
Subchondral Bone Apparent Density and Locomotor Behavior in Extant Primates and Subfossil Lemurs...
25
Subchondral Bone Apparent Density and Locomotor Behavior in Extant Primates and Subfossil Lemurs Hadropithecus and Pachylemur John D. Polk & Scott A. Williams & Jeffrey V. Peterson & Charles C. Roseman & Laurie R. Godfrey Received: 8 April 2009 / Accepted: 18 February 2010 / Published online: 11 April 2010 # Springer Science+Business Media, LLC 2010 Abstract We analyze patterns of subchondral bone apparent density in the distal femur of extant primates to reconstruct differences in knee posture, discriminate among extant species with different locomotor preferences, and investigate the knee postures used by subfossil lemur species Hadropithecus stenognathus and Pachylemur insignis. We obtained computed tomographic scans for 164 femora belonging to 39 primate species. We grouped species by locomotor preference into knuckle-walking, arboreal quadruped, terrestrial quadruped, quadrupedal leaper, suspensory and vertical clinging, and leaping categories. We reconstructed knee posture using an experimentally validated procedure of determining the anterior extent of the region of maximal subchondral bone apparent density on a median slice through the medial femoral condyle. We compared subchondral apparent density magnitudes between subfossil and extant specimens to ensure that fossils did not display substantial mineralization or degradation. Subfossil and extant specimens were found to have similar magnitudes of subchondral apparent density, thereby permitting comparisons of the density patterns. We observed significant differences in the position of maximum subchondral apparent density between leaping and nonleaping extant primates, with leaping primates appearing to use much more flexed knee postures than nonleaping species. The anterior placement of the regions of maximum subchondral bone apparent density in the subfossil specimens of Hadropithecus and Pachylemur suggests that both species differed from leaping primates and included in their broad range of knee Int J Primatol (2010) 31:275–299 DOI 10.1007/s10764-010-9401-y J. D. Polk (*) : S. A. Williams : C. C. Roseman Department of Anthropology, University of Illinois Urbana-Champaign, Urbana, IL 61801, USA e-mail: [email protected] J. V. Peterson Department of Anthropology, San Diego State University, San Diego, CA 92182, USA L. R. Godfrey Department of Anthropology, University of Massachusetts, Amherst, MA 01003, USA
Transcript of Subchondral Bone Apparent Density and Locomotor Behavior in Extant Primates and Subfossil Lemurs...

Subchondral Bone Apparent Densityand Locomotor Behavior in ExtantPrimates and Subfossil Lemurs Hadropithecusand Pachylemur
John D. Polk & Scott A. Williams &
Jeffrey V. Peterson & Charles C. Roseman &
Laurie R. Godfrey
Received: 8 April 2009 /Accepted: 18 February 2010 /Published online: 11 April 2010# Springer Science+Business Media, LLC 2010
Abstract We analyze patterns of subchondral bone apparent density in the distal femurof extant primates to reconstruct differences in knee posture, discriminate among extantspecies with different locomotor preferences, and investigate the knee postures used bysubfossil lemur species Hadropithecus stenognathus and Pachylemur insignis. Weobtained computed tomographic scans for 164 femora belonging to 39 primatespecies. We grouped species by locomotor preference into knuckle-walking, arborealquadruped, terrestrial quadruped, quadrupedal leaper, suspensory and vertical clinging,and leaping categories. We reconstructed knee posture using an experimentallyvalidated procedure of determining the anterior extent of the region of maximalsubchondral bone apparent density on a median slice through the medial femoralcondyle. We compared subchondral apparent density magnitudes between subfossiland extant specimens to ensure that fossils did not display substantial mineralizationor degradation. Subfossil and extant specimens were found to have similar magnitudesof subchondral apparent density, thereby permitting comparisons of the densitypatterns. We observed significant differences in the position of maximum subchondralapparent density between leaping and nonleaping extant primates, with leapingprimates appearing to use much more flexed knee postures than nonleaping species.The anterior placement of the regions of maximum subchondral bone apparent densityin the subfossil specimens of Hadropithecus and Pachylemur suggests that bothspecies differed from leaping primates and included in their broad range of knee
Int J Primatol (2010) 31:275–299DOI 10.1007/s10764-010-9401-y
J. D. Polk (*) : S. A. Williams : C. C. RosemanDepartment of Anthropology, University of Illinois Urbana-Champaign, Urbana, IL 61801, USAe-mail: [email protected]
J. V. PetersonDepartment of Anthropology, San Diego State University, San Diego, CA 92182, USA
L. R. GodfreyDepartment of Anthropology, University of Massachusetts, Amherst, MA 01003, USA

postures rather extended postures. For Hadropithecus, this result is consistent withother evidence for terrestrial locomotion. Pachylemur, reconstructed on the basis ofother evidence as a committed arboreal quadruped, likely employed extended kneepostures in other activities such as hindlimb suspension, in addition to occasionalterrestrial locomotion.
Keywords apparent density . locomotion . posture . subchondral bone .
subfossil lemur
Introduction
Interpreting, or testing hypotheses about, the locomotor behavior of extinct speciesrequires information to be accumulated from a variety of different types of analyses ofskeletal remains. In this study we focus on the locomotor behaviors of Hadropithecusstenognathus and Pachylemur insignis. The postcranial skeletons of these subfossillemurs are now well known (Godfrey and Jungers 2002). Discoveries of newassociated skeletal elements of Hadropithecus at Andrahomana cave in southeasternMadagascar (Burney et al. 2008; Godfrey et al. 2006a) confirmed the hind limbattributions made by Godfrey et al. (1997). The femora now ascribed toHadropithecus include a distal epiphysis from Andrahomana in perfect condition,analyzed here (now in the collections of the Naturhistorisches Museum, Vienna).There are also 5 partial femora of Hadropithecus in the collections of the University ofAntananarivo, 2 in the Muséum National d’Histoire Naturelle (Paris), and 1 illustratedby Grandidier (1905, originally misattributed to Palaeopropithecus; see Godfrey et al.1997) whose current whereabouts are unknown. Numerous whole femora ofPachylemur insignis exist in the collections of the University of Antananarivo.
The locomotor behaviors of Pachylemur insignis and Hadropithecus stenogna-thus have previously been reconstructed on the basis of elements of their axial andappendicular skeletons. Both were nonleaping quadrupeds, although Hadropithecusexhibits derived traits indicative of far greater terrestriality than Pachylemur, whichhas been described as a deliberate, quadrupedal climber. Both were stocky andrelatively short-limbed individuals (Godfrey et al. 2006b). Pachylemur andHadropithecus were also both larger in body size than any living lemur. At ca.35 kg, Hadropithecus was 3× as heavy as Pachylemur (ca. 11 kg), which was in turn3× heavier than the largest-bodied extant lemurid, Varecia (Jungers et al. 2008).Given its body size and lack of leaping adaptations, it is likely that Pachylemur, likeHadropithecus, used terrestrial routes for travel, particularly in the succulentwoodlands and spiny thickets of southern Madagascar.
Although gross bony morphology can provide important information aboutlocomotor behaviors, such information may be supplemented by new techniques.Our goal for this paper is to explore some of the ways in which analyses of boneapparent density patterns can be used to discriminate among the locomotor behaviorsused by extant primates and to make inferences about these behaviors in extinctsubfossil primates. We investigate patterns of apparent density in the femoraPachylemur insignis and Hadropithecus stenognathus, and use comparative data froma broad sample of primate species to discern gross patterns of locomotor behavior.
276 J. D. Polk et al.

Background to Analyses of Subchondral Bone Apparent Density
Subchondral bone is the thin layer of cortical bone immediately deep to thearticular cartilage in synovial joints. Subchondral bone serves to transmit anddistribute forces from the joint surface to the underlying trabecular bone, and itmay serve to dissipate a small portion of the force as well (Hoshino andWallace 1987; Radin and Paul 1971; Radin et al. 1970; Simon et al. 1972).Increasing subchondral bone density can alter the material properties of this tissueconferring increased compressive strength, compressive modulus, fatigue life, andresistance to crack initiation (Carter and Hayes 1977; Currey 1988, 2002; Rice etal. 1988; Wall et al. 1979; Wright and Hayes 1977). These alterations in materialproperties may develop as a consequence of altered loading magnitudes or loadingfrequencies (Carter and Beaupre 2001; Fischer et al., 1995), and thus variation inthe magnitudes of apparent density may provide some indication of differences inloading magnitudes across individuals, or across joint surfaces of the sameindividual (Ahluwalia 2000; Fischer et al. 1995).
Since the late 1980s a group of German researchers has applied new radiographicand imaging techniques to the spatial analysis of subchondral bone density in severalhuman joints (Eckstein et al. 1995, 1999; Müller-Gerbl et al. 1989, 1990; vonEisenhart et al. 1999). They suggested that the patterns of relative density that theyobserved in the joints of human cadavers, or living subjects reflected differences inload-bearing (Eckstein et al. 1995, 1999; von Eisenhart et al. 1999) and thetechnique has been clinically applied to understanding patterns of joint loading innormal and pathological human (Eckstein et al. 1995, 1999; Müller-Gerbl et al.1989, 1990) and animal subjects (Samii et al. 2002).
Comparative studies have analyzed patterns of bone apparent density in differentspecies of mammals to make inferences about joint posture, load magnitude, andload distribution (Ahluwalia 2000; Carlson and Patel 2006; Patel and Carlson 2006,2007, 2008; Polk et al. 2009). Polk et al. (2008) experimentally investigated therelationship between apparent density variation and joint posture. These researchersused differences in substrate orientation to alter the locomotor postures of 2 groupsof sheep during twice-daily bouts of exercise for a period of 45 d. They obtainedkinematic measurements for the knee postures during locomotion and comparedthese measurements to joint posture estimates obtained from bone apparent densitypatterns. They found that the position of the region of maximum subchondral boneapparent density could be used to reconstruct joint postures that closely matchedkinematically measured joint postures (Polk et al. 2008). Further, because this studydid not find differences in peak apparent density magnitudes between theexperimental groups, they inferred that the observed patterns of apparent densityresulted from the differences in habitual loading conditions instead of differences inloading magnitude. However, it remains possible that peak loads could influencepatterns of apparent density in the sample of specimens included in the current study.Polk et al. (2009) applied the same methods to test a biomechanical model for theeffects of body mass on primate locomotor postures. We used these methods in thecurrent study to discriminate between the bones of extant primates that engage indifferent locomotor modes, and draw inferences about the knee postures of 2subfossil lemurs, Pachylemur and Hadropithecus.
Subchondral Bone Apparent Density 277

Knee posture provides an informative window into the mechanisms of bodysupport and hindlimb use. The lower extremity is the primary propulsive limb forboth leaping and quadrupedal locomotion (Demes and Günther 1989; Demes et al.1994; Reynolds 1985). The hind limbs of primates also bear more weight thanprimate forelimbs, and the musculature crossing the knee is very important forresisting gravity (Demes et al. 1994; Hanna et al. 2006; Polk 2002, 2004). Thetorques necessary to propel most large-bodied leapers and quadrupeds are generatedat the hip and knee (Demes and Günther 1989; Demes et al. 1994; Reynolds 1985)and the loads placed on these joints during locomotion should influence someaspects of joint morphology. Because primates that use different modes oflocomotion, e.g., leaping vs. quadrupedalism, utilize different hip and knee posturesduring locomotion, and during nonlocomotor postural behavior, the mechanicalloads experienced by the bones will be located in different places across the jointsurfaces, and differences in joint loading should be detectable in spatial analyses ofthe apparent density patterns on the joint surface.
We here focus on the medial femoral condyle (MFC). We selected this surfacebecause the MFC should bear more weight than the lateral femoral condyle, and bemore likely to remodel in response to locomotor forces. This expectation issupported by experimental studies of primate and other terrestrial mammalianlocomotion where the peak ground reaction force vectors pass medial to the kneejoint during walking and running gaits (Demes and Carlson 2009; Polk unpubl. data;Taylor et al. 2006).
As a consequence, studying the patterns of apparent density on the medial femoralcondyle of subfossil lemurs can provide information about the load history and kneepostures used by these extinct species. Reconstructing density patterns in extinct speciesmust be done with caution and we must attempt to rule out nonbiological factors thatmay have influenced the variables we are attempting to measure. We assume that theapparent density patterns in the joint surface have not been modified by diagenetic ormineralization processes. This assumption is partially testable by comparing themagnitude and range of subchondral apparent densities in the subfossil specimens tomuseum specimens of extant species. If the density magnitudes in the subfossilspecimens overlap with those of extant species then we have no basis for rejecting thisassumption (see Discussion for some further qualifications).
Background to Hadropithecus and Pachylemur
Pachylemur insignis and Hadropithecus stenognathus both ranged throughout thespiny thicket and succulent woodland ecoregions of southern and southwesternMadagascar (Burgess et al. 2004; Muldoon et al. 2009). The former also inhabitedthe dry deciduous forests further to the north as well as more than one subhumidforest site in central Madagascar (Ampasambazimba). A larger Pachylemur,sometimes placed in its own species (P. jullyi) but sometimes considered ageographic variant of P. insignis, also lived there. Pachylemur occurs in greaterabundance in interior sites such as Taolambiby, Tsirave, and Ampasambazimba, whereasHadropithecus is better represented at coastal sites such as Andrahomana (SE) andAnavoha (SW), and is the only extinct lemur recorded at some dune sites (Ambovombe,extreme south) dominated by hippopotamuses, crocodiles, and giant tortoises (Decary
278 J. D. Polk et al.

1926). Niche separation of Pachylemur and Hadropithecus is confirmed by stablecarbon isotope analysis (Burney et al. 2004; Crowley et al. 2010). Pachylemur had adiet dominated by C3 vegetation (likely including a large amount of fruit; Crowley et al.2010; Godfrey et al. 2004, 2006b, 2008a), whereas Hadropithecus likely foraged on C4plants on the ground (Godfrey et al. 2006b; Ryan et al. 2008).
Both Pachylemur and Hadropithecus had postcranial proportions and morphol-ogies consistent with versatile quadrupedalism (Godfrey and Jungers 2002). InPachylemur, the forelimb was shorter than the hind limb but not markedly so,whereas, based on humerofemoral indices, the forelimb of Hadropithecus was likelyslightly longer than the hind limb (Godfrey et al. 2006a). Neither species hadspecializations for leaping or forelimb suspension. Researchers have reconstructedPachylemur as a slow or deliberate quadrupedal climber on the basis of its hindlimband vertebral anatomy (Ravololonarivo 1990; Shapiro et al. 2005; Tardieu andJouffroy 1979), and Hadropithecus as a semiterrestrial quadruped, less agile than itsclose relative, Archaeolemur (Godfrey 1988; Godfrey et al. 1997, 2006a).Pachylemur would have been considerably more arboreal than Hadropithecus.
The limb bones ofPachylemur are much like those of Varecia but the hind limb wasrelatively shorter, the forelimb relatively longer, and the knee shows none of thefeatures associated with leaping or bounding in Varecia and other lemurs. Mostrevealing are differences in the vertebral columns of Pachylemur and Varecia, whichsuggest greater spinal stability at the expense of mobility, and slower, more deliberatequadrupedalism in the former (Shapiro et al. 2005). As in Archaeolemur, Pachylemurhad lumbar vertebrae with somewhat elongated spinous processes, short vertebralbodies, narrow laminae and pedicles, and transverse processes that arise from thedorsal aspect of the vertebral body or pedicle, and are oriented laterally (instead ofventrally). The lumbar spinous processes are not as elongated as those of Varecia, andthey project dorsally instead of cranially as in Varecia. These traits suggest thatPachylemur lacked the bounding and leaping behaviors that characterize Varecia.
The traits revealing stronger terrestriality in Hadropithecus are highly derived, andinclude a weakly developed carpal tunnel, as evidenced by a weakly projectingscaphoid tubercle and a hamate with virtually no hook, and short digits (Lemelin et al.2008). The reconstructed hand posture of Hadropithecus is extended, pronated, andneutral, i.e., lacking the ulnar deviation typical of vertical clingers and leapers as wellas slow climbers (Lemelin et al. 2008). Godfrey et al. (1997, 2006a) provide detaileddescriptions of the long bones; their likenesses to those of semiterrestrialcercopithecoids have long been appreciated (Godfrey 1988; Walker 1974). A tendencyfor Hadropithecus to move slowly, in comparison to Archaeolemur and to morecursorial baboons and macaques, suggested particularly by its femoral morphology—robust, anteroposteriorly compressed diaphysis; shallow, proximodistally short andwide patellar groove; mediolaterally asymmetric femoral condyles— was recentlyconfirmed via an analysis of the relative sizes of the semicircular canals of living andextinct lemurs (Walker et al. 2008).
Predictions and Justification
The basic premise of our study is that differences in joint loading postures shouldcause differences in the position of the region of maximum apparent density on the
Subchondral Bone Apparent Density 279

femoral condyles. Primates that engage in vertical clinging and leaping use highlyflexed hip and knee postures while resting and feeding on vertical supports(Fleagle 1999; Walker 1983). In addition, when they leap between supports they mustgenerate high takeoff forces starting from these flexed postures (Demes et al. 1999;Demes and Günther 1989). Correspondingly, primates that engage in vertical clingingand leaping should exhibit a more posteriorly positioned region of maximum apparentbone density on the femoral condyle. Fleagle (1976) investigated morphologicallydifferences between arboreal quadrupeds and quadrupedal leapers in 2 colobine taxa.This study noted several differences in lower extremity morphology, whichpresumably relate to differences in hip and knee function between the 2 species.Differences in knee morphology also exist; e.g., Fleagle (1976) noted that the patellargroove of the quadrupedal leaper extended further proximally than in the arborealquadruped. This might imply the use of more extended knee postures in quadrupedalleapers. However, researchers have not investigated the biomechanics of leaping fromquadrupedal postures as thoroughly as other forms of primate locomotion, andquadrupedal leapers do use fairly flexed knee postures during propulsion (BrigitteDemes, pers. comm.). As a consequence, we predict that primates that engage insignificant amounts of leaping, whether as quadrupedal leapers or vertical clingers andleapers, should exhibit a region of maximum density that is posteriorly placed on themedial femoral condyle. Quadrupedal nonleapers are expected to adopt relativelymore extended knee postures and have correspondingly more anteriorly placed RMD.Arboreal primates may be expected to use more flexed limb postures than moreterrestrial species to improve limb compliance and to maintain above-branch stabilitybetter (Schmitt 1999). In another study using these same data, Polk et al. (2009)showed that body mass correlates significantly with the position of the RMD, withlarger animals using more extended knee postures. Thus we expect similar patternshere, and large-bodied knuckle walkers and Pongo are expected to use more extendedpostures than other smaller primate species do.
Our predictions about the postures used by the subfossil lemurs Hadropithecusand Pachylemur are informed by both their morphology and body size.Hadropithecus is a moderately large-bodied primate (35 kg; Jungers et al. 2008),and this, in combination with its inferred use of some terrestrial substrates, suggestsuse of fairly extended knee postures. Size-based predictions are less clear forPachylemur (11 kg; Jungers et al. 2008) because extant primates of similar sizeexhibit a broad range of variation in knee posture (Polk et al. 2009). Based onprevious morphological comparisons we predicted that the density patterns forPachylemur would be consistent with those of other arboreal quadrupedal,nonleaping species of primates.
Methods
We included 2 subfossil lemur specimens in this analysis. The University ofAntananarivo generously loaned the left femur of Pachylemur insignis to L. R.Godfrey. The second is a left distal femoral epiphysis of Hadropithecusstenognathus discovered in 2003 at Andrahomana Cave (site AHA-I), southeasternMadagascar (Burney et al. 2008; Godfrey et al. 2006a).
280 J. D. Polk et al.

We borrowed the comparative sample of primate femora from the Museum ofComparative Zoology at Harvard University. The sample represents 164 individualsof 39 primate species (Table I). All primate specimens were wild-shot adults. Themajority of femora were from the right side, but if a right femur was not available, aleft was substituted. Locomotor groupings in Table I follow Fleagle (1999). Ouranalysis is preliminary in the sense that many species are represented by smallsample sizes, but we anticipate that additional specimens will reinforce the patternswe describe.
Protocols for assessing bone density follow Polk et al. (2008, 2009). Wemeasured bone apparent density in all of the specimens using the same protocol onthe same computed tomography (CT) scanner (GE Lightspeed Plus) at the MountAuburn Hospital in Cambridge, MA. Medical CT scanning allows quantification ofapparent density of composite materials such as subchondral bone that can containdifferent types of tissues. True bone tissue density (TD) measurements differ frombone apparent density measurements in that TD can be calculated only for singlematerials, e.g., bone tissue itself, rather than some combination of bone tissue, air,residual soft tissue, etc. We obtained estimates of bone apparent density in thesubchondral bone of the medial femoral condyle for all specimens.
All bony specimens were oriented with the long axis perpendicular to the CTsource/sensor plane to maximize the number of slices obtained per specimen. Slicethickness was 0.625 mm, and pixel size was 0.234 mm×0.234 mm. We saved dataas 16-bit DICOM images. We used a standard soft-tissue CT algorithm to ensure thatthe intensity of gray in the images was linearly related to bone apparent density. Theintensity of gray levels, and therefore bone apparent density measurements, isquantified in Hounsfield units (H), and they are expressed relative to the densities ofknown materials (Ruff and Leo 1986). Air has apparent density of ca. –1000H,water is 0H, and subchondral bone has apparent densities of ca. 50H to >1500H.The Mount Auburn CT system is calibrated daily to ensure the accuracy of densitymeasurements. AMIRA (v. 4.1, Mercury Computing Systems) software was used tocreate virtual 3D representations of the femora from the CT images, therebypreserving the geometry of the entire bone.
Following Polk et al. (2008, 2009), we quantified the regions of maximumdensity (RMD) on a median slice through the medial femoral condyle (MFC). Toobtain these measurements, we obtained a single, oblique, 2-dimensional slicethrough the long axis of the MFC for each specimen (slice thickness=0.234 mm). Toensure consistency in slice orientation across individual subjects, we oriented theplane of this slice to pass through the long axis of the MFC.
Color maps were applied to the oblique slice image. These color maps help todistinguish variation in the 216 levels of gray in the 16-bit DICOM images. AMIRApermits manual selection of the range of Hounsfield units that can be displayed(comparable to setting the CT level and window) and to divide this range into amaximum of 256 separate colors (bins). We assessed maximum and minimumdensities along the subchondral surface to the nearest 25H. We assigned densitiesless than or equal to the minimum found on the subchondral surface to bin 0, anddensities greater than or equal to the maximum found on the subchondral surface tobin 255. We divided the remaining density range (254 bins) into 10 regions ofdiffering density (the highest and lowest density regions had 1 fewer bin than did the
Subchondral Bone Apparent Density 281

Table I Species, sample size, and locomotor mode
Species n Locomotor mode
Cercocebus sp. 2 Aq
Cercopithecus Diana 1 Aq
Cercopithecus l’hoesti 1 Aq
Chlorocebus aethiops 1 Tq
Piliocolobus badius 1 Ql
Colobus guereza 2 Ql
Colobus polykomos 2 Ql
Macaca fascicularis 21 Aq
Macaca mulatta 3 Tq
Macaca nemestrina 3 Tq
Mandrillus sp. 5 Tq
Miopithecus talapoin 2 Aq
Nasalis larvatus 11 ql
Presbytis hosei 3 ql
Trachypithecus cristata 13 ql
Trachypithecus phayrei 6 ql
Trachypithecus vetulus 1 ql
Gorilla gorilla 12 kw
Hylobates lar 24 s
Pan troglodytes 4 kw
Pongo pygmaeus 6 s
Alouatta belzebuth 2 aq
Alouatta fusca 4 aq
Ateles sp. 2 aq
Callithrix jaccus 2 aq
Leontopithecus rosalia 1 ql
Saguinus midas 1 ql
Daubentonia madagascariensis 1 aq
Eulemur fulvus 3 ql
Eulemur rubriventer 2 ql
Galago senegalensis 2 l
Hapalemur griseus 5 l
Indri indri 1 l
Lepilemur mustelinus 5 l
Nycticebus coucang 2 aq
Otolemur crassicaudatus 1 l
Perodicticus potto 1 aq
Propithecus verreauxi 2 l
Varecia variegate 3 ql
282 J. D. Polk et al.
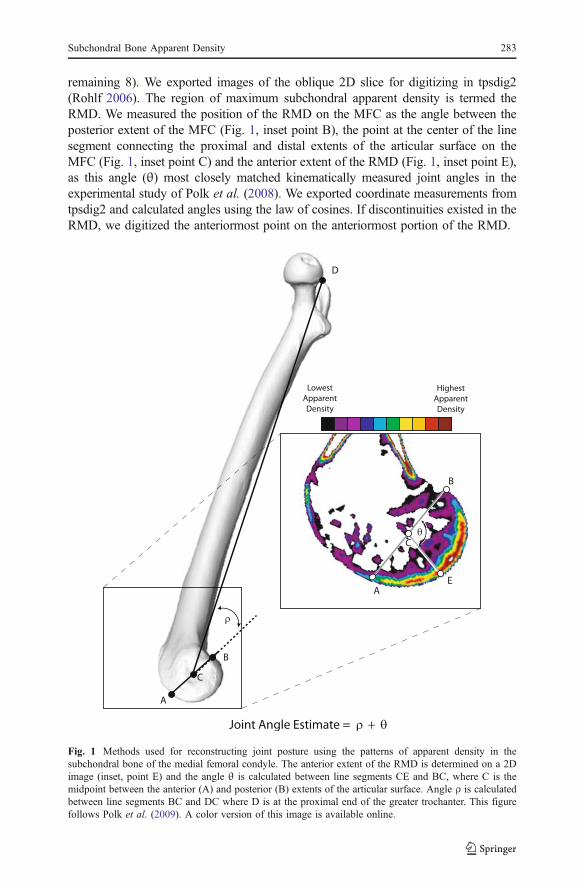
remaining 8). We exported images of the oblique 2D slice for digitizing in tpsdig2(Rohlf 2006). The region of maximum subchondral apparent density is termed theRMD. We measured the position of the RMD on the MFC as the angle between theposterior extent of the MFC (Fig. 1, inset point B), the point at the center of the linesegment connecting the proximal and distal extents of the articular surface on theMFC (Fig. 1, inset point C) and the anterior extent of the RMD (Fig. 1, inset point E),as this angle (θ) most closely matched kinematically measured joint angles in theexperimental study of Polk et al. (2008). We exported coordinate measurements fromtpsdig2 and calculated angles using the law of cosines. If discontinuities existed in theRMD, we digitized the anteriormost point on the anteriormost portion of the RMD.
D
A
B
C
A
B
E
C
HighestApparentDensity
LowestApparentDensity
Fig. 1 Methods used for reconstructing joint posture using the patterns of apparent density in thesubchondral bone of the medial femoral condyle. The anterior extent of the RMD is determined on a 2Dimage (inset, point E) and the angle θ is calculated between line segments CE and BC, where C is themidpoint between the anterior (A) and posterior (B) extents of the articular surface. Angle ρ is calculatedbetween line segments BC and DC where D is at the proximal end of the greater trochanter. This figurefollows Polk et al. (2009). A color version of this image is available online.
Subchondral Bone Apparent Density 283
S. A. Williams and J. V. Peterson digitized specimens without reference to thespecies to which they belonged, the size of the individual, or locomotor mode,thereby preventing observers from inadvertently biasing the results. Polk et al.(2009) reported the results of tests for interobserver error.
We estimated the position of the RMD on the entire femur for all completespecimens, i.e., all exceptHadropithecus, by combining the angle θ (obtained from the2D oblique slice), with angle ρ calculated from 3-dimensional coordinates placed onthe distal and proximal extents of the MFC articular surface (Fig. 1, points A and B)and a landmark on the proximalmost point on the greater trochanter (Fig. 1, point D).We obtained the 3-dimensional coordinates following the methodology of Polk et al.(2009), using AMIRA and Landmark (Wiley et al. 2005). For Hadropithecus, wecalculated the angle ρ from a 2D photograph of a complete, adult specimen(MNHN1910-33) from the collections of the Paris Museum of Natural History. We didnot obtained density measurements from this specimen because of some damage onthe condylar surfaces.
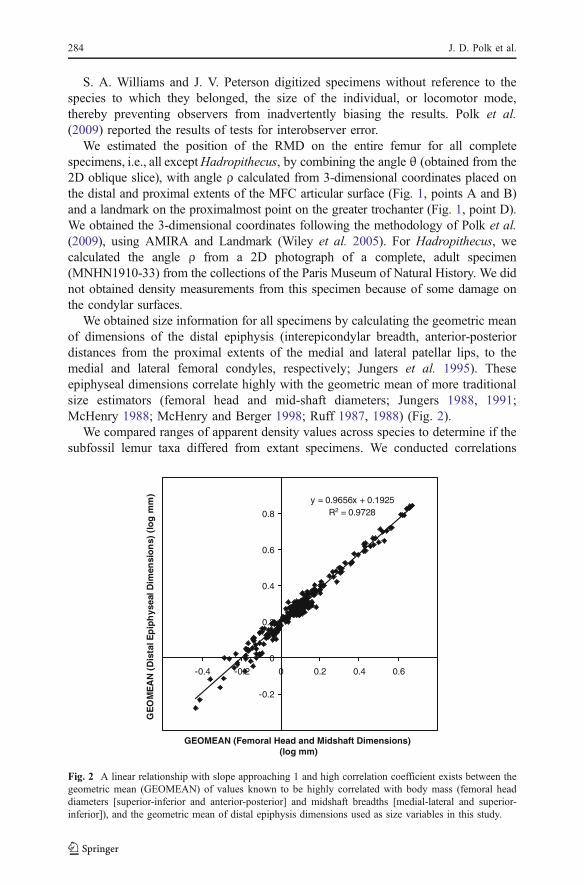
We obtained size information for all specimens by calculating the geometric meanof dimensions of the distal epiphysis (interepicondylar breadth, anterior-posteriordistances from the proximal extents of the medial and lateral patellar lips, to themedial and lateral femoral condyles, respectively; Jungers et al. 1995). Theseepiphyseal dimensions correlate highly with the geometric mean of more traditionalsize estimators (femoral head and mid-shaft diameters; Jungers 1988, 1991;McHenry 1988; McHenry and Berger 1998; Ruff 1987, 1988) (Fig. 2).
We compared ranges of apparent density values across species to determine if thesubfossil lemur taxa differed from extant specimens. We conducted correlations
y = 0.9656x + 0.1925R2 = 0.9728
-0.2
0
0.2
0.4
0.6
0.8
-0.4 -0.2 0 0.2 0.4 0.6
GEOMEAN (Femoral Head and Midshaft Dimensions)(log mm)
GE
OM
EA
N (
Dis
tal E
pip
hys
eal D
imen
sio
ns)
(lo
g m
m)
Fig. 2 A linear relationship with slope approaching 1 and high correlation coefficient exists between thegeometric mean (GEOMEAN) of values known to be highly correlated with body mass (femoral headdiameters [superior-inferior and anterior-posterior] and midshaft breadths [medial-lateral and superior-inferior]), and the geometric mean of distal epiphysis dimensions used as size variables in this study.
284 J. D. Polk et al.

between the upper or lower range boundary and the size variable via Pearsonproduct-moment correlation coefficients (Sokal and Rohlf 1995). We calculatedspecies means, or individual values where sample sizes required, for the threeangular measurements: θ, ρ, and θ+ρ.
In some cases, patterns of interspecific differences may be the result of evolutionaryprocesses that correlate with the phylogenetic relationships among groups (Felsenstein1985; Harvey and Pagel 1991; Lynch 1991). To assess whether or not interspecificdifferences in the size and angle traits were dependent on phylogenetic relationships,we estimated the proportion of variance among species accounted for via aphylogenetic mixed model (PMM). The PMM is a linear mixed model that canpartition the observed variance for any characteristic into a component that reflects thevariance associated with the effects of inheritance due to the nested sets of phylogeneticrelationships (phylogenetic parameter), and a component that reflects more rapid,taxon-specific changes (Housworth et al. 2004) that may be due to environmentaleffects or recent adaptive evolution. We estimated the phylogenetic and taxon-specificeffect estimates and the log-likelihood of the PMM (ln p) using an expectationmaximization algorithm using the compar.lynch function from the ape package (Paradis2006) for the R statistical computing environment (http://www.r-project.org/) modifiedto accept univariate data. We estimated the log-likelihood of the null model (ln p0) byestimating the probability density for drawing the observed means from a normaldistribution with a mean and variance equal to those estimated from the data on thetraits themselves. We calculated a likelihood ratio statistic (Λ) as Λ=2(ln p−ln p0),which is χ2 distributed with 1 degree of freedom. The phylogeny used in our study wasmodified from Smith and Cheverud (2002). Revision to the strepsirrhine phylogenyfollows Catlett et al. (2010), in turn based on Horvath et al. (2008), Orlando et al.(2008), and other research. The Newick formatted tree is provided in the Appendix.
The PMM, like other methods for studying the effects of phylogeny, can besensitive to departures from the distributional assumptions of the fitted models. Weassessed whether or not the data were acceptably represented by a normaldistribution using the Shapiro-Wilk test (Shapiro and Wilk 1965) and log-transformed (base e) when large departures were detected.
Differences between locomotor categories were evaluated using Kruskal Wallistests with post hoc pairwise Mann-Whitney comparisons in SPSS (v.16; SPSS Inc.).For the majority of statistical tests performed in this study, we assessed significanceusing a type I error rate of 0.05, but we used α=0.1 to assess significance for theMann-Whitney U tests. We used this lower threshold for significance to identifydifferences that are likely to be biologically meaningful, but that did not achievestatistical significance at the common α=0.05 level (cf. Cohen 1990).
Results
Density Magnitude
The ranges of apparent density magnitudes in subchondral bone on the medialfemoral condyle of several species of primates are shown in Fig. 3. The lowerboundaries of the range do not significantly correlate with body size (r=−0.196, n=
Subchondral Bone Apparent Density 285
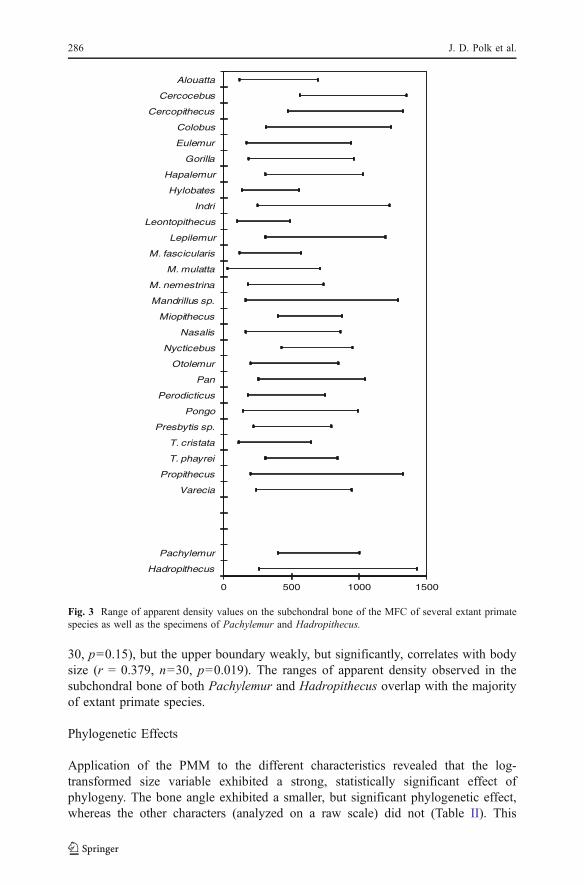
30, p=0.15), but the upper boundary weakly, but significantly, correlates with bodysize (r = 0.379, n=30, p=0.019). The ranges of apparent density observed in thesubchondral bone of both Pachylemur and Hadropithecus overlap with the majorityof extant primate species.
Phylogenetic Effects
Application of the PMM to the different characteristics revealed that the log-transformed size variable exhibited a strong, statistically significant effect ofphylogeny. The bone angle exhibited a smaller, but significant phylogenetic effect,whereas the other characters (analyzed on a raw scale) did not (Table II). This
0 500 1000 1500
Alouatta
Cercocebus
Cercopithecus
Colobus
Eulemur
Gorilla
Hapalemur
Hylobates
Indri
Leontopithecus
Lepilemur
M. fascicularis
M. mulatta
M. nemestrina
Mandrillus sp.
Miopithecus
Nasalis
Nycticebus
Otolemur
Pan
Perodicticus
Pongo
Presbytis sp.
T. cristata
T. phayrei
Propithecus
Varecia
Pachylemur
Hadropithecus
Fig. 3 Range of apparent density values on the subchondral bone of the MFC of several extant primatespecies as well as the specimens of Pachylemur and Hadropithecus.
286 J. D. Polk et al.

indicates that there is sufficient power to detect phylogenetic effects in the sampleand that any effect of phylogeny on the density-based angular measurements is smalland should not impart undue bias into ordinary statistical comparisons among groupsof the species. We did not attempt to correct for the phylogenetic effect in the boneangle measurement because this variable did not discriminate well among locomotorgroups.
The size traits were statistically different from the normal model on the raw scale,but were log-normally distributed. Both the density angle and sum of angles deviatedslightly from a normal distribution. These were not log-transformed because thedeviations from normality were small.
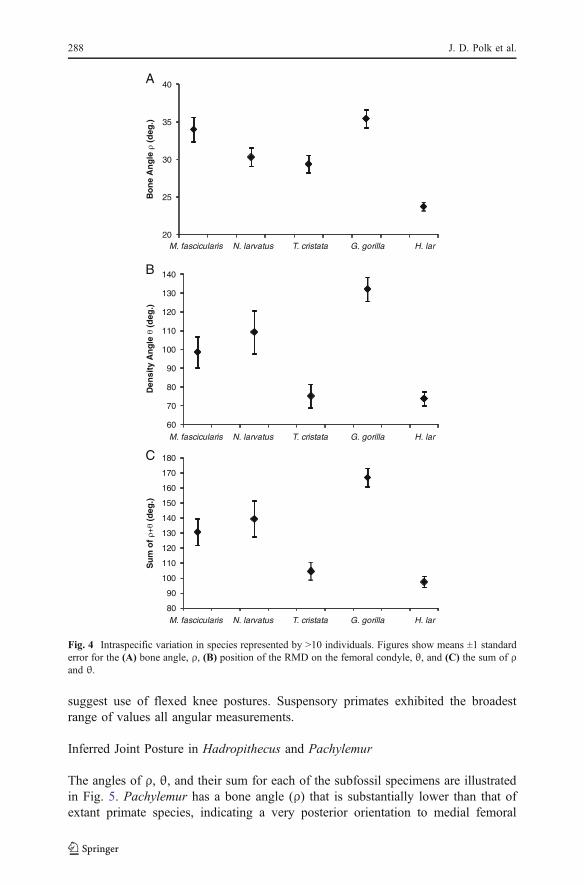
Intraspecific Variation
Figure 4 displays the means and standard errors for the 3 angular variables includedin this study. Species included in Fig. 4 were represented by ≥10 specimens andshould therefore provide a reasonable estimate of intraspecific variation. Standarderrors are low enough to permit meaningful discrimination between most of thesespecies. Results from a 1-way ANOVA reveal significant differences betweenspecies for each variable (F=18.1, F=10.3, F=13.7 for interspecific comparisons ofbone angle, density angle, and combined ρ+θ, respectively; df=4, p<0.001 for allcomparisons).
Locomotor Variation in the Position of the RMD
Figure 5 illustrates the differences among locomotor groups in the position of theRMD on the femur. The bone angle (ρ) did not show significant locomotor effects(χ2=4.929, df=5, p=0.425). Differences among locomotor groups approachedstatistical significance for the density angle (θ) (χ2=9.333, df=5, p=0.096), but thesum of the two angles did show significant differences among locomotor groups(χ2=11.401, df=5, p=0.044). Pairwise Mann-Whitney U tests reveal the pattern inthe differences among groups and these results are shown in Table III. Arborealquadrupeds, terrestrial quadrupeds, and knuckle-walking primates did not differsignificantly from each other, but all differed from quadrupedal leaping primates,with the quadrupedal leapers having RMD positions consistent with the use of flexedknee postures, Specialized leaping primates could be distinguished statistically fromknuckle-walkers but not from the other locomotor groups. In spite of this lack ofstatistical significance, specialized leaping primates did demonstrate RMD that
Table II Summary of tests for phylogenetic effect
Shapiro Wilk Shapiro Wilk p value Phylogenetic h2 likelihood ratio p value
Log size 0.970 0.368 0.982 46.155 0.000
Bone angle 0.961 0.192 0.623 6.447 0.011
Density angle 0.930 0.022 0.002 –0.013 1.000
Sum of angles 0.931 0.030 0.001 0.008 0.930
Subchondral Bone Apparent Density 287
suggest use of flexed knee postures. Suspensory primates exhibited the broadestrange of values all angular measurements.
Inferred Joint Posture in Hadropithecus and Pachylemur
The angles of ρ, θ, and their sum for each of the subfossil specimens are illustratedin Fig. 5. Pachylemur has a bone angle (ρ) that is substantially lower than that ofextant primate species, indicating a very posterior orientation to medial femoral
20
25
30
35
40
M. fascicularis N. larvatus T. cristata G. gorilla H. lar
M. fascicularis N. larvatus T. cristata G. gorilla H. lar
M. fascicularis N. larvatus T. cristata G. gorilla H. lar
Bo
ne
An
gle
ρ (
deg
.)
60
70
80
90
100
110
120
130
140
Den
sity
An
gle
θ (
deg
.)
80
90
100
110
120
130
140
150
160
170
180
Su
m o
f ρ+θ (
deg
.)
A
C
B
Fig. 4 Intraspecific variation in species represented by >10 individuals. Figures show means ±1 standarderror for the (A) bone angle, ρ, (B) position of the RMD on the femoral condyle, θ, and (C) the sum of ρand θ.
288 J. D. Polk et al.
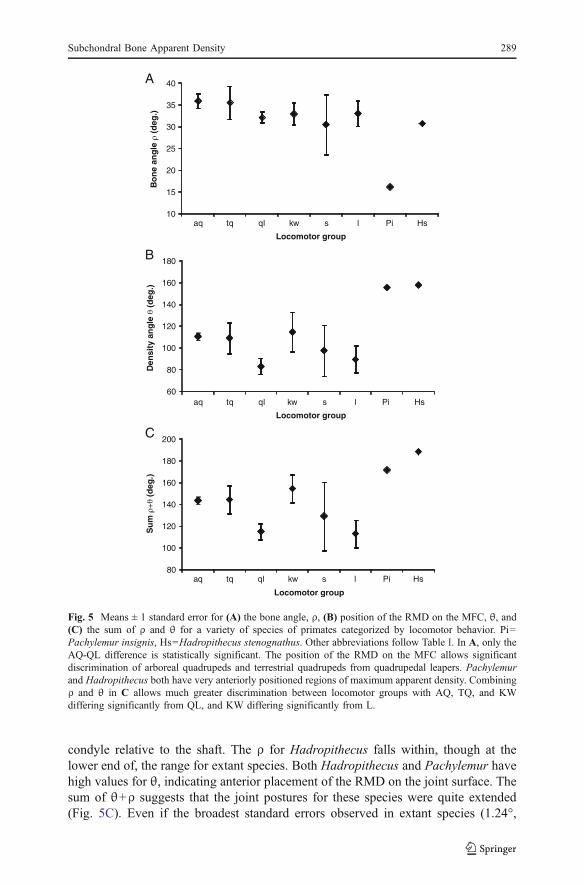
condyle relative to the shaft. The ρ for Hadropithecus falls within, though at thelower end of, the range for extant species. Both Hadropithecus and Pachylemur havehigh values for θ, indicating anterior placement of the RMD on the joint surface. Thesum of θ+ρ suggests that the joint postures for these species were quite extended(Fig. 5C). Even if the broadest standard errors observed in extant species (1.24°,
10
15
20
25
30
35
40
aq tq ql kw s l Pi Hs
Bo
ne
ang
le ρ
(d
eg.)
Locomotor group
aq tq ql kw s l Pi Hs
Locomotor group
Locomotor group
60
80
100
120
140
160
180
Den
sity
an
gle
θ (
deg
.)
80
100
120
140
160
180
200
aq tq ql kw s l Pi Hs
Su
m ρ+θ (
deg
.)
A
C
B
Fig. 5 Means ± 1 standard error for (A) the bone angle, ρ, (B) position of the RMD on the MFC, θ, and(C) the sum of ρ and θ for a variety of species of primates categorized by locomotor behavior. Pi=Pachylemur insignis, Hs=Hadropithecus stenognathus. Other abbreviations follow Table I. In A, only theAQ-QL difference is statistically significant. The position of the RMD on the MFC allows significantdiscrimination of arboreal quadrupeds and terrestrial quadrupeds from quadrupedal leapers. Pachylemurand Hadropithecus both have very anteriorly positioned regions of maximum apparent density. Combiningρ and θ in C allows much greater discrimination between locomotor groups with AQ, TQ, and KWdiffering significantly from QL, and KW differing significantly from L.
Subchondral Bone Apparent Density 289

11.5°, 11.8° observed in Nasalis for ρ θ, and their sum, respectively) were observedin these subfossil species, they would still have only minimal overlap with extanttaxa for this measurement.
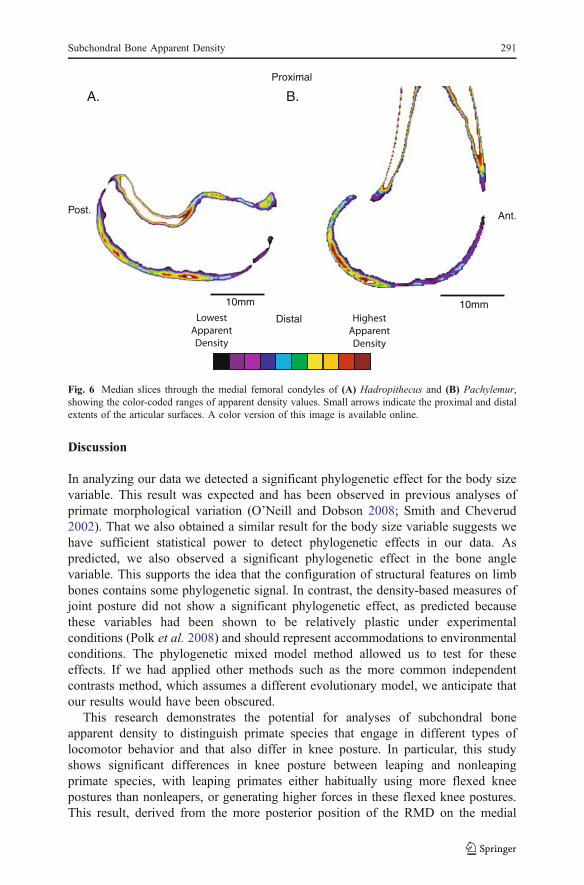
Inferring postural variation for extinct species requires more information thana single measurement. Indeed our summed angular estimate of knee posture is,by design, an overestimate of the actual postures that would have been used.For this reason we include images of the median slices through the MFC ofHadropithecus and Pachylemur (Fig. 6). These images show regions of highapparent density located in distal positions on their articular surfaces, but alsoregions of high apparent density more proximally or posteriorly on the articularsurfaces. This suggests that both of these species were loading their joints in arange of postures.
Table III Pairwise comparisons among locomotor groups
A. Bone angleLocomotor
Group aq tq ql Kw s lAq 0.617 0.045 0.551 0.551 0.150Tq 0.332 0.355 0.165 0.624Ql 0.715 0.715 0.752Kw 1.000 0.439S 0.857L
B. Density angleLocomotor
Group aq tq ql Kw s lAq 0.467 0.003 1.000 0.715 0.399Tq 0.090 0.643 0.355 0.462Ql 0.114 0.693 0.336Kw 0.439 0.439S 1.000L
C. SumLocomotor
Group aq tq ql Kw s lAq 0.361 0.003 0.236 0.844 0.117Tq 0.090 0.643 0.643 0.149Ql 0.048 0.844 0.794Kw 0.439 0.064S 0.800L
One-tailed exact probabilities. Predicted differences are underlined. Observed significant differences areshown in bold. Abbreviations follow Table I.
290 J. D. Polk et al.
Discussion
In analyzing our data we detected a significant phylogenetic effect for the body sizevariable. This result was expected and has been observed in previous analyses ofprimate morphological variation (O’Neill and Dobson 2008; Smith and Cheverud2002). That we also obtained a similar result for the body size variable suggests wehave sufficient statistical power to detect phylogenetic effects in our data. Aspredicted, we also observed a significant phylogenetic effect in the bone anglevariable. This supports the idea that the configuration of structural features on limbbones contains some phylogenetic signal. In contrast, the density-based measures ofjoint posture did not show a significant phylogenetic effect, as predicted becausethese variables had been shown to be relatively plastic under experimentalconditions (Polk et al. 2008) and should represent accommodations to environmentalconditions. The phylogenetic mixed model method allowed us to test for theseeffects. If we had applied other methods such as the more common independentcontrasts method, which assumes a different evolutionary model, we anticipate thatour results would have been obscured.
This research demonstrates the potential for analyses of subchondral boneapparent density to distinguish primate species that engage in different types oflocomotor behavior and that also differ in knee posture. In particular, this studyshows significant differences in knee posture between leaping and nonleapingprimate species, with leaping primates either habitually using more flexed kneepostures than nonleapers, or generating higher forces in these flexed knee postures.This result, derived from the more posterior position of the RMD on the medial
10mm10mm
Ant.Post.
Proximal
Distal HighestApparentDensity
LowestApparentDensity
B.A.
Fig. 6 Median slices through the medial femoral condyles of (A) Hadropithecus and (B) Pachylemur,showing the color-coded ranges of apparent density values. Small arrows indicate the proximal and distalextents of the articular surfaces. A color version of this image is available online.
Subchondral Bone Apparent Density 291

femoral condyle suggests that leaping primates are generating substantial joint loadswhen they leap from flexed knee postures. We had anticipated that vertical clinging andleaping (VCL) primates that habitually use more flexed postures than quadrupedalleapers (Demes et al. 1999; Fleagle 1999; Walker 1983), would have these posturaldifferences reflected in their patterns of subchondral apparent density. However, thefact that VCL and quadrupedal leapers have similar positions of their RMD (Fig. 4)suggests that joint loads incurred in more extended postures may be substantial. Forexample, VCL primates may incur high joint forces during landing on relativelyextended knees (Demes et al. 1999), or the use of fairly extended postures of sifakasduring bipedal locomotion may be sufficient to cause a somewhat anterior placementof their RMD. Increasing the sample size of leaping species in future studies may befruitful because we anticipate that large- and small-bodied leapers might differ in thepostures used during propulsion (cf. Demes and Günther 1989).
With respect to the nonleaping species, knuckle-walking apes and arborealquadrupeds were both significantly different from the leaping species in the positionof the RMD on the MFC. Terrestrial quadrupedal species did not differ significantlyfrom the leapers or from arboreal species. That apes use more extended postures thansmaller primate species may be a consequence of several factors but body size mustbe one of the most important. The relationship between body size and joint posturehas been described previously in diverse mammalian samples (Biewener 1989,1990) as well as in primates (Polk 2002; Polk et al. 2009). Body size differences arealso a likely explanation for the broad range of angular measurements for suspensoryprimates. Gibbons appear to use fairly flexed knee postures (Fig. 3), a resultconsistent with qualitative assessment of gibbon bipedalism (Fleagle 1980), whereasorangutans used more extended postures comparable to those of knuckle walkersand other quadrupeds. The fact that extant terrestrial quadrupeds did not differ fromleapers or more arboreal species may be explained by either of two possibilities.First, terrestrial primate species do make significant use of trees and other supportsthat require clambering and climbing, e.g., Chlorocebus aethiops: Fedigan andFedigan (1988). As a consequence, terrestrial species may also make habitual use offlexed hindlimb postures in these circumstances, and such loading may be sufficientto leave an osteological signal. There may also be differences between the forelimband hind limbs in compliance and weight support. Schmitt (1994, 1999) hasdemonstrated compliance in the forelimb but comparable data are lacking forprimate hind limbs. Second, terrestrial quadrupeds are poorly represented in oursample. The inclusion of more specimens, and more of the larger species of Papio,or more committed terrestrial genera such as Erythrocebus in future analyses mayhelp to clarify these relationships. However, exclusion of the Chlorocebus aethiopsspecimen from the terrestrial group did not alter the observed results.
The density magnitudes observed in the subchondral bone of the subfossilspecimens, overlap with the density magnitudes observed in extant primate species.This observation permits a preliminary rejection of the idea that there has beensignificant mineralization or degradation of the subchondral bone that would impairassessment of the patterns of apparent density. As a consequence, we believe it isreasonable to make comparisons between the subfossil and extant species and tomake inferences about possible differences in joint loading conditions duringlocomotion. It is important to note that this conclusion is preliminary because our
292 J. D. Polk et al.

method for quantifying patterns of density is relatively coarse, and diageneticchanges to bone structure may be difficult to detect using medical CT. For example,the bone may have increased in porosity, thereby reducing subchondral mass, but itis conceivable that such decreases in apparent density could have been offset byinfilling with sediment with densities similar to bone. We are not able to assess thesechanges without more detailed methods, e.g., micro-CT, for assessing eithersubchondral structure or the sediments from which the bones were extracted. Moreresearch characterizing the diagenetic changes to apparent density should beundertaken in the future. Nevertheless, our qualitative inspection of the boneexterior, combined with the fact that subfossil apparent density values for thesubchondral bone overlap with comparable measures for extant specimens, providesus with no evidence for a diagenetic effect.
In this study, based on the position of the region of maximal apparent density ontheir medial femoral condyles, we infer that both Pachylemur and Hadropithecusused very extended joint postures (171° and 188°, respectively). These estimates arecertainly overestimates of the actual knee postures that the animals used since it isunlikely that Hadropithecus would have walked with hyperextended knees, and bothspecies have elongated regions of high apparent density consistent with loading inmore flexed postures. We should discuss some caveats; for example, we obtained theρ and θ used for Hadropithecus from different specimens, and this may introducesome bias. Similarly, our overestimate of joint posture is a function of the method weused for estimating posture because we cannot know the position of the ankle andobtain estimates of posture that are completely comparable to kinematic measure-ments under experimental conditions. Nevertheless, the maximum postural estimatesobtained here provide useful estimates of the relative differences between species.When these posture estimates are compared to those of extant primates we can seethat Pachylemur falls at the upper end of the sampled extant primate range, whereasHadropithecus appeared to use even more extended postures than we observed inour extant sample. The postural estimate for Hadropithecus is consistent with thespecies’ body size because large-bodied extant quadrupeds tend to use moreextended postures than smaller-bodied species (Polk 2002; Polk et al. 2009). Inaddition, the medial femoral condyle has a distinctly flattened distal surface inHadropithecus. This flattening differs from most extant primate species and is mostreminiscent of the condition in humans (Lovejoy 2007). Although many othercharacteristics of the joint surface differ from humans, this flattening could help toenable greater stability of the tibiofemoral articulation in extended postures. The morerounded, posterior portion of the condyle would have enabled substantial flexion. Inthe Pachylemur specimen, we did not anticipate the reconstruction of such anextended knee posture. The more posterior orientation of the medial femoral condyle,as identified by the angle ρ, implies a very posterior “set” to the articular surface.However, the fact that the RMD was very distally positioned clearly indicates that thejoint surface was being loaded in extended postures at least some of the time. Figure 6illustrates use of a broader range of joint postures. That is, the RMD for Pachylemur isspread along a substantial portion of the articular surface, whereas the RMD forHadropithecus suggests loading in both extended and flexed postures.
The use of such extended postures by these 2 species clearly differentiates them bothfrom extant leaping primates. This is not surprising because previous studies of
Subchondral Bone Apparent Density 293

locomotor habits of both Pachylemur and Hadropithecus identified no leapingcharacteristics. Given that these species inhabited spiny thicket, succulent woodlandand dry deciduous forest environments (Godfrey and Jungers 2002; Muldoon et al.2009), where arboreal substrates can be widely spaced, it is likely that both of themwould have had to descend to the ground to move between trees and food resources. Itis likely that both species used extended postures while on terrestrial substrates butmore flexed postures while in the trees. The locomotor behavior of Pachylemur appearsto have been somewhat different from extant lemurs, e.g., Varecia, Lemur. Forexample, the vertebral morphology of Pachylemur (weak anticliny of the neural spines,shorter vertebral bodies) suggests the presence of a relatively stiff back and decreasedamplitude of flexion and extension (Ravololonarivo 1990; Shapiro et al. 2005), whichwould have diminished the ability of Pachylemur to bound and leap quadrupedally inthe manner of its closest living relative, Varecia. In comparison to Varecia and otherliving lemurids, Pachylemur limbs are shorter and more robust relative to the vertebralcolumn. The limb proportions (high humerofemoral index, high brachial index, andlow crural index) and associated interlimb robusticities (the humerus is not robust incomparison to the femur, as is typical of leapers) differ markedly from those extantlemurids and indriids. In addition, Tardieu and Jouffroy (1979) described differences inthe distal femoral proportions between Pachylemur and other lemurids which may berelated to its more deliberate slow climbing (see also Jouffroy 1960). Whereas theynote that, in some ways, the knee joint of Pachylemur was more like that of indriids,they do not relate these similarities to leaping. Godfrey (1988) suggested, on the basisof limb proportions and the morphology of the hip joint, especially the relatively largesize of the femoral head, that the locomotor repertoire of Pachylemur included morehind limb suspension than is typical for Varecia and living indriids, which also engagein hindlimb suspension, particularly while feeding. The behavioral differences betweenPachylemur and living lemurids and indriids may also lie in the degree to whichterrestrial pathways are or were used. Walker (1974) considered greater terrestriality inPachylemur a distinct possibility, based on its greater robusticity and roughly equalfore- and hind limb lengths; certainly, in the open habitats in which Pachylemur lived,assuming that leaping was not an important part of its locomotor repertory, it will havehad to descend to the ground to travel more often than agile leapers in the same habitat.However, in its overall morphology, and unlike Hadropithecus, there is little to signalterrestriality. In summary, Pachylemur appears to have been a relatively slow arborealquadruped, leaping and bounding less often but likely engaging in hind limbsuspension more often than its living relatives. The use of extended postures inPachylemur was not expected but might be explained either by more time spent in hindlimb suspension or in moving on the ground. However, it is clear that this species useda broad range of postures in arboreal settings.
We have observed a weak but significant correlation between our size variableand the maximum observed apparent subchondral density among the extant taxaincluded in this study. Because ground reaction forces (Alexander 1985), andpresumably knee joint reaction forces, correlate highly with body mass, our observedcorrelation between density and size suggests that there is a weak relationshipbetween joint loading and the osteological response of the subchondral bone. Thefact that this relationship is not stronger deserves further investigation. Some of thevariation may be due to differences in preferred or habitual speeds used during
294 J. D. Polk et al.

locomotion (cf. Alexander 1985), or there may be locomotor differences as well. Wewill investigate this variation in a future study.
Finally, it is important to discuss some of the limitations of the technique that weapply. We use the analyses of patterns of subchondral apparent density to makeinferences about the knee postures used by different species. We make this inferencebecause previous studies have suggested that subchondral apparent density appearsto respond to long-term patterns of joint loading (Carlson and Patel 2006; Müller-Gerbl et al. 1992; Patel and Carlson 2007; Polk et al. 2009). Additional tests need tobe performed to assess better whether rare but high-magnitude loads could causesimilar changes in the distribution of apparent density patterns across thesubchondral plate. In this study we examined only the anteriormost extent of theRMD as an indicator of habitual posture since this measure was experimentallyvalidated by Polk et al. (2008). There are other signals that remain to be examined,such as the breadth of the RMD along the condyle, or its posteriormost extent, bothof which could provide some indication of any limits to joint loading and kneeposture. Future studies could also incorporate apparent density patterns on the lateralfemoral condyle or on the femoral head to assess the degree of femoral abduction, orhip loading patterns, respectively.
In summary, in this study we analyzed patterns of subchondral apparent density in themedial femoral condyle of extant primates and two species of subfossil lemurs, in anattempt to reconstruct differences in knee posture. Our results show clear discriminationbetween the postures habitually used by leaping and nonleaping primates, with leapingspecies regularly using more flexed knee postures. The subfossil lemur species bothappear to have used a range of knee postures but seemed to have included ratherextended knee postures in their repertoire. This finding is most consistent with their lackof specialized adaptations for leaping, and in the case ofHadropithecus, with the use ofterrestrial substrates. Pachylemur likely employed extended knee postures in otheractivities such as hind limb suspension, in addition to occasional terrestrial locomotion.
Acknowledgments We thank Profs. E. Hirasaki, Y. Hamada, and T. C. Rae for their invitation to participatein the Primate FunctionalMorphology Symposium in August 2008.We also thank Judy Chupasko and her staffat Harvard’s Museum of Comparative Zoology for their generous loan of extant primate specimens for CTscanning. We thank Dr. Armand Rasoamiaramanana and the late Dr. Gisèle Randria (formerly Ravololonarivo)for the specimen (Pachylemur) loaned from the University of Antananarivo and for the paleontologicalresearch accord under which the Hadropithecus specimen was found. Kevin Reynolds and his staff at MountAuburn Hospital provided excellent assistance in CT scanning. Jennifer Swartz assisted with preliminaryanalyses of the patterns of subchondral density. Rebecca Stumpf provided excellent advice at all stages of theproject. This research was supported by grants from the National Science Foundation: BCS 0639630, withREU supplement (J. D. Polk), and BCS 0129185 (D. Burney, W. Jungers, and L. R. Godfrey), and theUniversity of Illinois, Urbana-Champaign (J. D. Polk). We also thank the editors, Bill Jungers, and 1anonymous reviewer for their substantial improvements to the manuscript.
Appendix
Phylogeny in Newick format
((((Nycticebus:35,Perodicticus:35):5,(Otolemur:20,Galago:20):20):33.7,(((((Eule-mur fulvus:9.2,Eulemur rubriventer:9.2):10.1,Hapalemur:19.3):4.1,Vare-
Subchondral Bone Apparent Density 295

cia:23.4):15.9,((Indri:29,Propithecus:29):6.9,Lepilemur: 35.9):3.4):26.9,Daubento-nia:66.2):7.5):1.5,((((Alouatta belzebuth: 4.1,A.fusca:4.1):9.5,Ateles:13.6): 6.5,((Saguinus:8.1,Leontopithecus:8.1):2,Callithrix: 10.1):10):19.9,((((((Macacamulatta:1.7,M. fascicularis:1.7):1.1,M.nemestrina:2.8):5,(Cercocebus: 6.5,Mandril-lus:6.5):1.3):1,((Miopithecus: 4,Chlorocebus:4):1,(Cercopithecus diana:5,Cercopithe-cus l’hoesti:5):0):3.3):4,(((Colobus guereza:0.1,C. polykomos:0.1):1.9,Piliocolobusbadius:2): 9,(((Trachypithecus cristata:1,t. vetulus:1,T. phayrei:1): 1,Presbytishosei:2):2.5,Nasalis:4.5):6.5):2):2,(((Pan:7.7,Gorilla:7.7):8.3,Pongo:16):3,Hylo-bates: 19):6):15):35.2);
References
Ahluwalia, K. (2000). Knee joint load as determined by tibial subchondral bone density: Its relationshipto gross morphology and locomotor behavior in catarrhines [PhD]. Stony Brook, NY: Stony BrookUniversity, 315 pp.
Alexander, R. M. (1985). The maximum forces exerted by animals. Journal of Experimental Biology, 115,231–238.
Biewener, A. A. (1989). Scaling body support in mammals: Limb posture and muscle mechanics. Science,245, 45–48.
Biewener, A. A. (1990). Biomechanics of mammalian terrestrial locomotion. Science, 250, 1097–1103.Burgess, N. D., D’Amico Hales, J., Underwood, E. C., Dinerstein, E., Olson, D., Itoua, I., et al. (2004).
Terrestrial ecoregions of Africa and Madagascar: A conservation assessment. Washington, DC:Island Press.
Burney, D. A., Burney, L. P., Godfrey, L. R., Jungers, W. L., Goodman, S. M., Wright, H. T., et al. (2004).A chronology for late prehistoric Madagascar. Journal of Human Evolution, 47, 25–63.
Burney, D. A., Vasey, N., Godfrey, L. R., Ramilisonina, Jungers, W. L., Ramarolahy, M., et al. (2008).New findings at Andrahomana Cave, Southeastern Madagascar. Journal of Cave and Karst Studies,70(1), 13–24.
Carlson, K. J., & Patel, B. A. (2006). Habitual use of the primate fore-limb is reflected in the materialproperties of subchondral bone in the distal radius. Journal of Anatomy, 208, 659–670.
Carter, D. R., & Beaupre, G. S. (2001). Skeletal form and function: Mechanobiology of skeletaldevelopment, aging and regeneration. Cambridge, UK: Cambridge University Press.
Carter, D. R., & Hayes, W. C. (1977). The compressive behavior of bone as a two phase porous structure.Journal of Bone and Joint Surgery, 59A, 954–962.
Catlett, K. K., Schwartz, G. T., Godfrey, L. R., & Jungers, W. L. (2010). “Life History Space”: Amultivariate analysis of life history variation in extant and extinct Malagasy lemurs. American Journalof Physical Anthropology. doi:10.1002/ajpa.21236.
Cohen, J. (1990). Things I have learned (so far). American Psychologist, 45, 1304–1312.Crowley, B. E., Godfrey, L. R., & Irwin, M. T. (2010). A glance to the past: Subfossils, stable isotopes, seed
dispersal, and lemur species loss in southern Madagascar. American Journal of Primatology, 71, 1–13.Currey, J. D. (1988). The effect of porosity and mineral content on the Young’s modulus of elasticity of
compact bone. Journal of Biomechanics, 21, 131–139.Currey, J. D. (2002). Bones: Structure and mechanics. Princeton, NJ: Princeton University Press.Decary, R. (1926). Une mission scientifique dans le S.E. de Madagascar. Bull. de l’Académie Malgache, 9,
79–86.Demes, B., & Günther, M. M. (1989). Biomechanics and allometric scaling in primate locomotion and
morphology. Folia Primatologica, 53318, 125–141.Demes, B., & Carlson, K. J. (2009). Locomotor variation and bending regimes of capuchin limb bones.
American Journal of Physical Anthropology, 139, 558–571.Demes, B., Larson, S. G., Stern, J. T., Jungers, W. L., Biknevicius, A. R., & Schmitt, D. (1994). The
kinematics of primate quadrupedalism: “Hindlimb drive” reconsidered. Journal of Human Evolution,26, 353–374.
Demes, B., Fleagle, J. G., & Jungers, W. L. (1999). Takeoff and landing forces of leaping strepsirhineprimates. Journal of Human Evolution, 37, 279–292.
296 J. D. Polk et al.

Eckstein, F., Müller-Gerbl, M., Steinlechner, M., Kierse, R., & Putz, R. (1995). Subchondral bone densityin the human elbow assessed by computed tomography osteoabsorptiometry: A reflection of theloading history of the joint surfaces. Journal of Orthopedic Research, 13, 268–278.
Eckstein, F., Merz, B., Schon, M., Jacobs, C. R., & Putz, R. (1999). Tension and bending, but notcompression alone, determine the functional adaptation of subchondral bone in incongruous joints.Anatomy and Embryology, 199(1), 85–97.
Fedigan, L, & Fedigan, L. M. (1988). Cercopithecus aethiops: a review of field studies. In A. Gautier-Hion, R. Fouliere, J-P. Gautier, & J. Kingdon (Eds.), A primate radiation: Evolutionary biology of theAfrican guenons. Cambridge, UK: Cambridge University Press.
Felsenstein, J. (1985). Phylogenies and the comparative method. American Naturalist, 125, 1–15.Fischer, K. J., Jacobs, C. R., & Carter, D. R. (1995). Computation method for determination of bone and
joint loads using bone density distributions. Journal of Biomechanics, 28, 1127–1135.Fleagle, J. G. (1976). Locomotor behavior and skeletal anatomy of sympatric Malaysian leaf monkeys
(Presbytis obscura and Presbytis melalophos). Yearbook of Physical Anthropology, 20, 440–453.Fleagle, J. G. (1980). Locomotion and posture. In D. J. Chivers (Ed.), Malayan Forest primates (191–
207). New York: Plenum Press.Fleagle, J. G. (1999). Primate adaptation and evolution. New York: Academic Press.Godfrey, L. R. (1988). Adaptive diversification of Malagasy strepsirrhines. Journal of Human Evolution,
17, 93–134.Godfrey, L. R., & Jungers, W. L. (2002). Quaternary fossil lemurs. In W. C. Hartwig (Ed.), The primate
fossil record (pp. 97–121). Cambridge, UK: Cambridge University Press.Godfrey, L. R., Jungers, W. L., Wunderlich, R. E., & Richmond, B. G. (1997). Reappraisal of the
postcranium of Hadropithecus (Primates, Indroidea). American Journal of Physical Anthropology,103, 529–556.
Godfrey, L. R., Semprebon, G. M., Jungers, W. L., Sutherland, M. R., Simons, E. L., & Solounias, N.(2004). Dental use wear in extinct lemurs: Evidence of diet and niche differentiation. Journal ofHuman Evolution, 47, 145–169.
Godfrey, L. R., Jungers, W. L., Burney, D. A., Vasey, N., Ramilisonina, Wheeler, W., et al. (2006a). Newdiscoveries of skeletal elements of Hadropithecus stenognathus from Andrahomana Cave,southeastern Madagascar. Journal of Human Evolution, 51(4), 395–410.
Godfrey, L. R., Jungers, W. L., & Schwartz, G. T. (2006b). Ecology and extinction of Madagascar’ssubfossil lemurs. In L. Gould & M. Sauther (Eds.), Lemurs: Ecology and adaptation (pp. 41–64).New York: Springer.
Godfrey, L. R., Jungers, W. L., Schwartz, G. T., & Irwin, M. T. (2008a). Ghosts and orphans:Madagascar’s vanishing ecosystems. In J. G. Fleagle & C. C. Gilbert (Eds.), Elwyn Simons: A searchfor origins (pp. 361–395). New York: Springer.
Godfrey, L. R., Crowley, B. E., Muldoon, K. M., King, S. J., & Burney, D. A. (2008b). TheHadropithecus conundrum. American Journal of Physical Anthropology (Supplement), 46, 105.
Grandidier, G. (1905). Recherches sur les lémuriens disparus et en particulier sur ceux qui vivaient àMadagascar. Nouvelles Archives du Muséum, 7, 1–142, plus 12 plates.
Hanna, J. B., Polk, J. D., & Schmitt, D. (2006). Forelimb and hindlimb forces in walking and gallopingprimates. American Journal of Physical Anthropology, 130(4), 529–535.
Harvey, P. H., & Pagel, M. D. (1991). The comparative method in evolutionary biology. Oxford: OxfordUniversity Press.
Horvath, J. E., Weisrock, D. W., Embry, S. L., Fiorentino, I., Balhoff, J. P., Kappeler, P., et al. (2008).Development and application of a phylogenomic toolkit: Resolving the evolutionary history ofMadagascar’s lemurs. Genome Research, 18, 489–499.
Hoshino, A., & Wallace, W. A. (1987). Impact-absorbing properties of the human knee. Journal of Boneand Joint Surgery, 69B, 807–811.
Housworth, E. A., Martins, E. P., & Lynch, M. (2004). The phylogenetic mixed model. AmericanNaturalist, 163, 84–96.
Jouffroy, F. K. (1960). Caractères adaptatifs dans les proportions des members chez les Lémurs fossiles.Comptes Rendus de l’Académie des Sciences Paris, 251, 2756–2757.
Jungers, W. L. (1988). Relative joint size and hominoid locomotor adaptations with implications for theevolution of hominid bipedalism. Journal of Human Evolution, 17, 247–265.
Jungers, W. L. (1991). Scaling of postcranial joint size in hominoid primates. Human Evolution, 6, 391–399.
Jungers, W. L., Falsetti, A. B., & Wall, C. E. (1995). Shape, relative size, and size-adjustments inmorphometrics. Yearbook of Physical Anthropology, 1995, 137–161.
Subchondral Bone Apparent Density 297

Jungers, W. L., Demes, B., & Godfrey, L. R. (2008). How big were the ‘giant’ extinct lemurs ofMadagascar? In J. G. Fleagle & C. C. Gilbert (Eds.), Elwyn Simons: A search for origins (pp. 343–360). New York: Springer.
Lemelin, P., Hamrick, M. W., Richmond, B. G., Godfrey, L. R., Jungers, W. L., & Burney, D. A. (2008).New hand bones of Hadropithecus stenognathus: Implications for the paleobiology of theArchaeolemuridae. Journal of Human Evolution, 54, 405–413.
Lovejoy, C. O. (2007). The natural history of human gait: Part 3. The knee. Gait & Posture, 25, 325–341.Lynch, M. (1991). Methods for the analysis of comparative data in evolutionary biology. Evolution, 45,
1065–1080.McHenry, H. M. (1988). New estimates of body weight in early hominids and their significance to
encephalization and megadontia in “robust” Australopithecines. In F. E. Grine (Ed.), Evolutionaryhistory of the “robust” Australopithecines (pp. 133–148). New York: Aldine de Gruyter.
McHenry, H. M., & Berger, L. R. (1998). Body proportions in Australopithecus afarensis and A. africanusand the origin of the genus Homo. Journal of Human Evolution, 35(1), 1–22.
Muldoon, K. M., Godfrey, L. R., & Jungers, W. L. (2009). Geographic patterning in subfossil primatecommunity dynamics in Madagascar. American Journal of Physical Anthropology (Supplement), 48,195.
Müller-Gerbl, M., Putz, R., Hodapp, N., Schulte, E., & Wimmer, B. (1989). Computed tomography-osteoabsorptiometry for assessing the density distribution of subchondral bone as a measure of long-term mechanical adaptation in individual joints. Skeletal Radiology, 18(7), 507–512.
Müller-Gerbl, M., Putz, R., Hodapp, N., Schulte, E., & Wimmer, B. (1990). Computed-tomographyosteoabsorptiometry – A method of assessing the mechanical condition of the major joints in a livingsubject. Clinical Biomechanics, 5(4), 193–198.
Muller-Gerbl, M., Putz, R., & Kenn, R. (1992). Demonstration of subchondral bone-density patterns by 3-dimensional Ct osteoabsorptiometry as a noninvasive method for in vivo assessment of individuallong-term stresses in joints. Journal of Bone and Mineral Research, 7, S411–S418.
O’Neill, M. C., & Dobson, S. D. (2008). The degree and pattern of phylogenetic signal in primate long-bone structure. Journal of Human Evolution, 54, 309–322.
Orlando, L., Calvignac, S., Schnebelen, C., Douady, C. J., Godfrey, L. R., & Hänni, C. (2008). DNA fromextinct giant lemurs links archaeolemurids to extant indriids. BMC Evolutionary Biology, 8, Articleno. 121.
Paradis, E. (2006). Analysis of phylogenetics and evolution with R. New York; Springer.Patel, B. A., & Carlson, K. J. (2006). Hand postures reflect bone apparent density patterns in the primate
distal radius. American Journal of Physical Anthropology, 144–145.Patel, B. A., & Carlson, K. J. (2007). Bone density spatial patterns in the distal radius reflect habitual hand
postures adopted by quadrupedal primates. Journal of Human Evolution, 52, 130–141.Patel, B. A., & Carlson, K. J. (2008). Apparent density patterns in subchondral bone of the sloth and
anteater forelimb. Biology Letters, 4(5), 486–489.Polk, J. D. (2002). Adaptive and phylogenetic influences on musculoskeletal design in cercopithecine
primates. Journal of Experimental Biology, 205(21), 3399–3412.Polk, J. D. (2004). The influence of body size and limb proportions on joint posture in terrestrial monkeys
and fossil hominins. Journal of Human Evolution, 47, 237–252.Polk, J. D., Blumenfeld, J., & Ahluwalia, K. (2008). Knee posture predicted from subchondral apparent
density in the distal femur: An experimental validation. Anatomical Record, 291, 293–302.Polk, J. D., Williams, S. A., & Peterson, J. V. (2009). Body size and joint posture in Primates. American
Journal of Physical Anthropology, 140, 359–375.Radin, E. L., & Paul, I. L. (1971). The importance of bone in sparing articular cartilage from impact.
Clinical Orthopaedics and Related Research, 78, 342–344.Radin, E. L., Paul, I. L., & Lowy, M. (1970). A comparison of the dynamic force transmitting properties
of subchondral bone and articular cartilage. Journal of Bone and Joint Surgery, 52A, 444–456.Ravololonarivo, G. F. N. (1990). Contribution à l’étude de la colonne vertébrale du genre Pachylemur
(Lamberton, 1946): Anatomie et analyse cladistique. 3 rd Cycle thesis, University of Antananarivo,Madagascar.
Reynolds, T. R. (1985). Mechanics of increased support of weight by the hindlimb in primates. AmericanJournal of Physical Anthropology, 67, 335–349.
Rice, J. C., Cowin, S. C., & Bowman, J. A. (1988). On the dependency of the elasticity and strength ofcancellous bone on apparent density. Journal of Biomechanics, 21, 155–168.
Rohlf, F. J. (2006). Tpsdig2: http://life.bio.sunysb.edu/morph/soft-dataacq.html.
298 J. D. Polk et al.

Ruff, C. (1987). Structural allometry of the femur and tibia in Hominoidea and Macaca. FoliaPrimatologica, 48, 9–49.
Ruff, C. (1988). Hindlimb articular surface allometry in Hominoidea and Macaca, with comparisons todiaphyseal scaling. Journal of Human Evolution, 17(7), 687–714.
Ruff, C. B., & Leo, F. P. (1986). Use of computed tomography in skeletal structure research. Yearbook ofPhysical Anthropology, 29, 181–196.
Ryan, T. M., Burney, D. A., Godfrey, L. R., Göhlich, U., Jungers, W. L., Vasey, N., et al. (2008). Areconstruction of the Vienna skull of Hadropithecus stenognathus. Proceedings of the NationalAcademy of Sciences of the United States of America, 105(31), 10698–10701.
Samii, V. F., Les, C. M., Schulz, K. S., Keyak, J. H., & Stover, S. M. (2002). Computed tomographicosteoabsorptiometry of the elbow joint in clinically normal dogs. American Journal of VeterinaryResearch, 63, 1159–1166.
Schmitt, D. (1994). Forelimb mechanics as a function of substrate type during quadrupedalism in twoanthropoid primates. Journal of Human Evolution, 26, 441–457.
Schmitt, D. (1999). Compliant walking in Primates. Journal of Zoology (London), 248, 149–160.Shapiro, L. J., Seiffert, C. V. M., Godfrey, L. R., Jungers, W. L., Simons, E. L., & Randria, G. F. N.
(2005). Morphometric analysis of lumbar vertebrae in extinct Malagasy strepsirrhines. AmericanJournal of Physical Anthropology, 128(4), 823–839.
Shapiro, S. S., & Wilk, M. B. (1965). An analysis of variance test for normality (complete samples).Biometrika, 52(3/4), 591–611.
Simon, S. R., Rose, R. M., Radin, E. L., & Paul, I. L. (1972). Response of joints to impact loading 2. In-vivo behavior of subchondral bone. Journal of Biomechanics, 5(3), 267–272.
Smith, R. J., & Cheverud, J. M. (2002). Scaling of sexual dimorphism in body mass: A phylogeneticanalysis of Rensch’s rule in Primates. International Journal of Primatology, 23, 1095–1135.
Sokal, R. R., & Rohlf, F. J. (1995). Biometry (3rd ed.). New York: W. H. Freeman.Tardieu, C., & Jouffroy, F. K. (1979). Les surfaces articulaires fémorales du genou chez les Primates:
étude préliminaire. Annales des Sciences Naturelles Zoologie (Paris), 13, 23–38.Taylor, W. R., Ehrig, R. M., Heller, M. O., Schell, H., Seebeck, P., & Duda, G. N. (2006). Tibio-femoral
joint contact forces in sheep. Journal of Biomechanics, 39(5), 791–798.von Eisenhart, R., Adam, C., Steinlechner, M., Muller-Gerbl, M., & Eckstein, F. (1999). Quantitative
determination of joint incongruity and pressure distribution during simulated gait and cartilagethickness in the human hip joint. Journal of Orthopaedic Research, 17(4), 532–539.
Walker, A. C. (1974). Locomotor adaptations in past and present prosimian primates. In F. A. Jenkins Jr.(Ed.), Primate locomotion (pp. 349–381). New York: Academic Press.
Walker, A. (1983). Prosimian locomotor behavior. In G. A. Doyle & R. D. Martin (Eds.), The study ofprosiman behavior (pp. 543–566). New York: Academic Press.
Walker, A., Ryan, T. M., Silcox, M. T., Simons, E. L., & Spoor, F. (2008). The semicircular canal systemand locomotion: The case of extinct lemuroids and lorisoids. Evolutionary Anthropology, 17(3), 135–145.
Wall, J. C., Chatterji, S. K., & Jeffery, J. W. (1979). Age-related changes in the density and tensile strengthof human femoral cortical bone. Calcified Tissue International, 27, 105–108.
Wiley, D. F., Amenta, N., Alcantara, D. A., Ghosh, D., Kil, Y. J., Delson, E., Harcourt-Smith, W., Rohlf, F.J., St. John, K., & Hamann, B. (2005). Evolutionary morphing. Proceedings IEEE Visualization,2005, 431–438.
Wright, T. M., & Hayes, W. C. (1977). Fracture mechanics parameters for compact bone – effects ofdensity and specimen thickness. Journal of Biomechanics, 10, 419–430.
Subchondral Bone Apparent Density 299




















