
Amyloid Precursor-Like Protein 2 Increases the Endocytosis, Instability, and Turnover of the H2-Kd...
11
of February 1, 2013. This information is current as MHC Class I Molecule d the H2-K the Endocytosis, Instability, and Turnover of Amyloid Precursor-Like Protein 2 Increases Solheim Talmadge, Naava Naslavsky, Steve Caplan and Joyce C. Amit Tuli, Mahak Sharma, Mary M. McIlhaney, James E. http://www.jimmunol.org/content/181/3/1978 2008; 181:1978-1987; ; J Immunol References http://www.jimmunol.org/content/181/3/1978.full#ref-list-1 , 33 of which you can access for free at: cites 59 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2008 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology at UNiversity of Nebraska Medical Center on February 1, 2013 http://jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Amyloid Precursor-Like Protein 2 Increases the Endocytosis, Instability, and Turnover of the H2-Kd...

of February 1, 2013.This information is current as
MHC Class I Moleculedthe H2-Kthe Endocytosis, Instability, and Turnover of Amyloid Precursor-Like Protein 2 Increases
SolheimTalmadge, Naava Naslavsky, Steve Caplan and Joyce C. Amit Tuli, Mahak Sharma, Mary M. McIlhaney, James E.
http://www.jimmunol.org/content/181/3/19782008; 181:1978-1987; ;J Immunol
Referenceshttp://www.jimmunol.org/content/181/3/1978.full#ref-list-1
, 33 of which you can access for free at: cites 59 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2008 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from

Amyloid Precursor-Like Protein 2 Increases the Endocytosis,Instability, and Turnover of the H2-Kd MHC Class IMolecule1
Amit Tuli,*† Mahak Sharma,* Mary M. McIlhaney,† James E. Talmadge,‡ Naava Naslavsky,*Steve Caplan,* and Joyce C. Solheim2*†‡
The defense against the invasion of viruses and tumors relies on the presentation of viral and tumor-derived peptides to CTL bycell surface MHC class I molecules. Previously, we showed that the ubiquitously expressed protein amyloid precursor-like protein2 (APLP2) associates with the folded form of the MHC class I molecule Kd. In the current study, APLP2 was found to associatewith folded Kd molecules following their endocytosis and to increase the amount of endocytosed Kd. In addition, increasedexpression of APLP2 was shown to decrease Kd surface expression and thermostability. Correspondingly, Kd thermostability andsurface expression were increased by down-regulation of APLP2 expression. Overall, these data suggest that APLP2 modulatesthe stability and endocytosis of Kd molecules. The Journal of Immunology, 2008, 181: 1978–1987.
T he presentation of antigenic peptides to T lymphocytes byMHC molecules is essential for recognition and killing ofinfected and malignant cells. Assembly of MHC class I H
chain with antigenic peptide and with the MHC class I L chain, �2-microglobulin (�2m)3 occurs in the endoplasmic reticulum (ER).Peptide processing and loading of MHC class I molecules involvesthe participation of several ER proteins: the transporter associatedwith Ag processing, tapasin, calreticulin, ERp57, Bap 29/31, pro-tein disulfide isomerase, and ER aminopeptidase (1–5). Evidencealso indicates that there is regulation of MHC class I traffickingbetween the ER and the plasma membrane (2, 6–11), although ourunderstanding of this process is relatively limited.
A protein not restricted to the ER that associates with the MHCclass I molecule is amyloid precursor-like protein 2 (APLP2) (12–14). APLP2 is a type I transmembrane protein which has a largeectodomain that can be cleaved off and secreted (15). APLP2 isubiquitously expressed (15), and has a variety of cellular functions,i.e., involvement in mitotic segregation, neurite outgrowth, andepithelial cell migration (16–20). APLP2 is closely related in se-
quence to amyloid precursor protein (APP), but does not have an�-amyloid peptide domain (21–22).
APLP2 was first identified as a protein coimmunoprecipitatingwith H2-Kd by microsequencing and serological methods (12–13).We have found that transient transfection with APLP2-specificsiRNA increases the cell surface expression of Kd, suggesting thatnormally APLP2 has a down-regulatory effect on Kd surface ex-pression (14). Furthermore, APLP2 interacts with folded Kd mol-ecules and not with open, peptide-free Kd molecules (14), and onlyassociates with Kd in the presence of �2m (13). Notably, APLP2can be displaced from Kd in cell lysates by the addition of Kd-binding peptides (12), suggesting APLP2 interacts with the �1/�2domain of Kd.
Our new studies have shown that increased expression ofAPLP2 reduces the quantity of Kd molecules present at the plasmamembrane. We also demonstrated that increased APLP2 expres-sion resulted in greater internalization and more rapid turnover ofKd. In addition, we found that APLP2 binds to the endocytosed Kd
molecules. Furthermore, we found that the overall stability of Kd
molecules is inversely related to the level of expression of APLP2in the cell. These data indicate that APLP2 can interact with en-docytosed Kd molecules and that it regulates the stability and sur-face expression of folded Kd molecules.
Materials and MethodsAntibodies
The 34-1-2 mAb recognizes the �1/�2 domain of Kd, Dd, and Dq, and itbinds weakly to Db and Ld, but strongly to Ld associated with human �2mand to some Ld mutants with amino acid substitutions in the peptide-bind-ing groove (23–25). The 64-3-7 mAb can detect open, peptide-free Ld (26)and can also detect open forms of other MHC class I H chains, such as Kd,into which the 64-3-7 epitope has been introduced. Introduction of the64-3-7 epitope does not impair peptide presentation, trafficking, or surfaceexpression of Kd or other MHC class I molecules (27–30). The 30-5-7 mAbrecognizes Ld molecules with a folded peptide-binding groove (26, 31–34),and the 28-14-8 mAb binds to the �3 domain of Ld, Db, Dq, and Lq (32, 33,35). The 34-1-2, 30-5-7, 28-14-8, and 64-3-7 Abs were donated by Dr. T.Hansen (Washington University, St. Louis, MO). The Ab used for APLP2detection was made against full-length APLP2 (Calbiochem). The Ab rec-ognizing �-actin (PanAb5) was purchased from Novus Biologicals.
*Department of Biochemistry and Molecular Biology, †Eppley Institute for Researchin Cancer and Allied Diseases, and ‡Department of Pathology and Microbiology,University of Nebraska Medical Center, Omaha, NE 68198
Received for publication July 17, 2007. Accepted for publication May 22, 2008.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grant GM57428 (toJ.C.S.) and GM74876 (to S.C.), the Nebraska Research Initiative Program in Trans-lational Biotechnology Research (to J.E.T. and J.C.S.), University of Nebraska Med-ical Center Graduate Studies Fellowships (to A.T. and M.S.), and a Nebraska Centerfor Cellular Signaling Fellowship (to M.S.). Core facilities at the University of Ne-braska Medical Center receive support from the National Institutes of Health CancerCenter Support Grant P30 CA036727.2 Address correspondence and reprint requests to Dr. Joyce C. Solheim, Eppley In-stitute for Research in Cancer and Allied Diseases, University of Nebraska MedicalCenter, 986805 Nebraska Medical Center, Omaha, NE 68198-6805. E-mail address:[email protected] Abbreviations used in this paper: �2m, � 2-microglobulin; ER, endoplasmic retic-ulum; APLP2, amyloid precursor-like protein 2; APP, amyloid precursor protein;CHAPS, 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate; Endo, en-doglycosidase; EHD1, Eps15 homology domain-containing protein.
Copyright © 2008 by The American Association of Immunologists, Inc. 0022-1767/08/$2.00
The Journal of Immunology
www.jimmunol.org
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from
Cell lines
Cell lines were grown in RPMI 1640 medium (Invitrogen) supplementedwith 15% FBS, glutamine, pyruvate, and penicillin/streptomycin. TheHeLa cell line was provided to us by Dr. W. Maury (University of Iowa,Iowa City, IA). HeLa cells were transfected with the cDNA encoding Kd
with the 64-3-7 epitope (27) in the pIRIS.puro2 vector (BD BiosciencesClontech) or with an Ld cDNA (33) in the RSV.5neo vector (36). Asmentioned above, previous studies have established the epitope-tagged Kd
(etKd) molecule as exhibiting normal assembly, folding, and trafficking(27). Selection in medium containing puromycin was performed to gener-ate the stable HeLa-etKd cell line. APLP2 siRNA and FLAG-taggedAPLP2 constructs were generated for this study. The C-terminally FLAG-tagged APLP2 cDNA in the pCMVTag4A vector (Stratagene) was trans-fected into HeLa-etKd cells to generate HeLa-etKd cells expressing APLP2at an elevated level. APLP2-specific short interfering RNA (siRNA) orinverse siRNA (the reverse sequence of the APLP2 siRNA) in the pSUPERvector was transfected into HeLa-etKd cells to generate the stable HeLa-etKd-APLP2 siRNA or HeLa-etKd-inverse siRNA cell line, respectively.The sequence used for APLP2 siRNA was confirmed to be unique bysearch of the National Center for Biotechnology Information internet site.Stable clones expressing transfected APLP2, APLP2-specific siRNA, orAPLP2 inverse siRNA were selected by incubation of the cells in mediumwith G418. Quantitative RT-PCR on samples from cells expressingAPLP2-specific siRNA or inverse siRNA confirmed that APLP2-specificsiRNA did not induce IFN expression more than the control inverse siRNA(data not shown). Western blotting for etKd was used to select for stabletransfectants that expressed the same level of etKd. Transient transfectionof FLAG-tagged APLP2 was done with analysis of the cells at 24 h fol-lowing the transfection procedure. A transferrin receptor cDNA (37) ex-pressing a C-terminal GFP tag in the Clontech EGFP vector (a gift from Dr.R. Lodge, Universite du Quebec, Montreal, Quebec) was transiently trans-fected into HeLa-etKd cells and confocal analysis was performed at 24 hafter transfection and flow cytometric analysis was performed at 48 h aftertransfection. GFP-tagged Rab5 and Rab5Q79L cDNAs (38) in the EGFPvector (Clontech) were also gifts from Dr. R. Lodge. The Rab5 orRab5Q79L construct was transiently transfected into HeLa-etKd cells andconfocal analysis was performed at 24 h after transfection. All transfections
were performed using Effectene (Qiagen) with 1 �g DNA per 0.5 � 106
cells.
Immunoprecipitations and Western blots
Immunoprecipitations and Western blotting were performed by a methodsimilar to a published protocol (39). For protein immunoprecipitations, thecells were washed in PBS containing 20 mM iodoacetamide (Sigma-Aldrich) three times and lysed in 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS) lysis buffer. The CHAPS buffer contained 1%CHAPS (Roche Applied Science) in Tris-buffered saline (pH 7.4) with freshlyadded 0.2 mM PMSF and 20 mM iodoacetamide and a saturating amount ofmAb. After 1 h on ice, the lysates were centrifuged to remove cell nuclei andincubated with protein A-Sepharose beads (GE Healthcare Biosciences). Thebeads were washed in 0.1% CHAPS/20 mM iodoacetamide in TBS (pH 7.4)four times and boiled in 0.125 M Tris (pH 6.8)/2% SDS/12% glycerol/0.02%bromphenol blue to elute the proteins.
The eluted immunoprecipitates were electrophoresed on SDS-PAGEgels (Invitrogen) and transferred to Immobilon-P membranes (Millipore)for Western blots. After overnight blocking in reconstituted dry milk,membranes were incubated in diluted Ab for 2 h, washed three times with0.05% Tween 20/PBS, and incubated for 1 h in a dilution of biotin-con-jugated goat anti-mouse or anti-rabbit IgG (Caltag Laboratories). Afterthree 0.05% Tween 20/PBS washes, the membranes were incubated withdiluted streptavidin-conjugated HRP (Zymed) for 1 h, washed with 0.3%Tween 20/PBS three times, and incubated with ECL Western blot devel-oping reagents (GE Healthcare Biosciences). The membranes were ex-posed to Kodak BioMax film (Eastman Kodak).
When Western blots were performed on cell lysates without an immu-noprecipitation step, the cells were washed in PBS containing 20 mMiodoacetamide (Sigma-Aldrich) three times and lysed in buffer containing0.125M Tris (pH 6.8)/2% (w/v) SDS/12% (v/v) glycerol/0.02% (w/v) bro-mophenol blue and fresh 0.2 mM PMSF. The lysates were incubated 1 h onice, then centrifuged to pellet nuclear material. Samples of the supernatantswere boiled before loading onto gels. Subsequent steps were performed asdescribed above.
For the endoglycosidase (Endo) H assay, immunoprecipitations werefirst performed as described above, except proteins were eluted from the
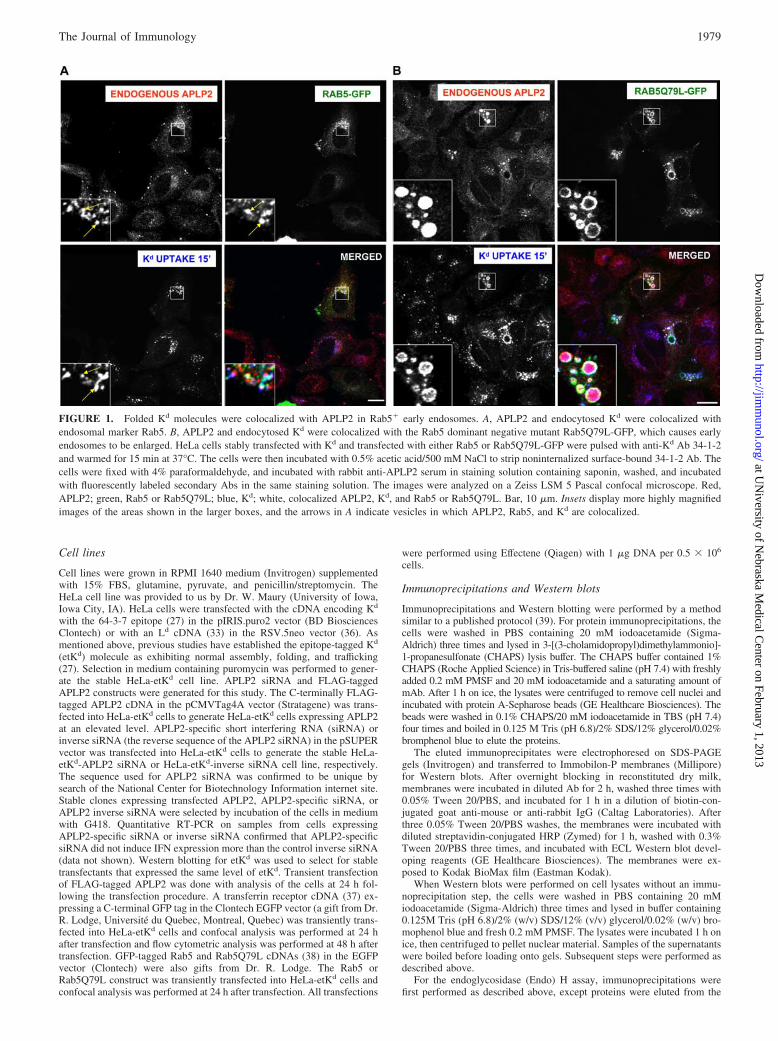
FIGURE 1. Folded Kd molecules were colocalized with APLP2 in Rab5� early endosomes. A, APLP2 and endocytosed Kd were colocalized withendosomal marker Rab5. B, APLP2 and endocytosed Kd were colocalized with the Rab5 dominant negative mutant Rab5Q79L-GFP, which causes earlyendosomes to be enlarged. HeLa cells stably transfected with Kd and transfected with either Rab5 or Rab5Q79L-GFP were pulsed with anti-Kd Ab 34-1-2and warmed for 15 min at 37°C. The cells were then incubated with 0.5% acetic acid/500 mM NaCl to strip noninternalized surface-bound 34-1-2 Ab. Thecells were fixed with 4% paraformaldehyde, and incubated with rabbit anti-APLP2 serum in staining solution containing saponin, washed, and incubatedwith fluorescently labeled secondary Abs in the same staining solution. The images were analyzed on a Zeiss LSM 5 Pascal confocal microscope. Red,APLP2; green, Rab5 or Rab5Q79L; blue, Kd; white, colocalized APLP2, Kd, and Rab5 or Rab5Q79L. Bar, 10 �m. Insets display more highly magnifiedimages of the areas shown in the larger boxes, and the arrows in A indicate vesicles in which APLP2, Rab5, and Kd are colocalized.
1979The Journal of Immunology
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from
protein A-Sepharose beads by boiling the samples for 5 min in 25 mM Tris(pH 8.3)/0.2 M glycine/0.1% SDS), centrifuging, and transferring the su-pernatants to fresh tubes. A 10� glycoprotein denaturing buffer (New En-gland Biolabs) was added to 9 �l of supernatant to a final concentration of
1�, and the sample was boiled for 10 min. The sample was then split inhalf, and the reaction volume of each half was increased by addition of 2�l of 10 G5 reaction buffer (New England Biolabs), 2 �l of Endo H (NewEngland Biolabs), or 2 �l water, for the mock digestion), and a quantity of
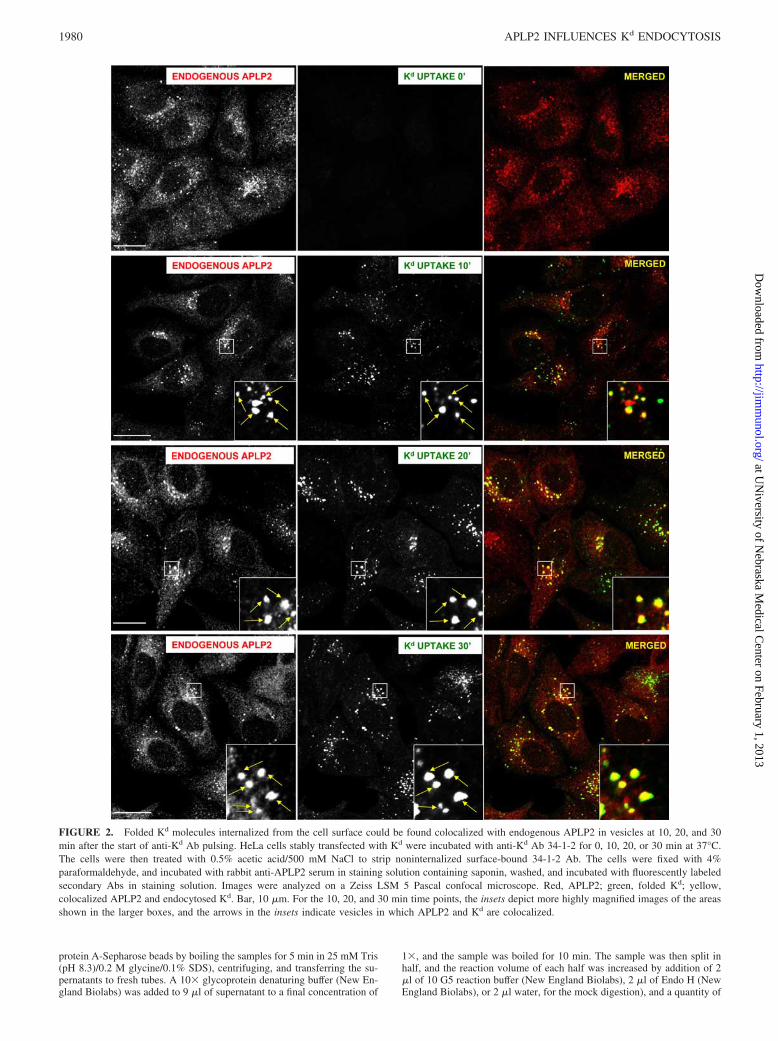
FIGURE 2. Folded Kd molecules internalized from the cell surface could be found colocalized with endogenous APLP2 in vesicles at 10, 20, and 30min after the start of anti-Kd Ab pulsing. HeLa cells stably transfected with Kd were incubated with anti-Kd Ab 34-1-2 for 0, 10, 20, or 30 min at 37°C.The cells were then treated with 0.5% acetic acid/500 mM NaCl to strip noninternalized surface-bound 34-1-2 Ab. The cells were fixed with 4%paraformaldehyde, and incubated with rabbit anti-APLP2 serum in staining solution containing saponin, washed, and incubated with fluorescently labeledsecondary Abs in staining solution. Images were analyzed on a Zeiss LSM 5 Pascal confocal microscope. Red, APLP2; green, folded Kd; yellow,colocalized APLP2 and endocytosed Kd. Bar, 10 �m. For the 10, 20, and 30 min time points, the insets depict more highly magnified images of the areasshown in the larger boxes, and the arrows in the insets indicate vesicles in which APLP2 and Kd are colocalized.
1980 APLP2 INFLUENCES Kd ENDOCYTOSIS
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from

water sufficient to yield a final volume of 20 �l. The tubes were incubatedfor 1 h at 37°C, and then 5 �l of 0.5 M Tris (pH 6.8)/8% SDS/48%glycerol/0.08% bromophenol blue/8% 2-ME were added. Samples wereelectrophoresed on 4–20% acrylamide Tris-glycine gels, and Western blotswere performed as described above.
Assessment of the Kd turnover rate
For analysis of Kd turnover, a method that was previously described wasused (40). Cells were treated with 10 �g/ml cycloheximide and then har-vested at 0, 1, 2, 4, or 8 h. Equivalent numbers of live cells were processedas described above for Western blots, and Ab 64-3-7 was used to detect theepitope-tagged Kd. Band intensity for Kd was normalized to �-actin bandintensity at the same time point. Values were expressed as the percentageof remaining Kd at the 0, 1, 2, 4, or 8 h time point.
Monitoring for induction of stress response
To test whether an increase in expression of APLP2 causes a cellular stressresponse, we monitored the expression of ER stress proteins, using a pub-lished approach (41). For this experiment, we used HeLa-etKd cells (stablyexpressing Kd) that had been transiently transfected with APLP2-FLAG,
transfected with vector only, or were left untransfected with either APLP2or vector. At 32 h posttransfection, the medium was removed and freshcomplete medium was added and the cells were incubated for another 16 h.To generate a positive control, during the 16 h, HeLa-etKd cells weretreated with 2 �g/ml tunicamycin (Sigma-Aldrich). The samples were elec-trophoresed on 4–20% Tris-glycine acrylamide gels, transferred to blottingmembranes, and Western blots of lysates of these cells were probed withan Ab specific for the KDEL sequence (Stressgen) present on the stressproteins GrP94 and BiP.
Biochemical analysis of the binding of APLP2 toendocytosed Kd
To demonstrate that APLP2 was bound to endocytosed Kd, HeLa-etKd
cells transiently transfected for 24 h with APLP2-FLAG were incubatedwith 34-1-2 Ab on ice for 20 min and then warmed at 37°C for 20 min. Any34-1-2 Ab still bound to cell surface Kd was removed by washing withstripping buffer (0.5% acetic acid and 500 mM NaCl), and the cells werelysed. After centrifugation, protein A-Sepharose was added to the lysatesupernatant. Several controls were included in the experiment: lysed HeLa-etKd cells, with no Ab added before or after lysis; HeLa and HeLa-etKd
FIGURE 3. Increased expression of APLP2 was found to enhance the endocytosis of Kd. A, HeLa-etKd cells (stably expressing Kd) were transientlytransfected with APLP2-FLAG for 24 h. Anti-Kd Ab 34-1-2 was added and the cells were warmed to 37°C for 20 min. Following Ab internalization, thecells were treated with 0.5% acetic acid/500 mM NaCl to strip off noninternalized surface-bound 34-1-2. The cells were fixed with 4% paraformaldehyde,incubated in staining solution (containing saponin) with rabbit anti-FLAG, washed, and incubated in staining solution and fluorescently labeled secondaryAbs, and visualized with a Zeiss LSM 5 Pascal confocal microscope. Red, APLP2; green, folded Kd; yellow, colocalized APLP2 and Kd. RepresentativeAPLP2-transfected cells are outlined with a dashed line. Bar, 10 �m. The insets display more highly magnified images of the areas depicted in the largerboxes. Arrows in the insets point to vesicles in which APLP2-FLAG and Kd are colocalized. B, Results confirming that APLP2 and endocytosed Kd arelocated together in vesicles were obtained by taking z-section images. Serial z-section images were acquired at 0.4 �m intervals of HeLa-etKd cellstransfected with APLP2-FLAG for 24 h, surface-labeled with 34-1-2, and incubated at 37°C for 15 min. The arrows point to common membrane structureson a representative photomicrograph. APLP2, red; Kd, green; merged green and red, yellow. These data confirm that the indicated APLP2- and Kd-containing structures are the same endocytic vesicles and not overlaid vesicles. C, APLP2 was bound to endocytosed Kd. Lane 1, Lysate of HeLa-etKd cellsplus protein A-Sepharose beads; lanes 2 and 3, 34-1-2 Ab was added to lysates of HeLa and HeLa-etKd for immunoprecipitation of Kd; lane 4, HeLa-etKd
cells transfected with APLP2-FLAG for 24 h were incubated with 34-1-2 Ab for 20 min on ice and then transferred to 37°C for 20 min, noninternalized34-1-2 Ab was removed by an acid wash, the cells were lysed and centrifuged, and protein A-Sepharose was added to the lysate supernatant; lanes 5 and6, Isotype control Ab (28-14-8) was added to lysates of HeLa and HeLa-etKd and an immunoprecipitation procedure was performed; lane 7, HeLa-etKd
cells transfected with APLP2-FLAG for 24 h were incubated with the isotype control Ab 28-14-8 for 20 min on ice and then transferred to 37°C for 20min, the cells were treated with an acid wash, lysed, and centrifuged, and protein A-Sepharose was added to the lysate supernatant. The samples wereelectrophoresed on 4320% acrylamide Tris-glycine gels, transferred to blots, and probed with mAb 64-3-7 that recognizes etKd (top panel) or with rabbitantiserum against APLP2 (bottom panel). The bands in the 2nd, 5th, and 6th lanes are nonspecific bands. D, Higher expression of APLP2 resulted in increasedKd endocytosis. Image J Software (http://rsb.info.nih.gov) was used to measure the fluorescence of the internalized Kd expressed by �80 cells transfectedwith APLP2-FLAG and �80 cells not transfected with APLP2-FLAG within the experiment for which confocal data is shown in A. Mean fluorescenceintensities and SEs of the mean were calculated, and p values were determined by the use of Student’s paired t test.
1981The Journal of Immunology
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from
cells lysed with 34-1-2 or with the 28-14-8 mAb (as an isotype control) inthe lysis buffer; and HeLa-etKd cells transiently transfected with APLP2-FLAG and treated with the surface-labeling procedure as described abovefor HeLa-etKd except that an isotype control Ab (28-14-8) was used insteadof 34-1-2. The samples were electrophoresed on 4320% acrylamide Tris-glycine gels and transferred to blotting membranes, which were probedwith mAb 64-3-7 to identify etKd or with rabbit antiserum to identifycoprecipitated APLP2.
Thermostability assays
The thermostability assay procedure used in this study was designed basedon a published procedure (42). For the thermostability assay, cells werewashed and lysed with CHAPS buffer and lysates were centrifuged to re-move cell nuclei just as described in the section above. Aliquots from thesupernatants were incubated at varied temperatures for 12 min on ice or in
a Biometra T3 gradient thermocycler (Whatman Biometra). After the ther-mocycler incubations, immunoprecipitations and Western blots were per-formed on the aliquots as described in the section above. The amount of Kd
in Western blot bands was quantified by densitometry using a Storm (Mo-lecular Dynamics). The relative percentage of folded Kd at each incubationtemperature between 25°C and 50°C was calculated after setting theamount of folded Kd at 4°C as 100%.
Flow cytometry assays
In flow cytometry assays, cells were suspended at 5 � 106/ml in PBS with0.2% BSA and 0.1% sodium azide. Cell suspension aliquots in volumes of0.1 ml were distributed to the wells of a 96-well plate. The cells wereincubated with excess mAb or with BSA/azide/PBS alone (as a control) at
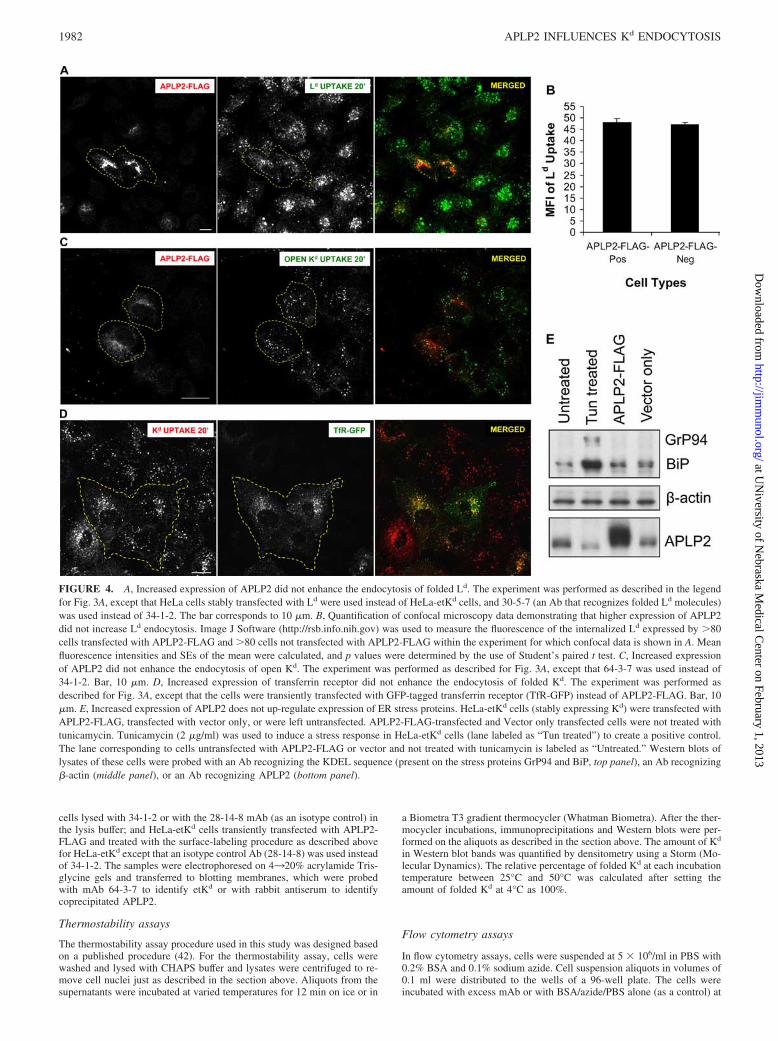
FIGURE 4. A, Increased expression of APLP2 did not enhance the endocytosis of folded Ld. The experiment was performed as described in the legendfor Fig. 3A, except that HeLa cells stably transfected with Ld were used instead of HeLa-etKd cells, and 30-5-7 (an Ab that recognizes folded Ld molecules)was used instead of 34-1-2. The bar corresponds to 10 �m. B, Quantification of confocal microscopy data demonstrating that higher expression of APLP2did not increase Ld endocytosis. Image J Software (http://rsb.info.nih.gov) was used to measure the fluorescence of the internalized Ld expressed by �80cells transfected with APLP2-FLAG and �80 cells not transfected with APLP2-FLAG within the experiment for which confocal data is shown in A. Meanfluorescence intensities and SEs of the mean were calculated, and p values were determined by the use of Student’s paired t test. C, Increased expressionof APLP2 did not enhance the endocytosis of open Kd. The experiment was performed as described for Fig. 3A, except that 64-3-7 was used instead of34-1-2. Bar, 10 �m. D, Increased expression of transferrin receptor did not enhance the endocytosis of folded Kd. The experiment was performed asdescribed for Fig. 3A, except that the cells were transiently transfected with GFP-tagged transferrin receptor (TfR-GFP) instead of APLP2-FLAG. Bar, 10�m. E, Increased expression of APLP2 does not up-regulate expression of ER stress proteins. HeLa-etKd cells (stably expressing Kd) were transfected withAPLP2-FLAG, transfected with vector only, or were left untransfected. APLP2-FLAG-transfected and Vector only transfected cells were not treated withtunicamycin. Tunicamycin (2 �g/ml) was used to induce a stress response in HeLa-etKd cells (lane labeled as “Tun treated”) to create a positive control.The lane corresponding to cells untransfected with APLP2-FLAG or vector and not treated with tunicamycin is labeled as “Untreated.” Western blots oflysates of these cells were probed with an Ab recognizing the KDEL sequence (present on the stress proteins GrP94 and BiP, top panel), an Ab recognizing�-actin (middle panel), or an Ab recognizing APLP2 (bottom panel).
1982 APLP2 INFLUENCES Kd ENDOCYTOSIS
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from
4°C for 30 min, washed twice, and incubated with a PE-conjugated, Fc-specific F(ab�)2 portion of goat anti-mouse IgG (Jackson ImmunoResearch) at 4°C for 30 min. The cells were washed three times, resus-pended in BSA/azide/PBS, and analyzed on a FACSCalibur flow cytometer(BD Biosciences). Statistical analyses were done with the Cell Quest soft-ware (BD Biosciences).
Immunofluorescence analysis
To assess APLP2 association with MHC molecules endocytosed from theplasma membrane, cells were grown on glass cover slips, in some casestransiently transfected with APLP2-FLAG using Effectene (Qiagen), andincubated with anti-Kd Ab 34-1-2 at 37°C to allow endocytosis of cellsurface Kd and bound Ab. Any Ab still bound to cell surface Kd moleculeswas then removed by incubation in stripping buffer (0.5% acetic acid/500mM NaCl) for 90 s and the cells were fixed with 4% (vol/vol) parafor-maldehyde in PBS for 10 min. Fixed cells were incubated with anti-FLAGor anti-APLP2 rabbit antiserum prepared in staining solution (0.2% sapo-nin wt/vol/0.5% wt/vol BSA/PBS) for 1 h at room temperature. After 3PBS washes (5 min/wash), the cells were incubated with a fluorochrome-conjugated mixture of secondary Abs (Alexa Fluor 568 goat anti-rabbit Aband Alexa Fluor 488 goat anti-mouse Ab) in staining solution for 30 minat room temperature. After three washes in PBS (5 min/wash), the cellswere mounted for image analysis. For all immunofluorescence experi-ments, the images were obtained with a Zeiss LSM 5 Pascal confocalmicroscope, using a 63� 1.4 numerical aperture lens with appropriatefilters.
To ascertain whether the vesicles in which internalized Kd moleculesassociated with endogenous APLP2 were endosomes, HeLa-Kd cells (sta-bly expressing Kd) were transfected for 24 h with Rab5 or the constitutivelyactive Rab5 mutant Q79L (both GFP-tagged), grown on cover slips, and
incubated with anti-Kd Ab 34-1-2 at 37°C for 15 min to allow endocytosisof Kd. After 15 min, any 34-1-2 Ab still bound to cell surface Kd moleculeswas removed by incubation in stripping buffer for 90 s so that the subse-quent immunofluorescence analysis would focus only on the internalizedKd molecules. The cells were fixed with 4% (vol/vol) paraformaldehyde inPBS for 10 min and incubated with anti-APLP2 rabbit antiserum (preparedin saponin-containing staining solution) for 1 h at room temperature. After3 PBS washes, the cells were incubated with a fluorochrome-conjugatedmixture of secondary Abs (Alexa Fluor 568 goat anti-rabbit Ab and AlexaFluor 405 goat anti-mouse Ab) in staining solution for 30 min at roomtemperature. After three washes in PBS (5 min/wash), the cells weremounted for image analysis.
ResultsColocalization of APLP2 with folded Kd molecules inendosomal vesicles
Previous studies from our laboratory suggested that APLP2 down-regulated the cell surface expression of Kd (14). To investigatewhether the effect of APLP2 on Kd might be mediated through anendocytic mechanism, we analyzed whether APLP2 colocalizedwith Kd molecules that had been endocytosed from the plasmamembrane and with an endosomal marker. An Ab uptake assaywas performed with an anti-Kd Ab, using HeLa cells stably trans-fected with Kd and transiently transfected with Rab5-GFP orRab5Q79L-GFP. Rab5 is an endosomal protein, and Rab5Q79L isa GTP-locked, nonhydrolyzable Rab5 mutant that stimulates en-dosomal membrane fusion (38). Expression of Rab5Q79L results
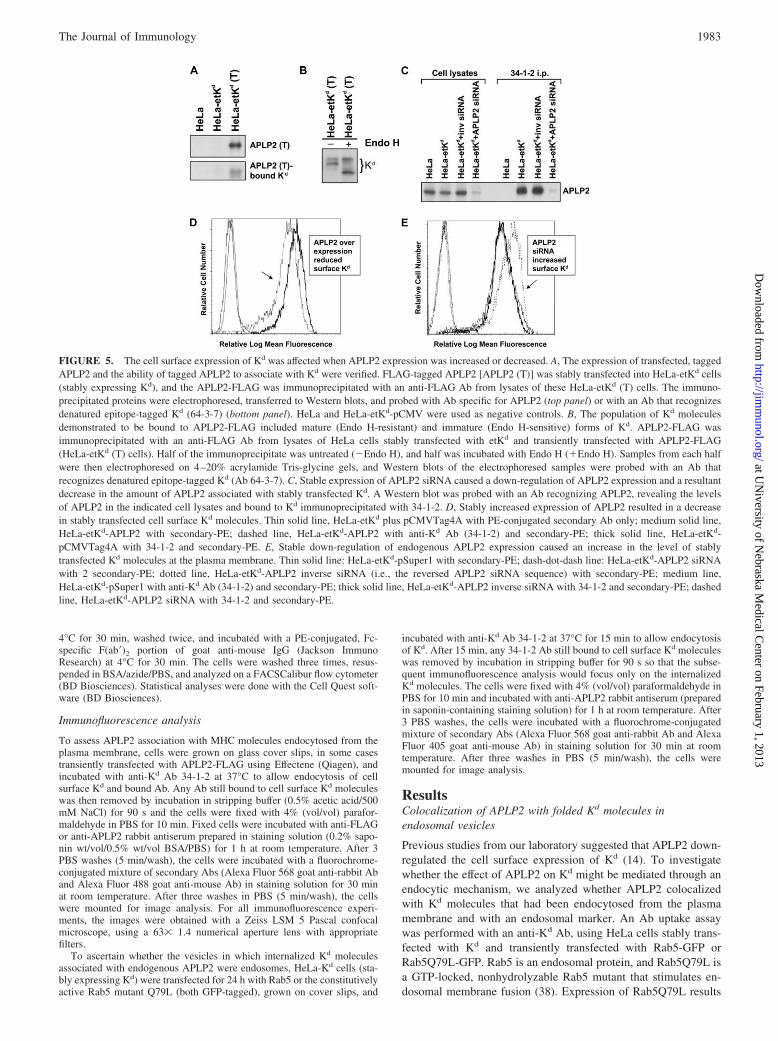
FIGURE 5. The cell surface expression of Kd was affected when APLP2 expression was increased or decreased. A, The expression of transfected, taggedAPLP2 and the ability of tagged APLP2 to associate with Kd were verified. FLAG-tagged APLP2 [APLP2 (T)] was stably transfected into HeLa-etKd cells(stably expressing Kd), and the APLP2-FLAG was immunoprecipitated with an anti-FLAG Ab from lysates of these HeLa-etKd (T) cells. The immuno-precipitated proteins were electrophoresed, transferred to Western blots, and probed with Ab specific for APLP2 (top panel) or with an Ab that recognizesdenatured epitope-tagged Kd (64-3-7) (bottom panel). HeLa and HeLa-etKd-pCMV were used as negative controls. B, The population of Kd moleculesdemonstrated to be bound to APLP2-FLAG included mature (Endo H-resistant) and immature (Endo H-sensitive) forms of Kd. APLP2-FLAG wasimmunoprecipitated with an anti-FLAG Ab from lysates of HeLa cells stably transfected with etKd and transiently transfected with APLP2-FLAG(HeLa-etKd (T) cells). Half of the immunoprecipitate was untreated (�Endo H), and half was incubated with Endo H (�Endo H). Samples from each halfwere then electrophoresed on 4–20% acrylamide Tris-glycine gels, and Western blots of the electrophoresed samples were probed with an Ab thatrecognizes denatured epitope-tagged Kd (Ab 64-3-7). C, Stable expression of APLP2 siRNA caused a down-regulation of APLP2 expression and a resultantdecrease in the amount of APLP2 associated with stably transfected Kd. A Western blot was probed with an Ab recognizing APLP2, revealing the levelsof APLP2 in the indicated cell lysates and bound to Kd immunoprecipitated with 34-1-2. D, Stably increased expression of APLP2 resulted in a decreasein stably transfected cell surface Kd molecules. Thin solid line, HeLa-etKd plus pCMVTag4A with PE-conjugated secondary Ab only; medium solid line,HeLa-etKd-APLP2 with secondary-PE; dashed line, HeLa-etKd-APLP2 with anti-Kd Ab (34-1-2) and secondary-PE; thick solid line, HeLa-etKd-pCMVTag4A with 34-1-2 and secondary-PE. E, Stable down-regulation of endogenous APLP2 expression caused an increase in the level of stablytransfected Kd molecules at the plasma membrane. Thin solid line: HeLa-etKd-pSuper1 with secondary-PE; dash-dot-dash line: HeLa-etKd-APLP2 siRNAwith 2 secondary-PE; dotted line, HeLa-etKd-APLP2 inverse siRNA (i.e., the reversed APLP2 siRNA sequence) with secondary-PE; medium line,HeLa-etKd-pSuper1 with anti-Kd Ab (34-1-2) and secondary-PE; thick solid line, HeLa-etKd-APLP2 inverse siRNA with 34-1-2 and secondary-PE; dashedline, HeLa-etKd-APLP2 siRNA with 34-1-2 and secondary-PE.
1983The Journal of Immunology
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from

in the formation of enlarged early endosomes that are accessible tointernalized cargo (38), facilitating colocalization analysis. In ourassay, we incubated nonpermeabilized cells with anti-Kd Ab 34-1-2 for 15 min at 37°C. After removal of noninternalized 34-1-2Ab with an acid wash, the cells were permeabilized and stainedwith Ab against APLP2. Colocalization of endogenous APLP2,internalized Kd, and Rab 5-GFP (Fig. 1A) or Rab5Q79L (Fig. 1B)was noticeable in the endosomes. Thus, APLP2 colocalizes withKd molecules that have been internalized from the cell surface andthat are present within endosomal vesicles.
APLP2 increased Kd endocytosis
We also examined the kinetics of the interaction of endogenousAPLP2 with Kd molecules endocytosed from the plasma mem-brane. Anti-Kd mAb 34-1-2 was added to label the cell surface Kd
molecules on HeLa-etKd cells (stably expressing Kd), and the cellswere incubated for varied amounts of time (0, 10, 20, or 30 min)at 37°C to allow internalization of Kd. The cells were then per-meabilized and incubated first with primary Ab against APLP2,washed, and incubated with secondary Abs recognizing the anti-Kd
and anti-APLP2 Abs. The 0 min time point is shown as evidenceof thorough stripping of noninternalized anti-Kd Ab (Fig. 2). Co-localization of endogenous APLP2 and internalized Kd was appar-ent by 10 min, and could still be visualized at 20 and 30 min(Fig. 2).
Kd was also colocalized with FLAG-tagged APLP2 (transientlyexpressed in HeLa-etKd cells) after Kd internalization from the cellsurface for 20 min (Fig. 3A). Confocal z-sectioning was done toconfirm that internalized Kd and APLP2-FLAG were present in thesame endocytic vesicles, and not merely within overlaid ones (Fig.3B). Furthermore, we demonstrated that APLP2 was bound to en-docytosed Kd molecules, as shown by isolation of internalized 34-1-2� Kd and demonstration of APLP2 coimmunoprecipitated withthe endocytosed Kd (Fig. 3C). In these experiments, 34-1-2 Ab wasincubated with HeLa-etKd cells transiently expressing APLP2-FLAG, the cells were warmed at 37°C for 20 min and then acidstripped and lysed, the samples were electrophoresed, and the 34-1-2-immunoprecipitated Kd and coimmunoprecipitated APLP2were identified by Western blotting. These data provide biochem-ical evidence for the binding of endocytosed Kd to APLP2.
Notably, from these confocal studies it could be seen that in-creased expression of APLP2 resulted in greater endocytosis of Kd.As shown in Fig. 3A, the Kd fluorescence in transiently APLP2-transfected cells (indicated by dashed lines) was greater than in thecells within the same field that were not transfected with APLP2.The graph shown in Fig. 3D displays the mean fluorescence in-tensity of internalized Kd in cells transiently expressing increased(FLAG-tagged) APLP2 versus cells that only expressed endoge-nous APLP2 (Fig. 3D). (The data shown in the graph are from the20-min point in the same Kd uptake assay for which confocal mi-croscopy results are shown in Fig. 3A.) Cells expressing the higherlevel of APLP2 had significantly more internalized Kd than didcells expressing endogenous levels of APLP2 (Fig. 3D). Thesefindings indicate that APLP2 not only associates with endocytosedKd, it potentiates its endocytosis. In contrast, elevated APLP2 ex-pression did not significantly increase endocytosis of a differentmouse MHC class I molecule, Ld (Figs. 4, A and B). APLP2 bindsLd more weakly than Kd and and has significantly less effect on thesurface expression of Ld than Kd (data not shown). The effect ofAPLP2 on Kd endocytosis was specific to the folded form of Kd;there was no increase in open (64-3-7�) etKd internalization whenAPLP2 was overexpressed (Fig. 4C). In addition, the effect onfolded Kd endocytosis was not seen when a different protein (trans-
ferrin receptor) instead of APLP2 was overexpressed in HeLa-Kd
cells (Fig. 4D).
Increased expression of APLP2 does not induce expression ofstress proteins
To confirm that expression of an increased amount of APLP2 didnot induce a cellular stress reaction, which could conceivably havesecondary effects on Kd endocytosis, we examined whether an in-crease in APLP2 expression led to heightened expression of stressproteins. To assess the stress response, we examined the cellularlevels of GrP94 and BiP following APLP2 overexpression. Thesestress proteins share a carboxy-terminal amino acid sequence(KDEL) that restricts them to the ER and, therefore, they can bedetected by an anti-KDEL Ab. HeLa cells stably expressing Kd
and untransfected with APLP2, transiently transfected with vectoronly, or transiently transfected with increased levels of APLP2were used in these experiments, along with HeLa-etKd cells thathad been treated with tunicamycin to induce a stress response (asa positive control). Western blots of lysates of these cell lines were
FIGURE 6. Overexpression of APLP2 decreased the stability of foldedKd molecules, and down-regulation of APLP2 expression increased thestability of folded Kd molecules. Cell lysates were incubated for 12 min onice or in a Biometra T3 gradient thermocycler (Whatman Biometra) at theindicated temperatures. After the thermocycler incubations, immunopre-cipitations with anti-folded Kd Ab 34-1-2 were performed. The immuno-precipitates were electrophoresed and probed on a Western blot with Ab64-3-7 to identify the tagged Kd H chain. Folded Kd was quantified bydensitometry, and is presented on the graphs as the relative percentage offolded Kd at each incubation temperature after setting the amount of foldedKd at 4°C as 100%. A, Increasing APLP2 expression by transfection ofAPLP2 into HeLa-etKd cells decreased folded Kd stability. The resultsillustrated in the graph were obtained with HeLa-etKd cells (stably ex-pressing Kd) that had been stably transfected with FLAG-tagged APLP2 inthe pCMVTag4A vector Œ, with the pCMVTag4A vector alone f, or withneither APLP2-pCMVTag4A nor pCMVTag4A F). B, Reduction ofAPLP2 expression by transfection of APLP2-siRNA into HeLa-etKd cellsimproved folded Kd stability. The cells used were HeLa-etKd cells (stablyexpressing Kd) that had been stably transfected with APLP2 siRNA in thepSuper vector �, with inverse siRNA (the reversed APLP2 siRNA se-quence) in the pSuper vector f, or the pSuper vector alone F. Error bars,SEM.
1984 APLP2 INFLUENCES Kd ENDOCYTOSIS
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from
probed with an Ab against the amino acid sequence KDEL, andproteins of the appropriate size to be GrP94 and BiP were detectedonly for the tunicamycin-treated positive control (Fig. 4E). Thus,increased expression of APLP2 does not induce expression of thestress proteins GrP94 and BiP.
Increased APLP2 expression decreases the quantity of Kd
molecules on the plasma membrane
Flow cytometric analysis of cells with stably overexpressed, aswell as stably down-regulated, APLP2 was performed to assess theability of APLP2 to regulate the amount of Kd expressed at the cellsurface. For these experiments, HeLa-etKd cells stably expressingFLAG-tagged APLP2 were generated, and the tagged APLP2’sexpression and ability to bind Kd were confirmed (Fig. 5A). ByEndo H assays, we determined that Kd coimmunoprecipitatingwith the transfected APLP2 included both mature (Endo H-resis-tant) and immature (Endo H-sensitive) forms (Fig. 5B). StableAPLP2 siRNA transfectants were also created (Fig. 5C). Overex-pression of APLP2 following transfection of FLAG-tagged APLP2reduced the amount of Kd present at the plasma membrane to�60% of the normal level (Fig. 5D). In contrast, HeLa-etKd cellsthat overexpressed the transferrin receptor had Kd expression lev-els at 99.6% of the normal level (data not shown). The conversewas also true, in that stable APLP2 siRNA transfectants expressedan increased level of Kd at the plasma membrane (�1.7-foldhigher than the level of Kd on the control cell lines) (Fig. 5E).These results indicate that the level of cellular expression ofAPLP2 influences the amount of folded Kd available at the cellsurface.
Kd thermostability and turnover was affected by the level ofAPLP2
To monitor for changes in Kd stability when the APLP2 level wasincreased, lysates of HeLa-etKd cells transfected with APLP2,with vector alone, or neither were incubated on ice or at 5°C tem-perature intervals between 25°C and 50°C. Following the incuba-tions, folded Kd molecules were immunoprecipitated, visualizedby probing the electrophoresed and transferred samples on a West-ern blot, and quantified by densitometry. The relative percentageof folded Kd at each incubation temperature (based on setting theamount of folded Kd at 4°C as 100%) was graphed. As shown inFig. 6A, in the presence of an increased level of APLP2 the sta-bility of folded Kd molecules was reduced. We also performed thesame type of experiment using lysates of HeLa-etKd cells trans-fected with APLP2 siRNA, inverse siRNA, or vector only. Thepresence of APLP2 siRNA increased the stability of Kd moleculesin the temperature range between 35°C and 50°C (Fig. 6B). Fur-thermore, by treating cells expressing normal or increased levels ofAPLP2 with cycloheximide for varied periods of time, we dem-onstrated that increased APLP2 causes a rise in the rate of Kd
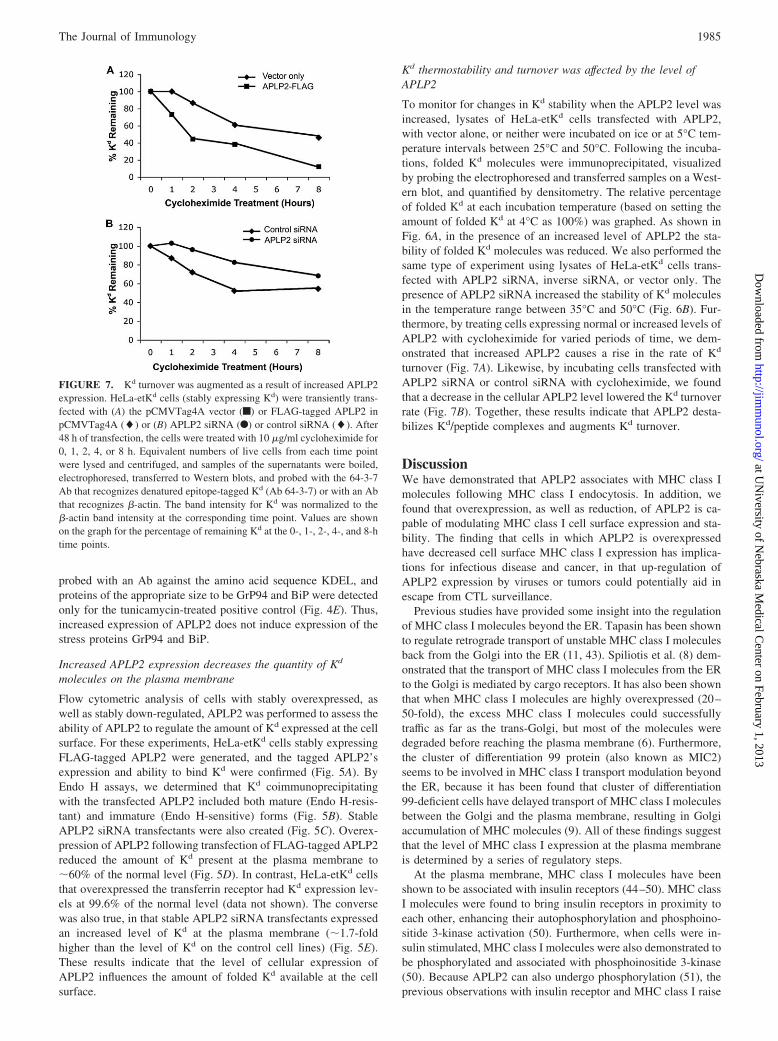
turnover (Fig. 7A). Likewise, by incubating cells transfected withAPLP2 siRNA or control siRNA with cycloheximide, we foundthat a decrease in the cellular APLP2 level lowered the Kd turnoverrate (Fig. 7B). Together, these results indicate that APLP2 desta-bilizes Kd/peptide complexes and augments Kd turnover.
DiscussionWe have demonstrated that APLP2 associates with MHC class Imolecules following MHC class I endocytosis. In addition, wefound that overexpression, as well as reduction, of APLP2 is ca-pable of modulating MHC class I cell surface expression and sta-bility. The finding that cells in which APLP2 is overexpressedhave decreased cell surface MHC class I expression has implica-tions for infectious disease and cancer, in that up-regulation ofAPLP2 expression by viruses or tumors could potentially aid inescape from CTL surveillance.
Previous studies have provided some insight into the regulationof MHC class I molecules beyond the ER. Tapasin has been shownto regulate retrograde transport of unstable MHC class I moleculesback from the Golgi into the ER (11, 43). Spiliotis et al. (8) dem-onstrated that the transport of MHC class I molecules from the ERto the Golgi is mediated by cargo receptors. It has also been shownthat when MHC class I molecules are highly overexpressed (20–50-fold), the excess MHC class I molecules could successfullytraffic as far as the trans-Golgi, but most of the molecules weredegraded before reaching the plasma membrane (6). Furthermore,the cluster of differentiation 99 protein (also known as MIC2)seems to be involved in MHC class I transport modulation beyondthe ER, because it has been found that cluster of differentiation99-deficient cells have delayed transport of MHC class I moleculesbetween the Golgi and the plasma membrane, resulting in Golgiaccumulation of MHC molecules (9). All of these findings suggestthat the level of MHC class I expression at the plasma membraneis determined by a series of regulatory steps.
At the plasma membrane, MHC class I molecules have beenshown to be associated with insulin receptors (44–50). MHC classI molecules were found to bring insulin receptors in proximity toeach other, enhancing their autophosphorylation and phosphoino-sitide 3-kinase activation (50). Furthermore, when cells were in-sulin stimulated, MHC class I molecules were also demonstrated tobe phosphorylated and associated with phosphoinositide 3-kinase(50). Because APLP2 can also undergo phosphorylation (51), theprevious observations with insulin receptor and MHC class I raise
FIGURE 7. Kd turnover was augmented as a result of increased APLP2expression. HeLa-etKd cells (stably expressing Kd) were transiently trans-fected with (A) the pCMVTag4A vector (f) or FLAG-tagged APLP2 inpCMVTag4A (�) or (B) APLP2 siRNA (F) or control siRNA (�). After48 h of transfection, the cells were treated with 10 �g/ml cycloheximide for0, 1, 2, 4, or 8 h. Equivalent numbers of live cells from each time pointwere lysed and centrifuged, and samples of the supernatants were boiled,electrophoresed, transferred to Western blots, and probed with the 64-3-7Ab that recognizes denatured epitope-tagged Kd (Ab 64-3-7) or with an Abthat recognizes �-actin. The band intensity for Kd was normalized to the�-actin band intensity at the corresponding time point. Values are shownon the graph for the percentage of remaining Kd at the 0-, 1-, 2-, 4-, and 8-htime points.
1985The Journal of Immunology
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from

the question of whether the interaction of APLP2 with MHC classI affects the phosphorylation of APLP2 or the MHC class Imolecule.
APLP2 is known to affect many intracellular pathways throughmechanisms which are as yet only poorly understood. Hence, it isconceivable that APLP2 may influence Kd expression and stabilityby modulating the action of chaperones in the MHC class I Agpathway. In this context, possible stress responses that might beinduced by APLP2 could be considered. However, our findings donot support any role for APLP2 in inducing stress responses, be-cause our data indicate that increased expression of APLP2 doesnot up-regulate expression of stress response markers (GrP94and BiP).
In our study, APLP2 was found to be associated with folded Kd
molecules endocytosed from the cell surface and to increase Kd
endocytosis. APLP2, and the closely related protein APP, havebeen shown to interact with the high-affinity choline transporter,and APP has been demonstrated to facilitate endocytosis of thistransporter, although whether APLP2 also facilitates its endocyto-sis was not examined (52). The association of APLP2 with Kd
molecules endocytosed from the cell surface suggests APLP2 mayinfluence Kd degradation and/or recycling. Our observation thatincreased APLP2 causes a rise in the turnover rate of Kd is con-sistent with APLP2 facilitating Kd degradation. Differing reportshave suggested that MHC class I molecules may be internalizedeither via clathrin-coated pits (53) or in a clathrin-independentmanner (54). A sequence within the cytoplasmic tail of MHC classI molecules has been shown to be required for their endocytosis(55). MHC class I molecules expressed by B lymphoblastoid cellsare continually endocytosed and recycled to the cell surface (56).The mechanism of MHC class I recycling appears to involve theEps15 homology domain-containing protein (EHD1). EHD1 in-duces the formation of tubules which contain internalized MHCclass I molecules, and overexpression of EHD1 up-regulates MHCclass I recycling (57), whereas siRNA down-regulation of EHD1delays the rate of MHC class I recycling (58).
Because APLP2 can associate with folded MHC class I mole-cules after their endocytosis, it may destabilize MHC/peptide com-plexes within the endocytic compartment. Following endocytosis,proteins can undergo deglycosylation and degradation (59). De-stabilization by APLP2 could facilitate MHC class I turnover, andperhaps also MHC class I rebinding of new peptides within thecell. Overall, our data support a model in which APLP2 binds toendocytosed Kd molecules and regulates Kd endocytosis and sta-bility, thereby modulating Kd cell surface expression.
Note added in proof. During the revision and resubmission ofthis manuscript, a comparative analysis of the variation in APLP2binding, colocalization, and effect among mouse MHC class I al-lotypes was reported (60).
AcknowledgmentsWe thank Dr. Ted Hansen and Dr. Wendy Maury for gifts of Abs and celllines, Dr. Robert Lodge for gifts of DNA constructs, and Dr. Kay-UweWagner for assistance with the thermocycler. We also gratefully acknowl-edge the assistance of Haley Capek, Vivek Gautam, Daniel McDermott,Miriam Menezes, and Phon T. Nguyen, and the personnel of the Universityof Nebraska Medical Center Cell Analysis Facility, mAb Facility, and Mo-lecular Biology Facility.
DisclosuresThe authors have no financial conflict of interest.
References1. Cresswell, P., N. Bangia, T. Dick, and G. Diedrich. 1999. The nature of the MHC
class I peptide loading complex. Immunol. Rev. 172: 21–28.
2. Paquet, M. E., M. Cohen-Doyle, G. C. Shore, and D. B. Williams. 2004.Bap29/31 influences the intracellular traffic of MHC class I molecules. J. Immu-nol. 172: 7548–7555.
3. Park, B., S. Lee, E. Kim, K. Cho, S. R. Riddell, S. Cho, and K. Ahn. 2006. Redoxregulation facilitates optimal peptide selection by MHC class I during antigenprocessing. Cell 127: 369–382.
4. Serwold, T., F. Gonzalez, J. Kim, R. Jacon, and N. Shastri. 2002. ERAAP cus-tomizes peptides for MHC class I molecules in the endoplasmic reticulum. Na-ture 419: 480–483.
5. Saric, T., S. C. Chang, A. Hattori, I. A. York, S. Markant, K. L. Rock,M. Tsujimoto, and A. L. Goldberg. 2002. An IFN-�-induced aminopeptidase inthe ER, ERAP1, trims precursors to MHC class I-presented peptides. Nat. Im-munol. 3: 1169–1176.
6. Joyce, S. 1997. Traffic control of completely assembled MHC class I moleculesbeyond the endoplasmic reticulum. J. Mol. Biol. 266: 993–1001.
7. Marguet, D., E. T. Spiliotis, T. Pentcheva, M. Lebowitz, J. Schneck, andM. Edidin. 1999. Lateral diffusion of GFP-tagged H2Ld molecules and of GFP-TAP1 reports on the assembly and retention of these molecules in the endoplas-mic reticulum. Immunity 11: 231–240.
8. Spiliotis, E. T., H. Manley, M. Osorio, M. C. Zuniga, and M. Edidin. 2000.Selective export of MHC class I molecules from the ER after their dissociationfrom TAP. Immunity 13: 841–851.
9. Sohn, H. W., Y. K. Shin, I. S. Lee, Y. M. Bae, Y. H. Suh, M. K. Kim, T. J. Kim,K. C. Jung, W. S. Park, C.-S. Park, et al. CD99 regulates the transport of MHCclass I molecules from the Golgi complex to the cell surface. J. Immunol. 166:787–794.
10. Pencheva, T., and M. Edidin. 2001. Clustering of peptide-loaded MHC class Imolecules for endoplasmic reticulum export imaged by fluorescence resonanceenergy transfer. J. Immunol. 166: 6625–6632.
11. Paulsson, K. M., M. J. Kleijmeer, J. Griffith, M. Jevon, S. Chen, P. O. Anderson,H. O. Sjogren, S. Li, and P. Wang. 2002. Association of tapasin and COPIprovides a mechanism for the retrograde transport of major histocompatibilitycomplex (MHC) class I molecules from the Golgi complex to the endoplasmicreticulum. J. Biol. Chem. 277: 18266–18271.
12. Feuerbach, D., and H.-G. Burgert. 1993. Novel proteins associated with MHCclass I antigens in cells expressing adenovirus protein E3/19K. EMBO J. 12:3153–3161.
13. Sester, M., D. Feuerbach, R. Frank, T. Preckel, A. Gutermann, and H.-G. Burgert.2000. The amyloid precursor-like protein 2 associates with the major histocom-patibility complex class I molecule Kd. J. Biol. Chem. 275: 3645–3654.
14. Morris, C. R., J. L. Petersen, S. E. Vargas, H. R. Turnquist, M. M. McIlhaney,S. D. Sanderson, J. T. Bruder, Y. Y. Yu, H.-G. Burgert, and J. C. Solheim. 2003.The amyloid precursor-like protein 2 and the adenoviral E3/19K protein bothbind to a conformational site on H-2Kd and regulate H-2Kd expression. J. Biol.Chem. 278: 12618–12623.
15. Slunt, H. H., G. Thinakaran, C. Von Koch, A. C. Lo, R. E. Tanzi, andS. S. Sisodia. 1994. Expression of a ubiquitous, cross-reactive homologue of themouse �-amyloid precursor protein. J. Biol. Chem. 269: 2637–2644.
16. Thinakaran, G., C. A. Kitt, A. J. Roskams, H. H. Slunt, E. Masliah, C. von Koch,S. D. Ginsberg, G. V. Ronnett, R. R. Reed, and D. L. Price. 1995. Distribution ofan APP homolog, APLP2, in the mouse olfactory system: a potential role forAPLP2 in axogenesis. J. Neurosci. 15: 6314–6326.
17. Rassoulzadegan, M., Y. Yang, and F. Cuzin. 1998. APLP2, a member of theAlzheimer precursor protein family, is required for correct genomic segregationin dividing mouse cells. EMBO J. 17: 4647–4656.
18. Guo, J., G. Thinakaran, Y. Guo, S. S. Sisodia, and F. X. Yu. 1998. A role foramyloid precursor-like protein 2 in corneal epithelial wound healing. Invest. Oph-thalmol. Visual Sci. 39: 292–300.
19. Cappai, R., S. S. Mok, D. Galatis, D. F. Tucker, A. Henry, K. Beyreuther,D. H. Small, and C. L. Masters. 1999. Recombinant human amyloid precursor-like protein 2 (APLP2) expressed in the yeast Pichia pastoris can stimulate neu-rite outgrowth. FEBS Lett. 442: 95–98.
20. Li, X. F., G. Thinakaran, S. S. Sisodia, and F. S. Yu. 1999. Amyloid precursor-like protein 2 promotes cell migration toward fibronectin and collagen IV. J. Biol.Chem. 274: 27249–27256.
21. Glenner, G. G., and C. W. Wong. 1984. Alzheimer’s disease: initial report of thepurification and characterization of a novel cerebrovascular amyloid. Biochem.Biophys. Res. Commun. 120: 885–890.
22. Masters, C. L., G. Simms, N. A. Weinman, G. Multhaup, B. L. McDonald,K. Beyreuther. 1985. Amyloid plaque core protein in Alzheimer disease andDown syndrome. Proc. Natl. Acad. Sci. USA 82: 4245–4249.
23. Ozato, K., G. A. Evans, B. Shykind, D. Margulies, and J. G. Seidman. 1983.Hybrid H-2 histocompatibility gene products assign domains recognized by al-loreactive T cells. Proc. Natl. Acad. Sci. USA 80: 2040–2043.
24. Nieto, M., E. S. Song, D. McKinney, M. McMillan, and R. S. Goodenow. 1989.The association of H-2Ld with human �-2 microglobulin induces localized con-formational changes in the �-1 and -2 superdomain. Immunogenetics 30:361–369.
25. Solheim, J. C., B. M. Carreno, N. B. Myers, D. R. Lee, and T. H. Hansen. 1995.Peptide-induced rescue of serologic epitopes on class I MHC molecules. J. Im-munol. 154: 1188–1197.
26. Smith, J. D., N. B. Myers, J. Gorka, and T. H. Hansen. 1993. Model for the invivo assembly of nascent Ld class I molecules and for the expression of unfoldedLd molecules at the cell surface. J. Exp. Med. 178: 2035–2046.
27. Yu, Y. Y., N. B. Myers, C. H. Hilbert, M. R. Harris, G. K. Balendiran, andT. H. Hansen. 1999. Definition and transfer of a serological epitope specific forpeptide-empty forms of MHC class I. Int. Immunol. 11: 1897–1906.
1986 APLP2 INFLUENCES Kd ENDOCYTOSIS
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from

28. Myers, N. B., M. R. Harris, J. M. Connolly, L. Lybarger, Y. Y. Yu, andT. H. Hansen. 2000. Kb, Kd, and Ld molecules share common tapasin dependen-cies as determined using a novel epitope tag. J. Immunol. 165: 5656–5663.
29. Harris, M. R., L. Lybarger, N. B. Myers, C. Hilbert, J. C. Solheim, T. H. Hansen,and Y. Y. Yu. 2001. Interactions of HLA-B27 with the peptide loading complexas revealed by heavy chain mutations. Int. Immunol. 13: 1275–1282.
30. Lybarger, L., Y. Y. Yu, T. Chun, C.-R. Wang, A. G. Grandea III, L. Van Kaer,and T. H. Hansen. 2001. Tapasin enhances peptide-induced expression of H2–M3molecules, but is not required for the retention of open conformers. J. Immunol.167: 2097–2105.
31. Ozato, K., T. H. Hansen, and D. Sachs. 1980. Monoclonal antibodies to mouseMHC antigens. II. Antibodies to the H-2Ld antigen, the product of a third poly-morphic locus of the mouse major histocompatibility complex. J. Immunol. 125:2473–2477.
32. Evans, G. A., D. H. Margulies, B. Shykind, J. G. Seidman, and K. Ozato. 1982.Exon shuffling: mapping polymorphic determinants on hybrid mouse transplan-tation antigens. Nature 300: 755–757.
33. Solheim, J. C., B. M. Carreno, J. D. Smith, J. Gorka, N. B. Myers, Z. Wen,J. M. Martinko, D. R. Lee, and T. H. Hansen. 1993. Binding of peptides lackingconsensus anchor anchor residue alters H-2Ld serologic recognition. J. Immunol.151: 5387–5397.
34. Smith, J. D., J. C. Solheim, B. M. Carreno, and T. H. Hansen. 1995. Character-ization of class I MHC folding intermediates and their disparate interactions withpeptide and �2-microglobulin. Mol. Immunol. 32: 531–540.
35. Ozato, K., and D. H. Sachs. 1981. Monoclonal antibodies to mouse MHC anti-gens. III. Hybridoma antibodies reacting to antigens of the H-2b haplotype revealgenetic control of isotype expression. J. Immunol. 126: 317–321.
36. Long, E. O., S. Rosen-Bronson, D. R. Karp, M. Malnati, R. P. Sekaly, andD. Jaraquemada. 1991. Efficient cDNA expression vectors for stable and transientexpression of HLA-DR in transfected fibroblasts and lymphoid cells. Hum. Im-munol. 31: 229–235.
37. Zerial, M., P. Melancon, C. Schneider, and H. Garoff. 1986. The transmembranesegment of the human transferrin receptor functions as a signal peptide. EMBOJ. 5: 1543–1550.
38. Stenmark, H., R. G. Parton, O. Steele-Mortimer, A. Lutcke, J. Gruenberg, andM. Zerial. 1994. Inhibition of rab5 GTPase activity stimulates membrane fusionin endocytosis. EMBO J. 13: 1287–1296.
39. Turnquist, H. R., and J. C. Solheim. 2001. Analysis of MHC class I interactionswith endoplasmic reticulum proteins. Methods Mol. Biol. 156: 165–173.
40. Rinderknecht, C. H., M. P. Belmares, T. L. Catanzarite, A. J. Bankovich,T. H. Holmes, K. C. Garcia, N. K. Nanda, R. Busch, S. Kovats, and E. D. Mellins.2007. Posttranslational regulation of I-Ed by affinity for CLIP. J. Immunol. 279:5907–5915.
41. Sato, T., S. Susuki, M. A. Suico, M. Miyata, Y. Ando, M. Mizuguchi,M. Takeuchi, M. Dobashi, T. Shuto, and H. Kai. 2007. Endoplasmic reticulumquality control regulates the fate of transthyretin variants in the cells. EMBO J.26: 2501–2512.
42. Williams, A. P., C. A. Peh, A. W. Purcell, J. McCluskey, and T. Elliott. 2002.Optimization of the MHC class I peptide cargo is dependent on tapasin. Immunity16: 509–520.
43. Paulsson, K. M., M. Jevon, J. W. Wang, S. Li, and P. Wang. 2006. The doublelysine motif of tapasin is a retrieval signal for retention of unstable MHC class Imolecules in the endoplasmic reticulum. J. Immunol. 176: 7482–7488.
44. Fehlmann, M., Y. Chvatchko, D. Brandenburg, E. van Obberghen, andN. Brossette. 1985. The subunit structure of the insulin receptor and molecularinteractions with the major histocompatibility complex antigens. Biochimie 67:1155–1159.
45. Fehlmann, M., J.-F. Peyron, M. A. Samson, E. van Obberghen, D. Brandenburg,and N. Brossette. 1985. Molecular association between major histocompatibilitycomplex class I antigens and insulin receptor in mouse liver membranes. Proc.Natl. Acad. Sci. USA 82: 8634–8637.
46. Due, C., M. Simonsen, and L. Olsson. 1986. The major histocompatibility com-plex class I heavy chain as a structural subunit of the human cell membraneinsulin receptor: implication for the range of biological functions of histocom-patibility antigens. Proc. Natl. Acad. Sci. USA 83: 6007–6011.
47. Phillips, M. L., M. L. Moule, T. L. Delovitch, and C. C. Yip. 1986. Class Ihistocompatibility antigens and insulin receptors: evidence for interactions. Proc.Natl. Acad. Sci. USA 83: 3474–3478.
48. Kittur, D., Y. Shimizu, R. DeMars, and M. Edidin. 1987. Insulin binding tohuman B lymphoblasts is a function of HLA haplotype. Proc. Natl. Acad. Sci.USA 84: 1351–1355.
49. Edidin, M., and J. Reiland. 1990. Dynamic measurements of the associationsbetween class I MHC antigens and insulin receptors. Mol. Immunol. 27:1313–1317.
50. Ramalingam, T. S., A. Chakrabarti, and M. Edidin. 1997. Interaction of class Ihuman leukocyte antigens (HLA-I) molecules with insulin receptors and its effecton the insulin-signaling cascade. Mol. Biol. Cell 8: 2463–2474.
51. Taru, H., and T. Suzuki. 2004. Facilitation of stress-induced phosphorylation of�-amyloid precursor protein family members by X11-like/Mint2 protein. J. Biol.Chem. 279: 21628–21636.
52. Wang, B., L. Yang, Z. Wang, and H. Zheng. 2007. Amyloid precursor proteinmediates presynaptic localization and activity of the high-affinity choline trans-porter. Proc. Natl. Acad. Sci. USA 104: 14140–14145.
53. Dasgupta, J. D., S. Watkins, H. Slayter, and E. J. Yunis. 1988. Receptor-likenature of class I HLA: endocytosis via coated pits. J. Immunol. 141: 2577–2580.
54. Radhakrishna, H., and J. G. Donaldson. 1997. ADP-ribosylation factor 6 regu-lates a novel plasma membrane recycling pathway. J. Cell Biol. 139: 49–61.
55. Vega, M. A., and J. L. Strominger. 1989. Constitutive endocytosis of HLA classI antigens requires a specific portion of the intracytoplasmic tail that shares struc-tural features with other endocytosed molecules. Proc. Natl. Acad. Sci. USA 86:2688–2692.
56. Reid, P. A., and C. Watts. 1990. Cycling of cell-surface MHC glycoproteinsthrough primaquine-sensitive intracellular compartments. Nature 346: 655–657.
57. Caplan, S., N. Naslavsky, L. M. Hartnell, R. Lodge, R. S. Polishchuk,J. G. Donaldson, and J. S. Bonifacino. 2002. A tubular EHD1-containing com-partment involved in the recycling of major histocompatibility complex class Imolecules to the plasma membrane. EMBO J. 21: 2557–2567.
58. Naslavsky, N., M. Boehm, P. S. Backlund, Jr., and S. Caplan. 2004. Rabenosyn-5and EHD1 interact and sequentially regulate protein recycling to the plasmamembrane. Mol. Biol. Cell 15: 2410–2422.
59. Schmitt, M., and T. Grand-Perret. 1999. Regulated turnover of a cell surface-associated pool of newly synthesized apolipoprotein E in HepG2 cells.J. Lipid Res. 40: 39–49.
60. Tuli, A., M. Sharma, X.Wang, N. Naslavsky, S. Caplan, and J. C. Solheim. 2008.Specificity of amyloid precursor-like protein 2 interactions with MHC class Imolecules. Immunogenetics 60: 303–313.
1987The Journal of Immunology
at UN
iversity of Nebraska M
edical Center on February 1, 2013
http://jimm
unol.org/D
ownloaded from




















