
Structure and functions of the human amyloid precursor protein: The whole is more than the sum of...
22
Structure and functions of the human amyloid precursor protein: The whole is more than the sum of its parts Matthias Gralle 1, * , Se ´rgio T. Ferreira * Instituto de Bioquı ´mica Me ´dica, Programa de Bioquı ´mica e Biofı ´sica Celular, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ 21944-590, Brazil Received 15 September 2006; received in revised form 26 October 2006; accepted 1 February 2007 Abstract The amyloid precursor protein (APP) is a transmembrane protein that plays major roles in the regulation of several important cellular functions, especially in the nervous system, where it is involved in synaptogenesis and synaptic plasticity. The secreted extracellular domain of APP, sAPPa , acts as a growth factor for many types of cells and promotes neuritogenesis in post-mitotic neurons. Alternative proteolytic processing of APP releases potentially neurotoxic species, including the amyloid-b (Ab) peptide that is centrally implicated in the pathogenesis of Alzheimer’s disease (AD). Reinforcing this biochemical link to neuronal dysfunction and neurodegeneration, APP is also genetically linked to AD. In this review, we discuss the biological functions of APP in the context of tissue morphogenesis and restructuring, where APP appears to play significant roles both as a contact receptor and as a diffusible factor. Structural investigation of APP, which is necessary for a deeper understanding of its roles at a molecular level, has also been advancing rapidly. We summarize recent progress in the determination of the structure of isolated APP fragments and of the conformations of full-length sAPPa , in both monomeric and dimeric states. The potential role of APP dimerization for the regulation of its biological functions is also discussed. # 2007 Elsevier Ltd. All rights reserved. Keywords: Alzheimer’s disease; Amyloid precursor protein; Adhesion protein; Structural determination; Dimerization; Functions Contents 1. Morphoregulatory molecules ....................................................................... 12 1.1. Adhesion proteins .......................................................................... 12 1.2. Secreted factors ........................................................................... 13 2. APP in brain function............................................................................ 13 2.1. Development ............................................................................. 13 2.2. Adult phase .............................................................................. 14 2.3. Secreted APP ............................................................................. 15 3. APP in Alzheimer’s disease ....................................................................... 15 3.1. Genesis of the amyloid-b peptide ............................................................... 15 3.2. Familial Alzheimer’s disease .................................................................. 16 3.3. APP and caspases .......................................................................... 16 3.4. Sporadic Alzheimer’s disease .................................................................. 16 4. Determination of protein structure ................................................................... 17 5. APP structure ................................................................................. 18 www.elsevier.com/locate/pneurobio Progress in Neurobiology 82 (2007) 11–32 Abbreviations: APP, b-amyloid precursor protein; APLP, APP-like protein; HCZ, heparin-, copper- and zinc-binding fragment of APP; KPI, Kunitz-type protease inhibitor domain of APP; LRP, low density lipoprotein receptor-related protein; NGF, nerve growth factor; NMR, nuclear magnetic resonance; sAPP, secreted APP; SAXS, small-angle X-ray scattering * Corresponding authors. Tel.: +55 21 2562 6789; fax: +55 21 2270 8647. E-mail addresses: [email protected] (M. Gralle), [email protected] (S.T. Ferreira). 1 Present address: Cell Biophysics Group, European Neuroscience Institute, Waldweg 33, 37073 Go ¨ttingen, Germany. 0301-0082/$ – see front matter # 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.pneurobio.2007.02.001
Transcript of Structure and functions of the human amyloid precursor protein: The whole is more than the sum of...

www.elsevier.com/locate/pneurobio
Progress in Neurobiology 82 (2007) 11–32
Structure and functions of the human amyloid precursor protein:
The whole is more than the sum of its parts
Matthias Gralle 1,*, Sergio T. Ferreira *
Instituto de Bioquımica Medica, Programa de Bioquımica e Biofısica Celular, Universidade Federal do Rio de Janeiro,
Rio de Janeiro, RJ 21944-590, Brazil
Received 15 September 2006; received in revised form 26 October 2006; accepted 1 February 2007
Abstract
The amyloid precursor protein (APP) is a transmembrane protein that plays major roles in the regulation of several important cellular functions,
especially in the nervous system, where it is involved in synaptogenesis and synaptic plasticity. The secreted extracellular domain of APP, sAPPa,
acts as a growth factor for many types of cells and promotes neuritogenesis in post-mitotic neurons. Alternative proteolytic processing of APP
releases potentially neurotoxic species, including the amyloid-b (Ab) peptide that is centrally implicated in the pathogenesis of Alzheimer’s
disease (AD). Reinforcing this biochemical link to neuronal dysfunction and neurodegeneration, APP is also genetically linked to AD. In this
review, we discuss the biological functions of APP in the context of tissue morphogenesis and restructuring, where APP appears to play significant
roles both as a contact receptor and as a diffusible factor. Structural investigation of APP, which is necessary for a deeper understanding of its roles
at a molecular level, has also been advancing rapidly. We summarize recent progress in the determination of the structure of isolated APP fragments
and of the conformations of full-length sAPPa, in both monomeric and dimeric states. The potential role of APP dimerization for the regulation of
its biological functions is also discussed.
# 2007 Elsevier Ltd. All rights reserved.
Keywords: Alzheimer’s disease; Amyloid precursor protein; Adhesion protein; Structural determination; Dimerization; Functions
Contents
1. Morphoregulatory molecules . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
1.1. Adhesion proteins. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
1.2. Secreted factors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2. APP in brain function. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2.1. Development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2.2. Adult phase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.3. Secreted APP. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
3. APP in Alzheimer’s disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
3.1. Genesis of the amyloid-b peptide . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
3.2. Familial Alzheimer’s disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
3.3. APP and caspases. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
3.4. Sporadic Alzheimer’s disease. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
4. Determination of protein structure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
5. APP structure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
Abbreviations: APP, b-amyloid precursor protein; APLP, APP-like protein; HCZ, heparin-, copper- and zinc-binding fragment of APP; KPI, Kunitz-type
protease inhibitor domain of APP; LRP, low density lipoprotein receptor-related protein; NGF, nerve growth factor; NMR, nuclear magnetic resonance; sAPP,
secreted APP; SAXS, small-angle X-ray scattering
* Corresponding authors. Tel.: +55 21 2562 6789; fax: +55 21 2270 8647.
E-mail addresses: [email protected] (M. Gralle), [email protected] (S.T. Ferreira).1 Present address: Cell Biophysics Group, European Neuroscience Institute, Waldweg 33, 37073 Gottingen, Germany.
0301-0082/$ – see front matter # 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.pneurobio.2007.02.001

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3212
5.1. Overview. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
5.2. Protease inhibitory domain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
5.3. N-Terminal domain. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
5.4. Central domains . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
5.5. Interactions between APP domains . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
6. Structure of full-length sAPPa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
6.1. Direct measurements on full-length sAPPa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
6.2. Topology and positioning of APP fragments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
7. Dimerization of sAPPa and APP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
8. Functional implications from structural studies with sAPPa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
9. Future perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
9.1. Possible questions to be addressed . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
9.2. APP and cholinergic transmission. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
9.3. Implications for APLP2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
9.4. Unexpected functions of APLP2 and APP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
10. Conclusions. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
1. Morphoregulatory molecules
Most of the research on the amyloid precursor protein (APP)
has aimed to understand its physiological and pathological roles
in the central nervous system (CNS). Because there are reasons
to believe that APP may have important functions in
intercellular communication in the brain, we initially present
a brief discussion of the molecular basis of tissue organization
and function, first in general and then in the specific case of the
CNS.
Because of the complexity of human tissues, a large part of
what is known about their functions, regulation and develop-
ment has been generated by studies carried out using
dissociated cellular models. Despite the wealth of information
such studies have provided, it is necessary to recognize the
limits posed by the use of cellular models and the types of
questions such models cannot answer. In the same way as the
large number of different molecular components of a cell,
connected by multiple non-linear relations, leads to the
emergence of new properties at the cellular level, the large
number of different cells in a tissue, connected by multiple
signaling pathways, can also give rise to new properties that
cannot be easily or at all derived from studies of isolated cells;
these so-called emergent properties are often among the most
interesting ones, both to the researcher and to the physician
(Nelson et al., 2005). The most complex human tissue is the
CNS, as it contains an immense number of cell types of distinct
morphologies, biochemical and electrophysiological properties
(Toledo-Rodriguez et al., 2004). In contrast to the immune
system, which also comprises a very large number of
functionally distinct cells, the nervous system additionally
has an exact spatiotemporal organization: each of its cells is
defined not only by its characteristic phenotype but also by
specific coordinates within the tissue and by precise temporally
regulated communication with distinct partners.
The most interesting properties of the CNS, among them the
formation and storage of memories and their later retrieval or
extinction, arise from interactions between specific nerve cells
localized in different parts of the brain. Although a vast number
of studies have examined different electrophysiological or
biochemical markers of neuronal plasticity as surrogates of
memory formation, tissue-level emergent phenomena such as
memory are not immediately intelligible from the study of
isolated neurons or glia cells—the effect of a given protein in
dissociated cells may often appear very distant from that in the
functional brain. Hence, two of the most important questions in
structural neuroscience—necessary for understanding complex
functional phenomena such as memory—are as follows: (1)
How do the cells initially organize to give rise to the exact
three-dimensional structure of the brain? (2) How do
intercellular connections restructure and reorganize during
life, responding to environmental stimuli?
During both development and adult phases, external
information in the form of sensorial stimuli interacts with
genetically encoded information in brain cells in order to guide
the (re)organization of the tissue. Two classes of molecules that
appear to play key roles in the development of the CNS and in
its plasticity are the cell adhesion and the substrate adhesion
molecules (Edelman and Cunningham, 1990; Washbourne
et al., 2004), while another important class is represented by
secreted molecules.
1.1. Adhesion proteins
In many cell types, from unicellular organisms to human
epithelia, adhesion regulates proliferation. In fact, considering
their diverse biological functions, the term ‘‘contact receptors’’
has been suggested as a substitute for ‘‘adhesion proteins’’,
which might suggest a merely mechanical role (Gebbink et al.,
1995). In the developing nervous system, as well as in the
neurogenesis that continues on a reduced scale in the adult
human brain, contact with other cells and with extracellular
molecules is an important regulator of proliferation. A well-
known example is the division of the neuroblasts of the

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 13
ventricular zone, which give rise to the cerebral cortex: some of
the adhesion proteins expressed in the apical membrane of the
neuroblast, probably at adhesion junctions, maintain the
neuroblast and its apical descendents in a proliferative state.
Thus, adhesion proteins play an important role in tissue
morphogenesis (Lien et al., 2006), a role that would be difficult
to infer from the study of neuroblast development in an in vitro
two-dimensional culture only.
Contact receptors, which in the proliferative state mediate
cell adhesion, continue to function in post-mitotic neurons.
Interaction between developing neurons and radial glia cells is
mediated by contact receptors and guides the future neurons
while they travel through the embryonic cortex (Trapp and
Hauer, 1994). After the developing neuronal cell reaches its
correct localization in the tissue, small processes arise from the
soma. Adhesion to the substrate will cause one of these
processes to become an axon, while the others later become
dendrites (Heidemann, 1996). At the tip of the processes,
specialized structures known as growth cones explore the
environment and guide the extension of the process. While
adhesion of the cell body inhibits cell migration, adhesion of
the tips of the filopodia in the growth cone to some types of
substrate promotes and is necessary for the extension of the
process, while repulsion by other types of substrate leads to
retraction or degeneration of the process. These different types
of cellular responses are not due to a simple biomechanical
effect of adhesion, but rather to signaling cascades set in motion
by contact receptors present in the growth cone (Heidemann,
1996). Finally, when the growth cone of an axon encounters an
adequate target cell, a synapse is formed. The central role of
adhesion molecules for general synaptogenesis, for the
specificity of synapses and for their continuing plasticity is
only now beginning to be understood (Washbourne et al.,
2004). Some of the same adhesion molecules that regulate the
cell cycle in mitotic cells regulate the extension and retraction
of axons and dendrites and the formation of synapses as well
(Goda, 2002).
While neuritogenesis and synaptogenesis reach a peak in
the perinatal period (Dunaevsky et al., 1999; Moya et al.,
1994), the extension and retraction of dendrites and axon
collaterals, the formation of synapses and the modification of
existing synapses continue during our whole life. Even in the
adult phase, there is considerable plasticity: in some areas of
the human brain, the mean total length of all dendrites of a
neuron increases 10% with age when individuals in their 30s
and 40s are compared to those in their 50s and 60s (Arendt
et al., 1998). This overall increase results from continuous
modifications, both extension and retraction, that take place in
a time scale of minutes (Dunaevsky et al., 1999; Engert and
Bonhoeffer, 1999). The formation and modification of long-
term memories and their extinction seem to depend on the
formation and modification of synapses, and, once again,
adhesion molecules, or rather contact receptors, contribute to
synaptic plasticity (Goda, 2002; Washbourne et al., 2004). It is
clear from this short discussion that contact receptors will
generally not function to generate a clear, environment-
independent phenotype; rather, they function as ‘‘resources’’,
i.e. they contribute to the ability of each cell in a tissue to
interact with its surroundings (Moss, 2003).
1.2. Secreted factors
Morphoregulatory communication between the cells of the
nervous system depends not only on adhesion molecules but
also on secreted factors. The latter include both proteins known
as growth factors and small molecule neurotransmitters. While
some secreted molecules diffuse freely through the extra-
cellular space and act over considerable distances, especially in
the embryonic nervous system, many others are released at the
synaptic cleft and act only in the immediate neighborhood.
Every synaptic transmission event may, in addition to its
immediate effect of transmitting information, also strengthen
(or sometimes weaken) the morphological connection between
pre- and postsynaptic cells, thereby allowing information
storage for longer times. While the effect of a secreted molecule
on a cell, even an isolated one, may be intense and obvious, it is
important to keep in mind that the effect of that molecule in the
functioning cellular network of the tissue may be more
complex.
2. APP in brain function
2.1. Development
The functions of APP are much better defined at the tissue
level than at the cellular and subcellular levels, and serve as
examples for many of the functions of contact receptors and
secreted factors described above. Full-length APP contains a
large extracellular domain (corresponding to �88% of the total
protein mass for the main neuronal isoform), a single
transmembrane region and a small cytoplasmic tail (Fig. 1).
This type of structure led its discoverers to suggest that APP
might function as a receptor (Kang et al., 1987). Though
functions, ligands and intracellular pathways have been well
established for many cellular receptors studied in vitro, this
kind of knowledge has been slow to emerge for APP, despite the
fact that an impressive body of immunohistochemical and
genetic data indicate the functional importance of APP in
nervous tissue.
Many studies using cells in culture have consistently shown
a role of APP in adhesion to other cells or to extracellular
matrix components (Kibbey et al., 1993; Qiu et al., 1995; Sabo
et al., 2001; Schubert et al., 1989; Soba et al., 2005). For
example, using an immortalized neuronal cell line, an early
study showed that APP increases adhesion to the substrate, to
other cells of the same line and to glia cells (Breen et al., 1991).
While these findings are interesting in themselves and
immediately suggest a role of APP in tissue maintenance, an
interesting aspect of the dynamic function of the neuron-glia
adherence promoted by APP emerged only from tissue-level
analysis: The discovery that APP is expressed at high
concentrations in the radial glia, which guides future neurons
to their correct positions in the embryonic cortex, suggests that
the adhesion of neurons to glia cells promoted by APP plays an

Fig. 1. Domain structure of APP. Grey ellipses: APP domains for which complete or partial structures are known. 1mwp, 1owt, 1aap, 1rw6, 1tkn correspond to the
PDB access codes for the fragments of known structure. (DE)n, RC: Intervening non-conserved stretches (Asp- and Glu-rich; random coil) suggested to be largely
unstructured or to have non-standard secondary structures (see Sections 5.3 and 3.4). KPI: Kunitz-type protease inhibitory domain (see Section 5.2). HBD1, HBD2:
Heparin-binding domains (see Sections 5.3 and 5.4). CuBD: Copper-binding domain (see Section 5.3). Ab: Amyloidogenic sequence (see Section 3.1). TM:
transmembrane domain. Cyt: Cytoplasmic domain. Cleavage by a-secretase (vertical arrow) releases sAPPa and the membrane-bound C-terminal fragment (CTFa),
as indicated (see Section 2.3). Amino acid numbering is for APP770 (see Section 5.1). Figure reprinted with permission from Gralle et al. (2006).
M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3214
important role in brain development (Trapp and Hauer, 1994).
Based on the findings described above, it seems warranted to
describe APP as a contact receptor. Indeed, several high-
specificity ligands of APP in the extracellular matrix have been
identified, though the mechanisms of signal transduction
triggered by such interactions are still unclear (Beher et al.,
1996; Ho and Sudhof, 2004; Kibbey et al., 1993; Williamson
et al., 1996; see also Sections 5 and 8).
As is also the case for other contact receptors, the adhesion-
promoting activity of APP in migratory cells is accompanied by
a neuritogenic activity in stationary, immature neurons in vitro
and in vivo (Allinquant et al., 1995; LeBlanc et al., 1992;
Majocha et al., 1994; Milward et al., 1992; Ohsawa et al., 1997;
Perez et al., 1997; Qiu et al., 1995; Small et al., 1994). The
biochemical basis for the involvement of APP in axonal growth
has recently been elucidated: The cytoplasmic tail of APP binds
to the actin regulatory proteins Fe65 and Mena, and this ternary
complex is localized to the dynamic adhesion sites in the
growth cone (Sabo et al., 2003). In non-neuronal cells, co-
expression of APP and Fe65 drastically increases their
migration velocity (Sabo et al., 2001). While studies with
cultured cells alone cannot fully reveal the spatial and temporal
importance of the involvement of APP, in vivo studies have shed
some light on where and when in brain development APP is
especially needed. A role of APP in the formation of long-
distance connections is supported indirectly by the fact that its
levels rise in the perinatal period, when axons rapidly extend in
the olfactory system of the rat and fall again after this period
(Clarris et al., 1995). During the peak period of axon extension,
APP and its homologues, the APP-like proteins APLP1 and
APLP2, comprise most of the protein mass that is rapidly
transported by the optical nerve (Lyckman et al., 1998; Moya
et al., 1994). Furthermore, the importance of APP for axon
arborization was demonstrated in Drosophila, where the
absence of the APP homologue leads to a failure of arborization
and, in consequence, to a reduced reproductive fitness of the
organism (Leyssen et al., 2005).
APP is also important for the next step of neuronal
morphogenesis, i.e., the formation of functional synapses. The
expression of APP in cultured hippocampal neurons increases
their response to glutamate (Tominaga-Yoshino et al., 2001),
and other studies have demonstrated electrophysiological
consequences of the lack of APP (Priller et al., 2006). On
the other hand, in vivo studies have focused on the importance
of APP for both morphological and electrophysiological
maturity of specific synapses. In double knock-out mice for
APP and its nearest homologue, APLP2, motoneuron axons
bypass their targets in muscle fibers and do not form synaptic
terminals at the correct sites, leading to a dramatic increase in
transmission failure on the day of birth (Wang et al., 2005) and
to the death of these animals a few days later (Heber et al.,
2000; Herms et al., 2004; Wang et al., 2005; Yang et al., 2005).
In the central nervous system, APP seems to be important for
the maturation of specific subtypes of neurons (Salbaum and
Ruddle, 1994).
2.2. Adult phase
In the adult brain, APP is expressed at high levels in those
regions that undergo more intense synaptic modifications
(Loffler and Huber, 1992; Ouimet et al., 1994), and its
expression level increases in animals reared in an enriched
environment (Huber et al., 1997). In an interesting example of
APP function in live, awake adult animals, specific knock-down

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 15
of APP in retinal ganglion cells leads to a reduction in
metabolic activity in the area innervated by those cells (Herard
et al., 2005). It is, therefore, not surprising that its permanent
absence in knock-out animals induces memory deficits
(Dawson et al., 1999).
Only a small fraction of the APP transported to the neuronal
plasma membrane remains there for any longer period of time
(Allinquant et al., 1994; Storey et al., 1999). This pool of APP
present at the plasma membrane probably mediates the
physiological functions described above. However, APP
metabolism in mature neurons is quite complex. Upon
emerging from the trans-Golgi network, APP is initially
inserted into fast axonal transport vesicles (Koo et al., 1990;
Sisodia et al., 1993). The hypothesis that APP itself anchors
these vesicles to kinesin, the molecular motor that transports
them along the axon to the synapses (Kamal et al., 2000), has
been broadly questioned (Lazarov et al., 2005). The details of
axonal transport of APP were very recently investigated using
tau mutant cells (Goldsbury et al., 2006). After the vesicles fuse
with the axonal or synaptic plasma membrane and APP appears
at the surface of the neuron, part of it is cleaved by proteases
collectively known as secretases and the extracellular domain is
secreted (see below); another fraction of APP returns via
endocytic vesicles to the cell body and to the dendrites
(Marquez-Sterling et al., 1997; Yamazaki et al., 1995). The
function of this return journey of APP is still unclear, though the
regulated endocytosis of other contact receptors has been
shown to be necessary for their functions (Schaefer et al.,
2002). The combination of partial proteolysis by the secretases
and re-internalization of uncleaved APP by the endocytic
pathway shortens the half-life of APP at the plasma membrane
to just �10 min (Koo and Squazzo, 1994).
2.3. Secreted APP
While the trafficking of transmembrane APP is still
incompletely understood, however, the biological effects of
the cleaved and secreted form of APP (known as sAPP, for
secreted APP) have been extensively investigated. Several
extracellular proteases of the ADAM (‘‘a disintegrin and
metalloproteinase’’) family cleave APP with the specificity
expected for the major secretase (‘‘a-secretase’’), 12 amino
acid residues upstream from the extracellular face of the
membrane, releasing the extracellular domain termed sAPPa
(Fig. 1). Among these proteases, ADAM10 is necessary for the
secretion of sufficient amounts of sAPPa in vivo (Postina et al.,
2004). In neuronal precursors in the adult brain, as well as in
non-neuronal cells, sAPPa acts as a proliferative factor
(Pietrzik et al., 1998; Saitoh et al., 1989; Caille et al., 2004).
In addition to stimulating proliferation, sAPPa also increases
the motility of keratinocytes (Kirfel et al., 2002) and of
melanocyte processes (Quast et al., 2003). Interestingly, sAPPa
increases neurite outgrowth in immortalized neuronal cell lines
(Milward et al., 1992). Not only the full-length transmembrane
form of APP but also sAPPa is transported through the axon
during the perinatal period (Lyckman et al., 1998; Moya et al.,
1994). At the synapse, sAPPa modulates transmission and is
neuroprotective against ischemic and excitotoxic injury
(Furukawa et al., 1996a,b; Mattson et al., 1993; Morimoto
et al., 1998; Smith-Swintosky et al., 1994). Probably as a
consequence of synaptic modulation, neuroprotection or both,
intracerebral injection of sAPPa enhances memory perfor-
mance in adult rats (Meziane et al., 1998).
Today it is clear that APP, acting both as a contact receptor in
its transmembrane form and as a proliferation and motility
factor in its secreted form, has important morphoregulatory
functions. The importance of APP is underscored by the
existence of two homologous proteins with partly redundant
functions that can compensate for part of the effects of eventual
APP loss (Heber et al., 2000). A more complete view of the
development of the brain and of synaptic plasticity will have to
await further advances in our knowledge of the dynamics of
neural circuits, on the one hand (Sakurai, 1999), and of the
exact modifications and relations of the participating mole-
cules, on the other. Elucidation of the structural basis of the
interactions of APP with its physiological partners will provide
important insight towards such an integrated view.
3. APP in Alzheimer’s disease
3.1. Genesis of the amyloid-b peptide
Retrospectively, it may now seem strange that APP was not
initially discovered because of its morphoregulatory roles in the
healthy brain, but rather because of the characteristic deposition
of plaques (known as senile plaques) that contain the amyloid-b
peptide (Ab), a proteolytic fragment derived from APP, in the
brains of demented patients (Glenner and Wong, 1984; Masters
et al., 1985). Following the elucidation of the amino acid
sequence of the Ab peptide, the cDNA of the principal neuronal
isoform of human APP (APP695) was cloned (Kang et al., 1987)
and, in the following year, the two isoforms that predominate in
other cell types (APP751 and APP770) were characterized
(Kitaguchi et al., 1988; Ponte et al., 1988; Tanzi et al., 1988).
The Ab peptide is derived from a quantitatively less important
cleavage pathway of APP (carried out by b-secretase; see
below) than the cleavage by a-secretase mentioned above. Ab
may serve neurotrophic and signaling functions when present at
low physiological concentrations (Kamenetz et al., 2003;
Yankner et al., 1990). However, amyloid aggregates that
accumulate in vivo in the brains of AD patients and that also
form in vitro in aqueous solutions of Ab are potent neurotoxins
(for an early study, see Pike et al., 1991; reviewed in Klein et al.,
2006). In spite of the notable presence of senile plaques in aged
brains and, particularly, in the brains of individuals affected by
Alzheimer’s disease, the degree of dementia in the patients
correlates better with the brain levels of soluble Ab than with
the quantity of Ab aggregated in plaques (McLean et al., 1999;
Lue et al., 1999). This important neuropathological observation
supports the notion that the species responsible for synaptic
dysfunction in AD patients may be soluble oligomers of Ab,
which interfere with synaptic plasticity (Lambert et al., 1998;
Walsh et al., 2002; Wang et al., 2002), rather than the fibrillar
amyloid aggregates that were initially considered to be the main

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3216
neurotoxic species. Soluble Ab oligomers are present at much
higher concentrations in AD brains than in unaffected brains
(Gong et al., 2003), and they bind specifically to a
subpopulation of dendritic spines (Lacor et al., 2004).
The Ab peptide is proteolytically generated by the
successive action of b-secretase at the N-terminus of the so-
called amyloidogenic sequence, and g-secretase, which cleaves
APP at one of several different peptide bonds in its
transmembrane domain to form the C-terminus of Ab (for
an overview of the secretases, see De Strooper and Annaert,
2000; Turner et al., 2003). b-Secretase was identified as an
aspartic protease (Vassar, 2002). On the other hand, g-secretase
activity is associated with a high-molecular-weigh protein
complex containing four necessary constituents (presenilin 1 or
2, nicastrin, Aph-1 and Pen-2), which cleave a large number of
substrates besides APP (De Strooper et al., 1999; Edbauer et al.,
2003; Kimberly et al., 2003; Wolfe et al., 1999). The existence
of other, accessory factors that might regulate the activity of g-
secretase and thereby potentially influence the genesis of Ab
was inferred from studies in Drosophila (Loewer et al., 2004);
two of these accessory factors have been recently reported to be
CD147 (Zhou et al., 2005) and TMP21 (Chen et al., 2006b).
Within these high-molecular-weight complexes, the active site
is localized in the presenilins (Esler et al., 2000; Tolia et al.,
2006; Wolfe et al., 1999). g-Secretase does not act on integral
APP, but only on fragments not containing the extracellular
domain, and therefore it cleaves APP following its initial
processing by a- or b-secretase (Goo and Park, 2004; Struhl
and Adachi, 2000).
3.2. Familial Alzheimer’s disease
A small percentage of AD patients develop clinical
symptoms before the age of 60 and show mutations in the
APP gene. This indicates that alterations in this protein are
sufficient to cause the entire spectrum of disease-associated
clinical symptoms and pathological hallmarks (Goate et al.,
1991). In those cases with mutations near the b-cleavage site,
dementia appears to be causally related to a large increase in the
production of Ab (Citron et al., 1992). In other cases, such as
mutations in amino acids 716 and 717 of APP, there is no
increase in total Ab concentration, but only in that of its longer
and more amyloidogenic form, Ab42, leading to the proposal
that pathogenesis in those patients is due to the higher
proportion of Ab42 relative to Ab40 (Suzuki et al., 1994).
Another proposed explanation, however, involves pathological
signaling by mutant transmembrane APP, without necessarily
involving processing of the protein by the secretases (Yamatsuji
et al., 1996b).
No cases of Alzheimer’s disease caused by mutations or
dysfunction of b-secretase, or by lack of a-secretase, are known
to date. On the other hand, mutations in the presenilin genes are
known to be causally associated with familial Alzheimer’s
disease, and these mutations are much more frequent than those
in APP (De Strooper and Annaert, 2000).
g-Secretase cleavage of the membrane-bound fragment of
APP releases not only the Ab peptide but also the C-terminal
cytoplasmic domain of APP. The cytoplasmic domain, which
retains the sites of interaction with signaling proteins present in
transmembrane APP, has been reported to translocate to the
nucleus and regulate gene transcription in vitro (Cao and
Sudhof, 2001). However, the in vivo target genes of this
possible regulatory pathway have not yet been discovered and,
therefore, the consequences of this signaling pathway for the
physiological actions of APP and/or for the genesis of
Alzheimer’s disease are still unclear (see also Section 8).
3.3. APP and caspases
Another interesting aspect of APP biology is its cytoplasmic
cleavage by proteases of the caspase family, which leads to the
release of fragments different from those produced by the g-
secretase complex (De Strooper and Annaert, 2000). As the
primary and best-known function of caspases is to promote cell
death by apoptosis, a connection between neuronal death by
apoptosis and neuronal death in the advanced stages of
Alzheimer’s disease has been the usual interpretation of these
data (De Strooper and Annaert, 2000). It is noteworthy,
however, that apoptosis induced by caspase activation typically
proceeds on a time scale of hours to days, whereas neuronal
death in AD takes place over the course of many years. It is also
important to remember that the highest expression level of APP
is during the development of the brain, and that selective
neuronal death by apoptosis is necessary for correct develop-
ment (Kuida et al., 1996). Therefore, if cleavage of APP by
caspases really causes nerve cell death, it will be important to
first understand the role of this signaling event in normal
development before discussing its possible role in slow
neurodegeneration.
Over the past few years, it has become clear that apoptotic
signaling is not the only function of caspases and that their
activation can lead to other cellular events as well (Arama et al.,
2003; Kennedy et al., 1999). In the specific case of neurons,
caspase signaling may have functions in synaptic plasticity and
axonal growth; for example, caspase 1 can negatively regulate
long-term potentiation (Lu et al., 2006), and caspase 3 must be
activated in order for the growth cone to change direction
(Campbell and Holt, 2003).
The recent evidence for functions of the caspases in synaptic
plasticity, one of the areas of physiological action suggested for
APP, suggests that it would be interesting to re-examine the
published results on caspase cleavage of APP. It is possible that,
rather than killing neurons, cleavage of APP by caspases might
be another of the signaling pathways used to regulate neuronal
plasticity. Still, very recent evidence indicates that cleavage of
APP by caspases might be necessary for the progression of AD
(Galvan et al., 2006).
3.4. Sporadic Alzheimer’s disease
In spite of considerable advances in understanding the
pathogenesis of AD, a still unanswered question is why soluble
Ab levels, and specifically Ab oligomer levels (Lacor et al.,
2004), rise in the brains of the great majority of AD patients that

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 17
have no mutations in the APP or secretase genes. While we will
not attempt to solve this question here, a few observations on
the etiology of sporadic AD are summarized in the following.
One suggestion that is completely independent of the
metabolism of APP itself is a deficiency in the proteases that
physiologically degrade Ab (such as neprilysin and insulin-
degrading enzyme), causing accumulation of the peptide
(Selkoe, 2001). In another series of observations, a link between
cholesterol and late-onset AD has been observed: lowering
plasma cholesterol levels can contribute to decreasing the risk
of dementia (Jick et al., 2000), and certain alleles of the lipid
transport protein ApoE (alleles e2 and e4) are associated with
an increase or decrease, respectively, of the age of onset of
dementia (Corder et al., 1994; Strittmatter et al., 1993a). These
findings, again, have been sometimes explained in terms of a
direct interaction of cholesterol and/or ApoE4 with Ab, without
taking into account possible changes in APP function or
neuronal physiology (Strittmatter et al., 1993b; Kakio et al.,
2001). Cholesterol may also affect the trafficking or the affinity
of APP for the secretases and thereby influence the release of
Ab (Kojro et al., 2001). On the other hand, the importance of
cholesterol and of different ApoE isoforms for neuronal
plasticity (Arendt et al., 1997; Mauch et al., 2001) indicates that
disturbances of neuronal plasticity lead to an adaptive change in
APP metabolism, which ultimately has deleterious conse-
quences, culminating, whether because of increased Ab
production or otherwise, in dementia. Cerebral traumatism is
also a risk factor for dementia (Jellinger, 2004), which suggests
that the regeneration of neurons after an insult may cause
neurodegeneration. In this regard, a relevant observation is that
trauma leads directly to a (possibly adaptive) increase in the
expression of APP and to the deposition of Ab (Blasko et al.,
2004; Roberts et al., 1991). Finally, in different mouse models,
a deficiency in nerve growth factor (NGF) has been reported to
cause neuronal degeneration and, as a secondary consequence
of this degeneration, the deposition of Ab (for a review, see
Isacson et al., 2002).
In conclusion, there can be no doubt that mutations in the
APP gene are sufficient to cause the entire spectrum of clinical
symptoms and neuropathology of Alzheimer’s disease. On the
other hand, it is unclear why so many different risk factors (age,
plasma cholesterol level, ApoE4 status, trauma, deficiency in
growth factors and other environmental factors) should lead to
neurotoxicity in certain brain regions. One or more of the
functions of APP that are important for synaptic plasticity may
be activated in response to insults, leading to pathological
deregulation of APP metabolism. Possibly, the ensuing
accumulation of soluble oligomers of Ab in the synaptic cleft
then causes synaptic dysfunction, neuronal damage and
dementia in all of those cases.
4. Determination of protein structure
As mentioned above, a deeper understanding of the actions
of APP in the healthy and demented brain will require both
identifying the dynamic neuronal circuits which depend on
APP and understanding the molecular/structural basis of the
interactions of APP with other molecules. With respect to
normal neuronal physiology, this may be obvious. As noted in
the previous section, however, in AD, too, and especially in
sporadic forms of the disease, the possibility that adaptive
interactions between APP and its physiological binding
partners may lead to dementia by Ab-dependent or Ab-
independent mechanisms highlights the need for understanding
the structure of APP outside the amyloidogenic domain. In the
following, we briefly outline a few basic principles that guide
protein structural investigation and review recent advances in
the structural biology of both full-length and secreted forms of
human APP.
In general, structure/function relationships for any given
protein cannot be completely understood by simply elucidating
the structures of isolated fragments. The temptation to regard a
protein as an object equivalent to the sum of small parts
possibly arises from our intuitive nature of comparing
biological molecules to macroscopic machines made of metal
and/or plastic pieces. In such macroscopic machines, each
constituent part is usually hard (i.e., it does not deform
significantly during work) and often moves independently from
neighbouring parts. In contrast, however, the final folded
structure of a polypeptide chain depends on a large number of
attractive and repulsive molecular interactions that very nearly
cancel out each other. As a result, the stability of proteins is
often quite small compared to thermal energy at physiological
temperatures. Consequently, proteins are not static objects, but
rather exhibit considerable conformational dynamics (or
plasticity) ranging from very fast (subnanosecond) residue
motions (for early examples, see Ferreira, 1989; Ferreira et al.,
1994) to larger domain movements in the millisecond to second
time scales (reviewed in Weber, 1992). In fact, even in the case
of proteins that yield well-diffracting crystals suitable for high-
resolution structural determination by X-ray crystallography,
the apparently rigid crystallographic structures hide substantial
and significant fluctuations and should be actually viewed as
time-averaged structures (DePristo et al., 2004).
Methods to determine protein structures in solution by
nuclear magnetic resonance (NMR) spectroscopy or small-
angle X-ray scattering (SAXS) offer the possibility to model
protein dynamics explicitly (Arai et al., 2004; Lindorff-Larsen
et al., 2005). In addition to the low-amplitude movements
present in all proteins, experiments on dynamics have revealed
the existence of protein domains, or even entire polypeptide
chains, that exhibit the large mobility typical of unfolded states
even in their biologically active states (Wright and Dyson,
1999). Such proteins (or protein domains) have been described
as natively unstructured, or natively unfolded, and they usually
have the capacity to acquire well-defined structures upon
interactions with other proteins or other cellular components
(e.g., lipid bilayers) (Wright and Dyson, 1999; McMahon and
Gallop, 2005).
In the case of large proteins that contain several distinct
structural domains, the acquisition of the biologically active
structure by one domain may depend on the presence of another
domain, even if the two domains are apart in the amino acid
sequence (Fedorov and Baldwin, 1999; Jansens et al., 2002), or

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3218
it may depend on the participation of other protein(s)
(Szepanski et al., 1994). In this way, different domains and/
or polypeptide chains cooperate to create proteins with more
complex structures and functions. Consequently, determining
the structure of an isolated fragment or domain of a protein does
not guarantee that that fragment maintains the same structure
when inserted into the full-length protein, and certainly does
not offer reliable information on the topological relations
between different domains in the full-length protein.
The energetics that rule protein folding are also very
important for understanding the interactions between proteins
and other molecules (ligands, substrates). In many cases,
intramolecular interactions are so weak and/or distributed over
so many residues that the entire protein moves and changes
conformation when it contacts a partner. For example,
mutational studies have shown that protease specificity towards
substrates is not necessarily localized to the handful of residues
present at the active site, but rather can be broadly distributed
throughout the protease (Hedstrom et al., 1992). Similarly,
studies on protein dimers have shown that the energy of
interaction between the monomers is not the sum of the
interactions of each residue at the dimer interface. Groups of
amino acids interact strongly and non-additively to build up the
interface (Reichmann et al., 2005).
5. APP structure
5.1. Overview
APP is a protein of up to 770 amino acids (in its longest
isoform). To date, it has not been crystallized satisfactorily, nor
have any of its two homologues in vertebrates, APLP1 and
APLP2 (Sprecher et al., 1993; Wasco et al., 1992), or any other
family member in invertebrates, which means that there is no
atomic resolution structure for the full-length, transmembrane
form of APP. This lack of a structure is actually not too
surprising, as the number of atomic resolution structures
available for transmembrane proteins is still much lower than
for soluble proteins. NMR studies have shown that the C-
terminal cytoplasmic tail of APP has no rigid structure
(Kroenke et al., 1997), while an a-helical structure is assumed
for the transmembrane segment (Kang et al., 1987) (see Fig. 1
for the overall structure of APP).
A variety of signaling adaptor proteins bind to the
cytoplasmic domain of APP, including G0 (Brouillet et al.,
1999), APP-BP1 (Chow et al., 1996), Fe65 (Borg et al., 1996),
X11 (Borg et al., 1996) and JIP-1 (Matsuda et al., 2001). The
structural basis of the interaction of the cytoplasmic domain
with some of these proteins has been inferred from solution
NMR studies on the isolated C-terminal tail (Ramelot et al.,
2000; Pastorino et al., 2006), while the interaction of the
transmembrane domain of APP with the g-secretase complex
has been inferred from biochemical experiments (Berezovska
et al., 2003) but not yet structurally elucidated. Investigating the
complexes of these proteins with APP fragments is an
important experimental challenge. However, definite knowl-
edge about the activation or the processing of APP will likely
depend on the availability of a structure of the entire
transmembrane APP, as the cytoplasmic and transmembrane
domains are both very small in relation to the full-length protein
and unable to fold independently into stable tertiary structures.
On the other hand, elucidation of the structure of the large
extracellular domain of APP (sAPP) would be a great step
forward in understanding the mechanisms mediated by the
intact transmembrane protein. Moreover, sAPP is produced in
vivo (Van Nostrand et al., 1990), is capable of independent
folding (Araki et al., 1991) and has potent biological activities
(as described in Section 2.3). For these reasons, obtaining the
structure of the extracellular portion of APP would also be of
significant value in itself.
The APP gene has 19 exons (Yoshikai et al., 1991). Several
isoforms of APP are generated by alternative splicing of exons
7, 8 and 15, all of them coding for domains localized in the
extracellular portion of the molecule; APLP2 is also
alternatively spliced in a very similar way (Sandbrink et al.,
1994; Yoshikai et al., 1990). In non-neuronal cells, the longer
isoforms containing exon 7 or exons 7 and 8 predominate, while
neurons express principally the isoform lacking exons 7 and 8
(Kang and Muller-Hill, 1990). APP isoforms lacking exon 15
serve as core proteins for a proteoglycan called appican
(Pangalos et al., 1995). Expression of these latter isoforms is
important in many cell types, but neurons express very little
APP- or APLP2-lacking exon 15 (Sandbrink et al., 1994).
After loss of the signal sequence and cleavage by a-
secretase, the extracellular portion of the neuronal isoform of
APP (sAPPa695) has 594 amino acid residues and an expected
molecular mass of 68 kDa, without accounting for eventual
post-translational modifications. Crystallization of the full-
length extracellular domain of APP has been attempted by
several groups, including our own, and to our knowledge has
not yet yielded satisfactory crystals. Moreover, the large
molecular mass of sAPPa still is a huge challenge for
successful three-dimensional structural determination by
NMR. In the following, we review structural information on
smaller fragments of APP in the context of available
biochemical knowledge on the full-length protein, before
proceeding to recent structural data on the full-length protein.
5.2. Protease inhibitory domain
The first APP fragment to be crystallized was the protease
inhibitory domain, present only in the non-neuronal isoforms
(sAPPa751 and sAPPa770; Hynes et al., 1990). As this domain is
coded for by a single exon and is inserted in the middle of the
APP sequence (see Fig. 1), and considering that it is quite
homologous to other Kunitz-type protease inhibitors (KPI;
Hynes et al., 1990), it seems likely that it constitutes an
independent folding unit and that the structure obtained for the
isolated domain represents well the structure of the domain in
the context of full-length APP. The protease inhibitory
properties of this isolated domain in vitro are very similar to
the inhibitory properties of full-length sAPPa, except for the
potentiation of inhibition by heparin in the case of the intact
protein (Wagner et al., 1992). APP and sAPP isoforms

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 19
containing the KPI domain are ligands of the LDL-receptor-
related protein (LRP) and can be internalized through
interaction with this receptor (Kounnas et al., 1995). Therefore,
the biological effects of those isoforms of APP and sAPP that
contain the KPI domain might be regulated by LRP. In the
longest isoform, APP770, the KPI domain is immediately
followed by a short stretch of 19 amino acids without significant
homology to other known proteins and without known structure
(Richards et al., 1995).
5.3. N-Terminal domain
One of the regions most conserved among APP and its
homologues in humans and other species is the N-terminal region
of 172 residues localized immediately after the signal sequence
(Daigle and Li, 1993). This region of APP contains 12 cysteine
residues, which suggests the presence of disulfide bonds. In 1999,
a 96-residue N-terminal subfragment was crystallized (HBD1 in
Fig. 1) and in fact showed three disulfide bonds that constrain a
very rigid tertiary structure rich in b-sheets (Rossjohn et al.,
1999). This isolated domain is capable of binding to heparin
(Mok et al., 1997), to the extracellular matrix protein fibulin
(Ohsawa et al., 2001) and also to Ab (Van Nostrand et al., 2002).
On the other hand, this subfragment has a rather hydrophobic
exposed surface that is prone to aggregation or interaction with
other protein surfaces, suggesting that it does not function as an
independent folding unit in full-length APP.
The sequence immediately following the N-terminal
subfragment contains a domain that binds Cu2+ in vitro (Hesse
et al., 1994). This domain is easily degraded in vitro (Rossjohn
et al., 1999), possibly as a consequence of free radicals
generated by Fenton reaction mediated by bound copper ions
(Multhaup et al., 1998). However, its structure was determined
by NMR and crystallization (Barnham et al., 2003; Kong et al.,
2007), localizing another three disulfide bonds and the copper-
binding site. A peptide sequence that binds Zn2+ in vitro is also
present in this amino acid sequence of APP, and Zn2+ binding
increases the affinity of APP for heparin (Bush et al., 1993).
However, recent evidence indicates that this sequence does not
contain all the amino acid residues necessary to fully chelate
Zn2+ (Ciuculescu et al., 2005). A possible solution to this
discrepancy would be the contribution of an additional amino
acid residue from another APP domain to Zn2+ chelation. These
observations suggest that the N-terminal fragments of APP, i.e.
the heparin, Cu2+- and Zn2+-binding regions, do not seem to be
independent structural or functional units in the APP molecule.
The stretch of APP contained between the Zn2+-binding
sequence and the KPI-domain insertion point [(DE)n in Fig. 1]
is also easily degraded in vitro (Rossjohn et al., 1999). It is very
rich in negatively charged residues and consists almost entirely
of a low complexity region (Wootton and Federhen, 1996).1
Because of the predominance of only a few types of amino
acids, a naıve use of alignment and structural prediction
1 Determined using the FASTA server on the site http://zeldia.cap.ed.ac.uk/
ncbi_blast.html.
algorithms would indicate high degrees of helicity and
sequence conservation among APP family members in this
stretch. However, low complexity domains do not usually
display permanent secondary structures and, therefore, only the
predominance of acidic amino acids may be conserved, not the
sequence itself (Wootton and Federhen, 1996). It has not yet
been possible to determine the structure of this part of APP in
isolated form or together with other domains, and it has also not
been determined if it interacts with other domains of the
protein, for example via electrostatic interactions. A possible
functional role of this region has recently been proposed in
regulating mitochondrial membrane translocation of APP
(Anandatheerthavarada et al., 2003).
5.4. Central domains
The second half of the extracellular domain of APP,
following the exons that are inserted in the non-neuronal
isoforms, consists of a 179-residue segment that is well
conserved in evolution (HBD2 in Fig. 1) followed by a 166-
residue segment that is specific for vertebrate APP (Daigle and
Li, 1993). The latter segment is rapidly degraded by proteases,
suggesting that it may exhibit a less compact conformation,
while the well-conserved region was predicted to have a high a-
helical content (Gralle et al., 2002; Sandbrink et al., 1994).
Indeed, the solution NMR structure of part of this region
(Dulubova et al., 2004) agrees perfectly with the crystal-
lographic structure of the entire conserved region (Wang and
Ha, 2004), which features a very long a-helix with two shorter
helix bundles at each end.
The helical domain contains sequences that bind heparin in
vitro (Mok et al., 1997), and the capacity of APP to bind
collagen and laminin also depends on sequences contained in
this domain (Beher et al., 1996; Narindrasorasak et al., 1992).
Binding of the morphoregulatory protein F-spondin to the
isolated helical domain is of special interest (Ho and Sudhof,
2004). With recent access to the crystallographic structure of
this domain, it may be expected that binding of these ligands
will soon be explored structurally. However, so far, there can be
no certainty as to whether the isolated highly elongated helical
domain behaves in the same way as within full-length APP or
sAPP. It is important to note that both heparin, as a substitute for
the side chains of proteoglycans such as glypican that might be
expected to constitute the real physiological ligands of APP
(Williamson et al., 1996), and laminin are large, extended
molecules and may, therefore, simultaneously interact with
more than one region of APP. In this regard, it is noteworthy
that binding of F-spondin inhibits cleavage of transmembrane
APP by b-secretase, and possibly by a-secretase as well (Ho
and Sudhof, 2004), which implies an interaction of the helical
domain with other regions of APP. Furthermore, the central
domain of APP contains an N-glycosylation site (Pahlsson and
Spitalnik, 1996), the effect of which could not be investigated in
the recombinant isolated domain.
All structural prediction algorithms employed so far indicate
that the region of extracellular APP nearest to the membrane
(indicated as RC in Fig. 1) does not possess standard secondary

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3220
structure (Gralle et al., 2002; Sandbrink et al., 1994). As
mentioned above, this segment of the protein is specific for
vertebrates and is easily degraded by proteases in vitro. No
physiological ligands of this region of APP have been
characterized. However, as the b- and a-secretase cleavage
sites are localized within this sequence, there must necessarily
be contact between it and the secretases. Indeed, the structural
coupling of APP and b-secretase has been investigated through
mutations in this region that influence cleavage of APP by b-
secretase (Citron et al., 1995; Qahwash et al., 2004). It is
possible that this natively unstructured (Wright and Dyson,
1999) region of APP acquires secondary and/or tertiary
structure in the presence of the secretases or of other
macromolecules.
5.5. Interactions between APP domains
In Section 2, several functions attributed to APP in nervous
tissue were discussed. Their underlying molecular bases are the
interactions that APP establishes with the various types of
ligands described in the preceding sections. An important
question that arises, then, is what are the mechanisms that
connect the biochemical binding interactions with the functions
of APP at the cell and tissue levels.
Homophilic interactions between APP and its homologues
have recently been shown to promote cell–cell adhesion (Soba
et al., 2005). This clearly indicates that APP functions as a cell
adhesion molecule. On the other hand, as described above, the
extracellular portion of APP binds to many components of the
extracellular matrix. APP binding to, e.g., heparan sulfate
moieties might be involved in its cell-adhesive function (see
below), and also in interactions with the extracellular matrix,
which would make APP a member of the family of substrate
adhesion molecules that are equally important for brain
development and plasticity (Edelman and Cunningham,
1990). Still, while for some adhesion proteins (e.g., NCAM)
signaling pathways beginning with interation with an extra-
cellular ligand and ending in specific intracellular effects have
been unraveled (Dityatev et al., 2004), in the case of APP there
is as yet no clear connection between its binding to specific
extracellular ligands and subsequent intracellular events.
At first glance, the large number of possible biological
ligands of APP may appear somewhat puzzling, as each ligand
is expected to induce a structural transition of APP that leads to
a given biological effect. However, even without knowledge of
the complete three-dimensional structure of transmembrane
APP or full-length sAPP, it was possible to show that some of
those interactions are not independent: For example, binding of
Zn2+ increases the affinity of APP for heparin in vitro
(Multhaup et al., 1994) and potentiates the inhibitory activity
of APP in the coagulation cascade, which also depends on
heparan sulfate (Van Nostrand, 1995). The reduction of Cu2+ by
APP, when combined with the binding of APP to proteoglycan
heparan sulfate chains, causes controled and regulated
degradation of these proteoglycans (Cappai et al., 2005).
These observations indicate spatial proximity between the
copper reduction site, where the free radicals are produced, and
the heparan sulfate binding site(s), where the free radicals act
on the proteoglycan chains. As concerns the cell biology of
APP, on the other hand, it becomes clear that the multiple in
vitro activities of APP, which had been originally regarded as
independent from one another, may, in live cells, take part in
common biological events.
These examples show that the many binding partners of APP
may not act in isolation from one another. Instead, each piece of
information obtained on the structure of full-length APP or
sAPP will further our understanding of the relations between
the many binding partners of APP and of how binding of one
ligand in a given domain may impact interactions that take
place in other APP domains. The glycosylation of APP provides
one further example of such interdomain interactions. During
its passage through the Golgi complex, APP is N-glycosylated,
O-glycosylated, tyrosine sulfated and phosphorylated on
several residues on both sides of the membrane (for a review,
see Turner et al., 2003). While in an ovary-derived cell line no
role of APP glycosylation could be demonstrated (Pahlsson and
Spitalnik, 1996), N- and O-glycosylations of the extracellular
portion of APP are a precondition for phosphorylation of a
threonine residue in its cytoplasmic domain during neuronal
differentiation (Ando et al., 1999). This is a good example of
how human neurons use one type of post-translational
modification to regulate another type of post-translational
modification. This regulatory mechanism also indicates the
existence of a still unknown mechanism to transfer information
from the extracellular to the cytoplasmic domains of APP.
Phosphorylation of Thr668 (in APP695 numbering), in turn,
seems to be a signal for APP to be transported to the axon and to
participate in neuronal differentiation (Ando et al., 1999). At
the same time, Thr668 phosphorylation inhibits the stabiliza-
tion of the cytoplasmic domain of APP by Fe65 and, therefore,
abruptly decreases the nuclear concentration of the cytoplasmic
domain in embryonic neurons, possibly terminating a transient
signaling mediated by this domain (Kimberly et al., 2005).
Homo-oligomerization of APP has been reported (Chen
et al., 2006a; Hashimoto et al., 2003; Lu et al., 2003;
Scheuermann et al., 2001; Soba et al., 2005). Dimerization (or
monomerization) of transmembrane APP might be part of a
mechanism to transfer information about ligand binding or
environmental changes from one side of the membrane to the
other. Regarding the pathogenesis of Alzheimer’s disease, it is
especially interesting that Ab binds to the most N-terminal
domain of APP (Van Nostrand et al., 2002) and that Ab binding
may cause dimerization of transmembrane APP (Lu et al.,
2003) and neurotoxicity (Lorenzo et al., 2000). From a
structural point of view, it would be important to clarify how a
binding event involving the N-terminal domain translates into
an approximation of the transmembrane domains of APP
(Scheuermann et al., 2001).
Based on the arguments presented above, it is clear that
while a structure for full-length transmembrane APP does not
become available, a major current goal is obtaining high-
resolution structural information on the full-length extracellular
domain of the protein. This would provide insight into the
relations between different domains within sAPP as well as into
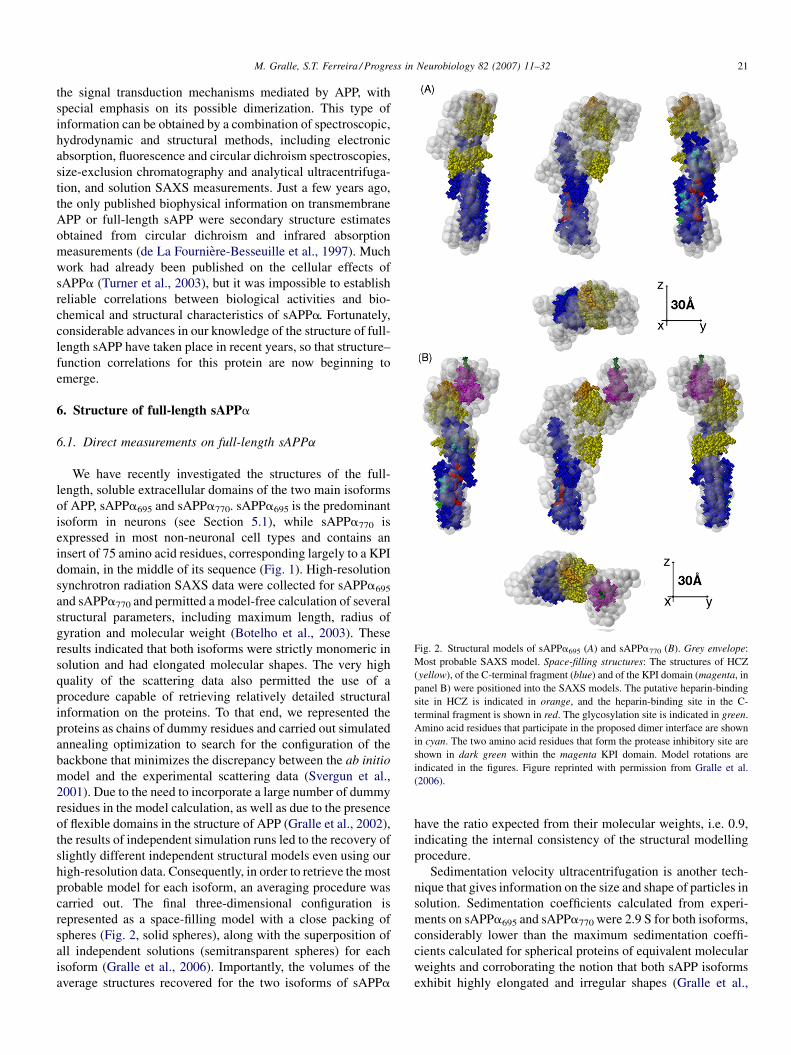
Fig. 2. Structural models of sAPPa695 (A) and sAPPa770 (B). Grey envelope:
Most probable SAXS model. Space-filling structures: The structures of HCZ
(yellow), of the C-terminal fragment (blue) and of the KPI domain (magenta, in
panel B) were positioned into the SAXS models. The putative heparin-binding
site in HCZ is indicated in orange, and the heparin-binding site in the C-
terminal fragment is shown in red. The glycosylation site is indicated in green.
Amino acid residues that participate in the proposed dimer interface are shown
in cyan. The two amino acid residues that form the protease inhibitory site are
shown in dark green within the magenta KPI domain. Model rotations are
indicated in the figures. Figure reprinted with permission from Gralle et al.
(2006).
M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 21
the signal transduction mechanisms mediated by APP, with
special emphasis on its possible dimerization. This type of
information can be obtained by a combination of spectroscopic,
hydrodynamic and structural methods, including electronic
absorption, fluorescence and circular dichroism spectroscopies,
size-exclusion chromatography and analytical ultracentrifuga-
tion, and solution SAXS measurements. Just a few years ago,
the only published biophysical information on transmembrane
APP or full-length sAPP were secondary structure estimates
obtained from circular dichroism and infrared absorption
measurements (de La Fourniere-Besseuille et al., 1997). Much
work had already been published on the cellular effects of
sAPPa (Turner et al., 2003), but it was impossible to establish
reliable correlations between biological activities and bio-
chemical and structural characteristics of sAPPa. Fortunately,
considerable advances in our knowledge of the structure of full-
length sAPP have taken place in recent years, so that structure–
function correlations for this protein are now beginning to
emerge.
6. Structure of full-length sAPPa
6.1. Direct measurements on full-length sAPPa
We have recently investigated the structures of the full-
length, soluble extracellular domains of the two main isoforms
of APP, sAPPa695 and sAPPa770. sAPPa695 is the predominant
isoform in neurons (see Section 5.1), while sAPPa770 is
expressed in most non-neuronal cell types and contains an
insert of 75 amino acid residues, corresponding largely to a KPI
domain, in the middle of its sequence (Fig. 1). High-resolution
synchrotron radiation SAXS data were collected for sAPPa695
and sAPPa770 and permitted a model-free calculation of several
structural parameters, including maximum length, radius of
gyration and molecular weight (Botelho et al., 2003). These
results indicated that both isoforms were strictly monomeric in
solution and had elongated molecular shapes. The very high
quality of the scattering data also permitted the use of a
procedure capable of retrieving relatively detailed structural
information on the proteins. To that end, we represented the
proteins as chains of dummy residues and carried out simulated
annealing optimization to search for the configuration of the
backbone that minimizes the discrepancy between the ab initio
model and the experimental scattering data (Svergun et al.,
2001). Due to the need to incorporate a large number of dummy
residues in the model calculation, as well as due to the presence
of flexible domains in the structure of APP (Gralle et al., 2002),
the results of independent simulation runs led to the recovery of
slightly different independent structural models even using our
high-resolution data. Consequently, in order to retrieve the most
probable model for each isoform, an averaging procedure was
carried out. The final three-dimensional configuration is
represented as a space-filling model with a close packing of
spheres (Fig. 2, solid spheres), along with the superposition of
all independent solutions (semitransparent spheres) for each
isoform (Gralle et al., 2006). Importantly, the volumes of the
average structures recovered for the two isoforms of sAPPa
have the ratio expected from their molecular weights, i.e. 0.9,
indicating the internal consistency of the structural modelling
procedure.
Sedimentation velocity ultracentrifugation is another tech-
nique that gives information on the size and shape of particles in
solution. Sedimentation coefficients calculated from experi-
ments on sAPPa695 and sAPPa770 were 2.9 S for both isoforms,
considerably lower than the maximum sedimentation coeffi-
cients calculated for spherical proteins of equivalent molecular
weights and corroborating the notion that both sAPP isoforms
exhibit highly elongated and irregular shapes (Gralle et al.,

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3222
2006). In fact, the sedimentation coefficients measured by
ultracentrifugation were even somewhat lower than the values
calculated from the SAXS models (3.77 S and 3.93 S for
sAPPa695 and sAPPa770, respectively). Consistent with our
previous observations (Botelho et al., 2003; Gralle et al., 2002),
this indicates that both sAPPa isoforms exhibit considerable
conformational flexibility. Thus, the filtered structural models
shown in Fig. 2 (dark spheres) should be considered time and
ensemble averages over the actual distributions of possible
conformations of sAPPa695 and sAPPa770 in solution.
Interestingly, the longer isoform, sAPPa770, contains an
additional structural domain that emerges laterally from one of
the ends of the molecule (Fig. 2B). That domain likely
corresponds to the KPI domain, which is present only in the
longer isoforms of APP (Kitaguchi et al., 1988), in addition to
other linker regions of the protein. The model thus indicates
that the KPI domain is freely exposed on the surface of APP,
facilitating its interaction with target proteases. The surface-
exposed location of the KPI domain is also in agreement with
the fact that its replacement by a yellow fluorescent protein
insert gives rise to a correctly localized APP chimera (Ehehalt
et al., 2003).
6.2. Topology and positioning of APP fragments
As described in Section 5, all the evolutionarily conserved
domains of APP have had their structures solved in isolation,
but it has not yet been possible to obtain suitable crystals or
perform NMR experiments that would solve the structure of
full-length sAPPa. In an attempt to overcome these difficulties,
in addition to the ab initio models of full-length sAPPa, we
performed SAXS measurements on the N-terminal fragment
that binds heparin, copper and zinc (see above; henceforth
denoted as HCZ) and showed that this fragment can best be
modelled as two rigid domains connected by a flexible linker
(Gralle et al., 2006). The available published data on high-
resolution structures of isolated sAPP fragments were then used
to establish topological correlations in the context of ab initio
SAXS models, based on a combination of (a) biochemical data
on the interaction of APP with known ligands, (b) a comparison
between the dimensional and conformational data available for
the crystallographic structures of isolated domains, (c) SAXS
models of the isolated HCZ domain and of the full-length
sAPPa isoforms, and (d) rigid body domain rotations and
translations guided by SAXS data.
The structures of HCZ and of the C-terminal domain (Wang
and Ha, 2004) were optimally positioned into the sAPPa695
model (Fig. 2A). Next, a similar procedure of rigid body
optimization was applied to position the KPI domain into the
salient protuberance of the sAPPa770 model, fixing the
positions of the other two domains as in the case sAPPa695
(Fig. 2B). The fits were found to be quite good in the angular
range conveying information on shape and quaternary
structure, though the existence of large structural gaps in the
models (i.e., regions of APP without defined structural
information) led to some deviation, notably at higher angles
(Gralle et al., 2006).
Positioning the high-resolution structures of individual
domains into the overall structure of sAPPa allowed the
localization of several functionally important sites of the protein.
For example, the sole N-glycosylation site contained within the
crystallized fragments is shown in green in Fig. 2. As expected, it
is fully exposed to the solvent in both isoforms, and its position
provides insight into the location of the glycan side chains in fully
glycosylated human APP. Two proposed heparan sulfate-binding
sites of APP (Rossjohn et al., 1999; Wang and Ha, 2004) are
shown in orange and red. The N-terminal-binding site (orange) is
localized in a groove between the N-terminal and C-terminal
fragments and may participate, along with nearby basic amino
acid residues on the C-terminal fragment, in heparin binding. On
the other hand, the C-terminal heparin-binding site (in red) is
fully exposed to the solvent in both isoforms. Interestingly, it has
been proposed that the isolated C-terminal domain forms dimers.
The amino acid residues participating in the suggested
hydrophobic dimerization interface (Wang and Ha, 2004) are
shown in cyan in Fig. 2. However, all the hydrodynamic data we
collected using analytical ultracentifugation, size-exclusion
chromatography and SAXS suggest that both sAPPa isoforms
are strictly monomeric in solution, in line with the fact that the
models indicate that part of the putative dimerization interface is
occluded in full-length sAPPa. This may be best appreciated in
the central panel of Fig. 2 for each isoform.
All the above-mentioned sites are present in the major
neuronal isoform, sAPPa695. An additional site of interest in the
structure of sAPPa770 is the protease inhibitory site in the KPI
domain (dark green in Fig. 2B), which corresponds to the
scissile bond in protease substrates and is expected to interact
most strongly with the proteases that are inhibited by sAPPa770
in vivo (Hynes et al., 1990). The structure shows that this
inhibitory site is fully exposed at one end of the sAPPa770
structure and, therefore, is poised for insertion at the active
center of a target protease.
7. Dimerization of sAPPa and APP
While it has recently been shown that a small fraction of
transmembrane APP may dimerize in intact cells (Soba et al.,
2005), the results described above indicate beyond doubt that the
extracellular domain of APP is monomeric in solution (Gralle
et al., 2002, 2006). Therefore, it is important to examine in more
depth the circumstances under which sAPP and APP might
dimerize and the possible consequences of this for the functions
of APP. Thewell-known binding of APP to heparan sulfate and of
APP-derived peptides to heparin (see Section 5.3) prompted us to
investigate the interaction of full-length sAPPa with heparin.
Combined size-exclusion chromatography, fluorescence spec-
troscopy, analytical ultracentrifugation and SAXS data sug-
gested that high-molecular-weight heparin chains induce the
formation of sAPP–heparin complexes at a sAPPa:heparin ratio
of 2:1 (Gralle et al., 2006). Structural models were also
calculated for heparin–APP dimers. From ab initio calculations
(imposing a two-fold symmetry axis), we obtained information
on the dimer shape. Several independent calculations of the
dimer model suggested that two configurations were possible,

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 23
with the two-fold axis parallel or perpendicular to the longer
molecular dimension. In order to investigate these possible
conformations, we used the structures of sAPPa695 monomers
previously calculated to generate the dimers and fit the models to
the experimental curve using rigid body modelling procedures to
obtain the best solutions. Parallel and anti-parallel dimer
configurations, both with approximately the same fit qualities,
were obtained. These results allowed the proposal of schematic
representations for the possible configurations of the heparin
(heparan sulfate)–APP complex at the cell membrane (Fig. 3A
and B; see below). The anti-parallel dimers connecting the
cellular membranes of neighboring cells may correspond to the
dimers discovered by cell aggregation assays, while co-
immunoprecipitation assays may have shown the presence of
both cis and trans dimers (Soba et al., 2005). Only biophysical
experiments such as those described above are capable of
indicating the presence of non-protein biomolecules, such as
heparan sulfate, in APP complexes.
Fig. 3. Alternative hypotheses for the conformation of transmembrane APP695 dimer
the sAPPa695–heparin complex, show the relations of scale between sAPPa695 m
intracellular and transmembrane domains of APP (yellow). As the SAXS data on the
sAPPa isoforms, the conformational changes that probably occur within each APP m
conformation of sAPPa and of heparin within the complex. Panel A: Parallel orient
molecular-weight heparin/heparin sulfate chain. The juxtaposition of the two trans
example by tyrosine phosphorylation of the cytoplasmic domain of APP. Note that, d
visible (yellow). Panel B: Anti-parallel orientation of two APP monomers present i
molecules, joined by a high-molecular-weight heparin/heparin sulfate chain, migh
reprinted with permission from Gralle et al. (2006).
8. Functional implications from structural studies with
sAPPa
Previous studies have tried to localize certain biological
activities of APP to smaller fragments of the protein (Jin et al.,
1994; Morimoto et al., 1998). However, as discussed in Section
4, proteins often function as coherent ‘‘machines’’, and their
workings cannot be completely understood by considering only
the activities of their constituent parts separately. The
importance of the recent structural results for full-length
sAPPa lies in that they allow the formulation of hypotheses on
the topological relations between the different domains of
sAPPa and APP and on the functions of these domains in the
context of the full-length protein.
The different types of receptors localized at the plasma
membrane can be classified as ion channel-coupled receptors,
G protein-coupled receptors, receptors with cytoplasmic
enzymatic activity (kinases, phosphatases and cyclases),
s. Both hypothetical conformations, derived from modeling of the SAXS data on
onomers (grey and red), heparin (green), the cellular membrane(s), and the
sAPPa695–heparin complex were of lower resolution than those on the isolated
onomer upon dimerization cannot be modeled. These changes may modify the
ation of two APP monomers in the same cellular membrane, joined by a high-
membrane and cytoplasmic domains might trigger intracellular signaling, for
ue to the tilt of the model, the cytoplasmic domain of only one APP monomer is
n the plasma membranes of different cells. The bridge formed by the two APP
t contribute to cell adhesion events as well as intercellular signaling. Figure

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3224
receptors associated to cytoplasmic enzymes (tyrosine kinases)
and receptors that signal by regulated intramembrane
proteolysis (Alberts et al., 2002). Transmembrane APP does
not seem to be directly coupled to channels, and no enzymatic
activity of its cytoplasmic domain is known either. Thus, in
what other ways might APP participate in signal transduction
pathways?
Regulated intramembrane proteolysis is the most recently
discovered basic mechanism of signal transduction, and the best
studied example is the proteolysis of the Notch receptor by the
g-secretase complex (De Strooper and Annaert, 2000). Results
obtained in immortalized cell lines indicated that cleavage of
APP by g-secretase releases a cytoplasmic domain (known as
AICD) that may regulate gene transcription in the nucleus (Cao
and Sudhof, 2001; Pardossi-Piquard et al., 2005). While these
are interesting observations, another recent study suggests that
the presence of AICD may be restricted to a short period in
neuronal development (Kimberly et al., 2005). Moreover, other
recent studies in mitotic cells have denied the importance of
APP processing by g-secretase for gene regulation (Cao and
Sudhof, 2004; Hass and Yankner, 2005; Hebert et al., 2006).
Indeed, expression of all target genes reported to be affected by
AICD signaling was shown to be scarcely affected by over- or
underexpression of APP, the absence of presenilin or the
inhibition of g-secretase (Hebert et al., 2006). Whether or not
AICD has a signal transduction function in vivo, its production
has been shown not to be affected by cross-linking of APP
(Kimberly et al., 2005) and, therefore, is probably not
physiologically regulated by dimerization of APP.
A marked feature of the cytoplasmic domain of APP is the
presence of three tyrosine residues that can be partially
phosphorylated (Russo et al., 2001). Therefore, it is possible
that APP is part of the family of tyrosine kinase-associated
receptors (Alberts et al., 2002). These receptors do not have an
enzymatic activity of their own in their cytoplasmic domain.
Instead, they contain a domain which, after activation of the
receptor, becomes capable of binding an intracellular tyrosine
kinase. The interaction between APP and the tyrosine kinase Abl
was proposed based on models of neural development and was
then experimentally demonstrated: activated Abl phosphorylates
the most C-terminal tyrosine residue of APP (Zambrano et al.,
2001). It is also clear that the intracellular domain of APP can
only bind to the intracellular signaling protein Shc when
phosphorylated on this tyrosine residue (Russo et al., 2002).
Recent results suggest that APP dimerization leading to signaling
via tyrosine phosphorylation may be typical of immune cells but
not of neurons (Sondag and Combs, 2006).
The recent report that APP causes the phosphorylation of the
transcription factor Tip60 in a membrane-bound complex that
also includes Fe65 and cyclin-dependent kinase (Hass and
Yankner, 2005) suggests a similar mode of action, where APP,
though not enzymatically active itself, facilitates post-transla-
tional modifications on associated proteins. The stimuli and
mechanism for APP signaling have not yet been described for
this system.
Finally, APP may signal by activation of G0, the most
abundant G protein in neurons. APP has been shown to bind and
modulate G0 activity in vitro (Nishimoto et al., 1993; Okamoto
et al., 1995; Brouillet et al., 1999). Moreover, in insects,
physiological G0 signaling has been suggested to involve an
APP homologue (Swanson et al., 2005). While the majority of
G protein-coupled receptors are seven-transmembrane helix
proteins, APP is not the only single transmembrane domain
protein coupled to G proteins, as there are several other such
unconventional G protein-coupled receptors (Patel, 2004). The
mechanism used by APP to transmit an extracellular signal to
the protein G0 has not yet been completely elucidated, but there
are reasons to believe it might involve dimerization of APP
(Brouillet et al., 1999; Hashimoto et al., 2003), similar to other
G protein-coupled receptors regulated by dimerization (Her-
rick-Davis et al., 2005). In this case, the results mentioned in
Section 7 raise the question whether binding of heparan sulfate
(present in the extracellular matrix or on the surface of a cell) to
APP might induce APP dimerization and therefore influence
the activation of G0. The proposed consequences of APP-
mediated G0 activation vary strongly according to the cellular
model used, and include Ca2+-mediated reduction of motility in
migrating enteric neurons in vivo (Swanson et al., 2005), gene
transcription changes and later apoptotic changes in COS-NK1
cells (Ikezu et al., 1996; Yamatsuji et al., 1996a), and apoptotic
cell death in immortalized neurohybrid cells (Yamatsuji et al.,
1996b; Hashimoto et al., 2003).
9. Future perspectives
9.1. Possible questions to be addressed
From the perspective of establishing the signaling path-
way(s) in which it is involved, it would now be quite useful to
express full-length APP harboring mutations at strategic
residues and to investigate the cellular effects of such APP
mutants, similar to what is being done for the paradigmatic
neuronal adhesion molecule, NCAM (Dityatev et al., 2004).
Indeed, part of an intracellular pathway triggered by
dimerization of APP has already been teased apart in
immortalized cells, but the exact mechanism of dimerization
or the physiological ligand of APP that induces dimerization
has not been determined (Hashimoto et al., 2003). Now, based
on our recent data, heparin (as a surrogate for naturally
occurring heparin sulfate glycosaminoglycans) could be used
as model physiological extracellular ligand for such studies.
The availability of the structure of a large fragment of sAPPa
(Wang and Ha, 2004) and the known topological relations
between the fragments of sAPPa (Gralle et al., 2006) should
allow a better analysis of the amino acid residues possibly
involved in dimerization.
In the blood coagulation cascade (Zhang et al., 1997) and in
other tissues, especially in the brain, the ligand glycosami-
noglycan is probably present as part of a proteoglycan of the
heparan sulfate family. APP might suffer dimerization by the
action of heparan sulfate moieties present in the extracellular
matrix or associated to the membrane of neighboring cells. It is
quite probable that there are other ligands, beyond heparin or
heparin sulfate, which influence the dimerization of sAPPa and

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 25
APP, notably Cu2+, collagen and Ab (Beher et al., 1996; Hesse
et al., 1994; Lu et al., 2003; Scheuermann et al., 2001). Maybe
the inhibition of the production of Ab by the binding of F-
spondin to APP (Ho and Sudhof, 2004) is also mediated by
dimerization of the latter. If dimerization of transmembrane
APP by any given ligand can be demonstrated, the subcellular
localization of this interaction must be determined, if possible
in neurons. A strong candidate localization is the growth cone
(Sabo et al., 2003).
9.2. APP and cholinergic transmission
The best available model for the role of APP (and its
homologue APLP2) in synaptogenesis is the formation of
cholinergic synapses at the neuromuscular junction (Wang
et al., 2005) and in the submandibular ganglion (Yang et al.,
2005). Today, in addition to the general roles of cell adhesion
molecules in synaptogenesis, the specific roles of different cell
adhesion molecules in the formation of different types of
synapses are being investigated (Washbourne et al., 2004). It
has been shown that APP is important for the formation of
distinct subtypes of synapses (Salbaum and Ruddle, 1994). An
interesting hypothesis would be a specific role of APP/APLP2
in the formation and/or maintenance of cholinergic synapses in
the central nervous system as well, as dysfunctions of the
cholinergic system have been implied in Alzheimer’s disease.
In this case, APP signaling might be mediated by members of
the thrombospondin family, recently discovered synaptogenesis
inductors, which bind to some ligands known to be involved in
APP metabolism, such as LRP and heparan sulfate (Wash-
bourne et al., 2004).
Lack of NGF induces degeneration of cholinergic forebrain
neurons, which is typically observed at early stages of
Alzheimer’s disease. Indeed, one transgenic mouse model
for the disease produces antibodies against NGF exclusively in
adult neurons, which leads to depletion of NGF during aging
and to many neuropathological and behavioral symptoms of
Alzheimer’s disease (Capsoni et al., 2002). If APP is indeed
important for the establishment and maintenance of cholinergic
synapses (Wang et al., 2005; Yang et al., 2005), then
overproduction of APP might be a compensatory mechanism
to re-establish cholinergic transmission (Isacson et al., 2002).
The overproduction and deposition of Ab in neuritic plaques
observed in these mice could, thus, be secondary toxic effects
arising from the overproduction of APP; it remains to be seen if
such a mechanism is operative in AD, too.
9.3. Implications for APLP2
All of the structural work described above was designed to
investigate the structure and function of APP, one of three very
similar and functionally redundant proteins present in humans
(Heber et al., 2000). Although there are no data on the structure
of APLP2, it is quite probable that the structural model of
sAPPa obtained from SAXS measurements is also largely
applicable to the extracellular domain of APLP2, the nearest
homologue of APP (APLP1 is a smaller protein). The cellular
metabolism of APLP2 follows the same general lines as that of
APP: APLP2 is transported from the endoplasmic reticulum to
the plasma membrane or, in neurons, to the synaptic membrane
(Lyckman et al., 1998), and can be cleaved by the same
secretases that cleave APP (Walsh et al., 2003; Eggert et al.,
2004). Only double knock-out mice for APP and APLP2 have a
phenotype of synaptic dysfunctions, while each of the single
knock-outs has an essentially normal phenotype (Wang et al.,
2005; Yang et al., 2005). The only clear functional difference
between APP and APLP2 is the impossibility to generate Ab
from APLP2, and APP single knock-out mice have some subtle
neurological defects (Dawson et al., 1999; Zheng et al., 1995).
The high-homology regions conserved between the two
proteins are the same regions that have already been crystal-
lized and which we localized within our models of full-length
sAPPa (Fig. 2). Taking into account both the sequence
homology and the redundancy of function, it is probable that the
extracellular domain of APLP2 is also monomeric in isolation.
Furthermore, the possibility of formation of homo- and
heterodimers of APP, APLP1 and APLP2 (Soba et al., 2005)
suggests that APLP2 may also dimerize in the presence of
heparin or other biomolecules.
9.4. Unexpected functions of APLP2 and APP
Because of the high structural similarity and functional
redundance between APP and APLP2, it would be surprising if
APLP2 had functions fundamentally different from those
described above for APP. However, in addition to those
functions of APP described in Section 2, which agree perfectly
with those expected for a protein inserted in the plasma
membrane, and which seem to be largely common to both APP
and APLP2, other functions for the two proteins have been
described that have not yet been integrated into this general
view.
For example, it has been reported that, in mice, APLP2
interacts with the protein Kd, which has the function of
facilitating peptide presentation by the histocompatibility
complex MHC2 (Rassoulzadegan et al., 1998). This interaction
was reported to begin in the endoplasmic reticulum and the
Golgi complex, and the APLP2–Kd complex would be
transported to the cell surface. It is clear that the majority of
both APLP2 and APP is localized in the endoplasmic reticulum
and the Golgi complex (Wasco et al., 1993). The usual
explanation for this fact is the short half-life of APP (and
possibly APLP2) at the cell surface (Koo and Squazzo, 1994).
If, on the other hand, APLP2 interacts with Kd well before it
arrives at the cell surface, the possibility cannot be discarded
that APP, too, may have functionally important interactions that
take place in the initial compartments of the secretory pathway.
For example, part of the production of Ab42 from APP occurs in
the endoplasmic reticulum, which suggests an interaction
between APP and b- and g-secretase activities already in this
compartment (Wilson et al., 2002).
It has also been reported that APLP2 interacts with a specific
DNA sequence and localizes to specific spots in the nucleus
during interphase (Blangy et al., 1995). These observations

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3226
imply that APLP2 might be imported into the nucleus instead of
being inserted into the endoplasmic reticulum, as the majority
of reports suggest. Which structural elements of APLP2 would
permit its nuclear import and whether APP could also be
imported into the nucleus are questions that have still not been
addressed. The conclusions obtained from microinjection
experiments indicating that APLP2 plays an essential role in
the first cellular divisions of the embryo (Blangy et al., 1995),
because it is strictly necessary for chromatide segregation
(Sester et al., 2000), are in conflict with transgenic animal
results which show largely redundant functions for APP,
APLP1 and APLP2 (Heber et al., 2000; Herms et al., 2004;
Wang et al., 2005; Yang et al., 2005). There may be technical
reasons for this difference between the different transgenic
models, but the results on nuclear localization of APLP2 are not
disqualified by this conflict.
Surprisingly, recent reports suggest that APP can be inserted
into the inner mitochondrial membrane import channels
because of an ambiguous signaling sequence. High levels of
mitochondrial APP impair mitochondrial energy metabolism
and also appear to generate oxidative stress via enhanced
production of H2O2 (Anandatheerthavarada et al., 2003; Devi
et al., 2006). APP and APLP2 are relatively large proteins and
their intracellular trafficking in differentiated cells has only
been investigated successfully in the last few years. It is quite
possible that a cell may put APP and APLP2 to good use in
different functions, in the same way as it does so with other
proteins. As mentioned in Section 5.5, glycosylation of APP
allows the phosphorylation of a specific threonine residue in
neuron-derived cells, and this phosphorylation guides APP to
the axon (Ando et al., 1999). Other modifications can be
imagined which target APP or APLP2 to other cellular fates and
explain the great variety of functions described for the APP
family. The recent results discussed here may contribute to our
further understanding of the structural bases of those functions.
10. Conclusions
In the past 20 years, a vast amount of research has focused on
the structure and metabolism of Ab. By comparison, much less
attention has been given to investigating the structure and
metabolism of APP, and the complexity of the interactions
between APP and other biomolecules has only recently begun
to be illuminated. This review has focused on the role of APP as
an adhesion protein/contact receptor, without excluding other
possible functions. At this stage of APP research, there is a
cogent need to combine the great number of single observations
on cellular aspects of the biology of APP into more
comprehensive pictures while avoiding oversimplification or
one-sidedness. We have approached this from two different
directions: (1) considering the functions of APP in the intact
nervous tissue as the result of complex interactions between
cells and (2) analyzing the structure of the intact protein as the
result of complex interactions between domains. It is hoped that
a more coherent picture will emerge which may help
researchers in designing relevant experimental approaches
for investigating the roles of APP in the healthy and in the
demented brain.
Acknowledgements
We wish to thank Luiz Henrique Guerreiro and Drs.
Michelle Gralle Botelho, Cristiano L.P. de Oliveira, Iris
Torriani and Carlos H.I. Ramos who contributed to the
experimental work on sAPP described in this review. During
the preparation of this review, M.G. was supported by a Bert
Sakmann Nobel Laureate Stipend from the Max Planck
Society. Research in S.T.F.’s laboratory was supported by grants
from Howard Hughes Medical Institute and by the Brazilian
agencies Conselho Nacional de Desenvolvimento Cientıfico e
Tecnologico (CNPq), Programa de Nucleos de Excelencia
(Pronex) and Fundacao de Amparo a Pesquisa do Estado do Rio
de Janeiro (FAPERJ). S.T.F. is a Howard Hughes Medical
Institute International Scholar.
References
Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., Walter, P., 2002. Cell
communication. Molecular Biology of the Cell, Garland Science, New
York, pp. 831–907.
Allinquant, B., Moya, K.L., Bouillot, C., Prochiantz, A., 1994. Amyloid
precursor protein in cortical neurons—coexistence of 2 pools differentially
distributed in axons and dendrites and association with cytoskeleton. J.
Neurosci. 14, 6842–6854.
Allinquant, B., Hantraye, P., Mailleux, P., Moya, K., Bouillot, C., Prochiantz,
A., 1995. Down-regulation of amyloid precursor protein inhibits neurite
outgrowth in vitro. J. Cell Biol. 128, 919–927.
Anandatheerthavarada, H.K., Biswas, G., Robin, M.A., Avadhani, N.G., 2003.
Mitochondrial targeting and a novel transmembrane arrest of Alzheimer’s
amyloid precursor protein impairs mitochondrial function in neuronal cells.
J. Cell Biol. 161, 41–54.
Ando, K., Oishi, M., Takeda, S., Iijima, K., Isohara, T., Nairn, A.C., Kirino, Y.,
Greengard, P., Suzuki, T., 1999. Role of phosphorylation of Alzheimer’s
amyloid precursor protein during neuronal differentiation. J. Neurosci. 19,
4421–4427.
Arai, R., Wriggers, W., Nishikawa, Y., Nagamune, T., Fujisawa, T., 2004.
Conformations of variably linked chimeric proteins evaluated by synchro-
tron X-ray small-angle scattering. Proteins 57, 829–838.
Araki, W., Kitaguchi, N., Tokushima, Y., Ishii, K., Aratake, H., Shimohama, S.,
Nakamura, S., Kimura, J., 1991. Trophic effect of beta-amyloid precursor
protein on cerebral cortical neurons in culture. Biochem. Biophys. Res.
Comm. 181, 265–271.
Arama, E., Agapite, J., Steller, H., 2003. Caspase activity and a specific
cytochrome c are required for sperm differentiation in Drosophila. Dev.
Cell 4, 687–697.
Arendt, T., Schindler, C., Bruckner, M.K., Eschrich, K., Bigl, V., Zedlick, D.,
Marcova, L., 1997. Plastic neuronal remodeling is impaired in patients with
Alzheimer’s disease carrying apolipoprotein epsilon 4 allele. J. Neurosci.
17, 516–529.
Arendt, T., Bruckner, M.K., Gertz, H.J., Marcova, L., 1998. Cortical distribution
of neurofibrillary tangles in Alzheimer’s disease matches the pattern of
neurons that retain their capacity of plastic remodelling in the adult brain.
Neuroscience 83, 991–1002.
Barnham, K.J., McKinstry, W.J., Multhaup, G., Galatis, D., Morton, C.J.,
Curtain, C.C., Williamson, N.A., White, A.R., Hinds, M.G., Norton,
R.S., Beyreuther, K., Masters, C.L., Parker, M.W., Cappai, R., 2003.
Structure of the Alzheimer’s disease amyloid precursor protein copper
binding domain—a regulator of neuronal copper homeostasis. J. Biol.
Chem. 278, 17401–17407.

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 27
Beher, D., Hesse, L., Masters, C.L., Multhaup, G., 1996. Regulation of amyloid
protein precursor (APP) binding to collagen and mapping of the binding
sites on APP and collagen type I. J. Biol. Chem. 271, 1613–1620.
Berezovska, O., Ramdya, P., Skoch, J., Wolfe, M.S., Bacskai, B.J., Hyman,
B.T., 2003. Amyloid precursor protein associates with a nicastrin-dependent
docking site on the presenilin 1-gamma-secretase complex in cells demon-
strated by fluorescence lifetime imaging. J. Neurosci. 23, 4560–4566.
Blangy, A., Vidal, F., Cuzin, F., Yang, Y.H., Boulukos, K.E., Rassoulzadegan,
M., 1995. CDEBP, s site-specific DNA-binding protein of the ‘APP-like’
family, is required during the early development of the mouse. J. Cell Sci.
108, 675–683.
Blasko, I., Beer, R., Bigl, M., Apelt, J., Franz, G., Rudzki, D., Ransmayr, G.,
Kampfl, A., Schliebs, R., 2004. Experimental traumatic brain injury in rats
stimulates the expression, production and activity of Alzheimer’s disease
beta-secretase (BACE-1). J. Neural Transm. 111, 523–536.
Borg, J.P., Ooi, J., Levy, E., Margolis, B., 1996. The phosphotyrosine interaction
domains of X11 and FE65 bind to distinct sites on the YENPTY motif of
amyloid precursor protein. Mol. Cell. Biol. 16, 6229–6241.
Botelho, M.G., Gralle, M., Oliveira, C.L.P., Torriani, I., Ferreira, S.T., 2003.
Folding and stability of the extracellular domain of the human amyloid
precursor protein. J. Biol. Chem. 278, 34259–34267.
Breen, K.C., Bruce, M., Anderton, B.H., 1991. Beta-amyloid precursor protein
mediates neuronal cell–cell and cell–surface adhesion. J. Neurosci. Res. 28,
90–100.
Brouillet, E., Trembleau, A., Galanaud, D., Volovitch, M., Bouillot, C., Valenza,
C., Prochiantz, A., Allinquant, B., 1999. The amyloid precursor protein
interacts with G(o) heterotrimeric protein within a cell compartment
specialized in signal transduction. J. Neurosci. 19, 1717–1727.
Bush, A.I., Multhaup, G., Moir, R.D., Williamson, T.G., Small, D.H., Rumble,
B., Pollwein, P., Beyreuther, K., Masters, C.L., 1993. A novel zinc(II)
binding site modulates the function of the beta-A4 amyloid protein pre-
cursor of Alzheimer’s disease. J. Biol. Chem. 268, 16109–16112.
Caille, I., Allinquant, B., Dupont, E., Bouillot, C., Langer, A., Muller, U.,
Prochiantz, A., 2004. Soluble form of amyloid precursor protein regulates
proliferation of progenitors in the adult subventricular zone. Development
131, 2173–2181.
Campbell, D.S., Holt, C.E., 2003. Apoptotic pathway and MAPKs differentially
regulate chemotropic responses of retinal growth cones. Neuron 37, 939–
952.
Cao, X., Sudhof, T.C., 2001. A transcriptionally active complex of APP with
Fe65 and histone acetyltransferase Tip60. Science 293, 115–120.
Cao, X., Sudhof, T.C., 2004. Dissection of amyloid-beta precursor protein-
dependent transcriptional transactivation. J. Biol. Chem. 279, 24601–
24611.
Cappai, R., Cheng, F., Ciccotosto, G.D., Needham, B.E., Masters, C.L.,
Multhaup, G., Fransson, L.A., Mani, K., 2005. The amyloid precursor
protein (APP) of Alzheimer disease and its paralog, APLP2, modulate the
Cu/Zn-nitric oxide-catalyzed degradation of glypican-1 heparan sulfate in
vivo. J. Biol. Chem. 280(14), 13913–13920.
Capsoni, S., Giannotta, S., Cattaneo, A., 2002. Beta-amyloid plaques in a model
for sporadic Alzheimer’s disease based on transgenic anti-nerve growth
factor antibodies. Mol. Cell. Neurosci. 21, 15–28.
Chen, C.D., Oh, S.Y., Hinman, J.D., Abraham, C.R., 2006a. Visualization of
APP dimerization and APP-Notch2 heterodimerization in living cells using
bimolecular fluorescence complementation. J. Neurochem. 97, 30–43.
Chen, F., Hasegawa, H., Schmitt-Ulms, G., Kawarai, T., Bohm, C., Katayama,
T., Gu, Y., Sanjo, N., Glista, M., Rogaeva, E., Wakutani, Y., Pardossi-
Piquard, R., Ruan, X., Tandon, A., Checler, F., Marambaud, P., Hansen, K.,
Westaway, D., St George-Hyslop, P., Fraser, P., 2006b. TMP21 is a pre-
senilin complex component that modulates gamma-secretase but not epsi-
lon-secretase activity. Nature 440, 1208–1212.
Chow, N.W., Korenberg, J.R., Chen, X.N., Neve, R.L., 1996. APP-BP1, a novel
protein that binds to the carboxyl-terminal region of the amyloid precursor
protein. J. Biol. Chem. 271, 11339–11346.
Citron, M., Oltersdorf, T., Haass, C., McConlogue, L., Hung, A.Y., Seubert, P.,
Vigopelfrey, C., Lieberburg, I., Selkoe, D.J., 1992. Mutation of the beta-
amyloid precursor protein in familial Alzheimer’s disease increases beta-
protein production. Nature 360, 672–674.
Citron, M., Teplow, D.B., Selkoe, D.J., 1995. Generation of amyloid beta
protein from its precursor is sequence specific. Neuron 14, 661–670.
Ciuculescu, E.D., Mekmouche, Y., Faller, P., 2005. Metal-binding properties of
the peptide APP(170–188): a model of the Zn-II-binding site of amyloid
precursor protein (APP). Chemistry 11, 903–909.
Clarris, H.J., Key, B., Beyreuther, K., Masters, C.L., Small, D.H., 1995.
Expression of the amyloid protein precursor of Alzheimer’s disease in
the developing rat olfactory system. Dev. Brain Res. 88, 87–95.
Corder, E.H., Saunders, A.M., Risch, N.J., Strittmatter, W.J., Schmechel, D.E.,
Gaskell, P.C., Rimmler, J.B., Locke, P.A., Conneally, P.M., Schmader, K.E.,
Small, G.W., Roses, A.D., Haines, J.L., Pericak-Vance, M.A., 1994. Pro-
tective effect of apolipoprotein E type 2 allele for late-onset Alzheimer
disease. Nat. Genet. 7, 180–184.
Daigle, I., Li, C., 1993. apl-1, a Caenorhabditis elegans gene encoding a protein
related to the human beta-amyloid protein precursor. Proc. Natl. Acad. Sci.
U.S.A. 90, 12045–12049.
Dawson, G.R., Seabrook, G., Zheng, H., Smith, D.W., Graham, S., O’Dowd, G.,
Bowery, B.J., Boyce, S., Trumbauer, M.E., Chen, H.Y., Van der Ploeg, L.H.,
Sirinathsinghji, D.J., 1999. Age-related cognitive deficits, impaired long-
term potentiation and reduction in synaptic marker density in mice lacking
the beta-amyloid precursor protein. Neuroscience 90, 1–13.
de La Fourniere-Besseuille, L., Grange, D., Buchet, R., 1997. Purification and
spectroscopic characterization of beta-amyloid precursor protein from
porcine brains. Eur. J. Biochem. 250, 705–711.
De Strooper, B., Annaert, W., 2000. Proteolytic processing and cell biological
functions of the amyloid precursor protein. J. Cell Sci. 113, 1857–
1870.
De Strooper, B., Annaert, W., Cupers, B., Saftig, P., Craessaerts, K., Mumm,
J.S., Schroeter, E.H., Schrijvers, V., Wolfe, M.S., Ray, W.J., Goate, A.,
Kopan, R., 1999. A presenilin 1-dependent gamma-secretase-like protease
mediates release of Notch intracellular domain. Nature 398, 518–522.
DePristo, M.A., de Bakker, P.I., Blundell, T.L., 2004. Heterogeneity and
inaccuracy in protein structures solved by X-ray crystallography. Structure
12, 831–838.
Devi, L., Prabhu, B.M., Galati, D.F., Avadhani, N.G., Anandatheerthavarada,
H.K., 2006. Accumulation of amyloid precursor protein in the mitochon-
drial import channels of human Alzheimer’s disease brain is associated with
mitochondrial dysfunction. J. Neurosci. 26, 9057–9068.
Dityatev, A., Dityateva, G., Sytnyk, V., Delling, M., Toni, N., Nikonenko, I.,
Muller, D., Schachner, M., 2004. Polysialylated neural cell adhesion
molecule promotes remodeling and formation of hippocampal synapses.
J. Neurosci. 24, 9372–9382.
Dulubova, I., Ho, A., Huryeva, I., Sudhof, T.C., Rizo, J., 2004. Three-dimen-
sional structure of an independently folded extracellular domain of human
amyloid-beta precursor protein. Biochemistry 43, 9583–9588.
Dunaevsky, A., Tashiro, A., Majewska, A., Mason, C., Yuste, R., 1999.
Developmental regulation of spine motility in the mammalian central
nervous system. Proc. Natl. Acad. Sci. U.S.A. 96, 13438–13443.
Edbauer, D., Winkler, E., Regula, J.T., Pesold, B., Steiner, H., Haass, C.,
2003. Reconstitution of gamma-secretase activity. Nat. Cell Biol. 5, 486–
488.
Edelman, G.M., Cunningham, B.A., 1990. Place-dependent cell adhesion,
process retraction, and spatial signaling in neural morphogenesis. Cold
Spring Harb. Symp. Quant. Biol. 55, 303–318.
Eggert, S., Paliga, K., Soba, P., Evin, G., Masters, C.L., Weidemann, A.,
Beyreuther, K., 2004. The proteolytic processing of the amyloid precursor
protein gene family members APLP-1 and APLP-2 involves alpha-, beta-,
gamma-, and epsilon-like cleavages. Modulation of APLP-1 processing by
N-glycosylation. J. Biol. Chem. 279, 18146–18156.
Ehehalt, R., Keller, P., Haass, C., Thiele, C., Simons, K., 2003. Amyloidogenic
processing of the Alzheimer beta-amyloid precursor protein depends on
lipid rafts. J. Cell Biol. 160, 113–123.
Engert, F., Bonhoeffer, T., 1999. Dendritic spine changes associated with
hippocampal long-term synaptic plasticity. Nature 399, 66–70.
Esler, W.P., Kimberly, W.T., Ostaszewski, B.L., Diehl, T.S., Moore, C.L., Tsai,
J.Y., Rahmati, T., Xia, W.M., Selkoe, D.J., Wolfe, M.S., 2000. Transition-
state analogue inhibitors of gamma-secretase bind directly to presenilin-1.
Nat. Cell Biol. 2, 428–434.

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3228
Fedorov, A.N., Baldwin, T.O., 1999. Process of biosynthetic protein folding
determines the rapid formation of native structure. J. Mol. Biol. 294, 579–
586.
Ferreira, S.T., 1989. Fluorescence studies of the conformational dynamics of
parvalbumin in solution lifetime and rotational motions of the single
tryptophan residue. Biochemistry 28, 10066–10072.
Ferreira, S.T., Stella, L., Gratton, E., 1994. Conformational dynamics of bovine
Cu, Zn superoxide dismutase revealed by time-resolved fluorescence spec-
troscopy of the single tyrosine residue. Biophys. J. 66, 1185–1196.
Furukawa, K., Barger, S.W., Blalock, E.M., Mattson, M.P., 1996a. Activation of
K+ channels and suppression of neuronal activity by secreted beta-amyloid
precursor protein. Nature 379, 74–78.
Furukawa, K., Sopher, B.L., Rydel, R.E., Begley, J.G., Pham, D.G., Martin,
G.M., Fox, M., Mattson, M.P., 1996b. Increased activity-regulating and
neuroprotective efficacy of alpha-secretase-derived secreted amyloid pre-
cursor protein conferred by a C-terminal heparin-binding domain. J.
Neurochem. 67, 1882–1896.
Galvan, V., Gorostiza, O.F., Banwait, S., Ataie, M., Logvinova, A.V., Sitaraman,
S., Carlson, E., Sagi, S.A., Chevallier, N., Jin, K., Greenberg, D.A.,
Bredesen, D.E., 2006. Reversal of Alzheimer’s-like pathology and behavior
in human APP transgenic mice by mutation of Asp664. Proc. Natl. Acad.
Sci. U.S.A. 103, 7130–7135.
Gebbink, M.F., Zondag, G.C., Koningstein, G.M., Feiken, E., Wubbolts, R.W.,
Moolenaar, W.H., 1995. Cell surface expression of receptor protein tyrosine
phosphatase RPTP mu is regulated by cell–cell contact. J. Cell Biol. 131,
251–260.
Glenner, G.G., Wong, C.W., 1984. Alzheimer’s disease—initial report of the
purification and characterization of a novel cerebrovascular amyloid pro-
tein. Biochem. Biophys. Res. Comm. 120, 885–890.
Goate, A., Chartier-Harlin, M.C., Mullan, M., Brown, J., Crawford, F., Fidani,
L., Giuffra, L., Haynes, A., Irving, N., James, L., et al., 1991. Segregation of
a missense mutation in the amyloid precursor protein gene with familial
Alzheimer’s disease. Nature 349, 704–706.
Goda, Y., 2002. Cadherins communicate structural plasticity of presynaptic and
postsynaptic terminals. Neuron 35, 1–3.
Goldsbury, C., Mocanu, M.M., Thies, E., Kaether, C., Haass, C., Keller, P.,
Biernat, J., Mandelkow, E., Mandelkow, E.M., 2006. Inhibition of APP
trafficking by tau protein does not increase the generation of amyloid-beta
peptides. Traffic 7, 873–888.
Gong, Y.S., Chang, L., Viola, K.L., Lacor, P.N., Lambert, M.P., Finch, C.E.,
Krafft, G.A., Klein, W.L., 2003. Alzheimer’s disease-affected brain: pre-
sence of oligomeric A beta ligands (ADDLs) suggests a molecular basis for
reversible memory loss. Proc. Natl. Acad. Sci. U.S.A. 100, 10417–10422.
Goo, J.H., Park, W.J., 2004. Elucidation of the interactions between C99,
presenilin, and nicastrin by the split-ubiquitin assay. DNA Cell Biol. 23, 59–
65.
Gralle, M., Botelho, M.M., de Oliveira, C.L.P., Torriani, I., Ferreira, S.T., 2002.
Solution studies and structural model of the extracellular domain of the
human amyloid precursor protein. Biophys. J. 83, 3513–3524.
Gralle, M., Oliveira, C.L.P., Guerreiro, L.H., McKinstry, W.J., Galatis, D.,
Masters, C.L., Cappai, R., Parker, M.W., Ramos, H.I., Torriani, I., Ferreira,
S.T., 2006. Solution conformation and heparin-induced dimerization of the
full-length extracellular domain of the human amyloid precursor protein. J.
Mol. Biol. 357, 493–508.
Hashimoto, Y., Niikura, T., Chiba, T., Tsukamoto, E., Kadowaki, H., Nishitoh,
H., Yamagishi, Y., Ishizaka, M., Yamada, M., Nawa, M., Terashita, K., Aiso,
S., Ichijo, H., Nishimoto, I., 2003. The cytoplasmic domain of Alzheimer’s
amyloid-beta protein precursor causes sustained apoptosis signal-regulating
kinase 1/c-Jun NH2-terminal kinase-mediated neurotoxic signal via dimer-
ization. J. Pharmacol. Exp. Ther. 306, 889–902.
Hass, M.R., Yankner, B.A., 2005. A gamma-secretase-independent mechanism
of signal transduction by the amyloid precursor protein. J. Biol. Chem. 280,
36895–36904.
Heber, S., Herms, J., Gajic, V., Hainfellner, J., Aguzzi, A., Rulicke, T.,
Kretzschmar, H., von Koch, C., Sisodia, S., Tremml, P., Lipp, H.P., Wolfer,
D.P., Muller, U., 2000. Mice with combined gene knock-outs reveal
essential and partially redundant functions of amyloid precursor protein
family members. J. Neurosci. 20, 7951–7963.
Hebert, S.S., Serneels, L., Tolia, A., Craessaerts, K., Derks, C., Filippov, M.A.,
Muller, U., De Strooper, B., 2006. Regulated intramembrane proteolysis of
amyloid precursor protein and regulation of expression of putative target
genes. EMBO Rep. 7, 739–745.
Hedstrom, L., Szilagyi, L., Rutter, W.J., 1992. Converting trypsin to chymo-
trypsin—the role of surface loops. Science 255, 1249–1253.
Heidemann, S.R., 1996. Cytoplasmic mechanisms of axonal and dendritic
growth in neurons. Int. Rev. Cytol. 165, 235–296.
Herard, A.S., Besret, L., Dubois, A., Dauguet, J., Delzescaux, T., Hantraye, P.,
Bonvento, G., Moya, K.L., 2006. siRNA targeted against amyloid precursor
protein impairs synaptic activity in vivo. Neurobiol. Aging 27(12), 1740–
1750.
Herms, J., Anliker, B., Heber, S., Ring, S., Fuhrmann, M., Kretzschmar, H.,
Sisodia, S., Muller, U., 2004. Cortical dysplasia resembling human type 2
lissencephaly in mice lacking all three APP family members. EMBO J. 23,
4106–4115.
Herrick-Davis, K., Grinde, E., Harrigan, T.J., Mazurkiewicz, J.E., 2005.
Inhibition of serotonin 5-hydroxytryptamine2C receptor function through
heterodimerization. Receptor dimers bind two molecules of ligand and one
G-protein. J. Biol. Chem. 280, 40144–40151.
Hesse, L., Beher, D., Masters, C.L., Multhaup, G., 1994. The beta-A4 amyloid
precursor protein binding to copper. FEBS Lett. 349, 109–116.
Ho, A., Sudhof, T.C., 2004. Binding of F-spondin to amyloid-beta precursor
protein: a candidate amyloid-beta precursor protein ligand that modulates
amyloid-beta precursor protein cleavage. Proc. Natl. Acad. Sci. U.S.A. 101,
2548–2553.
Huber, G., Bailly, Y., Martin, J.R., Mariani, J., Brugg, B., 1997. Synaptic beta-
amyloid precursor proteins increase with learning capacity in rats. Neu-
roscience 80, 313–320.
Hynes, T.R., Randal, M., Kennedy, L.A., Eigenbrot, C., Kossiakoff, A.A.,
1990. X-ray crystal structure of the protease inhibitor domain of
Alzheimer’s amyloid beta-protein precursor. Biochemistry 29, 10018–
10022.
Ikezu, T., Okamoto, T., Komatsuzaki, K., Matsui, T., Martyn, J.A.J., Nishimoto,
I., 1996. Negative transactivation of cAMP response element by familial
Alzheimer’s mutants of APP. EMBO J. 15, 2468–2475.
Isacson, O., Seo, H., Lin, L., Albeck, D., Granholm, A.-C., 2002. Alzheimer’s
disease and Down’s syndrome: roles of APP, trophic factors and ACh.
Trends Neurosci. 25, 79–84.
Jansens, A., van Duijn, E., Braakman, I., 2002. Coordinated nonvectorial
folding in a newly synthesized multidomain protein. Science 298, 2401–
2403.
Jellinger, K.A., 2004. Head injury and dementia. Curr. Opin. Neurol. 17, 719–
723.
Jick, H., Zornberg, G.L., Jick, S.S., Seshadri, S., Drachman, D.A., 2000. Statins
and the risk of dementia. Lancet 356, 1627–1631.
Jin, L.W., Ninomiya, H., Roch, J.M., Schubert, D., Masliah, E., Otero, D.A.C.,
Saitoh, T., 1994. Peptides containing the RERMS sequence of amyloid beta/
A4 protein precursor bind cell surface and promote neurite extension. J.
Neurosci. 14, 5461–5470.
Kakio, A., Nishimoto, S., Yanagisawa, K., Kozutsumi, Y., Matsuzaki, K., 2001.
Cholesterol-dependent formation of GM1 ganglioside-bound amyloid beta-
protein, an endogenous seed for Alzheimer amyloid. J. Biol. Chem. 276,
24985–24990.
Kamal, A., Stokin, G.B., Yang, Z.H., Xia, C.H., Goldstein, L.S.B., 2000. Axonal
transport of amyloid precursor protein is mediated by direct binding to the
kinesin light chain subunit of kinesin-I. Neuron 28, 449–459.
Kamenetz, F., Tomita, T., Hsieh, H., Seabrook, G., Borchelt, D., Iwatsubo, T.,
Sisodia, S., Malinow, R., 2003. APP processing and synaptic function.
Neuron 37, 925–937.
Kang, J., Muller-Hill, B., 1990. Differential splicing of Alzheimer’s disease
amyloid A4 precursor RNA in rat tissues—preA4(695) messenger RNA is
predominantly produced in rat and human brain. Biochem. Biophys. Res.
Comm. 166, 1192–1200.
Kang, J., Lemaire, H.G., Unterbeck, A., Salbaum, J.M., Masters, C.L.,
Grzeschik, K.H., Multhaup, G., Beyreuther, K., Muller-Hill, B., 1987.
The precursor of Alzheimer’s disease amyloid-A4 protein resembles a cell
surface receptor. Nature 325, 733–736.

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 29
Kennedy, N.J., Kataoka, T., Tschopp, J., Budd, R.C., 1999. Caspase activation is
required for T cell proliferation. J. Exp. Med. 190, 1891–1895.
Kibbey, M.C., Jucker, M., Weeks, B.S., Neve, R.L., Van Nostrand, W.E.,
Kleinman, H.K., 1993. Beta-amyloid precursor protein binds to the neur-
ite-promoting IKVAV site of laminin. Proc. Natl. Acad. Sci. U.S.A. 90,
10150–10153.
Kimberly, W.T., LaVoie, M.J., Ostaszewski, B.L., Ye, W., Wolfe, M.S., Selkoe,
D.J., 2003. Gamma secretase is a membrane protein complex comprised of
presenilin, nicastrin, Aph-1, and Pen-2. Proc. Natl. Acad. Sci. U.S.A. 100,
6382–6387.
Kimberly, W.T., Zheng, J.B., Town, T., Flavell, R.A., Selkoe, D.J., 2005.
Physiological regulation of the beta-amyloid precursor protein signaling
domain by c-Jun N-terminal kinase JNK3 during neuronal differentiation. J.
Neurosci. 25, 5533–5543.
Kirfel, G., Borm, B., Rigort, A., Herzog, V., 2002. The secretory beta-amyloid
precursor protein is a motogen for human epidermal keratinocytes. Eur. J.
Cell Biol. 81, 664–676.
Kitaguchi, N., Takahashi, Y., Tokushima, Y., Shiojiri, S., Ito, H., 1988. Novel
precursor of Alzheimer’s disease amyloid protein shows protease inhibitory
activity. Nature 331, 530–532.
Klein, W.L., Lacor, P.N., De Felice, F.G., Ferreira, S.T., 2006. Molecules that
disrupt memory circuits in Alzheimer’s disease: the attack on synapses by
Abeta oligomers (ADDLs). In: Bontempi, B., Silva, A., Christen, Y.
(Eds.), Memories: Molecules and Circuits. Springer, Paris.
Kojro, E., Gimpl, G., Lammich, S., Marz, W., Fahrenholz, F., 2001. Low
cholesterol stimulates the nonamyloidogenic pathway by its effect
on the alpha-secretase ADAM 10. Proc. Natl. Acad. Sci. U.S.A. 98,
5815–5820.
Kong, G.K., Adams, J.J., Harris, H.H., Boas, J.F., Curtain, C.C., Galatis, D.,
Masters, C.L., Barnham, K.J., McKinstry, W.J., Cappai, R., Parker, M.W.,
2007. Structural studies of the Alzheimer’s amyloid precursor protein
copper-binding domain reveal how it binds copper ions. J. Mol. Biol.
367, 148–161.
Koo, E.H., Squazzo, S.L., 1994. Evidence that production and release of
amyloid beta-protein involves the endocytic pathway. J. Biol. Chem.
269, 17386–17389.
Koo, E.H., Sisodia, S.S., Archer, D.R., Martin, L.J., Weidemann, A., Beyr-
euther, K., Fischer, P., Masters, C.L., Price, D.L., 1990. Precursor of
amyloid protein in Alzheimer disease undergoes fast anterograde axonal
transport. Proc. Natl. Acad. Sci. U.S.A. 87, 1561–1565.
Kounnas, M.Z., Moir, R.D., Rebeck, G.W., Bush, A.I., Argraves, W.S., Tanzi,
R.E., Hyman, B.T., Strickland, D.K., 1995. LDL receptor-related protein, a
multifunctional ApoE receptor, binds secreted beta amyloid precursor
protein and mediates its degradation. Cell 82, 331–340.
Kroenke, C.D., Ziemnicka-Kotula, D., Xu, J., Kotula, L., Palmer 3rd, A.G.,
1997. Solution conformations of a peptide containing the cytoplasmic
domain sequence of the beta amyloid precursor protein. Biochemistry
36, 8145–8152.
Kuida, K., Zheng, T.S., Na, S.Q., Kuan, C.Y., Yang, D., Karasuyama, H., Rakic,
P., Flavell, R.A., 1996. Decreased apoptosis in the brain and premature
lethality in CPP32-deficient mice. Nature 384, 368–372.
Lacor, P.N., Buniel, M.C., Chang, L., Fernandez, S.J., Gong, Y.S., Viola, K.L.,
Lambert, M.P., Velasco, P.T., Bigio, E.H., Finch, C.E., Krafft, G.A., Klein,
W.L., 2004. Synaptic targeting by Alzheimer’s-related amyloid beta oli-
gomers. J. Neurosci. 24, 10191–10200.
Lambert, M.P., Barlow, A.K., Chromy, B.A., Edwards, C., Freed, R., Liosatos,
M., Morgan, T.E., Rozovsky, I., Trommer, B., Viola, K.L., Wals, P., Zhang,
C., Finch, C.E., Krafft, G.A., Klein, W.L., 1998. Diffusible, nonfibrillar
ligands derived from A beta(1–42) are potent central nervous system
neurotoxins. Proc. Natl. Acad. Sci. U.S.A. 95, 6448–6453.
Lazarov, O., Morfini, G.A., Lee, E.B., Farah, M.H., Szodorai, A., DeBoer, S.R.,
Koliatsos, V.E., Kins, S., Lee, V.M.-Y., Wong, P.C., Price, D.L., Brady, S.T.,
Sisodia, S.S., 2005. Axonal transport, amyloid precursor protein, kinesin-1,
and the processing apparatus: revisited. J. Neurosci. 25, 2386–2395.
LeBlanc, A.C., Kovacs, D.M., Chen, H.Y., Villare, F., Tykocinski, M., Autilio-
Gambetti, L., Gambetti, P., 1992. Role of amyloid precursor protein (APP):
study with antisense transfection of human neuroblastoma cells. J. Neurosci.
Res. 31, 635–645.
Leyssen, M., Ayaz, D., Hebert, S.S., Reeve, S., De Strooper, B., Hassan, B.A.,
2005. Amyloid precursor protein promotes post-developmental neurite
arborization in the Drosophila brain. EMBO J. 24, 2944–2955.
Lien, W.H., Klezovitch, O., Fernandez, T.E., Delrow, J., Vasioukhin, V., 2006.
alpha E-Catenin controls cerebral cortical size by regulating the hedgehog
signaling pathway. Science 311, 1609–1612.
Lindorff-Larsen, K., Best, R.B., DePristo, M.A., Dobson, C.M., Vendruscolo,
M., 2005. Simultaneous determination of protein structure and dynamics.
Nature 433, 128–132.
Loewer, A., Soba, P., Beyreuther, K., Paro, R., Merdes, G., 2004. Cell-type-
specific processing of the amyloid precursor protein by Presenilin during
Drosophila development. EMBO Rep. 5, 405–411.
Loffler, J., Huber, G., 1992. Beta-amyloid precursor protein isoforms in various
rat brain regions and during brain development. J. Neurochem. 59, 1316–
1324.
Lorenzo, A., Yuan, M.L., Zhang, Z.H., Paganetti, P.A., Sturchler-Pierrat, C.,
Staufenbiel, M., Mautino, J., Sol Vigo, F., Sommer, B., Yankner, B.A.,
2000. Amyloid beta interacts with the amyloid precursor protein: a
potential toxic mechanism in Alzheimer’s disease. Nat. Neurosci. 3,
460–464.
Lue, L.F., Kuo, Y.M., Roher, A.E., Brachova, L., Shen, Y., Sue, L., Beach, T.,
Kurth, J.H., Rydel, R.E., Rogers, J., 1999. Soluble amyloid beta peptide
concentration as a predictor of synaptic change in Alzheimer’s disease. Am.
J. Pathol. 155, 853–862.
Lu, D.C., Shaked, G.M., Masliah, E., Bredesen, D.E., Koo, E.H., 2003. Amyloid
beta protein toxicity mediated by the formation of amyloid-beta protein
precursor complexes. Ann. Neurol. 54, 781–789.
Lu, C., Wang, Y., Furukawa, K., Fu, W., Ouyang, X., Mattson, M.P., 2006.
Evidence that caspase-1 is a negative regulator of AMPA receptor-mediated
long-term potentiation at hippocampal synapses. J. Neurochem. 97, 1104–
1110.
Lyckman, A.W., Confaloni, A.M., Thinakaran, G., Sisodia, S.S., Moya, K.L.,
1998. Post-translational processing and turnover kinetics of presynaptically
targeted amyloid precursor superfamily proteins in the central nervous
system. J. Biol. Chem. 273, 11100–11106.
Majocha, R.E., Agrawal, S., Tang, J.Y., Humke, E.W., Marotta, C.A., 1994.
Modulation of the PC12 cell response to nerve growth factor by antisense
oligonucleotide to amyloid precursor protein. Cell Mol. Neurobiol. 14, 425–
437.
Marquez-Sterling, N.R., Lo, A.C.Y., Sisodia, S.S., Koo, E.H., 1997. Trafficking
of cell-surface beta-amyloid precursor protein: evidence that a sorting
intermediate participates in synaptic vesicle recycling. J. Neurosci. 17,
140–151.
Masters, C.L., Simms, G., Weinman, N.A., Multhaup, G., McDonald, B.L.,
Beyreuther, K., 1985. Amyloid plaque core protein in Alzheimer’s disease
and Down Syndrome. Proc. Natl. Acad. Sci. U.S.A. 82, 4245–4249.
Matsuda, S., Yasukawa, T., Homma, Y., Ito, Y., Niikura, T., Hiraki, T., Hirai, S.,
Ohno, S., Kita, Y., Kawasumi, M., Kouyama, K., Yamamoto, T., Kyriakis,
J.M., Nishimoto, I., 2001. c-Jun N-terminal kinase (JNK)-interacting
protein-1b/islet-brain-1 scaffolds Alzheimer’s amyloid precursor protein
with JNK. J. Neurosci. 21, 6597–6607.
Mattson, M.P., Cheng, B., Culwell, A.R., Esch, F.S., Lieberburg, I., Rydel, R.E.,
1993. Evidence for excitoprotective and intraneuronal calcium-regulating
roles for secreted forms of the beta-amyloid precursor protein. Neuron 10,
243–254.
Mauch, D.H., Nagler, K., Schumacher, S., Goritz, C., Muller, E.C., Otto, A.,
Pfrieger, F.W., 2001. CNS synaptogenesis promoted by glia-derived cho-
lesterol. Science 294, 1354–1357.
McLean, C.A., Cherny, R.A., Fraser, F.W., Fuller, S.J., Smith, M.J., Beyreuther,
K., Bush, A.I., Masters, C.L., 1999. Soluble pool of Abeta amyloid as a
determinant of severity of neurodegeneration in Alzheimer’s disease. Ann.
Neurol. 46, 860–866.
McMahon, H.T., Gallop, J.L., 2005. Membrane curvature and mechanisms of
dynamic cell membrane remodelling. Nature 438, 590–596.
Meziane, H., Dodart, J.C., Mathis, C., Little, S., Clemens, J., Paul, S.M.,
Ungerer, A., 1998. Memory-enhancing effects of secreted forms of the beta-
amyloid precursor protein in normal and amnestic mice. Proc. Natl. Acad.
Sci. U.S.A. 95, 12683–12688.

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3230
Milward, E.A., Papadopoulos, R., Fuller, S.J., Moir, R.D., Small, D., Beyr-
euther, K., Masters, C.L., 1992. The amyloid protein precursor of Alzhei-
mer’s disease is a mediator of the effects of nerve growth factor on neurite
outgrowth. Neuron 9, 129–137.
Mok, S.S., Sberna, G., Heffernan, D., Cappai, R., Galatis, D., Clarris, H.J.,
Sawyer, W.H., Beyreuther, K., Masters, C.L., Small, D.H., 1997. Expression
and analysis of heparin-binding regions of the amyloid precursor protein of
Alzheimer’s disease. FEBS Lett. 415, 303–307.
Morimoto, T., Ohsawa, I., Takamura, C., Ishiguro, M., Nakamura, Y., Kohsaka,
S., 1998. Novel domain-specific actions of amyloid precursor protein on
developing synapses. J. Neurosci. 18, 9386–9393.
Moss, L., 2003. What Genes Can’t Do. MIT Press, Cambridge, MA.
Moya, K.L., Benowitz, L.I., Schneider, G.E., Allinquant, B., 1994. The amyloid
precursor protein is developmentally regulated and correlated with synap-
togenesis. Dev. Biol. 161, 597–603.
Multhaup, G., Bush, A.I., Pollwein, P., Masters, C.L., 1994. Interaction between
the zinc(II) and the heparin binding site of the Alzheimer’s disease beta-A4
amyloid precursor protein (APP). FEBS Lett. 355, 151–154.
Multhaup, G., Ruppert, T., Schlicksupp, A., Hesse, L., Bill, E., Pipkorn, R.,
Masters, C.L., Beyreuther, K., 1998. Copper-binding amyloid precursor
protein undergoes a site-specific fragmentation in the reduction of hydrogen
peroxide. Biochemistry 37, 7224–7230.
Narindrasorasak, S., Lowery, D.E., Altman, R.A., Gonzalez-de Whitt, P.A.,
Greenberg, B.D., Kisilevsky, R., 1992. Characterization of high-affinity
binding between laminin and Alzheimer’s disease amyloid precursor pro-
teins. Lab. Invest. 67, 643–652.
Nelson, C.M., Jean, R.P., Tan, J.L., Liu, W.F., Sniadescki, N.J., Spector, A.A.,
Chen, C.S., 2005. Emergent patterns of growth controlled by multicellular
form and mechanics. Proc. Natl. Acad. Sci. U.S.A. 102, 11594–11599.
Nishimoto, I., Okamoto, T., Matsuura, Y., Takahashi, S., Murayama, Y., Ogata,
E., 1993. Alzheimer amyloid protein precursor complexes with brain GTP-
binding protein G(o). Nature 362, 75–79.
Ohsawa, I., Takamura, C., Kohsaka, S., 1997. The amino-terminal region of
amyloid precursor protein is responsible for neurite outgrowth in rat
neocortical explant culture. Biochem. Biophys. Res. Commun. 236, 59–65.
Ohsawa, I., Takamura, C., Kohsaka, S., 2001. Fibulin-1 binds the amino-
terminal head of beta-amyloid precursor protein and modulates its physio-
logical function. J. Neurochem. 76, 1411–1420.
Okamoto, T., Takeda, S., Murayama, Y., Ogata, E., Nishimoto, I., 1995. Ligand-
dependent G protein coupling function of amyloid transmembrane pre-
cursor. J. Biol. Chem. 270, 4205–4208.
Ouimet, C.C., Baerwald, K.D., Gandy, S.E., Greengard, P., 1994. Immunocy-
tochemical localization of amyloid precursor protein in rat brain. J. Comp.
Neurol. 348, 244–260.
Pahlsson, P., Spitalnik, S.L., 1996. The role of glycosylation in synthesis and
secretion of beta-amyloid precursor protein by Chinese hamster ovary cells.
Arch. Biochem. Biophys. 331, 177–186.
Pangalos, M.N., Efthimiopoulos, S., Shioi, J., Robakis, N.K., 1995.
The chondroitin sulfate attachment site of appican is formed by splicing
out exon 15 of the amyloid precursor gene. J. Biol. Chem. 270, 10388–
10391.
Pardossi-Piquard, R., Petit, A., Kawarai, T., Sunyach, C., Alves da Costa, C.,
Vincent, B., Ring, S., D’Adamio, L., Shen, J., Muller, U., Hyslop, P.S.,
Checler, F., 2005. Presenilin-dependent transcriptional control of the Abeta-
degrading enzyme neprilysin by intracellular domains of betaAPP and
APLP. Neuron 46, 541–554.
Pastorino, L., Sun, A., Lu, P.J., Zhou, X.Z., Balastik, M., Finn, G., Wulf, G.,
Lim, J., Li, S.H., Li, X.J., Xia, W.M., Nicholson, L.K., Lu, K.P., 2006. The
prolyl isomerase Pin1 regulates amyloid precursor protein processing and
amyloid-beta production. Nature 440, 528–534.
Patel, T.B., 2004. Single transmembrane spanning heterotrimeric G protein-
coupled receptors and their signaling cascades. Pharmacol. Rev. 56, 371–
385.
Perez, R.G., Zheng, H., Van der Ploeg, L.H., Koo, E.H., 1997. The beta-amyloid
precursor protein of Alzheimer’s disease enhances neuron viability and
modulates neuronal polarity. J. Neurosci. 17, 9407–9414.
Pietrzik, C.U., Hoffmann, J., Stober, K., Chen, C.Y., Bauer, C., Otero, D.A.C.,
Roch, J.M., Herzog, V., 1998. From differentiation to proliferation: the
secretory amyloid precursor protein as a local mediator of growth in thyroid
epithelial cells. Proc. Natl. Acad. Sci. U.S.A. 95, 1770–1775.
Pike, C.J., Walencewicz, A.J., Glabe, C.G., Cotman, C.W., 1991. In vitro aging
of beta-amyloid protein causes peptide aggregation and neurotoxicity. Brain
Res. 563, 311–314.
Ponte, P., Gonzalez-de Whitt, P., Schilling, J., Miller, J., Hsu, D., Greenberg, B.,
Davis, K., Wallace, W., Lieberburg, I., Fuller, F., Cordell, B., 1988. A new
A4-amyloid messenger RNA contains a domain homologous to serine
proteinase inhibitors. Nature 331, 525–527.
Postina, R., Schroeder, A., Dewachter, I., Bohl, J., Schmitt, U., Kojro, E.,
Prinzen, C., Endres, K., Hiemke, C., Blessing, M., Flamez, P., Dequenne,
A., Godaux, E., van Leuven, F., Fahrenholz, F., 2004. A disintegrin-
metalloproteinase prevents amyloid plaque formation and hippocampal
defects in an Alzheimer disease mouse model. J. Clin. Invest. 113,
1456–1464.
Priller, C., Bauer, T., Mitteregger, G., Krebs, B., Kretzschmar, H.A., Herms, J.,
2006. Synapse formation and function is modulated by the amyloid pre-
cursor protein. J. Neurosci. 26, 7212–7221.
Qahwash, I., He, W., Tomasselli, A., Kletzien, R.F., Yan, R., 2004. Processing
amyloid precursor protein at the beta-site requires proper orientation to be
accessed by BACE1. J. Biol. Chem. 279, 39010–39016.
Qiu, W.Q., Ferreira, A., Miller, C., Koo, E.H., Selkoe, D.J., 1995. Cell-surface
beta-amyloid precursor protein stimulates neurite outgrowth of hippocam-
pal neurons in an isoform-dependent manner. J. Neurosci. 15, 2157–2167.
Quast, T., Wehner, S., Kirfel, G., Jaeger, K., De Luca, M., Herzog, V., 2003.
sAPP as a regulator of dendrite motility and melanin release in epidermal
melanocytes and melanoma cells. FASEB J. 17, 1739–1741.
Ramelot, T.A., Gentile, L.N., Nicholson, L.K., 2000. Transient structure of the
amyloid precursor protein cytoplasmic tail indicates preordering of struc-
ture for binding to cytosolic factors. Biochemistry 39, 2714–2725.
Rassoulzadegan, M., Yang, Y.H., Cuzin, F., 1998. APLP2, a member of the
Alzheimer precursor protein family, is required for correct genomic seg-
regation in dividing mouse cells. EMBO J. 17, 4647–4656.
Reichmann, D., Rahat, O., Albeck, S., Meged, R., Dym, O., Schreiber, G., 2005.
The modular architecture of protein–protein binding interfaces. Proc. Natl.
Acad. Sci. U.S.A. 102, 57–62.
Richards, S.J., Hodgman, C., Sharpe, M., 1995. Reported sequence homology
between Alzheimer amyloid(770) and the MRC OX-2 antigen does not
predict function. Brain Res. Bull. 38, 305–306.
Roberts, G.W., Gentleman, S.M., Lynch, A., Graham, D.I., 1991. Beta-A4
amyloid protein deposition in brain after head trauma. Lancet 338, 1422–
1423.
Rossjohn, J., Cappai, R., Feil, S.C., Henry, A., McKinstry, W.J., Galatis, D.,
Hesse, L., Multhaup, G., Beyreuther, K., Masters, C.L., Parker, M.W., 1999.
Crystal structure of the N-terminal, growth factor-like domain of Alzheimer
amyloid precursor protein. Nat. Struct. Biol. 6, 327–331.
Russo, C., Salis, S., Dolcini, V., Venezia, V., Song, X.H., Teller, J.K.,
Schettini, G., 2001. Identification of amino-terminally and phosphotyr-
osine-modified carboxy-terminal fragments of the amyloid precursor
protein in Alzheimer’s disease and Down’s syndrome brain. Neurobiol.
Dis. 8, 173–180.
Russo, C., Dolcini, V., Salis, S., Venezia, V., Zambrano, N., Russo, T., Schettini,
G., 2002. Signal transduction through tyrosine-phosphorylated C-terminal
fragments of amyloid precursor protein via an enhanced interaction with
Shc/Grb2 adaptor proteins in reactive astrocytes of Alzheimer’s disease
brain. J. Biol. Chem. 277, 35282–35288.
Sabo, S.L., Ikin, A.F., Buxbaum, J.D., Greengard, P., 2001. The Alzheimer
amyloid precursor protein (APP) and FE65, an APP-binding protein,
regulate cell movement. J. Cell Biol. 153, 1403–1414.
Sabo, S.L., Ikin, A.F., Buxbaum, J.D., Greengard, P., 2003. The amyloid
precursor protein and its regulatory protein, FE65, in growth cones and
synapses in vitro and in vivo. J. Neurosci. 23, 5407–5415.
Saitoh, T., Sundsmo, M., Roch, J.M., Kimura, N., Cole, G., Schubert, D.,
Oltersdorf, T., Schenk, D.B., 1989. Secreted form of amyloid beta protein
precursor is involved in the growth regulation of fibroblasts. Cell 58, 615–
622.
Sakurai, Y., 1999. How do cell assemblies encode information in the brain?
Neurosci. Biobehav. Rev. 23, 785–796.

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–32 31
Salbaum, J.M., Ruddle, F.H., 1994. Embryonic expression pattern of amyloid
protein precursor suggests a role in differentiation of specific subsets of
neurons. J. Exp. Zool. 269, 116–127.
Sandbrink, R., Masters, C.L., Beyreuther, K., 1994. Similar alternative splicing
of a non-homologous domain in betaA4-amyloid protein precursor-like
proteins. J. Biol. Chem. 269, 14227–14234.
Schaefer, A.W., Kamei, Y., Kamiguchi, H., Wong, E.V., Rapoport, I., Kirch-
hausen, T., Beach, C.M., Landreth, G., Lemmon, S.K., Lemmon, V., 2002.
L1 endocytosis is controlled by a phosphorylation–dephosphorylation cycle
stimulated by outside-in signaling by L1. J. Cell Biol. 157, 1223–1232.
Scheuermann, S., Hambsch, B., Hesse, L., Stumm, J., Schmidt, C., Beher, D.,
Bayer, T.A., Beyreuther, K., Multhaup, G., 2001. Homodimerization of
amyloid precursor protein and its implication in the amyloidogenic pathway
of Alzheimer’s disease. J. Biol. Chem. 276, 33923–33929.
Schubert, D., Lacorbiere, M., Saitoh, T., Cole, G., 1989. Characterization of an
amyloid beta precursor protein that binds heparin and contains tyrosine
sulfate. Proc. Natl. Acad. Sci. U.S.A. 86, 2066–2069.
Selkoe, D.J., 2001. Clearing the brain’s amyloid cobwebs. Neuron 32, 177–180.
Sester, M., Feuerbach, D., Frank, R., Preckel, T., Gutermann, A., Burgert, H.G.,
2000. The amyloid precursor-like protein 2 associates with the major
histocompatibility complex class I molecule K-d. J. Biol. Chem. 275,
3645–3654.
Sisodia, S.S., Koo, E.H., Hoffman, P.N., Perry, G., Price, D.L., 1993. Identifica-
tion and transport of full-length amyloid precursor proteins in rat peripheral
nervous system. J. Neurosci. 13, 3136–3142.
Small, D.H., Nurcombe, V., Reed, G., Clarris, H., Moir, R., Beyreuther, K.,
Masters, C.L., 1994. A heparin-binding domain in the amyloid protein
precursor of Alzheimer’s disease is involved in the regulation of neurite
outgrowth. J. Neurosci. 14, 2117–2127.
Smith-Swintosky, V.L., Pettigrew, L.C., Craddock, S.D., Culwell, A.R., Rydel,
R.E., Mattson, M.P., 1994. Secreted forms of beta-amyloid precursor
protein protect against ischemic brain injury. J. Neurochem. 63, 781–784.
Soba, P., Eggert, S., Wagner, K., Zentgraf, H., Siehl, K., Kreger, S., Lower, A.,
Langer, A., Merdes, G., Paro, R., Masters, C.L., Muller, U., Kins, S.,
Beyreuther, K., 2005. Homo- and heterodimerization of APP family
members promotes intercellular adhesion. EMBO J. 24, 3624–3634.
Sondag, C.M., Combs, C.K., 2006. Amyloid precursor protein cross-linking
stimulates beta amyloid production and pro-inflammatory cytokine release
in monocytic lineage cells. J. Neurochem. 97, 449–461.
Sprecher, C.A., Grant, F.J., Grimm, G., O’Hara, P.J., Norris, F., Norris, K.,
Foster, D.C., 1993. Molecular cloning of the cDNA for a human amyloid
precursor protein homolog: evidence for a multigene family. Biochemistry
32, 4481–4486.
Storey, E., Katz, M., Brickman, Y., Beyreuther, K., Masters, C.L., 1999.
Amyloid precursor protein of Alzheimer’s disease: evidence for a stable,
full-length, trans-membrane pool in primary neuronal cultures. Eur. J.
Neurosci. 11, 1779–1788.
Strittmatter, W.J., Saunders, A.M., Schmechel, D., Pericak-Vance, M., Enghild,
J., Salvesen, G.S., Roses, A.D., 1993a. Apolipoprotein E—high-avidity
binding to beta-amyloid and increased frequency of type 4 allele in late-
onset familial Alzheimer disease. Proc. Natl. Acad. Sci. U.S.A. 90, 1977–
1981.
Strittmatter, W.J., Weisgraber, K.H., Huang, D.Y., Dong, L.M., Salvesen, G.S.,
Pericak-Vance, M., Schmechel, D., Saunders, A.M., Goldgaber, D., Roses,
A.D., 1993b. Binding of human apolipoprotein E to synthetic amyloid beta
peptide: isoform-specific effects and implications for late-onset Alzhei-
mer’s disease. Proc. Natl. Acad. Sci. U.S.A. 90, 8098–8102.
Struhl, G., Adachi, A., 2000. Requirements for presenilin-dependent cleavage
of notch and other transmembrane proteins. Mol. Cell 6, 625–636.
Suzuki, N., Cheung, T.T., Cai, X.D., Odaka, A., Otvos, L., Eckman, C., Golde,
T.E., Younkin, S.G., 1994. An increased percentage of long amyloid beta
protein secreted by familial amyloid beta protein precursor (beta-APP(717))
mutants. Science 264, 1336–1340.
Svergun, D.I., Petoukhov, M.V., Koch, M.H.J., 2001. Determination of domain
structure of proteins from X-ray solution scattering. Biophys. J. 80, 2946–
2953.
Swanson, T.L., Knittel, L.M., Coate, T.M., Farley, S.M., Snyder, M.A., Copen-
haver, P.F., 2005. The insect homologue of the amyloid precursor protein
interacts with the heterotrimeric G protein G0 alpha in an identified
population of migratory neurons. Dev. Biol. 288, 160–178.
Szepanski, S., Veit, M., Pleschka, S., Klenk, H.D., Schmidt, M.F.G., Herrler, G.,
1994. Posttranslational folding of the influenza C virus glycoprotein HEF—
defective processing in cells expressing the cloned gene. J. Gen. Virol. 75,
1023–1030.
Tanzi, R.E., McClatchey, A.I., Lamperti, E.D., Villakomaroff, L., Gusella, J.F.,
Neve, R.L., 1988. Protease inhibitor domain encoded by an amyloid protein
precursor messenger RNA associated with Alzheimer’s disease. Nature 331,
528–530.
Toledo-Rodriguez, M., Blumenfeld, B., Wu, C.Z., Luo, J.Y., Attali, B., Good-
man, P., Markram, H., 2004. Correlation maps allow neuronal electrical
properties to be predicted from single-cell gene expression profiles in rat
neocortex. Cereb. Cortex 14, 1310–1327.
Tolia, A., Chavez-Gutierrez, L., De Strooper, B., 2006. Contribution of pre-
senilin transmembrane domains 6 and 7 to a water-containing cavity in the
gamma-secretase complex. J. Biol. Chem. 281, 27633–27642.
Tominaga-Yoshino, K., Uetsuki, T., Yoshikawa, K., Ogura, A., 2001. Neuro-
toxic and neuroprotective effects of glutamate are enhanced by introduction
of amyloid precursor protein cDNA. Brain Res. 918, 121–130.
Trapp, B.D., Hauer, P.E., 1994. Amyloid precursor protein is enriched in radial
glia—implications for neuronal development. J. Neurosci. Res. 37, 538–
550.
Turner, P.R., O’Connor, K., Tate, W.P., Abraham, W.C., 2003. Roles of amyloid
precursor protein and its fragments in regulating neural activity, plasticity
and memory. Prog. Neurobiol. 70, 1–32.
Van Nostrand, W.E., 1995. Zinc(II) selectively enhances the inhibition of
coagulation factor XIa by protease nexin-2/amyloid beta-protein precursor.
Thromb. Res. 78, 43–53.
Van Nostrand, W.E., Schmaier, A.H., Farrow, J.S., Cunningham, D.D., 1990.
Protease nexin-II (amyloid beta-protein precursor): a platelet alpha-granule
protein. Science 248, 745–748.
Van Nostrand, W.E., Melchor, J.P., Keane, D.M., Saporito-Irwin, S.M., Roma-
nov, G., Davis, J., Xu, F., 2002. Localization of a fibrillar amyloid beta-
protein binding domain on its precursor. J. Biol. Chem. 277, 36392–36398.
Vassar, R., 2002. Beta-secretase (BACE) as a drug target for Alzheimer’s
disease. Adv. Drug Deliv. Rev. 54, 1589–1602.
Wagner, S.L., Siegel, R.S., Vedvick, T.S., Raschke, W.C., Van Nostrand, W.E.,
1992. High-level expression, purification, and characterization of the
Kunitz-type protease inhibitor domain of protease nexin-2/amyloid beta
protein precursor. Biochem. Biophys. Res. Commun. 186, 1138–1145.
Walsh, D.M., Klyubin, I., Fadeeva, J.V., Cullen, W.K., Anwyl, R., Wolfe, M.S.,
Rowan, M.J., Selkoe, D.J., 2002. Naturally secreted oligomers of amyloid
beta protein potently inhibit hippocampal long-term potentiation in vivo.
Nature 416, 535–539.
Walsh, D.M., Fadeeva, J.V., LaVoie, M.J., Paliga, K., Eggert, S., Kimberly,
W.T., Wasco, W., Selkoe, D.J., 2003. Gamma-secretase cleavage and
binding to FE65 regulate the nuclear translocation of the intracellular C-
terminal domain (ICD) of the APP family of proteins. Biochemistry 42,
6664–6673.
Wang, Y., Ha, Y., 2004. The X-ray structure of an antiparallel dimer of the
human amyloid precursor protein E2 domain. Mol. Cell 15, 343–353.
Wang, H.W., Pasternak, J.F., Kuo, H., Ristic, H., Lambert, M.P., Chromy, B.,
Viola, K.L., Klein, W.L., Stine, W.B., Krafft, G.A., Trommer, B.L., 2002.
Soluble oligomers of beta amyloid (1-42) inhibit long-term potentiation but
not long-term depression in rat dentate gyrus. Brain Res. 924, 133–140.
Wang, P., Yang, G., Mosier, D.R., Chang, P., Zaidi, T., Gong, Y.D., Zhao, N.M.,
Dominguez, B., Lee, K.F., Gan, W.B., Zheng, H., 2005. Defective neuro-
muscular synapses in mice lacking amyloid precursor protein (APP) and
APP-like protein 2. J. Neurosci. 25, 1219–1225.
Wasco, W., Bupp, K., Magendantz, M., Gusella, J.F., Tanzi, R.E., Solomon, F.,
1992. Identification of a mouse brain cDNA that encodes a protein related to
the Alzheimer disease-associated amyloid-beta protein precursor. Proc.
Natl. Acad. Sci. U.S.A. 89, 10758–10762.
Wasco, W., Gurubhagavatula, S., Dparadis, M., Romano, D.M., Sisodia, S.S.,
Hyman, B.T., Neve, R.L., Tanzi, R.E., 1993. Isolation and characterization
of APLP2 encoding a homolog of the Alzheimer’s associated amyloid beta-
protein precursor. Nat. Genet. 5, 95–100.

M. Gralle, S.T. Ferreira / Progress in Neurobiology 82 (2007) 11–3232
Washbourne, P., Dityatev, A., Scheiffele, P., Biederer, T., Weiner, J.A., Chris-
topherson, K.S., El-Husseini, A., 2004. Cell adhesion molecules in synapse
formation. J. Neurosci. 24, 9244–9249.
Weber, G., 1992. Protein Interactions. Chapman & Hall, New York.
Yankner, B.A., Duffy, L.K., Kirschner, D.A., 1990. Neurotrophic and neuro-
toxic effects of amyloid beta protein: reversal by tachykinin neuropeptides.
Science 250, 279–282.
Williamson, T.G., Mok, S.S., Henry, A., Cappai, R., Lander, A.D., Nurcombe,
V., Beyreuther, K., Masters, C.L., Small, D.H., 1996. Secreted glypican
binds to the amyloid precursor protein of Alzheimer’s disease (APP) and
inhibits APP-induced neurite outgrowth. J. Biol. Chem. 271, 31215–31221.
Wilson, C.A., Doms, R.W., Zheng, H., Lee, V.M.-Y., 2002. Presenilins are not
required for Abeta42 production in the early secretory pathway. Nat.
Neurosci. 5, 849–855.
Wolfe, M.S., Xia, W., Ostaszewski, B.L., Diehl, T.S., Kimberly, W.T., Selkoe,
D.J., 1999. Two transmembrane aspartates in presenilin 1 required for
presenilin endoproteolysis and gamma secretase activity. Nature 398, 513–
517.
Wootton, J.C., Federhen, S., 1996. Analysis of compositionally biased regions
in sequence databases. Methods Enzymol. 266, 554–571.
Wright, P.E., Dyson, H.J., 1999. Intrinsically unstructured proteins: re-assessing
the protein structure–function paradigm. J. Mol. Biol. 293, 321–331.
Yamatsuji, T., Matsui, T., Okamoto, T., Komatsuzaki, K., Takeda, S., Fukumoto,
H., Iwatsubo, T., Suzuki, N., Asami-Odaka, A., Ireland, S., Kinane, T.B.,
Giambarella, U., Nishimoto, I., 1996a. G protein-mediated neuronal DNA
fragmentation induced by familial Alzheimer’s disease-associated mutants
of APP. Science 272, 1349–1352.
Yamatsuji, T., Okamoto, T., Takeda, S., Murayama, Y., Tanaka, N., Nishimoto,
I., 1996b. Expression of V642 APP mutant causes cellular apoptosis as
Alzheimer trait-linked phenotype. EMBO J. 15, 498–509.
Yamazaki, T., Selkoe, D.J., Koo, E.H., 1995. Trafficking of cell-surface beta-
amyloid precursor protein—retrograde and transcytotic transport in cul-
tured neurons. J. Cell Biol. 129, 431–442.
Yang, G., Gong, Y.D., Gong, K., Jiang, W.L., Kwon, E., Wang, P., Zheng, H.,
Zhang, X.F., Gan, W.B., Zhao, N.M., 2005. Reduced synaptic vesicle
density and active zone size in mice lacking amyloid precursor protein
(APP) and APP-like protein 2. Neurosci. Lett. 384, 66–71.
Yoshikai, S., Sasaki, H., Doh-ura, K., Furuya, H., Sakaki, Y., 1990. Genomic
organization of the human amyloid beta-protein precursor gene. Gene 87,
257–263.
Yoshikai, S., Sasaki, H., Doh-ura, K., Furuya, H., Sakaki, Y., 1991. Genomic
organization of the human-amyloid beta-protein precursor gene. Gene 102,
291–292.
Zambrano, N., Bruni, P., Minopoli, G., Mosca, R., Molino, D., Russo, C.,
Schettini, G., Sudol, M., Russo, T., 2001. The beta-amyloid precursor
protein APP is tyrosine-phosphorylated in cells expressing a constitu-
tively active form of the Abl protooncogene. J. Biol. Chem. 276, 19787–
19792.
Zhang, Y., Scandura, J.M., Van Nostrand, W.E., Walsh, P.N., 1997. The
mechanism by which heparin promotes the inhibition of coagulation factor
XIa by protease nexin-2. J. Biol. Chem. 272, 26139–26144.
Zheng, H., Jiang, M., Trumbauer, M.E., Sirinathsinghji, D.J., Hopkins, R.,
Smith, D.W., Heavens, R.P., Dawson, G.R., Boyce, S., Conner, M.W.,
Stevens, K.A., Slunt, H.H., Sisodia, S.S., Chen, H.Y., Van der Ploeg,
L.H., 1995. Beta-amyloid precursor protein-deficient mice show reactive
gliosis and decreased locomotor activity. Cell 81, 525–531.
Zhou, S., Zhou, H., Walian, P.J., Jap, B.K., 2005. CD147 is a regulatory
subunit of the gamma-secretase complex in Alzheimer’s disease
amyloid beta-peptide production. Proc. Natl. Acad. Sci. U.S.A. 102,
7499–7504.




















