
SENSITIZATION OF STUDENTS ON THE NEED FOR HEALTHY ENVIRONMENT FOR SUSTAINABLE DEVELOPMENT.

G
B
R
Is
Ha
b
c
a
ARRAA
KPNLILeR
1
ofhvs
otuatPn
0d
ARTICLE IN PRESSModel
BR-6648; No. of Pages 6
Behavioural Brain Research xxx (2010) xxx–xxx
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
esearch report
mpact of baseline prepulse inhibition on nicotine-induced locomotorensitization in rats
akan Kayira,∗, Gokhan Goktalayb, Oguzhan Yavuzc, Tayfun I. Uzbaya
Department of Medical Pharmacology, Psychopharmacology Research Unit, Gulhane Military Medical Academy, Ankara, TurkeyDepartment of Medical Pharmacology, Uludag University, Faculty of Medicine, Bursa, TurkeyDepartment of Pharmacology and Toxicology, Ondokuz Mayis University, Faculty of Veterinary, Samsun, Turkey
r t i c l e i n f o
rticle history:eceived 18 June 2010eceived in revised form 30 July 2010ccepted 5 August 2010vailable online xxx
eywords:repulse inhibitionicotineocomotor sensitizationndividual vulnerabilityocomotor reactivity to a novelnvironment
a b s t r a c t
The rats having high locomotor reactivity to a novel environment (LRNE) are known to be more vulner-able to develop locomotor sensitization, which reflects the initial neuroplastic changes in brain systemsrelated to addictive behaviours. The present study aimed to investigate whether sensorimotor gatinglevel, measured by prepulse inhibition (PPI) of acoustic startle reflex, also reflects vulnerability for nico-tine sensitization. A batch of rats was assigned into three groups according to their baseline PPI values.The highest 1/3 and the lowest 1/3 proportions were selected and defined as high-inhibitory (HI) and low-inhibitory (LI) groups. LRNE was measured in the rats, then they were treated with nicotine (1 mg/kg,tartrate salt, subcutaneously) or saline and locomotor activity (LMA) was immediately recorded for15 min. This procedure was performed daily for 5 successive days. After a 3-day drug-free period, allrats were challenged with nicotine (1 mg/kg) on 9th day and with saline on 12th day. Same sensitizationprotocol was applied in another batch of rats, except assigning them into the high-responder (HR) and
at(s)low-responder (LR) groups according to LRNE levels. There was no significant difference between HI and LIrats in LRNE. Although the acute effect of nicotine on LMA was higher in HI rats, a locomotor sensitizationdeveloped and expressed only in LI rats. In the following experiments, nicotine stimulated LMA both inHR and LR rats, but induced and expressed locomotor sensitization only in HR rats. The present studyshows that acute locomotor stimulant effect and locomotor sensitization developing effects of nicotineare associated with the baseline PPI and LRNE levels. But these two factors are independent from each
other.. Introduction
Many people smoke at least once in their lifetime, but only somef them keep on smoking and get addicted. In general, transitionrom drug use to abuse and development of addiction in humans areighly dependent on individual characteristics [9]. Identifying theulnerable individuals and personality types is important to under-tand basic mechanisms of addiction and preventive approaches.
One of the individual vulnerability factors for addiction devel-pment to nicotine, which is the major addictive ingredient ofobacco, is sensation seeking [7]. Smoking rate is higher in a pop-lation with high sensation seeking scores [37]. In experimental
Please cite this article in press as: Kayir H, et al. Impact of baseline prepulseBrain Res (2010), doi:10.1016/j.bbr.2010.08.004
nimals, sensation seeking can be evaluated by measuring locomo-or reactivity of the animals, exposed to a novel environment [3].iazza et al. [22] first indicated that high-responder (HR) rats in aovel environment were more vulnerable to develop amphetamine
∗ Corresponding author. Tel.: +90 312 304 4768; fax: +90 312 304 6091.E-mail address: [email protected] (H. Kayir).
166-4328/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2010.08.004
© 2010 Elsevier B.V. All rights reserved.
self-administration. Later, Suto et al. [30] reported that locomotorresponse to novelty was positively correlated with acquisition ofnicotine self-administration and number of infusions at progressiveratio in rats.
HR rats have elevated levels of dopamine (DA) in the nucleusaccumbens (NAcc) and matching lower levels of DA in the pre-frontal cortex compared to low-responder (LR) rats [23]. Moreover,HR rats, relative to their LR counterparts, show elevated levels of DAwhen responding to novelty [17]. Cellular and molecular changes inventral tegmental area (VTA)–NAcc system, together with the otherlimbic regions, are accepted to play a critical role in the develop-ment of addictive behaviour [21].
Mesocorticolimbic dopaminergic system also plays a role infunctioning of sensorimotor gating system, which is critical for theintegration of sensory and cognitive information processing and
inhibition on nicotine-induced locomotor sensitization in rats. Behav
execution of appropriate motor responses [18]. The performanceof this system, both in animals and humans, can be evaluated byusing a physiological phenomenon called prepulse inhibition (PPI)of startle reflex, which is referred to the attenuation of the startleresponse caused by a strong stimulus when it is abruptly preceded

ING
B
2 rain R
bd[[as
sepsitn
2
2
rIaWiqttfA
Mw
2
o
+mate1rdnfodfm
Lm
Laio2hs
2
RdgaAntgr
iiii
ARTICLEModel
BR-6648; No. of Pages 6
H. Kayir et al. / Behavioural B
y a weaker, non-startling stimulus [15]. Disruption of PPI in someisorders, such as schizophrenia, has been extensively investigated4,31] and used as a trusted research tool in experimental animals12,36]. On the other hand, baseline PPI levels may be considereds a biological marker and an endophenotype for schizophreniapectrum disorders [5,14].
Involvement of the similar brain areas in nicotine addiction andensorimotor gating deficits reminded us that individual differ-nces in baseline PPI levels might have an impact on addictiveroperties of nicotine. Thus, in the following experiments, we clas-ified rats according to their baseline PPI levels, and tested themn a nicotine-induced locomotor sensitization model. Also the rela-ionship between baseline PPI levels and locomotor reactivity to aovel environment (LRNE) was investigated.
. Materials and methods
.1. Animals, laboratory and drug
All procedures in the present study were performed in accordance with theules in the Guide for the Care and Use of Laboratory Animals adopted by Nationalnstitutes of Health (USA) and the Declaration of Helsinki. Local ethical committeepproval was also obtained (Date: October 13th, 2005; No.: 2005-73). Adult maleistar rats, weighing 260–320 g, and 12–16 weeks old at the beginning of exper-
ments were subjects. Four rats were housed in each cage. They were placed in auiet, temperature and humidity controlled room (22 ± 2 ◦C and 60 ± 5%, respec-ively) in which a 12 h:12 h light–dark cycle was maintained (lights on from 07:00o 19:00 h). Food and water were available ad libitum. All the experiments were per-ormed during the light phase of the light–dark cycle (between 08:30 and 11:30 h).ll the rats were handled for 2 days before the experiments start.
(−)-Nicotine hydrogen tartrate was purchased from Sigma Chemical (St. Louise,O, USA) and dissolved in saline at a concentration of 1 mg/ml (tartrate salt) andas injected subcutaneously (s.c.). Nicotine was prepared freshly every morning.
.2. Experimental design
The first experiment was designed to investigate the effects of baseline PPI levelsn LRNE and nicotine-induced locomotor sensitization.
A batch of 48 rats was evaluated for their baseline PPI levels at background16 dB prepulse and were ranked in descending order according to this baselineeasurement. Rats in the highest 1/3 and the lowest 1/3 proportions were selected
s high-inhibitory (HI) and low-inhibitory (LI), respectively (n = 16). Then, each ofhe HI and LI groups was assigned into two groups (n = 8), by attending the rats toither of the two groups in alternate order. After 1 week, LRNE was measured for5 min in all four groups. Twenty-four hours later, the rats were habituated to theegular locomotor activity (LMA) cages for 15 min. Beginning with the next day (1stay of locomotor sensitization protocol), one pair of the HI and LI group receivedicotine (1 mg/kg), and the other pair of HI and LI group received saline. Immediately
ollowing injections, LMA was measured for 15 min. This procedure was performedn daily basis for 5 sessions until a locomotor sensitization developed. After a 3-ay drug-free period, on 9th day, nicotine challenge injections were applied to allour groups and LMA was measured for 15 min. Saline challenge injections and LMA
easurements were applied to all groups on 11th day.The second experiment was designed to investigate the relationship between
RNE and baseline PPI levels. Later we also confirmed that our LRNE groupingethod was efficient in predicting nicotine sensitization.
A batch of 48 rats was ranked in a descending order according to their baselineRNE values. Rats in the highest 1/3 and the lowest 1/3 proportions were selecteds HR and LR, respectively (n = 16). Then, each of the HR and LR groups was assignednto two groups (n = 8), by attending the rats to either of the two groups in alternaterder. After 1 week, rats habituated to the acoustic startle cages for 15 min and4 h later baseline PPI was measured in all four groups. The next day, the rats wereabituated to the regular LMA cages for 15 min starting with the next day locomotorensitization protocol was applied as mentioned above.
.3. Prepulse inhibition of acoustic startle reflex test
PPI of acoustic startle reflex test was performed using Animal Acoustic Startleesponse System (AASR, Habitest Model E10-21, Coulbourn, PA, USA) as previouslyescribed [34]. This system consists of four sound proof boxes equipped with strainage load cell platforms, and speakers with a wide sound spectrum, mounted 12 cm
Please cite this article in press as: Kayir H, et al. Impact of baseline prepulseBrain Res (2010), doi:10.1016/j.bbr.2010.08.004
bove the animal holders. The system was controlled by a software (Coulbourncoustic Startle v4.530-00) run on an IBM-compatible computer. The backgroundoise levels were between 63 and 66 dB [A] sound pressure level (SPL) duringhe study. Prepulse stimulus levels, which are determined according to the back-round noise, were adjusted by monthly calibrations. The software recorded startleesponse of rats for 200 ms with a 1 ms resolution after the administration of pulse
PRESSesearch xxx (2010) xxx–xxx
stimuli with or without a prepulse stimulus, and expressed the peak startle responseas grams.
PPI measurements were performed between 09:00 and 11:00 h. Twenty-fourhours before the test sessions, rats were habituated to the animal holders of startletest system for 15 min. During this period, four pulse stimuli (110 dB) were appliedwith a random interval between 10 and 30 s. Rats having a startle reflex less than20 g in average were excluded from the study and replaced with other rats meetingthis criterion (this situation occurred only twice throughout the study). On the testday, rats were placed in the cages, and the test protocol was applied. These sessionspreceded by a 5-min acclimatization period, during which only background noisewas present. Then, eight blocks of five trials were administered. Following trailswere applied in random order within one block:
i. Pulse alone stimulus.i. Prepulse (4 dB [A] SPL above background noise) + pulse stimulus.i. Prepulse (8 dB [A] SPL above background noise) + pulse stimulus.v. Prepulse (16 dB [A] SPL above background noise) + pulse stimulus.v. No stimulus (only background noise was applied).
Pulse stimulus was a broadband noise stimulus at 110 dB [A] SPL for 40 ms,whereas all three prepulse stimuli were tone stimuli at 3 kHz frequency for 20 ms.Prepulse stimulus levels were selected at intensities which did not elicit significantstartle reflex when applied alone. Prepulse stimulus was applied 120 ms prior topulse stimulus (onset to onset). The inter-trial interval varied randomly in a range of10–30 s. This protocol approximately lasted for 20 min. PPI was defined as a decreasein the amplitude of startle reflex in the presence of the prepulse stimulus, and cal-culated for each of the three different prepulse intensities by using the followingformula:
PPI (%) = 100 − average startle reflex in presence of the prepulse × 100average startle reflex without a prepulse
2.4. Locomotor reactivity to a novel environment
LRNE was measured as distance travelled (cm) in black Plexiglas arenas(80 cm × 80 cm × 40 cm) during 15 min. Four cages were monitored at the sametime with a CCD camera (Safer, SF-2222) mounted 260 cm above from the floorand vertically aimed at the crossing of four cages. A video tracking software (Ethovi-sion v3.1.16, Noldus Information Technology, Wageningen, NL) was used to analyzethe visual data. Rats were detected due to their contrast according to the blackbackground. The parameter of distance travelled (cm) was computed with down-sampling filters (final frame-rate was 6 fps and minimum distance travelled was1 cm) to decrease the image noise.
2.5. Locomotor activity for sensitization development
Locomotor activity was measured with an open-field activity monitoring system(MAY 9908 model Activity Monitoring System: Commat Ltd., Turkey). This systemhad eight Plexiglas cages (42 cm × 42 cm × 30 cm) equipped with infrared photo-cells. Fifteen photocell emitter and detector pairs were located 4.5 cm above the floorat intervals of 2.5 cm on the both counter sides of each activity cage, and another15 photocell pairs were located 11.5 cm above the floor. Interruptions of photocellbeams were detected by an IBM-compatible computer system, and the location ofthe animal was calculated by the software at 0.1 s sensitivity. If the calculated loca-tions were completely changed, this was expressed as ambulatory activity. Otherbehavioural responses that caused interruptions of beams but not changes in loca-tion were presented as horizontal activity. Vertical activity, such as rearing, wasdetected by the photocells located 11.5 cm above the floor. Locomotor activitieswere calculated as the sum of ambulatory, horizontal and vertical activities.
2.6. Statistics
Difference between baseline PPI of HI and LI or HR and LR groups was shownby using a mixed design two-way analysis of variance (ANOVA) test (high or lowgroup as between subjects × prepulse intensity as within subjects factor) followedby Student’s t test. The acute effect of nicotine on locomotor activity in HI and LI or HRand LR groups (the 1st sessions of the sensitization protocols) was evaluated using atwo-way ANOVA test (treatment × high or low groups) followed by Tukey’s test. Thelocomotor activity data of following sessions were expressed as percent changes in
inhibition on nicotine-induced locomotor sensitization in rats. Behav
comparison to the first day values of each rat. Development of nicotine sensitizationwas evaluated by using a mixed design three-way ANOVA (treatment × high or lowgroups × sessions) followed by Bonferroni test for post hoc comparisons. Data ofnicotine or saline challenge injections were evaluated by two-way ANOVA tests(high or low groups × chronic treatment, both as between subjects factors) followedby Student’s t test. The levels of statistical significance were set at p < 0.05.
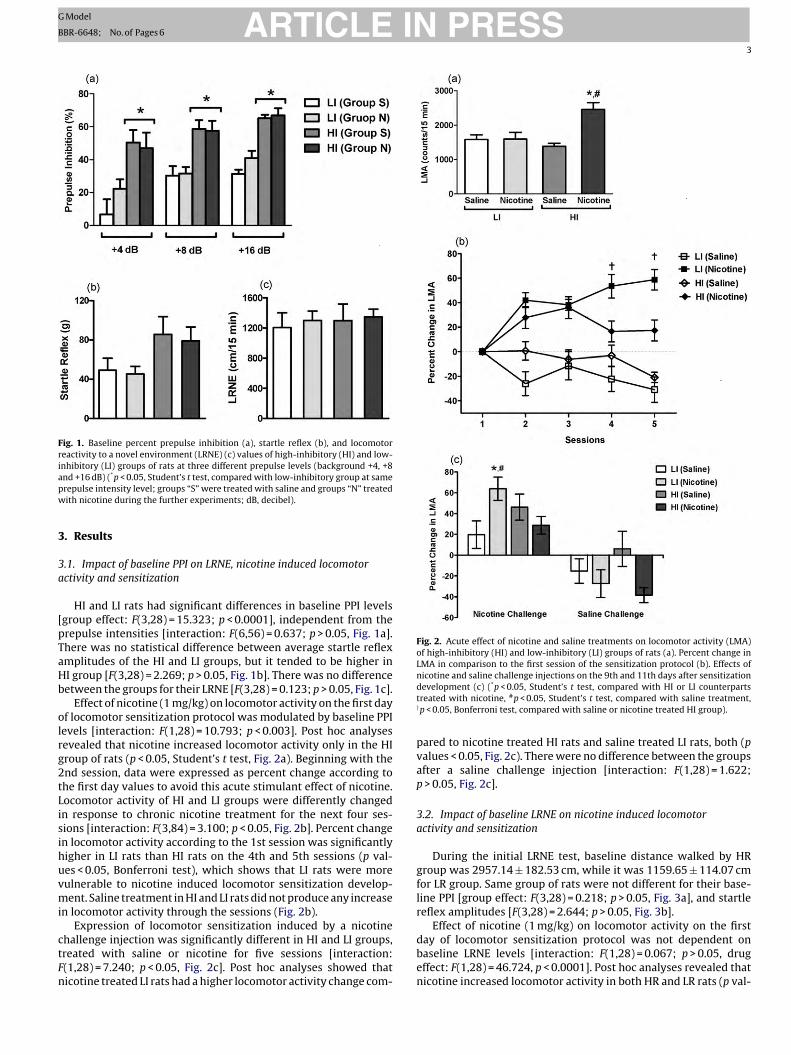
ARTICLE IN PRESSG Model
BBR-6648; No. of Pages 6
H. Kayir et al. / Behavioural Brain Research xxx (2010) xxx–xxx 3
Fig. 1. Baseline percent prepulse inhibition (a), startle reflex (b), and locomotorreactivity to a novel environment (LRNE) (c) values of high-inhibitory (HI) and low-inhibitory (LI) groups of rats at three different prepulse levels (background +4, +8apw
3
3a
[pTaHb
olrg2tLisihuvmi
ctFn
Fig. 2. Acute effect of nicotine and saline treatments on locomotor activity (LMA)of high-inhibitory (HI) and low-inhibitory (LI) groups of rats (a). Percent change inLMA in comparison to the first session of the sensitization protocol (b). Effects of
Effect of nicotine (1 mg/kg) on locomotor activity on the first
nd +16 dB) (*p < 0.05, Student’s t test, compared with low-inhibitory group at samerepulse intensity level; groups “S” were treated with saline and groups “N” treatedith nicotine during the further experiments; dB, decibel).
. Results
.1. Impact of baseline PPI on LRNE, nicotine induced locomotorctivity and sensitization
HI and LI rats had significant differences in baseline PPI levelsgroup effect: F(3,28) = 15.323; p < 0.0001], independent from therepulse intensities [interaction: F(6,56) = 0.637; p > 0.05, Fig. 1a].here was no statistical difference between average startle reflexmplitudes of the HI and LI groups, but it tended to be higher inI group [F(3,28) = 2.269; p > 0.05, Fig. 1b]. There was no differenceetween the groups for their LRNE [F(3,28) = 0.123; p > 0.05, Fig. 1c].
Effect of nicotine (1 mg/kg) on locomotor activity on the first dayf locomotor sensitization protocol was modulated by baseline PPIevels [interaction: F(1,28) = 10.793; p < 0.003]. Post hoc analysesevealed that nicotine increased locomotor activity only in the HIroup of rats (p < 0.05, Student’s t test, Fig. 2a). Beginning with thend session, data were expressed as percent change according tohe first day values to avoid this acute stimulant effect of nicotine.ocomotor activity of HI and LI groups were differently changedn response to chronic nicotine treatment for the next four ses-ions [interaction: F(3,84) = 3.100; p < 0.05, Fig. 2b]. Percent changen locomotor activity according to the 1st session was significantlyigher in LI rats than HI rats on the 4th and 5th sessions (p val-es < 0.05, Bonferroni test), which shows that LI rats were moreulnerable to nicotine induced locomotor sensitization develop-ent. Saline treatment in HI and LI rats did not produce any increase
n locomotor activity through the sessions (Fig. 2b).Expression of locomotor sensitization induced by a nicotine
Please cite this article in press as: Kayir H, et al. Impact of baseline prepulseBrain Res (2010), doi:10.1016/j.bbr.2010.08.004
hallenge injection was significantly different in HI and LI groups,reated with saline or nicotine for five sessions [interaction:(1,28) = 7.240; p < 0.05, Fig. 2c]. Post hoc analyses showed thaticotine treated LI rats had a higher locomotor activity change com-
nicotine and saline challenge injections on the 9th and 11th days after sensitizationdevelopment (c) (*p < 0.05, Student’s t test, compared with HI or LI counterpartstreated with nicotine, #p < 0.05, Student’s t test, compared with saline treatment,†p < 0.05, Bonferroni test, compared with saline or nicotine treated HI group).
pared to nicotine treated HI rats and saline treated LI rats, both (pvalues < 0.05, Fig. 2c). There were no difference between the groupsafter a saline challenge injection [interaction: F(1,28) = 1.622;p > 0.05, Fig. 2c].
3.2. Impact of baseline LRNE on nicotine induced locomotoractivity and sensitization
During the initial LRNE test, baseline distance walked by HRgroup was 2957.14 ± 182.53 cm, while it was 1159.65 ± 114.07 cmfor LR group. Same group of rats were not different for their base-line PPI [group effect: F(3,28) = 0.218; p > 0.05, Fig. 3a], and startlereflex amplitudes [F(3,28) = 2.644; p > 0.05, Fig. 3b].
inhibition on nicotine-induced locomotor sensitization in rats. Behav
day of locomotor sensitization protocol was not dependent onbaseline LRNE levels [interaction: F(1,28) = 0.067; p > 0.05, drugeffect: F(1,28) = 46.724, p < 0.0001]. Post hoc analyses revealed thatnicotine increased locomotor activity in both HR and LR rats (p val-
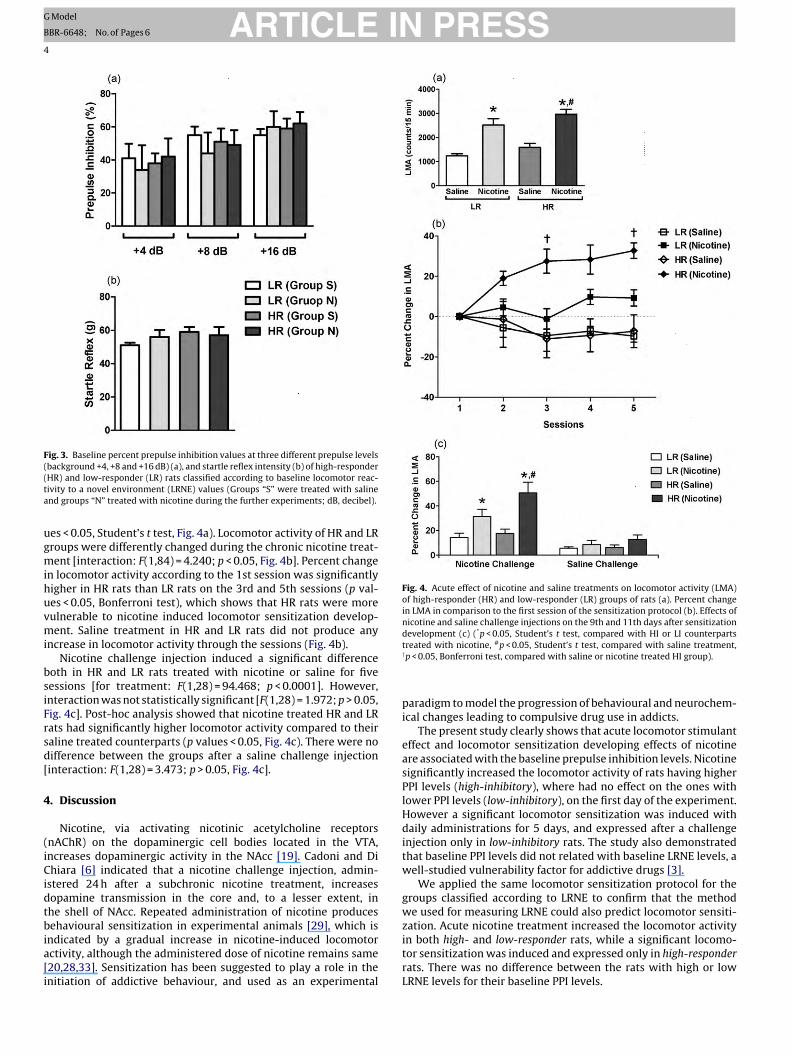
ARTICLE IN PRESSG Model
BBR-6648; No. of Pages 6
4 H. Kayir et al. / Behavioural Brain Research xxx (2010) xxx–xxx
Fig. 3. Baseline percent prepulse inhibition values at three different prepulse levels((ta
ugmihuvmi
bsiFrsd[
4
(iCidtbia[i
Fig. 4. Acute effect of nicotine and saline treatments on locomotor activity (LMA)of high-responder (HR) and low-responder (LR) groups of rats (a). Percent changein LMA in comparison to the first session of the sensitization protocol (b). Effects ofnicotine and saline challenge injections on the 9th and 11th days after sensitization
*
background +4, +8 and +16 dB) (a), and startle reflex intensity (b) of high-responderHR) and low-responder (LR) rats classified according to baseline locomotor reac-ivity to a novel environment (LRNE) values (Groups “S” were treated with salinend groups “N” treated with nicotine during the further experiments; dB, decibel).
es < 0.05, Student’s t test, Fig. 4a). Locomotor activity of HR and LRroups were differently changed during the chronic nicotine treat-ent [interaction: F(1,84) = 4.240; p < 0.05, Fig. 4b]. Percent change
n locomotor activity according to the 1st session was significantlyigher in HR rats than LR rats on the 3rd and 5th sessions (p val-es < 0.05, Bonferroni test), which shows that HR rats were moreulnerable to nicotine induced locomotor sensitization develop-ent. Saline treatment in HR and LR rats did not produce any
ncrease in locomotor activity through the sessions (Fig. 4b).Nicotine challenge injection induced a significant difference
oth in HR and LR rats treated with nicotine or saline for fiveessions [for treatment: F(1,28) = 94.468; p < 0.0001]. However,nteraction was not statistically significant [F(1,28) = 1.972; p > 0.05,ig. 4c]. Post-hoc analysis showed that nicotine treated HR and LRats had significantly higher locomotor activity compared to theiraline treated counterparts (p values < 0.05, Fig. 4c). There were noifference between the groups after a saline challenge injectioninteraction: F(1,28) = 3.473; p > 0.05, Fig. 4c].
. Discussion
Nicotine, via activating nicotinic acetylcholine receptorsnAChR) on the dopaminergic cell bodies located in the VTA,ncreases dopaminergic activity in the NAcc [19]. Cadoni and Dihiara [6] indicated that a nicotine challenge injection, admin-
stered 24 h after a subchronic nicotine treatment, increasesopamine transmission in the core and, to a lesser extent, inhe shell of NAcc. Repeated administration of nicotine produces
Please cite this article in press as: Kayir H, et al. Impact of baseline prepulseBrain Res (2010), doi:10.1016/j.bbr.2010.08.004
ehavioural sensitization in experimental animals [29], which isndicated by a gradual increase in nicotine-induced locomotorctivity, although the administered dose of nicotine remains same20,28,33]. Sensitization has been suggested to play a role in thenitiation of addictive behaviour, and used as an experimental
development (c) ( p < 0.05, Student’s t test, compared with HI or LI counterpartstreated with nicotine, #p < 0.05, Student’s t test, compared with saline treatment,†p < 0.05, Bonferroni test, compared with saline or nicotine treated HI group).
paradigm to model the progression of behavioural and neurochem-ical changes leading to compulsive drug use in addicts.
The present study clearly shows that acute locomotor stimulanteffect and locomotor sensitization developing effects of nicotineare associated with the baseline prepulse inhibition levels. Nicotinesignificantly increased the locomotor activity of rats having higherPPI levels (high-inhibitory), where had no effect on the ones withlower PPI levels (low-inhibitory), on the first day of the experiment.However a significant locomotor sensitization was induced withdaily administrations for 5 days, and expressed after a challengeinjection only in low-inhibitory rats. The study also demonstratedthat baseline PPI levels did not related with baseline LRNE levels, awell-studied vulnerability factor for addictive drugs [3].
We applied the same locomotor sensitization protocol for thegroups classified according to LRNE to confirm that the methodwe used for measuring LRNE could also predict locomotor sensiti-zation. Acute nicotine treatment increased the locomotor activity
inhibition on nicotine-induced locomotor sensitization in rats. Behav
in both high- and low-responder rats, while a significant locomo-tor sensitization was induced and expressed only in high-responderrats. There was no difference between the rats with high or lowLRNE levels for their baseline PPI levels.

ING
B
rain R
rtidh
rarcilb
mbiPobioin
tslapaitari
fdabfsifsnihs[
vvfldpasbsH[P
[
[
[
[
[
[
[
[
[
[
ARTICLEModel
BR-6648; No. of Pages 6
H. Kayir et al. / Behavioural B
Some studies indicated that an optimal level of dopamine neu-otransmission is needed for intact sensorimotor gating functions,hus baseline PPI levels might reflect the basal dopaminergic tonen the brain areas at concern [2,27]. Moreover, effects of someopaminergic and NMDA receptor active drugs were shown to beighly modulated by baseline PPI levels of the individuals.
In human studies, baseline PPI levels were shown to effectesponses to several different drugs, such as dopaminergic agonistspomorphine [27], amantadine and pergolide [2], or dopamine D2eceptor antagonist and a typical antipsychotic haloperidol [8], andlozapine, an atypical antipsychotic with a multi-receptor bind-ng profile [35]. Effects of memantine, an NMDA antagonist with aow or moderate receptor affinity, on PPI were also modulated byaseline PPI levels in humans [32].
Feifel [11] showed that effects of both haloperidol and apo-orphine on PPI were negatively correlated with the individual
aseline PPI levels of rats. Because, haloperidol increased PPI onlyn rats having low baseline PPI levels, and apomorphine disruptedPI also in the low PPI group. Effects of 5-HT1A agonist 8-OH-DPATn PPI also modulated by baseline PPI levels in rats [13]. In a doublelind placebo controlled study, nicotine nasal spray improved PPI
n both schizophrenia patients and healthy controls. Moreover, thebserved changes were positively correlated with symptom sever-ty that patients with more clinical symptoms benefited more fromicotine-induced PPI improvement [16].
The present study extends the current literature about the rela-ionship between baseline PPI levels and the effects of drugs in theame task (i.e. PPI itself). This was done by indicating that base-ine PPI levels were also associated with acute locomotor stimulantnd sensitization inducing effects of nicotine. Because sensitizationlays a role in the initiation and continuation of addictive behaviournd reflects the adaptive neuroplastic alterations in brain systemsnvolved in motivational processes [24,25] our findings suggesthat baseline PPI levels may predict individual vulnerability to theddictive properties of nicotine. Furthermore baseline PPI was notelated with LRNE, another vulnerability factor for the nicotine-nduced sensitization.
Impulsivity has been defined as other vulnerability factorsor drug addictions [1,10]. Among the others two different sub-imensions of impulsivity, poor inhibitory control (impulsivection) and impulsive decision making (delay discounting) haveeen widely studied in rats [1,10]. Those sub-dimensions wereound to be related with each-other, because high-impulsive ratselected by using 5-choice serial reaction time (5-CSRT) task, madempulsive choices in delayed reward task [26]. However they wereound to predict different stages of nicotine seeking, such as impul-ive action reflected a vulnerability to initiation and continuation oficotine taking, whereas impulsive choice was related to an inabil-
ty to inhibit nicotine seeking during abstinence [10]. Furthermore,igh impulsive phenotype has different characteristics from sen-ation seeking or high locomotor response in a novel environment1].
In the present study, we evaluated and did not find any rele-ance between LI and HR traits, both of which were found to predictulnerability to nicotine-induced locomotor sensitization in dif-erent groups. This finding is very interesting because the currentiterature indicates that basal dopaminergic neurotransmission andopaminergic D2 receptor function in NAcc are important for bothhenotypes [12,23]. Some possible explanations for this discrep-ncy may be the involvement of other receptor or neuroendocrineystems and/or other brain areas in mediating the expression of
Please cite this article in press as: Kayir H, et al. Impact of baseline prepulseBrain Res (2010), doi:10.1016/j.bbr.2010.08.004
oth phenotypes. For example, HR rats have lower basal levels oferotonin and its main metabolite, 5-hydroxyindoleacetic acid (5-IAA), in the prefrontal cortex, the NAcc, and the dorsal striatum
23]. On the other hand, serotonin agonists are known to disruptPI, and most of the atypical antipsychotics have antagonistic activ-
[
[
PRESSesearch xxx (2010) xxx–xxx 5
ity on serotonergic receptors [12]. Further studies investigating therelationship between LI phenotype and other vulnerable pheno-types such as high impulsivity in 5-CSRT and delayed reward tasks,together with HR phenotype, would improve our knowledge onindividual responses to nicotine and other addictive drugs. In orderto improve our findings and define the low inhibitory phenotypeas a vulnerability factor some further studies by using other addic-tive drugs, such as cocaine or amphetamine, and by using otherexperimental approaches, such as self-administration are needed.
In conclusion, our results suggest that nicotine-induced locomo-tor stimulation and sensitization are associated with both baselinePPI and LRNE levels without interacting with each other.
Acknowledgement
This study was supported by Scientific and TechnologicalResearch Council of Turkey (TUBITAK, Project Numbers: 105S387,SBAG-3194). Authors would like to thank Dr. Murat Yildirim andMr. Selami Alan for their technical assistance to the study, and Dr.Burak Bal for reviewing the language of the manuscript.
References
[1] Belin D, Mar AC, Dalley JW, Robbins TW, Everitt BJ. High impulsivity predictsthe switch to compulsive cocaine-taking. Science 2008;320:1352–5.
[2] Bitsios P, Giakoumaki SG, Frangou S. The effects of dopamine agonists onprepulse inhibition in healthy men depend on baseline PPI values. Psychophar-macology (Berl) 2005;182:144–52.
[3] Blanchard MM, Mendelsohn D, Stamp JA. The HR/LR model: further evi-dence as an animal model of sensation seeking. Neurosci Biobehav Rev2009;33:1145–54.
[4] Braff DL, Geyer MA, Light GA, Sprock J, Perry W, Cadenhead KS, et al. Impactof prepulse characteristics on the detection of sensorimotor gating deficits inschizophrenia. Schizophr Res 2001;49:171–8.
[5] Cadenhead KS, Light GA, Geyer MA, McDowell JE, Braff DL. Neurobiologicalmeasures of schizotypal personality disorder: defining an inhibitory endophe-notype? Am J Psychiatry 2002;159:869–71.
[6] Cadoni C, Di Chiara G. Differential changes in accumbens shell andcoredopamine in behavioral sensitization to nicotine. Eur J Pharmacol2000;387:R23–5.
[7] Cloninger CR. Temperament and personality. Curr Opin Neurobiol1994;4:266–73.
[8] Csomor PA, Stadler RR, Feldon J, Yee BK, Geyer MA, Vollenweider FX. Haloperi-dol differentially modulates prepulse inhibition and p50 suppression in healthyhumans stratified for low and high gating levels. Neuropsychopharmacology2008;33:497–512.
[9] de Wit H, Uhlenhuth EH, Johanson CE. Individual differences in the reinforcingand subjective effects of amphetamine and diazepam. Drug Alcohol Depend1986;16:341–60.
10] Diergaarde L, Pattij T, Poortvliet I, Hogenboom F, de Vries W, Schoffelmeer AN,et al. Impulsive choice and impulsive action predict vulnerability to distinctstages of nicotine seeking in rats. Biol Psychiatry 2008;63:301–8.
11] Feifel D. Individual differences in prepulse inhibition of startle as a measure ofindividual dopamine function. Behav Neurosci 1999;113:1020–9.
12] Geyer MA, Krebs-Thomson K, Braff DL, Swerdlow NR. Pharmacological studiesof prepulse inhibition models of sensorimotor gating deficits in schizophrenia:a decade in review. Psychopharmacology (Berl) 2001;156:117–54.
13] Gogos A, van den Buuse M. The importance of baseline in identifying 8-OH-DPAT-induced effects on prepulse inhibition in rats. Br J Pharmacol2007;150:750–7.
14] Gottesman II, Gould TD. The endophenotype concept in psychiatry: etymologyand strategic intentions. Am J Psychiatry 2003;160:636–45.
15] Graham FK. The more or less startling effects of weak prestimulation. Psy-chophysiology 1975;12:238–48.
16] Hong LE, Wonodi I, Lewis J, Thaker GK. Nicotine effect on prepulse inhibitionand prepulse facilitation in schizophrenia patients. Neuropsychopharmacology2008;33:2167–74.
17] Hooks MS, Colvin AC, Juncos JL, Justice Jr JB. Individual differences in basal andcocaine stimulated extracellular dopamine in the nucleus accumbens usingquantitative microdialysis. Brain Res 1992;587:306–12.
18] Kilts CD. The changing roles and targets for animal models of schizophrenia.Biol Psychiatry 2001;50:845–55.
19] Mansvelder HD, McGehee DS. Cellular and synaptic mechanisms of nicotine
inhibition on nicotine-induced locomotor sensitization in rats. Behav
addiction. J Neurobiol 2002;53:606–17.20] Miller DK, Harrod SB, Green TA, Wong MY, Bardo MT, Dwoskin LP. Lobeline
attenuates locomotor stimulation induced by repeated nicotine administrationin rats. Pharmacol Biochem Behav 2003;74:279–86.
21] Nestler EJ. Is there a common molecular pathway for addiction? Nat Neurosci2005;8:1445–9.

ING
B
6 rain R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ARTICLEModel
BR-6648; No. of Pages 6
H. Kayir et al. / Behavioural B
22] Piazza PV, Deminière JM, Le Moal M, Simon H. Factors that predict indi-vidual vulnerability to amphetamine self-administration. Science 1989;245:1511–3.
23] Piazza PV, Rouge-Pont F, Deminiere JM, Kharoubi M, Le Moal M, Simon H.Dopaminergic activity is reduced in the prefrontal cortex and increased in thenucleus accumbens of rats pre-disposed to develop amphetamine selfadmin-istration. Brain Res 1991;567:169–74.
24] Robinson TE, Berridge KC. The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res Brain Res Rev 1993;18:247–91.
25] Robinson TE, Berridge KC. Incentive-sensitization and addiction. Addiction2001;96:103–14.
26] Robinson ES, Eagle DM, Economidou D, Theobald DE, Mar AC, Murphy ER, etal. Behavioural characterisation of high impulsivity on the 5-choice serial reac-tion time task: specific deficits in ‘waiting’ versus ‘stopping’. Behav Brain Res2009;196:310–6.
27] Schellekens AF, Grootens KP, Neef C, Movig KL, Buitelaar JK, Ellenbroek B, et al.Effect of apomorphine on cognitive performance and sensorimotor gating in
Please cite this article in press as: Kayir H, et al. Impact of baseline prepulseBrain Res (2010), doi:10.1016/j.bbr.2010.08.004
humans. Psychopharmacology (Berl) 2010;207:559–69.28] Shim I, Kim HT, Kim YH, Chun BG, Hahm DH, Lee EH, et al. Role of nitric oxide
synthase inhibitors and NMDA receptor antagonist in nicotine-induced behav-ioral sensitization in the rat. Eur J Pharmacol 2002;443:119–24.
29] Stewart J, Badiani A. Tolerance and sensitization to the behavioral effects ofdrugs. Behav Pharmacol 1993;4:289–312.
[
[
PRESSesearch xxx (2010) xxx–xxx
30] Suto N, Austin JD, Vezina P. Locomotor response to novelty predictsa rat’s propensity to self-administer nicotine. Psychopharmacology (Berl)2001;158:175–80.
31] Swerdlow NR, Light GA, Cadenhead KS, Sprock J, Hsieh MH, Braff DL. Startlegating deficits in a large cohort of patients with schizophrenia: relationshipto medications, symptoms, neurocognition, and level of function. Arch GenPsychiatry 2006;63:1325–35.
32] Swerdlow NR, van Bergeijk DP, Bergsma F, Weber E, Talledo J. Theeffects of memantine on prepulse inhibition. Neuropsychopharmacology2009;34:1854–64.
33] Ulusu U, Uzbay IT, Kayir H, Alici T, Karakas S. Evidence for the role of nitricoxide in nicotine-induced locomotor sensitization in mice. Psychopharmacol-ogy (Berl) 2005;178:500–4.
34] Uzbay T, Kayir H, Goktalay G, Yildirim M. Agmatine disrupts prepulse inhibitionof acoustic startle reflex in rats. J Psychopharmacol 2010;24:923–9.
35] Vollenweider FX, Barro M, Csomor PA, Feldon J. Clozapine enhances prepulseinhibition in healthy humans with low but not with high prepulse inhibitionlevels. Biol Psychiatry 2006;60:597–603.
inhibition on nicotine-induced locomotor sensitization in rats. Behav
36] Weiss IC, Feldon J. Environmental animal models for sensorimotor gat-ing deficiencies in schizophrenia: a review. Psychopharmacology (Berl)2001;156:305–26.
37] Zuckerman M, Ball S, Black J. Influences of sensation seeking, gen-der, risk appraisal, and situational motivation on smoking. Addict Behav1990;15:209–20.
Copyright © 2022 FDOKUMEN




















