001).Spleen weights were measured as indicators of tumor involvement. The mean spleen weight ±SD of the mean in SCIDrecipients bearing established human leukemia post-i.v. MT-ALL challenge was 0.10 ±0.03 g, higher than the mean spleenweight of 0.06g ±0.04 g of animals treated with DA7 (P =0.16).
Effects of DA7 in SCID Mice with Human MT-ALL Givenby i.p. Injection. In additional studies which also tested the invivo effect of DA7, SCID mice that received i.p. injections ofMT-ALL cells were given i.p. injections of a daily dose of 10tig IT for 5 consecutive days starting 1 day after injection ofcells. All treated animals (10 of 10) survived until time ofsacrifice at 216-226 days (Fig. 4). Two of the 10 IT treatedanimals showed some involvement of the bone marrow (5 and6% human cells) detected by flow cytometric analysis usinganti-CD45 antibody. The remaining eight animals showed nodetectable tumor at time of sacrifice. The survival time inuntreated animals given i.p. injections was 68-95 days and allanimals died of tumor. Mice given i.p. injections of controlanti-melanoma IT developed terminal disease caused by humantumor in 10 of 10 animals (100%) after 82-112 days (Fig. 4).In an additional control group, SCID mice received the samequantity of unconjugated anti-CD7(3AlE) MoAb and dgRTAtoxin as treatment. Four of five animals died of human tumorafter 97-111 days. One mouse showed no evidence of disease216 days after injection of MT-ALL cells (Fig. 5). The productlimit esimale and SE of the probability of survival at 216 dayspost-i.p. injection of MT-ALL was 100 ±0% for DA7 whichwas significantly better than the O ±0% of all control groups(P< 0.001 for the treated group compared to any of the controlgroups). Also, spleen weights were measured as indicators oftumor involvement. The mean spleen weight ±SD of the mean
1.0
| 0.8-
£« 0.6-
0.4
OJO60 120
Days
180 240 300 360
Fig. 3. Survival curves of SCID mice given i.v. injections of human MT-ALLand treated i.v. with DA7. The product limit estimate and standard error of theprobability of survival at 348 days post i.v. MT-ALL challenge was 82 ±12% forDA7 treated mice ( , n = 11) which was significantly better than the O ±0% survival (median survival time, 68.9 days) of untreated control mice ( , n= 9) (P < 0.001) or the O ±0% survival (median survival time, 56.4 days) ofcontrol mice ( , n —¿�5) treated with a mixture of unconjugated 3A l E MoAband dgRTA (P < 0.001). DA7 treated animals received a dosage of 5 x 10 MgofIT starting on day 8 after injection of 5 x 10' MT-ALL cells.
1318
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

TREATMENT OF A HUMAN ACUTE T-CELL LEUKEMIA IN SCID MICE
1.0
o.e
oo 0.2C
0.060 120 180 240
Days
Fig. 4. Survival curves of SCID mice given i.p. injections of human MT-ALLand treated i.p. with DA7. The product limit estimate and SE of the probabilityof survival at 216 days after i.p. MT-ALL challenge was 100 ±0% for DA7treated mice ( , n = 10) which was significantly better than the O ±0%survival (median survival time, 82.5 days) of untreated control mice ( , n = 9)(P < 0.001), the O ±0% survival (median survival time, 103.8 days) of controlmice ( , n = 5) treated with a mixture of unconjugated anti-CD7(3A 1E)MoAb and dgRTA (P < 0.001) or the O ±0% survival (median survival time,95.4 days) of control mice ( , n = 10) treated with the control anti-humanmelanoma IT INDI (P < 0.001). DA7 treated animals received a dosage of 5 x10 »gof IT starting 1 day after injection of 5 x 10' MT-ALL cells.
in i.p. injected SCID recipients bearing human leukemia was0.20 ±0.03 g, significantly higher than the mean spleen weightof 0.11 ±0.06 g of animals treated with DA7 (P < 0.001).
DISCUSSION
The major findings to emerge from this study are: (a) establishment of a human T-ALL (MT-ALL) in SCID mice resultingin a reproducible animal model of disseminated human leukemia; (b) demonstration that our anti-CD7(3AlE)-dgRTA IT,DA7, had a marked in vitro effect on MT-ALL cells; and (c)the same IT was very effective in vivo, even when administeredsystemic-ally, leading to significant long term survival of IT
treated leukemia bearing animals.SCID mice given injections of human MT-ALL developed
disseminated disease with multiple organ involvement as seenin humans with T-cell leukemias. The establishment of humanleukemias in immunodeficient animals, particularly athymicnude mice, has been variable and difficult, often requiringirradiation and the coinjection of feeder cell lines (37, 38). Thelocalized tumor growth in these in vivo systems is artificial anddoes not reflect the distribution pattern of leukemia in humans,limiting the usefulness of nude mouse models for studies ofbiology and therapy of human leukemia. Thus, our model ofdisseminated T-cell leukemia, using the SCID mouse, an animaleven more immunodeficient than the nude mouse, has important advantages.
Our SCID mouse model provides the opportunity for testinga variety of new therapeutic agents, including immunotoxins.Immunotoxins vary considerably in their killing efficiency towards antigen positive cells. The absolute amount of antibodyinternalized was found to be more important in predicting theefficacy of ITs than the degree of antigen expression on targetcells. In an earlier study investigators (39) showed that anti-CD? had the fastest internalization rate followed by anti-CD3and anti-CD5. Anti-CD4 and anti-CD8 hardly internalized andshowed little or no cytotoxic effect even on cell lines with ahigh expression of the corresponding antigen. Efficient inhibition of protein synthesis was seen with anti-CD3, anti-CD5,and anti-CD7. On the basis of these in vitro studies and thenegative in vivo results using anti-CD5 (T101) ricin A chain IT(40), we chose to evaluate the therapeutic efficacy of an ITpreparation in which the anti-CD7(3AlE) MoAb is linked to a
deglycosylated ricin A chain. CD7, a pan-T-cell marker expressed on basically all T-cell leukemias, is expressed on 100%of MT-ALL target cells. We have studied and used intact ricinIT in clinical studies for in vitro bone marrow purging (34, 41,42). However, the intact toxin is unsuitable for systemic in vivouse. Removal of the B-chain (the native binding portion) andlinkage of the A-chain to antibody has shown clinical promise(43, 44). Deglycosylation of the ricin A chain renders a reagenteven more useful for in vivo application, since deglycosylatedIT are not removed as readily from the circulation by nonpar-enchymal cells of the liver that have receptors for the mannoserich and fucose containing oligosaccharides on the A-chain (28,45). Immunotoxins containing native A-chain were shown toclear 2-3-fold more rapidly from the bloodstream than thosecontaining dgRTA (46).
The gel analysis indicated that generally two or three dgRTAmolecules were linked to one MoAb molecule in DA7. Theconjugation of anti-CD7(3AlE) MoAb to dgRTA had no adverse effect on the ability of the MoAb to recognize the CD7antigen on MT-ALL cells. In vitro studies revealed that the ITwas effective in the killing of clonogenic MT-ALL and Jurkatcells. DA7 also selectively inhibited protein synthesis of MT-ALL cells.
Several publications discuss the in vivo usage of ITs for thetherapy of human leukemias established as localized tumors innude mice (47-50). Ricin or ricin A-chain have been usedfrequently as toxic components of immunotoxins (43-44). Onlyfew in vivo studies in humans have been done. Laurent et al.(40) injected a ricin A-chain containing IT into two leukemiapatients: one with acute T-lymphoblastic leukemia; and theother with chronic B-lymphocytic leukemia. The dose was low
and the IT was not effective in either patient. Until recently,preclinical studies were hampered by the lack of suitable animalmodels for disseminated human leukemia. The SCID mousehas been shown to provide an efficient and reproducible modelto study the pathogenesis of human leukemia and was introduced as a potentially useful system for preclinical studiesevaluating new therapeutic modalities (20-22). Studies evaluating the therapeutic efficacy of DA7 in our newly establishedmodel system were encouraging. More than 80% long termsurvival in mice given i.v. injections of human MT-ALL cellsand 100% long term survival in animals given i.p. injectionscould be observed after treatment with DA7. Only 2 of 11animals given injections and treated i.p. showed minimal residual disease in the bone marrow detected by flow cytometricanalysis; none of the i.v. injected and treated long term survivorsshowed detectable residual disease. The therapeutic effect wascaused by the intact IT; a mixture of anti-CD7(3AlE) MoAband dgRTA had no beneficial effect on long term survival orreduction of tumor mass. The irrelevant anti-melanoma ITIND1-RTA showed no significant killing of clonogenic MT-ALL cells in vitro and had only minimal effect on the survivaltime of leukemia bearing animals leading to no long termsurvival. Titration experiments indicated that as few as 5 x IO3MT-ALL cells caused disseminated human leukemia and thatthere is a time-dose effect for the development of the leukemiain SCID mice. These results suggest an in vivo killing of humanleukemia by DA7 in the range of several logs.
The SCID mouse system allows the evaluation of the therapeutic efficacy of immunotoxins without the interference ofanti-IT antibodies, a potential problem in clinical use whichwill require exact investigation. This is to our knowledge thefirst report of a reproducible animal model for disseminated
1319
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

TREATMENT OF A HUMAN ACUTE T-CELL LEUKEMIA IN SCID MICE
human T-cell leukemia upon i.v. cell transfer, the first successfuluse of an IT directed against CD7 in vivo, and the first successfuluse of any IT in an animal model with disseminated human T-ALL. DA7 is a potentially useful agent for the therapy ofhumans with refractory T-cell leukemia.
ACKNOWLEDGMENTS
We want to thank Jennifer Mattern and Tuyet Nguyen for excellenttechnical assistance; Lenise Burch and Michael Hupke for FACSanalysis; and Samuel Pirruccello, University of Nebraska, KazimieraGajl-Peczalska, Chien Shing Chen, and Bruce Bejcek for helpful discussions. We also thank members of the Immunophenotyping Laboratory for technical assistance; Andrew Sicheneder for DA7 synthesis;and Barton Haynes, Duke University, Durham, NC, who generouslyprovided the hybridoma cell line 3A1E 12H7.
REFERENCES
1. Bosma, G. <'.. Custer, R. P., and Bosma, M. J. A severe combined immu
nodeficiency in the mouse. Nature (Lond.), 301: 527-530, 1983.2. Bosma, G. C, Davisson. M. T., Reutsch, N. R., Sweet, H. O., Schultz, L.
I ).. and Bosma, M. J. The mouse mutation severe combined immune deficiency (SCID) is on chromosome 16. Immunogenetics, 29: 54-57, 1989.
3. Schuler, W., Weiler, I. J., Schuler, A., Phillips, R. A, Rosenberg, N., Mak,T. W., Kearney, J. F., Perry, R. P., and Bosma, M. J. Rearrangement ofantigen receptor genes is defective in mice with severe combined immunedeficiency. Cell, 46: 963-972, 1986.
4. Malynn, B. A., Blackwell, T. K., Fulop, G. M., Rathbun, G. A., Furley, A.J. W., Ferner, P., Heinke, L. B., Phillips, R. A., Yancopoulos, G., and Alt,F. W. The SCID defect affects the final step of the immunoglobulin VDJrecombinase mechanism. Cell, 54: 453-460, 1988.
5. Lieber, M. R., Hesse, J. E., Lewis, S., Bosma, G. C., Rosenberg, N, Mizuuchi,K., Bosma, M., and Geliert, M. The defect of murine severe combinedimmune deficiency: joining of signal sequences but not coding sequences inV(D)J recombination. Cell, 55: 7-16, 1988.
6. Fulop, G. M., and Phillips, R. A. The SCID mutation in mice causes ageneral defect in DNA repair. Nature (Lond.), 347: 479-482, 1990.
7. Dorschkind, K., Keller, G. M., Phillips, R. A., Miller, R. G., Bosma, G. C,O'Toole, M., and Bosma, M. J. Functional status of cells from lymphoid
and myeloid tissues in mice with severe combined immunodeficiency disease.J. Immunol., 132: 1804-1808, 1984.
8. Fulop, G. M., Wu, D. D., and Phillips R. A. The SCID mouse as a model toidentify and quantify myeloid and lymphoid stem cells. Curr. Top. Microbio!.Immunol.. 152: 173-180, 1989.
9. Dorshkind, K. Use of the SCID mouse transplantation system in studies oflymphocyte differentiation. Curr. Top. Microbio!. Immunol., 152: 169-172,1989.
10. McCune, J. M., Namikawa, R., Kaneshima, H., Shultz, L. D., Lieberman,M., and Weissman, I. L. The SCID-hu mouse: murine model for the analysisof human hematolymphoid differentiation and function. Science (Washington DC), 241: 1632-1639, 1988.
11. Mosier, D. E., Gulizia, R. J., Baird, S. M., and Wilson, D. B. Transfer of afunctional human immune system to mice with severe combined immunodeficiency. Nature (Lond.), 335: 256-259, 1988.
12. Mosier, D. E. Immunodeficient mice xenografted with human lymphoidcells: new models for in vivostudies of human immunobiology and infectiousdiseases. J. Clin. Immunol., 10: 185-191, 1990
13. Purtilo, D. T., Falk, K., Pirruccello, S. J., Nakamine, H., Kleveland, K.,Davis, J. R., Okano, M., Taguchi, Y., Sanger, W. G., and Beisel, K. W.SCID mouse model of Epstein-Barr virus induced lymphomagenesis ofimmunodeficient humans. Int. J. Cancer, 47: 510-517, 1991
14. Rowe, M., Young, L. S., Crocker, J., Stokes, H.. Henderson, S., and Rick-inson, A. B. Epstein-Barr virus (EBV)-associated lymphoproliferative diseasein the SCID mouse model: implications for the pathogenesis of EBV-positivelymphomas in man. J. Exp. Med., 173: 147-158, 1991.
15. Cannon, M. J., Pisa, P., Fox, R. I., and Cooper, R. I. Epstein-Barr virusinduces aggressive lymphoproliferative disorders of human B cell origin inSCID/hu chimeric mice. J. Clin. Invest., «5:1333-1337, 1990.
16. Namikawa. R., Kaneshima, H., Lieberman, M., Weissman, I. L., and McCune, J. M. Infection of the SCID-hu mouse by HIV-1. Science (WashingtonDC), 242: 1684-1687, 1988.
17. McCune, J. M., Namikawa, R., Shih, C-C, Rabin, L., and Kaneshima, H.Suppression of HIV infection in AZT-treated SCID-hu mice. Science (Washington DC), 247: 564-566, 1990.
18. Duchosal, M. A., McConahey, P. J., Robinson, C. A., and Dixon, F. J.Transfer of human systemic lupus erythematosus in severe combined immunodeficient (SCID) mice. J. Exp. Med., 172:985-988, 1990.
19. Charley, M. R., Tharp, M., Locker, J., Deng, J-S., Golsen, J. B., Mauro, T.,McCoy, P., Abell, E., and Jegasothy, B. Establishment of a human cutaneous
T-cell lymphoma in C.B-17 SCID mice. J. Invest. Dermatol., 94: 381-384,1990.
20. Kamel-Reid, S., Letarte, M., Sirard, C., Doedens, M., Grunberger, T., Fulop,G., Freedman, M. H., Phillips, R. A., and Dick, J. E. A model of humanacute lymphoblastic leukemia in immune deficient SCID mice. Science(Washington DC), 246: 1597-1600, 1989.
21. Ghetie, M-A., Tucker, T., Jones, D., Uhr, J. W., and Vitetta, E. S. Disseminated or localized growth of a human B-cell tumor (Daudi) in SCID mice.Int. J. Cancer, 45: 481-485, 1990.
22. Cesano, A., O'Connor, R., Lange, B., Finan, J., Rovera, G., and Santoli, D.
Homing and progression patterns of childhood acute lymph blast ic leukemiasin severe combined immunodeficiency mice. Blood, 77: 2463-2474, 1991.
23. Jansen, B., Uckun, F. M., Jaszcz, W. B., and Kersey, J. H. Establishment ofa human t(4;l 1) leukemia in SCID mice and successful treatment using anti-CD19(B43)-pokeweed anti viral protein immunotoxin. Cancer Res., 52:406-412, 1992.
24. Griesinger, F., Arthur, D. C., »running. R., Parkin, J. L., Ochoa, A. C.,Miller, W. J., Wilkowsky, C. W., Greenberg, J. M., Hurvitz, C., and Kersey,J. H. Mature T-lineage leukemia with growth factor-induced multilineagedifferentiation. J. Exp. Med., 169: 1101-1120, 1989.
25. Uckun, F. M., Gajl-Peczalska, K. J., Provisor, A. J., and Heerema, N. A.Immunophenotype-karyotype associations in human acute lymphoblasic leukemia. Blood, 73: 271-280, 1989.
26. Byers, V. S., Henslee, P. J., Keman, N., Blazar, B. R., Gingrich, R., Phillips,G. L., LeMaistre, C. R., Gilliland, G., Antin, J. H., Vogelsang, G., Martin,P., Tutschka, P. J., Trown, P., Ackerman, S. K., O'Reilly, R. J., and Scannon,P. J. Use of an anti-pan T lymphocyte ricin A chain immunotoxin as targetedimmunotherapy in steroid resistant acute graft rrrwi.v host disease. Blood,75:1426-1432, 1990.
27. Vitetta, E. S., Stone, M., Amlot, P., May, R., Till, M., Newman, J., Clark,P., Collins, R., Cunningham, D., Ghetie, V., Uhr, J. W., and Thorpe, P. E.Phase I immunotoxin trial in patients with B-cell lymphoma. Cancer Res.,51: 4052-4058, 1991.
28. Blakey, D. C., and Thorpe, P. E. Prevention of carbohydrate-mediatedclearance of ricin containing immunotoxins by the liver. In: A. E. Frankel(ed.), Immunotoxins, pp. 457-473. Boston: Kluwer Academic, 1988.
29. Scannon, P. J., Spitler, L. E., Lee, H. M., Kawahata, R. T., and Mischak, R.P. Human melanoma specific immunotoxins. United States Patent No.4,590,071; May 20, 1986.
30. Fulton, R. J., Blakey, D. C., Knowles, P. P, Uhr, J. W., Thorpe, P. E., andVitetta, E. S. Purification of ricin A,, A:. and B chains and characterizationof their toxicity. J. Biol. Chem., 261: 5314-5319, 1986.
31. Thorpe, P. E., Detre, S. L, Foxwell, B. M. J., Brown, A. N. F., Skilleter, D.N., Wilson, G., Forrester, J. A., and Stirpe, F. Modification of the carbohydrate in ricin with metaperiodate-cyanoborohydride mixtures. Eur. J.Biochem., 147: 197-206, 1985.
32. Vallera, D. A., Carrol, S. F., Snover, D., Carlson, G. J., and Blazar, B. R.Toxicity and efficacy of anti-T cell ricin toxin A chain immunotoxins in amurine model of established graft-vmiw-host disease induced across themajor histocompatibility barrier. Blood, 77: 182-194, 1991.
33. Myers, D. E, Uckun, F. M., Swaim, S. E., and Vallera, D. A. The effects ofaromatic and aliphatic maleimide crosslinkers on anti-CDS ricin immunotoxins. J. Immunol. Methods, 121: 129-142, 1989.
34. Uckun, F. M., Stong, R. C., Youle, R. J., and Vallera, D. A. Combined exvivo treatment with immunotoxins and mafosfamid: a novel immunochem-otherapeutic approach for elimination of neoplastic T cells from autologousmarrow grafts. J. Immunol., 134: 3504-3515, 1985.
35. Blazar, B. R., Hirsch, R., Cress, R. E., Carrol, S. F., and Vallera, D. A. Invivo administration of anti-CD3 monoclonal antibodies or immunotoxins inmurine recipients of allogeneic T-cell depleted marrow for the promotion ofengraftment. J. Immunol., 147: 1492-1503, 1991.
36. Mantel, N. Evaluation of survival data in two new rank order statistics arisingin its consideration. Cancer Chemother. Rep., 50: 163-168, 1966.
37. Ziegler, H. W., Fizzera, G., and Bach, F. H. Successful transplantation of ahuman leukemia cell line into nude mice: conditions optimizing graft acceptance. J. Nati. Cancer Inst., 68: 15-17, 1982.
38. Dillman, R. O., Johnson, D. E., Shawler, D. L., Halpern, S. E., Leonard, J.E., and Hagan, P. L. Athymic mouse model of a human T-cell tumor. CancerRes., 45: 5632-5636, 1985.
39. Preijers, F. W. M. B., Tax, W. J. M., De Witte, T., Janssen, A., v. d. Heijden,H., Vidal, H., Wessels, J. M. C., and Capei, P. J. A. Relationship betweenintemalization and cytotoxicity of ricin A chain immunotoxins. Br. J. Hae-mat., 70:289-294, 1988.
40. Laurent, G., Pris, J., Farcet, J-P., Carayon, P., Blythman, H., Casellas, P.,Poncelet, P., and Jansen, F. K. Effects of therapy with TlOl ricin A-chainimmunotoxin in two leukemia patients. Blood, 67: 1680-1687, 1986.
41. Vallera, D. A., Ash, R. C., Zanjani, E. D., Kersey, J. H., LeBien, T. W.,Beverly, P. C. L., Neville, D. M., Jr., and Youle, R. J. Anti-T cell reagentsfor human bone marrow transplantation: ricin linked to three monoclonalantibodies. Science (Washington DC), 222: 512-515, 1983.
42. Uckun, F. M., Kersey, J. H., Vallera, D. A., Ledbetter, J. A., Weisdorf, D.,Myers, D. E., Haake, R., and Ramsay, N. K. C. Autologous bone marrowtransplantation in high risk remission T-lineage ALL using immunotoxinsplus 4-1K for marrow purging: application of mult ¡parameterflow cytornei rvand colony assays for a quantitative analysis of autografts for the presenceof residual T-lineage leukemic lymphoid progenitor cells. Blood, 79: 1723-1733, 1990.
1320
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

TREATMENT OF A HUMAN ACUTE T-CELL LEUKEMIA IN SCID MICE
43. Vitetta, E. S. Immunotoxins: new therapeutic reagents for autoimmunity, 47. Leonard, J. £.,Johnson, D. E., Shawler, D. L., and Dillman, R. O. Inhibitioncancer, and AIDS. J. Clin. Inumino!.. IO: 15S-18S, 1990. of human T-cell tumor growth by TlOl-ricin A-chain in an athymic mouse
44. Hertler, A. A., and Frankel, A. E. Immunotoxins: a clinical review of their model. Cancer Res., 48:4862-4867, 1988.use in the treatment of malignancies. J. Clin. Oncol., 7: 1932-1942, 1989. 48- Hara- H •¿�Luo- Y- Haruta, Y., and Seon, B. K. Efficient transplantation of
45. Fulton, R. J., Tucker, T. F., Vitetta, E. S., and Uhr, J. W. Pharmacokinetics human non-T-leukemia cells into nude mice and induction of completeof tumor-reactive immunotoxins in tumor-bearing mice: effect of antibody regression of the transplanted distinct tumors by ricin A-chain conjugates ofvalency and deglycosylation of the ricin A chain on clearance and tumor ... mon°c'°nalant.bod.es SN5 and SN6. Cancer Res 48: 4673-4680 1988.._.••,,. f, „¿� ,„,,.„ ,,.,, .„„„ 49. Hará,H., and Seon, B. K. Complete suppression of in vivo growth of humanlocal.zat.on. Cancer Res ¥«:2618-2625, 1988. leukemia cells by specific immunotoxins. Proc. Nati. Acad. Sci. USA, 84:
46. Thorpe, P. E., Wallace, P. M., Knowles, P. P., Reif, M. G., Brown, A. N. 3390-3394, 1987F., Watson, G. J., Blakey, D. C., and Newell, D. R. Improved antitumor 50. Weil-Hillmà n, G., Uckun, F. M., Manske, J. M., and Vallera, D. A. Corn-effects of immunotoxins prepared with deglycosylated ricin A-chain and bined immunotherapy of human solid tumors in nude mice. Cancer Res., 47-hindered disulfide linkages. Cancer Res., 48:6396-6403, 1988. 579-585, 1987.
1321
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

1992;52:1314-1321. Cancer Res Burkhard Jansen, Daniel A. Vallera, Waclaw B. Jaszcz, et al. Immunotoxin DA7Mice Using the Anti-CD7-deglycosylated Ricin A-Chain Successful Treatment of Human Acute T-Cell Leukemia in SCID
Updated version
http://cancerres.aacrjournals.org/content/52/5/1314
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on January 4, 2015. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from




















