
Impact of the cycle-to-cycle variation of an internal combustion ...

94:1057-1065, 2005. First published Mar 30, 2005; doi:10.1152/jn.00991.2004 JNS. Yakovenko, D. A. McCrea, K. Stecina and A. Prochazka Control of Locomotor Cycle Durations
You might find this additional information useful...
39 articles, 13 of which you can access free at: This article cites http://jn.physiology.org/cgi/content/full/94/2/1057#BIBL
including high-resolution figures, can be found at: Updated information and services http://jn.physiology.org/cgi/content/full/94/2/1057
can be found at: Journal of Neurophysiologyabout Additional material and information http://www.the-aps.org/publications/jn
This information is current as of October 29, 2005 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the
publishes original articles on the function of the nervous system. It is published 12 times a yearJournal of Neurophysiology
on October 29, 2005
jn.physiology.orgD
ownloaded from

Control of Locomotor Cycle Durations
S. Yakovenko,1 D. A. McCrea,2 K. Stecina,2 and A. Prochazka3
1Department of Physiology, Universite de Montreal, Montreal, Quebec; 2Department of Physiology, University of Manitoba,Winnipeg, Manitoba; and 3Centre for Neuroscience, University of Alberta, Edmonton Alberta, Canada
Submitted 21 September 2004; accepted in final form 6 March 2005
Yakovenko, S., D. A. McCrea, K. Stecina, and A. Prochazka.Control of locomotor cycle durations. J Neurophysiol 94: 1057–1065,2005. First published April 7, 2005; doi:10.1152/jn.00991.2004. Inintact animals and humans, increases in locomotor speed are usuallyassociated with decreases in step cycle duration. Most data indicate thatthe locomotor central pattern generator (CPG) shortens cycle durationmainly by shortening the durations of extensor rather than flexor phasesof the step cycle. Here we report that in fictive locomotion elicited byelectrical stimulation of the midbrain locomotor region (MLR) in the cat,spontaneous variations in cycle duration were due more to changes inflexor rather than extensor phase durations in 22 of 31 experiments. Thelocomotor CPG is therefore not inherently extensor- or flexor-biased. Wecoined the term “dominant” to designate the phase (flexion or extension)showing the larger variation. In a simple half-center oscillator model,experimental phase duration plots were fitted well by adjusting twoparameters that corresponded to background drive (“bias”) and sensitivity(“gain”) of the oscillator’s timing elements. By analogy we argue thatvariations in background drive to the neural timing elements of the CPGcould produce larger variations in phase duration in the half-centerreceiving the lower background drive, i.e., background drive may deter-mine which half-center is dominant. The fact that data from normal catswere also fitted well by the model indicates that sensory input and centraldrive combine to determine locomotor phase durations. We conclude thatthere is a considerable flexibility in the control of phase durations inMLR-induced fictive locomotion. We posit that this may be explained bychanges in background excitation of neural timing elements in thelocomotor CPG.
I N T R O D U C T I O N
Many movements are rhythmical in nature, for examplebreathing, chewing, swimming, walking, and flying. In theseexamples, numerous muscles are cyclically activated in acoordinated sequence by neural commands. In mammals,rhythmical locomotor movements can be generated by thespinal cord even in the absence of supraspinal control (Brown1911). To explain the mechanism of rhythmogenesis, Brownposited a spinal intrinsic factor comprising mutually inhibitorygroups of neurons he named half-centers. The intrinsic factorwas later renamed the central pattern generator (CPG) (Grill-ner and Zangger 1975) and developed into a schematic of themammalian CPG in which interneurons involved in reorga-nized flexion reflexes functioned as half-centers coupled bymutual inhibition (Lundberg 1981). Although the precise loca-tion and structure of the locomotor network controlling thehind legs in vertebrates is poorly understood, a variety ofstudies indicate that it is widely distributed throughout thespinal cord (Cowley and Schmidt 1997; Kiehn and Kjaerulff1996) and can be activated pharmacologically using a variety
of neurotransmitters (Rossignol et al. 1998) or through reticu-lospinal pathways that in turn can be activated by stimulationof other brain stem areas such as the midbrain locomotorregion (MLR) (see (Jordan 1998; Noga et al. 2003).
Attempts have been made to deduce the functional organi-zation of the CPG on the basis of the mutability of the patternsof muscle activation during different locomotor behaviors, e.g.,comparing forward, backward, uphill, and downhill walking(Grillner 1981; Stein and Smith 1997). The results of theseinvestigations have been used to suggest that the CPG does notconsist of a simple flexion-extension oscillator controlling thewhole limb. Instead, a mechanism involving several oscillatorswas suggested (Grillner and Wallen 1985) in which each jointis controlled by its own “unit” oscillator that is flexibly coupledto the other unit oscillators of the limb. However, there is nocompelling evidence of localized regions of activation corre-sponding to separate oscillators controlling hip, knee, and ankle(Prochazka et al. 2002b). Furthermore, in fictive locomotion,preliminary evidence suggests there is a tight coupling betweenbursts of activity of extensor nerves of the entire limb and asimilar coupling of flexor bursts, hip flexors being slightly phase-advanced on ankle flexors (Quevedo et al. 2000b). Thus as a firstapproximation we consider the spinal CPG to be a bipartiteorganization that produces alternating and out-of-phase activationof flexor and extensor muscles throughout the hindlimb. Addi-tional circuitries also exist that shape the activity of individualmotoneuron pools, including those innervating bifunctional mus-cles, during various locomotor tasks (Burke 2001).
It is well known that locomotor cycle periods, phase transi-tions, and phase durations can be affected by descending inputs(Armstrong 1988; Drew 1991) as well as sensory input to thespinal cord (McCrea 2001; Pearson and Rossignol 1991; Ros-signol 1996). Afferent feedback from muscle proprioceptorsand low threshold cutaneous receptors is used continuously toshape and regulate motoneuron activity during stepping(Bouyer and Rossignol 1998; Prochazka et al. 2002a). Resultsfrom fictive locomotor preparations show that depending onwhich afferents are stimulated, segmental proprioceptive feed-back can be directed either to flexor or extensor components ofthe CPG (McCrea 2001). These observations suggest a flexibleorganization of the locomotor CPG that can be accessed andmanipulated by sensory feedback (as well as descending sys-tems) to control the activity and timing of both flexor andextensor phase motoneuron activity. Numerous conceptual andmathematical models of locomotor pattern generators and theirsensory control have been presented, e.g., Marder and Bucher2001; Pearson 2004; Yakovenko et al. 2004.
Address for reprint requests and other correspondence: A. Prochazka, Centrefor Neuroscience, 507 HMRC, University of Alberta, Edmonton, Alberta T6G2S2, Canada (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
J Neurophysiol 94: 1057–1065, 2005.First published April 7, 2005; doi:10.1152/jn.00991.2004.
10570022-3077/05 $8.00 Copyright © 2005 The American Physiological Societywww.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from

The timing of transitions between locomotor phases is cru-cial for matching the CPG output to actual limb forces andmotions. During normal overground gait in intact animals andhumans, an increase in the speed of locomotion is usuallyaccompanied by a decrease in the step cycle duration, mostlydue to the shortening of the stance phase. The duration of theswing phase changes much less (Arshavsky et al. 1965; Gos-low et al. 1973; Halbertsma 1983; Murray 1967). The same hasbeen reported in spinalized cats during treadmill locomotion(Forssberg and Grillner 1973; Prochazka et al. 2002b) and infictive locomotion (Dubuc et al. 1988; Leblond and Gossard1997; Pearson and Rossignol 1991). These results suggest thatthe CPG is hard-wired to produce adjustments of step cycleduration by mainly modulating the durations of extensor burstsrather than flexor bursts (Grillner and Dubuc 1988). This is incontrast to the flexibility of control of both the flexion andextension phases by afferent feedback.
In the present study, we sought to clarify whether thelocomotor CPG is extensor biased with a preferential control ofextensor phase duration. To this end, we used the fictivelocomotion preparation in adult decerebrate cats. The advan-tage of this reduced preparation is that without rhythmicafferent input and without cortical control, the locomotorpattern should reflect more of the operation of the basic spinalCPG than it would in more intact preparations. In this study,we report that in MLR-induced fictive locomotion in the cat,the locomotor cadence varied more as a result of changes inflexor rather than extensor phase durations in many prepara-tions. In a simple half-center model of the locomotor CPG,coupled changes in background excitation and sensitivity of thetiming elements reproduced the observed range of phase-duration properties. The results are consistent with the idea thatthe balance of drive to the flexor and extensor half-centers ofthe locomotor oscillator results either in flexor- or extensor-dominated control.
Portions of this work have appeared in Abstract form (Ya-kovenko et al. 2001).
M E T H O D S
Surgical procedures
All data reported in this paper originated from experiments done forother reasons in David McCrea’s laboratory, University of Manitoba(Gosgnach et al. 2000; Quevedo et al. 2000a). All surgical andexperimental protocols were in accordance with the guidelines set outby the Canadian Council for Animal Care. The experiments wereperformed on 23 cats (weight: 2.2–3.2 kg). The animals were anes-thetized with halothane delivered in a mixture of 30% oxygen and70% nitrous oxide for the duration of the surgery. Surgical proceduresconsisted of a tracheotomy, cannulation of arteries and veins, dissec-tions of various hindlimb nerves and a craniotomy. The position ofboth the ipsilateral and contralateral limbs was essentially the same inall preparations used in the present study. Furthermore, both limbswere extensively denervated by bilateral sectioning of the sciatic,femoral, and obturator nerves, and tendons surrounding the hips werecut. Mechanical, precollicular-postmamillary decerebration was per-formed by removing all the tissue rostral to the transection. Thisprocedure makes animals insentient (legally dead) and allows discon-tinuation of anesthetic. The neuromuscular blocker gallamine triethio-dide (Flaxedil, 2–3 mg � kg�1 � h�1) was used to abolish movementand the animals were artificially ventilated. Fictive locomotion waselicited by unilateral or bilateral monopolar electrical stimulation of
the MLR (30–200 �A, 0.5- to 1-ms pulses, 7–30 Hz) with tungstenmicroelectrodes. Selected hindlimb nerves were placed on conven-tional bipolar hook electrodes for recording. The following nerveswere included: the flexors sartorius (Sart), tibialis anterior (TA),extensor digitorum longus (EDL), peroneus longus (PerL), rectusfemoris (RF), biceps femoris posterior (BFp), or sometimes BFp andsemitendinosus together (PBST) and the extensors semimembranosusand biceps femoris anterior (SMAB), flexor hallucis longus (FHL),flexor digitorum longus (FDL), or sometimes FDL and FHL together(FDHL), gastrocnemius and soleus (GSol), plantaris (Plan), and quad-riceps (Quad).
Up to 12 nerves were recorded simultaneously in each cat. Alaminectomy was performed usually from the L4 to the S1 regions insome animals, but in others, it was more extensive, including thoracicand cervical segments. A lethal injection of pentobarbital anestheticwas administered at the end of the experiment.
Data recording and analysis
Automated analysis was applied to 31 episodes of MLR-evokedfictive locomotion from 23 different animals in which spontaneouschanges in the cycle timing occurred. Episodes in the same experi-ment were treated independently because they were recorded hoursapart, often with modified parameters of MLR stimulation. We ana-lyzed as many segments of ENG recording as were present in the datafrom a given episode. This amounted to an average of 117 individualcycles per episode. All individual cycles from a given run were thenanalyzed to produce phase-duration scatter plots of the type shown inFig. 2.
Electroneurogram (ENG) activity in the hindlimb muscle nerveslisted in the preceding text was filtered (30 Hz to 3 kHz), rectified,low-pass filtered (100 Hz), and digitized at 500 Hz. The waveformswere then analyzed to determine onsets and terminations of burstsusing an adaptive threshold crossing method described in the follow-ing text. Recordings with a signal-to-noise ratio �5 were rejected.Cycle period was determined as the time between two onsets of theENG envelope. We intentionally restricted the analysis to cycles withperiods between 0.3 and 1.5 s corresponding to speeds of overgroundlocomotion between 0.2 and 3.1 m/s (Halbertsma 1983). Regressionlines were fitted to scatter plots of burst duration and cycle period forindividual nerves. One-way ANOVA and multiple comparison ofregression line slopes with Tukey’s honestly significant differencecriterion was performed to determine significant differences (� �0.05).
ADAPTIVE THRESHOLD CROSSING METHOD. A novel means of de-termining the onsets and terminations of ENG bursts was developedfor this study. To understand the method, consider a portion of a noisysignal containing bursts of activity (Fig. 1A). Now consider a hori-zontal line representing a test level, moving upward through the signal(dashed lines Fig. 1A). Within the noise band, there are manyintersections of the signal and the test level. The number of intersec-tions reaches a peak somewhere within the noise band, as plotted inFig. 1B. As the test level moves above the noise band and into theregion of burst activity (dashed line b in Fig. 1, A and B), the numberof intersections drops and then remains constant. As the test levelmoves up into the region of peaks (line c) the number of intersectionsagain rises and then declines to zero (line d).
We developed a software algorithm to detect onsets and termina-tions of the ENG signal based on an examination of thresholdcrossings in the rectified-integrated ENG signal (Fig. 1B). For a givensegment of recording, the algorithm first constructs the intersectionsplot for test levels �20% of maximal signal amplitude (the 1st 8th ofthe plot in Fig. 1B). Next, the differential of this plot is computed(dotted line in Fig. 1B). A biphasic profile in this differential plot isnow sought. If it is absent, the noise is assumed to be �20% ofmaximal signal amplitude and the recording is discarded. If it is
1058 S. YAKOVENKO, D. A. MCCREA, K. STECINA, AND A. PROCHAZAKA
J Neurophysiol • VOL 94 • AUGUST 2005 • www.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from

present, the signal amplitude at which the differential returns to zeroafter the biphasic variation is determined. This is then used as apreliminary threshold to detect the times of burst onsets and termina-tions, which correspond to nerve activity immediately above the“noisy” background and depend on its peaks. Onsets are distinguishedfrom terminations according to the sign of slope of the signal as itcrosses threshold. Finally, more accurate burst timing is furthercalculated by adjusting each previously estimated onset and termina-tion to the nearest time the signal crosses the final threshold formed bythe summed mean and SD of “noise.” This last manipulation reducesthe error of underestimating the duration of activity common tostandard threshold methods and dependent on the slopes of activationand inactivation of each burst. The detection quality of the methodwas similar to that of manual selection with the obvious advantages ofautomation and objectivity.
The analysis of ENG activity may lead to underestimates of theduration of excitatory drive to the corresponding motoneuron pool.Significant errors in the estimation of the relationship between phaseand cycle duration may thereby emerge if the duration of the nerveactivity does not span all or most of the corresponding phase.However, in a separate analysis we confirmed that the mean ENG
profiles of the nerves analyzed in this preparation did in fact span allor most of the corresponding flexor or extensor phase. We estimatethat this limitation of the method only contributed small offsets to thelines of best fit reported in Figs. 2–4.
R E S U L T S
As discussed in the preceding text, previous studies oflocomotor control have demonstrated that in nearly all caseschanges in cycle period are due more to changes in stancephase duration than swing phase duration. Our study originatedfrom a chance observation that this was often reversed inMLR-induced fictive locomotion.
In a given cat, we found that either the extensor bursts or theflexor bursts occupied �50% of the step cycle period in nearlyall cycles (i.e., a given cat showed extensor- or flexor-domi-nated behavior). Furthermore, for a given change in cycleduration the amount of variation in burst duration depended onwhich of the two behaviors applied to that particular cat. Wetherefore decided to divide the data into two groups, corre-sponding to extensor- and flexor-dominated behavior.
Figure 2A shows a recording of nerve activity during fictivelocomotion evoked by the stimulation of MLR. Note that thebursts recorded in the flexor nerve (Sart) occupied �50% ofthe cycle durations, whereas those of the extensor nerve(SMAB) occupied �50% of the cycles. Using automaticthreshold detection as described in METHODS, the flexor andextensor phase durations were computed and plotted againstcycle duration in Fig. 2B. The - - - reference line has a slope of0.5 and passes through the origin. Data points on this linecorrespond to cycles in which phase duration is 50% of cycleduration. Data points above and below the - - - correspond tocycles in which phase duration is greater or �50% of cycleduration, respectively.
In Fig. 2B, flexor phase durations are mostly above the lineand extensor phase durations are mostly below. When theaverage of phase durations of all flexor nerves in an episode is�50% of cycle duration, we will refer to such patterns asflexor-dominated. Figure 2C shows recordings from anotherfictive locomotor cat. This time extensor bursts (GSol nerve)were longer than flexor bursts as in intact cats. Figure 2Dconfirms that in this case extensor phase durations are mostlyabove the line and flexor phase durations are mostly below.Because the average of phase durations of all extensor nervesin this episode is �50% of cycle duration, we call this anextensor-dominated pattern.
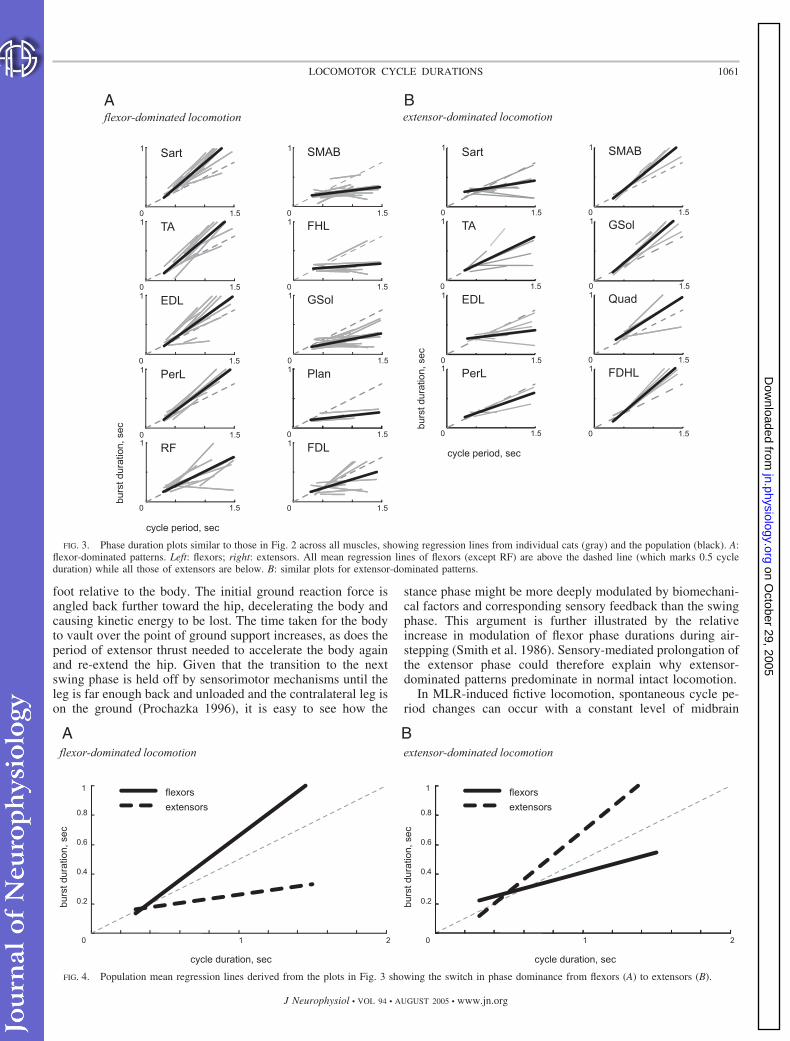
Figure 3 shows pooled data from 31 episodes in 23 fictiveMLR cats, 22 episodes showing flexor-dominated patterns(Fig. 3A) and 9 showing extensor-dominated patterns (Fig. 3B).The nerve recordings were divided into two groups, those fromthe flexors Sart, TA, EDL, PerL, and RF and those from theextensors SMAB, FHL, GSol, Plan, Quad, and FDL/FDHL.This grouping is based on the biomechanical action of thecorresponding muscles in flexor and crossed-extensor reflexesand the spatiotemporal activation of their motoneurons in thespinal cord during locomotion (Eccles and Lundberg 1958;Prochazka et al. 2002b; Sherrington 1910).
Each gray regression line represents a set of phase-durationdata for a given muscle nerve in a given cat. Solid lines are themeans of the individual regression lines. Most, though not all,lines corresponding to flexors in Fig. 3A are above the (dashed)
FIG. 1. Method of threshold detection. A: sample record with 4 completeneurogram bursts. The horizontal lines intersect the signal in the followingregions: background noise, many intersections (a), intersections at start andend of each burst (b), many intersections during bursts (c), few intersectionsclose to peak levels (d). B: The number of intersections (solid line) and itsdifferential (dashed line) plotted as a function of signal level in A. The detectedthreshold (vertical line b) is the level at which the differential returns to 0 aftera biphasic variation.
1059LOCOMOTOR CYCLE DURATIONS
J Neurophysiol • VOL 94 • AUGUST 2005 • www.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from

reference line and have slopes �0.5, whereas most, but againnot all, extensor lines are below the reference line and haveshallower slopes. The reverse is true in Fig. 3B.
Many of the individual regression lines (gray) did notextrapolate to the origin, meaning that phase duration was nota constant proportion of cycle duration. Several had negativeslopes (e.g., Sart in Fig. 3B) meaning that phase durationsdeclined as cycle durations increased. These findings haveimportant implications for the mechanism of central patterngeneration, as discussed later.
Figure 4, A and B, shows the means of all the individualregression lines from Fig. 3, A and B, combined into flexorand extensor groups. The regression slopes in Fig. 4A(flexor-dominated pattern) were 0.75 (flexor) and 0.14 (ex-tensor) and in Fig. 4B (extensor-dominated pattern), 0.27(flexor) and 0.82 (extensor). The flexor and extensor slopesin Fig. 4B (extensor-dominated pattern) are similar to thosepreviously reported for normal cats during overground gait(Halbertsma 1983) (see Fig. 8C), whereas the slopes in Fig.4A suggest a fundamental shift in the balance of rhythmgeneration in these experiments.
Figures 5 and 6 deal with the statistical analysis. Figure 5shows the results of a one-way ANOVA of the slopes of thephase adjustments during flexor and extensor-dominated fictivelocomotion. The mean slope of the bifunctional (knee extensor,hip flexor) RF was not significantly different from those offlexors or extensors. Figure 6 shows a comparison of the meansin the groups of Fig. 3, RF omitted. In flexor-dominated (Fig.6A) and extensor-dominated (Fig. 6B) fictive locomotion, theslopes of the flexor and extensor groups of nerves weresignificantly different.
D I S C U S S I O N
In intact animals and humans, changes in locomotor cadenceare usually due more to variations in stance (extensor phase)durations rather in swing (flexor phase) durations (Arshavskyet al. 1965; Goslow et al. 1973; Halbertsma 1983; Prochazka etal. 2002b). The present results demonstrate that during fictivelocomotion and without rhythmic afferent feedback, the loco-motor step cycle period can vary spontaneously and displayunequal flexion and extension phase durations. In 22 of the 31experiments involving MLR-induced fictive locomotion, weanalyzed, the variations in flexor burst durations were largerthan those of extensors. In the remaining 9 experiments,extensor bursts varied more. The term “dominant” correspondsto the phases showing the larger variation. Figure 4 shows thatfor the largest cycle durations, the dominant phase occupies�80% of the cycle. For cycle durations of 0.35–0.45 s, thephase durations converge to 50% of the cycle and for stilllower cycle durations the relative share reverse. In normal cats,flexor and extensor phase durations converge at gait velocitiesof 1.3–1.8 m/s (Goslow et al. 1973). In reviewing the literature,we found a few anecdotal reports of occasional flexor-domi-nated patterns in experiments involving fictive locomotion inthe cat (Berkinblit et al. 1978; Douglas et al. 1993; Gossard etal. 1994; Grillner and Zangger 1979; Noga et al. 2003).
What is it about MLR-induced fictive locomotion that oftenresults in flexor-dominated patterns? Or to put it the other way,what is it about locomotion in the intact animal that results inextensor-dominated patterns? During normal locomotion, bio-mechanical events elicit varying sensory inputs that modulatethe centrally generated rhythm, predominantly in the stancephase of the cycle. Consider the effect of a small increase inswing duration. This results in a more forward placement of the
FIG. 2. Examples of flexor-dominated fictive locomotor patterns (top) and extensor-dominated patterns (bottom). A: neurograms from a hip flexor nerve (Sart)and a hip extensor nerve (SMAB) during MLR-induced fictive locomotion. Sart bursts have consistently longer durations than SMAB bursts. B: correspondingplots of burst durations vs. cycle durations. Data points above and below - - - correspond to cycles in which phase duration was greater (Sart) or smaller (SMAB)than 50% of cycle duration respectively. C: neurograms from a flexor nerve (Sart) and an extensor nerve (GSol) during MLR-induced fictive locomotion. D:relative share of cycle duration between extensors and flexors is reversed compared with B.
1060 S. YAKOVENKO, D. A. MCCREA, K. STECINA, AND A. PROCHAZAKA
J Neurophysiol • VOL 94 • AUGUST 2005 • www.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from
foot relative to the body. The initial ground reaction force isangled back further toward the hip, decelerating the body andcausing kinetic energy to be lost. The time taken for the bodyto vault over the point of ground support increases, as does theperiod of extensor thrust needed to accelerate the body againand re-extend the hip. Given that the transition to the nextswing phase is held off by sensorimotor mechanisms until theleg is far enough back and unloaded and the contralateral leg ison the ground (Prochazka 1996), it is easy to see how the
stance phase might be more deeply modulated by biomechani-cal factors and corresponding sensory feedback than the swingphase. This argument is further illustrated by the relativeincrease in modulation of flexor phase durations during air-stepping (Smith et al. 1986). Sensory-mediated prolongation ofthe extensor phase could therefore explain why extensor-dominated patterns predominate in normal intact locomotion.
In MLR-induced fictive locomotion, spontaneous cycle pe-riod changes can occur with a constant level of midbrain
FIG. 3. Phase duration plots similar to those in Fig. 2 across all muscles, showing regression lines from individual cats (gray) and the population (black). A:flexor-dominated patterns. Left: flexors; right: extensors. All mean regression lines of flexors (except RF) are above the dashed line (which marks 0.5 cycleduration) while all those of extensors are below. B: similar plots for extensor-dominated patterns.
FIG. 4. Population mean regression lines derived from the plots in Fig. 3 showing the switch in phase dominance from flexors (A) to extensors (B).
1061LOCOMOTOR CYCLE DURATIONS
J Neurophysiol • VOL 94 • AUGUST 2005 • www.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from

stimulation. Presumably these variations result from fluctuat-ing synaptic input from local neuronal networks, fluctuations inintrinsic neuronal properties, or alterations in the excitability ofthe MLR-reticulospinal pathway system. During such varia-tions, phase durations co-vary in an orderly way, as shown bythe linearity of the phase duration plots. We wondered whetherthese properties might provide clues regarding basic oscillatormechanisms.
To explore this, we designed an oscillator model with theuse of Matlab Simulink software. Figure 7 shows the Simulinkmodel, with parameters chosen to model the phase durationplots of Figs. 2B and 8A (flexor-dominated data). The circuit istypical of electronic oscillators and hypothesized biologicaloscillators (Cohen et al. 1982; Marder and Bucher 2001) in thatit has two mutually inhibitory “half-centers” the duty cycles ofwhich are determined by timing elements, in this case a pair ofresettable integrators the outputs of which trigger bistable“flip-flops” to change state from low to high when they receivean input equal to 1.
Phase durations can be set to the longest and shortest phasesin a given set of experimental data by setting the offset andgain parameters to appropriate values. Consider offset A(0.864) and offset B (2.877) in Fig. 7. Integrator B outputs aramp that reaches the trigger value 1 after 1/0.864 (i.e., 1.2 s,the longest flexor phase duration in Fig. 8A). This triggersflip-flop B to go from 0 to 1, resetting integrator A, which inturn triggers flip-flop A after 1/2.877 � 0.35s, the longestextensor phase duration in Fig. 8A. Integrator B is reset and thenext cycle commences. In addition to the offsets, a very slowramp input is supplied to each of the integrators through gainsA and B. This gradually speeds up the integrators, decreasingphase durations. Thus at 100 s, the input to integrator A is2.459 � 0.864 � 3.32, resulting in a phase duration of1/3.32 � 0.30 s, the shortest flexor phase in Fig. 8A. Similarlythe input to integrator B is 1.476 � 2.877 � 4.35, resulting ina phase duration of 1/4.35 � 0.23 s, the shortest extensor phasein Fig. 8A.
FIG. 5. The results of 1-way ANOVAs of the slopes of phase duration plots in flexor-dominated (A) and extensor-dominated (B) fictive locomotion. Thewhiskers show the 95% confidence intervals about the medians.
FIG. 6. Results of the 1-way ANOVA and multiple comparisons of slopes of phase duration plots using Tukey’s honestly significant difference criterion inflexor(A) and extensor (B) dominated fictive locomotion. Whiskers show confidence intervals about the median.
1062 S. YAKOVENKO, D. A. MCCREA, K. STECINA, AND A. PROCHAZAKA
J Neurophysiol • VOL 94 • AUGUST 2005 • www.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from

Note that these oscillators produce a constant amplitudeoutput that represents the basic alternating flexion-extensionpattern. The model could be made more realistic by addingelements that distribute oscillator output to motoneurons andadjust their activity levels (Rybak et al. 2004).
Three sets of data were fitted: Fig. 8, A (flexor-dominatedMLR fictive), B (extensor dominated MLR fictive), and C [datafrom normal cats (Halbertsma 1983)]. We were surprised athow closely the modeled phase duration plots matched those ofthe raw data (in the case of the Halbertsma et al. data, rightdown to the slightly curved relationships which fitted the rawdata better than the linear regressions used by the authors).This suggests that the basic CPG oscillator behaves like recip-rocally coupled integrators. What other inferences can bedrawn? First, it is clear that to fit the various slopes and offsetsof the phase duration curves, separately adjustable timingelements for flexion and extension are required. This agreeswith the conclusion of a previous study on fictive scratchingand locomotion (Berkinblit et al. 1978). Second, becausechanges in flexor and extensor phase duration nearly alwaysoccur in parallel, it seems likely that there is common orparallel input to the timing elements. Third, because phasedurations co-vary by different amounts, either the commoninput is unequal, the sensitivity to the common input is un-equal, or both. Fourth, additional nonvarying input (“offset”) isrequired to match the mean phase durations in the data. Fifth,
the goodness of fit of the modeled data from normal cats (Fig.8C) indicates that the inclusion of rhythmic sensory input doesnot change the basic way in which phase durations within thestep cycle period are accomplished. For example, the phaseprolongation produced by extensor group I afferent input couldbe accomplished by adding an additional offset to integrator Ain Fig. 8.
Suppose that “offset” in the model corresponds to back-ground excitation of the neuronal networks of the CPG thatmake up the timing elements. This excitation would includesynaptic input such as that elicited by MLR stimulation as wellas other sources and could be preparation dependent. Theneuronal analogy of the “gain” term in the simple oscillator isthe range of variation of recruitment and spiking of timing-generating neuronal populations. Because both recruitment andfiring increases are saturable phenomena, the half-center withthe lower background excitation would have the higher gain.Thus variations in background drive would result in largervariations in phase duration in this half-center (i.e., this wouldbe the dominant one). Variability in the balance of backgroundexcitation between preparations could help explain why someof our cats were flexor-dominant and others were extensor-dominant. Note that anything that changed the excitability ofsome of the members of an oscillator could change the phasedominance. This could include random or preparation-specificchanges in either excitation or inhibition of any of the elements
FIG. 7. Oscillator model with 2 mutually in-hibitory “half-centers” the duty cycles of whichare determined by the resettable integrators. Themodulating input on the left is supplied to bothintegrators through separate gain stages with indi-vidual offsets. The inputs to the integrators deter-mine how quickly their outputs reach an arbitrarytrigger level and thereby reset the output of theother integrator (details in text). The model wasdeveloped in Matlab Simulink 6.5. The modifiedRunge-Kutta (odetb23tb) variable-step integrationmethod was used for simulations.
FIG. 8. Burst duration plots from Fig. 2 (A and B) and phase duration plots from normal cats [C: Halbertsma (1983)]. Filled circles: flexor burst or swing-phasedurations, open circles: extensor burst or stance-phase durations. Triangles show the trajectories of phase durations obtained from the model in Fig. 7, havingadjusted the gain and offset parameters to fit the start and endpoints of the regression lines (solid) fitted to the data points. Note the remarkable correspondenceof the actual and modeled phase duration plots.
1063LOCOMOTOR CYCLE DURATIONS
J Neurophysiol • VOL 94 • AUGUST 2005 • www.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from

affected by a variety of factors such as head, trunk or legposture (Pearson et al. 1992), neurotransmitter balance (Jacobsand Fornal 1999), the balance of descending drives (Noga et al.2003) and overall state.
In this regard, it is interesting to note that Pearson andRossignol showed that limb position can influence the intensityand relative durations of the flexor and extensor bursts ofactivity in fictive locomotion in chronic spinal cats. Moving thelegs from flexion to extension stretched flexor muscles andprogressively increased the duration of flexor bursts and de-creased the cycle period, though not to the extent of switchingfrom extensor- to flexor-dominated patterns, at least in the datashown (Pearson and Rossignol 1991).
Could the full range of phase duration plots presented abovebe obtained with a model that only had gain elements anddispensed with offsets? The answer is no. If gains were theonly contributing parameters, the curves would all passthrough the origin because each half-center’s share of the cycleduration would be constant. Many of the regression lines wepresented in this paper did not pass through the origin.
Several individual regression lines had negative slopes,meaning that phase durations declined as cycle durations in-creased. In the model, such lines can readily be matched simplyby setting the gain of the relevant integrator to an appropriatenegative value (the offset remaining positive). The neuralcorrelate of this would be would be a declining excitation or anincreasing inhibition of the drive to the relevant timing neu-rons.
One of the implications of the present findings is that whilemost preparations showed a particular bias for the flexion orextension phase during fictive locomotion, the structure of thelocomotor CPG does not appear to be inherently extensor orflexor biased. In other words, the flexor or extensor sides of thelocomotor oscillator can operate in similar fashion (compareFig. 8, A and B). This in turn supports models of CPG structurein which either the flexor and extensor phases can dominate(see Rybak et al. 2004).
To conclude, in this paper, we report a considerable flexi-bility in the control of phase durations in MLR-induced fictivelocomotion, which can be explained by changes in backgroundexcitation and sensitivity of timing elements in a simple half-center model of the locomotor CPG.
A C K N O W L E D G M E N T S
Present address of S. Yakovenko: Dept. of Physiology, University ofMontreal, CP 6128 Succ. A, Montreal, PQ H3C 3J7, Canada.
G R A N T S
This work was supported by grants from the Canadian Institutes of HealthResearch and the Alberta Heritage Foundation for Medical Research.
R E F E R E N C E S
Armstrong DM. The supraspinal control of mammalian locomotion. J Physiol405: 1–37, 1988.
Arshavsky YI, Kots YM, Orlovsky IM, Rodionov IM, and Shik ML.Investigation of the biomechanics of running by the dog (translation of RussBiofizika). Biophysics 10: 737–746, 1965.
Berkinblit MB, Deliagina TG, Feldman AG, Gelfand IM, and OrlovskyGN. Generation of scratching. II. Nonregular regimes of generation. J Neu-rophysiol 41: 1058–1069, 1978.
Bouyer LJ and Rossignol S. The contribution of cutaneous inputs to loco-motion in the intact and the spinal cat. Ann NY Acad Sci 860: 508–512,1998.
Brown TG. The intrinsic factors in the act of progression in the mammal. ProcRoy Soc Lond B Biol Sci 84: 308–319, 1911.
Burke RE. The central pattern generator for locomotion in mammals. AdvNeurol 87: 11–24, 2001.
Cohen AH, Holmes PJ, and Rand RH. The nature of the coupling betweensegmental oscillators of the lamprey spinal generator for locomotion: amathematical model. J Math Biol 13: 345–369, 1982.
Cowley KC and Schmidt BJ. Regional distribution of the locomotor pattern-generating network in the neonatal rat spinal cord. J Neurophysiol 77:247–259, 1997.
Douglas JR, Noga BR, Dai X, and Jordan LM. The effects of intrathecaladministration of excitatory amino acid agonists and antagonists on theinitiation of locomotion in the adult cat. J Neurosci 13: 990–1000, 1993.
Drew T. The role of the motor cortex in the control of gait modification in thecat. In: Neurobiological Basis of Human Locomotion, edited by ShimamuraM, Grillner S, and Edgerton VR. Tokyo, Japan: Japan Scientific SocietiesPress, 1991, p. 201–212.
Dubuc R, Cabelguen JM, and Rossignol S. Rhythmic fluctuations of dorsalroot potentials and antidromic discharges of primary afferents during fictivelocomotion in the cat. J Neurophysiol 60: 2014–2036, 1988.
Eccles RM and Lundberg A. Integrative pattern of Ia synaptic actions onmotoneurons of hip and knee muscles. J Physiol 144: 271–298, 1958.
Forssberg H and Grillner S. The locomotion of the acute spinal cat injectedwith clonidine i.v. Brain Res 50: 184–186, 1973.
Gosgnach S, Quevedo J, Fedirchuk B, and McCrea DA. Depression ofgroup Ia monosynaptic EPSPs in cat hindlimb motoneurones during fictivelocomotion. J Physiol 526: 639–652, 2000.
Goslow GE, Jr, Reinking RM, and Stuart DG. The cat step cycle: hind limbjoint angles and muscle lengths during unrestrained locomotion. J Morpholy141: 1–41, 1973.
Gossard JP, Brownstone RM, Barajon I, and Hultborn H. Transmission ina locomotor-related group Ib pathway from hindlimb extensor muscles inthe cat. Exp Brain Res 98: 213–228, 1994.
Grillner S. Control of locomotion in bipeds, tetrapods, and fish. In: Handbookof Physiology. The Nervous System. Motor Control. Bethesda, MD: Am.Physiol. Soc., 1981, sect. 1, vol. I, p. 1179–1236.
Grillner S and Dubuc R. Control of locomotion in vertebrates: spinal andsupraspinal mechanisms. Adv Neurol 47: 425–453, 1988.
Grillner S and Wallen P. Central pattern generators for locomotion, withspecial reference to vertebrates. Annu Rev Neurosci 8: 233–261, 1985.
Grillner S and Zangger P. How detailed is the central pattern generation forlocomotion? Brain Res 88: 367–371, 1975.
Grillner S and Zangger P. On the central generation of locomotion in the lowspinal cat. Exp Brain Res 34: 241–261, 1979.
Halbertsma JM. The stride cycle of the cat: the modelling of locomotion bycomputerized analysis of automatic recordings. Acta Physiol Scand Suppl521: 1–75, 1983.
Jacobs BL and Fornal CA. Activity of serotonergic neurons in behavinganimals. Neuropsychopharmacology 21: 9S–15S, 1999.
Jordan LM. Initiation of locomotion in mammals. Ann NY Acad Sci 860:83–93, 1998.
Kiehn O and Kjaerulff O. Spatiotemporal characteristics of 5-HT anddopamine-induced rhythmic hindlimb activity in the in vitro neonatal rat.J Neurophysiol 75: 1472–1482, 1996.
Leblond H and Gossard JP. Supraspinal and segmental signals can betransmitted through separate spinal cord pathways to enhance locomotoractivity in extensor muscles in the cat. Exp Brain Res 114: 188–192, 1997.
Lundberg A. Half-centres revisited. In: Advances in Physiological Sciences,edited by Szenthagothai J, Palkovits M, and Hamori J. New York: Pergamonand Akademiai Kiado, 1981, p. 155–167.
Marder E and Bucher D. Central pattern generators and the control ofrhythmic movements. Curr Biol 11: R986–996, 2001.
McCrea DA. Spinal circuitry of sensorimotor control of locomotion. J Physiol533: 41–50, 2001.
Murray MP. Gait as a total pattern of movement. Am J Phys Med 46:290–333, 1967.
Noga BR, Kriellaars DJ, Brownstone RM, and Jordan LM. Mechanism foractivation of locomotor centers in the spinal cord by stimulation of themesencephalic locomotor region. J Neurophysiol 90: 1464–1478, 2003.
Pearson KG. Generating the walking gait: role of sensory feedback. ProgBrain Res 143: 123–129, 2004.
Pearson KG, Jiang W, and Ramirez JM. The use of naloxone to facilitatethe generation of the locomotor rhythm in spinal cats. J Neurosci Methods42: 75–81, 1992.
1064 S. YAKOVENKO, D. A. MCCREA, K. STECINA, AND A. PROCHAZAKA
J Neurophysiol • VOL 94 • AUGUST 2005 • www.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from

Pearson KG and Rossignol S. Fictive motor patterns in chronic spinal cats.J Neurophysiol 66: 1874–1887, 1991.
Prochazka A. Proprioceptive feedback and movement regulation. In: Exer-cise: Regulation and Integration of Multiple Systems, edited by Rowell Land Sheperd JT. New York: Am. Physiol. Soc., 1996, sect. 12, p. 89–127.
Prochazka A, Gritsenko V, and Yakovenko S. Sensory control of locomo-tion: reflexes versus higher-level control. Adv Exp Med Biol 508: 357–367,2002a.
Prochazka A, Mushahwar V, and Yakovenko S. Activation and coordina-tion of spinal motoneuron pools after spinal cord injury. Prog Brain Res137: 109–124, 2002b.
Quevedo J, Fedirchuk B, Gosgnach S, and McCrea DA. Group I disynapticexcitation of cat hindlimb flexor and bifunctional motoneurons during fictivelocomotion. J Physiol 525: 549–564, 2000a.
Quevedo J, Stecina K, Gosgnach S, and McCrea DA. Activity of hip flexorsprecedes that of ankle flexors during fictive locomotion and scratching. SocNeurosci Abstr 747.746, 2000b.
Rossignol S. Neural control of stereotypic limb movements. In: Handbook ofPhysiology.Exercise: Regulation and Integration of Multiple Systems, editedby Rowell L and Sheperd JT. New York: Am. Physiol. Soc., 1996, sect. 12,p. 173–216.
Rossignol S, Chau C, Brustein E, Giroux N, Bouyer L, Barbeau H, andReader TA. Pharmacological activation and modulation of the central
pattern generator for locomotion in the cat. Ann NY Acad Sci 860: 346–359,1998.
Rybak IA, Lafreniere-Roula M, N.A. S, and McCrea DA. A two layermodel of the spinal locomotor CPG: insights from deletions of activityduring fictive locomotion. Soc Neurosci Abstr 883.883, 2004.
Sherrington CS. Flexion-reflex of the limb, crossed extension-reflex, andreflex stepping and standing. J Physiol 40: 28–121, 1910.
Smith JL, Bradley NS, Carter MC, Giuliani CA, Hoy MG, Koshland GF,and Zernicke RF. Rhythmical movements of the hindlimbs in spinal cat:considerations for a controlling network. In: Development and Plasticity ofthe Mammalian Spinal Cord, edited by Goldberger ME, Gorio A, andMurray M. Padova, Italy: Liviana, 1986, p. 347–361.
Stein PSG and Smith JL. Neural and biomechanical control strategies fordifferent forms of vertebrate hindlimb motor tasks. In: Neurons, Networks,and Motor Behavior, edited by Stein PSG, Grillner S, Selverston AI andStuart DG. Cambridge, MA: MIT Press, 1997, p. 61–73.
Yakovenko S, Gritsenko V, and Prochazka A. Contribution of stretchreflexes to locomotor control: a modeling study. Biol Cybern 90: 146–155,2004.
Yakovenko S, McCrea DA, Stecina K, and Prochazka A. Phase and cycledurations in MLR-induced fictive cat locomotion. Soc Neurosci Abstr625.625, 2001.
1065LOCOMOTOR CYCLE DURATIONS
J Neurophysiol • VOL 94 • AUGUST 2005 • www.jn.org
on October 29, 2005
jn.physiology.orgD
ownloaded from
Copyright © 2022 FDOKUMEN




















